psychrophilic bacteria1 - microbiology and molecular ...mmbr.asm.org/content/39/2/144.full.pdf ·...
TRANSCRIPT

BACrERIOLOGICAL REVIEWS, June 1975, p. 144-167Copyright 0 1975 American Society for Microbiology
Vol. 39, No. 2Printed in U.S.A.
Psychrophilic Bacteria1RICHARD Y. MORITA
Department of Microbiology and School of Oceanography, Oregon State University, Corvallis, Oregon 97331
INTRODUCTION ....................................................... 144
BACKGROUND INFORMATION ................................................
Definition used to describe psychrophiles ........................................ 146
ISOLATION OF PSYCHROPHILIC BACTERIA 147
ECOLOGICAL DISTRIBUTION OF PSYCHROPHILES ......................... 148
ECOLOGICAL SIGNIFICANCE OF PSYCHROPHILES 149
GROWTH OF PSYCHROPHILIC BACTERIA.150
Temperature characteristic of growth 153
PHYSIOLOGY OF PSYCHROPHILES ........................................... 153
Effect of temperature on respiration and viability .154
Proteins ....................................................... 155
Thermally induced leakage ..................................................... 156
Thermally induced lysis ....................................................... 157
Temperature effects of permeability .158
Temperature effects on macromolecular synthesis ............................... 158
Ecological factors affecting psychrophiles.159
CONCLUSION 161
LITERATURE CITED............................................... 162
INTRODUCTIONFor the purposes of this review, psychrophiles
are defined as organisms having an optimaltemperature for growth at about 15 C or lower, amaximal temperature for growth at about 20 C,and a minimal temperature for growth at 0 C orbelow. In order to avoid as much confusion aspossible concerning the term "psychrophile" inthis review, the term "psychrotrophic" will beused for organisms that do not meet the defini-tion as stated above, even though the citedauthor(s) may have referred to them as psychro-philes. Likewise, the term "psychrophile" willinclude those organisms previously referred toas obligate psychrophiles. Yeasts that meet theabove definition are known to exist but they willnot be discussed in this review.The main reasons for employing the above
definition are to eliminate future confusion andto adhere to the etymological significance of theterm psychrophile. Data are rapidly accumulat-ing on the existence of bacteria that meet theabove definition. The optimal temperaturelisted in this definition adheres to that given inthe Dictionary for Microbiology (88).Because of the recent isolation of psychro-
philes and the small number of investigatorsdealing with this unique group of bacteria, thereare many areas not yet investigated. Also,because of the restrictive definition employed inthis review there is a paucity of data from which
' Published as technical paper no. 4006, Oregon Agricul-tural Experiment Station.
to draw. Nevertheless the major objectives ofthis review are to make microbiologists awarethat "psychrophiles are psychrophiles" and toeliminate confusion in the literature concerningthe term.The general effects of temperature on mi-
crobial physiology and growth are well dis-cussed in many textbooks on microbiology andwill not be repeated in this review.
Hopefully this review will stimulate others touse the terms psychrophile and psychrotrophsas defined in this review. Although there maynever be a complete agreement on these terms,a start must be made to end the confusion aboutthe lower end of the thermal spectrum ofmicrobial life. The terms applied to the variousthermal groups are useful in microbiology, butnot when confusion about them exists.
BACKGROUND INFORMATIONTextbooks in general microbiology for many
years have classified microorganisms into vari-ous thermal groups based on their cardinaltemperatures. Of these groups, the cold-lovingbacteria are usually referred to as facultativepsychrophiles and psychrophiles. However, theterms used most often in the published papersto describe these groups are psychrophile andobligate psychrophile. Furthermore, textbooksgenerally list the optimum growth temperaturefor psychrophiles as 15 C. Yet, to most microbi-ologists, the term "psychrophile" does not referto the truly cold-loving bacteria but those thathave the ability to grow at low temperatures.
144
on August 4, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from

PSYCHROPHILIC BACTERIA
(The early literature would say "grow rapidly,"meaning growth in at least a week.) The termpsychrophile today is still misused, and themisnomer continues to be perpetuated in theliterature.That certain organisms can overlap any two
adjacent thermal groups is well recognized. Forthose mesophilic organisms that have the capa-bility to grow at 0 C, the term "psychrotoler-ant" (98) and "psychrotrophic" are perhaps thebest terms. The latter appears to be used morefrequently in the literature today.There are many review articles dealing with
"psychrophilic" bacteria (30, 83, 84, 85, 86, 159,173, 187); however, they cover the psychro-trophic bacteria, not the psychrophilic bacteria.There is a common misconception concerning
the maximum, minimum, and optimum growthtemperatures for psychrophilic bacteria. Thereports in the literature dealing with psychro-philic bacteria refer to organisms that are cold-tolerant rather than cold-loving. This was espe-cially true before the 1960's. Much of theconfusion in the early literature arose from thelack of adequate data to support the concept ofpsychrophily.The early investigators concentrated on the
ability to grow at or near 0 C. This ability togrow at 0 C was first reported by Forster (59) in1887. The organisms were isolated from fishpreserved by cold temperatures and were phos-phorescent (bioluminetent). Later Forster (60)
reported the isolation of other bacteria fromvarious environments (natural water, foods,surface and in intestines of freshwater fish, andespecially in sea water and ocean fish) that werecapable of growth at 0 C. From his studies in theAntarctic, Ekelof (51) proposed the existence ofpsychrophiles.The main deterrent to obtaining adequate
data in the early investigations was probablythe lack of refrigerated incubators. In addition,the investigators were unaware of the thermola-bility of psychrophilic organisms at laboratorytemperatures. Because of this, early investiga-tors (as listed in Table 1) did not determine thecardinal temperatures for their isolates. Thiswas the beginning of the confusion concerningthe cardinal temperatures for psychrophilicbacteria.
In 1902 Schmidt-Nielsen (161) grew microor-ganisms at 0 C which he termed "psychro-philes." A year later Muller (138) objected tothe term since the organisms described actuallygrew well at higher temperatures. The culturesisolated by early microbiologists actually grewbetter in the mesophilic temperature range(Table 1). As'a result of this inconsistency,various terms were coined by early investigatorsto describe organisms which had the ability togrow at 0 C (Table 2). A survey of the literaturefor the cardinal temperatures of psychrophileswas made by Morita (121). Unfortunately, thepsychrophiles described before 1960's were not
TABLE 1. Early investigators reporting bacterial growth at 0 C or lower
Investigator and year
Forster (1887, 1892)
Fischer (1888, 1893)Conradi and Vogt (1901)Baur (1902)Brandt (1902)Schmidt-Nielsen (1902)
Miller (1903)
Feitel (1903)Tsiklinsky (1908)Richardson (1908)Pennington (1908)Ravenel et al. (1910)Conn (1910,1912, and 1914)Brown and Smith (1912)Reed and Reynolds (1916)Vanderleck (1917 and 1918)Vass (1919)Fay and Olson (1924)Bardach (1924)Rubentschik (1925)
Source material or organism
Fish, natural waters, foods, wastes, rubbish, soil, surface and intestines offish
Harbor water and soils, certain pathogenic bacteria"proteus bacillus"Bacterium lobatumBacterium lobatumBacillus aquatile fluorescense non-liquefaciens, B. granulosum, B. paracoligasoformans anindolicum, B. radiatum, B. tarde fluorescens, B. pestis,and B. proteus fluorescens.
Sausage meat, fish, intestinal contents of fish, milk, vegetables, meal,garden soil and muck
Dentrifying bacteria from deep sea waterAntartic fishNonproteolytic bacteria on frozen meatMilkMilkFrozen soilsFrozen soilsMilkFrozen soilsFrozen soilsIce creamSewage, sludge and salt waterUrbacillus psychrocartericus, Urosarcina psychrocartericus
145VOL. 39, 1975
on August 4, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from

BACTERIOL. REV.
TABLE 2. Terms used to describe bacteria capable ofgrowing at or below 0 C
Term Reference
Glaciale Bakterien Forster (1892)Rhigophile Forster (1892)Psychrophile Schmidt-Nielsen (1902)Psychrotolerant Kruse (1910)Psychrocartericus Rubentschik (1925)Psychrobe Horowitz-Wlassowa and
Grinberg (1933)Thermophobic bacteria Edsall and Wetterlow
(1947)Cryophile Edsall and Wetterlow
(1947)Facultative psychrophile Hucker (1954)Obligate psychrophile Hucker (1954)Psychrotrophic Eddy (1960)
truly psychrophilic (with one exception [177]).After surveying the literature, Berry and Ma-goon (13) stated that "the contention that a
truly cold-loving or psychrophilic group of mi-croorganisms exists, appears unwarranted byavailable evidence. While organisms recordedas growing at 0 C or below may with proprietybe regarded as forming a cold-tolerant group,
their temperature requirements for best growthdefinitely place them in the mesophilic class."Other workers (69, 82, 83, 84, 85, 104, 105) alsoconcluded that there was a lack of adequatedata to substantiate the existence of psychro-philes. In 1963, Ingram (86) stated that the termpsychrophile was a misnomer. Since Schmidt-Nielsen (161) first used the term "psychro-phile," it has been employed indiscriminatelyso that there is much confusion in the literature.Hagen et al. (71) point out that the termpsychrophile has been used and defined inmany ways and most of the definitions ignorethe etymological significance of the word.
Definitions used to describe psychrophiles.Schmidt-Nielsen (161) defined psychrophiles as
organisms which are not only able to survive butalso multiply at 0 C. The term "psychrophile,"according to Horowitz-Wlassowa and Grinberg(81), should be used for the commonly isolatedbacteria that could grow at 0 C but actuallygrow better at higher temperatures; and theterm "psychrobe" should be used for the truepsychrophile if they existed.Without adequate data in the literature for
the existence of psychrophiles, the Dictionary forMicrobiology (88), published in 1957, definedpsychrophiles as those bacteria having an op-timum growth temperature of 15 C or lower.Organisms that grew at 0 C were separated
into two categories, obligate and facultative, byHucker (82). The former were capable of growth
at 0 C but not at 32 C, whereas the latter couldgrow at 0 C and 32 C. Many microgiologiststoday consider psychrophiles to be organismsthat more or less fit the facultative category.Ingraham (83) suggested that psychrophiles
be defined not by their maximum or optimumgrowth temperature but whether they growrapidly at 0 C. The term "rapidly" generallymeant 1 or 2 weeks, a period much too long forpsychrophiles. Later Ingraham and Stokes (85)redefined the term to refer to bacteria that growwell at 0 C within 2 weeks, disregarding thecardinal temperatures. The term "grow well"referred to a visable colony or turbidity to thenaked eye. They also suggested that the obli-gate psychrophile be defined as those bacteriathat grew rapidly at 0 C but also grew mostrapidly at temperatures below 20 C. The facul-tative psychrophile was capable of growth at 0C.Microorganisms capable of growing at 5 C
and below, according to Eddy (48), should beclassified as psychrotrophic regardless of theoptimum temperature and that the term psy-chrophile should be used only when a lowoptimum temperature is implied. There aremany microbiologists who employ the termpsychrotrophic (e.g., 47, 102, 114, 137, 186, 189),and this term is gaining wider acceptanceamong food and dairy microbiologists. Ingram(86) gives the best reasons published in theliterature as to why the term psychrotrophshould be employed for most of the organismslisted in the literature as psychrophiles. Baxterand Gibbons (10) defined psychrophiles as orga-nisms with a temperature optimum of 20 C orbelow. Rose (158) applied the term to organismshaving an optimum for growth at 25 C or belowwithout reference to growth at or near 0 C anddefined obligate psychrophiles as having a max-imum growth temperature of around 30 C. Thedefinition employed most often in the literatureis the one proposed by Stokes (173) in which hedefined psychrophiles as organisms which growat 0 C so that they can be detected visuallywithin 1 week; facultative psychrophiles havean optimum temperature of 20 C or higher,whereas the optimum growth temperature ofobligate psychrophiles is 20 C or lower.The more recent definitions for psychrophile
by Stokes (173) and Rose (158) are discussed byIngram (87), who points out the inadequacies oftheir definitions. Ingram (87) also states thatthe term psychrophile has been misapplied toorganisms which grow at 20 but not at 30 C andthat there is a preponderant group of organisms,with optimum temperatures between 20 and 30C, that are not "facultative psychrophiles" inthe true meaning of the term. This latter group
146 MORITA
on August 4, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from

PSYCHROPHILIC BACTERIA
cannot be named satisfactorily (87). For thebest discussion of the inconsistency in the logicof nomenclature dealing with psychrophiles,Ingram (87) should be consulted.Optimum growth temperature, ability to
grow at low temperature, method of enumera-tion, and criteria which are independent of thetemperature have been used to define psychro-philes (187). Different definitions are held byvarious areas of microbiology (e.g., dairy andfood microbiology). All this adds to the confu-sion in the literature and to the definition of theterm "psychrophile."For more discussion on the subject, Berry and
Magoon (13), Hucker (82), Ingraham andStokes (85), Witter (187), Ingraham (84), In-gram (87), and Morita (121) should be con-sulted. If the definition of psychrophile is to bechanged in the future, then it is my opinion thatthe cardinal temperatures will be lower thanthose that I have given in my definition.
ISOLATION OF PSYCHROPHILICBACTERIA
Tsiklinsky in 1908 (177) reported the exist-ence of a psychrophile. The psychrophile wasisolated from the excrement of a fish (environ-mental temperature was - 1.5 C) and had agrowth temperature range from 0 to 20 C, withan optimal temperature for growth at 16 C. Itappears that the isolate went into limbo and thesignificance of this isolate (probably a vibrioaccording to her description) has been neglectedby microbiologists. In 1948, Borg (15) isolatedthe etiological agent for "cold water" disease ofsilver salmon. The organism was assigned thename Cytophaga psychrophila and had an op-timum temperature between 15 and 20 C. Thisorganism, the first psychrophile to be wellcharacterized, fits the definition of psychrophilebetter than any organisms described prior to1963, but unfortunately Borg's thesis went un-noticed.
In 1964 Eimjhellen (71), Morita and Haight(135), and Sieburth (164) reported the existenceof psychrophilic bacteria. Since these reports,others have reported the existence of psychro-philic bacteria. All the isolates were from themarine environment. Eimjhellen isolated hisorganism from flounder eggs. Because the orga-nism produces a red pigment, it was tentativelydesignated as a Serratia sp. but was laterclassified as Vibrio psychroerythrus (41). Thered pigment of V. psychroerythrus is differentfrom five other prodigiosin-like compoundsfound in other organisms (39). Morita andHaight (135) isolated their organism fromoceanic waters 125 miles off the Oregon coast(1,200 meters, 3.24 C) and was identified as
Vibrio marinus, MP-1 (32). Sieburth's isolatesfrom Narragansett Bay, R.I., were Arthrobacterspp.The two main reasons why microbiologists in
the past were not successful in isolating psy-chrophilic bacteria are probably the improperselection of source material and the laboratoryprocedure for handling material. The sourcematerial must be selected carefully. The pasthistory of the source material is critical since itmust never reach warm temperature for anyextended period of time. For example, if suffi-cient radiant energy should fall on the sourcematerial it might be adequate to thermallyinactivate the psychrophilic bacteria, if present.Hence the environment from which the sourcematerial is taken must remain cold constantly.Llano (103) recorded a change from -15 to 27.8C on rock surfaces in the Horlick Mountains (40from the geographical South Pole) within 3 hdue to solar radiation even though the airtemperature was around 0 C. Boylen and Brock(17) were not able to demonstrate the develop-ment of a true psychrophilic flora in LakeWingra sediments which had a temperature of 4C for only 3 months of the year. Brock et al.(21), using proper techniques for the isolation ofpsychrophiles, were unable to obtain a psychro-phile from the cold springs in caves located insouthern Indiana. The above only indicates thatthe past history of the environment from whichsample material is taken is very important.Oceanic water has been cold for eons of time,and as a result it is easy to isolate psychrophilesin waters below the thermocline in the ocean aswell as in arctic and antarctic waters. Theisolation procedure for psychrophilic bacterialcells calls for the usual microbial methodsexcept that the source material, pipettes, me-dia, etc., are kept cold at all times (22, 121,185). The above emphasizes the abnormal ther-mal lability of the psychrophilic bacteria.The pour plate technique employing agar
medium cannot be recommended. It was shownthat most marine forms are thermosensitive tothe plating temperature of agar (192). Thepsychrophile, V. marinus MP-1, can be ren-dered nonviable when exposed to melted agar at42 C for 1 min (135).The enrichment technique, with the desired
medium, is recommended before trying to iso-late pure cultures. The inoculated enrichmentmedium should be incubated between 0 and 5 Cbefore proceeding with the isolation procedure.
Stock cultures of psychrophiles should betransferred frequently, and at no time shouldthe cultures be exposed to elevated tempera-tures for any extended period of time. Frequenttransfer is necessary since the temperature of
147VOL. 39, 1975
on August 4, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from

