receptor binding studies disclose a novel class of high-affinity inhibitors of the escherichia...
DESCRIPTION
A free research journal article on advances in nephrologic medicineTRANSCRIPT

Molecular Microbiology (2005)
55
(2) 441ndash455 doi101111j1365-2958200404415x
copy 2004 Blackwell Publishing Ltd
Blackwell Science LtdOxford UKMMIMolecular Microbiology1365-2958Blackwell Publishing Ltd 2004
2004
55
2441455
Original Article
A novel class of FimH high-affinity ligandsJ Bouckaert
et al
Accepted 30 September 2004 For correspondence E-mailhdegrevevubacbe Tel (
+
32) 2629 18 44 Fax (
+
32) 26 29 19 63E-mail stefanxraybmcuuse Tel (
+
46) 18 471 45 54 Fax(
+
46) 18 53 69 71 Present addresses
dagger
Centre for BiotechnologyJawaharlal Nehru University New Delhi 110067 India
Dagger
PharmAthene Inc 175 Admiral Cochrane Drive Annapolis MD21401 USA
sect
Both authors contributed equally to this work
Receptor binding studies disclose a novel class of high-affinity inhibitors of the
Escherichia coli
FimH adhesin
Julie Bouckaert
1sect
Jenny Berglund
2sect
Mark Schembri
3
Erwin De Genst
1
Lieve Cools
1
Manfred Wuhrer
4
Chia-Suei Hung
5
Jerome Pinkner
5
Rikard Slaumlttegaringrd
6
Anton Zavialov
2
Devapriya Choudhury
2dagger
Solomon Langermann
7Dagger
Scott J Hultgren
5
Lode Wyns
1
Per Klemm
8
Stefan Oscarson
6
Stefan D Knight
2
and Henri De Greve
1
1
Laboratorium voor Ultrastructuur Vrije Universiteit Brussel (VUB) and Vlaams Interuniversitair Instituut voor Biotechnologie (VIB) Building E Pleinlaan 2 1050 Brussels Belgium
2
Department of Molecular Biology Swedish University of Agricultural Sciences Uppsala Biomedical Center PO Box 590 SE-751 24 Uppsala Sweden
3
School of Molecular and Microbial Sciences University of Queensland Brisbane QLD 4072 Australia
4
Biomolecular Mass Spectrometry Unit Department of Parasitology Leiden University Medical Center Albinusdreef 2 2333 ZA Leiden the Netherlands
5
Department of Molecular Microbiology Box 8230 Washington University School of Medicine 660 S Euclid Ave St Louis MO 63110 USA
6
Department of Organic Chemistry Arrhenius Laboratory Stockholm University SE-10691 Stockholm Sweden
7
MedImmune 35 W Watkins Mill Road Gaithersburg MD 20878 USA
8
Microbial Adhesion Group Section for Molecular Microbiology Technical University of Denmark BioCentrum-DTU Building 301 DK-2800 Lyngby Denmark
Summary
Mannose-binding type 1 pili are important virulencefactors for the establishment of
Escherichia coli
uri-nary tract infections (UTIs) These infections are initi-ated by adhesion of uropathogenic
E coli
to uroplakin
receptors in the uroepithelium via the FimH adhesinlocated at the tips of type 1 pili Blocking of bacterialadhesion is able to prevent infection Here we pro-vide for the first time binding data of the molecularevents underlying type 1 fimbrial adherence by crys-tallographic analyses of the FimH receptor bindingdomains from a uropathogenic and a K-12 strain andaffinity measurements with mannose common mono-and disaccharides and a series of alkyl and aryl man-nosides Our results illustrate that the lectin domainof the FimH adhesin is a stable and functional entityand that an exogenous butyl
aaaa
-
D
-mannoside boundin the crystal structures exhibits a significantlybetter affinity for FimH (K
d
=
015
mmmm
M) than mannose(K
d
=
23
mmmm
M) Exploration of the binding affinities of
aaaa
-
D
-mannosides with longer alkyl tails revealed affin-ities up to 5 nM Aryl mannosides and fructose canalso bind with high affinities to the FimH lectindomain with a 100-fold improvement and 15-foldreduction in affinity respectively compared withmannose Taken together these relative FimH affini-ties correlate exceptionally well with the relative con-centrations of the same glycans needed for theinhibition of adherence of type 1 piliated
E coli
Weforesee that our findings will spark new ideas andinitiatives for the development of UTI vaccines andanti-adhesive drugs to prevent anticipated and recur-rent UTIs
Introduction
Although normally a commensal inhabitant of the intesti-nal tract of mammals
Escherichia coli
is also frequentlyassociated with a number of infectious diseases Likemost other Gram-negative pathogens
E coli
can expressa number of adhesive organelles assembled by severaldistinct pathways (Remaut and Waksman 2004) allowingthem to target and colonize specific niches in the hostType 1 fimbriae (
fim
) and pyelonephritis-associated pili(
pap
) are well characterized adhesins of uropathogenic
Ecoli
(UPEC) (Schilling
et al
2001 Mulvey 2002 Berglundand Knight 2003) The same UPEC strain often carriesdeterminants for both adhesins that are switched onoffin response to the local environment by a regulatory
442
J Bouckaert
et al
copy 2004 Blackwell Publishing Ltd
Molecular Microbiology
55
441ndash455
cross-talk between the
fim
and
pap
operons (Xia
et al
2000) As such specific binding is ensured by fimbrialadherence to distinct cell surface receptors Mannose-binding type 1 pili promote cystitis in the lower part of theurinary tract (Langermann
et al
1997 Ronald
et al
2001) by binding to a high-mannose glycoproteinuroplakin Ia (Old 1972 Firon
et al
1982 1984 Wu
et al
1996 Zhou
et al
2001) In ascending urinary tractinfections (UTIs) UPEC use P-pili to bind to galabiose-containing glycolipid receptors in the kidney and initiatepyelonephritis (Kallenius
et al
1980 Leffler andSvanborg-Eden 1980 Lund
et al
1987 Dodson
et al
2001) There has been controversy about the importanceand role of type 1 pili in the gastrointestinal tract (Bloch
et al
1992 Orndorff
et al
2004) and in biofilm formation(Pratt and Kolter 1998 Schembri
et al
2003) and itsreceptors are undefined in these environments
UPEC is the major causative agent of UTIs (Hooton andStamm 1997 Svanborg and Godaly 1997) UTIs are aserious health problem affecting almost one in every twowomen at least once in their lifetime The infections oftenrecur within the months after the primary infection despiteproper use of antibiotics UPEC can invade urothelial cellsin a type 1 pilus-dependent mechanism Once inside thecells they form intracellular bacterial communities suchas biofilms inside the bladder superficial umbrella cellscreating an environment protected from antibiotics and theimmune system of the host (Justice
et al
2004) Bacteriawithin these communities exhibit regional expression oftype 1 pili A subpopulation of UPEC can flux back out ofthe cells and reinvade to ultimately establish a quiescentreservoir that may serve as a seed for recurrent infections(Mulvey
et al
1998)Of the various UPEC adhesins type 1 pili are by far the
most prevalent (Brinton 1959 Buchanan
et al
1985Hultgren
et al
1985 Langermann
et al
1997 Bahrani-Mougeot
et al
2002) being expressed by a wide rangeof both pathogenic commensal and laboratory strains(Fukiya
et al
2004) Type 1 pili consist of a linear stringof repeating non-covalently linked immunoglobulin-like(Ig-like) FimA subunits wound into a rigid right-handedhelical rod followed by a short and stubby tip fibrillumcomposed of the FimF and FimG adaptor subunits andthe FimH adhesin The pilus is assembled by the chaper-oneusher pathway (Thanassi
et al
1998 Knight
et al
2000 Sauer
et al
2000) by complementing the incom-plete Ig fold of each constituent pilus subunit with anamino-terminal extension from a neighbouring subunit ina process termed lsquodonor strand exchangersquo (Choudhury
et al
1999 Sauer
et al
1999 2002 Zavialov
et al
2003) FimH is a two-domain adhesin protein at the endof the tip fibrillum responsible for the mannose-sensitivebacterial adhesion The amino-terminal lectin domain(residues 1ndash158) is joined to the carboxy-terminal pilin
domain (residues 159ndash279) that links the adhesin to therest of the pilus Bacterial two-domain adhesins such asFimH are not soluble unless their pilin domain is comple-mented by a lsquodonor strandrsquo However truncates ofadhesins corresponding to only the lectin domain havepreviously been shown to be properly folded and func-tional (Schembri
et al
2000 Dodson
et al
2001 Buts
et al
2003)The primary physiological receptor for FimH in the uri-
nary tract is the glycoprotein uroplakin Ia (Zhou
et al
2001 Min
et al
2002) that is abundantly present on dif-ferentiated uroepithelial cells However FimH recognizesa wide range of glycoproteins carrying one or more
N
-linked high-mannose structures FimH also binds yeastmannans and mediates agglutination of yeast cells FimH-mediated adhesion can be inhibited by
D
-mannose (Old1972 Hung
et al
2002) and a variety of natural andsynthetic saccharides containing terminal mannose resi-dues (Firon
et al
1982 1983 1984 1987 Neeser
et al
1986 Lindhorst
et al
1998 Nagahori
et al
2002) Block-ing of the FimHndashreceptor interaction has been shown toprevent bacterial adhesion to the bladder uroepitheliumand thereby infection (Langermann
et al
1997 2000Thankavel
et al
1997 Langermann and Ballou 2003)Increasing incidence of bacterial resistance to antibioticshas rejuvenated the interest in understanding the molec-ular basis for FimH-mediated adhesion to provide comple-mentary or alternative treatments of UTIs However forreasons becoming obvious in this article no binding anal-yses of FimHndashcarbohydrate interactions at the molecularlevel have previously been published Also no structuralinformation exists to explain the increased adhesion inhi-bition exhibited by oligomannosides (Firon
et al
19821983 1984 Neeser
et al
1986 Lindhorst
et al
1998) orby some aglycon-substituted mannosides (Firon
et al
1984 1987 Nagahori
et al
2002)In two independently determined crystal structures of
the FimH lectin domain of an
E coli
K-12 strain and auropathogenic strain respectively we made the serendip-itous discovery of a lsquostickyrsquo ligand butyl
a
-
D
-mannosidein the mannose binding site The heterogeneity caused bythis lsquostickyrsquo contaminant rendered several approaches formeasuring binding affinities of the purified FimH lectindomain for its natural carbohydrate receptors useless orat least highly unpredictable The origin of butyl
a
-
D
-man-noside was examined and this knowledge was used toproduce butyl
a
-
D
-mannoside-free FimH We determinedfor the first time FimH binding constants for several hex-oses including mannose two disaccharides two arylmannosides and synthetic alkyl mannosides with differentchain lengths We find that simple alkyl mannosides bindwith nanomolar affinities to FimH Also fructose bindsmore tightly to the FimH fimbrial adhesin than for exam-ple the PapGII adhesin to its natural receptor These
A novel class of FimH high-affinity ligands
443
copy 2004 Blackwell Publishing Ltd
Molecular Microbiology
55
441ndash455
highly specific small-molecule inhibitors targeting withhigh-affinity the FimH mannose binding site may preventUTIs by interfering with the attachment of
E coli
to theuroepithelial cells lining the bladder mucosa
Results
Affinity of FimH for mono- and disaccharides
Two different binding assays were developed one basedon surface plasmon resonance (SPR) and one based onequilibrium binding measurements using radioactivelylabelled mannose (Table 1) Inhibition by mannose of thebinding of FimH to the immobilized Fab fragments of themonoclonal antibody IC10 [k
a
= 147 yen 104 (plusmn417) M-1
s-1 kd = 411 yen 10-3 (plusmn565 yen 10-6)s-1 Kd = 280 yen 10-7
(plusmn88 yen 10-9) M] in SPR measurements allowed us toderive a solution affinity of mannose for FimH(Kd = 23 mM) (Table 1) Using either the Fabs of mono-clonal antibody IA7 or BSA-Man as the immobilized bind-ing partner in the competition experiment gave equivalentdissociation constants for the FimHndashmannose interaction(data not shown) To eliminate the possibility of differentbinding strength for full-length FimH and FimH truncatesthe binding of D-mannose to FimCFimH complex wasalso measured A value of Kd = 53 mM was obtained ingood agreement with the value measured using FimHtruncates
Removal of the 2-OH group of mannose leads to a 100-fold reduction in affinity for FimH (Table 2) Glucose theequatorial O2 epimer of mannose binds a 4000-foldweaker There is virtually no affinity for galactose the O4(axial) epimer and O2 (equatorial) epimer of mannoseFructose binds strongly to FimH (Kd = 31 mM) although a15-fold weaker than mannose The fructose-containingdisaccharides sucrose [Glc(a1ndash2)Frufb] and turanose[Glc(a1ndash3)Frub] have affinities for FimH very similar toglucose
Structure of the FimH lectin domain
The lectin domains FimH truncate 1 (FimHtr1) originatingfrom the UPEC strain J96 and FimH truncate 2 (FimHtr2)from the E coli K-12 laboratory strain MG1655 werecloned expressed and crystallized The structures weresolved by molecular replacement using the lectin domainfrom the FimCFimH complex (Choudhury et al 1999) asa search model and refined to 17 Aring (FimHtr1) and 21 Aring(FimHtr2) resolution respectively (Table S1 in Supplemen-tary material) Both models include residues 1ndash158 cor-responding to the entire lectin domain of FimH Twocopies of the lectin domain are present per asymmetricunit of the FimHtr2 crystals compared with only one unitfor the FimHtr1 crystals The carboxy-terminal 6-His tag ofFimHtr2 is not visible in the electron density maps and isnot included in the model
The structures of the FimH lectin domains are verysimilar to the structure for the same domain in theFimCFimH complex with a root mean square deviation(rmsd) over all 158 Ca positions of 036 Aring for FimHtr1056 Aring for FimHtr2 subunit A and 054 Aring for FimHtr2 subunitB The secondary structure elements arranged in an Igfold (Buts et al 2003) (Fig 1A) have almost interchange-able Ca traces in all of the FimH lectin domain structuresOnly Gly73 (in strand D1) and Gly79 (in loop connectingstrands D1 and Dcent) have slightly variable main chain con-formations while Val30 in the b3ndashb4 loop at the base ofthe domain clearly undergoes a peptide flip in all threesubunits of the truncate compared with the FimCFimHcomplex In the FimHtr1 structure the conformation of thecarboxy-terminal residue of the truncate Thr158 differsas it is packed against and hydrogen bonded to a glycerolmolecule via its carboxylate group In the FimHtr2 crystalstructure the main difference is in the b9ndashb10 loop withan rmsd of 29 Aring over Ca atoms 113ndash116 This loopadopts a different conformation because of crystal pack-ing contacts that are not present in the FimCFimH orFimHtr1 structures
Mannose binding pocket
Although no sugar was included in the crystallization set-ups strong positive electron density was present in the
Table 1 Kd and calculated DG0 for a series of O1 alkyl and arylmannosides
LigandKd SPR(nM)
DGinfin SPR (kcal mol-1)
Kd displace(nM)
DGinfin displace(kcal mol-1)
Mannose 23 yen 103 -76 41 yen 103 -73Methylaman 22 yen 103 -77 24 yen 103 -76Ethylaman 12 yen 103 -81 730 -83Propylaman 300 -89 400 -87Butylaman 151 -93 150 -93Pentylaman 25 -104 200 -91Hexylaman 10 -109 100 -95Heptylaman 5 -113 32 -102Octylaman 22 -104 28 -103PNPaMan 44 -100 26 -103MeUmbaMan 20 -105 12 -108
The SPR and displacement binding experiments define heptyl a-D-mannopyranoside as the best binder
Table 2 Kd and calculated DG0 for a series of mono- and disaccha-rides and a deoxymannose
Ligand Kd
DGinfin(kcal mol-1)
Methyl 2-deoxy-a-D-mannopyranoside 300 mM -48Glucose 924 mM -28Galactose 100 mM -14Fructose 31 mM -61Sucrose 128 mM -26Turanose 76 mM -29
444 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
initial FondashFc and 2FondashFc Fourier maps inside the mannose-binding pocket of both FimHtr1 and FimHtr2 This densityprecisely defined a-D-mannose with an additional elon-gated density shaped like a creased tail extending out-wards from the mannose O1 and towards tyrosines 48and 137 which make up part of the hydrophobic rimaround the mannose-binding pocket (Fig 1B) (Hunget al 2002) Further refinement strongly suggested thata butyl a-D-mannoside was bound in the mannose-bindingsite (Fig 1B and C) For FimHtr1 also a change in theconformation of the Tyr48 side-chain compared with theone from the model FimH used for molecular replacementwas immediately apparent As described previously (Hunget al 2002) the mannose ring makes 10 direct hydrogenbonds to the side-chains of residues Asp54 Gln133Asn135 and Asp140 and to the main chain of Phe1 andAsp47 and indirect water-mediated hydrogen bonds viaO2 to the side-chain of Glu133 and to the main chainoxygen of Phe1 and Gly14 The alpha-anomeric hydroxylgroup O1 of mannose is involved in a hydrogen-bondingwater network with the Asn138 and Asp140 side-chainsthrough a water molecule (wat 2 Fig 1B) that is con-
served between the two truncate structures The positionsof the other waters participating in this network dependon the crystal packing of FimHtr1 and FimHtr2
Mass spectrometric analysis of the bound ligand
To corroborate the nature of the ligand identified in thecrystal structures the complex of FimHtr1 and its pre-sumed butyl a-D-mannoside was analysed using massspectrometry The FimHtr1 protein sample used for crys-tallization was subjected to peracetylation and phase sep-aration The peracetylated ligand retrieved in the organicphase was analysed by nanoLC-online-ESI-MS The per-acetylated ligand eluted as a single peak and was regis-tered in its sodiated form at mz 427 corresponding tobutyl hexose (Fig 2A) No signals corresponding to otherpotential alkyl mannose species like ethyl hexose (mz399) propyl hexose (mz 413) or pentyl hexose (mz 441)could be observed indicating a homogeneous ligand witha four-carbon atoms alkyl chain Besides the sodiumadduct of the ligand a major signal was observed arisingfrom the loss of butanol (mz 331 Fig 2B) Cleavage at
Fig 1 Stereo views of FimHA Three-dimensional structure of the FimH lectin domain is an elongated 11-stranded b-barrel with an immunoglobulin fold Numbering is according to conventions established for antibody domains (Bork et al 1994) and for PapD (Sauer et al 1999) and loop identifications according to previous descriptions (reference strand numbers A1 = 1 Acent = 2 A2 = 3 B1 = 4 B2 = 4cent C = 5 D1 = 6 Dcent = 7 D2 = 8 E = 9 F = 10 G = 11) (Choudhury et al 1999 Hung et al 2002) The disulphide bridge between Cys3 and Cys44 is coloured cyanB Focus on the mannose-binding pocket in the same orientation as (A) showing the hydrophobic contacts (within 46 Aring orange) with butyl a-D-mannoside and the water network (salmon pink) present at O1C Surface presentation (wheat yellow) of the different alkyl conformations of the butyl a-D-mannoside in the crystal structures of FimH truncates 1 and 2 FimHtr1 butyl group (cyan) FimHtr1 Tyr48 (marine blue) FimHtr2 butyl group and Tyr48 (salmon pink) Tyr137 (violet) Ile52 (pink)
A B
C
A novel class of FimH high-affinity ligands 445
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
the anomeric carbon atom was often combined with elim-ination of one or more acetic acid molecules (massdecrease of 60 Da) or acetyl groups (mass decrease of42 Da) These eliminations gave rise to the major ions atmz 127 and mz 169 with a proposed aromatic ring struc-ture as well as to the ion at mz 109 with a proposedquinone-like ring structure Thus the mass spectrometricdata support the butyl hexose nature of the FimH ligandFimH was further characterized by carbohydrate constit-uent analysis (data not shown) which revealed only man-nose Mannose could only be detected after hydrolysisindicating that free mannose is not associated with theFimH truncate
Origin of butyl a-D-mannoside
As no sugar had been advertently added to the FimHtr
protein samples we hypothesized that the butyl manno-side ligand had been picked up from the LuriandashBertani(LB) medium used to grow bacteria during expression ofthe protein To test this FimHtr2 was expressed in bacteriagrown in minimal M9 medium Electro-spray ionizationmass spectrometry (ESI-MS) was performed on purifiedFimHtr2 samples from both types of media A clear peakat 259 Da could be detected (Fig 3A) with proteinexpressed in bacteria grown in LB medium correspondingto the molecular weight of butyl mannoside (236 Da) plus
a sodium ion (23 Da) As a control butyl mannoside wassynthesized and analysed in the same manner Again apeak at 259 Da was observed (Fig 3A inset) When ESI-MS was run on protein expressed in bacteria grown inminimal medium no such peak could be detected(Fig 3B) indicating that the ligand was not present andthat it indeed originated from the LB medium
Interactions of the butyl moiety with FimH
The butyl moiety of butyl a-D-mannoside extends out ofthe mannose-binding pocket towards Tyr48 and Tyr137making van der Waals contacts to both tyrosine rings andIle52 (Fig 1B and C) These residues are part of two loops(b4ndashb5 and b10ndashb11) that form the higher rear end of ahydrophobic collar around the binding site (Fig 1A andB) In both subunits of the FimHtr2 structure the Tyr48 andTyr137 side-chains are in the same almost parallel ori-entation as in the FimCFimH structure forming a gate-like structure at the back of the binding site (Fig 1C) Theclosest hydrophobic interactions (31ndash36 Aring) with the alkylchain are made between Tyr48 and the two non-terminalatoms of the butyl In the FimHtr1 structure a crystal con-tact to Val155 of a neighbouring molecule prevents theparallel orientation of the Tyr48 ring which instead ispacked edge-to-face with Tyr137 (Fig 1B) The reposition-ing of Tyr48 is accompanied by a change in the confor-
A
B
Fig 2 Mass spectrophotometric characteriza-tion of the butyl mannosideA Base peak chromatogram (BPC mass range mz 400ndash550) and extracted ion chromato-grams (EIC) of mz 427 and 331 obtained by nano-LC-ESI-MS of the derivatized FimH-ligand purified by peracetylation and organic-solvent extractionB ESI-IT mass spectrometric characterization of the peak species marked by a horizontal bar in (A) Ions are marked with mz-values and proposed schematic structures which is often only one out of several isobaric ion species Fragmentations already occurred in the source Ac acetyl
446 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
mation of the butyl chain of the ligand with the butyl chainin a more extended conformation than in the FimHtr2 struc-ture and with the four carbon atoms nearly in one plane(Fig 1B and C) The new conformations of Tyr48 and thebutyl moiety result in an almost parallel stacking of thearomatic ring of Tyr48 onto the butyl plane with the closesthydrophobic interactions (37 Aring) involving the two terminalcarbon atoms of the butyl (Fig 1B)
Affinity measurements for alkyl mannosides
The butyl mannoside ligand remained bound to FimHtr
despite extensive dialysis From a study of complex for-mation of monoclonal antibodies IC10 and IA7 withFimHtr1 it could be estimated that about 30 of the FimHtruncate retained its ligand after dialysis for 12 daysagainst 50 mM MES buffer at pH 65 100 mM NaCldespite daily refreshment of buffer (data not shown)Therefore to enable measurements of dissociation con-stants of FimH for mannosides FimH was produced inminimal medium Butyl a-D-mannoside binding is 15 timesimproved (Kd = 151 nM Fig 4A) compared with mannosebinding The equilibrium binding technique using [3H]-mannose led to a similar value (Kd = 41 mM Fig 4B) for
mannose and an almost identical value for butyl a-D-mannoside (Kd = 150 nM Fig 4C) To investigate theeffect of sequential addition of methyl groups to the O1oxygen of D-mannose a series of alkyl mannosides weresynthesized and the dissociation constants determinedusing the [3H]-mannose displacement assay as well as theSPR competition assay (Table 1) There is a linear corre-lation between the binding free energy as calculated fromthe measured dissociation constants and the number ofmethyl groups (between 1 and 7) in the alkyl mannosidewith each additional methyl group contributing on average-06 kcal mol-1 of binding energy (Fig 5)
Affinity measurements and modelling for aryl mannosides
Aromatically substituted mannosides have been reportedto be particularly potent inhibitors of FimH-mediated bac-terial adhesion (Firon et al 1987 Nagahori et al 2002)In accordance with the earlier results both pNPaMan andMeUmbaMan bind very tightly to FimH (Table 1) Despiteextensive trials no X-ray structures were obtained forcomplexes with aromatically substituted mannosides Inorder to model binding docking to both of the truncatestructures was carried out In all cases the mannose ring
Fig 3 Mass spectrometric analysis of the origin of the butyl mannosideA LB medium-produced FimH truncate The inset shows the mass spectrum of the synthesized butyl a-D-mannosideB Minimal media-produced FimH truncate
A novel class of FimH high-affinity ligands 447
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
docks in the binding pocket in a conformation very closeto the crystallographically observed mannose conforma-tion (Fig 6) while the aromatic substituent interacts withthe tyrosine gate The different conformation of Tyr48 in
our two FimH truncate structures leads to different pre-dicted interactions between the aromatic aglycon andTyr48 and Tyr137 Docking to the FimHtr2 structureresulted in somewhat tighter interactions between thetyrosine gate and the aglycon and hence in somewhatlower docking energies Calculated FimHtr2 docking ener-gies of Edoc = -104 kcal mol-1 for pNPaMan (Kd = 46 nM)and Edoc = -109 kcal mol-1 for MeUmbaMan (Kd = 20 nM)are in good agreement with the experimental binding data(Table 1) The docked umbelliferyl ring system is inserteddeep into the tyrosine gate in the FimHtr2 conformationwith the carbonyl group within hydrogen bonding distanceto Thr51 at the back of the gate (Fig 6A) Docking to theFimHtr1 structure instead stacks the umbelliferyl ring face-to-face with the Tyr48 ring (not shown) The phenyl ring ofpNPaMan docks similarly to both our structures It doesnot quite reach in between the tyrosine side-chains butinstead hydrogen bonds with its nitro group to the hydroxyloxygen of Tyr137 (Fig 6B)
Discussion
Most lectins bind naturally occurring carbohydrates withlow affinities often in the millimolar range for monosac-charides and micromolar range for oligosaccharides Thiswas recently exemplified to also hold true for theadhesins of F17 fimbriae (Kd = 12 mM for the F17a-G-N-acetylglucosamine interaction Buts et al 2003) and Ppili (Kd = 84 mM for the PapGII-globotetraoside interactionLarsson et al 2003) The recent crystal structure of FimHin complex with mannose illustrated that FimH almostcompletely envelops the monosaccharide in a deeppocket and involves it in an intense hydrogen bondingnetwork suggestive of an unusual lectin exerting highaffinities (Hung et al 2002)
Fig 4 FimH binding experimentsA Surface plasmon resonance competition experiment of FimHtr1 binding to monoclonal antibody IC10 Fab fragments with butyl a-D-mannosideB Equilibrium binding of [3H]-mannose to FimHtr2C Equilibrium displacement measurements of butyl a-D-mannoside to the FimHtr2 [3H]-mannose complex
log[Butyl a-D-mannoside] (M)
cpm
cpm
Butyl a-D-mannoside (mM)
Mannose (mM)
A
B
C
ndash64
ndash66
ndash68
ndash70
ndash72
ndash74
ndash76
ndash78
ndash80
ndash82
ndash8 ndash7 ndash6 ndash5 ndash4 ndash3
log
[Fim
H]
M
Fig 5 Linear free energy relationship for the alkyl chain on mannose A linear fit was made using 1 up to 7 methyl groups with the program MICROCAL ORIGIN 50
Number of carbon atoms0 2 4 6 8
ndash75
ndash80
ndash90
ndash95
ndash100
ndash105
ndash110
ndash115
ndash85
D G
(kc
al m
olndash1
)
slopecorrelation
ndash064ndash099
+ndash 003+ndash 018
448 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
The measured affinity confirm the predictions that FimHbinds the monosacharide mannose exceptionally well(Kd = 23ndash41 mM) To investigate the contribution of thedifferent hydroxyl groups of mannose in the interactionwith FimH the binding of a series of hexoses to butylmannoside-free FimH was examined using SPR (Table 2)The effect of deoxygenation of the O2 hydroxyl group(methyl 2-deoxy-a-D-mannopyranoside) is clearly lessdestructive (100-fold reduction) than its equatorial positionas in glucose O2 is the only hydroxyl group of mannosethat interacts indirectly with the FimH pocket via a con-served water molecule (Hung et al 2002) Glucose onlydisplays a weak affinity for FimH about 4000 times lowerthan methyl a-D-mannoside Repositioning of both the O2and O4 hydroxyl groups as in galactose abolishes sugarbinding by FimH Similar to these results the inhibitoryconcentrations for bacterial adherence of methyl a-D-glucoside and galactose derivatives have been estimatedto be respectively 104-fold and 105-fold weaker than formethyl a-D-mannoside (Nagahori et al 2002)
In stark contrast with the other monosaccharides fruc-tose has a binding affinity only about 15-fold less thanmannose for FimH (Table 2) The decrease in affinity for
fructose relative to mannose agrees with studies of inhi-bition of adherence of type 1 piliated E coli by fructosein which fructose was found to be the active compound infruit juices (present at about 5) (Zafriri et al 1989)Fructose binding to FimH is stronger than the binding ofthe physiological globotetraoside receptor to PapGII theother well-characterized adhesin of UPEC strains (Lars-son et al 2003) The pyranose form of fructose is verywell superimposable onto mannose in the FimH bindingsite where similar interactions are established with FimHas for mannose except for the loss of the anomeric O1and an extra hydroxyl group (O2) on the C5 position ofmannose Probably however both changes of fructopyr-anoside compared with mannopyranoside could be easilyaccommodated by FimH The mannose O1 does notmake direct interactions within 45 Aring of the FimH bindingpocket (Fig 1B) The extra O2 group of fructose would notclash or interfere significantly with the binding pocket butit would make a 27 Aring van der Waals contact with the Ile52side-chain under the condition that the Ile52 side-chaindoes not move The crystal structure of the Pseudomonasaeruginosa lectin LecB also showed that fructopyranosebut not fructofuranose can substitute for mannose (Loris
Fig 6 Stereo diagrams of docked MeUm-baMan (A) and pNPaMan (B) in the FimH bind-ing site The crystallographically determined position of a-D-mannose is shown in orange for comparison
A novel class of FimH high-affinity ligands 449
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
et al 2003a) Hydrogen bonds of the more specificfucose complex are replaced by van der Waals interac-tions involving a hydroxyl group in the complexes with thelower-affinity binders mannose and fructopyranose as forthe proposed fructopyranoside binding to FimH
The fructose-containing disaccharides sucrose andturanose do not have an inhibitory effect and bind withaffinities similar to glucose Sucrose is a non-reducingsugar that has fructose locked in its furanose form Tura-nose on the other hand allows the interconversion of b-D-fructofuranose and b-D-fructopyranose eg both formsoccur in complex with a plant lectin from Pterocarpusangolensis (Loris et al 2003b) It is predictable howeverfrom the crystal structures that the glycosidic linkage ofglucose with C3 of fructose equivalent in position to C4of mannose would be disallowed in the tight FimH pocketSucrose and turanose thus most probably interact with theFimH mannose binding pocket via their glucose moietyresulting in similar low affinities
Our mass spectrometry and chemical analyses con-firms the crystallographic identification of the ligand boundto the FimH lectin domains as butyl a-D-mannoside(Figs 1B and 2) The mannose ring of the butyl a-D-mannoside coincides with the mannose bound to theFimCFimH complex (Hung et al 2002) and makes thesame interactions The four-carbon alkyl tail exits towardsthe back wall (loop b4ndashb5 and b10ndashb11) of the pocket(Fig 1A and B) making hydrophobic contacts to Tyr48Tyr137 and Ile52 The mass spectrometry data also showthat the bound butyl a-D-mannoside is present in LBmedium but absent from minimal medium Hence thesource of the alkyl mannoside is possibly the yeast extractused in preparation of LB medium Although we do notwant to speculate about the significance of this findingalkyl mannoside production by yeast has not beenreported and might warrant investigation
The butyl mannoside was retained by FimH despiteextensive dialysis The phenomenon of ligand retention byprotein was described as early as 1975 for periplasmicbinding proteins (Silhavy et al 1975) Applied to dialysisof butyl mannoside away from FimH grown in richmedium half-lives of up to 30 h can be estimated whichis far beyond the duration of a normal buffer exchangedialysis experiment In the present case the retainedligand does not appear to seriously affect [3H]-mannosedisplacement equilibrium binding experiments as wecould detect no significant differences in dissociation con-stants obtained using FimHtr2 produced in LB or minimalmedia (data not shown) In contrast proper SPR bindingcurves could not be obtained when butyl mannoside occu-pied the binding site Hence the displacement techniquemay be of more general use to obtain binding constantsfor proteins that tend to inadvertently retain ligands whichis a common trait of lectins
Surface plasmon resonance as well as equilibrium bind-ing and displacement measurements using [3H]-mannosewere carried out to obtain dissociation constants for a-D-mannose and a linear series of alkyl-substituted manno-sides to FimH (Table 1) Both equilibrium binding experi-ments show a very good agreement in their measurementof the carbohydrate affinity of the adhesin FimH bindsbutyl a-D-mannoside 15ndash30 times stronger than a-D-mannose The importance of the added hydrophobiccontacts is clearly demonstrated by the binding of alkylmannosides (Table 1) The best binding alkyl mannosidesbind a few hundred times stronger than mannose equiv-alent in affinity to the most tightly binding aromaticallysubstituted mannosides for FimH (Firon et al 1987) andof mannose dendrimers for type 1 piliated E coli (Fironet al 1984 1987 Nagahori et al 2002)
Interestingly the butyl moiety takes on significantlydifferent conformations in FimHtr1 and FimHtr2 (Fig 1C)indicating that intrinsic conformational flexibility in theaglycon is allowed Indeed in spite of the larger flexibilityexpected for longer alkyl mannosides the binding energydecreases linearly with increasing length of the alkyl chainbetween one and seven carbon atoms (Fig 5) The affinityreaches its maximum for mannose with a seven-carbonalkyl and SPR measurements suggest that the affinitydecreases from eight carbon atoms on Evidently theeighth carbon atom goes beyond the hydrophobic interac-tion surface and protein edge which is penalized with adecrease in affinity We propose that the affinity increaseresults from an increased interaction surface with thehydrophobic gateway Alkyl aglycons fit well in the tyrosinegate and as observed in the two crystal structures in atleast two different conformations that depend on theconformation of Tyr48 This flexibility may indicate thataglycons can interact rather non-specifically with thehydrophobic gateway nevertheless resulting in a 100- to1000-fold increase in affinity
FimH binding to ligands other than just mannose ispossibly of importance in terms of the different environ-ments that E coli will encounter ndash the urinary tract gas-trointestinal tract and in biofilms Alkyl mannosides havenot been previously recognized as strong binders to FimHAs they are easily synthesized and highly soluble in waterthey may be interesting as potential blocking agents forFimH-mediated adhesion Moreover they have beenshown to be good drug vehicles themselves acting aspermeability enhancers on tight junctions (Eley and Triu-malashetty 2001) As such they might be able to pene-trate into deeper cell layers of the epithelium to alsoprevent those bacteria that form a quiescent reservoirfrom causing recurrent infections (Mulvey et al 1998Justice et al 2004)
On the other hand the two aromatically substitutedmannosides pNPaMan and MeUmbaMan are known to
450 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
be particularly potent inhibitors of FimH-mediated bacte-rial adhesion (Firon et al 1987 Nagahori et al 2002)Based on adhesion inhibition studies using type 1 piliatedE coli and guinea pig ileal epithelial cells the relativeinhibitory potential of pNPaMan and MeUmbaMan havebeen estimated to be about 70 and 600 times strongerrespectively than that of methyl a-D-mannose (Fironet al 1987) Our measurements indicate an affinity ofFimH for pNPaMan and MeUmbaMan between 50 and90 times and between 110 and 200 times higher respec-tively than for methyl a-D-mannose in good agreementwith the adhesion inhibition results (Firon et al 1987)Dissociation constants predicted using AutoDock3 (Morriset al 1998) are similar to experimental values Thecomputational docking predicts that tight binding of thearomatically substituted mannosides pNPaMan andMeUmbaMan is accomplished similarly as for butyl a-D-mannoside with the aglycon interacting with the tyrosinegate
This is the first systematic study providing molecularbinding data for the FimH adhesin the prerequisite forcolonization by UPEC (Langermann and Ballou 2003)We also show crystallographic evidence that the lectinor receptor binding domain of FimH is a stable andfunctional entity Its structure is virtually identical to thesame domain in the FimCFimH complex indicating thata truncate corresponding to the lectin domain could pro-vide a good alternative for the development of a UTIvaccine (Langermann et al 1997 Langermann and Bal-lou 2003) However progress of this interesting UTI vac-cine candidate into clinical trials has not been reportedThe binding affinities of FimH at the molecular levelrelate convincingly well with the inhibitory concentrationsused in previous bacterial adhesion assays thus endors-ing the direct and conclusive correlation of FimH withbacterial adherence Alkyl mannosides have nanomolaraffinities for FimH which may overcome the significantobstacle of low affinity generally encountered in attemptsto develop carbohydrate-based drugs which interferewith microbial attachment to host cell surface glycocon-jugates New anti-adhesive drugs could be based onthese leads to allow specific interaction and targeting tothe urinary tract in order to prevent anticipated andrecurrent infections
Experimental procedures
Strains and constructs
FimH proteins from two different E coli strains the UPECstrain J96 (Hull et al 1981) and the E coli K-12 strainMG1655 (Blattner et al 1997) were used in this study TheFimH adhesins encoded by these two E coli strains areidentical FimHtr1 (residues 1ndash158 of UPEC J96 FimH) wasexpressed from plasmid pMMB91 transformed into E coli
C600 cells FimHtr2 (residues 1ndash158 of E coli MG1655FimH + carboxy-terminal 6-His tag) was expressed fromplasmid pPKL241 (Schembri et al 2000) transformed into Ecoli host strain HB101
Purification and crystallization
FimHtr1 Bacteria were grown in LB medium at 37infinC inducedat OD600 = 08 with 1 mM IPTG and harvested 2ndash3 h laterThe cells were resuspended in 20 mM Tris-HCl pH 80 5 mMEDTA and 30 sucrose and the periplasmic content wasextracted by osmotic shock upon 10-fold dilution with 20 mMTris-HCl pH 80 Purification of FimHtr1 was performed by ionexchange chromatography with MonoS at pH 45 followed byMonoQ (Amersham Biosciences) at pH 8 FimHtr1 was crys-tallized using vapour diffusion in hanging drops composed of3 ml of the precipitant solution (01 M Hepes pH 75 12PEG 6000 and 5 vv methyl-24-pentanediol) and 3 ml of12 mg ml-1 protein solution in 20 mM Tris-HCl pH 80 with150 mM NaCl
FimHtr2 Cells were grown in LB medium containing50 mg ml-1 ampicillin at 37infinC At OD600 = 06 the cells wereinduced with 5 mM IPTG Cells were harvested 2 h afterinduction and the periplasmic content was extracted Theprotein was purified on a Pharmacia HiTrap Chelating HP5 ml column (Pharmacia) loaded with Ni-chloride and elutedwith a sharp 0ndash500 mM imidazole gradient Fractions con-taining FimHtr2 were pooled dialysed against 50 mM sodiumacetate pH 525 and loaded onto a Mono S HR 8 ml columnThe protein was eluted with a 0ndash500 mM NaCl gradient anddialysed overnight against 20 mM MES pH 65 before crys-tallization Crystallization was performed at room tempera-ture by the vapour diffusion method with hanging dropsmade by mixing of 2 ml of 15 mg ml-1 protein solution with 2 mlof the precipitant solution (65 methyl-24-pentanediol100 mM cacodylate pH 65) Microseeding was employed tooptimize crystal quality and reproducibility Crystals suitablefor X-ray data collection generally appeared within 48 h aftermicroseeding
