morphological analysis of the primary center receiving spatial information transferred by the waggle...
TRANSCRIPT

Morphological Analysis of the Primary CenterReceiving Spatial Information Transferred by theWaggle Dance of Honeybees
Hiroyuki Ai* and Hiromi Hagio
Division of Biology, Department of Earth System Science, Fukuoka University, Fukuoka 814-0180, Japan
ABSTRACTThe waggle dancers of honeybees encodes roughly the
distance and direction to the food source as the dura-
tion of the waggle phase and the body angle during the
waggle phase. It is believed that hive-mates detect air-
borne vibrations produced during the waggle phase to
acquire distance information and simultaneously detect
the body axis during the waggle phase to acquire direc-
tion information. It has been further proposed that the
orientation of the body axis on the vertical comb is
detected by neck hairs (NHs) on the prosternal organ.
The afferents of the NHs project into the prothoracic
and mesothoracic ganglia and the dorsal subesophageal
ganglion (dSEG). This study demonstrates somatotopic
organization within the dSEG of the central projections
of the mechanosensory neurons of the NHs. The termi-
nals of the NH afferents in dSEG are in close apposition
to those of Johnston’s organ (JO) afferents. The sensory
axons of both terminate in a region posterior to the
crossing of the ventral intermediate tract (VIT) and the
maxillary dorsal commissures I and III (MxDCI, III) in
the subesophageal ganglion. These features of the ter-
minal areas of the NH and JO afferents are common to
the worker, drone, and queen castes of honeybees.
Analysis of the spatial relationship between the NH
neurons and the morphologically and physiologically
characterized vibration-sensitive interneurons DL-Int-1
and DL-Int-2 demonstrated that several branches of DL-
Int-1 are in close proximity to the central projection of
the mechanosensory neurons of the NHs in the dSEG.
J. Comp. Neurol. 521:2570–2584, 2013.
VC 2013 Wiley Periodicals, Inc.
INDEXING TERMS: vibration; body angle; Johnston’s organ; neck hairs; subesophageal ganglion honeybee standard brain
Honeybees share information concerning profitable
flowers by a species-specific behavior, the waggle dance
(von Frisch, 1967). During waggle-dance communication,
hive-mates of the dancer are able to decipher the vector
information contained within the dance, and subse-
quently, they arrive at the indicated flower patch. Riley
et al. (2005) confirmed this phenomenon by recording
the actual flight paths of recruited bees.
It was suggested that Johnston’s organ (JO) on the ped-
icel of each antenna detects the vibration for decoding
the distance information (von Frisch, 1976) and that the
neck hairs (NHs) detect the body angle for decoding the
direction information (Fig. 1; Lindauer and Nedel, 1959).
More recently, the central projections of these two sen-
sory organs that receive this vector information have
been clarified. The afferents originating from the JO pro-
ject into the dorsal lobe (DL) through antennal nerve tract
6 (T6; Suzuki, 1975) and then bifurcate within the DL into
T6I and T6II. The JO afferents in T6I terminate in the
medial posterior protocerebral lobe (PPL) close to the
esophagus, whereas those in T6II terminate in the dorsal
subesophageal ganglion (dSEG; Ai et al., 2007). In con-
trast, the afferents of NH neurons project into the pro-
thoracic ganglion (pro-TG) through lateral nerve tract 1
and terminate ipsilaterally in a region close to the midline
of the pro-TG. From there, the NH afferents extend axons
into the posterior and anterior regions of the pro-TG.
These afferents extend axons through the ventral nerve
cord (VNC) posteriorly, terminating close to the midline of
the mesothoracic ganglion (meso-TG; Sandeman et al,
1997). Anteriorly, the axons extend through the VNC to
terminate at the dSEG (Brockmann and Robinson, 2007).
Thus, the JO and NH sensory afferents terminate in
Grant sponsor: Ministry of Education, Culture, Sports, Science andTechnology of Japan; Grant number: 22570079.
*CORRESPONDENCE TO: Hiroyuki Ai, Division of Biology, Department ofEarth System Science, Fukuoka University, 8-19-1 Nanakuma, Jonan-ku,Fukuoka 814-0180, Japan. E-mail: [email protected]
VC 2013 Wiley Periodicals, Inc.
Received November 15, 2012; Revised December 8, 2012; AcceptedDecember 27, 2012
DOI 10.1002/cne.23299
Published online January 8, 2013 in Wiley Online Library(wileyonlinelibrary.com)
2570 The Journal of Comparative Neurology | Research in Systems Neuroscience 521:2570–2584 (2013)
RESEARCH ARTICLE

diverse and different regions of the central nervous sys-
tem. However, it is noteworthy that terminations within
the dSEG are common to both JO and NH afferents, sug-
gesting a putative role for the dSEG as an integration cen-
ter for direction and distance information (Ai et al., 2007).
Coding of the topology of the receptors of the insect
auditory system is commonly achieved by somatotopic
organization of the projections of their sensory neurons in
the central nervous system (Ignell et al., 2005; Kamikou-
chi et al., 2006). In honeybees, somatotopic organization
of the JO is present in the PPL (Ai et al., 2007). Likewise,
to decode the orientation of the food source, honeybees
must recognize which of the NHs are compressed by the
posterior capsule of the head (Fig. 1). At least three sub-
regions of NHs have been described: the dorsal, central,
and ventral NHs (Sandeman et al., 1997). The dorsal and
ventral NHs are longer (>100 lm) than the central NHs
(<100 lm), so they can be conveniently identified from
both their position and their length. In this study, we
investigated whether there are differences among these
three NH groups with respect to the central projection
patterns of their afferents within the pro-TG and meso-TG
and the dSEG, i.e., whether there is a somatotopic organi-
zation of the dSEG indicative of a role in integration of
waggle dance sensory information.
