oritavancin kills stationary-phase and biofilm ...aac.asm.org/content/53/3/918.full.pdf ·...
TRANSCRIPT

ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Mar. 2009, p. 918–925 Vol. 53, No. 30066-4804/09/$08.00�0 doi:10.1128/AAC.00766-08Copyright © 2009, American Society for Microbiology. All Rights Reserved.
Oritavancin Kills Stationary-Phase and Biofilm Staphylococcus aureusCells In Vitro�
Adam Belley,1 Eve Neesham-Grenon,1 Geoffrey McKay,1 Francis F. Arhin,1 Robert Harris,2,3
Terry Beveridge,2,4 Thomas R. Parr, Jr.,1 and Gregory Moeck1*Targanta Therapeutics Incorporated, 7170 Frederick Banting, St. Laurent, Quebec H4S 2A1, Canada1; MicroTEM Inc., P.O. Box 1107,
101 Chalmers St., Elora, Ontario N0B 1S0, Canada2; Guelph Regional Integrated Imaging Facility, New Science Complex,488 Gordon St., University of Guelph, Guelph, Ontario N1G 2W1, Canada3; and Department of Molecular andCellular Biology, New Science Complex, 488 Gordon St., University of Guelph, Guelph, Ontario N1G 2W1, Canada4
Received 12 June 2008/Returned for modification 19 October 2008/Accepted 11 December 2008
Slow-growing bacteria and biofilms are notoriously tolerant to antibiotics. Oritavancin is a lipoglyco-peptide with multiple mechanisms of action that contribute to its bactericidal action against exponentiallygrowing gram-positive pathogens, including the inhibition of cell wall synthesis and perturbation ofmembrane barrier function. We sought to determine whether oritavancin could eradicate cells known tobe tolerant to many antimicrobial agents, that is, stationary-phase and biofilm cultures of Staphylococcusaureus in vitro. Oritavancin exhibited concentration-dependent bactericidal activity against stationary-phase inocula of methicillin-susceptible S. aureus (MSSA) ATCC 29213, methicillin-resistant S. aureus(MRSA) ATCC 33591, and vancomycin-resistant S. aureus (VRSA) VRS5 inoculated into nutrient-depletedcation-adjusted Mueller-Hinton broth. As has been described for exponential-phase cells, oritavancininduced membrane depolarization, increased membrane permeability, and caused ultrastructural defectsincluding a loss of nascent septal cross walls in stationary-phase MSSA. Furthermore, oritavancinsterilized biofilms of MSSA, MRSA, and VRSA at minimal biofilm eradication concentrations (MBECs)of between 0.5 and 8 �g/ml. Importantly, MBECs for oritavancin were within 1 doubling dilution of theirrespective planktonic broth MICs, highlighting the potency of oritavancin against biofilms. These resultsdemonstrate a significant activity of oritavancin against S. aureus in phases of growth that exhibittolerance to other antimicrobial agents.
Infections in which bacteria are either slow growing, dor-mant, or in a biofilm pose a serious clinical challenge fortherapy because cells in these states exhibit tolerance to theactivity of antimicrobial agents (15). Osteomyelitis, infectiveendocarditis, chronic wounds, and infections related to in-dwelling devices are examples of infections that harbor toler-ant cells (10, 16). Antimicrobial therapies for these infectionsare not optimal and thus require protracted treatment times. Amodel theory has been proposed to explain biofilm recalci-trance to chemotherapy (27): the diversity of the growth phasesof the biofilm community (44) and the composition of the slimematrix act to limit the effectiveness of otherwise useful antimi-crobial agents. It is believed that a population of slow-growing,stationary-phase, or “persister,” cells within the biofilm cantolerate the killing action of antibacterial agents. This has beendemonstrated with the fluoroquinolone antibiotic ofloxacin,for which a small population of cells within a biofilm was notkilled by this agent (43). Furthermore, it is thought that thesetolerant cells are protected from immune clearance in vivo bythe biofilm slime matrix and ultimately give rise to relapseinfections by reseeding the biofilm once drug levels drop belowtheir antibacterial concentration (27).
Oritavancin is a semisynthetic lipoglycopeptide in clinical
development for the treatment of serious gram-positive infec-tions. It exerts activity against both susceptible and methicillin-resistant Staphylococcus aureus (MRSA) and vancomycin-re-sistant enterococci. The rapidity of its bactericidal activityagainst exponentially growing S. aureus cells (�3-log reductionwithin 15 min to 2 h against methicillin-sensitive S. aureus[MSSA], MRSA, and vancomycin-resistant S. aureus [VRSA])is one feature that distinguishes it from the prototypic glyco-peptide vancomycin (31). Recent work demonstrated that ori-tavancin has multiple mechanisms of action that can contributeto the cell death of exponentially growing S. aureus cells, in-cluding the inhibition of cell wall synthesis by both substrate-dependent and -independent mechanisms (2, 4, 48), disruptionof membrane potential and increasing membrane permeability(32), and inhibition of RNA synthesis (4). The ability of ori-tavancin but not vancomycin to interact with the cell mem-brane, leading to a loss of membrane integrity and collapse oftransmembrane potential, correlates with the rapidity of orita-vancin bactericidal activity (32). Mechanisms of action beyondsubstrate-dependent cell wall synthesis inhibition have notbeen described to date for vancomycin; consequently, vanco-mycin typically requires 24 h and actively dividing cells to exertbactericidal activity (7, 31). With this in mind, we sought tocharacterize oritavancin activity in vitro against S. aureus inslow-growing and biofilm states.
(Part of this work was presented at the 47th InterscienceConference on Antimicrobial Agents and Chemotherapy, Chi-cago, IL, 17 to 20 September 2007 [7].)
* Corresponding author. Mailing address: Targanta TherapeuticsIncorporated, 7170 Frederick Banting, St. Laurent, Quebec H4S 2A1,Canada. Phone: (514) 332-1008, ext. 232. Fax: (514) 332-6033. E-mail:[email protected].
� Published ahead of print on 22 December 2008.
918
on May 30, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

MATERIALS AND METHODS
Bacterial strains. The strains used in this study were MSSA reference strainATCC 29213 (susceptible to daptomycin, linezolid, rifampin, and vancomycin asdetermined by CLSI broth microdilution guidelines) (11), MRSA isolates ATCC33591 and ATCC 43300 (both strains susceptible to daptomycin, linezolid, ri-fampin, and vancomycin), and VRSA isolate VRS5 (Network on AntimicrobialResistance in Staphylococcus aureus) (resistant to vancomycin and susceptible todaptomycin, linezolid, and rifampin). MSSA strain ATCC 29213 and MRSAstrains ATCC 33591 and ATCC 43300 were grown overnight to stationary phasein cation-adjusted Mueller-Hinton broth (CAMHB; Becton, Dickinson, andCompany, Sparks, MD) at 37°C with rotation at 225 rpm. VRSA VRS5 wasgrown overnight to stationary phase at 37°C with rotation at 225 rpm in brainheart infusion broth (Becton, Dickinson, and Company) containing 4 �g/ml ofvancomycin.
