quaternary 3d the - university of toronto t-space · receptor: quaternary 3d reconstruction of the...
TRANSCRIPT

STRUCTURE AND FUNC'MON OF THE HUMAN INSULM
RECEPTOR: QUATERNARY 3D RECONSTRUCTION OF THE
I N S ~ I N S U L I N RECEPTOR - COMPLEX AND THE
STRUCTURAL LOCALIZATLON OF THE INSULIN-BINDING SITE
Submitted to The School of Graduate Studies
of The University of Toronto
ROBERT ZECAO-TIAN LUO
In partial fiilfillment of reqainments
for the degree of
Doctor of Philosophy
O Robert Zhao-Tian Luo, 2000

~ u i s i i s and AcquWons et Bibliographk Services senrices bibriraphques
The author bas grantecl a non- exclusive Iicence ailowing the National Liirary of Caiiada to reproduce, logn, distniute or sell copies of this thesis in microform, paper or electronic fotmats.
L'auteur a accordé une licence non exclusive permettant à la Bibliothèque nationale du Canada de reproduiret prêter, distniuer ou v&e des copies de cette thèse sous la forme de m*Crofiche/5Im, de reproduction sur papier ou sur fonnat electroniqye.
The author tetains ownership of the L'auteur conserve la propriété du copyright in this thesis. Neither the droit d'auteur qoi protège cette thèse. thesis nor substantid extracts fkom it Ni la thèse ni des extraits substantiels may be printed or otherwise de celle-ci ne doivent être imprimés reproduced without the authds ou autrement reproduits sans son permission. autorisation-

TABLE OF CONTENTS
TABLE OF CONTENTS LIST OF FIGURES LIST OF ABREVIATIONS ACKNOWLEDGEMENT
1. INTERCELLULAR COMMUNICATION
2. TEE INSULIN FAMILY OF PEPTIDES 2.1. liisulin 2.2. -L&e Growth Factors 2.3- Other members ofthe Insulin fàmily
3. MOLECULAR AND CELL BIOLOGY OF INSUUN AND IGF-1 RECEPTOR
3 -1. The Receptor Tyrosine Kinase S u p a f d y 3.2. IasulinReceptor 3.3. IR, IGF-IR and Similarity and Wice
4, INSULIN BINDING AND S I G N a ~ S D U C T I O N 4.1. A f k i t y and. Kinetics of rilsulin Binding 4.2. Autophosphorylation 4.3, Post-Receptor Signal Transduction
5. MAPPING TEE INSULIN-BINDING SlTE 5.1 . Insulin Interaction with the IR 5.2. Insulia Binding Domain on the IR
CHAPTER 2 LOCALEATLON OF INSULIN-BINDING SITE@) ONTHE HUMAN INSULIN RECEPTOR BY BIOCHEMICAL APPROACHES
1. INTRODUCTION
2. MATERIAL AND METHODS 2.1. O v e r ~ i o n of the Extracellular Domain of the HIR 2.2. Purification of the Recombinant IR and Huaian Placenta Membrane
IR 53

3 e RESITLTS 63 3 -1, Recombinant HIR ExtraceiluIar Domain 63 3 2 - A Smaü Labeled Peptide Fragment on the Lnsulin Binding Region 69
4. DISCUSSION 76
-R 3 THE QUATERNARY 3D STRUCTURE OF THE
DE- BY SCANMKG TRANSMISSION ELECTRON MI%ROSCOPYr STRUCTURAL LOCALIZATION OF TEE ms-BINDING S m
1. INTRODUCTION 1.1. El-n M*~~~scupic 3-D Reco-=on Techniqges 1.2. Stnictural Detemunation of IR and Locaüzation of riisului Binding
Site
2. MATERIALS AND METHODS 2.1. BiologicalcalMateriais 2.2. Electron Microscopy and ïinage AnaIysis 2.3 . Three Dimensional Reconstruction
3 e RESULTS 3.1. Specimen Preparation for Electron Microscopy 3 -2- Image processing of NG-BI-HIR Complex 3 3 . Lacation of the uisului-Binding Region 3 -4- Domain Structutes of the HIR
4. DISCUSSION
C-R 4 CONCLUSION AND FUTURE RESEARCH 125
iii

LIST OF ETGITRES
Chapter 1 Page
Fig. 1-1. Predicted tertiary structures of the insalin family of peptides Fig. 1-2. The ptimary structure, the 3D crystai structure and the 3D
space-filiing mode1 of human Fnsulln Fig. 1-3. Schematic representation ofthe insulin receptor gene, the primary
and secondary structure of hIR Fig. 1-4. Human insuiin receptor amino acld sequence Fig. 1-5. A schematic diagram of the Insuiin signahg pathways
Chapter 2
Fig. 2-1, Expression of human insulin receptor extracellular domain with bacuiovirus expression system @ES)
Fig. 2-2. Construction of the BES transfection vectors for the bIR ectodomain Fig. 2-3, Insulin photoprobes Fig. 2-4. Tirne course of expression of =or, and Western blot of hlRa
and hlR Fig. 2-5. Immunoprecipitation of B29-MABI labeled 61Ra Fig. 2-6. Affinity purification of expressxi hIRa with anti-hlR monoclonal
antibody (&W5 1) column Fig. 2-7. Insulin-binding assay of hIRa compared with hIR Fig. 2-8. Tirne course of photolysis Fig. 2-9. Time course and products of trypsin digestion of B29-MABI or
AZAP Iabeled hiRa Fig. 2-10. The sensitivity of ELISA in detecting B-chain fragment of bovine
insulin before and after HPLC analysis Fig. 2-1 1. Trypsin digests of hIRa labeled with '=I-B~~-MABI Fig. 2- 12. Achromobacter protease 1 digests of hIRa Iabeled with
'~1-BBPa-riisuiin Fig. 2- 13. HPLC analysis of partiaiiy purified fractions as identified in Fig. 2-12
Chapter 3
Fig. 3-1. Different electron microscopes used in biomedicd studies Fig. 3-2. An overview of STEM 3D reconstruction Fig. 3-3. Outline of the preparation of NGBI Fig. 3-4. Purification of hIR Fig. 3 -5- Purification and characterization of NGBI

Fig. 3 6 . STEM images of purifid NGBI 101 Fig. 3-7. STEM àarkfield Images ofhWNGB1 cornpla~ 103 Fig. 3-8. Composite raw images ofhIR/NGBI mo1ecdes 104 Fig. 3 -9. Low band-pass nltasd images 105 Fig- 3-10. Finai images used for 3D - d o n 106 Fig. 3-1 1. Histogram ofthe. caiculated mass for 1,625 co1Iected images 107 Fig, 3-12. Caldateci relative anguiar orientation for indIvidual molecular images 108 Fig. 3-1 3.3D reconstruction of the hIWNGBI cornplex (&âce representation) 109 Fig. 3-14. The Seconaary domain stnicture ofhIR and the domain Idon in 3D
stnichae 112 Fig. 3 -1 5, Fitting of biochemkal domairis and their lmown =ray crystal dmctwes
to the 3 D mnstructim 113

OC 2D 3D AZAP BBpa BES BI BSA CD CNBr CR DAG DBI DIPEA DMA EGF ELISA EM FGF Fa1 Fn2 ETLC g HGF hIR hIRa HPLC ID IGF-1 IGF-II IGF-1 R IQAD IR IRS-1 IRR kDa kV L1 LI-CR-L2 L2 LDL M MABI
degree Celsnis twodlmensional tbtee-dimensional do-phenyb-aminopropyl biotinyl-benu,ylphen081anine .
baculovinis eqmssïngsystem bovineinsuIin boviaesenunaibumin connecting domain cyanogen bromide cysteine-rich domah diacylglycerol &-&--insulin di-iso-propylenthyhnhe bimethylacetamide epidermai pwthfactor enzyme-iinked immunosorbant assay eI-n microscope ~ b I a s t ~ w t h f i c t o r fibroneCtinîypeIIIrepeaî1 fibronectintypemrepeat2 rapt @ormance Iiquid chromatography P== hepatocyte growth f'actor humaninsulinreceptor human insuiin teceptor extracellular domain high perfiormance liquid chromatography insert domain insulin-like growth factor 1 insulul-like growthfactor II insulia-like growth factor 1 receptor itaative q--assisted angular daermmation insutinreceptor insulin receptor substtate-1 insulin receptor telated recepttor kiloDalton kilovolts large homologous domain 1 Ll-Cysteine-rich-L2 domain large homologous domain 2 low density fipoptein m o k monoazido-Wyl-insulin

mg Crg MIP ml
mM MOI Mr NG NGBI NGF nm NMR PAGE PBS PDB PDGF PTK RPTK SDS SEM SEI2 SK STEM TEM TK TM
microgram d o s c a n insulin-like pepîide
microlitet mrnimolar mdtipIIcity of Infection mofecular welght nanogold nanogold-bovine insulin nerve growthfactor nanometers nuclear magnetic resomnce polyacryl-de gel eIectmphoresis phosphate bufEer solutio~ protein data bank plateletderiveci g~owth fàcîor pmteimtyn,sine kinase recep~rpro~tyrosine kinase sodiumdoedecyl sulfiite scanniaig elcctron microscope Sn= homology 2 serindthreonine kinase .crnnning transmission electron micmscope transmission electron mic~oscope tyrosine kinase transmembrane domain

My greatest thanks are due to my thesis supevisor, Dr, Cecil C. Yip, It is Cecil's iavaIuabIe, enthusiastic inspiration, and never ending guidance that made my research experience at the University of Toronto most wonderfüi. Through hïs dedication and persistence to scientifïc research, Cecil not only showed me how to conduct great science but also taught me how to be a human bemg with great integrity. 1 am also exceedingiy gratefûi to my thesis co-supervisor Dr. F. Peter Ottensmeyer for his excellent gwgwdance, encouragement and giving me the opportunity to access the state of the art STEM and cornputer image processing technique in his lab.
1 am deeply grateful to other members of m y graduate committee: Dr. Daniel Drucker and Dr. Jennifer Dorrîngton. 1 appreciate their contriibutions and cntical suggestions to the fruition of this research-
1 would particularly thank Dr. Margaret Moule for her helpful discussion, encouragement., enthusiasticdy reviewïng m y thesis and her expertise in research during the project 1 would Iike to thank othet colleagues and members in Dr. Yip's lab: Helga Hsu, Guoqing Zhong, Christine Miasos, Elaine Jack and Dr. Teresa Tallenco-Melnyk. Their consistent encouragement, technihnical assistance and friendship helped me nni-sh my program. 1 wouid aiso iike to thank colleagues and members in Dr. Ottensmeyer's lab: Allen Fernandes, Dr. Yongyi Mao, Michael Burke, Dr. Daniel Beniac, Brenda Rutherford, Yew Meng Heng and David McAiduff. Without their technical assistance, inspiring discussion and noble friendship, my project wouid not get any progress.
1 acknowledge the financial support from the University Toronto, the Ontario Goverment and the Medical Research Council of Can&
FiaMy, 1 would like to give my special thanks to my Iovely wife, Dr. Hongqi Peng and daughter Nancy, for their endless support and companionship. Without their sacrifice, 1 wouldn't be able to stand where 1 am. 1 am extremely grateful to my parents, parents-in-law, brothers and sister for their long t h e encouragement and support.

RECEPTOR: QUA-NEY 3D RECONSTRUCTION OF THE
INSULININSULIN RECEPTUE COMPLEX AND THE
STRUCTURAL LOCALIZATIION OF THE INSULIN-BNDING SITE - PhD. Thesis, Robert Zhao-Tian Luo, hstitute of Medicai Science, University of Toronto,
2000
The elucidation of the mechanism of cellular signal transduction is one of the key
challenges in biology. The first step in this pathway has been particularly refiactory to
study. In the case of insuIin action, it is the binding of insulin to its ce11 s d a c e receptor,
a large integral trammembrane glycoprotein, The binding of insulin d t s in the
activation of the cytoplasmic domain of the receptor as a tyrosine kinase, While much is
known about the interaction of the activated receptor with its substrates Ieading to down-
stream signal transduction, how the binding of insului to the extracellular domain of the
receptor can lead to receptor activation remains unknown. The specific goal of this study
is to localize the insuiin-binding site of the insulin receptor (IR), and, if possible, to
elucidate in molecuiar details the insulin-IR interaction and the structure-fwiction
relationship of the insulin receptor,
To achieve this goal with biochemical approach, 1 used the baculovirus expression
system to produce in large quantity the extraceUuiar domain of the human insulin
receptor, and used various specific photoaffiity probes to iden- the insulin-contacting
sites. Peptide identification methods of high sensitivity were developed to detect the
labeled IR fiagment containing the putative insulin-bhding site by HPLC peptide
mapping analysis. A smali ftagment of 1-3 kDa that might contain the putative insulin-

binding or contacting site on the receptor a sabunit was âetected However, p d y due to
the lack of an insuiin photoprobe with a nigh efficiency of cross-linking, and a high
efficiency peptide mapping method, 1 failed to obtain a sufncient quantity of the fiagrnent
for amino acid sequencing.
In an alternative approach, usuig gold-labeIed insulin as a iigand, and sets of
eIectron micrographs obtained by low-dose low temperature dark field scanning
transmission electron microscopy (STEM) of the in&-IR complex, 1 successfiilly
reconstnicted the 3D quatemary structure of the whole 480-kDa human inSulln receptor
(KiR), complexed with Uisulin, at a resolution of 20 k Contiguous hi& densities within .
the 3D structure indicated a two-fold symmetry for this dimeric receptor and the presence
of subdomain structures which begin with the extracellular subdomain involved in insulin
biading and terminate with the inhracellular tyrosine kinase subdomains. These
subdomains were confinned by fitting known high-resolution crystai domain
substructures. In addition, the 3D insulin-HIR complex provides evidence for the
involvement of only a single insulin molecule in receptor activation. It also explains -
many of the known characteristics of insulia binding to its receptor and many
biochemicd fhdïngs in the studies of the insulin receptor.

INTRODUCTION
Io IiYIERCELLUIAR COMMUNICATLON
In mdticeUdar organisms communications are cruciai ta coordinate their fimction
in responduig to the e n . ~ ~ m e n t and maintahhg homeostasis. The pwth, migraîion
and differentiation of ceiis in the embryo, and theü organhtion nit0 specific tissues,
depend on signais transrmCtted h m one c d to another. In the adult, ce11 signahg
orchestrates normal cellular behavïor and responses to e x t d stimuli- The b i s for the
intercellular communication can be viewed as ligand-receptor interaction. The receptors,
either intraceiiular or ceii siirface, are an by ceiis to enabIe them to detect signal
molecules such as hormones and growth m o r s in their microenvironment, Signal
molecules such as neurotraaîmtters or growth fhctors may k limiteci to interacting with
the receptors on theù own cells or aeighbo~g ce&. Altematively, -y secrrted signal
molecules, including insuiin and many hormones, circulate in the blood and interact with
their target cells elsewhere in the organism. An extracellular ligand b * i to a specific
receptor on a target cell and converts the receptor to an active state. The activated
receptor then subse~uently stimulates intracelldar biochernical pathways leading to an
appropriate cellular response, which may involve changes in ceilular gene expression,
protein trafficking and metabolism. For example, the binding of mwilin to its receptor
activates the receptor on the ce11 surfàce, and subsequently the signal is transmitted to the
celi interior to activate various intraceîîuiar pathways to elicit the pmper biological
responses. Alterations of these signaiing pathways have been found in the pathogenesis of
many diseases such as cancer, diabetes meIlitus and disoders of immune and
cardiovascular systems.
There are genediy two caîegories of mceptors for the circulating signals based on
the lipid solubility of the ligands. Stemids, thyroid hormones as weil as rrtinoids are lipid

soluble signahg m o l d e s . They usualiy induce slowa, long~lasting responses in thek
target cens, *ch are cruciai in the regulation of ceU growth and m o n . Lipid-
soIub1e m o l d e s can cross the plasma membrane and bind to the cytosolic receptors thrt
fiinction as Iiganddependent tmmcriptïon fkctors. On the other hand, water-soIuble
polypeptide signal m01ecules. such as insuiin and growth fktors, are not ablt to cross the
plasma membrane k l y - They must bind to the specinc cell surfkce recepbrs on the
plasma membrane ofthe target tissues and signai via the transmembrane mceptors. These
glycopmfein receptors are stniftmally and biochermermcaIly very diverse, but they al1 have
an extraceiiular iigand-binding domain, mugh wbich the polypeptide si@ molecule is
recognized, and an internai effector domain. The e f f i o r may fimction as an enzyme,
such as a kinase, or a transporter, or it may interact with a cellular coastituent like the
subunit of G-protein, or an ion channel. The first response of the ligand-receptbr
- interaction may involve the production of second messengers intracelluiarly, typidy
small molecules like CAMP, cGMP, diacylglycer01 @AG) and ~ 2 % or, in the case of
the iosuün receptor, the activation of the protein kinase system. The kinase cascade
systems or charme1 Sictivities act as switches and amplifiers of one or more biochemid
pathways controlling ceU rnetabolism, gene expression, celI cycle or cell fate @aulieu,
1990; Fuller, 1991; Ullrich and Schldger, 1990).
Three major types of stmctudy and hctionally distinct receptors have been
intensively studied. They are the G-protein-coupled receptors, receptors with guanylyl
cyclase actIvity, and receptor protein-tyrosine kinases (RPTKs). Members of the 0-
protein-coupled receptor supedmdy famirr seven irausmembrane loops and comprise a
big receptor W y of appmhte ly 8oo/. of known homones and neurottansmitters
(Kobilka, 1992). Ligand-indd activation of these receptors results in the dissociation
of the a- and &- subunits of the heterotrimeric G protein, leading to the activation of
intracelldar e f fwr molecules for signai proproaiion (CIapham and Neer, 1993). The
receptors with guanylyl cyclase activity have been f o u to bind d u r e t i c peptide

hormones. The do- sigoal transduction of tfiese receptors is still not very clear
but an increased cGMP level was fomd foîlowing the activation of the receptor (Gsrbm
and Lowe, 1994). The third gmup of the receptors, whïch bind many polypeptide
homones and gmwth f=tors, such as insulin and epidermal growth f-or (EGF), have
an intrinsic, ligand-sensitive, pmtein-tyrosine kinase @TEE) activity @Jllrich and
Schlessinger, 1990; van der Gea, 1994). These StnrcWy related glycoprotein receptors
belong to a superfamily oi receptor-type PTKs (RPTKs). They feature a large
extracelIda. ligand bindïng domain, a singie transmembrane region and an intemai
tyrosine kinase domain. Ligand binding to the receptor leads to the autophosphorylation
of its kinase domain followed by the phosphorylation of the substrate prote&. Activation
of these receptors induces diverse bioch-cal pathways incIuding the generation of
second messmgers, alteraîion of etlzymatic activity, anaor expression of specinc genes.
As a principal polypeptide honnone controliing ceil metaboLism and growth, insulin
has pleiotropic effects on cells and in the whole organism. Insulin acts on cens to
stimulate glucose, protein, and tipid metabolism. In addition, it stimulates RNA and DNA
synthesis, by mo-g the expression or activity of a variety of enzymes and transport
processes. The giucoregulatory effects of insului at a whole body level are predominantly
exerted by its action on liver, fat, and muscle. Insulin stimulates glucose incorporation
into glycogen and inhibits the production of glucose by glycogenolysis and
gluconeogenesis in liver. By affecthg glycogen metabolism and gluconeogenesis in those
tissues insulin regulates glucose homeostasis. Insulin also stimulates amino acid uptake in
liver and muscle to increase protein synthesis. As weii insuiin acts as a growth factor to
mod@ or augment the action of other regdators of cellular metabolism. Altered insulin
action, such as insulin raistance, plays an important d e in the paîhogenesis of many
disorders, including diabetes meliitus, obesity, hypertension and athemsclerosis.
In the years since the discovery of insulin by Banting and Best in 1921, much work
has been done in an attempt to understand the molecuiar mechanism of insului action-

(Goldnne, 1987; Kahn, 1 W 9 White and Kahn, 1994). The identification of the lnsulin
reuqtor (Yip et ai., 1978, 1980; Yip and Moule, 1983; Pilch and Czech 1979), the
eluciàation of its Pnmary structure (Ebina et al., 1985; Ullrich et al., 1985) and its
characterization as a tyrosine kinase (Kaniga et al., 1982) served as the basis to cl- the
m o l d a r mechanism of insulin aCnacnon. This was followed by the identifidon of
do- transducers (White et al., 1985; Sun et al, 1991). The binding of insuiin to
the extraceliular domain of the receptor d t s in conformational changes leading to the
activation of a tyrosine kinase in the CytOpIamnic domain of the receptor- Activation of
the kinase is mediated by tyrosine phosphorylation of the receptor itseif. The Ievel of
tyrosine kinase activity refiects the senim concentration of insulin. The activated receptor
in tum phosphorylates a substrate calleci insuiin rrccptor substratbl (jRS-1). The
multiple phosphotyrosine residues in IRS-1 serve as docking sites for the signal
transduction proteins in the signaling cascade (White, 1994, Sun et al., 1995). Many
signal transduction pmteins, such as PI3'-kinase, SH-PTP-2, GRB-2, Syp and Nck,
containing a specifïc recognition domain, termed Sn: homology 2 (Sm) domain, can
specXcally recognize and bind to the phosphorylated tyrosines of IRS-1. The interactions
between activated receptors and SH2-containing proteins ampiify the signal produced by
the binding of insulin to its receptor and link insulin signaling to other kinase cascades
involving serindtbteonine kinases and other signal transduction pathwys, such as the ras
pathway. Wltimately, this process leads to stimuiation of glycogen, lipid and protein
synthesis, as well as translOC8fion of insulin-sensitive glucose transporters to the srirf8ce
of muscle and fât celis (Hiring, 1991; White and Kahn, 1994). Mdfûnction of this
signaiing pathwayy fi0111 the nrst step of ligand binding, through receptor
autophosphorylation, to any of the postreceptor signal transducing steps, wiîi lead ta the
manifestation of metabolic disorders in non-insulin dependent diabetes mellitus.

Ih this section, 1 wi i i reviewthe cumnt lmowiedge of the mnilin receptor fiimily in
terms of insulin-activated signai transduction, focusing on the sbirturr o f the xeceptor
and the ligand-receptor interaction.
2. THE INSULIN FAMILY OF PEPTIDES
Iasulin ïs a member of a family of hormones, growth fktors and n e u r ~ ~ ~ d e s ,
found in both vertehates and invertebrates. In addition to inmiin, the f d y includes
-like growth tirtor I (IGFQ and ïnsdhIike- growth f8ctor II QGF-n) as weiî as
the more distantly relateci peptides, the r&xï.ns, the bombysins and the molioscan
insulin-like peptides (MIPs). All members of the Eimily sime a high degree of secpence
homology and probabiy, a cornmon "insuiin fold" in the s tn ichnw. Apart fiom the IGFs,
dl comprise A and B chahs, joined by disuEde bonds, foiiowing the cleavage of the
conmcting (C) peptide in the processing of their precurso~~. Since the C -des in the
precwsors of IGFs are retained, the IGFs are analogous to proinsuiin, with the C peptide
luiking the C-temhd of the B chah to the A chah N-temiinus @ig. 1-1).
These hormones, though sharing very simiIar primary secpence and stmctwe, act
@te differentiy, wïth some overlaps, in their physiologic effécts. While insulin is
primariiy involved in regulating cerbohyàrate, fa and protein metabolian, the IGFs act
mainly as regdators of growth, development and Merentiation. IGF-1 and IGF-II, but
not insuh, circulate as complexes with specific binding proteins. The action of each
hormone is mediated by its distinct, specifïc hi&-afbiity ceii-surfâce receptor with
intrinsic tyrosine kinase activity (Czech 1982; Humbel, 1990).
Insulin: is produced exclusively by the fl cells of pancreas and has a remadcable -
array of biological effccts. Although liver, muscle and fàt tissue are considerrd as its
pNnary physiologic targets, insuiin exerts some reguiatory effectcl on virhially d ce11

types. These effects include the mobiiizatïon of transport systems for hexoses, amino
acids, ions, the acute modulation of enzyme systems sach as the homone-sensitive lipase
in adipocytes, and the modulation of the transcription of specinc genes, such as those
encoduig phosphoenolpyruvate carboxykinase. The time ofonset of these e f f i can Vary
fiom rapid (seconds to minutes) for alterations of enzyme activities to slow @ours) for
the synthesis of protein, lipid, and nucleic acid, and cell pm1Iferation. Iasulin acts through
its specinc insulin receptors expressed by almost all cell types, nomally at a level of 102
to 105 receptor rnoIeniles p a cell The amùio acid sequence of the ùwlùl receptor has
been deduced h m the nucleotide sequence of its cDNA (Ebina et al., 1985; Ullrich et ai,
1985).
One of the structural characteristics of insulin is that it is composed of two
polypeptide chains linked by two disulfide bridges. The shorter chain, caiied the A chain
because of its relatively acidic nature, has 21 amino acids residues and an internal
disulnde bridge. The less acidic and longer B chah has 29-31 amho acids, depending on
the species. There are 30 amino acids in the B chah of human insulin. This dual-chain
structure and the positions of the three disuEde bridges are invariant in al1 the dIfferent
species of insulin that have been isolated and characterized (Blundell and Wood, 1975).
Within the "insulin foid" hnework, the amino acid sequences of the A and B chains are
highly conserved as well. For example, human insulin differs h m porcine insuh by
only one, and fcom bovine by three amino acids.
The genes encoding uisulin have been sequenced and analyzed fiom several species,
including human. The immediate translation product of the gene is a single polypeptide,
preprouisullli. It contains a signal peptide at the N-terminus, whkh facilitates transit of
the precursor into the endoplasmic reticulum. nie signal peptide is cleaved away during
the process. The resuitant molecule, pminsnlin, consisting of Iùiked B, C, and A chains,

lnsulin Proinsulin
Fig. 1-1. Pmlictcd terthry striictiircr of the inrulin f d y af pcptidcr. AU peptides are presented to contain folding simihm to that of insaün as formai by its A and B citain The beavy line and the light line q m s a ~ mpactivcïy the A chah and the B cbain in insrilin, Double lines repriesent the C chain joining the A and B ch- in proinsulin, or the D extaision ofthe Achain found in the IGFs.

is mer plocessod into the mature insulin, a disulnde-Iinked polypeptide with a
moIecuIar weight of about 6 kDa
Since the amino acid quence of insulul was detennined by Sanger and CO-
workers more than 40 years ago @mwn et aL, 1955), om knowledge of insuün bas
greatly increased, In 1969, determination of the X-ray crystai structure of 2&c porcine
insuiin provideci the first direct expdnental evidence for the insulin fold (Adams et al.,
1969). In this stnicture, bulin protomers fonn a hexamer wmposed of t k e identicai
dimers assembled around a the-foId axis, In the dimer the two monomers have their A
and B chains foldeci to fonn compact globular structures with a number of cornmon
features Fig. 1-2a). 'Ihese include @ig. 1-2B): 1. an A chah with 2 helices (A2-7 and
A13-19), joined by an extended loop (A8-12); 2. a B chah with an extended N-terminus,
foliowed by a helix (B9-19), a sharp tum @2û-23) and an extended C-tennural section; 3.
three disulfide bridges (A6-Al l, A747, A2û-B 19); and 4. a hydrophobie CO=.
The crystal, and the later NMR structures, as well as the studies of insuün
sequences, mutagenesis and chernical- modifications have contributed ta our
understanding of the molecular mechauisms of receptor binding. Insulin binding to its
receptor exhiiits a curvÏiinear Scatcharci plot which indicates the presence ofat least two
classes ofbindiag sites or negative fooperativity between sites. It suggests that insului, as
a non-symmeec molecule, may contact its receptor at diffkrent surfàce sites. By
analyzing the interaction of insulin analogues with the insulin receptot, Schaffier pposed
a mode1 for insulin binding to the iiisulin receptor @ig. 1-2 C) (ScMer, 1994). He
hypothesized that insulin has two binding domains to contact its receptor. Binding site
one is the classical bimding ssit, shdied by most investigators and comprishg a number
of residues (kcludirig B24, B25, Al and Ml) in the dimer-fomiing surfàce of the insulin
molea.de. The second binding site wnsists of B 17 and AU, located in the hexamer-

Fig. .l-2. The primaq structure (A), the 3D crystai structure (B) and the 3D space-fig mode1 (C) of human insuih The 3D modd (C) shows the putative receptor contacthg sites PINDING S m 1 and 2). These two biuding sites located on m o separate SmIaces ofthe insuiin molecuie may be hvolved in the the high afîinïty binàing ofinsrilui to the IR.

forming d a c e on the opposite srde o f the molecule %rn the cl-& bindhg site.
Substitutions of these residues wouid dramaîicaily decrease insolln IifFinity to its receptor.
Thus, a hi&-afnnity bindhg of bulin to its receptor may reqaire the h d i n receptor to
contact a different bmding region on the insulin moIecdee
2.2. Insuiin-Like Growth Factors
U . e 0 t h members of insalin family* IGF-I and IGF-II are single-chain
polypeptides of about 7.5 kDa but share similar tertiary süucture and amho acid
homoIogies with inSnlin (LeRoith et al., 1992). Genes encodïng the IGFs have been
cloned and analyzed h m several species (Brisseaden et al., 1984; Humbel, 1990). The
mature fomi of IGF is a singIe-chain polypeptide of about 70 amino acid midues dMded
into contiguous B. C, A and D domains. Its structure is analogous to that of proinsuiin as
shown by computer-assisteci molecular modeling and NMR analysïs (Fig. 1-1). The A
and B domain exhibit about 42% amino acid identity to the correspondhg A and B chahs
of insulin. The C domain is analogous to the comecting (C) peptide of proinsulininsulin A
short D domain, which is not found in proinsulin, extends at the C-teminus.
IGF-1 and IGF-II, though sharing many stmcturai propertïes with insulin, differ
significantiy with respect to their sites of synthesis and biological effîts. The origin of
the circulating IGF-1 is the liver, where its expression is unda the control of growth
hormone. In the blood stream, the IGFs circulate at 1000 times higher concentration than
insulin but, unWre innilni, most if not aii of the cïÏculating IGFs are complexed with
specific binding proteins which are believed to modulate the action of the IGFs and to
prevent hypoglycemia by lowering their effective fke concentration in the circulation. Zn
addition, IGF-1 is also expressed at lower concentration in most tissues, both prenatally
and postnataüy, and thus fiinctions as an important paracrine or autocrine growth factor.
It is clear that IGF-1, by interacting with its own high-dlimity receptor, mediates most of
the effect of growth hormone on longitudinal growth. The IGFs also show insulin-like

metabolic effits hz vitro and bz vivo9 such as glucose transport. m adipose and muscle
tissue, but o d y at relatively high concentrations (Froesch et al, 1985). Like insiilin, IGF-
1 also plays a role in embryogenesis. It is expressed even before the development ofthe
embryonic Lnrer. As in the insului signahg system, the effits ofIGF-1 are mediated by a
specinc hïgh-affbity IGF-1 receptor which is a tyrosine kinase highly homologous to the
insulin receptor.
Though IGF-Iï rnimics most ofIGF-1 effects in vitro, its biological roles are stilI not
clear. It has been suggestd that IGF-II may be involveci in the fetal development since its
expression dramaticaliy decreases just before birth in both amount and the range of tissue
distniution. In contrast to IGF-& IGF-II receptor has no structural similarity to either the
insului or IGF-1 receptor. This receptor is in fact the same molecule as the cation-
dependent mannose-6-phosphate (Man-6-P) receptor, a protein that targets the Mm-6-P-
containhg proteins to lysosome (Massague et al., 1982), and its role in the action of IGF-
II remaius unlcnowm It has been proposed that most of IGF-II effects are mediated
through IGF-1 or insulin receptors (Czech, 1989).
2.3. Other members of the Insuiin famiïy
Several other polypeptides with higiiry homologous sequences to insulin have been
discovered. Some have been found in the vertebrates and 0th- have been identifiai in
proto-chordates, insects .md molluscs. They are bombyxins, relaxins and MIPs. The
sequence data suggest that they share similar three-dimensionai structures and
conformation. UnWEe insulin and the IGFs, these distantly related members show more
divergence. Interestingly, most residues in the extended receptor-binding region of insutin
are weU consmed in these family members, which suggests possible heterologous
ligand-receptor interactions. Relaxin has been identif?.ed in several species, including
hianan. In human, relaxùi has been identifieci rnaialy as a paracrine/autocrine hormone
regulating activities in the uro-reproductive systern (Schwabe et al., 1994). The

interaction of relaxin with its receptor at the molecdar level is still poorly mdersfood,
and the receptor itselfhas not been welf charackrkd.
3. MOLECULAR AND CELL BIOLOGY OF THE INSULIN AND IGF-1
RECEPTORS
As the proteins primarily responsi'ble for transducing the signds for msulin and
IGF-I, the insulin and IGF-1 receptors, Ure their Iigauds, are the products of homologous
genes (Abbott et al., 1992). The characterization ofcDNAs encodlag the uisalin and IGF-
1 receptors revealed a high degree of similarity in the primary nucleotide and amino acid
sequences, their overall structures, and enzymatic activities. Both receptors are
glycosylated hetemtetmmers, composed of two a subunits of -130 kDa and two f!
subunits of -90 kDa Ilnked by disuEde bonds, The a subunit, which is entirely
extracellular, contains the ligand-binding domain, while the fl subunit consists of an
extracellular domain, a trammembrane domain, and a tyrosine kinase (TK) domai..
Among the similarities between the insulin receptor (IR) and IGF-1 receptor (IGF-1 R),
the highest amko acid homology (84%) is fond in theù TIC domains, whiie the overall
amino acid homology between their P subunits is only 4460%. Both IR and IGF-1 R
belong to a membrane receptor superfamily calied the receptor tyrosine kinase @TI().
Members of this supdarnily possess inkirsic tyrosine kinase activity and are stnicturaIly
3.1. The Receptor Tyrosine Kinase Superfnmily
Tvrosine Kinase and Rece~tor Tvrosine Kinase
Insulin and IGF-1 receptors were among the nrSt RTKs identifkd in the early 1980s
due to their Ligand-stimulated autophosphorylation (Kasuga et al, 1982). More than LOO
proteins with the consensus sequences characteristic of TK have been cloned and
characterized h m a variety of eukaryotic species (Hunter and Cooper, 1985). Acting as

an onhff switch In signal transduction, the protein ph~sphory l~od âephosphorylation
cycle plays a key mie in wntrolling various ceUularprocesses- Protein kinases d y z e
the transfer of a phosphate group fhm a donor, usually the y phosphate of ATP, to an
acceptor amho acid in a substrate pmtek Serine/tsreOnine kinases (SKs) phosphoryIate
serine or threonùie residues of substrate proteins whereas tyrosine kinases (TE=)
specincally phosphorylate protein substrates on th& tyrosine residues. Hmter et ai have
demonstrated the existence also of protein-histidine kinases that phosphorylate histidine
residues of substrate protek (Hmter, 1991). Some protein kinases are able to
phosphorylate al l three hydroxy amino acid residues.
Members ofthe protein kinase f2ïmi.I~ share highly consewed amino acid sequences
in their catalytic domain. Based on their cellular location they can be divided into two
classes: cytoplasmic and membrane bound. Cytoplasrnic enzymes seme as molecules of
signal transduction inside the celis. Transmembrane kinases, such as the msulùl receptor
family, transfer signals across the ceïl membrane aud activate cytosolic kinases. It is
interesting that TKs are found only in multi-celluiar organisms, whereas SRs are found in
both single cell and multi-cellular organisms, This suggests that the tyrosine specincity
may have been an evolutionary development related to the acquisition of multi-cellularity
and cell-cell communication.
The overail architecture of iill RTKs is very similac a large glycosylated
extraceiiular domain that binds the polypeptide ligand; a single hydrophobie
transmembrane domain; and an intracellular domai. containing a highly conserved TK
catalytic region. Interestingiy, some cytoplasmic TKs are not transmembrane but anchor
at the Innet d a c e of the plasma membrane and are coupleci to transmembrane
glycopmtein receptors which do not have cytoplasmic kinase domain. This coupling
allows them function like a transmembrane R% T celï antigen receptor-fln, p w t h
hormone receptor-Jak2, and interleukùijaks are examples.

