nucleus and mitochondria: structure and...
TRANSCRIPT

Nucleus and Mitochondria: structure and disease
Peter TakizawaCell Biology

•Nuclear membrane structure
•Nuclear lamins and disease
•Mitochondria localization and dynamics
•Mitochondria fusion and disease

Nuclear membrane and disease

Nuclear envelope is a double lipid bilayer with supporting cytoskeleton and nuclear pores.
Pollard et al. Cell Biology 2nd Edition
The nuclear envelope separates nuclear space from cytoplasm. It protects DNA, provides mechanical support and regulates gene expression. The nuclear envelope consist of two membrane bilayers. The outer nuclear membrane is continuous with the endoplasmic reticulum and the inner membrane faces the nuclear space and DNA. Nuclear pores span both membranes to allow passage of proteins and RNAs between the cytoplasm and nucleus. Underneath the inner membrane is mesh of intermediate filaments called the nuclear lamina. It provides structural support for the nuclear membrane, anchors the nuclear pores and associates with chromosomes.
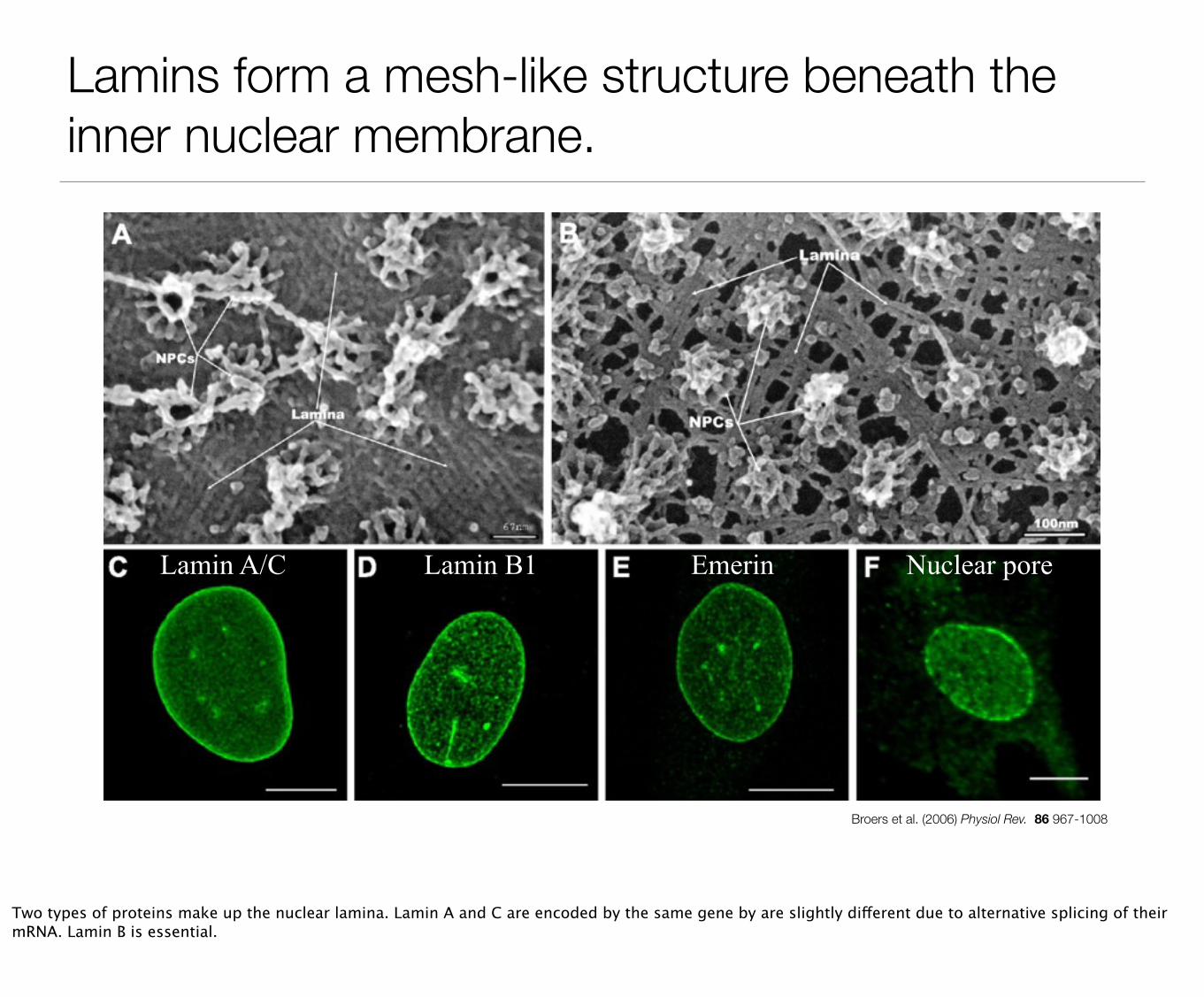
Lamins form a mesh-like structure beneath the inner nuclear membrane.
Broers et al. (2006) Physiol Rev. 86 967-1008
Lamin A/C Lamin B1 Emerin Nuclear pore
Two types of proteins make up the nuclear lamina. Lamin A and C are encoded by the same gene by are slightly different due to alternative splicing of their mRNA. Lamin B is essential.