Nutrient-depleted CAMHB. Nutrient-depleted CAMHB (29) from each re-spective strain was prepared by three rounds of inoculation of CAMHB withexponential-phase bacteria, incubation overnight at 37°C with rotation (225rpm), and centrifugation (8,000 � g for 30 min) to remove bacteria. After thefinal round of inoculation, growth, and centrifugation, the pH of the nutrient-depleted CAMHB was adjusted to pH 7.0, and the spent medium was filtersterilized using a 0.22-�m membrane (Corning Incorporated, Corning, NY).
Antibacterial agents and concentrations. Antibacterial agents were tested atpharmacologically achievable concentrations that have been determined fromclinical studies: concentrations were chosen to approximate free peak (fCmax)and free trough levels in plasma following administration of standard doses inhumans. For oritavancin, fCmax and free trough levels from a standard dose of200 mg (47) as well as an additional concentration that approximates the fCmax
following a single 800-mg dose (fCmax800) in humans were used (18). Oritavancindiphosphate powder (Targanta Therapeutics, Cambridge, MA) was dissolved inwater containing 0.002% (vol/vol) polysorbate 80; polysorbate 80 was also main-tained at 0.002% in assays to minimize oritavancin loss to the surface of vessels(3), except where indicated. Concentrations that approximate the fCmax and freetrough levels in plasma when administered at standard dosages for the proto-typical glycopeptide vancomycin, the oxazolidinone linezolid, and the lipopeptidedaptomycin were determined from pharmacokinetic data and protein bindingvalues reported in their respective package inserts (vancomycin, Vancocin; lin-ezolid, Zyvox; daptomycin, Cubicin). The approximation of the rifampin fCmax
was derived from data reported previously by Burman et al. (8).Time-kill studies. Nutrient-depleted CAMHB containing diluted antimicro-
bial agents was inoculated with stationary-phase bacteria from cultures of S.aureus strains grown overnight at approximately 107 CFU/ml. Other experimentscompared the killing of stationary- and exponential-phase S. aureus ATCC 29213cells when inoculated into nutrient-depleted CAMHB containing the test agents.For assays involving daptomycin, nutrient-depleted CAMHB was supplementedwith 50 �g/ml CaCl2 (11). All time-kill studies were performed using 96-welldeep-well plates at 37°C with rotation (225 rpm) in a total volume of 750 �l. Toprevent drug carryover during serial dilution plating, aliquots of the drug-chal-lenged culture were added to an equal volume of activated charcoal suspension(25 mg/ml) (6). Bactericidal activities of the antimicrobial agents were defined asa reduction in viable cell counts of �3 log at 24 h relative to cell counts in thestarting inoculum (35). Experiments were repeated at least three times andproduced similar results; results from one experiment are presented.
Short-duration (2-h) time-kill studies were performed using membrane assaybuffer (see below) to characterize the killing of S. aureus ATCC 29213 cells underconditions used in the membrane depolarization and permeability assays. Expo-nential- and stationary-phase S. aureus ATCC 29213 cells were diluted to anoptical density at 600 nm [OD600] of 0.005 (approximately 106 CFU/ml) inmembrane assay buffer (10 mM HEPES-Cl [pH 7.5], 50 �g/ml CaCl2) with andwithout 5 mM glucose, respectively (glucose was included or omitted to preventthe de-energization or energization of exponential- or stationary-phase cellmembranes, respectively). Experiments were initiated by the addition of antimi-crobial agents at the indicated concentrations, and bacteria were enumerated byserial dilution plating. Testing with oritavancin was done in the absence of0.002% polysorbate 80 to reflect conditions used in membrane depolarizationand permeability assays. Experiments were repeated three times and producedsimilar results; results from one assay are presented.
Measurements of membrane depolarization and permeability. Membrane de-polarization was monitored using the fluorescent probe 3,3�-dipropylthiadicar-bocyanine iodide [DiSC3(5)] (Invitrogen Corporation, Carlsbad, CA), whichpartitions into the plasma membrane in proportion to the membrane potential.Dissipation of the membrane potential releases the probe, leading to an increasein fluorescence. Previous studies with the glycopeptide telavancin (19) and the
lipopeptide daptomycin (42) have used this probe to demonstrate the mem-brane-perturbing activity of these drugs against exponential-phase cells. S. aureusATCC 29213 was chosen for testing in membrane studies. Bacteria were grownovernight in CAMHB and subcultured the following day in CAMHB to expo-nential phase (OD600 � 0.25). Exponential- and stationary-phase cells werewashed in membrane assay buffer with and without 5 mM glucose, respectively,and resuspended at an OD600 of 0.25. DiSC3(5) was added to a final concentra-tion of 1.5 �M, and the solution was incubated in the dark at ambient temper-ature for 30 min to allow the loading of the fluorescent dye into cell membranes.After the loading period, cells were diluted 50-fold (OD600 of 0.005) in depo-larization buffer with or without glucose for exponential- or stationary-phasecells, respectively. Assays were initiated by the addition of antimicrobial agentsover a range of concentrations and were monitored in real time by fluorescencespectroscopy (excitation wavelength of 612 nm and emission wavelength of 665nm) for a period of 30 min. Note that 0.002% polysorbate 80 was found tointerfere with fluorescence in these assays and was therefore omitted from theassay. Experiments were repeated three times and produced similar results;results from one assay are presented.
Changes in bacterial membrane permeability were quantified using the fluo-rescent dye pair Syto 9 and propidium iodide: bacterial membrane damage(increased permeability) allows the otherwise membrane-impermeable dye pro-pidium iodide to enter the cell and displace the permeative dye Syto 9, leadingto a loss of fluorescence. Bacteria were prepared as described above for themembrane depolarization assay, but Syto 9 and propidium iodide (InvitrogenCorporation) were added at 5 �M and 30 �M, respectively (32). Fluorescencespectroscopy (excitation wavelength of 485 nm and emission wavelength of 535nm) was monitored for 30 min following the addition of antimicrobial agents. Asmentioned above, 0.002% polysorbate 80 was omitted from the assay to preventinterference with fluorescence determinations. Experiments were repeated threetimes and produced similar results; results from one assay are presented.
Determination of ultrastructural effects of oritavancin and vancomycin onstationary-phase cells by transmission electron microscopy. Stationary-phaseMRSA ATCC 43300 cells (5 � 107 CFU/ml) were exposed to 1 �g/ml oritavancin(2� its broth microdilution MIC in the absence of 0.002% polysorbate 80) or 16�g/ml vancomycin (16� its broth microdilution MIC) in nutrient-depletedCAMHB for 3 h. Bacteria were fixed in 2.5% glutaraldehyde to cross-linkproteins and help preserve morphological structure. Prior to embedding, thesamples were treated with fresh 2.5% (vol/vol) glutaraldehyde in HEPES buffer(pH 6.8) for 2 h. The samples were then postfixed in 2.0% (wt/vol) osmiumtetroxide, followed by en bloc staining with 2.0% (wt/vol) uranyl acetate, as aheavy-metal stain, to add contrast to the cells. The cells were then dehydratedthrough a series of ethanol washes and then embedded in LR White resin. Oncepolymerized by curing, each culture sample was thin sectioned and stained byuranyl acetate and lead citrate to view the internal cellular constituents and thejuxtaposition of cell envelope layers such as the plasma membrane and cell wall.Transmission electron microscopy was used to view the thin sections using aPhilips CM10 microscope under standard operating conditions at 100 kV.