B a d on the wmprehdve SMaIysis of the prlmary -ces, RTKs are
subdivided into many subfhnilies acwrding to and shared stnictural f w
@ a d et al., 1993; van der Geer et al.,. 1994). More than ten subfkdies have been
classinecl over the last few years. The best c- s u b f d y mernbers iuc1~ :
- epidemial growth htor (-F) receptor, phlet-derived growth factor (PDGF) receptor,
insului and IGF-1 receptors, nenre growh f-r WGF) receptor, fibroblast growth nrtOr
(FGF') receptor, and hepaîocyte growth nwnOr (IIGF) rrceptor (c-met). Most of the RTK
f d y membexs are sïngle-chain polypeptide e r s , except for the msuIHi reœptor
subfamily. Members of the insuiin -or subfamily incide de -r, IGF-1
receptor and insulin receptor related receptor @SR) with th& unique a& tetnuneric
structure. The stnictural similarities within a subfàmiIiy suggest that receptors in a
s u b f d y s b similar mechanisms of molecular regulation and signal transduction
pathways, as in the case of insului and IGF-1 receptors.
The cataiytic domains ofprotein kinases are 250-300 amino acid resïdues in length
mostiy located neer the carboxyl terminus or as a subunit of multi-subunit enzymes
(Hanks et al., 1988; 1991). The tbreedimensional stnictine of the TIC domain of the
insulin receptor exhibits a highly conserved bi-lobular stnicture (Hubbd et d., 1999;
Hubbard, 1997). In the large lobe of the TK domain, an extendeci sequence containhg
Tyr1 158, Tyr1 162 and Tyr1 163 fomis a flexible bop, d e d the activation loop (A-
loop). During activation, the A-loop undergoes a major conformationai change upon
autophosphorylation of the three tyrosine residues within the Iwp, resulting in
uarestricted access of ATP and protein substrates to the kinase active sites. Although both
SKs and TKs share a basic struchm, TKs bave the extended A-loop sequeace in the
larger lobe. It may explain the specificity for tyrosine since serine and threonine
hydroxyIs may be too short to reach the phospho-tramfer site within the d y t i c cmter
(Hubbard et al., 1994).

I I a Subunit p Subunit
Fig. 1-3. Schemrtic nprcscatatioa Mthe inrmlin reccptor gene (A) and the primary (8) and secondary (C) structure of bIR Receptor dornains encoded by specinG exons are indicated (A and B), Labels: S=signal peptide, Ll,L2=Ll and L2 dbmahs, CR--teine-nch domain, CD/FnO=Çomiecting domain 0rfibromsi.n ïü repeaq Fnl,Fn2= fibroneah Eï repeats, ID=ïnsertdom8ïn, TM=transmembnme domain, JM+wûamedrane domain, TIC tyrosine b e domain, TL=C-temnîd tail, Disuifide bonds mmecting a-a and a+ subunits are iuriiated by- and possiile phosphorylation sites are mdicated
- b-(C)-

Gene strucîme and EsDression
The inailin receptor @I) is theproduct of a single gene IOcafed on the short ami of
chromosome 19 (I9p7) nearthe gene ofthe LDL receptor (Yang-Feng et al., 1985). The
structure of this gene has been eIucïdated by both restriction mapping @&der-Wieland et
al., 1989) and genomic cloning (Seino and Bell, 1989; Seino et al., 1990). The human IR
gene, composed of more than 150 Kb of DNA, has 22 exons and 21 htrons. Exon I
through 11 are spread over 90 Kb and code for the a subunit, Exons 12 through 22 span
30 Kb and code for the p subunit (Eig. 1-3). The promoter ofthe IR gene is ofthe typicd
-ch %ousekeeping" type with a few bmding sites for edancer protem (McKeon et
al., 1991)-
The receptor is synthesized in the mu& endoplasmic reticulum (RER) as a single
polypeptide cbain proreceptor of either 1339 qr 1351 amino acids, depending on the
differential splicing of exon 11. The pre-receptor has a 27 residue N-terminal signal
peptide that assists in the intraceUuiar translocation of the receptor precutsor. The signal
peptide is subsequently cleaved off. The proreceptor is transportecl through the Golgi
apparatus, where it rapidy undergoes extensive N- and O-linked glycosylation. Two
monomeric proreceptors are then linked by the formation of disuifide bonds into a
dimeric structure. Proteolytic removal of a tetrabasic amino acid sequence (Arg732-Lys-
Arg-Arg735, numberhg based on Ebina et al., 1985) h m each proreceptor monomer
gives nse to the a and f3 subunit Firrther glycosylation takes place before the mature
insulin receptor is inserted- into the plasma membrane (Olson et al., 1988). The ftnaI
active mature receptor is a fiilly giycosylated tetrameric transmembrane protein,
composed of two a subunits and two P subunits (Fig. 1-4). Each a subunit is comprised
of 719 or 731 amino acids. There are 620 arnino acid rtsidues in each $ subunit hcluding
an extcacellular portion of 194 residues, a trammembrane region of 23 amho acids, and a
cytoplasmic domain of 403 residues.

- -
Fig. 14. Human insuhm mceptor amho acid sequence<smmEbUu o t i ~ . 1985) Residues sl to s27 arc the signai seqence. Residucs 1 to 731 ccmstitute the a subunit, and maiducs 735 to 1355 wnstïtutc the B subunit, The polybasic residucm 732 to 735 arc tbe crupatio olenrage site to yield the a and the subunita UnderIined rcsiducs arc d d by Exaa 11-

The heîeroteîramer structure of the insufin receptor was initïaiiy elucidated in the
late 70s by cross-Ii'nkmg radioactive insnlin to either whole ceb o r plasma membranes
Orip et ai-, 1978; Pilch et aL, 1979). Later, pudication of the receptor led to cioning of its
cDNA @bina et al., 1985; UIIrich et al., 1985). While the prhnary sequence of the IR has
been known for some the (Fig- 1-4), knowIedge of the secondary and higher order
structures of the receptor are llmlted, Based on cacrent knowIedge of insulin receptor
hction, and the fûnction of RTK proteins, it seems reasonable to propose that sp&c
bctional properties of the IR reside m its separafe domains. A major advance in
understanding the overall receptor structure was the mode1 proposecl by Bajaj and co-
workers @ajaj et al., 1987)- They proposed that the N-texminal haIf of the IR wntained
two large homologous domains (21 and L2) sepacafed by the cysteine-rkh domain (CR) b
which itself was compnsed of three repeating d t s each containing eight cysteine
residues. A second major development was the observation that the C-terminal portion of
the extracellular region of IR contanis three fibmnecth type III repeats (here designateci
as FnO, Fnl and Fn2) linked to the L2 domain (Marino-Busije et aL, 1998; Malhem and
Booker, 1998; Ward 1999). The fkst repeat, FnO, or identifid as the comecting domain
(CD) by some authors, located in the a subunit wnsists of 123 amino acid residues. The
second repeat (Fnl) is a structure with contriiutions by both the a and the B subunit,
connected with a s o d e d imxî domain (ID) that includes both the a+ cleavage site
and the alternative-spliced exon 11 (O'Bryan et al., 1991). The third repeat (Fn2) is part
of the p subunit. The extraceliular region aiso provides the two disulfide-bonds that
covalently lïnk the a p monomers to form the constitutive IR dimer. One of them is in the
FnO region and the other is located in the a insert domain. The extracellular portion the
IR is foliowed by a short, single transmembrane domain 0 connected to the
intraceiiuiar portion of the B subunit that contains the TR domain. The crystaUographic

structure of the Tg fkgment ofthe fl subu& was detemiùied several years ago (Hiabbard
et al., 1994), but the t h e dimensional of the holoreceptor remaiaf miknown.
The generai structural domab layout of the IR agrees with the suggestion thaî
insulin binds to the extracelluî~ dom- 1-g to the transmembrane activation of the
intracelMar TK of the f3 submk
The a Sabunit
The a subunit of the IR is entireIy exfraceIIuIar wntahhg either 719 or 731 amho
acids depending on whether or not exon 11 is expressed (Seino and Bell, 1989). A
characteristic feature of the a subunit is its cysteine-rkh region which contains twenty
four of the thirty seven cysteine residues in the a subunit The fimction of the cysteine-
nch domain is wt M y understood, but it has been suggested that it is involved in
binding to insulin (Yip et al, 1988, Rafiisloff et d., 1989; GustafSon and Rutter, 1990).
Homologous cysteine-rich regions are also seen in the low density lipoprotein (DL)
receptor, IGF-L receptor and epideanal p w t h factor (EGF) receptor (Goldstein et al.,
1985; Ullrich et al., L984,1986). These regions in IGF-1 and LDL receptors h a . been
shown to bind to their ligands. However, in the case of the insulin receptor, it is d l
controversial as to whether the cysteine-nch domain is directly involved in insulin
binding.
There are 37 cysteine residues in each a subunit monomer- Most of them appear
to fonn intrachain dimifide bonds within the a subunïts, mainly in the Qs-nch region-
On the other hand, some cysteine residues fomi disulnde bonds joining a-a or a-P
subunits, which contribute to the important conformation of the fimctional receptor (Finn
et al., 1990). There are two classes of disulfide bonds in the IR. CIass 1 diSulfide bonds
link a+ subunits, while class 2 diSulfide bonds join the a-$ subunits. The latter is more
resistant to reduction than those of class 1. The mature iasulin receptor is a disalfide

llnked hetemtetramer, including one disulnde on each a-$ wmection and more than one
on the a-a cross-Ilnking8
Based on the structural analysis of the IR sribjected to mild tryptic pmteoLysis, the
C-temiinal25 kDa hgment of the a subunit was found to be involved in a-P didiide
bonding (Shoelson et ai., 1988; Xu et al., 1990)- Cys647 located in this region has been
reportai to be vital to the a-p bond formation (Cheatham and Kahn, 1992). Recently,
Sparmw et ai confirmed that Cys647 in the Fnl of the a subunit forms a diSulfide bond
with Cys872 in the Fn2 of the P subunit (Sparrow et al., 1997)-
Until recently the diSulfide bonds connectiag the two a a monomers together were
unknown. Schaffer and his CO-worker first noticed that the Cys524 upstream of the Fnl
domai. contributes an a-a interchah diSulfide bond (SchafEer and Ljunqvist, 1992).
Other investigators (Macaulay et al., 1994; Bilan and Yip, 1994) suggested that additional
disulfide bonds exist between the a* dimer, which is probably locatted in the cysteine-
rich domains, Surprisingly, the second dîsulnde bond connecting the a-a dimer has been
recently locaiized to one of the cysteine triplet, Cys682,683, and 685 at the end of the C-
terminai region of the a subunit (Spmw et al, 1997).
Bajaj et al @ajaj et al, 1987) proposed, in addition to the cys-rich region, two
similar but independently folded subdomains, LI (encoded by exonl-2, residues 1-1 56)
and L2 (encoded by exon 4-6, residues 3 10-470). each being mainiy B-pleated sheet in
structure. Between LL and L2 is the cysteine-rich (CR) domain Cresidues 157 to 309,
encoded by exon 3-4). Based on sequence alignment and secondary structure prediction,
Bajaj et al. proposed that the LI and L2 domains juxfaposed and extended away h m the
ce11 membrane, with the CR domain ~11der1ying and supporting them (Bajaj et al.. 1987).
The recently detennined crystal structure of a fiagrnent containhg the f b t three domains
(LI-CR-L2) of IGF-IR, a receptor closely related to IR (Garrett et al, 1998), shows that
the layout of the thcee domains was quite diffkent b m that proposeci by Bajaj et al. The
remaining regions of the a subunit (residues 471-731, encoded by exon 8 to 11) consist

of a l3ronectï.n III repeat @no, residues 471-593, also c d e d comectiag domain (CD)),
part of the second fIbronectln ï i I repeat (Fnl, residnes 594461) and a 71 residus insert
domain (ID) (O'Bryan et al., 1991, Schaefer et al., 1992). LI, L2 and CR domains have
been suggested to be involved in insulin bmding @Tapes et al., 1986; Yip,
l988,1992,l993; De Meyts, 1994) and in both a-a and a-p disulnde bond formation
(Cheatham and Kahn, 1992; Scbaffer and Ljunqist; 1992, Bilan and Yip, 1994; Spanow
et al, 1997). Little is known about the function of the insert and connecting domains.
The quatemaq structure of IR has not been determuid- Schaefer et al. (Schaefer et
al., 1992), using electron microscopie image techniqye, reported that the extraceUular
domains of the IR (the entire a subunits plus the extracellular domains of the B subunits)
in solution had a "Y" shape with giobular features at the end of each ann of the T",
Similar studies also showed the s o l u b ~ e d placenta holoreceptor as a "T" shaped
structure (Christiansen et ai., 1991). These studies suggest that the receptor has two
extracellular arms, each of which may represent a separate a subunit- The authors also
suggested that each of the subunits wodd contribute one independent insulin-bindiag
site. There has been no report of the successfiil crystallization of the insulin receptor- Its
large size (b& 450 kDa) and its glycoprotein nature may accokt for the lack of success in
its crystaUization. It is ais0 too large for NMR analysis,
Early studies using the technique of photoaffinity labeling (Yip et al., 1980) and
chemicai cross-linkiag vilch and Czech, 1980) have clearly shown that insalin binds to
the a subunit of IR. Although the entire a subunit could be involved in contacting
insulin, recent studies on -receptor interaction have graduaüy focused on the Ll-
CR-L2 region (White and Kahn, 1994). Since no structural Uiformation on the IR a
subunit or its hgments is available, the structure of the LI-CR42 domain of IR now can
only be inférred h m the x-ray structure of its counterpart of IGF-IR (Garrett et al, 1998).
The overall stmchne of the Ll-CR-L2 domains of IGF-IR was shown as a notch shape
structure. Each L domain is a singie-stranded right-handed P-hek The cysteine-rich

region is composed of eight disuEde-bonded modules, seven of which fonn an elongated
rod shape domain connecfing the LI and L2 dom- The three domains surround a cmter
space of sufncient size to accommodate a ligand moIecdee AIthough this hgment is
incapable ofbhding to KGF-& this structure does provide clear infiormation that the L1-
CR-L2 region is a putative region for ligand bindïng*
Two isoforms of the insulin receptor, différïng in length by 12 amîno &âs near
the C terminus of the a subunit, are daiveci h m the alternative splicmg of exon 11. This
results in a srnalier A isofonn of 1339 amino acids and a larger B isoform of 1351 amho
acids (UUrich et al., 1985; Ebina et al., 1985). Both isoforms (- and + exon 11) are
expraseci in most tissues in various proportions. The fimctionaL difEerences between
the two isofomis are not clear. 1t semm that isofomi A has a relatively higher affinity for
both insulùi and IGF-1 and is more rapidly uitemaiized and d o m regulated (Mosthaf et
al., 199 1; Voge et al., 199 1; Yamaguchi et al, 1991). The mechanian of this change in
mty by a structural variation distant h m the presumed binding site is uncertak
However, mutant insulin receptors with mutations at the cleavage site have been reported
with markedly reduced afftnity to insului (Wifiams et al, 1990; Sasaoka et al., 1993).
This suggests that the C-tenninal of the a subimit may contribute to a confionnation for
high afbity ligand bhding. hterestingiy, Fnisca et al recently reported that insulin
receptor isoform A binds to IGF IL with an &*ty close to that of insulin. It actualiy
serves as a high-aff?nity IGF II receptor affêcting growth of fetal and cancer cells (Frasca
et al., 1999; Sciacca et al., 1999). This finding also suggested that the small variation in
the C-terminal of the a subunit plays an important d e in the conformation for high
e t y ligand binding.
The B Snbnnit
The subunit ofthe Ïnsulin receptor contains an extracellular domain of 193 amho
acids, a single transmembrane domain of 23 amino *ds, and an inkacellular portion of

402 amho acid reslCdues- The htraceflular portion of the subunit can be fbrther
subdivided into three fima*onal domallis: a juxtamembrane domain, a tyrosine kinase
domain and a C-terminal domain. The tyrosine kinase domain bas been recently
c r y s and its stnicture detennined (Hubbard et al., 1994). The main hction of the
$ subunit is to react to the signal generated by extrace1luiar insuih binding.
Relatively littie is known about the fimcti*on of the extracellular domain of the $
subunit It con- a half (Fnl) and a fidi Pn2) fihnectin domain and four cysteine
residues. Cys872 in the Fn2 region fonns the interchain bonding *en the a and
subunits (Cheatham and Kahn, 1992, S p w et ai., 1997). There are also sites for both
N-linked and O-linked glycosylation (.ge et al., 1990). A monoclonal anh'body to this
region allows normal insulin binding but inhibits receptor autophosphorylation and
biologid signaling (Gherzi et al., 1987), suggesting that this domain IikeIy plays a role
in receptor signai transduction.
Besides the putative signai transduction fûnction., the extracellular domain of the
subunit may play a criticai role in the confonnation ofthe high aff?nity binding site of the
a subunits. Schaefer et al. (Schaefer et aI., 1990) demonstilated that truncated insulin
receptors with aU or part of the extracellular domain of the f3 subunit deleted dramaticaily
lost their insulin-binding af%ity. The minimai truncated receptor maintahhg fûli
binding Rffinity is the wmplete extraceiiuizu domain, inciuding both a subunits and the
entire extracellular part of the f3 subunits. They suggested that, even though the subunits
do not dinctly participate in insulin binding, their existence is critical in forming the nght
conformation for the high dûnity binding site in the a submïts.
The transmembrane domain of the subunit is a single a-helicai segment of 23
residues which links the extraceîlular and intracelluiax domains of the receptor. This
physical connection between the extraceiiuiar and intraceiiular region of the subunit is
impLicitly cruciai to signal transductio~ However, mutations or replacement of this
region with the trammembrane region of other receptor fhmiiy members did not afféct the

hc t ion of the receptor oramda et aL, 1992, FrattaIi et aI., 1991)- Thus, it appears that
the structural requirements of this region for sÏgnaL transduction are not restrich
The juxfamembrane domain is a short fbgment of48 -dues at the begùining of
the intraceilUlm portion ofthe B subunit Two essential fimctions have been found for the
juxfamembrane domain: receptor ïntennalization and signa1 transmission Interestingly,
both hct ions relate to the specinc NPXY motif in this region (Feener et ai., 1993). They
are NPEY at position 969-972 and GPLY at residues 962-965.
The NPXY motif bas been indicated to interact with adapttor proteins Ï n coaîed pits
to facilitate receptor endocytosis @ r m and Greene- 1991). Mutagenesis of this domai.
has shown that the ligand-induced IR endocytosis was abolished. More recently, the
NPXY-motif has been identified as the interacting site with ùisulin receptor substrates,
such as IRS-LIIRS-2, and Shc (Keegan et ai., 1994; O'Neill et ai., 1994). Mutations at or
near Tyr 972 inhi'bit insulin stimulated tyrosine phosphorylation of IRS-l/IRS-2 and Shc
(White et al., 1988, White & Khan 1994). The phosphorylation of IRS relies on the
engagement of the phosphorylated NPXY-motif in the insulin receptor (Sun et al., 1995;
Keegan et al., 1994).
The tyrosine kinase domain is the most conserveci region among the family of
RTK (White and Kahn. 1986; Ullrich and ScMessinger, 1990). It contains an ATP-
binding region and a so-called regulatory or catalytic region. The ATP-binding region
composed of an invananant sequence GXGXXGX located at residues 1003-1008 of the
insulin receptor, followed by a lysine residue located distally 10 to 20 residues toward the
C-tennind. The Lys1030 in this region provides an AT.-binding site through a sait
bridge. Point mutation of Lys1030 disrupts both ATP binding and kinase activïv
(Maegawa et ai., 1988), and 1eads to severe insulin resistance (J'ayIor et al. 1992).
There are 13 tyrosine residues in the cytoplasmic portion of each P subunit,
Phosphopeptide mapping demonstrated that 8 of the tyrosine residues are criticai for
exogenous tyrosine kinase activity and insulin receptor biological activities (White et al.,
24

1988; Kohanski et aI, 1993; Feener et al., 1993)- These tyrosine resïdues are IocaCed in
three clusters in the primary stnicture: Tyr96St- 972, and 984 m the juxfamembrane
region, Tyr1 158,1162, and 1163 in the kinase d y t i c domain, and Tyr1328 and 1334 in
the C-terminai taiI. This numbering system includes exon 1 1. The toree tyrosine residries,
Tyr1 158, Tyr1 162, and Tyrl163, lying in the b a s e activation loop (A-loop, residues
1149-1170) are the most important site of autophosphorylatim of the IR
Autophosphorylation of aii three tyrosine resïdues in the YXXXYY motif of the
activation Ioop occurs ~6thin seconds after insvim stimulation and stimidates tyrosine
kinase activity by 10-20-foId (White et al. 1988)- Mutation of one, two or three tyrosine
residues in this region prognssively duces hmlïmstimulated kinase activity and resuits
in a parallel 105s of biological actïvity (Vogt et al 1991; Wdden et al. 1992). The crystal
stninime ofthe tyrosine kinase domain of the inSulln receptor fbsubunit not onIy shows
the fine structure of the kinase domain but aiso provides important u i f o d o n on the
mechanism of kinase activation (Hubbard et al, 1994, Hubbard, 1997). In the inactive
state, Tyr1162 in the A-loop cornpetes with protein substrates for binding in the active
site. Dunng activation, the A-loop uudergoes a major codonnational change upon
autophosphorylation of Tyr1 158, 1 162 and 1 163, resulting in umestricteâ access by ATP
and protein substrates to the kinase active site. The crystai stnicture of the kinase domain
also indicates that autophosphorylated Try1163 plays a role in stabilizïng the
phospborylated A-loop and suggests that Tyr1 158 may provide a dockhg site for
downstream signahg proteins.
The C-terminal region of the IR exhi'bits maximai divergence between IR and
o t k RTKs -ch et ai., 1986). Therefore, it has ben proposed that it wntri'butes to
specificity in transmembrane simialinp. It also contains a second major region of
autophosphotylation uyr1328, and Tyr1334) in the IR B subunit. However, the exact role
of phosphorylaiïons of these tyrosines in insulin action is d l not clear since kinase
activity in the C-teuninai tnincaîed IR is normai Myers et al, 1991; Takata et d., 1992).

It also contains a few serhdthreonine phosphory1ation sites stimdated by several
stimulators, incIudIng insulin, The fimctïon of serine/hnine phosphoryiation in this
region remah controver~l*aL wonezewa et al., 1991; Hotami-sligil et al., 1993).
3.3. IR, LGF-IR and IRR: Simüarity and Divergence
Three receptors, insulin receptor w), IGF-1 receptor (IGF-IR) and insului receptor-
related receqtor W., in the RTK -y share a high level of overail structural
similarity. Although th& gens have been mapped to different chromosomes
(chromosome 19, 15 and 1, respectively)), IR, IGF-IR and IRR have similar amino acid
sequences and identical exodintron o q p h t i o n @bina et aL 1985, Ullnch et al. 1986,
Shier & Watt. 1989, Abbott et al. 1992). AU three receptors are synthesized as a single
polypeptide that is processeci to yield distinct a and P chahs joined by diSuldide bonds
into a 4 3 , heterotetramtxic cornplex. Whereas the IR and IGF-IR have a tetrabasic
cleavage site, RKRR, the IRR has an RHRR at thïs site. AU three have a completely
extracellular a subunit that contains a region of approximately 150 residues which is
particdarly nch in cysteines. However, these cysteine-rich regions show a lower
sequence homology than munding regions (-45% compared to 45%). difference
has been implicated in some shidies as playhg a role in detemiining ligand specificity
(Zhang and Roth, 1991; Kjeldsen et al., 1991; LeRoith et aL, 1994). The p subunit
consists of both extracelIular and intracelldar domains connecteci by a transmembrane
domain. The htracelldar tyrosine kinase domain is highly homologous in all s u b f d y
members.
Wlde the IR and IGF-IR share a very close sequence homology, IRR is less similar.
Most notably, the P subunit of IRR is only 551 amino acids long versus 620 and 627
amino acids for that of the IR and IGF-I& respectively (Shier and Watt, 1989). This sue
dinerence is in large part accounted for by the carboxyl-tail of I R . which laclcs 48 amino

acids including the two C-teLmlLlaI tyrosines- and the seraidthreonln phosphoryIation sites
present in other two receptors, h con- to the highiy comemed ATP-binding and
tyrosine kinase region in aii tbree receptors (79-84% identicai), the sequence homology in
the carboxyl--tail region of the th- receptors is remarkably low (1944%)-
Despite their structural similaütks, the msuIin and IGF-I receptors mediate dinerent
physiological effects. The insulin receptor regdates metaboiism as weU as some
development and growth (Lee and Piick 1994). The IGF-1 receptor, on the other han4 is
noted for rnediating the pwth-pmmoting action of IGF-I, as weU as some of the effects
of IGF-II (Humbel et al., 1990). The ligand for IRR is dl unknown. 1t is interesting that
the insullli receptor, besides binding insulin with high affrnity, a h bbinds IGF-1 and II
with about 100-fold weaka aflhity (Anderson et al., 1990; LeRoith et ai., 1994).
However, the IGF-1 receptor has a very low affcnity for insuiin. There is also a smaiï
amount of naturally occurring hybrid IGF-Ilinsuiin receptors which contani af3 hdves of
both the insulin and IGF-1 receptor (Mosthaf et ai.. 1994 Siddle et al., 1994) with low
aflkity to either ligand The hction of the hybnd receptor is not clear.
- Taking advantage of the similarities and diffèrences in the domain structures and
their big difference in e t y for their ligands, especially in the case of IR and IGF-IR,
investigators have an opportunity to study the iigand binding sites and specinc signaihg
pathways. Several chimera receptors with switched domains between IR and IGF-1 have
been studzed. These studies have revealed important information in the understanding of
the interaction of ligand and receptor (Anderson et al., 1990; Zhang and Roth, 1991;
Schumacher et al., 1993).
4. INSULIN BINDING AND SIGNAC TRANSDUCTION
4.1. Affhity and Kineties of Insaïin Bincihg
Insulin binds to its receptor in a rapid and reversii1e manner. The affinity constants
vary around 109 M-1, and dissociation occm h m within minutes to several hours with

increasïng insulin concentration De Meyts n o n " twenty years g that insalin binding
to its membrane receptor, as andyzed by Scatchard plots, showed an upwardly
curvilÏnear bindlng isothexm (De Meyts et al, 1973). These were Interpreted as indicative
of negatively c o o p d v e bmding, suggesthg that the a£E~Üty of the reaction is not
~ O R R over the saturation range. They pmposed that there were two "buiding sites" on
the receptor and consequently th& the IR c m bind insulin with high anàlor low afnnitty
based on the insulin concentration. The insulin receptor binds one insulin molecule with
hi& afiïnïty @Cd-19 M-1 ) in its high affinity site at physioIogicaI concentration of the
insulin. At saturation conditions, the receptor can bind auother insulin molecule with
lower affinity (~d-108 M-1). This intqretation has triggered numerous additional
binding studies h m many laboratories and resultted in considerable controversy over the
years.
De Meyts's mode1 also provideci an explanation for the kinetics of insuiin bhding.
When the binding site is empty, the receptor can bind to insullli with high afkity. This
binding ieads to additional insulin bhding occuning with lower af3bity and d t s in a
higher dissociation rate.
Considering the fact that the insulin receptor is a fûnctional a& hetemtetramer, it
is highly possible that high affinity insulin binduig requires that uisalin be in contact with
both a subunits, although the contact region in each a subunit may be different (Yip,
1992; Lee et al., 1993; Schaffer, 1993). Binding of one i . molecule to the receptor
may make it more dï.fi5cu.k to bind the second insulin molecule. The exact ligand contact
region on the a subunit is still inwmpletely defineci. The mechanism of negative
cooperative binding will be better explained when more details of the insuiin receptor
structure are hown.
4.2. Autophosphorylation

h s u ï h bhding to the receptor a subuüit rapidly stimulates aatophosphoryIation and
enhances tyrosine kinase actiw ofthe $ subunit of the receptot (White et al, 1988). The
signai of ligand binduig in the extraceiIdar domain is trammittecl through the
hydrophobie. transmernbrane domah, possiay hy a conformationai change. The
uwccupied insuiin receptor acaially inhibits the tyrosine kinase activity of the subunit
(Hubbard et ai., 1994)-
As mentioned preMousIy, aautphosphorylation of the receptor occurs in t h e
distinct regïons: the juxbmembrane region T-72, and possibly 965 and 984; the kinase
catalytic region, Tyr1 158, Tjn 1162, and Tyr1 163; and the C-temiinal region, Tyr1328
and Tyr1334 (White et ai., 1988, Kohanski, 1993; White and Kahn, 1994)- The
mechanism of iasulul receptor autophosphorylation likely occm by mutuai
transphosphorylation of the $ subunits (Lammers et ai., 1990; Lee n ai., 1993; Taouis et
al., 1994) although some cils-phosphorylation has been reported. The recent gystai
structure of the insuiin ~eceptor PTK domain suggests that a cls-autoinhibition is found in
the inactive kinase* Insulin binding moves two kinase domains closer to enable hm-
phosphorylation in the presence of ATP by disengaging Tyr1 162 nom the catalytic loop
(Hubbd et al, 1994; Hubbard, 1997).
The intracellular juxfamembrane domain of the insulin receptor P subunit contains
at least one phosphorylation site -72) in a NPxy972 motif (Feener et al., 1993).
Point mutation at this residue has no effict on the insuiin reccptor's tyrosine kinase
activity but impairs receptor signal transmission (White a al., 1988; Keburagi et al.,
1993). This NP2@72 sequence is also îhe binding site for the insuiin subsîrate IRS4
and Shc, which transduces the signal intraceiiularly (Gusiafson et al., 1995; -Pawson,
1995).
4.3. Post-Receptor Sigul Transduction

