of p. printedin icr-induced frameshift mutations …jb.asm.org/content/101/2/490.full.pdf ·...
TRANSCRIPT

JOURNAL OF BACTERIOLOGY, Feb. 1970, p. 490-504Copyright a 1970 American Society for Microbiology
Vol. 101, No. 2Printed in U.S.A.
ICR-Induced Frameshift Mutations in the HistidineOperon of Salmonella
NICOLE S. OESCHGER1 AND PHILIP E. HARTMAN
Department ofBiology, The Johns Hopkins University, Baltimore, Maryland 21218
Received for publication 19 November 1969
Both the acridine half-mustard, ICR191, and the nonalkylating azaacridine de-rivative, ICR364-OH, induce three classes of frameshift mutations in the histidineoperon of Salmonella typhimurium. (i) One class is completely stable in reversiontests and is presumed to represent deletion of one or a few critical nucleotide pairsor two nearby frameshifts. One extended deletion was found out of 11 stable muta-tions. (ii) Of two spontaneously reverting classes which also are considered to pre-
dominantly involve base deletions, one is unaffected in reversion with ICR191,nitrosoguanidine, and diethylsulfate, and the other is induced to revert with ICR191.(iii) A third class, considered to predominantly involve base additions, respondsin reversion tests with ICRl91 as well as with nitrosoguanidine and diethylsulfate.Other investigators have shown that one mutant of this class is a "plus" frameshiftand that nitrosoguanidine acts in reversion to delete a guanine plus cytosine basepair. Although such plus frameshifts are found with high frequency among muta-tions selected from acridine-treated bacteria or when strong selection pressure isapplied for their detection in reversion tests, data from this laboratory indicatethat this class of plus frameshifts is rare among mutations derived spontaneously orafter treatment with a variety of other mutagens. Finally, we demonstrate that thealkylating ICRl91 and the nonalkylating ICR364-OH preferentially cause muta-tions in different chromosome regions and that their spectra of activity only partiallyoverlap that found for spontaneous frameshift mutations.
Although simple acridine dyes have been re-ported to cause mutations in bacteria (28, 30),they are relatively ineffective on nonmating,haploid bacteria. Recently, a series of acridinehalf-mustards has been found to be very activein inducing mutations in Drosophila (11), Neu-rospora (13, 21), and bacteria (5, 36). The data,mainly obtained from reversion studies, indicatethat the acridine half-mustards predominantlyelicit frameshift mutations in bacteria.We elected to isolate histidine-requiring mu-
tants in Salmonella typhimurium induced byICR191 (Fig. 1), one of the more potent acridinehalf-mustards tested by Ames and Whitfield (5).In addition, an azaacridine derivative ICR-364-OH (Fig. 1), incapable of alkylation, wasused to induce a second group of histidine-requiring mutants. The 80 mutations wereassigned to subregions of the histidine operon bytransduction tests, and the mutations were classi-fied by the reversion patterns elicited by severalchemical mutagens. Some mutations also were
'Present address: Department of Molecular Biophysics andBiochemistry, Yale University, New Haven, Conn. 06520.
examined for effects on polarity and for behaviorin intragenic complementation. The propertiesof most of the mutants were those expected ofstrains carrying frameshift mutations, except thatmany reverted in the presence of N-methyl-N'nitro-N-nitrosoguanidine (NG) and diethyl-sulfate (DES). We hypothesize that these lattertwo mutagens, although highly effective ineliciting base substitutions, also can cause basedeletions. Data of Yourno and co-workers (39,40) substantiate this conclusion. We also ob-served that the two acridine and azaacridinederivatives exhibited differences with regard totheir ability to cause mutations in differentregions of the histidine operon and in nonsense-suppressor genes.
MATERIALS AND METHODS
Media. Difco nutrient broth and Vogel and Bonner(33) E mineral salts medium were used in most experi-ments. E medium was supplemented, where required,with 20 Ag of L-histidine -HCl per ml, 150 jug of L-his-tidinol-2HCI per ml, 2% (by volume) of a pool ofamino acids complete except for histidine, 1.25% (EM
490
on July 12, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

ICR-INDUCED FRAMESHIFTS
medium), or 2.5% (2 EM medium) liquid Difco nu-trient broth.
Chemicals. 2-Aminopurine nitrate, B grade, waspurchased from Calbiochem, DES from Fisher Scien-tific Co., NG from Aldrich Chemical Co., and strep-tomycin from E. R. Squibb and Sons. L-Histidinoland histidinol phosphate were from Cyclo ChemicalCorp., diphosphopyridine nucleotide from SigmaChemical Co., and phenazine methosulfate and 2-p-iodophenyl-3 -p - nitro - phenyl - 6 - phenyltetrazoliumchloride from Dajac Laboratories. Adenosine-5'-tri-phosphate was purchased from Schwarz BioResearchInc., and phosphoribosyl pyrophosphate from PabstLaboratories. 2-Chloro-6-methoxy-9- [3- (2-chloro-ethyl)aminopropylamino] acridine dihydrochloride(ICR191) and 2-chloro-6-methoxy-9-[2-(2-hydroxy-ethyl)aminoethylamino]-l-azaacridine dihydrochlo-ride (ICR364-OH) were the generous gifts of Hugh J.Creech, Institute for Cancer Research, Philadelphia,Pa.
Bacterial strains. Mutations leading to a histidinerequirement were induced in strain LT-2 of S. typhi-muriwn carrying either a stable arabinose mutation,ara-9 (22), or an operator constitutive mutation,hisO1242 (27). Other histidine-requiring mutants ofS. typhimurium mentioned are on stock in this lab-oratory (19). Strains SB391 and SB392 were kindlyprovided by David B. Berkowitz (9). Charles Yanofskysupplied a strain of Escherichia coli K-12, which isthr-, leui and carries a mutator gene (37).
Genetic crosses and complementation tests. Multi-step lysates of bacteriophage P22 mutants L3 and L4(29) were prepared in nutrient broth and stored at 4 Cin screw-cap tubes over chloroform. Mapping of mu-tants was done by transduction using spot tests (2)and more sensitive crosses (19). Complementationtests on hisD mutants utilized deletion strains carry-ing E. coli F'his- episomes, strains TR50 (Db2380),TR49 (Da2379), and TA46 (Dab2482) (Greeb et al.,submitted for publication).
Isolation of ICR-induced mutants. Bacteria weregrown to 1.5 X 108 cells/ml in minimal E mediumand diluted 1:100 into E medium supplemented with10 ,ug of L-histidine-HCl per ml. Either 30 or 40 Mug ofICR191 per ml or 20 or 30 Mg of ICR364-OH per mlwas added to experimental cultures. Each of the cul-tures grew exponentially at 37 C for at least five gen-erations, and reached stationary phase at 3 X 109 to4 X 10' cells/ml. The generation times were 48 mmnfor a control culture, 60 min for a culture grown with40 ;&g of ICRl91 per ml, and 54 min for a culturegrown with 20,ug of ICR364-OH per ml. The mutagen-treated cultures were centrifuged, washed, and resus-pended in T2 buffer (17). To enrich for histidine-re-quiring mutants, the treated bacteria were diluted1:10 and 1:100 into 2 ml of E medium supplementedwith histidine-free amino acid pool and 500 units ofpenicillin per ml; they were then incubated for 24 hr at37 C without aeration. Samples were plated on2 EM medium supplemented with histidine-free aminoacid pool, and incubated for 48 hr at 37 C. Underthese conditions, histidine-requiring mutants give riseto small colonies, and other amino acid auxotrophsand wild-type bacteria give rise to large colonies.
C I--,,- '"
1CR 191
HIHsoN > 0H
C l</ O \C3N
ICR 364-OHFIG. 1. Structures of ICR191 and ICR364-OH.
Small colonies were picked and tested for histidinerequirement, and single colonies were isolated. Toinsure the independence of his- mutants, usuallyonly one mutant was retained from each originaltreated culture. However, in eight cases, two mutantsshown by mapping and reversion studies to be ofindependent origin were selected from the same cul-ture. Tests of more than 200 small colonies fromcultures of untreated ara-9 bacteria selected as de-scribed above did not yield his- mutants.
Reversion tests. Mutants were grown in nutrientbroth overnight, and 0.1-ml portions were spread onEM plates in tests similar to ones performed previously(5, 23, 36). Crystals of 2-aminopurine, NG, or strep-tomycin were placed with sterile toothpicks directlyon separate plates; other plates were spotted withsingle drops of either DES or a 1 mg/ml solution ofICR191 or ICR364-OH. The plates were incubated at37 C for 2 to 3 days before scoring.
Detection of nonsense suppressors. His+ revertantswere tested for the presence of amber and ochre sup-pressors on medium lacking citrate and containinglactose in tests against strains SB391 and SB392 (9).An amber suppressor-carrying strain, TA132 (his-C434 supW502), was employed as a control.Enzyme assays. Histidine mutants were grown in E
medium containing 20 ,ug of L-histidine per ml forconditions of repression and on 150 ,ug of L-histidinolper ml for conditions of derepression. Late logarith-mic-phase bacteria (10' bacteria/ml) were harvestedby centrifugation for 10 min at 12,000 X g in a re-frigerated Sorvall centrifuge at 2 C. The pellet wasrinsed, and the cells were washed once with a chilledsolution of 0.85% NaCl. The bacteria were suspendedin 0.01 M tris(hydroxymethyl)aminomethane-hydro-chloride buffer (pH 7.5) and sonically treated, and theextract was centrifuged for 30 min at 39,000 X g. Theprotein concentration of the crude supernatant ex-
VOL. 101, 1970 491
on July 12, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

OESCHGER AND HARTMAN
tract, usually 1 to 3 mg of protein per ml, was meas-ured (20). Enzyme assays were usually performed onfreshly prepared extract.The histidinol phosphate phosphatase assay (1)
was modified by the omission of MgC92, and by theincubation of the molybdate-ascorbate reagent at 60rather than 45 C, for 30 min. One unit of activity isdefined as the amount of enzyme which will free,umole of inorganic phosphate from histidinol phos-phate in 1 hr under the assay conditions. A procedurefor the assay of histidinol dehydrogenase (3) was ap-plied to the crude extracts. Assay of phosphoribosyl-adenosine triphosphate (PR-ATP) pyrophosphorylase(35) was modified to include 45 ,umoles of KC1 per0.3 ml of reaction volume, and the reaction was fol-lowed at 295 nm. One unit of activity is defined as an
increase in optical density at 295 nm of 0.10 in 5 min.
