quaternary science reviews€¦ · the role of climate in the spread of modern humans into europe...
TRANSCRIPT

lable at ScienceDirect
Quaternary Science Reviews 30 (2011) 273e279
Contents lists avai
Quaternary Science Reviews
journal homepage: www.elsevier .com/locate/quascirev
The role of climate in the spread of modern humans into Europe
Ulrich C. Müller a,*, Jörg Pross a, Polychronis C. Tzedakis b,c, Clive Gamble d, Ulrich Kotthoff a,e,Gerhard Schmiedl e, Sabine Wulf f,g, Kimon Christanis h
a Institute of Geosciences, Goethe University Frankfurt, 60438 Frankfurt, GermanybDepartment of Geography, University College London, London WC1E 6BT, UKcDepartment of Environment, University of the Aegean, Mytilene 81100, GreecedDepartment of Geography, Royal Holloway, Egham, Surrey TW20 0EX, UKeGeological-Palaeontological Institute, University of Hamburg, 20146 Hamburg, Germanyf Institute for Geophysics, University of Texas at Austin, Austin TX78758, USAgDeutsches GeoForschungsZentrum GFZ, Sektion 5.2, 14473 Potsdam, GermanyhDepartment of Geology, University of Patras, 26500 Rio-Patras, Greece
a r t i c l e i n f o
Article history:Received 16 June 2010Received in revised form17 November 2010Accepted 19 November 2010Available online 15 December 2010
* Corresponding author. Tel.: þ49 69 798 40176; faE-mail address: [email protected]
0277-3791/$ e see front matter � 2010 Elsevier Ltd.doi:10.1016/j.quascirev.2010.11.016
a b s t r a c t
The spread of anatomically modern humans (AMH) into Europe occurred when shifts in the NorthAtlantic meridional overturning circulation triggered a series of large and abrupt climate changes duringthe last glacial. However, the role of climate forcing in this process has remained unclear. Here wepresent a last glacial record that provides insight into climate-related environmental shifts in the easternMediterranean region, i.e. the gateway for the colonisation of Europe by AMH. We show that the envi-ronmental impact of the Heinrich Event H5 climatic deterioration c. 48 kyr ago was as extreme as that ofthe glacial maximum of Marine Isotope Stage (MIS) 4 when most of Europe was deserted by Neander-thals. We argue that Heinrich H5 resulted in a similar demographic vacuum so that invasive AMHpopulations had the opportunity to spread into Europe and occupy large parts before the Neanderthalswere able to reoccupy this territory. This spread followed the resumption of the Atlantic meridionaloverturning circulation at the beginning of Greenland Interstadial (GIS) 12 c. 47 kyr ago that triggered anextreme and rapid shift from desert-steppe to open woodland biomes in the gateway to Europe. Weconclude that the extreme environmental impact of Heinrich H5 within a situation of competitiveexclusion between two closely related hominids species shifted the balance in favour of modern humans.
� 2010 Elsevier Ltd. All rights reserved.
1. Introduction
Located in an intermediate position between the higher-latitude(i.e., influenced by the westerlies) and lower-latitude (i.e., mon-soonally influenced) climate systems, the eastern Mediterraneanregion is highly sensitive to short-term climate variability. Thissensitivity is well documented for the early to fully developedinterglacial state of the Holocene (Rohling et al., 2002; Kotthoffet al., 2008a). The influence of short-term climatic forcing duringthat time is known to have affected late prehistoric to early historiccultures in the eastern Mediterranean region (Weiss et al., 1993;Pross et al., 2009). Markedly less information is available on thepotential role of climate forcing on human population dynamicsduring the last glacial, notably the dispersal of AMH fromAfrica andtheir subsequent colonisation of Eurasia. With regard to the
x: þ49 69 798 40185.e (U.C. Müller).
All rights reserved.
immigration of AMH into Europe, the easternMediterranean regionis of particular interest because it served as a gateway in thisprocess (Fig. 1). The archaeological evidence and the preference formigration routes along coastlines and rivers have shown that AMHused the south-eastern gateway to colonise Europe (Bar-Yosef,2002; van Andel et al., 2003; Mellars, 2006). The scarcity of suffi-ciently resolved, chronologically well constrained terrestrialclimate data from that region has, however, precluded insights intopotential relationships between climatic forcing and the migrationof AMH into Europe. In order to identify the environmentalconditions under which AMH migrated into Europe, we havegenerated a well-dated, centennial-scale-resolution pollen recordspanning MIS 2e4 based on a new core from the classical site ofTenaghi Philippon (Supplementary information) in NE Greece(Fig. 1). Given the proximity to glacial refugia of temperate plants(Tzedakis et al., 2002), the Tenaghi Philippon archive is ideallysuited to record abrupt climate improvements during glacials sincethe vegetation response to climatic forcing occurred withoutsignificant migration lags.

Fig. 1. Migration route of modern humans from Africa into Europe when sea-level was70 m below present (brown coastline) as realized at 48 kyr BP (Siddall et al., 2003)during early MIS 3. Black dots mark palaeoanthropological sites (Supplementary TableS2) relevant for the migration route: BT ¼ Boker Tachtit, Ke ¼ Kebara, KA ¼ Ksar ’Aqil,Üç ¼ Üça�gızlı, BK ¼ Bacho Kiro, Te ¼ Temnata, BB ¼ Brno-Bohunice. Black dots withnumbers indicate sites with dating results (in kyr before present) relevant for thetiming of the migration process. Black squares mark relevant palaeoenvironmentalreconstruction sites: TP ¼ Tenaghi Philippon, MD9501, So ¼ Soreq, ML ¼ Megali Limni,Io ¼ Ioannina, Mo ¼ Monticchio.