The dowastream elements respmsi'ble for the propagati*on of extract11uiar signais
through tyrosine Ianase rnessehger systems have amacted much attention and ban
extensively revïewed (Pawson and Gis& 1992; White and K h , 1994; White 1994;
1996). In this section, 1 briefly Summaaae the cumnt lmowfedge about the insulin
signahg pathways.
The bhding of insuiin stimulates the i n W c tyrosine kinase of the IR, *ch
d t s in autophosphoryIation of the subunits on tyrosine residues and the subsequent
phosphoryIation ofiasullli receptor sribstaite pmteias. including iasulio receptor substratc
proteins @RS-1 and IRS-2). IRS-1. and IRS-2 are proteins with molecular weight larger
than 180 kDa based on theh electrophoretic mobility SDS-PAGE. They err 43% identical
in their primary amino acid SeQuences (Sun et al. 1995). Both of them contain so-called
IRS-homology domains: Mly 2 and 3 (White, 1996). The Il32 domain, sharhg 75%
homology between IRS-1 and IRS-2, has been found to be similer to the phosphotyrosine
binding (PD) domain in Shc and can bind to the phosphoryIlatcd ~ ~ - 7 2 sapence in
the insulin nxqtor (GustafSon et al. 1995; Sun et al., 1995). While M1 and IEI2 are
important for recognkhg and binding to the insuiin refeptor, ïH3 1~3y m o d ï
downstream sigaals (Gustafin et al. 1995). IRS-proteins contain more than 20 potential
tyrosine and 30 serine/threonine phosphorylation sites, including many tyrosine
containing motifs such as YMXM and YXXM recognized by various kinases. Unlike
other RTK family members, such as EGF and PDGF receptors, the insuiin receptor does
not appear to have direct association with SH2-proteinS. Instead, in the case of the inmlin
receptor the phosphorylation sites and motifs in IRS-proteins provide docking sites for
SH.2-proteins, such as PI-3K, SH-PTP2(Syp), GRB-2 and Nck (White, 1994; 1996), and
many other kinases, such as ~8~15i.n kinase II, and MAP kinase (Sun et al., 1993;
Tanasijevic a al.. 1993). These multi-protein interactions provide a meam for signai.
ampiification by eliminating tht s to i~hiorn~c constraints enwuntered by TeCCPfOrs that
directly recnllt SH2-proteins to thQr autophosphorylation sites.

hteresthgîy, in addiition to intaadiag with the IR, IRS-pro* are also engageci
and phosphoryîated by the receptors of IGF-1 and various cfasses of cytokines and growth
f e r s (White a aL, 1985; Myers et al.. 1993, White. 1994; 1996). The shrned use of
IRS-proteins by diffaent recepfOrs likely suggests important connections previousiy
unknown, or observed but unexplained, between various homones and cytokines.
Severai intracellular pathways aie involved in the insulin signaihg system
through the phosphorylated IRS-proteins @ig. 1-5). For exampIe, phosphoryIated IRS-
protein am associate with and activafe PC3K and S&PTP2- The activateci PI-3K may
1ilÙE to p70 ribosomal S6 kinase and specinc isotypes of pte in kinase C Wyers et ai.,
1994)- Activated Pb3K was also found to be mvolved in GLUTQ trans1OC8fIon
(Cheatham et al, 1994). In addition, IRS-pmtehs, mostiy IRS-2, can actiV8fe Grb-2
which regulates a Ras guanine nucleotide exchange fktor called SOS. The Grb-2-
activated SOS initiates a mitogm-activated Lmasc cascade consisting of c-Raf; MAPKK,
MAP kinase. This pathway leads to alterations in gene transcription and 0 t h cellular
activities (Nishida and Gotoh. 1993). It is not yet clear how the metabolic effm of
insulin, such as glucose metabolism, are regdated in the insuiin-signaüng pathway.
Aithough th= is evidence that suggests that PI-3 kinase (Cheatham et al.. 1994) or a
kinase cascade simüar to the MAP kinase pathway may be involved in giucose
homeostasis (Suthedand et ai. 1993), many of the detaïis in this poithwa. are stül to be
elucidated.
Rotein tyrosine phosphatases (PTPs) also play a critical d e in regulating insulin
action in part through dephosphorylation of the active (autophosphorylated) fom of the
insuiin receptor and attenuation of its tyrosine kinase activity (Drake and Posner, 1998).
Recent studies have show that PTPlB knockout mice have increased iiisuün sensitivity
and are resistant to obesity. suggcsting this PTP is a negative regulator in insulin action
and related to the pathogenesis of diabetes and other diseases (EIchebiy et al., 1999).
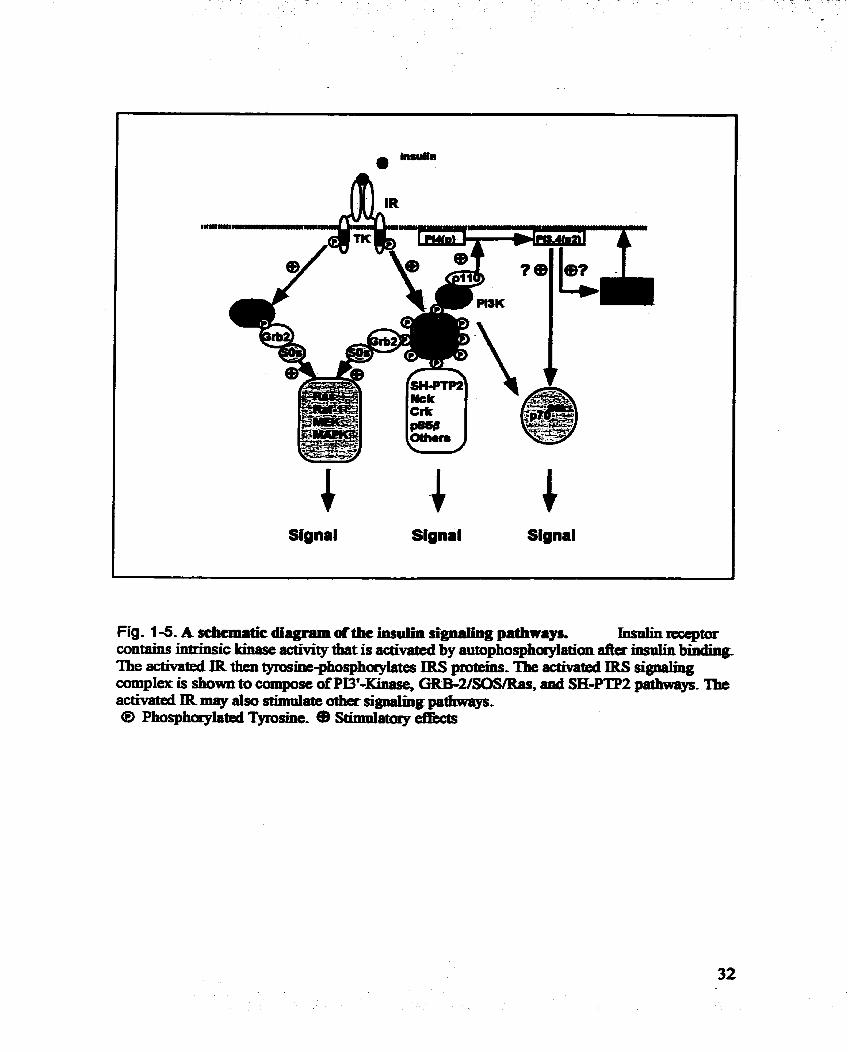
Sianal Signal Signal
Fig . 1 -5. A scbcmrtic &.gram ofthe insulin signaling pathways. Insalinrecepm contains intrinsic kinase activity that is activated by autophosphoay~ation a h mo~lin b & b g The activated IR then tyrosine-phosphoryiates IRS pmteïns. The rr tMted IRS signahg complex is show11 to compose ofPI3'-Klaase, GRB-2/SOSmaS, ami SH-PTP2 pathways. The activated IR may also stimuiaîe other signalïng pa6iwqrs- @ Phosphqlated Tyrosine- (B StimPlatory eftècts

B e s i k the priecipai IRS subsbeates, 0th- such as Shc, might aIso
exkt in the -signahg pathway. Altemative pdtways ofthe IR may be ptsponsible
for the multiple actions of uwlin. Shc is a substrate protein rapidiy tytghosphorylatcd
by insutin StEmulati'on. Shc contains an wkmimi PTB domain which b'î ta the
phosphoryiated NPxUs72 sequence ip the insuücl xeceptor and can also bd Grb-2 to
link these signais to Ras activation (Skolnik et al., 1993; h n k et al 199e Gustafbn et
ai-, 1995). It has been suggestedthaî Shc, rather than IRS-1, may be the main meàiator of
the mitogenic actions of insuiin and IGF- II Cyhaguchi and Pessia, 1994).
5. MAPPING THE INSULIN BINDING SITE
As discussed insulin initiates signai transduction in target ceUs by binding to a
specific ceil sunire receptor. Photoaffinity labeling Mes have impiicated the a subunit
as the location of inmlin-insulin receptor hbmctions (Yip et al., 1978). Subsequently,
several regions in the a subunits of the IR have been shown by different techniques to k
involveci in insulin binding (Yïp et al., 1988,1991; Anderson et al., 1990; Schumacher et
ai., 1993; Kjeldsen et al., 1991,1994; Schaefei. et al., 1992). The molecuiar details of
these events are still obscure and will require a detailed understanding of the structure
fitnction relaiionships of the IR, in particular those of the extracellular domain. The
purpose ofmy study is to explore the structure and hction of the IR., and particulat1y to
focus on the localization of the insuiin-binding site on the IR. Multiple appmaches were
used to achieve these goals. The information obtained would greatiy help us to
understand the mechanism of the insuhm interaction and signal transduction, the
pathogenesis of diabetes mefitus, and eventuaiiy, to assist in designing new medicine to
treat diabetic patients.
5-1- Insulin Interaction with the IR

As mentioned previously, human bulin cocoasists of two chains, the A-chaÏn (21
residues) and B-chah (30 residues), and it is assumed to bind to its receptor as a
monomer (Brange et aL, 1988). The X-ray crystal structure of porcine insulin provided
the fïrst direct experimental &dence for the insalin folding (Adams et aL, 1969).
Subseqpent studies have shown that hsdh A and B ch- fold to foxm a compact
globuiar structure, stabilîzed by the two interchain disulnde bridges.
A vast number of insulins b m diverse animal specïes, and chemically or
geneticaily moWed insulin analogues have been studied for their bioIogicaI properties
and binding to IR The results indicate that many residues of the insulin molecule may
contact orbe involved in the insulin-receptor bmding (Baker et al., 1988; Murray-Rust, et
ai., 1992; Mynarcik et al., 1997). On the putative weptor-bïnding surfie of uisulin,
GlyAl, IluA2, VaW,A19, AspA21, ValB12, PheB24 and B25, TyrB26 are the major
determinants of the receptor binding site, A few other residues make minor contri'butions
Meanwhile, the N-temiinus of the B chah being the most flexible region of insulùi as
suggested by X-ray and NMR studies, was comïdered to be a characteristic of insulin-
insulin interaction rather than being directly relevant to receptor binding W g e r et al.,
1990; Mul~y-Rust et al., 1992). Since mutations in the C-terminal region of the B chab
insulia have been reporteci to be associated with diabetes (Steiner et ai., 1990), residues in
this region has been suggested to play a critical role in the expression of insulin's
biological activity. Many studies have been done to elucidate the role of this region in the
interaction with the insulin receptor Wakagawa and Tager, 1986, 1987, 1993; Mirmira
and Tager, 1989, 1991; Mynarcrk, 1997). PheB24 and PheB25 have betn suggested to
provide much of the binding fke energy involved in inducing the site-site interaction @e
Meyte, et al., ! 978). Systemic studies of msulin analogs m&ed at B24, B25 (Mimina
and Tager, 1989; Nakagawa and Tager, 1986; 1987) have also supporteci the suggestion
that these residues are responsible for the initial interaction with the receptor.

CombinSng information ikrn the crystaï structure of insuiin and its biological
properties, Schaffer pmposed a model for insulin bindlng to the insulin receptor (Fig 1-2
b). He hypothesized that high afhïty insulÎn binding ropuirrd two receptor-binding sites
on insulin. The nrst bindihg site is the classical bhdïng site, mcluding B24, B25, AL and
A21 in the dimer-fomiing d a c e of the insulin molecule- The second binding site
consists of B I7 and A13, located in the hexamer-forming sudace on the opposite side of
the molecule k m the classical binding site (Schaffér, 1994)- Substitutions of these
residues woald be expected to dramaticaiiy decrease insuiin afE~Üty to its receptor. 0th-
snidies also suggest that insulin binding to its receptor may lead to a coaformational
change in the C-terminus of the B chai. a region which is essentid for high mty
receptor binding- The ernerging concept, therefore, is that receptor bindùig is not a simple
collision process, but rather a multiple step process that may involve successively
different residues in the transition towarâs to the active state-
5.2. The liisuïin-Binding Domain of the IR
Unlike insnlin, the exact regions of the receptor that directly contact its Ligand
remain incompletely defined and are stili the object of much study.
Evidence suggests that the binding of insulin at high aflhity involves multiple
regions in the a subunif and even the P subunit of the receptor- The fke single a subunit,
or the a+ monomer, as well as the a-a subunit dimer, has minimal, or no binding
iiffinity for insuiin (Boni-Schnetzler et ai., 1986; Schaefer et al., 1990; Roach et al, 1994).
The minimal tnuicated holoreceptor which still maintains the same high affinity for
insulin is the solubilized whole extracellular domain of the receptor, Uicluding the entire
a and all extracellular p subunit linked in tetrameric form (Schaefer et ai., 1990).
Although there is stili a considerable uncertainty about where the receptor binds to
hsulïn, these observations suggest that the hi&-affnity insului-binding pocket requires a
conformation consisting of the entire extracellular portion of IR It suggests that the

insulin-IR interaction mpks not only multiple specific contact sites but also some
supporting regions to keep the proper biedmg pocket in the right conformation.
A~nroaches to Studv the hsulin-Bhdhn Site
Attmpts to localize the specinc region for high affinity iiuului bmding on the a
subunit have involved severai different approaches, hluding affinity labeling,
recombinant DNA technology, such as sitedirected mutagenesis, deletion studies and the
construction of chimeric receptors comprised of different domairis of the msulùi and IGF-
1 receptors (Tavare and Siddle, 1993)- Although each approach has its advantages and
dîsadvantages, and the results may not agree, these studies do provide usefril information
about where on the IR insuIin binds.
Affinitv Labeling Chernical or photodlïnïty labeling was one of the earliest
methods used to study the insulin-receptor interaction. Photoaffinity labeling (Yip et al.,
1978; 1980) and chemical cross-linlrùig techniques (Pilch and Czech, 1979; 1980) have
been used to show that insulùi prbarily binds to the a subunit of the IR. These
pioneering experiments also revealed the overall a#, subunit structure of the insulin
holoreceptor CYip et al., 1978, 1980, 1982; Pilch and Czech, 1979, 1980; Yeung et al,
1980; Massague et al., 1980). Although they have advantages in the characterization of
the structure and hction of the IR, the chemical bihctionaf reagents or the early
photoaffinity labeling reagents for cross-linking of insulin to the receptor have low
intrinsic efficiency and/or specificiîy. The disadvautaga include the fact that the cross-
linked insulin-receptor complexes are either typicaliy a low percentage of the total LR and
the non-specific cross-linking products may Increase during attempts to increase cross-
hkhg efficiencies (Waugh et al., 1989). Later attempts to develop reagents with greater
efficiency and selectivity have focused primarily on derivatking the E- or a-amino groups
of LysB29 or PheB1, respectively, with additional modifiai aryl azides (Brandenburg et
al, 1980; Ng and Yip, 1985; Yip et ai., 1988; Fabry, et ai., 1992). These new reagents

have much hi* specificity in cross-Iinking and can provide more picciSc iofomation
on -receptor interaction, but their efficiencies are usuaiïy stül below 100& which
limit thek u s t ~ in fbtbnai studies.. Recentiy, a novel b e n z b y I p h e n y ~ B 2 5 -
iIisulur photoprobe was reportad to have an efnciency of cross-linlnnp as high as 60-
100% (Shoelson a ai., 1993). This new class o f photoafflnity reagents would kilitate
the studies ~ f ~ r e c e p t o r stnichnc and fimction.
m e technique ofphotdbity labehg bas been usedto iden* the insubbinding
site. The approach includes a photo-cn,ss-iinkîng process foiiowed by isolation of
receptor-i.nsuh fhgments to aüow the identification of the regions where the puîative
insulin-receptor contact occurs. Using radioactive photoreactive insuiin derivatives that
maintain mon than 75% ofthe bioectivty ofiilsulin, the Iaboratory of Yip found that the
cysteine-rich domain of the IR a subunit contains the insulin-binding site (Yïp et al.,
1988, 1991). This fhding has ban codkned by another sirniiar study (Waugh et al,
1988) and mutagenesis studies, using either site-directive mutagenesis (Rafiieloff et al.,
1989) or HIR/IGF-IR chimeras (Gustafson and Rutter, 1990). However, using different
photopmbes, other investigators later f o d that other regions rnight aiso be in contact
with î n s u h They are the L1 domain, residues 60-90, (Wedekind et ai, 1989), the L2
domain, residues 300-390 Fabry et al, 1992), and the carboxyL tenninal region of the a
subunit (residues 704-718)@Curose et al, 1994). This variation in the d t s indicates the
complicated nature o f the insulin- binding "pocket" and each of these regions may only
represent part of the whole receptor-binding site. The photoatbity labeling appmach
theoreticaily wuld provide the most accurate information for -receptor wn&ting
sites, but the generdy low cross-ünking efficiency and the -ment for a large
quantities of insulin receptor protein make the pst-Iabeling adysis ofthe products very
diff?cult
Recombinant DNA techniaues Recombinant DNA techniques have
fkequently been applied to mapping the iasulin-binding site. Takhg advantage of the
37

highly conserveci domain stnicturt but a large difference m the &nïty to their cognate
ligands, domain exchanged chim&c lnsullnna-1 receptors have been extensively
studied (Anderson et al., 1990; Schumacher et al., 1993; Kjelâsen et al., 1991,1994;
Schaefer et al., 1992). This technique is a usefiil approach for the study of domain
involvement in ligand binding. R d t s obtained have suggested that various regions of
the a subunit are involved in the üisulin binding- These include the N-tenuid region,
residues 1-68 or 1-137 (Schaffer et al., 1993; Kjeldsen et ai., 1991, Schumacher et al.,
1993), the cysteine-nch region (GustafSon et al, 1990, Anderson et ai., 1990), and
regions in the L2 domain (Schumacher et ai., 1993). 'Resalts fiam several cbimeric
studies also indicate that the Cysteine-rich region is specXc only for high afbity bmding
of IGF-1 but not of insulùl (Anderson et ai., 1992; Schumacher et aL, 1993; Zhang and
Roth, 1991).
Many mutations, both natudy occiming and r d t i n g h m siteairected
mutagenesis in the u subunit, also provide important information on insului binding of
the IR Site-directed mutations in the mino acid residues 240-250 of the cysteine-rich
domain ~-Cy~-Pro-Pro-Prr,~Tyr-Tyr-~s-Phe~ to Thr-Cys-Pro-Arg-Arg-Tyr-
Tyr-Asp-Phe-GIn-Asp) has increased the receptor's aflhity to insulin Waeloff et al.,
1989), while a mutation in Phe 89 of the LI domain almost abolishes insulin binding @e
Meyets et al., 1990). A naturaL mutation in Ser 323 (the beginning of the L2 domain) in a
diabetic patient markedly impairs insulin binding to its receptor, suggestiog its cntical
role in insulin binding (Roach et al., 1994). A series of deletions within the extracellular
domain (Schaefer et al., 1990) and alanine-scanning mutagenisis on the whole L l domain
and C-terminal region of the a subunit (Wiiïiams et al., 1995, Mynarik et al., 1996) have
also provided very usefûi information about the localization of the insulin-bïndïng site.
Mutation analysis also show that glycosylation of the a subunit does not &ect insulin
binding (Caro et ai., 1994).

The observations mentioned above sriggest tbt the mnilin-bmding site is a
complex multiple domain structureture It possiiIy involves the LI, C-R, L2, and C-tenninaZ
regions ofthe a subunit Both a subunits sean to be invovdin f o d g the high a£fjnïty
ligand-binding site, and even the $ subm-t may play a rofe m stabilizing the structure.
C~oe1ectrOnMicrosco~v Recently, with the progress of the eiectron
microscope technology, especiaUy the use of scamiing transmission electron microscope
(STEM) in biological studies, cryoelectron rnimscopy has become a new trend in the
study of structure- The concept and f-'biüty ofreconst~cting tbreedimensional images
fiom electron mimgraphs were introduced 30 years ago (DeRosier and Klug, 1968). A
convergence of technical advances in qoelectron miaoscopy and cornputhg image
processing has now allowed this technique to be used to analyze biomolecular structure
without crystaUization. It is clear that the electron cryornicruscopic and image processing
technique are an ideal and powernil tool to elucidate the structure of large proteins that
are quite dÏfEcult for ccconventi~nai" techniques, like x-ray crystallography or NMR. This
technique also c m be used to map out the fbctional domains in a large complex and to
study their conformational variations in mit hctional states. There is considerable
evidence that high resolution cryoelectron mïcroscopy is able to p&de the structure
resolution at below 10 A level (Bottcher et aï.. 1997; Conway et al., 1997; Chiu and
Schmid, 1997; Cheng et al., 1997; Walz et al., 1997). It is especialiy usefid in studying
large proteins, such as d e insulin receptor, which are very difIÏcult to analyze by x-ray
crystaliüation or NMR spectroscopy. This technique alsu has been successnilly applied
to stmcturally localize the ligand or substrate binding site in protein-protein / protein-
Wus interaction (Zhou et al., 1998; Paredes et al., 1998; Wagenknecht et al., 1994). This
new technique gives us a powerful approach in the study of the insuün receptor structure
and fimction-

During the course of my PaD program 1 have taecï biochemitcat and biophysicd
the insulin-insulin mxptor in-OIL In Chapter 2.1 describe my attempts to use the
conventionai biochemicai appmaches to Iocal iP the insulin-bindiag site on the human
insuün rswptor. A bacdovurus expressing system has also been estabfished to
overproduce the fimctionai extracelluiar domain of the HIR Mirent photoafnnty
labeiing and Iimited proteolysis, as weU as various peptide-mapping methods were used
in an attempt to identify the Pnmary site(s) ofthe însdhIR interaction- h Chapter 3,
using cryoelectron mlcroscopy and cornputer image pmcesshg technique, 1 determineci
the quate~1181y structure of the IR and whue the insulin receptor bhds to its ligand A
Nanogold-bovine iIwlui derivative was used as a mark- to locaüze the insuIin-b'îding
site on the 3D reconstruction of IR We have successnilly obtauied the 3D structure of
human insuün receptor and localized b u h in the 3D immbinsuiin receptor cornplex.
Finally, in the iast Chapter 1 discuss the significance and future perspectives on my thesis
work

LOCAUZKIXON OF INSULIN-BINDmG SITE@) ON THE EUMAN INSULIN RECEPTOR BY BIOCEEMICAL APPROACHES
1. LNTRODUCfION
As desc r i i in Chapter 1, the mature humaii insulin receptor @IR) consists of
two a and two subunits linked by diSulfide bonds, with a m a s ofappmxhateIy 450
kDa (Yip, 1988; Kahn and White, 1988). Revious aiXhïty cross-linlring studies using
photoreactive probes or bifimctionai reagents have identifid relativeIy large receptor
fhgments as putative insulin-binding (Wedekind et ai., 1988; Wsuigh a al., 1989; Faby
et al., 1992).
This Chapter descrii my attempt to identifif the insulin-binding site(s) on its
receptor by using various conventional biochemicd appmaches. The laboratory of Dr.
Yip originaiiy Iocalued the insulin-bindhg site to the a subunit of IR (Yeung, 1980). and
then more specifIcally to the Cys-rich domain of the a subunit by combining afnnity
labeling and immunological techniques (Yïp et al., 1988,1991)- My project was a
continuation of these studies The main objective was to use different insulin photoprobes
with high efficiency of cross-linking to label the insuiin-binding domain on the IR,
followed by proteolysis to obtain smailer cross-Iinked receptor fkgments for peptide
mapping and amino acid sequencing, thus to identify the putative iiwlin-binding domain.
This Chapter describes this appmach and the results obtaind

2. MATERLAC AND METHODS s
Intact rraptor or a tnnicated receptor with niII hsubbindiug activity of tée
intact IR was used in the study. Intact receptor protein has previously ban extraded and
pirn'f?ed h m himian tissues or mammalimammalian di iines (Cauh.cFasa9 a al, 1972; Fujita-
Y-hi et aL, 1983; Whittaker, 1987). However, a dEcient q-ntity of the intact
receptor is difncuit to obtain h m these sources for the identification of the insuiin-
binding site@). Since it is weU estabiished that the entire extraceiilar domain of IR is
requited for the high-aff?nity binding of insulki (Sweet et al., 1987; Schaefer et ai., 1990).
1 used the bacui0vVu.E expression system @ES) to express the entire ectodomain of HIR
to obtain substantial quantities of the pmteh for this study.
Construction of a cDNA ExDression Plasmid Encodine the ExtraceUuïar Domain of
HIR - The clonhg strategy of the BES expression v e r is shown in Fig. 2-1. To take
advantage of the bscuîovinis expression system, the gene of the HIR most be cloned into
a baculovinis transfer vector which is designed to position the EIIR gene immediately
dowmtream of the powerfbl bacuiovirus polyhedrin gene promotor. Ail the restriction
and rno&@hg enzymes in these experiments were purchased h m Pharmacia,
Boehringer-Mannheim, or New England Biolab. AU adapters and linkers were custom
synthesized by the ASC Biotechnology SeMce Center, Toronto
Pre~aration of eonstiucts Preparation of the transfer vector consisted of
several steps. cDNA encodîng the entire ectodomain of HIR (hcluding aii of the a

Signal Cys4ch a subunit p subunit
Encoded Protein
Recombinant Baculovirus
Recombinant HIR Extracellular Domain
Fig. 2-1. Expression of humm insuün mxptor crtrrcclluhr d d n with baculovinu erprtssion systcm. A 2.9 kb cDNA d g the extracclIular domam of hIR was cut than pRSV-IR plasmid and mbcioncd mto a bacdovirus expres~ingvector~ linsect cclls hkted with the recombinant virus were used to express the riecornbinant reœptorprotcin,

subunit and ail. of the extraceiiulairp~~on ofthe Jubmiif except for t h e amino m*dS
N-teaninal to the n'ansmcmôrane domain) war txcised h m the 9-7 Kb p M d pRSVIR
(a genemus gift fiom Dr. Ira Ooldfine, UCSF). Restriction endonuclease Srp I was useû
to digest the pRSVIR to remove tbe CD= sfuthg h m nuckotide 2999 of the receptor
cDNA, whÏch encodes the putative transmembrane domain and the eotin cytophsmic
portion of the subunit of the HIR. Ta fiditate the expression and the purification k,
a stop codon W G ) in axbd linka:
5'-CTArrTrTAGACTAG-3r
3 '-GATCAGATCTGATC-5'
L V s t o D
or, dtematively, an adapter encoduig an extra 6-Histidine amino acids pnor to the stop
codon was inaoduced at the SFpr site. In the latter case, the adapter and the expected
deduced protein sequence at the C-terminus of the protein is therefore:
5'-TCTCATCATCATCATCATCATGT-3'
3'-AGAGTAGTAGTAGTAGTAGTACAGATCS'
S H H H H H H V s e
The resulting intermediase plasmid, designateci as pIRX or piRXH (H represents paly-His
tag), was digested with NcoI, which cleaves at a single site at the beginning of the cDNA
(nucleotide 137) corresponding to the nrst metbiorùne of the signai peptide at position - 27 of the HIR protein. The digested plasmid was then blunted with the Klenow niigment
of DNA polymerase 1 to n11 in the ends. A M linker
5'-ATCGCTAGCGAT-3'
3'-TAGCGATCGCTA-S'

was then ietrodueai at the b I d Ncd site ta gcriercite the second p l d d
ofpIRXN or pïRXNH (4.0 KB)- Digestion of above the phsmid~ ~6th -1 aad X6rrI
generated a 2.9 Kb restriction fiagrnent of the tnmcated HIR, nucleotides 138-2998- It
encodes the entire extradular domain of& HIR, except the three amino acid LeSidues,
1 A Y, prior to the ttansmembrane domain of the $ subunit, wîth or without a 6-histidine
fag on the C-taminal enb The hrhef-Ba1 hgment was then subcloned into a l\rheI c i 9
cal€ intestinal phosphofase-treated bacdovirus transfer vector pBlueBac2 or pBlueBac4
(hvitrogen). The zight orientation of the insert CDNA m the transfér plasmicl, designated
as pHIRa or p H I R a was detemineci by vsrious restriction endonuclease digestions.
These transfer piasmids for M-transfdon were prepared by CsCl gradient cenpafugation
in a Beckman rotor VT801 at 80 K x g, at 22 OC for 4 hours. The purifid piasmid
. * coll~t~cts were steniized with 0 . 2 2 ~ filter and saved at 4 O C .
DNA seaaencin~ of the coastructs
In order to confinn that the nucleotide sequence was correct for the recombinant
protein, the DNA of the 2.9 Rb insert in the transfèr construct was sutscloned into
pBluescript (Stratagene) vector at the XoaI site (PBSHIRa, bp 138-2998), or crb with
restriction enzymes and subcloned into tbree sub-cunstructs in p B l d p t vector
(pBSHIR1, bp 559-1273; pBSHIR.2, bp 1101-1695; pBSHIR3, bp 1695-2606), thus
allowing the whoie 2.9 K . cDNA hgment to be sequenced Fige 2-2). AU constructs
were m a n d y (Pharmocia DNA sequencing Kit) or automatically (Perkin-Elma-AB1
Genetic Analyser) sequenced. AU sequenced DNA showed no e m r in the nucleotide
sequences.

