oxidation of acetaldehyde by acetobacter suboxydans · oxidation of acetaldehyde by acetobacter...
TRANSCRIPT

OXIDATION OF ACETALDEHYDE BY ACETOBACTER SUBOXYDANS *
BY TSOO l3:. KING AND VERNON H. CHELDELIN
(From the Department of Chemistry and the Science Research Institute, Oregon State College, Corvallis, Oregon)
(Received for publication, July 28, 1955)
Acetobacter suboxydans has a powerful system for the oxidation of ethanol to acetic acid, as might be expected from the fact that the acetic acid bacteria have long been used for the commercial production of vinegar.
Alcohol dehydrogenase in soluble extracts has been found to be DPlS- specific1 (1). It has now been observed that either TPN or DPN can be used as coenzyme for the oxidation of acetaldehyde. The concentration of DPN needed for optimal activity is, however, much higher than that of TPN. In this paper, the oxidative behavior, purification, and properties of the enzymes are presented.
EXPERIMENTAL
The A. suboxydans cells were harvested and lyophilized as reported previously (1). Protein was determined in crude preparation by the usual biuret reaction or turbidimetrically after the addition of trichloroacetic acid. The spectrophotometric method of Warburg and Christian was em- ployed (2) with preparations that had Ezso:EZ60 ratios of 1.6. Electro- phoreses were performed at 0” in a Perkin-Elmer Tiselius apparatus with a 2 ml. cell by both cylindrical lens and Longsworth scanning methods.
Materials-DPN and TPN were obtained commercially (Pabst; Sigma) either as the free acids or their sodium salts. The actual content was determined by use of crystalline yeast alcohol dehydrogenase for DPN and of purified glucose-6-phosphate dehydrogenase for TPN. A molar ab- sorbancy index of 6.22 X lo6 was used to convert optical density to molar concentrations. AElz4c represents the optical density change at wave-length 340 mp.
* Supported by grants from the Nutrition Foundnt,ion, Inc., and the Division of Research Grants, National Inst,itutes of Health, United States Public Health Serv- ice. Published with the approval of the Monographs Publications Committee, Re- search paper No. 289, School of Science, Department of Chemistry. Reported before the Third International Congress of Biochemistry, Brussels, August 1 to 6, 1955.
1 The following abbreviations are used throughout this paper: ADH, crystalline yeast alcohol dehydrogenase; CoA, coenzyme A; DPN and DPNH, oxidized and re- duced diphosphopyridine nucleotide; EDTA (Versene), ethylenediaminetetraacetic acid; TPN and TPNH, oxidized and reduced triphosphopyridine nucleotide; Tris, tris(hydroxymethyl)aminomethane.
177
by guest on June 16, 2018http://w
ww
.jbc.org/D
ownloaded from

178 ACETALDEHYDE OXIDATION BY A. SUBOXYDANS
Acetaldehyde was redistilled from Eastman’s white label material. Solu- tions were standardized according to Tomoda (3). Standardizations were always made at two concentrations of any acetaldehyde solution. The results from two concentrations usually checked within 1 per cent, whereas, in our hands, the method reported by Donnally (4) gave poorer agreement at different levels of acetaldehyde. This was probably due to insufficient control of the pH of the solutions.
ADH, twice crystallized and isolated from yeast according to Racker (5), was supplied by Dr. J. Wendell Davis. The sample was further re- crystallized twice from ammonium sulfate solutions.
Glucose-B-phosphate dehydrogenase was prepared from brewers’ yeast by the method of Kornberg (6).
Calcium phosphate gel, aged at least 6 months before use, was prepared according to Keilin and Hartree (7).
Other chemicals were obtained commercially. “Standard Procedure” for Routine Assay and Definition of Units-All
routine assays were made at room temperature (18-24”) in a Beckman spectrophotometer, model B, with 1 sq. cm. Corex cuvettes. However, during prolonged operation, excess heating was partially overcome by insulating the housing with asbestos boards and by leaving the cover open between readings.
Except when otherwise noted, the assay system contained 40 pmoles of Tris buffer, 40 pmoles of EDTA, 2.2 mg. of crystalline bovine serum albumin, either 1.34 pmoles of TPN or 12 pmoles of DPN, and an appro- priate amount of enzyme; final volume 2.2 ml., pH 8.6. The reaction was initiated by the addition of 8 pmoles of CH&HO. In determining ac- tivities with either TPN or DPN, a blank containing all constituents other than acetaldehyde was used. Blank values were usually less than 0.1 per cent of the experimental values. In the DPN experiments a solution of DPN was used in place of water to adjust the zero reading of the spectro- photometer, since DPN itself showed considerable absorption at the high concentrations used. When the AEs40 was less than 0.150, the reaction was strictly linear with time up to more than 4 minutes and directly pro- portional to the amount of the enzyme added.
1 unit of enzyme was defined as that amount which caused a change in optical density of 0.001 (AEgo = 0.001) per minute, calculated from the average of two cuvettes during the 2.5 minute period, between 30 and 180 seconds after the addition of acet,aldehyde. Specific activity was defined as the activity in units per mg. of protein.
Spectrophotometric determinations in kinetic studies were made in a Beckman spectrophotometer, model DU, with a photomultiplier attach- ment. The slit opening was maintained at 0.1 mm. by adjustment of
by guest on June 16, 2018http://w
ww
.jbc.org/D
ownloaded from

