phylogeographic structure and genetic variation in formica ants
TRANSCRIPT

Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Science and Technology 912
Phylogeographic Structureand Genetic Variation
in Formica Ants
BY
ANNA GOROPASHNAYA
ACTA UNIVERSITATIS UPSALIENSISUPPSALA 2003

Dissertation to be publicly examined in Lindahl-salen, EBC, Uppsala University, on Saturday,December, 20, 2003 at 10:00, for the degree of Doctor of Philosophy. The examination will beconducted in English.
AbstractGoropashnaya, A. 2003. Phylogeographic Structure and Genetic Variation in Formica Ants.Acta Universitatis Upsaliensis. Comprehensive Summaries of Uppsala Dissertations from theFaculty of Science and Technology 912. 36 pp. Uppsala. ISBN 91-554-5818-1.
The aim of this thesis is to study phylogeny, species-wide phylogeography and geneticdiversity in Formica ants across Eurasia in connection with the history of biotic responses toQuaternary environmental changes.
The mitochondrial DNA phylogeny of Palaearctic Formica species supported the subgenericgrouping based on morphological similarity. The exception was that F. uralensis formed aseparate phylogenetic group. The mitochondrial DNA phylogeny of the F. rufa group showedthe division into three major phylogenetic groups: one with the species F. polyctena and F.rufa, one with F. aquilonia , F. lugubris and F. paralugubris , and the third one with F.pratensis.
West-east phylogeographic divisions were found in F. pratensis suggesting post-glacialcolonization of western Europe and a wide area from Sweden to the Baikal Lake from separateforest refugia. In contrast, no phylogeographic divisions were detected in either F. lugubris orF. exsecta . Contraction of the distribution range to a single refugial area during the latePleistocene and the following population expansion could offer a general explanation for thelack of phylogeographic structure across most of Eurasia in these species.
Sympatrically distributed and ecologically similar species F. uralensis and F. candidashowed clear difference in the phylogeographic structure that reflected difference in theirvicariant history. Whereas no phylogeographic divisions were detected in F. uralensis acrossEurope, F. candida showed a well-supported phylogeographic division between the western,the central and the southern group.
In socially polymorphic F. cinerea , the overall level of intrapopulation microsatellitediversity was relatively high and differentiation among populations was low, indicating recenthistorical connections. The lack of correspondence between genetic affinities and geographiclocations of studied populations did not provide any evidence for differentiating betweenalternative hypotheses concerning the directions and sources of postglacial colonization ofFennoscandia.
Keywords: Formica ants, phylogeography, phylogeny, Pleistocene refugia, populationexpansion, social organization
Anna Goropashnaya, Department of Evolutionary Biology, Uppsala University, Norbyv. 18 D,SE-752 36 Uppsala, Sweden.
© Anna Goropashnaya 2003
ISSN 1104-232XISBN 91-554-5818-1URN:NBN:se:uu:diva-3803(http://urn.kb.se/resolve?urn=urn:nbn:se:uu:diva-3803)

A.Formica
b
A. V.Formica rufa
accepted
A. V.
Formica pratensis F. lugubris
A. V.Formica excecta
A.
Formica cinerea .

4
CONTENTS
Introduction 5
Pleistocene ice ages and their consequences 5
Formica ants 6
Genetic markers 7
Objectives 9
Results and Discussion 10
Phylogenetic relationships of Palaearctic Formica species 10
Limited phylogeographic structure across Eurasia in the ants Formica pratensis,
F. lugubris and F. exsecta 16
Mitochondrial DNA variation in Formica uralensis and F. candida 23
Genetic characteristics of northern European populations of Formica cinerea 26
Conclusions 29
Acknowledgments 31
References 32

5
INTRODUCTION
Pleistocene ice ages and their consequences
During the Pleistocene, the climate underwent dramatic fluctuations due to the orbital
eccentricity of the earth around the sun (cf. Bennett 1997). These climate oscillations were
expressed in ice ages that lasted from 10 to 100 kyr, and relatively short warm interglacials.
During the glacials, massive ice sheets partly covered the land, and vast continental areas
represented Arctic desert and tundra (Andersen & Borns 1997). As a consequence, all forest
biota could not survive in the changed environment but moved to more suitable habitats further
to the south or to the sheltered and moist valleys in mountain areas (Stewart & Lister 2001). The
last glaciation that started about 115 kyr before present (BP) was characterized by colder and
warmer periods with the last glacial maximum 22-18 kyr BP (Andersen & Borns 1997).
According to paleoecological data, the most of non-glaciated northern Eurasia during that stage
was covered by treeless vegetation (West 2000). Therefore, forest species were confined to small
areas with favorable conditions, i.e. refugia, where they could survive the glacials and then re-
colonize previously unsuitable habitats during interglacials. Recent paleoecological and genetic
studies suggest that refugia for boreal and temperate species occurred not only in Iberia, Italy
and the Balkans as previously suggested (Hewitt 1996) but also in Central and Northern Europe,
the southern Urals and Siberia, and on the coasts of the Azov and Black Seas (Lagercrantz &
Ryman 1990; Bilton et al. 1998; Tarasov et al. 2000; Schmitt & Seitz 2001; Jaarola & Searle
2002; Brunhoff et al. 2003; Haase et al. 2003;).
Past isolation in separate glacial refugia and following post-glacial colonization are reflected in
genes and gene pools of extant species (Hewitt 1996). Separation in different glacial refugia
generated intraspecific divergence, and the isolated gene pools were protected from mixing by
hybrid zones during interglacials. Throughout several glacial periods refugial separation became
reflected in the phylogeographic structure, i. e. significant association between the genealogical
relationship and geographic distribution of alleles. Since forest remained only in restricted areas
during the last glaciation, the refugial populations of many forest species were small. Therefore,
genetic drift was a strong factor in those populations and could lead to loss of genetic variation
within each refugium and genetic differentiation between different refugia in a short time. With
the advent of favorable conditions, forest fauna and flora spread over the continent resulting in
an admixture of different refugial populations and/or forming suture zones. Unlike

6
phylogeography based on relatively slow evolving insect mtDNA (2% per Myr; DeSalle et al.
1987), a population level analysis that takes into account not only the haplotype genealogy but
also differentiation in haplotype frequencies, can reveal refugial separation over shallow time
span of the last glaciation. The genetic signal of the past refugial separation is expected to be
stronger in species with limited dispersal abilities. Demographic history of populations is also
reflected in genes and gene pools (Rogers 1995; Kuhner et al. 1998).
While the refugial and colonization history has been studied by using genetic markers in a
number of boreal forest species in North America (cf. Arbogast &Kenagy 2001; Lessa et al.
2003), Eurasian species have received much less attention. Forest taxa studied to date across the
Palaearctic are mostly avian species. More phylogeographic studies are needed to elucidate the
post-glacial history of Eurasian forest biota. Although the routes of the re-colonization may have
been unique for different species, evidence of shared refugia has emerged from molecular data
on various taxa (Taberlet et al. 1998; Hewitt 1999).
Formica ants
Formica ants represent a large group of soil insects that occur mainly in the Holarctic. There are
about 150 species of this genus a bigger part of which is distributed in the Nearctic and a smaller
part in the Palaearctic. Many species are widespread and abundant, and they play an important
role in ecosystems being active predators, tending aphids and improving soil composition. Most
taxonomists have distinguished four subgenera in the European Formica species (e.g. Dlussky
1967): Raptiformica, Coptoformica, Serviformica, and Formica s. str. The subgeneric
subdivision of the Formica ants based on morphology has been questioned and remained
unclear, and an earlier study based on allozymes of 13 species did not help to solve the question.
Taxonomy of the F. rufa group ants that belong to Formica s. str. has also been unstable mainly
because of their morphological similarity and ability to hybridize and to form mixed colonies (cf.
Czechowski 1996). Since ants of this group are strictly associated with forest, estimating the
divergence time among the species can reveal the possible speciation effect of the Pleistocene
environmental changes.
Formica species demonstrate a great diversity of complex behavior and social organization. The
subgenus Raptiformica includes slave-making species, and the subgenera Formica s. str. and
Coptoformica use temporary parasitism as a mode of founding new colonies, while the species of