Structure determination
X-ray data for FimHtr1 were collected from a single crystalto 17 Aring resolution at the Advanced Photon SourceArgonne National Laboratory USA The crystal was flash-cooled to 100 K in the precipitant solution complementedwith 25 glycerol The data were processed and scaledusing the HKL2000 package (Otwinowski and Minor 1997)(Table S1 in Supplementary material) Data for FimHtr2
were collected from a single crystal under liquid-nitrogencryo conditions at 100 K at beamline ID29 ESRF Greno-ble France to 21 Aring Bragg spacing FimHtr2 data were pro-cessed and reduced using MOSFLM (Powell 1999) andscaled using SCALA (Collaborative computational project1994) (Table S1 in Supplementary material) The structuresof FimHtr1 and FimHtr2 were solved by molecular replace-ment using Amore (Navaza 2001) and MOLREP (Vaginand Teplyakov 1997) respectively with the FimH lectindomain from the FimCFimH complex (Choudhury et al
A novel class of FimH high-affinity ligands 451
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
1999) (PDB id 1QUN) as the search model Positional bulksolvent and isotropic temperature factor refinement wasperformed using CNS (Bruumlnger et al 1998) and REFMAC5(Murshudov et al 1997) respectively Model adjustmentand inspection were performed using Turbo-Frodo and O(Jones et al 1991) graphics programs respectively Refine-ment statistics are given in Table S1 Molecular graphicsillustrations were generated with Bobscript (Esnouf 1997)and Pymol 090 (DeLano Scientific LLC 1998ndash2003 httppymolsourceforgenet)
Mass spectrometry
The purified FimHtr1 protein (300 mg) that had been used forcrystallization was freeze-dried taken up in 100 ml of pyridineand 100 ml of acetic anhydride and incubated overnight atroom temperature in the dark for peracetylation The samplewas dried under nitrogen and taken up in chloroform (400 ml)The chloroform phase was extracted six times with waterdried and taken up in 200 ml of 50 aqueous methanol A1 ml aliquot was applied to a nano-scale RP column(75 mm yen 100 mm PepMap DionexLC Packings) using anUltimate nano-LC system (LC Packings) which was equili-brated with eluent A (H2Oacetonitrile 955 vv containing01 formic acid) at a flow rate of 100 nl min-1 After injectingthe sample the column was run isocratically for 5 min fol-lowed by a linear gradient to 100 eluent B (H2Oacetonitrile2080 vv containing 01 formic acid) in 20 min and a finalwash with 100 B for 5 min The system was directly coupledto an Esquire 3000 ESI-IT-MS (Bruker Daltonik) equippedwith an online nanospray source operating in the positive-ionmode For electrospray (900ndash1200 V) capillaries (360 mmOD 20 mm ID with 10 mm opening) from New Objective wereused The solvent was evaporated at 150infinC with a nitrogenstream of 6 l min-1
To verify the suspected source of the bound butyl manno-side mass spectrometry was performed on samples ofFimHtr2 grown in either LB or minimal M9 medium Proteinsamples were dialysed against water overnight before massspectrometry in order to reduce background from the Trisbuffer Samples were diluted to a concentration in the submi-cromolar range Electrospray mass spectra were recorded indirect inlet mode (50 aqueous methanol solutions) on aBruker Daltonics Esquire-LC (Billerica)
Carbohydrate constituent analysis
Samples were hydrolysed in 100 ml of 4 M aqueous trifluoro-acetic acid (Sigma) at 100infinC for 4 h and dried under astream of nitrogen Monosaccharides were converted intotheir anthranilic acid derivatives by reductive aminationresolved by RP-HPLC and detected by fluorescence (Anu-mula 1994)
Carbohydrates for binding studies
[3H]-mannose was obtained from Amersham Methyl a-D-mannoside glucose galactose fructose sucrose turanosep-nitrophenyl a-D-mannopyranoside (pNPaMan) and 4-methylumbelliferyl a-D-mannopyranoside (MeUmbaMan)
were obtained from Sigma Monomannosylated bovineserum albumin (BSA-Man) was kindly provided by MedIm-mune We synthesized methyl 2-deoxy-a-D-mannopyrano-side (Oscarson and Tedebark 1995) Alkyl mannosides weresynthesized through silver triflate-promoted couplings of thecorresponding alcohol with 2346-tetra-O-benzoyl-a-D-mannopyranosyl bromide followed by Zempleacuten deacylationof the obtained protected alkyl mannosides according to theprocedure reported for the octyl and tetradecyl mannosides(Oscarson and Tiden 1993)
Binding studies
Competition experiments using SPR To obtain butyl man-noside-free FimHtr1 the FimH truncate was overproducedwith a carboxy-terminal 6-His tag after transformation into Ecoli C43 (DE3) (Miroux and Walker 1996) and the bacteriagrown in minA minimal medium supplemented with the 20amino acids (40 mg l-1) thiamin (2 mg l-1) and biotin (2 mgl-1) Purification of the His-tagged protein was performedusing Ni-NTA affinity chromatography (Qiagen) in 50 mM Tris-HCl pH 8 and 300 mM NaCl Elution was in the same solutioncomplemented with 500 mM imidazole Subsequently pooledand concentrated FimH fractions were gel filtrated over aSuperdex G75 column (Pharmacia) in 20 mM Tris-HCl pH 8
We used Fab fragments of a monoclonal antibody againstthe mannose-binding site of FimH to determine the solutionaffinity of FimHndashcarbohydrate interactions The monoclonalantibody IC10 was produced by a mouse hybridoma cell lineat Medimmune and its Fab fragments were prepared andpurified The Fabs were covalently immobilized via aminecoupling at 1000 Resonance Units (1000 pg ligand mm-2) inflowcell Fc2 on a CM5 biasensor chip (BIAapplications Hand-book Biacore AB Uppsala Sweden) Flowcell Fc1 was usedas the reference surface and was treated in the same wayas Fc2 except that it was blocked immediately after activationThe immobilization buffer was 100 mM sodium acetatepH 50 with 100 mM NaCl All SPR measurements were per-formed on a Biacore3000trade
The kinetic constants of binding of FimH to the immobilizedantibody were measured with different concentrations ofFimH (2000 nM 1000 nM 500 nM 250 nM 125 nM625 nM 312 nM 156 nM 78 nM) in running buffer [phos-phate-buffered saline (PBS) with 0005 surfactant P20 and3 mM EDTA] on both flowcells Fc1 and Fc2 simultaneouslyat a flow rate of 30 ml min-1 and at 298 K Complete dissoci-ation of FimH was performed with running buffer beforestarting a new binding cycle For all measurements the asso-ciation time was 3 min and the dissociation time was 30 minAll binding cycles were performed in duplicate including azero concentration cycle of FimH (injection of running buffer)All analyses were performed with the BIAeval software ALangmuir binding isotherm with a 11 stoichiometry was fittedto the data from both the association phase and the dissoci-ation phase after subtraction of the reference and zero con-centration data The kinetic constants ka and kd and themaximal binding Rmax were obtained and the dissociationconstant Kd was calculated
The equilibrium dissociation constants Kd of FimHndashsaccha-ride interactions were determined in a competition experi-ment using the same chip as for the previous experiment
452 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
As a control the Fabs of monoclonal antibody IA7 as wellas BSA-Man were also immobilized Samples containing afixed concentration of FimH were used (close to the Kd of theFimHndashantibody interaction) in combination with varying con-centrations of saccharide First 10-fold dilutions of the sac-charide solution were used to determine the concentrationrange for binding of the saccharide to FimH A Langmuirbinding isotherm with a 11 stoichiometry was fitted to thedata using the kinetic constants and Rmax from the foregoingexperiment to obtain the concentrations of FimH that werefree ([FimH]free) to bind the antibody immobilized on the chipSecond the concentration range of the saccharide wasextended and adapted to ensure accurate fitting and theequilibrium binding constant of the FimHndashsaccharide interac-tion was derived from the curve of [FimH]free against concen-tration of saccharide Every measurement was repeated atleast twice including testing different protein batches andwhenever possible also different saccharide stock solutions(typically 200 mM)
Equilibrium binding studies using radioactively labelledmannose Mannose-free FimHtr2 was produced inM9 minimal medium and purified as described above forFimHtr2 Alkyl mannosides were dissolved in double distilledwater pNPaMan and MeUmbaMan (6 mg each) were dis-solved in 20 ml of DMSO and diluted to 20 mM using doubledistilled water Binding experiments were performed at 310 Kusing six different concentrations of [3H]-a-D-mannose (finalconcentrations 435 mM 290 mM 193 mM 129 mM 86 mMand 57 mM) Protein (180 ml) at a concentration of about500 nM was mixed with 20 ml of the radioactive ligand [3H]-a-D-mannose and incubated at 37degC for 20 min To separatefree ligand from bound the mixture was rapidly filtratedthrough a Protran BA 85 cellulose nitrate filter (Schleicherand Schuell) and washed once with 1 ml of ice-cold 1yen PBSFilter-bound radioactivity was measured by scintillation spec-trometry within 24 h Displacement experiments were per-formed using six different concentrations of inhibitor (finalconcentrations in the range 00ndash435 mM) in the presence of435 mM [3H]-a-D-mannose Twenty microlitres of radioactiveligand 20 ml of inhibitor and 160 ml of protein (500 nM) weremixed and the experiments were performed as describedabove All experiments were performed in duplicates and thedata presented are mean values of at least two independentmeasurements For the determination of Kd for a-D-mannosean hyperbolic curve (y = P1x(P2 + x) where P1 is an arbitraryscale factor and P2 = Kd) was fitted to the data For the dis-placement experiments the curve y = P1(P2 + x) where P1
is an arbitrary scale factor and P2 is the concentration of theinhibitor displacing 50 of the labelled ligand [I]05 was usedinstead To calculate the inhibitor dissociation constant (KI)the Cheng and Prusoff equation [KI = [I]05[L]KL + 1) KL is theconstant of dissociation for the ligand] (Cheng and Prusoff1973) was used when both the concentration of the radio-active ligand (L) and the displacing agent (I) are in excessover the protein (LT gtgt PT IT gtgt PT T indicates total concen-tration) For the two strong inhibitors pNPaMan and MeUm-baMan where IT was not in excess over PT the equationof KI = IT[(1 - Y)Y(LTKL) - 1] - PTKLYLT where Y is thefraction of the ligand bound in presence of the inhibitor wasused instead (Horovitz and Levitzki 1987)
Computational docking
Computational docking studies were performed usingAutoDock3 (Morris et al 1998) In AutoDock3 the proteinis represented by a grid of affinity potentials Hence theprotein conformation is not allowed to change during thedocking procedure Flexibility of ligands is taken intoaccount by allowing rotation around flexible torsion anglesThe docked energies are calculated as the sum of the inter-molecular interaction energy and the internal energy of theligand The search for the best interaction energy was car-ried out by means of the Lamarckian genetic algorithmLigand structures were obtained from the Protein DataBank and prepared for docking by adding hydrogens andcharges using AutoDockTools (Sanner et al 1996) Eachsimulation consisted of 100 independent runs with a popu-lation size of 200 500 generations and a maximum of25 000 000 energy evaluations Solutions were rankedbased on their docking energies and similar solutions wereclustered (cluster cut-off rmsd lt 1 Aring) For both ligandsthe top solutions were evaluated using the graphics pro-gram O (Jones et al 1991)
Acknowledgements
We thank Andrej Zavialov and Maringhns Ehrenberg for help withequilibrium binding experiments and Thomas Norberg forassistance with mass spectrometry Anthony DeFusco fromMedimmune provided us with mannosylated bovine serumalbumin We are grateful to Fred Sauer for assistance withdata collection Thanks to Maor Bar-Peled of the CCC at theUniversity of Georgia Athens USA for his suggestion toexpress FimH in minimal medium We acknowledge the useof beamline 19BM of the Structural Biology Center at theAdvanced Photon Source Argonne National LaboratoryChicago IL USA and of beamline ID29 ESRF GrenobleFrance JB is a postdoctoral fellow of the Fonds voor Weten-schappelijk Onderzoek-Vlaanderen which also supportedthe BiaCore3000 instrument (FWONL35) This work was sup-ported by grants from the Swedish Research Council andfrom the Strategic Research Foundation (Structural BiologyNetwork) to SDK
Supplementary material
The following material is available from httpwwwblackwellpublishingcomproductsjournalssuppmatmmimmi4415mmi4415smhtmTable S1 Crystallographic data and refinement statistics
References
Anumula KR (1994) Quantitative determination ofmonosaccharides in glycoproteins by high-performanceliquid chromatography with highly sensitive fluorescencedetection Anal Biochem 220 275ndash283
Bahrani-Mougeot FK Buckles EL Lockatell CV HebelJR Johnson DE Tang CM and Donnenberg MS(2002) Type 1 fimbriae and extracellular polysaccharidesare preeminent uropathogenic Escherichia coli virulence
A novel class of FimH high-affinity ligands 453
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
determinants in the murine urinary tract Mol Microbiol 451079ndash1093
Berglund J and Knight SD (2003) Structural basis forbacterial adhesion in the urinary tract Glycobiol Med 53533ndash52
Blattner FR Plunkett G Bloch CA Perna NT BurlandV Riley M et al (1997) The complete genome sequenceof Escherichia coli K-12 Science 277 1453ndashbullbull
Bloch CA Stocker BA and Orndorff PE (1992) A keyrole for type 1 pili in enterobacterial communicability MolMicrobiol 6 697ndash701
Bork P Holm L and Sander C (1994) The immunoglob-ulin fold ndash structural classification sequence patterns andcommon core J Mol Biol 242 309ndash320
Brinton CC (1959) Non-flagellar appendages of bacteriaNature 183 782ndash786
Bruumlnger AT Adams PD Clore GM DeLano WLGros P Grosse-Kunstleve RW et al (1998) Crystallog-raphy and NMR system a new software suite for macro-molecular structure determination Acta Crystallogr D BiolCrystallogr 54 905ndash921
Buchanan K Falkow S Hull RA and Hull SI (1985)Frequency among Enterobacteriaceae of the DNA-sequences encoding type-1 pili J Bacteriol 162 799ndash803
Buts L Bouckaert J De Genst E Loris R Oscarson SLahmann M et al (2003) The fimbrial adhesin F17-G ofenterotoxigenic Escherichia coli has an immunoglobulin-like lectin domain that binds N-acetylglucosamine MolMicrobiol 49 705ndash715
Cheng Y and Prusoff WH (1973) Relationship betweeninhibition constant (K1) and concentration of inhibitor whichcauses 50 per cent inhibition (I50) of an enzymatic reac-tion Biochem Pharmacol 22 3099ndash3108
Choudhury D Thompson A Stojanoff V LangermannS Pinkner J Hultgren SJ and Knight SD (1999) X-ray structure of the FimCndashFimH chaperonendashadhesin com-plex from uropathogenic Escherichia coli Science 2851061ndash1066
Collaborative computational project n4 (1994) The CCP4suite programs for crystallography Acta Crystallogr D BiolCrystallogr 50 760ndash763
Dodson KW Pinkner JS Rose T Magnusson G Hult-gren SJ and Waksman G (2001) Structural basis of theinteraction of the pyelonephritic Escherichia coli adhesin toits human kidney receptor Cell 105 733ndash743
Eley JG and Triumalashetty P (2001) In vitro assessmentof alkylglycosides as permeability enhancers AAPSPharmSciTech 2 (3) Article 19
Esnouf RM (1997) An extensively modified version of Mol-script that includes greatly enhanced coloring capabilitiesJ Mol Graph Model 15 132ndash134
Firon N Ofek I and Sharon N (1982) Interaction ofmannose-containing oligosaccharides with the fimbriallectin of Escherichia coli Biochem Biophys Res Commun105 1426ndash1432
Firon N Ofek I and Sharon N (1983) Carbohydrate spec-ificity of the surface lectins of Escherichia coli Klebsiellapneumoniae and Salmonella typhimurium Carbohydr Res120 235ndash249
Firon N Ofek I and Sharon N (1984) Carbohydrate-
binding sites of the mannose-specific fimbrial lectins ofEnterobacteria Infect Immun 43 1088ndash1090
Firon N Ashkenazis S Mirelman D Ofek I and SharonN (1987) Aromatic a-glycosides of mannose are powerfulinhibitors of the adherence of type 1 fimbriated Escherichiacoli to yeast and intestinal cells Infect Immun 55 472ndash476
Fukiya S Mizoguchi H Tobe T and Mori H (2004)Extensive genomic diversity in pathogenic Escherichia coliand Shigella strains revealed by comparative genomichybridization microarray J Bacteriol 186 3911ndash3921
Hooton TM and Stamm WE (1997) Diagnosis and treat-ment of uncomplicated urinary tract infection Infect DisClin North Am 11 551ndash581
Horovitz A and Levitzki A (1987) An accurate method fordetermination of receptor ligand and enzyme-inhibitor dis-sociation-constants from displacement curves Proc NatlAcad Sci USA 84 6654ndash6658
Hull RA Gill RE Hsu P Minshaw BH and Falkow S(1981) Construction and expression of recombinant plas-mids encoding type 1 and D-mannose-resistant pili from aurinary tract infection Escherichia coli isolate Infect Immun33 933ndash938
Hultgren SJ Porter TN Schaeffer AJ and Duncan JL(1985) Role of type 1 pili and effects of phase variation onlower urinary tract infections produced by Escherichia coliInfect Immun 50 370ndash377
Hung C-S Bouckaert J Hung DL Pinkner J WinbergC Defusco A et al (2002) Structural basis of tropism ofEscherichia coli to the bladder during urinary tract infec-tion Mol Microbiol 44 903ndash915
Jones TA Zou JY Cowan SW and Kjeldgaard M(1991) Improved methods for building protein models inelectron density maps and the location of errors in thesemodels Acta Crystallogr A 47 110ndash119
Justice SS Hung C-S Theriot JA Fletcher DAAnderson GG Footer MJ and Hulgren SJ (2004)Differentiation and developmental pathways of uropatho-genic Escherichia coli in urinary tract pathogenesis ProcNatl Acad Sci USA 101 1333ndash1338
Kallenius G Mollby R Svenson SB Windberg J Lun-dblud A Svenson S and Cedergen B (1980) The Pk
antigen as receptor for the haemagglutinin of pyelonephri-togenic Escherichia coli FEMS Microbiol Lett 8 297ndash302
Knight SD Berglund J and Choudhury D (2000) Bacte-rial adhesins structural studies reveal chaperone functionand pilus biogenesis Curr Opin Chem Biol 4 653ndash660
Langermann S and Ballou WR (2003) Development of arecombinant FimCH vaccine for urinary tract infectionsAdv Exp Med Biol 539 635ndash653
Langermann S Palaszynski S Barnhart M Auguste GPinkner JS Burlein J et al (1997) Prevention ofmucosal Escherichia coli infection by FimH-adhesin-basedsystemic vaccination Science 276 607ndash611
Langermann S Mollby R Burlein JE Palaszynski SRAuguste CG Defusco A et al (2000) Vaccination withFimH adhesin protects cynomolgus monkeys from coloni-zation and infection by uropathogenic Escherichia coli JInfect Dis 181 774ndash778
Larsson A Ohlsson J Dodson KW Hultgren SJ Nils-son U and Kihlberg J (2003) Quantitative studies of thebinding of the class II PapG adhesin from uropathogenic
454 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
Escherichia coli to oligosaccharides Bioorg Med Chem 112255ndash2261
Leffler H and Svanborg-Eden C (1980) Chemical identifi-cation of a glycosphingolipid receptor for Escherichia coliattaching to human urinary tract epithelial cells and agglu-tinating human erythrocytes FEMS Microbiol Lett 8 127ndash134
Lindhorst TK Kieburg C and Krallmann-Wenzel U(1998) Inhibition of the type 1 fimbriae-mediated adhesionof Escherichia coli to erythrocytes by multiantennary alpha-mannosyl clusters the effect of multivalency Glycoconj J15 605ndash613
Loris R Tielker D Jaeger KE and Wyns L (2003a)Structural basis of carbohydrate recognition by the lectinLecB from Pseudomonas aeruginosa J Mol Biol 331 861ndash870
Loris R Imberty A Beeckmans S Van Driessche ERead JS Bouckaert J et al (2003b) Crystal structureof Pterocarpus angolensis lectin in complex with glucosesucrose and turanose J Biol Chem 278 16297ndash16303
Lund B Lindberg F Marklund BI and Normark S(1987) The PapG protein is the a-D-galactopyranosyl-(1-4)-b-D-galactopyranose-binding adhesin of uropathogenicEscherichia coli Proc Natl Acad Sci USA 84 5898ndash5902
Min G Stolz M Zhou G Liang F Sebbel P StofflerD et al (2002) Localization of uroplakin Ia the urothelialreceptor for bacterial adhesin FimH on the six innerdomains of the 16 nm urothelial plaque particle J Mol Biol317 697ndash706
Miroux B and Walker JE (1996) Over-production of pro-teins in Escherichia coli mutant hosts that allow synthesisof some membrane proteins and globular proteins at highlevels J Mol Biol 260 289ndash298
Morris GM Goodsell DS Halliday RS Huey R HartWE Belew RK and Olson AJ (1998) Automateddocking using a Lamarckian genetic algorithm and anempirical binding free energy function J Comput Chem 191639ndash1662
Mulvey MA (2002) Adhesion and entry of uropathogenicEscherichia coli Cell Microbiol 4 257ndash271
Mulvey MA Lopez-Boado YS Wilson CL Roth RParks WC Heuser J and Hultgren SJ (1998) Induc-tion and evasion of host defenses by type 1-piliated uro-pathogenic Escherichia coli Science 282 1494ndash1497
Murshudov GN Vagin AA and Dodson EJ (1997)Refinement of macromolecular structures by the maxi-mum-likelihood method Acta Crystallogr D Biol Crystallogr53 240ndash255
Nagahori N Lee RT Nishimura S Pageacute D Roy Rand Lee YC (2002) Inhibition of adhesion of type 1 fim-briated Escherichia coli to highly mannosylated ligandsChembiochem 3 836ndash844
Navaza J (2001) Implementation of molecular replacementin AMoRe Acta Crystallogr D Biol Crystallogr 57 1367ndash1372
Neeser JR Koellreutter B and Wuersch P (1986)Oligomannoside-type glycopeptides inhibiting adhesion ofEscherichia coli strains mediated by type 1 pili preparationof potent inhibitors from plant glycoproteins Infect Immun52 428ndash436
Old DC (1972) Inhibition of the interaction between fimbrial
hemagglutinatinins and erythrocytes by D-mannose andother carbohydrates J Gen Microbiol 71 149ndash157
Orndorff PE Devapali A Palestrant S Wyse A EverettML Bollinger RR and Parker W (2004) Immunoglob-ulin-mediated agglutination of and biofilm formation byEscherichia coli K-12 require the type 1 pilus fiber InfectImmun 72 1929ndash1938
Oscarson S and Tedebark U (1995) Syntheses of deoxyanalogues of methyl 36-di-O-a-D-mannopyranosyl-a-D-mannopyranoside for studies of the binding site of con-canavalin A Carbohydr Res 278 271ndash287
Oscarson S and Tiden AK (1993) Syntheses of the octyland tetradecyl glycosides of 36-di-O-alpha-deuterium-mannopyranosyl-alpha-deuterium-mannopyranose and of34-di-O-alpha-deuterium-mannopyranosyl-alpha-deute-rium-mannopyranose ndash a new way for 24-di-O-protectionof mannopyranosides Carbohydr Res 247 323ndash328
Otwinowski Z and Minor W (1997) Processing of X-raydiffraction data collected in oscillation mode MethodsEnzymol 276 307ndash326
Powell HR (1999) The Rossmann Fourier autoindexingalgorithm in MOSFLM Acta Crystallogr D Biol Crystallogr55 1690ndash1695
Pratt LA and Kolter R (1998) Genetic analysis of Escher-ichia coli biofilm formation roles of flagella motility chemo-taxis and type I pili Mol Microbiol 30 285ndash293
Remaut H and Waksman G (2004) Structural biology ofbacterial pathogenesis Curr Opin Struct Biol 14 161ndash170
Ronald AR Nicolle LE Stamm E Krieger J WarrenJ Schaeffer A et al (2001) Urinary tract infection inadults research priorities and strategies Int J AntimicrobAgents 17 343ndash348
Sanner MF Olson AJ and Spehner JC (1996)Reduced surface an efficient way to compute molecularsurfaces Biopolymers 38 305ndash320
Sauer FG Futterer K Pinkner JS Dodson KW Hult-gren SJ and Waksman G (1999) Structural basis ofchaperone function and pilus biogenesis Science 2851058ndash1061
Sauer FG Knight SD Waksman GW and HultgrenSJ (2000) PapD-like chaperones and pilus biogenesisSemin Cell Dev Biol 11 27ndash34
Sauer FG Pinkner JS Waksman G and Hultgren SJ(2002) Chaperone priming of pilus subunits facilitates atopological transition that drives fiber formation Cell 111543ndash551
Schembri MA Hasman H and Klemm P (2000)Expression and purification of the mannose recognitiondomain of the FimH adhesin FEMS Microbiol Lett 188147ndash151
Schembri MA Kjaergaard K and Klemm P (2003) Glo-bal gene expression in Escherichia coli biofilms Mol Micro-biol 48 253ndash267
Schilling JD Mulvey MA Vincent CD Lorenz RGand Hultgren SJ (2001) Bacterial invasion augments epi-thelial cytokine responses to Escherichia coli through alipopolysaccharide-dependent mechanism J Immunol166 1148ndash1155
Silhavy TJ Szmelcman S Boos W and Schwartz M(1975) On the significance of the retention of ligand byprotein Proc Natl Acad Sci USA 72 2120ndash2124
A novel class of FimH high-affinity ligands 455
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
Svanborg C and Godaly G (1997) Bacterial virulence inurinary tract infection Infect Dis Clin North Am 11 513ndash529
Thanassi DG Saulino ET and Hultgren SJ (1998) Thechaperoneusher pathway a major terminal branch of thegeneral secretory pathway Curr Opin Struct Biol 1 223ndash231
Thankavel K Madison B Ikeda T Malaviya R ShahAH Arumugen PM and Abraham SN (1997) Local-ization of a domain in the FimH adhesin of Escherichia colitype 1 fimbriae capable of receptor recognition and use ofa domain-specific antibody to confer protection againstexperimental urinary tract infection J Clin Invest 1001123ndash1126
Vagin A and Teplyakov A (1997) MOLREP an automatedprogram for molecular replacement J Appl Crystallogr 301022ndash1025
Wu XR Sun TT and Medina JJ (1996) In vitro bindingof type 1-fimbriated Escherichia coli to uroplakins 1a and
1b relation to urinary tract infections Proc Natl Acad SciUSA 93 9630ndash9635
Xia Y Gally D Forsman-Semb K and Uhlin BE (2000)Regulatory cross-talk between adhesin operons in Escher-ichia coli inhibition of type 1 fimbriae expression by thePapB protein EMBO J 19 1450ndash1457
Zafriri D Ofek I Adar R Pocino M and Sharon N(1989) Inhibitory activity of cranberry juice on adherenceof type-1 and type-P fimbriated Escherichia coli to eukary-otic cells Antimicrob Agents Chemother 33 92ndash98
Zavialov AV Berglund J Pudney AF Fooks LJ Ibra-him TM MacIntyre S and Knight SD (2003) Structureand biogenesis of the capsular F1 antigen from Yersiniapestis preserved folding energy drives fiber formation Cell113 587ndash596
Zhou G Mo WJ Sebbel P Min G Neubert TA Glock-shuber R et al (2001) Uroplakin Ia is the urothelial recep-tor for uropathogenic Escherichia coli evidence from invitro FimH binding J Cell Sci 114 4095ndash4103

442
J Bouckaert
et al
copy 2004 Blackwell Publishing Ltd
Molecular Microbiology
55
441ndash455
cross-talk between the
fim
and
pap
operons (Xia
et al
2000) As such specific binding is ensured by fimbrialadherence to distinct cell surface receptors Mannose-binding type 1 pili promote cystitis in the lower part of theurinary tract (Langermann
et al
1997 Ronald
et al
2001) by binding to a high-mannose glycoproteinuroplakin Ia (Old 1972 Firon
et al
1982 1984 Wu
et al
1996 Zhou
et al
2001) In ascending urinary tractinfections (UTIs) UPEC use P-pili to bind to galabiose-containing glycolipid receptors in the kidney and initiatepyelonephritis (Kallenius
et al
1980 Leffler andSvanborg-Eden 1980 Lund
et al
1987 Dodson
et al
2001) There has been controversy about the importanceand role of type 1 pili in the gastrointestinal tract (Bloch
et al
1992 Orndorff
et al
2004) and in biofilm formation(Pratt and Kolter 1998 Schembri
et al
2003) and itsreceptors are undefined in these environments
UPEC is the major causative agent of UTIs (Hooton andStamm 1997 Svanborg and Godaly 1997) UTIs are aserious health problem affecting almost one in every twowomen at least once in their lifetime The infections oftenrecur within the months after the primary infection despiteproper use of antibiotics UPEC can invade urothelial cellsin a type 1 pilus-dependent mechanism Once inside thecells they form intracellular bacterial communities suchas biofilms inside the bladder superficial umbrella cellscreating an environment protected from antibiotics and theimmune system of the host (Justice
et al
2004) Bacteriawithin these communities exhibit regional expression oftype 1 pili A subpopulation of UPEC can flux back out ofthe cells and reinvade to ultimately establish a quiescentreservoir that may serve as a seed for recurrent infections(Mulvey
et al
1998)Of the various UPEC adhesins type 1 pili are by far the
most prevalent (Brinton 1959 Buchanan
et al
1985Hultgren
et al
1985 Langermann
et al
1997 Bahrani-Mougeot
et al
2002) being expressed by a wide rangeof both pathogenic commensal and laboratory strains(Fukiya
et al
2004) Type 1 pili consist of a linear stringof repeating non-covalently linked immunoglobulin-like(Ig-like) FimA subunits wound into a rigid right-handedhelical rod followed by a short and stubby tip fibrillumcomposed of the FimF and FimG adaptor subunits andthe FimH adhesin The pilus is assembled by the chaper-oneusher pathway (Thanassi
et al
1998 Knight
et al
2000 Sauer
et al
2000) by complementing the incom-plete Ig fold of each constituent pilus subunit with anamino-terminal extension from a neighbouring subunit ina process termed lsquodonor strand exchangersquo (Choudhury
et al
1999 Sauer
et al
1999 2002 Zavialov
et al
2003) FimH is a two-domain adhesin protein at the endof the tip fibrillum responsible for the mannose-sensitivebacterial adhesion The amino-terminal lectin domain(residues 1ndash158) is joined to the carboxy-terminal pilin
domain (residues 159ndash279) that links the adhesin to therest of the pilus Bacterial two-domain adhesins such asFimH are not soluble unless their pilin domain is comple-mented by a lsquodonor strandrsquo However truncates ofadhesins corresponding to only the lectin domain havepreviously been shown to be properly folded and func-tional (Schembri
et al
2000 Dodson
et al
2001 Buts
et al
2003)The primary physiological receptor for FimH in the uri-
nary tract is the glycoprotein uroplakin Ia (Zhou
et al
2001 Min
et al
2002) that is abundantly present on dif-ferentiated uroepithelial cells However FimH recognizesa wide range of glycoproteins carrying one or more
N
-linked high-mannose structures FimH also binds yeastmannans and mediates agglutination of yeast cells FimH-mediated adhesion can be inhibited by
D
-mannose (Old1972 Hung
et al
2002) and a variety of natural andsynthetic saccharides containing terminal mannose resi-dues (Firon
et al
1982 1983 1984 1987 Neeser
et al
1986 Lindhorst
et al
1998 Nagahori
et al
2002) Block-ing of the FimHndashreceptor interaction has been shown toprevent bacterial adhesion to the bladder uroepitheliumand thereby infection (Langermann
et al
1997 2000Thankavel
et al
1997 Langermann and Ballou 2003)Increasing incidence of bacterial resistance to antibioticshas rejuvenated the interest in understanding the molec-ular basis for FimH-mediated adhesion to provide comple-mentary or alternative treatments of UTIs However forreasons becoming obvious in this article no binding anal-yses of FimHndashcarbohydrate interactions at the molecularlevel have previously been published Also no structuralinformation exists to explain the increased adhesion inhi-bition exhibited by oligomannosides (Firon
et al
19821983 1984 Neeser
et al
1986 Lindhorst
et al
1998) orby some aglycon-substituted mannosides (Firon
et al
1984 1987 Nagahori
et al
2002)In two independently determined crystal structures of
the FimH lectin domain of an
E coli
K-12 strain and auropathogenic strain respectively we made the serendip-itous discovery of a lsquostickyrsquo ligand butyl
a
-
D
-mannosidein the mannose binding site The heterogeneity caused bythis lsquostickyrsquo contaminant rendered several approaches formeasuring binding affinities of the purified FimH lectindomain for its natural carbohydrate receptors useless orat least highly unpredictable The origin of butyl
a
-
D
-man-noside was examined and this knowledge was used toproduce butyl
a
-
D
-mannoside-free FimH We determinedfor the first time FimH binding constants for several hex-oses including mannose two disaccharides two arylmannosides and synthetic alkyl mannosides with differentchain lengths We find that simple alkyl mannosides bindwith nanomolar affinities to FimH Also fructose bindsmore tightly to the FimH fimbrial adhesin than for exam-ple the PapGII adhesin to its natural receptor These
A novel class of FimH high-affinity ligands
443
copy 2004 Blackwell Publishing Ltd
Molecular Microbiology
55
441ndash455
highly specific small-molecule inhibitors targeting withhigh-affinity the FimH mannose binding site may preventUTIs by interfering with the attachment of
E coli
to theuroepithelial cells lining the bladder mucosa
Results
Affinity of FimH for mono- and disaccharides
Two different binding assays were developed one basedon surface plasmon resonance (SPR) and one based onequilibrium binding measurements using radioactivelylabelled mannose (Table 1) Inhibition by mannose of thebinding of FimH to the immobilized Fab fragments of themonoclonal antibody IC10 [k
a
= 147 yen 104 (plusmn417) M-1
s-1 kd = 411 yen 10-3 (plusmn565 yen 10-6)s-1 Kd = 280 yen 10-7
(plusmn88 yen 10-9) M] in SPR measurements allowed us toderive a solution affinity of mannose for FimH(Kd = 23 mM) (Table 1) Using either the Fabs of mono-clonal antibody IA7 or BSA-Man as the immobilized bind-ing partner in the competition experiment gave equivalentdissociation constants for the FimHndashmannose interaction(data not shown) To eliminate the possibility of differentbinding strength for full-length FimH and FimH truncatesthe binding of D-mannose to FimCFimH complex wasalso measured A value of Kd = 53 mM was obtained ingood agreement with the value measured using FimHtruncates
Removal of the 2-OH group of mannose leads to a 100-fold reduction in affinity for FimH (Table 2) Glucose theequatorial O2 epimer of mannose binds a 4000-foldweaker There is virtually no affinity for galactose the O4(axial) epimer and O2 (equatorial) epimer of mannoseFructose binds strongly to FimH (Kd = 31 mM) although a15-fold weaker than mannose The fructose-containingdisaccharides sucrose [Glc(a1ndash2)Frufb] and turanose[Glc(a1ndash3)Frub] have affinities for FimH very similar toglucose
Structure of the FimH lectin domain
The lectin domains FimH truncate 1 (FimHtr1) originatingfrom the UPEC strain J96 and FimH truncate 2 (FimHtr2)from the E coli K-12 laboratory strain MG1655 werecloned expressed and crystallized The structures weresolved by molecular replacement using the lectin domainfrom the FimCFimH complex (Choudhury et al 1999) asa search model and refined to 17 Aring (FimHtr1) and 21 Aring(FimHtr2) resolution respectively (Table S1 in Supplemen-tary material) Both models include residues 1ndash158 cor-responding to the entire lectin domain of FimH Twocopies of the lectin domain are present per asymmetricunit of the FimHtr2 crystals compared with only one unitfor the FimHtr1 crystals The carboxy-terminal 6-His tag ofFimHtr2 is not visible in the electron density maps and isnot included in the model
The structures of the FimH lectin domains are verysimilar to the structure for the same domain in theFimCFimH complex with a root mean square deviation(rmsd) over all 158 Ca positions of 036 Aring for FimHtr1056 Aring for FimHtr2 subunit A and 054 Aring for FimHtr2 subunitB The secondary structure elements arranged in an Igfold (Buts et al 2003) (Fig 1A) have almost interchange-able Ca traces in all of the FimH lectin domain structuresOnly Gly73 (in strand D1) and Gly79 (in loop connectingstrands D1 and Dcent) have slightly variable main chain con-formations while Val30 in the b3ndashb4 loop at the base ofthe domain clearly undergoes a peptide flip in all threesubunits of the truncate compared with the FimCFimHcomplex In the FimHtr1 structure the conformation of thecarboxy-terminal residue of the truncate Thr158 differsas it is packed against and hydrogen bonded to a glycerolmolecule via its carboxylate group In the FimHtr2 crystalstructure the main difference is in the b9ndashb10 loop withan rmsd of 29 Aring over Ca atoms 113ndash116 This loopadopts a different conformation because of crystal pack-ing contacts that are not present in the FimCFimH orFimHtr1 structures
Mannose binding pocket
Although no sugar was included in the crystallization set-ups strong positive electron density was present in the
Table 1 Kd and calculated DG0 for a series of O1 alkyl and arylmannosides
LigandKd SPR(nM)
DGinfin SPR (kcal mol-1)
Kd displace(nM)
DGinfin displace(kcal mol-1)
Mannose 23 yen 103 -76 41 yen 103 -73Methylaman 22 yen 103 -77 24 yen 103 -76Ethylaman 12 yen 103 -81 730 -83Propylaman 300 -89 400 -87Butylaman 151 -93 150 -93Pentylaman 25 -104 200 -91Hexylaman 10 -109 100 -95Heptylaman 5 -113 32 -102Octylaman 22 -104 28 -103PNPaMan 44 -100 26 -103MeUmbaMan 20 -105 12 -108
The SPR and displacement binding experiments define heptyl a-D-mannopyranoside as the best binder
Table 2 Kd and calculated DG0 for a series of mono- and disaccha-rides and a deoxymannose
Ligand Kd
DGinfin(kcal mol-1)
Methyl 2-deoxy-a-D-mannopyranoside 300 mM -48Glucose 924 mM -28Galactose 100 mM -14Fructose 31 mM -61Sucrose 128 mM -26Turanose 76 mM -29
444 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
initial FondashFc and 2FondashFc Fourier maps inside the mannose-binding pocket of both FimHtr1 and FimHtr2 This densityprecisely defined a-D-mannose with an additional elon-gated density shaped like a creased tail extending out-wards from the mannose O1 and towards tyrosines 48and 137 which make up part of the hydrophobic rimaround the mannose-binding pocket (Fig 1B) (Hunget al 2002) Further refinement strongly suggested thata butyl a-D-mannoside was bound in the mannose-bindingsite (Fig 1B and C) For FimHtr1 also a change in theconformation of the Tyr48 side-chain compared with theone from the model FimH used for molecular replacementwas immediately apparent As described previously (Hunget al 2002) the mannose ring makes 10 direct hydrogenbonds to the side-chains of residues Asp54 Gln133Asn135 and Asp140 and to the main chain of Phe1 andAsp47 and indirect water-mediated hydrogen bonds viaO2 to the side-chain of Glu133 and to the main chainoxygen of Phe1 and Gly14 The alpha-anomeric hydroxylgroup O1 of mannose is involved in a hydrogen-bondingwater network with the Asn138 and Asp140 side-chainsthrough a water molecule (wat 2 Fig 1B) that is con-
served between the two truncate structures The positionsof the other waters participating in this network dependon the crystal packing of FimHtr1 and FimHtr2
Mass spectrometric analysis of the bound ligand
To corroborate the nature of the ligand identified in thecrystal structures the complex of FimHtr1 and its pre-sumed butyl a-D-mannoside was analysed using massspectrometry The FimHtr1 protein sample used for crys-tallization was subjected to peracetylation and phase sep-aration The peracetylated ligand retrieved in the organicphase was analysed by nanoLC-online-ESI-MS The per-acetylated ligand eluted as a single peak and was regis-tered in its sodiated form at mz 427 corresponding tobutyl hexose (Fig 2A) No signals corresponding to otherpotential alkyl mannose species like ethyl hexose (mz399) propyl hexose (mz 413) or pentyl hexose (mz 441)could be observed indicating a homogeneous ligand witha four-carbon atoms alkyl chain Besides the sodiumadduct of the ligand a major signal was observed arisingfrom the loss of butanol (mz 331 Fig 2B) Cleavage at