BACTERIOL. REV.
the laboratory refrigerator is nearly optimal forthe growth of some psychrophiles.
Psychrophiles are not rare in permanentlycold environments and can be isolated easily ifthe correct application of the above proceduresis followed.
ECOLOGICAL DISTRIBUTION OFPSYCHROPHILES
Most biologists tend to neglect the fact thatthe dominant environment of the biosphere iscold. The polar regions and the oceans are 14and 71% of the earth's surface, respectively.More than 90% (by volume) of the oceans is 5 Cor colder. The oceans are a natural habitat forpsychrophilic bacteria. Because of this situationit is relatively easy to isolate psychrophilicbacteria from below the thermocline of theoceans, in salt wedges (cold oceanic waterintrusion) of bays as well as both polar regions.
Psychrophilic bacteria have been isolated offthe Oregon coast as well as the Arctic andnorthern Pacific waters (130). For instance, 65miles off the Oregon coast 79 and 27 psychro-philes were found per 100 ml of sea waterobtained from depths of 350 and 2,800 meters,respectively. Freshwater psychrophilic bacteriaare readily isolated adjacent to melting gla-ciers in Alaska (Burton, personal communica-tion).
Sieburth (164, 166) examined various orga-nisms isolated at 0, 18, and 36 C from waters ofNarragansett Bay, R. I., and found that theredefinitely were microorganisms belonging to thepsychrophilic group. These psychrophiles were
only obtained on plates incubated at 0 C uponinitial isolation and only when the water tem-perature was below 10 C. Many of the isolateshad optimum temperatures for growth around10 C. They were tentatively identified as Ar-throbacter spp., Achromobacter spp., Flavobac-terium spp., Vibrio spp., and Pseudomonas spp.A seasonal selection for the various thermaltypes was also observed so that the minimal,optimal, and maximal growth temperature ofthe dominant microbial flora continuouslychange as a result of the continual seasonalchanges in water temperature. For example,certain isolates made at 0 C in the spring failedto grow above 16 C whereas isolates madeduring the summer could grow above 16 C.According to Sieburth (164), the life cycle ofArthrobacter spp. appear to be associated withdifferent temperatures (i.e., life cycle was a
function of water temperature).From the North Sea, Harder and Veldkamp
(75) isolated 69 strains of bacteria capable ofgrowth at low temperatures. Ten of these orga-
nisms had a maximum growth temperaturebelow 20 C, six of which were Pseudomonas,two were Vibrio, and two were Spirillum.Gounot (64, 65, 66) and Moiround and Gou-
not (120) investigated the microbial populationin sediments and water below arctic glaciers, inarctic caves, and other cold environments. Inher studies, she isolated an Arthrobacter sp.which had an optimum temperature for growthat 13 C. It also grew well at -5 and 18 C. Sheassigned the name Arthrobacter glacialis to theorganism.Approximately 800 isolates were obtained
from Arctic sediment samples (119), of whichtwo hundred fifty were found to grow well onagar at 0 but not at 25 C. Unfortunately, theseorganisms were never characterized thermally.Stanley and Rose (171) isolated psychrophilicbacteria from various lakes on Deception Is-land, Antarctica, and found 2, 9, and 18 bacte-ria with optimum growth temperatures of 5, 10,and 15 C, respectively.A preliminary examination of 488 cultures
isolated from the Antarctic was made by Wiebeand Hendricks (185) who found that 37.5% ofthe organisms would not grow at 15 C and 65%did not survive at 20 C. One of the species,designated as E5-4-4, had a maximal growthtemperature at 10 C (31).Matches and Liston (115) isolated 288 clos-
tridia from marine sediment collected in thePuget Sound area, of which 7 would not growabove 20 C. Five out of 29 clostridia studied byFinne and Matches (56) were psychrophiles.Approximately 400 isolates have been made
by my associates during Cruises 46 and 51 of theU.S.N.S. Eltanin in the Antarctic. One hun-dred fifty of the 400 isolates were selected forfurther characterization, and it was found that148 would grow at 10 C, 106 would grow at 15 Cand 98 would grow at 20 C; 144 were oxidasepositive, seven were agar digestors. All isolatesdemonstrated lipolytic activity and approxi-mately 50% were chitin digestors. Also presentwere several psychrophilic anaerobes.
Vishniac and Mainzar (183) investigated thedry valleys in southern Victorialand, Antarc-tica, for microbes. This area represents one ofthe driest regions on earth and the relativehumidity is about 10%. Although temperaturewas not mentioned in the paper, the in situ 1-hexperiments demonstrated the presence andactivity of bacteria inhabiting the dry soils. Thereason for the 1-h experiments was dictated bythe ability of the investigators to work comfort-ably with their hands at subfreezing tempera-tures coupled with a wind velocity of 60 km/h.Since miltiplication of the microorganisms were
148 MORITA
on August 4, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from

PSYCHROPHILIC BACTERIA
observed when water was available, the orga-
nisms could have been psychrophiles or psy-
chrotrophs. Their conclusion was that the bac-teria underwent rapid division within a fewhours of the day when water was available. Thiswater was the result of periodic snowfalls whichmelted when the radiant energy was present insufficient quantity. If water was not available,the bacteria laid dormant for a week or 10 daysat a time.
Psychrotrophic bacteria have been known toexist in the Antarctic region since the earlyinvestigations conducted by Ekelif (51) duringthe Swedish Antarctic Expedition of 1901-1903.Most bacteriological investigations (16, 26, 27,29, 42, 62, 117, 172) focused mainly on thedemonstration of various types of bacteria pres-
ent and not the existence of psychrophiles.Generally no precautions were taken to insurethat psychrophiles would not be inactivated.For instance Boyd and Boyd (16) used nutrientagar for dilution of their Antarctic samples aswell as an incubation temperature of 20 to 22 C,whereas Darling and Siple (42) used an incuba-tion temperature of 35 and 37 C. Cameron et al.(28) incubated their cultures at 2 and 20 C butin only two samples were there more bacteriagrowing at 2 than at 20 C. Starka and Stokes(172), using soil samples, demonstrated thepresence of psychrotrophic bacteria. It is diffi-cult to determine from their data whether or nota psychrophilic bacterium was isolated thatmeets the definition given in this review. Sincesoil samples were employed it is likely that thetrue psychrophiles may have expired in thesource material. For further details concerningthe early Antarctic microbiological literatureSieburth (165) should be consulted.
Unfortunately, there has never been an exten-sive effort made to determine ecological distri-
bution of psychrophiles in permanently coldenvironments. In the marine environment it iseasy to isolate psychrophilic bacteria as well asmany psychrotrophic forms. The latter gener-ally have a wide growth temperature range.
ECOLOGICAL SIGNIFICANCE OFPSYCHROPHILES
In the natural environment it is very difficultto assess the importance of psychrophiles in thecycle of matter. Generally within any sampleboth psychrophiles and psychrotrophs can beisolated. If sufficient substrates are present atlow temperatures near 0 C, a psychrophile canoutgrow the psychrotrophic forms (130). Evenat low substrate concentration and low temper-ature the psychrophiles will outgrow the psy-chrotrophic bacteria in continuous culture (77).These laboratory findings suggest that therewill be a continuous selection for bacteria whoseactivity is greatest near the environmental tem-perature, especially in environments showingseasonal temperature changes (140, 164, 166).
In Antarctic field experiments employing theheterotrophic activity method at temperaturesslightly above and below 0 C, the average Vmaxfor glutamate uptake in water samples taken atthe bottom of the euphotic zone was 1.09 x 10-2Ag/liter per h. A portion of the data obtainedduring Cruise 46 of the U.S.N.S. Eltanin isgiven in Table 3. The heterotrophic activitymethod (79, 148, 187) is a kinetic approachwhich is based on the principles of the Michae-lis-Menton theory. By using the kinetic ap-proach, the maximum velocity of uptake(Vmax), the turnover time (Ti), and the sum of a"transport constant" and the natural substrateconcentration (K, + SO) can be calculated. Thistype of data can be obtained in field studieswhere the water samples are immediately ana-
TABLE 3. Heterotrophic activity on "4C-labeled substrates by the indigenous microflora in water samples takenfrom various depths at Stations 8 and 9 during Cruise 46 of the U.S.N.S. Eltanina
Station Depth (m) Substrate Vma,,x(/g/liter per h) Tt(h) K, + S. %(jsg/liter) Respiration
8 10 Glu 1.8 x 10-2 570 10.4 36Glut 8.7 x 10-2 112 9.7 74
50 Glu 3.0 x 10-3 818 2.4 36Glut 1.3 x 10-2 382 5.0 70
500 Glut 1.1 x 10-3 12,350 13.8 779 10 Tyr 4.0 x 10-3 5,000 2.0 9
Lys 7.3 x 10-4 3,704 2.7 4Glu 3.1 x 10-3 4,800 11.8 24Glut 6.7 x 10-3 1,544 10.3 64
50 Glu 5.3 x 10-4 1,800 1.0 21Glut 1.6 x 10- 3 1,298 2.0 51
a The water temperature was 0 C ± 1. The percentage of respiration equals 14CO, x 100/"4C uptake + 14CO2.
149VOL. 39, 1975
on August 4, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from