T. E. KING AND V. H. CHELDELIN 179
the sensitivity control. Temperature control to &0.05” in the reaction mixture was effected by proper arrangement of double ‘LThermospacers” and by circulating liquids from a constant temperature bath supplied by the manufacturer together with an Aminco constant low temperature bath, model 486 E. Below room temperatures, the Aminco bath was used alone. Temperature equilibration usually required about 25 minutes.
RESULTS AND DISCUSSION
Oxidative Behavior of Acetaldehyde
Acetaldehyde was oxidized by molecular oxygen either in the presence of resting cells or of the soluble or the insoluble (particulate) fractions of disintegrated cells (see Step 1A under “Purification of acetaldehyde de- hydrogenase”). Under these conditions no additional exogenous coenzyme was required. The activity of terminal oxidative enzymes was much lower than that of the primary dehydrogenases, so that, under the experimental conditions used, the reduction of pyridine nucleotides could be measured spectrophotometrically.
When pyridine nucleotides were used as electron acceptors, it was found that both TPN and DPN were active. The concentration of DPN re- quired to show significant activity was, however, much higher than that of TPN (for example, Fig. 2, A and B). The apparent interchangeability of these coenzymes could be due to several possibilities: (a) pyridine nucle- otide transhydrogenase (8) might be present in the enzyme preparations; (b) TPN might be an impurity in the DPN samples used, and either might possess activity per se or feed the transhydrogenase reaction; (c) the pres- ence of two different enzymes; or (d) activity of both DPN and TPN for a single enzyme, as in lactic dehydrogenase (9).
To test possibility (a), assays of pyridine nucleotide transhydrogenase activity were made with a mixture of 0.3 pmole of TPNH, 10 pmoles of DPN, and 1.7 mg. of enzyme (Step 2) in a 2 ml. volume. As determined with ADH, negligible amounts of DPNH were produced after 5 minutes incubation of the mixture followed by boiling.
Commercial samples of DPN which were purified according to Kornberg and Pricer (10) still displayed the same activity. In addition, even com- mercial DPN did not oxidize glucose-g-phosphate in the presence of a purified yeast glucose-6-phosphate dehydrogenase, whereas the reaction proceeded immediately upon addition of TPN to the mixture. Finally, possibility (b) was ruled out in still another manner with DPN-specific yeast ADH, by quantitative estimation of DPNH formed in the oxidation of acetaldehyde by the A. suboxydans enzyme. The remaining possibilities (c) and (d) are considered in later sections.
by guest on June 16, 2018http://w
ww
.jbc.org/D
ownloaded from

180 ACETALDEHYDE OXIDATION BY A. SUBOXYDANS
Purification of Acetaldehyde Dehydrogenase
Soluble extracts prepared by either alumina grinding or sonic disinte- gration were active. After attempting various methods of protein frac- tionation, the following procedure was found to give about loo-fold con- centration.
Step 1. Preparation of Soluble Extracts-Disintegration by alumina grinding has been described previously (1). 0.02 M phosphate buffer at pH 6.0 was used for extraction. Alternatively, 5 gm. of lyophilized cells were suspended in 50 ml. of the phosphate buffer in a Waring blendor. The mixture was treated in a Raytheon 10 kc. sonic vibrator at the maxi- mal power output, for 30 minutes net elapsed time. (A 1 minute interlude followed each 2 minutes vibration.) The apparatus was cooled by running tap water rapidly through the jacket. The outlet temperature was usu- ally below 8”. Several batches from the treatment were combined. The pooled mixture was centrifuged for 1 hour in a Spinco model L centrifuge with a rotor No. 30.2 at a speed of 25,000 X g. About 45 ml. of a clear, pink, viscous liquid were obtained from 5 gm. of cells. The protein con- tent and specific activity varied wit)h the length of the disintegration and with each batch of cells, even when the conditions were controlled as rigidly as possible. As a rule, longer treatment yielded higher protein content in the extracts and less insoluble matter. However, no correlation was noted between the protein content and specific activities. Specific activities of the supernatant solutions from twenty-eight batches of alumina-ground and sonic-disintegrated cells varied from 500 to 2000 with TPN as coen- zyme and from 100 to 300 with DPN.
A typical run from 15 gm. of lyophilized cells, in which the residue was reextracted in a Waring blendor with phosphate buffer, furnished 440 ml. of soluble extract, containing 20 mg. of protein per ml,
Step 1A. Preparation of “Particulates”-In both sonic- and alumina- ground extracts, a pink layer was observed at the top of the pellet formed during centrifugation. This layer was carefully removed with a plastic spatula and suspended in 20 volumes of 0.02 M phosphate buffer, pH 6.0. The suspension was centrifuged for 30 minutes at 15,000 X g in a refrig- erated International No. 2 centrifuge. The upper, pink layer was again removed, suspended in buffer, and recentrifuged, and t,he process was again repeated. Complete separation from inorganic materials as well as from intact cells and soluble extract was achieved. The resulting pink solid, apparently homogeneous, was suspended in buffer and used within 24 hours. It catalyzed acetaldehyde oxidation in air.
Except as otherwise noted, all subsequent operations were performed at O-4”, with centrifugat,ion being carried out’ in a refrigerated International So. 2 model, at about 3000 r.p.m.
by guest on June 16, 2018http://w
ww
.jbc.org/D
ownloaded from