The spatial relationship between the termination
regions of these two sensory organs in honeybees is
unknown. Here we identify a putative integration area of
these sensory organs by two methods: by double staining
of the mechanosensory neurons of NHs and the JO and
by registering the positions of the NH and JO afferents
within an averaged three-dimensional honeybee brain
image (the ‘‘honeybee standard brain’’ [HSB]). For analy-
sis of the spatial relationships among the different neu-
rons using the HSB, laser scanning confocal microscopy
(LSM) images were transformed through two processes:
affine transformation and elastic registration within the
virtual space of the HSB (Rohlfing, et al., 2004). In addi-
tion, we investigated whether workers, drones, and
queens have similar projection areas of the JO and NH
afferents in the dSEG.
Previously, we have morphologically and physiologi-
cally characterized several types of vibration-sensitive
interneurons, which arborize in the dSEG, whose vibration
response patterns exhibit on-off phasic excitation, tonic
excitation, or tonic inhibition, suggesting a possible role
in coding of the duration of airborne vibrations (Ai et al.,
2009). These interneurons have dendritic arborization
close to the JO axon terminals (Ai, 2010). Here, we inves-
tigated the spatial relationship between morphologically
and physiologically characterized vibration-sensitive inter-
neurons and the NH terminals.
MATERIALS AND METHODS
Honeybees (Apis mellifera L.) were captured at the en-
trance of hives kept in Fukuoka University between April
and September, 2010, 2011, and 2012. The experiments
used more than 100 worker honeybees, two queens, and
six drones.
Anterograde staining of subgroups of NHsThere are three subgroups of NHs on each side of the
neck: the central, dorsal, and ventral subgroups (Fig.
1A,C). To stain subgroups of NHs specifically, a small
patch of the NHs was shaved off with a razor blade. Then,
a small incision was made to expose the cell bodies of
the NH neurons, onto which a single crystal of dextran
tetramethylrhodamine (D3308; Invitrogen, Carlsbad, CA)
was placed. The preparations were kept for 48 hours at
4�C. The success rate in obtaining the complete staining
of just a few sensory afferents of interest was about 10%.
Double-fluorescence dye injection using crystals of both
dextran-fluorescein (D3306; Invitrogen, Carlsbad, CA),
and dextran-tetramethylrhodamine was also applied to
different subgroups of NHs. The success rate for these
experiments was quite low, because the subgroups of the
NHs represented a relatively small area of the prosternal
organ.
Anterograde staining of the antennal nerveand the NH sensory nerve
It has been shown that axons within the antennal nerve
project into the antennal lobe, the DL, the subesophageal
ganglion (SEG), and the PPL (Pareto, 1972; Suzuki, 1975;
Mobbs, 1982; Arnold et al., 1985; Flanagan and Mercer,
1989; Maronde, 1991; Kloppenburg, 1995; Abel et al.,
2001; Kelber et al., 2006; Kirschner et al., 2006). The
Abbreviations
AL antennal lobeAN antennal nerveCA calyx of mushroom bodyCB central bodyDL-Int-1 dorsal lobe interneuron 1DL-Int-2 dorsal lobe interneuron 2dSEG dorsal subesophageal ganglionHSB honeybee standard brainEs esophagusJO Johnston’s organLo loburaMe medullameso-TG mesothoracic ganglionMxDCI, III maxillary dorsal commissure I and IIINHs neck hairsPC protocerebrumPPL posterior protocerebral lobepro-TG prothoracic ganglionSEG subesophageal ganglionT5 tract 5T6I, II tracts 6-1 and 2VIT ventral intermediate tract
Primary Center of Vector Information in Bees
The Journal of Comparative Neurology | Research in Systems Neuroscience 2571

central projection in the antennal lobe has been revealed
in more detail (Suzuki, 1975; Mobbs, 1982; Abel et al.,
2001; Kelber et al., 2006; Kirschner et al., 2006). In our
previous study, the central projections of the JO were
investigated by using three-dimensional reconstruction (Ai
et al., 2007). The sensory neurons originating from the
NHs have also been investigated, in this case by dye injec-
tion into the first lateral nerve of the pro-TG, a nerve that
innervates only the NHs (Lindauer and Nedel, 1959; San-
deman et al, 1997; Brockmann and Robinson, 2007). To
investigate the spatial relationship between the central
projections of JO afferents and those of NH afferents, dou-
ble-fluorescence dye injection was applied to the proximal
cut stump of the antennal nerve at the base of the pedicel
and to the first lateral nerve of the pro-TG. JO is located in
the pedicel. If the dye is injected proximal to the pedicel,
the sensory fibers running from both the pedicel and the
flagella will be stained in the brain. However, JO afferents
are easily distinguished from other afferents because of
their characteristic morphologies (Ai et al., 2007).
Figure 1. Neck hairs (NHs) and their correspondence with the body orientation on the vertical comb. A: The NHs on the left side. There
are three subregions, dorsal, central, and ventral NHs, recognized by the position and their length. B: Detection of the body angle on the
vertical comb. The bee detects the inclination of the head capsule against the thorax (gray arrows). C: Drawing of detection of body
angle during the waggle phase. The three subgroups of NHs are differentially subjected to pressure by the posterior head capsule. For
example, when the bees are oriented in the opposite direction from gravity (head upward, 0�), the ventral NHs contact the posterior head
capsule. When the bees are oriented to the right (90�), the central hairs on the right side make contact with the posterior head capsule.
Scale bar ¼ 500 lm.
Ai and Hagio
2572 The Journal of Comparative Neurology |Research in Systems Neuroscience

After anesthetizing the bees for 5–10 minutes, all wings
and legs were removed and the sensory nerves supplying
the NHs were exposed and cut at the base of the NHs.
The antenna was cut in the distal region of the scape. The
brain, pro-TG, and meso-TG complex, with the antennal
and NH nerves, was exposed and excised from the body.