RESULTSReversion of ICR191-induced mutations. Forty-
eight independently derived histidine auxotrophswere isolated from ICR191-treated cultures. Themutants were tested for their ability to revert toprototrophy spontaneously and in response tomutagens (Table 1) and for phenotypic suppres-
sion by streptomycin.The frequency of spontaneous reversion was
low in all the ICR191-induced mutants, andnone of the mutants was "leaky." None wasinduced to revert by 2-aminopurine nor was anyof the mutants phenotypically suppressed bystreptomycin, indicating that none of the ICR191-induced mutations is a transition mutation.
TABLE 1. Reversion pattern and map positions of ICR-induced histidine auxotrophs
Mutants obtained after treatment withReversion pattern
ICRl91 ICR364-OH
Stablea 3000(AII)b, 3015(FV), 3023(AVII-VIIIC), 3050(G through E), 3051(F), 3056(AII),3034(AVI), 3042(FI-II), 3045(DVB-VII) 3058(DIV-VA), 3075(AII through AIII)
Spontane- 3003(AVIB), "T"(F)d, 3008(DVIII), 3009- 3053(C), 3064(CIIB), 3073(DVIII), 3077-ously onlyc (DIIIB), 3020(FIII-IV), 3025(GII), (CIIB)
3027(A)
ICR 191 e 3001 (GIV), 3005(AVII-VIIIC), 3011 (FV- 3049(CIIB), 3054(FIII-IV), 3059(CIIB),VII), 3013(DIV-VA), 3028(DVB-VII), 3062(CIIB), 3063(CIIB), 3066(FVII),3030(AV), 3035(CIIB-III), 3036(DIII- 3067(CIIB), 3069(C), 3070(CIIB), 3071-VII) (CIIB), 3074(CIIB), 3079(CIIB)
ICR191, NG, 3002(AV), 3004(CIII), 3006(AV), 3007- 3052(DIV-VA), 3057(CIIB), 3060(CI), 3061-and DES (CIIB), 3010(A), 3012(DIIA), 3014(C), (AVII-VIIIC), 3065(CIIB), 3068(DIII-
3016(AII), 3017(AVII-VIIIC), 3018- VII), 3072(CIIA), 3076(CIIB), 3078-(DIIA), 3021(C), 3022(AV), 3024(G), (CIIB)3026(FII-III), 3029(DVIII), 3031 (FV),3032(C), 3033(HIIB), 3037(GIV), 3038-(FV-VII), 3039 (AVII-VIIIC), 3040-(DIII-VII), 3041 (FI-II), 3043(A), 3044-(FV-VII), 3046(FV-VII), 3047(FV-VII)
NG, DES, and 3048(GIII-IV)f, 3055(B)f2-amino-purine
a Stable spontaneously and to mutagens.b Map regions, indicated in parentheses after each mutant, where known, are those of Loper et al.
(19) and P. E. Hartman and R. C. Stahl (unpublished data). The exact map position has not been deter-mined for some mutants; therefore, a range of possible positions is given. Thus, the use of more thanone Roman numeral does not imply that the mutation is a multisite mutation spanning the region inquestion.
c Revert spontaneously, but none of the mutagens tested increases reversion frequency. In mostcases, two EM plates were used for each mutagen.
d Strain lost.e Reversion increased with ICR191 but not by NG, DES, or 2-aminopurine. All mutants tested that
reverted in response to ICR191 also reverted with ICR364-OH (e.g., data in Table 7).f Mutations of the transition type. Mutant his-3048 also is phenotypically reverted by streptomycin,
whereas his-3055 is not. Revertants of neither strain contain ochre or amber suppressors.
492 J. BAcTERioL.
on July 12, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
ICR-INDUCED FRAMESHIFTS
Of the 48 mutants, 6 did not revert spontane-ously or respond to any of the mutagens tested.Seven mutants reverted spontaneously, but thisfrequency was not increased by any of themutagens tested, and eight responded only toICR191. Twenty-seven mutants were induced torevert both by ICR191 and by the alkylatingagents DES and NG. The data reported in thefollowing sections establish that this is a class offrameshift mutations.
Reversion of ICR364-OH-induced mutations.Since ICR191 has a 2-chloroethylamino group inits alkyl side chain (Fig. 1), the compound couldhave multiple modes of action due to the acridinecomponent and to the alkylating moiety. It wasconsidered that the 27 mutants that respondedpositively to ICR191, DES, and NG in reversiontests were not frameshift mutants but werecaused by alkylation. Therefore, the mutagenicaction of a related compound, ICR364-OH,with a 2-hydroxyethylamino group instead of a2-chloroethylamino group in the side chain, wasstudied. The results show that the same classesof mutants are elicited by the nonalkylatingazaacridine as are elicited by ICR191.
Thirty-two independently derived histidineauxotrophs induced by ICR364-OH were studiedin reversion tests (Table 1). The frequency ofspontaneous reversion was relatively low in allmutants. Two presumed transition mutations(noted in Table 1) are discussed in later sections.The other 30 mutants fall into the same classes
of frameshift mutations as the ICR191-inducedmutants. Five did not revert spontaneously or inresponse to any of the mutagens tested, fourreverted spontaneously but did not respond tomutagens, and twelve responded to ICR191alone. Nine mutants responded to ICR191 aswell as to DES and NG. The latter mutant classthus appears to be a true class of frameshiftmutations and not due to alkylation.Mapping of mutational sites. Mapping by
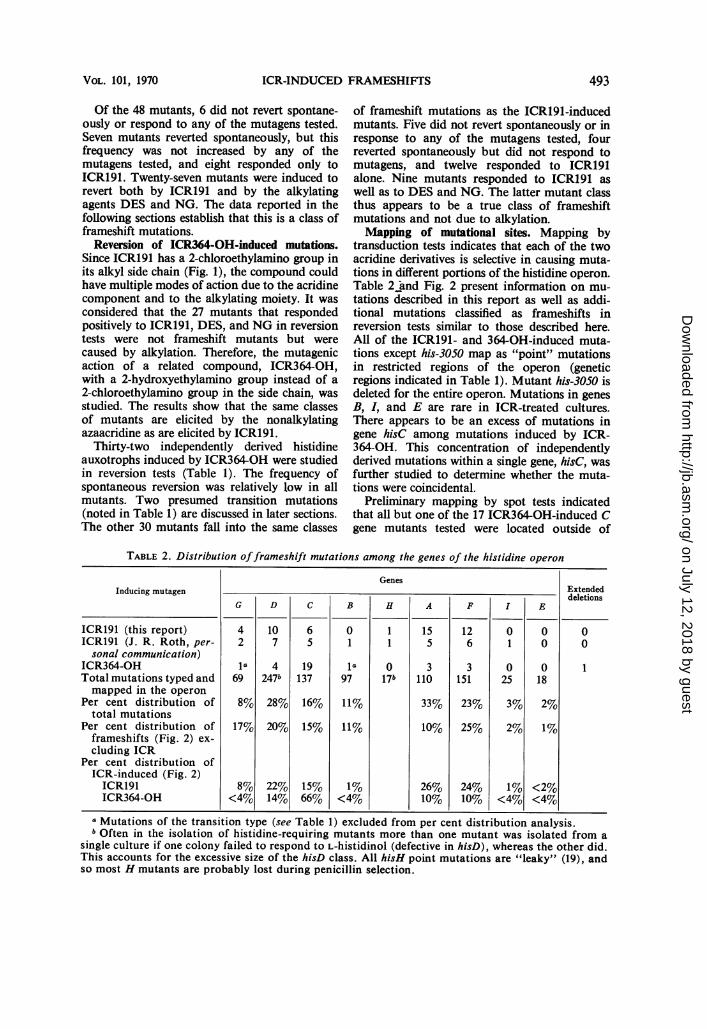
transduction tests indicates that each of the twoacridine derivatives is selective in causing muta-tions in different portions of the histidine operon.Table 2 and Fig. 2 present information on mu-tations described in this report as well as addi-tional mutations classified as frameshifts inreversion tests similar to those described here.All of the ICR191- and 364-OH-induced muta-tions except his-3050 map as "point" mutationsin restricted regions of the operon (geneticregions indicated in Table 1). Mutant his-3050 isdeleted for the entire operon. Mutations in genesB, I, and E are rare in ICR-treated cultures.There appears to be an excess of mutations ingene hisC among mutations induced by ICR-364-OH. This concentration of independentlyderived mutations within a single gene, hisC, wasfurther studied to determine whether the muta-tions were coincidental.
Preliminary mapping by spot tests indicatedthat all but one of the 17 ICR364-OH-induced Cgene mutants tested were located outside of
TABLE 2. Distribution offrameshift mutations among the genes of the histidine operon
GenesInducing mutagen Extended
G D C B H A F I E
ICR191 (this report) 4 10 6 0 1 15 12 0 0 0ICR191 (J. R. Roth, per- 2 7 5 1 1 5 6 1 0 0sonal communication)
ICR364-OH 1 4 19 la 0 3 3 0 0 1Total mutations typed and 69 247b 137 97 17b 110 151 25 18mapped in the operon
Per cent distribution of 8% 28% 16% 11% 33% 23% 3% 2%total mutations
Per cent distribution of 17% 20% 15% 11% 10% 25% 2% 1%frameshifts (Fig. 2) ex-cluding ICR
Per cent distribution ofICR-induced (Fig. 2)ICR191 8% 22% 15% 1% 26% 24% 1% <2%ICR364-OH <4% 14% 66% <4% 10% 10% <4% <4%
a Mutations of the transition type (see Table 1) excluded from per cent distribution analysis.b Often in the isolation of histidine-requiring mutants more than one mutant was isolated from a
single culture if one colony failed to respond to L-histidinol (defective in hisD), whereas the other did.This accounts for the excessive size of the hisD class. All hisH point mutations are "leaky" (19), andso most H mutants are probably lost during penicillin selection.