T. E. KING AND V. H. CHELDELIN 181
Step 2. Protamine Precipitation-440 ml. of the liquid from Step 1 were treated with 52 ml. of 2 per cent protamine sulfate. The precipitation was always first carried out with a small aliquot in order to find the minimal amount of protamine required. The mixture was allowed to stand for 30 minutes w&h occasional stirring and then centrifuged. The precipitate was discarded. 460 ml. of supernatant liquid were obtained with a protein concentration of 3.5 mg. per ml.
Step 3. Ammonium Sulfate Fractionation-The supernatant liquid from the foregoing step was readjusted to pH 6.0 with saturated Na2HP04. Solid ammonium sulfate was added with slow stirring, and the fraction precipitating between 57 and 72 per cent saturation was retained.2 This fraction was then dissolved in 95 ml. of 0.02 M phosphate buffer, pH 6.0, and dialyzed against the buffer for about 12 hours. A small amount of precipitate that formed during the dialysis was removed by centrifugation. 100 ml. of a clear solution were obtained with a protein concentration of 2.8 mg. per ml.
Step 4. Calcium Phosphate Gel Adsorption-49 ml. of calcium phosphate gel containing 8.0 mg. of solid per ml. were centrifuged. The solid was mixed with the product obtained from Step 3, stirred slowly for 20 min- utes, and centrifuged. 96 ml. of supernatant liquid were obtained with a protein concentration of 0.91 mg. per ml.
The optimal ratio of protein to gel varied considerably among different samples from Step 3. It was necessary to determine the preferred amount of gel required in each experiment to produce the best concentration of the enzyme, usually about 3-fold. Larger amounts of the gel did not enhance the specific activity of the enzyme, but did impair the yield.
Step 5. Acetone Fractionation-96 ml. of the effluent from the gel ad- sorption were cooled to - 1”. 48 ml. of acetone at - 10” were added drop- wise with stirring. After an additional 30 minutes stirring, the precipitate was centrifuged at -6” and discarded. 32 ml. of cold acetone were added dropwise to 126 ml. of the supernatant liquid with stirring. The precipi- tate was centrifuged, retained, and dissolved in 0.02 M phosphate buffer at pH 6.0. A small amount of insoluble material was removed by centrifu- gation. The solution was dialyzed against the buffer for about 12 hours, and any precipitates formed during dialysis were centrifuged off. 16 ml. of clear solution were obtained with a protein concentration of 2.0 mg. per ml.
2 The degree of ammonium sulfate saturation was calculated from the following equation: 1: = (50.6 (Sz - S,))/(l - 0.286&) modified from Kunitz (11) to give the saturation value at 0”. The derivation was made by employing 0.506 gm. of (NH,)*- SO4 per ml. as a saturated solution at O”, and 0.565 ml. per gm. as the specific volume of solid salt.
by guest on June 16, 2018http://w
ww
.jbc.org/D
ownloaded from