Fig. 2-2. Comstnrcioii of the BES transfiin vcctors br tht bïRcctodomain with (A) or w i ü ~ ~ u t @) 6 Eis tag. A29 kb ~ D N A t h g m a ~ t ~ a ~ cuî&mpSViR mdmodined üyaddingprop~adaptor~, with or without 6 Air rcriducs, and stop axbn. l%c modifiai cDNAhgmcas wat suôcloned into pBlueBac4 @BB4) or pBlueBac2 @BB2) Wovinis aansfection vcctor-

Log phase ùiseet Sp (S. fbgiperdzt) cek @.vitmgen) were p w n as anamiaent
culture at 27OC in modifled Grace's medium suppIemented with 0.33% lactalbumin
hydrolysate, 0.33% yeastolate, Iû% fetsl bovine sarmn (FBS), 10 ps/mL gentamycin a d
250 ps/ml amphotaich B &Se-Technofogy). In some experiments 0.1% of Pluronic F-
68 (L'Se-Technology) was addd to suspension culture of Sfl cells to d u c e the
sensitivity of the culture to mechanid shearing- Awthn insect ceii line, High-five (Hi3
ceiis @ichoplusia nl) (Tnvtrogen), was cuitured in Exce11 401 saum fia medium (JRH
Scientific) containhg 10 mdmi gentamycin and 0.25 m g f d amphotericin B to yield a
higher expression level- AU procedures involving celi culturîng, includhg routine
subculturing, and protein expression, were performed as d e s c n i in the M&~uals of
Methods for Bacuiovinis Expression h m Invitmgen.
Recombinant plasmi& were CO-transfected into SB cells dong with the linearized
wiid-type AcMiWV vual DNA (for p-) or Bac-N-Blue lineanzed Wal DNA (for
pHIRaH) by the technique of cationic liposome mediateci transfection (bvitrogen). In
vivo homologous recombination between the polyhedrin sequences in the Md-type Wal
DNA and the recombinant plasmids d t e d in the generation of recombinant viruses
coding for the expression of HlR ectodomaia, Briefiy, lpg virai DDNA, lpg of
recombinant tramfiet plasmid, and 20 PI of Insecth liposome @mitmgen) were mwd in
1 ml of Grace's media without FBS and suppiements, The mixture added to hshly
d e d Sfl celis in plates and incubated for 4 hours on a sIowly rocking platfom at 27OC.
Then, 1 ml of wmplete Grace's medium was aàded and incubated at 27" C for 48-72

hours. The medium b r n sueoessfiilIy co-traasfected, culnaed ails ans scrrmd by a
plaque assay for tht cccombinant vhs contrllning the msulin ncepor cDNA
Piaaue Assa. and Pre~antian of FliPb Titer Vira1 Stock
For the piaque assay, SP ceils in 60 mm plates were hfkcted with 1 ml of 1 x
102, 103, 104 diluteci media containhg CO-transfected virus for 1 hoia at 27°C. The
media were then removed and repiaced with a 4 ml overlay of 1.5% Sea-Phque agarose
(FMG Biopmducts) containing 0.01% X-Gal. Af€er 4-6 days' incubation at 2 7 O C, weil
separaîed, distinct blue piaqyes were isolatecl and used to hocuiaîe SiJ9 celis in multi-weil
plates for M e r screening and p r e h b r y virus ampMcatioa Usuaiiy, 612 plaques for
each recombinant protein were s e l m and 3 4 clones of the finai purifiecf virus were
saved at 4 O C as P-1 WaI stock for mer characterjzation and amplification. The P-1
virai stock was then used to infect log-phase Sfl celis in a T-25 flask for 5-10 days at 27O
C to generate P-2 virai stock The ampfified P-2 viral stock then was used to i n f i Sp
ceus at a ratio of 1: 10 (vfv) or MOI 0.5 - 1 (see detaüs later), either in adherent culture in
T-225 flash (Costar) or in suspension culture in 500 ml spinner flash (BeUco Glass) et
27O C for 5-8 days. This P-3 high titer vUal stock was analyzed in a plaque assay
following the same schebile as previody desctibed except the viral inoculate was
diluted up to 1 x 10-9, to measure the virus titer. The plaque forming units pet ml
@fu/ml) were calculated using the following formala to set the optimal conditions for
protein expression:
pfii/ml= l/dilution x mean number ofpleqyedplate x platelml