VOL. 101,) 1970 493
on July 12, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
OESCHGER AND HARTMAN
S, I 1, 1,1, 1,T, ,O/S3
D0oI II I
2 29 3
PP N
p 3 Sx x rx sI u u lxxp s x u s x t
I D S 3 3 S I 3 S X IX P SI 5 5 5 S S U S I
LS'IAIL I --AI WT ATSI I I wL
ISI,1,191,,0,0,A/29p
pp
pp
[tCXX
XS
p
00 .P
'00 p
X 00 pX 00 PI Ioo p
S A 00 XO A 00 X
0II IS
O -I aI
TA iD'sa m T M I IA 1 'KA '19 tC 'Z Ir 'TA'YS '= 'M = ^ a1z 331 C 9 9 2 1 2 2 4 23 3 11 41 I? 2 7 C I? 13 14 1t 2
6
N U
SN A
[SI I I3 II
I. P,P,P,P, X/I
H
p
p
p p
I I pIII
Il,S,SI',IAP.P,P,P,A
1 129
P., /i0
I I
XSI
II rI
x u x
o0 s s X XIX I . I I. I I II I S I X I X IT r ! ! ! 3 *1 1 t-i i i i1**i*?i i i i i
I I N T N M M Z IA IS EA US N N I N I T NAZS UA SCMD55 9 2 10 1 2 2 30 37 3 1 15 2 15 7 5 13 7
1604
zoo0
165*
P,PP,P,P,P,P,P,P,PP,P,P,P,P,P,P,t /5
iI,I,,I,I, I,,III,0,,P,P,P,,PP,P,P,P,P,P
F
I 0
3 3 5S x~~~~~~
N N X XU X S A N S
P X 3 X I s 8.rs s s T ~~s o P*~~
~
I/2
I
A A3
I I
E
I I1 4r S
I1S 11 12 14 S I 14 6 2 4 3 3 3 0 is1
a* Foil to recombinea, Precise mop position not yet determined
Wvvv' a Unmopped
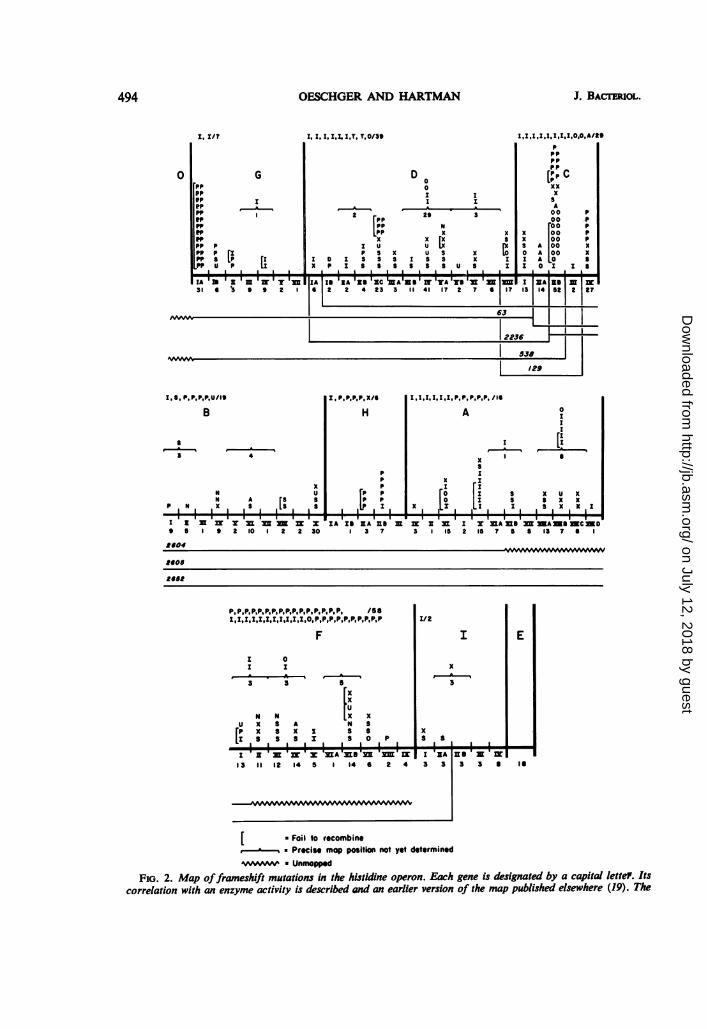
FiG. 2. Map offrameshift mutations in the hstidine operon. Each gene is designated by a capital lette7. Itscorrelation with an enzyme activity is described and an earlier version of the map published elsewhere (19). The
494
i, r/7
0 G
J. BACrERIOL.
ppppppIpp A
ppIeppppppp p
ppuppp [P ppp u P s1
I I I I I,4.
I,S,S, ,P,P,U/I1
B
-
3 4
IIIIIIIIpI
t:I
I 1
2256
558
on July 12, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

ICR-INDUCED FRAMESHIFTS
deletion mutant his-63 and covered by deletionhis-129, i.e., all but one were in regions II and IIIof the C gene (Fig. 2; reference 25). One muta-tion (3060) was located in Cl. Transducing phagegrown on each of the 16 CII-III mutants werespread on EM plates either with deletion mutanthis-538 or with his-2236 as recipient. Fifteen ofthe mutations gave recombination with 2236 butnot with 538, indicating that they were in geneticregion CIIB. Mutant 3072 did not recombinewith either strain, and crosses with extendeddeletion mutants 2652 and 2605 further indicatedthat 3072 was located in CIIA.The following experiment showed that 11 of
the 15 mutations in CIIB were coincidental withone another but separable by recombinationfrom a spontaneous frameshift "hotspot" inregion CIIB (14). Cultures of transducing phageand bacteria were prepared for all of the CII-IIImutants. Reciprocal crosses were performedamong the 15 mutants. As controls, each mutantwas crossed with itself, with phage grown onwild-type bacteria, and with phage grown eitheron hisB47 or on hisE35. The following mutantsgave no detectable recombination (less than0.05% recombination) in reciprocal crosses withone another: 3049, 3057, 3059, 3062, 3067, 3070,3071, 3074, 3076, and 3079. Mutant 3065 alsodid not show recombination when used as donorwith the above mutants as recipients. Of theabove mutants, seven revert spontaneously andwith ICR; two (3057 and 3076) are also inducedto revert with DES and NG (Table 1). Mutants3064, 3077, and 3078 were near but outside thehotspot region (about 1% recombination whencrossed with the other mutants), and 3063appeared to be even further removed (about 3%recombination). Crosses using mutant 3072 ofregion CIIA as recipient and the CIIB mutants asdonors gave 7 to 13% recombination. It is con-cluded that there is a hotspot in the C gene wherevarious kinds of frameshift mutations are prefer-entially induced by treatment with ICR364-OH(Fig. 2).Eight of the hotspot mutants were used as
donors for three spontaneous frameshift mutants
previously described as coincidental (14). Alleight ICR-induced mutations produced a lowfrequency of prototrophic recombinants with2123, 2124, and 2125. Also, one of the aboveeight ICR hotspot mutants and an additionalICR hotspot mutant gave a low frequency ofwild-type recombinants as recipients with phagegrown on two spontaneous hotspot mutants,2124 and 2126. This indicates that the spon-taneous and ICR364-OH hotspots are closelyspaced but not identical. On the other hand, theICR-induced mutation, 3064, failed to recombinewith 2123 and 2124, and gave a very low fre-quency of prototrophic recombinants with 2125.We conclude that 3064 is located in the spon-taneous hotspot and that 2125 is very closelyallied but not identically positioned. Both theICR364-OH and the spontaneous hotspots arelocated in a polynucleotide stretch that is highlysusceptible to frameshift mutations at a numberof positions.
In region IV of the hisC gene, the data ofMartin and Talal (25) show that five frameshiftmutations selected on the basis of their polareffects are localized in a region where one spon-taneous (hisCl22) and one X-ray-induced(hisC516) frameshift mutation lie, whereas muta-tions involving base substitutions map in othersections of the CIV region. None of the ICR-induced frameshifts tested maps in this frameshift-susceptible section.Hotspots of frameshift mutations have been
found in genes other than gene C, and there is apartial overlap between ICR-induced frame-shifts and others (Fig. 2). In gene G, none of theICR-induced frameshifts maps in a hotspot of 26spontaneous frameshifts (14), but 3025 fails torecombine with another spontaneous frameshift,2556, and two ICRI91-induced mutations withdiffering reversion patterns, 3001 and 3007, failto recombine to give wild-type recombinants.Although most of the ICR-induced frameshiftsin genes A and F are scattered, A3017 and A3039fail to recombine with each other, and F3041 failsto recombine with a spontaneous frameshift,F2118. A hotspot in region V of the hisA gene
FIG. 2-Continuedpositions offrameshift mutations are shown above the main horizontal line. Additional partially mappedframeshiftsare placed above brackets; mutations not mapped in detail but placed only as to gene affected are listed at the top.The genes are divided into subregions by deletion mapping, designated by Roman numerals and letters just beneaththe dark horizontal line. The total number of histidine-requiring auxotrophs in each region is indicated by anArabic number either below the gene region or bracket or after the list ofadditional mutants in each gene. Reason-ably up-to-date fine-structure maps and information relating to mutant characterization appear elsewhere(Greeb et al., submittedfor publication), [hisD hisC (25, 36) hisA (23)], and we have incorporated the informationso provided into the present resume. Except for three ICR-induced mutations in region AII, all multisite mutations(extended deletions) have been excludedfrom the listings offrameshifts. Map locations ofa few extended deletionsare shown as thin horizontal lines. Origin: I, ICR191; 0, ICR364-OH: T, ICR372; P. spontaneous, selected forpolarity at 42 C; S, spontaneous; U, ultraviolet; X, X-rays; N, fast neutrons; D, diethylsulfate; A, nitrous acid.
VOL. 101, 1970 495
on July 12, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
OESCHGER AND HARTMAN
included A3002, 3006, 3022, and 3030, whichfail to recombine with one another; again, theseICR-induced mutations constitute a hotspot butstill show differing patterns of reversion. Mu-tants A3000 and A3056 appear to be identicalmultisite mutations, either small deletions orclosely juxtaposed double frameshifts in regionII of the hisA gene; they both are stable and failto recombine with six point mutations, severalof which recombine with one another. Mutant3075 appears to be an even more extended multi-site mutation in this same region. All 14 of theICR-induced hisD mutants are recombinationallyseparable from a spontaneous hotspot of 6spontaneous frameshifts (at least 4 of which areindependent) isolated by Fink et al. (14) andJ. Yourno (personal communication). The 10tested ICR-induced hisD mutants- recombinewith one another.The above examples allow us to conclude that
ICR compounds preferentially induce mutationsin restricted gene regions which exhibit only apartial overlap with regions where spontaneousframeshifts tend to occur.