182 ACETALDEHYDIC OXIDATION BY A. SUBOXYDANS
Step 6. Second Ammonium Sulfate Precipitation-To 15 ml. of the acetone fraction, solid ammonium sulfate was added slowly to 60 per cent saturation. A small amount of precipitate was centrifuged off and dis- carded. 5 ml. of saturated ammonium sulfate in 0.02 M phosphate buffer at pH 6.0 were added to 15 ml. of the above supernatant liquid over a 2 hour period. The precipitate was collected, dissolved in the buffer, and dialyzed against the buffer for 8 hours. The resulting volume was Tj.6 ml., with a protein concentration of 3.35 mg. per ml.
This step was made in an attempt to crystallize the enzyme. Many variations in operation were used, but no indication of crystallization could be observed. All treatments were carried out at O”, since thermal de-
TABLE I
Purijkation of Acetaldehyde Dehydrogenase from A. suboxydans
TPN
Fraction
I I
Specific activity* Yield
DPN
Step 1,extract ...................... “ 2, protamine supernatant liquid ‘( 3, 1st (NH4)nSOa fraction ....... “ 4, gel effluent. ................. “ 5, acetone fraction. ............ “ 6, 2nd (NHI)zSOJ fraction. ..... . .
ger cent
1,100 4,600 76
15,000 57 42,000 87 81,000 70
108,800 84
Specific activity* Yield
-- per cent
170 800 86
3,800 84 11,500 97 18,400 57 25,000 85
6.5 5.8 4.0 3.6 4.4 4.1
* Units per mg. ; assayed by “standard procedure” (see “Experimental”). t Activity ratio, (specific activity with TPN/specific activity with DPN).
naturation at this step was found to be very great. Higher temperature might induce easier crystallization.
An electrophoretic pattern of this sample in phosphate buffer, pH 6.0, at ionic strength of 0.1, showed not more than five peaks during 6 hours electrophoresis. The activity remained in the components with lower mobilities. No satisfactory methods were developed for the mechanical separation of components from the 2 ml. cell used.
A summary of the specific activities and yields is presented in Table I. It should be mentioned that both yields and purity varied considerably among different batches of extracts. This variance was evidently due to the different degrees of disintegration of the cells, which at present cannot be quantitatively controlled. Likewise, the Ezso:E2,~ ratios in the crude extract (Step 1) varied from batch to batch, although they became con- stant at 1.6 after Step 2. In general, the specific activity of the final
by guest on June 16, 2018http://w
ww
.jbc.org/D
ownloaded from

T. E. KING AND V. H. CHELDELIN 183
preparation was at least 70,000 with TPN and 20,000 with DPN. This value is considerably higher than any previously recorded for acetaldehyde dehydrogenases from yeast or liver (12-14).
Two preparations gave specific activities as high as 140,000 with TPN and about 30,000 with DPN. However, the samples with lower specific activities were not improved by repeated treatment.
Seegmiller’s TPN-linked aldehyde dehydrogenase from yeast (12) dis- played high affinity toward aluminum hydroxide gel at pH 5.3 in acetate buffer, with an increase in enzyme activity of about 3-fold by adsorption and subsequent elution with phosphate buffer, pH 7.1. However, the i1. suboxydans enzyme was not adsorbed by alumina C-y even at a gel- protein ratio of 9:7 in dilute acetate buffers of pH 5.2 to 5.6 or in phos- phate buffer at pH 6.0.
Attempted Separation of Activity with TPN from L)PN-Ratios of specific activity with TPN compared to that with DPN (cf. Table I) in various fractions from the above procedure were not sufficiently different to tell whether one or two acetaldehyde dehydrogenases existed in the organism. The higher ratios in the early purification steps might have been due to the presence of DPN-linked alcohol dehydrogenase in the samples (1) OI to unequal activities of DPNH and TPNH oxidases. However, after Step 3, negligible amounts of the alcohol dehydrogenase and pyridine nucleotide oxidase were found.
Various methods were tried in order to separate one activity from the other. So far, these separations have been unsuccessful with the exception of zinc-ethanol fractionation (15, 16) described below.
An extract from Step 1 (42 ml.) was passed through a Dowex 1 column of 2 X 30 cm. The combined volume of the effluent and washings was 84 ml. To the solution were added 56 gm. of solid ammonium sulfate. After standing 20 minutes, the precipitate was collected by centrifugation, dissolved to 70 ml. with water, and dialyzed against water for 24 hours. Insoluble material was centrifuged off and discarded. The resulting vol- ume was 76 ml. (Fraction 1-A).
43 ml. of Fraction 1-A were diluted to 155 ml. with water to give 3 mg. of protein per ml. 10 ml. of 0.02 M zinc acetate in 0.1 M sodium acetate were added. A small amount of precipitate was formed and removed by centrifugation. 12 ml. of 70 per cent alcohol at -5” were added in 5 minutes to give an alcohol concentration of 4.8 per cent. The mixture was centrifuged immediately, and the supernatant liquid was retained for further treatment (see the next paragraph). The precipitate was sus- pended in 10 ml. of 0.01 M citrate buffer, pH 6.0. The mixture was di- alyzed against the buffer for 4 hours. The precipitate was collected by
by guest on June 16, 2018http://w
ww
.jbc.org/D
ownloaded from

