pleasant or unpleasant: attentional modulation of odor perception
TRANSCRIPT

Pleasant or Unpleasant: Attentional Modulationof Odor Perception
Jelena Djordjevic & Julie A. Boyle &
Marilyn Jones-Gotman
Received: 4 October 2011 /Accepted: 24 November 2011 /Published online: 20 January 2012# Springer Science+Business Media, LLC 2012
Abstract Using positron emission tomography, we investi-gated whether regional brain activations differ as a functionof attending to pleasant versus unpleasant components ofodors. There were two experimental (attention to pleasant-ness and attention to unpleasantness) and one control (base-line) condition. The stimuli presented during the twoexperimental conditions were exactly the same (three binarymixtures, each consisting of one pleasant and one unpleas-ant compound), but the affective property to which partic-ipants’ attention was directed was different: They indicatedwith a mouse click whether each stimulus contained a pleas-ant (during attention to pleasantness) or unpleasant (duringattention to unpleasantness) odor. During baseline, odorlessstimuli were presented, and participants pressed the mousebutton randomly after each one. Several brain regions wereinvolved in both types of attention, and these includedventral striatum, right orbitofrontal cortex, and anterior cin-gulate cortex. Subtle differences were also revealed:Attending to pleasantness was associated preferentially witha sensory/perceptual network (piriform cortex and amygdala),whereas attending to unpleasantness engaged a component ofthe attentional (right parietal) network. Thus, we delineateneural substrates of attending to olfactory pleasantness andunpleasantness, some of which are common to both and othersthat are specific to pleasantness or to unpleasantness. Ourresults suggest that the view of the mesolimbic dopaminergicsystem as the reward network that responds selectively topositive reinforcers is somewhat limited: Our findings are
more in keeping with a view of this set of structures as thesalience system of the brain.
Keywords Affective valence . Attention . Functionalneuroimaging . Olfaction
The topic of attention has attracted considerable noticeamong neuroscientists, and this interest has been growingduring recent years (Luck and Vicera 2001; Raz and Buhle2006). The majority of studies examining attention havebeen conducted in the visual, followed by the auditory andtactile domains, while questions of attentional mechanismsin chemical senses have been addressed by only a handful ofstudies. Behavioral studies demonstrated that directing atten-tion toward odors results in faster reaction times and greaterresponse accuracy, suggesting that attention can be directedselectively in the olfactory modality (Spence et al. 2000;Spence et al. 2001a,b). However, Ashkenazi and Marks(2004) showed that selective attention was not effective whenolfactory stimuli were presented retronasally, i.e., through themouth. The use of olfactory event-related potentials (OERPs)has resulted in several important contributions to our under-standing of complex interactions between attention and olfac-tion. Passive smelling versus pressing a button upon each odorpresentation (Pause et al. 1997) and directing attention towardversus away from odors (Geisler and Murphy 2000; Krauel etal. 1998) modify specific OERP components. In a functionalneuroimaging study, Zelano et al. (2005) demonstrated afunctional dissociation of subregions within the piriform cor-tex, such that attending to olfactory stimuli was associatedwith activity in the frontal piriform cortex and olfactorytubercle, but not in the temporal piriform cortex.Similarly, Rolls et al. (2008) showed functional specializationof regions within the orbito-frontal cortex: greater medial
J. Djordjevic (*) : J. A. Boyle :M. Jones-GotmanMontreal Neurological Institute, McGill University,3801 University Street,Montreal H3A 2B4, Québec, Canadae-mail: [email protected]
Chem. Percept. (2012) 5:11–21DOI 10.1007/s12078-011-9107-3

orbitofrontal and pregenual cingulate cortex activations dur-ing attention to odor pleasantness and greater inferior frontalgyrus activations during attention to odor intensity. Otherstudies demonstrated activation of the piriform cortex(Veldhuizen and Small 2011) and orbitofrontal cortex(Veldhuizen and Small 2011; Sabri et al. 2005) whenattending to odors. Plailly et al. (2008) recently showedthat mediodorsal thalamus and the human transthalamicpathway (the indirect projection from piriform to orbito-frontal cortex via thalamus) play an important role inselective attention to odors versus tones. Taken together,these findings show that directing attention toward theolfactory modality has specific behavioral, electrophysi-ological, and neural effects.
However, the neural mechanisms underlying attention tospecific odor properties are poorly understood. In the pres-ent study, we investigate the neural networks involved inattending to affective valence in olfaction. More specifically,we address the question of whether the same or differentbrain regions are involved in attending to odor pleasantnessversus attending to odor unpleasantness. We opted to exam-ine this question using binary mixtures composed of apleasant and an unpleasant odor and comparing neural acti-vations, while participants’ attention was directed towardeither the pleasant or the unpleasant component. Thisallowed us to isolate the effect of attention to one compo-nent or the other because the actual physical stimuli were thesame in both cases.
Materials and Methods
Participants
Twelve healthy volunteers participated in this study: six menand six women (mean age023 years; range, 20–28). Allsubjects reported normal ability to smell, which was con-firmed by an errorless performance on a seven-trial detectiontask: In each trial, three stimuli (two containing only diluantsand one containing a 10% dilution of phenyl ethyl alcohol)were presented, and the task was to indicate which one of thethree contained an odor. Exclusion criteria were respiratoryinfections, allergies leading to nasal congestion, history ofneurological or psychiatric disease, or other conditions asso-ciated with impaired sense of smell. All participants wereright-handed.
Stimuli
In the two experimental conditions of this study, we usedthree binary mixtures, each consisting of one pleasant andone unpleasant odor/compound. The three mixtures andtheir compounds are shown in Table 1.
Concentrations of each compound were selected basedon results of a pilot study (N051). Notably, all individualcompounds were within the same range of intensity (mod-erately strong): The mean ratings of the six compoundsvaried from 5.4 to 7.6 on a rating scale ranging from 0 to10. Importantly, the mean intensity of the three pleasantodors did not differ from the mean intensity of the threeunpleasant odors [M06.6 for pleasant and M06.5 forunpleasant odors, t(df050)00.43, p0 .67]. As expected, theodors that we had designated as pleasant were rated asmore pleasant than those designated unpleasant [M05.6for pleasant and M02.3 for unpleasant odors, t(df050)016.44, p<.001]. Other odor features, such as familiarityand edibility, were not considered during stimulus se-lection, but we note that most odors are ambiguous inquality and are not readily identified as “food odors.”All chosen odors had a moderate trigeminal componentin the concentrations used (Doty et al. 1978).
Each of the six compounds was diluted with propyleneglycol. Water (odorless stimulus) was used for the baselinecondition. All stimuli were presented in 60-ml amber bottlescontaining 10 ml of each binary mixture (obtained by mix-ing 5 ml of each compound) or 10 ml of water.