7
the subgenus Serviformica are used as slaves. The organization of colonies ranges from simple
monogynous (single- queen) societies to huge supercolonies, i.e. networks of connected
polygynous (multiple- queen) nests (Cherix et al. 1991; Chapuisat et al. 1997).
Variation of social characteristics has made Formica ants useful for ecological, behavioral and
evolutionary studies and good model objects for testing different theoretical implications of the
kin selection theory (e.g. Pamilo & Seppä 1994; Sundström et al. 2003). The phylogeny based on
DNA sequences provides a necessary evolutionary context in which the evolution of various
traits can be inferred and compared. Since most of the Formica species are confined to the forest
zone, comparative phylogeographic studies of selected species might shed light to the post-
glacial history of Eurasian forest biota.
Some of the Formica species have typically monogynous colonies, others build polygynous
colonies, and there are also species that show intraspecific social polymorphism and their
colonies are monogynous in some populations and polygynous in others. The colony type is
connected to dispersal of individuals at a local scale. Females of Formica species produced in
monogynous nests disperse by flight and found new colonies independently or through temporal
parasitism, while females from polygynous nests can remain in the natal nest or establish new
colonies with the help of workers in close neighborhood of the parental nest (Rosengren &
Pamilo 1983; Keller 1991). The latter strategy induces genetic differentiation between
geographically distant nests and may lead to population viscosity (Hamilton 1964). Limited
dispersal connected to social organization or habitat fragmentation increases genetic
differentiation among populations, particularly after dramatic declines in population size
(bottlenecks) caused by unfavorable environmental changes.
Genetic markers
Mitochondrial DNA (mtDNA) has been traditionally used for phylogenetic and phylogeographic
studies in animals. Its advantages such as maternal inheritance, lack of recombination, and
simple organization have made it especially useful and informative. Complete mtDNA genome
was sequenced for several species that makes it possible to study large variety of other species
from related taxa. Universal mtDNA primers were designed for different groups of animals,
particularly for ants on the basis of the sequence of the complete mtDNA genome of the
honeybee (Crozier & Crozier 1993). Specific primers can be designed with computer programs

8
using sequence of a target fragment. Since mtDNA is maternally inherited it has a smaller
effective population size than nuclear DNA which causes faster population differentiation in
mtDNA than in nuclear markers. In Hymenoptera, females (queens and workers) are diploid and
males are haploid (but see Pamilo et al. 1994), therefore the mtDNA genome has an effective
size three times smaller than the nuclear one if the sex ratio in population is 1:1 If the sex ratio is
biased then the effective population size for the nuclear genes is affected by the biased sex ratio.
The most commonly used nuclear markers nowadays are microsatellites, short sequences made
up of a simple sequence motif that is tandemly repeated (Goldstein & Schlötterer 1999).
Microsatellites show high levels of polymorphism and mutation rates, they are codominant and
widespread in most genomes. However, the mode of mutation for any given microsatellite locus
is generally unknown (Estoup & Cornuet 1998) that hampers testing population differentiation
with the use of estimates based on the variance in allele sizes (R ST). Moreover, the genetic
divergence between populations can be underestimated by microsatellites because of their high
degree of polymorphism (Hedrick 1999) and, to a lesser degree, homoplasy (Jarne & Lagoda
1996).
Since every genetic marker has advantages and disadvantages, the optimal choice is to use at
least two independent markers. In the present work, sequences of 1.5 and 2 kb mtDNA
fragments including cytochrome b were analyzed in phylogenetic and phylogeographic studies,
and several microsatellite loci were used to reveal the type of social organization in populations
and differentiation within and between localities.

9
OBJECTIVES
The study had the following objectives:
Reconstruct the phylogenetic relationships among Formica species and to evaluate the
extent of congruence between the phylogeny based on mtDNA sequences and
classifications based on morphological characters and allozymes.
Clarify the taxonomy of the Formica rufa group and to assess preliminarily the
phylogeographic structure within each morphologically defined species.
Infer evolution of social organization in the socially polymorphic Formica rufa group
and its possible effect on the speciation rate.
Assess continental phylogeography in ant species Formica pratensis, F. lugubris and
F. exsecta with Eurasian distribution, in order to reveal signs of possible vicariant
separation and routes of postglacial colonization from separate boreal forest refugia.
Examine genetic footprints of the demographic history in Formica pratensis, F. lugubris
and F. exsecta in order to reveal signs of possible demographic expansion from limited
refugial sources.
Test the correspondence between haplotype genealogical relationships and geographic
distribution of haplotypes in Formica uralensis and F. candida.
Estimate genetic differentiation in northern European populations of Formica cinerea in
order to reveal its possible colonization routes in Fennoscandia and to evaluate the effect
of isolation on the species’ genetic diversity in isolated populations.

10
RESULTS AND DISCUSSION
Phylogenetic relationships of Palaearctic Formica species
The previous allozyme study on 13 Palaearctic Formica species from all four subgenera (Pamilo
et al. 1979) agreed with the subgeneric division based on morphological and behavioral
characters with some exceptions. One of the exceptions was the topological position of
F. uralensis (subgenus Formica s. str.). This species was associated with Serviformica that
supported the subgeneric affiliation given by Dlussky (1967). In the present study, the
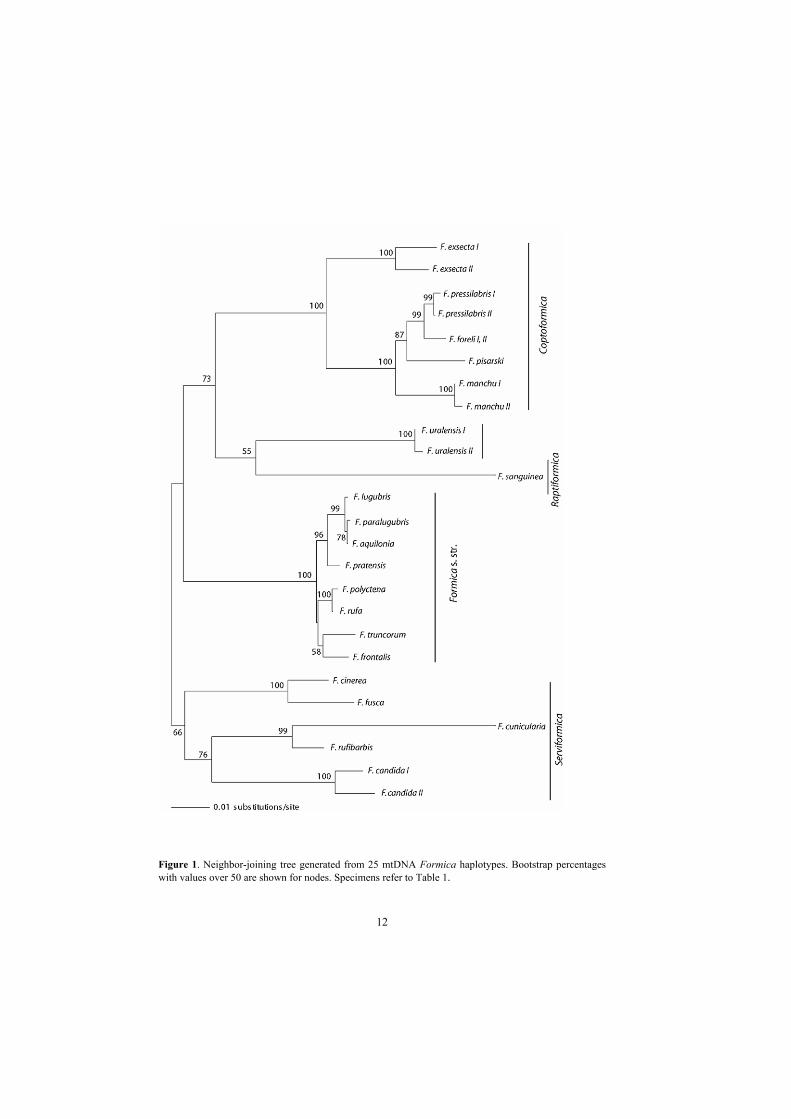
phylogenetic relationships of the Formica subgenera were examined using 25 mtDNA sequences
of 20 Eurasian species (Table 1). All sampled Formica species clustered according to the
subgeneric affiliation in the neighbor-joining (NJ) tree. The only species that did not belong to
any of the subgeneric clades and was loosely connected to F. sanguinea (subgenus Raptiformica)
was F. uralensis (Fig. 1). The independent position of F. uralensis in the phylogenetic tree
supported the results from the allozyme study (Pamilo et al. 1979) that this species represents a
separate phylogenetic lineage and might even be placed in a new subgenus. Notably, there have
been different opinions concerning the subgeneric affiliation of F. uralensis. One of the
arguments for not including this species in Serviformica but in Formica s. str. is the facultative
social parasitism of the females during nest founding, a common feature in all subgenera but
Serviformica. Furthermore, species of the latter subgenus do not build nest mounds, whereas
F. uralensis does.
In the Coptoformica clade, F. exsecta was the most genetically distant species from the others.
The subgenus Formica s. str. represented a very tight cluster of species with short branch
lengths. On the contrary, the Serviformica subgenus was highly diverged with possible
substructure, even though a reliable substructure is difficult to detect using only five Palaearctic
species of this subgenus with very many species.
One objective of the present study was to reconstruct the phylogeny of the subgenus Formica s.
str. Particularly, examination of phylogenetic relationships among closely related forest species
of the F. rufa group might clarify the taxonomy of this morphologically diverse group and
evaluate the possible speciation effect of the Pleistocene environmental changes.

