plant vira1 synergism: the potyviral genome encodes a ... of biology, texas a & m university,...
TRANSCRIPT

The Plant Cell, Vol. 9, 859-868, June 1997 O 1997 American Society of Plant Physiologists
Plant Vira1 Synergism: The Potyviral Genome Encodes a Broad-Range Pathogenicity Enhancer That Transactivates Replication of Heterologous Viruses
Gail Pruss,a Xin Ge,a Xing Ming Shi,a James C. Carrington,bsi and Vicki Bowman Vancea12 a Department of Biological Sciences, University of South Carolina, Columbia, South Carolina 29208
Department of Biology, Texas A & M University, College Station, Texas 77843
Synergistic viral diseases of higher plants are caused by the interaction of two independent viruses in the same host and are characterized by dramatic increases in symptoms and in accumulation of one of the coinfecting viruses. In po- tato vírus X (PVX)/potyviral synergism, increased pathogenicity and accumulation of PVX are mediated by the expression of potyviral5’ proximal sequences encoding Pl , the helper component proteinase (HC-Pro), and a fraction of P3. Here, we report that the same potyviral sequence (termed Pl/HC-Pro) enhances the pathogenicity and accumula- tion of two other heterologous viruses: cucumber mosaic virus and tobacco mosaic virus. In the case of PVX-potyviral synergism, we show that the expression of the HC-Pro gene product, but not the RNA sequence itself, is sufficient t o induce the increase in PVX pathogenicity and that both P1 and P3 coding sequences are dispensable for this aspect of the synergistic interaction. In protoplasts, expression of the potyviral Pl/HC-Pro region prolongs the accumulation of PVX (-) strand RNA and transactivates expression of a reporter gene from a PVX subgenomic promoter. Unlike the synergistic enhancement of PVX pathogenicity, which requires only expression of HC-Pro, the enhancement of PVX (-) strand RNA accumulation in protoplasts is significantly greater when the entire Pl/HC-Pro sequence is expressed. These results indicate that the potyviral Pl/HC-Pro region affects a step in disease development that is common to a broad range of virus infections and suggest a mechanism involving transactivation of viral replication.
INTRODUCTION
Mixed virus infections occur in both plant and animal sys- tems, and doubly infected organisms commonly display in- creases in disease symptoms and in the accumulation of one or both of the viruses. In animal hosts, mixed virus in- fections are relatively infrequent and are generally associ- ated with depression of the immune response (Calendar, 1986). In contrast, higher plants are commonly coinfected with multiple viruses, and a number of disease syndromes are caused by the interaction of two independent viruses (Matthews, 1991). Many such synergistic diseases involve a member of the potyvirus group of plant viruses. In potyvirus- associated synergisms, the other virus of the pair may be any of a broad range of unrelated viruses, including pararetro- viruses such as cauliflower mosaic virus (Khan and Demski, 1982) and RNA viruses of both the alphavirus supergroup (e.g., potato virus X [Pvx]; Rochow and Ross, 1955) and the picornavirus supergroup (e.g., cowpea mosaic virus; Anjos
Current address: lnstitute of Biological Chemistry, Washington State University, Pullman, WA 991 64. ‘To whom correspondence should be addressed. E-mail vance@biol. sc.edu; fax 803-777-4002.
et al., 1992). Severa1 such potyvirus-associated synergistic diseases have been examined in detail, and in each, a dra- matic increase in host symptoms has been observed in dou- bly infected plants compared with singly infected plants. The increase in symptoms is correlated with an increase in the ac- cumulation of the non-potyvirus, but there is no corresponding increase or decrease in the level of the potyvirus (Rochow and Ross, 1955; Calvert and Ghabrial, 1983; Goldberg and Brakke, 1987; Vance, 1991). Such synergistic interactions offer a unique opportunity to investigate the regulation of viral repli- cation and disease because the accumulation and pathoge- nicity of one virus are altered when it interacts with a relatively simple genetic element, the genome of the coin- fecting virus.
. lhe best characterized of the plant viral synergisms is the interaction between PVX and the potyvirus potato virus Y (Pw) in tobacco (Rochow and Ross, 1955; Goodman and Ross, 1974a, 1974b; Vance, 1991). Plants mechanically inoc- ulated with both viruses develop synergistic disease, which is characterized initially by severe vein clearing and then necro- sis of the first systemically infected leaf tissue, with a three- to 10-fold increase in the level of PVX compared with singly

860 The Plant Cell
infected plants. A similar synergistic response is induced incoinfections of PVX with several other potyviruses, includingthe well-studied tobacco vein mottling and tobacco etch vi-ruses (TVMV and TEV, respectively) (Vance et al., 1995).
We have recently shown that PVX-potyviral synergistic dis-ease does not require replication of the potyviral genomic RNAand that the response is mediated by expression of potyviralsequences in the 5' proximal one-third of the potyviral genome(Vance et al., 1995). This region of the potyviral genome in-cludes the viral genomic 5' untranslated region (UTR) as well asthe coding region for the N-terminal portion of the viral polypro-tein, including P1, the helper component proteinase (HC-Pro),and approximately one-quarter of P3 (termed P1/HC-Pro se-quence). Here, we report that expression of the TEV P1/HC-Pro sequence also enhances the pathogenicity and ac-cumulation of both tobacco mosaic virus (TMV) and cucum-ber mosaic virus (CMV), two unrelated plant viruses thatcause synergistic disease in conjunction with a potyvirus(Pio-Ribeiro et al., 1978; Clark et al., 1980; Poolpol and Inouye,1986). We show that in protoplasts, the potyviral sequenceprolongs both the accumulation of PVX (-) strand RNA andthe expression of a reporter gene from a PVX subgenomicpromoter, suggesting that it acts, at least in part, by transacti-vation of viral RNA replication.
Enhanced Accumulation of TMV and CMV Genomic RNAsin Transgenic Plants Expressing the TEVP1/HC-Pro Sequence
Accumulation of PVX RNA is enhanced in U-6B transgenicplants (Vance et al., 1995). To determine whether CMV andTMV RNA levels are similarly enhanced in this transgenicline, we compared the levels of viral genomic RNA in sys-temically infected U-6B plants with those in vector-onlytransformed plants. In the case of TMV infection, the level ofTMV genomic RNA in a severely symptomatic leaf of theU-6B transgenic plant (fourth leaf above the inoculated leaf;Figure 2A, lane 2) was >20-fold higher than that in the cor-responding leaf of the control plant (lane 1). Thus, accumu-lation of TMV genomic RNA, like that of PVX genomic RNA, isenhanced by the expression of the TEV P1/HC-Pro sequence.