Fig 1 Stereo views of FimHA Three-dimensional structure of the FimH lectin domain is an elongated 11-stranded b-barrel with an immunoglobulin fold Numbering is according to conventions established for antibody domains (Bork et al 1994) and for PapD (Sauer et al 1999) and loop identifications according to previous descriptions (reference strand numbers A1 = 1 Acent = 2 A2 = 3 B1 = 4 B2 = 4cent C = 5 D1 = 6 Dcent = 7 D2 = 8 E = 9 F = 10 G = 11) (Choudhury et al 1999 Hung et al 2002) The disulphide bridge between Cys3 and Cys44 is coloured cyanB Focus on the mannose-binding pocket in the same orientation as (A) showing the hydrophobic contacts (within 46 Aring orange) with butyl a-D-mannoside and the water network (salmon pink) present at O1C Surface presentation (wheat yellow) of the different alkyl conformations of the butyl a-D-mannoside in the crystal structures of FimH truncates 1 and 2 FimHtr1 butyl group (cyan) FimHtr1 Tyr48 (marine blue) FimHtr2 butyl group and Tyr48 (salmon pink) Tyr137 (violet) Ile52 (pink)
A B
C
A novel class of FimH high-affinity ligands 445
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
the anomeric carbon atom was often combined with elim-ination of one or more acetic acid molecules (massdecrease of 60 Da) or acetyl groups (mass decrease of42 Da) These eliminations gave rise to the major ions atmz 127 and mz 169 with a proposed aromatic ring struc-ture as well as to the ion at mz 109 with a proposedquinone-like ring structure Thus the mass spectrometricdata support the butyl hexose nature of the FimH ligandFimH was further characterized by carbohydrate constit-uent analysis (data not shown) which revealed only man-nose Mannose could only be detected after hydrolysisindicating that free mannose is not associated with theFimH truncate
Origin of butyl a-D-mannoside
As no sugar had been advertently added to the FimHtr
protein samples we hypothesized that the butyl manno-side ligand had been picked up from the LuriandashBertani(LB) medium used to grow bacteria during expression ofthe protein To test this FimHtr2 was expressed in bacteriagrown in minimal M9 medium Electro-spray ionizationmass spectrometry (ESI-MS) was performed on purifiedFimHtr2 samples from both types of media A clear peakat 259 Da could be detected (Fig 3A) with proteinexpressed in bacteria grown in LB medium correspondingto the molecular weight of butyl mannoside (236 Da) plus
a sodium ion (23 Da) As a control butyl mannoside wassynthesized and analysed in the same manner Again apeak at 259 Da was observed (Fig 3A inset) When ESI-MS was run on protein expressed in bacteria grown inminimal medium no such peak could be detected(Fig 3B) indicating that the ligand was not present andthat it indeed originated from the LB medium
Interactions of the butyl moiety with FimH
The butyl moiety of butyl a-D-mannoside extends out ofthe mannose-binding pocket towards Tyr48 and Tyr137making van der Waals contacts to both tyrosine rings andIle52 (Fig 1B and C) These residues are part of two loops(b4ndashb5 and b10ndashb11) that form the higher rear end of ahydrophobic collar around the binding site (Fig 1A andB) In both subunits of the FimHtr2 structure the Tyr48 andTyr137 side-chains are in the same almost parallel ori-entation as in the FimCFimH structure forming a gate-like structure at the back of the binding site (Fig 1C) Theclosest hydrophobic interactions (31ndash36 Aring) with the alkylchain are made between Tyr48 and the two non-terminalatoms of the butyl In the FimHtr1 structure a crystal con-tact to Val155 of a neighbouring molecule prevents theparallel orientation of the Tyr48 ring which instead ispacked edge-to-face with Tyr137 (Fig 1B) The reposition-ing of Tyr48 is accompanied by a change in the confor-
A
B
Fig 2 Mass spectrophotometric characteriza-tion of the butyl mannosideA Base peak chromatogram (BPC mass range mz 400ndash550) and extracted ion chromato-grams (EIC) of mz 427 and 331 obtained by nano-LC-ESI-MS of the derivatized FimH-ligand purified by peracetylation and organic-solvent extractionB ESI-IT mass spectrometric characterization of the peak species marked by a horizontal bar in (A) Ions are marked with mz-values and proposed schematic structures which is often only one out of several isobaric ion species Fragmentations already occurred in the source Ac acetyl
446 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
mation of the butyl chain of the ligand with the butyl chainin a more extended conformation than in the FimHtr2 struc-ture and with the four carbon atoms nearly in one plane(Fig 1B and C) The new conformations of Tyr48 and thebutyl moiety result in an almost parallel stacking of thearomatic ring of Tyr48 onto the butyl plane with the closesthydrophobic interactions (37 Aring) involving the two terminalcarbon atoms of the butyl (Fig 1B)
Affinity measurements for alkyl mannosides
The butyl mannoside ligand remained bound to FimHtr
despite extensive dialysis From a study of complex for-mation of monoclonal antibodies IC10 and IA7 withFimHtr1 it could be estimated that about 30 of the FimHtruncate retained its ligand after dialysis for 12 daysagainst 50 mM MES buffer at pH 65 100 mM NaCldespite daily refreshment of buffer (data not shown)Therefore to enable measurements of dissociation con-stants of FimH for mannosides FimH was produced inminimal medium Butyl a-D-mannoside binding is 15 timesimproved (Kd = 151 nM Fig 4A) compared with mannosebinding The equilibrium binding technique using [3H]-mannose led to a similar value (Kd = 41 mM Fig 4B) for
mannose and an almost identical value for butyl a-D-mannoside (Kd = 150 nM Fig 4C) To investigate theeffect of sequential addition of methyl groups to the O1oxygen of D-mannose a series of alkyl mannosides weresynthesized and the dissociation constants determinedusing the [3H]-mannose displacement assay as well as theSPR competition assay (Table 1) There is a linear corre-lation between the binding free energy as calculated fromthe measured dissociation constants and the number ofmethyl groups (between 1 and 7) in the alkyl mannosidewith each additional methyl group contributing on average-06 kcal mol-1 of binding energy (Fig 5)
Affinity measurements and modelling for aryl mannosides
Aromatically substituted mannosides have been reportedto be particularly potent inhibitors of FimH-mediated bac-terial adhesion (Firon et al 1987 Nagahori et al 2002)In accordance with the earlier results both pNPaMan andMeUmbaMan bind very tightly to FimH (Table 1) Despiteextensive trials no X-ray structures were obtained forcomplexes with aromatically substituted mannosides Inorder to model binding docking to both of the truncatestructures was carried out In all cases the mannose ring
Fig 3 Mass spectrometric analysis of the origin of the butyl mannosideA LB medium-produced FimH truncate The inset shows the mass spectrum of the synthesized butyl a-D-mannosideB Minimal media-produced FimH truncate
A novel class of FimH high-affinity ligands 447
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
docks in the binding pocket in a conformation very closeto the crystallographically observed mannose conforma-tion (Fig 6) while the aromatic substituent interacts withthe tyrosine gate The different conformation of Tyr48 in
our two FimH truncate structures leads to different pre-dicted interactions between the aromatic aglycon andTyr48 and Tyr137 Docking to the FimHtr2 structureresulted in somewhat tighter interactions between thetyrosine gate and the aglycon and hence in somewhatlower docking energies Calculated FimHtr2 docking ener-gies of Edoc = -104 kcal mol-1 for pNPaMan (Kd = 46 nM)and Edoc = -109 kcal mol-1 for MeUmbaMan (Kd = 20 nM)are in good agreement with the experimental binding data(Table 1) The docked umbelliferyl ring system is inserteddeep into the tyrosine gate in the FimHtr2 conformationwith the carbonyl group within hydrogen bonding distanceto Thr51 at the back of the gate (Fig 6A) Docking to theFimHtr1 structure instead stacks the umbelliferyl ring face-to-face with the Tyr48 ring (not shown) The phenyl ring ofpNPaMan docks similarly to both our structures It doesnot quite reach in between the tyrosine side-chains butinstead hydrogen bonds with its nitro group to the hydroxyloxygen of Tyr137 (Fig 6B)
Discussion
Most lectins bind naturally occurring carbohydrates withlow affinities often in the millimolar range for monosac-charides and micromolar range for oligosaccharides Thiswas recently exemplified to also hold true for theadhesins of F17 fimbriae (Kd = 12 mM for the F17a-G-N-acetylglucosamine interaction Buts et al 2003) and Ppili (Kd = 84 mM for the PapGII-globotetraoside interactionLarsson et al 2003) The recent crystal structure of FimHin complex with mannose illustrated that FimH almostcompletely envelops the monosaccharide in a deeppocket and involves it in an intense hydrogen bondingnetwork suggestive of an unusual lectin exerting highaffinities (Hung et al 2002)
Fig 4 FimH binding experimentsA Surface plasmon resonance competition experiment of FimHtr1 binding to monoclonal antibody IC10 Fab fragments with butyl a-D-mannosideB Equilibrium binding of [3H]-mannose to FimHtr2C Equilibrium displacement measurements of butyl a-D-mannoside to the FimHtr2 [3H]-mannose complex
log[Butyl a-D-mannoside] (M)
cpm
cpm
Butyl a-D-mannoside (mM)
Mannose (mM)
A
B
C
ndash64
ndash66
ndash68
ndash70
ndash72
ndash74
ndash76
ndash78
ndash80
ndash82
ndash8 ndash7 ndash6 ndash5 ndash4 ndash3
log
[Fim
H]
M
Fig 5 Linear free energy relationship for the alkyl chain on mannose A linear fit was made using 1 up to 7 methyl groups with the program MICROCAL ORIGIN 50
Number of carbon atoms0 2 4 6 8
ndash75
ndash80
ndash90
ndash95
ndash100
ndash105
ndash110
ndash115
ndash85
D G
(kc
al m
olndash1
)
slopecorrelation
ndash064ndash099
+ndash 003+ndash 018
448 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
The measured affinity confirm the predictions that FimHbinds the monosacharide mannose exceptionally well(Kd = 23ndash41 mM) To investigate the contribution of thedifferent hydroxyl groups of mannose in the interactionwith FimH the binding of a series of hexoses to butylmannoside-free FimH was examined using SPR (Table 2)The effect of deoxygenation of the O2 hydroxyl group(methyl 2-deoxy-a-D-mannopyranoside) is clearly lessdestructive (100-fold reduction) than its equatorial positionas in glucose O2 is the only hydroxyl group of mannosethat interacts indirectly with the FimH pocket via a con-served water molecule (Hung et al 2002) Glucose onlydisplays a weak affinity for FimH about 4000 times lowerthan methyl a-D-mannoside Repositioning of both the O2and O4 hydroxyl groups as in galactose abolishes sugarbinding by FimH Similar to these results the inhibitoryconcentrations for bacterial adherence of methyl a-D-glucoside and galactose derivatives have been estimatedto be respectively 104-fold and 105-fold weaker than formethyl a-D-mannoside (Nagahori et al 2002)
In stark contrast with the other monosaccharides fruc-tose has a binding affinity only about 15-fold less thanmannose for FimH (Table 2) The decrease in affinity for
fructose relative to mannose agrees with studies of inhi-bition of adherence of type 1 piliated E coli by fructosein which fructose was found to be the active compound infruit juices (present at about 5) (Zafriri et al 1989)Fructose binding to FimH is stronger than the binding ofthe physiological globotetraoside receptor to PapGII theother well-characterized adhesin of UPEC strains (Lars-son et al 2003) The pyranose form of fructose is verywell superimposable onto mannose in the FimH bindingsite where similar interactions are established with FimHas for mannose except for the loss of the anomeric O1and an extra hydroxyl group (O2) on the C5 position ofmannose Probably however both changes of fructopyr-anoside compared with mannopyranoside could be easilyaccommodated by FimH The mannose O1 does notmake direct interactions within 45 Aring of the FimH bindingpocket (Fig 1B) The extra O2 group of fructose would notclash or interfere significantly with the binding pocket butit would make a 27 Aring van der Waals contact with the Ile52side-chain under the condition that the Ile52 side-chaindoes not move The crystal structure of the Pseudomonasaeruginosa lectin LecB also showed that fructopyranosebut not fructofuranose can substitute for mannose (Loris
Fig 6 Stereo diagrams of docked MeUm-baMan (A) and pNPaMan (B) in the FimH bind-ing site The crystallographically determined position of a-D-mannose is shown in orange for comparison
A novel class of FimH high-affinity ligands 449
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
et al 2003a) Hydrogen bonds of the more specificfucose complex are replaced by van der Waals interac-tions involving a hydroxyl group in the complexes with thelower-affinity binders mannose and fructopyranose as forthe proposed fructopyranoside binding to FimH
The fructose-containing disaccharides sucrose andturanose do not have an inhibitory effect and bind withaffinities similar to glucose Sucrose is a non-reducingsugar that has fructose locked in its furanose form Tura-nose on the other hand allows the interconversion of b-D-fructofuranose and b-D-fructopyranose eg both formsoccur in complex with a plant lectin from Pterocarpusangolensis (Loris et al 2003b) It is predictable howeverfrom the crystal structures that the glycosidic linkage ofglucose with C3 of fructose equivalent in position to C4of mannose would be disallowed in the tight FimH pocketSucrose and turanose thus most probably interact with theFimH mannose binding pocket via their glucose moietyresulting in similar low affinities
Our mass spectrometry and chemical analyses con-firms the crystallographic identification of the ligand boundto the FimH lectin domains as butyl a-D-mannoside(Figs 1B and 2) The mannose ring of the butyl a-D-mannoside coincides with the mannose bound to theFimCFimH complex (Hung et al 2002) and makes thesame interactions The four-carbon alkyl tail exits towardsthe back wall (loop b4ndashb5 and b10ndashb11) of the pocket(Fig 1A and B) making hydrophobic contacts to Tyr48Tyr137 and Ile52 The mass spectrometry data also showthat the bound butyl a-D-mannoside is present in LBmedium but absent from minimal medium Hence thesource of the alkyl mannoside is possibly the yeast extractused in preparation of LB medium Although we do notwant to speculate about the significance of this findingalkyl mannoside production by yeast has not beenreported and might warrant investigation
The butyl mannoside was retained by FimH despiteextensive dialysis The phenomenon of ligand retention byprotein was described as early as 1975 for periplasmicbinding proteins (Silhavy et al 1975) Applied to dialysisof butyl mannoside away from FimH grown in richmedium half-lives of up to 30 h can be estimated whichis far beyond the duration of a normal buffer exchangedialysis experiment In the present case the retainedligand does not appear to seriously affect [3H]-mannosedisplacement equilibrium binding experiments as wecould detect no significant differences in dissociation con-stants obtained using FimHtr2 produced in LB or minimalmedia (data not shown) In contrast proper SPR bindingcurves could not be obtained when butyl mannoside occu-pied the binding site Hence the displacement techniquemay be of more general use to obtain binding constantsfor proteins that tend to inadvertently retain ligands whichis a common trait of lectins
Surface plasmon resonance as well as equilibrium bind-ing and displacement measurements using [3H]-mannosewere carried out to obtain dissociation constants for a-D-mannose and a linear series of alkyl-substituted manno-sides to FimH (Table 1) Both equilibrium binding experi-ments show a very good agreement in their measurementof the carbohydrate affinity of the adhesin FimH bindsbutyl a-D-mannoside 15ndash30 times stronger than a-D-mannose The importance of the added hydrophobiccontacts is clearly demonstrated by the binding of alkylmannosides (Table 1) The best binding alkyl mannosidesbind a few hundred times stronger than mannose equiv-alent in affinity to the most tightly binding aromaticallysubstituted mannosides for FimH (Firon et al 1987) andof mannose dendrimers for type 1 piliated E coli (Fironet al 1984 1987 Nagahori et al 2002)
Interestingly the butyl moiety takes on significantlydifferent conformations in FimHtr1 and FimHtr2 (Fig 1C)indicating that intrinsic conformational flexibility in theaglycon is allowed Indeed in spite of the larger flexibilityexpected for longer alkyl mannosides the binding energydecreases linearly with increasing length of the alkyl chainbetween one and seven carbon atoms (Fig 5) The affinityreaches its maximum for mannose with a seven-carbonalkyl and SPR measurements suggest that the affinitydecreases from eight carbon atoms on Evidently theeighth carbon atom goes beyond the hydrophobic interac-tion surface and protein edge which is penalized with adecrease in affinity We propose that the affinity increaseresults from an increased interaction surface with thehydrophobic gateway Alkyl aglycons fit well in the tyrosinegate and as observed in the two crystal structures in atleast two different conformations that depend on theconformation of Tyr48 This flexibility may indicate thataglycons can interact rather non-specifically with thehydrophobic gateway nevertheless resulting in a 100- to1000-fold increase in affinity
FimH binding to ligands other than just mannose ispossibly of importance in terms of the different environ-ments that E coli will encounter ndash the urinary tract gas-trointestinal tract and in biofilms Alkyl mannosides havenot been previously recognized as strong binders to FimHAs they are easily synthesized and highly soluble in waterthey may be interesting as potential blocking agents forFimH-mediated adhesion Moreover they have beenshown to be good drug vehicles themselves acting aspermeability enhancers on tight junctions (Eley and Triu-malashetty 2001) As such they might be able to pene-trate into deeper cell layers of the epithelium to alsoprevent those bacteria that form a quiescent reservoirfrom causing recurrent infections (Mulvey et al 1998Justice et al 2004)
On the other hand the two aromatically substitutedmannosides pNPaMan and MeUmbaMan are known to
450 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
be particularly potent inhibitors of FimH-mediated bacte-rial adhesion (Firon et al 1987 Nagahori et al 2002)Based on adhesion inhibition studies using type 1 piliatedE coli and guinea pig ileal epithelial cells the relativeinhibitory potential of pNPaMan and MeUmbaMan havebeen estimated to be about 70 and 600 times strongerrespectively than that of methyl a-D-mannose (Fironet al 1987) Our measurements indicate an affinity ofFimH for pNPaMan and MeUmbaMan between 50 and90 times and between 110 and 200 times higher respec-tively than for methyl a-D-mannose in good agreementwith the adhesion inhibition results (Firon et al 1987)Dissociation constants predicted using AutoDock3 (Morriset al 1998) are similar to experimental values Thecomputational docking predicts that tight binding of thearomatically substituted mannosides pNPaMan andMeUmbaMan is accomplished similarly as for butyl a-D-mannoside with the aglycon interacting with the tyrosinegate
This is the first systematic study providing molecularbinding data for the FimH adhesin the prerequisite forcolonization by UPEC (Langermann and Ballou 2003)We also show crystallographic evidence that the lectinor receptor binding domain of FimH is a stable andfunctional entity Its structure is virtually identical to thesame domain in the FimCFimH complex indicating thata truncate corresponding to the lectin domain could pro-vide a good alternative for the development of a UTIvaccine (Langermann et al 1997 Langermann and Bal-lou 2003) However progress of this interesting UTI vac-cine candidate into clinical trials has not been reportedThe binding affinities of FimH at the molecular levelrelate convincingly well with the inhibitory concentrationsused in previous bacterial adhesion assays thus endors-ing the direct and conclusive correlation of FimH withbacterial adherence Alkyl mannosides have nanomolaraffinities for FimH which may overcome the significantobstacle of low affinity generally encountered in attemptsto develop carbohydrate-based drugs which interferewith microbial attachment to host cell surface glycocon-jugates New anti-adhesive drugs could be based onthese leads to allow specific interaction and targeting tothe urinary tract in order to prevent anticipated andrecurrent infections
Experimental procedures
Strains and constructs
FimH proteins from two different E coli strains the UPECstrain J96 (Hull et al 1981) and the E coli K-12 strainMG1655 (Blattner et al 1997) were used in this study TheFimH adhesins encoded by these two E coli strains areidentical FimHtr1 (residues 1ndash158 of UPEC J96 FimH) wasexpressed from plasmid pMMB91 transformed into E coli
C600 cells FimHtr2 (residues 1ndash158 of E coli MG1655FimH + carboxy-terminal 6-His tag) was expressed fromplasmid pPKL241 (Schembri et al 2000) transformed into Ecoli host strain HB101
Purification and crystallization
FimHtr1 Bacteria were grown in LB medium at 37infinC inducedat OD600 = 08 with 1 mM IPTG and harvested 2ndash3 h laterThe cells were resuspended in 20 mM Tris-HCl pH 80 5 mMEDTA and 30 sucrose and the periplasmic content wasextracted by osmotic shock upon 10-fold dilution with 20 mMTris-HCl pH 80 Purification of FimHtr1 was performed by ionexchange chromatography with MonoS at pH 45 followed byMonoQ (Amersham Biosciences) at pH 8 FimHtr1 was crys-tallized using vapour diffusion in hanging drops composed of3 ml of the precipitant solution (01 M Hepes pH 75 12PEG 6000 and 5 vv methyl-24-pentanediol) and 3 ml of12 mg ml-1 protein solution in 20 mM Tris-HCl pH 80 with150 mM NaCl
FimHtr2 Cells were grown in LB medium containing50 mg ml-1 ampicillin at 37infinC At OD600 = 06 the cells wereinduced with 5 mM IPTG Cells were harvested 2 h afterinduction and the periplasmic content was extracted Theprotein was purified on a Pharmacia HiTrap Chelating HP5 ml column (Pharmacia) loaded with Ni-chloride and elutedwith a sharp 0ndash500 mM imidazole gradient Fractions con-taining FimHtr2 were pooled dialysed against 50 mM sodiumacetate pH 525 and loaded onto a Mono S HR 8 ml columnThe protein was eluted with a 0ndash500 mM NaCl gradient anddialysed overnight against 20 mM MES pH 65 before crys-tallization Crystallization was performed at room tempera-ture by the vapour diffusion method with hanging dropsmade by mixing of 2 ml of 15 mg ml-1 protein solution with 2 mlof the precipitant solution (65 methyl-24-pentanediol100 mM cacodylate pH 65) Microseeding was employed tooptimize crystal quality and reproducibility Crystals suitablefor X-ray data collection generally appeared within 48 h aftermicroseeding
Structure determination
X-ray data for FimHtr1 were collected from a single crystalto 17 Aring resolution at the Advanced Photon SourceArgonne National Laboratory USA The crystal was flash-cooled to 100 K in the precipitant solution complementedwith 25 glycerol The data were processed and scaledusing the HKL2000 package (Otwinowski and Minor 1997)(Table S1 in Supplementary material) Data for FimHtr2
were collected from a single crystal under liquid-nitrogencryo conditions at 100 K at beamline ID29 ESRF Greno-ble France to 21 Aring Bragg spacing FimHtr2 data were pro-cessed and reduced using MOSFLM (Powell 1999) andscaled using SCALA (Collaborative computational project1994) (Table S1 in Supplementary material) The structuresof FimHtr1 and FimHtr2 were solved by molecular replace-ment using Amore (Navaza 2001) and MOLREP (Vaginand Teplyakov 1997) respectively with the FimH lectindomain from the FimCFimH complex (Choudhury et al
A novel class of FimH high-affinity ligands 451
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
1999) (PDB id 1QUN) as the search model Positional bulksolvent and isotropic temperature factor refinement wasperformed using CNS (Bruumlnger et al 1998) and REFMAC5(Murshudov et al 1997) respectively Model adjustmentand inspection were performed using Turbo-Frodo and O(Jones et al 1991) graphics programs respectively Refine-ment statistics are given in Table S1 Molecular graphicsillustrations were generated with Bobscript (Esnouf 1997)and Pymol 090 (DeLano Scientific LLC 1998ndash2003 httppymolsourceforgenet)
Mass spectrometry
The purified FimHtr1 protein (300 mg) that had been used forcrystallization was freeze-dried taken up in 100 ml of pyridineand 100 ml of acetic anhydride and incubated overnight atroom temperature in the dark for peracetylation The samplewas dried under nitrogen and taken up in chloroform (400 ml)The chloroform phase was extracted six times with waterdried and taken up in 200 ml of 50 aqueous methanol A1 ml aliquot was applied to a nano-scale RP column(75 mm yen 100 mm PepMap DionexLC Packings) using anUltimate nano-LC system (LC Packings) which was equili-brated with eluent A (H2Oacetonitrile 955 vv containing01 formic acid) at a flow rate of 100 nl min-1 After injectingthe sample the column was run isocratically for 5 min fol-lowed by a linear gradient to 100 eluent B (H2Oacetonitrile2080 vv containing 01 formic acid) in 20 min and a finalwash with 100 B for 5 min The system was directly coupledto an Esquire 3000 ESI-IT-MS (Bruker Daltonik) equippedwith an online nanospray source operating in the positive-ionmode For electrospray (900ndash1200 V) capillaries (360 mmOD 20 mm ID with 10 mm opening) from New Objective wereused The solvent was evaporated at 150infinC with a nitrogenstream of 6 l min-1
To verify the suspected source of the bound butyl manno-side mass spectrometry was performed on samples ofFimHtr2 grown in either LB or minimal M9 medium Proteinsamples were dialysed against water overnight before massspectrometry in order to reduce background from the Trisbuffer Samples were diluted to a concentration in the submi-cromolar range Electrospray mass spectra were recorded indirect inlet mode (50 aqueous methanol solutions) on aBruker Daltonics Esquire-LC (Billerica)
Carbohydrate constituent analysis
Samples were hydrolysed in 100 ml of 4 M aqueous trifluoro-acetic acid (Sigma) at 100infinC for 4 h and dried under astream of nitrogen Monosaccharides were converted intotheir anthranilic acid derivatives by reductive aminationresolved by RP-HPLC and detected by fluorescence (Anu-mula 1994)
Carbohydrates for binding studies
[3H]-mannose was obtained from Amersham Methyl a-D-mannoside glucose galactose fructose sucrose turanosep-nitrophenyl a-D-mannopyranoside (pNPaMan) and 4-methylumbelliferyl a-D-mannopyranoside (MeUmbaMan)
were obtained from Sigma Monomannosylated bovineserum albumin (BSA-Man) was kindly provided by MedIm-mune We synthesized methyl 2-deoxy-a-D-mannopyrano-side (Oscarson and Tedebark 1995) Alkyl mannosides weresynthesized through silver triflate-promoted couplings of thecorresponding alcohol with 2346-tetra-O-benzoyl-a-D-mannopyranosyl bromide followed by Zempleacuten deacylationof the obtained protected alkyl mannosides according to theprocedure reported for the octyl and tetradecyl mannosides(Oscarson and Tiden 1993)
Binding studies
Competition experiments using SPR To obtain butyl man-noside-free FimHtr1 the FimH truncate was overproducedwith a carboxy-terminal 6-His tag after transformation into Ecoli C43 (DE3) (Miroux and Walker 1996) and the bacteriagrown in minA minimal medium supplemented with the 20amino acids (40 mg l-1) thiamin (2 mg l-1) and biotin (2 mgl-1) Purification of the His-tagged protein was performedusing Ni-NTA affinity chromatography (Qiagen) in 50 mM Tris-HCl pH 8 and 300 mM NaCl Elution was in the same solutioncomplemented with 500 mM imidazole Subsequently pooledand concentrated FimH fractions were gel filtrated over aSuperdex G75 column (Pharmacia) in 20 mM Tris-HCl pH 8
We used Fab fragments of a monoclonal antibody againstthe mannose-binding site of FimH to determine the solutionaffinity of FimHndashcarbohydrate interactions The monoclonalantibody IC10 was produced by a mouse hybridoma cell lineat Medimmune and its Fab fragments were prepared andpurified The Fabs were covalently immobilized via aminecoupling at 1000 Resonance Units (1000 pg ligand mm-2) inflowcell Fc2 on a CM5 biasensor chip (BIAapplications Hand-book Biacore AB Uppsala Sweden) Flowcell Fc1 was usedas the reference surface and was treated in the same wayas Fc2 except that it was blocked immediately after activationThe immobilization buffer was 100 mM sodium acetatepH 50 with 100 mM NaCl All SPR measurements were per-formed on a Biacore3000trade
The kinetic constants of binding of FimH to the immobilizedantibody were measured with different concentrations ofFimH (2000 nM 1000 nM 500 nM 250 nM 125 nM625 nM 312 nM 156 nM 78 nM) in running buffer [phos-phate-buffered saline (PBS) with 0005 surfactant P20 and3 mM EDTA] on both flowcells Fc1 and Fc2 simultaneouslyat a flow rate of 30 ml min-1 and at 298 K Complete dissoci-ation of FimH was performed with running buffer beforestarting a new binding cycle For all measurements the asso-ciation time was 3 min and the dissociation time was 30 minAll binding cycles were performed in duplicate including azero concentration cycle of FimH (injection of running buffer)All analyses were performed with the BIAeval software ALangmuir binding isotherm with a 11 stoichiometry was fittedto the data from both the association phase and the dissoci-ation phase after subtraction of the reference and zero con-centration data The kinetic constants ka and kd and themaximal binding Rmax were obtained and the dissociationconstant Kd was calculated
The equilibrium dissociation constants Kd of FimHndashsaccha-ride interactions were determined in a competition experi-ment using the same chip as for the previous experiment
452 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
As a control the Fabs of monoclonal antibody IA7 as wellas BSA-Man were also immobilized Samples containing afixed concentration of FimH were used (close to the Kd of theFimHndashantibody interaction) in combination with varying con-centrations of saccharide First 10-fold dilutions of the sac-charide solution were used to determine the concentrationrange for binding of the saccharide to FimH A Langmuirbinding isotherm with a 11 stoichiometry was fitted to thedata using the kinetic constants and Rmax from the foregoingexperiment to obtain the concentrations of FimH that werefree ([FimH]free) to bind the antibody immobilized on the chipSecond the concentration range of the saccharide wasextended and adapted to ensure accurate fitting and theequilibrium binding constant of the FimHndashsaccharide interac-tion was derived from the curve of [FimH]free against concen-tration of saccharide Every measurement was repeated atleast twice including testing different protein batches andwhenever possible also different saccharide stock solutions(typically 200 mM)
Equilibrium binding studies using radioactively labelledmannose Mannose-free FimHtr2 was produced inM9 minimal medium and purified as described above forFimHtr2 Alkyl mannosides were dissolved in double distilledwater pNPaMan and MeUmbaMan (6 mg each) were dis-solved in 20 ml of DMSO and diluted to 20 mM using doubledistilled water Binding experiments were performed at 310 Kusing six different concentrations of [3H]-a-D-mannose (finalconcentrations 435 mM 290 mM 193 mM 129 mM 86 mMand 57 mM) Protein (180 ml) at a concentration of about500 nM was mixed with 20 ml of the radioactive ligand [3H]-a-D-mannose and incubated at 37degC for 20 min To separatefree ligand from bound the mixture was rapidly filtratedthrough a Protran BA 85 cellulose nitrate filter (Schleicherand Schuell) and washed once with 1 ml of ice-cold 1yen PBSFilter-bound radioactivity was measured by scintillation spec-trometry within 24 h Displacement experiments were per-formed using six different concentrations of inhibitor (finalconcentrations in the range 00ndash435 mM) in the presence of435 mM [3H]-a-D-mannose Twenty microlitres of radioactiveligand 20 ml of inhibitor and 160 ml of protein (500 nM) weremixed and the experiments were performed as describedabove All experiments were performed in duplicates and thedata presented are mean values of at least two independentmeasurements For the determination of Kd for a-D-mannosean hyperbolic curve (y = P1x(P2 + x) where P1 is an arbitraryscale factor and P2 = Kd) was fitted to the data For the dis-placement experiments the curve y = P1(P2 + x) where P1
is an arbitrary scale factor and P2 is the concentration of theinhibitor displacing 50 of the labelled ligand [I]05 was usedinstead To calculate the inhibitor dissociation constant (KI)the Cheng and Prusoff equation [KI = [I]05[L]KL + 1) KL is theconstant of dissociation for the ligand] (Cheng and Prusoff1973) was used when both the concentration of the radio-active ligand (L) and the displacing agent (I) are in excessover the protein (LT gtgt PT IT gtgt PT T indicates total concen-tration) For the two strong inhibitors pNPaMan and MeUm-baMan where IT was not in excess over PT the equationof KI = IT[(1 - Y)Y(LTKL) - 1] - PTKLYLT where Y is thefraction of the ligand bound in presence of the inhibitor wasused instead (Horovitz and Levitzki 1987)
Computational docking
Computational docking studies were performed usingAutoDock3 (Morris et al 1998) In AutoDock3 the proteinis represented by a grid of affinity potentials Hence theprotein conformation is not allowed to change during thedocking procedure Flexibility of ligands is taken intoaccount by allowing rotation around flexible torsion anglesThe docked energies are calculated as the sum of the inter-molecular interaction energy and the internal energy of theligand The search for the best interaction energy was car-ried out by means of the Lamarckian genetic algorithmLigand structures were obtained from the Protein DataBank and prepared for docking by adding hydrogens andcharges using AutoDockTools (Sanner et al 1996) Eachsimulation consisted of 100 independent runs with a popu-lation size of 200 500 generations and a maximum of25 000 000 energy evaluations Solutions were rankedbased on their docking energies and similar solutions wereclustered (cluster cut-off rmsd lt 1 Aring) For both ligandsthe top solutions were evaluated using the graphics pro-gram O (Jones et al 1991)
Acknowledgements
We thank Andrej Zavialov and Maringhns Ehrenberg for help withequilibrium binding experiments and Thomas Norberg forassistance with mass spectrometry Anthony DeFusco fromMedimmune provided us with mannosylated bovine serumalbumin We are grateful to Fred Sauer for assistance withdata collection Thanks to Maor Bar-Peled of the CCC at theUniversity of Georgia Athens USA for his suggestion toexpress FimH in minimal medium We acknowledge the useof beamline 19BM of the Structural Biology Center at theAdvanced Photon Source Argonne National LaboratoryChicago IL USA and of beamline ID29 ESRF GrenobleFrance JB is a postdoctoral fellow of the Fonds voor Weten-schappelijk Onderzoek-Vlaanderen which also supportedthe BiaCore3000 instrument (FWONL35) This work was sup-ported by grants from the Swedish Research Council andfrom the Strategic Research Foundation (Structural BiologyNetwork) to SDK
Supplementary material
The following material is available from httpwwwblackwellpublishingcomproductsjournalssuppmatmmimmi4415mmi4415smhtmTable S1 Crystallographic data and refinement statistics
References
Anumula KR (1994) Quantitative determination ofmonosaccharides in glycoproteins by high-performanceliquid chromatography with highly sensitive fluorescencedetection Anal Biochem 220 275ndash283
Bahrani-Mougeot FK Buckles EL Lockatell CV HebelJR Johnson DE Tang CM and Donnenberg MS(2002) Type 1 fimbriae and extracellular polysaccharidesare preeminent uropathogenic Escherichia coli virulence
A novel class of FimH high-affinity ligands 453
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
determinants in the murine urinary tract Mol Microbiol 451079ndash1093
Berglund J and Knight SD (2003) Structural basis forbacterial adhesion in the urinary tract Glycobiol Med 53533ndash52
Blattner FR Plunkett G Bloch CA Perna NT BurlandV Riley M et al (1997) The complete genome sequenceof Escherichia coli K-12 Science 277 1453ndashbullbull
Bloch CA Stocker BA and Orndorff PE (1992) A keyrole for type 1 pili in enterobacterial communicability MolMicrobiol 6 697ndash701
Bork P Holm L and Sander C (1994) The immunoglob-ulin fold ndash structural classification sequence patterns andcommon core J Mol Biol 242 309ndash320
Brinton CC (1959) Non-flagellar appendages of bacteriaNature 183 782ndash786
Bruumlnger AT Adams PD Clore GM DeLano WLGros P Grosse-Kunstleve RW et al (1998) Crystallog-raphy and NMR system a new software suite for macro-molecular structure determination Acta Crystallogr D BiolCrystallogr 54 905ndash921
Buchanan K Falkow S Hull RA and Hull SI (1985)Frequency among Enterobacteriaceae of the DNA-sequences encoding type-1 pili J Bacteriol 162 799ndash803
Buts L Bouckaert J De Genst E Loris R Oscarson SLahmann M et al (2003) The fimbrial adhesin F17-G ofenterotoxigenic Escherichia coli has an immunoglobulin-like lectin domain that binds N-acetylglucosamine MolMicrobiol 49 705ndash715
Cheng Y and Prusoff WH (1973) Relationship betweeninhibition constant (K1) and concentration of inhibitor whichcauses 50 per cent inhibition (I50) of an enzymatic reac-tion Biochem Pharmacol 22 3099ndash3108
Choudhury D Thompson A Stojanoff V LangermannS Pinkner J Hultgren SJ and Knight SD (1999) X-ray structure of the FimCndashFimH chaperonendashadhesin com-plex from uropathogenic Escherichia coli Science 2851061ndash1066
Collaborative computational project n4 (1994) The CCP4suite programs for crystallography Acta Crystallogr D BiolCrystallogr 50 760ndash763
Dodson KW Pinkner JS Rose T Magnusson G Hult-gren SJ and Waksman G (2001) Structural basis of theinteraction of the pyelonephritic Escherichia coli adhesin toits human kidney receptor Cell 105 733ndash743
Eley JG and Triumalashetty P (2001) In vitro assessmentof alkylglycosides as permeability enhancers AAPSPharmSciTech 2 (3) Article 19
Esnouf RM (1997) An extensively modified version of Mol-script that includes greatly enhanced coloring capabilitiesJ Mol Graph Model 15 132ndash134
Firon N Ofek I and Sharon N (1982) Interaction ofmannose-containing oligosaccharides with the fimbriallectin of Escherichia coli Biochem Biophys Res Commun105 1426ndash1432
Firon N Ofek I and Sharon N (1983) Carbohydrate spec-ificity of the surface lectins of Escherichia coli Klebsiellapneumoniae and Salmonella typhimurium Carbohydr Res120 235ndash249
Firon N Ofek I and Sharon N (1984) Carbohydrate-
binding sites of the mannose-specific fimbrial lectins ofEnterobacteria Infect Immun 43 1088ndash1090
Firon N Ashkenazis S Mirelman D Ofek I and SharonN (1987) Aromatic a-glycosides of mannose are powerfulinhibitors of the adherence of type 1 fimbriated Escherichiacoli to yeast and intestinal cells Infect Immun 55 472ndash476
Fukiya S Mizoguchi H Tobe T and Mori H (2004)Extensive genomic diversity in pathogenic Escherichia coliand Shigella strains revealed by comparative genomichybridization microarray J Bacteriol 186 3911ndash3921
Hooton TM and Stamm WE (1997) Diagnosis and treat-ment of uncomplicated urinary tract infection Infect DisClin North Am 11 551ndash581
Horovitz A and Levitzki A (1987) An accurate method fordetermination of receptor ligand and enzyme-inhibitor dis-sociation-constants from displacement curves Proc NatlAcad Sci USA 84 6654ndash6658
Hull RA Gill RE Hsu P Minshaw BH and Falkow S(1981) Construction and expression of recombinant plas-mids encoding type 1 and D-mannose-resistant pili from aurinary tract infection Escherichia coli isolate Infect Immun33 933ndash938
Hultgren SJ Porter TN Schaeffer AJ and Duncan JL(1985) Role of type 1 pili and effects of phase variation onlower urinary tract infections produced by Escherichia coliInfect Immun 50 370ndash377
Hung C-S Bouckaert J Hung DL Pinkner J WinbergC Defusco A et al (2002) Structural basis of tropism ofEscherichia coli to the bladder during urinary tract infec-tion Mol Microbiol 44 903ndash915
Jones TA Zou JY Cowan SW and Kjeldgaard M(1991) Improved methods for building protein models inelectron density maps and the location of errors in thesemodels Acta Crystallogr A 47 110ndash119
Justice SS Hung C-S Theriot JA Fletcher DAAnderson GG Footer MJ and Hulgren SJ (2004)Differentiation and developmental pathways of uropatho-genic Escherichia coli in urinary tract pathogenesis ProcNatl Acad Sci USA 101 1333ndash1338
