rapid watet uptake and structural changes in imbibing seed · pdf filerapid water uptake and...
TRANSCRIPT

Protoplasma 77, 111--122 (1973)
�9 by Springer-Verlag 1973
Rapid Water Uptake and Structural Changes in Imbibing Seed Tissues
M. S. BUTTROSE
CSIRO, Division of Horticultural Research, Adelaide, South Australia
With 9 Figures
Received November 15, 1972
Summary
Studies on ultrastructure in tissues from air-dry seeds normally involve fixation in aqueous media. Recent freeze-etch studies suggest that water is taken up extremely rapidly by these tissues with attendant structurai change. Measures have now been made of water uptake by small (< 400 ~tg) pieces of Hordeum vulgare scutellum, Pisum sativum cotyledon and radicle, and Helianthus annum cotyledon. Measurable water uptake occurred within 10 seconds, and scutellum was already saturated (160% of initial air-dry weight) within 2 minutes. Tissues re-dried almost as rapidly. Barley scutellum was investigated by freeze- etching. Air-dry tissue, frozen direct in liquid nitrogen or surrounded by 100% glycerol, had convoluted cell walls, organelles with indented margins, and polyhedral lipid bodies which were in such close mutual proximity as to nearly form closed lipid sheets over plasmalemma and protein bodies. Within 8 seconds of exposing to water, cell walls had straightened and organelles and lipid bodies rounded out. It is likely that many descriptions of structure in "air-dry" seeds have been of this hydrated structure.
1. Introduction
A number of workers have used conventional fixation in aqueous media followed by embedding and sectioning in studying the ultrastructure of tissues from air-dry seeds. Examples of tissues are the scuteilum of wheat (SWIFT and O'BI~IEN 1972), the whole embryo of lettuce (PAuLsoN and SI~IVASTAVA 1968), and the cotyledons of cottonseed (YATsu 1965). Ultrastructure observed in these studies may differ from that in native air-dry seeds as water in aqueous fixatives could cause some swelling before fixation is effected. Evidence for this has been difficult to obtain. PERNEr, (1965) used osmium tetroxide vapour to fix radicles from air-dry pea seeds, and organelles in his preparations were indeed less rounded than is common in aqueously-fixed tissue. However, differences were not decisive. YATsu (1965) was unsuccess- ful in employing osmium vapour. Fixation by rapid freezing should render

112 M.S. BUTTROSE
swelling or shrinkage impossible, and consequently the technique of freeze- etching should be useful in studying structure in air-dry tissue. In routine freeze-etching, tissue is partly or wholly contained within a drop of water (or glycerol solution or pure glycerol) on a metal disk, then frozen rapidly by plunging into liquid Freon at approximately - - 1 5 0 ~ Thus the frozen liquid cements the tissue onto the disk, but the tissue is in contact with a liquid phase for some seconds before being frozen. Recent studies (S~,vIFT and BUTTROSE 1972 a and b, and unpublished) have shown that the structure of seed tissue exposed to water for 8 seconds before freezing differed from that exposed to pure glycerol. Where glycerol was used organdies had indented margins, oil droplets were polyhedral and cell walls convoluted; whereas where water was used organdies and lipid droplets were rounded and cell walls straightened. These results suggest on the one hand that water can enter cells of tissues dissected from air-dry seeds very rapidly, and on the other that organdies and walls can undergo shape changes within a few seconds. There appears to be a lack of information on water uptake by small pieces of seed embryo over short time intervals. This paper describes experi- ments designed to measure water uptake by small blocks of embryo tissue and to relate water uptake to structural changes.
2. Materia l and Methods 2.1. S e e d s
Tissue was taken from the scutellum of barley (Hordeurn vulgate cv. Himalaya), cotyledons and radicle of pea (Pisum sativurn cv. Greenfeast), and cotyledons of sunflower (Helianthus annuus cv. Polestar). In addition, whole seeds of barley, and of pea and sunflower after removal of the seed coat, were studied.
2.2. W a t e r U p t a k e
Pieces of tissue (150-400 ag) were dissected from seeds using scalpel and razor blades. Pieces of barley scutellum were thin and flat, with one of the flat faces and the four side faces freshly cut. Blo&s from pea and sunflower cotyledons were approximately cubic with either five or six freshly-cut sides. Pea radicles were removed from the embryo by making a single transverse cut. Immediately after dissection from the seed a sample piece was placed on the pan of a Cahn Electro-Microbalanee and weighed to the nearest 5 ~tg. The piece was then submerged in a drop of water at 22 ~ for a period ranging from 10 seconds to 15 minutes, and then removed and immediately blotted dry (no surface film of water visible at • 30 magnifi- cation) and reweighed. The time elapsing between removal from water and rebalancing on the balance was approximately I0 seconds. The imbibed weight was expressed as a percentage of the initial air-dry weight. All results presented are the mean of 10 different sample pieces for any time period. Water uptake over a 24-hour period by whole, submerged seeds was measured by periodically blotting dry and weighing.
2.3. W a t e r L o s s
Tissue pieces, as used for uptake measurements, were weighed, submerged in water for 15 minutes, then blotted dry and placed on the balance pan. Weights were recorded over