Determination of MBEC. In vitro biofilms were established using the MBECPhysiology & Genetics Assay plate (Innovotech; Edmonton, AB, Canada) ac-cording to the manufacturer’s protocol (20). The MBEC system also allowsthe determination of the MIC of the test agent for planktonic cells shed from thebiofilm as well as the minimum biofilm eradication concentration (MBEC), theconcentration of antimicrobial agent required to sterilize the biofilm after 24 h ofexposure. Briefly, 150 �l of bacterial inocula at 107 CFU/ml in tryptic soy brothwas aliquoted into each well of an MBEC plate. Biofilms were established on theMBEC peg lid for 24 h in a rotary incubator at 37°C and 150 rpm. For experi-ments involving 72-h biofilms, MBEC peg lids were transferred each day into96-well plates containing 150 �l/well of fresh tryptic soy broth and incubatedanother 24 h. MBEC peg lids with established biofilms were washed once insterile saline (200 �l/well) and then placed onto plates containing antimicrobialagents diluted in CAMHB (200 �l/well). Antimicrobial agents were seriallydiluted in CAMHB in 96-well plates, and MBEC peg lids were exposed for 24 hor for the indicated times. Note that 0.002% polysorbate 80 was found toadversely affect biofilm cell numbers and was therefore omitted from MBECdeterminations for oritavancin. Following antimicrobial challenge and determi-nation of the planktonic MICs, MBEC peg lids were washed once in sterile salineand then placed into recovery plates containing CAMHB (200 �l/well). Therecovery plates were sonicated for 5 min in an ultrasonic sonicating bath (VWRAquasonic model 550D) at the maximum setting and then incubated for 24 h,and the MBECs were recorded. MBECs were determined from at least threeindependent experiments; results represent the ranges of MBECs obtained. Toenumerate the biofilm CFU on individual control pegs, pegs were broken off theMBEC peg lid using sterile forceps placed into 1 ml of sterile saline, sonicated
VOL. 53, 2009 ORITAVANCIN KILLS S. AUREUS IN TOLERANT STATES 919
on May 30, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

for 5 min, and vortexed for 1 min at the highest setting. Bacteria were thenenumerated by serial dilution plating. CFU/peg counts were determined from atleast three independent experiments; results presented are the averages � stan-dard deviations.
RESULTS
Oritavancin retains activity against stationary-phase S. au-reus cells. Time-kill studies were performed using nutrient-depleted CAMHB to determine whether growth phase affectsthe antibacterial action of oritavancin and comparator agentsunder conditions of slow growth. In the first experiment, thekilling of stationary-phase MSSA ATCC 29213 (Fig. 1A) wascompared to the killing of an exponential-phase inoculum (Fig.1B). Nutrient-depleted CAMHB limited the growth of thebacterial inocula over a 24-h period (stationary-phase ATCC29213 increased by approximately 0.6 logs, whereas exponen-tial-phase cells increased by approximately 1.3 logs by 24 h)(Fig. 1A and B). Estimated fCmax levels of oritavancin, vanco-mycin, and daptomycin were bactericidal against exponential-phase MSSA ATCC 29213 in nutrient-depleted CAMHB atthe 24-h time point (Fig. 1B). In contrast, only oritavancinexerted bactericidal activity against stationary-phase MSSAstrain ATCC 29213 in nutrient-depleted CAMHB (Fig. 1A),with vancomycin being the most affected by growth phase (ap-
proximately 2.4-log-less killing activity against stationary-phasecells than against exponential-phase cells at the 24-h timepoint). The estimated free trough concentration derived froma 200-mg dose of oritavancin in humans (0.5 �g/ml) nearlyachieved bactericidal levels (approximately 2.9 logs) (Fig. 1A)against stationary-phase MSSA at the 24-h time point, whereasfree trough concentrations of vancomycin and daptomycin hadnegligible effects on cell numbers (data not shown).
In a follow-up experiment, the killing of stationary-phaseinocula of MRSA and VRSA in nutrient-depleted CAMHBwere determined (Fig. 2). As was seen with MSSA, limitedgrowth of MRSA ATCC 33591 (approximately a 0.6-log in-crease) (Fig. 2A) and VRSA VRS5 (approximately a 0.1-logincrease) (Fig. 2B) occurred in nutrient-depleted CAMHBover 24 h. Oritavancin exhibited concentration-dependent bac-tericidal activity against the MRSA and VRSA strains: orita-vancin at its fCmax and fCmax800 was bactericidal at the 24-htime point (Fig. 2), with the exception of the fCmax againstVRS5 (Fig. 2B), which nearly achieved bactericidal killing (ap-proximately a 2.2-log decrease). Vancomycin exhibited somekilling activity against the stationary-phase inocula of MRSA(approximately a 1.4-log reduction) and no killing againstVRSA in nutrient-depleted CAMHB after 24 h (Fig. 2). Dap-tomycin and rifampin exhibited bactericidal activity at their
FIG. 1. Time-kill kinetics of stationary- and exponential-phase MSSA ATCC 29213 in nutrient-depleted CAMHB. Viability was enumeratedat the indicated time points by serial dilution plating. Each point represents the mean of duplicate determinations. The limit of detection isindicated as a dashed line. (A) Stationary-phase inocula challenged with estimated free trough, fCmax, fCmax800 of oritavancin, and fCmax ofcomparators. (B) Exponential-phase inocula challenged with estimated free trough, fCmax, and fCmax800 of oritavancin and comparators. �,untreated control; �, 0.5 �g/ml oritavancin; E, 4 �g/ml oritavancin; F, 16 �g/ml oritavancin; f, 16 �g/ml vancomycin; �, 4 �g/ml daptomycin; Œ,8 �g/ml linezolid; ƒ, 2 �g/ml rifampin.
FIG. 2. Time-kill kinetics of stationary-phase MRSA ATCC 33591 and VRSA VRS5 by oritavancin and comparators in nutrient-depletedCAMHB. Viability was enumerated at the indicated time points by serial dilution plating. Each point represents the mean of duplicatedeterminations. The limit of detection is indicated as a dashed line. (A) MRSA ATCC 33591 challenged with estimated free trough, fCmax andfCmax800 of oritavancin, and fCmax of comparators. (B) VRSA VRS5 challenged with estimated free trough, fCmax and fCmax800 of oritavancin, andfCmax of comparators. �, growth control; �, 0.5 �g/ml oritavancin; E, 4 �g/ml oritavancin; F, 16 �g/ml oritavancin; f, 16 �g/ml vancomycin; �,4 �g/ml daptomycin; Œ, 8 �g/ml linezolid; ƒ, 2 �g/ml rifampin.
920 BELLEY ET AL. ANTIMICROB. AGENTS CHEMOTHER.
on May 30, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
respective fCmax values against the stationary-phase inoculumof MRSA (Fig. 2A) but were not bactericidal against theVRSA strain (Fig. 2B). The bacteriostatic agent linezolid (33)had no effect on the cell numbers of both strains over the 24-hincubation period. The estimated free trough concentration oforitavancin (0.5 �g/ml) reduced cell counts by approximately0.4 logs against the MRSA strain (Fig. 2A) and approximately1.2 logs against the VRSA strain (Fig. 2B), whereas the esti-mated free trough concentrations of vancomycin and dapto-mycin had no effect on the cell counts of these isolates (datanot shown).