In the case of CMV infection, accumulation of several ge-nomic RNAs and a satellite RNA potentially can be affectedby expression of the TEV P1/HC-Pro sequence. The CMVgenome is divided among three components: RNAs 1 and 2are monocistronic, whereas RNA 3 is bicistronic, with the in-ternally coded gene expressed from a subgenomic RNAtermed RNA 4. Many CMV strains, including the one usedhere, support the replication of a small satellite RNA. ThisCMV satellite RNA is completely dependent on the virus for
RESULTS
Enhanced Pathogenicity of TMV and CMV in TransgenicPlants Expressing the TEV P1/HC-Pro Sequence
The finding that transgenic plants expressing the P1/HC-Proregion of the potyviral genome develop synergistic diseasewhen infected with PVX (Vance et al., 1995) raised the possi-bility that many or all potyvirus-associated synergisms mightbe mediated by this same sequence. To test this hypothesis,we used two other viruses, TMV and CMV, to infect trans-genic tobacco line U-6B, which expresses the P1/HC-Pro re-gion of the TEV genome (Carrington et al., 1990). Both virusescan infect tobacco and are known to interact synergisticallywith a potyvirus. TMV and CMV are both (+) strand RNA vi-ruses in the alphavirus supergroup; however, CMV has a tri-partite genome, and many strains also support replication ofan associated satellite RNA, whereas the genome of TMV,like that of PVX, is monopartite. Developmentally matchedU-6B and vector-only transgenic tobacco plants were inoc-ulated with either TMV or CMV and monitored for symptomdevelopment. Both CMV and TMV induced severe symp-toms in the U-6B transgenic line, resulting in death of thehost plant after several weeks, as shown in Figures 1B and1D, respectively. In contrast, both viruses induced relativelymild symptoms in the control tobacco plants over the sametime period (Figures 1A and 1C, respectively). This resultsupports the idea that the P1/HC-Pro region of TEV acts asa pathogenicity enhancer for a broad range of viruses.
CMV - CONTROL CMV - U-6B TRANSGENIC
TMV - CONTROL TMV - U-6B TRANSGENIC
CHBk .̂ D
Figure 1. Enhanced Pathogenicity of CMV and TMV in TransgenicTobacco Expressing the Potyviral P1/HC-Pro Sequence.
(A) and (C) Vector-only transformed tobacco plants.(B) and (D) TEV-transformed line U-6B plants expressing the 5'proximal region of the TEV genome.Plants were infected with either CMV JA] and [B]) or TMV ([C] and [D]).
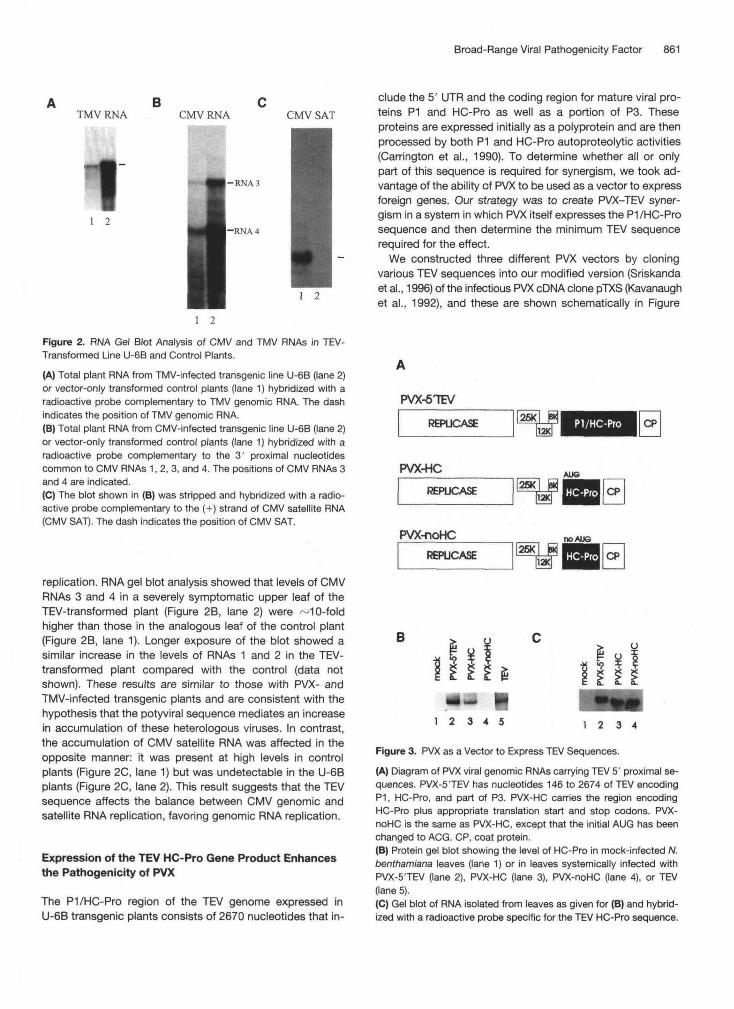
Broad-Range Viral Pathogenicity Factor 861
A B CTMV RNA CMV RNA CMV SAT
-RNA3
-RNA 41 2
1 2
1 2
Figure 2. RNA Gel Blot Analysis of CMV and TMV RNAs in TEV-Transformed Line U-6B and Control Plants.(A) Total plant RNA from TMV-infected transgenic line U-6B (lane 2)or vector-only transformed control plants (lane 1) hybridized with aradioactive probe complementary to TMV genomic RNA. The dashindicates the position of TMV genomic RNA.(B) Total plant RNA from CMV-infected transgenic line U-6B (lane 2)or vector-only transformed control plants (lane 1) hybridized with aradioactive probe complementary to the 3' proximal nucleotidescommon to CMV RNAs 1,2,3, and 4. The positions of CMV RNAs 3and 4 are indicated.(C) The blot shown in (B) was stripped and hybridized with a radio-active probe complementary to the (+) strand of CMV satellite RNA(CMV SAT). The dash indicates the position of CMV SAT.
replication. RNA gel blot analysis showed that levels of CMVRNAs 3 and 4 in a severely symptomatic upper leaf of theTEV-transformed plant (Figure 2B, lane 2) were ~10-foldhigher than those in the analogous leaf of the control plant(Figure 2B, lane 1). Longer exposure of the blot showed asimilar increase in the levels of RNAs 1 and 2 in the TEV-transformed plant compared with the control (data notshown). These results are similar to those with PVX- andTMV-infected transgenic plants and are consistent with thehypothesis that the potyviral sequence mediates an increasein accumulation of these heterologous viruses. In contrast,the accumulation of CMV satellite RNA was affected in theopposite manner: it was present at high levels in controlplants (Figure 2C, lane 1) but was undetectable in the U-6Bplants (Figure 2C, lane 2). This result suggests that the TEVsequence affects the balance between CMV genomic andsatellite RNA replication, favoring genomic RNA replication.
