molybdenum-containing nicotine hydroxylase genes …aem.asm.org/content/81/24/8330.full.pdf ·...
TRANSCRIPT

Molybdenum-Containing Nicotine Hydroxylase Genes in a NicotineDegradation Pathway That Is a Variant of the Pyridine andPyrrolidine Pathways
Hao Yu,a,b Hongzhi Tang,a Yangyang Li,a Ping Xua
State Key Laboratory of Microbial Metabolism and School of Life Sciences and Biotechnology, Shanghai Jiao Tong University, Shanghai, People’s Republic of Chinaa;College of Life Sciences, Qingdao Agricultural University, Qingdao, People’s Republic of Chinab
Ochrobactrum sp. strain SJY1 utilizes nicotine as a sole source of carbon, nitrogen, and energy via a variant of the pyridine and pyrrol-idine pathways (the VPP pathway). Several strains and genes involved in the VPP pathway have recently been reported; however, thefirst catalyzing step for enzymatic turnover of nicotine is still unclear. In this study, a nicotine hydroxylase for the initial hydroxylationstep of nicotine degradation was identified and characterized. The nicotine hydroxylase (VppA), which converts nicotine to 6-hy-droxynicotine in the strain SJY1, is encoded by two open reading frames (vppAS and vppAL [subunits S and L, respectively]). The vppAgenes were heterologously expressed in the non-nicotine-degrading strains Escherichia coli DH5� and Pseudomonas putidaKT2440; only the Pseudomonas strain acquired the ability to degrade nicotine. The small subunit of VppA contained a [2Fe-2S]cluster-binding domain, and the large subunit of VppA contained a molybdenum cofactor-binding domain; however, an FAD-binding domain was not found in VppA. Resting cells cultivated in a molybdenum-deficient medium had low nicotine transfor-mation activity, and excess molybdenum was detected in the purified VppA by inductively coupled plasma-mass spectrometryanalysis. Thus, it is demonstrated that VppA is a two-component molybdenum-containing hydroxylase.
Nicotine is the main toxic compound in tobacco, and it accu-mulates with tobacco wastes during the processing of tobacco
products. Nicotine can easily spread into the environment due toits water solubility and endanger the health of humans and otherorganisms (1). Microbial biodegradation is one of the best reme-diation strategies to remove nicotine from the environment (2–4).A number of bacteria and fungi which use nicotine as a sole sourceof carbon, nitrogen, and energy for their growth have been iso-lated and identified (3, 4). Three nicotine degradation pathwayshave been proposed in bacteria based on the identification of in-termediates: the pyridine pathway (3), the pyrrolidine pathway (2,5), and the variant of pyridine and pyrrolidine pathways (the VPPpathway) (Fig. 1) (6–9). The molecular mechanisms of nicotinedegradation by the pyridine pathway and the pyrrolidine pathwayhave been elucidated in detail by the research groups of Brandsch(3) and Xu et al. (2, 10, 11), respectively. However, the genesinvolved in several catalyzing steps of the VPP pathway are stillunknown.
Thus far, several bacteria that are able to degrade nicotine viathe VPP pathway have been reported, including Ochrobactrum sp.strain SJY1 (6, 7), Agrobacterium tumefaciens S33 (12), Shinella sp.strain HZN7 (8), and a Pusillimonas strain (13). In the VPP path-way, nicotine degradation is initiated by a hydroxylation reactionto form 6-hydroxynicotine (6HN), which is then converted to6-hydroxy-N-methylmyosmine (6HMM) and 6-hydroxypseu-dooxynicotine (6HPON). Subsequently, 6HPON is oxidized toform 6-hydroxy-3-succinoylpyridine (HSP), which is catabolizedto fumaric acid through 2,5-dihydroxypyridine (2,5-DHP), N-formylmaleamic acid, maleamic acid, and maleic acid (Fig. 1) (2).In an early study, Ochrobactrum sp. strain SJY1 was isolated, and a97.6-kb gene cluster (the vpp cluster, GenBank accession numberKM065745), which is responsible for nicotine degradation, wascharacterized (7). Six genes (vppBDEFGH) involved in the VPPpathway were identified in the vpp cluster, and three of them were
functionally characterized using in vitro experiments. VppB cata-lyzes the reaction from 6HN to 6HMM and 6HPON; VppD isresponsible for the reaction from HSP to 2,5-DHP; and VppE is a2,5-DHP dioxygenase (Fig. 1). Additionally, an HSP hydroxylase(GenBank accession number KJ129609) that catalyzes the oxida-tive decarboxylation of HSP to 2,5-DHP has been identified inAgrobacterium tumefaciens S33 (9). More recently, the function ofnctB (GenBank accession number AGS16700), a gene that en-codes (S)-6-hydroxynicotine oxidase, in Shinella sp. HZN7, wasalso investigated (14). However, the gene responsible for nicotinehydroxylation in the VPP pathway is still unknown.
Hydroxylation of the � position of the pyridine ring with theformation of 6HN is the first catalyzing step in the VPP pathway ofnicotine degradation (Fig. 1). Pyridine �-position hydroxylationreactions are essential for the degradation of pyridine derivatives(15, 16), and these kinds of reactions are typically catalyzed by afamily of bacterial molybdenum-containing hydroxylases withsimilar subunit structures, using molybdopterin dinucleotide,FAD, and [2Fe-2S] clusters as cofactors (17). Three pyridine�-position hydroxylases have been identified in nicotine degra-
Received 16 July 2015 Accepted 18 September 2015
Accepted manuscript posted online 25 September 2015
Citation Yu H, Tang H, Li Y, Xu P. 2015. Molybdenum-containing nicotinehydroxylase genes in a nicotine degradation pathway that is a variant of thepyridine and pyrrolidine pathways. Appl Environ Microbiol 81:8330 –8338.doi:10.1128/AEM.02253-15.
Editor: M. Kivisaar
Address correspondence to Hongzhi Tang, [email protected], orPing Xu, [email protected].
Supplemental material for this article may be found at http://dx.doi.org/10.1128/AEM.02253-15.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
8330 aem.asm.org December 2015 Volume 81 Number 24Applied and Environmental Microbiology
on August 18, 2018 by guest
http://aem.asm
.org/D
ownloaded from

dation: nicotine dehydrogenase (Ndh) (18, 19) and ketone dehy-drogenase (Kdh) (19, 20) from the pyridine pathway, and 3-succinoylpyridine dehydrogenase (Spm) (2) from the pyrrolidinepathway (Fig. 1). These enzymes are all composed of three sub-units; the small and middle subunits correspond to the [2Fe-2S]cluster and the FAD-binding motif, and the large subunit cor-responds to the molybdopterin-binding subunit. Molybdenum-containing enzymes are found in nearly all organisms and repre-sent a large growing class of enzymes that catalyze redox reactionsby taking advantage of the versatile redox chemistry of molybde-num. The molybdenum-containing hydroxylases use H2O insteadof O2 as the electron donor and are usually involved in the metab-olism of medicines (17). Molybdenum-containing hydroxylasesplay important roles in the biodegradation of aromatic N-hetero-cycles, such as nicotinic acid and nicotine (2, 15).