184 ACETALDEHYDE OXIDATION BY A. SUBOXYDANR
centrifugation, then suspended in 12 ml. of 0.1 M Tris buffer at pH 8.4, and dialyzed against 0.01 M Tris buffer for 16 hours. The resulting solu- tion was clear (Fraction 1-B).
TABLE II
Zinc-Ethanol Fractionation of Acetaldehyde Dehydrogenase from A. suboxydans*
I I I I
I Prorein Totie Specific activity* Ratia TPN DPN
____- -~
“g&t”’ d.
Fraction 1-A (Dowex 1 effluent). 11 43 2100 500 4.2 “ 1-B (4.8% ethanol ppt.). 3.2 14 0 54 0 “ 1-C (14% ethanol ppt.). . . 5.3 25 360 0 00
Per cent recovery
TPN DPN
5 10
* See Table I for an explanation of the units of measure and activity ratios.
TABLE III
Non-Additive Nature of TPN and DPN Activities in Acetaldehyde Dehydrogenase from A. suboxydans
Fractions
Step 1
(‘ 2
“ 3
(‘ 4
“ 5
TPN DPN TPN + DPN TPN + DPN
TPN
1,200 250 980 0.82 800 190 610 0.76
3,900 950 3,550 0.86 4,600 800 4,300 0.93
15,000 3,800 13,000 0.86 14,000 3,500 11,200 0.80 37,500 12,440 33,400 0.90 42,000 11,500 34,500 0.82 81,000 18,400 62,000 0.77 66,000 19,000 56,000 0.85 90,000 22,000 72,500 0.80
Specific activity*
* See Table I for an explanation of the units of measure.
170 ml. of the supernatant solution from the preceding alcohol fractiona- tion were mixed with 10 ml. of zinc acetate-sodium acetate buffer. Ethanol was added to 14 per cent concentration. The precipitate was suspended in citrate buffer and dialyzed against the buffer for 4 hours. The precipi- tate was then centrifuged off and discarded. The supernatant liquid was dialyzed against 0.01 M Tris buffer at pH 8.4 for 16 hours. The resulting solution was also clear (Fraction 1-C).
by guest on June 16, 2018http://w
ww
.jbc.org/D
ownloaded from

T. E. KING AND V. H. CHELDELIN 185
The summary of activities of the fractions from the zinc-ethanol treat- ment appears in Table II. Although the TPN and DPN activities were separated by this fractionation procedure, the specific activity with either nucleotide was greatly decreased, and the recoveries were very poor. In view of the low recoveries and the fact that the DPN and TPN activities were not additive, indeed were even mutually inhibitory (cj. Table III), it is perhaps unsafe to conclude definitely that two separate enzymes exist despite the apparent resolution with zinc and ethanol.
Properties of Acetaldehyde Dehydrogenase
E$ect of Potassium, Magnesium, Phosphate, CoA, Cysteine, and EDT8 -No stimulatory effect was found by additions of KC1 or MgClz up to 4 X 1V M, either alone or in combination. Thus this enzyme differs from the dehydrogenase reported by Black (13). Likewise, phosphate and CoA were not required. No evidence was found for the formation of acetyl CoA during the oxidation, in contrast to an acetaldehyde dehydrogenase from Clostridium (17).
Cysteine, glutathione, or EDTA was stimulatory. However, the activities were not additive. The extent of stimulation varied with dif- ferent preparations of the enzyme. This was evidently due to traces of heavy metals which combined with sulfhydryl groups on the enzyme and which could be released by large amounts of sulfhydryl compounds or chelating agents.
Optimal pH-The optimal pH for both TPN and DPN activity was found to be 8.7. No special effects were produced by the individual buf- fers for TPN. DPN activity varied with different buffers, although the optimal pH was similar for each. It was found that Tris was better than barbital, glycylglycine, or a-alanine. The nature of the buffer effect is not known at the present time. It may have been due to differences in chelating action by the buffers, because the enzyme was sensitive to heavy metal ions.
Michaelis Constants of Enzyme-Substrate Complexes-Michaelis con- stants were determined over substrate concentrations ranging from 3.8 X lo-5 t0 7.6 x 1o-4 M. The reaction was started by addition to the cuvette of 0.028 ml. of appropriate acetaldehyde solutions from a micro pipette to make the final volume 3.0 ml. The oxidation rate was expressed as the average during the 2.5 minute period between 30 and 180 seconds, except that at 3.8 X 1O-5 M the rate was taken for the 1 minute period between 30 and 90 seconds. V,,, and K, were determined according to Lineweaver and Burk (18) by the method of least squares. In Fig. I, A and R, the K, of the enzyme-acetaldehyde complex with TPN (9.8 X 1O-5 moles per liter) is smaller t,han that with DPN (12.8 X lop5 moles per
by guest on June 16, 2018http://w
ww
.jbc.org/D
ownloaded from