Expression of the TEV HC-Pro Gene Product Enhancesthe Pathogenicity of PVX
The P1/HC-Pro region of the TEV genome expressed inU-6B transgenic plants consists of 2670 nucleotides that in-
clude the 5' UTR and the coding region for mature viral pro-teins P1 and HC-Pro as well as a portion of P3. Theseproteins are expressed initially as a polyprotein and are thenprocessed by both P1 and HC-Pro autoproteolytic activities(Carrington et al., 1990). To determine whether all or onlypart of this sequence is required for synergism, we took ad-vantage of the ability of PVX to be used as a vector to expressforeign genes. Our strategy was to create PVX-TEV syner-gism in a system in which PVX itself expresses the P1 /HC-Prosequence and then determine the minimum TEV sequencerequired for the effect.
We constructed three different PVX vectors by cloningvarious TEV sequences into our modified version (Sriskandaet al., 1996) of the infectious PVX cDNA clone pTXS (Kavanaughet al., 1992), and these are shown schematically in Figure
PVX-5TEV
REPUCASE PI/HC-Pro CP
PVX-HC
REPUCASE
PVX-noHCREPUCASE
AUG
noAJJS
B
1 2 3 4
Figure 3. PVX as a Vector to Express TEV Sequences.(A) Diagram of PVX viral genomic RNAs carrying TEV 5' proximal se-quences. PVX-5'TEV has nucleotides 146 to 2674 of TEV encodingP1, HC-Pro, and part of P3. PVX-HC carries the region encodingHC-Pro plus appropriate translation start and stop codons. PVX-noHC is the same as PVX-HC, except that the initial AUG has beenchanged to ACG. CP, coat protein.(B) Protein gel blot showing the level of HC-Pro in mock-infected N.benthamiana leaves (lane 1) or in leaves systemically infected withPVX-5'TEV (lane 2), PVX-HC (lane 3), PVX-noHC (lane 4), or TEV(lane 5).(C) Gel blot of RNA isolated from leaves as given for (B) and hybrid-ized with a radioactive probe specific for the TEV HC-Pro sequence.

862 The Plant Cell
3A. In these constructs, the expression of the TEV insert isunder the control of the authentic PVX coat protein subge-nomic promoter, and expression of the coat protein is underthe control of an engineered repeated coat protein subge-nomic promoter. The vector PVX-5TEV carries the codingregion of the P1/HC-Pro sequence of TEV (nucleotides 146to 2674). PVX-HC carries only the region encoding HC-Pro(nucleotides 1057 to 2433), with a start codon followed byGCC added at the 5' end and a stop codon at the 3' end ofthe insert so that the mature HC-Pro protein (with two addi-tional N-terminal residues) is made without proteolytic pro-cessing. PVX-noHC carries the same TEV sequences asPVX-HC, except that the translation start site was mutatedfrom AUG to ACG.
These three engineered viruses were used to infect/V/cof/anabenthamiana plants. Protein gel blot analysis indicated thatHC-Pro accumulated in leaves infected systemically by eitherPVX-5TEV or PVX-HC (Figure 3B, lanes 2 and 3, respec-tively) but as expected was undetectable in leaves infectedwith PVX-noHC (Figure 3B, lane 4). Although PVX-noHC didnot express the HC-Pro gene at the protein level, it had notdeleted the sequence from the viral genomic RNA, as shownby gel blot analysis of RNA from systemically infectedleaves. A single genomic RNA was detected by using hy-bridization probes specific either for the TEV HC-Pro se-quence (Figure 3C, lane 4) or for the PVX (+) strand genomicRNA (data not shown). Plants infected with PVX-noHC dis-played mild symptoms (Figure 4, lower left) compared withthe mock-inoculated control plant (Figure 4, upper left). Incontrast, infection of plants with either of the viruses ex-pressing the HC-Pro gene product (PVX-5TEV or PVX-HC)initially caused vein clearing, which was followed by necro-sis of systemically infected leaves by 10 days after inocula-tion and usually killed the plant (Figure 4, upper and lowerright, respectively). Together, these results indicate that ex-pression of the HC-Pro gene product, but not the RNA se-quence itself, is sufficient to induce the increase in PVXpathogenicity. Furthermore, both P1 and P3 coding sequencesare dispensable for the enhanced pathogenicity.
Expression of the TEV P1/HC-Pro Sequence Prolongsthe Accumulation of PVX (-) Strand RNA in Protoplasts
In whole plants, PVX-potyviral synergism is characterized byan approximately threefold increase in the accumulation ofPVX coat protein and (+) strand RNA and M 0-fold increasein the level of PVX (-) strand RNA (Vance, 1991). This dis-proportionate increase in accumulation of PVX (-) strandRNA suggests that synergism involves a change in the regu-lation of viral RNA replication. However, infection of wholeplants is asynchronous and involves both replication andmovement of the virus. Thus, it is also possible that PVX RNAreplication is unaffected and that synergism results solelyfrom a change in the efficiency of viral movement. To deter-mine whether the potyviral sequence affects PVX replication
when viral movement is not a factor, we used the PVX vectorsdescribed above to examine the effect of TEV P1/HC-Pro ex-pression on the kinetics of accumulation of PVX (+) and (-)strand RNAs in tobacco protoplasts.