In the present study, the genes responsible for the first enzymaticcatalyzing step of nicotine degradation in Ochrobactrum sp. SJY1were cloned and characterized. VppA, the nicotine hydroxylasefor the hydroxylation of nicotine to 6HN in the VPP pathway, iscomposed of two subunits, a [2Fe-2S]-binding subunit and a mo-lybdenum cofactor (MoCo)-binding subunit, suggesting thatVppA is a two-component molybdenum-containing hydroxylase.
MATERIALS AND METHODSChemicals and media. L-(�)-Nicotine (99% purity) was purchased fromFluka Chemie GmbH (Buchs Corp., Switzerland). 6-Hydroxynicotinewas obtained from Roderich Brandsch. NAD(P)H, FAD, 2,6-dichloro-phenolindophenol sodium (DCIP), and phenazine ethosulfate (PES)were obtained from Sangon Biotech, Shanghai, China. The 6-hydroxynico-tine oxidase was purified according to the method described previously (7).All other reagents and solvents were of analytical or chromatographic gradeand were commercially available. Luria-Bertani (LB) medium and minimalsalt medium (MSM) were prepared as previously described (21). Molybde-num deficiency minimal salt medium (MoDM) was prepared using double-distilled H2O (ddH2O) by removing the molybdenum from MSM.
Bacterial strains, plasmids, and growth conditions. The bacterialstrains and plasmids used in this study are listed in Table 1. Escherichia colistrains were grown in LB medium at 37°C. Ochrobactrum sp. SJY1 andPseudomonas putida strains were grown at 30°C in LB medium (7) orMSM with different carbon and nitrogen sources (21).
Cloning and expression of the vppA genes and biotransformation ofnicotine. Genes with different open reading frames (ORFs) were ampli-fied from genomic DNA of Ochrobactrum sp. SJY1 by PCR using primersvppA-F/vppA-R (for cloning of vppAS and vppAL) and vppAplus-F/vppAplus-R (for cloning of vppAS, vppAL, and ORF3) (Table 1). The genefragment and plasmid pME6032 (22) were digested with the restriction
FIG 1 The pyridine pathway, VPP pathway, and pyrrolidine pathway of nicotine degradation in bacteria. The pyridine pathway is shown in the light greenportion at the top, and the enzymes reported in Arthrobacter nicotinovorans are indicated by green text. The VPP pathway is shown in the light pink portion inthe middle, and the enzymes reported in Ocrobactrum sp. SJY1 are indicated by red text. The pyrrolidine pathway is shown in the light blue portion at the bottom,and the enzymes reported in Pseudomonas putida S16 are indicated by blue text.
A Nicotine Hydroxylase in the VPP Pathway
December 2015 Volume 81 Number 24 aem.asm.org 8331Applied and Environmental Microbiology
on August 18, 2018 by guest
http://aem.asm
.org/D
ownloaded from

enzymes EcoRI and XhoI and ligated just downstream of the promoter ofthe vector to generate pME6032-vppA and pME6032-vppAplus, respec-tively (Table 1). Plasmids pME6032-vppA and pME6032-vppAplus weretransferred into P. putida KT2440 (23) to generate P. putida KTVppAand P. putida KTVppAplus, respectively. Plasmids were transferred intothe recipient strain by the Gene Pulser Xcell system (Bio-Rad, USA) ac-cording to the present protocol for Pseudomonas aeruginosa. The expres-sion of VppA was induced by adding 1 mM isopropyl-�-thiogalactopyra-noside (IPTG) and 25 �g/ml tetracycline (Tet) during the cultivation. Thecells were harvested by centrifugation at 6,000 � g for 5 min and washedtwice with 0.9% NaCl, and the cell pellets were resuspended with 50 mMphosphate buffer (pH 7.0) to an optical density at 600 nm (OD600) of 6.0(resting cells). Resting cells were determined by their ability for nicotinetransformation. Nicotine was added at final concentrations of 1 mg/ml inthe resting cells with shaking at 120 rpm under 30°C. Samples were takenat certain time intervals and subjected to spectrum scanning and high-pressure liquid chromatography (HPLC) analysis.
RT-PCR and RT-qPCR. Ochrobactrum sp. strain SJY1 was cultivatedin the presence and absence of nicotine as previously described (7). TotalRNA was extracted with the RNAprep pure cell/bacteria kit (TianGen,China) (7). The cDNA was prepared using random hexamers (TaKaRa,Japan) and SuperScript III reverse transcriptase (Invitrogen) according tothe manufacturers’ instructions. The boundaries of the vppA operon wereidentified by PCR of the intergenic regions between the adjacent genesusing cDNA as a template. Real-time reverse transcription-PCR (RT-qPCR) analysis was performed using the CFX96 real-time PCR detection
system (Bio-Rad, Hercules, CA) with RealMasterMix (SYBR green)(TianGen, China), using the following program: 2 min of initial denatur-ation at 94°C followed by 40 cycles of 20 s of denaturation at 94°C, 20 s ofprimer annealing at 56°C (target gene) or 60°C (16S rRNA gene), and 20s of elongation at 72°C. Fluorescence was measured at the end of eachcycle, and melting curve analysis was performed at the end of the PCR toverify the product specificity. Primers for reverse transcription-PCR (RT-PCR) and RT-qPCR are listed in Table 1.
