penelitian i: pendugaan ragam dan model genetik karakter … · hasil penelitian yang dilaporkan...
TRANSCRIPT

21
Penelitian I: Pendugaan Ragam dan Model Genetik Karakter Ketahananterhadap Penyakit Bulai pada Jagung
Pendahuluan
Kendala biotis yang paling sering terjadi dalam budidaya jagung di Indonesia adalah
penyakit bulai yang disebabkan oleh P. maydis. Patogen tersebut cukup berbahaya karena
dapat menyebabkan kehilangan hasil hingga 90 persen (Mikoshiba, 1983) dan bahkan
dapat menyebabkan kegagalan panen (Semangun, 1996; Subandi et al., 1996).
Hasil penelitian yang dilaporkan oleh Wakman dan Kontong (2000) di Lanrang
(Sulawesi Selatan) serta Azrai dan Kasim (2005a) di Maros, Bogor, dan Lampung
menunjukkan bahwa ketahanan dari beberapa varietas jagung unggul nasional terhadap
penyakit bulai masih bervariasi, yaitu dari sangat rentan hingga resisten. Hal ini
menunjukkan bahwa tingkat ketahanan jagung terhadap patogen penyebab penyakit bulai
cukup beragam, tergantung pada variabilitas genetik, variabilitas fenotipik, dan interaksi
antara genetik dengan lingkungannya. Pengetahuan mengenai keragaman tersebut sangat
penting terutama dalam penerapan program seleksi yang akan digunakan untuk
mendapatkan suatu karakter yang diinginkan (Prasanna, 2002).
Upaya perakitan jagung yang resisten terhadap penyakit bulai terus dilakukan
melalui penyaringan plasmanutfah yang dilanjutkan dengan kegiatan persilangan antara
tetua terpilih. Kegiatan tersebut telah lama dilakukan oleh pemulia jagung, akan tetapi
sejauh ini belum banyak dilaporkan mengenai model genetik ketahanan terhadap penyakit
bulai pada jagung di Indonesia.
Pengetahuan tentang sifat dan aksi gen yang mengendalikan suatu karakter sangat
penting terutama dalam hal keefektifan penerapan program seleksi yang akan digunakan
untuk mendapatkan karakter yang diinginkan. Beberapa peneliti melaporkan bahwa
karakter ketahanan terhadap penyakit bulai pada jagung dikendalikan oleh gen tunggal
(Chang dan Cheng, 1968; Chang, 1972; Handoo et al. 1970; Takdir et al., 2004),
sedangkan beberapa peneliti yang lain melaporkan melaporkan bahwa karakter ini
dikendalikan oleh banyak gen (polygenic) (Francis, 1967; Carangal et al. 1970; Hakim
dan Dahlan, 1972; Peerasak, 1974; Ruswandi, 2001).
Salah satu contoh kasus perbedaan hasil penelitian yang paling menonjol adalah
studi tentang kendali genetik terhadap P. philippinensis. Aday (1974), melaporkan bahwa
pola pewarisan ketahanan terhadap P. philippinensis dikendalikan oleh gen-gen dominan

22
dengan derajat dominansi berada dalam over dominan. Hal ini berbeda dengan yang
dilaporkan oleh Rifin (1983) yang menyatakan bahwa ketahanan tanaman jagung
terhadap P. philippinensis ditentukan oleh gen aditif dan dominan, tetapi aditif lebih
menonjol, sedangkan Ruswandi et. al., (2002) secara jelas menyatakan bahwa pola
pewarisan ketahanan tanaman jagung terhadap P. philippinensis bersifat kuantitatif
dengan efek aditif dan epistasis. Perbedaan tersebut diduga kuat karena perbedaan tingkat
ketahanan dari tetua persilangan, jumlah generasi, dan besarnya genotip yang diteliti.
Untuk itu, informasi tentang model genetik ketahanan terhadap penyakit bulai pada
jagung di Indonesia dirasa perlu untuk ditelaah lebih lanjut.
Berdasarkan hal tersebut, maka dilakukan penelitian yang bertujuan untuk
mengetahui variabilitas dan model genetik karakter ketahanan terhadap P. maydis pada
set persilangan MR10 x CML161 dan Nei9008 x CML161.
Bahan dan Metode Penelitian
Tempat dan WaktuPenelitian dilaksanakan di Kebun Percobaan Cikeumeuh, Bogor, terbagi atas dua
tahap, yaitu pembentukan genotip uji dan pengujian ketahanan genotip uji terhadap
penyakit bulai. Pembentukan genotip dilaksanakan dari Agustus 2004 sampai April 2005.
Pengujian ketahanan genotip uji terhadap penyakit bulai berlangsung dari bulan Januari-
Maret 2006.