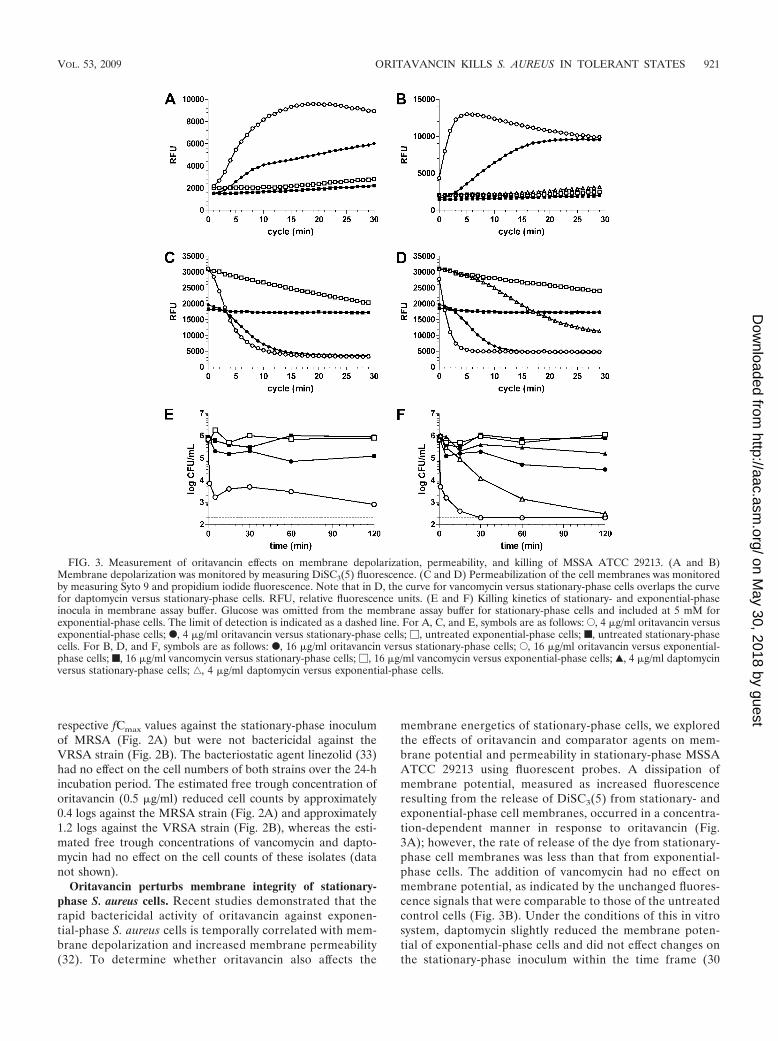
Oritavancin perturbs membrane integrity of stationary-phase S. aureus cells. Recent studies demonstrated that therapid bactericidal activity of oritavancin against exponen-tial-phase S. aureus cells is temporally correlated with mem-brane depolarization and increased membrane permeability(32). To determine whether oritavancin also affects the
membrane energetics of stationary-phase cells, we exploredthe effects of oritavancin and comparator agents on mem-brane potential and permeability in stationary-phase MSSAATCC 29213 using fluorescent probes. A dissipation ofmembrane potential, measured as increased fluorescenceresulting from the release of DiSC3(5) from stationary- andexponential-phase cell membranes, occurred in a concentra-tion-dependent manner in response to oritavancin (Fig.3A); however, the rate of release of the dye from stationary-phase cell membranes was less than that from exponential-phase cells. The addition of vancomycin had no effect onmembrane potential, as indicated by the unchanged fluores-cence signals that were comparable to those of the untreatedcontrol cells (Fig. 3B). Under the conditions of this in vitrosystem, daptomycin slightly reduced the membrane poten-tial of exponential-phase cells and did not effect changes onthe stationary-phase inoculum within the time frame (30
FIG. 3. Measurement of oritavancin effects on membrane depolarization, permeability, and killing of MSSA ATCC 29213. (A and B)Membrane depolarization was monitored by measuring DiSC3(5) fluorescence. (C and D) Permeabilization of the cell membranes was monitoredby measuring Syto 9 and propidium iodide fluorescence. Note that in D, the curve for vancomycin versus stationary-phase cells overlaps the curvefor daptomycin versus stationary-phase cells. RFU, relative fluorescence units. (E and F) Killing kinetics of stationary- and exponential-phaseinocula in membrane assay buffer. Glucose was omitted from the membrane assay buffer for stationary-phase cells and included at 5 mM forexponential-phase cells. The limit of detection is indicated as a dashed line. For A, C, and E, symbols are as follows: E, 4 �g/ml oritavancin versusexponential-phase cells; F, 4 �g/ml oritavancin versus stationary-phase cells; �, untreated exponential-phase cells; f, untreated stationary-phasecells. For B, D, and F, symbols are as follows: F, 16 �g/ml oritavancin versus stationary-phase cells; E, 16 �g/ml oritavancin versus exponential-phase cells; f, 16 �g/ml vancomycin versus stationary-phase cells; �, 16 �g/ml vancomycin versus exponential-phase cells; Œ, 4 �g/ml daptomycinversus stationary-phase cells; ‚, 4 �g/ml daptomycin versus exponential-phase cells.
VOL. 53, 2009 ORITAVANCIN KILLS S. AUREUS IN TOLERANT STATES 921
on May 30, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
min) and concentrations (4 �g/ml) used in the experiment(Fig. 3B).
In the membrane permeability assay, a quantitative differ-ence in initial fluorescence was observed in stationary-phasecells compared to exponential-phase cells (Fig. 3C). This find-ing may reflect that stationary-phase cells either have a loweruptake of Syto 9 or are initially more permeable to propidiumiodide. Oritavancin increased the membrane permeability ofstationary-phase cells in a concentration-dependent manner(data not shown), as evidenced by decreases in Syto 9 fluores-cence (Fig. 3C); however, the rate of loss of Syto 9 fluorescencefrom stationary-phase cells was less than that from exponen-tial-phase cells (Fig. 3C). Vancomycin had no effect on fluo-rescence within the time frame of the assay (30 min) (Fig. 3D),and daptomycin exposure caused a loss of fluorescence fromexponential-phase cells only and not from stationary-phasecells (Fig. 3D).
Time-kill studies using membrane assay buffer over a shortduration of exposure also showed that the rate of killing ofstationary-phase MSSA by oritavancin was decreased com-pared to that of the exponential-phase inoculum (Fig. 3E). Therapid bactericidal activity of oritavancin against exponential-phase cells was exemplified by a 3.2-log reduction in CFUwithin 15 min when tested at 16 �g/ml, its predicted fCmax800.Bacterial killing was also seen with the fCmax of oritavancin (4
�g/ml; 2.9-log reduction) and daptomycin (4 �g/ml; 3.4-logreduction) within 2 h of exposure (Fig. 3E and F). In contrast,stationary-phase cells exhibited approximately 1.5-log and 0.9-log reductions in CFU within the 2-h time period followingexposure to oritavancin at the fCmax800 and fCmax, respectively.Daptomycin activity was similarly reduced, as it exerted a 0.7-log reduction in CFU at its fCmax (Fig. 3F). Vancomycin didnot effect any change on bacterial counts of either inoculumover the short exposure time of the assay (Fig. 3F).