Purification and assays of VppA. The vppA genes were amplified byPCR using primers vppA-His-F/vppA-R, and the PCR product was used asthe template for the overlap extension PCR using primers vppA-His-F2/vppA-R. The product of overlap extension PCR was digested and theninserted into plasmid pME6032 to generate pME6032-vppA-His. P.putida KT2440 carrying pME6032-vppA-His (P. putida KTVppA-His)was grown at 30°C to an OD600 of 0.8 and induced at 25°C with 0.5 mMIPTG for 12 h. The cells were harvested, washed twice with 25 mM Tris-HCl (pH, 8.0), and then resuspended in the same buffer. The cell suspen-sion was broken by repetitive sonication, and the cell debris was removedby centrifugation at 12,000 � g for 20 min. The supernatant was loadedonto a HisTrap HP column, washed with 25 mM Tris-HCl (pH 8.0), andthen eluted with 25 mM Tris-HCl (pH 8.0) with 40 mM imidazole. Theeluted fractions were collected, and imidazole was removed by ultrafiltra-tion with a 10-kDa-cutoff-size Millipore filter. VppA activity was mea-sured at 600 nm using a UV-2550 spectrophotometer (Shimadzu, Kyoto,Japan) with the addition of nicotine, protein sample, and a different cofactor(NADH, PES, or DCIP). The products of enzyme catalytic reactions were
TABLE 1 Strains, plasmids, and primers used in this study
Strain, plasmid, orprimer Description or primer sequencea Source
StrainsOchrobactrum sp. SJY1 Nicotine-degrading strain, Gram negative CCTCC 2014136Pseudomonas putida
KT2440 Metabolically versatile saprophytic soil bacterium 23KT6032 KT2440 containing pME6032 This studyKTVppA KT2440 containing pME6032-vppA This studyKTVppAplus KT2440 containing pME6032-vppAplus This studyKTVppA-His KT2440 containing pME6032-vppA-His This study
Escherichia coliDH5� F� recA1 endA1 thi-1 hsdR17 supE44 relA1 deoR�(lacZYA-argF)U169 �80dlacZ�M15 TransGen, ChinaDHVppA DH5�, containing pME6032-vppA This study
PlasmidspME6032 Tetr, shuttle vector 22pME6032-vppA Tetr, pME6032 containing vppAS and vppAL This studypME6032-vppAplus Tetr, pME6032 containing vppAS, vppAL, and ORF3 This studypME6032-vppA-His Tetr, pME6032 containing vppAS and vppAL with a His tag sequence on the N terminus of vppAS This study
Primers This studyvppA-F ccgGAATTCatgaaagtcgattttactgttaa This studyvppA-R gtgCTCGAGcaatacagtaggaagcaggag This studyvppAplus-F ccgGAATTCatgaaagtcgattttactgttaa This studyvppAplus-R gtgCTCGAGtatttatgcctgggctccgtgct This studyvppA-His-F atcatcatcatcatcacagcagcaaagtcgattttactgtt This studyvppA-His-F2 ccgGAATTCatgggcagcagccatcatcatcatcatcacag This studyvppA-RTq-F cccgctcaggcgcgcagtc This studyvppA-RTq-R tttcccagtctgcatccaa This studyRT-reg1-F aaacaatctcggcgtggtc This studyRT-reg1-R gcgaacgacccaaagcagt This studyRT-reg2-F ttcaacaggcatggatcaagc This studyRT-reg2-R ccaactcttcggcaagcatct This studyRT-reg3-F gctggcactatcatcaatc This studyRT-reg3-R gccgctttgtttgtcattt This study
a Underlined, uppercase nucleotides indicate restriction sites.
Yu et al.
8332 aem.asm.org December 2015 Volume 81 Number 24Applied and Environmental Microbiology
on August 18, 2018 by guest
http://aem.asm
.org/D
ownloaded from

further determined by HPLC, and the reaction samples were terminated byadding three volumes of methanol and filtered with a 0.22-�m filter. All theenzyme activities were measured at 25°C in 50 mM Tris-HCl buffer (pH 8.0).VppB was purified according to the method described previously (7).
To identify the form of cofactor in VppA, the molybdenum concen-tration in partially purified VppA was determined after HNO3 digestionfor 1 h at 160°C, using an inductively coupled plasma-mass spectropho-tometer (ICP-MS). P. putida KTVppA was cultivated in the MSM andMoDM with 1 mg/ml (NH4)2SO4, 5 mg/ml sodium citrate, 1 mM IPTG,and 25 �g/ml Tet, respectively. The nicotine hydroxylation activities ofcells from the two growth media were detected by the resting-cell reac-tions. For further analysis, the recombinant pME6032-vppA was trans-ferred into Escherichia coli DH5� to generate E. coli DHVppA. Nicotinehydroxylation activity of E. coli DHVppA was induced by adding 1 mMIPTG, and the activity was also detected by the resting-cell reactions.
Analytical methods. Biotransformation of nicotine by resting cellswas analyzed by spectrum scanning using a UV-2550 spectrophotometer.The hydroxylation activities of the resting cells and cell extract were fol-lowed by measuring the concentration of nicotine and 6HN by HPLC.HPLC analysis was performed at 30°C with an Agilent 1200 system (Agi-lent, Santa Clara, CA, USA) equipped with an Eclipse XDB-C18 column (5�m, 4.6 by 250 mm; Keystone Scientific, Bellefonte, PA, USA). For anal-ysis of the product from nicotine biotransformation, the mobile phaseconsisted of 15% (vol/vol) methanol and 85% (vol/vol) 1 mM H2SO4 at aflow rate of 0.5 ml/min. The substrate and product were quantitativelymonitored at 259 nm and 300 nm, respectively. Liquid chromatography-mass spectrometry (LC-MS) analysis was performed as previously de-scribed on an Agilent 6230 time-of-flight (TOF)-MS equipped with elec-trospray ionization (ESI) sources (7). The mobile phase of LC-MSconsisted of 60% (vol/vol) ddH2O (0.05% formic acid, vol/vol) and 40%(vol/vol) methanol (0.1% formic acid, vol/vol) at a flow rate of 0.2 ml/min. After the addition of methanol, the samples were treated at 4°C for 10min, centrifuged at 12,000 � g for 2 min, and then filtered using a 0.22-�m-pore-size filter prior to HPLC or LC-MS analysis.
RESULTSThe vppA genes are responsible for biotransformation of nico-tine to 6HN. Our previous study indicated that the vpp cluster isresponsible for nicotine degradation in the VPP pathway (Fig. 1)in Ochrobactrum sp. SJY1. Two open reading frames (ORF2 andORF1) (7) are located in the vpp cluster, downstream of vppB, andshow 13.7% and 36.4% amino acid sequence identity with thelarge and the small subunits, respectively, of Ndh from Arthrobac-ter (Fig. 2A) (18, 19). To determine if these two ORFs are respon-sible for nicotine transformation, subcloning reactions were con-ducted. Based on the predicted ORFs, subclones containingORF1 and ORF2 or ORF1 to ORF3 were ligated into the vectorpME6032 and heterologously expressed in P. putida KT2440 (Fig.2A). Biotransformation was carried out with 0.5 mg/ml nicotineas the substrate using resting cells of P. putida KT6032, P. putidaKTVppA, and P. putida KTVppAplus (Table 1), respectively. Aftera 10-h reaction, the cells were removed by centrifugation and thesupernatant was analyzed by UV-visible scanning and HPLC.In the absence of subclones, the spectrum of the reaction sys-tem with resting cells of P. putida KT6032 did not change, and nonew peak was detected in the HPLC signal. During the reac-tions with resting cells of both P. putida KTVppA and P. putidaKTVppAplus (Table 1), the substrate peak, with a maximal absor-bance at 259 nm, gradually disappeared, giving rise to a new peakwith a maximal absorbance at 295 nm (Fig. 2B). A new peak, witha retention time of 5.05 min, was also detected in the HPLC signal,and the peak that was representative of nicotine disappeared. Theresults indicated that nicotine was transformed to a new product
in the resting-cell reactions. The new product was analyzed byHPLC and LC-MS and identified as 6HN, which has the samespectrum and retention time as the 6HN standard and a molecularweight of 179.1179 (Fig. 2C). There was no significant differencebetween the degradation rates of resting cells of P. putida KTVppAand P. putida KTVppAplus (data not shown). ORF3 has no homol-ogous sequence in the genome of P. putida KT2440 and is pre-dicted to be a pseudoazurin, which is an electron donor in thedenitrification pathway. Thirty-six coding DNA sequences fordenitrification were annotated in the genome of the strain SJY1(6), suggesting that ORF3 may be involved in denitrification reac-tions in this strain. These data indicate that ORF1 and ORF2,which were designated, respectively, vppAS and vppAL (S and Lindicate subunits S and L, respectively), are responsible for theconversion of nicotine to 6HN in the VPP pathway. ORF3 is notnecessary for the nicotine hydroxylation.