Kallenius G Mollby R Svenson SB Windberg J Lun-dblud A Svenson S and Cedergen B (1980) The Pk
antigen as receptor for the haemagglutinin of pyelonephri-togenic Escherichia coli FEMS Microbiol Lett 8 297ndash302
Knight SD Berglund J and Choudhury D (2000) Bacte-rial adhesins structural studies reveal chaperone functionand pilus biogenesis Curr Opin Chem Biol 4 653ndash660
Langermann S and Ballou WR (2003) Development of arecombinant FimCH vaccine for urinary tract infectionsAdv Exp Med Biol 539 635ndash653
Langermann S Palaszynski S Barnhart M Auguste GPinkner JS Burlein J et al (1997) Prevention ofmucosal Escherichia coli infection by FimH-adhesin-basedsystemic vaccination Science 276 607ndash611
Langermann S Mollby R Burlein JE Palaszynski SRAuguste CG Defusco A et al (2000) Vaccination withFimH adhesin protects cynomolgus monkeys from coloni-zation and infection by uropathogenic Escherichia coli JInfect Dis 181 774ndash778
Larsson A Ohlsson J Dodson KW Hultgren SJ Nils-son U and Kihlberg J (2003) Quantitative studies of thebinding of the class II PapG adhesin from uropathogenic
454 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
Escherichia coli to oligosaccharides Bioorg Med Chem 112255ndash2261
Leffler H and Svanborg-Eden C (1980) Chemical identifi-cation of a glycosphingolipid receptor for Escherichia coliattaching to human urinary tract epithelial cells and agglu-tinating human erythrocytes FEMS Microbiol Lett 8 127ndash134
Lindhorst TK Kieburg C and Krallmann-Wenzel U(1998) Inhibition of the type 1 fimbriae-mediated adhesionof Escherichia coli to erythrocytes by multiantennary alpha-mannosyl clusters the effect of multivalency Glycoconj J15 605ndash613
Loris R Tielker D Jaeger KE and Wyns L (2003a)Structural basis of carbohydrate recognition by the lectinLecB from Pseudomonas aeruginosa J Mol Biol 331 861ndash870
Loris R Imberty A Beeckmans S Van Driessche ERead JS Bouckaert J et al (2003b) Crystal structureof Pterocarpus angolensis lectin in complex with glucosesucrose and turanose J Biol Chem 278 16297ndash16303
Lund B Lindberg F Marklund BI and Normark S(1987) The PapG protein is the a-D-galactopyranosyl-(1-4)-b-D-galactopyranose-binding adhesin of uropathogenicEscherichia coli Proc Natl Acad Sci USA 84 5898ndash5902
Min G Stolz M Zhou G Liang F Sebbel P StofflerD et al (2002) Localization of uroplakin Ia the urothelialreceptor for bacterial adhesin FimH on the six innerdomains of the 16 nm urothelial plaque particle J Mol Biol317 697ndash706
Miroux B and Walker JE (1996) Over-production of pro-teins in Escherichia coli mutant hosts that allow synthesisof some membrane proteins and globular proteins at highlevels J Mol Biol 260 289ndash298
Morris GM Goodsell DS Halliday RS Huey R HartWE Belew RK and Olson AJ (1998) Automateddocking using a Lamarckian genetic algorithm and anempirical binding free energy function J Comput Chem 191639ndash1662
Mulvey MA (2002) Adhesion and entry of uropathogenicEscherichia coli Cell Microbiol 4 257ndash271
Mulvey MA Lopez-Boado YS Wilson CL Roth RParks WC Heuser J and Hultgren SJ (1998) Induc-tion and evasion of host defenses by type 1-piliated uro-pathogenic Escherichia coli Science 282 1494ndash1497
Murshudov GN Vagin AA and Dodson EJ (1997)Refinement of macromolecular structures by the maxi-mum-likelihood method Acta Crystallogr D Biol Crystallogr53 240ndash255
Nagahori N Lee RT Nishimura S Pageacute D Roy Rand Lee YC (2002) Inhibition of adhesion of type 1 fim-briated Escherichia coli to highly mannosylated ligandsChembiochem 3 836ndash844
Navaza J (2001) Implementation of molecular replacementin AMoRe Acta Crystallogr D Biol Crystallogr 57 1367ndash1372
Neeser JR Koellreutter B and Wuersch P (1986)Oligomannoside-type glycopeptides inhibiting adhesion ofEscherichia coli strains mediated by type 1 pili preparationof potent inhibitors from plant glycoproteins Infect Immun52 428ndash436
Old DC (1972) Inhibition of the interaction between fimbrial
hemagglutinatinins and erythrocytes by D-mannose andother carbohydrates J Gen Microbiol 71 149ndash157
Orndorff PE Devapali A Palestrant S Wyse A EverettML Bollinger RR and Parker W (2004) Immunoglob-ulin-mediated agglutination of and biofilm formation byEscherichia coli K-12 require the type 1 pilus fiber InfectImmun 72 1929ndash1938
Oscarson S and Tedebark U (1995) Syntheses of deoxyanalogues of methyl 36-di-O-a-D-mannopyranosyl-a-D-mannopyranoside for studies of the binding site of con-canavalin A Carbohydr Res 278 271ndash287
Oscarson S and Tiden AK (1993) Syntheses of the octyland tetradecyl glycosides of 36-di-O-alpha-deuterium-mannopyranosyl-alpha-deuterium-mannopyranose and of34-di-O-alpha-deuterium-mannopyranosyl-alpha-deute-rium-mannopyranose ndash a new way for 24-di-O-protectionof mannopyranosides Carbohydr Res 247 323ndash328
Otwinowski Z and Minor W (1997) Processing of X-raydiffraction data collected in oscillation mode MethodsEnzymol 276 307ndash326
Powell HR (1999) The Rossmann Fourier autoindexingalgorithm in MOSFLM Acta Crystallogr D Biol Crystallogr55 1690ndash1695
Pratt LA and Kolter R (1998) Genetic analysis of Escher-ichia coli biofilm formation roles of flagella motility chemo-taxis and type I pili Mol Microbiol 30 285ndash293
Remaut H and Waksman G (2004) Structural biology ofbacterial pathogenesis Curr Opin Struct Biol 14 161ndash170
Ronald AR Nicolle LE Stamm E Krieger J WarrenJ Schaeffer A et al (2001) Urinary tract infection inadults research priorities and strategies Int J AntimicrobAgents 17 343ndash348
Sanner MF Olson AJ and Spehner JC (1996)Reduced surface an efficient way to compute molecularsurfaces Biopolymers 38 305ndash320
Sauer FG Futterer K Pinkner JS Dodson KW Hult-gren SJ and Waksman G (1999) Structural basis ofchaperone function and pilus biogenesis Science 2851058ndash1061
Sauer FG Knight SD Waksman GW and HultgrenSJ (2000) PapD-like chaperones and pilus biogenesisSemin Cell Dev Biol 11 27ndash34
Sauer FG Pinkner JS Waksman G and Hultgren SJ(2002) Chaperone priming of pilus subunits facilitates atopological transition that drives fiber formation Cell 111543ndash551
Schembri MA Hasman H and Klemm P (2000)Expression and purification of the mannose recognitiondomain of the FimH adhesin FEMS Microbiol Lett 188147ndash151
Schembri MA Kjaergaard K and Klemm P (2003) Glo-bal gene expression in Escherichia coli biofilms Mol Micro-biol 48 253ndash267
Schilling JD Mulvey MA Vincent CD Lorenz RGand Hultgren SJ (2001) Bacterial invasion augments epi-thelial cytokine responses to Escherichia coli through alipopolysaccharide-dependent mechanism J Immunol166 1148ndash1155
Silhavy TJ Szmelcman S Boos W and Schwartz M(1975) On the significance of the retention of ligand byprotein Proc Natl Acad Sci USA 72 2120ndash2124
A novel class of FimH high-affinity ligands 455
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
Svanborg C and Godaly G (1997) Bacterial virulence inurinary tract infection Infect Dis Clin North Am 11 513ndash529
Thanassi DG Saulino ET and Hultgren SJ (1998) Thechaperoneusher pathway a major terminal branch of thegeneral secretory pathway Curr Opin Struct Biol 1 223ndash231
Thankavel K Madison B Ikeda T Malaviya R ShahAH Arumugen PM and Abraham SN (1997) Local-ization of a domain in the FimH adhesin of Escherichia colitype 1 fimbriae capable of receptor recognition and use ofa domain-specific antibody to confer protection againstexperimental urinary tract infection J Clin Invest 1001123ndash1126
Vagin A and Teplyakov A (1997) MOLREP an automatedprogram for molecular replacement J Appl Crystallogr 301022ndash1025
Wu XR Sun TT and Medina JJ (1996) In vitro bindingof type 1-fimbriated Escherichia coli to uroplakins 1a and
1b relation to urinary tract infections Proc Natl Acad SciUSA 93 9630ndash9635
Xia Y Gally D Forsman-Semb K and Uhlin BE (2000)Regulatory cross-talk between adhesin operons in Escher-ichia coli inhibition of type 1 fimbriae expression by thePapB protein EMBO J 19 1450ndash1457
Zafriri D Ofek I Adar R Pocino M and Sharon N(1989) Inhibitory activity of cranberry juice on adherenceof type-1 and type-P fimbriated Escherichia coli to eukary-otic cells Antimicrob Agents Chemother 33 92ndash98
Zavialov AV Berglund J Pudney AF Fooks LJ Ibra-him TM MacIntyre S and Knight SD (2003) Structureand biogenesis of the capsular F1 antigen from Yersiniapestis preserved folding energy drives fiber formation Cell113 587ndash596
Zhou G Mo WJ Sebbel P Min G Neubert TA Glock-shuber R et al (2001) Uroplakin Ia is the urothelial recep-tor for uropathogenic Escherichia coli evidence from invitro FimH binding J Cell Sci 114 4095ndash4103

A novel class of FimH high-affinity ligands
443
copy 2004 Blackwell Publishing Ltd
Molecular Microbiology
55
441ndash455
highly specific small-molecule inhibitors targeting withhigh-affinity the FimH mannose binding site may preventUTIs by interfering with the attachment of
E coli
to theuroepithelial cells lining the bladder mucosa
Results
Affinity of FimH for mono- and disaccharides
Two different binding assays were developed one basedon surface plasmon resonance (SPR) and one based onequilibrium binding measurements using radioactivelylabelled mannose (Table 1) Inhibition by mannose of thebinding of FimH to the immobilized Fab fragments of themonoclonal antibody IC10 [k
a
= 147 yen 104 (plusmn417) M-1
s-1 kd = 411 yen 10-3 (plusmn565 yen 10-6)s-1 Kd = 280 yen 10-7
(plusmn88 yen 10-9) M] in SPR measurements allowed us toderive a solution affinity of mannose for FimH(Kd = 23 mM) (Table 1) Using either the Fabs of mono-clonal antibody IA7 or BSA-Man as the immobilized bind-ing partner in the competition experiment gave equivalentdissociation constants for the FimHndashmannose interaction(data not shown) To eliminate the possibility of differentbinding strength for full-length FimH and FimH truncatesthe binding of D-mannose to FimCFimH complex wasalso measured A value of Kd = 53 mM was obtained ingood agreement with the value measured using FimHtruncates
Removal of the 2-OH group of mannose leads to a 100-fold reduction in affinity for FimH (Table 2) Glucose theequatorial O2 epimer of mannose binds a 4000-foldweaker There is virtually no affinity for galactose the O4(axial) epimer and O2 (equatorial) epimer of mannoseFructose binds strongly to FimH (Kd = 31 mM) although a15-fold weaker than mannose The fructose-containingdisaccharides sucrose [Glc(a1ndash2)Frufb] and turanose[Glc(a1ndash3)Frub] have affinities for FimH very similar toglucose
Structure of the FimH lectin domain
The lectin domains FimH truncate 1 (FimHtr1) originatingfrom the UPEC strain J96 and FimH truncate 2 (FimHtr2)from the E coli K-12 laboratory strain MG1655 werecloned expressed and crystallized The structures weresolved by molecular replacement using the lectin domainfrom the FimCFimH complex (Choudhury et al 1999) asa search model and refined to 17 Aring (FimHtr1) and 21 Aring(FimHtr2) resolution respectively (Table S1 in Supplemen-tary material) Both models include residues 1ndash158 cor-responding to the entire lectin domain of FimH Twocopies of the lectin domain are present per asymmetricunit of the FimHtr2 crystals compared with only one unitfor the FimHtr1 crystals The carboxy-terminal 6-His tag ofFimHtr2 is not visible in the electron density maps and isnot included in the model
The structures of the FimH lectin domains are verysimilar to the structure for the same domain in theFimCFimH complex with a root mean square deviation(rmsd) over all 158 Ca positions of 036 Aring for FimHtr1056 Aring for FimHtr2 subunit A and 054 Aring for FimHtr2 subunitB The secondary structure elements arranged in an Igfold (Buts et al 2003) (Fig 1A) have almost interchange-able Ca traces in all of the FimH lectin domain structuresOnly Gly73 (in strand D1) and Gly79 (in loop connectingstrands D1 and Dcent) have slightly variable main chain con-formations while Val30 in the b3ndashb4 loop at the base ofthe domain clearly undergoes a peptide flip in all threesubunits of the truncate compared with the FimCFimHcomplex In the FimHtr1 structure the conformation of thecarboxy-terminal residue of the truncate Thr158 differsas it is packed against and hydrogen bonded to a glycerolmolecule via its carboxylate group In the FimHtr2 crystalstructure the main difference is in the b9ndashb10 loop withan rmsd of 29 Aring over Ca atoms 113ndash116 This loopadopts a different conformation because of crystal pack-ing contacts that are not present in the FimCFimH orFimHtr1 structures
Mannose binding pocket
Although no sugar was included in the crystallization set-ups strong positive electron density was present in the
Table 1 Kd and calculated DG0 for a series of O1 alkyl and arylmannosides
LigandKd SPR(nM)
DGinfin SPR (kcal mol-1)
Kd displace(nM)
DGinfin displace(kcal mol-1)
Mannose 23 yen 103 -76 41 yen 103 -73Methylaman 22 yen 103 -77 24 yen 103 -76Ethylaman 12 yen 103 -81 730 -83Propylaman 300 -89 400 -87Butylaman 151 -93 150 -93Pentylaman 25 -104 200 -91Hexylaman 10 -109 100 -95Heptylaman 5 -113 32 -102Octylaman 22 -104 28 -103PNPaMan 44 -100 26 -103MeUmbaMan 20 -105 12 -108
The SPR and displacement binding experiments define heptyl a-D-mannopyranoside as the best binder
Table 2 Kd and calculated DG0 for a series of mono- and disaccha-rides and a deoxymannose
Ligand Kd
DGinfin(kcal mol-1)
Methyl 2-deoxy-a-D-mannopyranoside 300 mM -48Glucose 924 mM -28Galactose 100 mM -14Fructose 31 mM -61Sucrose 128 mM -26Turanose 76 mM -29
444 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
initial FondashFc and 2FondashFc Fourier maps inside the mannose-binding pocket of both FimHtr1 and FimHtr2 This densityprecisely defined a-D-mannose with an additional elon-gated density shaped like a creased tail extending out-wards from the mannose O1 and towards tyrosines 48and 137 which make up part of the hydrophobic rimaround the mannose-binding pocket (Fig 1B) (Hunget al 2002) Further refinement strongly suggested thata butyl a-D-mannoside was bound in the mannose-bindingsite (Fig 1B and C) For FimHtr1 also a change in theconformation of the Tyr48 side-chain compared with theone from the model FimH used for molecular replacementwas immediately apparent As described previously (Hunget al 2002) the mannose ring makes 10 direct hydrogenbonds to the side-chains of residues Asp54 Gln133Asn135 and Asp140 and to the main chain of Phe1 andAsp47 and indirect water-mediated hydrogen bonds viaO2 to the side-chain of Glu133 and to the main chainoxygen of Phe1 and Gly14 The alpha-anomeric hydroxylgroup O1 of mannose is involved in a hydrogen-bondingwater network with the Asn138 and Asp140 side-chainsthrough a water molecule (wat 2 Fig 1B) that is con-
served between the two truncate structures The positionsof the other waters participating in this network dependon the crystal packing of FimHtr1 and FimHtr2
Mass spectrometric analysis of the bound ligand
To corroborate the nature of the ligand identified in thecrystal structures the complex of FimHtr1 and its pre-sumed butyl a-D-mannoside was analysed using massspectrometry The FimHtr1 protein sample used for crys-tallization was subjected to peracetylation and phase sep-aration The peracetylated ligand retrieved in the organicphase was analysed by nanoLC-online-ESI-MS The per-acetylated ligand eluted as a single peak and was regis-tered in its sodiated form at mz 427 corresponding tobutyl hexose (Fig 2A) No signals corresponding to otherpotential alkyl mannose species like ethyl hexose (mz399) propyl hexose (mz 413) or pentyl hexose (mz 441)could be observed indicating a homogeneous ligand witha four-carbon atoms alkyl chain Besides the sodiumadduct of the ligand a major signal was observed arisingfrom the loss of butanol (mz 331 Fig 2B) Cleavage at
Fig 1 Stereo views of FimHA Three-dimensional structure of the FimH lectin domain is an elongated 11-stranded b-barrel with an immunoglobulin fold Numbering is according to conventions established for antibody domains (Bork et al 1994) and for PapD (Sauer et al 1999) and loop identifications according to previous descriptions (reference strand numbers A1 = 1 Acent = 2 A2 = 3 B1 = 4 B2 = 4cent C = 5 D1 = 6 Dcent = 7 D2 = 8 E = 9 F = 10 G = 11) (Choudhury et al 1999 Hung et al 2002) The disulphide bridge between Cys3 and Cys44 is coloured cyanB Focus on the mannose-binding pocket in the same orientation as (A) showing the hydrophobic contacts (within 46 Aring orange) with butyl a-D-mannoside and the water network (salmon pink) present at O1C Surface presentation (wheat yellow) of the different alkyl conformations of the butyl a-D-mannoside in the crystal structures of FimH truncates 1 and 2 FimHtr1 butyl group (cyan) FimHtr1 Tyr48 (marine blue) FimHtr2 butyl group and Tyr48 (salmon pink) Tyr137 (violet) Ile52 (pink)
A B
C
A novel class of FimH high-affinity ligands 445
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
the anomeric carbon atom was often combined with elim-ination of one or more acetic acid molecules (massdecrease of 60 Da) or acetyl groups (mass decrease of42 Da) These eliminations gave rise to the major ions atmz 127 and mz 169 with a proposed aromatic ring struc-ture as well as to the ion at mz 109 with a proposedquinone-like ring structure Thus the mass spectrometricdata support the butyl hexose nature of the FimH ligandFimH was further characterized by carbohydrate constit-uent analysis (data not shown) which revealed only man-nose Mannose could only be detected after hydrolysisindicating that free mannose is not associated with theFimH truncate
Origin of butyl a-D-mannoside
As no sugar had been advertently added to the FimHtr
protein samples we hypothesized that the butyl manno-side ligand had been picked up from the LuriandashBertani(LB) medium used to grow bacteria during expression ofthe protein To test this FimHtr2 was expressed in bacteriagrown in minimal M9 medium Electro-spray ionizationmass spectrometry (ESI-MS) was performed on purifiedFimHtr2 samples from both types of media A clear peakat 259 Da could be detected (Fig 3A) with proteinexpressed in bacteria grown in LB medium correspondingto the molecular weight of butyl mannoside (236 Da) plus
a sodium ion (23 Da) As a control butyl mannoside wassynthesized and analysed in the same manner Again apeak at 259 Da was observed (Fig 3A inset) When ESI-MS was run on protein expressed in bacteria grown inminimal medium no such peak could be detected(Fig 3B) indicating that the ligand was not present andthat it indeed originated from the LB medium
Interactions of the butyl moiety with FimH
The butyl moiety of butyl a-D-mannoside extends out ofthe mannose-binding pocket towards Tyr48 and Tyr137making van der Waals contacts to both tyrosine rings andIle52 (Fig 1B and C) These residues are part of two loops(b4ndashb5 and b10ndashb11) that form the higher rear end of ahydrophobic collar around the binding site (Fig 1A andB) In both subunits of the FimHtr2 structure the Tyr48 andTyr137 side-chains are in the same almost parallel ori-entation as in the FimCFimH structure forming a gate-like structure at the back of the binding site (Fig 1C) Theclosest hydrophobic interactions (31ndash36 Aring) with the alkylchain are made between Tyr48 and the two non-terminalatoms of the butyl In the FimHtr1 structure a crystal con-tact to Val155 of a neighbouring molecule prevents theparallel orientation of the Tyr48 ring which instead ispacked edge-to-face with Tyr137 (Fig 1B) The reposition-ing of Tyr48 is accompanied by a change in the confor-
A
B
Fig 2 Mass spectrophotometric characteriza-tion of the butyl mannosideA Base peak chromatogram (BPC mass range mz 400ndash550) and extracted ion chromato-grams (EIC) of mz 427 and 331 obtained by nano-LC-ESI-MS of the derivatized FimH-ligand purified by peracetylation and organic-solvent extractionB ESI-IT mass spectrometric characterization of the peak species marked by a horizontal bar in (A) Ions are marked with mz-values and proposed schematic structures which is often only one out of several isobaric ion species Fragmentations already occurred in the source Ac acetyl
446 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
mation of the butyl chain of the ligand with the butyl chainin a more extended conformation than in the FimHtr2 struc-ture and with the four carbon atoms nearly in one plane(Fig 1B and C) The new conformations of Tyr48 and thebutyl moiety result in an almost parallel stacking of thearomatic ring of Tyr48 onto the butyl plane with the closesthydrophobic interactions (37 Aring) involving the two terminalcarbon atoms of the butyl (Fig 1B)
Affinity measurements for alkyl mannosides
The butyl mannoside ligand remained bound to FimHtr
despite extensive dialysis From a study of complex for-mation of monoclonal antibodies IC10 and IA7 withFimHtr1 it could be estimated that about 30 of the FimHtruncate retained its ligand after dialysis for 12 daysagainst 50 mM MES buffer at pH 65 100 mM NaCldespite daily refreshment of buffer (data not shown)Therefore to enable measurements of dissociation con-stants of FimH for mannosides FimH was produced inminimal medium Butyl a-D-mannoside binding is 15 timesimproved (Kd = 151 nM Fig 4A) compared with mannosebinding The equilibrium binding technique using [3H]-mannose led to a similar value (Kd = 41 mM Fig 4B) for
mannose and an almost identical value for butyl a-D-mannoside (Kd = 150 nM Fig 4C) To investigate theeffect of sequential addition of methyl groups to the O1oxygen of D-mannose a series of alkyl mannosides weresynthesized and the dissociation constants determinedusing the [3H]-mannose displacement assay as well as theSPR competition assay (Table 1) There is a linear corre-lation between the binding free energy as calculated fromthe measured dissociation constants and the number ofmethyl groups (between 1 and 7) in the alkyl mannosidewith each additional methyl group contributing on average-06 kcal mol-1 of binding energy (Fig 5)
Affinity measurements and modelling for aryl mannosides
Aromatically substituted mannosides have been reportedto be particularly potent inhibitors of FimH-mediated bac-terial adhesion (Firon et al 1987 Nagahori et al 2002)In accordance with the earlier results both pNPaMan andMeUmbaMan bind very tightly to FimH (Table 1) Despiteextensive trials no X-ray structures were obtained forcomplexes with aromatically substituted mannosides Inorder to model binding docking to both of the truncatestructures was carried out In all cases the mannose ring
Fig 3 Mass spectrometric analysis of the origin of the butyl mannosideA LB medium-produced FimH truncate The inset shows the mass spectrum of the synthesized butyl a-D-mannosideB Minimal media-produced FimH truncate
A novel class of FimH high-affinity ligands 447
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
docks in the binding pocket in a conformation very closeto the crystallographically observed mannose conforma-tion (Fig 6) while the aromatic substituent interacts withthe tyrosine gate The different conformation of Tyr48 in
our two FimH truncate structures leads to different pre-dicted interactions between the aromatic aglycon andTyr48 and Tyr137 Docking to the FimHtr2 structureresulted in somewhat tighter interactions between thetyrosine gate and the aglycon and hence in somewhatlower docking energies Calculated FimHtr2 docking ener-gies of Edoc = -104 kcal mol-1 for pNPaMan (Kd = 46 nM)and Edoc = -109 kcal mol-1 for MeUmbaMan (Kd = 20 nM)are in good agreement with the experimental binding data(Table 1) The docked umbelliferyl ring system is inserteddeep into the tyrosine gate in the FimHtr2 conformationwith the carbonyl group within hydrogen bonding distanceto Thr51 at the back of the gate (Fig 6A) Docking to theFimHtr1 structure instead stacks the umbelliferyl ring face-to-face with the Tyr48 ring (not shown) The phenyl ring ofpNPaMan docks similarly to both our structures It doesnot quite reach in between the tyrosine side-chains butinstead hydrogen bonds with its nitro group to the hydroxyloxygen of Tyr137 (Fig 6B)
Discussion
Most lectins bind naturally occurring carbohydrates withlow affinities often in the millimolar range for monosac-charides and micromolar range for oligosaccharides Thiswas recently exemplified to also hold true for theadhesins of F17 fimbriae (Kd = 12 mM for the F17a-G-N-acetylglucosamine interaction Buts et al 2003) and Ppili (Kd = 84 mM for the PapGII-globotetraoside interactionLarsson et al 2003) The recent crystal structure of FimHin complex with mannose illustrated that FimH almostcompletely envelops the monosaccharide in a deeppocket and involves it in an intense hydrogen bondingnetwork suggestive of an unusual lectin exerting highaffinities (Hung et al 2002)
Fig 4 FimH binding experimentsA Surface plasmon resonance competition experiment of FimHtr1 binding to monoclonal antibody IC10 Fab fragments with butyl a-D-mannosideB Equilibrium binding of [3H]-mannose to FimHtr2C Equilibrium displacement measurements of butyl a-D-mannoside to the FimHtr2 [3H]-mannose complex
log[Butyl a-D-mannoside] (M)
cpm
cpm
Butyl a-D-mannoside (mM)
Mannose (mM)
A
B
C
ndash64
ndash66
ndash68
ndash70
ndash72
ndash74
ndash76
ndash78
ndash80
ndash82
ndash8 ndash7 ndash6 ndash5 ndash4 ndash3
log
[Fim
H]
M
Fig 5 Linear free energy relationship for the alkyl chain on mannose A linear fit was made using 1 up to 7 methyl groups with the program MICROCAL ORIGIN 50
Number of carbon atoms0 2 4 6 8
ndash75
ndash80
ndash90
ndash95
ndash100
ndash105
ndash110
ndash115
ndash85
D G
(kc
al m
olndash1
)
slopecorrelation
ndash064ndash099
+ndash 003+ndash 018
448 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
The measured affinity confirm the predictions that FimHbinds the monosacharide mannose exceptionally well(Kd = 23ndash41 mM) To investigate the contribution of thedifferent hydroxyl groups of mannose in the interactionwith FimH the binding of a series of hexoses to butylmannoside-free FimH was examined using SPR (Table 2)The effect of deoxygenation of the O2 hydroxyl group(methyl 2-deoxy-a-D-mannopyranoside) is clearly lessdestructive (100-fold reduction) than its equatorial positionas in glucose O2 is the only hydroxyl group of mannosethat interacts indirectly with the FimH pocket via a con-served water molecule (Hung et al 2002) Glucose onlydisplays a weak affinity for FimH about 4000 times lowerthan methyl a-D-mannoside Repositioning of both the O2and O4 hydroxyl groups as in galactose abolishes sugarbinding by FimH Similar to these results the inhibitoryconcentrations for bacterial adherence of methyl a-D-glucoside and galactose derivatives have been estimatedto be respectively 104-fold and 105-fold weaker than formethyl a-D-mannoside (Nagahori et al 2002)
In stark contrast with the other monosaccharides fruc-tose has a binding affinity only about 15-fold less thanmannose for FimH (Table 2) The decrease in affinity for
fructose relative to mannose agrees with studies of inhi-bition of adherence of type 1 piliated E coli by fructosein which fructose was found to be the active compound infruit juices (present at about 5) (Zafriri et al 1989)Fructose binding to FimH is stronger than the binding ofthe physiological globotetraoside receptor to PapGII theother well-characterized adhesin of UPEC strains (Lars-son et al 2003) The pyranose form of fructose is verywell superimposable onto mannose in the FimH bindingsite where similar interactions are established with FimHas for mannose except for the loss of the anomeric O1and an extra hydroxyl group (O2) on the C5 position ofmannose Probably however both changes of fructopyr-anoside compared with mannopyranoside could be easilyaccommodated by FimH The mannose O1 does notmake direct interactions within 45 Aring of the FimH bindingpocket (Fig 1B) The extra O2 group of fructose would notclash or interfere significantly with the binding pocket butit would make a 27 Aring van der Waals contact with the Ile52side-chain under the condition that the Ile52 side-chaindoes not move The crystal structure of the Pseudomonasaeruginosa lectin LecB also showed that fructopyranosebut not fructofuranose can substitute for mannose (Loris
Fig 6 Stereo diagrams of docked MeUm-baMan (A) and pNPaMan (B) in the FimH bind-ing site The crystallographically determined position of a-D-mannose is shown in orange for comparison
A novel class of FimH high-affinity ligands 449
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
et al 2003a) Hydrogen bonds of the more specificfucose complex are replaced by van der Waals interac-tions involving a hydroxyl group in the complexes with thelower-affinity binders mannose and fructopyranose as forthe proposed fructopyranoside binding to FimH
The fructose-containing disaccharides sucrose andturanose do not have an inhibitory effect and bind withaffinities similar to glucose Sucrose is a non-reducingsugar that has fructose locked in its furanose form Tura-nose on the other hand allows the interconversion of b-D-fructofuranose and b-D-fructopyranose eg both formsoccur in complex with a plant lectin from Pterocarpusangolensis (Loris et al 2003b) It is predictable howeverfrom the crystal structures that the glycosidic linkage ofglucose with C3 of fructose equivalent in position to C4of mannose would be disallowed in the tight FimH pocketSucrose and turanose thus most probably interact with theFimH mannose binding pocket via their glucose moietyresulting in similar low affinities
Our mass spectrometry and chemical analyses con-firms the crystallographic identification of the ligand boundto the FimH lectin domains as butyl a-D-mannoside(Figs 1B and 2) The mannose ring of the butyl a-D-mannoside coincides with the mannose bound to theFimCFimH complex (Hung et al 2002) and makes thesame interactions The four-carbon alkyl tail exits towardsthe back wall (loop b4ndashb5 and b10ndashb11) of the pocket(Fig 1A and B) making hydrophobic contacts to Tyr48Tyr137 and Ile52 The mass spectrometry data also showthat the bound butyl a-D-mannoside is present in LBmedium but absent from minimal medium Hence thesource of the alkyl mannoside is possibly the yeast extractused in preparation of LB medium Although we do notwant to speculate about the significance of this findingalkyl mannoside production by yeast has not beenreported and might warrant investigation
The butyl mannoside was retained by FimH despiteextensive dialysis The phenomenon of ligand retention byprotein was described as early as 1975 for periplasmicbinding proteins (Silhavy et al 1975) Applied to dialysisof butyl mannoside away from FimH grown in richmedium half-lives of up to 30 h can be estimated whichis far beyond the duration of a normal buffer exchangedialysis experiment In the present case the retainedligand does not appear to seriously affect [3H]-mannosedisplacement equilibrium binding experiments as wecould detect no significant differences in dissociation con-stants obtained using FimHtr2 produced in LB or minimalmedia (data not shown) In contrast proper SPR bindingcurves could not be obtained when butyl mannoside occu-pied the binding site Hence the displacement techniquemay be of more general use to obtain binding constantsfor proteins that tend to inadvertently retain ligands whichis a common trait of lectins
Surface plasmon resonance as well as equilibrium bind-ing and displacement measurements using [3H]-mannosewere carried out to obtain dissociation constants for a-D-mannose and a linear series of alkyl-substituted manno-sides to FimH (Table 1) Both equilibrium binding experi-ments show a very good agreement in their measurementof the carbohydrate affinity of the adhesin FimH bindsbutyl a-D-mannoside 15ndash30 times stronger than a-D-mannose The importance of the added hydrophobiccontacts is clearly demonstrated by the binding of alkylmannosides (Table 1) The best binding alkyl mannosidesbind a few hundred times stronger than mannose equiv-alent in affinity to the most tightly binding aromaticallysubstituted mannosides for FimH (Firon et al 1987) andof mannose dendrimers for type 1 piliated E coli (Fironet al 1984 1987 Nagahori et al 2002)
Interestingly the butyl moiety takes on significantlydifferent conformations in FimHtr1 and FimHtr2 (Fig 1C)indicating that intrinsic conformational flexibility in theaglycon is allowed Indeed in spite of the larger flexibilityexpected for longer alkyl mannosides the binding energydecreases linearly with increasing length of the alkyl chainbetween one and seven carbon atoms (Fig 5) The affinityreaches its maximum for mannose with a seven-carbonalkyl and SPR measurements suggest that the affinitydecreases from eight carbon atoms on Evidently theeighth carbon atom goes beyond the hydrophobic interac-tion surface and protein edge which is penalized with adecrease in affinity We propose that the affinity increaseresults from an increased interaction surface with thehydrophobic gateway Alkyl aglycons fit well in the tyrosinegate and as observed in the two crystal structures in atleast two different conformations that depend on theconformation of Tyr48 This flexibility may indicate thataglycons can interact rather non-specifically with thehydrophobic gateway nevertheless resulting in a 100- to1000-fold increase in affinity
FimH binding to ligands other than just mannose ispossibly of importance in terms of the different environ-ments that E coli will encounter ndash the urinary tract gas-trointestinal tract and in biofilms Alkyl mannosides havenot been previously recognized as strong binders to FimHAs they are easily synthesized and highly soluble in waterthey may be interesting as potential blocking agents forFimH-mediated adhesion Moreover they have beenshown to be good drug vehicles themselves acting aspermeability enhancers on tight junctions (Eley and Triu-malashetty 2001) As such they might be able to pene-trate into deeper cell layers of the epithelium to alsoprevent those bacteria that form a quiescent reservoirfrom causing recurrent infections (Mulvey et al 1998Justice et al 2004)
On the other hand the two aromatically substitutedmannosides pNPaMan and MeUmbaMan are known to
450 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
be particularly potent inhibitors of FimH-mediated bacte-rial adhesion (Firon et al 1987 Nagahori et al 2002)Based on adhesion inhibition studies using type 1 piliatedE coli and guinea pig ileal epithelial cells the relativeinhibitory potential of pNPaMan and MeUmbaMan havebeen estimated to be about 70 and 600 times strongerrespectively than that of methyl a-D-mannose (Fironet al 1987) Our measurements indicate an affinity ofFimH for pNPaMan and MeUmbaMan between 50 and90 times and between 110 and 200 times higher respec-tively than for methyl a-D-mannose in good agreementwith the adhesion inhibition results (Firon et al 1987)Dissociation constants predicted using AutoDock3 (Morriset al 1998) are similar to experimental values Thecomputational docking predicts that tight binding of thearomatically substituted mannosides pNPaMan andMeUmbaMan is accomplished similarly as for butyl a-D-mannoside with the aglycon interacting with the tyrosinegate
This is the first systematic study providing molecularbinding data for the FimH adhesin the prerequisite forcolonization by UPEC (Langermann and Ballou 2003)We also show crystallographic evidence that the lectinor receptor binding domain of FimH is a stable andfunctional entity Its structure is virtually identical to thesame domain in the FimCFimH complex indicating thata truncate corresponding to the lectin domain could pro-vide a good alternative for the development of a UTIvaccine (Langermann et al 1997 Langermann and Bal-lou 2003) However progress of this interesting UTI vac-cine candidate into clinical trials has not been reportedThe binding affinities of FimH at the molecular levelrelate convincingly well with the inhibitory concentrationsused in previous bacterial adhesion assays thus endors-ing the direct and conclusive correlation of FimH withbacterial adherence Alkyl mannosides have nanomolaraffinities for FimH which may overcome the significantobstacle of low affinity generally encountered in attemptsto develop carbohydrate-based drugs which interferewith microbial attachment to host cell surface glycocon-jugates New anti-adhesive drugs could be based onthese leads to allow specific interaction and targeting tothe urinary tract in order to prevent anticipated andrecurrent infections
Experimental procedures
Strains and constructs
FimH proteins from two different E coli strains the UPECstrain J96 (Hull et al 1981) and the E coli K-12 strainMG1655 (Blattner et al 1997) were used in this study TheFimH adhesins encoded by these two E coli strains areidentical FimHtr1 (residues 1ndash158 of UPEC J96 FimH) wasexpressed from plasmid pMMB91 transformed into E coli
C600 cells FimHtr2 (residues 1ndash158 of E coli MG1655FimH + carboxy-terminal 6-His tag) was expressed fromplasmid pPKL241 (Schembri et al 2000) transformed into Ecoli host strain HB101
Purification and crystallization
FimHtr1 Bacteria were grown in LB medium at 37infinC inducedat OD600 = 08 with 1 mM IPTG and harvested 2ndash3 h laterThe cells were resuspended in 20 mM Tris-HCl pH 80 5 mMEDTA and 30 sucrose and the periplasmic content wasextracted by osmotic shock upon 10-fold dilution with 20 mMTris-HCl pH 80 Purification of FimHtr1 was performed by ionexchange chromatography with MonoS at pH 45 followed byMonoQ (Amersham Biosciences) at pH 8 FimHtr1 was crys-tallized using vapour diffusion in hanging drops composed of3 ml of the precipitant solution (01 M Hepes pH 75 12PEG 6000 and 5 vv methyl-24-pentanediol) and 3 ml of12 mg ml-1 protein solution in 20 mM Tris-HCl pH 80 with150 mM NaCl
FimHtr2 Cells were grown in LB medium containing50 mg ml-1 ampicillin at 37infinC At OD600 = 06 the cells wereinduced with 5 mM IPTG Cells were harvested 2 h afterinduction and the periplasmic content was extracted Theprotein was purified on a Pharmacia HiTrap Chelating HP5 ml column (Pharmacia) loaded with Ni-chloride and elutedwith a sharp 0ndash500 mM imidazole gradient Fractions con-taining FimHtr2 were pooled dialysed against 50 mM sodiumacetate pH 525 and loaded onto a Mono S HR 8 ml columnThe protein was eluted with a 0ndash500 mM NaCl gradient anddialysed overnight against 20 mM MES pH 65 before crys-tallization Crystallization was performed at room tempera-ture by the vapour diffusion method with hanging dropsmade by mixing of 2 ml of 15 mg ml-1 protein solution with 2 mlof the precipitant solution (65 methyl-24-pentanediol100 mM cacodylate pH 65) Microseeding was employed tooptimize crystal quality and reproducibility Crystals suitablefor X-ray data collection generally appeared within 48 h aftermicroseeding
Structure determination
X-ray data for FimHtr1 were collected from a single crystalto 17 Aring resolution at the Advanced Photon SourceArgonne National Laboratory USA The crystal was flash-cooled to 100 K in the precipitant solution complementedwith 25 glycerol The data were processed and scaledusing the HKL2000 package (Otwinowski and Minor 1997)(Table S1 in Supplementary material) Data for FimHtr2
were collected from a single crystal under liquid-nitrogencryo conditions at 100 K at beamline ID29 ESRF Greno-ble France to 21 Aring Bragg spacing FimHtr2 data were pro-cessed and reduced using MOSFLM (Powell 1999) andscaled using SCALA (Collaborative computational project1994) (Table S1 in Supplementary material) The structuresof FimHtr1 and FimHtr2 were solved by molecular replace-ment using Amore (Navaza 2001) and MOLREP (Vaginand Teplyakov 1997) respectively with the FimH lectindomain from the FimCFimH complex (Choudhury et al
A novel class of FimH high-affinity ligands 451
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
1999) (PDB id 1QUN) as the search model Positional bulksolvent and isotropic temperature factor refinement wasperformed using CNS (Bruumlnger et al 1998) and REFMAC5(Murshudov et al 1997) respectively Model adjustmentand inspection were performed using Turbo-Frodo and O(Jones et al 1991) graphics programs respectively Refine-ment statistics are given in Table S1 Molecular graphicsillustrations were generated with Bobscript (Esnouf 1997)and Pymol 090 (DeLano Scientific LLC 1998ndash2003 httppymolsourceforgenet)