Fig. 2. Stratigraphy and chronology of the core Tenaghi Philippon TP-2005. (A) SPECMAP c(Pross et al., 2007); taxa are grouped according to their climatic requirements (Fig. 3). (C)converted into calendar ages (Table 1). White squares indicate reported ages of identified tepfrom Campanian Ignimbrite eruption in southern Italy (Figs. S1, S2). Note, the MIS 4/5 bou
Table 1AMS 14C dates from Tenaghi Philippon core TP-2005 converted into calendar agesaccording Weninger and Jöris (2008).
Depth (m) 14C age (yrsBP)
Calendar age(yrs BP)
Laboratory code Material
mean std dev mean std dev
0.76 1950 30 1906 30 Poz-15890 Viviparus contectus1.79 4200 40 4743 81 Poz-15891 Oxyloma elegans3.41 5790 40 6591 53 Poz-15894 Oxyloma elegans4.20 6350 50 7297 67 Beta-244646 wood4.59 7600 50 8411 32 Beta-244647 wood5.56 8820 50 9924 154 Beta-244650 wood6.15 9890 60 11325 75 Beta-244651 wood6.98 13570 70 16574 353 Beta-244654 peat bulk7.18 16560 90 19861 306 Beta-244655 peat bulk8.20 20220 100 24167 285 Beta-244637 peat bulk8.86 23330 150 28089 201 Poz-16295 peat bulk9.30 24310 160 29067 439 Beta-244638 peat bulk9.85 25120 150 30050 251 Beta-244639 peat bulk10.60 27760 190 32317 292 Beta-244640 peat bulk11.25 28680 230 33155 417 Beta-244641 peat bulk11.85 32390 260 36926 764 Beta-244642 peat bulk13.30 35290 350 40219 886 Beta-244643 peat bulk13.83 36520 400 41625 335 Beta-244644 peat bulk14.65 39570 570 43526 633 Beta-244645 peat bulk15.28 43100 1200 46812 1842 Beta-246628 peat bulk
U.C. Müller et al. / Quaternary Science Reviews 30 (2011) 273e279274
2. Stratigraphy and chronology
The new Tenaghi Philippon core (40�5802400 N, 24�1302600 E,40 m asl) holds a terrestrial archive that is preserved mainly infen-peat and comprises the last c. 300 kyr BP continuously (Fig. 2).The chronology of the core interval covering MIS 1e4 is based on20 AMS 14C dates (Table 1), tephrostratigraphy, and, beyond therange of 14C dating, tuning of its pollen record to the SPECMAPcurve (Martinson et al., 1987). All age data yield an internally
hronology (Imbrie et al., 1984). (B) Overview pollen record of the 60 m core TP-2005Age-depth curve for the interval from MIS 1 to 4; Black dots indicate AMS 14C dateshra layers, i.e. Y-2 tephra derived from Cape Riva eruption on Santorini, and Y-5 tephrandary is situated at 19 m depth.
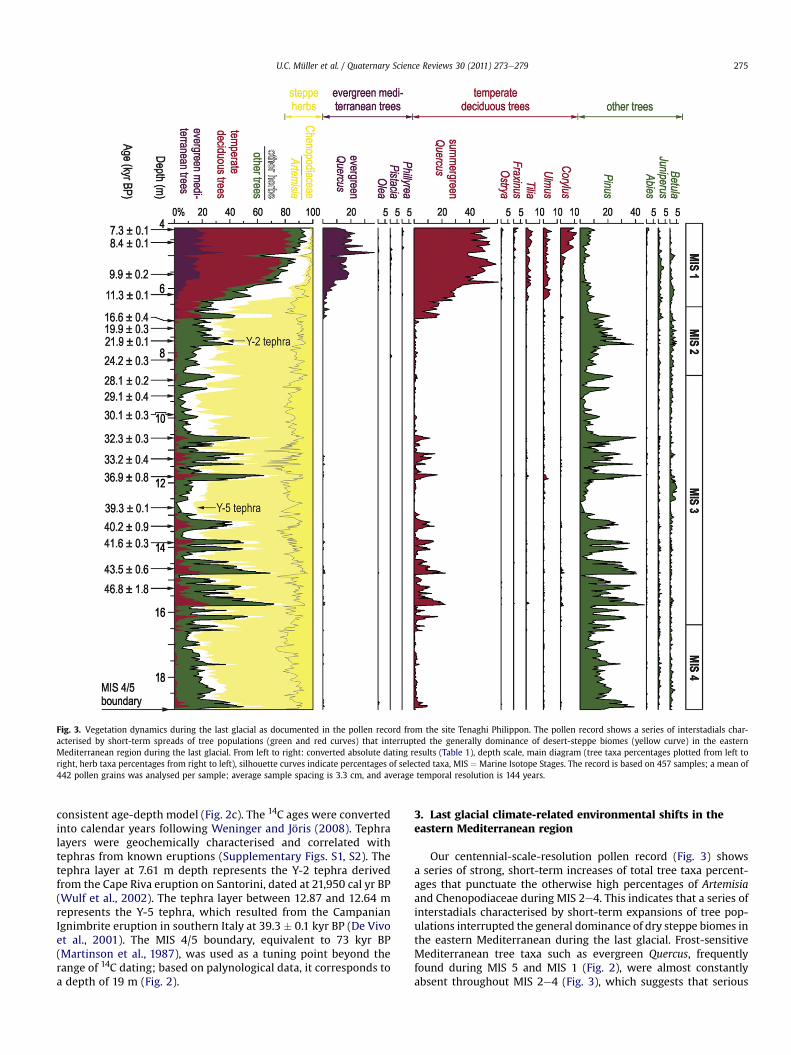
Fig. 3. Vegetation dynamics during the last glacial as documented in the pollen record from the site Tenaghi Philippon. The pollen record shows a series of interstadials char-acterised by short-term spreads of tree populations (green and red curves) that interrupted the generally dominance of desert-steppe biomes (yellow curve) in the easternMediterranean region during the last glacial. From left to right: converted absolute dating results (Table 1), depth scale, main diagram (tree taxa percentages plotted from left toright, herb taxa percentages from right to left), silhouette curves indicate percentages of selected taxa, MIS ¼ Marine Isotope Stages. The record is based on 457 samples; a mean of442 pollen grains was analysed per sample; average sample spacing is 3.3 cm, and average temporal resolution is 144 years.