11
Table 1. List of Formica species used in the phylogenetic study, their subgeneric groupings and sampling
localities.
Species and Groupings Locality
Subgenus Raptiformica
F. sanguinea Sweden
Subgenus Coptoformica
F. pressilabris-I Urals, Russia
F. pressilabris-II Urals, Russia
F. foreli-I Öland, Sweden
F. foreli-II Öland, Sweden
F. pisarkii Eastern Siberia, Russia
F. manchu-I Eastern Siberia, Russia
F. manchu-II Eastern Siberia, Russia
F. exsecta-I Germany
F. exsecta-II Tibet, China
Subgenus Serviformica
F. cinerea Sweden
F. fusca Sweden
F. candida-I Sweden
F. candida-II Kyrgyztan
F. cunicularia Western Siberia, Russia
F. rufibarbis Sweden
Subgenus Formica s. str.
F. paralugubris Switzerland
F. aquilonia Sweden
F. lugubris Switzerland
F. pratensis Finland
F. polyctena Urals, Russia
F. rufa Belgium
F. truncorum Sweden
F. frontalis Spain
F. uralensis-I Finland
F. uralensis-II Urals, Russia
12
Figure 1. Neighbor-joining tree generated from 25 mtDNA Formica haplotypes. Bootstrap percentages with values over 50 are shown for nodes. Specimens refer to Table 1.

13
Recent speciation in the Formica rufa group ants
According to the Pleistocene speciation hypothesis, separation in different glacial refugia
generated intraspecific divergence, and isolated gene pools were protected from mixing by
hybrid zones during interglacials leading to allopatric speciation (Hewitt 1996). The opposite
view is that Pleistocene environmental changes inhibited allopatric speciation by repeatedly
altering species distributions and thus prevented accumulation of evolutionary changes (Zink &
Slowinski 1995). The Pleistocene speciation hypothesis can be evaluated by reconstructing
phylogeny and estimating species divergence time, which is expected to be less than two million
years under this hypothesis.
The Formica rufa group includes several morphologically similar species of mound-building red
wood ants: F. rufa, F. polyctena, F. lugubris, F. paralugubris, F. aquilonia, and F. pratensis. A
total of 44 individuals including all six species of the Formica rufa group were sampled from
different localities in Eurasia over most of their distribution (Fig. 2, 3). One individual of F.
truncorum and two individuals of F. frontalis were used as outgroups representing the same
subgenus Formica s. str.
Figure 2. Map showing the sampling localities and species distribution of the Formica rufa group (Dlussky 1967).

14
Figure 3. Neighbor-joining phylogenetic tree of 30 mtDNA haplotypes of the Formica rufa group species. The tree is rooted using F. truncorum and two F. frontalis sequences. Numbers in parenthesis indicate localities for one specimen of a particular haplotype and refer to Fig. 2. Bootstrap percentages with values over 60 are shown for nodes. Monogyny is indicated on the tree as M, polygyny as P.
The mtDNA phylogeny shows that the six Formica rufa group species form a very tight group
compared to the outgroups (Fig. 3). The clade of F. polyctena / F. rufa (II) is basal within this
group in the rooted phylogenetic tree and F. pratensis (clade IB) is clearly distinct from all the
other species. The branching pattern does not support the suggestion based on morphological
differences that F. pratensis diverged before the separation of the other F. rufa group species
(Seifert, personal communication) and agrees better with the earlier allozyme results (Pamilo et
al. 1979) in associating F. pratensis with the species F. lugubris and F. aquilonia. The species
F. rufa and F. polyctena belong to one undivided monophyletic group and this implies recent

15
divergence and incomplete lineage sorting between them. The lack of hiatus in the mtDNA
phylogeny is consistent with the occurrence of frequent hybridization between F. rufa and
F. polyctena in nature (Seifert 1991; Czechowski 1996). Although F. rufa and F. polyctena are
not distinct as regards mtDNA, we do not suggest that their taxonomic status should be revised
as there are pronounced differences in morphological, social and population characteristics
(Collingwood 1979). Genotype studies of sympatric populations also suggested that they form
two separate gene pools (Gyllenstrand et al. in press). The species F. paralugubris that was only
recently described morphologically (Seifert 1996) is phylogenetically close to F. aquilonia. So
far F. paralugubris is known only from the Alps and the Jura mountains in Switzerland and
adjacent regions. This species is morphologically similar to F. lugubris, but the allozyme study
(Pamilo et al. 1992) and behavioral experiments based on workers’ discrimination against the
sexual pupae of an alien type (Rosengren et al. 1994) showed that F. lugubris and
F. paralugubris represent different gene pools. In Switzerland, all three species F. lugubris,
F. aquilonia and F. paralugubris are highly polygynous and differentiated at a set of allozyme
and microsatellite loci (Pamilo et al. 1992; Chapuisat et al. 1997). It is possible that
F. paralugubris has arisen as a result of hybridization between F. lugubris and F. aquilonia and
consequent isolation of a highly polygynous population for a long time. Thus the mtDNA
phylogeny generally supports the division of the Formica rufa group into distinct species
suggested on the morphological basis.
Despite morphological similarity, the F. rufa group species have different types of social
organization. Formica polyctena, F. aquilonia and F. paralugubris are obligatorily highly
polygynous species often forming large networks of interconnected nests (Crozier & Pamilo
1996, pp.114-115; Chapuisat et al. 1997). Formica rufa and F. pratensis are mainly monogynous
though polygynous nests have been recorded for both species. Formica lugubris is polygynous
on the British Isles and Switzerland and mainly monogynous in Ireland and Fennoscandia. The
distribution of the social types in the phylogenetic tree shows that the transition between
monogyny and a very high level of polygyny has taken place more than once during the
evolutionary time (Fig. 3). This result agrees with the general conclusion that polygyny has
multiple origins (Ward 1989; Ross & Carpenter 1991) and gives no strong phylogenetic evidence
supporting the importance of polygyny for speciation, except perhaps for F. aquilonia and
F. paralugubris both of which build large supercolonies and show little sequence variation.