Bahan Penelitian
Materi genetik yang digunakan pada penelitian ini masing-masing terdiri atas 7
macam populasi pada dua set persilangan dan progeninya, yaitu: masing-masing 10
genotip (tongkol) tetua persilangan yaitu P1 (CML161) sebagai tetua rentan (donor gen
opaque-2) dan P2 (MR10 dan Nei9008) sebagai tetua resisten (silang balik = recurrent),
masing-masing 20 genotip dari generasi F1, F2, BC1P1, dan BC1P2 serta 100 genotip dari
generasi F3. CML161 merupakan galur yang bermutu protein tinggi (QPM = Quality
Protein Maize) yang diintroduksi dari CIMMYT-Mexico. Nei9008 diintroduksi dari
Thailand melalui Jaringan Kerjasama Bioteknologi Jagung Asia (AMBIONET = Asian
Maize Biotechnology Network) dan MR10 merupakan salah satu galur elit Balitseral yang
digunakan sebagai tetua hibrida Semar 8. Selain itu, juga digunakan Varietas Anoman-1
sebagai cek rentan dan tanaman baris penyebar konidia bulai. Varietas Anoman-1

23
merupakan salah satu varietas bersari bebas milik Balitsereal yang sangat rentan terhadap
penyakit bulai (Azrai, 2006).
Pembentukan Genotip Uji
Pembentukan genotip uji dilakukan dengan cara melakukan silang balik antara
tanaman F1 dengan kedua tetuanya untuk membentuk masing-masing 20 genotip BC1P1
dan BC1P2 dan mensegregasikan tanaman F1 untuk membentuk 20 genotip F2. Benih
genotip F2 masing-masing ditanam dua baris kemudian dilakukan penyerbukan sendiri
sebanyak 10 tanaman per genotip. Setelah panen, dipilih 5 tongkol per genotip sehingga
diperoleh 100 tongkol benih F3. Dengan demikian telah tersedia masing-masing 7 macam
populasi dari dua set persilangan untuk dievaluasi sifat ketahanannya terhadap penyakit
bulai.
Pengujian Karakter Ketahanan terhadap P. maydis
Masing-masing set persilangan diuji karakter ketahanannya terhadap P. maydis
dengan teknik inokulasi pada tanaman baris penyebar dan pada genotip uji. Saat tanaman
baris penyebar >80% terinfeksi bulai, masing-masing set persilangan ditanam sesuai
dengan rancangan acak kelompok (RAK), dua ulangan (Petersen, 1994), kemudian 5 hari
setelah kecambah muncul dipermukaan tanah, genotip uji disemprot dengan konidia bulai
dan 3 hari setelah penyemprotan pertama dulangi lagi dengan cara yang sama dengan
penyemprotan sebelumnya. Faktor pertama adalah dua set persilangan dan faktor kedua
adalah 7 macam populasi uji. Evaluasi dilakukan dengan menghitung persentase tanaman
terinfeksi per genotip, sedangkan metode penyiapan inokulum sampai evaluasi tingkat
ketahanannya mengikuti metode yang dilakukan oleh Azrai et al. (2000). Tata letak
percobaan di lapangan disajikan pada Lampiran 3.
PengamatanPengamatan dilakukan terhadap jumlah tanaman yang tumbuh dan terinfeksi
konidia bulai pada tiap genotip yang diuji. Waktu pengamatan yaitu saat tanaman
berumur 14, 21, 28, 35 dan 42 hari setelah tanam (hst). Data yang diperoleh merupakan
data komulatif dari pengamatan setiap pengamatan tersebut, kemudian dikonversi ke
dalam persentase tanaman terinfeksi (P) patogen P. maydis dengan menggunakan rumus :
ba
P x 100%

24
keterangan :P = persentase tanaman terinfeksi penyakit bulaia = jumlah kumulatif tanaman terinfeksi penyakit bulaib = jumlah tanaman tumbuh
Pengamatan tanaman terinfeksi dimulai pada umur 14 hst karena pada umur tersebut
biasanya penyakit bulai mulai menular pada daun tanaman jagung dan setelah berumur 42
hst penularannya sudah jarang terjadi.
Analisis Data
Analisis Ragam
Analisis ragam meliputi ragam genetik per generasi dari masing-masing set
persilangan. Estimasi ragam genetik dan fenotip dianalisis berdasarkan nilai kuadrat
tengah genotip (M2), nilai tengah galat (M1), dan ulangan (r), dengan persamaan sebagai
berikut, (Bernardo, 2002):
2g =
rMM 12
2e = M12f = 2
g + 2e
Luas sempitnya nilai keragaman genetik suatu karakter ditentukan berdasarkan
variabilitasgenetik ( 2g ) dan standar deviasi variabilitasgenetik ( 2
g ) menurut Anderson
dan Brancoff (1952), dikutip Wahdah et al. (1995) sebagai berikut :
2g
=
22
2 21
22
2 dbgalatM
dbgenotipeM
r
keragaman genetik luas jika 2g >2 2
g dan sempit jika 2
g 2 2g
Analisis Data Sebaran Frekuensi
Untuk mempelajari pola sebaran frekuensi terhadap pola distribusi berlanjut,
dilakukan dengan uji normalitas berdasarkan aturan Sturgers (Siregar, 2004):
1
22
i i
ii
eef
keterangan :fi = jumlah fenotip ke i menurut hasil pengamatanei = jumlah fenotip ke-i yang diharapkan

25
Kriteria keputusan sesuai dengan hipotesis yakni jika nilai peluang (p-v) > 0.05 maka data
dinyatakan berdistribusi normal.