U.C. Müller et al. / Quaternary Science Reviews 30 (2011) 273e279 275
consistent age-depth model (Fig. 2c). The 14C ages were convertedinto calendar years following Weninger and Jöris (2008). Tephralayers were geochemically characterised and correlated withtephras from known eruptions (Supplementary Figs. S1, S2). Thetephra layer at 7.61 m depth represents the Y-2 tephra derivedfrom the Cape Riva eruption on Santorini, dated at 21,950 cal yr BP(Wulf et al., 2002). The tephra layer between 12.87 and 12.64 mrepresents the Y-5 tephra, which resulted from the CampanianIgnimbrite eruption in southern Italy at 39.3 � 0.1 kyr BP (De Vivoet al., 2001). The MIS 4/5 boundary, equivalent to 73 kyr BP(Martinson et al., 1987), was used as a tuning point beyond therange of 14C dating; based on palynological data, it corresponds toa depth of 19 m (Fig. 2).
3. Last glacial climate-related environmental shifts in theeastern Mediterranean region
Our centennial-scale-resolution pollen record (Fig. 3) showsa series of strong, short-term increases of total tree taxa percent-ages that punctuate the otherwise high percentages of Artemisiaand Chenopodiaceae during MIS 2e4. This indicates that a series ofinterstadials characterised by short-term expansions of tree pop-ulations interrupted the general dominance of dry steppe biomes inthe eastern Mediterranean during the last glacial. Frost-sensitiveMediterranean tree taxa such as evergreen Quercus, frequentlyfound during MIS 5 and MIS 1 (Fig. 2), were almost constantlyabsent throughout MIS 2e4 (Fig. 3), which suggests that serious
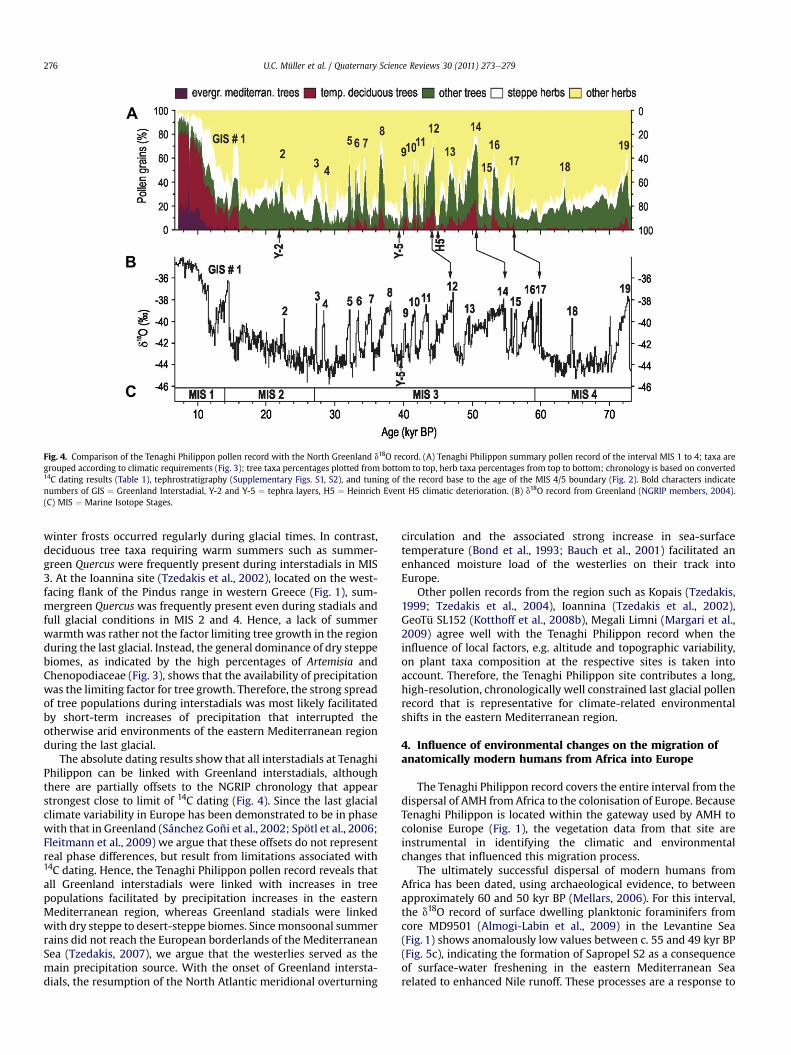
Fig. 4. Comparison of the Tenaghi Philippon pollen record with the North Greenland d18O record. (A) Tenaghi Philippon summary pollen record of the interval MIS 1 to 4; taxa aregrouped according to climatic requirements (Fig. 3); tree taxa percentages plotted from bottom to top, herb taxa percentages from top to bottom; chronology is based on converted14C dating results (Table 1), tephrostratigraphy (Supplementary Figs. S1, S2), and tuning of the record base to the age of the MIS 4/5 boundary (Fig. 2). Bold characters indicatenumbers of GIS ¼ Greenland Interstadial, Y-2 and Y-5 ¼ tephra layers, H5 ¼ Heinrich Event H5 climatic deterioration. (B) d18O record from Greenland (NGRIP members, 2004).(C) MIS ¼ Marine Isotope Stages.