Procedure
Protocol during scanning This experiment consisted ofthree conditions, two experimental and one control. Thetwo experimental conditions were attention to pleasantness(AttP) and attention to unpleasantness (AttU), and an odor-less baseline (Base) was the control. Each condition was runonce, for a total of three scans, and the order was counter-balanced across participants. About ten minutes prior to theexperimental scans, examples of the relevant compoundswere presented: Isoamyl acetate, linalool, and citral werepresented preceding the AttP condition as examples ofpleasant components, while butyric acid, allyl sulfide andpyridine were introduced before the AttU condition asexamples of the unpleasant components. These odorantswere each presented just once for about 2–3 s, and the
Table 1 Binary mixtures usedin scanning, and their individualcompounds
Binary mixture Unpleasant compound Pleasant compound
BI B: Butyric acid 10% v/v I: Isoamyl acetate 10% v/v
AL A: Allyl sulfide 1% v/v L: Linalool 32% v/v
PC P: Pyridine 2% v/v C: Citral 14% v/v
12 Chem. Percept. (2012) 5:11–21

interval between presentations was about 20 s. An essentialfeature of the experimental design was that the stimulipresented in the two experimental conditions (AttP andAttU) were exactly the same: three binary mixtures. Theonly difference between the two experimental conditionswas that the stimulus feature to which participants attendedwas different. During AttP, the participants’ task was toindicate whether each presented stimulus contained apleasant odor, while during AttU, the task was to indi-cate whether the stimulus contained an unpleasant odor.The response was made by pressing the left mouse key toindicate “yes” and the right mouse key to indicate “no.” Thestimulus order within the experimental scans was the same forall participants to ensure that the physical stimulation pre-sented in those two conditions was exactly the same withinand between participants. The stimulus order was pseudor-andomized so that each mixture was always followed by adifferent mixture. The control (base) condition consisted ofpresentations of odorless water, and participants pressed theleft or right mouse key at random. They were instructed tosniff in exactly the same way during the baseline as during thetwo experimental conditions. Each stimulus was presented for2 s, followed by an interstimulus interval of 8 s. The odorswere presented birhinally by placing the odor bottles under theparticipant’s nose. The accurate timing of odor presen-tation was assured in two ways. First, all participantswere tested by the same experimenter, who practiced the2-s odor presentations before the study and before eachparticipant. Second, her timing was controlled by com-puter, with visual prompts appearing on a computer screenthat was visible to the experimenter but not to participants.The total duration of each scan was 60 s. Task presentationstarted before the scan and continued for about 10–15 s after itended. This was done to ensure that participants would be inthe “task mode” at the beginning of the scan and also to avoidcompletion of stimulation/task presentation before the scanended.
Image acquisition and analysis Positron emission tomogra-phy (PET) scans were obtained using a CTI Siemens ExactECAT HR + scanner operating in a three-dimensional mode.The distribution of rCBF was measured during three 60-sscans using the 15O-labelled water bolus method. MRI scans(1 mm slices) were obtained with a 1.5-T Philips MRIscanner. Each participant’s PET and MRI scans were core-gistered and transformed into the Montreal NeurologicalInstitute standardized (MNI-305) proportional stereotaxicspace that is based on the Talairach and Tournoux atlas(Talairach and Tournoux 1988). Dot, an in-house (MontrealNeurological Institute) software designed for analysis of PETdata, was used for PETsubtraction analyses. PET images wereblurred using a 14-mm Hanning filter and normalized fordifferences in global blood flow. PET data were averaged
across subjects for each scanning condition; subtractions ofinterest yielded mean change image volumes and were con-verted to t-statistic maps. In calculation of t-statistics for eachvoxel, a standard deviation pooled across all voxels was used,yielding a very high (practically infinite) value of degrees offreedom (Worsley et al. 1996). To localize the observed peaks,these statistical maps were superimposed on the average MRIimage of the 12 participants. For the subtractions of interest,the statistical significance of rCBF changes was assessedusing three-dimensional Gaussian random-field theory(Worsley et al. 1996). Significant changes were estab-lished using two threshold values: For the exploratorysearch of the gray matter volume of approximately500 cc (182 resolution elements), the threshold was setat t03.52, corresponding to an uncorrected probabilityof p<.0002. For the directed search within regionspreviously activated by odor perception (piriform cortex,posterior orbitofrontal cortex, and insula), the thresholdwas lowered to t03.00, representing an uncorrected p<0.004.Given that there were three conditions (two experimental andone control), we conducted three subtraction analyses: In two,each experimental condition was contrasted against baseline,and in the third, the two experimental conditions were con-trasted to each other. As direct comparison of two tasks (ratherthan task versus baseline) typically yields sparse findings, thelatter analysis was followed by post hoc volume ofinterest analyses (VOI). We determined two regions ofinterest, namely, piriform cortex and amygdala, andextracted the blood flow values of these VOIs with aradius of 4 and 3 mm, respectively, around the anatomicallydefined coordinates, following which we conducted a t testcomparing mean blood flow values of all participants in theseregions across the two experimental conditions.
We also determined which regions were activated duringboth attention to pleasantness and attention to unpleasant-ness in olfaction, by conducting a conjunction analysis. Infunctional neuroimaging, conjunction refers to the occur-rence of an activity change at the same location in two ormore independent three-dimensional brain images (Worsleyand Friston, 2000). There are different types of conjunctionanalyses. Price and Friston (1997) defined conjunction as asignificant main effect in the absence of any differences orinteractions among the simple effects in two pairs of tasks.Friston et al. (1999) introduced a minimum statistic thatbecame a common test for conjunction in the neuroimagingliterature. Here, we conducted a conjunction of two subtrac-tions: AttP minus Base and AttU minus Base. Conjunctionanalysis was performed using a command from the fmristatprogram (Worsley et al. 2002), a set of matlab toolsdesigned to analyze functional neuroimaging data includingPET. In this application, conjunction analysis yields mini-mum activations that are consistently found in all includedimages, whereas subtraction analysis relies on the average
Chem. Percept. (2012) 5:11–21 13

(Friston et al., 1999). The probability that two t-statisticsfrom two different subtractions (the conjunction analysis)exceed a threshold is lower than the probability that one t-statistic does (subtraction analysis). Therefore, the thresholdfor the conjunction analysis was set at t03.00, p<0.05 (forthe search region of 500 cc, conjunction of twosubtractions).
Results
Behavioral Results During Scanning
Behavioral responses were recorded during each scan.Because the mixtures always contained one pleasant and oneunpleasant compound in the two experimental conditions, thecorrect response was “yes,” as indicated by the left mousebutton press, on every trial (100%). The overall percentage ofcorrect responses across conditions was comparable: 63%correct responses were observed for the pleasant, and 50%for the unpleasant, conditions. As the left mouse button waspressed 64% of the time in the baseline condition, the motoroutput was comparable across conditions.
PET Results
PET results were analyzed with two types of contrast: Eachexperimental condition was compared to the baseline andagainst each other, and a conjunction analysis of the twoexperimental conditions was performed.
In the first subtraction, blood flow during attending topleasantness was compared with the baseline (AttP-Base),Table 2.