Uji Kesesuaian Model Genetik
Untuk menentukan model genetik yang sesuai terhadap sifat ketahanan dari kedua
set persilangan, dilakukan analisis rata-rata generasi. Model genetik aditif dominan yang
sesuai untuk pola pewarisan kuantitatif, digunakan untuk menduga pengaruh gen-gen
yang mengendalikan karakter ketahanan untuk kedua genotip yang digunakan.
Selanjutnya model aditif dominan diuji untuk menentukan kesesuaiannya dengan uji t
(Hill et al., 1998). Jika hasil uji skala menunjukkan adanya pengaruh interaksi antar
lokus, maka model interaksi ditentukan kesesuaiannya dengan uji skala gabungan (Joint
Scalling Test) dengan menggunakan seluruh generasi secara bersama-sama (Mather dan
Jinks, 1982).
Enam parameter genetik dari model yang menyertakan pengaruh interaksi adalah
m = pengaruh rata-rata generasi, [d] = pengaruh aditif, [h] = pengaruh dominan, [I] =
pengaruh interaksi aditif x aditif, [j] = pengaruh interaksi aditif x dominan, dan [l] =
pengaruh interaksi dominan x dominan. Jadi model genetik yang menyertakan pengaruh
interaksi adalah [m][d][h][I]; [m][d][h][j]; [m][d][h][l]; [m][d][h][i][j]; [m][d][h][i][l] dan
[m][d][h][i][j][l]. Setiap model diuji kebaikan suainya dengan 2 terboboti. Koefisien dari
parameter genetik yang digunakan dalam model lengkap disajikan pada Tabel 1.
Tabel 1. Koefisien parameter genetik yang digunakan dalam analisis rata-rata generasi.
Parameter GenetikGenerasi m [d] [h] [i] [j] [l]
P1 1 1 0 1 0 0P2 1 -1 0 1 0 0F1 1 0 1 0 0 1F2 1 0 ½ 0 0 ¼F3 1 0 ¼ 0 0 1/16
BCP1 1 ½ ½ ¼ ¼ ¼BCP2 1 - ½ ½ ¼ - ¼ ¼
Analisis selanjutnya adalah pendugaan komponen ragam yang ditentukan sesuai
dengan persamaan menurut Kearsey dan Pooni, (1996) sebagai berikut :
VP1 = E1
VP2 = E2
VF1 = E3
VF2 = E4

26
VF3 = VA + VD +¼VE2 + ½VE3
VBCP1 = ½VA + VD –½VI + ½VE1 + ½VE3
VBCP2 = ½VA + VD + ½ VI + ½VE2 + ½VE3
VP1 , VP2 , VF1 , VF2, VF3, VBCP1 , VBCP2 masing-masing adalah ragam P1, P2, F1, F2,
F3, BCP1 dan BCP2. Awal pendugaan parameter dilakukan dengan menduga nilai E = (VP1
+ VP2 + VF1 + VF2 )/4. Selanjutnya Nilai E disubstitusikan ke persamaan, sehingga
diperoleh nilai VA, VD dan VF. Nilai VE adalah jumlah ragam lingkungan, VA adalah
jumlah ragam aditif, VD adalah jumlah ragam dominan, dan VI adalah jumlah ragam
interaksi aditif dan dominan.
Parameter yang diduga adalah :
a. Heritabilitas arti sempit (narrow sense heritability):
h2ns = VA
VA + VD + E
b. Heritabilitas arti luas (broad sense heritability):
h2bs = VA+ VD
VA + VD + E
HASIL DAN PEMBAHASAN
Keragaman Genetik
Hasil analisis ragam genetik ketahanan dari set persilangan MR10 x CML161 dan
Nei9008 x CML161 terhadap P. maydis disajikan pada Tabel 2. Kedua tetua dari masing-
masing set persilangan memperlihatkan reaksi ketahanan yang sangat berbeda. Galur
MR10 dan Nei9008 sebagai tetua donor gen resisten memperlihatkan reaksi yang sangat
resisten, sedangkan galur CML161 sebagai tetua donor gen opaque-2 (gen pengendali
lisin dan triptofan tinggi) sangat rentan terhadap patogen P. maydis. Perbedaan karakter
ketahanan penyakit bulai yang ekstrim antara kedua tetua sangat bermanfaat untuk
melihat frekuensi sebaran karakter kuantitatif ketahanan genotip terhadap patogen
penyakit bulai yang terekspresi pada progeninya (Namkong, 1979; Hoisington dan Coe,
1990; Prasanna, 2002).