U.C. Müller et al. / Quaternary Science Reviews 30 (2011) 273e279276
winter frosts occurred regularly during glacial times. In contrast,deciduous tree taxa requiring warm summers such as summer-green Quercus were frequently present during interstadials in MIS3. At the Ioannina site (Tzedakis et al., 2002), located on the west-facing flank of the Pindus range in western Greece (Fig. 1), sum-mergreen Quercus was frequently present even during stadials andfull glacial conditions in MIS 2 and 4. Hence, a lack of summerwarmth was rather not the factor limiting tree growth in the regionduring the last glacial. Instead, the general dominance of dry steppebiomes, as indicated by the high percentages of Artemisia andChenopodiaceae (Fig. 3), shows that the availability of precipitationwas the limiting factor for tree growth. Therefore, the strong spreadof tree populations during interstadials was most likely facilitatedby short-term increases of precipitation that interrupted theotherwise arid environments of the eastern Mediterranean regionduring the last glacial.
The absolute dating results show that all interstadials at TenaghiPhilippon can be linked with Greenland interstadials, althoughthere are partially offsets to the NGRIP chronology that appearstrongest close to limit of 14C dating (Fig. 4). Since the last glacialclimate variability in Europe has been demonstrated to be in phasewith that in Greenland (Sánchez Goñi et al., 2002; Spötl et al., 2006;Fleitmann et al., 2009) we argue that these offsets do not representreal phase differences, but result from limitations associated with14C dating. Hence, the Tenaghi Philippon pollen record reveals thatall Greenland interstadials were linked with increases in treepopulations facilitated by precipitation increases in the easternMediterranean region, whereas Greenland stadials were linkedwith dry steppe to desert-steppe biomes. Sincemonsoonal summerrains did not reach the European borderlands of the MediterraneanSea (Tzedakis, 2007), we argue that the westerlies served as themain precipitation source. With the onset of Greenland intersta-dials, the resumption of the North Atlantic meridional overturning
circulation and the associated strong increase in sea-surfacetemperature (Bond et al., 1993; Bauch et al., 2001) facilitated anenhanced moisture load of the westerlies on their track intoEurope.
Other pollen records from the region such as Kopais (Tzedakis,1999; Tzedakis et al., 2004), Ioannina (Tzedakis et al., 2002),GeoTü SL152 (Kotthoff et al., 2008b), Megali Limni (Margari et al.,2009) agree well with the Tenaghi Philippon record when theinfluence of local factors, e.g. altitude and topographic variability,on plant taxa composition at the respective sites is taken intoaccount. Therefore, the Tenaghi Philippon site contributes a long,high-resolution, chronologically well constrained last glacial pollenrecord that is representative for climate-related environmentalshifts in the eastern Mediterranean region.
4. Influence of environmental changes on the migration ofanatomically modern humans from Africa into Europe
The Tenaghi Philippon record covers the entire interval from thedispersal of AMH from Africa to the colonisation of Europe. BecauseTenaghi Philippon is located within the gateway used by AMH tocolonise Europe (Fig. 1), the vegetation data from that site areinstrumental in identifying the climatic and environmentalchanges that influenced this migration process.