To obtain a synchronous Mixtion for e q m s h g the recombinant receptor* a
particular multiplicity of infection WOI) h c k of 5-10 was iiscd to caicuiated the
volume of the Mirr stock with thefoiIowhg fomuk
[MOI ( v i r o d d ) x number ofcelIs]/titer ofvirus (Vli.oIIS/ml)=ml ofinocuium stock
Hi@-titer virai stock was kept at 4 O C in the dadc mtil use. A smd aliquot was
hzen at -70° C for long-term stomge.
ExDression and Characterimation of the Recombinant IR
MOI o~timization for vinw iafiition High titer viral stock was useâ to
infect the Hi5 ceils in 2 ml of Exceii-401 medium in T-25 flasks at -90% confluence at
vaqing MOIS (2, 5, 8, 10 and 15) for 1 hour. Three ml of Eresh media were then added
and the ceUs were cuItured for 4 days at 27O C. Anti-HIR immunoblotting and B29
~~~-[125IJ-iodoinsulin photoafkity labeling ( see details later in this chapter ) were
used to check the levels of protein expression. An MOI of 5-10 was chosen in our
expression infection. On the other hanci, for generating hi*titer virai stock, an MOI of
0.5-1 was used to avoid any mwatlted interference in virus production. Western blots
with anti-IR a subunit antibody and r n o n o - [ 1 2 ~ 1 binding assays were carrieci
out to confimi the expression of protein and protein fiinction (see detaiis inter).
Time course and lime sale exDression of the recombinant rece~tor Hï5
cells at -90% confluence were Ilifected with viral stock at an MOI of 10 in 6 weii plates
at 27O C for up to 7 &YS. Media was rexnoved at O hout h m one weii and at eveiy 24
hours h m other wells to m o d r the infidon and expression process. The media h m
the infected ce& as weil as the corresponding ah, washed with pH 7.4 PBS with

cuitme, ~i5ceiIs weniaféctedatacel1daisityof2r 10~ce~swithviralstock atan
MOI of 10 in a 500 ml spinner flasL at 110 rpm at 27' C. One ml of suspension was
collected at zero time and each day foilowing for 7 days, h m the cuituze. These samples
were treated the same as the adherent culture-
In preparetve culture fôr protein expression, HiS cells were cuitured in Exceli-
401 medium to -90% confluence in T a Basks at 27O (=. A f k removing 25 mi of
media, the ceiis were infectecl with viral stock aî an MOI of 10 for 1 hour. Fresh medium
was repiaced to a -38 ml and the ceiis were cultured at 27O C for 4 or 5 days. The
cultured media were coliected for purification of the recombinant protein.
Pre~arinvceU lvsates and lasm ma membranes of the infecteà insect cells
Cell lysate samples for the tirne course sbudy were prepad by M y adding
SDS-PAGE sample bufEa (Laemmli et al., 1970) containkg 100 m M D'IT to the pre-
washed cells for subsequent SDS-PAGE analysis.
The plasma membrane and miczosome fiactions of the celis were prepared as
foilows: Hi5 cells in 25 ml of adherent culture were coliected at Day 4 after infection and
resuspended in 5 ml of pH 7.4 solubilization b s e r (Bu81er A) containing 50 m M Hepes,
150 m M NaCl, 0.1% Triton X-LOO, 1.5 m g / d Bacitricin, I O m M b e d d i n e HCI,
2mM pepstatin, 2mM leupeptin, 5 mg/mI aprotinin and 1 m M PMSF. The cells were
homogenized with an ice-cooled glass-Teflon tissue grinder (Kontes Glass) for 50 stmkes
then centrifiiged in an SS34 rotor -nt) at 1,000 x g, at 4 O C for 10 minutes. The
pellet containing the nuclei was rrsuspended in Bunet B (8uner A without the 0.1%
Triton X-100). The supernatant then was centrifitged in an SS34 rotor at 35,000 x g, at 4 O

C for 30 min, The pellets were resuspended in B e é r B to yield the sample of plasma
membrane- The supernatant was subjected to M e r ultracentüfügation in a Ti 50 rotor
(8eck;mm) at 250,000 x g at 4' C for 60 min. The pellet was resuspended with buffér B >
as the microsorne sample-
Anti-ER]Immunoblo~g Samples of cultumi media, cell Iysata, nuclei,
plasma membrane and lm-msome wne solubilized by boiiing for 5 min. in LaemmIils
sampie b s e r (La& et al., 1970) containing 100 mM DTT, and then separated by
electrophoresis on 7.5% SDS-PAGE, followed by t r a n s f d to 1mmobilon-P PVDF
membranes @dïUpore)- After transfet the membrane was blocked with 5% skim milk
powder in pH 8.0 TBST buffer (10 mM Tns-HCI, 150 m M NaCl, 0.1% Tween-20) for 60
min at room temperature and then incubated with rabbit anti-IR a subunit anti'body
(Santa Cruz) in blocking bufk over night at 4 O C- After washing with TBST buffer, the
aembrane was incubated with goat anti-rabbit alkaline phosphatase conjugated annibody
(Sigma) in blocking b s e r for 60 min at room temperature- The recombinant proteins
were visualized using the AP substrate color kit pharmacia).
Insulin Binding Insulia binding was measured by incubating - 50 ng of
human placenta membrane insulin receptor or recombinant insului receptor protein, 20-
30 h o 1 of m o n o - [ 1 2 5 I J ~ 1 4 - ( final concentration of -50 pM ), and varying
concentrations ( h m O - 1 x 10-5 M ) of unlabeled insulin in 200 ml of buffer, pH 7.6,
containing 50 m . Tris-HC1, 150 mM NaCI, 011% bovine serum albumin (BSA), 1.5
mghi bacitricin (0.1% Triton x-100 in the butEr for placenta membrane rezeptor) at 4 O
C over night. The receptor-insului complex was separated from unbound insuiin by
adding 50 pl of 0.4% bovine y-globulin and 250 pl of 20% polyethylene glycol 6000

f o i l o d by centnfiigation at 1,SW x g for 20 min at 4" C. Radi08Cfivity in the pellets
was determined in a PJmmmcïeLKB y-counter- The lgdioactbity daccted h the
presence of 10-5 M of uniaôeled iasulin was consicid nomspecific binding and was
always less than 5% o f the total activity. ALI binâing experiinents w m e pediomed in
duplicate on thme seprirate o c c a s î ~ ~ . The cornpetition binding curves were compiied
h m several experiinents, and each point was reported as the mean +/- SD- nie
Scatchard plot was a n a l . with EBDA cornputer software (Muoson and Rodmi,
1980)-
For samp1es of time course and subce11uïar fhctions, insuiin binâhg was
measiued as foiiows: 50 pl of harvested media or resuspended M o n samples were
dispersed in a total volume of 200 pi of the above binding bu&r plus 0.1% Triton X-LOO,
and - 20-30 h o 1 of mono-P2S1]~14ins~lin, and assayed as descnbed above.
Photoaffinitv Iabetinn and autoradionnuhv - Purifieci or Cnide protein
samples, such as harvested media or plasma membrane, were incubated with about 1 x
106 cpm of ~2g-MAB1:-[~~51]-iodoinsuün-o (Yip et al., 1980) in 200 pl insuiin binding
buffer with or without au excess (10 mM) UnlabeIed insuiin overnight in the dark at 4 O C.
Photolysis was carieci out for 15 second using a focused iight source h m a 100 W hi@-
pressure mercury lamp (Yip and Moule, 1983). Photolyzed samples were reduced and
s o l u b i i in Laemmli's sample bUner containhg LOO mM DIT- Cross-iinked rrceptors
were either separateci by SDS-PAGE directly or precipitated with anti-insulin receptor
antibodies pnor to SDS-PAGE. Cross-linked, radiolabeled proteins were identifid by
autoradiography of the nx.d and dried gels. The radioactive bands on the dned gels were
cut out and wunted in the y-counter.

MoliniLr m a d o n SampIes - aarilyzcd by 3012% lïnear
gradient SDS-PAGE imda reduced (10 mM DIT) or no~reâuced conditions. 'Ihe gels
were nxed in Io./; acetic acid and 25% isopmpsinol for 30 min Proteins wae visuaüad
using either a dver stain kit @bRad') or by C o o d e BSUiant Blue staining. The
molecular mass of the detected proteins was eshated by cornparhg the relaîîve mobility
of the receptor bads to molecular mass standards (Bio-Rad, orNovex)
2.2. Purification of the Recombinant IR and Human Phcenta Membrane IR
Prenvrtion of iasulin iffinitv coiumn Twenty five mi of m g e l 10 (Bio-
Rad) were washed with one volume of cold isopmpauoi, foliowed by 3 volumes of cold
EQO and were then equilr'brated with the coupling buffer, pH 7.4, containhg OJM
phosphate and 6 M urea Twenty five mg of zinc bovine insulin (a generous gift h m
Connaught Canada) in 25 ml of coupiing buffer was mixed with AfE-gel10 for 24 heurs
at 4" C. A trace amount ofrnono-p2I]~l4iiwlin was added to m o d r the coupling
process. The suspended gel then was transferred to a coiumn and washed with 20
volumes of the coupling buEer and fkther washed extensively with bufEer containing 50
mM Tris-HCl, pH 7.4, 1 M NaCI and 0.02% NaN3, Total radioactivity in the washed
bufEer and the wlumn was counted to calculate the couphg efficiency.
Pre~aration of MA51 monoclonal antibodv rtninitv column Mouse
anti-insutin recqtor monoclonal anti'body, MA51 (IN1 subclass, a generous gift k m
Dr. Ira Goldfine, UCSF) was prepared h m the mouse ascites the mAb MAS1 at - 1-3
mg/ml. The p H ofthe ascites was adjusted to 9.0 (wlth borate buffer) and NaCl was
added to a concentration of 3 M. Twenty ml of ascites containing MA5 1 were mixeci with

15 ml of protein &Sephamse beads @bunaci&) and hcubated for 60 min at m m
temperatureperature The btads were washed twiCce with 10 volumes of 3 M NaCl, 50 m M
sodium borate @H 9.0) by centrifugalion and aspMollc The beads were muspeded in
IO voiumes of 3 M NaCi, 0.2 M Murn bonite (pH 9.0) and mked with
dimethylpimelimidate @MP) to a final concentration of 20 mM- Afkr a 30 min
incubation at m m temperatme, the coupling was stopped by wasbing the beads once
with 0-2 M ethanolamine @H 8.û) and incubated with the same buffer at m m
temperature for 2 h o w with gentie mirring. The beads w m then extensively washed
with a birffe~ containÏng 50 m . Tris-HC1, pH 7.6, 150 mM NaCl and 0.02% NaN3 at 4
Purification of the Recombinant Eetodomain of IR
ARer 4-5 days infecton with viral stock, low-senim-containing medium was
hanrested and adjusted to pH 7.4 with 1 M Tris-HCI bu&r. PMSF and NaNj were addd
to a finai concentration of 1 mM and 0.02% respectively. The conditioned medium was
centrifugecl in a JA14 rotor meclanan) at 4" C, at 10,000 x g for 25 min to remove m y
cells or debris. The supetnatant was concentrateci -50 fold using a Minitan Uttafiltration
System (Mülipcwe) with a 100 LDa cut-off membrane to remove proteins of lower
moIecuiar as the nrst step of purification, The concentrated medium was then adjusted to
50% ammonium sulfate by dropwise addition of saturateci ammonium sulfate at 4 O C.
M e r ovemight stimng, the mixture was centrifiiged at 10,000 x g for 30 min at 4 O C, to
obtain a peiiet which was resuspeaded in 20 ml of 50 mM Tris b&er @H 7.4) and
dialyzed exhaustively at 4 O C against either the His-binding b&er (20 mM TnS-HCI, pH

8.5,O.S M NaCi, 10 mM imidazole, 10% giycerol, 0.02% NaN3 and 1 m . PMSF for 6-
Ris tagged ectod~main)~ or the buffer containhg 50 mM Tnis-HCI, pH 7.6, 150 mM
NaCî, 0.02% NaN3 and 1 mM PMSF for the ectodomain without the o-His tag.
Ammonium d a t e precipitation folIowed by diaïysis M e r rernoved Iow molecuiar
mass molecules and up to 40% of the BSA in the concentrated medium to facilitate
subsequent purification-
Purification of the ectodomain of the IR ais GHk tag Ni-NTA r e s h
(Qiagen) was washed andpre-charged with 100 mM NiCl and then equilr'brated with His-
binding buffer, pH 8.5. Twenty five mi of concentrated, dialyzeâ was mixed by rotation
with 300-500 pl Ni-NTA resin over night at 4 O C. The resin was t r a n s f d to a 2 ml
mini-column (Bio-Rad) and washed with 10 volumes ofHïs-binding buffer, followed by
50 volumes of washing buffer (20 mM Tris-HC1 pH 7.8, 0.5 M NaCl, 20 m M
immidazole, 10% glycerol,0.02% NaN3 and 1 mM PEIISF). AU b & i were kept on ice
when in use except that the Ni-NTA column was used at mom temperature. Recombinant
protein was eluted by washing the resin 5 tirnes with 0.5 ml of elution buffer (20 mM
Tris-HCI, pH 7.8,0.15 M NaCI, 250 mM immidazole, 10% glycerol, 0.02% NaN3 and 1
mM PMSF). The pooled eluted protein was uitrafiltered in Ultramax-50 (Miiiipore) to
concentrate the protein and change the bUner to 50 mM Tris-HC1, pH 7.5,lSO mM NaCl,
1 mM PMSF and 0.02% NaN3. M e d protein samples were aliquoted into severai
siliconized microfuge tubes and stored at -70" C.
Puriecation of the ectodomain of the IR without dHis tag Twenty five to
fîfty ml of concentrated and dialyzed medium was mixeci with 12 ml of MA51-protein A
beads by rotation ovemight at 4 O C. The mixture was transferred to a column and washed

wita 500 mi bUna containirig 50 m M Trk-HCl, pH 7-4,150 mM NaCL and 1 mM PMSF*
The bound protein was eluted with an- elution bu&r conta- 2.5 M MgC12, 0 2 M
concenfrated in an Amïwn ulaafiIttaSrCon device with a PM-30 membrane- The
wncentrated protein then was loaded on a Dextraa G-25 wlumn to ranove the high
concentration of sait The punfied protein h m the MA51 af'hity column was saved in
siliwnized microfuge tubes at -70" C,
Both Ni-NTA and MAS 1-protein A column purifid proteins were purined M e r
by gel filtration chmmstagrqhy on a FPLC Superdex-200 HR 16/60 column
(Pharmacia) in 20 mM Tris-HCl, pH 7-6, wntain. 150 m M NaCl, and 0.02% NaN3 to
remove either giycerol or/and the small amount of coeluted contamimieci proteinS. The
purifiai ectodomains of IR were a d y z e d by SDS-PAGE (Laemmli ct al., 1970) with or
without reducing agent (100 m . D m to check the punty, and by 12~1-insulin binding
for their activim
Puritication of haman insulin rece~tor
The methods of p r e p a ~ g the placenta membrane human insuiin receptot wae
modlfied h m Fujita-Yamaguchi a aL(1982). Briefly, a fresh n o d human placenta
was obtained within 1 hour after delivery, kept on ice, washed with normal saline (150
mM NaCL) 3 times and 0.25 M se once, and then cut into 4 pieces. The pieces
were tramferreci to 1 volume of 50 mM Tns-HCl buffer, pH 7.4, containhg 0.25 M
sucrose and 1 mM PMSF, homogenipd for 90 seconds with a Polytron homogenizer
(??olytron), and centtifiiged at 15,000 x g in a JA14 rotor (Beckman) for 35 min at 4* C.

MgCl and NaCl were added to a final concentration of 0.041 mg/ml and 5.84 mg/m.i,
respectively, The supematant was then centrsfiiged in a SW28 rotor @eckman) at
100,000 x g for 25 min at 4 O C- The peilets obtained were resuspended in homogenization
b&er and the spin/resuspend steps were repeated three times at the same speed but with
increased times (30,35 and 45 min). The final peUets were saved at -70" C until analyses
or fbrther purification
Placenta membranes (0.8-1.2 g of protein) fbm two fksh placentas was used to
obtain the pUnned human insulin receptor. The placenta membrane pellets were
solubiiized in 25 ml of 50 mM Tris-HCl, pH 7.4, buffer containùig 2% Triton X-100,100
mM EDTA, 1.5 m g h l bacitracin, I O mM benzamidlne HCi, 21x1. pepstatin, 2mM
Leupeptin, 5 p s / d apmti'nin and 1 mM PMSF. The suspension was c e n e g e d in a
SW28 rotor at 100,000 x g for 90 min at 4 O C, The supernatant was then loaded on the
insulin column which had been pre-eqyiiibrated with the washing b u f k of 50 m . Tns-
HCI, pH 7.4, containing 1 M NaCl, 0.1% Triton X-100 and 1 mM PMSF. The coIumn
was rotated ovemight at 4" C. It was then washed with 400 ml of washing b a e r and
eluted with 50 rnM acetate buffer, pH 5.0, containing 1 M NaCl, 0.1% Triton X-100 and
1 mM PMSF. The neutralized hctions with insulin-binding activity were pooled and
fùrther concentrated and desalted by ultrafiltration using Diaflo membrane PM-30
(Amicon). The partialiy purified receptor was M e r purifiecl by gel filtration
chromatography on a FPLC Supercryl-200 HR 16/60 column (Pharmacia) as previously
described, and saved at -70 OC.

The amount of protein was m e d by a modified Lo-s method (Bensadoun
and Weinston, 1976) or Btsdford's methad (Bio-Rsd)* using BSA as a protein standad
Amino acid adysis was pebormed by the SHC BiotecbnoIogy Service Center, Toronto.
Insuün Photoprobes and Photoafünitv Inbelhg
N-e(4'-Azid0-3'-~~iodopheaylau,~3-aminopmpyl iiisuün ( p q - A Z A P -
insuiin) (Fig.2-3) is a cleavabIe radioactive insulùi photoprobe *ch aiiows the iiisuün
moiety to be cleaved h m the labe1ed receptor protein a f k photoaf3hity labeling. Ne-
Monoazido-benzoyl-['Y ]-iodoinsulin (829-[IV-MABr) is iilJulin dgivatizcd near its
putative receptor-binding region, aud it retains more than 75Y0 of the bio-activity of
insulin CYip et al., 1980; Ng and Yip, 1985). Both [13+AZAP-insulin and B 2 9 - [ q
MABI were synthesized in Dr. Yip's laboratory* [q-Iodo-biotinm-p
benzoy1phenylaianinP-insulin (m-BBpa-insulin) was provideci by Dr. Steven
Shoeïsoq Harvard Medical School. It was a new photoprobe with two markets
(radioactivity and biotin) for detestion, and was reported to have much higher cmss-
ünking efficiency of 60-100% (Shoeison et al., 1993). In prrparative photolabeling, the
photoprobes used in photolysis were a mixture of trace arnounts of radioactive '9 labeled
photoprobe and non-iodinated photoprobe.
Photoiysis conditions were optirniPd for high photolabelhg efficiency with the
least damage to the protein. In p r e p d v e photoafbity labeling, the mixture of non-
iodinated and 'DI labled photoprobe and receptor protein at a 5: 1 molsr ratio (for 4

A Chain: GNEQC!CASV&LYQLEWCN \
\ B Chain: FVNQHLCGSHLEALYLV 4 GERGFFITP r
A Chain: G W E ~ ASV~SLYQLEWCN F * C * B Chain: FVNQHLCGSHLVEALVLV GERGFF'VTPW
A Chain: G N E Q ~ * B Chah:
Fig. 23. bul in photoprobcs uscd AZAPBi is a cleavable phoîoproble. Anowhead. indicates cleavage site m the probe. BBpiuBI has a biotin madra on the B29 Lysine. * iiadicates Radioia- beiing Ptcs on the B chin -1 and BBpauBI) a the cross-Iinting p u p (AZAPBI).

HiR) were incribiited atpH 7.5 Tris/NapCT binding biiffi with protease inhibitors at 4OC in
the dark for 20 hours. Photdysis Ïn prepaestive was canïed out imdg W
Iight with 1 0 W m powa for 15 seconds or with a 365 mn nIter for 45 minutes. Dtiring
photolysis the pr~te~photoprobe mhmewas keptonice.
Reduction and AUNhtion
The photoIyzed IR mixture was adjusteci to p H 82,O.l M TSs, 10 m M EDTA
and 8 M urai and kept at 50DC under N, and da& for 30 minutes. A 25-fold molar excess
dithiothreitol @Ti') then was d e d and iacubgtsd at 3PC under N,for 2 hours. Fresbly
prepared iodoacetamide in 8 M inca, 0.1 M Tris, pH 8.2 b u f k was added and the
preparation was incubated at m m temperature for 30 miautes. The duction and
alkylation were Camed out in the da&
To remove excess fke photoprobes, the subunit portion o f the receptor and
salts, the reaction mixture was separated by electmphoresis in SDS-PAGE under
reducing condition. After electmphoresis, the gel was irnmediately h z e n in dry ice and
an autoradiograph of the gel was obtained to 1- the photolabeled receptor band on the
gel. This gel band was then excised, cut into smaii pieces anci soaked in a pH 7.6,20 mM
NH4HC03 bu&r at 4 O C overnight. The extract was lyophilîzed and the gel pieces were
DiSemnt proteases or chemicai reagents were tned to obtain complete
hgmentation of the labeled receptor. Dirrct digestion of the extracted labeled protein or

the electmblotted protein on PVDE membrane was triai to avoid loss &ring s a q I t -
transfer. Before ptadysis, SDS wa+ rcmovcd by using the 8CCtOne-triethanolami'ne
precipitaîion m e & d Typicaiiy, the enzyme to protein ratio was kept at M O (wh) to
maiatain sufEcient digestion while the amount of enzyme was at a minimum, The
digestion tirne for each enzyme or c 1 e a . e with CNBrwas tested. Control digestion with
enzyme only was always cam*ed out at the t h e of a proteolysis experiment Roteh
fhgmntation was.monitored by 9-26% gtadi'kt SDS-PAGE, 40% a h h e PAGE and
autoradiography, AU enzymes used were of sequence grade purity.
For digestion with trypsin, 10-30 pi of TPCT-treated trypeùi, depending on the
quantity of protein, was added to the extmc&d protein dissollved in 250 pl of fkshly
prepared 2 M urea 0-4 M -CO3 bufk(pIi 8.0) and incubaîed at 37 "C for 24 hours.
Endopmteinase G1u.E (Va) was used unda sunilsr conditions to digest the receptor.
Digestion with endoproteinase Lys< or Achromobacter protease 1 (\Kako Chernicals
USA Inc) was d e d out by addïng a srnail volume to the protein solution to give a 150
to 1:lOO (wlw) pmtease: receptor ratio and the murtinr was incubateci at 37 O C for 6 8
hours. The &on was stopped by imrnediately W g the solution at -70 O C .
Cyanogen bromlde (CNBr) cleavage of the receptor protein was d e d out by
dissolving the protein in 200 pl of7W f o e c acid followed by the addition of CNBr at
100-fold m o k excess over the methionine residues of the protein- The mixture was kept
in the under nitmgen, at m m temperature for 24 hours, The sample was diluted 10-
fold with water and lyophilized

Hï?LC Pe~tide Isolation
The enzymatic or chernical digest was ac~*difïed with 10 pi of 10%
trifluornacetic acid ('EA) and then injected into a 2.1 mm x 25 cm Vydac Cl8
narrowbore 300A reverse phase HPLC column in a Waters 410 HPLC system (Waters)
with a detector at a Ieadnig wave length of 214 nm, The co1um.u was eluted at a flow rate
of 0-15 d m i n with a linear gradient h m 0-70% acetonitrile/O0065% TFA in 120
minutes, The fiactions were h z e n immedïateLy after coUection.
Identification of Labeled Fra~ments
Identification by rntibody to the B chah of insalin The m e n t of the receptor
that cross-Linked to the insulin photopmbe should contain a fiagrnent of the B chain of
insulin fier reduction - aikylation and enzyme digestion. To iden- this peptide whïch
spdcalIy cross-linked to the B chain of insulin, bovine insnlin B chain was used to
immunize a rabbit to generate a poIyclonal antiiody for an enzyme-linked
immunosorbant assay (ELISA) experiment to detect HPLC fiactions that contain B chah
or B chain-receptor hgments.
ELISA was established for detecihg peptides conaining the B chain of insulin.
To increase the reduced antigen absorption to the soiid phase of the multiwell plates due
to the acetic, organic solvent h m KPLC @a 2.0, up to 70% acetonittde), a covalent
cross-luiking plate, Reacti-Plates (Pierce) was used to nX the peptide on the plates.
Brïefly, 10 pl of the ftaction h m HPLC was added to the welï containhg 100 pl of PBS,
pH 7.6, and incubated at 4 OC over night. Then the plate was blocked with 3% BSA in
PBS for 60 minutes at rnom temperature. 25 pl ofanti-insulin B ch& antiiody at 1:1000
62

dilution was added and 100 fl o f f ((alkalie phosphatase&mnjugated second antibody
at 1:25,000 düution was used for the color reaction Varied concentrations of insulia B
chain were measured at the same time as the standard curve.
Identitication based on biotin-a~din intera&on To take advantage of the second
rnadcer of biotin on the BBpa-insulln, a sensitive biotimavidin assay method was
established to detect the fiaction contalliing the biotin-B chain signal. Similar sample
fixation was done for the ELISA. AAer blocking, A P - c o n t e d streptavidin wiis added
to bind the biotin-contained peptide onto the plates. The color reaction with the enzyme
then was carried out. Altematïvely, to obtain higher sensitivity, extra biotin was added to
bùid the remaimng binding sites of streptavidin, resuiting in more streptaviidin-enzymes
bound, which, in turn, led to an amplifid signal.
3. RESULTS
3.1. Recombinant HIR ExtraceMar Domain
AU inserts in the virus constructs were found to have the correct DNA sequences
by manmi sequencing (data not shown). The titer of purifid recombinant vinises was 5 x
8 9 10 to 1 x 10 as obtained by plaque assay. Within 24 hours of infection of Hi5 ceîls with
the recombinant virus, uisulin-binding activity was detectable in the culture media and
increased with time until Day 4 or 5 aAer infecton. Then the secreted protein in the
culture media degraded gradualiy when the ceils started to lyse (Fig. 2-4). While the

m u -
Hl- HIR H I b HIR
Fig. 24. Time course d ~ ~ i o n ofthe rccombiiunt atrrcdluiu domain of hIR (bIRar) (A), rnd Western blot (antia subunit of UR) of IüRa rad bnmuiplrnnW inniiin ribocpdor @IR) (B). The haerotetramaic hIR bris a molecuiar mus of450 kDa, rmd a mIacolar mass of 130 kDa fot iis asnbimit 'IhedinnntblRcrsbo~amolecolar~of-280 Wa,andamolearlarxuassofllO Wa fiir its a subunit. A s d amount of uncIcavcd recombinant ~icccptar (440 kDa) am be seen an the duced gel, which tcgreseats the uncleaved siugle chain containhg the a mbanit and the erdnctllurar domain of the p sub11111-t-
ot HIR
: of HlRa


-116 a subunit
4 7
49
4 p ' subunit
Fig. 24. Aftinity p- afexpmreâ tüRatnccUular domah with mti-hlR monoclonal antibody (MASI) columa. A) The purification profile manitami by radtradtcmctive tracer of *=I labeled bovine insalin- E x p d insulin receptor extraceilular domain was eluted with 2.5 M MgCI2. B) Purifiecl recombinant hiRexmceiluiar domain was d y z e d by SDS-PAGE underredacimg conditio11 The geIs were stalned either with silver or Coomassie blrie- Tmœ amotint ofimcleaved ap' single chah receptors and bovine albumin can be seen ni silver stained gel.


O 20 40 60 80 100
lime ( 0 min, O sec)
Fig. 2-8. Time caursc of photdyris. Radioactk IabeIed insuiin photoprobe was incubaîed with hIR in bïnding b-er in the da& at 4 4 for 20 hours, foiiowd by nporiirr to UV Iighî (100 Watts) for 5,10,15,20,40,60,80 and 100 se&, or witha 365 nm filter fbr 5, IO, 20,40,60, 80 and 100 minates- Higher cross-iinking eflilciency and lcss protein damage were obstrved with 365 nmfilter,

seaaed fUy procesd ectodomain of HIR accumdated in the culture media, many
mpmxmd precutsors accumulated in the piasma membrane (data not shown). The
secreîed recombinant receptor was mgniPd by human aur~immune anti-HIR antibody
and monoc10nai anti-HIR anti-es Vig. 2-5). More t h 90% of secmted protein was
shown to have a fbiiy processed a2Ss2 heterotetramer structure (two - 110 kDa a
subunits and two -30 kDa truncaîed $' submits), A smali amouut of protein was secreted
as non-cleaved (ap'h dimer (two -140 kDa af3' monomcrs) (Fig. 2-6). The yield of
expression was - 1 mgfiter of culture media as meesursd by immunoassay (data not
shown). The affjnïty p d e d receptor binds iasulin with a high afflnity @Cd = 1 x IO-'
M) which is oomparabie wîth that of wild-type human placenta IR (Fig. 2-1). This
receptor exhr'bits a linear Scatchad plot.
3.2. A S m d Labeled Peptide Fragment on the Insulia Binding Region
To optimirc photolysis 1 tested m i n t photolabeling conditions. With~ut the
filter, photolysis occ& very fast in seconds. Protein demage could be seen with
exposwes longer than 20 seconds. mth a narrow band optical nIter et 365 which
eliminates most short-wave UV light, the cross-lllikiag efficiency was i n c d 2-fold
and the protein darnages resulting mostly h m short wave W light, was reduced (Fig. 2-
8)-
In a preliminary sady, both the cleavable (rq-AZAP-insulin) and the
uncleavable (829-rq-MABI) photopmbes were used to characterize the pmteolyzed
peptide containhg the putative insulh binding site. In the case of the cleavable

photoprobe, k s u b B chah shouid k cleaved a£€er photolysis, and oniy radioactive
iodine left cross-- to the mxptor. A single radioactive bard containhg no attached
insulin B chah appead aRer dpeptidase digestÎ011, In the case of the other
photoprobes, shce since B chah has haso iodinabion sites, Te16 and T-26, bwo
radioactive hgments should be generaîed in the enzyme digests. A small 1-3 kDa
radioactive iabeled fhgment was gaierated by either trypsh or V8 m e n t of HIR
Iabeled with B29-Cq-MABI (Eig. 2-9). h the case of ELIR labeled with m-AZAP-
insulin trypsin digestion generafed a single small labeled peptide. Most of the digestion
o c c d in the fk t few ho- but wmplete digestion took 20 hom. Similar results were
obtained when using endopeptidase Lys-C digestion. CNBr trratment failed to
completely cleave the labeled receptor (Aatsi not shown).
Antibody against the B chia of insulin was shown to reco- with high affinity
the intact insulin B chain or its tryptic digest at pmole levels (Fig. 2-10). However, when
the same method was used to detect hrigments in HPLC Grattions, the sensitivity of
detection decreased several thousand fol& To obtain enough sensitivity to recognize the
peptide(s) ofinterest in HPLC fkactions, 1 cross-Linked the peptides in the HPLC fiactions
to ELISA plates instead of using the conventional coaîing method. The conventional
sample preparation method showed a dramatic drop in the semitivity of ELISA analysis
due to the very low pH (-pH 2) and high concentration of organic solvent (-30-70% of
acetonitile). The modified method wuld detect the peptide containing B chah hgments
in HPLC hctions (Fig. 2-10) or in digests of receptor labeled with B29-MABI (Fig. 2-
1 1) at the pmole level.

0 2 0 2 0 TTmm (hrs)
Fig. 2-9. Time course and products attrypsin digestion dB29-MABL or AZAP iabckd MR -cellular domain- Panels A d B show that trypsm digestion starts wïdiin an hom and is completed in 20 hours. Complete digestion generstes ladeied @pan& smriller tbrin 3 Id)a (A and C) wïîh both B29- MABI a d AZAP photopmbcs, niE 45% alLaline gel analpis shows that BW-MABI labeling genenites two radiolabeled bgments by aypsin digestion, SnSIng k m thepresence oftwo radioactive iodhe label- h g sites on the msPtm B chah One o f t k labelai ftrrgmats contaïus the patative insulln-bhdhg sité, Ih the case of AZAP photoprobe labehg, since radioactive iodirr labelcd on the cross-lnik gnnip instcad of the insulin B &ah, wbich is c i e a . after photolabebg, a single radolakled rccqtor hgpcnt is seen on the gel @)- Bovine insrilin and the iasulia B chab insulin wen Imded on the nght Iancs of the gel as a control (B),

Fig. 2-1 0. The sensithity of ELISA in detccting B chah fhgmcnt of bovine InniUn More (a and d) and 8ftœ HPLC Pnrlysis @, G 0 and f). Adramatically reduœd sensi* @, c) was obsaved undcr the acidic c œ ï ~ o n m tk HPLC ~~ when asmg regiilarpl&es- Using Reacti-Bind plates merce) restarrs the sensitivity &ELISA to pgkcU 1-1 (e, f), Synthetic B22-30 hgmm d the insulin B chain was used as s t d d ma, b, d and e. Trgptic digests of the insulin B chain containhg the f i a m dB1-21 and B22-30 ~~ wne d a s HPLC sa?npIe.


Fio. 2-1 2. Achromobe pcotcmc 1 digests dla5 1-BBp~Inmlia Wcd hIBoL A) HPLC B) An aliqiiot (10 ml) o f d hcticm was assa- by ELISA ushg a h h c pbosphatasmxmjugaod streptavidin, C) Raâïoadvityofeaeh~(0.5 ml) was de6ermmed Oae SbcptaViddin-positnrepeiiL a n d t w o r s d i & e ~ ~ ~ B a s e d a r t h e E L I S A r e s a l m , ~ - 74(mwherdimnA) aud 75 wem paoled and d y z c â again by HPLC.

pmok 0.2
Fig. 2-1 3 HPLC rndysia ofpooled tractions as iQiidilled h Eîgi 2-12 (A). A) HPLC profile obtainaî using a mobiîe phase dincrenthm tbt used to obtam the prome shown in Fis 2-12- B) An riliqaot (10 ml) of cach M o n was assayed by ELISA ushg altcaline phosphaîaseumjugated streptavidk The ~tre~tavi~pos i t i ivc M o n s showed very poor rtcovexy.

In the case of HLR labeled with BB~insuIin, 1 iiscd bot6 th antibody and thc
streptavidinmahod of detection. Radi08CfÉYity was also used inthe dcttction when
fadlfadloactive photoprobes were uscd, A typicai HPLC profile is show in Fig.2-12. Ibc
fiaction containing the peptiCde o f interest was nniner p&ed by HPLC under diffkrent
conditions, resulting in a great l o s of the peptide (Fig. 2-13). I was d l e to coiiect a
sunicient quantity ofthe peptide for amino acid seqyencing.
4. DISCUSSION
The bacuiovinis expression system @ES) bas ban succcssfiilly useâ to express
many fùnctional proteins &uckow and Summers, 1986). It is a very powerfid eukaryotic
protein expression system, and it is -or to bacterial, yeast and mammalian
expression systems for many reasons. Most recomb'i t pmteins fcom BES are soluble
and fully h c t i o d . hsect ceiis used in the protein expression many ofthe protein
modification, processing and transport systems present in higher eukaryotic ceiis that are
essential for fbll biological actîvity ofthe recombinant protein. Like 0 t h investigators, 1
was not able to express soluble intact HIR in M a or yeast (unpublished dm).
Mammalian cell systems, on the other han& usuaiïy express Iimited puantities of
recombinant protek. My prehinary study and the results h m other imrestigators
suggest that the extraceiïular domain of the HIR expssed h m BES main- full
binding aff?nity for its ligand @ad et al., 1990; Sissom and Ellis, 1989).
In this study, I have successfûlly used the baculovuUS system to overproduce the
truncated, soluble, secretory ectodomain of HIR which reEains fuli insulin-binding
afEity cornparrd to native placenta insuiin receptor (Fig. 2-8). Whik a dsf icmry

quantity (a few mgfiter) was s e d m the media with full fiaidon, a sigdîcant
amount of uucIeaved av precursor accumulated mside the ceU @ig. 2-9, Otha
investigators (Paul et ai., 1990; Sissom and Ellis, 1989) have previously reported the
expression of similar versions of the ectodomains of HIR in the bacuiovirus -*on
system, and mlmimalian expresion system (Cosgrove et aï., 1995). The molecular mas
of the a subunit expresd in the baculovinis system is d e r (1 10 B a versus 130 kDa
for the its native cotmterpert), likely due to d i f f i giycosylation dirruig the pst-
translation process @ad a al., 1990; Sissom and Ellis, 1989). Nevertheles, the
carbohydrae content of the receptor has been demoIlSfrZIted to have no effèct on the
ligand-receptor interaction (Caro et al., 1994).
S e v d investigators have reported that the sccraed HIR ectodomrrin pduced by
BES &'bits a hear Scatchard plot (one c h of hi&-afEnity site only) whereas the
native membrane HIR exhibits a curviiinear plot, suggesting one high-affinity and one
low-atFinity site. Interestingiy, the Scatchard plot obtained with the secmted HIR
ectodomain expressed in ' celis was curvilinear (Whittaker and Okamoto,
1988). This difference in the Iigand-binding behavior is not understood. It might d t
fiom the different pmcessing of the heterotetilamer in the d i f f i t expression systems.
AIternaîively, the relative abundance of the uncleaved prorecepfor produced by the insect
cell may e t the binding of a second iasulin m o l d e . Tt is hown that the af! half
receptor exhibits a linear Scatchard plot, but with low affinity for iasulùl (Sweet et al.,
1997). The fact th& only the heterotetrarner stnictute of the HIR can bind insulin with
high afnnty suggests that high afnnity hsuihbinding sites require both halves of the
receptor to participate. Insulin itselfhas no obvious intdnsic symmetry but must interact

with two identicaï a$-receptor halves for hi& affiaitp K i - 1 beiieve thet mat may
exisc identicai but more than one contact site in each a subunit of IR Hi@ afIinity
insului bi~lding Fepairrs contacting diBiirent sites on each half of the tecepfor. At low
physiologid insuijn concentrations, hmib can contact Ïts biading sites on both a
subunits for hi@-affinity binding- At higher concentrations of hsuiin, a second insulin
m o l d e can ody- contact some of the sites requïred for high afnnlty binding, -l&g to
Iow aff?nity biading. The fhct that the IR b i i one insulin with high efnmty and the
second with low affinity suggests that asymmetry may k induceci in the receptor by
insulin binding. It also suggests that one insulin moIecule biadmg to the recep&~ with
high affinity e i t k leads to a conforrnatioI181 change of the receptor, or simply one
insulin ocaipies the space dowing for high atnnity biiding making it dBicult to bind
the second insulin. The d i f f i t pst-transIational processes in the bacdovirus
expression system may lead to some c o d o d o n a l varhtion in the recombinant
receptor to allow either one insuiin molecuie oniy to bind or two insulin molecules to
bind with similar af£ïnity.
M t y labeling of the receptor with photoreactive insulin derivatives followed
by isolation of ~eceptor-inmlin fkagments has ailowed the identification of regions of the
a subunit as potential hormonerecepf~r contact sites. S e v d regions have beai
suggested by vaxhs investigators (Wedekind et ai., 1989; W b et al., 1989; Yip et ai.,
1988; Fabray et al., 1992). Most ofthe studies redted in relatively large hgments ( k m
14-55 kDa), making it diffIcuit to pinpoint accurately the contacting location. Resuits
obtained in the present study indicate tbat a very small -ent (-1-3 LDa) containai
the insuiin biidhg site, or part of the bhding site on the a subu& ResUlts obtained h m

HPLC -de mapphg based onthehighly d e detecthg metho& suggesteâ that it
may be possible to identifjr this hgment by it9 amino acid sequence if dficient
qyantities of the peptide could be obtained, ûn the other hand, d e r peptides are more
dScult to isolate and idenMy. West+m blots used by many investigators to daect their
larger receptor fragments, would be difncuit to use to isolate such a smd peptide for
sequencing. Reversephase HPLC adysis has been succediliy used to isoIate a s d
peptide &action h m otherpeptiCdesdes However. the low photoIabeIing efficlency makes it
difficuit to load enough protein digests on the column to yield a d c i e n t q d î y in the
Graction wntaininp the labeled fiagrnent Iae large percentage of non-productive, non-
labe1ed receptor also contri'butes to the di86cuity of purification. It will require a second
or third re-pudication of the HPLC -011s to obtain a pun hgment for amho acid
sequence anaiysis. The cuxrent amino acid sequencing method aiso requirts up to ten
times more to have reasonabIe d t s for a smaller peptide compared to larger