BACTERIOL. REV.
lyzed at an in situ temperature. Other measure-
ments with other amino acids were also made(134). Employing the best field results obtainedduring Cruise 46 of the U.S.N.S. Eltanin, one
can extrapolate these figures to a longer periodof time and for a larger volume of water. Byextrapolation 2.47 x 107 g/nautical mile3 per
day of carbon dioxide can be produced by theindigenous microorganisms in the Antarcticwater (-1.0 C) (131). If one could legitimatelyextrapolate this figure to the entire volume ofthe ocean, then about 1015 g of carbon dioxidewould be produced per day. The foregoing alsoindicates that organisms can function very wellin cold environments where the temperature is0 C.There are approximately 129 x 1018 g of
carbon dioxide in the ocean (160). Although theconditions necessary to produce the ex-
trapolated amount of carbon dioxide by mi-crobial activity may not be present, the poten-tial to produce this quantity of carbon dioxide ispresent. In consideration of the importance ofthe entire carbon dioxide system on earth, thefunction of microbes in the environment shouldnot be taken lightly (125).Many of our psychrophilic isolates in our
laboratory are extremely good chitin digestorsat temperatures slightly above and below 0 C.The supernatant culture fluid of psychrophile18-500 (tentatively identified as a Vibrio sp.)displays chitinase activity at 0 C. Although theorganism has a maximum temperature forgrowth at 13 C, its chitinase functions at a
much higher temperature (Table 4). ZoBell andRittenberg (193) estimated that the annualchitin production from copepods is several bil-lion tons. This does not take into considerationthe amount of chitin produced by other marineforms (e.g., shrimps, crabs, etc.). Since fish eatorganisms having exoskeletons composed of chi-tin, the supernatant fluid of a sculpin's (Lep-tocottus armatus) stomach content was checkedfor the presence of chitinoclastic bacteria. Permilliliter of supernatant fluid, 1.5 x 1010chitinoclasts were isolated. The sculpin wasobtained in the winter where its environmentaltemperature was 6 C. When a portion of thesupernatant material from the fish was treatedwith toluene and placed in the presence ofchitin, 162 gg of N-acetylglucosamine/g (dryweight) per h at 5 C was produced. Otherexoenzymes are also produced by psychrophilesso that the cycle of matter can continue inenvironments close to 0 C.Whether or not there are pathogenic psychro-
philic bacteria in the aquatic environment hasyet to be investigated. The organism Cytophaga
TABLE 4. Chitinase activity by supernatant culturefluid resulting growth of 18-500 at 5 C for 5 daysa
Temp (C) N-acetylglucosamine producedTemp(C) ~~~(;&g/h per ml)
0 2.03 2.46 3.510 3.513.5 5.018.5 6.620 8.027 9.535 6.840 1.1
a The culture medium contained: yeast extract(Difco), 1.2 g; Trypticase (BBL), 2.3 g; sodiumcitrate, 0.3 g; L-glutamic acid, 0.3 g; sodium nitrate,0.05 g; ferrous sulfate, 0.005 g; sodium chloride, 24 g;magnesium sulfate, 7 g; magnesium chloride, 5.3 g;potassium chloride, 0.7 g; and purified chitin, 1 g.
psychrophila (psychrotroph) does have the abil-ity to cause cold-water disease (15, 148). Shelldisease, burned spot disease, and rust disease(necrotic lesions in the exoskeleton of crus-taceans) may be directly or indirectly caused bychitinoclastic bacteria since these organisms arenot always isolated from the necrotic lesions(37). Apparently no precautions have beentaken to isolate psychrophilic bacteria fromshell disease.The role that psychrophiles play as producers
of growth factors (e.g., B12), as food for higherforms, or as producers of hormonal-like com-pounds has yet to be assessed.
GROWTH OF PSYCHROPHILICBACTERIA
Growth. The psychrotrophic bacteria de-scribed in the literature are actually slow grow-ing at low temperatures and therefore are notrepresentative of the psychrophiles that meetthe definition in this review. In the figures andtables presented by various authors (e.g., 52, 78,79, 83, 168, 174), the growth of the psychro-trophic bacteria at low temperatures (0 to 10 C)may be in the order of many hours, days, or evenweeks. Because of the past and current data inthe literature, the commonly accepted conceptis that low temperature is iminical to the growthof "psychrophilic" bacteria. This concept is notvalid when applied to psychrophiles.
In order to refute the common concept thatlow cell yields and slow grow rates are manifes-tations of low incubation temperatures for bac-teria, Morita and Albright (126) demonstratedthat the psychrophile V. marinus MP-1 grows
150 MORITA
on August 4, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from
PSYCHROPHILIC BACTERIA
well at low temperature. The best cell yieldsfound in the literature for previously describedpsychrotrophic strains of Pseudomonas fluores-cens are by Hess (79) and Sinclair and Stokes(168). The former demonstrated that 9 x 108cells could be obtained at 20 C in 72 h, 9.0 x 104at5 C in72h, and3.4 x 103at0 Cin72h.Thelatter was able to obtain 1.8 x 1010 cells/ml at30 C in 30 h and 1.7 x 1010 cells/ml at 10 C in 72h. These data can be contrasted with the cellyields of V. marinus (126) which were 1.3 x 1012cells/ml at 15 C in 24 h and 9 x 109 cells/ml at3 C in 24 h. It can be readily seen that lowtemperatures of incubation for psychrophilesare not detrimental to large cell yields.Table 5 lists some of the generation times for
previously reported psychrotrophic bacteria. Itcan be noted that the generation times are verylong, especially for temperatures close to 0 C. Incontrast the psychrophiles grow very well at lowtemperatures (31, 71, 123, 154, 166). The gener-ation times for V. marinus at 15 and 3 C,respectively, are 80.7 and 226 min (126). Unfor-tunately the growth characteristics of all psy-
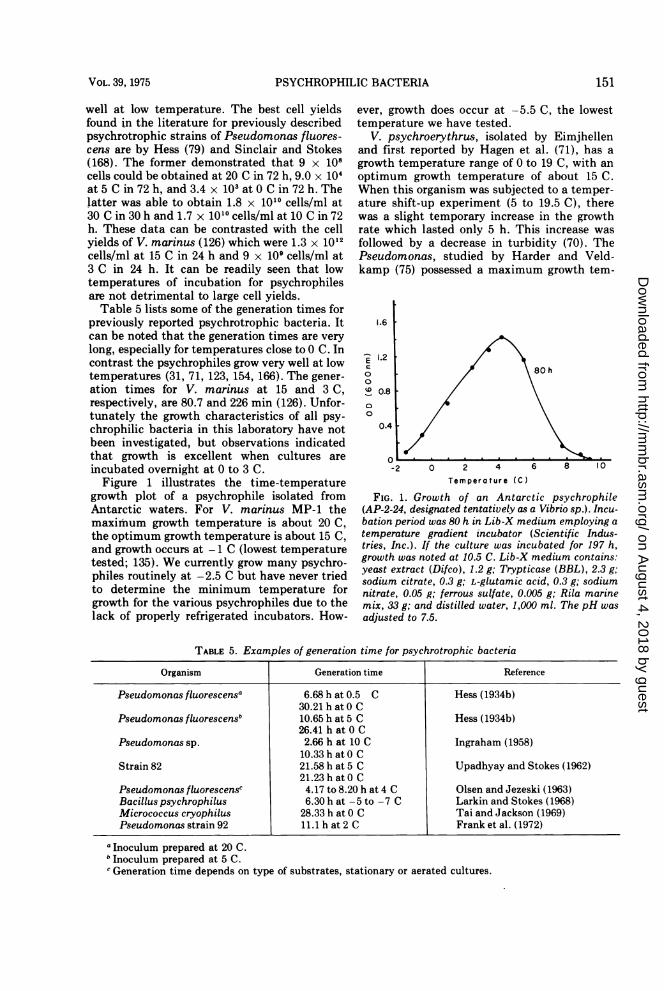
chrophilic bacteria in this laboratory have notbeen investigated, but observations indicatedthat growth is excellent when cultures areincubated overnight at 0 to 3 C.Figure 1 illustrates the time-temperature
growth plot of a psychrophile isolated fromAntarctic waters. For V. marinus MP-1 themaximum growth temperature is about 20 C,the optimum growth temperature is about 15 C,and growth occurs at - 1 C (lowest temperaturetested; 135). We currently grow many psychro-philes routinely at -2.5 C but have never triedto determine the minimum temperature forgrowth for the various psychrophiles due to thelack of properly refrigerated incubators. How-
ever, growth does occur at -5.5 C, the lowesttemperature we have tested.
V. psychroerythrus, isolated by Eimjhellenand first reported by Hagen et al. (71), has a
growth temperature range of 0 to 19 C, with an
optimum growth temperature of about 15 C.When this organism was subjected to a temper-ature shift-up experiment (5 to 19.5 C), therewas a slight temporary increase in the growthrate which lasted only 5 h. This increase was
followed by a decrease in turbidity (70). ThePseudomonas, studied by Harder and Veld-kamp (75) possessed a maximum growth tem-
1.6
O 1.2
00
@ 0.8
0
0.4
0
-2 0 2 4 6 8 10Temperature (C)
FIG. 1. Growth of an Antarctic psychrophile(AP-2-24, designated tentatively as a Vibrio sp.). Incu-bation period was 80 h in Lib-X medium employing a
temperature gradient incubator (Scientific Indus-tries, Inc.). If the culture was incubated for 197 h,growth was noted at 10.5 C. Lib-X medium contains:yeast extract (Difco), 1.2 g; Trypticase (BBL), 2.3 g;sodium citrate, 0.3 g; L-glutamic acid, 0.3 g; sodiumnitrate, 0.05 g; ferrous sulfate, 0.005 g; Rila marinemix, 33 g; and distilled water, 1,000 ml. The pH was
adjusted to 7.5.
TABLE 5. Examples of generation time for psychrotrophic bacteria
Organism Generation time Reference
Pseudomonas fluorescensa 6.68 h at 0.5 C Hess (1934b)30.21hatO C
Pseudomonas fluorescens0 10.65 h at 5 C Hess (1934b)26.41 h at 0 C
Pseudomonas sp. 2.66 h at 10 C Ingraham (1958)10.33hat0 C
Strain 82 21.58 h at 5 C Upadhyay and Stokes (1962)21.23hatO C
Pseudomonas fluorescensc 4.17 to 8.20 h at 4 C Olsen and Jezeski (1963)Bacillus psychrophilus 6.30 h at -5 to -7 C Larkin and Stokes (1968)Micrococcus cryophilus 28.33 h at 0 C Tai and Jackson (1969)Pseudomonas strain 92 11.1 h at 2 C Frank et al. (1972)
a Inoculum prepared at 20 C.b Inoculum prepared at 5 C.c Generation time depends on type of substrates, stationary or aerated cultures.
151VOL. 39, 1975
on August 4, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from
BACTERIOL. REV.
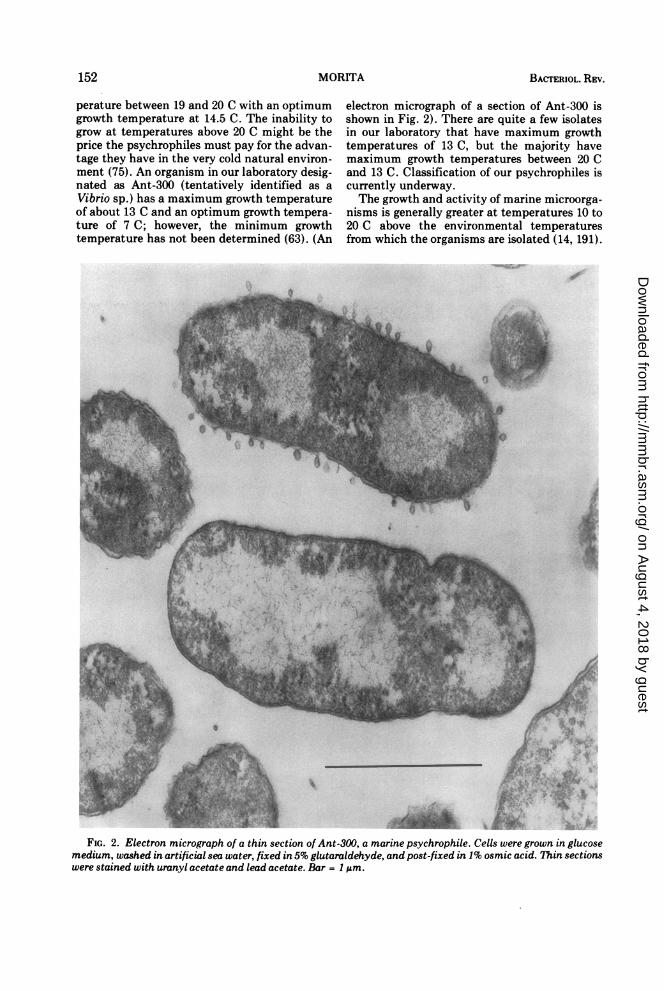
perature between 19 and 20 C with an optimumgrowth temperature at 14.5 C. The inability togrow at temperatures above 20 C might be theprice the psychrophiles must pay for the advan-tage they have in the very cold natural environ-ment (75). An organism in our laboratory desig-nated as Ant-300 (tentatively identified as aVibrio sp.) has a maximum growth temperatureof about 13 C and an optimum growth tempera-ture of 7 C; however, the minimum growthtemperature has not been determined (63). (An
electron micrograph of a section of Ant-300 isshown in Fig. 2). There are quite a few isolatesin our laboratory that have maximum growthtemperatures of 13 C, but the majority havemaximum growth temperatures between 20 Cand 13 C. Classification of our psychrophiles iscurrently underway.The growth and activity of marine microorga-
nisms is generally greater at temperatures 10 to20 C above the environmental temperaturesfrom which the organisms are isolated (14, 191).
FIG. 2. Electron micrograph of a thin section of Ant-300, a marine psychrophile. Cells were grown in glucosemedium, washed in artificial sea water, fixed in 5% glutaraldehyde, and post-fixed in 1% osmic acid. Thin sectionswere stained with uranyl acetate and lead acetate. Bar = 1 jAm.
152 MORITA
on August 4, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from

PSYCHROPHILIC BACTERIA
Although the maximum growth rate is attainedat optimal growth temperature, the maximumcell crop is obtained when organisms are incu-bated below the optimum for growth for ex-tended periods of incubation, probably due tothe efficient use of substrates (167). This situa-tion holds true for Ant-300 (63). Generally theocean environment is low in substrates, and thelow temperature of the environment allows theutilization of the available substrates efficiently.Nevertheless the ability of psychrophiles tofunction well in a cold environment has beennoted (131, 132, 133, 134). Ten to 20 C above theorganism's original environmental temperaturemay be too high for many of the currentlyknown psychrophiles. However, this generalconcept probably does apply to the psychro-trophic bacteria. Many laboratory studies onmarine bacteria do not generally employ envi-ronmental temperatures, and therefore the datais difficult to extrapolate to the environment.Temperature characteristic for growth. It
was suggested that the temperature characteris-tic for growth (Au) could be used to determinewhether or not microorganisms were psychro-philic or mesophilic (83). The bacterial growthrate is substituted for the reaction rate in thevan't Hoff-Arrhenius equation to obtain the Atvalue; g is then analogous to the activationenergy. Hence psychrophiles should have alower AL value than mesophiles. This conceptwas found to be erroneous (73, 162). Ingraham(83) and Shaw (162) used a mesophile and apsychrotroph to obtain their results, whereasHanus and Morita (74) employed a psychro-phile, a psychrotroph, and a mesophile allbelonging to the same genus. Baig and Hopton(6) found that organisms isolated from naturalwater having the fastest growth rates at 5 and10 C had the lowest g values and that thecorrelation is best when temperatures from 15 to25 C are employed. Nevertheless, they statethat a low A value and psychrophilic propertiesmay be unrelated attributes of the bacteriacapable of persisting in natural waters. Workingwith a thermophile and comparing their datawith Hanus and Morita (74), Ward and Cockson(184) could find no correlation between g andthe optimal growth temperatures.Harder and Veldkamp (77) determined the
maximum specific growth rates of four psychro-trophic and four psychrophilic bacteria. Theirdata show that the difference is found in thetemperature range in which the Arrhenius plotsare linear, rather than in different temperaturecharacteristics of growth. Deviation from linear-ity was at -4.5 to -5.0 C and 4 to 5 C for thepsychrophiles and the psychrotrophs, respec-
tively, at the lower end of the thermal spectrumof growth. Their data helps establish the factthat making an Arrhenius plot for a microorga-nism's growth response to temperatures is ex-cessive work when simple growth curves of theorganism at different temperatures will giveessentially the same results.
PHYSIOLOGY OF PSYCHROPHILESThe genetic constitution of the cell deter-
mines its cardinal temperatures for growth.Although this premise is generally accepted, thedata in the literature are completely lacking inorder to postulate the exact genetic basis ofpsychrophily. Most of the research emphasishave been placed on physiological studies.Nevertheless, there is genetic evidence for theconversion of a mesophile to a psychrotroph bytransduction (145), ultraviolet light (5), and themutation by ultraviolet light from a psychro-troph to a mesophile (176). Since phage havebeen isolated from psychrotrophic bacteria (43,44, 99) transduction between cold-loving orga-nisms is possible. Unfortunately none of thehost cells for the phage are psychrophiles. Itappears unlikely that either mutagenesis byultraviolet irradiation or transduction wouldchange the total enzymic make-up of cellssufficiently to promote psychrophily, sincethere appears to be more than one enzyme in apsychrophilic cell that is abnormally thermola-bile.One of the hypotheses of microbial evolution
is that thermophiles were the first to evolve(20), followed by the mesophiles and then thepsychrophiles. The evolution of psychrophilicbacteria is probably due to many genetic eventssince a psychrophile has more than one enzymethat is abnormally thermolabile and has differ-ences in the membrane. According to Allen (4)organisms that have lived in a cold environmentfor generations may have experienced mutationthat caused some of their critical proteins orprotein complexes to become relatively heatlabile. There are probably many pathways lead-ing to psychrophily, since we have found that aspecific abnormally thermolabile enzyme in onepsychrophile may not be abnormally thermola-bile in another psychrophile.
In the eighth edition of Bergey (25), V.marinus MP-1 is classified as V. fisherii. Bio-chemical characteristics used in taxonomy donot characterize the thermal properties of en-zymes or membranes. There is at present nodata in the literature that indicate how thecode (probably sequence of codons) for "meso-philic" enzyme differs from the code for "psy-
VOL. 39, 1975 153
on August 4, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from