Rapid Water Uptake and Structural Changes in Imbibing Seed Tissues 113
a 15 minutes period as the pieces lost water under conditions of approximately 22 ~ and 60% relative humidity. Results were expressed as a percentage of the initial air-dry weight. Similar measurements were made with pieces of tissue dissected from whole seeds which had been submerged in water for 16 hours. Results were expressed as a percentage of the final air-dry weight.
2.4. L i p i d a n d M o i s t u r e C o n t e n t o f B a r l e y S c u t e l l u m
Tissue was dissected from air-dry seeds, weighed on a microbalance, and extracted in chloroform (four changes) at 22 ~ for 4 days. The weight lost by the tissue was taken as a measure of lipid content. Moisture content of air-dry scutellum was measured as weight lost after drying at 105 ~ for.24 hours.
2.5. L i g h t M i c r o s c o p y
Scutellum tissue was dissected from air-dry barley seeds, placed on a microscope slide, covered with a coverslip, and photographed. Water was presented at the edge of the coverslip and the tissue re-photographed at known intervals after the water front was observed to make first contact with the tissue. Ten pieces of scutellum tissue were also taken as above, placed on a microscope slide and one axis of each piece was measured using a micrometer eyepiece. The same axis was remeasured after covering the piece with water for 2 minutes. Final measurements were expressed as a percentage of the initial measurements.
2.6. F r e e z e - E t c h i n g
The following preparations of barley scutellum were examined:
a) pieces dissected from air-dry seeds and mounted in water or glycerol before immediate freezing; b) pieces as above but allowed to remain in contact with liquid water for 2 minutes before freezing; c) pieces dissected from air-dry seeds and frozen by plunging the air-dry piece into liquid nitrogen; d) pieces dissected from whole seeds which had been allowed to imbibe 40% of their own weight of water over a 24-hour period followed by return to air-dryness ("hardened") --these pieces were mounted in water or glycerol before immediate freezing.
For a), b), and d) the tissue was in contact with liquid (water or glycerol) on gold specimen disks for approximately 8 seconds before freezing by plunging into tiquid Freon at approximately - -150 ~ For c), frozen pieces were cemented to disks by placing them in a drop of liquid trichloro-ethylene held on the specimen disks at - - 7 0 ~ (JoHNsoN 1968), followed by plunging into liquid nitrogen. Specimens were freeze-fractured and replicas made in a Balzers apparatus according to the method of Moor et al. (1961). Fractured faces of specimens frozen in a water droplet were etched for 2 minutes at - - 100 ~ before replication.
2.7. T o l e r a n c e t o F r e e z i n g
Twenty seeds of barley, 10 of pea, and 10 of sunflower were plunged into liquid nitrogen and held there for 5 minutes. Seeds were transferred to a bench and allowed to come to room temperature, then placed on moist filter paper in Petri dishes. Unfrozen seeds, as controls, were also placed on moist filter paper. After 7 days a count was made of the number of seeds which had germinated.
Protoplasma 77/1 8