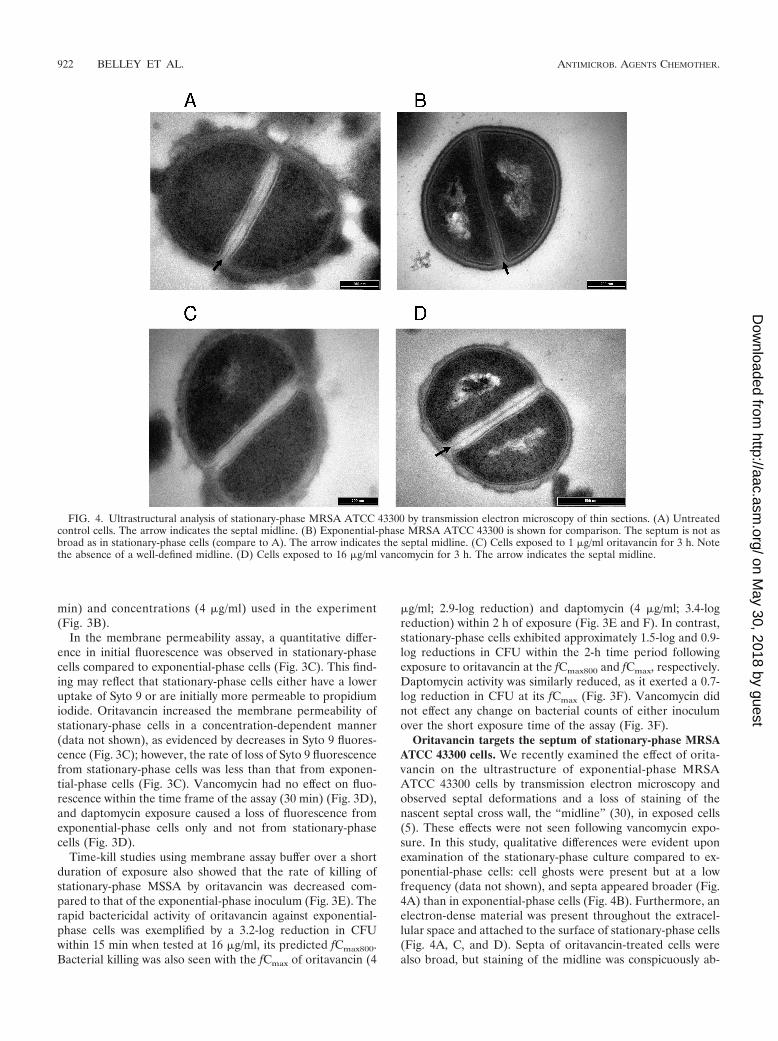
Oritavancin targets the septum of stationary-phase MRSAATCC 43300 cells. We recently examined the effect of orita-vancin on the ultrastructure of exponential-phase MRSAATCC 43300 cells by transmission electron microscopy andobserved septal deformations and a loss of staining of thenascent septal cross wall, the “midline” (30), in exposed cells(5). These effects were not seen following vancomycin expo-sure. In this study, qualitative differences were evident uponexamination of the stationary-phase culture compared to ex-ponential-phase cells: cell ghosts were present but at a lowfrequency (data not shown), and septa appeared broader (Fig.4A) than in exponential-phase cells (Fig. 4B). Furthermore, anelectron-dense material was present throughout the extracel-lular space and attached to the surface of stationary-phase cells(Fig. 4A, C, and D). Septa of oritavancin-treated cells werealso broad, but staining of the midline was conspicuously ab-
FIG. 4. Ultrastructural analysis of stationary-phase MRSA ATCC 43300 by transmission electron microscopy of thin sections. (A) Untreatedcontrol cells. The arrow indicates the septal midline. (B) Exponential-phase MRSA ATCC 43300 is shown for comparison. The septum is not asbroad as in stationary-phase cells (compare to A). The arrow indicates the septal midline. (C) Cells exposed to 1 �g/ml oritavancin for 3 h. Notethe absence of a well-defined midline. (D) Cells exposed to 16 �g/ml vancomycin for 3 h. The arrow indicates the septal midline.
922 BELLEY ET AL. ANTIMICROB. AGENTS CHEMOTHER.
on May 30, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

sent (Fig. 4C), corroborating observations of exponential-phase cells (5). The midline was evident in vancomycin-treatedcells (Fig. 4D), which overall had an ultrastructural appearancesimilar to that of the untreated cells.
Oritavancin is active against in vitro S. aureus biofilms. Thecommercially available MBEC system (20) was used to estab-lish in vitro biofilms of S. aureus and to determine MBECs oforitavancin and comparator antimicrobial agents. To preventthe loss of oritavancin due to binding to vessel surfaces (3), theClinical and Laboratory Standards Institute (CLSI; formerlyNCCLS) recommends the inclusion of 0.002% polysorbate 80for oritavancin broth microdilution MIC determinations (12).Initial experiments determined that the inclusion of polysor-bate 80 during S. aureus biofilm establishment or antimicrobialchallenge caused a significant reduction in CFU/peg densities(data not shown). Therefore, polysorbate 80 was omitted fromthe planktonic MIC and MBEC determinations in the MBECsystem. This omission is predicted to result in an underestima-tion of oritavancin potency.
The capacity of each strain to form a biofilm on the pegs ofthe MBEC plate was determined by enumerating the CFUattached to the peg surface (CFU/peg). Biofilm cell densitieson the pegs for each strain were (2.9 � 2.4) � 105 CFU/peg forMSSA ATCC 29213, (2.2 � 1.7) � 105 CFU/peg for MRSAATCC 33591, and (2.6 � 1.1) � 105 CFU/peg for VRSA VRS5after 24 h of incubation. Planktonic MICs determined for com-parator antimicrobial agents in the MBEC assay were withinthe CLSI quality control ranges (Table 1). Although the testingmethodologies were not identical to the conventional brothmicrodilution MIC method based on CLSI guidelines (10),oritavancin planktonic MICs were also within the quality con-trol range (0.5 to 2 �g/ml) for MSSA ATCC 29213 as deter-mined in the absence of 0.002% polysorbate 80 (13). Thegrowth of S. aureus in a biofilm resulted in dramatic decreasesin the antimicrobial activities of vancomycin and linezolid asmeasured by the concentration of antimicrobial agent neededto sterilize the 24-hour biofilm (MBEC) compared to theirrespective planktonic MICs (Table 1): MBECs for both agentswere �128 �g/ml against all three strains. In contrast, orita-vancin MBECs ranged from 2 to 8 �g/ml against the S. aureusstrains (Table 1) and were within 1 doubling dilution of theirrespective planktonic MICs in each experiment.