Lamins are structurally similar to intermediate filaments.
NH2 COOH
Lamins are similar in structure to intermediate filaments. They contain a long alpha helical domain that mediates dimerization. The globular ends provide functions such as importing lamins into the nucleus and mediating association with protein in the inner nuclear membrane. Lamin dimers associated end to end to form polymers.
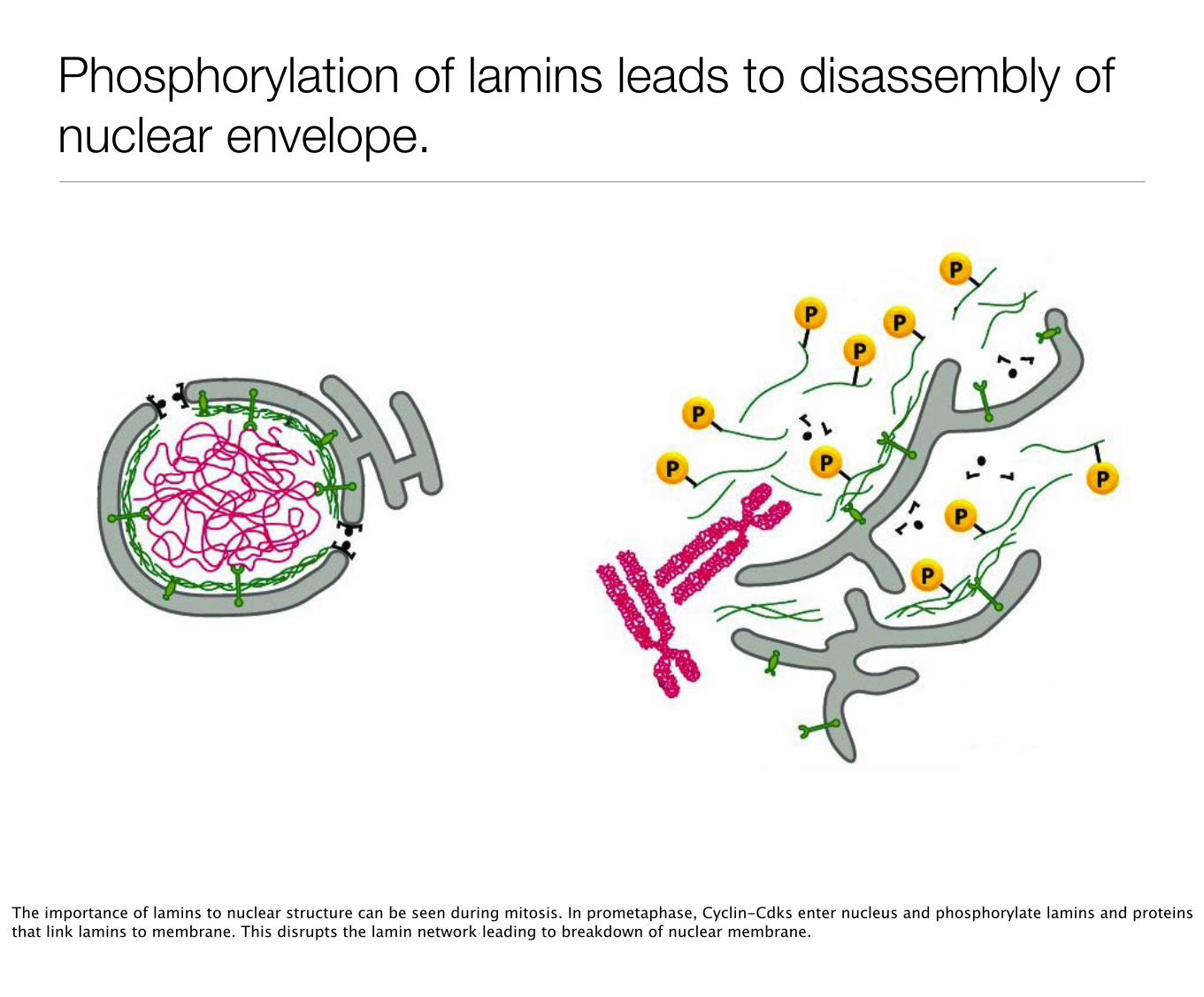
Phosphorylation of lamins leads to disassembly of nuclear envelope.
The importance of lamins to nuclear structure can be seen during mitosis. In prometaphase, Cyclin-Cdks enter nucleus and phosphorylate lamins and proteins that link lamins to membrane. This disrupts the lamin network leading to breakdown of nuclear membrane.