BACTERIOL. REV.
crophilic" enzyme. The thermal properties ofpsychrophiles should be taken into considera-tion in bacterial classification, if we accept thehypothesis that the psychrophiles evolved frommesophiles.
Krajewska and Szer (97) conducted compara-tive studies of amino acid incorporation in acell-free system of a psychrotrophic Pseudo-monas and Escherichia coli and found thatthe ribosomes and supernatant fractions ofthe psychrotrophic Pseudomonas and E. coliare interchangeable at 30 to 45 C in the polyU-promoted phenylalanine incorporation. At 2to 9 C only the psychrotrophic Pseudomonasribosomes are active with both supernatantfractions, and therefore the ability to performprotein synthesis at low temperature is relatedto ribosomal specificity. These authors alsopoint out that the psychrotrophic system oper-ates at a relatively high fidelity compared to themesophilic and thermophilic subcellular sys-tems. Does this indicate that through evolutionthe psychrophiles are more stable in terms oftheir genetic fidelity compared to the meso-philes and thermophiles?A positive correlation was found between the
thermal stability of the ribosomes and themaximal growth temperature of the organismsstudied. The percentage of guanine plus cyto-sine content of ribosomal ribonucleic acid(RNA) for V. marinus MP-1 and Bacillusstearothermophilus was 52.1 and 59.3 mol%,respectively. The percentage of guanine pluscytosine content of the deoxyribonucleic acid(DNA) was reported for the two organisms justmentioned as 40 and 51.5 mol%, respectively.Pace and Campbell (147) state that the thermalstability of the ribosome lies in the interactionbetween the RNA and the ribosomal protein.Numerous reasons have been put forth to
explain why microorganisms expire when ex-posed to temperatures above their maximalgrowth temperature, but most of the researchhas been done with mesophiles. These explana-tions include the accelerated use of the intracel-lular amino acid pool (70), enzyme inactivationby elevated temperature (50, 100), changes inextent of cellular lipid saturation (91), disrup-tion of the intracellular organization (84), inac-tivation of an enzyme-forming system (138,178), the loss of permeability control (129), andthermally induced leakage of ribonucleotides,ammonia, ninhydrin-positive material, potas-sium, and phosphate (173).
Effect of temperature on respiration andviability. Psychrophilic bacteria function wellin terms of growth and respiration at environ-mental temperatures from which they were
isolated. V. marinus MP-1 (grown at 15 C) wasshown to be able to respire very well (13 ,u ofoxygen uptake/mg of cell protein per 30 min) atits optimal growth temperature (15 C) and nearits environmental temperature (4 C; 6.5 qI ofoxygen uptake/mg of cell protein per 30 min).Cells grown at 4 C utilized 7.25 ,ul of oxygen/mgof cell protein per 30 min at 15 C and 3.5 ,ul ofoxygen/mg of cell protein per 30 min at 4 C (72).The temperature at which the cells were growngoverns their oxygen uptake. This same patternof results was also shown by Christian andWiebe (31). This situation could be attributedto the fluidity of the membrane at differenttemperatures, which, in turn, is reflected in theability of the cells to transport various sub-strates at different temperatures.The respiratory ability of psychrophiles in-
creases during the initial heat treatment attemperatures near and slightly above the max-imum temperature for growth (31, 63, 75).Harder and Veldkamp (75), working with apsychrophilic Pseudomonas in continuous cul-ture, found that the maximum growth tempera-ture for the organism was 20 C, yet the max-imum rate for respiration was at 23 C. Althoughthe actual Q02 increased towards the maximumand minimum growth temperatures, there was aconcomitant decrease in the molar growthyields. This decrease in the molar growth yieldssuggested to Harder and Veldkamp (75) thatthe energy needed per unit weight per unit timeto maintain a certain growth increased withincreasing deviation from the optimum growthtemperature. Christian and Wiebe (31) restateHarder and Veldkamp's (75) and Brown's (23)results by saying that the portion of energycapable of being produced via respiration,which is utilized for reproduction, decreases astemperature increases.At 13 C (maximum growth temperature for
Ant-300) and at slightly higher temperatures,the amount of carbon dioxide respired de-creased with exposure time in the presence ofexogenous glucose (Table 6). This decrease inrespired carbon dioxide appeared to be theresult of heat-induced restriction on glucoseuptake which, in turn, accelerates starvation ofthe cells (63).An exposure of 110 min to 20.4 C will greatly
reduce the ability of V. marinus MP-1 to takeup oxygen endogenously or in the presence ofglucose, thereby indicating that a thermal le-sion(s) in the cells has taken place (157). Thisthermal injury appears to be a sequence ofevents since V. marinus MP-1 goes from anuninjured state rapidly through a state of re-versible injury and then to a condition of
154 MORITA
on August 4, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from
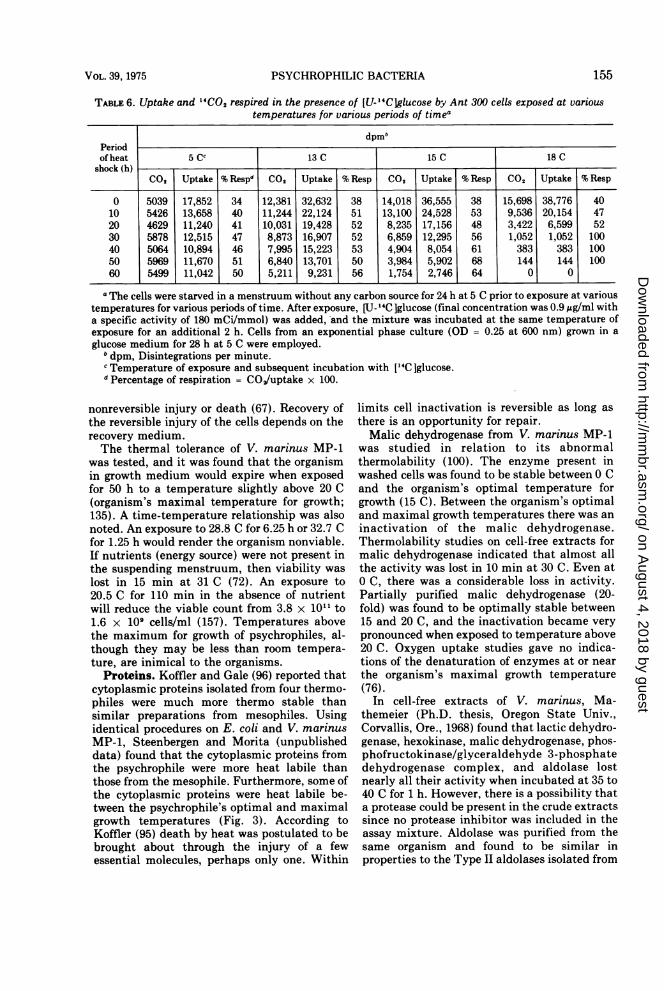
TABLE 6. Uptake and "CO2 respired in the presence of [U-14Clglucose by Ant 300 cells exposed at varioustemperatures for various periods of timea
dpmbPeriodof heat 5 cc 13 C 15 C 18 C
shock (h)00o Uptake % Respd CO, Uptake % Resp CO,2 Uptake % Resp CO, Uptake % Resp
0 5039 17,852 34 12,381 32,632 38 14,018 36,555 38 15,698 38,776 4010 5426 13,658 40 11,244 22,124 51 13,100 24,528 53 9,536 20,154 4720 4629 11,240 41 10,031 19,428 52 8,235 17,156 48 3,422 6,599 5230 5878 12,515 47 8,873 16,907 52 6,859 12,295 56 1,052 1,052 10040 5064 10,894 46 7,995 15,223 53 4,904 8,054 61 383 383 10050 5969 11,670 51 6,840 13,701 50 3,984 5,902 68 144 144 10060 5499 11,042 50 5,211 9,231 56 1,754 2,746 64 0 0
a The cells were starved in a menstruum without any carbon source for 24 h at 5 C prior to exposure at varioustemperatures for various periods of time. After exposure, [U- 14C ]glucose (final concentration was 0.9 Ag/ml witha specific activity of 180 mCi/mmol) was added, and the mixture was incubated at the same temperature ofexposure for an additional 2 h. Cells from an exponential phase culture (OD = 0.25 at 600 nm) grown in aglucose medium for 28 h at 5 C were employed.
b dpm, Disintegrations per minute.Temperature of exposure and subsequent incubation with [4C ]glucose.
dPercentage of respiration = CO/uptake x 100.
nonreversible injury or death (67). Recovery ofthe reversible injury of the cells depends on therecovery medium.The thermal tolerance of V. marinus MP-1
was tested, and it was found that the organismin growth medium would expire when exposedfor 50 h to a temperature slightly above 20 C(organism's maximal temperature for growth;135). A time-temperature relationship was alsonoted. An exposure to 28.8 C for 6.25 h or 32.7 Cfor 1.25 h would render the organism nonviable.If nutrients (energy source) were not present inthe suspending menstruum, then viability waslost in 15 min at 31 C (72). An exposure to20.5 C for 110 min in the absence of nutrientwill reduce the viable count from 3.8 x 1011 to1.6 x 109 cells/ml (157). Temperatures abovethe maximum for growth of psychrophiles, al-though they may be less than room tempera-ture, are inimical to the organisms.
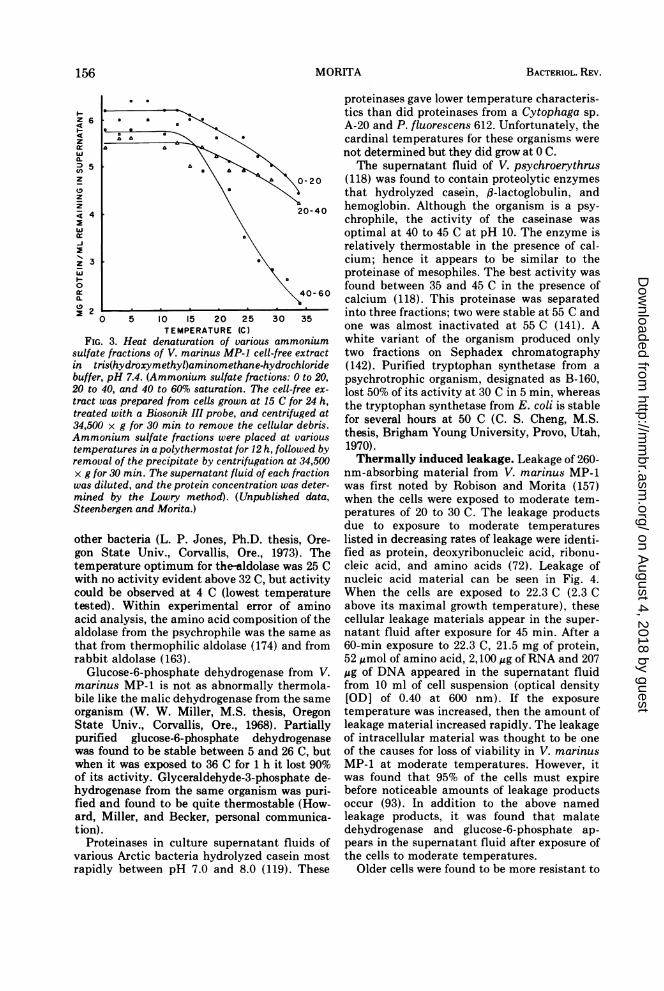
Proteins. Koffler and Gale (96) reported thatcytoplasmic proteins isolated from four thermo-philes were much more thermo stable thansimilar preparations from mesophiles. Usingidentical procedures on E. coli and V. marinusMP-1, Steenbergen and Morita (unpublisheddata) found that the cytoplasmic proteins fromthe psychrophile were more heat labile thanthose from the mesophile. Furthermore, some ofthe cytoplasmic proteins were heat labile be-tween the psychrophile's optimal and maximalgrowth temperatures (Fig. 3). According toKoffler (95) death by heat was postulated to bebrought about through the injury of a fewessential molecules, perhaps only one. Within
limits cell inactivation is reversible as long asthere is an opportunity for repair.Malic dehydrogenase from V. marinus MP-1
was studied in relation to its abnormalthermolability (100). The enzyme present inwashed cells was found to be stable between 0 Cand the organism's optimal temperature forgrowth (15 C). Between the organism's optimaland maximal growth temperatures there was aninactivation of the malic dehydrogenase.Thermolability studies on cell-free extracts formalic dehydrogenase indicated that almost allthe activity was lost in 10 min at 30 C. Even at0 C, there was a considerable loss in activity.Partially purified malic dehydrogenase (20-fold) was found to be optimally stable between15 and 20 C, and the inactivation became verypronounced when exposed to temperature above20 C. Oxygen uptake studies gave no indica-tions of the denaturation of enzymes at or nearthe organism's maximal growth temperature(76).
In cell-free extracts of V. marinus, Ma-themeier (Ph.D. thesis, Oregon State Univ.,Corvallis, Ore., 1968) found that lactic dehydro-genase, hexokinase, malic dehydrogenase, phos-phofructokinase/glyceraldehyde 3-phosphatedehydrogenase complex, and aldolase lostnearly all their activity when incubated at 35 to40 C for 1 h. However, there is a possibility thata protease could be present in the crude extractssince no protease inhibitor was included in theassay mixture. Aldolase was purified from thesame organism and found to be similar inproperties to the Type II aldolases isolated from
VOL. 39, 1975 PSYCHROPHILIC BACTERIA 155
on August 4, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from
BACTERIOL. REV.
Z 6
CLi
z -20-20.az
-2 4 20-40
- \0
C: . 40- 60
2
CLl
0 5 10 15 20 25 30 35TEMPERATURE (C)
FIG. 3. Heat denaturation of various ammoniumsulfate fractions of V. marinus MP-1 cell-free extractin tris(hydroxymethyl)aminomethane-hydrochloridebuffer, pH 7.4. (Ammonium sulfate fractions: 0 to 20,20 to 40, and 40 to 60% saturation. The cell-free ex-
tract was prepared from cells grown at 15 C for 24 h,treated with a Biosonik III probe, and centrifuged at34,500 x g for 30 min to remove the cellular debris.Ammonium sulfate fractions were placed at varioustemperatures in a polythermostat for 12 h, followed byremoval of the precipitate by centrifugation at 34,500x g for 30 min. The supernatant fluid of each fractionwas diluted, and the protein concentration was deter-mined by the Lowry method). (Unpublished data,Steenbergen and Morita.)
other bacteria (L. P. Jones, Ph.D. thesis, Ore-gon State Univ., Corvallis, Ore., 1973). Thetemperature optimum for the-aldolase was 25 Cwith no activity evident above 32 C, but activitycould be observed at 4 C (lowest temperaturetested). Within experimental error of aminoacid analysis, the amino acid composition of thealdolase from the psychrophile was the same asthat from thermophilic aldolase (174) and fromrabbit aldolase (163).
Glucose-6-phosphate dehydrogenase from V.marinus MP-1 is not as abnormally thermola-bile like the malic dehydrogenase from the sameorganism (W. W. Miller, M.S. thesis, OregonState Univ., Corvallis, Ore., 1968). Partiallypurified glucose-6-phosphate dehydrogenasewas found to be stable between 5 and 26 C, butwhen it was exposed to 36 C for 1 h it lost 90%of its activity. Glyceraldehyde-3-phosphate de-hydrogenase from the same organism was puri-fied and found to be quite thermostable (How-ard, Miller, and Becker, personal communica-tion).Proteinases in culture supernatant fluids of
various Arctic bacteria hydrolyzed casein mostrapidly between pH 7.0 and 8.0 (119). These
proteinases gave lower temperature characteris-tics than did proteinases from a Cytophaga sp.A-20 and P. fluorescens 612. Unfortunately, thecardinal temperatures for these organisms werenot determined but they did grow at 0 C.The supernatant fluid of V. psychroerythrus
(118) was found to contain proteolytic enzymesthat hydrolyzed casein, f3-lactoglobulin, andhemoglobin. Although the organism is a psy-chrophile, the activity of the caseinase wasoptimal at 40 to 45 C at pH 10. The enzyme isrelatively thermostable in the presence of cal-cium; hence it appears to be similar to theproteinase of mesophiles. The best activity wasfound between 35 and 45 C in the presence ofcalcium (118). This proteinase was separatedinto three fractions; two were stable at 55 C andone was almost inactivated at 55 C (141). Awhite variant of the organism produced onlytwo fractions on Sephadex chromatography(142). Purified tryptophan synthetase from apsychrotrophic organism, designated as B-160,lost 50% of its activity at 30 C in 5 min, whereasthe tryptophan synthetase from E. coli is stablefor several hours at 50 C (C. S. Cheng, M.S.thesis, Brigham Young University, Provo, Utah,1970).Thermally induced leakage. Leakage of 260-
nm-absorbing material from V. marinus MP-1was first noted by Robison and Morita (157)when the cells were exposed to moderate tem-peratures of 20 to 30 C. The leakage productsdue to exposure to moderate temperatureslisted in decreasing rates of leakage were identi-fied as protein, deoxyribonucleic acid, ribonu-cleic acid, and amino acids (72). Leakage ofnucleic acid material can be seen in Fig. 4.When the cells are exposed to 22.3 C (2.3 Cabove its maximal growth temperature), thesecellular leakage materials appear in the super-natant fluid after exposure for 45 min. After a60-min exposure to 22.3 C, 21.5 mg of protein,52 Atmol of amino acid, 2,100 ,g of RNA and 207jig of DNA appeared in the supernatant fluidfrom 10 ml of cell suspension (optical density[OD] of 0.40 at 600 nm). If the exposuretemperature was increased, then the amount ofleakage material increased rapidly. The leakageof intracellular material was thought to be oneof the causes for loss of viability in V. marinusMP-1 at moderate temperatures. However, itwas found that 95% of the cells must expirebefore noticeable amounts of leakage productsoccur (93). In addition to the above namedleakage products, it was found that malatedehydrogenase and glucose-6-phosphate ap-pears in the supernatant fluid after exposure ofthe cells to moderate temperatures.
Older cells were found to be more resistant to
156 MORITA
on August 4, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from
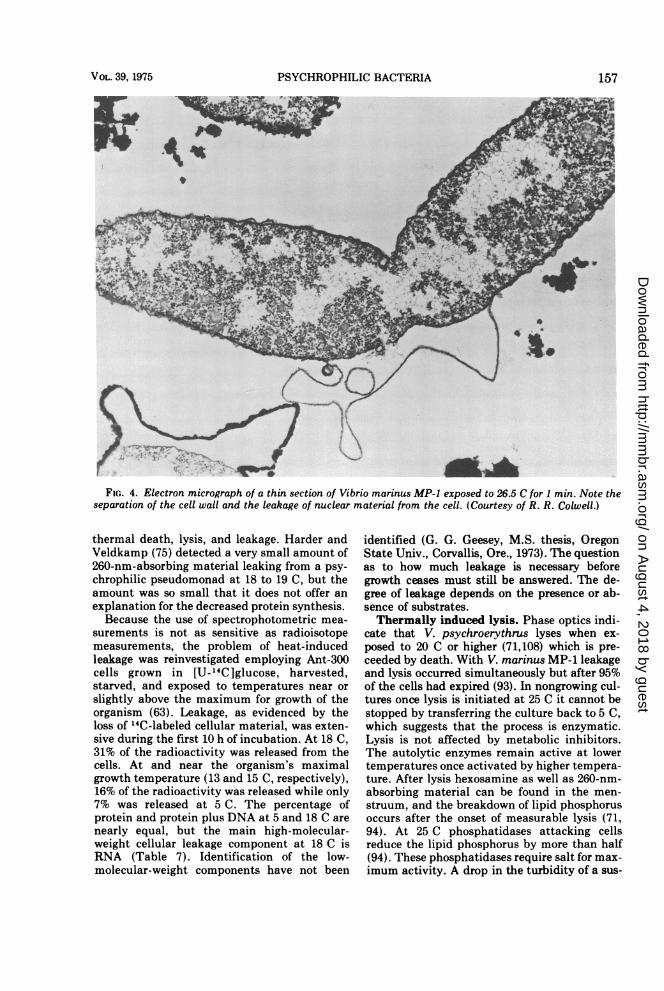
PSYCHROPHILIC BACTERIA
FIG. 4. Electron micrograph of a thin section of Vibrio marinus MP-1 exposed to 26.5 C for I min. Note theseparation of the cell wall and the leakage of nuclear material from the cell. (Courtesy of R. R. Colwell.)
thermal death, lysis, and leakage. Harder andVeldkamp (75) detected a very small amount of260-nm-absorbing material leaking from a psy-chrophilic pseudomonad at 18 to 19 C, but theamount was so small that it does not offer anexplanation for the decreased protein synthesis.Because the use of spectrophotometric mea-
surements is not as sensitive as radioisotopemeasurements, the problem of heat-inducedleakage was reinvestigated employing Ant-300cells grown in [U-_4C]glucose, harvested,starved, and exposed to temperatures near orslightly above the maximum for growth of theorganism (63). Leakage, as evidenced by theloss of "C-labeled cellular material, was exten-sive during the first 10 h of incubation. At 18 C,31% of the radioactivity was released from thecells. At and near the organism's maximalgrowth temperature (13 and 15 C, respectively),16% of the radioactivity was released while only7% was released at 5 C. The percentage ofprotein and protein plus DNA at 5 and 18 C arenearly equal, but the main high-molecular-weight cellular leakage component at 18 C isRNA (Table 7). Identification of the low-molecular-weight components have not been
identified (G. G. Geesey, M.S. thesis, OregonState Univ., Corvallis, Ore., 1973). The questionas to how much leakage is necessary beforegrowth ceases must still be answered. The de-gree of leakage depends on the presence or ab-sence of substrates.Thermally induced lysis. Phase optics indi-
cate that V. psychroerythrus lyses when ex-posed to 20 C or higher (71,108) which is pre-ceeded by death. With V. marinus MP-1 leakageand lysis occurred simultaneously but after 95%of the cells had expired (93). In nongrowing cul-tures once lysis is initiated at 25 C it cannot bestopped by transferring the culture back to 5 C,which suggests that the process is enzymatic.Lysis is not affected by metabolic inhibitors.The autolytic enzymes remain active at lowertemperatures once activated by higher tempera-ture. After lysis hexosamine as well as 260-nm-absorbing material can be found in the men-struum, and the breakdown of lipid phosphorusoccurs after the onset of measurable lysis (71,94). At 25 C phosphatidases attacking cellsreduce the lipid phosphorus by more than half(94). These phosphatidases require salt for max-imum activity. A drop in the turbidity of a sus-
VOL. 39, 1975 157
on August 4, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from