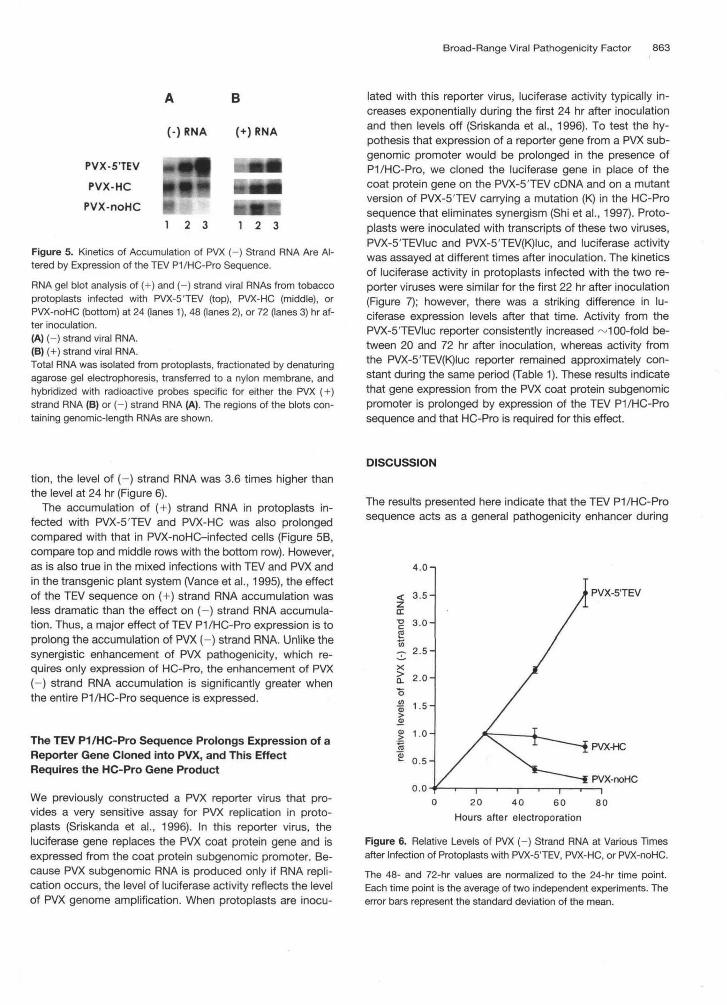
Protoplasts were inoculated by electroporation with tran-scripts of the three engineered PVX cDNAs (Figure 3A, PVX-5'TEV, PVX-HC, and PVX-noHC), and the accumulation of(+) and (-) strand RNAs was assayed by RNA gel blot anal-ysis at different times after inoculation. Because determiningkinetics involves comparing RNA levels only within the sameinoculated protoplast culture and not between protoplastcultures infected with different PVX vectors, the analysis is in-dependent of infection efficiency. The kinetics of (-) strandRNA accumulation in protoplasts infected with PVX-noHCwere similar to those previously reported for the parentalPVX strain (Sriskanda et al., 1996), with the level of (-)strand RNA peaking at 24 hr after inoculation and then de-clining to ~0.2 times the peak level over the next 48 hr, asshown by the RNA gel blot in Figure 5A (bottom row) andgraphically in Figure 6. In protoplasts infected with PVX-HC,the level of (-) strand RNA declined only slightly during thesame period to a level ~0.8 times the 24-hr peak (Figure 5A,middle row, and Figure 6). The most striking result, however,was obtained in protoplasts infected with PVX-5TEV. In thiscase, the level of PVX (-) strand RNA increased over the en-tire 72-hr period (Figure 5A, top gel); at 72 hr postinocula-
Figure 4. Enhanced Pathogenicity of PVX Expressing the HC-ProGene Product.
(Upper left) Mock-inoculated N. benthamiana plant.(Upper right) N. benthamiana plant infected with PVX-5TEV.(Lower left) N. benthamiana plant infected with PVX-noHC.(Lower right) W. benthamiana plant infected with PVX-HC.
Broad-Range Viral Pathogenicity Factor 863
B
(-)RNA (+)RNA
PVX-5'TEVPVX-HC
PVX-noHC1 2 3 2 3
Figure 5. Kinetics of Accumulation of PVX (-) Strand RNA Are Al-tered by Expression of the TEV P1/HC-Pro Sequence.
RNA gel blot analysis of (+) and (-) strand viral RNAs from tobaccoprotoplasts infected with PVX-5'TEV (top), PVX-HC (middle), orPVX-noHC (bottom) at 24 (lanes 1), 48 (lanes 2), or 72 (lanes 3) hr af-ter inoculation.(A) (-) strand viral RNA.(B) (+) strand viral RNA.Total RNA was isolated from protoplasts, fractionated by denaturingagarose gel electrophoresis, transferred to a nylon membrane, andhybridized with radioactive probes specific for either the PVX (+)strand RNA (B) or (-) strand RNA (A). The regions of the blots con-taining genomic-length RNAs are shown.
lated with this reporter virus, luciferase activity typically in-creases exponentially during the first 24 hr after inoculationand then levels off (Sriskanda et al., 1996). To test the hy-pothesis that expression of a reporter gene from a PVX sub-genomic promoter would be prolonged in the presence ofP1/HC-Pro, we cloned the luciferase gene in place of thecoat protein gene on the PVX-5'TEV cDNA and on a mutantversion of PVX-5'TEV carrying a mutation (K) in the HC-Prosequence that eliminates synergism (Shi et al., 1997). Proto-plasts were inoculated with transcripts of these two viruses,PVX-5'TEVIuc and PVX-5'TEV(K)luc, and luciferase activitywas assayed at different times after inoculation. The kineticsof luciferase activity in protoplasts infected with the two re-porter viruses were similar for the first 22 hr after inoculation(Figure 7); however, there was a striking difference in lu-ciferase expression levels after that time. Activity from thePVX-5'TEVIuc reporter consistently increased ~100-fold be-tween 20 and 72 hr after inoculation, whereas activity fromthe PVX-5TEV(K)luc reporter remained approximately con-stant during the same period (Table 1). These results indicatethat gene expression from the PVX coat protein subgenomicpromoter is prolonged by expression of the TEV P1/HC-Prosequence and that HC-Pro is required for this effect.
tion, the level of (-) strand RNA was 3.6 times higher thanthe level at 24 hr (Figure 6).
The accumulation of (+) strand RNA in protoplasts in-fected with PVX-5TEV and PVX-HC was also prolongedcompared with that in PVX-noHC-infected cells (Figure 5B,compare top and middle rows with the bottom row). However,as is also true in the mixed infections with TEV and PVX andin the transgenic plant system (Vance et al., 1995), the effectof the TEV sequence on (+) strand RNA accumulation wasless dramatic than the effect on (-) strand RNA accumula-tion. Thus, a major effect of TEV P1/HC-Pro expression is toprolong the accumulation of PVX (-) strand RNA. Unlike thesynergistic enhancement of PVX pathogenicity, which re-quires only expression of HC-Pro, the enhancement of PVX(-) strand RNA accumulation is significantly greater whenthe entire P1/HC-Pro sequence is expressed.
The TEV P1/HC-Pro Sequence Prolongs Expression of aReporter Gene Cloned into PVX, and This EffectRequires the HC-Pro Gene Product
We previously constructed a PVX reporter virus that pro-vides a very sensitive assay for PVX replication in proto-plasts (Sriskanda et al., 1996). In this reporter virus, theluciferase gene replaces the PVX coat protein gene and isexpressed from the coat protein subgenomic promoter. Be-cause PVX subgenomic RNA is produced only if RNA repli-cation occurs, the level of luciferase activity reflects the levelof PVX genome amplification. When protoplasts are inocu-
DISCUSSION
The results presented here indicate that the TEV P1/HC-Prosequence acts as a general pathogenicity enhancer during
4.0-
3.5-
3.0-
2.5-
2.0-
1.5-
1.0-
0.5-
0.0
PVX-5TEV
0 20 40 60 80
Hours after electroporation
Figure 6. Relative Levels of PVX (-) Strand RNA at Various Timesafter Infection of Protoplasts with PVX-5'TEV, PVX-HC, or PVX-noHC.The 48- and 72-hr values are normalized to the 24-hr time point.Each time point is the average of two independent experiments. Theerror bars represent the standard deviation of the mean.