114 M.S. BUTTROSE
3. Resul ts
3.1. W a t e r U p t a k e
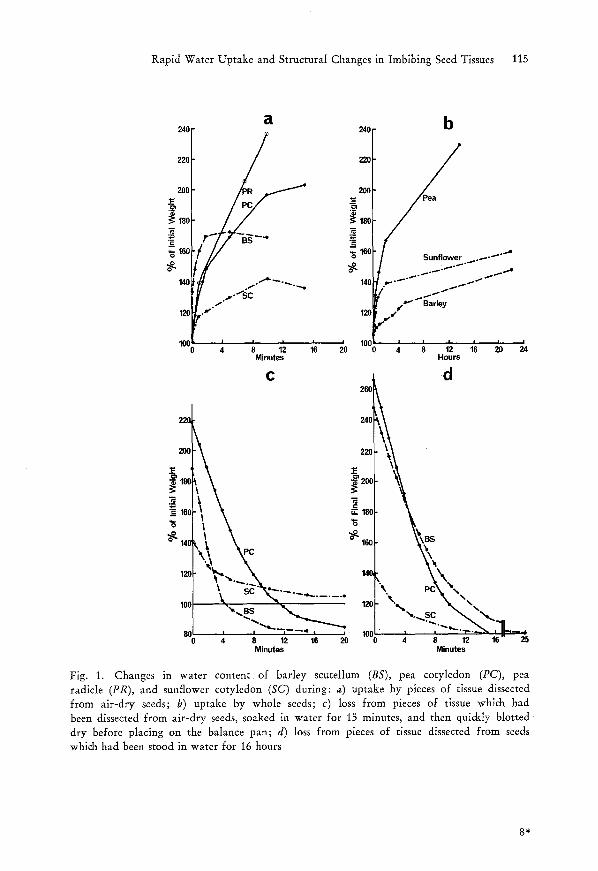
Pieces of barley scutellum took up water for only 2 minutes, and half this uptake occurred during the first 10 seconds (Fig. 1 a). The other tissues examined took up water for approximately 10 minutes. Water uptake by whole seeds was much slower (Fig. 1 b).
3.2. W a t e r L o s s
Pieces of barley scutel!um and pea cotyledon, which had been soaked in water for 15 minutes, lost their water within 15 minutes, and in fact ended with a net loss in air-dry weight (Fig. 1 c). This net loss was presumably due to dry matter in cut surface cells dissolving in water or being mechanically displaced. Water loss from pieces of sunflower cotyledon was slower (Fig. 1 c). Rates of water loss from pieces which were taken from imbibed whole seeds were similar to those from corresponding pieces cut initially from dry seeds, except that loss from barley scutellum was slower over the first few minutes (Fig. 1 d).
3.3. L i p i d a n d W a t e r C o n t e n t o f B a r l e y S c u t e l l u m
Chloroform solubles were 22.20/o (_+ 1.0~ of air-dry weight, and water 7.90/0 (_+ 0.2)0/0.
3.4. L i g h t M i c r o s c o p y
Swelling of barley scutellum is illustrated in Fig~ 8. Pieces of scutellum, after immersion in water for 2 minutes, showed dimensional increases of 17.5~ (_+ 1.5~
3.5. F r e e z e - E t c h i n g
Replicas from barley scutellum that had been frozen in a drop of glycerol showed extensive areas with little obvious differentiation (Fig. 2). The basic structure was scale-like, a scale often being resolved as a pentagon or hexagon. Fractures followed a very irregular path, and cell walls were deeply folded (Fig. 2). Areas representing cytoplasm were seldom seen, and protein bodies (Fig. 2 inset) had indented margins. Replicas from scutellum frozen direct in liquid nitrogen and cemented to disks with trichloroethylene appeared similar to those frozen in glycerol. Fig. 3, which was taken from such a replica, was chosen because it showed clearly the scale-like profiles of poly- hedral bodies. A ridge surrounded each profile. Exposure to liquid water for 8 seconds before freezing caused substantial changes in appearance of replicas (Fig. 4). Fracture planes were relatively flat, and prominent features were profiles of rounded protein bodies surrounded by rounded lipid bodies,
Rapid Water Uptake and Structural Changes in Imbibing Seed Tissues 115
24(]
220
280
18(3
_= 16C
140
~C
a
PR J PC
.p~ , f
188
"~ Is8
140
b
12( r d'~'" Barley
240
C
220
20O
18[1
.:E_- _c "6 16(~
; '8 ;2 1'6 ~o % ; ~ ;2 1~ ~o 2', Minutes Hours
d 260
220, 240 ~
200 ~ 22O ~
, \ ~ 206
leo %\,~
\ ........... .IX "- "~ 120]- "~ ~ %
" - . - ~ I. - - . - . _ . _ \ "1. 80 I " ~ 4 , i. 100 U I I - - " P - ~ , - - ~
4 8 12 16 20 0 4 8 12 16' 25 Minutes Minutes
Fig. 1. Changes in water content of barley scutellum (BS), pea cotyledon (PC), pea radicle (PR), and sunflower cotyledon (SC) during: a) uptake by pieces of tissue dissected from air-dry seeds; b) uptake by whole seeds; c) loss from pieces of tissue which had been dissected from air-dry seeds, soaked in water for 15 minutes, and then quickly blotted dry before placing on the balance pan; d) loss from pieces of tissue dissected from seeds which had been stood in water for 16 hours
8*