TABLE 7. High-molecular-weight components released from Ant 300 cells at 5 and 18 C for 4 ha
Heatshock ~~~~~~~~~~~~~~~~~~~~~~TotalmaterialHeat shock Protein (%) Protein + DNA (%) RNA (%) released
(dpm/ml)b
5 17 (397 dpm) 18 (409 dpm) 10 (218 dpm) 2,26418 16 (848 dpm) 17 (886 dpm) 14 (744 dpm) 5,161
a Cells were grown at 5 C for 22 h in a medium containing [U- "4C ]glucose (final concentration was 6.0 ,g/mlwith an activity of 180 mCi/ml). The cells were harvested by centrifugation and suspended in artificial seawater to give a final concentration of 2 x 107 cells/ml.
b dpm, Disintegrations per minute.
pension of water-lysed cells or cell envelopes at25 C was also salt dependent.The ultrastructure of V. marinus MP-1 and
V. psychroerythrus appear typical of gram-neg-ative bacteria and both possess a trilaminarmembrane structure (40, 55). Degradation ofthe cell wall and liberation of wall fragmentsoccurs when lysis of the cell takes place at 37 C,and the trilaminar structure is rarely observed(40). This destruction occurs in the presence ofall salt solutions tested.Temperature effects of permeability. The
ability of the psychrophile to function well atcold temperature depends to a great degree onthe availability of substrates. When the temper-ature is raised above the maximum temperaturefor growth, the transport of substrates is im-paired. An apparent increase in malic dehydro-genase activity is noted in a psychrotrophicbacterium when the temperature of the systemis raised above the maximum growth tempera-tures (129). In other words, near-maximal activ-ity occurred even when the intracellular enzymewas being inactivated. This apparent increasein activity with whole cells is due to thealteration in permeability which permits diffu-sion rather than active transport of the sub-strate into the cells.When the temperature is lowered below the
optimal growth temperature, the uptake ofsubstrates is decreased (9, 92, 128, 150, 152).The foregoing data definitely indicate that
changes in transport systems occur when thetemperature is altered. B616hradek (11, 12)suggested that thermal death may be due to thesolution of certain lipids and there was a corre-lation with the melting points of the lipids of thecell and death. Since that time, much researchhas been concentrated on membrane lipids inrelation to growth temperature. As the tempera-ture of growth is lowered, the proportion ofunsaturated fatty acids (hexadecenoic and oc-todecenoic) increases, and this occurs whencells are grown at different temperatures in bothminimal and complex medium (111). This situ-
ation also occurs in a psychrotrophic strain of V.marinus PS 207 (143) and with yeasts (90). Thefatty acid composition of V. psychroerythrus(thought to be a Serratia sp. at the time ofpublication) was analyzed and compared toSerratia marcescens (90). The psychrophile didhave more unsaturated fatty acids than S.marcescens.The percentage of unsaturated fatty acids is
lower in V. marinus PS 207 than in many formsthat have higher cardinal temperatures (143).The relative percentage of unsaturated fattyacids in V. marinus MP-1 (145) is 59.8% whenthe organism is grown at its optimum tempera-ture for growth. The percentage of unsaturatedfatty acids for V. marinus PS 207 grown at 25 C(optimum growth temperature) and 15 C are68.5 and 72.0, respectively. Growth of bothstrains of V. marinus at their temperatureoptima reveal similar fatty acid patterns, butnot when PS 207 is grown at 15 C (143).
It therefore appears that within a speciesthere is a shift toward more unsaturated fattyacids with a decrease in the temperature underwhich the organism is grown. Since V. marinusMP-1 has a much lower percentage of unsatu-rated fatty acid composition (146) compared tomany of the other marine bacteria that are notpsychrophiles (143), the question still remainsas to how psychrophiles can maintain theirmembrane fluidity so that transport systemsare still operative at 0 C or below.Temperature effects on macromolecular
synthesis. When a psychrophilic Pseudomonaswas grown above its optimal temperature forgrowth, there was an increase in the synthesis ofRNA in the cells (75). This increased RNAsynthesis was noted until a temperature of 18 Cwas reached (maximal growth temperature was19 C). Between 18 and 19 C a drop in RNAcontent was noted. If the continuous culturesystem was elevated to 20 C, protein synthesisceased and the culture was washed out of thecontinuous culture apparatus. The increasedRNA of cells at the higher temperature was
158 MORITA BACTERIOL. REV.
on August 4, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from
PSYCHROPHILIC BACTERIA
mainly ribosomal RNA.The ability of V. marinus MP-1 to take up
[U-14C ]proline, synthesize protein (measuredby incorporation of [U-14C]proline), and synthe-size RNA (incorporation of [U-5H]uracil) wasfound to be a function of salinity and tempera-ture. The critical temperature of the lesion(s) inprotein synthesis increased with increasing sa-linity of the growth medium. At a salinity of25%o and 22 C, protein synthesis was signifi-cantly inhibited, but not at a salinity of 35%o(maximal growth temperature at a salinity of35%o is 21.2 C). When cells were incubated at24 C at a salinity of 35%o, protein synthesiscontinues at a linear rate for 20 minutes beforeit begins to decrease. Total RNA synthesiscontinues at a linear rate for 20 min beforesalinities between 15 and 35%o. A salinity-tem-perature relationship is noted for both RNA andprotein synthesis (38). However, it should benoted that both RNA and protein synthesiscontinue in growth media when cells are incu-bated a few degrees above the maximum forgrowth of a psychrophilic vibrio (127). At 5 Cabove the maximal growth temperature, thecessation of DNA and protein synthesis occursapproximately 30 min after exposure to the ele-vated temperature. An increase in protein andDNA was noted at the maximal temperature forgrowth over the amount produced at the op-timum temperature for growth (75).No increase in protein, RNA, and DNA could
be noted by Harder and Veldkamp (76) whencells were exposed in the presence or absence ofsubstrate at 2.5 to 3.5 C above the maximaltemperature for growth. Since no RNA synthe-sis could be detected at 2.5 to 3.5 C above themaximum growth temperature, they concludedthat this may be the basis for the cessation ofgrowth. However, as they point out, this doesnot account for the decline in the growth of theorganism between the optimum and maximumgrowth temperatures. When a cell-free extractprepared from the psychrotrophic bacteriumMicrococcus cryophilus was subjected to 5 Cabove the maximal growth temperature for theorganism, three aminoacyl-tRNA synthetases(glutamic acid, histidine, and proline) werefound to be temperature sensitive (109, 110, 111,112). Unfortunately, the experiments were notcarried out 1 or 2 C above the maximal growthtemperature for M. cryophilus.At temperatures of 1 or 2 C above the maxi-
mal for growth, the cessation of macromolecularsynthesis does not appear to be the cause ofthermal death. Most thermal death experi-ments are carried out at 5 to 10 C above themaximal temperature for growth which is much
too high to determine the subtle changes in theorganism which results in the cessation ofgrowth.Ecological factors affecting psychrophiles.
As stated previously, psychrophiles functionwell at environmental temperatures eventhough their optimum activity and growth maybe several degrees higher than the environmen-tal temperature. The main environmental fac-tors other than the amount of substrates presentthat affect the psychrophiles' ability to functionare salinity, temperature, and hydrostatic pres-sure.The effect of salinity and the various cations
making up the salinity of the ocean on marinebacteria are well reviewed by MacLeod (106,107).Membrane permeability has been shown to
be salt dependent in marine bacteria (45, 46,107).
After washing cells of a marine pseudomonadfree of medium components with 0.05 MMgSO4, there was an immediate increase in ODwhen the cells were suspended in solutionscontaining 200 mM of various salts (116). Thisincrease in OD was followed by a slow decreasein OD. The decrease in OD can be prevented byomitting K+ from the suspending solution. TheOD changes were not attributed to osmoticeffects. The OD of cell envelopes prepared fromthe organism also increased with increases inNaCl but not in the presence of Mg2+. Thisinitial increase in OD was attributed to aninteraction of the salts with the components ofthe cell envelope, thereby causing the contrac-tion of the envelopes and shrinkage of the cells.
.05
.04
.03E .02oC .01
0 S%O 151o100< -.01O -.024 -.03 S= 150
O 2 3 4 5 6 7TIME (MIN) EXPOSED TO VARIOUS SALINITIES
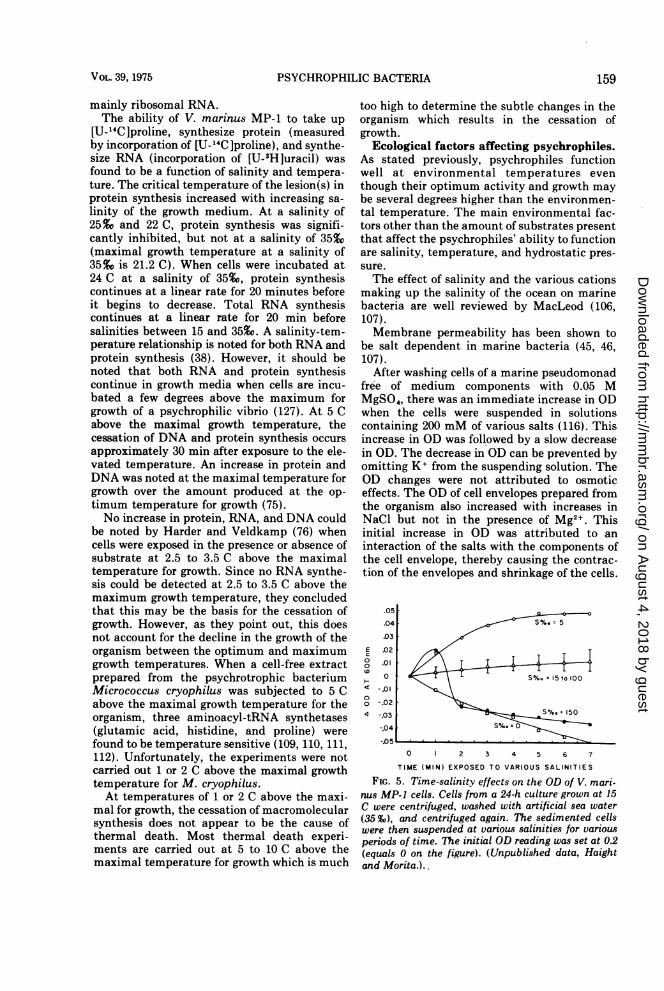
FIG. 5. Time-salinity effects on the OD of V. mari-nus MP-1 cells. Cells from a 24-h culture grown at 15C were centrifuged, washed with artificial sea water(35 Vo), and centrifuged again. The sedimented cellswere then suspended at various salinities for variousperiods of time. The initial OD reading was set at 0.2(equals 0 on the figure). (Unpublished data, Haightand Morita.)..
VOL. 39, 1975 159
on August 4, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from
BACTERIOL. REV.
Salinity changes can also do the same for V.marinus MP-1 (Fig. 5).
Salt and low temperature are necessary forthe stability of the cellular structure of V.psychroerythrus (40, 94), and lysis can occur atlow ionic strength (108). V. psychroerythrus isstable in cold sea water or in a solution contain-ing 0.1 M Mg2+ and 0.5 M NaCl. When sub-jected to water and in various monovalent salts(including 3.0 M NaCl), the turbidity of the cellsuspension fell and some intracellular materialswere released into the menstruum. Phase con-trast microscopy revealed that the cell disinte-grated into amorphous aggregates after firstbecoming swollen. The effectiveness of divalentsalts in maintaining cell structure was in theorder Cu2+ > Ni2+ > Ca2+ > Mn2+ > Mg2+(94).Depending on the salt concentration in the
medium, V. marinus MP-1 can possess a max-imum growth temperature from 10.5 to 21.2 C(170). The lower the salt concentration, thelower the maximum growth temperature of theorganism. This situation holds true for all thesalt-requiring bacteria we have tested in ourlaboratory. In ecological situations where thesalinity and temperature can change due tofreshwater intrusions, evaporation, and diurnalinsolation, the temperature-salinity regime maybe an important factor in the enumeration ofthe microbial flora; hence an important thermalgroup may have been overlooked in microbialecological studies (136). Figure 6 illustrates theco-effect of salinity and temperature variationson the growth of Vibrio marinus MP-1.The change in the maximum growth temper-
ature was shown to be dependent upon thevarious ions present in the growth medium. Theorder of effectiveness in restoring the normalgrowth temperature (21.2 C), when added todilute sea water, was Na+, Li+, Mg2+, K+, Rb+,NH,+. In a completely defined medium, thehighest maximal growth temperature was20.0 C at 0.40 M NaCl. A decrease in themaximal growth temperature was observed atboth low and high concentrations of NaCl.
In order to try to explain the change in themaximal growth temperature in relation to thesalt concentration, studies were initiated todetermine the effect of a salinity-temperatureregime on macromolecular (see section on tem-perature effects on macromolecular synthesis).Rhodes and Payne (155) have found that the
induction of washed cells of Pseudomonasnatriegens (marine bacterium) to the oxidationof lactose and mannitol was dependent on thepresence of Na+ with marine levels of Mg2+ andK+ in the suspending medium. A temperature-salinity relationship was shown for the induc-
50
40 I
30Co
20
10
00 4 8 12 16 20
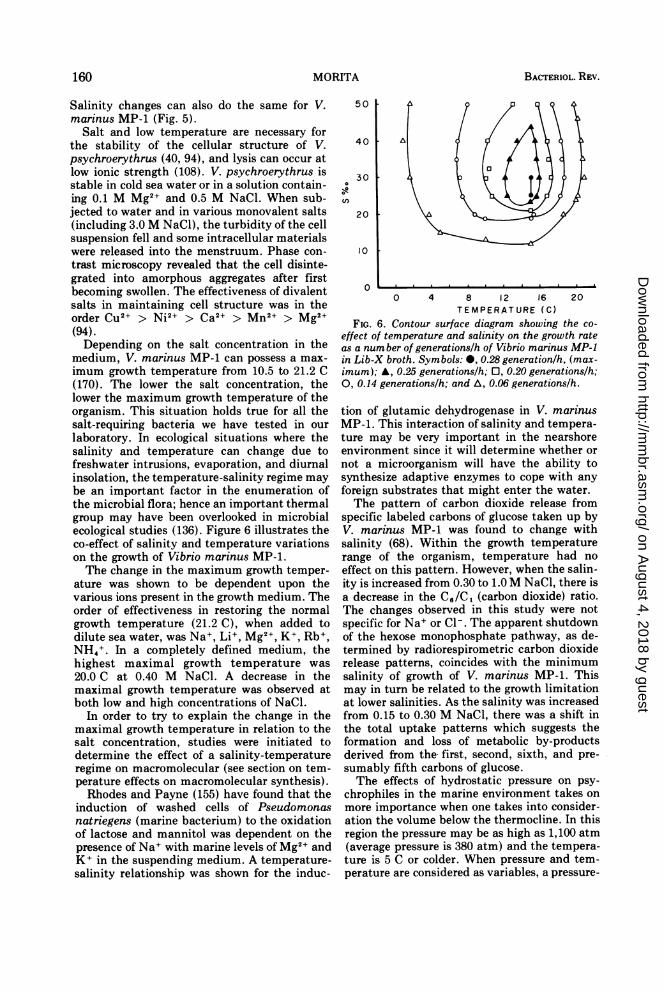
TEMPERATURE (C)FIG. 6. Contour surface diagram showing the co-
effect of temperature and salinity on the growth rateas a number of generations/h of Vibrio marinus MP-1in Lib-X broth. Symbols: 0, 0.28 generation/h, (max-imum); A, 0.25 generations/h; 0, 0.20 generations/h;0, 0.14 generations/h; and A, 0.06 generations/h.
tion of glutamic dehydrogenase in V. marinusMP-1. This interaction of salinity and tempera-ture may be very important in the nearshoreenvironment since it will determine whether ornot a microorganism will have the ability tosynthesize adaptive enzymes to cope with anyforeign substrates that might enter the water.The pattern of carbon dioxide release from
specific labeled carbons of glucose taken up byV. marinus MP-1 was found to change withsalinity (68). Within the growth temperaturerange of the organism, temperature had noeffect on this pattern. However, when the salin-ity is increased from 0.30 to 1.0M NaCl, there isa decrease in the C6/C, (carbon dioxide) ratio.The changes observed in this study were notspecific for Na+ or Cl-. The apparent shutdownof the hexose monophosphate pathway, as de-termined by radiorespirometric carbon dioxiderelease patterns, coincides with the minimumsalinity of growth of V. marinus MP-1. Thismay in turn be related to the growth limitationat lower salinities. As the salinity was increasedfrom 0.15 to 0.30 M NaCl, there was a shift inthe total uptake patterns which suggests theformation and loss of metabolic by-productsderived from the first, second, sixth, and pre-sumably fifth carbons of glucose.The effects of hydrostatic pressure on psy-
chrophiles in the marine environment takes onmore importance when one takes into consider-ation the volume below the thermocline. In thisregion the pressure may be as high as 1,100 atm(average pressure is 380 atm) and the tempera-ture is 5 C or colder. When pressure and tem-perature are considered as variables, a pressure-
160 MORITA
on August 4, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from