16
The average net divergence estimate for the main phylogenetic groups IA, IB and II (Fig. 3) was
0.98 ± 0.15 %. Within the clade IA, the net divergence estimate between F. lugubris and
F. paralugubris / F. aquilonia was 0.20 ± 0.09%. Assuming the divergence rate of 2% per Myr
(DeSalle et al. 1987), the time of divergence among the main phylogenetic groups (IA, IB and II)
in the F. rufa group is about 490 thousand years (kyr) before present (BP), and between
F. lugubris and F. paralugubris / aquilonia about 100 kyr BP. Even though these time estimates
include uncertainty, they imply that speciation took place during the Pleistocene.
Despite the extensive geographic sampling, no phylogeographic structure was detected within
species by this preliminary study. The only association between the genealogies and geographic
distribution of the haplotypes was found in F. lugubris (Fig. 2, 3). The eastern group included
haplotypes (A-D) from two rather distant Siberian localities (13 and 14). The western group
included haplotypes (J-N) from Pyrenees (8), Switzerland (7) and Britain (5). Lack of
phylogeographic structure in F. pratensis could be due to the limited sampling size. A population
level study with larger sample sizes of F. pratensis and F. lugubris is required to reveal the
geographic structure and historical relationships among their populations.
Limited phylogeographic structure across Eurasia in the ants Formica pratensis,
F. lugubris and F. exsecta
Phylogeography
In the present study, mtDNA variation was examined in three ant species with a distribution
range covering the forest zone throughout Eurasia. Relatively high mtDNA variation was found
in closely related species F. pratensis and F. lugubris (see above) as well as in F. exsecta (see
e.g. Liautard & Keller 2001). This makes it possible to study mtDNA phylogeography and
population structure of these species in connection with biotic responses to Quaternary
environmental changes.
In total, 49 different haplotypes were found among 125 F. pratensis and F. lugubris ants (Fig. 4,
5). The majority of haplotypes, with a few exceptions, affiliated to their species clades. Three
haplotypes found in individuals morphologically regarded as F. pratensis from the Pyrenees
(h46, h47, h48; locality 1) belonged to the lugubris clade and clustered together with a haplotype
of F. lugubris (h49) from the same locality. Three individuals of F. pratensis from the Urals
(locality 9) also had haplotypes that belonged to the lugubris clade (Table 1). Two of these

17
individuals shared the haplotype h28 which was common in F. lugubris. One individual of
F. pratensis from the Urals had a unique haplotype (h27) that differed by one nucleotide
substitution from h28. Six other individuals of F. pratensis from the Urals had haplotypes that
clustered clearly in the pratensis clade (Fig. 5). These samples were carefully identified
morphologically and showed no hybrid traits, and the discordance between morphology and
mtDNA phylogeny most probably indicated old interspecific hybridization.
Figure 4. Map showing sampling localities of Formica pratensis and F. lugubris and their distribution. Both species occur throughout Eurasia. The distribution of F. pratensis coincides with the zone of mixed forest and steppe-forest, and the distribution of F. lugubris covers the taiga zone, European mountains and deciduous forest in England and Primorskiy Krai (Dlussky 1967).

18
Figure 5. Neighbor-joining tree of Formica pratensis and F. lugubris mtDNA sequences. Locality codes are in parentheses and refer to Fig. 4. Bootstrap percentages with values greater than 50 are shown on nodes. Haplotypes affiliated to a different species clades are indicated in bold. Haplotype h28 found in both species and four localities from the Pyrenees to the Urals is designated by an asterisk.
The only phylogeographic division found in these two species of red wood ants across Eurasia
was between the western (clade B) and the eastern (clade A) groups of F. pratensis. The amount
of divergence between the phylogeographic groups A and B suggests that these two lineages
were separated around 350 kyr BP. This time estimate is highly approximate but suggests
separation of the two phylogroups before the last glaciation (10 – 115 kyr), probably during one
of the previous two 100 kyr glacial periods (Andersen & Borns 1997). Haplotypes from the

19
pratensis group B were found only in the three western populations sampled, two of which,
Romania and Öland (localities 4 and 5), entirely consisted of haplotypes from this phylogroup.
This phylogeographic pattern suggests that two old lineages of F. pratensis survived glaciations
in separate refugia and colonized the continent during the Holocene via different routes. The
population from Romania (locality 4) is situated close to the glacial forest refugium in Hungary
revealed by the paleoecological evidence (Willis et al. 2000; Stewart & Lister 2001; Sumegi &
Krolopp 2002). The eastern phylogeographic group (clade A) of F. pratensis probably colonized
a vast area of Eurasia, ranging from Sweden to the Baikal Lake, from putative forest refugia that
were located according to the paleoecological evidence on the north east coast of the Azov and
Black Seas, the southern Ural, south Siberian mountains and Mongolia (Grichuk 1984; Efimik
1996; Tarasov et al. 2000). The samples from the westernmost locality (the Pyrenees) shed no
light to this lineage differentiation as they showed lugubris haplotypes indicating past
hybridization. No phylogeographic divisions were detected in F. lugubris. This pattern, similar
to the eastern phylogroup (clade A) of F. pratensis, gives no evidence for refugial separation
over several glacial periods.
Among 80 specimens of F. exsecta and four specimens of a sister species F. mesasiatica, 29
different haplotypes were found (Fig. 6, 7). In the NJ tree (Fig 6), one haplotype from Tibet
(locality 15) was clearly distant to all the other F. exsecta and F. mesasiatica haplotypes.
Notably, all F. mesasiatica haplotypes grouped together (h7 – h9) with low bootstrap support
among the F. exsecta haplotypes. A weak structure was found in the phylogenetic tree. Two
haplotypes from Northern Sweden (h27, h28, locality 8) subdivided in a separate cluster with
high bootstrap support. Most of the Asian haplotypes from the Urals to Kamchatka (localities 6,
7, 11-14) formed a separate cluster (h23 - h26). Western haplotypes (localities 1-5, 8-10)
grouped together in several small clusters representing different localities. The results suggest
that F. exsecta colonized the most part of the continent after the last glaciation from one source.
One highly diverged haplotype from Tibet indicates that another mitochondrial lineage occurs in
the species. More samples from this area are needed to make any conclusions concerning
haplotype variation in that population. These data do not enable to determine any area where
F. exsecta survived the last glaciation. The refugium might have been in the European part of the

20
Figure 6. The map showing the sampling localities of Formica exsecta and F. mesasiatica. The distribution area of F. exsecta is shown according to Dlussky (1967).
present-day range of F. exsecta, possibly close to the Urals because the Asian part of the
distribution was represented by the haplotypes from a small clade in the phylogenetic tree, and in
the Urals the mtDNA diversity was relatively high. Notably, one haplotype (h24) was extremely
common in the Asian part of the distribution. This could mean that its frequency was high in the
source population that spread towards east, giving also rise to several new haplotypes derived
from h24. Other haplotypes with eastern distribution (h12, h13, h18 and h29) are also derived
from ancestral types close to h24. Haplotypes of F. mesasiatica that occurred only in Central
Asia formed a separate cluster in the phylogenetic tree and, according to the divergence estimate
(0.12 ± 0.07% from the neighboring clade) this species separated 25-95 kyr BP, i.e. during the
last glaciation. With the climate warming after the last ice age, its ancestor population colonized
mountain pastures and remained there isolated by a vast area without suitable habitats around the
mountains.