The stumps of the antennal nerve and NH nerve were
carefully placed into the tips (internal diameter �70 lm)
of tapered separate glass electrodes filled, respectively,
with 0.8% dextran-fluorescein and with 0.8% dextran-tetra-
methylrhodamine. Alternatively, crystals of these fluores-
cence dyes were placed directly onto the stump of the
antennal nerve and the NHs. The dye was then distributed
by axonal transport into the brain. After incubation at 4�C
for 24–48 hours, the brain, pro-TG and meso-TG com-
plexes were dissected out, rinsed in PBS (pH 6.7) several
times, fixed in 4% paraformaldehyde in 0.1 M PBS for 12
hours, dehydrated, and cleared in methyl salicylate.
Confocal microscopy and three-dimensionalreconstruction
The posterior aspects of stained and cleared specimens
of the central projections of antennal and NH afferents
were viewed with a confocal laser scanning microscope
(LSM 510; Carl Zeiss, Jena, Germany) using a Zeiss Plan-
Apochromat �10/0.45 dry lens objective for low-magnifi-
cation images and a Zeiss Plan-APO LD �25/0.8 Imm
Korr DIC or a Zeiss Plan-Neofluar �40/1.30 oil lens objec-
tives at high magnifications. Optical sections were made
at 3 lm (low magnification) or 1 lm (high magnification)
throughout the entire depth of each specimen. Dextran-
tetramethylrhodamine was excited by the 543-nm line of a
HeNe laser, whereas dextran-fluorescein was excited by
488-nm line of an argon laser. The stained sensory fibers
were reconstructed three-dimensionally from optical sec-
tions by using the label field editor in Amira 4.1 software
(Mercury Computer Systems Inc., San Diego, CA). The out-
lines of the neuropil and the brain/SEG complex were also
traced with the label field editor to identify the location of
the stained sensory fibers in the brain/SEG complex.
Registration of the morphologies of sensoryafferents and interneurons into thehoneybee standard brain
In addition to double-staining analyses, we used the
honeybee standard brain (HSB) to analyze the spatial rela-
tionships among JO afferents, NH afferents, and
morphologically and physiologically characterized vibra-
tion-sensitive interneurons. First, the neuronal profiles of
stained JO afferents, NH afferents, and interneurons
DL-Int-1 and DL-Int-2, obtained from different prepara-
tions, were reconstructed in Amira 4.1 (Evers et al.,
2005). Subsequently, the neuropilar outlines were traced
manually and segmented with the Amira 4.1 label field edi-
tor. These neuropilar label fields were used to register the
segmented neuron of each preparation into the HSB fol-
lowing the method described by Brandt et al. (2005). This
registration process involves both affine transformations
and elastic geometric deformations. The affine transforma-
tion matrixes and the deformation vector fields were
applied to the segmented neurons, and these were fitted
into the HSB. To identify a possible overlap region
between the NH afferents and each morphologically and
physiologically characterized interneuron, Amira’s ‘‘surface
distance’’ was used to calculate the distance between
neurons. As landmarks for identifying the common projec-
tion areas of the sensory afferents of JO and NHs, the ven-
tral intermediate tract (VIT) and the maxillary dorsal com-
missures I and III (MxDCI and -III) were also registered
into the HSB. Adobe Photoshop 5.0 was used to enhance
the contrast of images and to store all photographs.
RESULTS
Central projection of three neuralsubgroups of NHs
Dye injection specifically into the central (n ¼ 7), dor-
sal (n ¼ 7), and ventral (n ¼ 6) NHs revealed projection
patterns corresponding to the three neural subgroups of
the NHs (Fig. 2). In general, the projection patterns of the
three subgroups were similar; afferents from each of the
neural subgroups supplied fine collaterals in the pro-TG
and sent axons both through the anterior VNC toward the
SEG and through the posterior VNC toward the meso-TG
(Fig. 2). In the dSEG, all afferents from different sub-
groups bifurcated into the dorsal and ventral branches
and then ramified to form fine collaterals with blebby ter-
minals (Fig. 2A–C). In the pro-TG, the NH afferents bifur-
cated into two main tracts close to the proximal region of
nerve N1. In the thicker tract, the axons extended close
to the midline of the pro-TG toward the posterior ipsilat-
eral VNC. The axons running in the VNC terminated along
the midline of the meso-TG (Fig. 2G–I). The other thinner
tract also sent axons to the midline of the pro-TG and
then bifurcated into two subtracts. One tract sent axons
toward the anterior ipsilateral VNC, and the other was
elongated posteriorly along the midline of the pro-TG,
with fine ramifications with blebby terminals (Fig. 2D–F).
To compare spatial projection patterns of the different
NH subgroups, pairs of the three subgroups were stained
differentially with fluorescent dyes. No dye coupling was
detected between the subgroups (Figs. 3C,D). However,
because the central NHs were very close to both the dor-
sal and the ventral NHs, dye mixing always occurred
between this group and the other groups during the
Primary Center of Vector Information in Bees
The Journal of Comparative Neurology | Research in Systems Neuroscience 2573

incubation following dye injection. Therefore, double
staining was carried out on the dorsal and ventral NH
groups. There were no clear differences between the pro-
jection areas of these two subgroups in the pro-TG and in
the meso-TG (data not shown). In the pro-TG and the
meso-TG, the afferents of the dorsal NHs and the ventral
NHs were in close apposition to each other. However, in
the dSEG, the bifurcating points of the dorsal and ventral
branches were clearly different (arrows in Fig. 3A,B; n ¼5). The bifurcating point of the afferents originating from
the dorsal NHs was more anterior to that of the ventral
NHs (arrows in Fig. 3B). The three-dimensional represen-
tation of axon terminals showed that their termination
fields were spatially well segregated in the dSEG: axon
Figure 2. Central projections of the afferents originating from the right-side NHs of the three subregions (dorsal NHs, central NHs and
ventral NHs). A,D,G show those of the dorsal NHs; B,E,H those of the central NHs; and C,F,H those of ventral NHs. The NHs project into
the subesophageal (SEG; A–C), prothoracic (pro-TG; D–F), and mesothoracic (meso-TG; G–I) ganglia. There were no clear differences in the
branching patterns of the afferents originating from NHs of the three subregions in these ganglia. The dSEG are outlined by dashed lines,
which is the terminal regions of the NH afferents in SEG. In F, the ventral NHs of the left side were also weakly stained. See Results for
details. Scale bars ¼ 50 lm in C (applies to A–C); 100 lm in F (applies to D–F); 100 lm in I (applies to G–I).