The ultimately successful dispersal of modern humans fromAfrica has been dated, using archaeological evidence, to betweenapproximately 60 and 50 kyr BP (Mellars, 2006). For this interval,the d18O record of surface dwelling planktonic foraminifers fromcore MD9501 (Almogi-Labin et al., 2009) in the Levantine Sea(Fig. 1) shows anomalously low values between c. 55 and 49 kyr BP(Fig. 5c), indicating the formation of Sapropel S2 as a consequenceof surface-water freshening in the eastern Mediterranean Searelated to enhanced Nile runoff. These processes are a response to
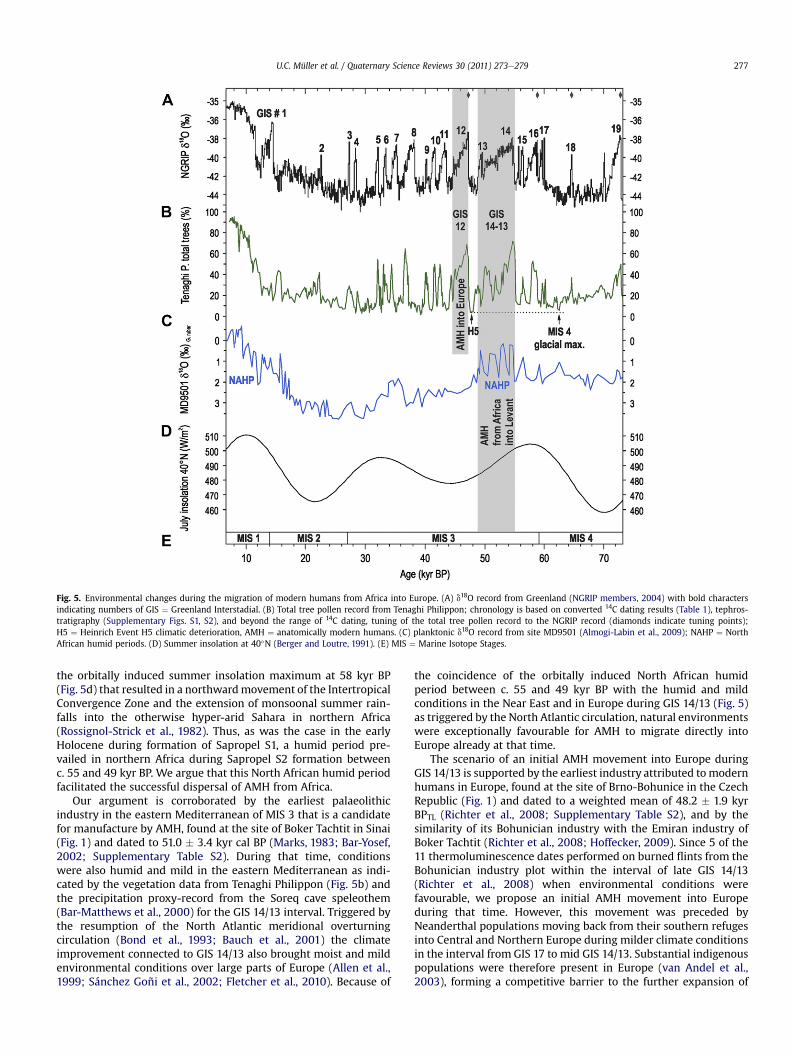
Fig. 5. Environmental changes during the migration of modern humans from Africa into Europe. (A) d18O record from Greenland (NGRIP members, 2004) with bold charactersindicating numbers of GIS ¼ Greenland Interstadial. (B) Total tree pollen record from Tenaghi Philippon; chronology is based on converted 14C dating results (Table 1), tephros-tratigraphy (Supplementary Figs. S1, S2), and beyond the range of 14C dating, tuning of the total tree pollen record to the NGRIP record (diamonds indicate tuning points);H5 ¼ Heinrich Event H5 climatic deterioration, AMH ¼ anatomically modern humans. (C) planktonic d18O record from site MD9501 (Almogi-Labin et al., 2009); NAHP ¼ NorthAfrican humid periods. (D) Summer insolation at 40�N (Berger and Loutre, 1991). (E) MIS ¼ Marine Isotope Stages.
U.C. Müller et al. / Quaternary Science Reviews 30 (2011) 273e279 277
the orbitally induced summer insolation maximum at 58 kyr BP(Fig. 5d) that resulted in a northwardmovement of the IntertropicalConvergence Zone and the extension of monsoonal summer rain-falls into the otherwise hyper-arid Sahara in northern Africa(Rossignol-Strick et al., 1982). Thus, as was the case in the earlyHolocene during formation of Sapropel S1, a humid period pre-vailed in northern Africa during Sapropel S2 formation betweenc. 55 and 49 kyr BP. We argue that this North African humid periodfacilitated the successful dispersal of AMH from Africa.
Our argument is corroborated by the earliest palaeolithicindustry in the eastern Mediterranean of MIS 3 that is a candidatefor manufacture by AMH, found at the site of Boker Tachtit in Sinai(Fig. 1) and dated to 51.0 � 3.4 kyr cal BP (Marks, 1983; Bar-Yosef,2002; Supplementary Table S2). During that time, conditionswere also humid and mild in the eastern Mediterranean as indi-cated by the vegetation data from Tenaghi Philippon (Fig. 5b) andthe precipitation proxy-record from the Soreq cave speleothem(Bar-Matthews et al., 2000) for the GIS 14/13 interval. Triggered bythe resumption of the North Atlantic meridional overturningcirculation (Bond et al., 1993; Bauch et al., 2001) the climateimprovement connected to GIS 14/13 also brought moist and mildenvironmental conditions over large parts of Europe (Allen et al.,1999; Sánchez Goñi et al., 2002; Fletcher et al., 2010). Because of
the coincidence of the orbitally induced North African humidperiod between c. 55 and 49 kyr BP with the humid and mildconditions in the Near East and in Europe during GIS 14/13 (Fig. 5)as triggered by the North Atlantic circulation, natural environmentswere exceptionally favourable for AMH to migrate directly intoEurope already at that time.
The scenario of an initial AMH movement into Europe duringGIS 14/13 is supported by the earliest industry attributed tomodernhumans in Europe, found at the site of Brno-Bohunice in the CzechRepublic (Fig. 1) and dated to a weighted mean of 48.2 � 1.9 kyrBPTL (Richter et al., 2008; Supplementary Table S2), and by thesimilarity of its Bohunician industry with the Emiran industry ofBoker Tachtit (Richter et al., 2008; Hoffecker, 2009). Since 5 of the11 thermoluminescence dates performed on burned flints from theBohunician industry plot within the interval of late GIS 14/13(Richter et al., 2008) when environmental conditions werefavourable, we propose an initial AMH movement into Europeduring that time. However, this movement was preceded byNeanderthal populations moving back from their southern refugesinto Central and Northern Europe during milder climate conditionsin the interval from GIS 17 to mid GIS 14/13. Substantial indigenouspopulations were therefore present in Europe (van Andel et al.,2003), forming a competitive barrier to the further expansion of

U.C. Müller et al. / Quaternary Science Reviews 30 (2011) 273e279278
AMH during late GIS 14/13. The decisive environmental event inthis situation of competition between two closely related hominidspecies was the sharp climatic deterioration of Heinrich Event H5 atc. 48 kyr BP.