Mutations in nuclear lamins associated with a diverse set of diseases.
Striated Muscle Diseases
Emery-Dreifuss muscular dystrophy
Limb-girdle muscular dystrophy
Dilated cardiomyopathy
Congenital muscular dystrophy
Heart-hand syndrome
Lipodystrophy Syndromes
Dunnigan-type familial partial lipodystrophy
Mandibuloacral dysplasia
Lipoatrophy
Partial lipodystrophy
Accelerated Aging Disorders
Atypical Werner syndrome
Hutchinson-Gilford progeria syndrome
Restrictive dermopathy
Atypical progeria syndrome
Peripheral Nerve Disorders
Charcot-Marie-Tooth disease
Adult-onset leukodystrophy
Spinocerebellar ataxia type 8
Bone Diseases
Buschke-Ollendorff syndrome
Melorheostosis
Osteopoikilosis
Greenberg skeletal dysplasia
Over 200 diseases have been associated with mutations in lamin genes. Most mutations are in lamin A and lead to wide variety of phenotypes. How do mutations in one gene lead to such diverse phenotypes? Currently, we don’t know the answer. Mutations in lamin A likely affect nuclear structure and possible gene expression.

Mutations in nuclear lamins associated with a diverse set of diseases.
DNA
Lamin A
HGPNormal
Over 200 diseases have been associated with mutations in lamin genes. Most mutations are in lamin A and lead to wide variety of phenotypes. How do mutations in one gene lead to such diverse phenotypes? Currently, we don’t know the answer. Mutations in lamin A likely affect nuclear structure and possible gene expression.

Nuclear lamins are linked to cytoskeleton and may integrate mechanical stress with gene regulation.
Recent work has connected lamins in the nucleus with the cytoskeleton in the cytoplasm, including both actin filaments and microtubules. The connections are thought to help cells position the nucleus within the cell via molecular motor proteins. In addition, the link between cytoskeleton and lamins allows communication of mechanical forces on the cell with the nucleus, which may allow cells to activate gene expression in response to changes in mechanical stress.

Mitochondria transport and dynamics

Mitochondria contain two lipid bilayers.
Mitochondria contain two membranes: an outer membrane and an inner membrane that invaginates to form cristae.

Cells localize mitochondria to regions of high energy consumption.
Cells localize mitochondria to areas of high energy use. Epithelial cells move mitochondria to regions where there are pumps actively removing or taking up material. In neurons, mitochondria localize throughout dendrites and axons to provide energy for release of synaptic vesicles at axon terminus or processing of signals in dendrites. Given the size of cells, especially neurons, motor proteins are needed to transport mitochondria.