polypeptides. Since most photoaffkity probes have low cross-linking efficiency (cl-
IO%), either more starting material or a probe with higher efficiency is required to isolate
enough m e n t AIteniatively, a higher efficiency purification method or a more
sensitive amino acïd sequencing method n&ds to be developad The new BBPa-insuiin
photoprobe with bïgher cross-linlo'ng efficiency has shed new light on this project
aithough we failed to achieve the efficiency claimed by the onginai investigators @ee et
al., 1993). Unfortwiately, the new BBPa-insuiin photoprobe we obtained was only
dEcient for developing the method01ogy. It is clear that more wodr would need to be
done to overcome the obstacles in this biochemical approach,

QUATERNARY 3D STRUCTURE OF THE INSULIN- INSULIN RECEPTOR COMPLEX DETERMINED BY
SCANMNG TRANSMTSSTON ELECTRON MICROSCOPE: STRUCTURAL LOCALIZATION OF
INSULIN-BINDING SITE ON THE HIR
1. INTRODUCTION
Rotein-protein intcractons et the molecular Ievel are largely reletcd to th&
three-dimensional (3-D) structures Elucidation of 3-D structures of proteins at high
resolution is cruciai to the understanding of many fimdamental fimctions of
macromolecules and macfomoldar complexes. Besides its significance in
understanding the chemisiry of Mie, the d n e d stmctme o f a protein also provides highly
valuable infîodon for medical appkations, such as new drug design (Gisanda a al.,
The most commonly used methods for solWig macromolecuiar stnicturr are x-ray
crystallography, nuclear magnetic resomnce (NMR) spectroscopy and electron
microscopy (EM). While the 3-D stmcturt of a great number of proteins has been
determineci by x-ray crystallography, if they crystaljize, and by NMR spectroscopy, if
they are sdicientiy small, the vast majority of biologïcai 1118~10molecuies have not been
crystallized or are too large for these appmaches. X-ray crystailography has been
successfÛUy employed to determine 3-D structure at a nsohrtion of 1.5 - 3 k However,
due to the di86culty in growing crystals of proteins suitable for x-ray crysnillography,
most proteins whose 3-D stnwtme have been sohred by this techaique are d e r than
100 kDa Using NMR spectroscopy, investigaiors can solve the struchne of a protcin at

atomic resolution but the molecde must be wen d e r , around 25 - 30 kDa, and hlghly
purined. Howevr, electron microscopy, combined with cornputer pmcessing algorithms,
has successfirlly salved the 3-D structure of macromolecules with molecdar weight Grom
several to several thousand kDa at A to nm level (ûttensmeyer, 1982; Schrag et al., 1989;
Chiu and Schmid, 1997; Rosenbag et al., 1997). This technique offm a powedul hîgh-
resolution tool in structural biology, and Ïs especially useful for large protein molecules
that cannot be studied by either x-ray crystallography or NMR method, The EM-based
3D reconstruction enabIes investigators to solve the molecular architecture of proteins,
and to detennine the overali size and shape of th& constituent protein subunits with
atomic details. In addition, this technique ais0 allows researchers to investigate distinct
fbctional States by monitoring conformational changes or structural reorganizations
(Whittaker et al., 1995; Jontes et al., 1995; Unwin N, 1995, 1998)- Such studies have
provided insight into the structure and fünction of many proteins, including virus
structure and mocphology (Shaw et al., 1993; Zhou et al., 1994). localization of
individual protein components (Zhou et al, 1995), identification of cellular receptors
(Olson et al., 1993) or monoclonal antibody binding sites (Wang et al., 1994; Chiu and
Smith, 1994) and protein-protein interactions (8eckmann et al., 1997).
The electron microscope provides much higher magnincation than the iight
microscope by the combination of magnetic lenses and short wavelength electmns at high
voltage. However, compared to the case in materiai science, biological specimens are
more diffi.cult to image in the electron microscope because of their low contrast and their
sensitivity to radiation. Much effort has been made in the Iast few decades to make EM
more useful to biology. High contrast heavy atoms (osmium, lead, uranium, etc) are

uçuaILy required to stain or coat the bioIogical sampfes in a controlled way to make them
become "visible". However, the object being imaged is acbially an "ertif&ct'' cornposed of
the distn'bution of heavy atoms suuoming the features of hiterest.
Scannin~EIecfrOn M ~ ~ O S C O D ~ One of the collzmody used eiectron
microscopes in biology is the scanning electron microscope (SEM). SEM images arz
obtained by using a very narrow electron beam to scan over the surface of the specimens
and by mapping the signal of emitted secondary efectrons (or of back-scaîtered electrons)
into the corresponding pixel of the recording/display image (Fig. 3-1). Although SEM
provides much higher resolution and a much greater depth of field than the light
microscope, biological specimens usuaiiy need to be coated with a conducting layer of
heavy atoms to avoid charging under the scanning electron beam. This approach limits
the detailed interpretation of smali structures and provides only the surfie feature of the
specimen. In addition, although improvernents are being made, the resolution commody
at about 20 nm is also not high enough to reveal detail in individuai protein structures,
which usudy requires resolution at nrn or sub-nm level. SEM has been mostly used in
the studies of organ or tissue structure in biomedical research.
Transmissio~ Etectron Microsco~e and Scanninn Transmission Electron
Microsco~e Compared with the SEM, the transmission electron microscope (TEM) is a
more powerfiil tool available for the investigation of the microstructure of bio-
macromolecules (Fig. 3-1)- The TEM is somewhat similar to the SEM in that both use a
hi&-energy beam of electrons and a series of magnetic lensa to control the beam.
However, the electron beam voltage is much higher (LOO KeV or pater; shorter
wavelength) in the TEM, and the high energy beam passing thmugh the specimen is used

Fig. 3-1. Dienmt ckctmn miemscqm Prcd ia biomedid rtudier, Tbe samning electron microscope (SEM) is maïdy used to saidy the strrictme oforgan or tissue at relative low tesolution The transmission electron ~ÏQOSCOP~ (TEW is usai to investigate the micro- structure ofbioniolecriles. The scanning aansmissioa elatrm mïczos~ope (STEM) camb' ï the ahmages ofboth SEM and TEM and is more efficient in the use of scat&& elecbons, and CO- can reduce specimcn damage STEM has beai uscd fbr high resolution 3D reconsbniction of profein ~lcculcs.

to form a direct image. A magoi6ication of about 500,000 x is typical and the resolution
can srpproach L mn or bet-
Within the last few years, tremendous pmgress in EM technology and computer
image processihg have been made. The introductfon of low-dose and cryo-presemation
technipes into electron microscopy has opened a new era for 3-D structure
determination of biomacromolecules, Cryo-fixation, ice-embedding or fkze-drying the
protein sample in its physiological b&i7 permit d k t Miaging of the protein itseif m a
near-physiological state. It has been widely accepted that these methods reduce
adsorption artifacts and reproduce structural details more accurately and faithfdly- Most
importantly9 in addition to mapping out the overaiL size and shape ofbiomacrornolecules,
cryo-transmission electron microsmpy, and cryo-scanning transmission electron
microscopy (STEM) enables investigators to look "uiside" biomacromolecules and is
beginning to image theV secondary structural elements, or even thw atomic detaüs. The
3-D EM structural information can, under some conditions, be obtained at a resolution of
as good as severai hgstroms9 sunilar to the resolution in X-ray crystallography, to
directly reveal the a-hek structural details of snme proteins @ü.hlbrandt et al., 1994;
Cheng et al., 1997; Walz et al., 1997). Conseq~ently~ more researchers are beginning to
use electron cryo-microscopy for structural and fiuictional studies of bÏomacromoIecuieses
The 3-D image reconstruction of EM has become a new tool and a very powerfùi
approach in biomedical research.
The uniqueness of the STEM is its ability to visualize individual biological
molecules directly without staining, fixing or shad~wing~ Ih the STEM, image acquisition
is digitized as a small electron probe (3 A beam size in the HB601UX STEM), produced

by a cold-field emiemissÎon gun, a condenser and objective lem, sans in raster fasbion
across the specUnen* The microscope operates in a dark-field mode with high efficiency
amular detectors coilecting virtuaily aU the scattered electrons. The high efficiency of
STEM pennits da&-field images to be obtained d g Iower radiation doses (ten to
hundreds times less than by dark-field imaging in 'ïEl& resuitïng in less damage to
protein structures)- The number of scattered electrons received by the detector is directiy
proportional to the mass thickness ofthe scanned molecules, Thus the molecular mass of
the individual molecule c m be caIcuIated by subtracting the background and using a
scattering cross-section or cali'bration standard- Another unique feature of STEM is that
individual heavy atoms can be visualized and thus specinc sites on biologicd molecules
can be mapped @aidielcl., 1992). Recently, a monofimctional reactive gold cluster (1.4
nm in size), Nanogold, has been successfùliy used to localize specinc fùnctionai sites
with STEM in several proteins (Boisset et al-, 1992; Wilkinson et al., 1994). With low-
dose electron beam scanning, high contrast dark-field imaging and digitally recordhg
features, STEM can collect hi&-resolution images fiom a specirnen as s m d as a single
heavy atom to proteins larger than a thousand kDa. Combined with novel computer
processuig techniques developed for the microscope, STEM has beeo successfüily used
to reconstruct 3-D structures of several proteins, such as SRP54 and Klenow fitapent of
DNA polymerase 1 (Farrow and Ottemmeyer, 1992; 1993; Czarnota et al., 1994) at a
resolution of 12 - 20 A.
1.1. Electron Microscopie 3-D ReeoutrPction Techniques
3-D electron cryomicroscopy of biological macromolecule consists of several

l!lllh 111 STEM El.cboir B..m
A IIR
Specimen Pmparation &i Digital lmage Collection
Cornputer Image Processing
30 structure visualkation and intwpretation
Fig. 3-2. An ovemew of STEM 3D reconstruction Single particles ofthe insulin/insulin receptor cornplex, represented as a han4 were subjected to STEU, Images were collected and analyzed and processed by cornputer involving (A) background subdon, (8) low pass filtering, and (C) alignment F m e r processed images were back-pmjected and displayed as a 3D structure.

major steps: specimen pqmdon;. image coiiectio11, nomielization and pn>assiag; and
3-D reconstnrtion and -*on @ig- 3-2.). Briefly, msny large images of di-
single molecules are coiiected and placed in a cornputer file with himdrods or thousamis
of individual particles. The individuai images then undergo twodbensional(2D) image
dysis , including variauce nonnalization, backgn,und subtraction, low band pas
filtering, as well as statisticai d y s i s . The pmcesed 2.D images thm are d y for 3D
image processingins The backgmumi has been completely removed d&g ?bis pn>cess. A
sinogram, a stack of one-dimensional projections of the 2-D iinage rotated 3609 amund
its Y axis, of each singîe particle is dculated and is used to reiate the orientation of one
image to any other. A f k the relative angular orientation of tach image is calCUI8ted, all
images are back-ected to produce the final 3-D reconstru~*on.
A number of recent developments in wmputational techniqys and experimentai
approaches have greatly improved this pnness and for speciLai cases, increased the
resolution of the electron microscopy reconstruction h m the early 30 A (Dryden et al,
1993) to a few ~ngstroms (Bottcher et al., 1997; TIUS a al., 1997). One of th
approaches, which greatly increases the structure resolution in 3-D feconstruction, is the
use of 2-D electron crystallogmphy. In Tii methd each image contains thousands of
identical units in identical orientations, wbich can be accinately and easily averagd As a
consequence, structurai analysis by electmn cryomicroscopy of 2-D crystais can extend
the resolution to th atomic level. For example, by using this technique, the structurai
resolution of bacteriorhodopsin with a redution approaching 3.5 A peLLlZits visualization
of a-helical bundles in this membrane protein @enderson et al., 1990). Anothet
technique, hown as the random c o n i d tilt method, is able to use samples in which

molecules assmne one, or a few. preferred orienmCons so that-a hi* resoIubLon cau be
attained @&macher et al., 1992; Serysheva et al., 1995). Ho-, tb technicpe is
Iimited by the reqpimnent that the specimen adopt recognizable p r e f d on-ons
withrespectto the supportnIm.
NoveL rnethods *ch do not nquirr crystauization, shuchaal symmetry or
preferred orientations have ban developed and iinplemented as weil, and used in om
work (van Heel, 1987; Ottensmeyer and Farrow, 1992; F-w and Ottaismeyer, 1993;
ûttensmeyer et al, 1994). This approach was designed for random orientations of non-
symmetric molecdes, and maLes use of qmemîon mathematics to sdve the stnicbat of
biomacromoIecules even h m noisy images of single particles. An advantage of tbis
method is that it pemiits stnietural detenninations of macnrmolecuIes under many
different conditions incIuding diffgicnt ionic envbnments, molecule s b or pst-
translational modifications, wbich ovacome many limits in crystallization or pre-
requirement in sample p r e p a d o ~ ~ Recently, a signal stqyence binding protein has been
successfiilly resolved to its 3-D at 12 A res01irtion with this technique (Czarnota
et al., 1994)-
1.2. Strilcturaï Determination of IRand Loaliutbn of riwniin Bindîng Site
Insuiin binding to the ce11 siiraice IR activaies its in%c cytoplasdc tyrosine
kinase through the autophosphorylation of specïfïc tyrosine midues, initiatUig the
intraceUular signal transàuction cascade, wbich is essential to insulin actions such as
glucose homeosmis, protein syntbesis, p w t b a d development (Taylor, 1991)-
However, IR IR not been crystallized, probably due in pazt to its molecular ma98 of 450
88

kDa, M g a membrane protein, eild being highiy glywsylaîed. A f k more than 25 yëars
of intensive stuâies by many Investi*gators, the quatcmey sinidure of IR and the
stmcaaal basis for the mechemsm of IR activadion by b s u h binding remains u n l c l M a It
is known that insulin binds to the extrace11ular a subunit of the whole receptor, and also
with unaltered afflnity to the isolated ectodomain (Schaefer et al., 1990). Several large
regions of the ectodomain have been impLicafPA in binding hdh, but a more detailed
description of the bindhg site is not yet maiiab1e becarise the crysîai structure ofneither
IR nor its ectodomain is biown. On the other hend, for an anaiogous feceptor, IGF-IR,
the crystai structure of a m e n t containhg the first three domains &l-Cy~-ric~L2)
involved in ligand binding has been solved (Garrett et al., 1998). IGF-IR and IR are
closely rdated members of the tyrosine kinase superfamiiy and highly homologous in
their primary stmctwe (ülirïch et al., 1986). The structure of the ment, which Iacks
Ligand-binding activity, shows that the tbree domains form a centrai notch sdïcient for
ligand binding. In addition, the structure of the hgment of the intracellular tyrosine
kinase domain of IR has been detemUned by x-ray crydography, both in the
unphosphoryiated and phosphorylated state (Rubbard et ai., 1994; Hubbard, 1997).
Since the IR is a mode1 protein in signal transduction system, and is involved in
basic metabolism, in development and in many human diseases, solving L structure is
very important, Since detesminiig IR by x-ray crystalîogxaphy or by NMR analysis hes
proven ciifficuit, electron cryomicroscopy seems to be a possible alternative appmach to
solve this problem. As mentioned before, the large molecular s h and the high degree of
gIycosyIaîion of IR should not present a problem in STEM d y s i s . Another advantage
of STEM is the visualization of heavy metal atoms, such as the 70 atom gold cluster,

Nanogold This havy metaI marlrer can be risad to map specinc hctionai sites on
protein moIecules shce it will appear microscopidy as siilgle "spots" of about 1.4 mn
in diameter m e I d and Furuya, 1992). If insulin is labeled with NanogoId, this iiisuliri
derivative can then be used to Iocate the hsu&binâing site on the IR with electron
cryomicroscopy. In addition, available structural information on the TK domain of IR
and the L1-CM2 domain of IGF 1 receptor (Habbard et ai., 1994; Garrett et al., 1997;
Baron et al., 1992) couid be helpfirl in analyzÏng the results obtained fiom STEM sndy
ofthe whole recepfOr.
In this Chapter, 1 descfl'be the result h m experiments d e d outto daamùie the
3-D qusteniary structure ofthe human insuün receptor by 3-D reconstruction fFom STEM
images of singie molemles. Nanogold-bovine insuiin as an EM market was pqared and
used to localize the insulin-binding site in the structure of the intact insulin-insulin
receptor cornplex. Results describeci in this Chapter have been published in abbreviated
form in Science (Lu0 et al., 1999).
2.1. Biological Materials
Preparation of Nanoeold Bode Insuiin =BE)
Nanogold @Janopmbes, Stony Brook, NY) is a newly developed gold duster
label, prepared by using a discrete gold cornpoumi rather thaii the traditional coiloid
Nanogold is an exîremely U13iform 1.4 nm diameter gold clusta particle wntaining
appmximately 70 gold atoms with a molecular mass of 15 kDa This heavy metaï particle

BOC 81 DBI NO NO-DBI NG-BI

can be easily sccn as a bright baU shape spot unàer the electmn at a
magnification firtor higher than 200.000 r The sulfo-whydrov-do (NEIS)
NmogoId vig.3-3a) used to label bovine insulin has a monocreactive p u p which is able
to cross-iink to an accessible reacthe amine group on a protein,
An iosuün moIecule contams three reactive amine groups, one each at -*due,
Al, B I and B29. Since the amino termiripl of the B chain (BI) is known not to be
involveci in binding to the receptor (Nakagaw8 and Taga, 1986, 1987. 1993; Schaffer,
1994) the amino group of PheI was derivatïzed with a gold cluster. This was done using
bis-(t-Bot)-bovine irisului @BI) in which the d o group of Al and B29 are protected.
DBI was prepared in Dr. Yip's laboratory (Yetmg et al., 1980). One mg of DBI (-150
nmole) and 30 nmole of mono-NHS-Nanogold were dissolved in 200 pi of f-
dimethylacetarnide @MA) containing 2 pl di-iso-propylethylamine PIPEA), pH 7.6-
nie mixture was vigorously mixed at room temperatme for 60 minutes thm dned in a
SpeedVac (Savant Instruments). The pellet obtained was dissolved in 20 pl of
tci£iuoroacetic a d , kept et m m taperahue for 5 minutes, and then dried in the
SpeedVac. The peliet was washed twïce with 100 pi of 1 M acetk acid, resuspended in
120 pl of 1 M acetic acid and cbmatographed on a Bi&l P-10 column (1.7 x 25 cm)
in 1 M acetic ad. Fractions of 0.5 ml were collected, and their absorbency at 280 n m
was measrned. An aliquot of each M o n was subjected to electrophonsis in 10-200h
SDS-tricine gel in tricine/glycine b u f k (pH 8.8). The fiactions containhg NGBL (Fig. 3-
3b) wem pooled and rechromatographed on the Bio-Gel P-10 colimui. The purifiecl
NGBI was storeci at a concentration of 0.5 pmoYml ai -700 C.
Preyrition of Nia0~01d-ib~~lWiiimlh ~ C C C D ~ ~ CO-

Insulin receptor protem @IR) was solubilized h m human placenta membranes
and pmffied by afnmty cinomatography on an iiwlin co11m.n followed by M e r FPLC
purification on a Sephacryl S-200 HR column (1.6 x 60 cm, Phannacïa FPLC set and
column) as d e s c n i in Chapter 2. The p d e d HIR was immedïately quick fiozen in
liquid nitrogen in aliquots of 20 pl at a protein concentration of 20 pg/jd and kept at -
70°C mtil needd For gold labehg, EIIR was incubated at 4OC oveniight in 20 mM
=ES buffa @H 7.5) with NGBI (final concentration of - 0.5 x 1 0 ~ M) at a mdar
ratio of insulin= of - 10:l- Free NGBI was removed h m the mixture by
d ~ t r a t i o n through a microfilter with a cut-off of 300 kDa (Sigma) aAer incubation-
The NGBI-hIR cornplex was then diluted to 7.5 pg of receptor proteintml with 20 mM
HEPES buffer, pH 7.5, prior to loading on the microscope grid support. The presence of
one NGBI per one hlR in the majorïty of the complex particles was confirmeci by
preliminary TEM and STEM imaging before loading the specimen for the final STEM
analysis.
2.2. Electron Microscopy and Image Anaiysis
Pre~aration of s~ecimen for STEM
Copper grids (300 mesh) coated with a holey plastic nIm (pore size 5-10 p)
overlaid with a carbon film 23 A thick were used to support the protek in both TEM and
STEM analysis. The thin carbon film was prepared by high vacuum electron gun
evaporation of carbon onto h h l y cleaved mica The thin carbon nIm was then floated
on a dish of double distilleci, de-ionized water that had been passed through a 0.2 p m

mUïpore £iI= The tfiin carbon film wis picked up with the piastic coated gnd and th
grid was air*-ed.
To determine the optimal concentration of protein for STEM analysis, 20,10,7.5,
5, and 2.5 nfil of HIRdiluted with 20 mM HEPES, pH 7.5 b e , were testad in
902 TEM using electmn energy l o s dadceld condition at the magnifïcation fhctor of
20,000 r The specimen (5 pl) was injected into 5 pl ofdilution M e r preloaded on the
carbon film of the grid. The specimen was washed with 3 drops of 20 mM, pH 7.5
HEPES buffg and 1 drop o f 10 mM ammonia acetaie bu@- eH7.5). For purposes of
concentration optimizaton, the grid was Ieft to air dry. The concentration of75 ng/ jd of
HIR was then chosen for the specimen concentration for STEM study.
For STEM anaiysis, the same sample preparation was perfomed, except that a&r
washing, the grid was draineci by wicking with Whatmam No. I £ilter paper to luive a
very thin layer of solution, as o b d with an opt id microscope. The sample grid was
immediately quick-hzen by plungïng the Md into liquid ethane at -150" C. Quick-
fÏeezing to liqud nitmgen temperature assud immobilization of the molecules in a solid
glas-like environment similar to the aqueous state. The hzen specimen was trmderred
at liqyid nitrogen temperatme into the STEM (Vacuum Generators, Mode1 HB601UX).
The specimen was fieeze-dried within the cold-stage holder of the STEM at -140 OC.
Freezedrybg at this temperatme resulted in a contrast-rich specimen that has not been
distoaed structuraUy by phase bomdary forces during dehydration.
Imane acauisition and anriysig
mes in a 480 x 480 pixel format were acqpbd in the STEM with the
specimen at -lSOS=, using culd-field anission at en acceleraîing voltage of 100 kV, a

dose of 6e/A2, and a magnïfication of 252,ûûû x, conespondnig to a pixel size of 65 k
The beam size was 3 A per pixel. Both melastic and elastk antlular dadc field s i w s
were detected and dïgitally recordeci Smidtaneously- The sharp Nanogold signal was
used to adjustthe stigmatism and focus on the images.
The paired electron images acquired h m the eIastic and inelastic detectors were
combined to increase the signal-to-noise ratio two-fold compared to the elastic signal
alone. To process molecular images: singie particles were ùiteractively selected in 64 x
64 pixel windows using the program WEB (Wadsworth Laboratones, Albany, NY). In
preparation for orientation determination the selected images were low-pass filteteci to 11
A using a Gaussian filter in the program SPIDER (\Kadsworth Laboratories). The
molecular mass was calculated from the seIected particle images in relation ta the mass of
the 23 A thick carbon support with a carbon density of 2.0 g/&. The particles had a
Gaussian mass distribution with a modal mass of 570 kDa, which inchded the mass of
480 kDa for HIR and NGBI plus the weight of an estimated 150 Triton X-100 molecules.
To detennine the molecde bomdary, particle images were CCgrown7' fkm a center high
density point in expanding contiguous contour levels until a global cut-off was reached
that corresponded to contiguous structures with the average mass. 704 Particles with
apparent structural integnty, appropnate relative molecuiar mass and showing the
presence of the high-density Nanogold spots were selected for M e r processing.
2.3. Tbree Dimensional Reconstruction
Iterative Ouaternaw-assisted Aneolar determination

Three-dimensionai teconstructions were. wmputed h m the mes of randomly
oriented RIR/NGBI: cornplex particles ushg Iteratbe Quatemary-assisted Angufar
Deteminafion VQAD) method @amw and Ottensmeyer, 1992; 1993a; 1993b; Czarnota
et al., 1994; 1997) based on skogram correlation and the common axk theorem (van
Heel, 1987; Crowther et al., 1970; Crowthe~~ 1971).
The IQAD approach to determinïng macromolecular structure permits the
determination of the tbree-di.-ol structure of a biological macromolede by
cdculating a poste no^ the anguiar relationship of a biological macn>molecule in a
particular image with respect to many other images of an identical macromolecule at
different random orientations. Briefly, in Fourier space any 2D Fourier transforms
(fiequency space representations) of corresponnding 2D projections of a 3D density
distribution will be centrai sections of the 3D Fourier transfoun of the 3D density
distniution, and thus will share a cornmon line of intersection in 3D Fourier space. By
calculating the relative position of common lines to one another in a 3D coordinate
systern, the relative orientations of the images can be determine& The addition of IQAD
processing, sinograms, sinogram correlation fûnctions and qyaternion mathematics to
o p e e the fitting togeheer makes it possible to use even sets of noisy image to obtain
the optimal orientation arnong them of any individual image @arrow and Onemneyer,
1992, 1993). 3D reconstnicctions then are performed by fI1tered back-projection using an
angular distniution-dependent filter. Calculations were carrieci out on an SGI Indigo
work station (Silicon Graphks hc.)-
Resolution measurements and results dis~tav

Mea~u~ements of resoIutïon for ~ns t ruc t ions were dculated by cornputmg 3D
Po- transforms for two independent n x o ~ * o n s and h m a set of haif of the
images. From these 3D transfonns the average differenca in phase between them is
calculated in radiai shelIs h m the center of each transfonn outwards. This phase
differences related to the correlation of StnicturaL detail at each shelî. Since the radius of
the sheiî corresponds to a resolution level (rager radius, better resolution) the loss of
correlation at a given radius mdicates the resolution E t -
The program IRIS EXPLORER 2-0 (Silicon Graphics hc) was used to display the
3D reconstructions at a chosen intensity-based contour levei, a threshold which
corresponds to the calculated volume of a 450 B a protein, assuming a protein density of -
1.3~/crn~. To show domah relationships and structural b, the collStNctions were
displayed with intermediate densities 5% to 10% higher than the average density of the
fùll volume.
Crystallographic modd or co-oduiates of LI-Cys-rich-L2 domains of the IGF-IR
(Garrett, et al., 1998), the tyrosine kinase domain of IR mubbard, et al., 1994, Hubbard,
1997; PDB: L I E X , 1IRp) and human nbronectin III repeats domain (Baron et al., 1992;
PDB: I W ) were fined into the reconstruction volume of the insulin-HIR complex by
modeling the electron density of the x-ray crystallographic structures into the volume of the
assignecl domains of the 3D remntruction using program INSIGHT II (Molecular
Simultations Inc.). Since the CO-ordinates of the first three domains of the IGF-1R were
not available, a cornputer draw 3D mode1 of approximate cylinders based on the x-ray
structure idormation in the publication of Garrett et al. (1998) was used instead.

3. RESULTS
3.1. Specimen Preparatioa for ElQfffOn Mfcrooropy
The purity of the receptor protein @IR) was found to be better than 95% as detennined
by sodium dodecyL solfate polyacryIamïde gel electrophoresis (Fig. 3-4). The punfied
HIR showed the same binding a£hity as placental membrane IR for insulin. (Fig. 3-4).
In the preparation of nanogold-bovine insullli, we carried out the reaction in organic
solvent instead of aqueous solvent as suggested by the manufacturerturer Our method
significantly increased the efficiency of derïvatization to greater than 70% as compared to
about 15% in aqueous condition (data not shown). The derivatkation and purification
processes wae monitored by SDS-PAGE and acid-urea gel electrophoresis (Fig. 3-5).
The purifÏed NG-BI was found to be more than 95% pure and to have a molecular mass
of 19796 Da by manix-assisted laser desoxption/iozation time-of-fîight (MALDI-TOF)
mass spectrometry Pig. 3-9, a mass consistent with one insulin molecule having been
derivatized with one Nanogold cluster. Compared with native bovine insulin, NG-BI
showed only a slight decrease in binding affinity to human placental membrane insulin
receptor (Fig. 3-5). As a compact particle containhg about 70 gold atoms surromded by
an organic matrk, Nanogold was visible as a bEght ball shape spot in the STEM at a
magnincation higher than 200,000 x. The purifieci NGBI showed no visible damage in
the STEM at 2,000,000 x (Fig. 3-6). The insulin signal was too weak to display when
compared to Nanogold at the same time.
3.2. Image Processing of NGBI-HIR Cornplex

A SDS-PAGE of HIR
F ig. 3 4 Purification of hunirto placeataol insulin nxeptor @IR)* hIR protein was pmi- fied by aflFiniîy chromatography using an insulin-AIE-gel column, foiiowed by gel filtration A) SDS-PAGE of the pmified hIR, stained with Coomassie blue- Lam 1 shows a band corre- spondmg to the unredusedhIR of450 kDa- Lane2 shows two stained bands conespondkg
obtained showthai there was is no detet&abk diffei.ence in me binding &ty for bovine insu- ün between the purifieci hIR and thehIR in placenta membrane (HPPM hIR)-

Fig. 3 4 Purificatfan aad chmtccizatkm of bdll~gdd imuün (NGBr). A) Gel-fiIîratlon sepdm of the reaction mixture- Solid bars represent the OB- OfNGBL and fb NG, and open bars reptesent the OD- of i k inSnlin, B) SDS-PAGE in 10120% tricine gradient gel ofthe reactïon mixtint and hctions obtained in (A). Tbe gei was stained with Coomassie Blue, The staiued band seen in fhction 1 L shows that NGBI can be partdiy purineci fiom the reaction mixîure. C) MALDI-TOFF ana@s established that the NGBI obtaiiëed h m repeat geF nIiraîion was more than 90% pure, rmd that the mass of the NGBI was cansisteat with a stoichionietry of one Nanogold per one Uisulin molecule- D) Receptor- binding assay s h o d that NGBI binds to hIR wiîh a siightly redlKXd af6nity compared to nonderiva- tized nisalin.

Fig. 3 4 STEM images of puriîied NGBI at a msi&nification of2,000,000 X ( i i r t ) . The brïght m d images seen are signals h m Nanogold, Signals h m insplin wodd be very weak compared m*th Nanogold There is no obvious damage of Nanogold particfes as evident i2om their weii-round sphericai .chape.

To increase the si* to noise --O for a given dose, the simuitaneously recordai elastic
and inelastic detector signais of the STEM were combind A representative unprocesseâ
STEM datk-field image containhg a few NGBI-HIR complexes is shown in Fig. 3-7A
As indicated by arrows, obvious intense sigaals can be seen h m the Nanogold marka.
Randody orientated complex parficles have an average size of about 15 nm. Most of the
complexes exhibit one brïght spot (or two in less than 10% of the complexes) on the
particles, that can be seen directly or can be visualized by varying the threshold on the
displayed intensity. Figures 3-7B and 3-7C show that brÏght NGBI locations are even
more strikùig in the high-density threshold representations of the extracteci particles after
low pass filtering. When two NGBI particles are detected, they are in close proximity to
each other (Fîg. 3-7B, lower two iinages),
Figure 3-8 shows a group of selected complexes extracted into a circular rnask
h m the raw images. Figures 3-9 and 3-10 show two m e r processing steps for the
same group of particles: low pass filtering to 11 A using a Gaussian filter, and molecular
boundary determination. The latter images are ready for 3D image processing çuch as
orientation deteunination.
The relative molecuiar weights for approximately 1,600 particles, ushg the 23 A
carbon support film as a m a s standard, are shown in the histogram in Fig. 3-1 1. Only the
particles having a calculated molecular weight no greater than a difference of 18 % fiom
the average were chosen for reconstruction. The calculated average mode1 mass is about
570 kDa, which ïncludes the mass of 480 kDa for AIR and NGBI plus the weight for an
estimated 150 Triton X-100 molecules.

Fig. 3-7. STEM dark6ehï images of human insuün nccpttor bound to Nanogdd insulin @RNGBI)- A) A field of view showing severaï complexes- Anowheads point to the Nanogold markers, Scale bar = 20 nm- B) IRNGBI images extracted fbm Mage fields, low-pss filterd to 1.0 nm (lefi oolumn), and obtaiaed at high daisi*ty thrrshold (nght cd-) showing one or two Nanogold matkers indicating the binding of one or two Naaogold-insulin (Luo et al. Science 285:1077,1999),


Fig. 3-9. Low band-pasr nItercd images. The sime images as in Fig. 3-8 were low-pass fil- tend to 11 A ushg a Gussian filter and background was subs~raded to increase bu signal-to- noise ratio, Scale bar = 20 ma,

FIg. 34 0. Final images used for 3D recoustrucîion. To finthet r&e the images for 3D reconstruction, the ssme images as in Fig, 3-9 wece processeci using "grown" software to detennine the molecuie mass center and boundaqundarv Various shapes of the images were obtaMed which repre- sent the same molecule king vie& in different on-ans, The location of NGBI is seen as a bnghter spot on the image- After tbïs refinement, images were ready for 3D image processlng. Scale bar = 20 nm,
S.

mîaüve Molecular MISS
Fig. 3-1 1. Histogram of crilcuïatcâ mass for 1,625 coilectcâ images. llv nlrtive molecalar weights werc calcuiaaed using the canboa supportmg EiIm as a mas standard. The partides in the shndad area reptesent the ones having a calcalated rnoIccuk weight witbin 18% of the average. 704 IWNGBI particies k m the shaâedarea were chosen fbr reconstruction-


Fig. 3-13. 3D reconstruction of the IR/NGBI complu (surhce repfesentation). A) Density threshold representing the tutal expexted volume for the complex (AL); intermediate density tbreshold, unsymme- trized (A2); high density threshold of (A2) showing only the Nawgold label (A3). Circles indiate loca- tion of the gold marker within the ~ ~ t i o n s . Resolution was 20 A by phase residual analysis. B) Reconstruction with two-fold symmetry shown at -70% of full volume, indicating relationship and con- wctivity of structurai domab. Labels, for only one af3 monomer, d e r fo biochemical domallis. Arrow- head indicates proposed plane of ceii membrane. LI, C-R, L2 = Ll-Cysteine-rich-L2 domains; CD =mm nechg domain; Fnl, Fn2 = nbronectin Hi repeats 1 and 2; TK = tyrosine kinase; TM = trammembrane domain (Luo, et al. Science 285:1077, 1999).

The rcletive orientations of the selected m o l d e mes of NGBYEW aim
O#ensmeya, 1993). They are plotted in Hg. 3-12, iadicating a -y rdom didriibuaon
of angles for these images with some tendency t o m a p r e f d orientation- Such
tendencies away h m riBILdOmness in orientation are compematd prier to 3D
r e c o d o n by orientation dcpendcnt weighting ofthe images.
The 3D recomtruction ofthe NGBI/HIR wmplex is shown in Figure 3-13k The
shucnne at the fWy expeskd volume, shown in Figure 3-13A (top panel), is compact
and giobular in overall shape. The domain-like fcatines of the structiin are shown in
Figure 3-13B at intermediaie densîty thresholds-
33. Location of the hnlCn-Binding Region
The location of NGBI on îhe recoLiStNCtion was detcnnineed by increasing the
density tbreshold without imposiag symmetzy (Fig. 3-13A, pane1 2 and 3) to pinpoint the
site of highest density @me1 3). Based on the d t s of biochemical evidence, insuIin
binds to the ectodomain of the nceptor, picucularly in the region of Ll-Cys-nch-L2, the
srst three contiguous domains of the IR (Fabry et al., 1992; Murray-Rut et al., 1992).
The hi- derisity in the 3-D volume indicates the l d o n of the NG clusta and, with it,
the location of insuün binding @ig. 3-13A, panel 3). It identifies this region of the
structure as the receptot ectodomain and, more specifIcally, part of the LI-Cys-rich-L2
domains.
When we fit a NGBI molecule into the tentative region (see details in Cbapter 3.3.
and 3.4), the best fit ïs o h h e d with a molenile of iasulin king in contact with the L1-

Cys-rich domaias of one a subunit and with the L2 domah of the other a subunit, T&is
isw mistent with the previously proposeû mode1 invohnDg both a subtlIUlfS in the high-
3.4. Donuin Structures of the HIR
The domain-like features of the sbcuctiae becorne evident d e n the stnrcture is