Ai and Hagio
2574 The Journal of Comparative Neurology |Research in Systems Neuroscience

terminals of the dorsal NHs were located anteriorly to
those of the ventral NHs (Fig. 3A,B). In the anterior region
of the dSEG, there were blebby terminals of the dorsal
NH afferents but not of the ventral NH afferents (arrow-
heads in Fig. 3C). In the posterior region of the dSEG, the
blebby terminals of both dorsal and ventral NH afferents
were intermingled (arrowheads in Fig. 3D). To visualize
the spatial maps of the terminal regions of the NH sub-
groups, we registered these afferents into the HSB
(Fig. 4A–C). The central projections of the NH subgroups
segregated in the dSEG along the anterior to posterior
axis (arrowheads in Fig. 4E,F). The JO afferents send
axons in each terminal region of the NH subgroups;
however, they do not cover the whole terminal regions of
the NHs (Fig. 4D–F).
Spatial relationship between the centralprojections of the antennal afferents andthe NH afferents in the dSEG
Both JO and NH afferents extend their axons into the
dSEG (Ai et al., 2007; Brockmann and Robinson, 2007).
The spatial relationship between these afferents was
examined in double-stained preparations. The axons of
the NH afferents pass through the ipsilateral VNC and ter-
minate in the ipsilateral dSEG (Fig. 2). They then bifurcate
into dorsal and ventral branches, which produce fine
arborizations in the dSEG terminating in close proximity to
the terminals of the antennal mechanosensory afferents
(magenta in Fig. 5A,B; n ¼ 11). Afferents from the hair
plates on the pedicel and from JO pass through T5 and
T6, respectively (Ai et al., 2007). The pedicel-hair-plate
Figure 3. Central projection of the dorsal (magenta) and ventral (green) NHs in the dSEG. A,B: Different locations of the bifurcation points
of the dorsal and ventral branches (A, frontal view; B, lateral view). The bifurcation point of the afferents originating from the dorsal
NHs was anterior to that of the ventral NHs (arrows). C,D: Frontal images in the optical planes indicated in B. C: In the anterior region of
the dSEG (left blue line in B), there were blebby terminals of the dorsal NH afferents, but there were no terminals of the ventral NH
afferents (arrowheads). D: In the posterior region of the dSEG (right blue line in B), blebby terminals of both dorsal and ventral NH affer-
ents are intermingled (arrowheads). Scale bars ¼ 20 lm.
Primary Center of Vector Information in Bees
The Journal of Comparative Neurology | Research in Systems Neuroscience 2575

afferents terminate in an anterior arborization within the
dSEG, whereas JO afferents terminate in a posterior arbo-
rization within the dSEG (Fig. 5D; Ai et al., 2007). The LSM
and three-dimensional images indicated that the sensory
afferents of the NHs overlapped with the posterior arbori-
zation of the antennal mechanosensory afferents T6II in
the dSEG, strongly suggesting that they were in close
proximity to the terminals of JO afferents (Fig. 5B–E).
We also compared the central projections of NH affer-
ents in worker honeybees with those in queens and
drones (Fig. 6; queens, n ¼ 2; drones n ¼ 6). Those in
queen and drone also projected into the dSEG. Overlap-
ping of JO and NH afferents was observed in all three hon-
eybee castes, indicating that the central projection of the
NHs was not unique to workers.
Spatial relationships between centralprojections of JO and NH afferents
To identify the common projection area of JO and NH
afferents in the dSEG, we referred to the HSB (Brandt
et al., 2005). The morphologies of JO afferents were ana-
lyzed in our previous studies (Ai et al., 2007) and were reg-
istered into the HSB (Ai, 2010). The confocal serial images
of the NH afferents and JO afferents obtained in the pres-
ent study were segmented and skeletonized and then reg-
istered into the HSB. This process demonstrated that the
axons of the NH afferents terminate close to axon termi-
nals T6II of the JO afferents in a location posterior to the
crossing of ventral intermediate tract (VIT) and maxillary
dorsal commissure I and III (MxDCI and -III; Fig. 7).
Spatial relationships between centralprojections of the NH afferents andmorphologically and physiologicallycharacterized vibration-sensitiveinterneurons
Previously, we morphologically and physiologically char-
acterized two vibration-sensitive interneurons (DL-Int-1
and DL-Int-2), which had dendritic arborizations in the
vibration primary center in the DL and the dSEG (Ai et al.,
2009). These interneurons had arborizations close to JO
terminals in the DL (Ai, 2010; Ai and Itoh, 2012), and both
Figure 4. Spatial relationships of the central projections in the dSEG of the NHs in the three subregions. Light blue, dorsal NHs; magenta,
central NHs; blue, ventral NHs; green, JO. A–C: Each of these central projections of the afferents was registered in the HSB (transparent
gray images). A, frontal view; B, lateral view; C, dorsal view. D–F: Magnified images of the boxed areas in A–C, respectively. These topolog-
ical projections reflect the peripheral NH subregions. E,F: The dorsal NHs (light blue arrow) terminated anterior to those of the central
NHs (magenta arrow), and the ventral NHs (blue arrow) terminated posterior to those of the central NHs. The transparent green-shaded
area shows the terminal region of JO.
Ai and Hagio
2576 The Journal of Comparative Neurology |Research in Systems Neuroscience

Figure 5. Double staining of antennal mechanosensory afferents and NH afferents. A: The NH afferents (magenta) terminate in close
proximity to the terminals of the antennal mechanosensory afferents (green) in dSEG. B: The sensory afferents of NHs bifurcate into dorsal
and ventral branches with fine arborizations in the dSEG (magenta). A,B: Frontal views. C–E: Three-dimensional reconstructed images of
NH afferents and antennal mechanosensory afferents. (C, frontal view; D, lateral view; E, dorsal view). The sensory afferents of NHs over-
lap the posterior arborization of the antennal mechanosensory afferents (T6II) in the dSEG, which are the terminals of JO afferents (Ai
et al., 2007). Scale bars ¼ 100 lm in A; 50 lm in B.