Kinesin and dynein transport mitochondria bidirectionally in axons.
Mitochondria move bidirectionally in axons. Kinesins move mitochondria toward the terminus of an axon whereas dynein move mitochondria toward the cell body.

Mitochondria undergo fusion and fission.
Chan, DC (2006) Cell 125 1241- 1252
Besides movement along microtubules, mitochondria also fuse with each other and then divide into two mitochondria.

Drp1 mediates fission of mitochondria.
Fission of mitochondria is driven by Drp1 which binds and polymerizes on the surface of mitochondria. Drp1 undergoes a conformational change to squeeze and divide mitos into two.

Mitofusins and Opa1 mediate mitochondrial fusion.
Fusion of mitochondria occurs in two steps: fusion of the outer membrane and fusion of the inner membrane. Mitofusins reside on the outer membrane mediate fusion of the outer membrane. Mitofusins have a structure similar to SNAREs with extended helical regions that might pair to drive membrane fusion. Fusion of the inner membrane fusion is driven by Opa1. Complete fusion of the outer and inner membranes allows for mixing of proteins and DNA
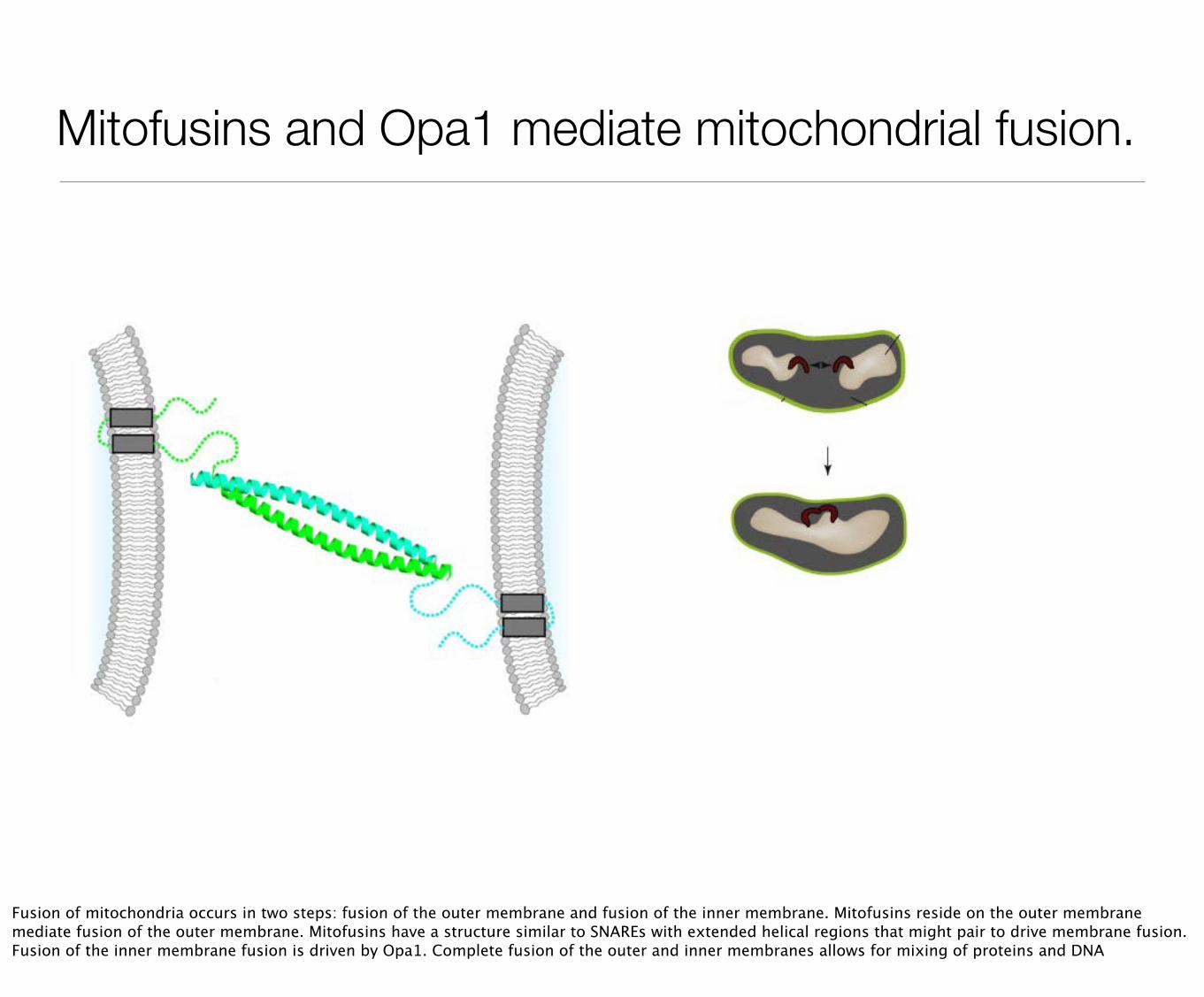
Mitofusins and Opa1 mediate mitochondrial fusion.
Fusion of mitochondria occurs in two steps: fusion of the outer membrane and fusion of the inner membrane. Mitofusins reside on the outer membrane mediate fusion of the outer membrane. Mitofusins have a structure similar to SNAREs with extended helical regions that might pair to drive membrane fusion. Fusion of the inner membrane fusion is driven by Opa1. Complete fusion of the outer and inner membranes allows for mixing of proteins and DNA