21
Figure 7. The neighbor-joining tree of Formica exsecta and F. mesasiatica mtDNA sequences. Locality numbers are in parentheses and refer to Fig. 6. Bootstrap percentages with values greater than 50 are shown on nodes. Haplotypes of F. mesasiatica are indicated in bold. Haplotype h24 found in six localities from Romania to Kamchatka is designated by an asterisk.
To summarize, limited phylogeographic structure was found in three ant species F. pratensis,
F. lugubris and F. exsecta from the forest zone across the most of their distribution range
implying one major refugial resource for re-colonization of Eurasia after the Pleistocene
glaciations.
Demographic history
Two methods, the likelihood method and the mismatch distribution analysis, were applied to
infer demographic history of F. pratensis, F. lugubris and F. exsecta. Estimates of exponential
22
growth rate obtained by the likelihood method for F. pratensis (g = 5115 with 99.9% CI 4334 -
5895), F. lugubris (g = 5677 with 99.9% CI 4467 - 6887) and F. exsecta (g = 1797 with 99.9%
CI 1073 – 2520) significantly exceeded zero, indicating clear signs of demographic expansion.
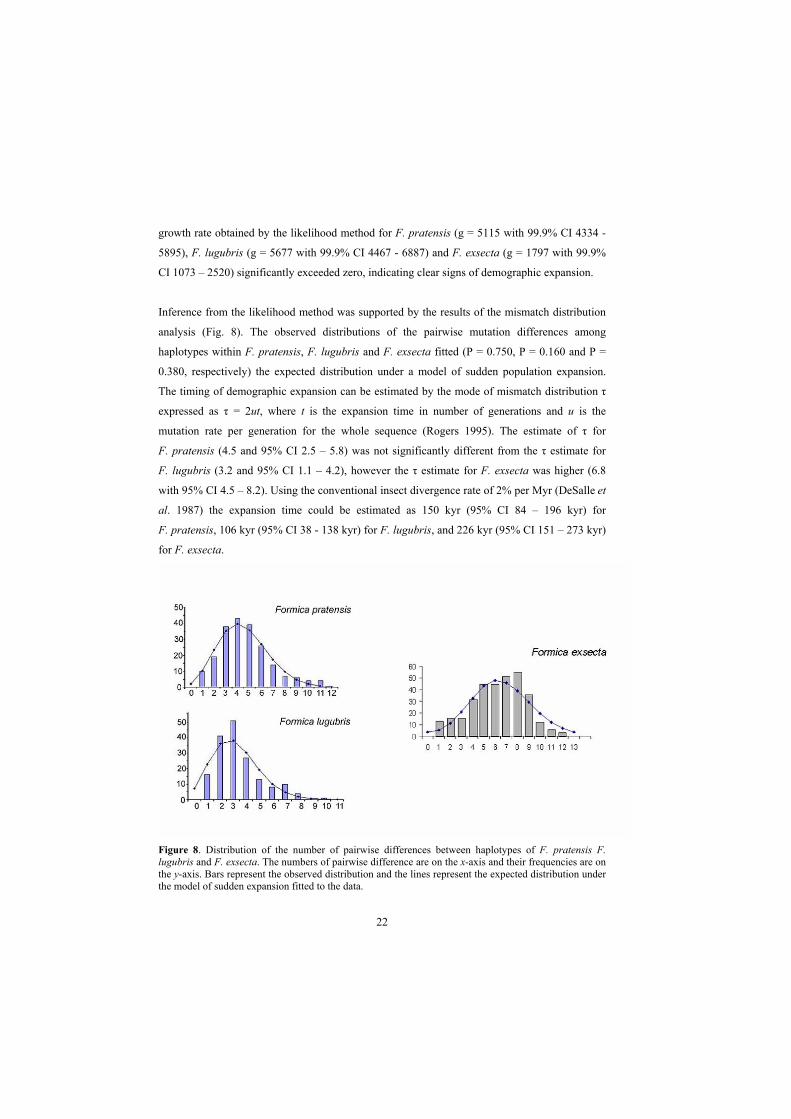
Inference from the likelihood method was supported by the results of the mismatch distribution
analysis (Fig. 8). The observed distributions of the pairwise mutation differences among
haplotypes within F. pratensis, F. lugubris and F. exsecta fitted (P = 0.750, P = 0.160 and P =
0.380, respectively) the expected distribution under a model of sudden population expansion.
The timing of demographic expansion can be estimated by the mode of mismatch distribution
expressed as = 2ut, where t is the expansion time in number of generations and u is the
mutation rate per generation for the whole sequence (Rogers 1995). The estimate of for
F. pratensis (4.5 and 95% CI 2.5 – 5.8) was not significantly different from the estimate for
F. lugubris (3.2 and 95% CI 1.1 – 4.2), however the estimate for F. exsecta was higher (6.8
with 95% CI 4.5 – 8.2). Using the conventional insect divergence rate of 2% per Myr (DeSalle et
al. 1987) the expansion time could be estimated as 150 kyr (95% CI 84 – 196 kyr) for
F. pratensis, 106 kyr (95% CI 38 - 138 kyr) for F. lugubris, and 226 kyr (95% CI 151 – 273 kyr)
for F. exsecta.
Figure 8. Distribution of the number of pairwise differences between haplotypes of F. pratensis F.lugubris and F. exsecta. The numbers of pairwise difference are on the x-axis and their frequencies are on the y-axis. Bars represent the observed distribution and the lines represent the expected distribution under the model of sudden expansion fitted to the data.

23
The estimates of population expansion time of these species indicate that the forest ants
underwent expansions during the Pleistocene when the climate experienced cyclical variations
on a time scale of 10 to 100 kyr (Bennett 1997). The species probably went through range
contractions and further expansions many times but the mismatch distribution method assesses
the time of the earliest population expansion (Rogers 1995).
Other Eurasian boreal forest species, i.e. tits (Kvist et al. 1999a, 1999b, 2001), woodpeckers
(Zink et al. 2002a, 2002b), the greenfinch (Merilä et al. 1997) also showed genetic signs of
population expansion and genetic homogeneity over the most of their distribution ranges. The
concordance in the phylogeographic pattern and similarity in demographic histories of different
species associated with boreal forests suggest that the general explanation for the lack of
phylogeographic structure across the most of Eurasia includes contraction of the distribution
range to a single refugial area at different times during the late Pleistocene, followed by
population expansion.
Mitochondrial DNA variation in Formica uralensis and F. candida
Eurasian species Formica uralensis and F. candida are widely distributed and exist as two
ecotypes with different habitat preferences. Both species inhabit bogs in Europe but in southern
Siberia, Mongolia and China they occur in dry steppe areas. Moreover, F. uralensis in the Urals
and F. candida on the vast area from the Volga River to Primorkiy Krai, can be found in both
types of habitats (cf. Dlussky 1967). One of hypotheses explains this pattern by strong
competition with other ant species that occupy the same ecological niche in non-steppe areas
where F. uralensis and F. candida presumably originated and then colonized the rest of the
continent. Another hypothesis suggests that these species occurred in European steppes before
glaciations, and with the changing environment they occupied open areas, e.g. bogs. Testing the
correspondence between haplotype genealogical relationships and geographic distribution of
haplotypes might reveal phylogeographic structure reflecting difference in the history of the two
ecotypes of these species and the putative origin of the source populations that colonized the
continent.
24
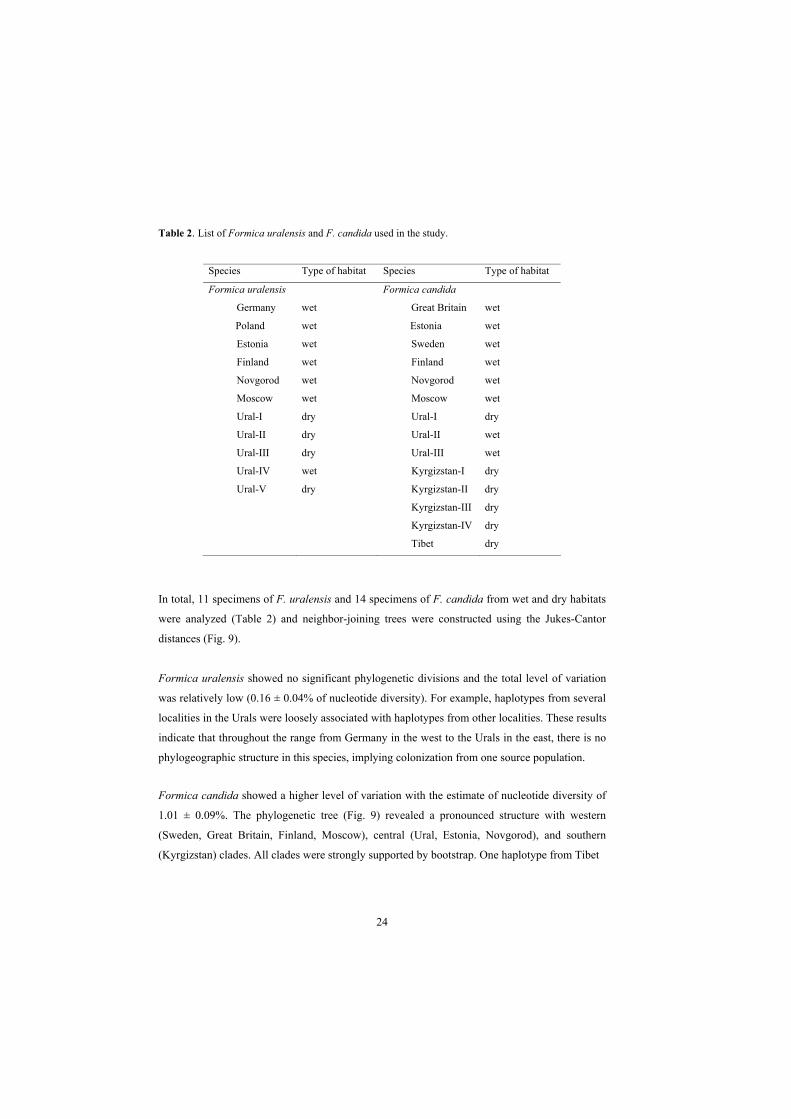
Table 2. List of Formica uralensis and F. candida used in the study.
Species Type of habitat Species Type of habitat
Formica uralensis Formica candida
Germany wet Great Britain wet
Poland wet Estonia wet
Estonia wet Sweden wet
Finland wet Finland wet
Novgorod wet Novgorod wet
Moscow wet Moscow wet
Ural-I dry Ural-I dry
Ural-II dry Ural-II wet
Ural-III dry Ural-III wet
Ural-IV wet Kyrgizstan-I dry
Ural-V dry Kyrgizstan-II dry
Kyrgizstan-III dry
Kyrgizstan-IV dry
Tibet dry
In total, 11 specimens of F. uralensis and 14 specimens of F. candida from wet and dry habitats
were analyzed (Table 2) and neighbor-joining trees were constructed using the Jukes-Cantor
distances (Fig. 9).
Formica uralensis showed no significant phylogenetic divisions and the total level of variation
was relatively low (0.16 ± 0.04% of nucleotide diversity). For example, haplotypes from several
localities in the Urals were loosely associated with haplotypes from other localities. These results
indicate that throughout the range from Germany in the west to the Urals in the east, there is no
phylogeographic structure in this species, implying colonization from one source population.
Formica candida showed a higher level of variation with the estimate of nucleotide diversity of
1.01 ± 0.09%. The phylogenetic tree (Fig. 9) revealed a pronounced structure with western
(Sweden, Great Britain, Finland, Moscow), central (Ural, Estonia, Novgorod), and southern
(Kyrgizstan) clades. All clades were strongly supported by bootstrap. One haplotype from Tibet