Figure 6. Central projections of antennal mechanosensory afferents (green) and NH afferents (magenta) in the dSEG of a drone (upper
images) and a queen honeybee (lower images). A,B: Image stack from the LSM (anterior views). C–H: Three-dimensional reconstructed
images of a drone (C, frontal view; D, lateral view; E, dorsal view) and a queen (F, frontal view; G, lateral view; H, dorsal view). In both
castes, NH afferents also projected to the terminals of T6II (arrows) in the dSEG, which is similar to that in worker honeybees. Scale bars
¼ 100 lm.

responded to vibration stimuli with tonic inhibitory, excita-
tory, or on–off phasic excitatory responses. These proper-
ties indicate that either neuron would be capable of moni-
toring the length of the vibration phase that encodes
distance information (Ai et al., 2009). In the present study,
we observed that the NH afferents terminated close to the
terminal region of JO afferents in the dSEG. To investigate
the spatial relationship between the NH afferents and
these vibration-sensitive interneurons, these interneurons
and the NH afferents were registered into the HSB (see
Figs. 8–10). This analysis revealed that the finest several
branches of the dorsal branch of DL-Int-1 are in close prox-
imity to the ventral branch of the NHs in the dSEG (Fig. 8).
It was also revealed that DL-Int-1 had more branches close
to the terminals of the dorsal NH subgroups than those
close to the other NH subgroups (Fig. 9). DL-Int-2 appears
to be an output neuron, whose axons extend from the DL-
dSEG to the lateral and posterior protocerebrum (Ai et al.,
2009). DL-Int-2 did not have dendritic arborization close to
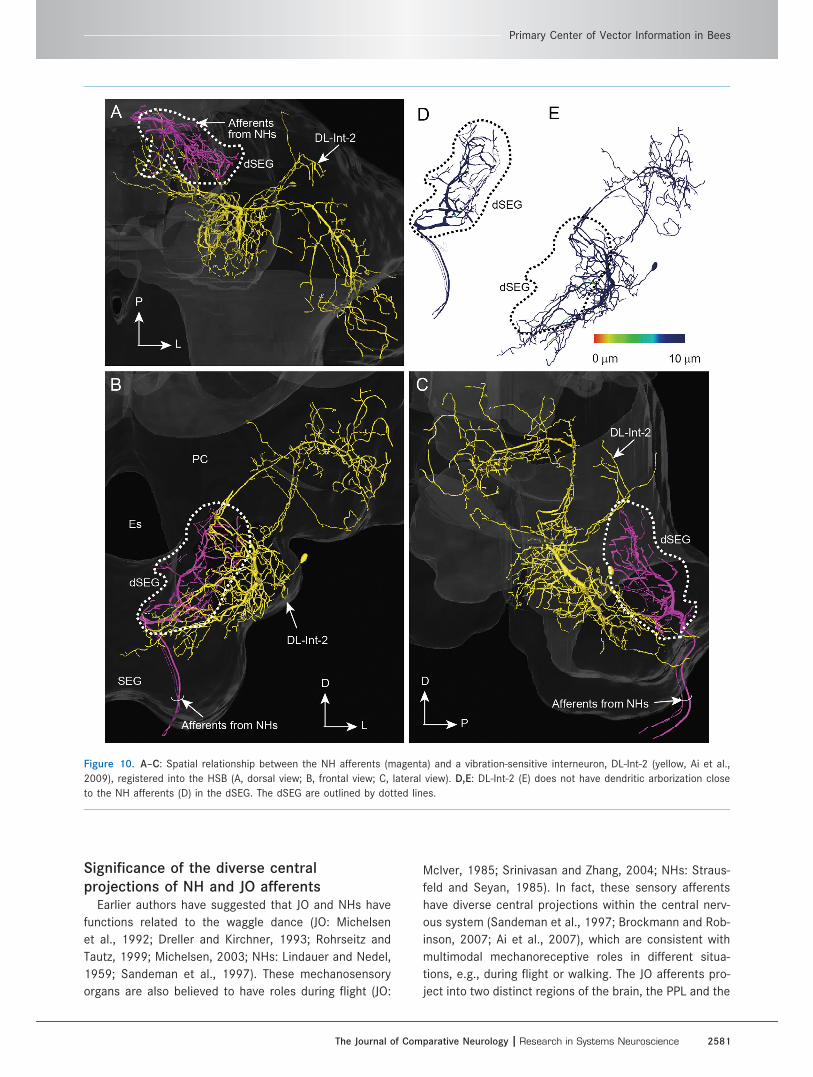
the NH afferents in the dSEG (Fig. 10).
Figure 7. A: NH afferents and JO afferents registered into the honeybee standard brain (HSB). B–D: Three-dimensional images of the NH
afferents and JO afferents (B, frontal view; C, dorsal view; D, lateral view) in the HSB (transparent image). B is a magnified image of the
boxed area in A. The axons of the NH afferents (magenta) terminate close to the axon terminals T6II of the JO afferents (green), enclosed
by yellow dotted lines. The dSEGs are outlined by white dotted lines. The common terminal region is situated posterior to the crossing of
VIT and MxDCI and -III.
Ai and Hagio
2578 The Journal of Comparative Neurology |Research in Systems Neuroscience

Figure 8. A–C: Spatial relationships between the NH afferents (magenta) and a vibration-sensitive interneuron, DL-Int-1 (orange; Ai et al.,
2009), registered into the HSB (A, dorsal view; B, frontal view; C, lateral view). D,E: The minor arborization of the dorsal branch of DL-Int-1
(arrowheads in E) is in close proximity to the ventral branch of the NHs in dSEG (arrowheads in D). The distances between the NH affer-
ents and the DL-Int-1 show in pseudocolors. The dSEG are outlined by dotted lines.