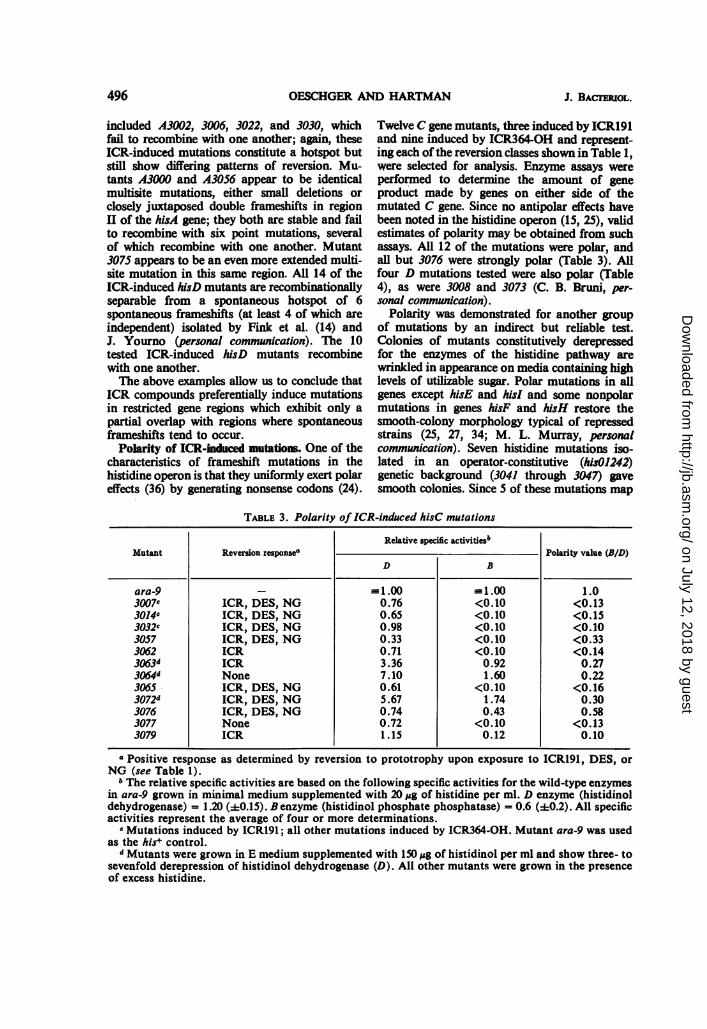
Polarity of ICR-induced mutations One of thecharacteristics of frameshift mutations in thehistidine operon is that they uniformly exert polareffects (36) by generating nonsense codons (24).
Twelve C gene mutants, three induced by ICR191and nine induced by ICR364-OH and represent-ing each of the reversion classes shown in Table 1,
were selected for analysis. Enzyme assays wereperformed to determine the amount of geneproduct made by genes on either side of themutated C gene. Since no antipolar effects havebeen noted in the histidine operon (15, 25), validestimates of polarity may be obtained from suchassays. All 12 of the mutations were polar, andall but 3076 were strongly polar (Table 3). Allfour D mutations tested were also polar (Table4), as were 3008 and 3073 (C. B. Bruni, per-
sonal communication).Polarity was demonstrated for another group
of mutations by an indirect but reliable test.Colonies of mutants constitutively derepressedfor the enzymes of the histidine pathway arewrinkled in appearance on media containing highlevels of utilizable sugar. Polar mutations in allgenes except hisE and hisl and some nonpolarmutations in genes hisF and hisH restore thesmooth-colony morphology typical of repressedstrains (25, 27, 34; M. L. Murray, personalcommunication). Seven histidine mutations iso-lated in an operator-constitutive (hisO1242)genetic background (3041 through 3047) gavesmooth colonies. Since 5 of these mutations map
TABLE 3. Polarity of ICR-induced hisC mutations
Relative specific activitiesbMutant Reversion response" Polarity value (BID)
D B
ara-9 --1.00 - 1.00 1.03007c ICR, DES, NG 0.76 <0.10 <0.133014c ICR, DES, NG 0.65 <0.10 <0.153032" ICR, DES, NG 0.98 <0.10 <0.103057 ICR, DES, NG 0.33 <0.10 <0.333062 ICR 0.71 <0.10 <0.143063d ICR 3.36 0.92 0.273064d None 7.10 1.60 0.223065 ICR, DES, NG 0.61 <0.10 <0.163072d ICR, DES, NG 5.67 1.74 0.303076 ICR, DES, NG 0.74 0.43 0.583077 None 0.72 <0.10 <0.133079 ICR 1.15 0.12 0.10
Positive response as determined by reversion to prototrophy upon exposure to ICR191, DES, orNG (see Table 1).
b The relative specific activities are based on the following specific activities for the wild-type enzymesin ara-9 grown in minimal medium supplemented with 20 pg of histidine per ml. D enzyme (histidinoldehydrogenase) = 1.20 (±0.15). B enzyme (histidinol phosphate phosphatase) = 0.6 (±0.2). All specificactivities represent the average of four or more determinations.
e Mutations induced by ICR191; all other mutations induced by ICR364-OH. Mutant ara-9 was usedas the hit control.
d Mutants were grown in E medium supplemented with 150,ug of histidinol per ml and show three- tosevenfold derepression of histidinol dehydrogenase (D). All other mutants were grown in the presenceof excess histidine.
IGA J. BAamEiuoL.
on July 12, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
ICR-INDUCED FRAMESHIFTS
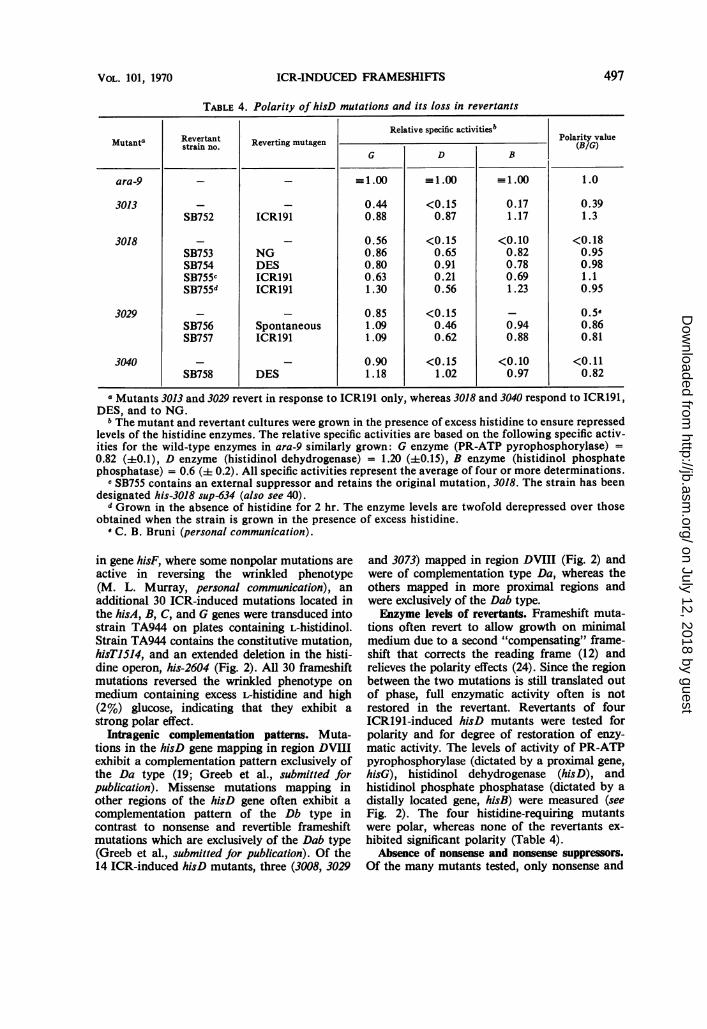
TABLE 4. Polarity of hisD mutations and its loss in revertants
Relative specific activitiesbMutanta Revertant Reverting______mutagen_______ Polarity value
strain no. verting mutagen | G D B|-(BIG)G D B
ara-9 _ _ _1.00 _1.00 -1.00 1.0
3013 _ - 0.44 <0.15 0.17 0.39SB752 ICR191 0.88 0.87 1.17 1.3
3018 _ _ 0.56 <0.15 <0.10 <0.18SB753 NG 0.86 0.65 0.82 0.95SB754 DES 0.80 0.91 0.78 0.98SB755c ICR191 0.63 0.21 0.69 1.1SB755d ICRl91 1.30 0.56 1.23 0.95
3029 _ - 0.85 <0.15 - 0.5sSB756 Spontaneous 1.09 0.46 0.94 0.86SB757 ICR191 1.09 0.62 0.88 0.81
3040 - - 0.90 <0.15 <0.10 <0.11SB758 DES 1.18 1.02 0.97 0.82
aMutants 3013 and 3029 revert in response to ICRl91 only, whereas 3018 and 3040 respond to ICR191,DES, and to NG.
b The mutant and revertant cultures were grown in the presence of excess histidine to ensure repressedlevels of the histidine enzymes. The relative specific activities are based on the following specific activ-ities for the wild-type enzymes in ara-9 similarly grown: G enzyme (PR-ATP pyrophosphorylase) =0.82 (40.1), D enzyme (histidinol dehydrogenase) = 1.20 (a0.15), B enzyme (histidinol phosphatephosphatase) = 0.6 (i 0.2). All specific activities represent the average of four or more determinations.
c SB755 contains an external suppressor and retains the original mutation, 3018. The strain has beendesignated his-3018 sup-634 (also see 40).
d Grown in the absence of histidine for 2 hr. The enzyme levels are twofold derepressed over thoseobtained when the strain is grown in the presence of excess histidine.
e C. B. Bruni (personal communication).
in gene hisF, where some nonpolar mutations areactive in reversing the wrinkled phenotype(M. L. Murray, personal communication), anadditional 30 ICR-induced mutations located inthe hisA, B, C, and G genes were transduced intostrain TA944 on plates containing L-histidinol.Strain TA944 contains the constitutive mutation,hisT514, and an extended deletion in the histi-dine operon, his-2604 (Fig. 2). All 30 frameshiftmutations reversed the wrinkled phenotype onmedium containing excess L-histidine and high(2%) glucose, indicating that they exhibit astrong polar effect.