Heinrich Event H5 brought a collapse of the Laurentide ice sheetand the release of a huge amount of icebergs into the North Atlantic(Broecker, 1994) which even reached the coastlines of Europe(Sánchez Goñi et al., 2002). This event triggered a disruption of theNorth Atlantic meridional overturning circulation and an extremeclimate deterioration with cold and/or dry conditions over Europe(Allen et al., 1999; Sánchez Goñi et al., 2002; Müller et al., 2003).Our vegetation data from Tenaghi Philippon reveal that the impactof Heinrich H5 was as extreme as the glacial maximum of MIS 4(Fig. 5b) whenmost parts of Europewere deserted by Neanderthals(van Andel et al., 2003). Therefore, we argue that there wasa demographic vacuum in most of Europe during the centuries-long Heinrich H5 event. In the mid-latitudes of Europe, the impactof H5 was mainly through a strong temperature decline (Mülleret al., 2003), whereas drought was the critical issue in the Medi-terranean as documented by the records Tenaghi Philippon (Figs. 4and 5), Megali Limni (Margari et al., 2009), and MD95-2043 in theAlboran Sea (d’Errico and Sánchez Goñi, 2003). The lake level datafor Lake Lisan (Bartov et al., 2002), the huge precursor of the DeadSea, suggest, however, that the freshwater capacity in the NWLevant was still sufficient to sustain large AMH populations close tothe gateway to Europe.
With the subsequent climate improvement at the onset of GIS12, c. 47 kyr BP, triggered by the rapid resumption of the NorthAtlantic meridional overturning circulation (Bond et al., 1993),environments in the gateway to Europe changed quickly fromdesert-steppe into open forest biomes (Fig. 4), which comprisedapart from Pinus mainly summergreen Quercus, but also minorpercentages of Corylus, Ulmus, Tilia, and even evergreen Quercus(Fig. 3). The occurrence of these taxa indicates warm summers witha mean temperature of>17 �C during the warmest month and mildwinters with a mean temperature of >3 �C during the coldestmonth (Frenzel, 1991; Barbero et al., 1992). Due to this rapid andextreme climate improvement the invasive AMH populations thatpersisted through Heinrich H5 in the refuge of the NW Levant hadthe opportunity to spread into Europe and occupy large parts in thecentre and north of the continent before the sedentary Neander-thals were able to reoccupy this territory. This scenario is consistentwith the finding that migration and settlement choices of earlyAMH show clear preferences for mild climate conditions (Daviesand Gollop, 2003), and the similarity of artefact assemblagesfound at Bacho Kiro and Temnata Cave in SE Europe (Fig. 1) withthose of Boker Tachtit, Ksar ’Aqil, and Üça�gızlı in the Levant(Hoffecker, 2009).
In contrast to the situation after the glacial maximum of MIS 4when understrengthed Neanderthal populations could recoverwithout competition by AMH during the milder climate conditionsin the interval from GIS 17 to mid GIS 14/13 (Fig. 5b), the HeinrichH5 climate deterioration resulted in a demographic vacuum thatoccurred when an invasive human species that had already rapidlycolonised SE Asia and Australia was at the gateway to Europe. Weconclude that the extreme environmental impact of Heinrich EventH5 at c. 48 kyr BP as documented at Tenaghi Philippon withina situation of competitive exclusion between two closely relatedhominids species shifted the balance in favour of modern humans.
During the further colonisation of Europe by AMH, the impact ofsubsequent extreme climate changes caused environmentalconditions to switch repeatedly e in some cases within one humangeneration e from those supporting open forest biomes duringGreenland interstadials to dry steppe or tundra biomes duringGreenland stadials and vice versa. Adaptation to such abrupt
environmental changes is a hallmark of AMH and was achievedthrough innovation in both technology and social organisation tocreate a dispersal specialist with a global distribution (Gamble,2009).
Acknowledgements
We thank N.J. Conard for comments on an earlier version of thispaper. This study was financially supported by the Deutsche For-schungsgemeinschaft (project PR 651/3), the Biodiversity andClimate Research Center (BIK-F) of the Hessian Initiative forScientific and Economic Excellence (LOEWE), the Wilhelm Schuler-Foundation (all Germany), The Royal Society (United Kingdom), andthe Jackson School of Geosciences (USA). We thank A. Almogi-Labinfor providing planktonic isotope data from core MD9501. Technicalsupport by S. Liner and S. Kalaitzidis is gratefully acknowledged.
Appendix. Supplementary information
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.quascirev.2010.11.016.
References
Allen, J.R.M., Brandt, U., Brauer, A., Hubberten, H.-W., Huntley, B., Keller, J.,Kraml, M., Mackensen, A., Mingram, J., Negendank, J.F.W., Nowaczyk, N.R.,Oberhänsli, H., Watts, W.A., Wulf, S., Zolitschka, B., 1999. Rapid environmentalchanges in southern Europe during the last glacial period. Nature 400, 740e742.