864 The Plant Cell
108 7
1 0 ’ 1
h
E 8 1 0 6 j Y
10‘ 4- O 20 40 60 80 100 120 140
Hours after electroporation
Figure 7. Time Course of Luciferase Activity in Protoplasts lnfected with PVX Reporter Viruses.
PVX reporter viruses expressed the TEV PVHC-Pro sequence (5’TEV) ora mutant version of that sequence (5’TEV(K)) that encodes an altered HC-Pro that fails to support synergism.
infection by viruses from three different groups (PVX, potex- virus; TMV, tobamovirus; CMV, cucumovirus). Because each of these viruses is capable of inducing synergistic dis- ease in mixed infections with a member of the potyvirus group, this result supports the hypothesis that all such poty- virus-associated synergistic diseases, which occur in an evolutionarily diverse range of host plants and involve inter- actions with many viral groups, are mediated by the same potyviral sequence. The fact that expression of this se- quence alters disease induction by each of these unrelated heterologous viruses suggests that it affects a step in viral infection that is common to all of these viruses and thus of general significance.
Our results using PVX as a vector to express TEV se- quences indicate that the processed form of HC-Pro is both necessary and sufficient for enhancement of PVX pathoge- nicity and that it also plays an essential role in the transacti- vation of PVX replication. However, additional sequences from the Pl/HC-Pro region are required for the dramatic ef- fect on the accumulation of PVX (-) strand RNA in proto- plasts. We can obtain clues for how HC-Pro and P1 might enhance the pathogenicity and transactivate the replication of heterologous viruses by examining the functions they per- form in the TEV infection process.
Both HC-Pro and P1 are multifunctional proteins. P1 has proteinase activity that cleaves the potyviral polyprotein, creating the C terminus of P1 and the N terminus of HC-Pro (Verchot et al., 1991). P1 also functions in trans as an acces- sory factor for genome replication (Verchot and Carrington, 1995) and has RNA binding activity (Brantley and Hunt, 1993). One possibility is that P1 plays a direct role in syner- gism, perhaps analogous to its role as an accessory factor in TEV replication. Alternatively, it might enhance synergism only indirectly by producing the authentic HC-Pro N termi- nus by means of its proteinase activity.
HC-Pro has at least three functional domains: an N-termi- na1 domain required for aphid transmission; a central domain involved in pathogenicity, RNA replication, and leaf-to-leaf movement of the virus through the phloem; and a C-terminal domain required for autoproteolytic processing of the HC-Pro C terminus (reviewed in Maia et al., 1996). The central domain of HC-Pro is of particular interest because it is involved in the regulation of both pathogenicity and RNA replication of potyviruses (Atreya et al., 1992; Atreya and Pirone, 1993; Klein et al., 1994; Cronin et al., 1995; Kasschau et al., 1997), and these are the characteristics that are altered in the het- erologous virus during synergism. Mutations within the cod- ing region for the central domain of TEV HC-Pro eliminate the ability of the sequence to mediate PVX-potyviral syner- gistic disease in transgenic plants (Shi et al., 1997). Further- more, mutations within the central region result in a premature shutoff of TEV RNA amplification, suggesting that HC-Pro may function to prolong potyviral RNA replication (Kasschau et al., 1997). Because expression of Pl/HC-Pro prolongs PVX (-) strand RNA accumulation, it is possible that the ba-
Table 1. Luciferase Activity (Counts per Minute) in Protoplasts lnfected with PVX Reporter Viruses
Experiment 1 Experiment 2 Experiment 3 Experiment 4
Virus 22 Hr 70 Hr 23 Hr 73 Hr 21 Hr 70 Hr 21 Hr 70 Hr
PVX-5’TEVluc 3.6 x 105 3.7 x 107 1.1 x 105 5.0 x 106 5.3 x 103 7.9 x 105 9.2 x 104 8.7 x 106 normalizeda 1 103 1 45 1 149 1 95
normalizeda 1 0.98 1 0.83 1 0.76 1 1.19 PVX-5’TEV(K)luc 4.6 x I05 4.5 x I05 2.3 X 104 1.9 x 104 1.2 X I05 9.1 x 104 1.6 x 105 1.9 X 105
aln each infection, the counts per minute at the two times are normalized to the earlier time.

Broad-Range Viral Pathogenicity Factor 865
sis of potyvirus-associated synergism is the ability of HC- Pro to prolong replication not only of potyviruses but also of a broad range of heterologous viruses.
Although our results suggest that synergism involves transactivation of viral replication, they do not rule out the possibility that the TEV sequence also facilitates viral move- ment. In fact, because the central domain of HC-Pro is re- quired for both movement and replication of the potyvirus (Cronin et al., 1995; Kasschau et al., 1997) and the same do- main is also required for the synergistic effect on PVX (Shi et al., 1997), a role in movement in addition to the role in repli- cation remains quite possible.
Expression of the TEV Pl/HC-Pro region affects TEV as well as a broad range of heterologous viruses. Therefore, we propose that synergism may occur via an indirect mecha- nism involving an interaction of the TEV-encoded proteins with one or more host factors common to the different viral infections rather than a direct mechanism involving interac- tions with different RNAs or proteins from three heterolo- gous viruses.
Two different indirect mechanisms could explain transac- tivation of viral replication by Pl/HC-Pro. The TEV sequence might increase the activity or availability of a positive regulator of viral replication that affects both TEV and heterologous vi- ruses. The stimulation of one virus by a host factor induced by another virus has been shown in mixed infections with hu- man cytomegalovirus and human immunodeficiency virus-1 , in which human cytomegalovirus can induce expression of host transcription factor NF-KB, which then activates human immunodeficiency virus-1 replication (Chinnadurai, 1991 ; Kim et al., 1996).
Alternatively, the potyviral sequence might interfere with the activity or availability of a negative regulator of viral repli- cation, perhaps part of a host defense system that normally limits viral accumulation. Because HC-Pro enhances the ac- cumulation of a broad range of viruses and acts at the level of viral replication, the putative host defense system would necessarily be general in nature and act at the single-cell level. A host system consistent with these requirements has been proposed as the underlying mode of action in sense RNA-mediated virus resistance in transgenic plants (Lindbo et al., 1993; Smith et al., 1994; Mueller et al., 1995; Baulcombe, 1996). In this model, an RNA targeting system is activated by high-leve1 expression of a transgene that contains viral sequences. Once activated, the system rapidly destroys the specific viral RNA target, whether the RNA is expressed from the transgene or from a viral RNA template during viral replication (Dougherty et al., 1994; Dougherty and Parks, 1995; Mueller et al., 1995; English et al., 1996; Goodwin et al., 1996). The same cellular system is also thought to be in- volved in post-transcriptional gene silencing of nonviral transgenes in plants (Ingelbrecht et al., 1994; de Carvalho Niebel et al., 1995; Baulcombe, 1996). It is possible that syn- ergism impacts this RNA targeting system as well. In this case, the Pl/HC-Pro sequence might interfere with the in- duction or the action of the pathway and thus allow a broad
range of viruses to overstep the normal host-imposed limits of RNA accumulation.