Mass spectrometry
The purified FimHtr1 protein (300 mg) that had been used forcrystallization was freeze-dried taken up in 100 ml of pyridineand 100 ml of acetic anhydride and incubated overnight atroom temperature in the dark for peracetylation The samplewas dried under nitrogen and taken up in chloroform (400 ml)The chloroform phase was extracted six times with waterdried and taken up in 200 ml of 50 aqueous methanol A1 ml aliquot was applied to a nano-scale RP column(75 mm yen 100 mm PepMap DionexLC Packings) using anUltimate nano-LC system (LC Packings) which was equili-brated with eluent A (H2Oacetonitrile 955 vv containing01 formic acid) at a flow rate of 100 nl min-1 After injectingthe sample the column was run isocratically for 5 min fol-lowed by a linear gradient to 100 eluent B (H2Oacetonitrile2080 vv containing 01 formic acid) in 20 min and a finalwash with 100 B for 5 min The system was directly coupledto an Esquire 3000 ESI-IT-MS (Bruker Daltonik) equippedwith an online nanospray source operating in the positive-ionmode For electrospray (900ndash1200 V) capillaries (360 mmOD 20 mm ID with 10 mm opening) from New Objective wereused The solvent was evaporated at 150infinC with a nitrogenstream of 6 l min-1
To verify the suspected source of the bound butyl manno-side mass spectrometry was performed on samples ofFimHtr2 grown in either LB or minimal M9 medium Proteinsamples were dialysed against water overnight before massspectrometry in order to reduce background from the Trisbuffer Samples were diluted to a concentration in the submi-cromolar range Electrospray mass spectra were recorded indirect inlet mode (50 aqueous methanol solutions) on aBruker Daltonics Esquire-LC (Billerica)
Carbohydrate constituent analysis
Samples were hydrolysed in 100 ml of 4 M aqueous trifluoro-acetic acid (Sigma) at 100infinC for 4 h and dried under astream of nitrogen Monosaccharides were converted intotheir anthranilic acid derivatives by reductive aminationresolved by RP-HPLC and detected by fluorescence (Anu-mula 1994)
Carbohydrates for binding studies
[3H]-mannose was obtained from Amersham Methyl a-D-mannoside glucose galactose fructose sucrose turanosep-nitrophenyl a-D-mannopyranoside (pNPaMan) and 4-methylumbelliferyl a-D-mannopyranoside (MeUmbaMan)
were obtained from Sigma Monomannosylated bovineserum albumin (BSA-Man) was kindly provided by MedIm-mune We synthesized methyl 2-deoxy-a-D-mannopyrano-side (Oscarson and Tedebark 1995) Alkyl mannosides weresynthesized through silver triflate-promoted couplings of thecorresponding alcohol with 2346-tetra-O-benzoyl-a-D-mannopyranosyl bromide followed by Zempleacuten deacylationof the obtained protected alkyl mannosides according to theprocedure reported for the octyl and tetradecyl mannosides(Oscarson and Tiden 1993)
Binding studies
Competition experiments using SPR To obtain butyl man-noside-free FimHtr1 the FimH truncate was overproducedwith a carboxy-terminal 6-His tag after transformation into Ecoli C43 (DE3) (Miroux and Walker 1996) and the bacteriagrown in minA minimal medium supplemented with the 20amino acids (40 mg l-1) thiamin (2 mg l-1) and biotin (2 mgl-1) Purification of the His-tagged protein was performedusing Ni-NTA affinity chromatography (Qiagen) in 50 mM Tris-HCl pH 8 and 300 mM NaCl Elution was in the same solutioncomplemented with 500 mM imidazole Subsequently pooledand concentrated FimH fractions were gel filtrated over aSuperdex G75 column (Pharmacia) in 20 mM Tris-HCl pH 8
We used Fab fragments of a monoclonal antibody againstthe mannose-binding site of FimH to determine the solutionaffinity of FimHndashcarbohydrate interactions The monoclonalantibody IC10 was produced by a mouse hybridoma cell lineat Medimmune and its Fab fragments were prepared andpurified The Fabs were covalently immobilized via aminecoupling at 1000 Resonance Units (1000 pg ligand mm-2) inflowcell Fc2 on a CM5 biasensor chip (BIAapplications Hand-book Biacore AB Uppsala Sweden) Flowcell Fc1 was usedas the reference surface and was treated in the same wayas Fc2 except that it was blocked immediately after activationThe immobilization buffer was 100 mM sodium acetatepH 50 with 100 mM NaCl All SPR measurements were per-formed on a Biacore3000trade
The kinetic constants of binding of FimH to the immobilizedantibody were measured with different concentrations ofFimH (2000 nM 1000 nM 500 nM 250 nM 125 nM625 nM 312 nM 156 nM 78 nM) in running buffer [phos-phate-buffered saline (PBS) with 0005 surfactant P20 and3 mM EDTA] on both flowcells Fc1 and Fc2 simultaneouslyat a flow rate of 30 ml min-1 and at 298 K Complete dissoci-ation of FimH was performed with running buffer beforestarting a new binding cycle For all measurements the asso-ciation time was 3 min and the dissociation time was 30 minAll binding cycles were performed in duplicate including azero concentration cycle of FimH (injection of running buffer)All analyses were performed with the BIAeval software ALangmuir binding isotherm with a 11 stoichiometry was fittedto the data from both the association phase and the dissoci-ation phase after subtraction of the reference and zero con-centration data The kinetic constants ka and kd and themaximal binding Rmax were obtained and the dissociationconstant Kd was calculated
The equilibrium dissociation constants Kd of FimHndashsaccha-ride interactions were determined in a competition experi-ment using the same chip as for the previous experiment
452 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
As a control the Fabs of monoclonal antibody IA7 as wellas BSA-Man were also immobilized Samples containing afixed concentration of FimH were used (close to the Kd of theFimHndashantibody interaction) in combination with varying con-centrations of saccharide First 10-fold dilutions of the sac-charide solution were used to determine the concentrationrange for binding of the saccharide to FimH A Langmuirbinding isotherm with a 11 stoichiometry was fitted to thedata using the kinetic constants and Rmax from the foregoingexperiment to obtain the concentrations of FimH that werefree ([FimH]free) to bind the antibody immobilized on the chipSecond the concentration range of the saccharide wasextended and adapted to ensure accurate fitting and theequilibrium binding constant of the FimHndashsaccharide interac-tion was derived from the curve of [FimH]free against concen-tration of saccharide Every measurement was repeated atleast twice including testing different protein batches andwhenever possible also different saccharide stock solutions(typically 200 mM)
Equilibrium binding studies using radioactively labelledmannose Mannose-free FimHtr2 was produced inM9 minimal medium and purified as described above forFimHtr2 Alkyl mannosides were dissolved in double distilledwater pNPaMan and MeUmbaMan (6 mg each) were dis-solved in 20 ml of DMSO and diluted to 20 mM using doubledistilled water Binding experiments were performed at 310 Kusing six different concentrations of [3H]-a-D-mannose (finalconcentrations 435 mM 290 mM 193 mM 129 mM 86 mMand 57 mM) Protein (180 ml) at a concentration of about500 nM was mixed with 20 ml of the radioactive ligand [3H]-a-D-mannose and incubated at 37degC for 20 min To separatefree ligand from bound the mixture was rapidly filtratedthrough a Protran BA 85 cellulose nitrate filter (Schleicherand Schuell) and washed once with 1 ml of ice-cold 1yen PBSFilter-bound radioactivity was measured by scintillation spec-trometry within 24 h Displacement experiments were per-formed using six different concentrations of inhibitor (finalconcentrations in the range 00ndash435 mM) in the presence of435 mM [3H]-a-D-mannose Twenty microlitres of radioactiveligand 20 ml of inhibitor and 160 ml of protein (500 nM) weremixed and the experiments were performed as describedabove All experiments were performed in duplicates and thedata presented are mean values of at least two independentmeasurements For the determination of Kd for a-D-mannosean hyperbolic curve (y = P1x(P2 + x) where P1 is an arbitraryscale factor and P2 = Kd) was fitted to the data For the dis-placement experiments the curve y = P1(P2 + x) where P1
is an arbitrary scale factor and P2 is the concentration of theinhibitor displacing 50 of the labelled ligand [I]05 was usedinstead To calculate the inhibitor dissociation constant (KI)the Cheng and Prusoff equation [KI = [I]05[L]KL + 1) KL is theconstant of dissociation for the ligand] (Cheng and Prusoff1973) was used when both the concentration of the radio-active ligand (L) and the displacing agent (I) are in excessover the protein (LT gtgt PT IT gtgt PT T indicates total concen-tration) For the two strong inhibitors pNPaMan and MeUm-baMan where IT was not in excess over PT the equationof KI = IT[(1 - Y)Y(LTKL) - 1] - PTKLYLT where Y is thefraction of the ligand bound in presence of the inhibitor wasused instead (Horovitz and Levitzki 1987)
Computational docking
Computational docking studies were performed usingAutoDock3 (Morris et al 1998) In AutoDock3 the proteinis represented by a grid of affinity potentials Hence theprotein conformation is not allowed to change during thedocking procedure Flexibility of ligands is taken intoaccount by allowing rotation around flexible torsion anglesThe docked energies are calculated as the sum of the inter-molecular interaction energy and the internal energy of theligand The search for the best interaction energy was car-ried out by means of the Lamarckian genetic algorithmLigand structures were obtained from the Protein DataBank and prepared for docking by adding hydrogens andcharges using AutoDockTools (Sanner et al 1996) Eachsimulation consisted of 100 independent runs with a popu-lation size of 200 500 generations and a maximum of25 000 000 energy evaluations Solutions were rankedbased on their docking energies and similar solutions wereclustered (cluster cut-off rmsd lt 1 Aring) For both ligandsthe top solutions were evaluated using the graphics pro-gram O (Jones et al 1991)
Acknowledgements
We thank Andrej Zavialov and Maringhns Ehrenberg for help withequilibrium binding experiments and Thomas Norberg forassistance with mass spectrometry Anthony DeFusco fromMedimmune provided us with mannosylated bovine serumalbumin We are grateful to Fred Sauer for assistance withdata collection Thanks to Maor Bar-Peled of the CCC at theUniversity of Georgia Athens USA for his suggestion toexpress FimH in minimal medium We acknowledge the useof beamline 19BM of the Structural Biology Center at theAdvanced Photon Source Argonne National LaboratoryChicago IL USA and of beamline ID29 ESRF GrenobleFrance JB is a postdoctoral fellow of the Fonds voor Weten-schappelijk Onderzoek-Vlaanderen which also supportedthe BiaCore3000 instrument (FWONL35) This work was sup-ported by grants from the Swedish Research Council andfrom the Strategic Research Foundation (Structural BiologyNetwork) to SDK
Supplementary material
The following material is available from httpwwwblackwellpublishingcomproductsjournalssuppmatmmimmi4415mmi4415smhtmTable S1 Crystallographic data and refinement statistics
References
Anumula KR (1994) Quantitative determination ofmonosaccharides in glycoproteins by high-performanceliquid chromatography with highly sensitive fluorescencedetection Anal Biochem 220 275ndash283
Bahrani-Mougeot FK Buckles EL Lockatell CV HebelJR Johnson DE Tang CM and Donnenberg MS(2002) Type 1 fimbriae and extracellular polysaccharidesare preeminent uropathogenic Escherichia coli virulence
A novel class of FimH high-affinity ligands 453
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
determinants in the murine urinary tract Mol Microbiol 451079ndash1093
Berglund J and Knight SD (2003) Structural basis forbacterial adhesion in the urinary tract Glycobiol Med 53533ndash52
Blattner FR Plunkett G Bloch CA Perna NT BurlandV Riley M et al (1997) The complete genome sequenceof Escherichia coli K-12 Science 277 1453ndashbullbull
Bloch CA Stocker BA and Orndorff PE (1992) A keyrole for type 1 pili in enterobacterial communicability MolMicrobiol 6 697ndash701
Bork P Holm L and Sander C (1994) The immunoglob-ulin fold ndash structural classification sequence patterns andcommon core J Mol Biol 242 309ndash320
Brinton CC (1959) Non-flagellar appendages of bacteriaNature 183 782ndash786
Bruumlnger AT Adams PD Clore GM DeLano WLGros P Grosse-Kunstleve RW et al (1998) Crystallog-raphy and NMR system a new software suite for macro-molecular structure determination Acta Crystallogr D BiolCrystallogr 54 905ndash921
Buchanan K Falkow S Hull RA and Hull SI (1985)Frequency among Enterobacteriaceae of the DNA-sequences encoding type-1 pili J Bacteriol 162 799ndash803
Buts L Bouckaert J De Genst E Loris R Oscarson SLahmann M et al (2003) The fimbrial adhesin F17-G ofenterotoxigenic Escherichia coli has an immunoglobulin-like lectin domain that binds N-acetylglucosamine MolMicrobiol 49 705ndash715
Cheng Y and Prusoff WH (1973) Relationship betweeninhibition constant (K1) and concentration of inhibitor whichcauses 50 per cent inhibition (I50) of an enzymatic reac-tion Biochem Pharmacol 22 3099ndash3108
Choudhury D Thompson A Stojanoff V LangermannS Pinkner J Hultgren SJ and Knight SD (1999) X-ray structure of the FimCndashFimH chaperonendashadhesin com-plex from uropathogenic Escherichia coli Science 2851061ndash1066
Collaborative computational project n4 (1994) The CCP4suite programs for crystallography Acta Crystallogr D BiolCrystallogr 50 760ndash763
Dodson KW Pinkner JS Rose T Magnusson G Hult-gren SJ and Waksman G (2001) Structural basis of theinteraction of the pyelonephritic Escherichia coli adhesin toits human kidney receptor Cell 105 733ndash743
Eley JG and Triumalashetty P (2001) In vitro assessmentof alkylglycosides as permeability enhancers AAPSPharmSciTech 2 (3) Article 19
Esnouf RM (1997) An extensively modified version of Mol-script that includes greatly enhanced coloring capabilitiesJ Mol Graph Model 15 132ndash134
Firon N Ofek I and Sharon N (1982) Interaction ofmannose-containing oligosaccharides with the fimbriallectin of Escherichia coli Biochem Biophys Res Commun105 1426ndash1432
Firon N Ofek I and Sharon N (1983) Carbohydrate spec-ificity of the surface lectins of Escherichia coli Klebsiellapneumoniae and Salmonella typhimurium Carbohydr Res120 235ndash249
Firon N Ofek I and Sharon N (1984) Carbohydrate-
binding sites of the mannose-specific fimbrial lectins ofEnterobacteria Infect Immun 43 1088ndash1090
Firon N Ashkenazis S Mirelman D Ofek I and SharonN (1987) Aromatic a-glycosides of mannose are powerfulinhibitors of the adherence of type 1 fimbriated Escherichiacoli to yeast and intestinal cells Infect Immun 55 472ndash476
Fukiya S Mizoguchi H Tobe T and Mori H (2004)Extensive genomic diversity in pathogenic Escherichia coliand Shigella strains revealed by comparative genomichybridization microarray J Bacteriol 186 3911ndash3921
Hooton TM and Stamm WE (1997) Diagnosis and treat-ment of uncomplicated urinary tract infection Infect DisClin North Am 11 551ndash581
Horovitz A and Levitzki A (1987) An accurate method fordetermination of receptor ligand and enzyme-inhibitor dis-sociation-constants from displacement curves Proc NatlAcad Sci USA 84 6654ndash6658
Hull RA Gill RE Hsu P Minshaw BH and Falkow S(1981) Construction and expression of recombinant plas-mids encoding type 1 and D-mannose-resistant pili from aurinary tract infection Escherichia coli isolate Infect Immun33 933ndash938
Hultgren SJ Porter TN Schaeffer AJ and Duncan JL(1985) Role of type 1 pili and effects of phase variation onlower urinary tract infections produced by Escherichia coliInfect Immun 50 370ndash377
Hung C-S Bouckaert J Hung DL Pinkner J WinbergC Defusco A et al (2002) Structural basis of tropism ofEscherichia coli to the bladder during urinary tract infec-tion Mol Microbiol 44 903ndash915
Jones TA Zou JY Cowan SW and Kjeldgaard M(1991) Improved methods for building protein models inelectron density maps and the location of errors in thesemodels Acta Crystallogr A 47 110ndash119
Justice SS Hung C-S Theriot JA Fletcher DAAnderson GG Footer MJ and Hulgren SJ (2004)Differentiation and developmental pathways of uropatho-genic Escherichia coli in urinary tract pathogenesis ProcNatl Acad Sci USA 101 1333ndash1338
Kallenius G Mollby R Svenson SB Windberg J Lun-dblud A Svenson S and Cedergen B (1980) The Pk
antigen as receptor for the haemagglutinin of pyelonephri-togenic Escherichia coli FEMS Microbiol Lett 8 297ndash302
Knight SD Berglund J and Choudhury D (2000) Bacte-rial adhesins structural studies reveal chaperone functionand pilus biogenesis Curr Opin Chem Biol 4 653ndash660
Langermann S and Ballou WR (2003) Development of arecombinant FimCH vaccine for urinary tract infectionsAdv Exp Med Biol 539 635ndash653
Langermann S Palaszynski S Barnhart M Auguste GPinkner JS Burlein J et al (1997) Prevention ofmucosal Escherichia coli infection by FimH-adhesin-basedsystemic vaccination Science 276 607ndash611
Langermann S Mollby R Burlein JE Palaszynski SRAuguste CG Defusco A et al (2000) Vaccination withFimH adhesin protects cynomolgus monkeys from coloni-zation and infection by uropathogenic Escherichia coli JInfect Dis 181 774ndash778
Larsson A Ohlsson J Dodson KW Hultgren SJ Nils-son U and Kihlberg J (2003) Quantitative studies of thebinding of the class II PapG adhesin from uropathogenic
454 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
Escherichia coli to oligosaccharides Bioorg Med Chem 112255ndash2261
Leffler H and Svanborg-Eden C (1980) Chemical identifi-cation of a glycosphingolipid receptor for Escherichia coliattaching to human urinary tract epithelial cells and agglu-tinating human erythrocytes FEMS Microbiol Lett 8 127ndash134
Lindhorst TK Kieburg C and Krallmann-Wenzel U(1998) Inhibition of the type 1 fimbriae-mediated adhesionof Escherichia coli to erythrocytes by multiantennary alpha-mannosyl clusters the effect of multivalency Glycoconj J15 605ndash613
Loris R Tielker D Jaeger KE and Wyns L (2003a)Structural basis of carbohydrate recognition by the lectinLecB from Pseudomonas aeruginosa J Mol Biol 331 861ndash870
Loris R Imberty A Beeckmans S Van Driessche ERead JS Bouckaert J et al (2003b) Crystal structureof Pterocarpus angolensis lectin in complex with glucosesucrose and turanose J Biol Chem 278 16297ndash16303
Lund B Lindberg F Marklund BI and Normark S(1987) The PapG protein is the a-D-galactopyranosyl-(1-4)-b-D-galactopyranose-binding adhesin of uropathogenicEscherichia coli Proc Natl Acad Sci USA 84 5898ndash5902
Min G Stolz M Zhou G Liang F Sebbel P StofflerD et al (2002) Localization of uroplakin Ia the urothelialreceptor for bacterial adhesin FimH on the six innerdomains of the 16 nm urothelial plaque particle J Mol Biol317 697ndash706
Miroux B and Walker JE (1996) Over-production of pro-teins in Escherichia coli mutant hosts that allow synthesisof some membrane proteins and globular proteins at highlevels J Mol Biol 260 289ndash298
Morris GM Goodsell DS Halliday RS Huey R HartWE Belew RK and Olson AJ (1998) Automateddocking using a Lamarckian genetic algorithm and anempirical binding free energy function J Comput Chem 191639ndash1662
Mulvey MA (2002) Adhesion and entry of uropathogenicEscherichia coli Cell Microbiol 4 257ndash271
Mulvey MA Lopez-Boado YS Wilson CL Roth RParks WC Heuser J and Hultgren SJ (1998) Induc-tion and evasion of host defenses by type 1-piliated uro-pathogenic Escherichia coli Science 282 1494ndash1497
Murshudov GN Vagin AA and Dodson EJ (1997)Refinement of macromolecular structures by the maxi-mum-likelihood method Acta Crystallogr D Biol Crystallogr53 240ndash255
Nagahori N Lee RT Nishimura S Pageacute D Roy Rand Lee YC (2002) Inhibition of adhesion of type 1 fim-briated Escherichia coli to highly mannosylated ligandsChembiochem 3 836ndash844
Navaza J (2001) Implementation of molecular replacementin AMoRe Acta Crystallogr D Biol Crystallogr 57 1367ndash1372
Neeser JR Koellreutter B and Wuersch P (1986)Oligomannoside-type glycopeptides inhibiting adhesion ofEscherichia coli strains mediated by type 1 pili preparationof potent inhibitors from plant glycoproteins Infect Immun52 428ndash436
Old DC (1972) Inhibition of the interaction between fimbrial
hemagglutinatinins and erythrocytes by D-mannose andother carbohydrates J Gen Microbiol 71 149ndash157
Orndorff PE Devapali A Palestrant S Wyse A EverettML Bollinger RR and Parker W (2004) Immunoglob-ulin-mediated agglutination of and biofilm formation byEscherichia coli K-12 require the type 1 pilus fiber InfectImmun 72 1929ndash1938
Oscarson S and Tedebark U (1995) Syntheses of deoxyanalogues of methyl 36-di-O-a-D-mannopyranosyl-a-D-mannopyranoside for studies of the binding site of con-canavalin A Carbohydr Res 278 271ndash287
Oscarson S and Tiden AK (1993) Syntheses of the octyland tetradecyl glycosides of 36-di-O-alpha-deuterium-mannopyranosyl-alpha-deuterium-mannopyranose and of34-di-O-alpha-deuterium-mannopyranosyl-alpha-deute-rium-mannopyranose ndash a new way for 24-di-O-protectionof mannopyranosides Carbohydr Res 247 323ndash328
Otwinowski Z and Minor W (1997) Processing of X-raydiffraction data collected in oscillation mode MethodsEnzymol 276 307ndash326
Powell HR (1999) The Rossmann Fourier autoindexingalgorithm in MOSFLM Acta Crystallogr D Biol Crystallogr55 1690ndash1695
Pratt LA and Kolter R (1998) Genetic analysis of Escher-ichia coli biofilm formation roles of flagella motility chemo-taxis and type I pili Mol Microbiol 30 285ndash293
Remaut H and Waksman G (2004) Structural biology ofbacterial pathogenesis Curr Opin Struct Biol 14 161ndash170
Ronald AR Nicolle LE Stamm E Krieger J WarrenJ Schaeffer A et al (2001) Urinary tract infection inadults research priorities and strategies Int J AntimicrobAgents 17 343ndash348
Sanner MF Olson AJ and Spehner JC (1996)Reduced surface an efficient way to compute molecularsurfaces Biopolymers 38 305ndash320
Sauer FG Futterer K Pinkner JS Dodson KW Hult-gren SJ and Waksman G (1999) Structural basis ofchaperone function and pilus biogenesis Science 2851058ndash1061
Sauer FG Knight SD Waksman GW and HultgrenSJ (2000) PapD-like chaperones and pilus biogenesisSemin Cell Dev Biol 11 27ndash34
Sauer FG Pinkner JS Waksman G and Hultgren SJ(2002) Chaperone priming of pilus subunits facilitates atopological transition that drives fiber formation Cell 111543ndash551
Schembri MA Hasman H and Klemm P (2000)Expression and purification of the mannose recognitiondomain of the FimH adhesin FEMS Microbiol Lett 188147ndash151
Schembri MA Kjaergaard K and Klemm P (2003) Glo-bal gene expression in Escherichia coli biofilms Mol Micro-biol 48 253ndash267
Schilling JD Mulvey MA Vincent CD Lorenz RGand Hultgren SJ (2001) Bacterial invasion augments epi-thelial cytokine responses to Escherichia coli through alipopolysaccharide-dependent mechanism J Immunol166 1148ndash1155
Silhavy TJ Szmelcman S Boos W and Schwartz M(1975) On the significance of the retention of ligand byprotein Proc Natl Acad Sci USA 72 2120ndash2124
A novel class of FimH high-affinity ligands 455
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
Svanborg C and Godaly G (1997) Bacterial virulence inurinary tract infection Infect Dis Clin North Am 11 513ndash529
Thanassi DG Saulino ET and Hultgren SJ (1998) Thechaperoneusher pathway a major terminal branch of thegeneral secretory pathway Curr Opin Struct Biol 1 223ndash231
Thankavel K Madison B Ikeda T Malaviya R ShahAH Arumugen PM and Abraham SN (1997) Local-ization of a domain in the FimH adhesin of Escherichia colitype 1 fimbriae capable of receptor recognition and use ofa domain-specific antibody to confer protection againstexperimental urinary tract infection J Clin Invest 1001123ndash1126
Vagin A and Teplyakov A (1997) MOLREP an automatedprogram for molecular replacement J Appl Crystallogr 301022ndash1025
Wu XR Sun TT and Medina JJ (1996) In vitro bindingof type 1-fimbriated Escherichia coli to uroplakins 1a and
1b relation to urinary tract infections Proc Natl Acad SciUSA 93 9630ndash9635
Xia Y Gally D Forsman-Semb K and Uhlin BE (2000)Regulatory cross-talk between adhesin operons in Escher-ichia coli inhibition of type 1 fimbriae expression by thePapB protein EMBO J 19 1450ndash1457
Zafriri D Ofek I Adar R Pocino M and Sharon N(1989) Inhibitory activity of cranberry juice on adherenceof type-1 and type-P fimbriated Escherichia coli to eukary-otic cells Antimicrob Agents Chemother 33 92ndash98
Zavialov AV Berglund J Pudney AF Fooks LJ Ibra-him TM MacIntyre S and Knight SD (2003) Structureand biogenesis of the capsular F1 antigen from Yersiniapestis preserved folding energy drives fiber formation Cell113 587ndash596
Zhou G Mo WJ Sebbel P Min G Neubert TA Glock-shuber R et al (2001) Uroplakin Ia is the urothelial recep-tor for uropathogenic Escherichia coli evidence from invitro FimH binding J Cell Sci 114 4095ndash4103

444 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
initial FondashFc and 2FondashFc Fourier maps inside the mannose-binding pocket of both FimHtr1 and FimHtr2 This densityprecisely defined a-D-mannose with an additional elon-gated density shaped like a creased tail extending out-wards from the mannose O1 and towards tyrosines 48and 137 which make up part of the hydrophobic rimaround the mannose-binding pocket (Fig 1B) (Hunget al 2002) Further refinement strongly suggested thata butyl a-D-mannoside was bound in the mannose-bindingsite (Fig 1B and C) For FimHtr1 also a change in theconformation of the Tyr48 side-chain compared with theone from the model FimH used for molecular replacementwas immediately apparent As described previously (Hunget al 2002) the mannose ring makes 10 direct hydrogenbonds to the side-chains of residues Asp54 Gln133Asn135 and Asp140 and to the main chain of Phe1 andAsp47 and indirect water-mediated hydrogen bonds viaO2 to the side-chain of Glu133 and to the main chainoxygen of Phe1 and Gly14 The alpha-anomeric hydroxylgroup O1 of mannose is involved in a hydrogen-bondingwater network with the Asn138 and Asp140 side-chainsthrough a water molecule (wat 2 Fig 1B) that is con-
served between the two truncate structures The positionsof the other waters participating in this network dependon the crystal packing of FimHtr1 and FimHtr2
Mass spectrometric analysis of the bound ligand
To corroborate the nature of the ligand identified in thecrystal structures the complex of FimHtr1 and its pre-sumed butyl a-D-mannoside was analysed using massspectrometry The FimHtr1 protein sample used for crys-tallization was subjected to peracetylation and phase sep-aration The peracetylated ligand retrieved in the organicphase was analysed by nanoLC-online-ESI-MS The per-acetylated ligand eluted as a single peak and was regis-tered in its sodiated form at mz 427 corresponding tobutyl hexose (Fig 2A) No signals corresponding to otherpotential alkyl mannose species like ethyl hexose (mz399) propyl hexose (mz 413) or pentyl hexose (mz 441)could be observed indicating a homogeneous ligand witha four-carbon atoms alkyl chain Besides the sodiumadduct of the ligand a major signal was observed arisingfrom the loss of butanol (mz 331 Fig 2B) Cleavage at
Fig 1 Stereo views of FimHA Three-dimensional structure of the FimH lectin domain is an elongated 11-stranded b-barrel with an immunoglobulin fold Numbering is according to conventions established for antibody domains (Bork et al 1994) and for PapD (Sauer et al 1999) and loop identifications according to previous descriptions (reference strand numbers A1 = 1 Acent = 2 A2 = 3 B1 = 4 B2 = 4cent C = 5 D1 = 6 Dcent = 7 D2 = 8 E = 9 F = 10 G = 11) (Choudhury et al 1999 Hung et al 2002) The disulphide bridge between Cys3 and Cys44 is coloured cyanB Focus on the mannose-binding pocket in the same orientation as (A) showing the hydrophobic contacts (within 46 Aring orange) with butyl a-D-mannoside and the water network (salmon pink) present at O1C Surface presentation (wheat yellow) of the different alkyl conformations of the butyl a-D-mannoside in the crystal structures of FimH truncates 1 and 2 FimHtr1 butyl group (cyan) FimHtr1 Tyr48 (marine blue) FimHtr2 butyl group and Tyr48 (salmon pink) Tyr137 (violet) Ile52 (pink)
A B
C
A novel class of FimH high-affinity ligands 445
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
the anomeric carbon atom was often combined with elim-ination of one or more acetic acid molecules (massdecrease of 60 Da) or acetyl groups (mass decrease of42 Da) These eliminations gave rise to the major ions atmz 127 and mz 169 with a proposed aromatic ring struc-ture as well as to the ion at mz 109 with a proposedquinone-like ring structure Thus the mass spectrometricdata support the butyl hexose nature of the FimH ligandFimH was further characterized by carbohydrate constit-uent analysis (data not shown) which revealed only man-nose Mannose could only be detected after hydrolysisindicating that free mannose is not associated with theFimH truncate
Origin of butyl a-D-mannoside
As no sugar had been advertently added to the FimHtr
protein samples we hypothesized that the butyl manno-side ligand had been picked up from the LuriandashBertani(LB) medium used to grow bacteria during expression ofthe protein To test this FimHtr2 was expressed in bacteriagrown in minimal M9 medium Electro-spray ionizationmass spectrometry (ESI-MS) was performed on purifiedFimHtr2 samples from both types of media A clear peakat 259 Da could be detected (Fig 3A) with proteinexpressed in bacteria grown in LB medium correspondingto the molecular weight of butyl mannoside (236 Da) plus
a sodium ion (23 Da) As a control butyl mannoside wassynthesized and analysed in the same manner Again apeak at 259 Da was observed (Fig 3A inset) When ESI-MS was run on protein expressed in bacteria grown inminimal medium no such peak could be detected(Fig 3B) indicating that the ligand was not present andthat it indeed originated from the LB medium
Interactions of the butyl moiety with FimH
The butyl moiety of butyl a-D-mannoside extends out ofthe mannose-binding pocket towards Tyr48 and Tyr137making van der Waals contacts to both tyrosine rings andIle52 (Fig 1B and C) These residues are part of two loops(b4ndashb5 and b10ndashb11) that form the higher rear end of ahydrophobic collar around the binding site (Fig 1A andB) In both subunits of the FimHtr2 structure the Tyr48 andTyr137 side-chains are in the same almost parallel ori-entation as in the FimCFimH structure forming a gate-like structure at the back of the binding site (Fig 1C) Theclosest hydrophobic interactions (31ndash36 Aring) with the alkylchain are made between Tyr48 and the two non-terminalatoms of the butyl In the FimHtr1 structure a crystal con-tact to Val155 of a neighbouring molecule prevents theparallel orientation of the Tyr48 ring which instead ispacked edge-to-face with Tyr137 (Fig 1B) The reposition-ing of Tyr48 is accompanied by a change in the confor-
A
B
Fig 2 Mass spectrophotometric characteriza-tion of the butyl mannosideA Base peak chromatogram (BPC mass range mz 400ndash550) and extracted ion chromato-grams (EIC) of mz 427 and 331 obtained by nano-LC-ESI-MS of the derivatized FimH-ligand purified by peracetylation and organic-solvent extractionB ESI-IT mass spectrometric characterization of the peak species marked by a horizontal bar in (A) Ions are marked with mz-values and proposed schematic structures which is often only one out of several isobaric ion species Fragmentations already occurred in the source Ac acetyl
446 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
mation of the butyl chain of the ligand with the butyl chainin a more extended conformation than in the FimHtr2 struc-ture and with the four carbon atoms nearly in one plane(Fig 1B and C) The new conformations of Tyr48 and thebutyl moiety result in an almost parallel stacking of thearomatic ring of Tyr48 onto the butyl plane with the closesthydrophobic interactions (37 Aring) involving the two terminalcarbon atoms of the butyl (Fig 1B)
Affinity measurements for alkyl mannosides
The butyl mannoside ligand remained bound to FimHtr
despite extensive dialysis From a study of complex for-mation of monoclonal antibodies IC10 and IA7 withFimHtr1 it could be estimated that about 30 of the FimHtruncate retained its ligand after dialysis for 12 daysagainst 50 mM MES buffer at pH 65 100 mM NaCldespite daily refreshment of buffer (data not shown)Therefore to enable measurements of dissociation con-stants of FimH for mannosides FimH was produced inminimal medium Butyl a-D-mannoside binding is 15 timesimproved (Kd = 151 nM Fig 4A) compared with mannosebinding The equilibrium binding technique using [3H]-mannose led to a similar value (Kd = 41 mM Fig 4B) for
mannose and an almost identical value for butyl a-D-mannoside (Kd = 150 nM Fig 4C) To investigate theeffect of sequential addition of methyl groups to the O1oxygen of D-mannose a series of alkyl mannosides weresynthesized and the dissociation constants determinedusing the [3H]-mannose displacement assay as well as theSPR competition assay (Table 1) There is a linear corre-lation between the binding free energy as calculated fromthe measured dissociation constants and the number ofmethyl groups (between 1 and 7) in the alkyl mannosidewith each additional methyl group contributing on average-06 kcal mol-1 of binding energy (Fig 5)
Affinity measurements and modelling for aryl mannosides
Aromatically substituted mannosides have been reportedto be particularly potent inhibitors of FimH-mediated bac-terial adhesion (Firon et al 1987 Nagahori et al 2002)In accordance with the earlier results both pNPaMan andMeUmbaMan bind very tightly to FimH (Table 1) Despiteextensive trials no X-ray structures were obtained forcomplexes with aromatically substituted mannosides Inorder to model binding docking to both of the truncatestructures was carried out In all cases the mannose ring
Fig 3 Mass spectrometric analysis of the origin of the butyl mannosideA LB medium-produced FimH truncate The inset shows the mass spectrum of the synthesized butyl a-D-mannosideB Minimal media-produced FimH truncate
A novel class of FimH high-affinity ligands 447
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
docks in the binding pocket in a conformation very closeto the crystallographically observed mannose conforma-tion (Fig 6) while the aromatic substituent interacts withthe tyrosine gate The different conformation of Tyr48 in
our two FimH truncate structures leads to different pre-dicted interactions between the aromatic aglycon andTyr48 and Tyr137 Docking to the FimHtr2 structureresulted in somewhat tighter interactions between thetyrosine gate and the aglycon and hence in somewhatlower docking energies Calculated FimHtr2 docking ener-gies of Edoc = -104 kcal mol-1 for pNPaMan (Kd = 46 nM)and Edoc = -109 kcal mol-1 for MeUmbaMan (Kd = 20 nM)are in good agreement with the experimental binding data(Table 1) The docked umbelliferyl ring system is inserteddeep into the tyrosine gate in the FimHtr2 conformationwith the carbonyl group within hydrogen bonding distanceto Thr51 at the back of the gate (Fig 6A) Docking to theFimHtr1 structure instead stacks the umbelliferyl ring face-to-face with the Tyr48 ring (not shown) The phenyl ring ofpNPaMan docks similarly to both our structures It doesnot quite reach in between the tyrosine side-chains butinstead hydrogen bonds with its nitro group to the hydroxyloxygen of Tyr137 (Fig 6B)
Discussion
Most lectins bind naturally occurring carbohydrates withlow affinities often in the millimolar range for monosac-charides and micromolar range for oligosaccharides Thiswas recently exemplified to also hold true for theadhesins of F17 fimbriae (Kd = 12 mM for the F17a-G-N-acetylglucosamine interaction Buts et al 2003) and Ppili (Kd = 84 mM for the PapGII-globotetraoside interactionLarsson et al 2003) The recent crystal structure of FimHin complex with mannose illustrated that FimH almostcompletely envelops the monosaccharide in a deeppocket and involves it in an intense hydrogen bondingnetwork suggestive of an unusual lectin exerting highaffinities (Hung et al 2002)
Fig 4 FimH binding experimentsA Surface plasmon resonance competition experiment of FimHtr1 binding to monoclonal antibody IC10 Fab fragments with butyl a-D-mannosideB Equilibrium binding of [3H]-mannose to FimHtr2C Equilibrium displacement measurements of butyl a-D-mannoside to the FimHtr2 [3H]-mannose complex
log[Butyl a-D-mannoside] (M)
cpm
cpm
Butyl a-D-mannoside (mM)
Mannose (mM)
A
B
C
ndash64
ndash66
ndash68
ndash70
ndash72
ndash74
ndash76
ndash78
ndash80
ndash82
ndash8 ndash7 ndash6 ndash5 ndash4 ndash3
log
[Fim
H]
M
Fig 5 Linear free energy relationship for the alkyl chain on mannose A linear fit was made using 1 up to 7 methyl groups with the program MICROCAL ORIGIN 50
Number of carbon atoms0 2 4 6 8
ndash75
ndash80
ndash90
ndash95
ndash100
ndash105
ndash110
ndash115
ndash85
D G
(kc
al m
olndash1
)
slopecorrelation
ndash064ndash099
+ndash 003+ndash 018
448 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
The measured affinity confirm the predictions that FimHbinds the monosacharide mannose exceptionally well(Kd = 23ndash41 mM) To investigate the contribution of thedifferent hydroxyl groups of mannose in the interactionwith FimH the binding of a series of hexoses to butylmannoside-free FimH was examined using SPR (Table 2)The effect of deoxygenation of the O2 hydroxyl group(methyl 2-deoxy-a-D-mannopyranoside) is clearly lessdestructive (100-fold reduction) than its equatorial positionas in glucose O2 is the only hydroxyl group of mannosethat interacts indirectly with the FimH pocket via a con-served water molecule (Hung et al 2002) Glucose onlydisplays a weak affinity for FimH about 4000 times lowerthan methyl a-D-mannoside Repositioning of both the O2and O4 hydroxyl groups as in galactose abolishes sugarbinding by FimH Similar to these results the inhibitoryconcentrations for bacterial adherence of methyl a-D-glucoside and galactose derivatives have been estimatedto be respectively 104-fold and 105-fold weaker than formethyl a-D-mannoside (Nagahori et al 2002)
In stark contrast with the other monosaccharides fruc-tose has a binding affinity only about 15-fold less thanmannose for FimH (Table 2) The decrease in affinity for
fructose relative to mannose agrees with studies of inhi-bition of adherence of type 1 piliated E coli by fructosein which fructose was found to be the active compound infruit juices (present at about 5) (Zafriri et al 1989)Fructose binding to FimH is stronger than the binding ofthe physiological globotetraoside receptor to PapGII theother well-characterized adhesin of UPEC strains (Lars-son et al 2003) The pyranose form of fructose is verywell superimposable onto mannose in the FimH bindingsite where similar interactions are established with FimHas for mannose except for the loss of the anomeric O1and an extra hydroxyl group (O2) on the C5 position ofmannose Probably however both changes of fructopyr-anoside compared with mannopyranoside could be easilyaccommodated by FimH The mannose O1 does notmake direct interactions within 45 Aring of the FimH bindingpocket (Fig 1B) The extra O2 group of fructose would notclash or interfere significantly with the binding pocket butit would make a 27 Aring van der Waals contact with the Ile52side-chain under the condition that the Ile52 side-chaindoes not move The crystal structure of the Pseudomonasaeruginosa lectin LecB also showed that fructopyranosebut not fructofuranose can substitute for mannose (Loris
Fig 6 Stereo diagrams of docked MeUm-baMan (A) and pNPaMan (B) in the FimH bind-ing site The crystallographically determined position of a-D-mannose is shown in orange for comparison
A novel class of FimH high-affinity ligands 449
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
et al 2003a) Hydrogen bonds of the more specificfucose complex are replaced by van der Waals interac-tions involving a hydroxyl group in the complexes with thelower-affinity binders mannose and fructopyranose as forthe proposed fructopyranoside binding to FimH