Intragenic complementation patterns. Muta-tions in the hisD gene mapping in region DVIIIexhibit a complementation pattern exclusively ofthe Da type (19; Greeb et al., submitted forpublication). Missense mutations mapping inother regions of the hisD gene often exhibit acomplementation pattern of the Db type incontrast to nonsense and revertible frameshiftmutations which are exclusively of the Dab type(Greeb et al., submitted jbr publication). Of the14 ICR-induced hisD mutants, three (3008, 3029
and 3073) mapped in region DVIII (Fig. 2) andwere of complementation type Da, whereas theothers mapped in more proximal regions andwere exclusively of the Dab type.Enzyme levels of revertants. Frameshift muta-
tions often revert to allow growth on minimalmedium due to a second "compensating" frame-shift that corrects the reading frame (12) andrelieves the polarity effects (24). Since the regionbetween the two mutations is still translated outof phase, full enzymatic activity often is notrestored in the revertant. Revertants of fourICRl91-induced hisD mutants were tested forpolarity and for degree of restoration of enzy-matic activity. The levels of activity of PR-ATPpyrophosphorylase (dictated by a proximal gene,hisG), histidinol dehydrogenase (hisD), andhistidinol phosphate phosphatase (dictated by adistally located gene, hisB) were measured (seeFig. 2). The four histidine-requiring mutantswere polar, whereas none of the revertants ex-hibited significant polarity (Table 4).
Absence of nonsense and nonsense suppressors.Of the many mutants tested, only nonsense and
497VOL. 101,) 1970
on July 12, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

OESCHGER AND HARTMAN
frameshift mutations are polar in the histidineoperon (14, 15, 24, 36). Even among the polarhistidine prototrophs isolated by Voll (34) andanalyzed more fully, some appear to be muta-tions in a "non-essential" gene, hisH (19), andone appears to be a frameshift very close to thedistal end of the hisD gene (M. M. Rechler andR. G. Martin, personal communication). Becauseof the presence of weak UGA [uridine (U),guanosine (G), adenosine (A)] suppressors inour stocks, few UGA mutants are recoveredunless special screening is made for "leaky"mutants (J. Ferretti and P. E. Hartman, unpub-lished data; J. R. Roth, personal communication),so that it seems unlikely that the ICR-inducedmutations are base substitutions to UGA.Nonsense mutations in the histidine operon
are generally highly revertible by 2-aminopurineand phenotypically suppressed by streptomycin.None of the ICR-induced mutations (with theexception of two mutants, his-3048 and 3055)responds to 2-aminopurine or to streptomycin(Table 1). This makes it unlikely that the ICR-induced mutations, including members of the NGand DES revertible class, are nonsense mutations.Furthermore, the following data indicate thatDES and NG do not allow reversion to the his+phenotype by inducing suppressor mutations thatact on nonsense codons generated by frameshiftmutations.The test of Berkowitz et al. (9) for amber or
ochre suppressors was applied to a total of 112plates of DES-, NG-, ICR191-, and ICR364-OH-induced revertants. Of the several thousandcolonies tested involving 37 different mutants,only two colonies from hisD3018 appeared tocontain nonsense suppressors. One colony camefrom a culture treated with ICRI91 and onefrom a culture treated with NG. Both coloniesexhibited growth only after 5 days of incubation.The hisD3018 mutation was recovered fromthese strains, and new cultures were grown andtreated with NG and with ICRl91. No furthernonsense suppressors were detected, even thoughboth mutagens were active in inducing revertants.Although the significance of the two suppressorsremains obscure, these tests permit us to con-clude that ICR-induced mutations do not showthe reversion properties of amber and ochremutations.
Evidence for intragenic reversion. Direct evi-dence for reversion due to genetic change withinthe same gene locus was obtained by transduc-tional analysis of revertant clones. Phage wasgrown on 24 revertants of hisD mutants (3008,3009, 3012, 3013, 3028, 3029, 3036, 3040, 3052,3068, and 3073). Mutant his-63, deleted for genesG, D, and part of C (Fig. 2), served as recipient
in transduction tests. Plating was performed onminimal E medium containing histidinol, allow-ing the growth of hisD+ clones, whether mutantin other histidine genes or not. Approximately1,000 to 2,000 colonies resulted in each of thetests, corresponding to the value obtained whenphage grown on wild-type bacteria was used. Allclones printed onto minimal medium, indicatingthe absence of defects in histidine genes otherthan hisD. We conclude that the mutationsengendering the 24 hisD+ revertants occurred inthe hisD gene. In contrast, phage prepared on thetwo hisD3018 revertants suspected of carryingsuppressor mutations confirmed the continuedpresence of the 3018 mutation.NG as a mutagen. Of 78 suspected frameshift
mutants, 36 reverted to prototrophy whentreated with NG (Table 1). In contrast, none of101 NG-induced histidine auxotrophs respondedin reversion tests with ICR191 (Hartman et al.,manuscript in preparation). The apparent con-flict in these contrasting observations could beaccounted for if NG predominantly inducedbase substitutions and only comparatively rarelyinduced frameshifts. The following observationsshow that NG is much more effective in causingreversion of mutants originally obtained withNG than it is in causing reversion of mutantscollected after ICR treatment.ICRl91-induced mutants 3006, 3014, 3017,
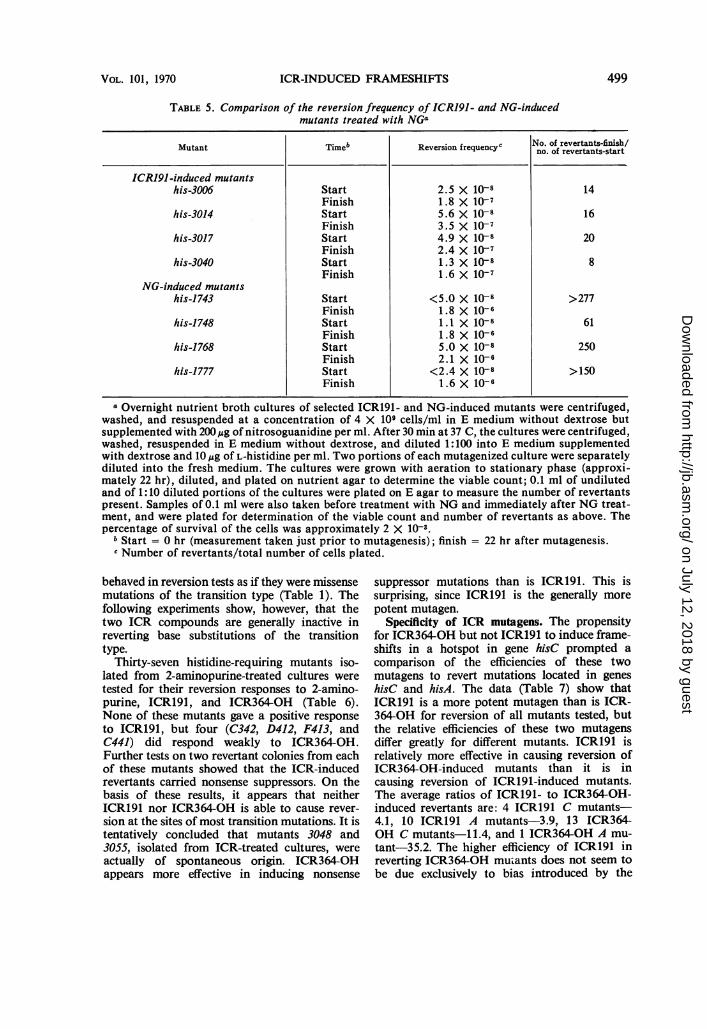
and 3040, and NG-induced mutants 1743, 1748,1768, and 1777 gave strong positive responseswhen a crystal of NG was placed on an EMplate spread with the mutant. The four ICRmutants were selected from among the ICRmutants as giving the strongest responses to NGin such spot tests. Liquid cultures of these eightmutants were tested for their quantitative re-sponses to NG-induced reversion (Table 5).The ICR191-induced mutants showed an
average increase in the number of revertants overthe course of the experiment of about 15-fold. Incontrast, the NG-induced mutants had an averageincrease of greater than 184-fold the number ofrevertants present initially. This observationsupports the hypothesis that NG-inducedreversions of frameshift mutants represent arelatively infrequent mode of action of NG.We have compared hundreds of spot tests for
reversion patterns between ICR induced mu-tants (Table 1) and mutants induced by otheragents. The tests uniformly indicate that rever-sion response to NG is strong for all nonsenseand for most missense mutants but weak ornegative for frameshift mutants.
Failure of ICR to revert 2-aminopurine-inducedmutants. Two mutants that appeared afterICR364-OH treatment, G3048 and B3055,
498 J. BACrERioL.
on July 12, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
ICR-INDUCED FRAMESHIFTS
TABLE 5. Comparison of the reversion frequency of ICRJ91- and NG-inducedmutants treated with NG"
Mutant Timeb Reversion frequencyc No. of revertants-finish/no. of revertants-start
ICRJ91-induced mutanitshis-3006 Start 2.5 X 10-8 14
Finish 1.8 X 10-7his-3014 Start 5.6 X 10-8 16
Finish 3.5 X 10-7his-3017 Start 4.9 X 10-8 20
Finish 2.4 X 10-7his-3040 Start 1.3 X 10-8 8
Finish 1.6 X 10-7NG-induced mutants
his-1743 Start <5.0 X 10-8 >277Finish 1.8 X 10-6
his-1748 Start 1.1 X 10-8 61Finish 1.8 X 10-6
his-1768 Start 5.0 X 10-8 250Finish 2.1 X 10-6
his-1777 Start <2.4 X 10-8 >150Finish 1.6 X 10-6
aOvernight nutrient broth cultures of selected ICR191- and NG-induced mutants were centrifuged,washed, and resuspended at a concentration of 4 X 109 cells/ml in E medium without dextrose butsupplemented with 200,pg of nitrosoguanidine per ml. After 30 min at 37 C, the cultures were centrifuged,washed, resuspended in E medium without dextrose, and diluted 1:100 into E medium supplementedwith dextrose and 10 ug of L-histidine per ml. Two portions of each mutagenized culture were separatelydiluted into the fresh medium. The cultures were grown with aeration to stationary phase (approxi-mately 22 hr), diluted, and plated on nutrient agar to determine the viable count; 0.1 ml of undilutedand of 1:10 diluted portions of the cultures were plated on E agar to measure the number of revertantspresent. Samples of 0.1 ml were also taken before treatment with NG and immediately after NG treat-ment, and were plated for determination of the viable count and number of revertants as above. Thepercentage of survival of the cells was approximately 2 X 10-3.
b Start = 0 hr (measurement taken just prior to mutagenesis); finish = 22 hr after mutagenesis.c Number of revertants/total number of cells plated.
behaved in reversion tests as if they were missensemutations of the transition type (Table 1). Thefollowing experiments show, however, that thetwo ICR compounds are generally inactive inreverting base substitutions of the transitiontype.