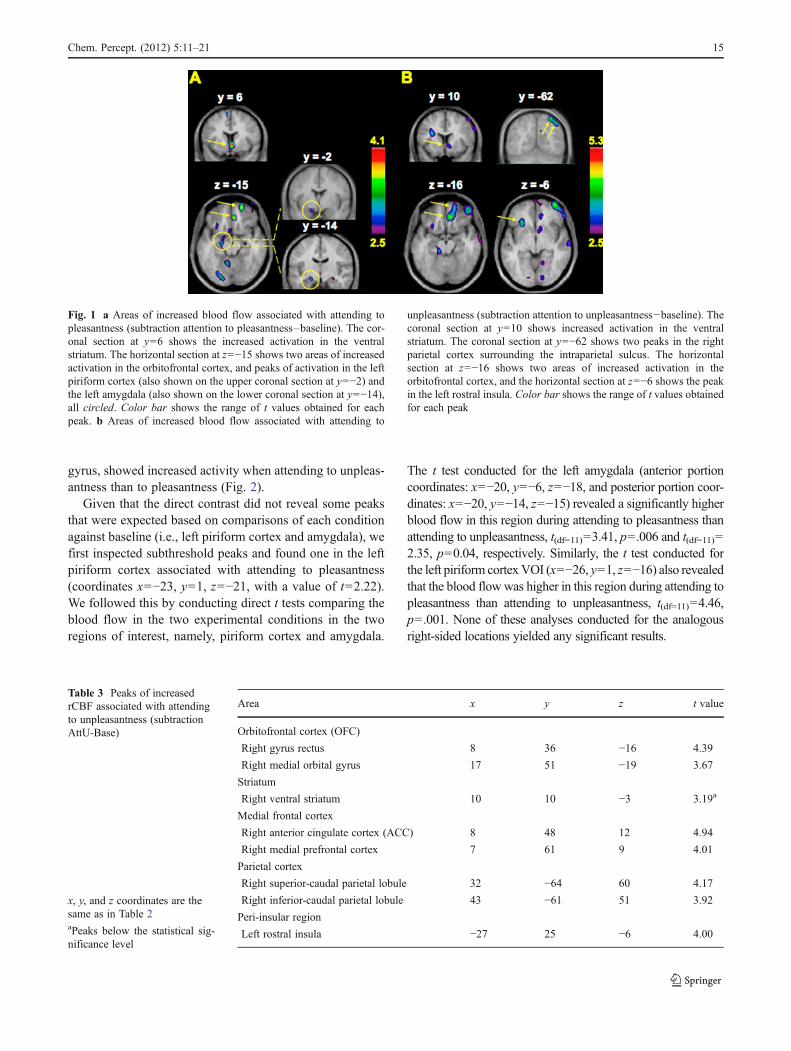
This contrast revealed a subthreshold activation in the leftpiriform cortex. This peak in the piriform cortex was adja-cent to another one in the left amygdala (in fact two peaks inthe left posterior amygdala with nearly identical locationwere found) (Fig. 1a). Two distinct regions of the rightorbitofrontal cortex (OFC) also showed significant activity:a medial (gyrus rectus) and a rostral part of the medialorbital gyrus (Fig. 1a). In addition, activation in the rightventral striatum, including ventral putamen, ventral caudatenucleus, and nucleus accumbens, was seen in this contrast(Fig. 1a).
To determine which regions show increased activity asso-ciated with attending to unpleasantness, Attending to unpleas-antness was contrasted with the baseline (AttU-Base). Thiscontrast also revealed two peaks in the right OFC, one in gyrusrectus and another in the medial orbital gyrus (Table 3 andFig. 1b).
A location in the ventral striatum, similar to the oneobserved in the AttP-Base contrast, was also seen. In addi-tion to these, two peaks of activation were found in the rightparietal cortex caudally: One fell just above, and the otherjust below the intraparietal sulcus. An increase in activationwas also seen in the left rostral insula (Fig. 1b).
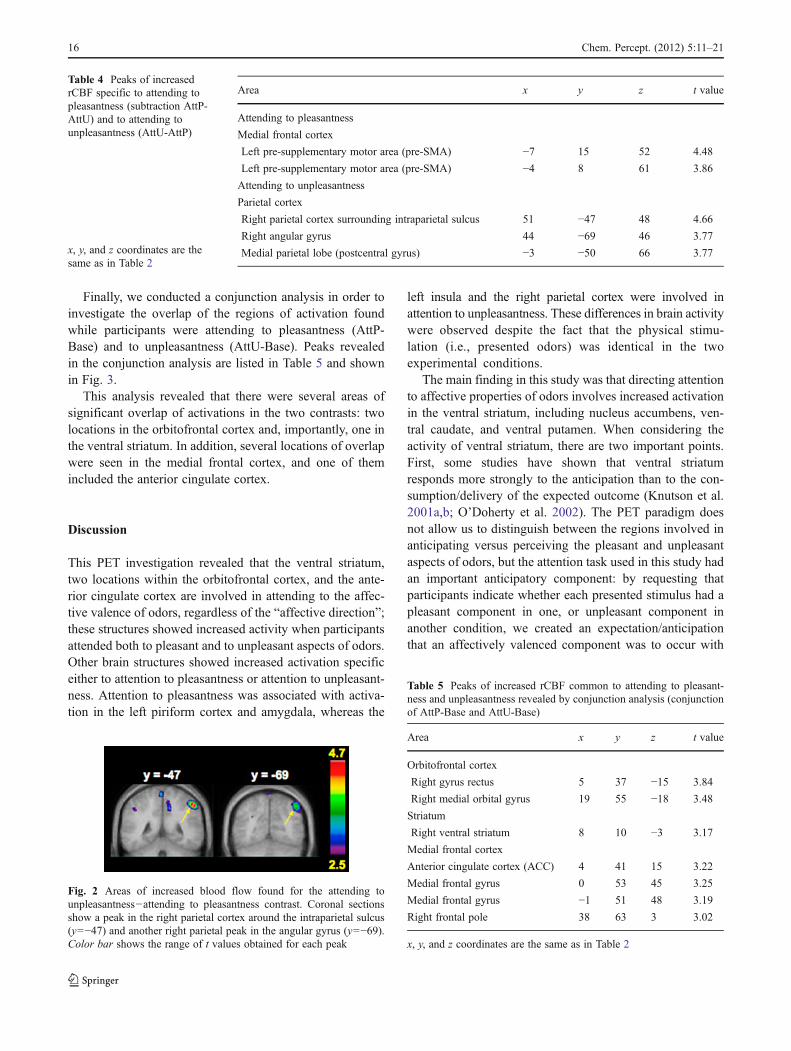
In order to determine which of these activations werespecific to attending to affective aspects of odors and notto smelling odors, we made a direct comparison between thetwo experimental conditions (AttP-AttU and AttU-AttP).There were a few significant peaks in this direct comparison(Table 4). A region in the left frontal medial gyrus (twopeaks) corresponding to the pre-supplementary motor area(pre-SMA) showed higher activity when attending to pleas-antness than when attending to unpleasantness. In contrast,two regions in the right parietal cortex, one in the areaaround the intraparietal sulcus and the other in the angular
Table 2 Peaks of increasedrCBF associated with attendingto pleasantness (subtractionAttP-Base)
x, y, and z coordinates are inMNI space, and reported x, y andz values denote the distance(in millimeters) from the anteriorcommissureaBelow the statistical signifi-cance level
Area x Y z t value
Primary olfactory cortical region
Left piriform cortex −16 −2 −18 2.80a
Orbitofrontal cortex (OFC)
Right gyrus rectus 5 37 −15 3.84
Right medial orbital gyrus 20 58 −16 3.67
Striatum
Right ventral striatum 4 6 −3 4.06
Medial frontal cortex
Left pre-supplementary motor area (pre-SMA) −8 10 63 3.58
Amygdala
Left amygdala −20 −14 −15 3.12
Left amygdala −20 −16 −15 3.12
Right amygdala 27 −18 −15 2.71a
Occipital cortex
Left extrastriate cortex −20 −85 −9 3.74
14 Chem. Percept. (2012) 5:11–21
gyrus, showed increased activity when attending to unpleas-antness than to pleasantness (Fig. 2).