The fructose-containing disaccharides sucrose andturanose do not have an inhibitory effect and bind withaffinities similar to glucose Sucrose is a non-reducingsugar that has fructose locked in its furanose form Tura-nose on the other hand allows the interconversion of b-D-fructofuranose and b-D-fructopyranose eg both formsoccur in complex with a plant lectin from Pterocarpusangolensis (Loris et al 2003b) It is predictable howeverfrom the crystal structures that the glycosidic linkage ofglucose with C3 of fructose equivalent in position to C4of mannose would be disallowed in the tight FimH pocketSucrose and turanose thus most probably interact with theFimH mannose binding pocket via their glucose moietyresulting in similar low affinities
Our mass spectrometry and chemical analyses con-firms the crystallographic identification of the ligand boundto the FimH lectin domains as butyl a-D-mannoside(Figs 1B and 2) The mannose ring of the butyl a-D-mannoside coincides with the mannose bound to theFimCFimH complex (Hung et al 2002) and makes thesame interactions The four-carbon alkyl tail exits towardsthe back wall (loop b4ndashb5 and b10ndashb11) of the pocket(Fig 1A and B) making hydrophobic contacts to Tyr48Tyr137 and Ile52 The mass spectrometry data also showthat the bound butyl a-D-mannoside is present in LBmedium but absent from minimal medium Hence thesource of the alkyl mannoside is possibly the yeast extractused in preparation of LB medium Although we do notwant to speculate about the significance of this findingalkyl mannoside production by yeast has not beenreported and might warrant investigation
The butyl mannoside was retained by FimH despiteextensive dialysis The phenomenon of ligand retention byprotein was described as early as 1975 for periplasmicbinding proteins (Silhavy et al 1975) Applied to dialysisof butyl mannoside away from FimH grown in richmedium half-lives of up to 30 h can be estimated whichis far beyond the duration of a normal buffer exchangedialysis experiment In the present case the retainedligand does not appear to seriously affect [3H]-mannosedisplacement equilibrium binding experiments as wecould detect no significant differences in dissociation con-stants obtained using FimHtr2 produced in LB or minimalmedia (data not shown) In contrast proper SPR bindingcurves could not be obtained when butyl mannoside occu-pied the binding site Hence the displacement techniquemay be of more general use to obtain binding constantsfor proteins that tend to inadvertently retain ligands whichis a common trait of lectins
Surface plasmon resonance as well as equilibrium bind-ing and displacement measurements using [3H]-mannosewere carried out to obtain dissociation constants for a-D-mannose and a linear series of alkyl-substituted manno-sides to FimH (Table 1) Both equilibrium binding experi-ments show a very good agreement in their measurementof the carbohydrate affinity of the adhesin FimH bindsbutyl a-D-mannoside 15ndash30 times stronger than a-D-mannose The importance of the added hydrophobiccontacts is clearly demonstrated by the binding of alkylmannosides (Table 1) The best binding alkyl mannosidesbind a few hundred times stronger than mannose equiv-alent in affinity to the most tightly binding aromaticallysubstituted mannosides for FimH (Firon et al 1987) andof mannose dendrimers for type 1 piliated E coli (Fironet al 1984 1987 Nagahori et al 2002)
Interestingly the butyl moiety takes on significantlydifferent conformations in FimHtr1 and FimHtr2 (Fig 1C)indicating that intrinsic conformational flexibility in theaglycon is allowed Indeed in spite of the larger flexibilityexpected for longer alkyl mannosides the binding energydecreases linearly with increasing length of the alkyl chainbetween one and seven carbon atoms (Fig 5) The affinityreaches its maximum for mannose with a seven-carbonalkyl and SPR measurements suggest that the affinitydecreases from eight carbon atoms on Evidently theeighth carbon atom goes beyond the hydrophobic interac-tion surface and protein edge which is penalized with adecrease in affinity We propose that the affinity increaseresults from an increased interaction surface with thehydrophobic gateway Alkyl aglycons fit well in the tyrosinegate and as observed in the two crystal structures in atleast two different conformations that depend on theconformation of Tyr48 This flexibility may indicate thataglycons can interact rather non-specifically with thehydrophobic gateway nevertheless resulting in a 100- to1000-fold increase in affinity
FimH binding to ligands other than just mannose ispossibly of importance in terms of the different environ-ments that E coli will encounter ndash the urinary tract gas-trointestinal tract and in biofilms Alkyl mannosides havenot been previously recognized as strong binders to FimHAs they are easily synthesized and highly soluble in waterthey may be interesting as potential blocking agents forFimH-mediated adhesion Moreover they have beenshown to be good drug vehicles themselves acting aspermeability enhancers on tight junctions (Eley and Triu-malashetty 2001) As such they might be able to pene-trate into deeper cell layers of the epithelium to alsoprevent those bacteria that form a quiescent reservoirfrom causing recurrent infections (Mulvey et al 1998Justice et al 2004)
On the other hand the two aromatically substitutedmannosides pNPaMan and MeUmbaMan are known to
450 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
be particularly potent inhibitors of FimH-mediated bacte-rial adhesion (Firon et al 1987 Nagahori et al 2002)Based on adhesion inhibition studies using type 1 piliatedE coli and guinea pig ileal epithelial cells the relativeinhibitory potential of pNPaMan and MeUmbaMan havebeen estimated to be about 70 and 600 times strongerrespectively than that of methyl a-D-mannose (Fironet al 1987) Our measurements indicate an affinity ofFimH for pNPaMan and MeUmbaMan between 50 and90 times and between 110 and 200 times higher respec-tively than for methyl a-D-mannose in good agreementwith the adhesion inhibition results (Firon et al 1987)Dissociation constants predicted using AutoDock3 (Morriset al 1998) are similar to experimental values Thecomputational docking predicts that tight binding of thearomatically substituted mannosides pNPaMan andMeUmbaMan is accomplished similarly as for butyl a-D-mannoside with the aglycon interacting with the tyrosinegate
This is the first systematic study providing molecularbinding data for the FimH adhesin the prerequisite forcolonization by UPEC (Langermann and Ballou 2003)We also show crystallographic evidence that the lectinor receptor binding domain of FimH is a stable andfunctional entity Its structure is virtually identical to thesame domain in the FimCFimH complex indicating thata truncate corresponding to the lectin domain could pro-vide a good alternative for the development of a UTIvaccine (Langermann et al 1997 Langermann and Bal-lou 2003) However progress of this interesting UTI vac-cine candidate into clinical trials has not been reportedThe binding affinities of FimH at the molecular levelrelate convincingly well with the inhibitory concentrationsused in previous bacterial adhesion assays thus endors-ing the direct and conclusive correlation of FimH withbacterial adherence Alkyl mannosides have nanomolaraffinities for FimH which may overcome the significantobstacle of low affinity generally encountered in attemptsto develop carbohydrate-based drugs which interferewith microbial attachment to host cell surface glycocon-jugates New anti-adhesive drugs could be based onthese leads to allow specific interaction and targeting tothe urinary tract in order to prevent anticipated andrecurrent infections
Experimental procedures
Strains and constructs
FimH proteins from two different E coli strains the UPECstrain J96 (Hull et al 1981) and the E coli K-12 strainMG1655 (Blattner et al 1997) were used in this study TheFimH adhesins encoded by these two E coli strains areidentical FimHtr1 (residues 1ndash158 of UPEC J96 FimH) wasexpressed from plasmid pMMB91 transformed into E coli
C600 cells FimHtr2 (residues 1ndash158 of E coli MG1655FimH + carboxy-terminal 6-His tag) was expressed fromplasmid pPKL241 (Schembri et al 2000) transformed into Ecoli host strain HB101
Purification and crystallization
FimHtr1 Bacteria were grown in LB medium at 37infinC inducedat OD600 = 08 with 1 mM IPTG and harvested 2ndash3 h laterThe cells were resuspended in 20 mM Tris-HCl pH 80 5 mMEDTA and 30 sucrose and the periplasmic content wasextracted by osmotic shock upon 10-fold dilution with 20 mMTris-HCl pH 80 Purification of FimHtr1 was performed by ionexchange chromatography with MonoS at pH 45 followed byMonoQ (Amersham Biosciences) at pH 8 FimHtr1 was crys-tallized using vapour diffusion in hanging drops composed of3 ml of the precipitant solution (01 M Hepes pH 75 12PEG 6000 and 5 vv methyl-24-pentanediol) and 3 ml of12 mg ml-1 protein solution in 20 mM Tris-HCl pH 80 with150 mM NaCl
FimHtr2 Cells were grown in LB medium containing50 mg ml-1 ampicillin at 37infinC At OD600 = 06 the cells wereinduced with 5 mM IPTG Cells were harvested 2 h afterinduction and the periplasmic content was extracted Theprotein was purified on a Pharmacia HiTrap Chelating HP5 ml column (Pharmacia) loaded with Ni-chloride and elutedwith a sharp 0ndash500 mM imidazole gradient Fractions con-taining FimHtr2 were pooled dialysed against 50 mM sodiumacetate pH 525 and loaded onto a Mono S HR 8 ml columnThe protein was eluted with a 0ndash500 mM NaCl gradient anddialysed overnight against 20 mM MES pH 65 before crys-tallization Crystallization was performed at room tempera-ture by the vapour diffusion method with hanging dropsmade by mixing of 2 ml of 15 mg ml-1 protein solution with 2 mlof the precipitant solution (65 methyl-24-pentanediol100 mM cacodylate pH 65) Microseeding was employed tooptimize crystal quality and reproducibility Crystals suitablefor X-ray data collection generally appeared within 48 h aftermicroseeding
Structure determination
X-ray data for FimHtr1 were collected from a single crystalto 17 Aring resolution at the Advanced Photon SourceArgonne National Laboratory USA The crystal was flash-cooled to 100 K in the precipitant solution complementedwith 25 glycerol The data were processed and scaledusing the HKL2000 package (Otwinowski and Minor 1997)(Table S1 in Supplementary material) Data for FimHtr2
were collected from a single crystal under liquid-nitrogencryo conditions at 100 K at beamline ID29 ESRF Greno-ble France to 21 Aring Bragg spacing FimHtr2 data were pro-cessed and reduced using MOSFLM (Powell 1999) andscaled using SCALA (Collaborative computational project1994) (Table S1 in Supplementary material) The structuresof FimHtr1 and FimHtr2 were solved by molecular replace-ment using Amore (Navaza 2001) and MOLREP (Vaginand Teplyakov 1997) respectively with the FimH lectindomain from the FimCFimH complex (Choudhury et al
A novel class of FimH high-affinity ligands 451
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
1999) (PDB id 1QUN) as the search model Positional bulksolvent and isotropic temperature factor refinement wasperformed using CNS (Bruumlnger et al 1998) and REFMAC5(Murshudov et al 1997) respectively Model adjustmentand inspection were performed using Turbo-Frodo and O(Jones et al 1991) graphics programs respectively Refine-ment statistics are given in Table S1 Molecular graphicsillustrations were generated with Bobscript (Esnouf 1997)and Pymol 090 (DeLano Scientific LLC 1998ndash2003 httppymolsourceforgenet)
Mass spectrometry
The purified FimHtr1 protein (300 mg) that had been used forcrystallization was freeze-dried taken up in 100 ml of pyridineand 100 ml of acetic anhydride and incubated overnight atroom temperature in the dark for peracetylation The samplewas dried under nitrogen and taken up in chloroform (400 ml)The chloroform phase was extracted six times with waterdried and taken up in 200 ml of 50 aqueous methanol A1 ml aliquot was applied to a nano-scale RP column(75 mm yen 100 mm PepMap DionexLC Packings) using anUltimate nano-LC system (LC Packings) which was equili-brated with eluent A (H2Oacetonitrile 955 vv containing01 formic acid) at a flow rate of 100 nl min-1 After injectingthe sample the column was run isocratically for 5 min fol-lowed by a linear gradient to 100 eluent B (H2Oacetonitrile2080 vv containing 01 formic acid) in 20 min and a finalwash with 100 B for 5 min The system was directly coupledto an Esquire 3000 ESI-IT-MS (Bruker Daltonik) equippedwith an online nanospray source operating in the positive-ionmode For electrospray (900ndash1200 V) capillaries (360 mmOD 20 mm ID with 10 mm opening) from New Objective wereused The solvent was evaporated at 150infinC with a nitrogenstream of 6 l min-1
To verify the suspected source of the bound butyl manno-side mass spectrometry was performed on samples ofFimHtr2 grown in either LB or minimal M9 medium Proteinsamples were dialysed against water overnight before massspectrometry in order to reduce background from the Trisbuffer Samples were diluted to a concentration in the submi-cromolar range Electrospray mass spectra were recorded indirect inlet mode (50 aqueous methanol solutions) on aBruker Daltonics Esquire-LC (Billerica)
Carbohydrate constituent analysis
Samples were hydrolysed in 100 ml of 4 M aqueous trifluoro-acetic acid (Sigma) at 100infinC for 4 h and dried under astream of nitrogen Monosaccharides were converted intotheir anthranilic acid derivatives by reductive aminationresolved by RP-HPLC and detected by fluorescence (Anu-mula 1994)
Carbohydrates for binding studies
[3H]-mannose was obtained from Amersham Methyl a-D-mannoside glucose galactose fructose sucrose turanosep-nitrophenyl a-D-mannopyranoside (pNPaMan) and 4-methylumbelliferyl a-D-mannopyranoside (MeUmbaMan)
were obtained from Sigma Monomannosylated bovineserum albumin (BSA-Man) was kindly provided by MedIm-mune We synthesized methyl 2-deoxy-a-D-mannopyrano-side (Oscarson and Tedebark 1995) Alkyl mannosides weresynthesized through silver triflate-promoted couplings of thecorresponding alcohol with 2346-tetra-O-benzoyl-a-D-mannopyranosyl bromide followed by Zempleacuten deacylationof the obtained protected alkyl mannosides according to theprocedure reported for the octyl and tetradecyl mannosides(Oscarson and Tiden 1993)
Binding studies
Competition experiments using SPR To obtain butyl man-noside-free FimHtr1 the FimH truncate was overproducedwith a carboxy-terminal 6-His tag after transformation into Ecoli C43 (DE3) (Miroux and Walker 1996) and the bacteriagrown in minA minimal medium supplemented with the 20amino acids (40 mg l-1) thiamin (2 mg l-1) and biotin (2 mgl-1) Purification of the His-tagged protein was performedusing Ni-NTA affinity chromatography (Qiagen) in 50 mM Tris-HCl pH 8 and 300 mM NaCl Elution was in the same solutioncomplemented with 500 mM imidazole Subsequently pooledand concentrated FimH fractions were gel filtrated over aSuperdex G75 column (Pharmacia) in 20 mM Tris-HCl pH 8
We used Fab fragments of a monoclonal antibody againstthe mannose-binding site of FimH to determine the solutionaffinity of FimHndashcarbohydrate interactions The monoclonalantibody IC10 was produced by a mouse hybridoma cell lineat Medimmune and its Fab fragments were prepared andpurified The Fabs were covalently immobilized via aminecoupling at 1000 Resonance Units (1000 pg ligand mm-2) inflowcell Fc2 on a CM5 biasensor chip (BIAapplications Hand-book Biacore AB Uppsala Sweden) Flowcell Fc1 was usedas the reference surface and was treated in the same wayas Fc2 except that it was blocked immediately after activationThe immobilization buffer was 100 mM sodium acetatepH 50 with 100 mM NaCl All SPR measurements were per-formed on a Biacore3000trade
The kinetic constants of binding of FimH to the immobilizedantibody were measured with different concentrations ofFimH (2000 nM 1000 nM 500 nM 250 nM 125 nM625 nM 312 nM 156 nM 78 nM) in running buffer [phos-phate-buffered saline (PBS) with 0005 surfactant P20 and3 mM EDTA] on both flowcells Fc1 and Fc2 simultaneouslyat a flow rate of 30 ml min-1 and at 298 K Complete dissoci-ation of FimH was performed with running buffer beforestarting a new binding cycle For all measurements the asso-ciation time was 3 min and the dissociation time was 30 minAll binding cycles were performed in duplicate including azero concentration cycle of FimH (injection of running buffer)All analyses were performed with the BIAeval software ALangmuir binding isotherm with a 11 stoichiometry was fittedto the data from both the association phase and the dissoci-ation phase after subtraction of the reference and zero con-centration data The kinetic constants ka and kd and themaximal binding Rmax were obtained and the dissociationconstant Kd was calculated
The equilibrium dissociation constants Kd of FimHndashsaccha-ride interactions were determined in a competition experi-ment using the same chip as for the previous experiment
452 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
As a control the Fabs of monoclonal antibody IA7 as wellas BSA-Man were also immobilized Samples containing afixed concentration of FimH were used (close to the Kd of theFimHndashantibody interaction) in combination with varying con-centrations of saccharide First 10-fold dilutions of the sac-charide solution were used to determine the concentrationrange for binding of the saccharide to FimH A Langmuirbinding isotherm with a 11 stoichiometry was fitted to thedata using the kinetic constants and Rmax from the foregoingexperiment to obtain the concentrations of FimH that werefree ([FimH]free) to bind the antibody immobilized on the chipSecond the concentration range of the saccharide wasextended and adapted to ensure accurate fitting and theequilibrium binding constant of the FimHndashsaccharide interac-tion was derived from the curve of [FimH]free against concen-tration of saccharide Every measurement was repeated atleast twice including testing different protein batches andwhenever possible also different saccharide stock solutions(typically 200 mM)
Equilibrium binding studies using radioactively labelledmannose Mannose-free FimHtr2 was produced inM9 minimal medium and purified as described above forFimHtr2 Alkyl mannosides were dissolved in double distilledwater pNPaMan and MeUmbaMan (6 mg each) were dis-solved in 20 ml of DMSO and diluted to 20 mM using doubledistilled water Binding experiments were performed at 310 Kusing six different concentrations of [3H]-a-D-mannose (finalconcentrations 435 mM 290 mM 193 mM 129 mM 86 mMand 57 mM) Protein (180 ml) at a concentration of about500 nM was mixed with 20 ml of the radioactive ligand [3H]-a-D-mannose and incubated at 37degC for 20 min To separatefree ligand from bound the mixture was rapidly filtratedthrough a Protran BA 85 cellulose nitrate filter (Schleicherand Schuell) and washed once with 1 ml of ice-cold 1yen PBSFilter-bound radioactivity was measured by scintillation spec-trometry within 24 h Displacement experiments were per-formed using six different concentrations of inhibitor (finalconcentrations in the range 00ndash435 mM) in the presence of435 mM [3H]-a-D-mannose Twenty microlitres of radioactiveligand 20 ml of inhibitor and 160 ml of protein (500 nM) weremixed and the experiments were performed as describedabove All experiments were performed in duplicates and thedata presented are mean values of at least two independentmeasurements For the determination of Kd for a-D-mannosean hyperbolic curve (y = P1x(P2 + x) where P1 is an arbitraryscale factor and P2 = Kd) was fitted to the data For the dis-placement experiments the curve y = P1(P2 + x) where P1
is an arbitrary scale factor and P2 is the concentration of theinhibitor displacing 50 of the labelled ligand [I]05 was usedinstead To calculate the inhibitor dissociation constant (KI)the Cheng and Prusoff equation [KI = [I]05[L]KL + 1) KL is theconstant of dissociation for the ligand] (Cheng and Prusoff1973) was used when both the concentration of the radio-active ligand (L) and the displacing agent (I) are in excessover the protein (LT gtgt PT IT gtgt PT T indicates total concen-tration) For the two strong inhibitors pNPaMan and MeUm-baMan where IT was not in excess over PT the equationof KI = IT[(1 - Y)Y(LTKL) - 1] - PTKLYLT where Y is thefraction of the ligand bound in presence of the inhibitor wasused instead (Horovitz and Levitzki 1987)
Computational docking
Computational docking studies were performed usingAutoDock3 (Morris et al 1998) In AutoDock3 the proteinis represented by a grid of affinity potentials Hence theprotein conformation is not allowed to change during thedocking procedure Flexibility of ligands is taken intoaccount by allowing rotation around flexible torsion anglesThe docked energies are calculated as the sum of the inter-molecular interaction energy and the internal energy of theligand The search for the best interaction energy was car-ried out by means of the Lamarckian genetic algorithmLigand structures were obtained from the Protein DataBank and prepared for docking by adding hydrogens andcharges using AutoDockTools (Sanner et al 1996) Eachsimulation consisted of 100 independent runs with a popu-lation size of 200 500 generations and a maximum of25 000 000 energy evaluations Solutions were rankedbased on their docking energies and similar solutions wereclustered (cluster cut-off rmsd lt 1 Aring) For both ligandsthe top solutions were evaluated using the graphics pro-gram O (Jones et al 1991)
Acknowledgements
We thank Andrej Zavialov and Maringhns Ehrenberg for help withequilibrium binding experiments and Thomas Norberg forassistance with mass spectrometry Anthony DeFusco fromMedimmune provided us with mannosylated bovine serumalbumin We are grateful to Fred Sauer for assistance withdata collection Thanks to Maor Bar-Peled of the CCC at theUniversity of Georgia Athens USA for his suggestion toexpress FimH in minimal medium We acknowledge the useof beamline 19BM of the Structural Biology Center at theAdvanced Photon Source Argonne National LaboratoryChicago IL USA and of beamline ID29 ESRF GrenobleFrance JB is a postdoctoral fellow of the Fonds voor Weten-schappelijk Onderzoek-Vlaanderen which also supportedthe BiaCore3000 instrument (FWONL35) This work was sup-ported by grants from the Swedish Research Council andfrom the Strategic Research Foundation (Structural BiologyNetwork) to SDK
Supplementary material
The following material is available from httpwwwblackwellpublishingcomproductsjournalssuppmatmmimmi4415mmi4415smhtmTable S1 Crystallographic data and refinement statistics
References
Anumula KR (1994) Quantitative determination ofmonosaccharides in glycoproteins by high-performanceliquid chromatography with highly sensitive fluorescencedetection Anal Biochem 220 275ndash283
Bahrani-Mougeot FK Buckles EL Lockatell CV HebelJR Johnson DE Tang CM and Donnenberg MS(2002) Type 1 fimbriae and extracellular polysaccharidesare preeminent uropathogenic Escherichia coli virulence
A novel class of FimH high-affinity ligands 453
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
determinants in the murine urinary tract Mol Microbiol 451079ndash1093
Berglund J and Knight SD (2003) Structural basis forbacterial adhesion in the urinary tract Glycobiol Med 53533ndash52
Blattner FR Plunkett G Bloch CA Perna NT BurlandV Riley M et al (1997) The complete genome sequenceof Escherichia coli K-12 Science 277 1453ndashbullbull
Bloch CA Stocker BA and Orndorff PE (1992) A keyrole for type 1 pili in enterobacterial communicability MolMicrobiol 6 697ndash701
Bork P Holm L and Sander C (1994) The immunoglob-ulin fold ndash structural classification sequence patterns andcommon core J Mol Biol 242 309ndash320
Brinton CC (1959) Non-flagellar appendages of bacteriaNature 183 782ndash786
Bruumlnger AT Adams PD Clore GM DeLano WLGros P Grosse-Kunstleve RW et al (1998) Crystallog-raphy and NMR system a new software suite for macro-molecular structure determination Acta Crystallogr D BiolCrystallogr 54 905ndash921
Buchanan K Falkow S Hull RA and Hull SI (1985)Frequency among Enterobacteriaceae of the DNA-sequences encoding type-1 pili J Bacteriol 162 799ndash803
Buts L Bouckaert J De Genst E Loris R Oscarson SLahmann M et al (2003) The fimbrial adhesin F17-G ofenterotoxigenic Escherichia coli has an immunoglobulin-like lectin domain that binds N-acetylglucosamine MolMicrobiol 49 705ndash715
Cheng Y and Prusoff WH (1973) Relationship betweeninhibition constant (K1) and concentration of inhibitor whichcauses 50 per cent inhibition (I50) of an enzymatic reac-tion Biochem Pharmacol 22 3099ndash3108
Choudhury D Thompson A Stojanoff V LangermannS Pinkner J Hultgren SJ and Knight SD (1999) X-ray structure of the FimCndashFimH chaperonendashadhesin com-plex from uropathogenic Escherichia coli Science 2851061ndash1066
Collaborative computational project n4 (1994) The CCP4suite programs for crystallography Acta Crystallogr D BiolCrystallogr 50 760ndash763
Dodson KW Pinkner JS Rose T Magnusson G Hult-gren SJ and Waksman G (2001) Structural basis of theinteraction of the pyelonephritic Escherichia coli adhesin toits human kidney receptor Cell 105 733ndash743
Eley JG and Triumalashetty P (2001) In vitro assessmentof alkylglycosides as permeability enhancers AAPSPharmSciTech 2 (3) Article 19
Esnouf RM (1997) An extensively modified version of Mol-script that includes greatly enhanced coloring capabilitiesJ Mol Graph Model 15 132ndash134
Firon N Ofek I and Sharon N (1982) Interaction ofmannose-containing oligosaccharides with the fimbriallectin of Escherichia coli Biochem Biophys Res Commun105 1426ndash1432
Firon N Ofek I and Sharon N (1983) Carbohydrate spec-ificity of the surface lectins of Escherichia coli Klebsiellapneumoniae and Salmonella typhimurium Carbohydr Res120 235ndash249
Firon N Ofek I and Sharon N (1984) Carbohydrate-
binding sites of the mannose-specific fimbrial lectins ofEnterobacteria Infect Immun 43 1088ndash1090
Firon N Ashkenazis S Mirelman D Ofek I and SharonN (1987) Aromatic a-glycosides of mannose are powerfulinhibitors of the adherence of type 1 fimbriated Escherichiacoli to yeast and intestinal cells Infect Immun 55 472ndash476
Fukiya S Mizoguchi H Tobe T and Mori H (2004)Extensive genomic diversity in pathogenic Escherichia coliand Shigella strains revealed by comparative genomichybridization microarray J Bacteriol 186 3911ndash3921
Hooton TM and Stamm WE (1997) Diagnosis and treat-ment of uncomplicated urinary tract infection Infect DisClin North Am 11 551ndash581
Horovitz A and Levitzki A (1987) An accurate method fordetermination of receptor ligand and enzyme-inhibitor dis-sociation-constants from displacement curves Proc NatlAcad Sci USA 84 6654ndash6658
Hull RA Gill RE Hsu P Minshaw BH and Falkow S(1981) Construction and expression of recombinant plas-mids encoding type 1 and D-mannose-resistant pili from aurinary tract infection Escherichia coli isolate Infect Immun33 933ndash938
Hultgren SJ Porter TN Schaeffer AJ and Duncan JL(1985) Role of type 1 pili and effects of phase variation onlower urinary tract infections produced by Escherichia coliInfect Immun 50 370ndash377
Hung C-S Bouckaert J Hung DL Pinkner J WinbergC Defusco A et al (2002) Structural basis of tropism ofEscherichia coli to the bladder during urinary tract infec-tion Mol Microbiol 44 903ndash915
Jones TA Zou JY Cowan SW and Kjeldgaard M(1991) Improved methods for building protein models inelectron density maps and the location of errors in thesemodels Acta Crystallogr A 47 110ndash119
Justice SS Hung C-S Theriot JA Fletcher DAAnderson GG Footer MJ and Hulgren SJ (2004)Differentiation and developmental pathways of uropatho-genic Escherichia coli in urinary tract pathogenesis ProcNatl Acad Sci USA 101 1333ndash1338
Kallenius G Mollby R Svenson SB Windberg J Lun-dblud A Svenson S and Cedergen B (1980) The Pk
antigen as receptor for the haemagglutinin of pyelonephri-togenic Escherichia coli FEMS Microbiol Lett 8 297ndash302
Knight SD Berglund J and Choudhury D (2000) Bacte-rial adhesins structural studies reveal chaperone functionand pilus biogenesis Curr Opin Chem Biol 4 653ndash660
Langermann S and Ballou WR (2003) Development of arecombinant FimCH vaccine for urinary tract infectionsAdv Exp Med Biol 539 635ndash653
Langermann S Palaszynski S Barnhart M Auguste GPinkner JS Burlein J et al (1997) Prevention ofmucosal Escherichia coli infection by FimH-adhesin-basedsystemic vaccination Science 276 607ndash611
Langermann S Mollby R Burlein JE Palaszynski SRAuguste CG Defusco A et al (2000) Vaccination withFimH adhesin protects cynomolgus monkeys from coloni-zation and infection by uropathogenic Escherichia coli JInfect Dis 181 774ndash778
Larsson A Ohlsson J Dodson KW Hultgren SJ Nils-son U and Kihlberg J (2003) Quantitative studies of thebinding of the class II PapG adhesin from uropathogenic
454 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
Escherichia coli to oligosaccharides Bioorg Med Chem 112255ndash2261
Leffler H and Svanborg-Eden C (1980) Chemical identifi-cation of a glycosphingolipid receptor for Escherichia coliattaching to human urinary tract epithelial cells and agglu-tinating human erythrocytes FEMS Microbiol Lett 8 127ndash134
Lindhorst TK Kieburg C and Krallmann-Wenzel U(1998) Inhibition of the type 1 fimbriae-mediated adhesionof Escherichia coli to erythrocytes by multiantennary alpha-mannosyl clusters the effect of multivalency Glycoconj J15 605ndash613
Loris R Tielker D Jaeger KE and Wyns L (2003a)Structural basis of carbohydrate recognition by the lectinLecB from Pseudomonas aeruginosa J Mol Biol 331 861ndash870
Loris R Imberty A Beeckmans S Van Driessche ERead JS Bouckaert J et al (2003b) Crystal structureof Pterocarpus angolensis lectin in complex with glucosesucrose and turanose J Biol Chem 278 16297ndash16303
Lund B Lindberg F Marklund BI and Normark S(1987) The PapG protein is the a-D-galactopyranosyl-(1-4)-b-D-galactopyranose-binding adhesin of uropathogenicEscherichia coli Proc Natl Acad Sci USA 84 5898ndash5902
Min G Stolz M Zhou G Liang F Sebbel P StofflerD et al (2002) Localization of uroplakin Ia the urothelialreceptor for bacterial adhesin FimH on the six innerdomains of the 16 nm urothelial plaque particle J Mol Biol317 697ndash706
Miroux B and Walker JE (1996) Over-production of pro-teins in Escherichia coli mutant hosts that allow synthesisof some membrane proteins and globular proteins at highlevels J Mol Biol 260 289ndash298
Morris GM Goodsell DS Halliday RS Huey R HartWE Belew RK and Olson AJ (1998) Automateddocking using a Lamarckian genetic algorithm and anempirical binding free energy function J Comput Chem 191639ndash1662
Mulvey MA (2002) Adhesion and entry of uropathogenicEscherichia coli Cell Microbiol 4 257ndash271
Mulvey MA Lopez-Boado YS Wilson CL Roth RParks WC Heuser J and Hultgren SJ (1998) Induc-tion and evasion of host defenses by type 1-piliated uro-pathogenic Escherichia coli Science 282 1494ndash1497
Murshudov GN Vagin AA and Dodson EJ (1997)Refinement of macromolecular structures by the maxi-mum-likelihood method Acta Crystallogr D Biol Crystallogr53 240ndash255
Nagahori N Lee RT Nishimura S Pageacute D Roy Rand Lee YC (2002) Inhibition of adhesion of type 1 fim-briated Escherichia coli to highly mannosylated ligandsChembiochem 3 836ndash844
Navaza J (2001) Implementation of molecular replacementin AMoRe Acta Crystallogr D Biol Crystallogr 57 1367ndash1372
Neeser JR Koellreutter B and Wuersch P (1986)Oligomannoside-type glycopeptides inhibiting adhesion ofEscherichia coli strains mediated by type 1 pili preparationof potent inhibitors from plant glycoproteins Infect Immun52 428ndash436
Old DC (1972) Inhibition of the interaction between fimbrial
hemagglutinatinins and erythrocytes by D-mannose andother carbohydrates J Gen Microbiol 71 149ndash157
Orndorff PE Devapali A Palestrant S Wyse A EverettML Bollinger RR and Parker W (2004) Immunoglob-ulin-mediated agglutination of and biofilm formation byEscherichia coli K-12 require the type 1 pilus fiber InfectImmun 72 1929ndash1938
Oscarson S and Tedebark U (1995) Syntheses of deoxyanalogues of methyl 36-di-O-a-D-mannopyranosyl-a-D-mannopyranoside for studies of the binding site of con-canavalin A Carbohydr Res 278 271ndash287
Oscarson S and Tiden AK (1993) Syntheses of the octyland tetradecyl glycosides of 36-di-O-alpha-deuterium-mannopyranosyl-alpha-deuterium-mannopyranose and of34-di-O-alpha-deuterium-mannopyranosyl-alpha-deute-rium-mannopyranose ndash a new way for 24-di-O-protectionof mannopyranosides Carbohydr Res 247 323ndash328
Otwinowski Z and Minor W (1997) Processing of X-raydiffraction data collected in oscillation mode MethodsEnzymol 276 307ndash326
Powell HR (1999) The Rossmann Fourier autoindexingalgorithm in MOSFLM Acta Crystallogr D Biol Crystallogr55 1690ndash1695
Pratt LA and Kolter R (1998) Genetic analysis of Escher-ichia coli biofilm formation roles of flagella motility chemo-taxis and type I pili Mol Microbiol 30 285ndash293
Remaut H and Waksman G (2004) Structural biology ofbacterial pathogenesis Curr Opin Struct Biol 14 161ndash170
Ronald AR Nicolle LE Stamm E Krieger J WarrenJ Schaeffer A et al (2001) Urinary tract infection inadults research priorities and strategies Int J AntimicrobAgents 17 343ndash348
Sanner MF Olson AJ and Spehner JC (1996)Reduced surface an efficient way to compute molecularsurfaces Biopolymers 38 305ndash320
Sauer FG Futterer K Pinkner JS Dodson KW Hult-gren SJ and Waksman G (1999) Structural basis ofchaperone function and pilus biogenesis Science 2851058ndash1061
Sauer FG Knight SD Waksman GW and HultgrenSJ (2000) PapD-like chaperones and pilus biogenesisSemin Cell Dev Biol 11 27ndash34
Sauer FG Pinkner JS Waksman G and Hultgren SJ(2002) Chaperone priming of pilus subunits facilitates atopological transition that drives fiber formation Cell 111543ndash551
Schembri MA Hasman H and Klemm P (2000)Expression and purification of the mannose recognitiondomain of the FimH adhesin FEMS Microbiol Lett 188147ndash151
Schembri MA Kjaergaard K and Klemm P (2003) Glo-bal gene expression in Escherichia coli biofilms Mol Micro-biol 48 253ndash267
Schilling JD Mulvey MA Vincent CD Lorenz RGand Hultgren SJ (2001) Bacterial invasion augments epi-thelial cytokine responses to Escherichia coli through alipopolysaccharide-dependent mechanism J Immunol166 1148ndash1155
Silhavy TJ Szmelcman S Boos W and Schwartz M(1975) On the significance of the retention of ligand byprotein Proc Natl Acad Sci USA 72 2120ndash2124
A novel class of FimH high-affinity ligands 455
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
Svanborg C and Godaly G (1997) Bacterial virulence inurinary tract infection Infect Dis Clin North Am 11 513ndash529
Thanassi DG Saulino ET and Hultgren SJ (1998) Thechaperoneusher pathway a major terminal branch of thegeneral secretory pathway Curr Opin Struct Biol 1 223ndash231
Thankavel K Madison B Ikeda T Malaviya R ShahAH Arumugen PM and Abraham SN (1997) Local-ization of a domain in the FimH adhesin of Escherichia colitype 1 fimbriae capable of receptor recognition and use ofa domain-specific antibody to confer protection againstexperimental urinary tract infection J Clin Invest 1001123ndash1126
Vagin A and Teplyakov A (1997) MOLREP an automatedprogram for molecular replacement J Appl Crystallogr 301022ndash1025
Wu XR Sun TT and Medina JJ (1996) In vitro bindingof type 1-fimbriated Escherichia coli to uroplakins 1a and
1b relation to urinary tract infections Proc Natl Acad SciUSA 93 9630ndash9635
Xia Y Gally D Forsman-Semb K and Uhlin BE (2000)Regulatory cross-talk between adhesin operons in Escher-ichia coli inhibition of type 1 fimbriae expression by thePapB protein EMBO J 19 1450ndash1457
Zafriri D Ofek I Adar R Pocino M and Sharon N(1989) Inhibitory activity of cranberry juice on adherenceof type-1 and type-P fimbriated Escherichia coli to eukary-otic cells Antimicrob Agents Chemother 33 92ndash98
Zavialov AV Berglund J Pudney AF Fooks LJ Ibra-him TM MacIntyre S and Knight SD (2003) Structureand biogenesis of the capsular F1 antigen from Yersiniapestis preserved folding energy drives fiber formation Cell113 587ndash596
Zhou G Mo WJ Sebbel P Min G Neubert TA Glock-shuber R et al (2001) Uroplakin Ia is the urothelial recep-tor for uropathogenic Escherichia coli evidence from invitro FimH binding J Cell Sci 114 4095ndash4103

A novel class of FimH high-affinity ligands 445
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
the anomeric carbon atom was often combined with elim-ination of one or more acetic acid molecules (massdecrease of 60 Da) or acetyl groups (mass decrease of42 Da) These eliminations gave rise to the major ions atmz 127 and mz 169 with a proposed aromatic ring struc-ture as well as to the ion at mz 109 with a proposedquinone-like ring structure Thus the mass spectrometricdata support the butyl hexose nature of the FimH ligandFimH was further characterized by carbohydrate constit-uent analysis (data not shown) which revealed only man-nose Mannose could only be detected after hydrolysisindicating that free mannose is not associated with theFimH truncate
Origin of butyl a-D-mannoside
As no sugar had been advertently added to the FimHtr
protein samples we hypothesized that the butyl manno-side ligand had been picked up from the LuriandashBertani(LB) medium used to grow bacteria during expression ofthe protein To test this FimHtr2 was expressed in bacteriagrown in minimal M9 medium Electro-spray ionizationmass spectrometry (ESI-MS) was performed on purifiedFimHtr2 samples from both types of media A clear peakat 259 Da could be detected (Fig 3A) with proteinexpressed in bacteria grown in LB medium correspondingto the molecular weight of butyl mannoside (236 Da) plus
a sodium ion (23 Da) As a control butyl mannoside wassynthesized and analysed in the same manner Again apeak at 259 Da was observed (Fig 3A inset) When ESI-MS was run on protein expressed in bacteria grown inminimal medium no such peak could be detected(Fig 3B) indicating that the ligand was not present andthat it indeed originated from the LB medium
Interactions of the butyl moiety with FimH
The butyl moiety of butyl a-D-mannoside extends out ofthe mannose-binding pocket towards Tyr48 and Tyr137making van der Waals contacts to both tyrosine rings andIle52 (Fig 1B and C) These residues are part of two loops(b4ndashb5 and b10ndashb11) that form the higher rear end of ahydrophobic collar around the binding site (Fig 1A andB) In both subunits of the FimHtr2 structure the Tyr48 andTyr137 side-chains are in the same almost parallel ori-entation as in the FimCFimH structure forming a gate-like structure at the back of the binding site (Fig 1C) Theclosest hydrophobic interactions (31ndash36 Aring) with the alkylchain are made between Tyr48 and the two non-terminalatoms of the butyl In the FimHtr1 structure a crystal con-tact to Val155 of a neighbouring molecule prevents theparallel orientation of the Tyr48 ring which instead ispacked edge-to-face with Tyr137 (Fig 1B) The reposition-ing of Tyr48 is accompanied by a change in the confor-
A
B
Fig 2 Mass spectrophotometric characteriza-tion of the butyl mannosideA Base peak chromatogram (BPC mass range mz 400ndash550) and extracted ion chromato-grams (EIC) of mz 427 and 331 obtained by nano-LC-ESI-MS of the derivatized FimH-ligand purified by peracetylation and organic-solvent extractionB ESI-IT mass spectrometric characterization of the peak species marked by a horizontal bar in (A) Ions are marked with mz-values and proposed schematic structures which is often only one out of several isobaric ion species Fragmentations already occurred in the source Ac acetyl
446 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
mation of the butyl chain of the ligand with the butyl chainin a more extended conformation than in the FimHtr2 struc-ture and with the four carbon atoms nearly in one plane(Fig 1B and C) The new conformations of Tyr48 and thebutyl moiety result in an almost parallel stacking of thearomatic ring of Tyr48 onto the butyl plane with the closesthydrophobic interactions (37 Aring) involving the two terminalcarbon atoms of the butyl (Fig 1B)