27
Nilai tengah antar generasi dari set persilangan yang sama maupun dari set
persilangan yang berbeda memperlihatkan reaksi ketahanan terhadap penyakit bulai yang
beragam. Progeni Nei9008 x CML161 lebih resisten dibandingkan dengan progeni MR10
x CML161. Hal ini diduga karena pada pengujian ini, Nei9008 sangat resisten dan bahkan
tidak terinfeksi bulai sama sekali. Dari beberapa pengujian sebelumnya, Nei9008 sangat
resisten bulai di Sulawesi Selatan, Bogor, dan Lampung (Kasim et al., 2004). Selain
resisten terhadap P. maydis, Nei9008 juga resisten terhadap P. zeae dan P. philippinensis
(Ruswandi, 2002 dan Grudloyma et al, 2004).
Tabel 2. Nilai tengah, keragaman genetik, dan heritabiltas persentasepenularan terhadap P. maydis pada set persilangan galur jagung
Set PersilanganMR10 x CML161 Nei9008 x CML161
Gene-rasi
Rerata± SD
Interval(%)
2g
2 x2g
Rerata± SD
Interval(%)
2g
2 x2g
P1 2.2 ± 0.2tn 1.3 - 3 0.1 0.4 0.0 ± 0.0 tn 0.0–0.0 - -
P2 99.6 ± 0.6 tn 95.7–100 0 2.3 99.0 ± 0.3 tn 98.0–100 0 0.5
F1 60.0 ± 2.0 tn 36.4–70.8 35.1 53.5 28.6 ± 1.3 tn 21.0 - 39.1 6.4 17.2
F2 59.0 ± 1.5 tn 47.4–67.0 1.6 19.4 35.1 ± 1.4 tn 25.2 - 47.9 12.5 22.9
F3 54.2 ± 0.8** 6.3–100 324.8 101.9 41.8 ± 0.6** 4.2–85.7 249.5 76.3
BC1P1 32.5 ± 1.0 tn 26.1–41.2 3.9 10.0 23.9 ± 0.9 tn 20.0 - 31.8 4.8 8.7
BC1P 2 78.2 ± 1.7 tn 68.6–88.8 7.0 28.3 56.7 ± 1.3 tn 50.3–64.6 3.0 15.2
Keterangan: Data diolah berdasarkan persentase tanaman yang terinfeksi
2g = Keragaman genetik;
2g = standar deviasi keragaman genetik; Hbs = Heritabilitas dalam arti
luas; - = data tidak dianalisis karena tidak ada infeksi bulai
Nilai duga ragam genetik yang disajikan pada Tabel 2, menunjukkan bahwa
variabilitas genetik dari setiap generasi pada kedua set persilangan menurut kriteria
Anderson dan Brancoff (1952) tergolong sempit, kecuali pada generasi F3 yang tergolong
luas. Hal ini menunjukkan bahwa segregasi puncak terjadi pada generasi F3. Kejadian ini
disebabkan karena data yang digunakan merupakan data persentase antar famili, bukan
berupa skoring terhadap individu tanaman, sehingga secara teoritis variabilitas genetik
yang timbul pada tetua, F1, generasi silang balik, dan F2 adalah keragaman antara individu
dalam famili yang sama yang ditimbulkan oleh ragam lingkungan. Hasil penelitian ini
mendukung hasil penelitian Takdir (2003) yang melaporkan bahwa tidak ditemukan

28
0
5
10
15
20
25
30
35
0
Fre
kue
nsi MR10
Nei
adanya keragaman genetik yang nyata antar famili pada generasi P1, P2, F1, F2, BC1P1,
dan BC1P2. Variabiliatas genetik dapat terjadi karena adanya gen-gen yang bersegregasi
dan berinteraksi dengan gen pada generasi tersebut sehingga tingkat heterosigositasnya
tinggi (Crowder, 1988).
Sebaran Frekuensi Generasi F3
Uji normalitas untuk pola sebaran frekuensi terhadap pola distribusi berlanjut
tingkat penularan penyakit bulai menurut aturan Sturgers (Siregar, 2004) disajikan pada
Tabel 3. Sebaran frekuensi generasi F3 dari kedua set persilangan disajikan pada Gambar
6. Tabel 3 menunjukkan bahwa persentase penularan terhadap P. maydis pada generasi F3
dari kedua set persilangan menyebar normal
Tabel 3. Uji normalitas persentase penularan terhadap P. maydis pada generasi F3
set persilangan galur jagung
Set PersilanganParameter
MR10 x CML161 Nei9008 x CML161
Rerata 54.2 41.8
Simpangan baku (S) 19.0 17.0
χ2 8.2tn 7.4 tn
p-v 0.3 0.4Keterangan: Data diolah berdasarkan persentase tanaman yang terinfeksi; t
n = tidak nyata; Nilai chi-square0.05: db 7 = 14.07; Nilai chi-square0.01: db 7 = 12.02; p-v =nilai peluang (p-v > 0.05 = sesuai dengan hipotesis)
MR10Nei9008
CML161
Gambar 6. Sebaran frekuensi penularan terhadap P. maydis generasi F3 pada setpersilangan MR10 x CML161 dan Nei9008
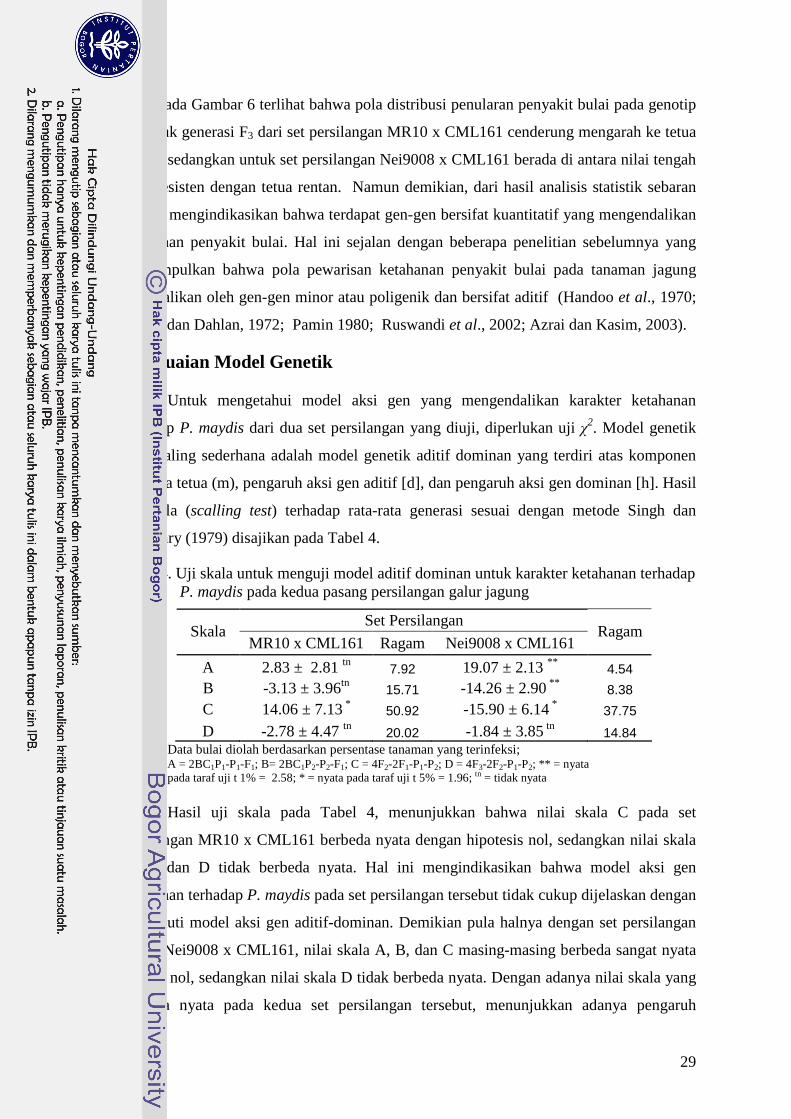
29
Pada Gambar 6 terlihat bahwa pola distribusi penularan penyakit bulai pada genotip
uji untuk generasi F3 dari set persilangan MR10 x CML161 cenderung mengarah ke tetua
rentan, sedangkan untuk set persilangan Nei9008 x CML161 berada di antara nilai tengah
tetua resisten dengan tetua rentan. Namun demikian, dari hasil analisis statistik sebaran
normal mengindikasikan bahwa terdapat gen-gen bersifat kuantitatif yang mengendalikan
ketahanan penyakit bulai. Hal ini sejalan dengan beberapa penelitian sebelumnya yang
menyimpulkan bahwa pola pewarisan ketahanan penyakit bulai pada tanaman jagung
dikendalikan oleh gen-gen minor atau poligenik dan bersifat aditif (Handoo et al., 1970;
Hakim dan Dahlan, 1972; Pamin 1980; Ruswandi et al., 2002; Azrai dan Kasim, 2003).
Kesesuaian Model Genetik
Untuk mengetahui model aksi gen yang mengendalikan karakter ketahanan
terhadap P. maydis dari dua set persilangan yang diuji, diperlukan uji χ2. Model genetik
yang paling sederhana adalah model genetik aditif dominan yang terdiri atas komponen
rata-rata tetua (m), pengaruh aksi gen aditif [d], dan pengaruh aksi gen dominan [h]. Hasil
uji skala (scalling test) terhadap rata-rata generasi sesuai dengan metode Singh dan
Chaudary (1979) disajikan pada Tabel 4.