viewed at intermediate density threshoids (Fik 343& panel 2), and, except for the NO-
BI region, indicaie a strong 2-fold verticai r o t ~ * o d symmeky as would be acpeaed
h m the oligotetramerlc primary stnicture of IR. This symmetry was used to d u c e noise
in the recomtmctions and render the stmcbaes shown in panel 1 and in Figure 3-13B.
This Figure shows the top, bottom and side views of the stmdurr. putatively as being
viewed in the piane of the membrane, and in the extraceflular (top) and intracellular
(boîiom) perspectives.
In the side views, the top part of the stmctwe. where NG is Iocaîed, is identifiexi as
the ectodomain of the a subunit The dog-bone-shaped substnicture of the 3D reconscniction
(Fig. 3-13B, top view), and the equivaient topmost, bw-tie-shapecl stnicture (Fig. 3-13B,
OO), are designated as the two L1 domains of the dimeric receptor on the bask of the x+ay
stnichire of the Ll-Qs-rich-W d0main.C. The si& view at 65" shows the putative LL-Cys-
rich-12 domains as contiguous su- ~CLTOSS the upper centrai region ofthe molecule.
with enough additional volume in this region to sccount for most of the remahhg mass of
the two a subunits. primarily the ibnnecth-like comecting d0msin.c (CD) (O'Bryan et ai.,
1991) and the _cmaller insert domain (ZD) where the second diSulfide bond fom between
the two a subunits.

I a subunit
Fig. 3-14. The secondary domah structure of LIR (A) and the domain location in 3D structure. A) Linear locations of domauis in the dimeric hIR B) Schematic representation of the domain structure of one a p monomer based on the comectivity in 3D reconsûuction, the primary domain ~equence, the symmetry reqyirement for two disulndes on two-foid axis, the fit of known domain struchues, and on the principle of keeping unknown domains compact Di-stances between modeled locaticms of CD, Fnl, and symmetrid diadfides cornmensurate with numbers ofintemening amino acids (stn>caires not to scale; alpha subunit: red; beta subunit: blue and green; unknown structures: spheres or Lines): Labels: A = TK activation loop; 1 = Cys524; 2 = -82,683,685; 3 = diSulfiide bond betweea Cys647 and Cys872; arrowhead = cleavage site of pecursor recepta protein (A and B); other labels as d e s c n i in Fig. 3- 13)- C) Approximate locations of the domains on the 3D structure ofhlWNGB1 cornplex
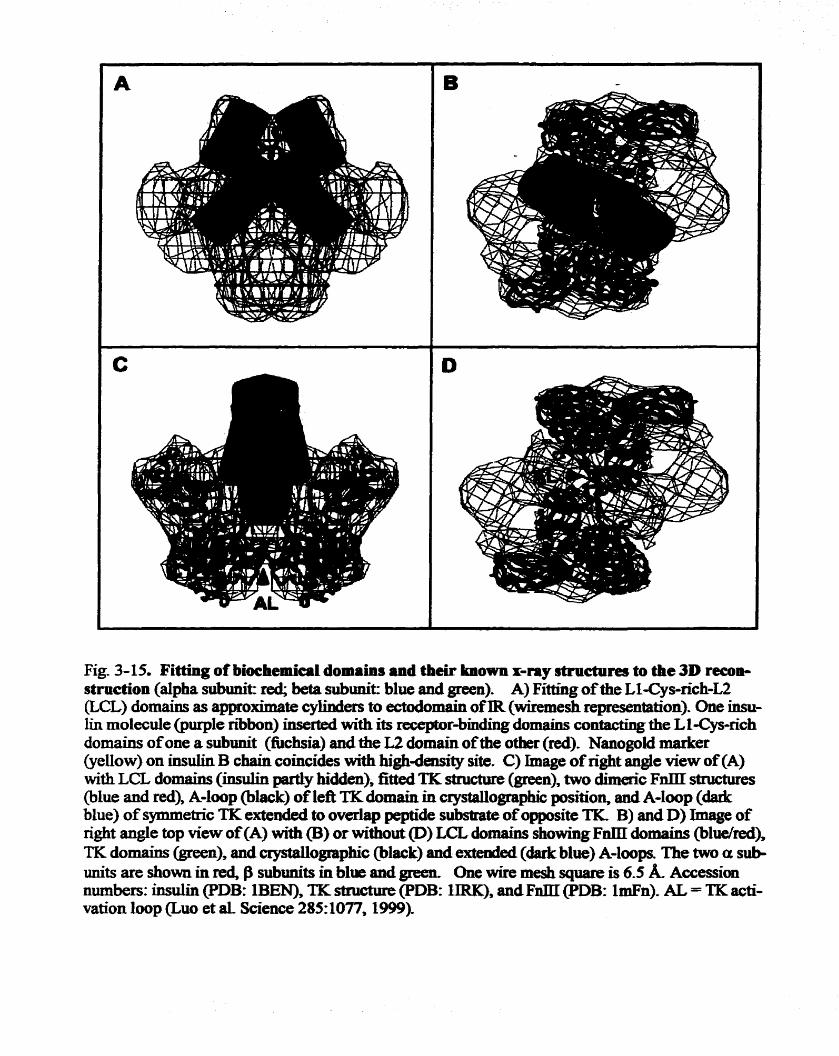
Fig. 3-15. Fitting o f bioehemical domaios and their h o w n r-my structures to the 3D recon- struction (alpha subunit: red; beta subunit: blue and green). A) Fitting of the Ll-Cys-richu &CL) domains as approximate cyllnders to ectodomain of IR (wiremesh representation). One insu- lin rnolecule @urpIe nibon) inserted with its receptor-biuding domains contacting the LlICys-rich domains of one a subunit (fuchsia) and the L2 domah ofthe other (mi). Nanogold market (yeilow) on insulin B chiain wincides with highaensity site. C) hage of right angle view of (A) with LCL domains (iiisului @y hidden), fitted TIC stnicture (green), two dimeric F a structures @lue and red), A-hop @lack) of left TK domain in crystauographic position, and A-Loop (dadc blue) of symmetric TK extended to overlap peptide substrate ofopposite TK B) and D) Image of right angle top view of (A) with (B) or without @) LCL damains showing F m domains (bludred), TK domaius (green), and crystallogmphic (bIack) and exteded (dark blue) A-loops. The two a sut+ unie are show11 in rd, f3 subunits in blue and green. One wke me& square is 6.5 k Accession nubers: insulin PDB: l B W TK stnicaae (PDB: lIRK), andFm (Pm: 1mFn). AL = TK acti- vation loop 6110 et al- Science 285:1077,1999).

The contiguity ofthe domain stmctme @ig. 3-13B, top ami side view 9û"), dong with tbe
residuesftomboththea andB subunit, is assignedtdteupperleftendoftûecres~enr(side
view, 0°) where it is contiguous with the CD portion of the a submit @op view). It bas been
s u g g d xecentiy that the CD has a fibmI1CCfiil-like stnicbae ~~ et al, L998). Fig. 345C and 3-15D (cf Fig. 3-138, 9û0, top vKw, iespcctively) show the fïtting of the
crystal stnichaeoftheTKdomain(gr0en) ofthe $ subunitdofthetwo FnIlIiepeats
@lue/.ed) modeled as the canonid ~~ type ï ï I stnictims (O'Bryan et aï., 1991;
P a s p i e , 1991).
The spatial relationsbip beîween the domains of the a and f3 subunits (e-g. side view,
90°) suggests the location of the celi membxane lipid bilayer as the space below the a
subunits and above the bidge linking the two dgned TK domains- Iosiead of a flat open
@on, this space in the 3D rrcoastnrction foms a thick dome-like slab above the bridge
with a thichess variation of 2.2 to 2.7 nu^
Starhg fbm the putative transmembrane domain, the $ subunits &me slightly
twisted towarü each other- H a t the intracelïularr TK domains of IR would then occupy this
portion of the (ide view, 6650, bottom view). There is a sepration of several
nanometers between the fkes of the masses of the two putative kinase domains. Around the

4. DISCUSSEON
By blocking two ofthe three fkee amine grow on iasulin m o l d e , we d e r i .
the B1 phenylalanine of bovine hsdh wïthNaaogold. The amino temiiaal region ofthe B
chah of insulin is beliwed to be minimaBy invoLved in receptor binding. Birxiing
experimemts showed that the Zeceptot-binding afniiity of NG-BI a f h dablocking was
decreased only slightly compared to native bovine insrilin. The con&ions ofderiVaSiZafion
were gentle enough for Nanogold since the pochrt showed a0 stmctud damage of the gold
clusta aftrrward î l e bigh pirrity ofNG-BI, mm> c o n î m b h g k e insulùi or Nanogold
molecules, ensured the specificity o f NG-BI bindiirg to HIR Aiîhough we iwbated both
NGBI and the bIR at relatively h i g k concentmtïon (10% " to to maximal bbiadirig
of NGBI to the IR, the rnibsequent removal of fine ligand a d the dilution of the bound
wmplex establishexi a new equili'bntun concütï011 The new equilibrium wodd result in the
binding of one NGBI to one hZR STEM h@ng confinnecl the presence of one NGBI
bouud to one hlR in the of the complex @cles. There was aïso very few fine
NGBI particles seen in the background (Fig. 3-7A).
To achieve hi@-redution reconstruction of the NGBI-HIR complex, we applied
the recent advances in elecûon cryo-rnicro~copy to out study, including the instrument,
sample -on and image processing. To pqyre the specimen, we d d out the
Ligand-receptor bmding in physiologicai biinir, followed by qyick freezing, and fnac-
drying inside the STEM at -130 to -140. C. This maintainecl the proteIn in the near

phNoIogicaI hydrateci w-011. dmhg image aqiidion. At a vezy low dose (+d'A!),
imagnig at -1500 C marLsdy rrdueed the radiation damage. Th use of both elastic a d
inelastic detectors to captine morr signais in the S m nir(ha breased sienoise ratio.
A sharp Nanogold signai was used to mainiein di the images m sharp f m - These
improvements are fidamentai for the successfiil3D remuüudon. As a dt, for the frst
tune, the 20 A resolution obSamd in this study d e d clear domain stnrctims of the
HIWNG-BI cornplex. A strong two-fold verticai rom*onal symmetry as anticipated h m the
dimeric configuration of the oiigotetrameric (apb dmctme of IR can be seen in Fgmr 3-
13B. A clear Namgold-insulh sisignal can be . * * on the 3D cornplex stmdme and
localized on the assignai L1-Cys-richL2 domah
The o v e d 3D structure of the ~ - m S a h rece~tor eomnles
In this shdy, we have showun thaL the ovaaIL shape of the 3D reconstniction of the
insulin-IR compIex by STEM is compact and g l o b e (Eg. 3-13A-1,). In studies by othas,
the HIR images obtained by e l w n mic~os~opy have been shown as X-. T- or Y-shaped
particles (Chnsnansen . et al., 1991; Schaefer a ai, 1992). Following the pubIicaîion of our
present study (Luo et al, 1999) Woldin et ai. Folclin a ai., 1999) reported their TEM
imaging of vesicle-reconstituted HIR cross-Iinked to a biotinyl daivatiaed insuiin
photoprobe @Bpa insulin) wtiose localbtion was visuaüpd by b i i to Nanogold
streptavidin. The TEM images were rather fûzzy and not clearly discemable h m the
background. Nonefheless, they showed the image of the nxeptor as Y or T shape with very
thin cW-Wa" Jhuctinw of O@ 1.5 nm in width for the armg the putative extracellular
domains of the receptor. 'Ibis is signifïdy d e r than that reporteâ (about 3 m) for the

appsrrntly d i f f i nOm ours- Nevextbless, Waen Wewiqg the domablike dmctmes m
some orientdons at mtmnediate deaüty ttii.esho1ds (Fik 3-13B), we do fbd images with
shapes- * . of X- or Yshspsd particIes- AIthough Woldin et aL used about 7 times
kger dose Ur o h i n h g their TEM mipses, the difference between our observations made in
this stuciy and observations by others ising TEM may be the d t of sample prepadon
For example, the negative-stain methoà, which was used in TEM studies may intduce
artifacts byhighlighang thebomdaûes or nIliiig i n d spaceq or alteringthe shape of
m o l d e s by iimoduaiig a nomphysiologicai ionic environment, Fiaehennore, stabhg a h
limits the potential resolution by the large sîze ofthe staining ions,
Superimposiilg h w n aysEal stmctms of d e r domains of the q t o r on
substnictures of the ~econstniction has macle it possiale to deduce the spaaal relationship
among the domainw in the overail cornplex. There is no crystal structure available for a
protein with such a big moleculat s iz . However, crystai d m û w û m s of severai domains of
the human insuiin receptor are available. Tbey are either h m the insuiin receptor or its
closely homologous family members. These hclude the TK domain of the insulin zeceptor
@ubbard et ai., 1974; Hubbard 1977), Ll-Cys-rich-L2 domain of IGF-1 receptor (Ganrett et
ai., 1998) and fibrrctin III repeats domaias (O'Bryan et al, 1991; Pasquale et al., 1991). Al1
these domains cau be properiy Mted into the correspondhg domain volume of our 3D
reconstniction Taken together, the o v d stmctue shows the division of the complex into
the extracellular and the cytopWc segments dong a plane, the putative cell membranes on
which the fibn,nectïn type ilI npa ts lie. These repeats appear pontoonlike to support the

d y 1- insulin-binding segment of the ectodomaim Undarvash the putative
tnmsmedllbraae mon, TK domaihs oc- the major voIirme of the htracellular portions
of the f3 subhts @ig- 343B),
Stnictunl correlation wîth the ~rinrrirv stm- of HIR
As a complex Ileteroîeîratner membrane protein, the tW CO- svei.l fîmctional
domallis. 1t bas ban proposecl that the specific fimctionai properties of the IR reside in its
separate domains. B a d on itp Pnmay stnicaae @ig. 3-14A)¶ the stmctud relationship
of the various domains in one or$ monomer is schematïcaliy show in Figure 3-14B.
Beginnins at the N4erminaI, the LL-Cys-n'ch-L2 domain (tadi contains about 150 amino
acid residues) le& into a comectïng domain (a9 a structure of Fibnectin III repeat (see
Ward, 1999), assigned FnO) ofabout 140 amino acid residues, foilowed by part ofone F a
repeat @nl) and the insert domain. The comecting and insert domains contain -24,682,
683 and 685 that form at least two disulfrde bonds between two a subutüts (Spamw et al.,
1997). The FnIU repeat in general is a p süucture of s e v a strands (Baron et aL, 1%
Leahy et al., 1992). The second F a repeat is constitiited by amiao acid sequences nMn
both a and subunits, and is htempkd by the ckavage site between the two subunits. The
N-terminai ofthe $ subunit contains about the of the seven f3 straods ofthe second rrpeat
(Fnl) and the entire third repeat Wn2). Fmm symmetry consideration, the disuüïdes am
located at the center portion of the 3D stnictiae. The two a p dimers are linLod by the
disuifide bonds between FnO and insert domains and the a anci subunits are connected by
a disuifide bond between Fnl and Fd? repeat domains. The third repeat is îinked to the TK
domain by a secpnce of 70 amino acids, incliidinp the 23-residue transmembrane sequence.

acid residues) oOcuplLes the majorvolinne ofthe mtracc1lu.k portion of aie subunit
S-on of known crystal stmchnes of d e r domains of tbe meptot on
substnrcaae~ of* 3D zec~OStNCtion would codkm the domain asSigment as cfisaissed
above and thus hlps to deduce the spahi relationship among th do& in the cornplex.
Availeble x-ray coodhates weze fitted to the respective subshuetine by modehg the
electron density of the xiay crystsuographic s&u%ms mto the volume of the assigned
domains of the 3 0 riecontrutïon. Figures 3-15C and 3-1!5D (CE Fig. 3-13B, 9û0, and top
Vimy mspectively) show the Mtiag ofthe crystai stmctme ofthe TK domain Cgreen) ofthe
subunit and of the F a ~epcais (bldred) to the reconstriIction. When fitted into the top
of the two crescent-shaped ap rnommers, the type III repeats Jie on a horizontal plane
£lankhg on two sides, pontoomlikey the nmaining strucûm of the a subuuits.
Two symmetrical TIC crystal stnictures (Fig. 3-15C a d D) can be fitted into the
putative intraceiiular space of the receptor stmctwe. nie masses of the putative kinase
domains are connatcd via a slender hotizontai bridge (Fig. 3-13B, side view 90") that was
not ob~exrved in the x-ray stnichms ofthe TKs The prrsaice of this bridge can be explained
in temis of the reconstniction being in a transition between fk IR aad its ligand-activateci
form. In this situation, the TK donains of the IR have undergone a confomrsitionai change
such that the A loop of one TK domain wouid k reaching out to the utalytic loop of the
opposing TIC domein However, since thae was no ATP in the biading buffer in the
prepouaton of the NGBI-IR complex for STEM, the activation of the IR stopped in this

stage and no bm-phosphorylation occumd The catalytic loops in the two fitted TIC
domains are separaid by 4 ma hterestingîy, this distance ik just SuffiCient to pemiit the
tyrosine triplet m 1 5 8 , 1 1 6 2 a d 1163) m a fiüly extendeci flexible activation loop ofone
TR, to reach the catalytic loop of the opposite TK as modeIïed Ecm the x-ray coordmates
(PDB lIRp). The extemsion of the activation loops wuid account for the Lmking deaSity
observeci between the lower portions of the subunîts m the structure assignments.
Our 3D stnictine showed a Iimited space for the transmembrane region of the
putative plasma membrane- This spacing is a change in shape, and a decrease in the
thickness h m that expected for a ~anbrane bilayer that would accommodate an aipha-
helicai transmembrane domain CTM) of 23-26 hydrophobie amim acids. However, since
the p d e d IR in the absence of its native membrane was Wly active, the relative positions
of the extraceiidar and intraceiiular domains must still represent a close to native
arrangement-
The mssing LI-Cys-rich-L2 domains of the dimerk a subunits are presented in
the reconstruction only in their general shape (Fig. 3-154 B and C), since the known x-
ray coordinates were not available (Garrett et al., 1998). An approximate mode1
composed of cylinders for the LI-Cys-rkh-L2 domains was generated instead. This
approximate structure was fitted into the extraceiiular portion of the 3D structure. Based
on the location of the Nanogold signal in the cornplex structure, one, or two if the
condition alIowed, molenile(s) of insulin should be fitted into this region discussed
below.
Structural Iocalizrrtion of the insnlin-bindinn site on the insulin rece~tor

By labeiïng the receptor withNGBr, we cleady demonstrated the stoichiometric
relation of inSulln to its receptor and showed the stnrcalral IOC8fI*on of insulin on the
receptor in the pnicess of-on. As me~ltiolied above, the crossing LI-Cys~ric~L;!
domains of the dimeric a subunits are presented in the mconstNCtion only in their
general shape (Fig. 3-15A, B and C). Nonetheles, using this sfnietirrr, the localizaton of
the gold cluster, and the known receptor-biading domain of ùrniün (Murry-Rust et aL,
L992), a NGBI molecule is tentatively fitted i . this region. The ntted insulin molecule
is in contact with the L1-Cys-icch domains of one a subunit and with the L2 domain of
the other a subunit. A mode1 imrolving both a subm*ts in the high9tnnty bindhg of
insulin has previously been proposed based on studies of insuiin analogues binding to the
IR and WiGF-1 R chimeras (Yip 1992; S c W i 1993; De Meyts et al., 1993). The 3D
recoIIStNCfion provides the structural evidence for th ïnvolvemmt Although two
mo1ecuies of ursulin can be fiîted to tbis configuration, two molecules of Nanogold-
labeled insulin were observed only rarely in the STEM images. The h i ~ ~ t y binding
of the fïrst insuün molecule to the IR has likely induced a conformational change in the
binding domain so that the second msuün molecule would bmd only et low affinity.
Likewise the binding of a second moIecule of insulin could effect a conformational
change that enhances the dissociation of the bound insulie Thus the cwyilitlear
Scatchad plot and the negative cooperatvity of insulin binding (De Meyts et al., 1976)
can be explained on the b i s of the 3D ieconstmdon. The reconsûuction also explains
why only low-afEdty binding is obtained with pmified a p monomer (Boni-Scnetzier et
al., 1987; Sweet et al., 1987)-

Evïdence of insiilUi. rcüvatson of the HIR
The 3D dmcture zweais that a siender horizontai bridge Vig. 3-13B, side view 9@')
wmects the masses of the prdative lrinaJe domain?c. It was not observed in the xiay
stmctms ofthe TKs, but, as stated above, can be expiaïned in terms of îhe r e m ~ C o n
being in a transition between the fke IR and ito Iigand-BCfivafed f o m h the two
symmetndy filied TK crystal structrires @ig. 3-L4C and D) the catalytic loops are
separated by 4 nm. This distance is just d c i e n t to pemnit the tyrosine triplet flyr1158,
1 162 and 1163) in a fully extended flexi'ble -on lmp of one TIC to reach the d y t i c
bop of the opposite TK as modeleci k m the x-ray coordlnates (PDB lIRp). The extendon
of the activation loops, eqiiident in mdon to four adadod po1ypeptide chains,
d y accounts for the iïnkïng Iuiloag oeasiryed betuem the lowerportions of the subuds
(Fige 13B, 90"). Ti& is an important dinkence h m the =ray stm%mes of the inactive and
activateci TKs as discussed below.
Monomeric inactive receptor TKs such as EGFR are brought together by ligand
binding and becorne activated as dimers resulting in TIC a u t o p h o h o o n (Canaals,
1992; Schlessinger aad UllrichJ992). Ln the inainsicaüy dune8c I R - M y mxptors, the
distance between the two cytopasmic ksubunït TKs within the dimer must be too great
without ligand b i g for the ectivetion of the kinase. Hubbard and bis coileagues
pubbard et ai., 1974; Hubbard 1997) suggested thet insulin binding to IR d e a e a d this
distance by dknpgkg Tyr1162 hmthe cataiytic loop to enable tram phosphorylation in
the presence of ATP. Zn the reconsûuction a good fit to the ligand-receptor complex is
obtained when the two TK domains are orienteci with th& caîalytic Ioops juxtapod In
this orientation the exîeded fiexiie activation loop of each TK, which moves 30 A

Hubbd lm, canjustreachthe catdytic loop ofthe opps&TKto be a c t i . 'Ibese
two l o o p s c a n d y f o r n t n e l i n l a n g m a p s ~ b c t a n a n t b e ~ s e n i p t h e 3 D
recollstrtlcttConin the absence of ATP.
The 3D stmctme obtsiaed nwi images of the HIR-compIex contains ody a suigîe
NGBI. This suggests that one m o l d e of insulin is sufncient to bring the two a$
monomas to an activating confTgiiration, Ln light of this supposition, it is signifiant to note
that the dimexic q r with a Sei.323- mutation in the L2 domain of both a suburiits
showed a sevete Hiipaimiem in insulin bhding (Roach a al, 1994), whereas a hytxid
reoeptor with ody one ofthe two a subunits miitattd was found to bhd insuiin with high
nffinity and was M y active as a tyrosine kinase Umuis a al., 19W). Based on the 3D
reconstru&on we would suggest that insuiin is bound to the L1 domain of the mutant a
subunit and the wild-type L2 domain ofthe hybrid IR and that the bindïng of only a sïngïe
molecule of insuiin is sufEcient for TK a c t i . o a
In this shdy the 3D guetemary stmctwe of the Ikinsulia cornplex f o d in the
absence of ATP was obtained Thus the smrhnr was expected to be an intemediate
between the insuiin-- IR and the fXiy activated, phosphorylated IR. 'Ibe recoostruction is
d y interpretd as a receptor poised for activation by *uns-phosphoryiaîio~. The fidi
extent of conformational changes induced by insuiin b i i remains conjecbiÿa, without
simiiar reconstnictions ofthe initial state of insulln-iiee IR and of the W activated state for
cornparison. Nonetheless, in the absence of a aystailographic stnicane of the insuiin
recepttor, the 3D recoastnrction presented here provides concrete stnicttnaI information
towards the fidi undersCanding of trammembrane signai trairpmission in insulin action.


CONCLUSION AND FUTURE RESEARCH
The nntstep inthepathwayofmwilinactionis thebindingofirrpulinto its ce11
sinnice receptor; an integrai transmembrane glycoprotein, leadhg to the activation ofthe
cytoplasmic domain of the nxeptor tyrosine kinase- Mueh is lcnown about the iateraction
of the acfivated receptor with its dsûaks leadhg to dom-stmam signal transduction.
On the o t k han& how binding of insuiin to the extraceliuiar domain of the receptor can
lead to receptor activation rernains unknow11, Understanding w h e insulin binds to the
exbraceiluiar domain would provide the foundation to anmer this Question.
Many approaches, biochemicai and biophysîcai, have ken applied to elucidate
the site(s) of insulin-insulin zeceptor interaction Diffbnt d t s were reported, and
various regions in the a subunit of the IR have ken suggested to be involvesi in the
insulin-IR interaction. These regions inchde almost every major domain, such as N-
terminai, cysteine-rich domain and C-tenninal of the a subunit, An accurate location of
the insulin-binding site on the xeceptor has not been fortbcoming.
The specific goal of th& study was to locaüze the insulin-binding site in the IR,
and, if possible, to elucidate the molecuiar detaifs of insulia-IR interaction and the
relatiomhip of the stmctme and fbnction of the iilJulin receptor. To achieve this goal by
the biochemicai appmach, 1 have expred the human insulin receptot extraceîlular
domain in a large s a l e in the baculoinis expression system and used various specinc
photoa5nity probes to identifil the insalui contact sites. The photolabeled receptor
protein was digested with proteases, and high-sensitivity identification methods wae
developed to detect the labeled IR fbgment contahihg the putative insulin-binding site

by HPLC -&mappiag d y & - 1 have fOCUS4d on a fbgment of 1-3 kDa that mi*
contain the putative -binding or 0 0 ~ site on the reccpf~r a subunit;
However, 1 I d to obEain SLlfncient qymtin'es of the fkagment for emino acid
~ e ~ ~ e ~ l c i a g , partiy drr to the ladr ofa high efficiency photoprobe of iosului and a high
efficiency peptide mapping method AvaiMiIity of inoalin photoprobes with higher
efficiency is a kcy in fÙture studies ushg this approach. The bcnzoylphenylalsinine
photoreactive group in a anv msulin photoprobe has shown a much hi* cross-lnilring
efficiency than the traditionai phenyIazido photoreactive group. A cross-luiliag
efficiency of more than 50.A has been reportecl for this photoprobe compared to Iess t h
10 % reportai for other insuiin photoprobes (Shoelson et al., 1993). The use of this high
efficiency photoprobe should lead to a higher yield of the cross-hkhg productsC Tii
addition, methods for the purification of the cross-linked fhpnent and for higher
sensitivity in amino acid sequeaciag need to be developed The use of MALDI-TOF to
determine amino acid sequence of srnall peptides is clearly a p d c a l alternative.
M y next approach to the goal was cryoelectzon microscopy, spedically STEAK
By using insulin labeled with an electrondense gold cluster to bind to pirrified human
placenta membrane insulin receptor, 1 successfiilly detennined the 3D structure of
insulin-iasulin receptor complex at a resolution of 20 A and localized the site of insuiin
binding on the complex. The stnifam was fitted with available known high redution
crystal domain substnicanes to obtain a detailed contiguous mode1 of the
heteitotetrameric (aph transmembrane receptor. The 3D ieconstniction shows that the
two a subunits jointly participate in insuiin Endhg and that the two S s u b d kinases in
this receptor-ligand complex are in a jiatsposition thet permits trans-autophosphorylation

on finding only one molccule of insulin (as visiiaüzed by the gold-labeled insulin), ami
oniy rarely two. the zecoIlStNCtion provides shactmal evidence fot the involvement of
only a single iasulinm01ecuIe in receptor activation.
The <iuiitcmery stnrane ofhuman insaün receptor wmplex detnmined bythu
study provides the shuctiiral expianaiion for many of the known c-CS of insulin
bmding to its receptor* Fitting the crystai stnictirrG of hdh into the fec~nstructioa
mode1 shows that the L1, CR and L2 domains of both a subunits contili'bute multiple
contacthg sites for high9ninty insuiin-IR intaacton. The stnictiirr a b shows that
exbraceIlular Qmains of the B subunits play d e in s t a b i i the highdlh& insulin.
binding site. Moreover the stnicbne provides a mechmistic basis for the mechankm of
The stmctue ais0 e x p W many biochemicai findings in the studies ofthe insulin
receptor. A recent study indicateà that insutin receptor isofonn A in which exon 11 is not
expressed, besides its high dlinity to insulin, binds to IGF II with an affinity close to that
of iosulin. It appears to serve as a high-afEnity IGF II receptor affecting the growth of
fetal and cancer ceb (Frasca et aï., 1999; Sciacca a al., 1999). lkis Gnding suggests that
the 4 variation in the C-terminal o f the a subunit of the insuiin receptor plays an
important role in the wnfonnation for high-ety ligand binding. Interestingly, binnding
prolacth, are composed of the two fibronectin repeats of each monomer that fomio the
dimeric receptor Web, 1996; BoleFeysot et al., 1998). The skucture we bave obtained
for the wmplex shows that the C-terminai ofthe or subunit and îhe adjacent N-terminal of

thep subunitwntsinsasùnilarstniehncoffibro~rrpeatlidlmer~Howthe~
or the absence of these 12 rcsïdaes ccxied by the exon LI, in the C-tenainal region d d
change the IR'S affinity to IGF-II is uuknown. H o m , the stnictural similPr@
between the IR C-terminus and growth fhctor receptots Qes provide a stniduraI b i s for
a suggestion that the C te rmï~L region of the insuiin receptor isoform A could serve as
&other Iigand binding site in acUti011 b the LL-CR-L2 region for insalia This
hypothesis wuid be confimeci by STEM Smüar to the present study, but using
Nanogold-IGF-II-
Insuiin and impaireci insuün d o n play a key d e in the pathogenesis of di-,
in type II diabetes which account for more tfLen 85% of the diabetic
population. While a combination of genetic and environmentai fktors contn'butes to the
development af type II II, defects in the insulin si@ transduction pathway
beginning with the intetaction of iasulia with its receptor play a major d e in the
manifestation of the disease. The 3-D recoIIStNCtion of the insuijn-IR complex describecl
here presents the fïrst mode1 to explain signaLing process fion ligand binding to tyrosine
Ianase activation The sirucairal localization of the insulin binding sites in the IR
provides evidence of how and where iiisulin interacts with its receptor. Combining with
information hrn crystai structure ofthe i s o W receptot domains, such as LI-CsL2 of
the IGFI-R, it wodd be possible to obtain more details ofthe insulin-IR interaction. This
would eventuaiiy heip to develop (1i. new antï-diabetic medicine.
Recently, a non-peptidyl fimgal compound L-783,281, was reporteci to have anti-
diabetic activity in mice (Zhang et al., 1999). M e r than interacting with the insulin-
binding sites on the a subuints, this compound appears to interact with the cataiytic

stül mimic the efEcts of the ligand of the Lcccpbr. Our ramnstniction mode1 may
provide a shucaiiril basis for sbuiying the potential mechaniSm by wbich tbu fimgai
compound actives tyrosine Linase ofthe IR. Otheroral (or non-oral) antidiabetiCc agents
have aiso ban reporteci to have insuiin mimimimic e f f i such as vanadates, PTP inhibitors
and phosphoinositoIgLycaus Frick et ai., 1998; Shdev, 1999; Chen et ai., 1999; Iversen
et ai., 2000). Some of them dirrctly ïnteract with the IR. It is still not very clear how these
cornpo~~~ds mimic insulin action at the m o l e leve1. Neverthelesss more detailed
structure and fiinction studies and the nhecpent stmctwe-based design for new dmgs
wouid help to develop rational therapies for trrating insulin resistance-
Besides the principai IRS substrates, other nhtmtes, such as Shc, might also
exist in the insulin-signaling pathway. Altemative pathwmys of the IR may be responsible
for the multiple actions of insulia Shc is a substrate protein rapidly tyr-phosphorylated
by insuiin stimuiation. Shc contains an Noteminai PTB domain which binds to the
phosphorylated N P X Y ~ ~ ~ sequence in the insului receptor md can also bind Grb-2 to
link these signais to Ras activation (Skolaüc et aL, 1993; RonL et al 1994; Gusta£Eon et
al., 1995). It has ken suggested that Shc, rather than IRS-1, may be the main mediator of
the mitogenic actions of insuiin and IGF- II (Yamaguchi and Pessin, 1994).
Aithough our observation represents a signiîïcant advance in elucidating the
structure of the insulin receptor, and provides a structural basis for the mechanism of
receptor activation, additional saidies remain to be done. First, the present study was
d e d out on detergent-solubiï membrane protein in the absence of a bilayer

membranet Althoughtht s o l u b i i protein has been show11 to k fUy active in ligand-
binding and autophosphorylstion when cornpprcd tu placenta membrane receptor, the
sauctine of the comp1ex obtained did show a cornpresseci tninsmembrane domain in the
3D r e c o ~ o ~ ~ A receptor in a biIayer q q o x t would p v i & a more naturai
recomtruction. S W e s on 2D crystai of the protein supported by a iïpid monolayer or
bilayex would greatly increase the final res01ution of the 3D rewnetnicdon. Severai 3D
reconstructions obtained by electron cryomicroscopy based on 2D crystd mes have
resolved the stmdure of a nunber of proteins et resolutions below 7 A ( m g et al.,
1997; Walz a ai., 1997), very close to the resolution obtalned by x-tay crystdlography.
Since the insulin receptor has so fiu not been crysîaked the 3D - d o n of this
protein at a higher resolution is especially miportant to elucidate the molecular daails of
the fiulction related stnictun, such as the insulhbinding site. Understandiog the
molecular mechanism of the insuh-IR interaction would help in designhg new drug foc
treatment of diabetes meIlitus.
Insuiin bi~lding to the receptor leads to codonnation change of the receptor. This
conformafion change is the key step to receptor activation. A reanstruction of the fke
receptor without bomd iiwlin would provide very usefhi information Cornparison of the
activated and the pre-activateci conformation of the receptor wiii provide the information
required for undexstandhg the mechankm of receptor activation. Cn this study we
obtained the structure of a receptor bound to iasulin at the pre-phosphorylation stage in
the absence of ATP. Addition of ATP to the insulin-binding b s e r could reveai the
conformation of W y phosphorylated receptor kinase. Combining informaîion obtained
h m different activation states should help us to understand the mechanism of insulin

receptot a c t i . 0 1 ~ Furthermore, in addition to monitoring the conformatio~ change
during the receptor dvafi*on, electron ~ ~ ~ O I I I I * C I O S C O ~ ~ wodd also k an exceiient t d
for visuali;nng * . the c o n f o ~ * o n change ofthe receptor resulttîng h m mutagenesis, This
visuaiization wouId provide the direct explanlition ofthe chan@ relationship in stnictun
and firnctïon Visdhtion of clown-stream interaction with signal transduction elenicnts,
such as IRS proteins is also v a p important for mkstanding the mechanism of insuiin
receptor activation and the uisulin s i m g systemstem IRS proteins are another grwp of
proteins that phy important des in signal traasdm*on but are dficult to crystaUize
structure for d y i n g their stnichae and fiincti011, W e c a n imegine that th cornbidon
of molecular g d c s in sampIe preparation, stateof-the-art electron cryomimscopy for
data acquisition, and pownful image proceshg will open a new era of structure and
fiindon studies for the iiwlin receptor at close to the atomic resolution level.

Abbott AM, Buew R, Pcdrini MT* Muaay JhiZ Smith RL (1992) hdim-like growth fiictot 1 receptor gene stmctme- J Bi01 Chcm 26?(lS):lO759-63
Adham Ih& Blundeii TL. Dodsen U, Dodson GG, Vijayaa Il& Bakr EN, HodgLin DC, Rimmer B and Sheaî S. (1%9) Stnrtiire of rhombohedrai 2 zinc insuiin crystals. Nahue 224:49 1-5
Andersen AS, Kjeldsen T, W~berg FC, Christensen PM, Rasmussen JS, No* K, M o l k KB, Molier NP. (1990) Chrmging the iiwün roceptor ?O possess insulin-like growth h t o r 1 ligand specificitp Biochemistry 29(32):7363-6
Bajaj U, W d e I d MD, Schlessinger J, Taylor WR, BhmdeU T. (1987) On the tatiary stnicture of the extracellular domains of the epidetmal p w t h fiilctor and insuiin recepttors. Biochim Biophys Acta 916(2):220-6
Baker EN, Blundeli TL, CiJtfIeld JF, Cutfield SU, Dodson ET, Dodson GG, Hodgkin DM, Hubbard RE, bacs NW, Reynolds CD, Sakabe K and VijayanM. (1988) The structure of 2Znpig insulin crystals at 1.5 A resolution. Philos Tram R Soc L o d B Bi01 Sci 3 l9(L l95):3694S6
Baron M, Miin AL, DnscoU PC, Mimion EEJ, Boyd J, Campbell ID. (1992) 1H NMR assignment and secondary sûuctme of the ceLi adhesion type III module of fibmnectin. Biochemistry 3 L('7):2068-73
Baulieu EE (1990) Hormones: a wmplex communication network. Iii: Hormones: h m molecules to disease. Badieu EE Keliy PA (eds), C h a p m and Hall, New York, ppl- 169.
Beckmann R, Bubeck D, Grassucci R, Penczek P, Vershoor& Blobel G, Frank J. (1997) Alignment of conduits for the nascent polypeptide chah in the n'bosome-Sec61 cornplex. Science 278(5346):S 123 -6
Bilan PI and Yip CC. (1994) Unusual insuiin bmding to ceils expressing an insulin receptor mutated Blundeli TL. Cibneld JF, Dodson GG, Dodson E, Hodgkin DCy Mercola D. 0971) The structure and biology of insulin. Biochem J 125(3):50-1-
Blundell TL, Wood SP. (1975) 1s the evolution of insulin Darwinian or due to selectively neutrai mutation? Nature 257(SS23): 197-203
Boisset N, Grassucci R, Penczek P, D e b E, Pochon F, Frank J, Lamy JN. (1992 ) Three-dimensionai ~econstruction of a cornplex of hunüm alpha 2-macroglobulul with monomaleimido Nanogold (Aul.4nm) embedded in ice. J Stnict Bi01 109(1):3945

BoIe-Feysot C, Oonm V, Edeÿy lb& Binart N, W y Pk (1998) Rolaetin (PRL) aud ifs recqtor: actions, signai tramdudon pathways andphenotypes observeci in PRL receptot knockout micece Enàacr RCv 19(3):22!548
Boni-Schnetzler Scott W, Waugh Sh& DiBeIla E, Pilch PF- (1987 ) The insulin receptor. S m basis for high affinity ligand binding. I Bi01 Chem 262(17):8395-401
Bottcher B, KiscIty NA, Stel'Mashchuk VY, Perevozchikova NA, Borisov AV, Cmwther RA, (1997) Threeimensional stnmcane ofiirfectEous bursal disease virus determined by e1ecfrOn ayomicroscopy- J V i l 7l(f ):325-30
Boni-SchnetzIer U, Rubin JB, PiIch PF. (1986) Strucniral requitemeents for the trammembrane actiVBfion ofthe insufin receptor kihase J Bi01 Chem 261(32):15281-7-
Brandenburg D, Dhconescu C, Saunders D, Thamm P, (1980) Covalent Iinking of photoreactive insulin to adipocytes produces a prolongeci signal- Nature 286(5775):82I -2
Brange I, RibeL U, Hansen JI?, Dodson G, Hansen MT, Havelund S, Melberg SG, Norris F, Noms K, Snel L, et al, (1988) Monomeric insulins obtained by protein engineering and their medicd implidons, Nature 333(6174):679-82
Brissenden JE, Ulhich A, Fmcke U. (1984) Human chmmosomal mapping of genes for insuiin-Iike growth factors 1 and 11 and e p i d e d growth factor. Nature 3 lû@980):78 1-4
Brown H, Sanger F and Kitai R (1955) The structure of sheep and pig insulin. Biochem J 60: 556-65.
Brown VI and Greene MI. (1991) Molecular and ceIIular mechankm of ~eceptor- rnediated endocytosis- DNA Ceil Bi01 10(6):399-409
Canais F. (1992) Signal transmission by epidennai growth factor receptot: coincidence of activation and dimerization. Biochemistry 3 l(lS):U93-50 1
Caro LH, Oh& A, Goden P, Coîüer E. (1994) Mutational analysis of the NID-terminal glycosylation sites ofthe insuiin receptor alpha-subunit. Diabetes 43(2):240-6
Cheng A, van Hoek AN, Yeager M, Verkman AS, Mitra AK (1997 ) Three-dimensional organization of a human water chamel- Nature 387:62?-30
Cheatham B and Kahn CR (1992) Cysteine 647 in the insulin receptor is required for normal covalent interaction between alpha- and beta-subunits and signal ttansduction J Biot Chem 267(lO):7lOS-lS
Cheatham B and Kahn CR (1995) Insuiin action and the insulin signahg network. Endm Rev l6('): 1 1742