The time required for oritavancin to sterilize 24-h biofilms ofMSSA ATCC 29213 was determined by measuring the MBECs
after shorter exposure times. Oritavancin sterilized the biofilmafter a 1-h exposure at an MBEC of 4 �g/ml. As expected,MBECs for the comparator agents were �128 �g/ml at thisexposure time. To further test the ability of oritavancin toeradicate biofilm-associated S. aureus in vitro, biofilms ofMSSA ATCC 29213 were grown for 72 h to increase thecellular density of the peg biofilms. Indeed, CFU/peg increasedto (4.6 � 1.3) � 106 CFU/peg (approximately a 1.2-log in-crease compared to the 24-h biofilm cellular density), andoritavancin planktonic MIC and MBEC values were concom-itantly affected, ranging from 4 to 32 �g/ml and 8 to 32 �g/ml,respectively. Importantly, within each experiment, oritavancinMBECs were no more than 1 doubling dilution higher thantheir respective planktonic MICs. Planktonic MICs for vanco-mycin and linezolid were also affected by the increased cellularpeg density and were 2 to 16 �g/ml and 8 to 16 �g/ml, respec-tively. MBECs for both agents were �128 �g/ml.
DISCUSSION
Oritavancin has been defined as a lipoglycopeptide, distin-guishing it structurally from vancomycin (12). Its 4�-chlorobi-phenylmethyl group confers the ability to disrupt membranepotential and to increase the permeability of exponential-phase gram-positive pathogens (32). Because membrane bar-rier function is essential to the viability of all cells, we hypoth-esized that oritavancin would be active against slowly dividingcells and biofilms. The finding that oritavancin exerted bacte-ricidal activity in vitro against stationary-phase cells, as shownin the present study, suggests that its additional mechanism ofaction relative to vancomycin, namely, its effect on membraneintegrity, is its principal antibacterial mechanism against bac-teria in these states. The membrane-perturbing antimicrobialagent daptomycin (1, 42) also exhibited antibacterial activityagainst stationary-phase cells in our study, confirming findingsfrom a previous report (29). In contrast, the reduced activity ofvancomycin against stationary-phase S. aureus, reported hereand in other studies (34, 45), suggests the dispensability of cellwall synthesis for viability in this test system and is likely afunction of its single mode of action: vancomycin inhibits cellwall synthesis, which occurs in S. aureus only during septumformation in dividing cells (41). Thus, infections in which cellgrowth is limited by nutrient limitation, quorum sensing, oranaerobiosis may render cells tolerant to the killing action ofbactericidal antimicrobial agents, but these cells remain vul-nerable to membrane-perturbing agents such as oritavancinand daptomycin.
A reduced rate of killing of stationary-phase cells by orita-vancin was observed in either nutrient-depleted CAMHB ormembrane assay buffer and was concomitant with a reducedrate of depolarization and permeabilization of cell membranes.Although we cannot exclude the possibility that oritavancintargets are less abundant in stationary-phase cells, these find-ings are likely an indication that the bactericidal mechanism ofaction of oritavancin is linked to the energized state of the cell:membrane potential is known to be greater in exponential-phase S. aureus cells (�167 mV) than in stationary-phase cells(�123 mV) (23). Membrane potential and the protonatedstate of the cell wall are thought to regulate cell wall hydrolase(autolysin) activity (24), and oritavancin-induced changes in
TABLE 1. Oritavancin exhibits antibiofilm activity in vitro againstS. aureus strains of different resistance phenotypesa
Antimicrobialagent
MSSA ATCC29213
MRSA ATCC33591 VRSA VRS5
MIC(�g/ml)
MBEC(�g/ml)
MIC(�g/ml)
MBEC(�g/ml)
MIC(�g/ml)
MBEC(�g/ml)
Oritavancinb 2 2–4 0.5–4 0.5–4 2–8 2–8Linezolid 8 �128 2–4 �128 4–8 �128Rifampin 0.02 4 0.03 0.25–4 0.03–0.06 4Vancomycin 1 �128 1–2 �128 �128 �128
a MICs (�g/ml) were determined using MBEC plates and represent the anti-bacterial activity against planktonic cells shed from the biofilms. MBECs (�g/ml)were determined according to the manufacturer’s protocol.
b Oritavancin MICs and MBECs were determined in the absence of 0.002%polysorbate 80.
VOL. 53, 2009 ORITAVANCIN KILLS S. AUREUS IN TOLERANT STATES 923
on May 30, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

the membrane potential of stationary-phase cells may accountfor the observed loss of the septal midline: the decreasedstaining intensity of the septal midline in stationary-phase S.aureus cells may have reasonably resulted from a loss of thechemically reactive sites that are exposed (and that bind theheavy metal stain) when cell wall hydrolases cleave septal poly-mers of the nascent cross walls during division (30, 46). Such aloss of the septal midline in stationary-phase cells corroboratespreviously reported observations of exponential-phase cells(5). Another possibility is that the inhibition of nascent cross-wall synthesis could also account for the loss of the midline:oritavancin inhibits cell wall synthesis (4) and can directlyinhibit the transglycosylase activity of S. aureus penicillin-bind-ing protein 2 (48), an essential transglycosylase-transpeptidasethat is localized to the division septum during cell division (36).Thus, while a full understanding of the means by which orita-vancin causes cell death requires further investigation, we showin the current work that it likely involves a common mechanismagainst stationary- and exponential-phase cells.
The potency of oritavancin against S. aureus in vitro biofilmsis highlighted by (i) MBEC values within 1 doubling dilution oftheir planktonic MICs, (ii) sterilization of MSSA biofilmswithin 1 h, and (iii) sterilization of 72-h biofilms of MSSA thatalso had increased cellular densities at MBECs within 1 dou-bling dilution of its planktonic MIC against the test strain. Aswas seen against stationary-phase cells, vancomycin and lin-ezolid did not sterilize the biofilms, confirming previously re-ported findings of greatly reduced activities of these agents inin vitro biofilm models (9, 17, 21, 39). Thus, in the in vitrobiofilms described here, a tolerant population of cells waspresent. The finding that the stratification of DNA synthesisactivity in an in vitro colony biofilm model in which the vastmajority of cells synthesizing DNA (and, therefore, activelydividing) was at the air interface (37) indicates that biofilms arecomposed of a mixed population of cells with different levels ofmetabolic activity. Furthermore, another study reported thatonly a subpopulation of cells in a biofilm were readily killed bythe fluoroquinolone ofloxacin (43). Regardless of whether thein vitro (24- or 72-h) biofilms of S. aureus described here werecomposed of cells of different metabolic activities, oritavancinsterilized the biofilms at a concentration that was 1 doublingdilution higher than that needed to kill planktonic cells. Al-though rifampin exhibited reduced activity against the in vitroS. aureus biofilms, its MBEC (4 �g/ml) approached pharma-cologically achievable concentrations (fCmax � 2 �g/ml) (8).However, development of rifampin resistance has been ob-served when this agent is used alone in in vitro biofilm models(32) and in vivo (22, 28), which therefore should restrict its useto combination therapy.