Chen, et al, (2003) J Cell Biol 160 189-200
Mitofusin mutants fail to fuse and have reduced size.
Wild-Type
mitofusin mutant
Mutations in mitofusins result in smaller mitochondria. To demonstrate the role of mitofusins in mitochondria fusion, two cells with differently labeled mitochondria were fused. In wild-type cells the green and red mitochondria fuse producing yellow mitochondria. In the mitofusin mutant the green and red mitchondria remain distinct. Mitochondria in cells that lack mitofusin have reduced activity.

Mitochondrial fusion mutants associated with neurodegenerative diseases.
Wild-type
mitofusin mutant
Several neurodegenerative diseases are associated with mitochondria fusion defects. Mutation in mitofusins cause of Charcot-Marie-Tooth disease, peripheral neuropathy. Mutations in Opa1 cause dominant optic atrophy, a degeneration of neurons in the retina, which results in a loss of visual acuity. Recent work suggests a role for mitochondria in both Parkinson’s and Alzheimer’s diseases.

Mitochondrial fusion mutants associated with neurodegenerative diseases.
Several neurodegenerative diseases are associated with mitochondria fusion defects. Mutation in mitofusins cause of Charcot-Marie-Tooth disease, peripheral neuropathy. Mutations in Opa1 cause dominant optic atrophy, a degeneration of neurons in the retina, which results in a loss of visual acuity. Recent work suggests a role for mitochondria in both Parkinson’s and Alzheimer’s diseases.
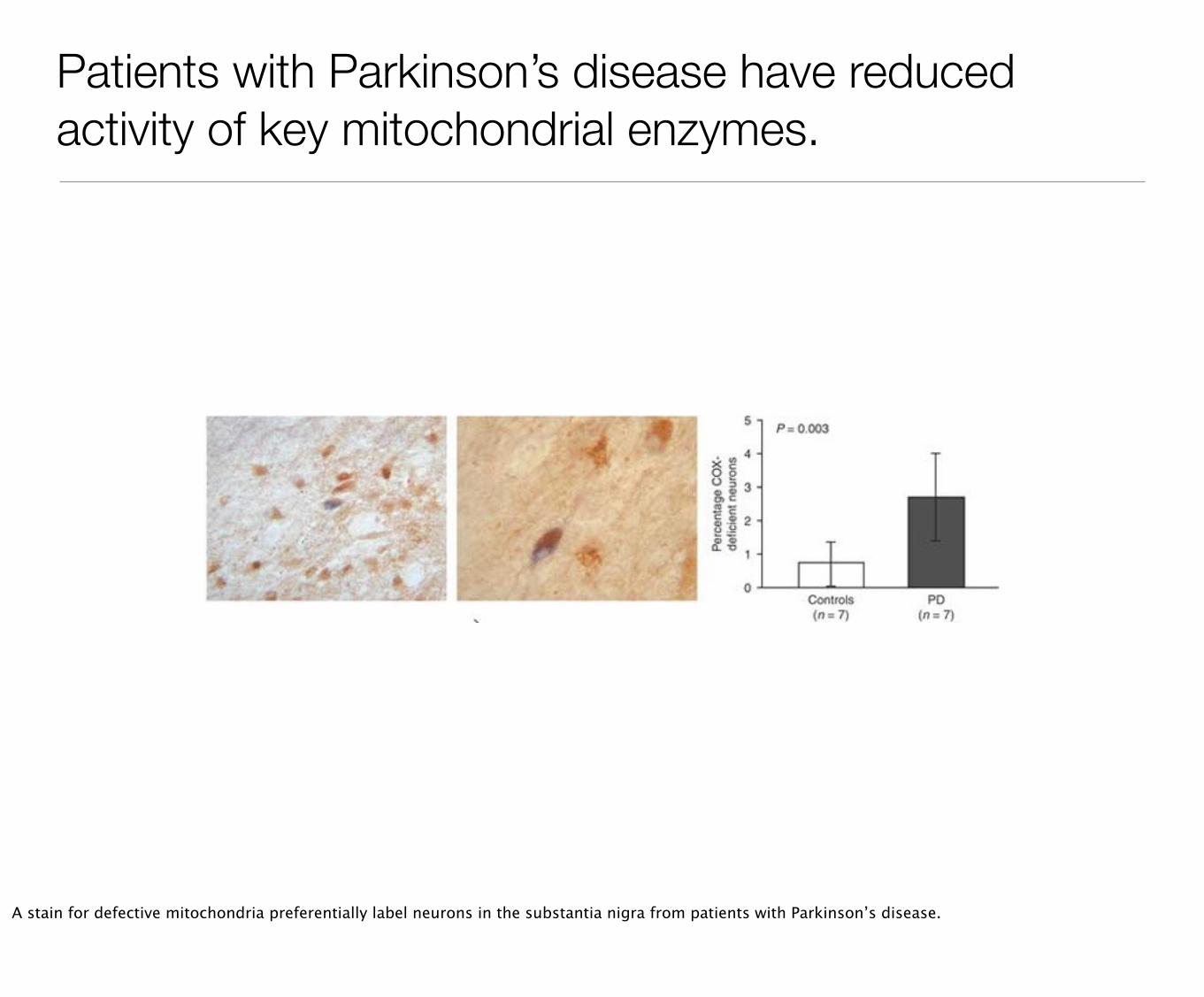
Patients with Parkinson’s disease have reduced activity of key mitochondrial enzymes.