PSYCHROPHILIC BACTERIA
temperature interaction is found as would bepredicted by the Ideal Gas Law. The pressureincreases approximately 1 atm for every 10 m indepth. There have been many reviews dealingwith hydrostatic pressure effects on microorga-nisms (19, 89, 122, 124, 169, 190). However, thedata on pressure effects on psychrophiles islimited.
V. marinus MP-1 was isolated from watertaken from a depth of 1,200 m (135) where thetemperature was 3.24 C. It has the ability togrow slightly better at 100 and 200 atm at 3 Cthan at 1 atm at 3 C. With increasing pressureabove 300 atm, growth decreases and at 1,000atm the organism expires. Death ensues at 800atm when the temperature is 15 C (127).The response to pressure depends upon the
environmental temperature from which the or-ganisms were obtained. Hydrostatic pressurestimulated or supressed L-serine deaminationby washed cells of V. marinus MP-1 dependingupon the temperature at which the cells weregrown and the incubation temperature of thereaction mixture (3). Cells grown at 15 or 4 Chad deamination stimulated under pressure inthe following cases: (i) cells grown at 15 C andtested for deamination at 15 C; (ii) cells grownat 4 C and tested at 4 C; and (iii) cells grown at4 C at tested and 15 C. When cells were grown at15 C and tested at 4 C, no stimulation ofdeamination activity due to pressure was ob-served. Unfortunately there is a tendency toneglect this fact when working on the physiolog-ical and biochemical reactions under pressure.The ability of a psychrophile to synthesize
protein, RNA, and DNA under pressure wastested, and it was found that at 200 atmmacromolecular synthesis was not affected. At400 atm protein and RNA but not DNA synthe-sis was adversely affected, whereas at 500 atmthere was a rather sharp decline in the syntheticrates. At 1,000 atm virtually no synthesis oc-curred (2). The effect of alternate pressuriza-tion-depressurization effects on growth and netsynthesis of protein, RNA, and DNA was stud-ied by Albright (1). He found that the applica-tion of 544 atm caused cell division and synthe-sis of RNA and protein to cease immediately.Upon release of the pressure cell division andsynthesis of RNA and protein resumed theoriginal rate. The second application of pressurehad a less drastic effect on growth, and synthesisof RNA and protein. There was no immediateeffect on DNA synthesis with the application ofpressure, but this synthetic process slowed andceased with time under pressure.
Pressure was found to inhibit the transport ofamino acids into a psychrotrophic bacterium(150). Once the amino acids are transported
into the cells, the respiratory mechanismneeded to produce carbon dioxide from theamino acids does not appear to be inhibited.Immediate pressurization of cells in the pres-ence of an amino acid brings about inhibition oftransport, and this transport inhibition is re-versible when the pressure is released. If thecells were incubated in the presence of an aminoacid, 3 h prior to pressurization, the ability ofthe transport mechanism for the amino acids isnot impaired.The uptake of uracil was followed in two
antarctic psychrophiles, V. marinus MP-1 andV. alginolyticus, and E. coli under pressure (8).Between 100 to 300 atm at 15 C, the ability of E.coli and V. alginolyticus to take up uracil wasreduced approximately 70%. The same reduc-tion was noted for V. marinus MP-1 at 5 C. Thetwo antarctic psychrophiles were found to bemore barotolerant since they had the ability totake up considerable uracil at 500 atm. It wasshown that the mechanism for uracil uptake ismore pressure sensitive than the RNA-syntheticmechanism. Growth studies were also con-ducted and correlated with uracil uptake at thesame pressures and temperatures. In each orga-nism there was a correlation between the organ-ism's ability to divide and the rate of uraciluptake. The growth patterns, determined underpressures, of each psychrophile differed mark-edly. Morphological changes (spheroplast-likebodies and elongated forms) were commonlyobserved in cells obtained from cultures sub-jected to pressures limiting their growth. Thesemorphological changes indicate that pressureprofoundly influences the physiology of thecells.The above data on the pressure effects on
psychrophiles strongly implicates the mem-brane as the primary site of pressure damage.
CONCLUSIONThe data available concerning psychrophiles
are too fragmentary to explain the biologicalbasis of psychrophily. Nevertheless, there aresufficient data accumulated to use the term"psychrophile" in a meaningful way. By use ofthe definition employed in this review, a startcan be made to end the confusion concerningthe term. Furthermore, I believe that futurescientific data on psychrophilic bacteria willmore than justify this definition.
ACKNOWLEDGMENTSI wish to thank my associates (J. A. Baross, P. A.
Gillespie, R. P. Griffiths, G. G. Geesey, T. D.Goodrich, F. J. Hanus, S. S. Hayasaka, and L. P.Jones) for use of unpublished scientific data.
This work was supported by National ScienceFoundation research grant GA 38583X.
161VOL. 39, 1975
on August 4, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from

BACTERIOL. REV.
LITERATURE CITED
1. Albright, L. J. 1969. Alternate pressurization-depressurization effects on growth and netprotein, RNA and DNA synthesis by Esche-richia coli and Vibrio marinus. Can. J. Micro-biol. 15:1237-1240.
2. Albright, L. J., and R. Y. Morita. 1968. Effects ofhydrostatic pressure on the synthesis of protein,ribonucleic acid and deoxyribonucleic acid bythe marine psychrophilic bacterium, Vibriomarinus. Limnol. Oceanogr. 13:637-643.
3. Albright, L. J., and R. Y. Morita. 1972. Effect ofenvironmental parameters of low temperatureand hydrostatic pressure on L-serine deamina-tion by Vibrio marinus. J. Oceanogr. Soc. Jpn.28:63-70.
4. Allen, M. B. 1953. The thermophilic aerobicsporeforming bacteria. Bacteriol. Rev.17:125-173.
5. Azuma, Y., S. B. Newton, and L. D. Witter.1962. Production of psychrophilic mutantsfrom mesophilic bacteria by ultraviolet irradi-ation. J. Dairy Sci. 45:1529-1530.
6. Baig, I. A., and J. W. Hopton. 1969. Psychro-philic properties and the temperature charac-teristic of growth of bacteria. J. Bacteriol.100:552-553.
7. Bardach, I. 1924. Ueber die Lebensprozesse derBakterien bei Temperaturen, die unter 0°stehen. Ber. Wiss. Forsch. Inst. Odessa 1:1-5.
8. Baross, J. A., F. J. Hanus, and R. Y. Morita.1974. The effects of hydrostatic pressure onuracil uptake, ribonucleic acid synthesis, andgrowth of three obligately psychrophilic ma-rine vibrios, V. alginolyticus and Escherichiacoli. In R. R. Colwell and R. Y. Morita (ed.),Effect of the ocean environment on microbialactivities. University Park Press, Baltimore.
9. Baur, E. 1902. Ueber zwei denitrifizierende Bak-terien aus der Ostee. Wiss. Meeresuntersu-chungen, Kiel, N. F. 6:9-22.
10. Baxter, R. M., and N. E. Gibbons. 1962. Obser-vations on the physiology of psychrophilism ina yeast. Can. J. Microbiol. 8:511-517.
11. B6lehradek, J. 1931. Le mecanisme physico-chimique de l'adaptation thermique. Proto-plasma 12:406-434.
12. B6lhradek, J. 1935. Temperature and livingmatter. Gebruder Borntraeger, Berlin.
13. Berry, J. A., and C. A. Magoon. 1934. Growth ofmicroorganisms at and below 0C. Phytopa-thology 24:780-796.
14. Braarud, T. 1961. Cultivation of marine orga-nisms as a means of understanding environ-mental influences on populations, p. 271-298.Oceanography. American Association for theAdvancement of Science Publication no. 67.American Association for the Advancement ofScience, Washington, D.C.
15. Borg, A. F. 1960. Studies on myxobacteria asso-ciated with diseases in salmonid fishes. Wildl.Dis. 8:1-85. (2 Microcards.)
16. Boyd, W. L., and J. W. Boyd. 1962. Soil microor-ganisms of the McMurdo Sound Area, Antarc-
tica. Appl. Microbiol. 11:116-121.17. Boylen, C. W., and T. D. Brock. 1973. Bacterial
decomposition processes in Lake Wingra sedi-ments during winter. Limnol. Oceanogr.18:628-634.
18. Brandt, K. 1902. Ueber den Stoffwechsel imMeere. 2. Abhandlung. Wiss. Meeresuntersu-chungen, Kiel, N.F. 6:23-79.
19. Brauer, R. (ed.). 1972. Barobiology and theexperimental biology of the deep sea. NorthCarolina Sea Grant Program, University ofNorth Carolina, Chapel Hill.
20. Brock, T. D. 1967. Life at high temperatures.Science 158:1012-1019.
21. Brock, T. D., F. Passman, and I. Yoder. 1973.Absence of obligately psychrophilic bacteriain constantly cold springs associated withcaves in southern Indiana. Am. Midl. Nat.90:240-246.
22. Brock, T. D., and A. H. Rose. 1969. Psychro-philes and thermophiles, p. 161-168. In J. R.Norris and D. W. Ribbons (ed.), Methods inmicrobiology, vol. 3B. Academic Press Inc.,London.
23. Brown, A. D. 1957. Some general properties of apsychrophilic pseudomonad: the effects oftemperature on some of these properties andthe utilization of glucose by this organism andPseudomonas aeruginosa. J. Gen. Microbiol.17:640-648.
24. Brown, P. E., and R. E. Smith. 1912. Bacterialactivities in frozen soils. Iowa Agric. Exp. Stn.Res. Bull. 4:155-184.
25. Buchanan, R. E., and N. E. Gibbons. 1974.Bergey's manual of determinative bacteriol-ogy, 8th ed. Williams and Wilkins, Baltimore.
26. Bunt, J. S. 1971. Microbial productivity in polarregions, p. 333-354. In D. E. Hughes and A. H.Rose (ed.), Microbes and biological productiv-ity. Symp. Soc. Gen. Microbiol. 21:333-354.
27. Bunt, J. S., and A. D. Rovira. 1955. Microbiolog-ical studies of some sub Antarctic soils. J. SoilSci. 6:119-128.
28. Cameron, R. E., and R. E. Benoit. 1970. Mi-crobial and ecological investigations of recentcinder cones. Deception Island. Antarctica-apreliminary report. Ecology 51:802-809.
29. Cameron, R. E., J. King, and C. N. David. 1970.Microbiology, ecology, and microclimatologyof soil sites in dry valleys of Southern VictoriaLand, Antarctica, p. 702-716. In M. W. Hold-gate (ed.), Antarctic ecology, vol. 2. AcademicPress Inc., London.
30. Campbell Soup Company. 1961. Proceedings ofthe low temperature microbiology sympo-sium-1961. Campbell Soup Co., Camden, N.J.
31. Christian, R. R., and W. J. Wiebe. 1974. Theeffects of temperature upon the reproductionand respiration of a marine obligate psychro-phile. Can. J. Microbiol. 20:1341-1345.
32. Colwell, R. R., and R. Y. Morita. 1964. Reisola-tion and emendation of description of Vibriomarinus (Russell) Ford. J. Bacteriol.88:831-837.
33. Conn, H. J. 1910. Bacteria of frozen soil. Cent.
162 MORITA
on August 4, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from