186 ACETALDEHYDE OXIDATION BY A. SUBOXYDrlNS
liter). Both values are in the same order of magnitude as a TPN-linked dehydrogenase from yeast (12). V,,, with TPN (0.0789 pmole per minute per 8 y of enzyme) is much larger than that with DPN (0.104 pmole per minute per 42 y of enzyme).
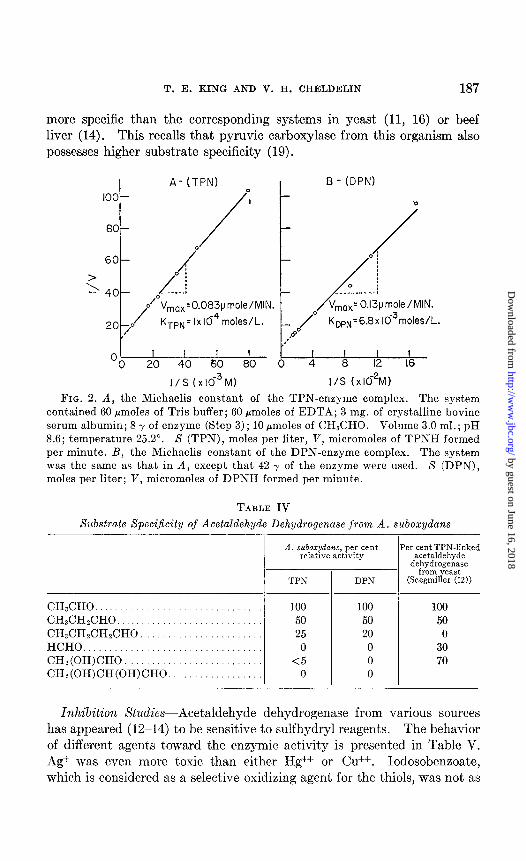
Michaelis Constants of Enzyme-Coenzyme Complexes-These were also determined by Lineweaver and Burk’s plot, by measuring the initial rate against concentrations of coenzymes. The reaction was started by the addition of acetaldehyde. As shown in Fig. 2, A and B, K,,, is about 70 times larger than KTPN. KTPN for the enzyme from A. suboxydans
A - (TPN) I B- (DPN)
FIG. 1. A, the Michaelis constant of the enzyme (TPN)-substrate complex. The system contained 60 pmoles of barbital buffer, 60 pmoles of EDTA, 3 mg. of crystal- line bovine serum albumin, 8 y of enzyme (Step 3), 1.83 pmoles of TPN. Volume 3.0 ml.; pH. 8.6; temperature 25”. S (acetaldehyde), moles per liter; V, micromoles of TPNH formed per minute. B, the Michaelis constant of the enzyme (DPN)-sub- strate complex. The system was the same as that in A, except that 20 pmoles of DPN and 42 y of the enzyme were used.
is higher than that of the yeast enzymes (12). Among pyridinoproteins, the present enzyme (with DPN) has one of the lowest recorded affinities for a coenzyme.
Substrate Specificity-Purified aldehyde dehydrogenase from either yeast or beef liver is relatively unspecific toward various aldehydes. As depicted in Table IV, the enzyme from A. suboxyclans catalyzed the oxida- t’ion of propionaldehyde, and n-butyraldehyde among various aldehydes tested, whereas DL-glyceraldehyde was completely inert. The slight. activity toward glycolaldehyde was doubtful. The lower activity toward the latter aldehydes was not due to low enzyme-substrate affinities, since a 3-fold increase in concentration of the aldehydes failed to increase their oxidation rates. It is evident that the enzyme from A. suboxydans is
by guest on June 16, 2018http://w
ww
.jbc.org/D
ownloaded from
T. E. KING AND V. H. CHELDELIN 187
more specific than the corresponding systems in yeast (11, 16) or beef liver (14). This recalls that pyruvic carboxylase from this organism also possesses higher substrate specificity (19).
IOO-
A- (TPNI o
a
B - (DPN)
‘0
80-
>
b 40- / i
0 : __ -___--. 1 0 =O.O83~mole/MIN.
2. -,/ 2;;; ,x 104 mo,es,L. - /’ _ p ~~~1~k%:~lz.
/ *e” ,’
I I I I I I 1 I ‘0 20 40 60 80 0 4 8 12 16
I/S (xIO-~M) I/S (x102M)
FIG. 2. A, the Michaelis constant of the TPN-enzyme complex. The system contained 60 Bmoles of Tris buffer; 60 pmoles of EDTA; 3 mg. of crystalline bovine serum albumin; 8 y of enzyme (Step 3); 10 Imoles of CH&HO. Volume 3.0 ml.; pH 8.6; temperature 25.2”. S (TPN), moles per liter, V, micromoles of TPNH formed per minute. B, the Michaelis constant of the DPN-enzyme complex. The system was the same as that in A, except that 42 y of the enzyme were used. S (DPN), moles per liter; V, micromoles of DPNH formed per minute.
TABLE IV
Substrate Specificity of Acetaldehyde Dehydrogenase from A. suboxydans
A. subozydam, per cent relative activity
TPN DPN
Per cent TPN-linked acetaldehyde
dehydrogenase from yeast
(Seegmiller (12))
CHsCHO. CH,CH&HO CH,CH,CH&HO HCHO. CH2(0H)CHO. CH:!(OH)CH(OH)CHO.
100 100 50 50 25 20
0 0 <5 0
0 0
100 50
0 30 70
Inhibition Studies-Acetaldehyde dehydrogenase from various sources has appeared (12-14) to be sensitive to sulfhydryl reagents. The behavior of different agents toward the enzymic activity is presented in Table V. Ag+ was even more toxic than either Hg++ or Cu++. Iodosobenzoate, which is considered as a selective oxidizing agent for the thiols, was not as
by guest on June 16, 2018http://w
ww
.jbc.org/D
ownloaded from