Thirty-seven histidine-requiring mutants iso-lated from 2-aminopurine-treated cultures weretested for their reversion responses to 2-amino-purine, ICR191, and ICR364-OH (Table 6).None of these mutants gave a positive responseto ICR191, but four (C342, D412, F413, andC441) did respond weakly to ICR364-OH.Further tests on two revertant colonies from eachof these mutants showed that the ICR-inducedrevertants carried nonsense suppressors. On thebasis of these results, it appears that neitherICRl91 nor ICR364-OH is able to cause rever-sion at the sites of most transition mutations. It istentatively concluded that mutants 3048 and3055, isolated from ICR-treated cultures, wereactually of spontaneous origin. ICR364-OHappears more effective in inducing nonsense
suppressor mutations than is ICRl91. This issurprising, since ICR191 is the generally morepotent mutagen.
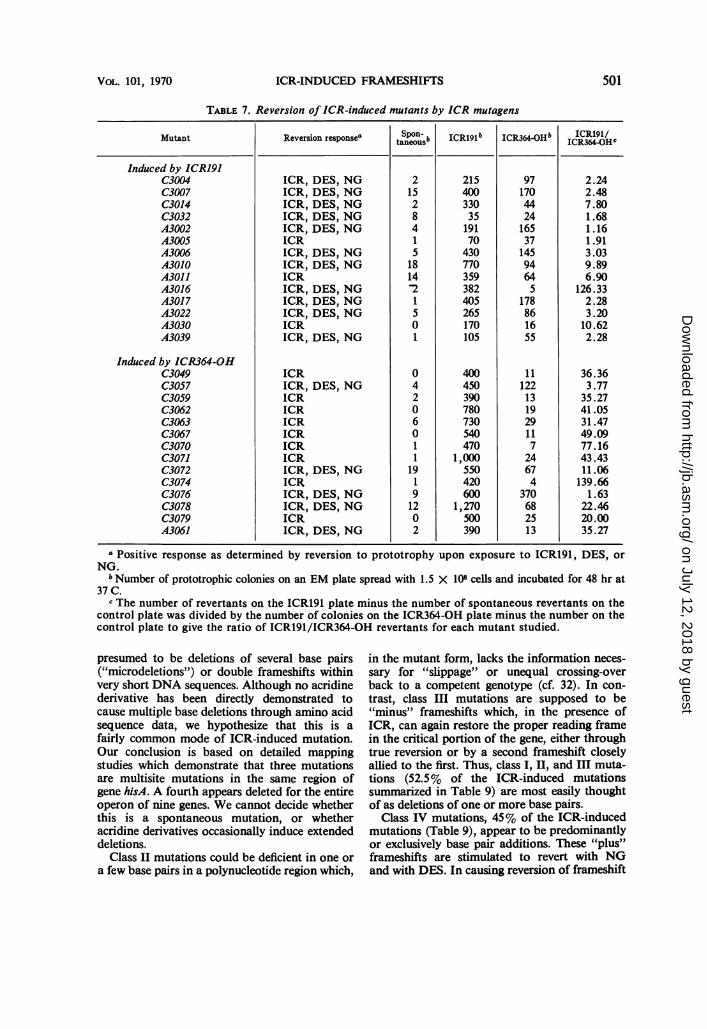
Specificity of ICR mutagens. The propensityfor ICR364-OH but not ICR191 to induce frame-shifts in a hotspot in gene hisC prompted acomparison of the efficiencies of these twomutagens to revert mutations located in geneshisC and hisA. The data (Table 7) show thatICR191 is a more potent mutagen than is ICR-364-OH for reversion of all mutants tested, butthe relative efficiencies of these two mutagensdiffer greatly for different mutants. ICR191 isrelatively more effective in causing reversion ofICR364-OH-induced mutants than it is incausing reversion of ICR191-induced mutants.The average ratios of ICR191- to ICR364-OH-induced revertants are: 4 ICR191 C mutants-4.1, 10 ICRl91 A mutants-3.9, 13 ICR364-OH C mutants-11.4, and 1 ICR364-OH A mu-tant-35.2. The higher efficiency of ICR191 inreverting ICR364-OH mutants does not seem tobe due exclusively to bias introduced by the
499VOL. 101,1 1970
on July 12, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

OESCHGER AND HARTMAN
TABLE 6. Reversion properties of 2-aminopurine-induced mutants"
Induced to revert byMutantb
2-Amino- ICR364-OH ICR191purinec
F336a, B353, C354o, B35S, C357, C358, C367m, C368m, 414, C417, D420, + _G421o, A423m, 1427m, F440mn, D443, A444n, C446a, 454, C455, CSOIa,C502o, CS03, I505a, C863, A864m, F8680
C342o, D412n, F413n, C441n + +d
A338m, 1352m, D362, C364a, A430m, AS12m | _ -
a Approximately 1.5 X 108 bacteria from overnight broth cultures of each mutant were spread onEM plates to which were then added a drop or a few crystals of the mutagen tested.
b Suffix: a-amber, m-missense, n-nonsense (amber or ochre), o-ochre. Other mutations are not yetclassified.
¢ Some mutants that fail to show a positive response to 2-aminopurine in spot tests on plates do sounder altered conditions in tube tests (R. C. Stahl and P. E. Hartman, personal communication).
d Nonsense suppressor mutations.
mutants of the hisC hotspot, since the nonallelic3063, 3072, and 3078 mutations show highvalues, as does A3061.The data (Table 7) also indicate that ICRl91
is relatively more potent than ICR364-OH ininducing reversions among mutants that do notrespond to NG and DES than it is among thosemutants that do. It appears that the two ICRcompounds have somewhat different specificities.Failue of ICR191 to cause CG -+ AT trans-
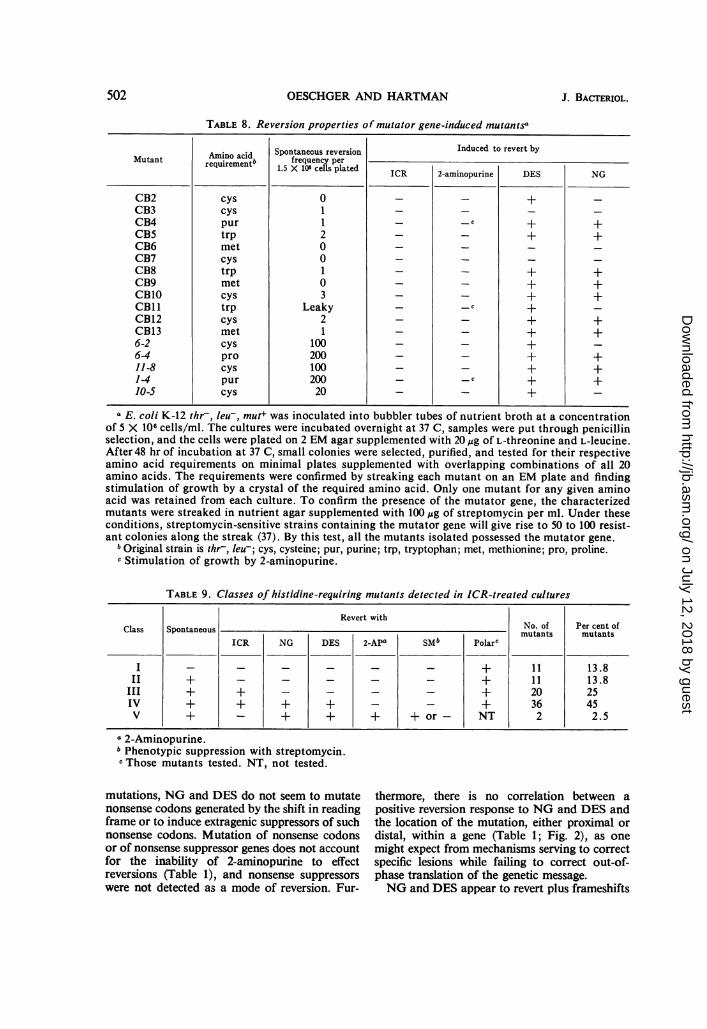
versions. A mutator gene of E. coli K-12 exclu-sively causes adenosine (A)-thymidine (T)cytidine (C)-guanosine (G) transversion muta-tions (37). We show below that ICR191 does notcause reversion of mutator gene-induced mu-tants, and we conclude that ICRl91 is incapableof eliciting CG -. AT transversions.
Seventeen spontaneous mutants isolated bypenicillin selection from 12 cultures of themutator K-12 strain were tested for their responseto various mutagens by spot tests on EM plates(Table 8). The response to DES was in all posi-tive cases stronger than that to NG. In no casewas ICRl91 or 2-aminopurine effective ineliciting reversion. None of the mutants wasphenotypically suppressed by streptomycin (notshown in Table 8).