PSYCHROPHILIC BACTERIA
Bakt. Abt. 2. 28:422-434.34. Conn, H. J. 1912. Bacteria of frozen soil. II. Cent.
Bakt. Abt. 2. 28:70-97.35. Conn, H. J. 1914. Bacteria of frozen soil. III.
Cent. Bakt. Abt. 2. 42:510-519.36. Conradi, H., and H. Vogt. 1901. Ein Beitrag zur
Aetiologie der Weil'schen Krankheit. Z. Hyg.37:283-293.
37. Cook, D. W., and S. R. Lofton. 1973. Chitino-clastic bacteria associated with shell diseasein Penaeus shrimp and the blue crab. J. Wildl.Dis. 9:154-159.
38. Cooper, M. F., and R. Y. Morita. 1972. Interac-tion of salinity and temperature on net proteinsynthesis and viability of Vibrio marinus. Lim-nol. Oceanogr. 17:556-565.
39. D'Aoust, J. Y., and N. N. Gerber. 1974. Isolationand purification of prodigiosin from Vibriopsychroerythrus. J. Bacteriol. 118:756-757.
40. D'Aoust, J. Y., and D. J. Kushner. 1971. Struc-tural changes during lysis of a psychrophilicmarine bacterium. J. Bacteriol. 108:916-927.
41. D'Aoust, J. Y., and D. J. Kushner. 1972. Vibriopsychroerythrus sp. n.: classification of thepsychrophilic marine bacterium, NRC 1004. J.Bacteriol. 111:340-342.
42. Darling, C. A., and Siple, P. A. 1941. Bacteria ofAntarctica. J. Bacteriol. 42:83-98.
43. Delisle, A. L., and R. E. Levin. 1969a. Bacterio-phages of psychrophilic pseudomonads. I.
Host range of phage pools active against fishspoilage and fish-pathogenic psuedomonads.Antonie van Leeuwenhoek J. Microbiol. Serol.35:307-317.
44. Delisle, A. L., and R. E. Levin. 1969b. Bacterio-phages of psychrophilic pseudomonads. II.
Host range of phage active against Pseudomo-nas putrefaciens. Antonie van Leeuwenhoek J.Microbiol. Serol. 35:318-324.
45. Drapeau, G. R., and R. A. MacLeod. 1963. Na+dependent active transport of a-
aminoisobutyric acid into cells of a marinepseudomonad. Biochem. Biophys. Res. Com-mun. 12:111-115.
46. Drapeau, G. R., T. I. Matula, and R. A. Mac-Leod. 1966. Nutrition and metabolism of ma-
rine bacteria. XV. Relation of Na+-activatedtransport to the Na+ requirement of a marinepseudomonad for growth. J. Bacteriol.92:63-71.
47. Druce, R. G., and S. B. Thomas. 1970. Anecological study of psychrotrophic bacteria ofsoil, water, grass and hay. J. Appl. Bacteriol.33:420-435.
48. Eddy, B. P. 1960. The use and meaning of theterm "psychrophilic". J. Appl. Bacteriol.23:189-190.
49. Edsall, G., and L. H. Wetterlow. 1947. Missedcontaminations in biologic products: the roleof cryophile bacteria. J. Bacteriol. 54:31-32.
50. Edwards, 0. F., and L. R. Rettger. 1937. Therelationship of certain respiratory enzymes tothe maximum growth temperatures of bacte-ria. J. Bacteriol. 34:489-515.
51. Ekelof, E. 1908, Bakteriologische Studien wah-
rend der Schwedischen Siidpolar-Expedition1901-1903. W. Erg. der schwed. StidpolarExpedition, 1901-1903, vol. 4. Bonnier, Stock-holm.
52. Elliott, R. P., and H. D. Michener. 1965. Factorsaffecting the growth of psychrophilic microor-ganisms in food-a review. U.S. Departmentof Agriculture Technical Bulletin 1320. U.S.Government Printing Office, Washington,D.C.
53. Fay, A. C., and N. E. Olsen. 1924. How toproduce ice cream with a low.bacterial con-tent. Kansas Agricultural Experiment StationCircular 103.
54. Feitel, R. 1903. BeitrAge zur Kenntniss deni-trifizierender Meeresbakterien. Wiss. Meere-suntersuchungen, Kiel, N. F. 7:91-110.
55. Felter, R. A., R. R. Colwell, and G. B. Chapman.1969. Morphology and round body formationin Vibrio marinus. J. Bacteriol. 99:326-335.
56. Finne, G., and J. R. Matches. 1974. Low-temper-ature-growing clostridia from marine sedi-ments. Can. J. Microbiol. 20:1639-1645.
57. Fischer, B. 1888. Bakterienwachsthum bei OC,sowie uber das Photographieren von Kulturenleuchtender Bakterien in ihrem eigenenLichte. Centr. Bakt. 4:89-92.
58. Fischer, B. 1893. Ueber einige bemerkenswertheBefunde bei der Untersuchung choleraverda-chtigen Materials. Deutsch. Med. Woch.19:541-543, 575-577, 598-599, and 627-629.
59. Forster, J. 1887. Ueber einige Eigenschaftenleuchtender Bakterien. Centr. Bakteriol. Pa-rasitenk. 2:337-340.
60. Forster, J. 1892. Ueber die Entwicklung vonBakterien bei niederen Temperaturen. Centr.Bakt. 12:431-436.
61. Frank, H. A., A. Reid, L. M. Santo, N. A. Lum,and S. T. Sandler. 1972. Similarity in severalproperties of psychrophilic bacteria grown atlow and moderate temperatures. Appl. Micro-biol. 24:571-574.
62. Gazert, H. 1912. Untersuchungen fiber Meeres-bakterien und ihren Einfluss auf den Stoff-wechsel in Meere. Deutsche Siidpolar Expedi-tion, 1901-1903. 7:1-296.
63. Geesey, G. G., and R. Y. Morita. 1975. Somephysiological effects of near maximum growthtemperatures on an obligately psychrophilicmarine bacterium. Can. J. Microbiol. (Inpress).
64. Gounot, A.-M. 1968a. Etude microbiologique deboues glaciaires arctiques. C. R. Acad. Sci.Ser. D. 266:1437-1438.
65. Gounot, A.-M. 1968b. etude microbiologiquedes limons de deux grottes arctique. C. R.Acad. Sci. Ser. D. 266:1619-1620.
66. Gounot, A.-M. 1973. Importance of the tempera-ture factor in the study of cold soils microbiol-ogy. Bull. Ecol. Res. Commun. (Stockholm)17: 172-173.
67. Griffiths, R. P., and R. D. Haight. 1973. Reversi-ble heat injury in the marine psychrophilicbacterium Vibrio marinus MP-1. Can. J. Mi-crobiol. 19:557-561.
VOL. 39, 1975 163
on August 4, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from

BACTERIOL. REV.
68. Griffiths, R. P., and R. Y. Morita. 1973. Salinityeffects on glucose uptake and catabolism inthe obligately psychrophilic bacterium, Vibriomarinus. Mar. Biol. 23:177-182.
69. Gubitz, H. 1928. Kaltebakterien, mit besondererBerticksichtigung ihrer Bedeutung fUr dieMilchwirtschaft. Milchw. Forsch. 5:407-456.
70. Hagen, P. O., and A. H. Rose. 1961. A psychro-philic cryptococcus. Can. J. Microbiol.7:287-294.
71. Hagen, P. D., D. J. Kushner, and N. E. Gibbons.1964. Temperature induced death and lysis ina psychrophilic bacterium. Can. J. Microbiol.10:813-823.
72. Haight, J. J., and R. Y. Morita. 1966. Somephysiological differences in Vibrio marinusgrown at environmental and optimal tempera-ture. Limnol. Oceanogr. 11:470-474.
73. Haight, R. D., and R. Y. Morita. 1966. Ther-mally induced leakage from Vibrio marinus,an obligately psychrophilic bacterium. J. Bac-teriol. 92:1388-1393.
74. Hanus, F. J., and R. Y. Morita. 1968. Signifi-cance of the temperature characteristic ofgrowth. J. Bacteriol. 95:736-737.
75. Harder, W., and H. Veldkamp. 1967. A continu-ous culture study of an obligately psychro-philic Pseudomonas species. Arch. Mikrobiol.59: 123-130.
76. Harder, W., and H. Veldkamp. 1968. Physiologyof an obligately psychrophilic marinePseudomonas species. J. Appl. Bacteriol.31:12-23.
77. Harder, W., and H. Veldkamp. 1971. Competi-tion of marine psychrophilic bacteria at lowtemperatures. Antonie van Leeuwenhoek J.Microbiol. Serol. 37:51-63.
78. Hess, E. 1934a. Cultural characteristics of ma-
rine bacteria in relation to low temperatureand freezing. Can. Biol. Fish. Ser. C.8:461-474.
79. Hess, E. 1934b. Effects of low temperature on
the growth of marine bacteria. Contrib. Can.Biol. Fish. Ser. C. 8:491-505.
80. Hobbie, J. E., and C. C. Crawford, 1969. Respira-tion corrections for bacterial uptake of dis-solved organic compounds in natural waters.Limnol. Oceanogr. 14:528-532.
81. Horowitz-Wlassowa, L. M., and L. D. Grinberg.1933. Zur Frage fiber psychrophile Mikroben.Zentr. Bakteriol. Parasitenkd. Infektionskr.Hyg. Abt. II. 89:54-62.
82. Hucker, G. J. 1954. Low temperature organismsin frozen vegetables. Food Technol. 8:79-108.
83. Ingraham, J. L. 1958. Growth of psychrophilicbacteria. J. Bacteriol. 76:75-80.
84. Ingraham, J. L. 1962. Temperature relation-ships, p. 265-296. In I. C. Gunsalus and R. Y.Stanier (ed.). The Bacteria, vol. 4 AcademicPress Inc., New York.
85. Ingraham, J. L., and J. L. Stokes. 1959. Psychro-philic bacteria. Bacteriol. Rev. 23:97-108.
86. Ingram, M. 1963. Chairman's remarks, p.
185-186. In N. E. Gibbons (ed.), Recent prog-
ress in microbiology. University of TorontoPress, Toronto.
87. Ingram, M. 1965. Psychrophilic and psychro-trophic micro-organisms. Ann. Inst. Pasteur(Lille) 16:111-118.
88. Jacobs, M. B., M. J. Gerstein, and W. G. Walter.1957. Dictionary of microbiology. Van Nos-trand, New York.
89. Johnson, F. H., H. Eyring, and M. J. Polissar.1954. The kinetic basis of molecular biology.John Wiley & Sons, New York.
90. Kates, M., and R. M. Baxter. 1962. Lipidcomposition of mesophilic and psychrophilicyeasts (Candida species) as influenced byenvironmental temperature. Can. J. Biochem.Physiol. 40:1213-1227.
91. Kates, M., and P. 0. Hagen. 1964. Influence oftemperature on fatty acid composition of psy-chrophilic and mesophilic Serratia species.Can. J. Biochem. 42:481-488.
92. Kawasaki, T., I. Miyata, K. Esaki, and Y. Nose.1969. Thiamine uptake in Escherichia coli. I.General properties of thiamine uptake systemin Escherichia coli. Arch. Biochem. Biophys.131:223-230.
93. Kenis, P. R., and R. Y. Morita. 1968. Thermallyinduced leakage of cellular material and via-bility in Vibrio marinus, a psychrophilic ma-rine bacterium. Can. J. Microbiol. 14:1239-1244.
94. Korngold, R. R., and D. J. Kushner. 1968.Responses of a psychrophilic marine bacte-rium to changes in its ionic environment. Can.J. Microbiol. 14:253-263.
95. Koffler, H. 1957. Protoplasmic differences be-tween mesophiles and thermophiles. Bacte-riol. Rev. 21:227-240.
96. Koeffler, H., and G. 0. Gale. 1957. The relativethermostability of cytoplasmic proteins fromthermophilic bacteria. Arch. Biochem. Bio-phys. 76:249-251.
97. Krajewska, E., and W. Szer. 1967. Comparativestudies of amino acid incorporation in a cell-free system from psychrophilic Pseudomonassp. 412. Eur. J. Biochem. 2:250-256.
98. Kruse, W. 1910. Allgemeine Mikrobiologie. F. C.W. Vogel, Leipzig.
99. Kulpa, C. F., Jr., and R. H. Olsen. 1971.Properties of psychrophilic bacteriophage spe-cific for Micrococcus cryophilus. Can. J. Mi-crobiol. 17:157-160.
100. Langridge, P., and R. Y. Morita. 1966.Thermolability of malic dehydrogenase fromthe obligate psychrophile, Vibrio marinus. J.Bacteriol. 92:418-423.
101. Larkin, J. M., and J. L. Stokes. 1968. Growth ofpsychrophilic microorganisms at subzero tem-peratures. Can. J. Microbiol. 14:97-101.
102. Liston, J. 1966. p. 180. In C. H. Oppenheimer(ed.), Marine biology. IV. Unresolved problemin marine microbiology. Proceedings of the 4thInternational Interdisciplinary Conference.New York Academy of Sciences, New York.
103. Llano, G. A. 1962. The terrestrial life of the
164 MORITA
on August 4, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from