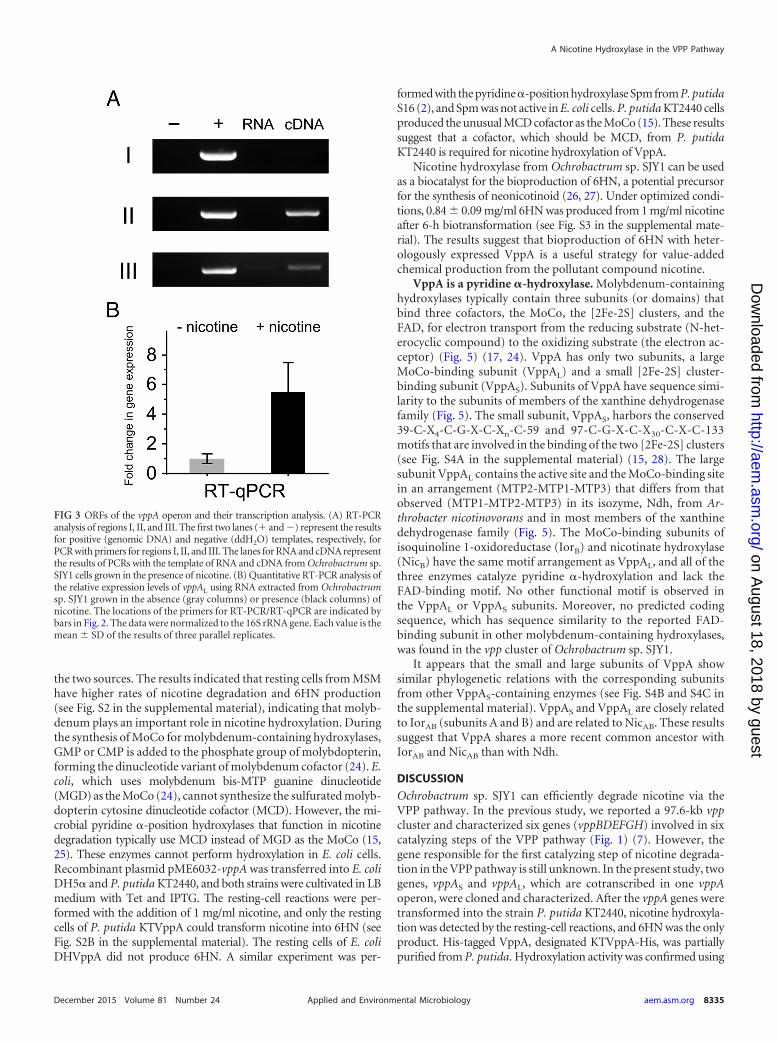
The vppA genes are upregulated in the presence of nicotine.Despite its unknown function, ORF3 is located immediately down-stream of ORF2 in the same orientation. ORFr, a predicted regulator,is located adjacent to, and is transcribed divergently to, ORF1 (Fig.2A). All four ORFs may be translationally coupled. In order to iden-tify the complete operon of vppA sequences, three primer pairs weredesigned to target the flanking regions between ORFr and ORF3. Weobtained amplicons for the intergenic regions between ORF1 andORF2 and between ORF2 and ORF3 but not for the intergenic re-gions between ORFr and ORF1 (Fig. 3A), indicating that, in Ochro-bactrum sp. SJY1, the vppA operon consists of three ORFs (vppAS,vppAL, and ORF3). Genes involved in nicotine catabolism are usu-ally upregulated when nicotine is present in the growth medium(2, 7). To prove the connection between nicotine degradation andthe vppA genes, the expression of vppAL in the presence and absenceof nicotine was studied using RT-qPCR. The results indicated that theexpression level of vppAL transcription significantly increased inthe presence of nicotine compared to the vppAL transcription level inthe absence of nicotine (fold change 5.49) (Fig. 3B), suggesting thatvppAL expression was induced by nicotine or other intermediates ofnicotine degradation. Increased expression correlates with previousdata, showing that nicotine-induced resting cells of strain SJY1 havehigher nicotine transformation rates (7).
VppA is a two-component molybdenum-containing hydrox-ylase. A His tag was added to the N terminus of vppAS, and therecombinant plasmid pME6032-vppA-His was transformed intoP. putida KT2440 to generate P. putida KTVppA-His. His-taggedVppA expression was induced in P. putida KTVppA-His (Table 1)and purified by Ni-Sepharose affinity chromatography under nonde-naturing conditions. The partially purified VppA was confirmed bySDS-PAGE, and two extra bands of approximately 75 kDa and 17kDa, which were absent for the control-purified protein from P.putida KT6032, were identified (Fig. 4A). The 17-kDa protein was theHis-tagged, small subunit of VppA, which was further confirmed bystaining using the InVision His tag in-gel stain (Fig. 4C). A nicotinehydroxylase assay using partially purified VppA indicated that hy-droxylation could be detected when external electron acceptors, suchas DCIP and PES, were added. A decrease in absorption was observedat 600 nm when using DCIP as the electron acceptor (see Fig. S1A inthe supplemental material). HPLC analysis confirmed that nicotinewas transformed to 6HN by partially purified VppA in the catalyticreaction (see Fig. S1B in the supplemental material).