Affinity measurements for alkyl mannosides
The butyl mannoside ligand remained bound to FimHtr
despite extensive dialysis From a study of complex for-mation of monoclonal antibodies IC10 and IA7 withFimHtr1 it could be estimated that about 30 of the FimHtruncate retained its ligand after dialysis for 12 daysagainst 50 mM MES buffer at pH 65 100 mM NaCldespite daily refreshment of buffer (data not shown)Therefore to enable measurements of dissociation con-stants of FimH for mannosides FimH was produced inminimal medium Butyl a-D-mannoside binding is 15 timesimproved (Kd = 151 nM Fig 4A) compared with mannosebinding The equilibrium binding technique using [3H]-mannose led to a similar value (Kd = 41 mM Fig 4B) for
mannose and an almost identical value for butyl a-D-mannoside (Kd = 150 nM Fig 4C) To investigate theeffect of sequential addition of methyl groups to the O1oxygen of D-mannose a series of alkyl mannosides weresynthesized and the dissociation constants determinedusing the [3H]-mannose displacement assay as well as theSPR competition assay (Table 1) There is a linear corre-lation between the binding free energy as calculated fromthe measured dissociation constants and the number ofmethyl groups (between 1 and 7) in the alkyl mannosidewith each additional methyl group contributing on average-06 kcal mol-1 of binding energy (Fig 5)
Affinity measurements and modelling for aryl mannosides
Aromatically substituted mannosides have been reportedto be particularly potent inhibitors of FimH-mediated bac-terial adhesion (Firon et al 1987 Nagahori et al 2002)In accordance with the earlier results both pNPaMan andMeUmbaMan bind very tightly to FimH (Table 1) Despiteextensive trials no X-ray structures were obtained forcomplexes with aromatically substituted mannosides Inorder to model binding docking to both of the truncatestructures was carried out In all cases the mannose ring
Fig 3 Mass spectrometric analysis of the origin of the butyl mannosideA LB medium-produced FimH truncate The inset shows the mass spectrum of the synthesized butyl a-D-mannosideB Minimal media-produced FimH truncate
A novel class of FimH high-affinity ligands 447
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
docks in the binding pocket in a conformation very closeto the crystallographically observed mannose conforma-tion (Fig 6) while the aromatic substituent interacts withthe tyrosine gate The different conformation of Tyr48 in
our two FimH truncate structures leads to different pre-dicted interactions between the aromatic aglycon andTyr48 and Tyr137 Docking to the FimHtr2 structureresulted in somewhat tighter interactions between thetyrosine gate and the aglycon and hence in somewhatlower docking energies Calculated FimHtr2 docking ener-gies of Edoc = -104 kcal mol-1 for pNPaMan (Kd = 46 nM)and Edoc = -109 kcal mol-1 for MeUmbaMan (Kd = 20 nM)are in good agreement with the experimental binding data(Table 1) The docked umbelliferyl ring system is inserteddeep into the tyrosine gate in the FimHtr2 conformationwith the carbonyl group within hydrogen bonding distanceto Thr51 at the back of the gate (Fig 6A) Docking to theFimHtr1 structure instead stacks the umbelliferyl ring face-to-face with the Tyr48 ring (not shown) The phenyl ring ofpNPaMan docks similarly to both our structures It doesnot quite reach in between the tyrosine side-chains butinstead hydrogen bonds with its nitro group to the hydroxyloxygen of Tyr137 (Fig 6B)
Discussion
Most lectins bind naturally occurring carbohydrates withlow affinities often in the millimolar range for monosac-charides and micromolar range for oligosaccharides Thiswas recently exemplified to also hold true for theadhesins of F17 fimbriae (Kd = 12 mM for the F17a-G-N-acetylglucosamine interaction Buts et al 2003) and Ppili (Kd = 84 mM for the PapGII-globotetraoside interactionLarsson et al 2003) The recent crystal structure of FimHin complex with mannose illustrated that FimH almostcompletely envelops the monosaccharide in a deeppocket and involves it in an intense hydrogen bondingnetwork suggestive of an unusual lectin exerting highaffinities (Hung et al 2002)
Fig 4 FimH binding experimentsA Surface plasmon resonance competition experiment of FimHtr1 binding to monoclonal antibody IC10 Fab fragments with butyl a-D-mannosideB Equilibrium binding of [3H]-mannose to FimHtr2C Equilibrium displacement measurements of butyl a-D-mannoside to the FimHtr2 [3H]-mannose complex
log[Butyl a-D-mannoside] (M)
cpm
cpm
Butyl a-D-mannoside (mM)
Mannose (mM)
A
B
C
ndash64
ndash66
ndash68
ndash70
ndash72
ndash74
ndash76
ndash78
ndash80
ndash82
ndash8 ndash7 ndash6 ndash5 ndash4 ndash3
log
[Fim
H]
M
Fig 5 Linear free energy relationship for the alkyl chain on mannose A linear fit was made using 1 up to 7 methyl groups with the program MICROCAL ORIGIN 50
Number of carbon atoms0 2 4 6 8
ndash75
ndash80
ndash90
ndash95
ndash100
ndash105
ndash110
ndash115
ndash85
D G
(kc
al m
olndash1
)
slopecorrelation
ndash064ndash099
+ndash 003+ndash 018
448 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
The measured affinity confirm the predictions that FimHbinds the monosacharide mannose exceptionally well(Kd = 23ndash41 mM) To investigate the contribution of thedifferent hydroxyl groups of mannose in the interactionwith FimH the binding of a series of hexoses to butylmannoside-free FimH was examined using SPR (Table 2)The effect of deoxygenation of the O2 hydroxyl group(methyl 2-deoxy-a-D-mannopyranoside) is clearly lessdestructive (100-fold reduction) than its equatorial positionas in glucose O2 is the only hydroxyl group of mannosethat interacts indirectly with the FimH pocket via a con-served water molecule (Hung et al 2002) Glucose onlydisplays a weak affinity for FimH about 4000 times lowerthan methyl a-D-mannoside Repositioning of both the O2and O4 hydroxyl groups as in galactose abolishes sugarbinding by FimH Similar to these results the inhibitoryconcentrations for bacterial adherence of methyl a-D-glucoside and galactose derivatives have been estimatedto be respectively 104-fold and 105-fold weaker than formethyl a-D-mannoside (Nagahori et al 2002)
In stark contrast with the other monosaccharides fruc-tose has a binding affinity only about 15-fold less thanmannose for FimH (Table 2) The decrease in affinity for
fructose relative to mannose agrees with studies of inhi-bition of adherence of type 1 piliated E coli by fructosein which fructose was found to be the active compound infruit juices (present at about 5) (Zafriri et al 1989)Fructose binding to FimH is stronger than the binding ofthe physiological globotetraoside receptor to PapGII theother well-characterized adhesin of UPEC strains (Lars-son et al 2003) The pyranose form of fructose is verywell superimposable onto mannose in the FimH bindingsite where similar interactions are established with FimHas for mannose except for the loss of the anomeric O1and an extra hydroxyl group (O2) on the C5 position ofmannose Probably however both changes of fructopyr-anoside compared with mannopyranoside could be easilyaccommodated by FimH The mannose O1 does notmake direct interactions within 45 Aring of the FimH bindingpocket (Fig 1B) The extra O2 group of fructose would notclash or interfere significantly with the binding pocket butit would make a 27 Aring van der Waals contact with the Ile52side-chain under the condition that the Ile52 side-chaindoes not move The crystal structure of the Pseudomonasaeruginosa lectin LecB also showed that fructopyranosebut not fructofuranose can substitute for mannose (Loris
Fig 6 Stereo diagrams of docked MeUm-baMan (A) and pNPaMan (B) in the FimH bind-ing site The crystallographically determined position of a-D-mannose is shown in orange for comparison
A novel class of FimH high-affinity ligands 449
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
et al 2003a) Hydrogen bonds of the more specificfucose complex are replaced by van der Waals interac-tions involving a hydroxyl group in the complexes with thelower-affinity binders mannose and fructopyranose as forthe proposed fructopyranoside binding to FimH
The fructose-containing disaccharides sucrose andturanose do not have an inhibitory effect and bind withaffinities similar to glucose Sucrose is a non-reducingsugar that has fructose locked in its furanose form Tura-nose on the other hand allows the interconversion of b-D-fructofuranose and b-D-fructopyranose eg both formsoccur in complex with a plant lectin from Pterocarpusangolensis (Loris et al 2003b) It is predictable howeverfrom the crystal structures that the glycosidic linkage ofglucose with C3 of fructose equivalent in position to C4of mannose would be disallowed in the tight FimH pocketSucrose and turanose thus most probably interact with theFimH mannose binding pocket via their glucose moietyresulting in similar low affinities
Our mass spectrometry and chemical analyses con-firms the crystallographic identification of the ligand boundto the FimH lectin domains as butyl a-D-mannoside(Figs 1B and 2) The mannose ring of the butyl a-D-mannoside coincides with the mannose bound to theFimCFimH complex (Hung et al 2002) and makes thesame interactions The four-carbon alkyl tail exits towardsthe back wall (loop b4ndashb5 and b10ndashb11) of the pocket(Fig 1A and B) making hydrophobic contacts to Tyr48Tyr137 and Ile52 The mass spectrometry data also showthat the bound butyl a-D-mannoside is present in LBmedium but absent from minimal medium Hence thesource of the alkyl mannoside is possibly the yeast extractused in preparation of LB medium Although we do notwant to speculate about the significance of this findingalkyl mannoside production by yeast has not beenreported and might warrant investigation
The butyl mannoside was retained by FimH despiteextensive dialysis The phenomenon of ligand retention byprotein was described as early as 1975 for periplasmicbinding proteins (Silhavy et al 1975) Applied to dialysisof butyl mannoside away from FimH grown in richmedium half-lives of up to 30 h can be estimated whichis far beyond the duration of a normal buffer exchangedialysis experiment In the present case the retainedligand does not appear to seriously affect [3H]-mannosedisplacement equilibrium binding experiments as wecould detect no significant differences in dissociation con-stants obtained using FimHtr2 produced in LB or minimalmedia (data not shown) In contrast proper SPR bindingcurves could not be obtained when butyl mannoside occu-pied the binding site Hence the displacement techniquemay be of more general use to obtain binding constantsfor proteins that tend to inadvertently retain ligands whichis a common trait of lectins
Surface plasmon resonance as well as equilibrium bind-ing and displacement measurements using [3H]-mannosewere carried out to obtain dissociation constants for a-D-mannose and a linear series of alkyl-substituted manno-sides to FimH (Table 1) Both equilibrium binding experi-ments show a very good agreement in their measurementof the carbohydrate affinity of the adhesin FimH bindsbutyl a-D-mannoside 15ndash30 times stronger than a-D-mannose The importance of the added hydrophobiccontacts is clearly demonstrated by the binding of alkylmannosides (Table 1) The best binding alkyl mannosidesbind a few hundred times stronger than mannose equiv-alent in affinity to the most tightly binding aromaticallysubstituted mannosides for FimH (Firon et al 1987) andof mannose dendrimers for type 1 piliated E coli (Fironet al 1984 1987 Nagahori et al 2002)
Interestingly the butyl moiety takes on significantlydifferent conformations in FimHtr1 and FimHtr2 (Fig 1C)indicating that intrinsic conformational flexibility in theaglycon is allowed Indeed in spite of the larger flexibilityexpected for longer alkyl mannosides the binding energydecreases linearly with increasing length of the alkyl chainbetween one and seven carbon atoms (Fig 5) The affinityreaches its maximum for mannose with a seven-carbonalkyl and SPR measurements suggest that the affinitydecreases from eight carbon atoms on Evidently theeighth carbon atom goes beyond the hydrophobic interac-tion surface and protein edge which is penalized with adecrease in affinity We propose that the affinity increaseresults from an increased interaction surface with thehydrophobic gateway Alkyl aglycons fit well in the tyrosinegate and as observed in the two crystal structures in atleast two different conformations that depend on theconformation of Tyr48 This flexibility may indicate thataglycons can interact rather non-specifically with thehydrophobic gateway nevertheless resulting in a 100- to1000-fold increase in affinity
FimH binding to ligands other than just mannose ispossibly of importance in terms of the different environ-ments that E coli will encounter ndash the urinary tract gas-trointestinal tract and in biofilms Alkyl mannosides havenot been previously recognized as strong binders to FimHAs they are easily synthesized and highly soluble in waterthey may be interesting as potential blocking agents forFimH-mediated adhesion Moreover they have beenshown to be good drug vehicles themselves acting aspermeability enhancers on tight junctions (Eley and Triu-malashetty 2001) As such they might be able to pene-trate into deeper cell layers of the epithelium to alsoprevent those bacteria that form a quiescent reservoirfrom causing recurrent infections (Mulvey et al 1998Justice et al 2004)
On the other hand the two aromatically substitutedmannosides pNPaMan and MeUmbaMan are known to
450 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
be particularly potent inhibitors of FimH-mediated bacte-rial adhesion (Firon et al 1987 Nagahori et al 2002)Based on adhesion inhibition studies using type 1 piliatedE coli and guinea pig ileal epithelial cells the relativeinhibitory potential of pNPaMan and MeUmbaMan havebeen estimated to be about 70 and 600 times strongerrespectively than that of methyl a-D-mannose (Fironet al 1987) Our measurements indicate an affinity ofFimH for pNPaMan and MeUmbaMan between 50 and90 times and between 110 and 200 times higher respec-tively than for methyl a-D-mannose in good agreementwith the adhesion inhibition results (Firon et al 1987)Dissociation constants predicted using AutoDock3 (Morriset al 1998) are similar to experimental values Thecomputational docking predicts that tight binding of thearomatically substituted mannosides pNPaMan andMeUmbaMan is accomplished similarly as for butyl a-D-mannoside with the aglycon interacting with the tyrosinegate
This is the first systematic study providing molecularbinding data for the FimH adhesin the prerequisite forcolonization by UPEC (Langermann and Ballou 2003)We also show crystallographic evidence that the lectinor receptor binding domain of FimH is a stable andfunctional entity Its structure is virtually identical to thesame domain in the FimCFimH complex indicating thata truncate corresponding to the lectin domain could pro-vide a good alternative for the development of a UTIvaccine (Langermann et al 1997 Langermann and Bal-lou 2003) However progress of this interesting UTI vac-cine candidate into clinical trials has not been reportedThe binding affinities of FimH at the molecular levelrelate convincingly well with the inhibitory concentrationsused in previous bacterial adhesion assays thus endors-ing the direct and conclusive correlation of FimH withbacterial adherence Alkyl mannosides have nanomolaraffinities for FimH which may overcome the significantobstacle of low affinity generally encountered in attemptsto develop carbohydrate-based drugs which interferewith microbial attachment to host cell surface glycocon-jugates New anti-adhesive drugs could be based onthese leads to allow specific interaction and targeting tothe urinary tract in order to prevent anticipated andrecurrent infections
Experimental procedures
Strains and constructs
FimH proteins from two different E coli strains the UPECstrain J96 (Hull et al 1981) and the E coli K-12 strainMG1655 (Blattner et al 1997) were used in this study TheFimH adhesins encoded by these two E coli strains areidentical FimHtr1 (residues 1ndash158 of UPEC J96 FimH) wasexpressed from plasmid pMMB91 transformed into E coli
C600 cells FimHtr2 (residues 1ndash158 of E coli MG1655FimH + carboxy-terminal 6-His tag) was expressed fromplasmid pPKL241 (Schembri et al 2000) transformed into Ecoli host strain HB101
Purification and crystallization
FimHtr1 Bacteria were grown in LB medium at 37infinC inducedat OD600 = 08 with 1 mM IPTG and harvested 2ndash3 h laterThe cells were resuspended in 20 mM Tris-HCl pH 80 5 mMEDTA and 30 sucrose and the periplasmic content wasextracted by osmotic shock upon 10-fold dilution with 20 mMTris-HCl pH 80 Purification of FimHtr1 was performed by ionexchange chromatography with MonoS at pH 45 followed byMonoQ (Amersham Biosciences) at pH 8 FimHtr1 was crys-tallized using vapour diffusion in hanging drops composed of3 ml of the precipitant solution (01 M Hepes pH 75 12PEG 6000 and 5 vv methyl-24-pentanediol) and 3 ml of12 mg ml-1 protein solution in 20 mM Tris-HCl pH 80 with150 mM NaCl
FimHtr2 Cells were grown in LB medium containing50 mg ml-1 ampicillin at 37infinC At OD600 = 06 the cells wereinduced with 5 mM IPTG Cells were harvested 2 h afterinduction and the periplasmic content was extracted Theprotein was purified on a Pharmacia HiTrap Chelating HP5 ml column (Pharmacia) loaded with Ni-chloride and elutedwith a sharp 0ndash500 mM imidazole gradient Fractions con-taining FimHtr2 were pooled dialysed against 50 mM sodiumacetate pH 525 and loaded onto a Mono S HR 8 ml columnThe protein was eluted with a 0ndash500 mM NaCl gradient anddialysed overnight against 20 mM MES pH 65 before crys-tallization Crystallization was performed at room tempera-ture by the vapour diffusion method with hanging dropsmade by mixing of 2 ml of 15 mg ml-1 protein solution with 2 mlof the precipitant solution (65 methyl-24-pentanediol100 mM cacodylate pH 65) Microseeding was employed tooptimize crystal quality and reproducibility Crystals suitablefor X-ray data collection generally appeared within 48 h aftermicroseeding
Structure determination
X-ray data for FimHtr1 were collected from a single crystalto 17 Aring resolution at the Advanced Photon SourceArgonne National Laboratory USA The crystal was flash-cooled to 100 K in the precipitant solution complementedwith 25 glycerol The data were processed and scaledusing the HKL2000 package (Otwinowski and Minor 1997)(Table S1 in Supplementary material) Data for FimHtr2
were collected from a single crystal under liquid-nitrogencryo conditions at 100 K at beamline ID29 ESRF Greno-ble France to 21 Aring Bragg spacing FimHtr2 data were pro-cessed and reduced using MOSFLM (Powell 1999) andscaled using SCALA (Collaborative computational project1994) (Table S1 in Supplementary material) The structuresof FimHtr1 and FimHtr2 were solved by molecular replace-ment using Amore (Navaza 2001) and MOLREP (Vaginand Teplyakov 1997) respectively with the FimH lectindomain from the FimCFimH complex (Choudhury et al
A novel class of FimH high-affinity ligands 451
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
1999) (PDB id 1QUN) as the search model Positional bulksolvent and isotropic temperature factor refinement wasperformed using CNS (Bruumlnger et al 1998) and REFMAC5(Murshudov et al 1997) respectively Model adjustmentand inspection were performed using Turbo-Frodo and O(Jones et al 1991) graphics programs respectively Refine-ment statistics are given in Table S1 Molecular graphicsillustrations were generated with Bobscript (Esnouf 1997)and Pymol 090 (DeLano Scientific LLC 1998ndash2003 httppymolsourceforgenet)
Mass spectrometry
The purified FimHtr1 protein (300 mg) that had been used forcrystallization was freeze-dried taken up in 100 ml of pyridineand 100 ml of acetic anhydride and incubated overnight atroom temperature in the dark for peracetylation The samplewas dried under nitrogen and taken up in chloroform (400 ml)The chloroform phase was extracted six times with waterdried and taken up in 200 ml of 50 aqueous methanol A1 ml aliquot was applied to a nano-scale RP column(75 mm yen 100 mm PepMap DionexLC Packings) using anUltimate nano-LC system (LC Packings) which was equili-brated with eluent A (H2Oacetonitrile 955 vv containing01 formic acid) at a flow rate of 100 nl min-1 After injectingthe sample the column was run isocratically for 5 min fol-lowed by a linear gradient to 100 eluent B (H2Oacetonitrile2080 vv containing 01 formic acid) in 20 min and a finalwash with 100 B for 5 min The system was directly coupledto an Esquire 3000 ESI-IT-MS (Bruker Daltonik) equippedwith an online nanospray source operating in the positive-ionmode For electrospray (900ndash1200 V) capillaries (360 mmOD 20 mm ID with 10 mm opening) from New Objective wereused The solvent was evaporated at 150infinC with a nitrogenstream of 6 l min-1
To verify the suspected source of the bound butyl manno-side mass spectrometry was performed on samples ofFimHtr2 grown in either LB or minimal M9 medium Proteinsamples were dialysed against water overnight before massspectrometry in order to reduce background from the Trisbuffer Samples were diluted to a concentration in the submi-cromolar range Electrospray mass spectra were recorded indirect inlet mode (50 aqueous methanol solutions) on aBruker Daltonics Esquire-LC (Billerica)
Carbohydrate constituent analysis
Samples were hydrolysed in 100 ml of 4 M aqueous trifluoro-acetic acid (Sigma) at 100infinC for 4 h and dried under astream of nitrogen Monosaccharides were converted intotheir anthranilic acid derivatives by reductive aminationresolved by RP-HPLC and detected by fluorescence (Anu-mula 1994)
Carbohydrates for binding studies
[3H]-mannose was obtained from Amersham Methyl a-D-mannoside glucose galactose fructose sucrose turanosep-nitrophenyl a-D-mannopyranoside (pNPaMan) and 4-methylumbelliferyl a-D-mannopyranoside (MeUmbaMan)
were obtained from Sigma Monomannosylated bovineserum albumin (BSA-Man) was kindly provided by MedIm-mune We synthesized methyl 2-deoxy-a-D-mannopyrano-side (Oscarson and Tedebark 1995) Alkyl mannosides weresynthesized through silver triflate-promoted couplings of thecorresponding alcohol with 2346-tetra-O-benzoyl-a-D-mannopyranosyl bromide followed by Zempleacuten deacylationof the obtained protected alkyl mannosides according to theprocedure reported for the octyl and tetradecyl mannosides(Oscarson and Tiden 1993)
Binding studies
Competition experiments using SPR To obtain butyl man-noside-free FimHtr1 the FimH truncate was overproducedwith a carboxy-terminal 6-His tag after transformation into Ecoli C43 (DE3) (Miroux and Walker 1996) and the bacteriagrown in minA minimal medium supplemented with the 20amino acids (40 mg l-1) thiamin (2 mg l-1) and biotin (2 mgl-1) Purification of the His-tagged protein was performedusing Ni-NTA affinity chromatography (Qiagen) in 50 mM Tris-HCl pH 8 and 300 mM NaCl Elution was in the same solutioncomplemented with 500 mM imidazole Subsequently pooledand concentrated FimH fractions were gel filtrated over aSuperdex G75 column (Pharmacia) in 20 mM Tris-HCl pH 8
We used Fab fragments of a monoclonal antibody againstthe mannose-binding site of FimH to determine the solutionaffinity of FimHndashcarbohydrate interactions The monoclonalantibody IC10 was produced by a mouse hybridoma cell lineat Medimmune and its Fab fragments were prepared andpurified The Fabs were covalently immobilized via aminecoupling at 1000 Resonance Units (1000 pg ligand mm-2) inflowcell Fc2 on a CM5 biasensor chip (BIAapplications Hand-book Biacore AB Uppsala Sweden) Flowcell Fc1 was usedas the reference surface and was treated in the same wayas Fc2 except that it was blocked immediately after activationThe immobilization buffer was 100 mM sodium acetatepH 50 with 100 mM NaCl All SPR measurements were per-formed on a Biacore3000trade
The kinetic constants of binding of FimH to the immobilizedantibody were measured with different concentrations ofFimH (2000 nM 1000 nM 500 nM 250 nM 125 nM625 nM 312 nM 156 nM 78 nM) in running buffer [phos-phate-buffered saline (PBS) with 0005 surfactant P20 and3 mM EDTA] on both flowcells Fc1 and Fc2 simultaneouslyat a flow rate of 30 ml min-1 and at 298 K Complete dissoci-ation of FimH was performed with running buffer beforestarting a new binding cycle For all measurements the asso-ciation time was 3 min and the dissociation time was 30 minAll binding cycles were performed in duplicate including azero concentration cycle of FimH (injection of running buffer)All analyses were performed with the BIAeval software ALangmuir binding isotherm with a 11 stoichiometry was fittedto the data from both the association phase and the dissoci-ation phase after subtraction of the reference and zero con-centration data The kinetic constants ka and kd and themaximal binding Rmax were obtained and the dissociationconstant Kd was calculated
The equilibrium dissociation constants Kd of FimHndashsaccha-ride interactions were determined in a competition experi-ment using the same chip as for the previous experiment
452 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
As a control the Fabs of monoclonal antibody IA7 as wellas BSA-Man were also immobilized Samples containing afixed concentration of FimH were used (close to the Kd of theFimHndashantibody interaction) in combination with varying con-centrations of saccharide First 10-fold dilutions of the sac-charide solution were used to determine the concentrationrange for binding of the saccharide to FimH A Langmuirbinding isotherm with a 11 stoichiometry was fitted to thedata using the kinetic constants and Rmax from the foregoingexperiment to obtain the concentrations of FimH that werefree ([FimH]free) to bind the antibody immobilized on the chipSecond the concentration range of the saccharide wasextended and adapted to ensure accurate fitting and theequilibrium binding constant of the FimHndashsaccharide interac-tion was derived from the curve of [FimH]free against concen-tration of saccharide Every measurement was repeated atleast twice including testing different protein batches andwhenever possible also different saccharide stock solutions(typically 200 mM)
Equilibrium binding studies using radioactively labelledmannose Mannose-free FimHtr2 was produced inM9 minimal medium and purified as described above forFimHtr2 Alkyl mannosides were dissolved in double distilledwater pNPaMan and MeUmbaMan (6 mg each) were dis-solved in 20 ml of DMSO and diluted to 20 mM using doubledistilled water Binding experiments were performed at 310 Kusing six different concentrations of [3H]-a-D-mannose (finalconcentrations 435 mM 290 mM 193 mM 129 mM 86 mMand 57 mM) Protein (180 ml) at a concentration of about500 nM was mixed with 20 ml of the radioactive ligand [3H]-a-D-mannose and incubated at 37degC for 20 min To separatefree ligand from bound the mixture was rapidly filtratedthrough a Protran BA 85 cellulose nitrate filter (Schleicherand Schuell) and washed once with 1 ml of ice-cold 1yen PBSFilter-bound radioactivity was measured by scintillation spec-trometry within 24 h Displacement experiments were per-formed using six different concentrations of inhibitor (finalconcentrations in the range 00ndash435 mM) in the presence of435 mM [3H]-a-D-mannose Twenty microlitres of radioactiveligand 20 ml of inhibitor and 160 ml of protein (500 nM) weremixed and the experiments were performed as describedabove All experiments were performed in duplicates and thedata presented are mean values of at least two independentmeasurements For the determination of Kd for a-D-mannosean hyperbolic curve (y = P1x(P2 + x) where P1 is an arbitraryscale factor and P2 = Kd) was fitted to the data For the dis-placement experiments the curve y = P1(P2 + x) where P1
is an arbitrary scale factor and P2 is the concentration of theinhibitor displacing 50 of the labelled ligand [I]05 was usedinstead To calculate the inhibitor dissociation constant (KI)the Cheng and Prusoff equation [KI = [I]05[L]KL + 1) KL is theconstant of dissociation for the ligand] (Cheng and Prusoff1973) was used when both the concentration of the radio-active ligand (L) and the displacing agent (I) are in excessover the protein (LT gtgt PT IT gtgt PT T indicates total concen-tration) For the two strong inhibitors pNPaMan and MeUm-baMan where IT was not in excess over PT the equationof KI = IT[(1 - Y)Y(LTKL) - 1] - PTKLYLT where Y is thefraction of the ligand bound in presence of the inhibitor wasused instead (Horovitz and Levitzki 1987)
Computational docking
Computational docking studies were performed usingAutoDock3 (Morris et al 1998) In AutoDock3 the proteinis represented by a grid of affinity potentials Hence theprotein conformation is not allowed to change during thedocking procedure Flexibility of ligands is taken intoaccount by allowing rotation around flexible torsion anglesThe docked energies are calculated as the sum of the inter-molecular interaction energy and the internal energy of theligand The search for the best interaction energy was car-ried out by means of the Lamarckian genetic algorithmLigand structures were obtained from the Protein DataBank and prepared for docking by adding hydrogens andcharges using AutoDockTools (Sanner et al 1996) Eachsimulation consisted of 100 independent runs with a popu-lation size of 200 500 generations and a maximum of25 000 000 energy evaluations Solutions were rankedbased on their docking energies and similar solutions wereclustered (cluster cut-off rmsd lt 1 Aring) For both ligandsthe top solutions were evaluated using the graphics pro-gram O (Jones et al 1991)
Acknowledgements
We thank Andrej Zavialov and Maringhns Ehrenberg for help withequilibrium binding experiments and Thomas Norberg forassistance with mass spectrometry Anthony DeFusco fromMedimmune provided us with mannosylated bovine serumalbumin We are grateful to Fred Sauer for assistance withdata collection Thanks to Maor Bar-Peled of the CCC at theUniversity of Georgia Athens USA for his suggestion toexpress FimH in minimal medium We acknowledge the useof beamline 19BM of the Structural Biology Center at theAdvanced Photon Source Argonne National LaboratoryChicago IL USA and of beamline ID29 ESRF GrenobleFrance JB is a postdoctoral fellow of the Fonds voor Weten-schappelijk Onderzoek-Vlaanderen which also supportedthe BiaCore3000 instrument (FWONL35) This work was sup-ported by grants from the Swedish Research Council andfrom the Strategic Research Foundation (Structural BiologyNetwork) to SDK
Supplementary material
The following material is available from httpwwwblackwellpublishingcomproductsjournalssuppmatmmimmi4415mmi4415smhtmTable S1 Crystallographic data and refinement statistics
References
Anumula KR (1994) Quantitative determination ofmonosaccharides in glycoproteins by high-performanceliquid chromatography with highly sensitive fluorescencedetection Anal Biochem 220 275ndash283
Bahrani-Mougeot FK Buckles EL Lockatell CV HebelJR Johnson DE Tang CM and Donnenberg MS(2002) Type 1 fimbriae and extracellular polysaccharidesare preeminent uropathogenic Escherichia coli virulence
A novel class of FimH high-affinity ligands 453
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
determinants in the murine urinary tract Mol Microbiol 451079ndash1093
Berglund J and Knight SD (2003) Structural basis forbacterial adhesion in the urinary tract Glycobiol Med 53533ndash52
Blattner FR Plunkett G Bloch CA Perna NT BurlandV Riley M et al (1997) The complete genome sequenceof Escherichia coli K-12 Science 277 1453ndashbullbull
Bloch CA Stocker BA and Orndorff PE (1992) A keyrole for type 1 pili in enterobacterial communicability MolMicrobiol 6 697ndash701
Bork P Holm L and Sander C (1994) The immunoglob-ulin fold ndash structural classification sequence patterns andcommon core J Mol Biol 242 309ndash320
Brinton CC (1959) Non-flagellar appendages of bacteriaNature 183 782ndash786
Bruumlnger AT Adams PD Clore GM DeLano WLGros P Grosse-Kunstleve RW et al (1998) Crystallog-raphy and NMR system a new software suite for macro-molecular structure determination Acta Crystallogr D BiolCrystallogr 54 905ndash921
Buchanan K Falkow S Hull RA and Hull SI (1985)Frequency among Enterobacteriaceae of the DNA-sequences encoding type-1 pili J Bacteriol 162 799ndash803
Buts L Bouckaert J De Genst E Loris R Oscarson SLahmann M et al (2003) The fimbrial adhesin F17-G ofenterotoxigenic Escherichia coli has an immunoglobulin-like lectin domain that binds N-acetylglucosamine MolMicrobiol 49 705ndash715
Cheng Y and Prusoff WH (1973) Relationship betweeninhibition constant (K1) and concentration of inhibitor whichcauses 50 per cent inhibition (I50) of an enzymatic reac-tion Biochem Pharmacol 22 3099ndash3108
Choudhury D Thompson A Stojanoff V LangermannS Pinkner J Hultgren SJ and Knight SD (1999) X-ray structure of the FimCndashFimH chaperonendashadhesin com-plex from uropathogenic Escherichia coli Science 2851061ndash1066
Collaborative computational project n4 (1994) The CCP4suite programs for crystallography Acta Crystallogr D BiolCrystallogr 50 760ndash763
Dodson KW Pinkner JS Rose T Magnusson G Hult-gren SJ and Waksman G (2001) Structural basis of theinteraction of the pyelonephritic Escherichia coli adhesin toits human kidney receptor Cell 105 733ndash743
Eley JG and Triumalashetty P (2001) In vitro assessmentof alkylglycosides as permeability enhancers AAPSPharmSciTech 2 (3) Article 19
Esnouf RM (1997) An extensively modified version of Mol-script that includes greatly enhanced coloring capabilitiesJ Mol Graph Model 15 132ndash134
Firon N Ofek I and Sharon N (1982) Interaction ofmannose-containing oligosaccharides with the fimbriallectin of Escherichia coli Biochem Biophys Res Commun105 1426ndash1432
Firon N Ofek I and Sharon N (1983) Carbohydrate spec-ificity of the surface lectins of Escherichia coli Klebsiellapneumoniae and Salmonella typhimurium Carbohydr Res120 235ndash249
Firon N Ofek I and Sharon N (1984) Carbohydrate-
binding sites of the mannose-specific fimbrial lectins ofEnterobacteria Infect Immun 43 1088ndash1090
Firon N Ashkenazis S Mirelman D Ofek I and SharonN (1987) Aromatic a-glycosides of mannose are powerfulinhibitors of the adherence of type 1 fimbriated Escherichiacoli to yeast and intestinal cells Infect Immun 55 472ndash476
Fukiya S Mizoguchi H Tobe T and Mori H (2004)Extensive genomic diversity in pathogenic Escherichia coliand Shigella strains revealed by comparative genomichybridization microarray J Bacteriol 186 3911ndash3921
Hooton TM and Stamm WE (1997) Diagnosis and treat-ment of uncomplicated urinary tract infection Infect DisClin North Am 11 551ndash581
Horovitz A and Levitzki A (1987) An accurate method fordetermination of receptor ligand and enzyme-inhibitor dis-sociation-constants from displacement curves Proc NatlAcad Sci USA 84 6654ndash6658
Hull RA Gill RE Hsu P Minshaw BH and Falkow S(1981) Construction and expression of recombinant plas-mids encoding type 1 and D-mannose-resistant pili from aurinary tract infection Escherichia coli isolate Infect Immun33 933ndash938
Hultgren SJ Porter TN Schaeffer AJ and Duncan JL(1985) Role of type 1 pili and effects of phase variation onlower urinary tract infections produced by Escherichia coliInfect Immun 50 370ndash377
Hung C-S Bouckaert J Hung DL Pinkner J WinbergC Defusco A et al (2002) Structural basis of tropism ofEscherichia coli to the bladder during urinary tract infec-tion Mol Microbiol 44 903ndash915
Jones TA Zou JY Cowan SW and Kjeldgaard M(1991) Improved methods for building protein models inelectron density maps and the location of errors in thesemodels Acta Crystallogr A 47 110ndash119
Justice SS Hung C-S Theriot JA Fletcher DAAnderson GG Footer MJ and Hulgren SJ (2004)Differentiation and developmental pathways of uropatho-genic Escherichia coli in urinary tract pathogenesis ProcNatl Acad Sci USA 101 1333ndash1338
Kallenius G Mollby R Svenson SB Windberg J Lun-dblud A Svenson S and Cedergen B (1980) The Pk
antigen as receptor for the haemagglutinin of pyelonephri-togenic Escherichia coli FEMS Microbiol Lett 8 297ndash302
Knight SD Berglund J and Choudhury D (2000) Bacte-rial adhesins structural studies reveal chaperone functionand pilus biogenesis Curr Opin Chem Biol 4 653ndash660
Langermann S and Ballou WR (2003) Development of arecombinant FimCH vaccine for urinary tract infectionsAdv Exp Med Biol 539 635ndash653
Langermann S Palaszynski S Barnhart M Auguste GPinkner JS Burlein J et al (1997) Prevention ofmucosal Escherichia coli infection by FimH-adhesin-basedsystemic vaccination Science 276 607ndash611
Langermann S Mollby R Burlein JE Palaszynski SRAuguste CG Defusco A et al (2000) Vaccination withFimH adhesin protects cynomolgus monkeys from coloni-zation and infection by uropathogenic Escherichia coli JInfect Dis 181 774ndash778
Larsson A Ohlsson J Dodson KW Hultgren SJ Nils-son U and Kihlberg J (2003) Quantitative studies of thebinding of the class II PapG adhesin from uropathogenic
454 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
Escherichia coli to oligosaccharides Bioorg Med Chem 112255ndash2261
Leffler H and Svanborg-Eden C (1980) Chemical identifi-cation of a glycosphingolipid receptor for Escherichia coliattaching to human urinary tract epithelial cells and agglu-tinating human erythrocytes FEMS Microbiol Lett 8 127ndash134
Lindhorst TK Kieburg C and Krallmann-Wenzel U(1998) Inhibition of the type 1 fimbriae-mediated adhesionof Escherichia coli to erythrocytes by multiantennary alpha-mannosyl clusters the effect of multivalency Glycoconj J15 605ndash613
Loris R Tielker D Jaeger KE and Wyns L (2003a)Structural basis of carbohydrate recognition by the lectinLecB from Pseudomonas aeruginosa J Mol Biol 331 861ndash870
Loris R Imberty A Beeckmans S Van Driessche ERead JS Bouckaert J et al (2003b) Crystal structureof Pterocarpus angolensis lectin in complex with glucosesucrose and turanose J Biol Chem 278 16297ndash16303
Lund B Lindberg F Marklund BI and Normark S(1987) The PapG protein is the a-D-galactopyranosyl-(1-4)-b-D-galactopyranose-binding adhesin of uropathogenicEscherichia coli Proc Natl Acad Sci USA 84 5898ndash5902
Min G Stolz M Zhou G Liang F Sebbel P StofflerD et al (2002) Localization of uroplakin Ia the urothelialreceptor for bacterial adhesin FimH on the six innerdomains of the 16 nm urothelial plaque particle J Mol Biol317 697ndash706
Miroux B and Walker JE (1996) Over-production of pro-teins in Escherichia coli mutant hosts that allow synthesisof some membrane proteins and globular proteins at highlevels J Mol Biol 260 289ndash298
Morris GM Goodsell DS Halliday RS Huey R HartWE Belew RK and Olson AJ (1998) Automateddocking using a Lamarckian genetic algorithm and anempirical binding free energy function J Comput Chem 191639ndash1662
Mulvey MA (2002) Adhesion and entry of uropathogenicEscherichia coli Cell Microbiol 4 257ndash271
Mulvey MA Lopez-Boado YS Wilson CL Roth RParks WC Heuser J and Hultgren SJ (1998) Induc-tion and evasion of host defenses by type 1-piliated uro-pathogenic Escherichia coli Science 282 1494ndash1497
Murshudov GN Vagin AA and Dodson EJ (1997)Refinement of macromolecular structures by the maxi-mum-likelihood method Acta Crystallogr D Biol Crystallogr53 240ndash255
Nagahori N Lee RT Nishimura S Pageacute D Roy Rand Lee YC (2002) Inhibition of adhesion of type 1 fim-briated Escherichia coli to highly mannosylated ligandsChembiochem 3 836ndash844
Navaza J (2001) Implementation of molecular replacementin AMoRe Acta Crystallogr D Biol Crystallogr 57 1367ndash1372
Neeser JR Koellreutter B and Wuersch P (1986)Oligomannoside-type glycopeptides inhibiting adhesion ofEscherichia coli strains mediated by type 1 pili preparationof potent inhibitors from plant glycoproteins Infect Immun52 428ndash436
Old DC (1972) Inhibition of the interaction between fimbrial
hemagglutinatinins and erythrocytes by D-mannose andother carbohydrates J Gen Microbiol 71 149ndash157
Orndorff PE Devapali A Palestrant S Wyse A EverettML Bollinger RR and Parker W (2004) Immunoglob-ulin-mediated agglutination of and biofilm formation byEscherichia coli K-12 require the type 1 pilus fiber InfectImmun 72 1929ndash1938