METHODS
Viruses and Plants
The virus isolates used in these experiments are as follows: the com- mon strain of tobacco mosaic virus (TMV) (ATCC PV-135) and cu- cumber mosaic virus (CMV-S), which was the kind gifl of S. Scott (Clemson University, Clemson, SC). The U-6B and vector-only con- trol plants are transgenic lines in Nicotiana fabacum cv Havana 425 and have been previously described (Carrington et al., 1990). Engi- neered potato virus X (PVX) viruses were prepared by infecting to- bacco protoplasts with in vitro transcripts of the corresponding infectious cDNAs. Experiments using these engineered viruses were performed in N. benfhamiana.
lnoculation and Sampling
Transgenic lines U-6B and vector-only control plants were mechani- cally inoculated with purified CMV or TMV at a concentration of 0.5 mg/mL. All plants were germinated and grown in an insect-free growth chamber, and plants for a given experiment were of the same size and developmental stage at the time of inoculation. Plants in- fected with TMV were incubated at 30 to 32°C to allow systemic spread of the virus, because N. f. Havana 425 carries the N gene, which mediates a hypersensitive response and localizes TMV at tem- peratures <28"C (Whitham et al., 1994). Total nucleic acid was iso- lated from equivalent systematically infected leaves of the U-6B line and vector-only control plants by using a guanidine-HC1 procedure (Vance, 1991). Levels of viral RNAs were determined by RNA gel blot analysis, as described below.
lnfectious Viral cDNAs
An infectious cDNA clone (pTXS; Kavanagh et al., 1992) of the UK-3 strain of PVX was the kind gift of D. Baulcombe (The Sainsbuty Lab- oratoty, Norwich, UK) and served as the starting point for construc- tion of infectious PVX cDNAs carlying foreign inserts. To construct PVX-HC, the tobacco etch virus (TEV) HC-Pro gene was amplified from TEV cDNA clone pTL7SN.3-0027 (Verchot et al., 1991) by using polymerase chain reaction (PCR) with high-fidelity Vent polymerase (New England Biolabs, Beverly, MA). Two PCR primers were used: one was complementary to the 3' proximal region of the HC-Pro coding region, but with an added translation stop codon (TGA) fol- lowed by a sequence specifying an Xhol restriction enzyme site. The other primer was complementary to the 5' proximal portion of the proteolytically processed HC-Pro, but with 12 added upstream nu- cleotides, GAUAUCAUGGCC, encoding an EcoRV restriction site for cloning purposes and a translation start site followed by an alanine codon to provide a good context for translational initiation in plants (Lutcke et al., 1987).
Thus, translation of the sequence encoding HC-Pro on PVX-HC produces a protein with two extra N-terminal amino acids (methionine

866 The Plant Cell
and alanine) compared with authentic HC-Pro. The PCR fragment was cut with EcoRV and Xhol and ligated into EcoRV-Xhokut pTXS-luc, a derivative of pTXS (Sriskanda et al., 1996), to put the TEV HC-Pro gene under control of the PVX coat protein promoter in place of the coat protein gene. To put the coat protein gene back into the construct, a DNA fragment containing 40 nucleotides of coat protein subgenomic promoter sequence, the PVX coat protein coding se- quence, the entire PVX 3‘ untranslated region (UTR) plus poly(A) tail, and vector sequence up to the Sacl site was amplified from pTXS by using the amplification primers to add a Sal1 site upstream of the PVX coat protein promoter and replace the Spel site in the vector with an Mlul site. This amplified Sall-Sacl fragment was ligated into the pre- vious construct at the Xhol and Sacl sites to produce pTXS-HC.
The Spel site, which is located at the end of the PVX sequence in pTXS and used to linearize that plasmid for in vitro transcription, was replaced with an Mlul site in pTXS-HC, because the HC-Pro coding sequence contains two Spel sites. Linearization of pTXS-HC with Mlul and transcription from the T7 promoter result in an infectious RNA transcript with 5’ and 3’ termini identical to those of authentic PVX genomic RNA.
Thus, the PVX-HC cDNA clone has the original pTXS sequence up to the EcoRV site, followed by the TEV HC-Pro coding region with added translation start and stop sites, followed by the pTXS se- quence comprising the region 40 nucleotides upstream of the coat protein AUG through the coat protein coding region and including the Sacl site in the downstream polylinker of the vector. Because both HC-Pro and PVX coat protein are expressed from the coat pro- tein subgenomic promoter, nucleotides 5620 to 5660 of pTXS are duplicated in the construct.
The plasmid pTXS-noHC was constructed in exactly the same manner as pPVX-HC, except that the PCR primer used to amplify TEV HC-Pro had ACG in place of the translation start site ATG to eliminate translation of the HC-Pro gene. pTXS-5’TEV was produced from pTXS-HC by replacing the HC-Pro gene and most of the coat protein gene with a PCR-amplified EcoRV-Xhol fragment containing nucleotides 146 to 2674 of the TEV genome. The PVX sequence specifying the coat protein promoter, coat protein coding sequence, and 3’ UTR was then replaced as described for pTXS-HC. The orga- nization of the infectious viral RNAs derived by in vitro transcription of these cDNAs is shown in Figure 3A.
The PVX reporter virus PVX-5’TEVluc was constructed by using PCR to amplify the luciferase encoding region of pTXS-luc (Sriskanda et al., 1996) and then digesting the PCR-amplitied fragment with Sal1 and Xhol and cloning it into the Xhol site of a precursor of pTXS-5’TEV lacking most of the PVX coat protein coding sequence. The K muta- tion was introduced by replacing an interna1 Spel fragment within the HC-Pro coding region of pTXS-5’TEVluc with the corresponding fragment from a TEV cDNA containing a nine-nucleotide insertion that introduced an Ncol site and resulted in the insertion of the amino acid triplet Thr-Met-Ala immediately after amino acid 426 of the e&- pressed TEV polyprotein (Shi et al., 1997).