Sequence analysis indicated that the small subunit of VppA hastwo [2Fe-2S] cluster-binding domains. The partially purified
A Nicotine Hydroxylase in the VPP Pathway
December 2015 Volume 81 Number 24 aem.asm.org 8333Applied and Environmental Microbiology
on August 18, 2018 by guest
http://aem.asm
.org/D
ownloaded from

VppA turned tinged brown and exhibited maximum absorptionat 417 nm, which confirmed that VppA is an iron-sulfur protein(Fig. 4B). The previously reported enzymes for pyridine ring�-position hydroxylation are all molybdenum-containing hy-droxylases that require a petrin cofactor. The large subunit ofVppA, encoded by vppAL, is predicted to be a MoCo-binding sub-unit, which suggests that VppA is also a molybdenum-containinghydroxylase. In order to identify the role of molybdenum inVppA, the concentration of molybdenum in 80 �g/ml partially
purified VppA (8.51 0.11 ng/ml molybdenum) and 80 �g/mlcell extract of P. putida KTVppA (3.01 0.09 ng/ml molybde-num) were determined by ICP-MS. The excess molybdenum sug-gested that molybdenum constitutes the cofactor for VppA. Also,resting cells of P. putida KTVppA were prepared from the onescultivated in MSM and MoDM, with (NH4)2SO4 and sodium ci-trate as the nitrogen and carbon sources, respectively, and withIPTG and Tet in the media. Reactions with an initial concentra-tion of 0.5 mg/ml nicotine were carried out with resting cells from
FIG 2 Biotransformation of nicotine to 6-hydroxynicotine (6HN) by heterologously expressed vppA genes. (A) Transcription orientation of 5 ORFs in the vppcluster (GenBank accession numbers AIH15808 to AIH15804 from left to right, respectively). (B) Spectrophotometric changes during the transformation ofnicotine by resting cells of P. putida KTVppA. The reaction was initiated by the addition of 1 mg/ml nicotine to resting cells of strain KTVppA with an OD600 of6.0, and the spectra were recorded every 20 min. Arrows indicate the direction of spectral changes. (C) Identification of 6HN by LC-MS analysis.
Yu et al.
8334 aem.asm.org December 2015 Volume 81 Number 24Applied and Environmental Microbiology
on August 18, 2018 by guest
http://aem.asm
.org/D
ownloaded from
the two sources. The results indicated that resting cells from MSMhave higher rates of nicotine degradation and 6HN production(see Fig. S2 in the supplemental material), indicating that molyb-denum plays an important role in nicotine hydroxylation. Duringthe synthesis of MoCo for molybdenum-containing hydroxylases,GMP or CMP is added to the phosphate group of molybdopterin,forming the dinucleotide variant of molybdenum cofactor (24). E.coli, which uses molybdenum bis-MTP guanine dinucleotide(MGD) as the MoCo (24), cannot synthesize the sulfurated molyb-dopterin cytosine dinucleotide cofactor (MCD). However, the mi-crobial pyridine �-position hydroxylases that function in nicotinedegradation typically use MCD instead of MGD as the MoCo (15,25). These enzymes cannot perform hydroxylation in E. coli cells.Recombinant plasmid pME6032-vppA was transferred into E. coliDH5� and P. putida KT2440, and both strains were cultivated in LBmedium with Tet and IPTG. The resting-cell reactions were per-formed with the addition of 1 mg/ml nicotine, and only the restingcells of P. putida KTVppA could transform nicotine into 6HN (seeFig. S2B in the supplemental material). The resting cells of E. coliDHVppA did not produce 6HN. A similar experiment was per-
formed with the pyridine�-position hydroxylase Spm from P. putidaS16 (2), and Spm was not active in E. coli cells. P. putida KT2440 cellsproduced the unusual MCD cofactor as the MoCo (15). These resultssuggest that a cofactor, which should be MCD, from P. putidaKT2440 is required for nicotine hydroxylation of VppA.
Nicotine hydroxylase from Ochrobactrum sp. SJY1 can be usedas a biocatalyst for the bioproduction of 6HN, a potential precursorfor the synthesis of neonicotinoid (26, 27). Under optimized condi-tions, 0.84 0.09 mg/ml 6HN was produced from 1 mg/ml nicotineafter 6-h biotransformation (see Fig. S3 in the supplemental mate-rial). The results suggest that bioproduction of 6HN with heter-ologously expressed VppA is a useful strategy for value-addedchemical production from the pollutant compound nicotine.
VppA is a pyridine �-hydroxylase. Molybdenum-containinghydroxylases typically contain three subunits (or domains) thatbind three cofactors, the MoCo, the [2Fe-2S] clusters, and theFAD, for electron transport from the reducing substrate (N-het-erocyclic compound) to the oxidizing substrate (the electron ac-ceptor) (Fig. 5) (17, 24). VppA has only two subunits, a largeMoCo-binding subunit (VppAL) and a small [2Fe-2S] cluster-binding subunit (VppAS). Subunits of VppA have sequence simi-larity to the subunits of members of the xanthine dehydrogenasefamily (Fig. 5). The small subunit, VppAS, harbors the conserved39-C-X4-C-G-X-C-Xn-C-59 and 97-C-G-X-C-X30-C-X-C-133motifs that are involved in the binding of the two [2Fe-2S] clusters(see Fig. S4A in the supplemental material) (15, 28). The largesubunit VppAL contains the active site and the MoCo-binding sitein an arrangement (MTP2-MTP1-MTP3) that differs from thatobserved (MTP1-MTP2-MTP3) in its isozyme, Ndh, from Ar-throbacter nicotinovorans and in most members of the xanthinedehydrogenase family (Fig. 5). The MoCo-binding subunits ofisoquinoline 1-oxidoreductase (IorB) and nicotinate hydroxylase(NicB) have the same motif arrangement as VppAL, and all of thethree enzymes catalyze pyridine �-hydroxylation and lack theFAD-binding motif. No other functional motif is observed inthe VppAL or VppAS subunits. Moreover, no predicted codingsequence, which has sequence similarity to the reported FAD-binding subunit in other molybdenum-containing hydroxylases,was found in the vpp cluster of Ochrobactrum sp. SJY1.
It appears that the small and large subunits of VppA showsimilar phylogenetic relations with the corresponding subunitsfrom other VppAS-containing enzymes (see Fig. S4B and S4C inthe supplemental material). VppAS and VppAL are closely relatedto IorAB (subunits A and B) and are related to NicAB. These resultssuggest that VppA shares a more recent common ancestor withIorAB and NicAB than with Ndh.