Given that the direct contrast did not reveal some peaksthat were expected based on comparisons of each conditionagainst baseline (i.e., left piriform cortex and amygdala), wefirst inspected subthreshold peaks and found one in the leftpiriform cortex associated with attending to pleasantness(coordinates x0−23, y01, z0−21, with a value of t02.22).We followed this by conducting direct t tests comparing theblood flow in the two experimental conditions in the tworegions of interest, namely, piriform cortex and amygdala.
The t test conducted for the left amygdala (anterior portioncoordinates: x0−20, y0−6, z0−18, and posterior portion coor-dinates: x0−20, y0−14, z0−15) revealed a significantly higherblood flow in this region during attending to pleasantness thanattending to unpleasantness, t(df011)03.41, p0 .006 and t(df011)02.35, p00.04, respectively. Similarly, the t test conducted forthe left piriform cortexVOI (x0−26, y01, z0−16) also revealedthat the blood flowwas higher in this region during attending topleasantness than attending to unpleasantness, t(df011)04.46,p0 .001. None of these analyses conducted for the analogousright-sided locations yielded any significant results.
Fig. 1 a Areas of increased blood flow associated with attending topleasantness (subtraction attention to pleasantness–baseline). The cor-onal section at y06 shows the increased activation in the ventralstriatum. The horizontal section at z0−15 shows two areas of increasedactivation in the orbitofrontal cortex, and peaks of activation in the leftpiriform cortex (also shown on the upper coronal section at y0−2) andthe left amygdala (also shown on the lower coronal section at y0−14),all circled. Color bar shows the range of t values obtained for eachpeak. b Areas of increased blood flow associated with attending to
unpleasantness (subtraction attention to unpleasantness−baseline). Thecoronal section at y010 shows increased activation in the ventralstriatum. The coronal section at y0−62 shows two peaks in the rightparietal cortex surrounding the intraparietal sulcus. The horizontalsection at z0−16 shows two areas of increased activation in theorbitofrontal cortex, and the horizontal section at z0−6 shows the peakin the left rostral insula. Color bar shows the range of t values obtainedfor each peak
Table 3 Peaks of increasedrCBF associated with attendingto unpleasantness (subtractionAttU-Base)
x, y, and z coordinates are thesame as in Table 2aPeaks below the statistical sig-nificance level
Area x y z t value
Orbitofrontal cortex (OFC)
Right gyrus rectus 8 36 −16 4.39
Right medial orbital gyrus 17 51 −19 3.67
Striatum
Right ventral striatum 10 10 −3 3.19a
Medial frontal cortex
Right anterior cingulate cortex (ACC) 8 48 12 4.94
Right medial prefrontal cortex 7 61 9 4.01
Parietal cortex
Right superior-caudal parietal lobule 32 −64 60 4.17
Right inferior-caudal parietal lobule 43 −61 51 3.92
Peri-insular region
Left rostral insula −27 25 −6 4.00
Chem. Percept. (2012) 5:11–21 15
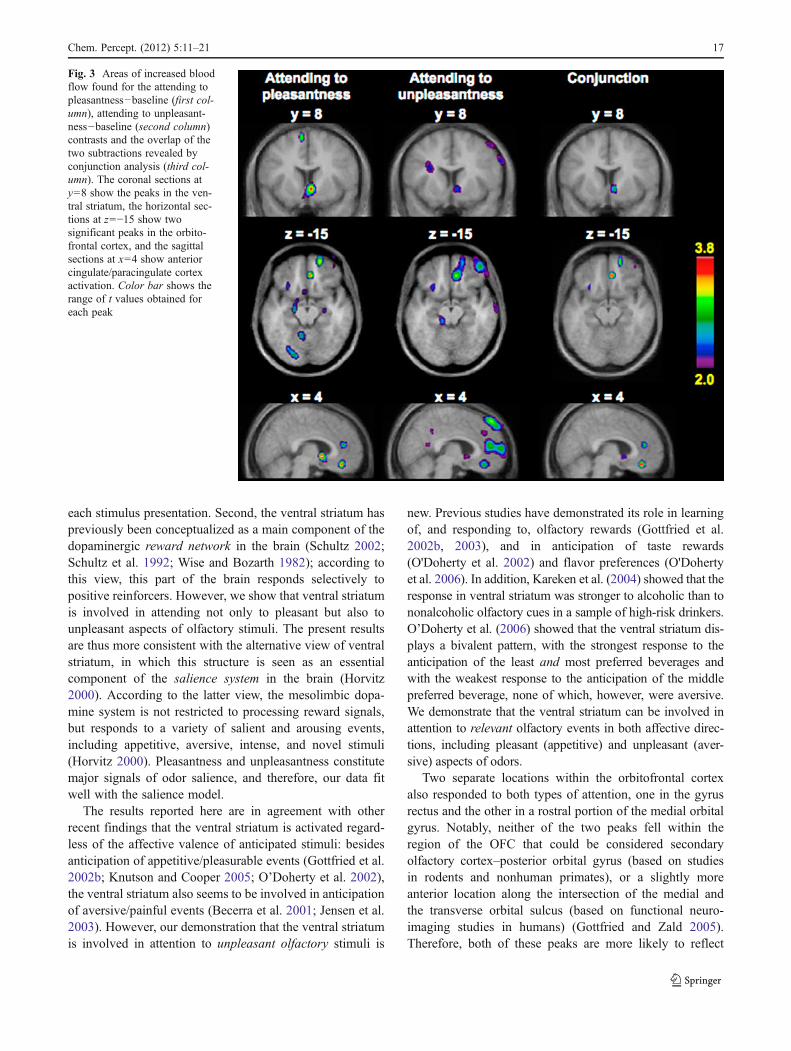
Finally, we conducted a conjunction analysis in order toinvestigate the overlap of the regions of activation foundwhile participants were attending to pleasantness (AttP-Base) and to unpleasantness (AttU-Base). Peaks revealedin the conjunction analysis are listed in Table 5 and shownin Fig. 3.