Protoplast lnfection System
Protoplasts from NT-I tobacco suspension culture cells (An, 1985) were infected with in vitro transcripts of the infectious engineered PVX cDNAs. Capped transcripts were synthesized using the Ri- bomax T7 transcription kit (Promega) with the GTP concentration re- duced to 3.75 mM and the addition of 3.75 mM CAP analog (New England Biolabs). Protoplasts were prepared from NT-1 cells at the
logarithmic stage of growth and electroporated with viral transcripts exactly as previously described (Sriskanda et al., 7996). The levels of PVX (+) and (-) strand RNAs at various times after electroporation were determined by RNA gel blot analysis with strand-specific probes, as described below. Luciferase activity from protoplasts in- fected with either PVX-5‘TEVluc or PVX-S’TEV(K)luc was assayed as previously described (Sriskanda et al., 1996) by using a luciferase as- say kit (Promega). Luciferase activity was determined for cell lysates from -30,000 protoplasts.
RNA and Protein Gel Blot Analyses
Total nucleic acids were isolated from leaves or protoplasts and an- alyzed by using RNA gel blots exactly as previously described (vance, 1991). Radioactive probes complementary to the entire PVX or TMV (+) strand RNA were generated by using reverse tran- scriptase, hexanucleotide random primers, and either PVX or TMV genomic RNA isolated from purified virus. Probe specific to the (-) strand of PVX was generated by limited base hydrolysis (10 min at 65°C in Tris-HCI, pH 9.5) of purified PVX genomic RNA, followed by ethanol precipitation and 5‘ end labeling with T4 polynucleotide ki- nase and y3*P-ATP (Maizels, 1976). Specificity of the PVX (-) strand probe was confirmed by its failure to hybridize to a sample contain- ing microgram amounts of PVX genomic RNA included on the blot (vance, 1991). Radioactive transcripts specific for CMV viral RNAs were generated by in vitro transcription of a CMV cDNA clone from the T7 promoter to produce a 200-nucleotide transcript complemen- tary to the 3‘ proximal region common to CMV viral RNAs 1, 2,3, and 4 (Rizzo and Palukaitis, 1990). Radioactive transcripts complemen- tary to the (+) strand of CMV satellite RNA were generated by in vitro transcription of a cDNA clone (ATCC 451 25) from the T7 promoter. The levels of viral RNAs on RNA gel blots were quantitated by densi- tometric scanning of the x-ray film (at the appropriate exposure to be within the linear range of the film) by using a Microscan 1 O00 scanner (Technology Resources, Inc., Nashville, TN).
Levels of HC-Pro in systemically infected leaves were assayed by protein gel blot analysis. Total protein was extracted, quantitated, separated by SDS-PAGE (vance, 1991), and transferred to polyvi- nylidene difluoride membrane (Bio-Rad) by using a Bio-Rad mini- trans-blot apparatus at 80 V for 1 hr. Rabbit polyclonal antiserum specific to TEV HC-Pro (Carrington et al., 1990) was used as primary antiserum, and immunoreactive proteins were visualized by using a goat anti-rabbit antiserum conjugated to alkaline phosphatase (Bio-Rad).
ACKNOWLEDGMENTS
We thank Drs. Lewis Bowman and William Dawson for helpful dis- cussions and critical reading of the manuscript and Trent Smith for help with figures. This work was supported by Grant No. 9304447 from the U.S. Department of Agriculture (USDA) Biotechnology Risk Assessment Program, Grant No. 9501 106 from the USDA National Research lnitiative (NRI) Competitive Grants Program to V.B.V., and Grant No. 95-37303-1867 from the USDA NRI Competitive Grants Program to J.C.C.
Received January 30, 1997; accepted March 31,1997.

Broad-Range Viral Pathogenicity Factor 867
REFERENCES
An, G. (1985). High efficiency transformation of cultured tobacco cells. Plant Physiol. 79, 568-570.
Anjos, J.R., Jarlfors, U., and Ghabrial, S.A. (1992). Soybean mosaic potyvirus enhances the titer of two comoviruses in dually infected soybean plants. Phytopathology 82, 17-23.
Atreya, C.D., and Pirone, T.P. (1993). Mutational analysis of the helper component-proteinase gene of a potyvirus: Effects of amino acid substitutions, deletions, and gene replacement on vir- ulence and aphid transmissibility. Proc. Natl. Acad. Sci. USA 90,
Atreya, C.D., Atreya, P.L, Thornbury, D.W., and Pirone, T.P. (1 992). Site-directed mutations in the potyvirus HC-PRO gene affect helper component activity, virus and symptom expression in infected tobacco plants. Virology 191, 106-1 11.
Baulcombe, D.C. (1996). Mechanisms of pathogen-derived resis- tance to viruses in transgenic plants. Plant Cell8, 1833-1 844.
Brantley, J.D., and Hunt, A.G. (1993). The N-terminal protein of the polyprotein encoded by the potyvirus tobacco vein mottling virus is an RNA-binding protein. J. Gen. Virol. 74, 1157-1162.
Calendar, R. (1 986). Viral transactivation. Bio/Technology 4,
Calvert, L.A., and Ghabrial, S.A. (1983). Enhancement by soybean mosaic virus of bean pod mottle virus titer in doubly infected soy- bean. Phytopathology 73,992-997.
Carrington, J.C., Freed, D.D., and Oh, C.-S. (1990). Expression of potyviral polyproteins in transgenic plants reveals three pro- teolytic activities required for complete processing. EMBO J. 9,
Chinnadurai, G. (1 991). Modulation of HIV-enhancer activity by het- erologous agents: A minireview. Gene 101,165-170.
Clark, R.L., Hill, J.H., and Ellis, M.D. (1980). Tomato scorch, a new virus disease of tomatoes. Phytopathology 70, 131-134.
Cronin, S., Verchot, J., Haldeman-Cahill, M.C., Schaad, M.C., and Carrington, J.C. (1 995). Long-distance movement factor: A transpor3 function of the potyvirus helper component proteinase. Plant Cell7,549-559.
de Carvalho Niebel, F., Frendo, P., Van Montagu, M., and Cornelissen, M. (1 995). Post-transcriptional cosuppression of p-1,3-glucanase genes does not affect accumulation of transgene nuclear mRNA. Plant Cell7,347-358.
Dougherty, W.G., and Parks, T.D. (1995). Transgenes and gene suppression: Telling us something new? Curr. Opin. Cell Biol. 7, 399-405.
Dougherty, W.G., Lindbo, J.L., Smith, H.A., Parks, T.D., Swaney, S., and Proebsting, W.M. (1994). RNA-mediated virus resistance in transgenic plants: Exploitation of a cellular pathway possibly involved in RNA degradation. MOI. Plant-Microbe Interact. 7,
English, J.J., Mueller, E., and Baulcombe, D.C. (1 996). Suppres- sion of virus accumulation in transgenic plants exhibiting silencing of nuclear genes. Plant Cell8, 179-1 88.