The development of antimicrobial agents and therapies thatare active against biofilms and tolerant cells would benefit thetreatment of infections that harbor cells in these states (14).Unfortunately, these types of infections are often complicatedby ischemia (i.e., sequestra of osteomyelitis and chronic woundof diabetic foot ulcers), compromising antimicrobial exposureand activity at the infection site. Thus, new agents and thera-pies must overcome significant hurdles. The present studyshows that oritavancin exerts bactericidal activity against sta-tionary-phase S. aureus cells, likely by its capacity to disrupt themembrane integrity of susceptible bacteria. Furthermore, its
potency against the in vitro biofilm was remarkable in light ofthe observed significant tolerance of the biofilm to comparatorantimicrobial agents. To date, oritavancin has shown efficacy ininfections that likely harbor tolerant cells, including rat centralvenous catheter (38), rat granuloma pouch (26), and rabbitendocarditis (25, 40) models. Studies to examine the efficacy oforitavancin in in vivo models of biofilm infections are war-ranted to confirm its promising activity against cells in a toler-ant state in vitro.
ACKNOWLEDGMENTS
We thank the scientific support personnel at Targanta Therapeuticsfor their contributions to this research. We also thank Norris Allen forhelpful discussions and critical review of the manuscript.
With the exception of R.H and T.B., all authors of the manuscriptare Targanta Therapeutics employees.
REFERENCES
1. Alborn, W. E., Jr., N. E. Allen, and D. A. Preston. 1991. Daptomycin disruptsmembrane potential in growing Staphylococcus aureus. Antimicrob. AgentsChemother. 35:2282–2287.
2. Allen, N. E., and T. I. Nicas. 2003. Mechanism of action of oritavancin andrelated glycopeptide antibiotics. FEMS Microbiol. Rev. 26:511–532.
3. Arhin, F. F., I. Sarmiento, A. Belley, G. A. McKay, D. C. Draghi, P. Grover,D. F. Sahm, T. R. Parr, Jr., and G. Moeck. 2008. Effect of polysorbate 80 onoritavancin binding to plastic surfaces: implications for susceptibility testing.Antimicrob. Agents Chemother. 52:1597–1603.
4. Arhin, F. F., I. Sarmiento, T. R. Parr, Jr., and G. Moeck. 2007. Mechanismsof action of oritavancin in Staphylococcus aureus, abstr. C1-1471. Abstr. 47thIntersci. Conf. Antimicrob. Agents Chemother., Chicago, IL, 17 to 20 Sep-tember 2007.
5. Belley, A., R. Harris, T. Beveridge, T. Parr, Jr., and G. Moeck. 2009. Ultra-structural effects of oritavancin on methicillin-resistant Staphylococcus au-reus and vancomycin-resistant Enterococcus. Antimicrob. Agents Che-mother. 53:800–804.
6. Belley, A., E. Neesham-Grenon, F. F. Arhin, G. A. McKay, T. R. Parr, Jr.,and G. Moeck. 2008. Assessment by time-kill methodology of the synergisticeffects of oritavancin in combination with other antimicrobial agents againstStaphylococcus aureus. Antimicrob. Agents Chemother. 52:3820–3822.
7. Belley, A., E. Neesham-Grenon, T. R. Parr, Jr., and G. Moeck. 2007. Phar-macokinetic concentrations of oritavancin kill stationary-phase and biofilmStaphylococcus aureus in vitro, abstr. E-1620. Abstr. 47th Intersci. Conf.Antimicrob. Agents Chemother., Chicago, IL, 17 to 20 September 2007.
8. Burman, W. J., K. Gallicano, and C. Peloquin. 2001. Comparative pharma-cokinetics and pharmacodynamics of the rifamycin antibacterials. Clin. Phar-macokinet. 40:327–341.
9. Ceri, H., M. E. Olson, C. Stremick, R. R. Read, D. Morck, and A. Buret.1999. The Calgary Biofilm Device: new technology for rapid determinationof antibiotic susceptibilities of bacterial biofilms. J. Clin. Microbiol. 37:1771–1776.
10. Ciampolini, J., and K. G. Harding. 2000. Pathophysiology of chronic bacte-rial osteomyelitis. Why do antibiotics fail so often? Postgrad. Med. J. 76:479–483.
11. CLSI. 2006. Methods for dilution antimicrobial susceptibility tests for bac-teria that grow aerobically. Approved standard, CLSI document M7-A7, 7thed. Clinical and Laboratory Standards Institute, Wayne, PA.
12. CLSI. 2008. Performance standards for antimicrobial susceptibility testing;eighteenth informational supplement, CLSI document M100-S18, 7th ed.Clinical and Laboratory Standards Institute, Wayne, PA.
13. CLSI. 2006. Performance standards for antimicrobial susceptibility testing;sixteenth informational supplement, CLSI document M100-S16, 7th ed.Clinical and Laboratory Standards Institute, Wayne, PA.
14. Coates, A., Y. Hu, R. Bax, and C. Page. 2002. The future challenges facingthe development of new antimicrobial drugs. Nat. Rev. Drug Discov. 1:895–910.
15. Costerton, J. W., P. S. Stewart, and E. P. Greenberg. 1999. Bacterial biofilms:a common cause of persistent infections. Science 284:1318–1322.
16. Donlan, R. M., and J. W. Costerton. 2002. Biofilms: survival mechanisms ofclinically relevant microorganisms. Clin. Microbiol. Rev. 15:167–193.
17. El-Azizi, M., S. Rao, T. Kanchanapoom, and N. Khardori. 2005. In vitroactivity of vancomycin, quinupristin/dalfopristin, and linezolid againstintact and disrupted biofilms of staphylococci. Ann. Clin. Microbiol.Antimicrob. 4:2.
18. Fetterly, G. J., C. M. Ong, S. M. Bhavnani, J. S. Loutit, S. B. Porter, L. G.Morello, P. G. Ambrose, and D. P. Nicolau. 2005. Pharmacokinetics oforitavancin in plasma and skin blister fluid following administration of a
924 BELLEY ET AL. ANTIMICROB. AGENTS CHEMOTHER.
on May 30, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

200-milligram dose for 3 days or a single 800-milligram dose. Antimicrob.Agents Chemother. 49:148–152.
19. Higgins, D. L., R. Chang, D. V. Debabov, J. Leung, T. Wu, K. M. Krause, E.Sandvik, J. M. Hubbard, K. Kaniga, D. E. Schmidt, Jr., Q. Gao, R. T. Cass,D. E. Karr, B. M. Benton, and P. P. Humphrey. 2005. Telavancin, a multi-functional lipoglycopeptide, disrupts both cell wall synthesis and cell mem-brane integrity in methicillin-resistant Staphylococcus aureus. Antimicrob.Agents Chemother. 49:1127–1134.
20. Innovotech. 2008. The MBEC Physiology & Genetics assay: instruction man-ual. Innovotech Inc., Edmonton, Canada.
21. Jones, S. M., M. Morgan, T. J. Humphrey, and H. Lappin-Scott. 2001. Effectof vancomycin and rifampicin on meticillin-resistant Staphylococcus aureusbiofilms. Lancet 357:40–41.
22. Kadurugamuwa, J. L., L. V. Sin, J. Yu, K. P. Francis, R. Kimura, T. Purchio,and P. R. Contag. 2003. Rapid direct method for monitoring antibiotics in amouse model of bacterial biofilm infection. Antimicrob. Agents Chemother.47:3130–3137.