This analysis revealed that there were several areas ofsignificant overlap of activations in the two contrasts: twolocations in the orbitofrontal cortex and, importantly, one inthe ventral striatum. In addition, several locations of overlapwere seen in the medial frontal cortex, and one of themincluded the anterior cingulate cortex.
Discussion
This PET investigation revealed that the ventral striatum,two locations within the orbitofrontal cortex, and the ante-rior cingulate cortex are involved in attending to the affec-tive valence of odors, regardless of the “affective direction”;these structures showed increased activity when participantsattended both to pleasant and to unpleasant aspects of odors.Other brain structures showed increased activation specificeither to attention to pleasantness or attention to unpleasant-ness. Attention to pleasantness was associated with activa-tion in the left piriform cortex and amygdala, whereas the
left insula and the right parietal cortex were involved inattention to unpleasantness. These differences in brain activitywere observed despite the fact that the physical stimu-lation (i.e., presented odors) was identical in the twoexperimental conditions.
The main finding in this study was that directing attentionto affective properties of odors involves increased activationin the ventral striatum, including nucleus accumbens, ven-tral caudate, and ventral putamen. When considering theactivity of ventral striatum, there are two important points.First, some studies have shown that ventral striatumresponds more strongly to the anticipation than to the con-sumption/delivery of the expected outcome (Knutson et al.2001a,b; O’Doherty et al. 2002). The PET paradigm doesnot allow us to distinguish between the regions involved inanticipating versus perceiving the pleasant and unpleasantaspects of odors, but the attention task used in this study hadan important anticipatory component: by requesting thatparticipants indicate whether each presented stimulus had apleasant component in one, or unpleasant component inanother condition, we created an expectation/anticipationthat an affectively valenced component was to occur with
Table 4 Peaks of increasedrCBF specific to attending topleasantness (subtraction AttP-AttU) and to attending tounpleasantness (AttU-AttP)
x, y, and z coordinates are thesame as in Table 2
Area x y z t value
Attending to pleasantness
Medial frontal cortex
Left pre-supplementary motor area (pre-SMA) −7 15 52 4.48
Left pre-supplementary motor area (pre-SMA) −4 8 61 3.86
Attending to unpleasantness
Parietal cortex
Right parietal cortex surrounding intraparietal sulcus 51 −47 48 4.66
Right angular gyrus 44 −69 46 3.77
Medial parietal lobe (postcentral gyrus) −3 −50 66 3.77
Fig. 2 Areas of increased blood flow found for the attending tounpleasantness−attending to pleasantness contrast. Coronal sectionsshow a peak in the right parietal cortex around the intraparietal sulcus(y0−47) and another right parietal peak in the angular gyrus (y0−69).Color bar shows the range of t values obtained for each peak
Table 5 Peaks of increased rCBF common to attending to pleasant-ness and unpleasantness revealed by conjunction analysis (conjunctionof AttP-Base and AttU-Base)
Area x y z t value
Orbitofrontal cortex
Right gyrus rectus 5 37 −15 3.84
Right medial orbital gyrus 19 55 −18 3.48
Striatum
Right ventral striatum 8 10 −3 3.17
Medial frontal cortex
Anterior cingulate cortex (ACC) 4 41 15 3.22
Medial frontal gyrus 0 53 45 3.25
Medial frontal gyrus −1 51 48 3.19
Right frontal pole 38 63 3 3.02
x, y, and z coordinates are the same as in Table 2
16 Chem. Percept. (2012) 5:11–21
each stimulus presentation. Second, the ventral striatum haspreviously been conceptualized as a main component of thedopaminergic reward network in the brain (Schultz 2002;Schultz et al. 1992; Wise and Bozarth 1982); according tothis view, this part of the brain responds selectively topositive reinforcers. However, we show that ventral striatumis involved in attending not only to pleasant but also tounpleasant aspects of olfactory stimuli. The present resultsare thus more consistent with the alternative view of ventralstriatum, in which this structure is seen as an essentialcomponent of the salience system in the brain (Horvitz2000). According to the latter view, the mesolimbic dopa-mine system is not restricted to processing reward signals,but responds to a variety of salient and arousing events,including appetitive, aversive, intense, and novel stimuli(Horvitz 2000). Pleasantness and unpleasantness constitutemajor signals of odor salience, and therefore, our data fitwell with the salience model.
The results reported here are in agreement with otherrecent findings that the ventral striatum is activated regard-less of the affective valence of anticipated stimuli: besidesanticipation of appetitive/pleasurable events (Gottfried et al.2002b; Knutson and Cooper 2005; O’Doherty et al. 2002),the ventral striatum also seems to be involved in anticipationof aversive/painful events (Becerra et al. 2001; Jensen et al.2003). However, our demonstration that the ventral striatumis involved in attention to unpleasant olfactory stimuli is
new. Previous studies have demonstrated its role in learningof, and responding to, olfactory rewards (Gottfried et al.2002b, 2003), and in anticipation of taste rewards(O'Doherty et al. 2002) and flavor preferences (O'Dohertyet al. 2006). In addition, Kareken et al. (2004) showed that theresponse in ventral striatum was stronger to alcoholic than tononalcoholic olfactory cues in a sample of high-risk drinkers.O’Doherty et al. (2006) showed that the ventral striatum dis-plays a bivalent pattern, with the strongest response to theanticipation of the least and most preferred beverages andwith the weakest response to the anticipation of the middlepreferred beverage, none of which, however, were aversive.We demonstrate that the ventral striatum can be involved inattention to relevant olfactory events in both affective direc-tions, including pleasant (appetitive) and unpleasant (aver-sive) aspects of odors.
Two separate locations within the orbitofrontal cortexalso responded to both types of attention, one in the gyrusrectus and the other in a rostral portion of the medial orbitalgyrus. Notably, neither of the two peaks fell within theregion of the OFC that could be considered secondaryolfactory cortex–posterior orbital gyrus (based on studiesin rodents and nonhuman primates), or a slightly moreanterior location along the intersection of the medial andthe transverse orbital sulcus (based on functional neuro-imaging studies in humans) (Gottfried and Zald 2005).Therefore, both of these peaks are more likely to reflect
Fig. 3 Areas of increased bloodflow found for the attending topleasantness−baseline (first col-umn), attending to unpleasant-ness−baseline (second column)contrasts and the overlap of thetwo subtractions revealed byconjunction analysis (third col-umn). The coronal sections aty08 show the peaks in the ven-tral striatum, the horizontal sec-tions at z0−15 show twosignificant peaks in the orbito-frontal cortex, and the sagittalsections at x04 show anteriorcingulate/paracingulate cortexactivation. Color bar shows therange of t values obtained foreach peak
Chem. Percept. (2012) 5:11–21 17

higher order processing rather than sensory processing.Similar regions of OFC have been reported previously inassociation with olfactory learning (Gottfried et al. 2002b),olfactory working memory (Dade et al. 2001) and in responseto affective features of odors (Anderson et al. 2003;Grabenhorst et al. 2007; Rolls et al. 2003).
Finally, another area of increased activation common toattending to pleasantness and unpleasantness was revealedin anterior cingulate cortex (ACC). This rostral ACC acti-vation fell within the so-called affective subdivision of theACC, shown to participate in assessing the salience ofemotional and motivational information (Bush et al. 2000).The rostral ACC has direct anatomical links with the nucleusaccumbens in the ventral striatum and with the orbitofrontalcortex, which puts it in an ideal position for a role in mediatingmotivated attention (Carter et al. 1999) and emotional regula-tion (Bush et al. 2000; Devinsky et al. 1995).
Besides the areas of common activation, attending topleasantness versus unpleasantness in olfaction also showedsubtle, yet interesting differences. First, peaks of activationwithin the amygdala and the primary olfactory cortex werefound in association with attending to pleasantness but notunpleasantness, both when experimental conditions werecontrasted against baseline and in post hoc volume of inter-est analyses. Second, two peaks in the right caudal parietalcortex and one in the left insula were seen in associationwith attending to unpleasantness but not pleasantness. Inaddition to this, comparable peaks in the right parietal cortexwere associated with attending to unpleasantness and notpleasantness in the direct comparison between the two exper-imental conditions. Overall, these results show that the brainregions involved in attending to pleasantness and unpleasant-ness are partially dissociable.
While attending to odor pleasantness, piriform cortexwas recruited, implying that this process loads on odorperception. Furthermore, the amygdala was involved duringattention to pleasantness but not unpleasantness of odors.This finding is consistent with some other findings showingthat the amygdala can respond to positively valenced stimuli(Dolan 2002; Zald 2003). It remains unclear why amygdalaactivation was not found during attention to unpleasantodors. Some findings suggested that the amygdala plays arole in processing of all odors (Anderson et al. 2003;Gottfried et al. 2002a).Moreover, the anatomical link betweenthe amygdala and the olfactory brain is much tighter than forany other sensory system. Namely, several subregions of theamygdala receive direct projections from the olfactory bulband as such constitute part of the primary olfactory area(Carmichael et al. 1994). Such intimate connection betweenthe olfactory system and the amygdala is consistent with theview that this structure plays a role both in sensory andemotional processing of odors. This idea is consistent withour finding that the amygdala and the piriform cortex showed
a differential significance in the contrasts of each attentionalcondition against baseline and that the direct comparisonbetween the two attentional conditions did not yield signifi-cance in these regions. This was to be expected, given thatboth attentional conditions involved smelling odors and there-fore required activity of the piriform cortex and the amygdala,both of which are components of the sensory network; thus,the activations in these structures cancelled each other out inthe direct comparison. Therefore, we conclude that theseolfactory regions were activated during both conditions butthat there was a relative, subtle difference in that piriformcortex and the amygdala were more strongly activated inassociation with attending to pleasant than to unpleasantodors, as confirmed by the presence of a subthresholdpeak in the piriform cortex in the direct contrast andfurther confirmed by post hoc analyses. A possibilitythat the amygdala was activated more in associationwith attending to pleasant than to unpleasant odorsbecause of a difference in their intensity (i.e., the pleas-ant odors being stronger than the unpleasant ones) isnot likely, as the overall intensity of the pleasant andunpleasant odors was equivalent.
Another area suggested to play some role in attending topleasant but not unpleasant odors is the occipital cortex, asrevealed by its increased activity when baseline was sub-tracted from the attention to pleasantness condition.Although it is very tempting to interpret recruitment ofoccipital peristriate regions as an indication of some in-volvement of visual perception and/or visual imagery, morestudies will be needed to uncover this link. We note thatsome other functional neuroimaging studies to date havereported increased activity in the visual regions of the brainwhile smelling odors (e.g., Dade et al. 2002; Djordjevic etal. 2005; Qureshy et al. 2000; Royet et al. 2001; Zatorre etal. 2000).
Attending to odor unpleasantness recruited regions im-plicated in attention and working memory, whereas the mainregion implicated in odor perception (piriform cortex) becameless involved while participants attended to unpleasantnessversus pleasantness. The locations in the lateral-caudal parie-tal cortex observed in this study are in keeping with previousstudies showing participation of those same regions in olfac-tory working memory (Dade et al. 2001) and olfactory short-term memory (Dade et al. 2002). The fact that parietal peakswere found to be associated with attending to odor unpleas-antness both when it was contrasted against baseline andagainst attending to odor pleasantness provides a strong sup-port to the notion that attending to unpleasant components ofodors activates an attentional network more strongly than doesattending to pleasant components. Finally, activation in theleft insula/frontal operculumwas seen only in association withattending to unpleasant odors, consistent with previous find-ings associating activity in the left insula with disgusting odors
18 Chem. Percept. (2012) 5:11–21

(Royet et al. 2003; Wicker et al. 2003; Zald and Pardo 2000).Furthermore, our finding that the left rostral insula is activatedby simply attending to odor unpleasantness fits well with aprevious report that simply seeing somebody else experiencedisgusting odors activates the same region in the brain asmuch as does experiencing them oneself (Wicker et al. 2003).
Previous studies have shown that different brain regionsare involved in processing pleasant versus unpleasant odors.In contrast to other senses, several studies suggested that theprimary (including piriform cortex) and the secondary (orbi-tofrontal cortex) olfactory cortices are also involved in affec-tive evaluation of odorants. Gottfried et al. (2002a) were thefirst to show dissociable responses in the piriform cortex, bywhich posterior piriform responded to all odors, while theanterior piriform cortex responded selectively to pleasantand unpleasant odors. Zelano et al. (2007) showed that theright frontal piriform cortex responded more strongly tounpleasant than to pleasant odorants. Several studiesalso reported regional subspecialization with the OFC:More medial regions of the OFC responded to pleasant,while more lateral OFC regions responded to unpleasantodors (Anderson and Sobel 2003; Gottfried et al. 2002a;Grabenhorst et al. 2007; Rolls et al. 2003). Finally,findings have been mixed with respect to the role ofthe amygdala in the affective processing of odorants.The amygdala was found to respond preferentially toaversive odors (Zald and Pardo 1997) or odor valence(Royet et al. 2000). Gottfried et al. (2002a) reportedthat the amygdala had a more pronounced activity inresponse to an unpleasant odor, even though this struc-ture responded to all odors regardless of their valence.However, other findings suggested that the amygdalaresponded more to odor intensity than to odor valence(Anderson et al. 2003; Wicker et al. 2003), and Plaillyet al. (2005) found that the amygdala also responded toodor familiarity. Finally, Winston et al. (2005) showed that theamygdala responded to odor intensity of valenced odors (bothpleasant and unpleasant), but not to odor intensity of neutralodors. Overall, these findings suggest that affective processingin olfaction may be more complex than in other senses butalso that there is a clear disparity of brain structures implicatedin encoding pleasant versus unpleasant odors. In the presentinvestigation, the focus was not to determine which regionsrespond to pleasant versus unpleasant odors, as the odors ofour two experimental conditions were exactly the same andtherefore had the same affective quality. Similarly, it is lessrelevant whether a pleasant or unpleasant component wasdominant in any mixture, or whether their trigeminal aspectsinfluenced detection of pleasant versus unpleasant compo-nents. The main point is that the stimuli in the two experi-mental conditions were identical. Therefore, different brainactivations cannot be accounted for by differences within oramong the mixtures; instead, the only difference between
them was the ongoing cognitive activity, i.e., attending topleasant versus unpleasant odors. Our findings speak moreto the fact that just by selectively directing our attentiontoward the pleasant or unpleasant aspects of odors, the brainactivity subtly changes. We consider these fine differences tobe important because the physical stimulation was identical.Only a few previous studies have explicitly demonstrated thatbrain response to the same odors can vary as a function ofcognitive activity (de Araujo et al. 2005; Rolls et al. 2008;Zatorre et al. 2000) while physical stimulation remained thesame. Our findings thus confirm an important mediating roleof attention in olfactory sensory processing.