25
Formica uralensis Formica candida
Figure 9. The neighbor-joining tree of Formica uralensis and F. candida mtDNA sequences. Specimens refer to Table 2. Bootstrap percentages are shown on nodes.
did not cluster to any group but was distantly connected to the western clade. There was no
association between the haplotype position on the NJ tree and type of the habitat, i.e. the samples
from wet and dry habitats clustered together in the central clade. The net divergence estimates
between the western and the central groups was 0.8 ± 0.2%, and between the pooled western-
central group and the southern group 1.2 ± 0.3%. Using the conventional insect divergence rate
as 2% per Myr enables to estimate the approximate divergence time for the three lineages. The
southern clade diverged from the western and the central groups 450 – 750 kyr, approximately at
the same time as the F. exsecta Tibetan sample from the rest of the conspecific samples (see
above). These results imply the existence of old Asian lineages that did not disperse over large
areas. The F. candida haplotype from Tibet however, was separate from the other clades that
might indicate the existence of another Asian lineage highly diverged from the eastern clade in
the present data. The western and the central clades were separated 300 – 500 kyr BP which
exceeds the time span of the last glaciation (115 kyr BP) and indicates existence of at least two
refugia during the last glaciation.
Although F. uralensis and F. candida demonstrate similar habitat preferences, their
phylogeographic patterns are different, even if we take into account differences in the
geographical sampling of the two species. More studies with extensive sampling across the entire
distribution ranges of both species and population sampling for F. uralensis should further
clarify their histories.

26
Genetic characteristics of northern European populations of Formica cinerea
Ants were collected from 23 localities (Fig. 1) that can be divided into two groups based on the
distribution and abundance of the species. Group A includes samples from southernmost
Sweden, Denmark, Estonia and south-eastern Finland, where the species is abundant and the
populations are not isolated. Group B includes the remaining populations in Fennoscandia which
are considered isolated on the basis of long distances to other known populations. Five
microsatellite loci used in the present study showed different level of polymorphism. The
average number of alleles per locus ranged from 2.8 in Öland (10) to 5.6 in Estonia-A (14) and
was significantly higher in populations of Group A than of Group B. A similar pattern applied to
the expected heterozygosity.
Figure 10. Map showing sampling localities. Open triangles correspond to populations of Group A, filled triangles to populations of Group B (see text).

27
Mean genetic relatedness among nestmate workers varied widely from -0.003 in Estonia-B (15)
to 0.68 in Skåne (11) (Table 3). The populations could be classified in three groups on the basis
of worker relatedness: highly polygynous (relatedness estimate not significantly different from
zero), putatively monogynous (relatedness higher than 0.59; Pamilo 1993) and of intermediate
levels of polygyny. There was a trend of monogyny being more common in geographically
isolated populations. The effective population size could be fairly small in most populations with
monogynous colonies that makes the populations vulnerable to demographic stochasticity.
The pairwise exact test of population differentiation (Goudet et al. 1996) revealed highly
significant differences in allele frequencies between most of the populations across most loci.
Table 3. Mean genetic relatedness (r) among nestmate workers with standard errors over nests and loci in Formica cinerea populations. Data marked with asterisk are from Zhu et al. (in press).
Population Mean r S.E. over nests S.E. over loci Highly polygynous 15 Estonia-B -0.003 0.030 0.033 16 Ruhnu 0.009 0.032 0.015 4 Hällefors 0.021 0.042 0.076 5 Brattfors-A 0.058 0.046 0.060 6 Brattfors-B 0.058 0.048 0.041 20 Hanko-A 0.062 0.027 0.043 14 Estonia-A 0.070 0.054 0.018 18* Sotkamo 0.08 0.03 2 Ambjörby 0.099 0.055 0.052 19* Kontiolahti 0.10 0.05
Intermediate 8 Bollnäs-A 0.164 0.130 0.056 17* Kalajoki 0.25 0.07 21 Hanko-B 0.269 0.255 0.061 13 Jutland-B 0.397 0.053 0.046 10 Öland 0.423 0.017 0.099 12 Jutland-A 0.464 0.067 0.048 22 Hanko-C 0.467 0.079 0.069
Monogynous 3 Sysslebäck 0.593 0.075 0.047 9 Bollnäs-B 0.617 0.077 0.034 1 Elverum 0.633 0.061 0.019 23 Hanko-D 0.660 0.054 0.057 7 Mora 0.677 0.058 0.046 11 Skåne 0.684 0.035 0.026
Fixation indices FST and RST among all localities were 0.111 and 0.082 respectively. Smaller
value of RST than FST indicates large length variation of alleles that reflects the ancient
polymorphism in a source population before colonization of northern Europe. Some alleles could

28
be lost due to founder effects and stochastic events, so the pairwise RST estimate is sensitive to the
length difference of the remaining alleles. There was no correlation between genetic (pairwise
FST) and geographical distances for all populations (coef. corr. = 0.000, P = 0.160 with the Mantel
test). The multidimensional scale plot did not show any geographical pattern among different
localities (data not shown).
It has been hypothesized that different dispersal strategies, connected to the level of polygyny,
can lead to different spatial patterns of genetic differentiation (Pamilo & Rosengren 1984; Seppä
& Pamilo 1995; Ross et al. 1997). New females from monogynous nests mate and disperse by
flight whereas those from polygynous nests can stay in their natal colony or disperse by budding
at limited distances. Allozyme studies have indicated that subdivision is stronger among
polygynous populations than among monogynous populations of the same or closely related
species in F. truncorum and in Myrmica ants (Sundström 1993; Seppä & Pamilo 1995). The
present results support to limited extent the hypothesis that polygyny can lead to genetic
differentiation. For example, closely situated (8 km) populations Hanko-A (20) and -B (21) were
polygynous and genetically differentiated from each other (FST = 0.13), suggesting restricted
gene flow. However, low differentiation between other closely located populations with
polygynous colonies did not support the hypothesis.
It seems plausible that the connectivity of F. cinerea populations has been higher in the past than
it seems to be today. The species has been observed to colonize disturbed habitats, such as old
sand mining areas, and it is likely that similar open sandy habitats have earlier been created for
example by forest fires. This would have allowed the species to expand its distribution and could
explain the present pattern of genetic diversity and differentiation. Today, the paucity of suitable
habitats has led to separation of populations and it is probable that the populations are no more
connected to a similar extent they used to be.
Even though the level of heterozygosity was slightly reduced in the group B populations, the
relatively high polymorphism in all populations studied supports the idea that there has been a
large panmictic, or at least non-differentiated, F. cinerea population in northern Europe and that
the time of isolation of the present populations has not been long enough to decrease genetic
diversity. The lack of clustering among all studied populations did not allow to conclude whether
F. cinerea colonized Fennoscandia from the south, possibly via a land bridge that connected
Scandinavia to the continent, or from the north-east, or from both directions.