A stain for defective mitochondria preferentially label neurons in the substantia nigra from patients with Parkinson’s disease.

Disruption of mitochondrial fusion leads to neurodegeneration.
Chen et al., (2007) Cell 130 548-562
Mice with mitofusins knocked out in cells of the cerebellum lack ability to move and coordinate movement. Mutant mice display an atrophied cerebellum with reduced numbers of neurons. A stain for apoptotic cells reveals increase numbered of dying cells in the cerebellum of mutant mice.

Disruption of mitochondrial fusion leads to neurodegeneration.
Chen et al., (2007) Cell 130 548-562
Mice with mitofusins knocked out in cells of the cerebellum lack ability to move and coordinate movement. Mutant mice display an atrophied cerebellum with reduced numbers of neurons. A stain for apoptotic cells reveals increase numbered of dying cells in the cerebellum of mutant mice.

Disruption of mitochondrial fusion leads to neurodegeneration.
Chen et al., (2007) Cell 130 548-562
Mice with mitofusins knocked out in cells of the cerebellum lack ability to move and coordinate movement. Mutant mice display an atrophied cerebellum with reduced numbers of neurons. A stain for apoptotic cells reveals increase numbered of dying cells in the cerebellum of mutant mice.

Disruption of mitochondrial fusion leads to neurodegeneration.
Chen et al., (2007) Cell 130 548-562
Mice with mitofusins knocked out in cells of the cerebellum lack ability to move and coordinate movement. Mutant mice display an atrophied cerebellum with reduced numbers of neurons. A stain for apoptotic cells reveals increase numbered of dying cells in the cerebellum of mutant mice.