DISCUSSIONHistidine-requiring mutants isolated from cul-
tures treated with ICRl91 or ICR364-OH are offive classes (Table 9). We consider that classes Ithrough IV are frameshift mutations, i.e., one ormore base pairs have been added or deleted fromthe deoxyribonucleic acid (DNA; 12). The twomissense mutations (class V) could well bespontaneous mutations present in the treated
cultures. However, one cannot exclude the possi-bility that acridine derivatives occasionallyelicit mutations that mimic base substitutions,possibly by causing two closely allied frame-shifts in a favorable nucleotide sequence. Such amechanism is consistent with the nucleotidesequence known to surround an unusual, ICR-revertible missense mutation in the tryptophansynthetase gene (8). We show here that ICR191and ICR364-OH fail to cause intragenic reversionof Salmonella histidine mutants obtained after2-aminopurine treatment (Table 6), and fail torevert auxotrophs presumably obtained asAT -- CG transversions in E. coli K-12 carryinga mutator gene (37; Table 7). ICR191 also failsto revert 101 histidine mutants obtained afterNG treatment, although all but five revert inresponse to NG treatment, and ICR191 fails torevert nine mutants obtained after growth with5-bromodeoxyuridine (Hartman et al., personalcommunication).Over 97% of the ICR-induced mutations are
considered to be true frameshift mutations basedon: (i) inability of 2-aminopurine to elicit rever-sion and inability of streptomycin to elicitphenotypic suppression (Table 6); (ii) ability ofICR compounds to engender reversion of 70%of the mutants (classes m and IV in Table 9);(iii) polarity of the mutations (Tables 3 and 4);(iv) essential lack of external monsense suppres-sors in revertants and direct demonstration ofintragenic reversion; (v) elimination of polarityin revertants; (vi) absence of intragenic comple-mentation; (vii) absence of "leaky" mutantswhich generally are found as about 10% ofpenicillin-screened mutants in our collection.Some of the class I mutations (Table 9) are
500 J. BACmERioL.
on July 12, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
ICR-INDUCED FRAMESHIFTS
TABLE 7. Reversion of ICR-induced mutants by ICR mutagens
Mutant Reversion responsea Spon-b ICR191b ICR3640Hb ICR191/taneous6R4OH ICR364-OH0
Induced by ICRl9JC3004 ICR, DES, NG 2 215 97 2.24C3007 ICR, DES, NG 15 400 170 2.48C3014 ICR, DES, NG 2 330 44 7.80C3032 ICR, DES, NG 8 35 24 1.68A3002 ICR, DES, NG 4 191 165 1.16A3005 ICR 1 70 37 1.91A3006 ICR, DES, NG 5 430 145 3.03A3010 ICR, DES, NG 18 770 94 9.89A3011 ICR 14 359 64 6.90A3016 ICR, DES, NG 2 382 5 126.33A3017 ICR, DES, NG 1 405 178 2.28A3022 ICR, DES, NG 5 265 86 3.20A3030 ICR 0 170 16 10.62A3039 ICR, DES, NG 1 105 55 2.28
Induced by ICR364-OHC3049 ICR 0 400 11 36.36C3057 ICR, DES, NG 4 450 122 3.77C3059 ICR 2 390 13 35.27C3062 ICR 0 780 19 41.05C3063 ICR 6 730 29 31.47C3067 ICR 0 540 11 49.09C3070 ICR 1 470 7 77.16C3071 ICR 1 1,000 24 43.43C3072 ICR, DES, NG 19 550 67 11.06C3074 ICR 1 420 4 139.66C3076 ICR, DES, NG 9 600 370 1.63C3078 ICR, DES, NG 12 1,270 68 22.46C3079 ICR 0 500 25 20.00A3061 ICR, DES, NG 2 390 13 35.27
a Positive response as determined by reversion to prototrophyNG.
b Number of prototrophic colonies on an EM plate spread with 1.537C.
upon exposure to ICR191, DES, or
X 108 cells and incubated for 48 hr at
c The number of revertants on the ICR191 plate minus the number of spontaneous revertants on thecontrol plate was divided by the number of colonies on the ICR364-OH plate minus the number on thecontrol plate to give the ratio of ICR191/ICR364-OH revertants for each mutant studied.
presumed to be deletions of several base pairs("microdeletions") or double frameshifts withinvery short DNA sequences. Although no acridinederivative has been directly demonstrated tocause multiple base deletions through amino acidsequence data, we hypothesize that this is afairly common mode of ICR-induced mutation.Our conclusion is based on detailed mappingstudies which demonstrate that three mutationsare multisite mutations in the same region ofgene hisA. A fourth appears deleted for the entireoperon of nine genes. We cannot decide whetherthis is a spontaneous mutation, or whetheracridine derivatives occasionally induce extendeddeletions.
Class II mutations could be deficient in one ora few base pairs in a polynucleotide region which,
in the mutant form, lacks the information neces-sary for "slippage" or unequal crossing-overback to a competent genotype (cf. 32). In con-trast, class III mutations are supposed to be"minus" frameshifts which, in the presence ofICR, can again restore the proper reading framein the critical portion of the gene, either throughtrue reversion or by a second frameshift closelyallied to the first. Thus, class I, II, and III muta-tions (52.5% of the ICR-induced mutationssummarized in Table 9) are most easily thoughtof as deletions of one or more base pairs.
Class IV mutations, 45% of the ICR-inducedmutations (Table 9), appear to be predominantlyor exclusively base pair additions. These "plus"frameshifts are stimulated to revert with NGand with DES. In causing reversion of frameshift
501VOL. 101,1 1970
on July 12, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
OESCHGER AND HARTMAN
TABLE 8. Reversion properties of mutator genze-induced mutantsa
Spontanecus reversionfrequency per
1.5 X 108 cells plated
011200103
Leaky21
10020010020020
Induced to revert by
ICR 2-aminopurine
_ c
_ c
_ c
DES
+
+++++
NG
+
- E. coli K-12 thr-, leu-, mut+ was inoculated into bubbler tubes of nutrient broth at a concentrationof 5 X 106 cells/ml. The cultures were incubated overnight at 37 C, samples were put through penicillinselection, and the cells were plated on 2 EM agar supplemented with 20,g of L-threonine and L-leucine.After 48 hr of incubation at 37 C, small colonies were selected, purified, and tested for their respectiveamino acid requirements on minimal plates supplemented with overlapping combinations of all 20amino acids. The requirements were confirmed by streaking each mutant on an EM plate and findingstimulation of growth by a crystal of the required amino acid. Only one mutant for any given aminoacid was retained from each culture. To confirm the presence of the mutator gene, the characterizedmutants were streaked in nutrient agar supplemented with 100 jug of streptomycin per ml. Under theseconditions, streptomycin-sensitive strains containing the mutator gene will give rise to 50 to 100 resist-ant colonies along the streak (37). By this test, all the mutants isolated possessed the mutator gene.
I Original strain is thr-, leu-; cys, cysteine; pur, purine; trp, tryptophan; met, methionine; pro, proline.c Stimulation of growth by 2-aminopurine.
TABLE 9. Classes of histidine-requiring mutants detected in ICR-treated cultures
Revert withClass___Spontaneous ._______ No. of Per cent ofClass Spontaneous mutants mutants
ICR NG DES 2-APa SMb Polarc
IV + + + - - - + 11 13.8II + + - - --+ 20 25
IV + + + + --+ 36 45V + + + + or- NT 2 2.5
2-Aminopurine.bPhenotypic suppression with streptomycin.c Those mutants tested. NT, not tested.
mutations, NG and DES do not seem to mutatenonsense codons generated by the shift in readingframe or to induce extragenic suppressors of suchnonsense codons. Mutation of nonsense codonsor of nonsense suppressor genes does not accountfor the inability of 2-aminopurine to effectreversions (Table 1), and nonsense suppressorswere not detected as a mode of reversion. Fur-
thermore, there is no correlation between apositive reversion response to NG and DES andthe location of the mutation, either proximal ordistal, within a gene (Table 1; Fig. 2), as onemight expect from mechanisms serving to correctspecific lesions while failing to correct out-of-phase translation of the genetic message.NG and DES appear to revert plus frameshifts
Mutant
CB2CB3CB4CB5CB6CB7CB8CB9CB10CB11CB12CB136-26411-81-410-5
Amino acidrequirementb
cyscyspurtrpmetcystrpmetcystrpcysmetcysprocyspurcys
502 J. BACTERIOL.
on July 12, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

ICR-INDUCED FRAMESHIFTS
by causing point deletions, possibly as a result ofdepurination (6), as a result of repair of lesionsin DNA (31), or by some other mechanism.Most recently, Yourno and Heath (39) provideddirect evidence that NG treatment of a demon-strated plus frameshift (hisD3018) leads torestoration of the proper reading frame bydeletion of a GC base pair. It is not yet clear ifAT base pairs also can be deleted by NG treat-ment. Thus, although NG predominantly elicitsbase substitutions (Table 5), it and DES areeffective in causing frameshift mutations withreasonable frequency.
Plus frameshifts appear frequently in acridine-treated material (mutants J12, J17, and J44 inreferences 26 and 32; J201 in reference 18;hisD3018 in reference 39) and among revertantsof frameshift mutants where strong selectionmay be present (mutants JDl1 and JD12 inreferences 26 and 32; A23 PRI in reference 8).However, in the only other three cases carefullyanalyzed, one spontaneous mutation (JD10 inreference 32) and two ultraviolet-induced muta-tions (A21 in reference 7; A9813 in reference 10)all appear to be single-base deletions. Studieswith Salmonella histidineless mutants indicatethat there may be only one mutation susceptibleto intragenic reversion both with ICR and withNG from among over 158 spontaneous muta-tions classified as frameshifts (see Fig. 2). Thisclass of mutations also is absent among 9 frame-shift mutations induced with ultraviolet irradia-tion, 8 with nitrous acid, 36 with X-rays, and 5with fast neutrons (Hartman et al., manuscript inpreparation). That is, whereas mutations com-parable to classes I to III (Table 9) are commonin this material, the few mutations initiallyclassified as class IV in reversion tests have beenfound to be nonsense mutations whose ICR-induced revertants carry nonsense suppressors(Table 6 and reference 15). Thus, NG-revertibleframeshifts occur relatively infrequently in theabsence of acridine derivatives. They also areabsent in a series of 23 mutations of the frame-shift type isolated after proflavine treatment ofmating bacteria (28). We postulate that in theabsence of highly potent ICR compounds orstrong selection for their specific detection, plusframeshifts (i.e., NG and DES revertible frame-shifts) are rare as compared with minus frame-shifts.Martin (24) isolated, by recombination, four
spontaneous secondary frameshifts that allowhisC function when combined with frameshifthisC2126. Neither the original frameshift nor thefour secondary frameshifts responds to NG, aresult we have confirmed in the case of three ofthe mutants. One of the secondary frameshifts is
stable to ICR191, whereas the original parentand the other secondary frameshifts respond toICRI91. It seems to us that all of these frame-shifts might be of the minus variety, but thatNG-revertible plus frameshifts might be foundamong revertants of hisC2126 were ICR191-induced revertants examined.We agree that an NG-positive, ICR-negative
reversion pattern is indicative of a mutationinvolving a base substitution, and that a doublynegative or NG-negative, ICR-positive responseis indicative of a frameshift mutation (8, 36).However, in cases where a positive response isobtained with both mutagens, further genetictests are required for mutation classification.ICRl91 and ICR364-OH (Fig. 1) have similar
but not identical activities in mutagenesis. Bothmutagens produce four classes of frameshiftmutations (classes I through IV in Table 9).However, histidine-requiring mutations inducedby ICR191 are largely located in the hisD, A,and F genes, whereas most ICR364-OH-inducedmutations are located in region II of the hisC gene(Table 2; Fig. 2). Eleven of the CII mutationsappear to be allelic in transduction tests, esti-mated to be sensitive enough to detect recom-bination over a distance estimated as less than10 nucleotide pairs (38). Members of this hotspotdiffer from one another in their spontaneousreversion frequency (Table 7) and response tomutagens (Table 1). This clustering phenomenonis common among frameshift mutations in thehistidine operon (Fig. 2; reference 14) andpresumably occurs in regions rich in repeatingnucleotide sequences where mispairing and"slippage" occur (32). Our results show that thetwo acridine and azaacridine derivatives testedboth create new frameshift hotspots and enrichsome but not all spontaneous hotspots. Eachderivative exhibits some unique discrimination,whether measured by induction of auxotrophs(Table 2; Fig. 2), suppressor mutations (Table 6),or in reversion tests of frameshift mutations(Table 7). Frameshifts induced by irradiationappear to be more widely scattered than thoseoccurring spontaneously or after ICR mutagene-sis.