DISCUSSION
Ochrobactrum sp. SJY1 can efficiently degrade nicotine via theVPP pathway. In the previous study, we reported a 97.6-kb vppcluster and characterized six genes (vppBDEFGH) involved in sixcatalyzing steps of the VPP pathway (Fig. 1) (7). However, thegene responsible for the first catalyzing step of nicotine degrada-tion in the VPP pathway is still unknown. In the present study, twogenes, vppAS and vppAL, which are cotranscribed in one vppAoperon, were cloned and characterized. After the vppA genes weretransformed into the strain P. putida KT2440, nicotine hydroxyla-tion was detected by the resting-cell reactions, and 6HN was the onlyproduct. His-tagged VppA, designated KTVppA-His, was partiallypurified from P. putida. Hydroxylation activity was confirmed using
FIG 3 ORFs of the vppA operon and their transcription analysis. (A) RT-PCRanalysis of regions I, II, and III. The first two lanes (� and �) represent the resultsfor positive (genomic DNA) and negative (ddH2O) templates, respectively, forPCR with primers for regions I, II, and III. The lanes for RNA and cDNA representthe results of PCRs with the template of RNA and cDNA from Ochrobactrum sp.SJY1 cells grown in the presence of nicotine. (B) Quantitative RT-PCR analysis ofthe relative expression levels of vppAL using RNA extracted from Ochrobactrumsp. SJY1 grown in the absence (gray columns) or presence (black columns) ofnicotine. The locations of the primers for RT-PCR/RT-qPCR are indicated bybars in Fig. 2. The data were normalized to the 16S rRNA gene. Each value is themean SD of the results of three parallel replicates.
A Nicotine Hydroxylase in the VPP Pathway
December 2015 Volume 81 Number 24 aem.asm.org 8335Applied and Environmental Microbiology
on August 18, 2018 by guest
http://aem.asm
.org/D
ownloaded from

a UV-visible spectrometer and HPLC with nicotine as the substrateand DCIP as the electric acceptor. In addition, the vppA operon, lo-cated in the vpp cluster, is approximately 100 bp downstream of thevppB gene. Based on these results, we concluded that the vppA genes,which encode the nicotine hydroxylase, are responsible for the hy-droxylation of nicotine to 6HN in the VPP pathway.
The pyridine hydroxylases can be subclassified as �-, �-, or�-position hydroxylases according to the hydroxylation positionon the pyridine ring (10). VppA is classified as a pyridine ring�-position hydroxylase based on its catalytic reaction. This kind ofenzyme typically belongs to the family of molybdenum cofactor,Fe-S cluster, and FAD-dependent hydroxylases, such as Ndh (theisozyme of VppA), Spm, and NicAB, which are involved in pyri-dine degradation (18–20). However, VppA is a two-componentenzyme, which is different from its isozyme Ndh (18). A MoCo-binding subunit and Fe-S clusters were identified in VppA, but theFAD-binding subunit was not. Subcloning experiments showedthat two coding sequences are sufficient for hydroxylation, and
only two extra bands were recognized in an SDS-PAGE gel ofpartially purified VppA compared with that of the control. Theexistence of molybdenum was confirmed by ICP-MS and Mo de-ficiency cultivation experiments. Fe-S clusters can be identifiedfrom the characteristic absorption peak of purified VppA, andHis-tag-specific staining indicated the existence of a [2Fe-2S]-binding subunit. However, no other extra bands were observed bySDS-PAGE, and the UV-visible spectrum did not show the char-acteristic peak of a flavin cofactor (10). These results suggestedthat VppA does not have an FAD-binding subunit.
Molybdenum-containing enzymes are ubiquitous and foundin nearly all organisms (17, 24). Of interest to drug research are themolybdenum-containing hydroxylases, which not only need theMoCo but also usually require an FAD cofactor for catalytic activ-ity (28, 29). In contrast to flavin-dependent monooxygenases,molybdenum hydroxylases use water instead of oxygen as the ul-timate source of the oxygen atom to be incorporated into thesubstrate, and they generate reducing equivalents in the hydroxy-
FIG 4 Partially purified His-tagged nicotine hydroxylase from P. putida KTVppA-His. (A) SDS-PAGE of protein markers (lane M), cell extract of P. putidaKTVppA-His (lane CE), purified protein from P. putida KT6032 (lane CK), and purified VppA (lane VppA) stained with Coomassie blue. VppA and the controlwere purified by the same procedures. (B) UV-visible spectra of purified VppA. (C) Gel visualized with Coomassie blue (left) and InVision His tag in-gel stain(right) of markers (lane M), VppB (lane CK), and partially purified nicotine hydroxylase (lane VppA).
Yu et al.
8336 aem.asm.org December 2015 Volume 81 Number 24Applied and Environmental Microbiology
on August 18, 2018 by guest
http://aem.asm
.org/D
ownloaded from

lation reaction (30). FAD is an intermediate electron transfercomponent. In xanthine oxidase, electrons transfer from the Fe-Scenter to the flavin center to reduce the flavin center (31). VppAdoes not have an FAD-binding subunit, and neither nicotinatedehydrogenase (NicAB) nor isoquinoline 1-oxidoreductase (IorAB)has this kind of subunit (15, 32). Also, VppAL has a different motifarrangement (MTP2-MTP1-MTP3) than the MoCo-bindingsubunits of those molybdenum-containing hydroxylases thathave the FAD-binding subunit. It is interesting that the MoCo-binding subunit of NicAB and IorAB has the same domain arrange-ment as VppA. Therefore, rearrangement of the motifs in theMoCo-binding subunit may reconstruct the electron transfer chain,which does not require participation of the FAD cofactor. VppA,NicAB, and IorAB, therefore, can be subclassified as a group within themolybdenum-containing hydroxylase family that only has theMoCo-binding and the Fe-S cluster-binding components and thathas a specific arrangement of motifs in the MoCo-binding subunit.
Three nicotine degradation pathways have been characterizedin bacteria (2–4, 7). The pyridine pathway was mainly discoveredin Arthrobacter species (3), and the pyrrolidine pathway was typ-ically carried by Pseudomonas strains (2, 33). In contrast, the VPPpathway was reported in at least four strains belonging to fourdifferent genera (7, 8, 12, 13), and genes of the VPP pathway fromdifferent strains have high sequence identity to each other (9, 14).On the other hand, VppA shows low sequence identity to itsisozyme Ndh from Arthrobacter (13.7% and 36.4% sequence iden-tity to the large and small subunits, respectively). However, VppAL
shows 22.1% amino acid sequence identity to the MoCo-binding
motif of NicB (from positions 1 to 743), which is higher than thatto NdhL. This is in accordance with previous results showing thatgenes of the VPP pathway, which may have evolved indepen-dently, are not a simple combination of genes from the pyridinepathway and the pyrrolidine pathway (7). Genes of the VPP path-way appear to be distributed across different genera; this supportsour previous speculation that strain SJY1 may have obtained thevpp cluster via horizontal gene transfer (7).
The pyridine heterocycles continue to play vital roles in thedevelopment of agricultural chemicals and human medicines(27). Microbial degradation of pyridine derivatives, such as nico-tine, may produce numerous useful intermediates for agrochemi-cal and pharmaceutical synthesis (26). Here, we demonstratedthat the recombinant strain P. putida KTVppA is an efficient bio-catalyst for the biotransformation of nicotine to 6HN, which caneasily be modified by replacing the hydroxide radical with a halo-gen element. 6HN can potentially serve as a precursor for thesynthesis of a neonicotinoid. In contrast, the formation of2-substituted pyridine derivatives requires strict conditions forhigh product yields in organic chemistry methods (26). Thisstudy, therefore, not only extends the understanding of molybde-num-containing hydroxylases but also provides a useful potentialbiocatalyst preparation for nicotine biotransformation.