116 M.S. BuTTgos~
with areas of cytoplasm in between. Cell walls were normally straight in these preparations. Tubular membrane structures, lying between the circular lipid profiles, were common (Figs. 4 and 5). These preparations showed no evidence of ice crystal damage. After exposing scutellum tissue to water for 60 seconds before freezing, replicas showed ice crystal damage both in the ground cytoplasm and within the protein bodies (Fig. 6); otherwise features were similar to those observed after 8 seconds exposure. The structure of scutellum from seeds which had been dried down after a 24-hours imbibition period was similar to the structure of the corresponding preparation (glycerol- or water-frozen) of unimbibed air-dry seeds. Occasionally finely-corrugated, rather than straight, cell walls were observed in these replicas (Fig. 7).
3.6. T o l e r a n c e t o F r e e z i n g
Submerging air-dry seeds in liquid nitrogen for 5 minutes had no deleterious effect on viability (Table 1).
Table 1. Percent Germination of Control Air-Dry Seeds and of Seeds after Freezing in Liquid Nitrogen for 5 Minutes
Seed Control Frozen
Barley 90 95 Pea 90 100 Sunflower 70 70
4. D i s c u s s i o n
Small pieces of tissue dissected from embryos of mature air-dry seeds can imbibe water very rapidly, such that those of barley scutellum, for instance, can accommodate half their own weight of water within 30 seconds and saturate within 2 minutes. The rate of uptake may increase with increase in the cut surface: volume ratio, as dissected whole rye embryos, which have tissues comparable to barley scutellum, took 13 minutes to saturate (HALLAM
Fig. 2. Fracture face from scutellum tissue taken from an air-dry seed and exposed to 100% glycerol for 8 seconds before freezing. W = cell wail. • Inset: Margin (arrows) of protein body (P) from a similar preparation. • 20,000 Fig. 3. Fracture face from scutellum tissue taken from an air-dry seed and frozen direct in liquid nitrogen before cementing with triehloroethylene. • 12,000 Fig. 4. Fracture face from tissue taken from an air-dry seed and exposed to water for 8 seconds before freezing. P = protein body, L = lipid droplet, C = cytoplasm. Note tubular membrane structures (arrows). • 24,000