29
CONCLUSIONS
The mitochondrial DNA phylogeny of Palaearctic Formica species supports the conventional
subgeneric grouping based on morphological characteristics. The exception is that F. uralensis
formed a separate phylogroup. The mitochondrial DNA phylogeny is to a large extent congruent
with the phylogenetic tree inferred from allozymes.
The mitochondrial DNA phylogeny of the Formica rufa group shows the division of the group
into three major phylogenetic groups: one with the species F. polyctena and F. rufa, one with
F. aquilonia, F. lugubris and F. paralugubris, and the third one with F. pratensis.
The mitochondrial DNA phylogeny of the Formica rufa group gives evidence for multiple
evolutionary origin of polygyny and does not clearly support the importance of social
organization for speciation, except perhaps for F. aquilonia and F. paralugubris that build large
supercolonies and cluster closely together in the phylogenetic tree.
The mitochondrial DNA phylogeny shows west-east phylogeographic divisions in Formica
pratensis. This division suggests post-glacial colonization of western Europe and of a wide area
from Sweden to the Baikal Lake from separate forest refugia. In contrast, no phylogeographic
divisions were detected in either F. lugubris or F. exsecta. Comparison of species-wide
phylogeography between these three sympatrically distributed species demonstrates a difference
in the phylogeographic structure that implies different vicariant history. However, over most of
the distribution ranges, similar signs of demographic expansion predating the last glaciation and
the lack of phylogeographic structure were found in the eastern phylogroup of F. pratensis, in
F. lugubris and in F. exsecta. Contraction of the distribution range of each species to a single
refugial area during the late Pleistocene and the following population expansion seem to offer a
general explanation for the lack of the phylogeographic structure across the most of Eurasia.
Two species, Formica uralensis and F. candida, show clear difference in the phylogeographic
structure. While no phylogeographic divisions were detected in F. uralensis across Europe,
F. candida showed well supported phylogeographic divisions between the western group
(Sweden, Great Britain, Finland, Moscow), the central group (Ural, Estonia, Novgorod), and the
southern group (Kyrgizstan). This difference implies that the two sympatrically distributed and
ecologically similar species had different vicariant history.

30
In Formica cinerea, the geographically isolated populations showed slightly reduced levels of
genetic diversity, although the overall level of intrapopulation microsatellite diversity was
relatively high and differentiation among populations low, indicating recent historical
connections. The genetic clustering of the populations does not follow clear geographical
patterns, and the data do not allow inferring the routes used to colonize Fennoscandia.

31
ACKNOWLEDGMENTS
I thank my supervisor Pekka Pamilo for help and support, for the opportunity to meet interesting
people and to travel. I thank Vadim Fedorov for his contribution to this study. I am grateful to
Bernhard Seifert for close collaboration.
I also thank A. Alexeev, V.Baglione, A. Belyaev, S.-Å. Berglind, I. Bortnikova, M. Chapuisat,
D. Cherix, G. Dlussky, N. Dokuchaev, A. Gilev, N. Gyllenstrand, A. Kaluzhnikov, A. Lazutkin,
K. Liautard, M. Lund, B. Marko, A.-J. Martin, T. Monnin, P. Neumann, G. Orledge, I.
Sarapultsev, V. Semerikov, P. Seppä, P. Smith, D. Stradling, A. Tinaut, E. Van Walsum, and A.
Zakharov for providing sampling material. Many thanks to C. Vila, V. Semerikov and M.
Griesser for the help with computer programs, to J. A. Cook and K. G. McCracken for providing
laboratory space in the Institute of Arctic Biology, Fairbanks, USA for a part of the work. I thank
J. Wallén for laboratory assistance.
This study has been supported by grants from the Natural Science Research Councils of Sweden
and Finland, Sven and Lilly Lawski’s Foundation and from the European Comission.
Finally, I would like to thank all my friends and my family for love and support, all people at our
department for creating a nice working environment.

32
REFERENCES
Anderson BG, Borns HW Jr (1997) The Ice Age World. An Introduction to Quaternary History
and Research with Emphasis on North America and Europe during the Last 2.5 Million
Years. Scandinavian University Press, Oslo, Norway.
Arbogast BS, Kenagy GJ (2001) Comparative phylogeography as an integrative approach to
historical biogeography. Journal of Biogeography, 28, 819-825.
Bennett KD (1997) Evolution and Ecology: the Pace of Life. Cambridge University Press.
Bilton DT, Mirol PM, Mascheretti S, Fredga K, Zima J, Searle JB (1998) Mediterranean Europe
as an area of endemism for small mammals rather than a source for northwards
postglacial colonization. Proceedings of the Royal Society of London, Series B, 265,
1219-1226.
Brunhoff C, Galbreath KE, Fedorov VB, Cook JA, Jaarola M (2003) Holarctic phylogeography
of the root vole (Microtus oeconomus): implications for late Quaternary biogeography of
high latitudes. Molecular Ecology, 12, 957-968.
Chapuisat M, Goudet J, Keller L (1997) Microsatellites reveal high population viscosity and
limited dispersal in the ant Formica paralugubris. Evolution, 51, 475-482.
Cherix D, Chautems D, Fletcher DJC, Fortelius W, Gris G, Keller L, Passera L, Rosengren R,
Vargo EL, Walter F (1991) Alternative reproductive strategies in Formica lugubris Zett
(Hymenoptera, Formicidae). Ethology, Ecology & Evolution, Special Issue, 1, 61-66.
Collingwood CA (1979) The Formicidae (Hymenoptera) of Fennoscandia and Denmark. Fauna
Entomologica Scandinavica, 8, 1-174.
Crozier RH, Crozier YC (1993) The mitochondrial genome of the honeybee Apis mellifera:
complete sequence and genome organization. Genetics, 133, 97-117.
Crozier RH, Pamilo P (1996) Evolution of Social Insect Colonies. Oxford University Press,
Oxford New York Tokyo.
Czechowski W (1996) Colonies of hybrids and mixed colonies; interspecific nest takeover in
wood ants (Hymenoptera, Formicidae). Memorabilia zoologica, 50, 1-116.
DeSalle R, Freedman T, Prager E, Wilson AC (1987) Tempo and mode of sequence evolution in
miochondrial DNA of Hawaiian Drosophila. Journal of Molecular Evolution, 26, 157-
164.
Dlussky GM (1967) The Ants of Genus Formica (in Russian). Nauka, Moscow.
Efimik VE (1996) Pliocene and Pleistocene relict species in spider fauna of the South Urals.
Zoologichesky Zhurnal, 75, 1138-1148.

33
Estoup A, Cornuet J-M (1998) Microsatellites evolution: inferences from population data. In:
Microsatellites: Evolution and Applications (eds Goldstein DB, Schlötter C), pp. 50-65.
Oxford University Press, Oxford New York.
Goldstein DB, Schlötterer C (1999) Microsatellites: Evolution and Applications. Oxford
University Press, Oxford.
Goudet J, Raymond M, De Meeus T, Rousset F (1996) Testing differentiation in diploid
populations. Genetics, 144, 933-1940.
Grichuk VP (1984) Late Pleistocene vegetation history. In: Late Quaternary environments of the
Soviet Union (eds Velichko AA, Wright JrHE, Barnosky CW), pp. 155-179. University
of Minnesota Press. Minneapolis.
Goropashnaya AV, Fedorov VB, Pamilo P. Recent speciation in the Formica rufa group ants
(Hymenoptera, Formicidae): inference from mitochondrial DNA phylogeny. Molecular
Phylogenetics & Evolution (in press).
Gyllenstrand N, Seppä P, Pamilo, P. Genetic differentiation in sympatric wood ants, Formica
rufa and F. polyctena. Insectes Sociaux (in press).
Haase M, Misof B, Wirth T, Baminger H, Baur B (2003) Mitochondrial differentiation in a
polymorphic land snail: evidence for Pleistocene survival within the boundaries of
permafrost. Journal of Evolutionary Biology, 16, 415-428.
Hamilton WD (1964) The genetical evolution of social behaviour, II. Journal of Theoretical
Biology, 7, 17-52.
Hedrick PW (1999) Perspective: Highly variable loci and their interpretation in evolution and
conservation. Evolution, 53, 313–318.
Hewitt GM (1996) Some genetic consequences of ice ages, and their role in divergence and
speciation. Biological Journal of the Linnean Society, 58, 247-276.
Hewitt GM (1999) Post-glacial re-colonization of European biota. Biological Journal of the
Linnean Society, 68, 87-112.
Jaarola M, Searle JB (2002) Phylogeography of field voles (Microtus agrestis) in Eurasia
inferred from mitochondrial DNA sequences. Molecular Ecology, 11, 2613-2621.
Jarne P, Lagoda PJL (1996) Microsatellites, from molecules to populations and back. Trends of
Ecology & Evolution, 11, 424-429.
Keller L (1991) Queen number, mode of colony founding and queen reproductive success in ants
(Hymenoptera, Formicidae). Ethology, Ecology & Evolution, 3, 307-316.
Kuhner MK, Yamato J, Felsenstein J (1998) Maximum likelihood estimation of population
growth rates based on the coalescent. Genetics, 149, 429-434.