Goldberg, K.-B., and Brakke, M.K. (1987). Concentration of maize chlorotic mottle virus is increased in mixed infections with maize dwarf mosaic virus, strain B. Phytopathology 77, 162-1 67.
11 91 9-1 1923.
1074-1 077.
1347-1 353.
544-552.
Goodman, R.M., and Ross, A.F. (1974a). Enhancement of potato virus X synthesis in doubly infected tobacco occurs in doubly infected cells. Virology 58, 16-24.
Goodman, R.M., and Ross, A.F. (1974b). Enhancement by potato virus Y of potato virus X synthesis in doubly infected tobacco depends on the timing of invasion by the viruses. Virology 58,
Goodwin, J., Chapman, K., Swaney, S., Parks, T.D., Wernsman, E.A., and Dougherty, W.G. (1996). Genetic and biochemical dis- section of transgenic RNA-mediated virus resistance. Plant Cell8,
Ingelbrecht, I., Van Houdt, H., Van Montagu, M., and Depicker, A. (1 994). Posttranscriptional silencing of reporter transgenes in tobacco correlates with DNA methylation. Proc. Natl. Acad. Sci.
Kassachau, K.D., Cronin, S., and Carrington, J.C. (1 997). Genome amplification and long-distance movement functions associated with the central domain of tobacco etch potyvirus helper compo- nent-proteinase (HC-Pro). Virology 228,251 -262.
Kavanaugh, T., Goulden, M., Santa Cruz, S., Chapman, S., Barker, I., and Baulcombe, D. (1992). Molecular analysis of a resistance- breaking strain of potato virus X. Virology 189,609-61 7.
Khan, M.A., and Demski, J.W. (1982). ldentification of turnip mosaic and cauliflower mosaic viruses naturally infecting collards. Plant Dis. 66,253-256.
Kim, S., Yu, S., and Kim, V.N. (1996). Essential role of NF-kappa B in transactivation of the human immunodeficiency virus long ter- minal repeat by human cytomegalovirus 1 E1 protein. J. Gen. Virol.
Klein, P.G., Klein, R.R., Rodrigues-Cerezo, E., Hunt, A.G., and Shaw, J.G. (1994). Mutational analysis of the tobacco vein mot- tling virus genome. Virology 204, 759-769.
Lindbo, J.A., Silva-Rosales, L., Proebsting, W.M., and Dougherty, W.G. (1993). lnduction of a highly specific antiviral state in trans- genic plants: lmplications for regulation of gene expression and virus resistance. Plant Cell 5, 1749-1 759.
Lutcke, H.A., Chow, K.C., Mickel, F.S., Moss, K.A., Kern, H.F., and Scheele, G.A. (1987). Selection of AUG initiation codons dif- fers in plants and animals. EMBO J. 6, 43-48.
Maia, I.G., Haenni, A.-L., and Bernardi, F. (1996). Potyviral HC- Pro: A multifunctional protein. J. Gen. Virol. 77, 1335-1341.
Maizels, N. (1976). Dictyostelium 17S, 25S, and 5s rDNAs lie within a 38,000 base pair repeated unit. Cell9,431-438.
Matthews, R.E.F. (1991). Plant Virology, 3rd ed. (San Diego, CA: Academic Press).
Mueller, E., Gilbert, J.E., Davenport, G., Brigneti, G., and Baulcombe, D.C. (1995). Homology-dependent resistance: Transgenic virus resistance in plants related to homology depen- dent gene silencing. Plant J. 7, 1001-1013.
Pio-Ribeiro, G., Wyatt, S.D., and Kuhn, C.W. (1978). Cowpea stunt: A disease caused by a synergistic interaction of two viruses. Phytopathology 68, 1260-1 265.
Poolpol, P., and Inouye, T. (1986). Enhancement of cucumber mosaic virus multiplication by zucchini yellow mosaic virus in doubly infected cucumber plants. Ann. Phytopathol. SOC. Jpn. 52,2230.
263-271.
95-1 05.
USA 91,10502-1 0506.
77,83-91.

868 The Plant Cell
Riuo, T.M., and Palukaitis, P. (1990). Construction of full-length cDNA clones of cucumber mosaic virus RNAs 1, 2, and 3: Genera- tion of infectious RNA transcripts. MOI. Gen. Genet. 222, 249-256.
Rochow, W.F., and Ross, A.F. (1955). Virus multiplication in plants doubly infected by potato viruses X and Y. Virology 1, 10-27.
Shi, X.M., Miller, H., Verchot, J., Carrington, J.C., and Vance, V.B. (1 997). Mutations in the helper component-proteinase (HC- Pro) coding region eliminate potato virus Wpotyviral synergism. Virology, in press.
Smith, H.A., Swaney, S.L., Parks, T.D., Wernsman, E.A., and Dougherty, W.G. (1 994). Transgenic plant virus resistance medi- ated by nontranslatable sense RNAs: Expression, regulation, and fate of nonessential RNAs. Plant Cell6, 1441-1453.
Sriskanda, V.S., Pruss, G., Ge, X., and Vance, V.B. (1996). An eight nucleotide sequence in the potato virus X 3'-UTR is required for both host protein binding and vira1 multiplication. J. Virol. 70, 5266-5271.
Vance, V.B. (1991). Replication of potato virus X RNA is altered in coinfections with potato virus Y. Virology 182, 486-494.
Vance, V.B., Berger, P.H., Carrington, J.C., Hunt, A.G., and Shi, X.M. (1995). 5' proximal potyviral sequences mediate potato virus Wpotyviral synergistic disease in transgenic tobacco. Virology 206,583-590.
Verchot, J., and Carrington, J.C. (1 995). Evidence that the potyvi- rus P1 proteinase functions in trans as an accessory factor for genome amplification. J. Virol. 69, 3668-3674.
Verchot, J., Koonin, E.V., and Carrington, J.C. (1991). The 35-kDa protein from the N terminus of the potyviral polyprotein functions as a third virus-encoded proteinase. Virology 185, 527-535.
Whitham, S., Dinesh-Kumar, S.P., Choi, D., Hehl, R., Corr, C., and Baker, B. (1994). The product of the tobacco mosaic virus resistance gene N: Similarity to Toll and the interleukin-1 receptor. Cell78,1101-1115.

DOI 10.1105/tpc.9.6.859 1997;9;859-868Plant Cell
G Pruss, X Ge, X M Shi, J C Carrington and V Bowman Vancetransactivates replication of heterologous viruses.
Plant viral synergism: the potyviral genome encodes a broad-range pathogenicity enhancer that
This information is current as of July 6, 2018
Permissions X
https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists