production of high-titer stocks of the english strain of rat cytomegalovirus
TRANSCRIPT

Journal of Virological Methods 86 (2000) 173–177
Short communication
Production of high-titer stocks of the English strain of ratcytomegalovirus
Alice Adams a,*, Lois Jane Heller b
a Department of Medical Microbiology and Immunology, School of Medicine, Uni6ersity of Minnesota, 10 Uni6ersity Dri6e, Duluth,MN 55812, USA
b Department of Medical and Molecular Physiology, School of Medicine, Uni6ersity of Minnesota, 10 Uni6ersity Dri6e, Duluth,MN 55812, USA
Received 4 October 1999; received in revised form 23 December 1999; accepted 10 January 2000
Abstract
A simple technique to increase dramatically the yield of rat cytomegalovirus (RCMV) from infected monolayers ofa rat embryo fibroblast-derived cell line is described. The method, which involves daily changes of the cell culturemedium, can result in a 50 000-fold amplification of virus from cell monolayers inoculated with only a few RCMVparticles. This modification of the standard in vitro culture technique to amplify viral yield can be used to increasethe sensitivity of the plaque assay for detecting very small amounts of infectious virus in organ homogenates ofRCMV infected animals. © 2000 Elsevier Science B.V. All rights reserved.
Keywords: b-Herpesvirus; In vitro culture; Plaque assay
www.elsevier.com/locate/jviromet
Human cytomegalovirus (HCMV) infection istypically asymptomatic in healthy adults. For im-munocompromised individuals, however, eitherprimary infection or activation of latent HCMVgenomes can result in serious sequelae. A numberof species specific cytomegaloviruses have beendescribed and used in animal models to understandvarious aspects of CMV-associated disease.
Two rat b-herpesvirus models for HCMV havebeen reported (Bruggeman et al., 1982; Priscott and
Tyrell, 1982). The Dutch or Maastricht strain of ratcytomegalovirus (RCMV) has been extensivelyused by C. Bruggeman and coworkers in Europe toprobe the involvement of CMV in organ transplan-tation. The English strain, which was likewiseisolated from a wild brown rat (Rattus nor6egicus)and once available from the American type culturecollection (ATCC), is the RCMV commonly usedin the United States. DNA sequence analysis of themajor immediate early gene locus of these twoRCMV isolates indicates that they are distinct ratCMV-like, b-herpesviruses rather than closely re-lated variants of the same virus (Beisser et al., 1998).
* Corresponding author. Tel.: +1-218-7267561; fax: +1-218-7267937.
0166-0934/00/$ - see front matter © 2000 Elsevier Science B.V. All rights reserved.
PII: S 0166 -0934 (00 )00136 -1

A. Adams, L.J. Heller / Journal of Virological Methods 86 (2000) 173–177174
In order to investigate further in vivo aspects ofCMV infection using a rat model, a method toproduce high titer stocks of infectious RCMV(English) was sought. Standard in vivo and invitro methods of propagating cytomegaloviruseswere evaluated. This report describes a modifica-tion of the tissue culture technique used to pro-duce virus for the molecular characterization ofRCMV (English) (Burns et al., 1988) that consis-tently yields high titer stocks (\106 PFU/ml) ofinfectious RCMV (English) even when the initialinfecting dose of virus is low.
Tissue culture-derived RCMV (English) wasobtained from G. Sandford (Medical College ofWI) along with a continuous rat embryo fibrob-last-derived cell line (REF) which was used toculture the virus in vitro and to quantitate virusby plaque assay. The REF cells were subculturedtwice weekly in MEM (minimal essential mediawith Earle’s salts and L-glutamine supplied byGibco) supplemented with 10% normal calf serum(Sigma) and 100 IU/ml penicillin and 100 mg/mlstreptomycin. Confluent monolayers of REF cellsin individual wells of 24 well titer plates (Costar)were used to quantitate virus by the plaque assayas follows. Duplicate aliquots (150 ml) of serialdilutions (10-fold) of virus were added to the cellmonolayers in the titer plates and incubated for 1h at 37°C. Unabsorbed virus was then washedaway and the cells overlaid with 1.0 ml completemedium that contained 0.9% methylcellulose(Sigma). The plates were incubated for 4–5 daysat 37°C after which the overlay was removed, thecells fixed with 95% methanol and stained with0.25% methylene blue. The plaques, which aresmall, were counted at a magnification of 50×with the aid of an inverted microscope. A linearrelationship where the number of plaque formingunits (PFU) was proportional to the virus dilutionwas consistently observed. Thus, there was noobvious indication that defective particles wereaccumulating with any of the virus productiontechniques tested.
Several attempts were made to propagateRCMV (English) utilizing the standard in vivomethod. This method involves intraperitoneal in-jection of a sublethal dose of virus into younganimals and recovery of progeny virions from
homogenates of the submaxillary salivary glandsharvested 2–4 weeks post infection. While over a1000-fold amplification of mouse cytomegaloviruscan be achieved by this method when highlysusceptible Balb/c mice are used (Adams et al.,1992), only modest yields of RCMC (Dutch) werereported following infection of Lewis (100-fold) orNorwegian brown rats (10-fold) (Bruggeman etal., 1983). In the present study, 3-week-old maleSprague Dawley (Harlan) rats (n=19) were in-jected i.p. with 1–3×105 PFU of RCMV (En-glish) and were killed after 2–6 weeks.Submaxillary salivary glands were harvested andthe total amount of virus in the tissue was deter-mined by plaque assay. The total yield of virusfrom the submaxillary salivary glands of the 19rats used for these experiments was only 0.8 timesthat of the combined input. The amount of virusrecovered was greater than that initially injectedfor only 6 of the 19 animals with the single bestyield only five times that of the infecting dose.Infection of Lewis and Norwegian brown ratswith RCMV (English) resulted in similar disap-pointing yields (unpublished observation). Theamount of RCMV (Dutch) in the salivary glandsremained at near plateau values for several weeks(Bruggeman et al., 1983) and the differences inyield observed with individual rats was not at-tributable to time of harvest of the glands duringthe experimental period. The concentration ofvirus (PFU/ml) in the 10% w/v salivary glandhomogenates was 1/10 of that of the infectingdose. Because of these low titers, no attempt wasmade to infect additional rats with the virus re-covered from the salivary glands. The plaquesproduced by virus obtained from organ ho-mogenates were indistinguishable from those ofthe in vitro produced inoculum, suggesting that asingle in vivo passage does not select more viru-lent progeny.
RCMV (English) has been propagated success-fully in vitro by infecting monolayers of the REFcell line with virus and harvesting the cell culturesupernatant 5–7 days later (Burns et al., 1988).However, in order to obtain high titer superna-tants (\106 PFU/ml), it was found that cellsmust be infected at a high multiplicity of infection(MOI) similar to the 0.1 PFU per cell used by
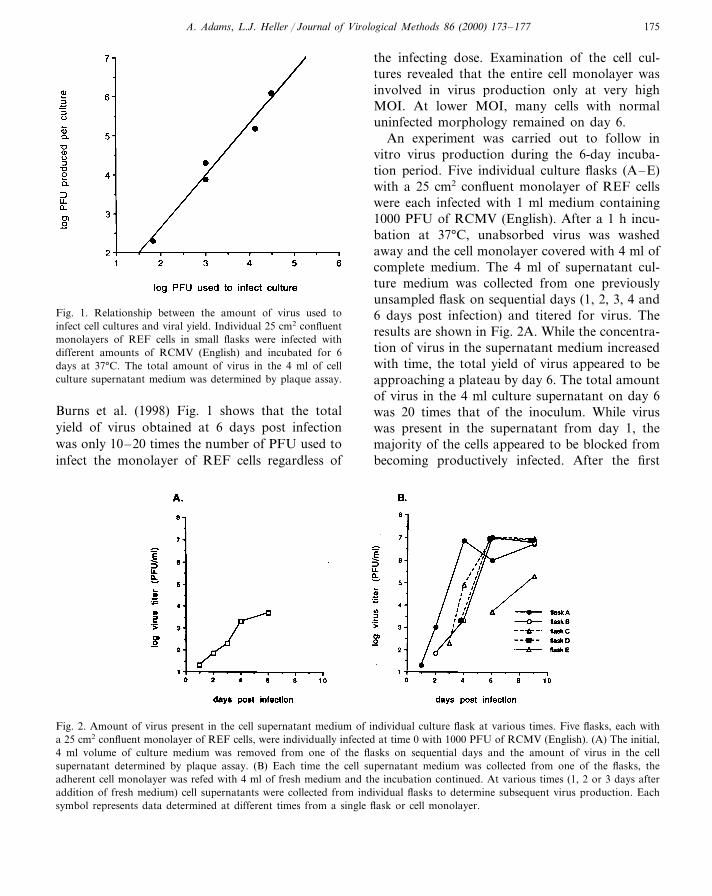
A. Adams, L.J. Heller / Journal of Virological Methods 86 (2000) 173–177 175
Fig. 1. Relationship between the amount of virus used toinfect cell cultures and viral yield. Individual 25 cm2 confluentmonolayers of REF cells in small flasks were infected withdifferent amounts of RCMV (English) and incubated for 6days at 37°C. The total amount of virus in the 4 ml of cellculture supernatant medium was determined by plaque assay.
the infecting dose. Examination of the cell cul-tures revealed that the entire cell monolayer wasinvolved in virus production only at very highMOI. At lower MOI, many cells with normaluninfected morphology remained on day 6.
An experiment was carried out to follow invitro virus production during the 6-day incuba-tion period. Five individual culture flasks (A–E)with a 25 cm2 confluent monolayer of REF cellswere each infected with 1 ml medium containing1000 PFU of RCMV (English). After a 1 h incu-bation at 37°C, unabsorbed virus was washedaway and the cell monolayer covered with 4 ml ofcomplete medium. The 4 ml of supernatant cul-ture medium was collected from one previouslyunsampled flask on sequential days (1, 2, 3, 4 and6 days post infection) and titered for virus. Theresults are shown in Fig. 2A. While the concentra-tion of virus in the supernatant medium increasedwith time, the total yield of virus appeared to beapproaching a plateau by day 6. The total amountof virus in the 4 ml culture supernatant on day 6was 20 times that of the inoculum. While viruswas present in the supernatant from day 1, themajority of the cells appeared to be blocked frombecoming productively infected. After the first
Burns et al. (1998) Fig. 1 shows that the totalyield of virus obtained at 6 days post infectionwas only 10–20 times the number of PFU used toinfect the monolayer of REF cells regardless of
Fig. 2. Amount of virus present in the cell supernatant medium of individual culture flask at various times. Five flasks, each witha 25 cm2 confluent monolayer of REF cells, were individually infected at time 0 with 1000 PFU of RCMV (English). (A) The initial,4 ml volume of culture medium was removed from one of the flasks on sequential days and the amount of virus in the cellsupernatant determined by plaque assay. (B) Each time the cell supernatant medium was collected from one of the flasks, theadherent cell monolayer was refed with 4 ml of fresh medium and the incubation continued. At various times (1, 2 or 3 days afteraddition of fresh medium) cell supernatants were collected from individual flasks to determine subsequent virus production. Eachsymbol represents data determined at different times from a single flask or cell monolayer.

A. Adams, L.J. Heller / Journal of Virological Methods 86 (2000) 173–177176
week of incubation, uninfected cells started tocontract and detach from the culture flask and itis unlikely that continued incubation would haveyielded more virus.
Each time the cell supernatant medium wascollected from one of the five flasks for analysis, 4ml of fresh medium was added and incubation ofthe cell monolayer continued. As is shown in Fig.2B, the addition of fresh medium stimulated dra-matically the cultures to produce additional virus.For example, the collective yield of virus in thefive supernatants collected from the cells of flaskA that was first sampled on day 1 was 50 000times that of the initial inoculum. All the cells ofthe monolayer of this flask on day four exhibitedthe morphology of typical CMV-infected cells. Onfurther incubation, these large round cells beganto detach from the flask and few cells remainedafter the final supernatant was collected on day 9.Thus, these data indicate that repetitive replace-ment of the supernatant medium promotes theproductive infection of more REF cells in theculture flask.
The best yields were obtained when the culturemedium was changed daily. Flask B that was firstsampled on day 2 produced about the sameamount of virus on day 4 (48 h after the firstaddition of fresh medium) as the culture in flaskA had produced on day 2. By day 4, cells of flaskA had produced nearly 10 000 times more virusthan those of flask B.
Fig. 2B also demonstrates that the level ofstimulation achieved by a given media change isrelated to the amount of virus in the culturesupernatant at the time the medium is changed.For example, flask A with 20 PFU/ml on day 1yielded 50 times more virus on the next day whileflask C with 200 PFU/ml on day 3 yielded 400times more virus the following day. A similartrend was seen for the 2-day incubation. Eventu-ally the cells lost their potential to amplify virusproduction as seen with flask E. When first titeredon day 6, flask E had over 1000 PFU/ml but onlya 100-fold increase was found 3 days later ascompared to the more than 10 000-fold increaseseen in 3 days for Flasks A, B, C and D. Frequentchanges of cell culture medium has consistentlyincreased the yield of virus and, while the mecha-
nism is not understood, the procedure is nowstandard for producing high titer stocks ofRCMV (English) in the laboratory.
The mechanism by which fresh medium stimu-lates virus infection is not known. It has beenreported that when primary rat embryo-fibrob-lasts are infected with small amounts of in vivo-derived RCMV (English), the cytopathic effecteventually progresses to involve the entire mono-layer (Priscott and Tyrell, 1982). However, pro-gression of infection on monolayers of REF cellswas blocked unless the supernatant medium waschanged. The difference between present findingsand the previous study could be associated with(a) the use of an in vitro- instead of an in vivo-derived inoculum; (b) attenuation of the virussince originally characterized in 1982; or (c) theuse of the REF cells rather than a primary cultureof rat embryo fibroblasts. The first hypothesisseems least likely because of the observations thatplaques produced by in vivo-derived virus are ofthe same small size as those produced with invitro-derived virus and the lack of progression ofcytopathic effects observed with the in vivo-derived virus is similar to that observed with thein vitro-derived virus.
It is suggested that, in addition to obtaininghigh titer stocks of RCMV (English), this methodof virus amplification is also useful for detectingvery small amounts of infectious virus in organhomogenates of infected animals. Early data ob-tained with RCMV (Dutch) (Bruggeman et al.,1983, 1985) indicate that a variety of different ratstrains, including Lewis and Norwegian Brownare similar to humans in that they are quiteresistant to infection with CMV and the amountof virus in various organs during the first week ofinfection is low. Because in vitro infection of REFcells with only one or few virus particles does notprogress to involve the entire monolayer, it isdifficult to detect the small amounts of infectiousvirus present in homogenates of various organsharvested during the acute phase of infection. Theminimum amount of virus detected by the stan-dard plaque assay is about 20 PFU/ml of 10%(w/v) organ homogenate. With the in vitro culturetechnique to amplify viruses described above, thesensitivity of the test can be increased 10-fold to

A. Adams, L.J. Heller / Journal of Virological Methods 86 (2000) 173–177 177
detect one or two virus particles per ml of organhomogenate.
The following describes the protocol developedand used to amplify virus present in organ ho-mogenates to detectable levels. The homogenate(1 ml) is added to a 25 cm2 monolayer of REFcells in a single culture flask. After 1 h, themonolayers are rinsed to remove debris and unab-sorbed virus. Medium (4 ml) is added to theculture flask and the cells are incubated. At 24and 48 h post infection, the supernatant mediumis discarded and 4 ml fresh medium is added. The72 h supernatants are collected and saved. Astandard plaque assay of the original organ ho-mogenate is conducted as described above to testfor presence of virus. If there are five or fewerplaques per well from the organ homogenate, the72-h supernatants in which virus production hasbeen amplified are then tested in a second plaqueassay. Within any given test series, the amount ofvirus observed in this second assay is roughlyproportional to that present in the initial ho-mogenate. The sensitivity could be increased fur-ther by infecting either more flasks or more cellsin a larger flask with a larger volume of organhomogenate and changing the medium for 2 or 3additional days before titering the final super-natant.
Thus, this simple modification of a standardtechnique for in vitro propagation of virus wasfound to be useful for producing high titer stocksof RCMV (English) and for detecting low levelsof infectious virus in tissue homogenates. Becauseof the growing importance of studies on CMV’s
role in various chronic diseases, it is useful tohave this rat model more readily available.
Acknowledgements
We thank Julie Smith for technical assistance.This work was supported by funds from the Min-nesota Medical Foundation and the University ofMinnesota Graduate School.
References
Adams, A., Mohrman, M., Johnson, A.G., Morin, A.,Deschamps de Paillette, E., 1992. Polyadenylic:polyuridylicacid-induced protection of Balb/c mice against acutemurine cytomegalovirus infection. J. Gen. Virol. 73, 2409–2413.
Beisser, P.S., Kaptein, S.J.F., Beuden, E., Bruggeman, C.A.,Vink, C., 1998. The Maastricht strain and England strainof rat cytomegalovirus represent different beta-herpesvirusspecies rather than strains. Virology 246, 341–351.
Bruggeman, C.A., Meijer, H., Dormans, P.H.J., Debie,W.M.H., Grauls, G.E.L.M., van Boven, C.P.A., 1982.Isolation of a cytomegalovirus-like agent from wild rats.Arch. Virol. 73, 231–241.
Bruggeman, C.A., Debie, W.M.H., Grauls, G., Majoor, G.,van Boven, C.P.A., 1983. Infection of laboratory rats witha new cytomegalo-like virus. Arch. Virol. 76, 189–199.
Bruggeman, C.A., Meijer, H., Bosman, F., van Boven, C.P.A.,1985. Biology of rat cytomegalovirus infection. Intervirol-ogy 24, 1–9.
Burns, W.H., Barbour, G.M., Sandford, G.R., 1988. Molecu-lar cloning and mapping of rat cytomegalovirus DNA.Virology 166, 140–148.
Priscott, P.K., Tyrell, D.A.J., 1982. The isolation and partialcharacterisation of a cytomegalovirus from the brown rat,Rattus nor6egicus. Arch. Virol. 73, 145–160.
.