34
Kvist L, Ruokonen M, Lumme J, Orell M (1999a) Different population structures in northern
and southern populations of the European blue tit (Parus caeruleus). Journal of
Evolutionary Biology, 12, 798-805.
Kvist L, Ruokonen M, Lumme J, Orell M (1999b) The colonisation history and present day
colonisation structure of the European great tit (Parus major). Heredity, 82, 495-502.
Kvist L, Martens J, Ahoa A, Orell M (2001) Phylogeography of a Palaearctic sedentary
passerine, the willow tit (Parus montanus). Journal of Evolutionary Biology, 14, 930-
941.
Lagercrantz U, Ryman N (1990) Genetic structure of Norway spruce (Picea abies): concordance
of morphological and allozymic variation. Evolution, 44, 38-53.
Lessa EP, Cook JA, Patton JL (2003) Genetic footprints of demographic expansion in North
America, but not Amazonia, during the Late Quaternary. Proceedings of the National
Academy of Sciences USA, 100, 10331-10334.
Liautard C, Keller L (2001) Restricted effective queen dispersal at a microgeographic scale in
polygynous populations of the ant Formica exsecta. Evolution, 55, 2484-2492.
Merilä J, Björklund M, Baker AJ (1997) Historical demography and present day population
structure of the greenfinch, Carduelis chloris – an analysis of mtDNA control-region
sequences. Evolution, 51, 946-956.
Pamilo P (1993) Polyandry and allele frequency differences between the sexes in the ant
Formica aquilonia. Heredity, 70, 472-480.
Pamilo P, Rosengren R (1984) Evolution of nesting strategies of ants: genetic evidence from
different population types of Formica ants. Biological Journal of the Linnean Society, 21,
331-348.
Pamilo P, Seppä P (1994) Reproductive competition and conflicts in colonies of the ant Formica
sanguinea. Animal Behaviour, 48, 1201-1206.
Pamilo P, Vepsäläinen K, Rosengren R, Varvio-Aho S-L, Pisarski B (1979) Population genetics
of Formica ants II. Genic differentiation between species. Annales Entomologica Fennici,
45, 65-76.
Pamilo P, Chautems D, Cherix D (1992) Genetic differentiation of disjunct populations of the
ants Formica aquilonia and Formica lugubris in Europe. Insectes Sociaux, 39, 15-29.
Pamilo P, Sundström L, Fortelius W, Rosengren R (1994) Diploid males and colony-level
selection in Formica ants. Ethology Ecology & Evolution, 6, 221-235.
Rogers AR (1995) Genetic evidence for a Pleistocene population explosion. Evolution, 49, 608-
615.

35
Rosengren R, Pamilo P (1983) The evolution of polygyny and polydomy in mound-building
Formica ants. Acta Entomologica Fennica, 42, 65-77.
Rosengren R, Chautems D, Cherix D, Fortelius W, Keller L (1994) Separation of two sympatric
sibling species of Formica L. ants by behavioural choice test based on brood
discrimination. Memorabilia Zoologica, 48, 237-249.
Ross KG, Carpenter JM (1991) Phylogenetic analysis and the evolution of queen number in
eusocial Hymenoptera. Journal of Evolutionary Biology, 4, 117-130.
Ross KG, Krieger MJB, Shoemarker DD, Vargo EL, Keller L (1997) Hierarchical analysis of
genetic structure in native fire ant populations: results from three classas of molecular
markers. Genetics, 147, 643-655.
Schmitt T, Seitz A (2001) Intraspecific allozymatic differentiation reveals the glacial refugia and
the postglacial expansions of European Erebia medusa (Lepidoptera: Nymphalidae).
Biological Journal of the Linnean Society, 74, 429-458.
Seifert B (1991) The phenotypes of the Formica rufa complex in East Germany. Abh. Ber.
Naturkundemus, 65, 1-27.
Seifert B (1996) Formica paralugubris nov. spec. – a sympatric sibling species of Formica
lugubris from the western Alps (Insecta: Hymenoptera: Formicoidea: Formicidae).
Reichenbachia, 31, 193-201.
Seppä P, Pamilo P (1995) Gene flow and population viscosity in Myrmica ants. Heredity, 74,
200-209.
Stewart JR, Lister AM (2001) Cryptic northern refugia and the origin of modern biota. Trends
Ecology Evolution, 16, 608-613.
Sumegi P, Krolopp E (2002) Quatermalacological analyses for modeling of the Upper
Weichselian palaeoenvironment changes in the Carpathian Basin. Quaternary
International, 91, 53-63.
Sundström L (1993) Genetic population structure and sociogenetic organisation in Formica
truncorum (Hymenoptera, Formicidae). Behavioral Ecology & Sociobiology, 33, 345-
354.
Sundström L, Keller L, Chapuisat M (2003) Inbreeding and sex-biased gene flow in the ant
Formica exsecta. Evolution, 57, 1552-1561.
Taberlet P, Fumagalli L, Wust-Saucy A-G, Cosson J-F (1998) Comparative phylogeography and
postglacial colonization routes in Europe. Molecular Ecology, 7, 453-464.
Tarasov PE, Volkova VS, Webb T, Guiot J, Andreev AA, Bezusko LG, Bezusko TV, Bykova
GV, Dorofeyuk NI, Kvavadze EV, Osipova IM, Panova NK, Sevastyanov DV (2000)

36
Last glacial maximum biomes reconstructed from pollen and plant macrofossil data from
northern Eurasia. Journal of Biogeography, 27, 609-620.
Ward PS (1989) Genetic and social changes associated with ant speciation. In: The Genetics of
Social Evolution (eds Breed MD, Page RE Jr) pp. 123-148.Westview Press.
West KJ (2000) Plant Life of the Quaternary Cold Stages. Cambridge University Press,
Cambridge.
Willis KJ, Rudner E, Sümegi P (2000) The full-glacial forests of central and southeastern
Europe. Quaternary Research, 53, 203-213.
Zink RM, Slowinski JB (1995) Evidence from molecular systematics for decreased avian
diversification in the Pleistocene epoch. Proceedings of the National Academy of Science
USA, 92, 5832-5835.
Zink RM, Drovetski SV, Rohwer S (2002a) Phylogeographic patterns in the great spotted
woodpecker Dendrocopos major across Eurasia. Journal of Avian Biology, 33, 175-178.
Zink RM, Rohwer S, Drovetski SV, Blackwell-Rago RC, Farrell SL (2002b) Holarctic
phylogeography and species limits of three-toed woodpeckers. Condor, 104, 167-170.
Zhu D, Chapuisat M, Pamilo P. Highly variable social organisation of colonies in the ant
Formica cinerea. Hereditas (in press).


Acta Universitatis UpsaliensisComprehensive Summaries of Uppsala Dissertations
from the Faculty of Science and TechnologyEditor: The Dean of the Faculty of Science and Technology
Distribution:Uppsala University Library
Box 510, SE-751 20 Uppsala, Swedenwww.uu.se, [email protected]
ISSN 1104-232XISBN 91-554-5818-1
A doctoral dissertation from the Faculty of Science and Technology, UppsalaUniversity, is usually a summary of a number of papers. A few copies of thecomplete dissertation are kept at major Swedish research libraries, while thesummary alone is distributed internationally through the series ComprehensiveSummaries of Uppsala Dissertations from the Faculty of Science and Technology.(Prior to October, 1993, the series was published under the title “ComprehensiveSummaries of Uppsala Dissertations from the Faculty of Science”.)