23. Kashket, E. R. 1981. Proton motive force in growing Streptococcus lactis andStaphylococcus aureus cells under aerobic and anaerobic conditions. J. Bac-teriol. 146:369–376.
24. Kemper, M. A., M. M. Urrutia, T. J. Beveridge, A. L. Koch, and R. J. Doyle.1993. Proton motive force may regulate cell wall-associated enzymes ofBacillus subtilis. J. Bacteriol. 175:5690–5696.
25. Lefort, A., A. Saleh-Mghir, L. Garry, C. Carbon, and B. Fantin. 2000.Activity of LY333328 combined with gentamicin in vitro and in rabbit ex-perimental endocarditis due to vancomycin-susceptible or -resistant Entero-coccus faecalis. Antimicrob. Agents Chemother. 44:3017–3021.
26. Lehoux, D., F. F. Arhin, I. Fadhil, K. Laquerre, V. Ostiguy, I. Sarmiento, G.Moeck, and T. R. Parr, Jr. 2006. Oritavancin demonstrates rapid and sus-tained bactericidal activity in the rat granuloma pouch model of Staphylo-coccus aureus infection, abstr. B-0404. Abstr. 46th Intersci. Conf. Antimi-crob. Agents Chemother., San Francisco, CA, 27 to 30 September 2006.
27. Lewis, K. 2007. Persister cells, dormancy and infectious disease. Nat. Rev.Microbiol. 5:48–56.
28. Lucet, J. C., M. Herrmann, P. Rohner, R. Auckenthaler, F. A. Waldvogel,and D. P. Lew. 1990. Treatment of experimental foreign body infectioncaused by methicillin-resistant Staphylococcus aureus. Antimicrob. AgentsChemother. 34:2312–2317.
29. Mascio, C. T., J. D. Alder, and J. A. Silverman. 2007. Bactericidal action ofdaptomycin against stationary-phase and nondividing Staphylococcus aureuscells. Antimicrob. Agents Chemother. 51:4255–4260.
30. Matias, V. R., and T. J. Beveridge. 2007. Cryo-electron microscopy of celldivision in Staphylococcus aureus reveals a mid-zone between nascent crosswalls. Mol. Microbiol. 64:195–206.
31. McKay, G. A., S. Beaulieu, A. Belley, F. F. Arhin, T. R. Parr, Jr., and G.Moeck. 2007. In vitro time-kill studies of oritavancin against drug-resistantisolates of Staphylococcus aureus and enterococci, abstr. E-1614. Abstr. 47thIntersci. Conf. Antimicrob. Agents Chemother., Chicago, IL, 17 to 20 Sep-tember 2007.
32. McKay, G. A., I. Fadhil, S. Beaulieu, S. Ciblat, A. R. Far, G. Moeck, andT. R. Parr, Jr. 2006. Oritavancin disrupts transmembrane potential andmembrane integrity concomitantly with cell killing in Staphylococcus aureusand vancomycin-resistant enterococci, abstr. C1-682. Abstr. 46th Intersci.
Conf. Antimicrob. Agents Chemother., San Francisco, CA, 27 to 30 Sep-tember 2006.
33. Moellering, R. C. 2003. Linezolid: the first oxazolidinone antimicrobial. Ann.Intern. Med. 138:135–142.
34. Murillo, O., A. Domenech, A. Garcia, F. Tubau, C. Cabellos, F. Gudiol, andJ. Ariza. 2006. Efficacy of high doses of levofloxacin in experimental foreign-body infection by methicillin-susceptible Staphylococcus aureus. Antimicrob.Agents Chemother. 50:4011–4017.
35. NCCLS. 1999. Methods for determining bactericidal activity of antimicrobialagents; approved guideline, NCCLS document M26-A. National Committeefor Clinical Laboratory Standards, Wayne, PA.
36. Pinho, M. G., and J. Errington. 2005. Recruitment of penicillin-bindingprotein PBP2 to the division site of Staphylococcus aureus is dependent onits transpeptidation substrates. Mol. Microbiol. 55:799–807.
37. Rani, S. A., B. Pitts, H. Beyenal, R. A. Veluchamy, Z. Lewandowski, W. M.Davison, K. Buckingham-Meyer, and P. S. Stewart. 2007. Spatial patterns ofDNA replication, protein synthesis, and oxygen concentration within bacte-rial biofilms reveal diverse physiological states. J. Bacteriol. 189:4223–4233.
38. Rupp, M. E., P. D. Fey, and G. M. Longo. 2001. Effect of LY333328 againstvancomycin-resistant Enterococcus faecium in a rat central venous catheter-associated infection model. J. Antimicrob. Chemother. 47:705–707.
39. Saginur, R., M. Stdenis, W. Ferris, S. D. Aaron, F. Chan, C. Lee, and K.Ramotar. 2006. Multiple combination bactericidal testing of staphylococcalbiofilms from implant-associated infections. Antimicrob. Agents Chemother.50:55–61.
40. Saleh-Mghir, A., A. Lefort, Y. Petegnief, S. Dautrey, J. M. Vallois, D. LeGuludec, C. Carbon, and B. Fantin. 1999. Activity and diffusion of LY333328in experimental endocarditis due to vancomycin-resistant Enterococcus fae-calis. Antimicrob. Agents Chemother. 43:115–120.
41. Scheffers, D. J., and M. G. Pinho. 2005. Bacterial cell wall synthesis: newinsights from localization studies. Microbiol. Mol. Biol. Rev. 69:585–607.
42. Silverman, J. A., N. G. Perlmutter, and H. M. Shapiro. 2003. Correlation ofdaptomycin bactericidal activity and membrane depolarization in Staphylo-coccus aureus. Antimicrob. Agents Chemother. 47:2538–2544.
43. Spoering, A. L., and K. Lewis. 2001. Biofilms and planktonic cells of Pseudo-monas aeruginosa have similar resistance to killing by antimicrobials. J.Bacteriol. 183:6746–6751.
44. Stewart, P. S., and M. J. Franklin. 2008. Physiological heterogeneity inbiofilms. Nat. Rev. Microbiol. 6:199–210.
45. Svensson, E., H. Hanberger, and L. E. Nilsson. 1997. Pharmacodynamiceffects of antibiotics and antibiotic combinations on growing and nongrowingStaphylococcus epidermidis cells. Antimicrob. Agents Chemother. 41:107–111.
46. Touhami, A., M. H. Jericho, and T. J. Beveridge. 2004. Atomic force mi-croscopy of cell growth and division in Staphylococcus aureus. J. Bacteriol.186:3286–3295.
47. Van Bambeke, F., Y. Van Laethem, P. Courvalin, and P. M. Tulkens. 2004.Glycopeptide antibiotics: from conventional molecules to new derivatives.Drugs 64:913–936.
48. Wang, T.-S. A., D. Kahne, and S. Walker. 2007. Probing the mechanism ofinhibition of bacterial peptidoglycan glycosyltransferases by glycopeptideanalogs, abstr. C1-1474. Abstr. 47th Intersci. Conf. Antimicrob. Agents Che-mother., Chicago, IL, 17 to 20 September 2007.
VOL. 53, 2009 ORITAVANCIN KILLS S. AUREUS IN TOLERANT STATES 925
on May 30, 2018 by guest
http://aac.asm.org/
Dow
nloaded from