(3heatham B, Vlahos CJ, Cbeiitbam L, Wang L. BlenïsJ, ïhhn CR (1994) Ph~sp~dyIinositoL 3-Linesc activation is mcp id for insuiin stimulation ofpp70 S6 kinase, DNA synihesis, and gfucose transporfer transiocatÎon. Mol Cell Bi01 14(7):4902- II
Chen & Cong-LN, Li Y, Yao ZT, W u L, Zhang ZY, Burke TR Ji, Quon MJ. (1999) A phosphotyrosyl mi"metic peptide reverses impairment o f i n s d b s t i m ~ translocation of GLUT4 caused by overexpression ofPTPlB in ratadipose celis. Biochemistry 38(1)384-9
Chiu W, and Schmid MF. (1997) Pushing badr the lirnits ofelectmn cryomicroscopy. Nat Stmct Bi01 4(5):33 1-3
Cbristiansen K, Tmum-Jensen I. Carisen J9 Vinten 1. (199 1) A mode1 for the cluatemary stnicture ofhmnen pfacental h s u h raeptor d e d d h m electron micmscopy. Roc Nat1 Acad Sci U S A 88(1):249-52
Clapham DE and Neer U. (1993) New mles for Goprotein beta gamma-dimers in trammembrane signdbg- Nature 365(6445):403-6
Conway JF, Cheng N, ZLotnick A, WingfieIdPT, Stahl SJ, Steven AC. (1997) V Ï Ï o n of a 4-helix bundie in the hepatith B virus capsid by cryo-eiectron rnicroscopy, Nature 386(6620):914
Cuatrecasas P, (1971) Insulin-receptor interactions in adipose tissue ceUs: direct measurement and properties- Roc Nat1 A d Sci U S A 68@):1264-8
Cosgrove L, Lovrecz GO, Verkuylen A, C a . e R L, BIadc LA, Bentiey JD, Howlett GJ, Gray PP, Wad CW, McKem NU (1995) Purification and pmperties of iiwlin receptor ectodomain h m large-de mammalian celi culture. Protein Expr Purif 6(#:789-98
Crowther RA, Amos LA, Finch JT, De Rosier DJ, Klug A. (1970) Three dimensional reconstructions of sphericai vinises by fourier synthesis Eiom electron micrographs. Naîure 226(244):421-5
Crowther RA. (1971) niree-dimensionai reconstruction and the architecture of spherical viruses, Endeavour 30(111): 124-9
Czamota GJ, Andrews DW, Fattow NA, ûttensmeyer FP. (1994) A structure for the signal sequeace binding protein SRP54: 3D reconstruction fbm STEM images of single molecules. J Struct Biol L l3{1):35-46
C~a~lota GJ, Bazett-Jones DP, Menda E, AMey VG, Ottensmeyer FP. (1997) High resolution microanaiysis and thxee-dimensional nucleosome structure associated with transcribbing cbromatin. Micron 28(6):419-3 1

C s h MP. (1982) Stnichaal and fimctional hom010gies in the ~eceptors for insulin and the hsublike p w t h f"actors. Ceii 3 I(1):8-10
Czech Mi?. (1989) Signai tmmmkion by the insuiblike growîh ktors. CelL 59(2)235- 8
Takata Y, Webster NJ, Ole- JM, (1992) TnûaceUuk simialinp by a mutant human insulin receptor lacking the carboxyl-termid tyrosine autophosphory1ation sites. I Bi01 Chem 267(13):9065-70
DeFnxao RA (1992) Pathognwis of type 2 (non-bdk dependent) diabetes mellitus: a balanced overview. Ihbetologïa 3 5(4):3 89-97
Daosier RA and Klug k (1968). Nature 217A304,
De Meyts P. (1976) Cooperative piopeaies ofhomone reaptors in ceil membranes. J Supramol Struct 4(2):241-58
De Meyts P. (1993) The diabetogenes concept 0fNIDDM- Adv Exp Md Biol 33489- 100
De Meyts P. (1994) The structural b i s of insulin end insulin-like growth factor4 receptor binding and negetve ~~~~perativiity, and its reievance to mitogenic versu metabolic sigxdhg. Diabetologia 37 Suppl2:SI35-48
De Meyts P de, Roth J, Neville DM Ir, Gavin JR 3 4 Lesniak Mk (1973) Insulin interactions with its receptors: arpnimental evidence for negative cooperativity. Biochem Biophys Res Commun 55(l):154dL
De Meyts P, Vaa Obberghen E, Roth J. (1978) hkpping of the residues responsibIe for the negative oooperativity of the receptor-binding region of iiwlul Nature 273 (5663):504-9
De Meyts P. Gu JL, Shymko RM, Kaplan BE, Bell G1, W h i a L a J. (1990) Identification of a ligand-binding region of the human iosulin receptor encodeci by the second exon of the gene. Mol Endocrino1 4(3):409-16
Drake PG, Posner BI. (1998) riisulin receptor-8ssociated protein tyrosine phosphaîase(s): role in insulin action. Mol Cell Biochem 182(1-2):79-89
Dryden KA, Wmg G, Yeager M, Nibert ML, Cwmbs K.b& Furlong DBy Fields BN, Baker TS. (1993) Early steps in reovinis infection are 8ssociated with dramatic changes in supram01ecular sîmcture and protein conformation: ~ y s i s of virions and subvirai particles by cryoelectron microscopy and image reconstruction. I CeU Bi01 lZ(5): 1023- 41

Elcheblyw P a y e P, MicWkyn E, Cromlish W, CoCallùw S, Loy AL, Normandin D, Cheng A, Himms-Hagen J, Chan CC, Ramachandmin C, Gresser UT. Trembla. MLy Kennedy BP. (1999) hcreased insulin sensitivity and obesity mistance in mice iacking the protein tyrosine phosphatase-1B gent; Science 283(5407):1544-8.
Engl J, Moule M, Yip CC. (1994) Dithiothreitoi stimulates msaünteceptor autophosp60ryIation at the jutamembrane domain, Biochem Biophys Res Commun 201(3):1439-44
Fantl WJ, Johnson DE, Williams LT. (1993) Signaüing by receptor tyrosine kinases. Annu Rev Biochem 62:453-8 1
Fabry M, Brandenburg D. (1992) Andysis of the human insuiinreceptor. Biol Chan Hoppe Seyler 373(9):9 15-23
Fabry M, Schaefer Ey EIlis L, Kojro E, Fahrenhoiz F, Brandenburg D. (1992) Detection of a new hormone contact site within the iasuüareceptor ectodomain by the use of a novel photoreactive insulin. J Bi01 Chem 267(13):8950-6
Farrow NA and Ottensmeyer-FP, (1993) Automstic 3D alignment of projection images of randomly oriented objects. Ultramicroscopy 52,14146,
Farrow NA and Ottensmeyer FP, (1992) A posterioi deteunination of relative projection directions of arbitarily orienteci macromolecules. L. Opt Soc. Am. Ag, 174960
Feener EP, Backer JM, King GL, Wilden PA, Sun XJ, Kahn CR, White MF. (1993) Insulin stimulates serine and tyrosiae phosphorylation in the juxtamembrane region of the insuiin receptor. J Bi01 Chem 268(15): 1 1256-64
Finn Fw Ridge KD, Hofimm K. (1990) Labile disulnde bonds in humau placenta1 insuiin receptor. PIoc Nd Acad Sci U S A 87(1):419-23
Frasca F, Pandini G, Scalia P, Sciacca L, Mine0 R, Costantho A, Golâfhe ID, Beifiore A, Vigneri R (1999) Insulh receptor isoform A, a newly recognized, hi@-aninity insulin-like p w t h fmor II receptor in fetai and cancer cells. Mol Celi Bi01 19(5):3278- 88

F e AL, "ïteadway JL, Pessin JE (1991) Evïdence supporting apassivt d e fôr the insuijn receptot transmembrane domaïa inihdk~pendentsignaL transduction. IBiol Chem 266(15)9829-34
Fndc W, K, A, Bauer Jy Wied S, MderG. (1998) Structure-activity relationship of synthetic phoJphomosit01glycans ~ k k i n g metabolie insuüa action. Biocbmiistrg 3 7(3 8): 13421-36
Fn>esch ER aadZapf J. (1985) -1ike growth tbtors and iMulin: comparative aspects. Diabetologia 28(8):485-93
Fuller PI. (1991) The steroid receptor saperfamily: rnechmisms of diversity. FASEB I 5(15):3092-9
Fujita-Yamagucht Y, Choi S, Sakamoto Y, Itahaa K. (1983) Purification of insuiin receptor with fidl binding d t y . J Biol Chem 258(8):504S-9
Fujita-Yamaguchi Y, Kathuria S. (1985) The monomeric alpha beta form of the insulin receptor exhl'bits much higher -dependent tyrosine-specific protein kinase activity than the intact alpha 2 beta 2 form of the receptor- Roc Nd A d Sci U S A 82(l8):6095-9
Garbers DL and Lowe DG. (1994) Guanylyl cyclase receptors. I Biol Chem 269(49):307414
Garrett TP, McKcmNk& Lou lbl, FrenkeL MI, Bentley JD, Lovrea GO, Ellemau TC, Cosgrove U, Ward CW. (1998) Crystai stmctme ofthe first thr# domains of the type-l insuIin-like growth factor receptor. Nature 394(6691):395-9
Gherzi R, Russeli DS, Taylor SI, Rosen O U (1987) Reevaluation of the evidence that an mtibodyto theinsuliniaeptoris' msdhmimetic without activaîïng the protein tyrosine kinase activity of the receptor- J Bi01 Chem 262(3 5): 16Wû-5
Giranda VL. (1994) Stnictracbased drug design of antirhinovirai wmpounds. Structure 2(8):695-8
Goldfine ID. (1987) The insuiin recepto~ molecuiar biology and transmembrane signaling. Endocr Rev 8(3):235-55
Goldste'm JL and Brown MS. (1985) The LDL receptor and the regdation of cellular cholesterol metabolism. J Ceii Sci Suppl3:l3 1-7
Gustafson TA, He W. Crapm A, Schaub CD, O'Neill TJ. (1995) Phosphotyrosine- dependent interaction of SHC and insulin receptor substrate 1 with the NPEY motif of the insulin receptor via a novel non-SH2 domah Mol Ceii Bi01 15(5):2500-8

HanLs SK, QPùmAM, Hmter T. (1988) TkprotelnLmeseEamiy: conservcdfeaiurrs and deducedphylogeny ofthe d y t i c &mainsC Science 241(4861):42-52
Hainfeld JF, Fiiniya FR (1992) A 1.4-nm gold cluster ~0vaIentIy attacheci to anti'bodies improves immunolabefing. J Hhchem Cytochem 40(2):177-84
Waring W, (L99 1) The insulin recepfOr, siPnlling mechwiism and contn'busion to the pathogenesis of insulin resktance- Diabetol~gia 34(-2):848-61
Haynes FI, Hehethorst E, T p CC. (ï986) The stnictiirr ofthe hepatic insuijn rrceptor and insuiin binding- Biochern 1 239(1):127-33
Kenderson R., Baldwin JM, Ceska TA, Zemiin F, Bec- E, Downing KR (1990) Mode1 for the stmctme of bacteriorhoàopsin based on high-resolution e1ectton cryo-mimscopy- J Mol Biol 213(4):899-929
Hotarnisligii GS, Shsrgill NS, Spiegelman B U (1993) Adipose expsion of -or necrosis ktor-alpha= direct d e in obesity-linked insuiin resktauce- Science 2Sg(SO9 L):87-91
Hubbard SR (1997) Cystd stnictrae ofthe activateci iiwün rezeptor tyrosine kinase in cornpiex with peptide substrate and ATP analog, EMBO J 16(18):5572-81
Hubbard SR, Wei L, Ellis L, Hendrickson WA (1994) Crystai stnichirr of the tyrosine kinase domain ofthe human iasulin receptor. Nature 372(6508):746-54
Humbel RE. (1990) Insulin-like growth fbctors 1 and II, Eur J Biochem 190(3):445-62
Hunter T. (199 1) Protein kinase classification. Methods Enzymol200:3-37
Hunter T, Cooper JA, (1985) Pmtebtyrosine kinases. Annu Rev Biochem 54:897-930
Hotamisiigd GS, P d & P, Budavari A, EKs R, White MF, S p i e g e h BU. (1996) IRS-1-mediated inhibition of insuliP receptor tyrosine kinase activity in TNF-alpha- and obesity-induced insulin resistance. Science 27 l(5249):665-8
Ivemen LF, Andersen HS, Brauner S, Mortemen SB, Peters GH, No& K, Olsen O& Jeppesen CB, Lundt BF, Ripka W, M o l k KB, Moller NP. (2000) Structure-based design of a low rnolecuiar weight, nonphosphorus, nonpeptide, and highiy selective inhibitor of pr~tei~tyrosine phosphatase 1B. J Biol Chem 27S(M):lO3OG7

Jontes JD, Whn-Kubalek MilliCgan Rk (1995) A 32 degree triil swing in bmsh border myosin E on ADP re1ease Nature 378(6558):751-3
Kaburagi Y, MomomuraK, Yairsmoto-Hoda R, Tobe R, TamoR Y, Sakura& Akauuma Y, Y& Y, KadowakC T- (1993) S i k d h $ e d mutagenesis ofthe juxtamembraae &main of the human insuiin receptor. I Bi01 Chem268(22):166lO-22
Kasuga Zick Y, Büthe DL, Cxettazw Kahn CR (1982) h s u h dinulates tyrosine phosphorylationof the insuün receptor in a ceii-frre system, N- 298(5875):667-9
Keegan AD, N e h K, White ib& Wang LU, Pierce JH, Paul WE. (1994) An L G 4 feceptor region containing an insulin receptor motif îs important for IL4medïated IRS-1 phosphorylation and celi growth. Ceii 76(5):811-20
Kahn CR (1978) Insulinresisîance, iasPlin insensitivity, and insulîn ulltesponsivmess: a necessary distinction. MetaboliSm 27(12 Suppl2):1893-902
Kahn CR (1994) Banting Lecture. riisullli action, diabetogenes, and the cause oftype II diabetes. Diabetes 43(8):1û66-84
Rahn CR, White MF. (1988) The insulin receptor and the molecuïar mechanism o f insulin action. J CIin Invest 82(4): 1 151-6
Kjeldsen T, Andersen AS, Wïberg FC, Rasmussen JS, Schaffer L, BalSchmidt P, M o k KB, Moller NP. (1991) The ligand specificities of the hdh receptor and the insulin- like growth htor 1 receptor reside in Hèrent regions ofa cornmon binding site. Roc Nati Acad Sci U S A 88(10):4404-8
Kjeldsen T, Wiberg FCy Andersen AS. (1994) Chimerïc receptors indicaîe that phenylaianine 39 is a major contributor to insulin specificity of the insulin receptor. J Biol Chem 269(52):32942-6
Kobilka B. (1992) Adrenergic receptors as models for G pmtein-coupled receptors. Annu Rev Nanosci LSr87-114
Kohanski RA. (1993) Insulin receptor autophosphorylatio~~ II. Iletennination of autophosphoryIation sites by chemid sequence analysis and identification of the juxtamembrane sites. Biochemistry 32(22):5773-80
Kruger P, GiIge G, Cabuk Y, WoIlmer A. (1990) Cooperativity and intermediate states in the T-R-structural ttansfonnation of insulin. Bi01 Chem Hoppe Seyler 37 1 (8):669-73
Kuhlbrandt W, Wang DN. Fujiyoshi Y. (1994) Atomic mode1 ofplant light-harvesting complex by electmn crystailography. N m 367(6464):6lM 1

- 7
...,
Kurose T, PaPhmforoush Yosbùaasa Y, Carn,li R, Schwartz GP, Burke GT, Katsoyannis PCf. Stcina DF. (lm) Cross-[iiiLingofa B25 azidophenytrilsnine msulin derivative to the c a r b o x y L ~ m o n ofthe dpha-subunit of the insulh mxptor. Identification ofa new ïnsdbbindllig domain in the msiilin cecepfor- 1 Bi01 Chan 269(-329190-7
L a d UK, (1970) C 1 e a v a g e o f ~ proteins duhg the assemblyofthe head of bacteriophage T4. Naturt m259):68û-5
Lammers R, Van Obberghen E, Ballotti R, Schtessinger J, Ullrich A. (1990) Tramphosphorylation as a possible mechanhm for insulin and epidennal growth nictor receptor activation. J Bi01 Chem 265(28):16886-90
Leahy DI, HendiÏckson WA, AuWI & Erickson HP. (1992) Stmcture of a f i b r o n d type III domriin h m tenascin phased by MAD anaiysis ofthe seIenomethiony1 ppr0tei.n. Science 258(5084):98%91
Lee J and Piich PF. (1994) The iiisulinreceptor= stmctum, hction, and signaiing. Am I Physiol26q2 Pt l):C3 19-34
Lee J, 0- T, P M PF, Shoelsw SE. (1993) Iiisulin receptor autophosphorylation occm asymmeûically~ J Bi01 Chem 268(6):4092-8
Lemmon MA, Bu 2, Ladbury JE, Zhou M, Pinchasi D, Lax 1, EngeIrnan DU, Schlesshger J- (1997) Two EGF moIecules conûibute additively to stabilhtion ofthe EGFR dimer- EMBO J 16(2)281-94
LeRoith D, Werner H, Burguera B, Roberts CT Jr, Mulroney S, Haramati A. (1992) The insulin-like gn,wth fiictor family of peptides, binding proteins and receptots: their potentiai role in tissue regeneration. Adv Exp Mi Bi01 321:21-8; discussion 29-30
LeRoith D, Sampson PC, Roberts CT Ji. (1994) How does the mitogenic insulin-like growth factor 1 rezeptor M e r b m the metabolic insuiin receptor? Horm Res 41 Suppl 2:74-8
Luo RZ, Beniac DR, Fernandes A, Yip CC, Ottetlsmeyet FP. 1999 Quatemary structure of the insulin-insuiin mceptor complex. Science 285(543O): 1077-80
Macaulay SL, Polites M, Hewish DR, W d CW. (1994) Cysteine-524 is not the only residue involvecl in the forniaton of disulphide-bonded dimers of the iasului receptot. Biochem J 303 (Pt 2):575-81
Maddux BA, Goldnne ID. (1991) Evidence that insulin p1us ATP may induce a confonnaîional change in the beta subunit of the insulin receptor without inducing receptor autophosphorylation. J Bi01 Chem 266(11):673 1-6

~ B u s l j e C, Mlzuguchi K, Siddk BlundelI TL. (1998) A third fibmncctii~type III domain in tht exîraceiIuIarregion ofthe insului reœptor -yC FEBS Lett 441(2):33 1-6
hbsagw i, Piich PF, Czech MP. (I980) Electrophonetic resolution ofthrœ -or uisuün receptor stnrcbacs with uni- subunit stoichiometrieses Proc Nati Acad Sci U S A 77(l2) 9 1 37-4
M&sague I and Czech MP, (1982) The subunit stnrturrs oftwo distinct receptors for ïasuh-Iike growth fàctors 1 and II and th& relationship to the insuiin receptor- l Biol Chem 257(9):503845
McKeon C and Pham T. (1991) T d v a t i o n of the himien insulinreceptor gme by the CAAT/eahancei. binding protein. Bîochem Biophys Res Coa3mun I7*2):721-8
Mïrmira RG, Nakagawa S& Tager HS. (1991) importance of the character and configuration ofresidues B24, B25, and B26 in insulin-qtor interactions. 1 Bi01 Chan 266(3): 1428-3 6
Mirmira RG and Tager HS. (1989) Rok ofthe phenylalanine B24 side chah in dirrcting insuiin interaction with its xeceptor. Itnportance ofmainchah confodon. J Bi01 Chem 264(29): 17613
Mosthaf L, Vogt B, Haring HU, U1Irich A. (1991) Altered expression of insufin zeceptor types A and B in the siceletal muscle of non.-insuiin-dépendent diabetes meilitus patients. Roc Natl Acad Sci U S A880 1):4728-30
Mulhem TD, Booker GW (1998) A third fibronectin-typee-I domah in the insulin- famiiy receptors. Trends Biochem Sci 23(12):4656
Muller-Wieland D, Taub R, Tewari DS, Kriauciunas KM, Sethu S, Reddy K, Kahn CR (1989) Insuiin-receptor gene and its expression in patients with insuiin resistance- Diabetes 38(1):3 1-8
Mumon PJ, Rodbard D. (1980) Ligand: a versatile wmputerized approach for cbaraçterization of Ligand-binding systems. Anal Biochem 107(1):220-39
Murray-Rust J, McLeod AN. Blundeli TL, Wood SP. (1992) Structure and evolution of insulins: impIications for zeceptor bï~lding. Biocssays 14(5):325-3 1
Myers MG, Backer JM, Siddle K, White MF. (1991) The insuiin receptot functions normally in Chinese hamster ovary celis afkr -cation of the C terminusus J Bi01 Chem 266(L6): 10616-23

My- MGJr, Sun=, Cbesthsm B. JachnaBR, G l n s h a n w B a c h JU. White MF. (1993) IRS-1 is a common dement in hdih and ïnsdbIiIrepwth fhctor-1 signahg to the phospMdylinositol 3'-khase- EndocrinoIogy I32(4):1421-30
Myers MG Ir, Sun=, White MF. (1994) The IRS-L signsling system- Tmds Biochem Sci 19(n:289-93
Mynarcik DC, Yu GQ, Whittaker L (1996) A l a n i n e d g mutagenesis of a C- tgminai iigand buding domah of the uisuün teceptor alpha subunit I Bi01 Chem 27 1 (5):243942
Mynarcik DC, Williams PF, Scha&t L, Yu GQ, Wbi#akerL (1997) Identification of cornmon ligaud binding detemhnts ofthe insulin and -& p w t h firtor 1 receptors- Iiisights into mechanisms of ligaud binding- J Bi01 Chem 272(3O):l86SO-S
Nakagawa SH a d Tager EIS. (1986) Role ofthe phenylalanine B25 side chah in directhg insuiin interaction with its receptor. Stenc and w d i o d o d effocts. J Bi01 Chem 261(16):733241
Nakagawa SH and Tager HS. (1987) Rok of the COOH-termhaL B-chain domain in insulin receptor interactions. Identi£icahCon of perturbations involving th insulin mainchain. J Bi01 Chem 262(25):12054-8
Nakagawa S H and Tager HS. (1993) hportance of mah-chain flexibility and the uwlin fold in insulin-recepf~r interactions. Biochemistry 32(28):723743
Ng DS, Yîp CC. (1985) Peptide mapping of the iasulia-binding site of the 130-kDa subunit of the insulinreceptor by means ofa novel cleavabIe radioactive photoprobe. Biochem Biophys Res Commun l33(l): lH-6û
Nishida E, Gotoh Y. (1993) The MAP kinase cascade U essential for diverse signal transduction pathways. Trends Biochem Sci l8(4):128-3 1
O'Bryan JP, Frye RA, Cogswell PC, Neubauer A, Kitch B, Pmkop C, Espinosa R 3 4 Le Beau MM, Earp EIS, Liu ET. (1991) ad, a tnudionning geme isolateci k m primary human myeloid leukemia celis, encodes a novel receptor tyrosine kinase. Mol Celi Bi01 1 1(10):SOl6-3 1
Olefsky JM. (198 1) LIUy lecture 1980, Insulin resis&nce and insulin action. An in vitro and in vivo perspective. Diabetes 30(2):148-62
O'Neili TI, Craparo A, Gustafson TA. (1994) Chatacterization of an interaction between insuiin receptor &strate 1 ând the insuiin receptor by using the two-hybnd ~ystem- Mol CeIl Bi01 14(10):6433-42

Oison TS, BambergerW9 Lane MD. (1988) Post+aansIationaL -es in tatiary and quaternary striicture ofthe msulin pmreceptot- C o d a t i o n w i l h ~ o n offûnctior~ J Bi01 Chem 263(12):7342-51
O l s o n m KolatLar PR, OliveiraMA, Cheng- Grerem McCleUaad A, Baker TS, Rossmann MG. (i.993) Stmctine ofahuman rbinovims complexedwith its reccptor moIecule. Proc Natl Acad Sci U S A 90(2):507-11
Ottensmeyer FI?. (1982 ) Sca#ered electrons in microscopy d m i c m d y i s Science 2 l5(4532):46 1-6
Ottensmeyer FP, Czarnota, GL, Andrews DW and Farrow Nk (1994) Proceedings 1 3 ~ Int Congress on EIectron Microscopy, 509-510
Pasqyle EB. (199 1) Identifscation of chicken embryo kinase 5, a developmentally regtdated receptor-type tyrosine kinase of the Eph M y - CelI Regui 2(7):523-34
Paui Ji, Tavare Jy Daiton RiH, Steiner DF. (1990) B$C010virusdkcted expression of the human insulin xeceptor and an insuiin-indicihg ectodornain. J Biol Chem 265(22): 13074- 83
Paredes a Heidner I& Thuman-Cornmilce P, Prasad BVy Jobnston RE, Chiu W. (1998) Structurai localization ofthe E3 giycoprotein in aüenuaîed Sindbis virus mutants. J V h l 72(2): 1534-4 L
Pawson T and Gish GD, (1992) SH.2 and SH3 domliins= h m stmctme to fiinction. Cell 71 (3):3 59-62
Pawson T, (1995) Protein modules and signalhg networks. Nahue 373(6S l5):573-80
Pelicci Gy Ladimcone L, Grignani Fy McGlade Jy C a 4 0 Fy Fomi G, Nicoletti 1, Grignani Fy Pawson T, Pelicci PG. (1992) A novel transfonning protein (SHC) with an SH2 domain is irnpLicated in mitogenic signal transduction. Ce11 70(l):93-104
Pilch PF and Czcch MP. (1979) Interaction of cross-iinking agents with the insulin effector system of isolated £ht tek. Covalent iinkage of 12!X-insuiin to a plasma membrane receptor protein of 140,000 daItons, 1 Bi01 Chem 254(9):3375-8 1
Pilch PF and Czcb MP. (1980) Hormone binding alters the conformation ofthe insulin receptor. Science XO(4474): 1152-3
Porte D Jr, (1991) Banting lecture 1990- Beta-cells in type 11 diabetes meIlitus. Diabetes 40(2): 166.80
Pronk GJ, McGlade J, Pelicci Gy Pawson T, Bos Ji,. (1993) hsuh-induced phosphorylation ofthe 46- and 52-kDa Shc proteins. J Bi01 Chem 268(8):5748-53

Rademacherhil, Wagenlrnecht T, Grassucci& E h I, Inin -ckC, Flekher S. (1992) Crgo-EM of the native stmeturt ofthe calcium nlease chatmei/Ïyanodine receptor fbm s a r c o p ~ c reticulum, Biophys 1 61(4):9393640
RafaeIoff R Patd R, Yip C, GoldheID, Hawlqr DM- (1989) Mutation ofthe high cysteine -on ofthe hucnan insuiin recepbr alpha-subuait in- iaoaün zeceptor binding affkïty and transmembrane signaüng. l Bi01 Chem 2w27):lS9W
Roach P, Zick Y, Formisano P, AcciIi D, Taylor SI, Gorden P. (1994) A novel human insuiin receptor gene mutatiDon uniqucly inhibits hdh binding without mipairing posttranslationai processing. Diabetes 43(9):1096-102
Rosenberg MF, Cakghau R, Ford RC, =gghs CF. (1997) Sîmcîure of the mdtidrug resista~ce P-giycoprotein to 2.5 nm resoIution detennined by eledron ~nicroscopy and image d y s i s . J Biol Chem 272(16):10685-94
Roth RA, Casseli DJ, Wong KY, Maddux BA, Goldfine ID. (1982) Monoclonaï anti'bodies to the human insulinreceptor block iapuluibinding and inhiit insuiin action Proc Nat1 Acad Sci U S A 79(23):73 12-6
Sasaoka T, Takata Y, K d J, Anderson CU. Langlois WJ, Olefkky Jh+l. (1993) Transmembrane signaLing by an insulin receptor lacking a cytopIasmic beta-subunit domain, Prcx: Nati Acad Sci U S A 90(10):4379-83
Schwabe C, Bullesbach EE. (1994) Re1ax.k skuctutes, fûnctions, promises, and nonevolution FASEB J 8(14):1152-60
Seino S, Bell GI. (1989) Alternative splicing of human insulin receptor messenger RNA. Biochem Biophys Res Commun lS(l):3 12-6
Seino S, Seino U, Bell GI. (1990) Humen insulin-receptor gene. Diabetes 39(2): 129-33
Schaefer EU, Siddie K, EUis L. (1990) Deletion analysis ofthe human iasulin roceptor ecudornain reveais independietly folded soluble subdomauis and insulin binding by a monomeric alpha-subunit, J Bi01 Chem 26S(22): 13248-53
Schaefer EU, Erickson HP, Federwisch U, Wollmer A, Ellis L. (1992) Structurai organization of the human insulin receptor ectodomain_ J Biol Chem 267(32):23393-402
Schaffer L, Ljungqvist L. (1992) Identification of a disulfide bridge c01111ecting the alpha- subunits of the extracellular domain of the ùwlin receptor- Biochern Biophys Res Commun 189(2):650-3
SchafEer L, Kjeldsen T, Andersen AS, Wiberg FC, Larsen UD, Cera JF, Mimllra RG, Nakagawa SH, Tager HS. (1993) Interactions of a hybrid i n s u W i i - l i k e p w t h

factor-1 analog with cbimen'c ùisnIinltype 1 -me growth fàctot receptors. J Biol Chem 268(5):3044-7
Scaaffer L (1994 ) A mode1 for insrilin b i i to the irisuün nceptor. Eur J Biochem 221(3):1127-32
Schrag JD, Prasad BV, Rixon FJ, J,u W. (1989) Thtee-dimensÎonaI stnrturr ofthe HSVl nucleocapsid, Ce11 S6(4):65 160
Schumacher R, Soos MA, Schlessinger I, Brandenburg D, Siddle K, Ullrich A. (1993) Signaling-competent receptor chimeras d o w mapping ofmqCot insulin receptor binding domaiu determinants, J Bi01 Chem 268(2):108?-94
Serysheva II, OrIova EV, Chiu W, Sherman MB, Hamilton SL, van Heei M. (1995) Electron cryomicrosoopy and anguiar reconstitution d to visualize the ske1e~al muscle calcium release channel. Nat Struct Biol2(1):18-24
Shaw AL, Samai SK, Subramanian K, Rasad BV. (1996) The stnicturr o f aquareovirus shows how the mirent geometries of the two layers of the capsid are reconciled to provide symmetrical interactions and s tab i ion . Stnicture 4(8):95747
Skolnik EY, BatPr A, Li N, Lee CH2, Lowenstein E, Mohammadi Y Uargolis B. Schlessinger JO (1993) The fiinctionofGRB2 in linking the insulin receptor to Ras signaiing path-YS. Science 26OG 1 16): 1953-5
Sciacca L, Costantino A, Pandini G, Mine0 R, Frasca F, Scalia P, Sbraccia P, Goldfine ID, Vigneri R, Beffiore A. (1999) Insulin receptor activation by IGF-II in breast cancers: evidence for a new autocrindparacrine mechaism. Oncogene 18(15):2471-9
Shalev A (1999) Hope for insuün mimetic orai antidiabetic dnigs. Eut J Endocrino1 141(6):561-2
Shier P, Watt W. (1989) Primary structure of aputative teceptor for a ligand of the insuiin farnily- J Bi01 Chem S64(25): 14605-8
Shoelson SE, White MF, Kahn CR (1988) Tryptic activation of the insuiin receptor0 Proteolytic tnincation of the alpha-subunit releases the beta-subunit h m inhibitory control. J Bi01 Chem 263(10):4852-60
Shoelson SE, Lee I, Lynch CS, Backer Pilch PF. (1993) BpaE525 iasulins. Photoactivaiable anaiogues that qmtitatively cross-liok, radiolabel, and activaie the insulin recepttor. J Bi01 Chem 268(6):M85-9 1
Siddle K, Soos MA, Field CE, Nave BT. (1994) Hybrid and mical inml in / ' i - l ike growth factor 1 receptors. Horm Res Suppl2:5664; discussion 65

SisSom J, Ellis L.(1989) Secretion ofthe extrsce1IuIar domsin of the human insuiin receptor fimm insect ceils by use ofa b a c a l o ~ vector- Biochem J 261(1):119-26
S ~ W L O . M C K ~ ~ W , 00-JJ. strürtp~, ~obinsoncp, Beritlqm, wardcw. (1997 ) The disuIfide bords in the C-taminel domains ofthe human insulin receptor ectodomain_ P Biol Chem272(47):2946û-7
Staubs PA, Reichart DR, Saltiel AR, M . KL, Mkegawa H, Berhanu P, Ole- JïM, Seely BL. (1994) Localizationofthe iasulin lleceptor bindiilg sites for the SH2 domain proteins p85, Syp, and GAP. J Biol Chem 269(44)272718692
Steiner DF, Tager HS, Chan SJ, Nanjo K. Sanice T, RubenStein AH. (1990) Lessons leamed h m molecular biology of -genie mutations. Diabetes Care 13(6):600-9
Sutherland. C, Campbeli DG, Cohen P. (1993) Identification ofinsulln-stimuiated protein kinase4 as the raôbit eqivalent of rskmo-2. Identificatiion oftwo threonines phosphorylated during activation by mitogen-activated protein kinase. Euc J Biochem 212(2):58 1-8
Sun XJ, Rothenberg P, Kahn CR, Backer M, Araki E, Wdden PA, Cahill DA, Goldstein BJ, White MF. (1991) Structme of the iosulia receptor substrate IRS-1 defines a unique signal transduction protein. Nature 3 52(6330):73-7
Sun XT, Crimmins DL, Myers MG k, Miralpeix White MF. (1993) Pleiotropic insulin signals are engaged by multisite phosphorylation of IRS-1. Mol Cell Bi01 l3(12):7418- 28
Sun XT, Wang LM, Zhang Y, Yenush L, Myers MG Jr, Glasheen E, Lane WS, Pierce JET, White MF. (1995) Role ofIRS-2 in insulin and cytokhe signalIhgt Nature Sep l4;3 77(6545): 193-7
Sung CK, Wong KY, Yip CC, Hawley DnL, &Idfine ID. (1994) Deletion of tesidues 485-599 h m the human insuiin receptor aboiishes antireceptor antibody binding and influences tyrosine k h s e activation. Mo1 Endocrino1 8(3):3 15-24
Sweet W, Wilden PA, Pessin JE. (1986) Dithiothreitol activation of the insulin recpqtor/kinase does not involve subunit dissociation ofthe native alpha 2 beta 2 insulin receptor subunit cornplex. Biochemistry 25(22):7068-74
Sweet W, Momïson BD, Wilden PA, Pessin JE. (1987 ) Insulin-dependent intennoleculat subunit co~l l l~l~cat ion between isolated alpha beta heterodimeric iilsulin receptor complexes. J Bi01 Chem 262(34): 16730-8
Tager Ii, Thomas N, Assoian R, Rubenstein A, Saekow M, Olefiiq J, Kaiser ET. (1980) Semïsynthesis and biological activity of porcine ~uB24]hsuih and ~uB25]insulin. Proc Nat1 A d Sci U S A 77(6):3 181-5

Taouis U. Levy-Toledam R, Roaeh P, Taylor SI9 Go* P. (1994) Rescur d activation of a b ~ d e f i c i e n t ~ r e c e p t o r . Evidence for inferm01ecular transphospharylation. J Bioi Chem 269(44.27762-6
Tavare JM, Siddle K. (1993) Mutaficonai analysls of iasalin receptor fhctio11: consensus and wntn,versyerSy Biochim Biophys Acta 1 I78(l):X-39
Taylor R (199 1) Insulin action 199 1, Clin EndocrinoL (OxQ Fe6;34(2):159-71
Taylor SI, Cama A, Accili D, Barbetîi F, Quon UT, de la Luz Sicrra M, Suzuici Y, Kolier E, Levy-Toledano R, Wertheimer E, et al. (1992) Mutatiotls in the insulul receptor gene. End- Rev 13(3):566-95
Taouis n& Levy-Toledano R, RoachP, Taylor SI, Goden P. (1994) Rescue and activation of a binding-deficient insuün receptor. Evidence for intermolecular transphosphorylaîion. 3 Bi01 Chem 269(44):277626
Tanasijevic MJ. My- MG Ir, ThomaRS, Crimmins DL, White MF, Sa& DB. (1993) Phosphorylation ofthe insuiin receptor substrate IRS-1 by casein kinase II, J Bi01 Chem 268(24): 18 157456
Trus BL, Roden RB, Greenstone HL, VrheL hiZ, Schiller JT, Bwy FP. (1997) NoveL structural fe- of bovine papillomavirus capsid revealed by a threedimensional reconstruction to 9 A resolution. Nat Stmct Bi01 4(5):413-20
Turner R Hattersley A, Cook J. (1992) Type II diabetes: search for primary defects. AM Med 24(6):511-6
Ulirich A, Berman CH, Duil TJ, Gray A, Lee JU (1984) IsoIation of the human insulin- like growth factor 1 gene using a single synthetic DNA probe- EMBO J 3(2):3614
Ullrich A, Bell JR, Chen EY, Haraa R, Petruzzelii LlM, h31 TJy Gray A, Coussens L, Liao YC, Tsubokawa M, et al. 0985) Human insulin receptor and its relationship to the tyrosine kinase family ofoncogenes. Naîure 3 13(6005):75641
Ullrich A, Gray A, Tarn AW, Yang-Feng T, Tsubokawa M. Collins C, Heozel W, L+e Bon T, Kathuria S, Chen E, a al. (1986) hdin-like p w t h factor 1 receptor primary stnictue: cornparison with insulin receptor suggests stnictural detcrmhnts that define fbctionai specificity. EMBO J 5(10):2503-12
Uiirïch A, Schlessinger 1. (1990) Signal transduction by teceptors with tyrosine kinase activity. Ce11 61(2):203-12
Unwin N. 1995 Acetylcholine receptor charme1 imagecl in the open state. Nsture 373(6509):3743

Unwîn N. 1998 The nicotinïc acetylcholine receptor ofthe Torpedo e l d c ray. J Struct Bi01 121(2):181-90
Van Horn DI, Myers MG Jr, B a c k MW (L994) D k c t activation ofthe phosphatidyhositoL 3'-kinase by the uwlin receptor- I Bi01 Chem 269(1):29-32
van der Geer P, Himter T, Lindberg RA.( 1994) Receptor protein-tyrosine kinases aud th& signal transduction padiways. Annu Rev CeU Bi01 10251-337
Van Hed U. (1981) Angularreeonstruai*~~ a posteriori assigrment ofprojection directions for 3D reconshuctîon. Ultramicroscopy 21:l L 1-24
Wagenknecht T, Berkowitz J, Grasma5 R, Tirneman Al?, Flekher S. (1994) Locaüzabion of caImodulin binding sites on the ryanodine receptOr h m ske1etal muscle by electron micmscopy~ Biophys 1 67(6):228695
Wak T, Hirai T, Murata K, H e m JB, Mitsuoka K. Fujiyoshi Y, S m . BL. Agre P, Engel k(1997) The thxeedimensioaal stnrtint ofaqriaip~rin-1. Nature 387@33):624-7
Ward CW. (1999) Manbers of the iasulin receptor fimily contain tbree fibronectin type rn do-. ~rowth F$IC~OZS i6(4)3 15-22
Waugh S M DiBeïïa EE, Pilch PF. (1989) IsoIation of a proteoIyticaliy derived domain of the iosulin receptor containhg the major site of cross-linkinglbinding. Biochemistry 28(8):3448-55
Wedekind F, Baer-Pontzen K, Bala-Mohan S, Choli D, Zahn H, Bdenburg D. (1989 ) Honnone binding site of the insuiin receptor: anaiysis using photoaffinity-mediated avidin compleràng. Bi01 Chem Hoppe Seyler 370(3):251-8
Wells IA (1996) Hommne mimicry. Science 273(5274):449-50
White MF. (1994) The IRS-1 signaling system. Curr Opin Genet Dev 4(1):47-54
White MF. (1996) The IRS-signalhg system in insuün and cytokine action. Philos Trans R Soc Lond B Bi01 Sci 3Sl(l336):18 1-9
White MF, Kahn CR (1994) The insulin signaüng system J Bi01 Chem 269(1):1-4
White MF. Maron R, Kahn CR. (1985) Insuiin rapidy stimulaies tyrosine phosphoryIation o f a Mt-185.000 protein in intact ceUs. Nature 3 l8(6042): 183-6
White W, Livingston JN, BackaJM, Laiiris V, Dull TJ, W c h A, Kahn C R (1988) Mutation ofthe insului receptor at tyrosine 96û inhi'bits signal transmision but does not affect its tyrosine kinase activity. Celi 54<5):641-9

White MFy Kahn CR (1994) The uwlin signaihg system. J Bi01 Chrm 269(1):14
Whittaker U. WilsonXubalek Eh& Smith JE, Faust L, Milligan RA, Sweeney HC. (1995) A 35-A movement ofsmooth rnusdemyosin on ADP release. N e 378(6558):748-51
Wilden PA, Kahn CR, Siddle K, White MF. (1992) îkdin receptot Liasse domain sutophosphoryIation reguiates receptor enymatic hction J Biol Chem 267(23):16660- 8
Wdden PA, Boyle TR, Swanson ML, Sweeî W; Pessin JE- (1986 ) Alteration of - intramolecuIar disulfides in insulin receptor/kinase by msalin and dit)llCothreitol: insulin potentiates the apparent ditfiIcothreito1depencfent subimit redttction ofinsaIul receptor. BiochemiShy 2!j(lS):438 1-8
whittaker J, Okamoto AK, Thys R, Bell GI, Steiner DFy H o i Ï n a ~ CA. (1987 ) High- level expression ofhuman iasulin zeceptor cDNA in mouseNIH 313 ails. Roc Natl Acad Sci U S A 84(15):523741
Whittaker J, Okamoto k (1988) Secdon ofsoluble fuactional insuiin receptors by transfected NIEZ3T3 cds. J Bi01 Chem 263(7):3063-6
Wfiams JF, McClain DA, hill TI, Ullrich A, Olefslry M. (1990) -on of an insulin receptor mutant lacking the subunit p~ocessing site. J Bi01 Chem 265(15):8463-9
WiUrinson DA, Marion TN, Tiiiman Dw Norcum MT, H i e I d JF, Seyer 1M, Carlson GM. (1992) An epitope proximal to the carboxyl terminus of the alpha-subunit is locetcd near the lobe tips of the phosph~ry~ase kinase hexadecamer- J Mol Biol 23 5(3):974-82
Wiuiams PF, Myiiarcüc DC, Yu GQ, Whittaker J. (1995) Mapping of an NH2-terminal ligand binding site of the insulin receptor by alanine scanning mutagenesis. J Bi01 Chem 27O(î):3O 12-6
Yang-Feng TL, Francke U, U ' c h k (1985) Gene for human insulin receptor: localization to site on chromosome 19 involved in prr-B-ceU ledcemia Science 228(4700):728-3 1
Yamada K, Goncalves E, Kahn CR, Shoelson SE- (1992) Substitution of the insulin receptor transmeembrane domain with the c-neulerbB2 transmembrane domain constitutively activates the insulin receptor kinase in vitro. J Bi01 Chem 26ï(l8): 12452- 61

YamauchiK, PessinIE (1994) riisulinreceptor-1 ml) d S h e compete for a limited pool of Grb2 in mcdiatuig hdiit do- sigdhg. J Biol Chem 269(?9):3 1 107-14
Yamagrichi Y, Flier JS. Yokota 4 Benecke & Baekerm Moiler DE. (1991) Fundional properties oftwo nahinlly occming isofomw ofthehumaninsului reoeptor in Chinese hamster ovsry cells. EnQcanol~gy 129(4):205866
Yeung CW, Mode ML, Yi CC. (L980) Photoaffinity Labelhg ofmsUün rraptor with an insuiin d o g u e seIectively modified at the amho terminai ofthe B chain. BiochmiWtry 19(10):21%-203
Yip CC. (1991) Iosuiùi receptor, aspects of its stnicturr and fiinction. Adv Exp Med BioL - 3 3&79-8&
Yip CC- (1992) The insulin-binding domain of iasulin q t o r is emcoded by exon 2 and exon 3. J CeU Biochem 48(1):19-25
Yip CC. (1993) The -binding domain of insuiin receptor is encodeci by exon 2 and exon 3. J Ce11 Biochem 1992 48(1):19-25 Yip CC, Yeung CW, MouleML (1978)-PhotoaEnity labeiingofinsuiin receptot of rat adiopocyte plasma membrane- J Bi01 Chem 253(6):1743-5
Yip CC, Yewig CW, Moule ML (1980) Photoaffbity iabeling of insuiin receptor proteins
Yip CC, Mode ML, Yeung CW. (1982) Subunit stnicdure of iiisulin receptor ofrat adipocytes as demoIlSftafed by photoafEnity labeiing. Biochemistry 21(12):2940-5
Yip CC, Hsu EE, Patel RG, Hawtey Dw Uaddux BA, Goldfine ID. (1988) Localization of the insulin-b'iding site to the cysteke-rich region ofthe insulin receptor alpha- subunit Biochem Biophys Res Commun lS7(l)32 1-9
Yip CC, Gninfeld C, Goldfïne ID. (1991) Identification and characterization of the ligand-binding domain of insulùl receptor by use of an anti-peptide antisenim agahst amino acid sequence 241-25 1 of the alpha subuait Biochemistry 30(3):695-701
Yip CC, Jack E. (1992) Insulin receptors are bivalent as d e m o u by photoafhity iabeling. J Biol Chem 267(f 9): 13 13 1-4
Yonezawa K, Roth U (1991) Assessment of the in situ tyrosine kinase activity of mutant insuün receptors lacking tyrosine autophosphorylation sites 1162 and 1 163. Mol Endocrinol S(2): L94-2ûû

Zhang B, Saüturo G. Szallrowski D, Li 2, Zhang Y, Royo I, VileIlaD, D i a MT, PeIaez F,RubyC.KendalLRL,Mao~GrifnnP,CalaycayI,ZiaathJR,HodcN,SmithRG, Molk DE, (1999) Discovery of a s d moiecule insulin mimetk wïth 81itidEabetttc activity in mice- Science 284(5416):974-7
Zhou ZtF, He J, JeLana J, Tatman JD, Rixon FI, Chiu W. (1995) AssembLy of VP26 in herpes simp1ex ~~IUS-1 Inferred fkm stnrctans ofwild-typ and recomb'tnzult apsids. Nat Struct Bi01 2(11): 1026-30
Zhou ZH, Prasad BV, Jàkana J, Rixon FJ, Chni W- (1994) Ekotein subunit stnictutes in the herpes simplex vinis A q s i d detemiined h m 400 kV spot-scan el-n cryomicrosc~py. J Mol Bi01 242(4):456-69
Zhou ZI!& M;acnab SJ, Jakana J, Scott LR, Chiu W, Rixon FJ. (1998) Identification of the sites of interaction between the d o i d and outer Shen in éerpes simplex vims-1 capsids by difference electron imaging- Roc Nat1 Acad Sci U S A 95(6)2778-83