Mitochondrial genome contains genes essential for function.
Mitochondria contain own DNA that encodes rRNAs, tRNAs and 13 proteins involved in oxidative phosphorylation and ATP synthesis. These genes are not replaceable by nuclear genes, so they are essential for functioning of mitochondria. Mitochondria contain several copies of their genome probably to protect the integrity of their genome. The rate of DNA mutations and deletions is higher in mitochondria due to electron transport chain and less robust repair system.

Each mitochondria contains several copies of its own genome.
Most mitochondria contain multiple genomes some of which will contain few mutation and others that may contain several mutations which result in non-functioning DNA. If a mitochondria contains too many mutated genomes, it will lose the ability to express proteins that are essential to its function.

Mitochondria fusion allows for exchange of damaged mutated DNA.
Mutated DNA and protein
Normal DNA and protein
One way that mitochondria fusion may increase their viability is by allowing repair of mitochondrial genomes. Fusion between mitochondria allows for mixing of components of both mitochondrion, including DNA, protein and lipid. If a mitochondrion that has a mostly mutated genomes fuses with a mitochondrion with several copies of wild-type genome, the genomes would mix and distribute throughout the fused mitochondrion. After fusion, each mitochondrion would receive wild-type and mutated genomes and both mitochondrion might now contain a sufficient number of wild-type genomes to maintain viability.

Mitochondria fusion allows for exchange of damaged mutated DNA.
Mutated DNA and protein
Normal DNA and protein
One way that mitochondria fusion may increase their viability is by allowing repair of mitochondrial genomes. Fusion between mitochondria allows for mixing of components of both mitochondrion, including DNA, protein and lipid. If a mitochondrion that has a mostly mutated genomes fuses with a mitochondrion with several copies of wild-type genome, the genomes would mix and distribute throughout the fused mitochondrion. After fusion, each mitochondrion would receive wild-type and mutated genomes and both mitochondrion might now contain a sufficient number of wild-type genomes to maintain viability.

Inhibition of mitochondrial fusion increases mutation rate in mitochondrial DNA.
Controlmitofusin mutant
These results show that mitochondria that cannot fuse, have more mutations and deletions in their genomes than mitochondria that can fuse.

Inhibiting mitochondrial fusion leads to loss of mitochondrial DNA.
Wild-type
mitofusin mutant
Chen et al., (2007) Cell 130 548-562
Over time mitochondria that cannot fuse, lose their genomes and become non-functional.

Mitochondria fusion allows healthy mitochondria to complement damaged mitochondria.
Fusion
Fission
Mitochondrial fusion allows a healthy mitochondria to share wild-type genomes and proteins with damaged mitochondria.

Mitochondria fission may allow for removal of debris and damaged mitochondria.
Fission of mitochondria may allow cells to partition damaged material into one mitochondria and then dispose of that mitochondria via mitophagy.

Genetic causes of parkinsonism Genetic causes of parkinsonism Genetic causes of parkinsonismGene Age at onsetGene productPARK1/4 40s α-SynucleinPARK2 20s+ ParkinPARK3 60s ..UCH-L1 50s Ubiquitin thiolesterasePINK1 30s PTEN-induced putative kinase 1PARK7 30s DJ-1LRRK2 Variable Leucine-rich repeat kinase 2ATP13A2 Variable ATPase type 13A2PARK10 .. ..PARK11 .. ..
Pink1 and Parkin bind to damaged mitochondria and target them form mitophagy and destruction.
Parkin = E3 ligase
Two proteins, Pink1 and Parkin, are required to target proteins for mitophagy and destruction in the lysosome. Mutations in both proteins cause Parkinson’s disease.

Take home points...
• Lamins provide structural support to nuclear membrane
• Mutations in lamin A lead to a disparate set of diseases
• Mitochondria undergo rounds of fusion and fission to maintain pool of highly active mitochondria
• Defects in mitochondria fusion and fission are associated with neurodegeneration
• Mitophagy destroys old and damaged mitochondria and is defective in patients with Parkinson’s disease