Almogi-Labin, A., Bar-Matthews, M., Shriki, D., Kolosovsky, E., Paterne, M.,Schilman, B., Ayalon, A., Aizenshtat, Z., Matthews, A., 2009. Climatic variabilityduring the last ∼90 ka of the southern and northern LevantineBasin as evidentfrom marine records and speleothems. Quaternary Science Reviews 28,2882e2896.
Barbero, M., Loisel, R., Quézel, P., 1992. Biogeography, ecology and history ofMediterranean Quercus ilex ecosystems. Vegetatio 99e100, 19e34.
Bar-Matthews, M., Ayalon, A., Kaufman, A., 2000. Timing and hydrological condi-tions of sarpropel events in the Eastern Mediterranean, as evident from spe-leothems, Soreq cave. Israel. Chemical Geology 169, 145e156.
Bartov, Y., Stein, M., Enzel, Y., Agnon, A., Reches, Z., 2002. Lake levels and sequencestratigraphy of Lake Lisan, the late pleistocene precursor of the Dead sea.Quaternary Research 57, 9e21.
Bar-Yosef, O., 2002. The Upper paleolithic revolution. Annual Review of Anthro-pology 31, 363e393.
Bauch, H.A., Erlenkeuser, H., Spielhagen, R.F., Struck, U., Matthiessen, J., Thiede, J.,Heinemeier, J., 2001. A multiproxy reconstruction of the evolution of deep andsurface waters in the subarctic Nordic seas over the last 30,000 yr. QuaternaryScience Reviews 20, 659e678.
Berger, A., Loutre, M.F., 1991. Insolation values for the climate of the last 10 millionyears. Quaternary Science Reviews 10, 297e317.
Bond, G., Broecker, W., Johnsen, S., McManus, J., Labeyrie, L., Jouzel, J., Bonani, G.,1993. Correlations between climate records from North Atlantic sediments andGreenland ice. Nature 365, 143e147.
Broecker, W., 1994. Massive iceberg discharges as triggers for global climate change.Nature 372, 421e424.
Davies, W., Gollop, P., 2003. The human presence in Europe during the last glacialPeriod II: climate tolerance and climate preferences of mid- and LateGlacialhominids. In: van Andel, T.H., Davies, W. (Eds.), Neanderthals and ModernHumans in the European Landscape During the Last Glaciation. McDonaldInstitute Monographs, Cambridge, pp. 131e146.
d’Errico, F., Sánchez Goñi, M.F., 2003. Neandertal extinction and the millennial scaleclimatic variability of OIS 3. Quaternary Science Reviews 22, 769e788.
De Vivo, B., Rolandi, G., Gans, P.B., Calvert, A., Bohrson, W.A., Spera, F.J., Belkin, H.E.,2001. New constraints on the pyroclastic eruptive history of the Campanianvolcanic Plain (Italy). Mineralogy and Petrology 73, 47e65.
Fleitmann, D., Cheng, H., Badertscher, S., Edwards, R.L., Mudelsee, M., Göktürk, O.M.,Fankhauser, A., Pickering, R., Raible, C.C., Matter, A., Kramers, J., Tüysüz, O.,2009. Timing and climatic impact of Greenland interstadials recorded instalagmites from northern Turkey. Geophysical Research Letters 36, L19707.doi:10.1029/2009GL040050.
Fletcher, W., Sánchez Goñi, M.F., Allen, J.M.R., Cheddadi, R., Combourieu-Nebout, N.,Huntley, B., Lawson, I., Londeix, L., Magri, D., Margari, V., Müller, U.C.,Naughton, F., Novenko, E., Roucoux, K., Tzedakis, P.C., 2010. Millennial-scalevariability during the last glacial in vegetation records from Europe. QuaternaryScience Reviews 29, 2839e2864.

U.C. Müller et al. / Quaternary Science Reviews 30 (2011) 273e279 279
Frenzel, B., 1991. Das Klima des Letzten Interglazials in Europa. In: Frenzel, B. (Ed.),Klimageschichtliche Probleme der letzten 130000 Jahre. Paläoklimaforschung.Band, 1. Fischer Verlag, Stuttgart, New York, pp. 51e78.
Gamble, C., 2009. Human display and dispersal: a case study from biotidal Britain inthe middle and upper pleistocene. Evolutionary Anthropology 18, 144e156.
Hoffecker, J.F., 2009. The spread of modern humans in Europe. Proceedings of theNational Academy of Sciences 106, 16040e16045.
Imbrie, J., Hays, J.D., Martinson, D.G., McIntyre, A., Mix, A.C., Morley, J.J., Pisias, N.G.,Prell, W.L., Shackleton, N.J., 1984. The orbital theory of Pleistocene climate:support from a revised chronology of the marine d18O record. Part 1. In:Berger, A.L., et al. (Eds.), Milankovitch and Climate. Reidel Publishing Company,pp. 269e305.
Kotthoff, U., Pross, J., Müller, U.C., Peyron, O., Schmiedl, G., Schulz, H., Bordon, A.,2008a. Climate dynamics in the borderlands of the Aegean Sea during forma-tion of sapropel S1 deduced from a marine pollen record. Quaternary ScienceReviews 27, 832e845.
Kotthoff, U., Müller, U.C., Pross, J., Schmiedl, G., Lawson, I.T., van de Schootbrugge, B.,Schulz, H., 2008b. Lateglacial and Holocene vegetation dynamics in the Aegeanregion: an integrated view based on pollen data from marine and terrestrialarchives. The Holocene 18, 1019e1032.