Seven genes in the VPP pathway have been identified; neverthe-less, identification of the enzyme for conversion of 6HPON to HSP isstill needed for complete understanding of this pathway. The genesthat are involved in nicotine regulation and degradation are usuallyclustered on the megaplasmid or chromosome (2, 7, 19). Analysis of
FIG 5 Molecular architecture of different molybdenum-containing hydroxylases. NdhLMS (GenBank accession numbers CAA53088, CAA53087, andCAA53086), nicotine dehydrogenase from A. nicotinovorans; KdhLMS (WP_016359451, WP_016359456, and WP_016359457), ketone dehydrogenase from A.nicotinovorans; SpmABC (AEJ14617 and AEJ14616), 3-succinoylpyridine dehydrogenase from P. putida; QoxLMS (GenBank accession numbers CAD61045,CAD61046, and CAD61047), quinaldine 4-oxidase from Arthrobacter ilicis; Hxa (GenBank accession number Q12553), xanthine dehydrogenase from Aspergillusnidulans; NicAB (AAN69541 and AAN69542), nicotinate dehydrogenase from P. putida; and IorAB (GenBank accession numbers CAA88753 and CAA88754),isoquinoline 1-oxidoreductase from Brevundimonas diminuta. The letters depicted below the proteins indicate the subunit names of the corresponding proteins.The conserved domains are as follows: [FeS], ferredoxin-like [2Fe-2S]-binding domain; FAD, FAD-binding domain; SRPBCC, SRPBCC ligand-binding domain;MPT, domains for binding to the MoCo; CytC, cytochrome c binding domain.
A Nicotine Hydroxylase in the VPP Pathway
December 2015 Volume 81 Number 24 aem.asm.org 8337Applied and Environmental Microbiology
on August 18, 2018 by guest
http://aem.asm
.org/D
ownloaded from

the vpp cluster may provide novel information, enabling us to furtherunderstand the molecular mechanism of the VPP pathway.
ACKNOWLEDGMENTS
This work was supported in part by grants from the Chinese NationalNatural Science Foundation (31270154 and 31230002), the Chinese Na-tional Science Foundation for Excellent Young Scholars (31422004), andthe National Basic Research Program of China (2013CB733901).
We thank the Chen Xing project from Shanghai Jiaotong University. We alsothankRoderichBrandsch(InstituteofBiochemistryandMolecularBiology,Uni-versity of Freiburg) for providing the compound used in this study.
REFERENCES1. Benowitz NL, Hukkanen J, Jacob P, III. 2009. Nicotine chemistry, metabolism,
kinetics and biomarkers, p 29–60. In Henningfield JE, Calvento E, Pogun S (ed),Nicotine psychopharmacology. Springer, San Francisco, CA.
2. Tang HZ, Wang LJ, Wang WW, Yu H, Zhang KZ, Yao YX, Xu P. 2013.Systematic unraveling of the unsolved pathway of nicotine degradation inPseudomonas. PLoS Genet 9:e1003923. http://dx.doi.org/10.1371/journal.pgen.1003923.
3. Brandsch R. 2006. Microbiology and biochemistry of nicotine degrada-tion. Appl Microbiol Biotechnol 69:493– 498. http://dx.doi.org/10.1007/s00253-005-0226-0.
4. Gurusamy R, Natarajan S. 2013. Current status on biochemistry andmolecular biology of microbial degradation of nicotine. Sci World J 2013:125385. http://dx.doi.org/10.1155/2013/125385.
5. Yu H, Tang HZ, Wang LJ, Yao YX, Wu G, Xu P. 2011. Completegenome sequence of the nicotine-degrading Pseudomonas putida strainS16. J Bacteriol 193:5541–5542. http://dx.doi.org/10.1128/JB.05663-11.
6. Yu H, Li YY, Tang HZ, Xu P. 2014. Genome sequence of a newly isolatednicotine-degrading bacterium Ochrobactrum sp. SJY1. Genome Announc2:e00720-14. http://dx.doi.org/10.1128/genomeA.00720-14.
7. Yu H, Tang HZ, Zhu XY, Li YY, Xu P. 2015. Molecular mechanism ofnicotine degradation by a newly isolated strain, Ochrobactrum sp. strainSJY1. Appl Environ Microbiol 81:272–281. http://dx.doi.org/10.1128/AEM.02265-14.
8. Ma Y, Wei Y, Qiu JG, Wen RT, Hong J, Liu WP. 2014. Isolation,transposon mutagenesis, and characterization of the novel nicotine-degrading strain Shinella sp. HZN7. Appl Microbiol Biotechnol 98:2625–2636. http://dx.doi.org/10.1007/s00253-013-5207-0.
9. Wang SN, Huang HY, Xie KB, Xu P. 2012. Identification of nicotinebiotransformation intermediates by Agrobacterium tumefaciens strain S33suggests a novel nicotine degradation pathway. Appl Microbiol Biotech-nol 95:1567–1578. http://dx.doi.org/10.1007/s00253-012-4007-2.
10. Yu H, Hausinger RP, Tang HZ, Xu P. 2014. Mechanism of the 6-hy-droxy-3-succinoyl-pyridine 3-monooxygenase flavoprotein from Pseu-domonas putida S16. J Biol Chem 289:29158 –29170. http://dx.doi.org/10.1074/jbc.M114.558049.
11. Tang HZ, Yao YX, Wang LJ, Yu H, Ren YL, Wu G, Xu P. 2012. Genomicanalysis of Pseudomonas putida: genes in a genome island are crucial fornicotine degradation. Sci Rep 2:377. http://dx.doi.org/10.1038/srep00377.
12. Wang SN, Liu Z, Xu P. 2009. Biodegradation of nicotine by a newlyisolated Agrobacterium sp. strain S33. J Appl Microbiol 107:838 – 847.http://dx.doi.org/10.1111/j.1365-2672.2009.04259.x.
13. Ma Y, Wen R, Qiu J, Hong J, Liu M, Zhang D. 2014. Biodegradation ofnicotine by a novel strain Pusillimonas. Res Microbiol 166:67–71.
14. Qiu JG, Wei Y, Ma Y, Wen RT, Wen YZ, Liu WP. 2014. A novel (S)-6-hydroxynicotine oxidase gene from Shinella sp. strain HZN7. Appl EnvironMicrobiol 80:5552–5560. http://dx.doi.org/10.1128/AEM.01312-14.