Rapid Water Uptake and Structural Changes in Imbibing Seed Tissues 117
Figs. 2-4

118 M.S. Bu'r'rRos~
et al. 1972). Whole seeds, with no cut surfaces, imbibed at a much slower rate than dissected pieces. The rapidity with which pieces of tissue could lose water suggests that most of the water remains unbound. This lack of binding must likewise apply to tissues in the imbibed whole seeds which had presum- ably begun germination activity. Water could only enter the non-lipid proportion of the barley scutellum, and the changes in water content of this portion may be calculated taking into account the initial lipid content of 220/0, the initial water content of 80/0, and values from Fig. 1 a. Thus this portion had a water content of 11~ initially, of 38% after 10 seconds exposure to water and of 530/0 after 2 minutes exposure. Light microscope observations suggested that water taken up permeated the entire tissue block with attendant increase in total volume. It is noted that the measured increase in dimension of pieces of scutellum after 2 minutes in water (100-+ 117.5) corresponds to a relative increase in volume of from 100 to 162. This is comparable to the increase of from 100 to 170 in fresh weight (Fig. I a). Freeze-etch results support the suggestion that water rapidly permeated the whole tissue blo&. Thus there was a consistent difference in appearance between preparations from blo&s frozen in glycerol and those from blo&s frozen in water, even though fracture paths are at random through frozen blocks. The similarity between the "glycerol" and the "trichloroethylene" preparations is evidence that glycerol did not affect the structure of tissue prior to freezing. Freeze-etch results also indicate that water incorporation was uniform within cells; if it were not, one would expect to see variations in fracture texture due to formation of ice crystals where there were local concentrations of water (compare ice crystallization in Fig. 6 with its absence in Fig. 4). The results show that there is a latent potential for increase in volume by tissues of air-dry seeds. Folded cell walls can straighten to allow cells to enlarge, and indented organelles can round out, without rupturing, as they imbibe. Presumably these changes are as readily reversible as is the passage of water, because freeze-etch replicas of scutellum from "hardened" barley seeds could not be distinguished from those of control tissue. There are other changes in cell ultrastructnre which may not be reversible (MAI~INos
Fig. 5. Lipid bodies (L) with interspersed tubular membrane structures (arrows) from air- dry scutellum exposed to water for 8 seconds before freezing. X42,000 Fig. 6. As for Fig. 4 bur exposed to water for 2 minutes before freezing. • 12,000 Fig. 7. Fracture face from tissue dissected from an air-dry seed, which had previously imbibed 40% of its own weight of water before redrying, and exposed to water for 8 seconds before freezing. W = cell wall, L = lipid body, PI = plasmalemma. • 12,000 Fig. 8. Piece of barley scutellum in air-dry state (O), and at 2, 10, and 60 seconds after contact with water. X 80

Rapid Water Uptake and Structural Changes in Imbibing Seed Tissues 119
Figs. 5-8

120 M.S . BUTTROSE
and FIFe 1972). Fig. 7 may represent a cell wall frozen just before full straightening. An interpretation of these shape changes is illustrated in Fig. 9. Abundant lipid bodies are congregated over the plasmalemma and over the surfaces of protein bodies in cells of the scutellum of barley (Ni~uwDoRv 1963) and of wheat (SWIFT and O'BRIEN 1972). Freeze-fracture replicas
a b
Detail of protein body
Fig. 9. Diagram i l lustrat ing structure in a barley scutellum cell a) in the air-dry state and b ) a f t e r exposure to water for 8 seconds. The increase in area f rom a) to b) represents an increase in volume of f rom 100 to 17(3. The dark lines breaking across the cells represent fracture paths as in freeze-etching. W = cell wall, P = prote in body, L = lipid body, N = nucleus
showed that these lipid bodies, in glycerol-frozen wheat scutellum, were close-packed polyhedrons (SWIFT and BUTTI~OSE 1972 a). The small scale-like or polyhedral bodies, with common straight edges, observed after freezing in the absence of water in the present work, can consequently be identified as lipid bodies. It is suggested that a polyhedral, rather than spherical, form is due purely to pressure arising during dehydration contraction, and has no physiological basis. Resumption of spherical shape would be a result of relaxation of pressure, not water uptake. These bodies must have occupied a major proportion of the uneven fracture plane in non-aqueousty-frozen