Conclusions
Taken together, our results show that the neural correlates ofattention to odor pleasantness and odor unpleasantness over-lap partially. Regions involved in attending to affectiveproperties of odors regardless of the valence include ventralstriatum, orbitofrontal cortex, and anterior cingulate cortex.However, brain responses to those same physical stimulivary as a function of the content toward which our attentionis directed. We show that the type of cognitive activity ourmind is engaged in modifies our neural activity even whenexternal, physical stimulation remains constant. These dif-ferences in neural activity that arise depending on whetherwe are attending to pleasant or unpleasant smells are con-sistent with an evolutionary view of olfaction, by whichunpleasant odors are more alerting to us than pleasant odors,i.e., unpleasant odors more readily engage an attentionalnetwork in the brain. Thus, unpleasant odors are perceivedmainly as warning signals to which we need to attend inorder to maintain our safety and wellbeing.
Acknowledgment This study was supported in part by grant 57846,awarded to MJG by the Canadian Institutes of Health Research (CIHR)and by grant 355938-08, awarded to JD by the he Natural Sciences andEngineering Research Council of Canada (NSERC). We are grateful toSidonie Pénicaud for testing participants, Michael Petrides for hisanatomical expertise, and the staff of the Brain Imaging Center at theMontreal Neurological Institute.
References
Anderson AK, Sobel N (2003) Dissociating intensity from valence assensory inputs to emotion. Neuron 39(4):581–583
Anderson AK, Christoff K, Stappen I, Panitz D, Ghahremani DG,Glover G, Gabrieli JD, Sobel N (2003) Dissociated neural repre-sentations of intensity and valence in human olfaction. NatNeurosci 6(2):196–202
Ashkenazi A, Marks LE (2004) Effect of endogenous attention ondetection of weak gustatory and olfactory flavors. PerceptPsychophys 66(4):596–608
Chem. Percept. (2012) 5:11–21 19

Becerra L, Breiter HC, Wise R, Gonzalez RG, Borsook (2001) Rewardcircuitry activation by noxious thermal stimuli. Neuron 32(5):927–946
Bush G, Luu P, Posner M (2000) Cognitive and emotional influencesin anterior cingulate cortex. Trends Cogn Sci 4(6):215–222
Carmichael ST, Clugnet MC, Price JL (1994) Central olfactory con-nections in the macaque monkey. J Comp Neurol 346(3):403–434
Carter CS, Botvinick MM, Cohen JD (1999) The contribution of theanterior cingulate cortex to executive processes in cognition. RevNeurosci 10(1):49–57
Dade LA, Zatorre RJ, Evans AC, Jones-Gotman M (2001) Workingmemory in another dimension: functional imaging of humanolfactory working memory. NeuroImage 14(3):650–660
Dade LA, Zatorre RJ, Jones-Gotman M (2002) Olfactory learning:convergent findings from lesion and brain imaging studies inhumans. Brain 125(Pt 1):86–101
de Araujo IE, Rolls ET, Velazco MI, Margot C, Cayeux I (2005) Cog<nitive modulation of olfactory processing. Neuron 46(4):671–679
Devinsky O, Morrell MJ, Vogt BA (1995) Contributions of anteriorcingulate cortex to behaviour. Brain 118(Pt 1):279–306
Djordjevic J, Zatorre RJ, Petriedes M, Boyle JA, Jones-Gotman M(2005) Functional neuroimaging of odor imagery. NeuroImage24:791–801
Dolan RJ (2002) Emotion, cognition, and behavior. Science 298(5596):1191–1194
Doty RL, Brugger WE, Jurs PC, Orndorff MA, Snyder PJ, Lowry LD(1978) Intranasal trigeminal stimulation from odorous volatiles:psychometric responses from anosmic and normal humans.Physiol Behav 20(2):175–185
Friston KJ, Holmes AP, Price CJ, Buchel C, Worsley KJ (1999)Multisubject fMRI studies and conjunction analyses. NeuroImage10(4):385–396
Geisler MW, Murphy C (2000) Event-related brain potentials toattended and ignored olfactory and trigeminal stimuli. Int JPsychophysiol 37(3):309–315
Gottfried JA, Zald DH (2005) On the scent of human olfactoryorbitofrontal cortex: meta-analysis and comparison to non-humanprimates. Brain Res Brain Res Rev 50(2):287–304
Gottfried JA, Deichmann R, Winston JS, Dolan RJ (2002a) Functionalheterogeneity in human olfactory cortex: an event-related functionalmagnetic resonance imaging study. J Neurosci 22(24):10819–10828
Gottfried JA, O'Doherty J, Dolan RJ (2002b) Appetitive and aversiveolfactory learning in humans studied using event-related functionalmagnetic resonance imaging. J Neurosci 22(24):10829–10837
Gottfried JA, O'Doherty J, Dolan RJ (2003) Encoding predictivereward value in human amygdala and orbitofrontal cortex.Science 301(5636):1104–1107
Grabenhorst F, Rolls ET, Margot C, da Silva MA, Velazco MI (2007)How pleasant and unpleasant stimuli combine in different brainregions: odor mixtures. J Neurosci 27(49):13532–13540
Horvitz JC (2000) Mesolimbocortical and nigrostriatal dopamine res-ponses to salient non-reward events. Neuroscience 96(4):651–656
Jensen J, McIntosh AR, Crawley AP, Mikulis DJ, Remington G, KapurS (2003) Direct activation of the ventral striatum in anticipation ofaversive stimuli. Neuron 40(6):1251–1257
Kareken DA, Claus ED, Sabri M, Dzemidzic M, Kosobud AE,Radnovich AJ, Hector D, Ramchandani VA, O'Connor SJ, LoweM (2004) Alcohol-related olfactory cues activate the nucleusaccumbens and ventral tegmental area in high-risk drinkers: pre-liminary findings. Alcohol Clin Exp Res 28(4):550–557
Knutson B, Cooper JC (2005) Functional magnetic resonance imagingof reward prediction. Curr Opin Neurol 18(4):411–417
Knutson B, Adams CM, Fong GW, Hommer D (2001a) Anticipation ofincreasing monetary reward selectively recruits nucleus accum-bens. J Neurosci 21(16):RC159