Margari, V., Gibbard, P.L., Bryant, C.L., Tzedakis, P.C., 2009. Character of vegetationaland environmental changes in southern Europe during the last glacial period;evidence from Lesvos Island, Greece. Quaternary Science Reviews 28,1317e1339.
Marks, A.E., 1983. The sites of Boker Tachtit and Boker: a brief introduction. Part 3.In: Marks, A.E. (Ed.), Prehistory and Palaeoenvironments in the Central Negev,Israel. The Avdat/Aqev Area, vol. III. Southern Methodist University Press, Dal-las, pp. 15e37.
Martinson, D.G., Pisias, N.G., Hays, J.D., Imbrie, J., Moore Jr., T.C., Shackleton, N.J.,1987. Age dating and the orbital theory of the ice ages: development of a high-resolution 0 to 300,000-year chronostratigraphy. Quaternary Research 27, 1e29.
Mellars, P., 2006. A new radiocarbon revolution and the dispersal of modernhumans in Eurasia. Nature 439, 931e935.
Müller, U.C., Pross, J., Bibus, E., 2003. Vegetation response to rapid climate change inCentral Europe during the past 140,000 yr based on evidence from the Für-amoos pollen record. Quaternary Research 59, 235e245.
NGRIP members, 2004. High-resolution record of northern hemisphere climateextending into the last interglacial period. Nature 431, 147e151.
Pross, J., Tzedakis, C., Schmiedl, G., Christanis, K., Hooghiemstra, H., Müller, U.C.,Kotthoff, U., Kalaitzidis, S., Milner, A., 2007. Tenaghi Philippon re-visited: dril-ling a continuous lower-latitude terrestrial climate archive of the last 250,000years. Scientific Drilling 5, 30e32.
Pross, J., Kotthoff, U., Müller, U.C., Peyron, O., Dormoy, I., Schmiedl, G., Kalaitzidis, S.,Smith, A.M., 2009. Massive perturbation in terrestrial ecosystems of the EasternMediterranean region associated with the 8.2 ka climatic event. Geology 37,887e890.
Richter, D., Tostevin, G., Skrdla, P., 2008. Bohunician technology and thermolumi-nescence dating of the type locality of Brno-Bohunice (CzechRepublic). Journalof Human Evolution 55, 871e885.
Rohling, E.J., Mayewski, P.A., Abu-Zied, R.H., Casford, J.S.L., Hayes, A., 2002. Holoceneatmosphere-ocean interactions: records from Greenland and the Aegean Sea.Climate Dynamics 18, 587e593.
Rossignol-Strick, M., Nesteroff, W., Olive, P., Vergnaud-Grazzini, C., 1982. After thedeluge: Mediterranean stagnation and sapropel formation. Nature 295,105e110.
Sánchez Goñi, M.F., Cacho, I., Turon, J.-L., Guiot, J., Sierro, F.J., Peypouquet, J.-P.,Grimalt, J.O., Shackleton, N.J., 2002. Synchroneity between marine and terres-trial responses to millennial scale climatic variability during the last glacialperiod in the Mediterranean region. Climate Dynamics 19, 95e105.
Siddall, M., Rohling, E.J., Almogi-Labin, A., Hemleben, Ch., Meischner, D.,Schmelzer, I., Smeed, D.A., 2003. Sea-level fluctuations during the last glacialcycle. Nature 423, 853e858.
Spötl, C., Mangini, A., Richards, D.A., 2006. Chronology and paleoenvironment ofmarine isotope stage 3 from two high-elevation speleothems, Austrian Alps.Quaternary Science Reviews 25, 1127e1136.
Tzedakis, P.C., 1999. The last climatic cycle at Kopais, central Greece. Journal of theGeological Society 156, 425e434.
Tzedakis, P.C., 2007. Seven ambiguities in the Mediterranean palaeoenvironmentalnarrative. Quaternary Science Reviews 26, 2042e2066.
Tzedakis, P.C., Lawson, I.T., Frogley, M.R., Hewitt, G.M., Preece, R.C., 2002. Bufferedtree population changes in a Quaternary refugium: evolutionary implications.Science 297, 2044e2047.
Tzedakis, P.C., Frogley, M.R., Lawson, I.T., Preece, R.C., Cacho, I., de Abreu, L., 2004.Ecological thresholds and patterns of millennial-scale climate variability: theresponse of vegetation in Greece during the last glacial period. Geology 32,109e112.
van Andel, T.H., Davies, W., Weninger, B., 2003. The human presence in Europeduring the last glacial Period I: human Migrations and the changing climate. In:van Andel, T.H., Davies, W. (Eds.), Neanderthals and Modern Humans in theEuropean Landscape During the Last Glaciation. McDonald Institute Mono-graphs, Cambridge, pp. 31e56.
Weiss, H., Courty, M.-A., Wetterstrom, W., Guichard, F., Senior, L., Meadow, R.,Curnow, A., 1993. The genesis and collapse of third millennium North Meso-potamian civilization. Science 261, 995e1004.
Weninger, B., Jöris, O., 2008. A 14C age calibration curve for the last 60 ka: theGreenland-Hulu U/Th timescale and its impact on understanding the Middle toUpper Paleolithic transition in Western Eurasia. Journal of Human Evolution 55,772e781.
Wulf, S., Kraml, M., Kuhn, T., Schwarz, M., Inthorn, M., Keller, J., Kuscu, I., Halbach, P.,2002. Marine tephra from the Cape Riva eruption (22 ka) of Santorini in the Seaof Marmara. Marine Geology 183, 131e141.