Primary Center of Vector Information in Bees
The Journal of Comparative Neurology | Research in Systems Neuroscience 2579

DISCUSSION
This study provides a morphological analysis of the pri-
mary center concerned with the vector information trans-
ferred by the waggle dance of honeybees. We demon-
strate that the topological organization of sensory
afferents within the dSEG reflects the subregions of the
peripheral NHs. Furthermore, the terminals of the NH
afferents within the dSEG are in close apposition to those
of JO afferents and to arborizations of the morphologically
and physiologically characterized vibration-sensitive inter-
neuron DL-Int-1. These features are common to the
worker, queen, and drone castes of honeybees.
Figure 9. A: Spatial relationships among the terminal region of three NH subgroups and a DL-Int-1, registered into the HSB (lateral
view). Light blue, dorsal NHs; magenta, central NHs; blue, ventral NHs; orange, DL-Int-1. B–D: The area close to the afferents of three
NH subgroups (B, dorsal NHs; C, central NHs; D, ventral NHs) on the DL-Int-1 in the HSB (frontal views). The distances between each
NH subgroup and the DL-Int-1 show in pseudocolors. The dSEG is outlined by dotted line. The arrowheads indicate the areas close to
the afferents of three NH subgroups.
Ai and Hagio
2580 The Journal of Comparative Neurology |Research in Systems Neuroscience
Significance of the diverse centralprojections of NH and JO afferents
Earlier authors have suggested that JO and NHs have
functions related to the waggle dance (JO: Michelsen
et al., 1992; Dreller and Kirchner, 1993; Rohrseitz and
Tautz, 1999; Michelsen, 2003; NHs: Lindauer and Nedel,
1959; Sandeman et al., 1997). These mechanosensory
organs are also believed to have roles during flight (JO:
McIver, 1985; Srinivasan and Zhang, 2004; NHs: Straus-
feld and Seyan, 1985). In fact, these sensory afferents
have diverse central projections within the central nerv-
ous system (Sandeman et al., 1997; Brockmann and Rob-
inson, 2007; Ai et al., 2007), which are consistent with
multimodal mechanoreceptive roles in different situa-
tions, e.g., during flight or walking. The JO afferents pro-
ject into two distinct regions of the brain, the PPL and the
Figure 10. A–C: Spatial relationship between the NH afferents (magenta) and a vibration-sensitive interneuron, DL-Int-2 (yellow, Ai et al.,
2009), registered into the HSB (A, dorsal view; B, frontal view; C, lateral view). D,E: DL-Int-2 (E) does not have dendritic arborization close
to the NH afferents (D) in the dSEG. The dSEG are outlined by dotted lines.
Primary Center of Vector Information in Bees
The Journal of Comparative Neurology | Research in Systems Neuroscience 2581

dorsal lobe-dorsal subesophageal ganglion neuromere
(DL-dSEG; Ai et al, 2007). Proposed roles for JO include
velocity detection during flight (Srinivasan and Zhang,
2004) and airborne vibration detection in the hive (Dreller
and Kirchner, 1995). The PPL is believed to be concerned
with the integration of vision with movements and veloc-
ity during flight because it is a region in which the visual
interneurons have terminal arborization. Correspondingly,
the DL-dSEG may be involved in the integration of me-
chanical (e.g., tactile and airborne vibrations) and gusta-
tory stimuli in the hive because it is a region in which the
antennal mechanosensory and gustatory neurons have
terminal arborizations (Haupt, 2007).
The NH afferents, on the other hand, project into three
distinct regions of the proto- and mesothoracic ganglia,
as well as into dSEG (Fig. 2), a pattern common to the
worker, queen, and drone castes (Fig. 6). It has been sug-
gested that the NHs monitor the angle between the head
and the thorax during flight in blowflies (Strausfeld and
Seyan, 1985) and the orientation of the body axis to the
vertical comb in honeybees (Sandeman et al., 1997). In
locusts (Braunig et al., 1983) and blowflies (Strausfeld
and Seyan, 1985), the sensory afferents of the cervical
hair plate and prosternal organ project into the pro- and
mesothoracic ganglia but not into the dSEG. These obser-
vations suggest that the termination of NH afferents in
the dSEG is unique to honeybees. One unique behavior of
honeybees is the waggle dance. The waggle dancer enco-
des the direction toward the food source as the body
angle on the vertical comb during the waggle phase of
the dance and the follower detects the body angle by
NHs (detector of direction; von Frisch, 1963). In this
study, the terminals of the NH afferents within the dSEG
are in close apposition to afferents from JO (which
detects distance). This suggests that the dSEG could
have a role in the integration of spatial information trans-
ferred by the waggle dance. However, the JO terminals
overlap only parts of the terminal regions of the NHs (Fig.
4D–F). The terminal regions of the NH subgroups might
therefore be related to other roles in addition to integrat-
ing spatial information transferred by the waggle dance.
Honeybees maintain negative geotaxis on the vertical
comb, and gravity-sensing organs, which include the NHs,
are able to detect the body’s angle (Sandeman et al.,
1997). The neuroanatomical features of the dSEG identi-
fied in this study might be related to these specific
aspects of honeybee behavior.
The present study shows that the NH afferents in
queen and drone also projected into the dSEG. Overlap-
ping of JO and NH afferents was observed in all three hon-
eybee castes, indicating that the central projection of the
NHs was not unique to workers (Fig. 6). The waggle dance
is used for transferring the vector information not only to
the profitable flowers but also to the swarming place to
the hivemates. Therefore, queen and drones might also
be able to decipher the vector information in their brains.
Significance of somatotopic organization ofthe NH afferents
Honeybees live on vertical combs within the hive.
Therefore, the direction of gravity sensed by individuals
varies depending on their orientation on the comb (Fig.