15. Jiménez JI, Canales A, Jimenez-Barbero J, Ginalski K, Rychlewski L,Garcia JL, Diaz E. 2008. Deciphering the genetic determinants for aerobicnicotinic acid degradation: the nic cluster from Pseudomonas putidaKT2440. Proc Natl Acad Sci U S A 105:11329 –11334. http://dx.doi.org/10.1073/pnas.0802273105.
16. Zientek M, Kang P, Hutzler MJ, Obach SR. 2012. Molybdenum-containing hydroxylases, p 1–59. In Lyubimov AV (ed), Encyclopedia ofdrug metabolism and interactions, 1st ed. John Wiley & Sons, Inc, Hobo-ken, NJ, USA.
17. Schwarz G, Mendel RR, Ribbe MW. 2009. Molybdenum cofactors,
enzymes, and pathways. Nature 460:839 – 847. http://dx.doi.org/10.1038/nature08302.
18. Grether-Beck S, Igloi GL, Pust S, Schilz E, Decker K, Brandsch R. 1994.Structural analysis and molybdenum-dependent expression of the pAO1-encoded nicotine dehydrogenase genes of Arthrobacter nicotinovorans.Mol Microbiol 13:929 –936. http://dx.doi.org/10.1111/j.1365-2958.1994.tb00484.x.
19. Igloi GL, Brandsch R. 2003. Sequence of the 165-kilobase catabolic plas-mid pAO1 from Arthrobacter nicotinovorans and identification of a pAO1-dependent nicotine uptake system. J Bacteriol 185:1976 –1986. http://dx.doi.org/10.1128/JB.185.6.1976-1986.2003.
20. Schenk S, Hoelz A, Krauss B, Decker K. 1998. Gene structures andproperties of enzymes of the plasmid-encoded nicotine catabolism of Ar-throbacter nicotinovorans. J Mol Biol 284:1323–1339. http://dx.doi.org/10.1006/jmbi.1998.2227.
21. Wang SN, Xu P, Tang HZ, Meng J, Liu XL, Huang J, Chen H, Du Y,Blankespoor HD. 2004. Biodegradation and detoxification of nicotine intobacco solid waste by a Pseudomonas sp. Biotechnol Lett 26:1493–1496.http://dx.doi.org/10.1023/B:BILE.0000044450.16235.65.
22. Heeb S, Itoh Y, Nishijyo T, Schnider U, Keel C, Wade J, Walsh U,O’Gara F, Haas D. 2000. Small, stable shuttle vectors based on the min-imal pVS1 replicon for use in gram-negative, plant-associated bacteria.Mol Plant Microbe Interact 13:232–237. http://dx.doi.org/10.1094/MPMI.2000.13.2.232.
23. Nelson KE, Weinel C, Paulsen IT, Dodson RJ, Hilbert H, Martins dosSantos VA, Fouts DE, Gill SR, Pop M, Holmes M, Brinkac L, Beanan M,DeBoy RT, Daugherty S, Kolonay J, Madupu R, Nelson W, White O,Peterson J, Khouri H, Hance I, Chris Lee P, Holtzapple E, Scanlan D,Tran K, Moazzez A, Utterback T, Rizzo M, Lee K, Kosack D, Moestl D,Wedler H, Lauber J, Stjepandic D, Hoheisel J, Straetz M, Heim S,Kiewitz C, Eisen JA, Timmis KN, Dusterhoft A, Tummler B, Fraser CM.2002. Complete genome sequence and comparative analysis of the meta-bolically versatile Pseudomonas putida KT2440. Environ Microbiol 4:799 –808. http://dx.doi.org/10.1046/j.1462-2920.2002.00366.x.
24. Iobbi-Nivol C, Leimkuhler S. 2013. Molybdenum enzymes, their matu-ration, and molybdenum cofactor biosynthesis in Escherichia coli.Biochim Biophys Acta 1827:1086–1101. http://dx.doi.org/10.1016/j.bbabio.2012.11.007.
25. Sachelaru P, Schiltz E, Brandsch R. 2006. A functional mobA gene formolybdopterin cytosine dinucleotide cofactor biosynthesis is required foractivity and holoenzyme assembly of the heterotrimeric nicotine dehydro-genases of Arthrobacter nicotinovorans. Appl Environ Microbiol 72:5126 –5131. http://dx.doi.org/10.1128/AEM.00437-06.
26. Yu H, Tang HZ, Xu P. 2014. Green strategy from waste to value-added-chemical production: efficient biosynthesis of 6-hydroxy-3-succinoyl-pyridine by an engineered biocatalyst. Sci Rep 4:5397. http://dx.doi.org/10.1038/srep05397.
27. Schmid A, Dordick JS, Hauer B, Kiener A, Wubbolts M, Witholt B.2001. Industrial biocatalysis today and tomorrow. Nature 409:258 –268.http://dx.doi.org/10.1038/35051736.
28. Hille R. 2005. Molybdenum-containing hydroxylases. Arch Biochem Bio-phys 433:107–116. http://dx.doi.org/10.1016/j.abb.2004.08.012.
29. Kitamura S, Sugihara K, Ohta S. 2006. Drug-metabolizing ability ofmolybdenum hydroxylases. Drug Metab Pharmacokinet 21:83–98. http://dx.doi.org/10.2133/dmpk.21.83.
30. Huijbers MM, Montersino S, Westphal AH, Tischler D, van Berkel WJ.2014. Flavin-dependent monooxygenases. Arch Biochem Biophys 544:2–17. http://dx.doi.org/10.1016/j.abb.2013.12.005.
31. Hille R, Anderson RF. 2001. Coupled electron/proton transfer in com-plex flavoproteins: solvent kinetic isotope effect studies of electron trans-fer in xanthine oxidase and trimethylamine dehydrogenase. J Biol Chem276:31193–31201. http://dx.doi.org/10.1074/jbc.M100673200.
32. Lehmann M, Tshisuaka B, Fetzner S, Lingens F. 1995. Molecular cloningof the isoquinoline 1-oxidoreductase genes from Pseudomonas diminuta 7,structural analysis of iorA and iorB, and sequence comparisons with othermolybdenum-containing hydroxylases. J Biol Chem 270:14420 –14429.http://dx.doi.org/10.1074/jbc.270.24.14420.
33. Qiu JG, Ma Y, Wen YZ, Chen LS, Wu LF, Liu WP. 2012. Functionalidentification of two novel genes from Pseudomonas sp. strain HZN6 in-volved in the catabolism of nicotine. Appl Environ Microbiol 78:2154 –2160. http://dx.doi.org/10.1128/AEM.07025-11.
Yu et al.
8338 aem.asm.org December 2015 Volume 81 Number 24Applied and Environmental Microbiology
on August 18, 2018 by guest
http://aem.asm
.org/D
ownloaded from