188 ACETALDEHYDE OXIDATION BY A. SUBOXYDANS
potent as the heavy metals. The quantitative differences in the effects of iodosobenzoate and Zn++ on TPN and DPN reduction suggest that the essential apoenzyme sites for these two activities may not be identical. However, this does not necessarily mean that two enzymes are present.
TABLE V
Behavior of Sulfhydryl Reagents toward Acetaldehyde Dehydrogenase from A. suboxydans
Inhibitor
-
CUSOl
AgNO,
HgW&)?
ZnClr
pmChloromercuribenzoate
Iodosobenzoate
Concentration
M 2 x 10-d 2 x 10-S 1 x 10-t 2 x 10-S 1.5 x 10-S 2 x 10-e 2 x 10-h 2 x 10-s 3.6 X lo-” 7.3 x 10-c 1 x 10-a 1 x 10-d 2 x 10-S 1 x 10-S 2 x 10-z 1 x 10-z 2 x 10-a
-
l- Per cent inhibition
TPN DPN
87 39 98 86 60
0 98 53 86 50 97 64
98 53
0
64
94
92
52 5
100 97 70 38
86 35
The basal system for the activity with TPN contained 80 pmoles of barbital buffer; 1.34 rmoles of TPN; 2 mg. of crystalline bovine serum albumin; 10 y of en- zyme (Step 3); 8 pmoles of acetaldehyde. Volume 2.2 ml., pH 8.6; room tempera- ture, The basal system for the activity with DPN was the same as above, except that 12 pmoles of DPN and 45 y of the enzyme were used.
Chelating agents such as EDTA or thiols such as cysteine or glutathione could overcome the inhibitions. Tris alone gave only partial reversal.
Activation Energy-The temperature effect on enzymic activity was studied by measuring the initial rate, i.e. zero order, with respect to either the coenzyme or acetaldehyde. Thus the reaction constant of the Ar- rhenius equation will be the rate of the reaction. In Table VI, the activa- tion energy? was about 13 t)o 14 kilocalories with either coenzyme (SW
3 By plotting log k versus l/T, the slope would be equal to - ~/(2.303 R) of tho ;1rrhenius equation. Instend of direct, plotting, the least, squaresmethod forthe best. $1 raigbt lint, was llsetl.
by guest on June 16, 2018http://w
ww
.jbc.org/D
ownloaded from

T. E. KING AND V. II. CHELDELIRT 189
Table VII for a statistical analysis of the data). The ,.L values are ob- viously much lower than those in the absence of the enzyme, in view of
TABLE VI
Activation Energy for Oxidation of Acetaldehyde by TPN and DPN in Presence of Acetaldehyde Dehydrogenase from A. suboxydans
TPN* DPNt
Experiment 1 Experiment 2 Experiment 3 Experiment 4
,u (activation energy), Cal- ories.. 14,240 14,310 14,320 12,700
Standard error of p. 293 232 796 1,020
Confidence limits of p$ 14,960 and 13,530 14,900 and 13,710 16,270 and 12,370 15,390 and 10,150
* The system contained 60 pmoles of Tris buffer, 60 rmoles of EDTA, 3 mg. of crystalline bovine serum albumin, 1.93 pmoles of TPN, 6.0 y of enzyme (Step 3); volume 3.0 ml. The reaction was started by the addition of 10 pmoles of CH&HO. Eight temperatures between 275.4-310.4” K. were used. Individual values of k were obtained after determination at several time intervals and extrapolation to zero time. This was necessitated by the partial denaturation of the enzyme during tem- perature equilibration (up to 25 minutes to reach the last degree of temperature).
t The system and conditions were the same as those for TPN, except that 20 pmoles of DPN and 32 y of the enzyme were used.
$ With a confidence coefficient of 0.95.
TABLE VII
Statistical Analysis of p Values for Reaction with TPN and with DPN
Source of variation
Among Slopes b1 to b4t. Slopes b, and b? vs. b3 and b4. Slope b1 vs. bz.
“ bavs.bq Error... .._.......
Sum of squares
0.008495 0.002066 0.000011 0.006418 0.052711
I
fr
3 1 1 1
22 -
0.002832 0.002066 0.000011 0.006418 0.002396
1.18 (F0.05 = 3.05) 0.86 0.00 2.68 (Fo.os = 4.30)
* F, variance ratio. t Slopes bi, bq, bl, and br are the best slopes of the straight lines from Experi-
ments 1, 2, 3, and 4, respectively.
the fact that acetaldehyde is very stable in the presence of pyridine nucleo- tides even at elevated temperatures.
Xtability-The oxidative activity with either coenzyme in crude extracts
by guest on June 16, 2018http://w
ww
.jbc.org/D
ownloaded from