1). The inclination of the head relative to the thorax
changes with its orientation (Fig. 1B; Lindauer and Nedel,
1959). The NHs are arranged around the prosternal
appendage, forming a U shape on each side (Fig. 1C),
with long hairs in the dorsal and ventral subregions and
short hairs in the central subregion (Fig. 1A; Sandeman
et al., 1997). This distribution of hairs around the neck
suggests that honeybees may determine their orientation
with respect to gravity by sensing the inclination of the
head from the differential activation of specific subregions
of the NHs (Lindauer and Nedel, 1959). This hypothesis is
consistent with the present demonstration of a somato-
topic map in the dSEG that reflects the topological distri-
bution of the NHs. Thus, the aspects of honeybee’s spa-
tial orientation are represented in spatially organized
sensory projections in the dSEG. Although bilateral topo-
logical maps must reflect the topology of left and right
NHs in the bilateral dSEG, it is unclear whether interneur-
ons connect the two fields. In addition, the NH afferents
bifurcate into dorsal and ventral branches within the
dSEG (Fig. 2), but it remains to be established whether
these two branches are also related to the peripheral to-
pology of NHs or whether there are functional differences
between the dorsal and the ventral branches.
Hypothesis of the integration in thecommon projection area of JO and NHs
We observed that JO and NH afferents both project
into the dSEG. The afferents of JO project into the PPL
and the DL-dSEG, but a somatotopic map is present only
in the PPL (Ai et al., 2007). This suggests that there is no
spatial map of airborne vibrations in the dSEG. However,
as noted, a spatial map of the NHs in the dSEG appears
to be concerned with the bee’s orientation on the vertical
comb. Based on these observations, a model of the pri-
mary processing of the waggle dance information is
hypothesized (Fig. 11). In the example shown, a forager
(waggle dancer) has just returned from a feeder at a bear-
ing 90� west of the bearing of the sun from the hive and
performs a waggle dance oriented 90� to the left. The in-
hive honeybees (followers) that follow the waggle run
from position angles between 630� of the run are
successfully recruited (Judd, 1995), suggesting that
Ai and Hagio
2582 The Journal of Comparative Neurology |Research in Systems Neuroscience

successfully recruited followers follow the dancer from its
tail end and align themselves parallel to the dancer to
detect the dancer’s body angle (Rohrseitz and Tautz,
1999). In this example, the left central NHs of the follower
are activated, and these afferents send signals to the cor-
responding subregion in the dSEG (in the example, ‘‘–90�’’
in the dSEG), whereas the afferents of JO also send signals
to the part of dSEG. In the dSEG, only the �90� subregion
is able to be activated by signals received from JO and the
left central NHs. We analyzed the spatial relationship
between the NH terminals and two morphologically and
physiologically characterized vibration-sensitive interneur-
ons DL-Int-1 and DL-Int-2 in the HSB (Figs. 8–10). The den-
dritic arborization of DL-Int-1 is close to the terminals of
the NHs in dSEG (Figs. 8, 9). In our previous study, the DL-
Int-1 was also close to the terminals of the JOs in the DL-
dSEG (Ai and Itoh, 2012), and DL-Int-1 has responsiveness
to the olfactory stimulus to the contralateral antenna (Ai
et al., 2009). HSB analysis does not allow visualization of
the synaptic contacts between different neurons, but it is
possible that these interneurons have synaptic inputs from
JO, NH afferents, and olfactory interneurons in the DL-
dSEG. Moreover, it has been found that there are other dif-
ferent types of vibration-sensitive interneurons arborized in
the DL-dSEG on our intracellular recording and staining
experiments and that some of them have dendritic arbori-
zation close to the terminals of the JO and NH afferents
(Ai, unpublished observations). By analyzing the morpholo-
gies and response properties of these vibration-sensitive
interneurons during stimulation of different NH subregions
and during the olfactory stimulation, it should be possible
to clarify whether these interneurons are concerned not
only with vibration processing but also with the integration
of sensory information contained in the waggle dance.
ACKNOWLEDGMENTS
I thank Prof. Dr. Randolf Menzel, Institut fur Neurobio-
logie, Freie Universit€at Berlin, and Dr. Jurgen Rybak, Max
Planck Institute for Chemical Ecology, for help with the
registration of the confocal images of neurons into the
HSB and for helpful comments on this research.
CONFLICT OF INTEREST STATEMENTThe authors declare no conflict of interest.
ROLE OF AUTHORSAll authors had full access to all the data in the study and
take responsibility for the integrity of the data and the
Figure 11. Putative model of the primary processing of the waggle dance information. In this example, the forager (waggle dancer) has
just returned from a feeder whose bearing is 90� west of the bearing of the sun to the hive. The bee performs a waggle dance oriented
90� to the left. The in-hive honeybee (follower) follows the dancer from its tail end. The left central NHs are activated, and the afferents
send signals to the corresponding subregion in the dSEG (shown as �90� in the dSEG). The afferents of JO also send signals to the part
of the �90� degree subregion in dSEG. In the dSEG, only the part of the �90� degree subregion can be activated by signals received
from JO and the left central NHs.
Primary Center of Vector Information in Bees
The Journal of Comparative Neurology | Research in Systems Neuroscience 2583

accuracy of the data analysis. Study concept and design:
HA. Acquisition of data:HA,HH. Analysis and interpretation
of data: HA,HH.Drafting of themanuscript: HA,HH.Critical
revision of the manuscript for important intellectual
content: HA. Obtained funding: HA. Administrative, tech-
nical, and material support: HA. Study supervision: HA.
LITERATURE CITEDAi H. 2010. Vibration-processing interneurons in the honeybee
brain. Front Syst Neurosci doi: 10.3389/neuro.06.019.2009.Ai H, Itoh T. 2012. The auditory system of the honeybee. In:
Eisenhardt D, Galizia CG, Giurfa M, editors. Honeybee neu-robiology and behaviors, 2nd ed. Berlin: Springer Verlag.p 269–284.
Ai H, Nishino H, Itoh T. 2007. Topographic organization ofsensory afferents of Johnston’s organ in the honeybeebrain. J Comp Neurol 502:1030–1046.