Tabel 4. Uji skala untuk menguji model aditif dominan untuk karakter ketahanan terhadapP. maydis pada kedua pasang persilangan galur jagung
Set PersilanganSkala
MR10 x CML161 Ragam Nei9008 x CML161Ragam
A 2.83 ± 2.81 tn7.92 19.07 ± 2.13 **
4.54B -3.13 ± 3.96tn
15.71 -14.26 ± 2.90 **8.38
C 14.06 ± 7.13 *50.92 -15.90 ± 6.14 *
37.75
D -2.78 ± 4.47 tn20.02 -1.84 ± 3.85 tn
14.84Data bulai diolah berdasarkan persentase tanaman yang terinfeksi;A = 2BC1P1-P1-F1; B= 2BC1P2-P2-F1; C = 4F2-2F1-P1-P2; D = 4F3-2F2-P1-P2; ** = nyatapada taraf uji t 1% = 2.58; * = nyata pada taraf uji t 5% = 1.96; tn = tidak nyata
Hasil uji skala pada Tabel 4, menunjukkan bahwa nilai skala C pada set
persilangan MR10 x CML161 berbeda nyata dengan hipotesis nol, sedangkan nilai skala
A, B, dan D tidak berbeda nyata. Hal ini mengindikasikan bahwa model aksi gen
ketahanan terhadap P. maydis pada set persilangan tersebut tidak cukup dijelaskan dengan
mengikuti model aksi gen aditif-dominan. Demikian pula halnya dengan set persilangan
antara Nei9008 x CML161, nilai skala A, B, dan C masing-masing berbeda sangat nyata
dengan nol, sedangkan nilai skala D tidak berbeda nyata. Dengan adanya nilai skala yang
berbeda nyata pada kedua set persilangan tersebut, menunjukkan adanya pengaruh

30
interaksi antar lokus (non alelik) sehingga untuk menguji kesesuaian model interaksinya
diperlukan uji skala gabungan dengan menggunakan seluruh generasi secara bersama-
sama (Mather dan Jinks, 1982). Hasil uji skala gabungan untuk karakter ketahanan
terhadap P. maydis untuk kedua set persilangan yang digunakan disajikan pada Tabel 5.
Tabel 5. Uji χ2 dua persilangan galur jagung menggunakan beberapa model genetik
Persilangan Model genetik χ2
MR10 x CML161 m [d] 101.62**
m [d] [h] 90.85**
m [d] [h] [i] 7.49tn
m [d] [h] [j] 7.77 tn
m [d] [h] [l] 8.14*
m [d] [h] [i] [j] 4.93 tn
m [d] [h] [i] [l] 7.38 *
m [d] [h] [j] [l] 6.11*
m [d] [h] [i] [j] [l] 11.65*
Nei9008 x CML161 m [d] 355.66 **
m [d] [h] 138.26 **
m [d] [h] [i] 5.06 tn
m [d] [h] [j] 14.03*
m [d] [h] [l] 102.78 **
m [d] [h] [i] [j] 2.40 tn
m [d] [h] [i] [l] 0.34 tn
m [d] [h] [j] [l] 10.71 *
m [d] [h] [i] [j] [l] 0.01tn
Data bulai diolah berdasarkan persentase tanaman yang terinfeksi;** = nyata pada taraf uji chi-square 1%; * = nyata pada taraf uji chi-square 5%
tn = tidak nyata; Nilai chi-square0.05: db 5 = 11.07; db 4 = 9.49; db 3 = 7.82;db 2 = 5.99; db 1 = 3.84
Beberapa model genetik memperlihatkan nilai kebaikan suai yang cukup kontras
antara kedua set persilangan pada generasi yang sama. Dari hasil analisis χ2 diketahui
bahwa penyebab perbedaan tersebut disebabkan karena adanya penyimpangan nilai yang
cukup besar antara nilai pengamatan dengan nilai duganya sehingga menimbulkan selisih
nilai yang tinggi.
Dari uji kebaikan suai untuk set persilangan MR10 x CML161 diperoleh tiga
model genetik yang sesuai yaitu: m [d] [h] [i]; m [d] [h] [j]; m [d] [h] [i] [j]. Hal ini
menunjukkan bahwa komponen genetik karakter ketahanan terhadap P. maydis untuk set
persilangan tersebut terdiri atas gen aditif, dominan, interaksi aditif x aditif, dan interaksi
aditif x dominan. Model aksi gen yang sesuai dengan set persilangan Nei9008 x CML161
yaitu: m [d] [h] [i]; m [d] [h] [i] [j]; m [d] [h] [i] [l], dan m [d] [h] [i] [j] [l]. Model aksi
gen tersebut menunjukkan bahwa aksi gen aditf, dominan, interaksi aditif x aditif,

31
interaksi aditif x dominan, dan interaksi dominan x dominan berkontribusi untuk
ketahanan genetik pada set persilangan Nei9008 x CML161 terhadap P. maydis.