190 ACETALDEHYDE OXIDATION BY A. SUBOXYDANS
(Step 1) was not impaired by heating at 56” for 10 minutes, at 30” for 4 hours, by standing at room temperature for 14 hours, by repeated freezing and thawing, or by lyophilization. A sample stored at - 10” for 20 months retained more than 80 per cent of its original activity. However, thermal inactivation was rapid in fractions after Step 2. These lost activity gradually over a few weeks time, even when stored at -10”.
An attempt to determine the kinetics of the denaturation was not suc- cessful. The denaturation rate was determined between 300.9-310” IL Rate constants changed with time and were thus impossible to calculate (cl. Fig. 3). It has been found (cf. (20)) that the denaturation of protein
O71
FIG. 3. Stability of acetaldehyde dehydrogenase with TPN from A. suboxydans. The conditions were the same as in Table VI.
follows simple orders of reaction. In this dehydrogenase under the condi- tions tested, the whole course of the denaturation was evidently complex. ‘l’his fact suggested that more than one reaction may have taken place. When the initial denaturation was treated as a first order reaction, a provi- sional value of over 100 kilocalories was obtained for the activation energy of denaturation.
Acetobacter suboxydans has a powerful acetaldehyde oxidation system. The primary dehydrogenase, after concentration over IOO-fold from a soluble cell-free extract, exhibited activity of over 100,000 spectrophoto- metric “units” per mg. per minute, with TPN as coenzyme.
At all stages of purification, the apoenzyme could be reactivated with either TPN or DPX, At the highest stages of purity, TPN was approxi-
by guest on June 16, 2018http://w
ww
.jbc.org/D
ownloaded from

T. E. KING Ai\‘D V. H. CHELDELIN 191
mstely four times as effective as DPN. Evidence indicated that DPN was effective per se, although it was not possible to state with certainty whether two dehydrogenases were present.
Various physical constants and other properties of the dehydrogenase are described.
The authors are grateful to Rowena G. Knudson and Margaret G. Thome for their valuable technical assistance, and to Dr. Jerome C. R. Li for performing the statistical analyses of the data.
BIBLIOGRAPHY
1. King, T. E., and Cheldelin, V. H., Biochim. et biophys. acta, 14, 108 (1954). 2. Warburg, O., and Christian, W., Biochem. Z., 310, 384 (1941-42). 3. Tomoda, Y., .I. Sac. Chem. Ind., 48, 76 T (1929). 4. Donnally, L. H., Ind. and Eng. Chem., Anal. Ed., 5, 91 (1933). 5. Racker, E., .I. Biol. Chem., 184, 313 (1950). 6. Kornberg, A., .I. Biol. Chem., 182, 805 (1950). 7. Keilin, D., and Hartree, E. F., Proc. Roy. Sac. London, Series B, 124, 397 (1938). 8. Kaplan, N. O., Colowick, S. P., and Neufeld, E. F., .I. Biol. Chem., 205, 1 (1953). 9. Schlenk, F., in Sumner, J. B., and Myrblck, K., The enzymes, New York, 2, pt.
1, 250 (1951). 10. Kornberg, A., and Pricer, W. E., Jr., Biochem. Preparations, 3, 20 (1953). 11. Kunitz, M., .I. Gen. Physiol., 35, 423 (1952). 12. Seegmiller, J. E., J. Biol. Chem., 201, 629 (1953). 13. Black, S., Arch. Biochenz. and Biophys., 34, 86 (1951). 14. Racker, E., J. Biol. Chem., 177, 883 (1949). 15. Tallan, H. H., Jones, M. E., and Fruton, J. S., .I. Biol. Chem., 194, 793 (1952). 16. Cohn, E. J., Gurd, F. R. N., Surgenor, D. M., Barnes, B. A., Brown, R. K.,
Derouaux, G., Gillespie, J. M., Kahnt, F. W., Lever, W. F., Liu, C. H., Mittel- man, D., Mouton, R. F., Schmid, K., and Uroma, E., .I. Am. Chem. Sot., 72, 465 (1950).
17. Burton, R. M., and Stadtman, E. R., I. Biol. Chem., 202, 873 (1953). 18. Lineweaver, H., and Burk, D., .I. Am. Chem. Sot., 66, 658 (1934). 19. King, T. E., and Cheldelin, V. H., .I. BioZ. Chem., 208, 821 (1954). 20. Putnam, F. W., in Neurath, H., and Bailey, K., The proteins, New York, 1, pt.
B, 807 (1953).
by guest on June 16, 2018http://w
ww
.jbc.org/D
ownloaded from

Tsoo E. King and Vernon H. CheldelinACETOBACTER SUBOXYDANS
OXIDATION OF ACETALDEHYDE BY
1956, 220:177-191.J. Biol. Chem.
http://www.jbc.org/content/220/1/177.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
ml#ref-list-1
http://www.jbc.org/content/220/1/177.citation.full.htaccessed free atThis article cites 0 references, 0 of which can be
by guest on June 16, 2018http://w
ww
.jbc.org/D
ownloaded from