Ai H, Rybak J, Menzel R, Itoh T. 2009. Response characteris-tics of vibration-sensitive interneurons related to John-ston’s organ in the honeybee, Apis mellifera. J CompNeurol 515:145–160.
Abel R, Rybak J, Menzel R. 2001. Structure and response pat-terns of olfactory interneurons in the honeybee, Apis melli-fera. J Comp Neurol 437:363–383.
Arnold G, Masson C, Budharugsa S. 1985. Comparative studyof the antennal lobes and their afferent pathway in theworker bee and the drone (Apis mellifera). Cell Tissue Res242:593–605.
Brandt R, Rohlfing T, Rybak J, Krofczik S, Maye A, WesterhoffM, Hege H, Menzel R. 2005. Three-dimensional average-shape atlas of the honeybee brain and its applications.J Comp Neurol 492:1–19.
Braunig P, Pfluger H-J, Hustert R. 1983. The specificity ofcentral nervous projections of locust mechanoreceptors.J Comp Neurol 218:197–207.
Brockmann A, Robinson GE. 2007. Central projections ofsensory systems involved in honey bee dance languagecommunication. Brain Behav Evol 70:125–136.
Dreller C, Kirchner WH. 1993. How honeybees perceive theinformation of the dance language. Naturwisshenschaften80:319–321.
Dreller C, Kirchner WH. 1995. The sense of hearing in honeybees. Bee World 76:6–17.
Evers J-F, Schmitt S, Sibilia M, Duch C. 2005. Progress infunctional neuroanatomy: precise automatic geometricreconstruction of neuronal morphology from confocalimage stacks. J Neurophysiol 93:2331–2342.
Flanagan D, Mercer AR 1989. Morphology and response char-acteristics of neurones in the deutocerebrum of the brainin the honeybee Apis mellifera. J Comp Physiol A 164:483–494.
Frisch von K. 1967. The tail-wagging dance as a means ofcommunication when food sources are distant. In: vonFrisch K, editor. The dance language and orientation ofbees. Cambridge, MA: Belknap Press of Harvard UniversityPress. p 57–235.
Haupt SS. 2007. Central gustatory projections and side-speci-ficity of operant antennal muscle conditioning in the hon-eybee. J Comp Physiol A 193:523–535.
Ignell R, Dekker T, Ghaninia M, Hansson BS. 2005. Neuronalarchitecture of the mosquito deutocerebrum. J Comp Neu-rol 493:207–240.
Judd TM. 1995. The waggle dance of the honey bee: whichbees following a dancer successfully acquire the informa-tion? J Insect Behav 8:343–354.
Kamikouchi A, Shimada T, Itoh K. 2006. Comprehensive clas-sification of the auditory sensory projections in the brainof the fruit fly Drosophila melanogaster. J Comp Neurol499:317–356.
Kelber C, R€ossler W, Kleineidam CJ. 2006. Multiple olfactoryreceptor neurons and their axonal projections in the anten-nal lobe of the honeybee Apis mellifera. J Comp Neurol496:395–405.
Kirschner S, Kleineidam CJ, Zube C, Rybak J, Grunewald B,R€ossler W. 2006. Dual olfactory pathway in the honeybeeApis mellifera. J Comp Neurol 499:933–952.
Kloppenburg P. 1995. Anatomy of the antennal motoneuronsin the brain of the honeybee (Apis mellifera). J Comp Neu-rol 363:333–343.
Lindauer M, Nedel JO. 1959. Ein Schweresinnesorgan derHonigbiene. Z Physiol 42:334–364.
Maronde U. 1991. Common projection areas of antennal andvisual pathways in the honeybee brain, Apis mellifera.J Comp Neurol 309:328–340.
McIver SB. 1985. Mechanoreception. In: Kerkut GA, Gilbert LI,editors. Comprehensive insect physiology, biochemistryand pharmacology, vol 6. Oxford: Pergamon Press.p 71–132.
Michelsen A. 2003. Signals and flexibility in the dance com-munication of honeybees. J Comp Physiol A 189:165–174.
Michelsen A, Andersen BB, Storm J, Kirchner WH, Lindauer M.1992. How honeybees perceive communication dances,studies by means of a mechanical model. Behav Ecol Soci-obiol 30:143–150.
Mobbs PG. 1982. The brain of the honeybee Apis mellifera. I.The connections and spatial organization of the mushroombodies. Philos Trans R Soc Lond B Biol Sci 298:309–354.
Pareto A. 1972. Die Zentrale Verteilung der Fuehlerafferenzbei Arbeiterinnen der Honigbiene, Apis mellifera L. Z Zell-forsch 131:109–140.
Rehder V. 1988. A neuroanatomical map of the suboesopha-geal and prothoracic ganglia of the honey bee (Apis melli-fera). Proc R Soc Lond B Biol Sci 235:179–202.
Riley J, Greggers U, Smith AD, Reynolds DR, Menzel R. 2005.The flight paths of honeybees recruited by the waggledance. Nature 435:205–207.
Rohlfing T, Brandt R, Menzel R, Maurer CR Jr. 2004. Evalua-tion of atlas selection strategies for atlas-based image seg-mentation with application to confocal microscopy imagesof bee brains. Neuroimage 21:1428–1442.
Rohrseitz K, Tautz J. 1999. Honeybee dance communication:waggle run direction coded in antennal contacts? J CompPhysiol A 184:463–470.
Sandeman D, Sandeman R, Tautz J. 1997. Head andabdominal posture and the orientation of honeybees onvertical combs. Zoology 100:85–97.
Srinivasan MV, Zhang S. 2004. Visual motor computations ininsects. Annu Rev Neurosci 27:679–696.
Strausfeld NJ, Seyan HS. 1985. Convergence of visual, hal-tere, and prosternal inputs at neck motor neurons of Calli-phora erythrocephala. Cell Tissue Res 240:601–615.
Suzuki H. 1975. Antennal movements induced by odour andcentral projection of the antennal neurons in the honey-bee. J Insect Physiol 21:831–847.
Ai and Hagio
2584 The Journal of Comparative Neurology |Research in Systems Neuroscience