ACKNOWLEDGMENTS
We are indebted to Hugh J. Creech for supplying ICRI91 andICR364-OH, to Charles Yanofsky for supplying an E. coli K-12mutator strain, and to Joseph Yourno and John C. Loper fordiscussions on work with hisD mutants prior to publication.
This investigation was supported by Public Health Serviceresearch grant AI-01650 from the National Institute of Allergyand Infectious Diseases. During 1963 to 1964, N.S.O. was acooperative fellow of the National Science Foundation, and from1964 to 1967, a trainee on Public Health Service grant GM 57.
VOL. 101) 1970 503
on July 12, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

OESCHGER AND HARTMAN
LITERATURE CITED
1. Ames, B. N., B. Garry, and L. A. Hertzenberg. 1960. Thegenetic control of the enzymes of histidine biosynthesis inSalmonella typhimurium. J. Gen. Microbiol. 22:369-378.
2. Ames, B. N., and P. E. Hartman. 1962. Genes, enzymes, andcontrol mechanisms in histidine biosynthesis, p. 322-345.In The molecular basis of neoplasia. 15th Annu. Symp.Fundamental Cancer Res. University of Texas M.D.Anderson Hospital and Tumor Institute, University ofTexas Press, Austin.
3. Ames, B. N., P. E. Hartman, and F. Jacob. 1963. Chromo-somal alterations affecting the regulation of histidinebiosynthetic enzymes in Salmonella. J. Mol. Biol. 7:23-42.
4. Ames, B. N., R. G. Martin, and B. Garry. 1961. The first stepof histidine biosynthesis. J. Biol. Chem. 236:2019-2026.
5. Ames, B. N., and H. J. Whitfield, Jr. 1966. Frameshiftmutagenesis in Salmonella. Cold Spring Harbor Symp.Quant. Biol. 31:211-225.
6. Bautz, E., and E. Freese. 1960. On the mutagenic effect ofalkylating agents. Proc. Nat. Acad. Sci. U.S.A. 46:1585-1594.
7. Berger, H., W. J. Brammar, and C. Yanofsky. 1968a. Analysisof amino acid replacements resulting from frameshift andmissense mutations in the tryptophan synthetase A geneof Escherichla coll. J. Mol. Biol. 34:219-238.
8. Berger, H., W. J. Brammar, and C. Yanofsky. 1968b. Spon-taneous and ICRl91-A-induced frameshift mutations inthe A gene of Escherichia coll tryptophan synthetase. J.Bacteriol. 96:1672-1679.
9. Berkowitz, D., J. M. Hushon, H. J. Whitefield, Jr., J. Roth,and B. N. Ames. 1968. Procedure for identifying nonsense
mutations. J. Bacteriol. 96:215-220.10. Bramznar, W. J., H. Berger, and C. Yanofsky. 1967. Altered
amino acid sequences produced by reversion of frameshiftmutants of tryptophan synthetase A gene of E. coli. Proc.Nat. Acad. Sci. U.S.A. 58:1499-1506.
11. Carlson, E. A., R. Sederoff, and M. Cogan. 1967. Evidencefavoring a frameshift mechanism for ICR-170 inducedmutations in Drosophila melanogaster. Genetics 55:295-313.
12. Crick, F. H. C., L. Barnett, S. Brenner, and R. J. Watts-Tobin.1961. General nature of the genetic code for proteins.Nature 192:1227-1232.
13. deSerres, F. J., and H. E. Brockman. 1968. Homology tests onpresumed multilocus deletions in the ad-3 region ofNeurospora crassa induced by the acridine mustard ICR-170.Genetics 58:79-83.
14. Fink, G. R., T. Klopotowski, and B. N. Ames. 1967. Histidineregulatory mutants in Salmonella typhimurium. IV. Apositive selection for polar histidine-requiring mutants fromhistidine operator constitutive mutants. J. Mol. Biol. 30:81-95.
15. Fink, G. R., and R. G. Martin. 1967. Translation and polarityin the histidine operon. II. Polarity in the histidine operonJ. Mol. Biol. 30:97-107.
16. Hartman, P. E. 1956. Linked loci in the control of consecutivesteps in the primary pathway of histidine synthesis inSalmonella typhimurium, p. 35-62. Genetic studies withbacteria. Carnegie lnst. Wash. Publ. no. 612.
17. Hershey, A. D., and M. Chase. 1952. Independent functions ofviral protein and nucleic acid in growth of bacteriophage.J. Gen. Physiol. 36:39-56.
18. Inouye, M., E. Akaboshi, A. Tsugita, G. Streisinger, and Y.Okada. 1967. A frame-shift mutation resulting in thedeletion of two base pairs in the lysozyme gene of bacteri-ophage T4. J. Mol. Biol. 30:39-47.
19. Loper, J. C., M. Grabnar, R. C. Stahl, Z. Hartman, and P. E.Hartman. 1964. Genes and proteins involved in histidinebiosynthesis in Salmonella. Brookhaven Symp. Biol. 17:15-52.
20. Lowry, 0. H., N. J. Rosenberg, A. L. Farr, and R. J. Randall,1951. Protein measurement with the Folin phenol reagent.
J. Biol. Chem. 193:265-275.21. Malling, H. V. 1967. The mutagenicity of the acridine mustard
(ICR-170) and the structurally related compounds inNeurospora. Mutat. Res. 4:265-274.
22. Margolin, P. 1963. Genetic fine structure of the leucine operonin Salmonella. Genetics 48:441-457.
23. Margolies, M. N., and R. F. Goldberger. 1968. Correlationbetween mutation type and the production of cross-re-
acting material in mutants of the A gene of the histidineoperon in Salmonella typhimurium. J. Bacteriol. 95:507-519.
24. Martin, R. G. 1967. Frameshift mutants in the histidineoperon of Salmonella typhimuriun. J. Mol. Biol. 26:311-328.
25. Martin, R. G., and N. Talal. 1968. Translation and polarityin the histidine operon. IV. Relation of polarity to map
position in hisC. J. Mol. Biol. 36:219-229.26. Okada, Y., G. Streisinger, J. Ernich, J. Newton, A. Tsugita,
and M. Inouye. 1968. Frame shift mutations near thebeginning of the lysozyme gene of bacteriophage T4.Science 162:807-808.
27. Roth, J. R., D. N. Ant6n, and P. E. Hartman. 1966. Histidineregulatory mutants in Salmonella typhimurium. I. Isolationand general properties. J. Mol. Biol. 22:305-323.
28. Sesnowitz-Horn, S., and E. A. Adelberg. 1968. Proflavintreatment of Escherichia coll: generation of frameshiftmutations. Cold Spring Harbor Symp. Quant. Biol. 33:393-402.
29. Smith, H. O., and M. Levine. 1967. A phage P22 gene con-
trolling integration of prophage. Virology 31:207-216.30. Stewart, C. R. 1968. Mutagenesis by acridine yellow in
Bacillus subtilis. Genetics 59:23-31.31. Strauss, B., M. Coyle, and M. Robbins. 1968. Alkylation
damage and its repair. Cold Spring Harbor Symp. Quant.Biol. 33:277-287.
32. Streisinger, G., Y. Okada, J. Emrich, J. Newton, A. Tsugita,E. Terzaghi, and M. Inouye. 1966. Frameshift mutationsand the genetic code. Cold Spring Harbor Symp. Quant.Biol. 31 :77-84.
33. Vogel, H. J., and D. M. Bonner. 1956. Acetylornithinase ofEscherichia coli: partial purification and some properties.J. Biol. Chem. 218:97-106.
34. Voll, M. J. 1967. Translation and polarity in the histidineoperon. III. The isolation of prototrophic polar mutations.J. Mol. Biol. 30:109-124.
35. Voll, M. J., E. Appella, and R. G. Martin. 1967. Purificationand composition studies of phosphoribosyladenosinetriphosphate: pyrophosphate phosphoribosyltransferase,the first enzyme of histidine biosynthesis. J. Biol. Chem.242:1760-1767.
36. Whitfield, H. J., Jr., R. G. Martin, and B. N. Ames. 1966.Classification of aminotransferase (C gene) mutants in thehistidine operon. J. Mol. Biol. 21:335-355.
37. Yanofsky, C., E. C. Cox, and V. Horn. 1966. The unusualmutagenic specificity of an E. coll mutator gene. Proc. Nat.Acad. Sci. U.S.A. 55:274-281.
38. Yanofsky, C., G. R. Drapeau, J. R. Guest, and B. C. Carlton.1967. The complete amino acid sequence of the tryptophansynthetase A protein (a subunit) and its colinear relation-ship with the genetic map of the A gene. Proc. Nat. Acad.Sci. U.S.A. 57:296-298.
39. Yourno, J., and S. Heath. 1969. Nature of the hisD3018frameshift mutation in Salmonella typhimurium. J. Bacteriol.100:460-468.
40. Yourno, J., D. Barr, and S. Tanemura. 1969. Externallysuppressible frameshift mutant of Salmonella typhimurium.J. Bacteriol. 100:453-459.
504 J. BACTERIOL.
on July 12, 2018 by guesthttp://jb.asm
.org/D
ownloaded from