Rapid Water Uptake and Structural Changes in Imbibing Seed Tissues 121
tissue (Figs. 2 and 3), but only a minor proportion in partially-hydrated tissue (Fig. 4), where they were rounded and separated. The diagram in Fig. 9 illustrates the conclusions drawn from the results. In air-dry tissue, con- gregated lipid bodies could form nearly continuous interweaving sheets, and BRANTON (1969) has pointed out that fractures occur preferentially through lipid layers. During hydration, the mutual separation of lipid bodies would destroy the continuity and fractures would tend to be more direct and to pass through organelles and areas of cytoplasm. The concept of closed sheets of lipid, as in Fig. 9 a, suggests a mechanism for retention of that water which remains when cell volume is reduced to a point where all pores between lipid bodies are closed. Freezing of whole seeds in liquid nitrogen had no adverse effect on their viability. Consequently it is concluded that structure observed in barley scutellum after freezing small pieces direct in liquid nitrogen represented structure in the native air-dry state. Some aspects of this structure, for the case of barley scutellum, are shown in Fig. 9 a. Water-uptake results, in conjunction with other freeze-etch results (SwrrT and BUTTROSE 1972 b), indicate that this diagram could be a valid generalization for seed embryos. It is likely that all studies of structure in air-dry seeds, where aqueous fixa- tives have been used, have resulted in descriptions of modified structure as in Fig. 9 b.
Miss C. L. ROWNTREE gave technical assistance.
References
BRANTON, D., 1969: Membrane structure. Ann. Rev. Plant Physiol. 20, 209--238. HALLA~a, N. D., B. E. ROBERTS, and D. J. OSBORNE, 1972: Embryogenesis and germination
in rye (SecaIe cereaIe L.). II. Biochemical and fine structural changes during germination. Planta 105, 293--309.
JOHNSON, R. P. C., 1968: Microfilaments in pores between frozen-etched sieve elements. Planta 81, 314--332.
MARINOS, N. G., and D. N. FIFE, 1972: Ultrastructural changes in wheat embryos during a "presowing drought hardening" treatment. Protoplasma 74, 381--396.
MooR, H., K. Mi3HLETHALER, I-I. WALDNER, and A. FREY-WYSSLING, 1961: A new freezing ultramicrotome. J. biophys, biochem. Cytoh 10, 1--13.
NlrUWDORV, P. J., 1963: Electron microscopic structure of the epithelial cells of the scutellum of barley. The structure of the epithelial cells before germination. Acta Bot. Neerl. 12, 295--301.
PAISLSON, R. E., and L. M. SRIVASTAVA, 1968: The fine structure of the embryo of Lactuca sativa. I. Dry embryo. Canad. J. Bot. 46, 1437--1446.
P~I~NER, E., 1965: Elektronenmikroskopische Untersuchungen an Zellen yon Embryonen im Zustand v/511iger Samenruhe. I. Die zellulS.re Strukturordnung in der Radicula luft- trockener Samen yon Pisurn sativum. Planta 65, 334--357.
SWIFT, J. G., and M. S. BIJTTROSE, 1972 a: Freeze-etch studies of protein bodies in wheat scutellum. J. Ultrastruct. Res. 4,9, 378--390.

122 BUTTROSI~: Rapid Water Uptake and Structural Changes in Imbibing Seed Tissues
SWIFT, J. G., and M. S. BUTTROS~, 1972 b: Lipid layers, protein bodies, and plastids in freeze- etched pea cotyledons. Planta 109, 61--72.
- - and T. P. O'BRIEN, 1972: The fine structure of the wheat scutelium before germination. Aust. J. Biol. Sci. 25, 9--22.
YATSU, L. Y., 1965: The ultrastructure of cotyledonary tissue from Gossypium hirsutum L. seeds. J. Cell Biol. 25, 193--199.
Author's address: Dr. M. S. B~3TTI~ose, CSIRO, Division of Horticultural Research, G. P. O. Box 350, Adelaide 5001, South Australia.