PSYCHROPHILIC BACTERIA
Antarctic. Sci. Amer. 207:212-230.104. Lochhead, A. G. 1924. Microbiological studies on
frozen soils. Trans. R. Soc. Can. Sect. III, Ser.III. 18:75.
105. Lochhead, A. G. 1926. The bacterial types occur-
ring in frozen soil. Soil Sci. 21:225-231.106. MacLeod, R. A. 1965. The question of the
existence of specific marine bacteria. Bacte-riol. Rev. 29:9-23.
107. MacLeod, R. A. 1971. Salinity, bacteria, fungi,and blue-green algae, p. 689-703. In 0. Kinne(ed.). Marine ecology, vol. 1, part 2. WileyInterscience Publishing Co., London.
108. Madeley, J. R., R. R. Korngold, D. J. Kushner,and N. E. Gibbons. 1967. The lysis of a
psychrophilic marine bacterium as studied bymicroelectrophoresis. Can. J. Microbiol.13:45-55.
109. Malcolm, N. L. 1968a. Synthesis of protein andribonucleic acid in a psychrophile at normaland restrictive growth temperatures. J. Bac-teriol. 95:1388-1399.
110. Malcolm, N. L. 1968b. A temperature-inducedlesion in amino acid-transfer ribonucleic acidattachment in a psychrophile. Biochim. Bio-phys. Acta 157:493-503.
111. Malcolm, N. L. 1969a. Subunit structure andfunction of Micrococcus cryophilus glutamyltransfer RNA synthetase. Biochim. Biophys.Acta 190:347-357.
112. Malcolm, N. L. 1969b. Molecular determinantsof obligate psychrophily. Nature (London)221:1031-1033.
113. Marr, A. G., and J. L. Ingraham. 1962. Effect oftemperature on the composition of fatty acidsin Escherichia coli. J. Bacteriol. 84: 1260-1267.
114. Marshall, B. J., and D. F. Ohye. 1966. Bacillusmacquariensis n. sp. a psychrotophic bacte-rium from sub-Antarctic soil. J. Gen. Micro-biol. 44:41-46.
115. Matches, J. R., and J. Liston. 1973. Methodsand techniques for the isolation and testing ofclostridia from estuarine environment, p.345-361. In L. H. Stevenson and R. R. Colwell(ed.), Estuarine microbial ecology. Universityof South Carolina Press, Columbia.
116. Matula, T. I., and R. A. MacLeod. 1969. Mecha-nism of optical effects in suspensions of a
marine pseudomonad. J. Bacteriol. 100:403-410.
117. McLean, A. L. 1918. Bacteria of ice and snow inAntarctica. Nature (London) 102:35-39.
118. McDonald, I. J., and A. K. Chambers. 1963.Some characteristics of proteinases of an obli-gately psychrophilic red-pigmented bacteriumand of Serratia marcescens. Can. J. Microbiol.9:871-877.
119. McDonald, I. J., C. Quadling, and A. K. Cham-bers. 1963. Proteolytic activity of some cold-tolerant bacteria from Arctic sediments. Can.J. Microbiol. 9:303-315.
120. Moiroud, A., and A-M. Gounot. 1969. Sur une
bacterie psychrophile obligatoire isolee de li-mons glaciaires. C. R. Acad. Sci. Ser. D.
269:2150-2152.121. Morita, R. Y. 1966. Marine psychrophilic bacte-
ria. Oceanogr. Mar. Biol. Annu. Rev.4: 105-121.
122. Morita, R. Y. 1967. Effects of hydrostatic pres-sure on marine bacteria. Oceanogr. Mar. Biol.Annu. Rev. 5:187-203.
123. Morita, R. Y. 1968. The basic nature of marinepsychrophilic bacteria. Bull. Misaki Mar.Biol. Inst. Kyoto Univ. 12:163-168.
124. Morita, R. Y. 1972. Pressure-bacteria, fungi,and blue-green algae, p. 1381-1388. In 0.Kinne (ed.), Marine ecology, vol. 1, pt. 3.Wiley Interscience Publishing Co., London.
125. Morita, R. Y. 1975. Microbial contribution to theevolution of the "steady state" carbon dioxidesystem. Origins Life 6:37-44.
126. Morita, R. Y., and L. J. Albright. 1965. Cellyields of Vibrio marinus, an obligate psychro-phile, at low temperature. Can. J. Microbiol.11:221-227.
127. Morita, R. Y., and L. J. Albright. 1968. Moder-ate temperature effects on protein, ribonucleicacid and deoxyribonucleic acid synthesis byVibrio marinus, an obligately psychrophilicmarine bacterium. Z. Allg. Mikrobiol.8:269-273.
128. Morita, R. Y., and G. E. Buck. 1974. Lowtemperature inhibition of substrate uptake, p.124-129. Inn R R. Colwell and R. Y. Morita(ed.), Effect of ocean environment on mi-crobial activities. University Park Press, Bal-timore.
129. Morita, R. Y., and S. D. Burton. 1963. Influenceof moderate temperature on the growth andmalic dehydrogenase of a marine psychro-phile. J. Bacteriol. 86:1025-1029.
130. Morita, R. Y., and S. D. Burton. 1970. Occur-rence, possible significance, and metabolismof obligate psychrophiles in marine waters, p.275-285. In D. W. Hood (ed.), Organic matterin natural waters. Publication no. 1, Instituteof Marine Science, University of Alaska, Fair-banks.
131. Morita, R. Y., G. G. Geesey, and T. D. Goodrich.1974. Potential microbial contribution to thecarbon dioxide system in the sea, p. 386-391.In R. R. Colwell and R. Y. Morita (ed.), Effectof the ocean environment on microbial activi-ties. University Park Press, Baltimore.
132. Morita, R. Y., P. A. Gillespie, and L. P. Jones.1971. Microbiology of antarctic sea water.Antarct. J. U.S. 6:157.
133. Morita, R. Y., R. P. Griffiths, and S. S. Hayasa-ka. 1972. Heterotrophic potential of antarcticmarine microorganisms. Antarct. J. U.S. 7:180-181.
134. Morita, R. Y., R. P. Griffiths, and S. S. Hayasa-ka. 1975. Heterotrophic potential of microor-ganisms in Antarctic waters. Third SCAR/IUBS Symposium on Antarctic BiologyN.A.S. (In press).
135. Morita, R. Y., and R. D. Haight. 1964. Tempera-ture effects on the growth of an obligate
165VOL. 39, 1975
on August 4, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from

BACTERIOL. REV.
psychrophilic marine bacterium. Limnol.Oceanogr. 9:102-106.
136. Morita, R. Y., L. P. Jones, R. P. Griffiths, and T.E. Staley. 1973. Salinity and temperatureinteractions and their relationship to the mi-crobiology of the estuarine environment, p.
221-232. In L. H. Stevenson and R. R. Colwell(ed.). Estuarine microbial ecology. Universityof South S. Carolina Press, Columbia.
137. Mossel, D. A. A., and A. van de Mooskijk. 1964.The practical significance in the microbiologi-cal examination of cold stores foods of theallegedly low heat resistance among psychro-trophic organisms. J. Appl. Bacteriol.27:221-223.
138. Muller, M. 1903. Ueber das Wachstum und dieLebenstatigkeit von Bakterien sowie denAblauf fermentativer Prozesse bei niedererTemperatur unter spezieller Beruicksichtigungdes Fleisches als Nahrungsmittel. Arch. Hyg.47:127-193.
139. Nashif, S. A., and F. E. Nelson. 1953. The lipaseof Pseudomonas fragi. II. Fractors affectinglipase production. J. Dairy Sci. 36:471-480.
140. Nedwell, D. B., and G. D. Floodgate. 1971. Theseasonal selection by temperature of hetero-trophic bacteria in an intertidal sediment.Mar. Biol. 11:306-310.
141. Nunokawa, Y., and I. J. McDonald. 1968a.Extracellular proteolytic enzymes of psychro-philic bacteria. I. Purification and some prop-erties of enzymes of an obligately psychro-philic red-pigmented bacterium. Can. J. Mi-crobiol. 14:215-231.
142. Nunokawa, Y., and I. J. McDonald. 1968b.Extracellular proteolytic enzymes of psychro-philic bacteria. II. Production and heterogene-ity of enzymes of an obligately psychrophilicred-pigmented bacterium and of a white vari-ant. Can. J. Microbiol. 14:225-231.
143. Oliver, J. D., and R. R. Colwell. 1973. Extracta-ble lipids of gram-negative marine bacteria:fatty-acid composition. Int. J. Syst. Bacteriol.23:442-458.
144. Olsen, R. H., and J. J. Jezeski. 1963. Someeffects of carbon source, aeration, and temper-ature on growth of a psychrophilic strain ofPseudomonas fluorescens. J. Bacteriol.86:429-433.
145. Olsen, R. H., and E. S. Metcalf. 1968. Conver-sion of mesophilic to psychrophilic bacteria.Science 162:1288-1289.
146. Or6, J., T. G. Tornabene, D. W. Nooner, and E.Gelpi. 1967. Aliphatic hydrocarbons and fattyacids of some marine and freshwater microor-ganisms. J. Bacteriol. 93:1811-1818.
147. Pace, B., and L. L. Campbell. 1967. Correlationof maximal growth temperature and ribosomeheat stability. Proc. Natl. Acad. Sci. U.S.A.57:1110-1116.
148. Pacha, R. E. 1968. Characteristics of Cytophagapsychrophila (Borg) isolated during outbreaksof bacteria cold-water disease. Appl. Micro-biol. 16:97-101.
149. Parsons, R. R., and J. D. H. Strickland. 1962. On
the production of particulate organic carbonby heterotrophic processes in sea water. Deep-Sea Res. 8:211-222.
150. Paul, K. L., and R. Y. Morita. 1971. The effect ofhydrostatic pressure on the uptake and respi-ration of amino acids by a facultatively psy-chrophilic marine bacterium. J. Bacteriol.108:835-843.
151. Pennington, M. E. 1908. Bacterial growth andchemical changes in milk kept at low tempera-ture. J. Biol. Chem. 4:353-393.
152. Piperno, J. R., and D. L. Oxender. 1968. Aminoacid transport systems in Escherichia coliK12. J. Biol. Chem. 243:5914-5920.
153. Ravenel, M. P., E. G. Hastings, and B. W.Hammer. 1910. The bacterial flora of milkheld at low temperatures. J. Infect. Dis.7:38-46.
154. Reed, H. S., and R. R. Reynolds. 1916. Someeffects of temperature upon the growth andactivity of bacteria in milk. Virginia Agricul-tural Experimental Station Technical Bul-letin 10.
155. Rhodes, M. E., and W. J. Payne. 1962. Furtherobservations on effects of cations on enzymeinduction in marine bacteria. Antonie vanLeeuwenhoek J. Microbiol. Serol. 28:302-314.
156. Richardson, W. D. 1908. The cold storage of beefand poultry, p. 261-316. In Premier CongressInternational du Froid, Paris, 1908. Tome 2.Rapports et Communications des Sections I,II, et III.
157. Robison, S. M., and R. Y. Morita. 1966. Theeffect of moderate temperature on the respira-tion and viability of Vibrio marinus. Z. Allg.Mikrobiol. 6:181-187.
158. Rose, A. H. 1962. Temperature relationshipsamong microorganisms. Wallerstein Lab.Commun. 25:5-18.
159. Rubentschik, L. 1925. Ueber die LebenstAtigkeitder Urobakterien bie einer Temperatur unter0°. Zentr. Bakteriol. Parasitenkd. Infek-tionsk. Hyg. Abt. II. 64:166-174.
160. Rubey, W. W. 1951. Geological history of seawater. Bull. Geol. Soc. Am. 62:1111-1147.
161. Schmidt-Nielsen, S. 1902. Ueber einige psychro-phile Mikroorganismen und ihr Vorkommen.Zentr. Bakteriol. Parasitenkd. Infektionsk.Hyg. Abt. II. 9:145-147.
162. Shaw, M. K. 1967. Effect of abrupt temperatureshifts on the growth of mesophilic and psy-chrophilic yeasts. J. Bacteriol. 93:1332-1336.
163. Shimizu, H., and H. Ozawa. 1967. Studies onaldolases. I. Amino acid composition andsubunit structure of rabbit-muscle aldolase.Biochim. Biophys. Acta 133:195-205.
164. Sieburth, J. McN. 1964. Polymorphism of amarine bacterium (Arthrobacter) as a func-tion of multiple temperature optima and nu-trition, p. 11-16. Symposium on ExperimentalMarine Ecology, University of Rhode IslandOccasional Publication no. 2. University ofRhode Island, Kingston.
165. Sieburth, J. McN. 1965. Microbiology of theAntarctic, p. 267-295. In P. V. Oye and J. V.
166 MORITA
on August 4, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from

PSYCHROPHILIC BACTERIA
Mieghem (ed.), Biogeography and ecology inAntarctica. Dr. W. Junk, Pub., The Hague.
166. Sieburth, J. McN. 1967. Seasonal selection ofestuarine bacteria by water temperature. J.Exp. Mar. Biol. Ecol. 1:98-121.
167. Sieburth, J. McN. 1968. Observations on bacte-ria planktonic in Narragansett Bay, RhodeIsland; a resume. Bull. Misaki Mar. Biol. Inst.Kyoto Univ. 12:49-64.
168. Sinclair, N. A., and J. L. Stokes. 1963. Role ofoxygen in the high yields of psychrophiles andmesophiles at low temperature. J. Bacteriol.85:164-167.
169. Sleigh, M. A., and A. G. MacDonald (ed.). 1972.The effects of pressure on organisms. Symp.Soc. Exp. Biol., vol. 26. Academic Press Inc.,New York.
170. Stanley, S. O., and R. Y. Morita. 1968. Salinityeffect on the maximum growth temperature ofsome bacteria isolated from marine environ-ments. J. Bacteriol. 95:169-173.
171. Stanley, S. O., and A. H. Rose. 1967. Bacteriaand yeasts from lakes on Deception Island. R.Soc. (London) Phil. Trans. B. 252:199-207.
172. Starka, R. P., and J. L. Stokes. 1960. Psychro-philic bacteria from Antarctica. J. Bacteriol.80:622-625.
173. Stokes, J. L. 1963. General biology and nomen-
clature of psychrophilic bacteria, p. 187-192.In N. E. Gibbons (ed.), Recent progress inmicrobiology. University of Toronto Press,Toronto.
174. Stokes, J. L. 1968. Nature of psychrophilicmicroorganisms, p. 221-233. In J. Hawthorne(ed.), Low temperature biology of foodstuffs.Pergamon Press, Oxford.
175. Sugimoto, S., and Y. Nosoh. 1971. Thermalproperties of fructose 1,6-diphosphate aldolasefrom thermophilic bacteria. Biochim. Bio-phys. Acta 235:210-221.
176. Tai, P.-C., and H. Jackson. 1969. Mesophilicmutants of an obligate psychrophile, Micro-coccus cryophilus. Can. J. Microbiol.15:1145-1150.
177. Tsiklinsky, M. 1908. La flore microbienne dansles regions du Pole Sud. Expedition antarc-tique francaise, 1903-1905. Masson et cie,Paris.
178. Upadhyay, J., and J. L. Stokes. 1962. Anaerobicgrowth of psychrophilic bacteria. J. Bacteriol.83:270-275.
179. Upadhyay, J., and J. L. Stokes. 1963. Tempera-ture-sensitive formic hydrogenlyase in a psy-chrophilic bacterium. J. Bacteriol.85:177-185.
180. Vanderleck, J. 1917. Bacteria of frozen soils inQuebec. Trans. R. Soc. Can. Ser. III, Sect. IV.11:15-39.
181. Vanderleck, J. 1918. Bacteria of frozen soils inQuebec. II. Trans. R. Soc. Can. Ser. III, Sect.IV. 12:15-39.
182. Vass, A. F. 1919. The influence of low tempera-ture on soil bacteria. N.Y. Agric. Exp. Stn.(Ithaca) Mem. 27:1039-1074.
183. Vishniac, W. V., and S. E. Mainzer. 1973.Antarctica as a Martian model, p. 3-31. In P.H. A. Sneath (ed.), Cospar life sciences andspace Research XI. Akademie-Verlag, Berlin.
184. Ward, J., and A. Cockson. 1972. Studies on athermophilic bacillus: its isolation, properties,and temperature coefficient of growth. J. Bac-teriol. 112:1040-1042.
185. Wiebe, W. J., and C. W. Hendricks. 1971.Simple, reliable cold tray for the recovery andexamination of thermosensitive organisms.Appl. Microbiol. 22:734-735.
186. Wilkins, P. O., R. Bourgeois, and R. G. E.Murray. 1972. Psychrotrophic properties ofListeria monocytogenes. Can. J. Microbiol.18:543-551.
187. Witter, L. D. 1961. Psychrophilic bacteria-areview. J. Dairy Sci. 44:983-1015.
188. Wright, R. T., and J. E. Hobbie. 1965. Theuptake of organic solutes in lake water. Lim-nol. Oceanogr. 10:22-28.
189. Zachariah, P., and J. Liston. 1973. Temperatureadaptability of psychrotrophic Pseudomonas.Appl. Microbiol. 26:437-438.
190. Zimmerman, A. M. (ed.). 1970. High pressureeffects on cellular processes. Academic PressInc., New York.
191. ZoBell, C. E. 1961. Importance of microorga-nisms in the sea, p. 107-132. In Proceedings ofthe low temperature microbiology symposium.Campbell Soup Co., Camden, N.J.
192. ZoBell, C. E., and J. E. Conn. 1940. Studies onthe thermal sensitivity of marine bacteria. J.Bacteriol. 40:223-238.
193. ZoBell, C. E., and S. C. Rittenberg. 1938. Theoccurrence and characteristics of chitinoclas-tic bacteria in the sea. J. Bacteriol.35:275-287.
167VOL. 39, 1975
on August 4, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from