Knutson B, Fong GW, Adams CM, Varner JL, Hommer D (2001b)Dissociation of reward anticipation and outcome with event-related fMRI. Neuroreport 12(17):3683–3687
Krauel K, Pause BM, Mueller C, Sojka B, Mueller-Ruchholtz W, FerstlR (1998) Central nervous correlates of chemical communicationin humans. Ann N YAcad Sci 855:628–631
Luck SJ, Vicera SP (2001) Attention. In: Pashler H, Yantis S, Medin D,Gallistel R, Wixted J (eds) Steven's handbook of experimentalpsychology, 3rd edn. Wiley, New York, pp 235–286
O'Doherty JP, Deichmann R, Critchley HD, Dolan RJ (2002) Neuralresponses during anticipation of a primary taste reward. Neuron33(5):815–826
O'Doherty JP, Buchanan TW, Seymour B, Dolan RJ (2006) Predictiveneural coding of reward preference involves dissociable responsesin human ventral midbrain and ventral striatum. Neuron 49(1):157–166
Pause BM, Sojka B, Ferstl R (1997) Central processing of odorconcentration is a temporal phenomenon as revealed by chemo-sensory event-related potentials (CSERP). Chem Senses 22(1):9–26
Plailly J, Bensafi M, Pachot-Clouard M, Delon-Martin C, Kareken DA,Rouby C, Segebarth C, Royet JP (2005) Involvement of rightpiriform cortex in olfactory familiarity judgments. NeuroImage 24(4):1032–1041
Plailly J, Howard JD, Gitelman DR, Gottfried JA (2008) Attention toodor modulates thalamocortical connectivity in the human brain. JNeurosci 28(20):5257–5267
Price CJ, Friston KJ (1997) Cognitive conjunction: a new approach tobrain activation experiments. NeuroImage 5(4 Pt 1):261–270
Qureshy A, Kawashima R, Imran MB, Sugiura M, Goto R, Okada K,Inoue K, Itoh M, Schormann T, Zilles K, Fukuda H (2000)Functional mapping of human brain in olfactory processing: aPET study. J Neurophysiol 84(3):1656–1666
Raz A, Buhle J (2006) Typologies of attentional networks. Nat RevNeurosci 7(5):367–379
Rolls ET, Kringelbach ML, de Araujo IE (2003) Different representa-tions of pleasant and unpleasant odours in the human brain. Eur JNeurosci 18(3):695–703
Rolls ET, Grabenhorst F, Margot C, da Silva MA, Velazco MI(2008) Selective attention to affective value alters how thebrain processes olfactory stimuli. J Cogn Neurosci 20(10):1815–1826
Royet JP, Zald D, Versace R, Costes N, Lavenne F, Koenig O, GervaisR (2000) Emotional responses to pleasant and unpleasant olfac-tory, visual, and auditory stimuli: a positron emission tomographystudy. J Neurosci 20(20):7752–7759
Royet JP, Hudry J, Zald DH, Godinot D, Gregoire MC, Lavenne F,Costes N, Holley A (2001) Functional neuroanatomy of differentolfactory judgments. NeuroImage 13:506–519
Royet JP, Plailly J, Delon-Martin C, Kareken DA, Segebarth C (2003)fMRI of emotional responses to odors: influence of hedonicvalence and judgment, handedness, and gender. NeuroImage 20(2):713–728
Sabri M, Radnovich AJ, Li TQ, Kareken DA (2005) Neural correlatesof olfactory change detection. NeuroImage 25:969–974
Schultz W (2002) Getting formal with dopamine and reward. Neuron36(2):241–263
Schultz W, Apicella P, Scarnati E, Ljungberg T (1992) Neuronalactivity in monkey ventral striatum related to the expectation ofreward. J Neurosci 12(12):4595–4610
Spence C, Kettenmann B, Kobal G, McGlone FP (2000) Selectiveattention to the chemosensory modality. Percept Psychophys 62(6):1265–1271
Spence C, Kettenmann B, Kobal G, McGlone FP (2001a) Sharedattentional resources for processing visual and chemosensoryinformation. Q J Exp Psychol A 54(3):775–783
20 Chem. Percept. (2012) 5:11–21

Spence C, McGlone FP, Kettenmann B, Kobal G (2001b) Attention toolfaction. A psychophysical investigation. Exp Brain Res 138(4):432–437
Talairach J, Tournoux P (1988) Co-planar stereotaxic atlas of thehuman brain. Thieme, New York
Veldhuizen MG, Small DM (2011) Modality-specific neural effects ofselective attention to taste and odor. Chem Senses 36:747–760
Wicker B, Keysers C, Plailly J, Royet JP, Gallese V, Rizzolatti G(2003) Both of us disgusted in My insula: the common neuralbasis of seeing and feeling disgust. Neuron 40(3):655–664
Winston JS, Gottfried JA, Kilner JM, Dolan RJ (2005) Integratedneural representations of odor intensity and affective valence inhuman amygdala. J Neurosci 25(39):8903–8907
Wise RA, Bozarth MA (1982) Action of drugs of abuse on brainreward systems: an update with specific attention to opiates.Pharmacol Biochem Behav 17(2):239–243
Worsley KJ, Friston KJ (2000) A test for a conjunction. Stat ProbabLett 47:135–140
Worsley KJ,Marrett S, Neelin P, Vandal AC, Friston KJ, Evans AC (1996)A unified statistical approach for determining significant signals inimages of cerebral activation. Human Brain Mapping 4:58–83
Worsley KJ, Liao CH, Aston J, Petre V, Duncan GH, Morales F, EvansAC (2002) A general statistical analysis for fMRI data.NeuroImage 15(1):1–15
Zald DH (2003) The human amygdala and the emotional evalua-tion of sensory stimuli. Brain Res Brain Res Rev 41(1):88–123
Zald DH, Pardo JV (1997) Emotion, olfaction, and the human amyg-dala: amygdala activation during aversive olfactory stimulation.Proc Natl Acad Sci U S A 94(8):4119–4124
Zald DH, Pardo JV (2000) Functional neuroimaging of the olfactorysystem in humans. Int J Psychophysiol 36(2):165–181
Zatorre RJ, Jones-Gotman M, Rouby C (2000) Neural mechanisms 823involved in odor pleasantness and intensity judgments. 824NeuroReport 11:2711–2716
Zelano C, Bensafi M, Porter J, Mainland J, Johnson B, BremnerE, Telles C, Khan R, Sobel N (2005) Attentional modulationin human primary olfactory cortex. Nat Neurosci 8(1):114–120
Zelano C, Montag J, Johnson B, Khan R, Sobel N (2007) Dissociatedrepresentations of irritation and valence in human primary olfac-tory cortex. J Neurophysiol 97(3):1969–1976
Chem. Percept. (2012) 5:11–21 21