Untuk mengetahui komponen genetik yang berkontribusi nyata terhadap karakter
ketahanan terhadap P. maydis dari kedua set yang digunakan, dilakukan uji t. Uji t untuk
komponen dari model-model genetik yang digunakan adalah model genetik yang
mempunyai komponen yang paling lengkap seperti yang disajikan pada Tabel 6.
Tabel 6. Komponen genetik dan galat baku dari model genetik yang sesuai pada uji X2
untuk karakter ketahanan terhadap P. maydis pada dua set persilangan galurjagung.
Persilangan Model yang sesuai Komponen genetikMR10 x CML161 m [d] [h] [i] [j] m = 53.07 ± 1.26
[d] = -48.67 ± 0.35**
[h] = 7.12 ± 2.14**
[i] = -2.20 ± 1.11*
[j] = 5.22 ± 3.26tn
Nei9008 x CML161 m [d] [h] [i] [j] [l] m = 50.94 ± 3.30[d] = -37.37 ± 9.07**
[h] = -41.06 ± 15.03**
[i] = 20.72 ± 6.33 **
[j] = -10.96 ± 18.76 tn
[l] = 18.76 ± 12.11 tn
Data bulai diolah berdasarkan persentase tanaman yang terinfeksi;** = nyata pada taraf uji t 1% = 2.58; * = nyata pada taraf uji t 5% = 1.96; tn = tidak nyata
Hasil uji t pada Tabel 6, menunjukkan bahwa gen-gen aditif [d] dan dominan [h]
berkontribusi sangat nyata pada kedua set persilangan yang digunakan. Selain itu,
komponen interaksi gen aditif x aditif [i] masing-masing sangat nyata dan nyata pada set
persilangan MR10 x CML161 dan Nei9008 x CML161. Nilai komponen genetik aditif
yang bertanda sama dengan interaksi aditif x aditif pada set persilangan MR10 x CML161
menunjukkan adanya interaksi gen yang bersifat epistasis komplomenter, sedangkan pada
set persilangan Nei9008 x CML161 komponen genetik aditif bertanda sama dengan
interaksi aditif x aditif menunjukkan adanya interaksi gen yang bersifat epistasis duplikat
(Hill et al, 1998)
Kontribusi gen-gen aditif untuk karakter ketahanan terhadap P. maydis lebih
tinggi daripada gen-gen dominan pada set persilangan MR10 x CML161 dan sebaliknya
pada set persilangan Nei9008 x CML161 relatif seimbang (Tabel 6). Selain itu, interaksi
gen aditif x aditif masing-masing berkontribusi nyata dan sangat nyata pada set
persilangan MR10 x CML161 dan set persilangan Nei9008 x CML161. Kontribusi yang
nyata dari interaksi gen aditif x aditif pada set persilangan tersebut dapat meningkatkan

32
variasi antara famili generasi F3 dan ini dapat difiksasi (Singleton, 1967). Ragam aditif
mencerminkan nilai pewarisan (breeding value) yang merupakan penyebab utama
kemiripan antara famili sehingga menjadi penentu utama dalam penurunan suatu sifat,
sedangkan ragam dominan dapat dimanfatkan jika varietas hibrida yang menjadi tujuan
program pemuliaan (Hallauer dan Miranda, 1981).
Parameter Genetik Gabungan Generasi
Nilai parameter genetik dari kedua set persilangan disajikan pada Tabel 7. Pada
set persilangan MR10 x CML161 memiliki nilai ragam genetik (dominan dan aditif),
ragam lingkungan dan fenotip serta ragam interaksi genetik x lingkungan yang lebih luas
dibandingkan dengan ragam pada set persilangan Nei9008 x CML161.
Kedua set persilangan yang digunakan juga mempunyai nilai duga heritabilitas
dalam arti luas yang tergolong tinggi berdasarkan kriteria yang digunakan oleh
McWhirter (1979) yaitu masing-masing sebesar 0.78 untuk set persilangan MR10 x
CML161 dan 0.76 untuk set persilangan Nei9008 x CML161. Berbeda halnya dengan
nilai duga heritabilitas dalam arti sempit, dimana kedua set persilangan menunjukkan
nilai heritabilitas yang tergolong sedang yaitu masing-masing sebesar 0.46 untuk set
persilangan MR10 x CML161 dan 0.47 untuk set persilangan Nei9008 x CML161. Nilai
duga heritabilitas merupakan suatu ukuran sampai sejauh mana fenotip yang tampak
sebagai refleksi genotip, atau hubungan antara keragaman genetik dengan keragaman
fenotipiknya (Fehr, 1987). Nilai duga heritabilitas yang tinggi menunjukkan bahwa faktor
genetik lebih berperan dari faktor lingkungan. Begitu pula sebaliknya bila nilai duga
heritabilitas rendah, menunjukkan bahwa faktor lingkungan lebih berperan daripada
faktor genetik (Sjamsudin, 1990).
Nilai heritabilitas dalam arti sempit yang tergolong sedang pada kedua set
persilangan yang digunakan menunjukkan bahwa faktor lingkungan cukup berpengaruh
pada pewarisan ketahanan terhadap P. maydis sehingga seleksi akan lebih efektif jika
dilakukan pada generasi lebih lanjut. Dengan keberadaan interaksi gen yang bersifat
epistasis duplikat pada set persilangan Nei9008 x CML161, maka disarankan proses
seleksi dilakukan secara hati-hati dan tidak dilakukan pada generasi awal yang masih
bersegregasi (Stoskpf, 1993).

33
Tabel 7. Parameter genetik untuk karakter ketahanan pada dua pasangpersilangan galur jagung terhadap P. maydis
Set PersilanganParameterMR10 x CML161 Nei9008 x CML161
VA 65.12 37.40VD 26.03 14.38VE 31.63 18.82VP 122.78 70.60VI 19.41 8.51Hbs 0.78 0.76Hns 0.46 0.47Hns/Hbs (%) 58.90 62.03
Data bulai diolah berdasarkan persentase tanaman yang terinfeksi;VA = jumlah ragam aditif; VD = jumlah ragam dominan; VE = jumlah ragam lingkungan;VI = jumlah ragam interaksi genetik x lingkungan; Vp = jumlah ragam fenotip;Hbs = Heritabilitas dalam arti luas; Hns = Heritabilitas dalam arti sempit
Efek dominansi berdasarkan rasio heritabilitas disajikan pada Tabel 7. Menurut
Moeljopawiro (1986), nisbah antara nilai heritabilitas dalam arti sempit terhadap nilai
heritabilitas dalam arti luas menggambarkan besarnya porsi ragam aditif terhadap total
ragam genetik. Karakter yang mempunyai nisbah Hns/Hbs < 50%, berarti karakter tersebut
dikendalikan oleh aksi gen dominan, dan bila mempunyai nilai nisbah Hns/Hbs > 50%,
berarti karakter tersebut dikendalikan oleh aksi gen aditif. Berdasarkan kriteria tersebut,
aksi gen-gen aditif mempunyai porsi yang lebih besar dalam mengendalikan karakter
ketahanan terhadap P. maydis pada kedua set persilangan yang digunakan. Oleh karena
kedua set persilangan yang digunakan memiliki ragam aditif yang lebih tinggi daripada
ragam dominan, maka peluang pewarisan karakter ketahanan terhadap P. maydis dari
kedua persilangan tersebut cukup tinggi sehingga sangat bermanfaat dalam proses seleksi
untuk mengintrogresikan gen resisten ke galur jagung rentan terhadap penyakit tersebut,
namun punya nilai ekonomis tinggi, seperti pada galur-galur QPM .
KESIMPULAN
1. Estimasi variabilitas genetik dari tiap generasi pada kedua set persilangan
tergolong sempit, kecuali generasi F3 yang tergolong luas.
2. Nilai duga heritabilitas dalam arti sempit dari tiap generasi pada kedua set
persilangan tergolong rendah sampai sedang, kecuali generasi F3 yang tergolong
luas. Dari hasil analisis gabungan rata-rata generasi pada kedua pasang
persilangan diperoleh nilai duga heritabilitas dalam arti luas tergolong tinggi,
sedangkan dalam arti sempit tergolong sedang.

34
3. Pola distribusi penularan konidia patogen bulai genotip uji pada generasi F3 dari
set persilangan MR10 x CML161 cenderung mengarah ke tetua rentan, sedangkan
dari set persilangan Nei9008 x CML161 berada di antara nilai tengah tetua
resisten dengan tetua rentan, namun keduanya berdistribusi normal. Terdapat gen-
gen yang bersifat kuantitatif dalam mengontrol ketahanan penyakit bulai.
4. Model genetik karakter ketahanan terhadap P. maydis yang sesuai untuk set
persilangan MR10 x CML161 adalah m [d] [h] [i] [j], sedangkan untuk set
persilangan Nei9008 x CML161 yaitu: m [d] [h] [i] [j] [l].
5. Aksi gen aditif dan dominan dari kedua set persilangan berkontribusi nyata untuk
karakter ketahanan terhadap P. maydis pada kedua set persilangan. Set MR10 x
CML161 diperankan oleh aksi gen aditif dominan dengan pengaruh interaksi non
alelik aditif komplementer epistasis dan non alelik aditif duplikat epistasis pada
set persilangan Nei9008 x CML161.