Oscarson S and Tedebark U (1995) Syntheses of deoxyanalogues of methyl 36-di-O-a-D-mannopyranosyl-a-D-mannopyranoside for studies of the binding site of con-canavalin A Carbohydr Res 278 271ndash287
Oscarson S and Tiden AK (1993) Syntheses of the octyland tetradecyl glycosides of 36-di-O-alpha-deuterium-mannopyranosyl-alpha-deuterium-mannopyranose and of34-di-O-alpha-deuterium-mannopyranosyl-alpha-deute-rium-mannopyranose ndash a new way for 24-di-O-protectionof mannopyranosides Carbohydr Res 247 323ndash328
Otwinowski Z and Minor W (1997) Processing of X-raydiffraction data collected in oscillation mode MethodsEnzymol 276 307ndash326
Powell HR (1999) The Rossmann Fourier autoindexingalgorithm in MOSFLM Acta Crystallogr D Biol Crystallogr55 1690ndash1695
Pratt LA and Kolter R (1998) Genetic analysis of Escher-ichia coli biofilm formation roles of flagella motility chemo-taxis and type I pili Mol Microbiol 30 285ndash293
Remaut H and Waksman G (2004) Structural biology ofbacterial pathogenesis Curr Opin Struct Biol 14 161ndash170
Ronald AR Nicolle LE Stamm E Krieger J WarrenJ Schaeffer A et al (2001) Urinary tract infection inadults research priorities and strategies Int J AntimicrobAgents 17 343ndash348
Sanner MF Olson AJ and Spehner JC (1996)Reduced surface an efficient way to compute molecularsurfaces Biopolymers 38 305ndash320
Sauer FG Futterer K Pinkner JS Dodson KW Hult-gren SJ and Waksman G (1999) Structural basis ofchaperone function and pilus biogenesis Science 2851058ndash1061
Sauer FG Knight SD Waksman GW and HultgrenSJ (2000) PapD-like chaperones and pilus biogenesisSemin Cell Dev Biol 11 27ndash34
Sauer FG Pinkner JS Waksman G and Hultgren SJ(2002) Chaperone priming of pilus subunits facilitates atopological transition that drives fiber formation Cell 111543ndash551
Schembri MA Hasman H and Klemm P (2000)Expression and purification of the mannose recognitiondomain of the FimH adhesin FEMS Microbiol Lett 188147ndash151
Schembri MA Kjaergaard K and Klemm P (2003) Glo-bal gene expression in Escherichia coli biofilms Mol Micro-biol 48 253ndash267
Schilling JD Mulvey MA Vincent CD Lorenz RGand Hultgren SJ (2001) Bacterial invasion augments epi-thelial cytokine responses to Escherichia coli through alipopolysaccharide-dependent mechanism J Immunol166 1148ndash1155
Silhavy TJ Szmelcman S Boos W and Schwartz M(1975) On the significance of the retention of ligand byprotein Proc Natl Acad Sci USA 72 2120ndash2124
A novel class of FimH high-affinity ligands 455
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
Svanborg C and Godaly G (1997) Bacterial virulence inurinary tract infection Infect Dis Clin North Am 11 513ndash529
Thanassi DG Saulino ET and Hultgren SJ (1998) Thechaperoneusher pathway a major terminal branch of thegeneral secretory pathway Curr Opin Struct Biol 1 223ndash231
Thankavel K Madison B Ikeda T Malaviya R ShahAH Arumugen PM and Abraham SN (1997) Local-ization of a domain in the FimH adhesin of Escherichia colitype 1 fimbriae capable of receptor recognition and use ofa domain-specific antibody to confer protection againstexperimental urinary tract infection J Clin Invest 1001123ndash1126
Vagin A and Teplyakov A (1997) MOLREP an automatedprogram for molecular replacement J Appl Crystallogr 301022ndash1025
Wu XR Sun TT and Medina JJ (1996) In vitro bindingof type 1-fimbriated Escherichia coli to uroplakins 1a and
1b relation to urinary tract infections Proc Natl Acad SciUSA 93 9630ndash9635
Xia Y Gally D Forsman-Semb K and Uhlin BE (2000)Regulatory cross-talk between adhesin operons in Escher-ichia coli inhibition of type 1 fimbriae expression by thePapB protein EMBO J 19 1450ndash1457
Zafriri D Ofek I Adar R Pocino M and Sharon N(1989) Inhibitory activity of cranberry juice on adherenceof type-1 and type-P fimbriated Escherichia coli to eukary-otic cells Antimicrob Agents Chemother 33 92ndash98
Zavialov AV Berglund J Pudney AF Fooks LJ Ibra-him TM MacIntyre S and Knight SD (2003) Structureand biogenesis of the capsular F1 antigen from Yersiniapestis preserved folding energy drives fiber formation Cell113 587ndash596
Zhou G Mo WJ Sebbel P Min G Neubert TA Glock-shuber R et al (2001) Uroplakin Ia is the urothelial recep-tor for uropathogenic Escherichia coli evidence from invitro FimH binding J Cell Sci 114 4095ndash4103
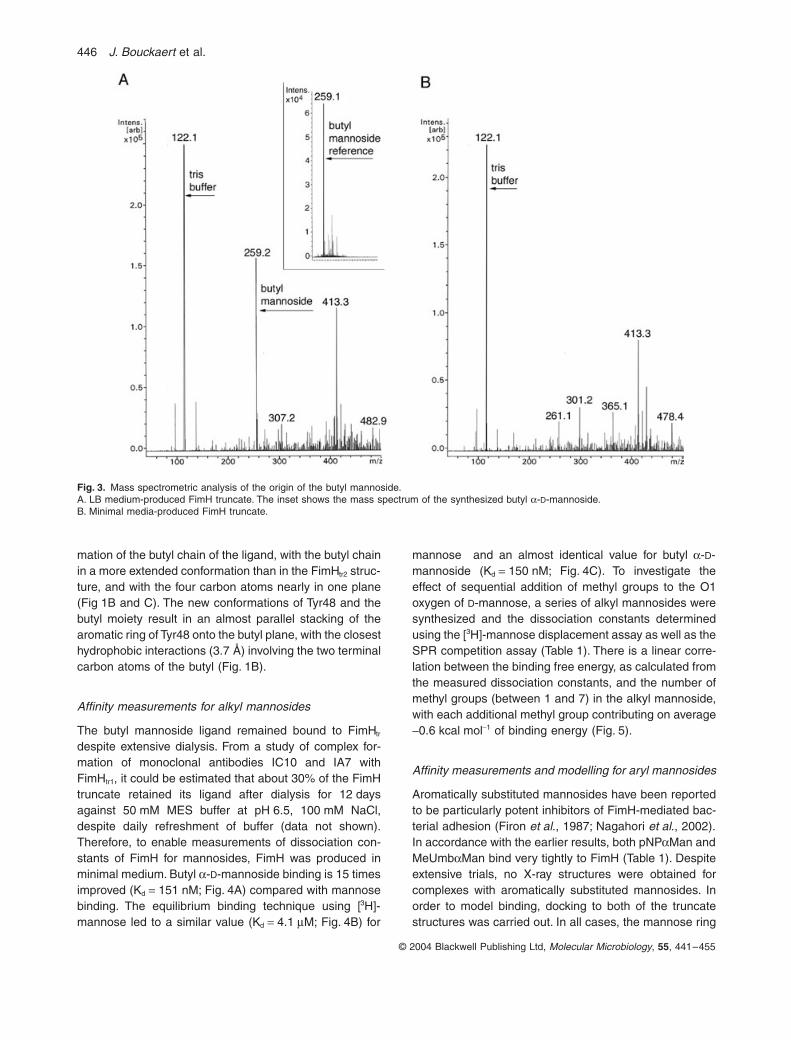
446 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
mation of the butyl chain of the ligand with the butyl chainin a more extended conformation than in the FimHtr2 struc-ture and with the four carbon atoms nearly in one plane(Fig 1B and C) The new conformations of Tyr48 and thebutyl moiety result in an almost parallel stacking of thearomatic ring of Tyr48 onto the butyl plane with the closesthydrophobic interactions (37 Aring) involving the two terminalcarbon atoms of the butyl (Fig 1B)
Affinity measurements for alkyl mannosides
The butyl mannoside ligand remained bound to FimHtr
despite extensive dialysis From a study of complex for-mation of monoclonal antibodies IC10 and IA7 withFimHtr1 it could be estimated that about 30 of the FimHtruncate retained its ligand after dialysis for 12 daysagainst 50 mM MES buffer at pH 65 100 mM NaCldespite daily refreshment of buffer (data not shown)Therefore to enable measurements of dissociation con-stants of FimH for mannosides FimH was produced inminimal medium Butyl a-D-mannoside binding is 15 timesimproved (Kd = 151 nM Fig 4A) compared with mannosebinding The equilibrium binding technique using [3H]-mannose led to a similar value (Kd = 41 mM Fig 4B) for
mannose and an almost identical value for butyl a-D-mannoside (Kd = 150 nM Fig 4C) To investigate theeffect of sequential addition of methyl groups to the O1oxygen of D-mannose a series of alkyl mannosides weresynthesized and the dissociation constants determinedusing the [3H]-mannose displacement assay as well as theSPR competition assay (Table 1) There is a linear corre-lation between the binding free energy as calculated fromthe measured dissociation constants and the number ofmethyl groups (between 1 and 7) in the alkyl mannosidewith each additional methyl group contributing on average-06 kcal mol-1 of binding energy (Fig 5)
Affinity measurements and modelling for aryl mannosides
Aromatically substituted mannosides have been reportedto be particularly potent inhibitors of FimH-mediated bac-terial adhesion (Firon et al 1987 Nagahori et al 2002)In accordance with the earlier results both pNPaMan andMeUmbaMan bind very tightly to FimH (Table 1) Despiteextensive trials no X-ray structures were obtained forcomplexes with aromatically substituted mannosides Inorder to model binding docking to both of the truncatestructures was carried out In all cases the mannose ring
Fig 3 Mass spectrometric analysis of the origin of the butyl mannosideA LB medium-produced FimH truncate The inset shows the mass spectrum of the synthesized butyl a-D-mannosideB Minimal media-produced FimH truncate
A novel class of FimH high-affinity ligands 447
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
docks in the binding pocket in a conformation very closeto the crystallographically observed mannose conforma-tion (Fig 6) while the aromatic substituent interacts withthe tyrosine gate The different conformation of Tyr48 in
our two FimH truncate structures leads to different pre-dicted interactions between the aromatic aglycon andTyr48 and Tyr137 Docking to the FimHtr2 structureresulted in somewhat tighter interactions between thetyrosine gate and the aglycon and hence in somewhatlower docking energies Calculated FimHtr2 docking ener-gies of Edoc = -104 kcal mol-1 for pNPaMan (Kd = 46 nM)and Edoc = -109 kcal mol-1 for MeUmbaMan (Kd = 20 nM)are in good agreement with the experimental binding data(Table 1) The docked umbelliferyl ring system is inserteddeep into the tyrosine gate in the FimHtr2 conformationwith the carbonyl group within hydrogen bonding distanceto Thr51 at the back of the gate (Fig 6A) Docking to theFimHtr1 structure instead stacks the umbelliferyl ring face-to-face with the Tyr48 ring (not shown) The phenyl ring ofpNPaMan docks similarly to both our structures It doesnot quite reach in between the tyrosine side-chains butinstead hydrogen bonds with its nitro group to the hydroxyloxygen of Tyr137 (Fig 6B)
Discussion
Most lectins bind naturally occurring carbohydrates withlow affinities often in the millimolar range for monosac-charides and micromolar range for oligosaccharides Thiswas recently exemplified to also hold true for theadhesins of F17 fimbriae (Kd = 12 mM for the F17a-G-N-acetylglucosamine interaction Buts et al 2003) and Ppili (Kd = 84 mM for the PapGII-globotetraoside interactionLarsson et al 2003) The recent crystal structure of FimHin complex with mannose illustrated that FimH almostcompletely envelops the monosaccharide in a deeppocket and involves it in an intense hydrogen bondingnetwork suggestive of an unusual lectin exerting highaffinities (Hung et al 2002)
Fig 4 FimH binding experimentsA Surface plasmon resonance competition experiment of FimHtr1 binding to monoclonal antibody IC10 Fab fragments with butyl a-D-mannosideB Equilibrium binding of [3H]-mannose to FimHtr2C Equilibrium displacement measurements of butyl a-D-mannoside to the FimHtr2 [3H]-mannose complex
log[Butyl a-D-mannoside] (M)
cpm
cpm
Butyl a-D-mannoside (mM)
Mannose (mM)
A
B
C
ndash64
ndash66
ndash68
ndash70
ndash72
ndash74
ndash76
ndash78
ndash80
ndash82
ndash8 ndash7 ndash6 ndash5 ndash4 ndash3
log
[Fim
H]
M
Fig 5 Linear free energy relationship for the alkyl chain on mannose A linear fit was made using 1 up to 7 methyl groups with the program MICROCAL ORIGIN 50
Number of carbon atoms0 2 4 6 8
ndash75
ndash80
ndash90
ndash95
ndash100
ndash105
ndash110
ndash115
ndash85
D G
(kc
al m
olndash1
)
slopecorrelation
ndash064ndash099
+ndash 003+ndash 018
448 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
The measured affinity confirm the predictions that FimHbinds the monosacharide mannose exceptionally well(Kd = 23ndash41 mM) To investigate the contribution of thedifferent hydroxyl groups of mannose in the interactionwith FimH the binding of a series of hexoses to butylmannoside-free FimH was examined using SPR (Table 2)The effect of deoxygenation of the O2 hydroxyl group(methyl 2-deoxy-a-D-mannopyranoside) is clearly lessdestructive (100-fold reduction) than its equatorial positionas in glucose O2 is the only hydroxyl group of mannosethat interacts indirectly with the FimH pocket via a con-served water molecule (Hung et al 2002) Glucose onlydisplays a weak affinity for FimH about 4000 times lowerthan methyl a-D-mannoside Repositioning of both the O2and O4 hydroxyl groups as in galactose abolishes sugarbinding by FimH Similar to these results the inhibitoryconcentrations for bacterial adherence of methyl a-D-glucoside and galactose derivatives have been estimatedto be respectively 104-fold and 105-fold weaker than formethyl a-D-mannoside (Nagahori et al 2002)
In stark contrast with the other monosaccharides fruc-tose has a binding affinity only about 15-fold less thanmannose for FimH (Table 2) The decrease in affinity for
fructose relative to mannose agrees with studies of inhi-bition of adherence of type 1 piliated E coli by fructosein which fructose was found to be the active compound infruit juices (present at about 5) (Zafriri et al 1989)Fructose binding to FimH is stronger than the binding ofthe physiological globotetraoside receptor to PapGII theother well-characterized adhesin of UPEC strains (Lars-son et al 2003) The pyranose form of fructose is verywell superimposable onto mannose in the FimH bindingsite where similar interactions are established with FimHas for mannose except for the loss of the anomeric O1and an extra hydroxyl group (O2) on the C5 position ofmannose Probably however both changes of fructopyr-anoside compared with mannopyranoside could be easilyaccommodated by FimH The mannose O1 does notmake direct interactions within 45 Aring of the FimH bindingpocket (Fig 1B) The extra O2 group of fructose would notclash or interfere significantly with the binding pocket butit would make a 27 Aring van der Waals contact with the Ile52side-chain under the condition that the Ile52 side-chaindoes not move The crystal structure of the Pseudomonasaeruginosa lectin LecB also showed that fructopyranosebut not fructofuranose can substitute for mannose (Loris
Fig 6 Stereo diagrams of docked MeUm-baMan (A) and pNPaMan (B) in the FimH bind-ing site The crystallographically determined position of a-D-mannose is shown in orange for comparison
A novel class of FimH high-affinity ligands 449
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
et al 2003a) Hydrogen bonds of the more specificfucose complex are replaced by van der Waals interac-tions involving a hydroxyl group in the complexes with thelower-affinity binders mannose and fructopyranose as forthe proposed fructopyranoside binding to FimH
The fructose-containing disaccharides sucrose andturanose do not have an inhibitory effect and bind withaffinities similar to glucose Sucrose is a non-reducingsugar that has fructose locked in its furanose form Tura-nose on the other hand allows the interconversion of b-D-fructofuranose and b-D-fructopyranose eg both formsoccur in complex with a plant lectin from Pterocarpusangolensis (Loris et al 2003b) It is predictable howeverfrom the crystal structures that the glycosidic linkage ofglucose with C3 of fructose equivalent in position to C4of mannose would be disallowed in the tight FimH pocketSucrose and turanose thus most probably interact with theFimH mannose binding pocket via their glucose moietyresulting in similar low affinities
Our mass spectrometry and chemical analyses con-firms the crystallographic identification of the ligand boundto the FimH lectin domains as butyl a-D-mannoside(Figs 1B and 2) The mannose ring of the butyl a-D-mannoside coincides with the mannose bound to theFimCFimH complex (Hung et al 2002) and makes thesame interactions The four-carbon alkyl tail exits towardsthe back wall (loop b4ndashb5 and b10ndashb11) of the pocket(Fig 1A and B) making hydrophobic contacts to Tyr48Tyr137 and Ile52 The mass spectrometry data also showthat the bound butyl a-D-mannoside is present in LBmedium but absent from minimal medium Hence thesource of the alkyl mannoside is possibly the yeast extractused in preparation of LB medium Although we do notwant to speculate about the significance of this findingalkyl mannoside production by yeast has not beenreported and might warrant investigation
The butyl mannoside was retained by FimH despiteextensive dialysis The phenomenon of ligand retention byprotein was described as early as 1975 for periplasmicbinding proteins (Silhavy et al 1975) Applied to dialysisof butyl mannoside away from FimH grown in richmedium half-lives of up to 30 h can be estimated whichis far beyond the duration of a normal buffer exchangedialysis experiment In the present case the retainedligand does not appear to seriously affect [3H]-mannosedisplacement equilibrium binding experiments as wecould detect no significant differences in dissociation con-stants obtained using FimHtr2 produced in LB or minimalmedia (data not shown) In contrast proper SPR bindingcurves could not be obtained when butyl mannoside occu-pied the binding site Hence the displacement techniquemay be of more general use to obtain binding constantsfor proteins that tend to inadvertently retain ligands whichis a common trait of lectins
Surface plasmon resonance as well as equilibrium bind-ing and displacement measurements using [3H]-mannosewere carried out to obtain dissociation constants for a-D-mannose and a linear series of alkyl-substituted manno-sides to FimH (Table 1) Both equilibrium binding experi-ments show a very good agreement in their measurementof the carbohydrate affinity of the adhesin FimH bindsbutyl a-D-mannoside 15ndash30 times stronger than a-D-mannose The importance of the added hydrophobiccontacts is clearly demonstrated by the binding of alkylmannosides (Table 1) The best binding alkyl mannosidesbind a few hundred times stronger than mannose equiv-alent in affinity to the most tightly binding aromaticallysubstituted mannosides for FimH (Firon et al 1987) andof mannose dendrimers for type 1 piliated E coli (Fironet al 1984 1987 Nagahori et al 2002)
Interestingly the butyl moiety takes on significantlydifferent conformations in FimHtr1 and FimHtr2 (Fig 1C)indicating that intrinsic conformational flexibility in theaglycon is allowed Indeed in spite of the larger flexibilityexpected for longer alkyl mannosides the binding energydecreases linearly with increasing length of the alkyl chainbetween one and seven carbon atoms (Fig 5) The affinityreaches its maximum for mannose with a seven-carbonalkyl and SPR measurements suggest that the affinitydecreases from eight carbon atoms on Evidently theeighth carbon atom goes beyond the hydrophobic interac-tion surface and protein edge which is penalized with adecrease in affinity We propose that the affinity increaseresults from an increased interaction surface with thehydrophobic gateway Alkyl aglycons fit well in the tyrosinegate and as observed in the two crystal structures in atleast two different conformations that depend on theconformation of Tyr48 This flexibility may indicate thataglycons can interact rather non-specifically with thehydrophobic gateway nevertheless resulting in a 100- to1000-fold increase in affinity
FimH binding to ligands other than just mannose ispossibly of importance in terms of the different environ-ments that E coli will encounter ndash the urinary tract gas-trointestinal tract and in biofilms Alkyl mannosides havenot been previously recognized as strong binders to FimHAs they are easily synthesized and highly soluble in waterthey may be interesting as potential blocking agents forFimH-mediated adhesion Moreover they have beenshown to be good drug vehicles themselves acting aspermeability enhancers on tight junctions (Eley and Triu-malashetty 2001) As such they might be able to pene-trate into deeper cell layers of the epithelium to alsoprevent those bacteria that form a quiescent reservoirfrom causing recurrent infections (Mulvey et al 1998Justice et al 2004)
On the other hand the two aromatically substitutedmannosides pNPaMan and MeUmbaMan are known to
450 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
be particularly potent inhibitors of FimH-mediated bacte-rial adhesion (Firon et al 1987 Nagahori et al 2002)Based on adhesion inhibition studies using type 1 piliatedE coli and guinea pig ileal epithelial cells the relativeinhibitory potential of pNPaMan and MeUmbaMan havebeen estimated to be about 70 and 600 times strongerrespectively than that of methyl a-D-mannose (Fironet al 1987) Our measurements indicate an affinity ofFimH for pNPaMan and MeUmbaMan between 50 and90 times and between 110 and 200 times higher respec-tively than for methyl a-D-mannose in good agreementwith the adhesion inhibition results (Firon et al 1987)Dissociation constants predicted using AutoDock3 (Morriset al 1998) are similar to experimental values Thecomputational docking predicts that tight binding of thearomatically substituted mannosides pNPaMan andMeUmbaMan is accomplished similarly as for butyl a-D-mannoside with the aglycon interacting with the tyrosinegate
This is the first systematic study providing molecularbinding data for the FimH adhesin the prerequisite forcolonization by UPEC (Langermann and Ballou 2003)We also show crystallographic evidence that the lectinor receptor binding domain of FimH is a stable andfunctional entity Its structure is virtually identical to thesame domain in the FimCFimH complex indicating thata truncate corresponding to the lectin domain could pro-vide a good alternative for the development of a UTIvaccine (Langermann et al 1997 Langermann and Bal-lou 2003) However progress of this interesting UTI vac-cine candidate into clinical trials has not been reportedThe binding affinities of FimH at the molecular levelrelate convincingly well with the inhibitory concentrationsused in previous bacterial adhesion assays thus endors-ing the direct and conclusive correlation of FimH withbacterial adherence Alkyl mannosides have nanomolaraffinities for FimH which may overcome the significantobstacle of low affinity generally encountered in attemptsto develop carbohydrate-based drugs which interferewith microbial attachment to host cell surface glycocon-jugates New anti-adhesive drugs could be based onthese leads to allow specific interaction and targeting tothe urinary tract in order to prevent anticipated andrecurrent infections
Experimental procedures
Strains and constructs
FimH proteins from two different E coli strains the UPECstrain J96 (Hull et al 1981) and the E coli K-12 strainMG1655 (Blattner et al 1997) were used in this study TheFimH adhesins encoded by these two E coli strains areidentical FimHtr1 (residues 1ndash158 of UPEC J96 FimH) wasexpressed from plasmid pMMB91 transformed into E coli
C600 cells FimHtr2 (residues 1ndash158 of E coli MG1655FimH + carboxy-terminal 6-His tag) was expressed fromplasmid pPKL241 (Schembri et al 2000) transformed into Ecoli host strain HB101
Purification and crystallization
FimHtr1 Bacteria were grown in LB medium at 37infinC inducedat OD600 = 08 with 1 mM IPTG and harvested 2ndash3 h laterThe cells were resuspended in 20 mM Tris-HCl pH 80 5 mMEDTA and 30 sucrose and the periplasmic content wasextracted by osmotic shock upon 10-fold dilution with 20 mMTris-HCl pH 80 Purification of FimHtr1 was performed by ionexchange chromatography with MonoS at pH 45 followed byMonoQ (Amersham Biosciences) at pH 8 FimHtr1 was crys-tallized using vapour diffusion in hanging drops composed of3 ml of the precipitant solution (01 M Hepes pH 75 12PEG 6000 and 5 vv methyl-24-pentanediol) and 3 ml of12 mg ml-1 protein solution in 20 mM Tris-HCl pH 80 with150 mM NaCl
FimHtr2 Cells were grown in LB medium containing50 mg ml-1 ampicillin at 37infinC At OD600 = 06 the cells wereinduced with 5 mM IPTG Cells were harvested 2 h afterinduction and the periplasmic content was extracted Theprotein was purified on a Pharmacia HiTrap Chelating HP5 ml column (Pharmacia) loaded with Ni-chloride and elutedwith a sharp 0ndash500 mM imidazole gradient Fractions con-taining FimHtr2 were pooled dialysed against 50 mM sodiumacetate pH 525 and loaded onto a Mono S HR 8 ml columnThe protein was eluted with a 0ndash500 mM NaCl gradient anddialysed overnight against 20 mM MES pH 65 before crys-tallization Crystallization was performed at room tempera-ture by the vapour diffusion method with hanging dropsmade by mixing of 2 ml of 15 mg ml-1 protein solution with 2 mlof the precipitant solution (65 methyl-24-pentanediol100 mM cacodylate pH 65) Microseeding was employed tooptimize crystal quality and reproducibility Crystals suitablefor X-ray data collection generally appeared within 48 h aftermicroseeding
Structure determination
X-ray data for FimHtr1 were collected from a single crystalto 17 Aring resolution at the Advanced Photon SourceArgonne National Laboratory USA The crystal was flash-cooled to 100 K in the precipitant solution complementedwith 25 glycerol The data were processed and scaledusing the HKL2000 package (Otwinowski and Minor 1997)(Table S1 in Supplementary material) Data for FimHtr2
were collected from a single crystal under liquid-nitrogencryo conditions at 100 K at beamline ID29 ESRF Greno-ble France to 21 Aring Bragg spacing FimHtr2 data were pro-cessed and reduced using MOSFLM (Powell 1999) andscaled using SCALA (Collaborative computational project1994) (Table S1 in Supplementary material) The structuresof FimHtr1 and FimHtr2 were solved by molecular replace-ment using Amore (Navaza 2001) and MOLREP (Vaginand Teplyakov 1997) respectively with the FimH lectindomain from the FimCFimH complex (Choudhury et al
A novel class of FimH high-affinity ligands 451
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
1999) (PDB id 1QUN) as the search model Positional bulksolvent and isotropic temperature factor refinement wasperformed using CNS (Bruumlnger et al 1998) and REFMAC5(Murshudov et al 1997) respectively Model adjustmentand inspection were performed using Turbo-Frodo and O(Jones et al 1991) graphics programs respectively Refine-ment statistics are given in Table S1 Molecular graphicsillustrations were generated with Bobscript (Esnouf 1997)and Pymol 090 (DeLano Scientific LLC 1998ndash2003 httppymolsourceforgenet)
Mass spectrometry
The purified FimHtr1 protein (300 mg) that had been used forcrystallization was freeze-dried taken up in 100 ml of pyridineand 100 ml of acetic anhydride and incubated overnight atroom temperature in the dark for peracetylation The samplewas dried under nitrogen and taken up in chloroform (400 ml)The chloroform phase was extracted six times with waterdried and taken up in 200 ml of 50 aqueous methanol A1 ml aliquot was applied to a nano-scale RP column(75 mm yen 100 mm PepMap DionexLC Packings) using anUltimate nano-LC system (LC Packings) which was equili-brated with eluent A (H2Oacetonitrile 955 vv containing01 formic acid) at a flow rate of 100 nl min-1 After injectingthe sample the column was run isocratically for 5 min fol-lowed by a linear gradient to 100 eluent B (H2Oacetonitrile2080 vv containing 01 formic acid) in 20 min and a finalwash with 100 B for 5 min The system was directly coupledto an Esquire 3000 ESI-IT-MS (Bruker Daltonik) equippedwith an online nanospray source operating in the positive-ionmode For electrospray (900ndash1200 V) capillaries (360 mmOD 20 mm ID with 10 mm opening) from New Objective wereused The solvent was evaporated at 150infinC with a nitrogenstream of 6 l min-1
To verify the suspected source of the bound butyl manno-side mass spectrometry was performed on samples ofFimHtr2 grown in either LB or minimal M9 medium Proteinsamples were dialysed against water overnight before massspectrometry in order to reduce background from the Trisbuffer Samples were diluted to a concentration in the submi-cromolar range Electrospray mass spectra were recorded indirect inlet mode (50 aqueous methanol solutions) on aBruker Daltonics Esquire-LC (Billerica)
Carbohydrate constituent analysis
Samples were hydrolysed in 100 ml of 4 M aqueous trifluoro-acetic acid (Sigma) at 100infinC for 4 h and dried under astream of nitrogen Monosaccharides were converted intotheir anthranilic acid derivatives by reductive aminationresolved by RP-HPLC and detected by fluorescence (Anu-mula 1994)
Carbohydrates for binding studies
[3H]-mannose was obtained from Amersham Methyl a-D-mannoside glucose galactose fructose sucrose turanosep-nitrophenyl a-D-mannopyranoside (pNPaMan) and 4-methylumbelliferyl a-D-mannopyranoside (MeUmbaMan)
were obtained from Sigma Monomannosylated bovineserum albumin (BSA-Man) was kindly provided by MedIm-mune We synthesized methyl 2-deoxy-a-D-mannopyrano-side (Oscarson and Tedebark 1995) Alkyl mannosides weresynthesized through silver triflate-promoted couplings of thecorresponding alcohol with 2346-tetra-O-benzoyl-a-D-mannopyranosyl bromide followed by Zempleacuten deacylationof the obtained protected alkyl mannosides according to theprocedure reported for the octyl and tetradecyl mannosides(Oscarson and Tiden 1993)
Binding studies
Competition experiments using SPR To obtain butyl man-noside-free FimHtr1 the FimH truncate was overproducedwith a carboxy-terminal 6-His tag after transformation into Ecoli C43 (DE3) (Miroux and Walker 1996) and the bacteriagrown in minA minimal medium supplemented with the 20amino acids (40 mg l-1) thiamin (2 mg l-1) and biotin (2 mgl-1) Purification of the His-tagged protein was performedusing Ni-NTA affinity chromatography (Qiagen) in 50 mM Tris-HCl pH 8 and 300 mM NaCl Elution was in the same solutioncomplemented with 500 mM imidazole Subsequently pooledand concentrated FimH fractions were gel filtrated over aSuperdex G75 column (Pharmacia) in 20 mM Tris-HCl pH 8
We used Fab fragments of a monoclonal antibody againstthe mannose-binding site of FimH to determine the solutionaffinity of FimHndashcarbohydrate interactions The monoclonalantibody IC10 was produced by a mouse hybridoma cell lineat Medimmune and its Fab fragments were prepared andpurified The Fabs were covalently immobilized via aminecoupling at 1000 Resonance Units (1000 pg ligand mm-2) inflowcell Fc2 on a CM5 biasensor chip (BIAapplications Hand-book Biacore AB Uppsala Sweden) Flowcell Fc1 was usedas the reference surface and was treated in the same wayas Fc2 except that it was blocked immediately after activationThe immobilization buffer was 100 mM sodium acetatepH 50 with 100 mM NaCl All SPR measurements were per-formed on a Biacore3000trade
The kinetic constants of binding of FimH to the immobilizedantibody were measured with different concentrations ofFimH (2000 nM 1000 nM 500 nM 250 nM 125 nM625 nM 312 nM 156 nM 78 nM) in running buffer [phos-phate-buffered saline (PBS) with 0005 surfactant P20 and3 mM EDTA] on both flowcells Fc1 and Fc2 simultaneouslyat a flow rate of 30 ml min-1 and at 298 K Complete dissoci-ation of FimH was performed with running buffer beforestarting a new binding cycle For all measurements the asso-ciation time was 3 min and the dissociation time was 30 minAll binding cycles were performed in duplicate including azero concentration cycle of FimH (injection of running buffer)All analyses were performed with the BIAeval software ALangmuir binding isotherm with a 11 stoichiometry was fittedto the data from both the association phase and the dissoci-ation phase after subtraction of the reference and zero con-centration data The kinetic constants ka and kd and themaximal binding Rmax were obtained and the dissociationconstant Kd was calculated
The equilibrium dissociation constants Kd of FimHndashsaccha-ride interactions were determined in a competition experi-ment using the same chip as for the previous experiment
452 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
As a control the Fabs of monoclonal antibody IA7 as wellas BSA-Man were also immobilized Samples containing afixed concentration of FimH were used (close to the Kd of theFimHndashantibody interaction) in combination with varying con-centrations of saccharide First 10-fold dilutions of the sac-charide solution were used to determine the concentrationrange for binding of the saccharide to FimH A Langmuirbinding isotherm with a 11 stoichiometry was fitted to thedata using the kinetic constants and Rmax from the foregoingexperiment to obtain the concentrations of FimH that werefree ([FimH]free) to bind the antibody immobilized on the chipSecond the concentration range of the saccharide wasextended and adapted to ensure accurate fitting and theequilibrium binding constant of the FimHndashsaccharide interac-tion was derived from the curve of [FimH]free against concen-tration of saccharide Every measurement was repeated atleast twice including testing different protein batches andwhenever possible also different saccharide stock solutions(typically 200 mM)
Equilibrium binding studies using radioactively labelledmannose Mannose-free FimHtr2 was produced inM9 minimal medium and purified as described above forFimHtr2 Alkyl mannosides were dissolved in double distilledwater pNPaMan and MeUmbaMan (6 mg each) were dis-solved in 20 ml of DMSO and diluted to 20 mM using doubledistilled water Binding experiments were performed at 310 Kusing six different concentrations of [3H]-a-D-mannose (finalconcentrations 435 mM 290 mM 193 mM 129 mM 86 mMand 57 mM) Protein (180 ml) at a concentration of about500 nM was mixed with 20 ml of the radioactive ligand [3H]-a-D-mannose and incubated at 37degC for 20 min To separatefree ligand from bound the mixture was rapidly filtratedthrough a Protran BA 85 cellulose nitrate filter (Schleicherand Schuell) and washed once with 1 ml of ice-cold 1yen PBSFilter-bound radioactivity was measured by scintillation spec-trometry within 24 h Displacement experiments were per-formed using six different concentrations of inhibitor (finalconcentrations in the range 00ndash435 mM) in the presence of435 mM [3H]-a-D-mannose Twenty microlitres of radioactiveligand 20 ml of inhibitor and 160 ml of protein (500 nM) weremixed and the experiments were performed as describedabove All experiments were performed in duplicates and thedata presented are mean values of at least two independentmeasurements For the determination of Kd for a-D-mannosean hyperbolic curve (y = P1x(P2 + x) where P1 is an arbitraryscale factor and P2 = Kd) was fitted to the data For the dis-placement experiments the curve y = P1(P2 + x) where P1
is an arbitrary scale factor and P2 is the concentration of theinhibitor displacing 50 of the labelled ligand [I]05 was usedinstead To calculate the inhibitor dissociation constant (KI)the Cheng and Prusoff equation [KI = [I]05[L]KL + 1) KL is theconstant of dissociation for the ligand] (Cheng and Prusoff1973) was used when both the concentration of the radio-active ligand (L) and the displacing agent (I) are in excessover the protein (LT gtgt PT IT gtgt PT T indicates total concen-tration) For the two strong inhibitors pNPaMan and MeUm-baMan where IT was not in excess over PT the equationof KI = IT[(1 - Y)Y(LTKL) - 1] - PTKLYLT where Y is thefraction of the ligand bound in presence of the inhibitor wasused instead (Horovitz and Levitzki 1987)
Computational docking
Computational docking studies were performed usingAutoDock3 (Morris et al 1998) In AutoDock3 the proteinis represented by a grid of affinity potentials Hence theprotein conformation is not allowed to change during thedocking procedure Flexibility of ligands is taken intoaccount by allowing rotation around flexible torsion anglesThe docked energies are calculated as the sum of the inter-molecular interaction energy and the internal energy of theligand The search for the best interaction energy was car-ried out by means of the Lamarckian genetic algorithmLigand structures were obtained from the Protein DataBank and prepared for docking by adding hydrogens andcharges using AutoDockTools (Sanner et al 1996) Eachsimulation consisted of 100 independent runs with a popu-lation size of 200 500 generations and a maximum of25 000 000 energy evaluations Solutions were rankedbased on their docking energies and similar solutions wereclustered (cluster cut-off rmsd lt 1 Aring) For both ligandsthe top solutions were evaluated using the graphics pro-gram O (Jones et al 1991)
Acknowledgements
We thank Andrej Zavialov and Maringhns Ehrenberg for help withequilibrium binding experiments and Thomas Norberg forassistance with mass spectrometry Anthony DeFusco fromMedimmune provided us with mannosylated bovine serumalbumin We are grateful to Fred Sauer for assistance withdata collection Thanks to Maor Bar-Peled of the CCC at theUniversity of Georgia Athens USA for his suggestion toexpress FimH in minimal medium We acknowledge the useof beamline 19BM of the Structural Biology Center at theAdvanced Photon Source Argonne National LaboratoryChicago IL USA and of beamline ID29 ESRF GrenobleFrance JB is a postdoctoral fellow of the Fonds voor Weten-schappelijk Onderzoek-Vlaanderen which also supportedthe BiaCore3000 instrument (FWONL35) This work was sup-ported by grants from the Swedish Research Council andfrom the Strategic Research Foundation (Structural BiologyNetwork) to SDK
Supplementary material
The following material is available from httpwwwblackwellpublishingcomproductsjournalssuppmatmmimmi4415mmi4415smhtmTable S1 Crystallographic data and refinement statistics
References
Anumula KR (1994) Quantitative determination ofmonosaccharides in glycoproteins by high-performanceliquid chromatography with highly sensitive fluorescencedetection Anal Biochem 220 275ndash283
Bahrani-Mougeot FK Buckles EL Lockatell CV HebelJR Johnson DE Tang CM and Donnenberg MS(2002) Type 1 fimbriae and extracellular polysaccharidesare preeminent uropathogenic Escherichia coli virulence
A novel class of FimH high-affinity ligands 453
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
determinants in the murine urinary tract Mol Microbiol 451079ndash1093
Berglund J and Knight SD (2003) Structural basis forbacterial adhesion in the urinary tract Glycobiol Med 53533ndash52
Blattner FR Plunkett G Bloch CA Perna NT BurlandV Riley M et al (1997) The complete genome sequenceof Escherichia coli K-12 Science 277 1453ndashbullbull
Bloch CA Stocker BA and Orndorff PE (1992) A keyrole for type 1 pili in enterobacterial communicability MolMicrobiol 6 697ndash701
Bork P Holm L and Sander C (1994) The immunoglob-ulin fold ndash structural classification sequence patterns andcommon core J Mol Biol 242 309ndash320
Brinton CC (1959) Non-flagellar appendages of bacteriaNature 183 782ndash786
Bruumlnger AT Adams PD Clore GM DeLano WLGros P Grosse-Kunstleve RW et al (1998) Crystallog-raphy and NMR system a new software suite for macro-molecular structure determination Acta Crystallogr D BiolCrystallogr 54 905ndash921
Buchanan K Falkow S Hull RA and Hull SI (1985)Frequency among Enterobacteriaceae of the DNA-sequences encoding type-1 pili J Bacteriol 162 799ndash803
Buts L Bouckaert J De Genst E Loris R Oscarson SLahmann M et al (2003) The fimbrial adhesin F17-G ofenterotoxigenic Escherichia coli has an immunoglobulin-like lectin domain that binds N-acetylglucosamine MolMicrobiol 49 705ndash715
Cheng Y and Prusoff WH (1973) Relationship betweeninhibition constant (K1) and concentration of inhibitor whichcauses 50 per cent inhibition (I50) of an enzymatic reac-tion Biochem Pharmacol 22 3099ndash3108
Choudhury D Thompson A Stojanoff V LangermannS Pinkner J Hultgren SJ and Knight SD (1999) X-ray structure of the FimCndashFimH chaperonendashadhesin com-plex from uropathogenic Escherichia coli Science 2851061ndash1066
Collaborative computational project n4 (1994) The CCP4suite programs for crystallography Acta Crystallogr D BiolCrystallogr 50 760ndash763
Dodson KW Pinkner JS Rose T Magnusson G Hult-gren SJ and Waksman G (2001) Structural basis of theinteraction of the pyelonephritic Escherichia coli adhesin toits human kidney receptor Cell 105 733ndash743
Eley JG and Triumalashetty P (2001) In vitro assessmentof alkylglycosides as permeability enhancers AAPSPharmSciTech 2 (3) Article 19
Esnouf RM (1997) An extensively modified version of Mol-script that includes greatly enhanced coloring capabilitiesJ Mol Graph Model 15 132ndash134
Firon N Ofek I and Sharon N (1982) Interaction ofmannose-containing oligosaccharides with the fimbriallectin of Escherichia coli Biochem Biophys Res Commun105 1426ndash1432
Firon N Ofek I and Sharon N (1983) Carbohydrate spec-ificity of the surface lectins of Escherichia coli Klebsiellapneumoniae and Salmonella typhimurium Carbohydr Res120 235ndash249
Firon N Ofek I and Sharon N (1984) Carbohydrate-
binding sites of the mannose-specific fimbrial lectins ofEnterobacteria Infect Immun 43 1088ndash1090
Firon N Ashkenazis S Mirelman D Ofek I and SharonN (1987) Aromatic a-glycosides of mannose are powerfulinhibitors of the adherence of type 1 fimbriated Escherichiacoli to yeast and intestinal cells Infect Immun 55 472ndash476
Fukiya S Mizoguchi H Tobe T and Mori H (2004)Extensive genomic diversity in pathogenic Escherichia coliand Shigella strains revealed by comparative genomichybridization microarray J Bacteriol 186 3911ndash3921
Hooton TM and Stamm WE (1997) Diagnosis and treat-ment of uncomplicated urinary tract infection Infect DisClin North Am 11 551ndash581
Horovitz A and Levitzki A (1987) An accurate method fordetermination of receptor ligand and enzyme-inhibitor dis-sociation-constants from displacement curves Proc NatlAcad Sci USA 84 6654ndash6658
Hull RA Gill RE Hsu P Minshaw BH and Falkow S(1981) Construction and expression of recombinant plas-mids encoding type 1 and D-mannose-resistant pili from aurinary tract infection Escherichia coli isolate Infect Immun33 933ndash938
Hultgren SJ Porter TN Schaeffer AJ and Duncan JL(1985) Role of type 1 pili and effects of phase variation onlower urinary tract infections produced by Escherichia coliInfect Immun 50 370ndash377
Hung C-S Bouckaert J Hung DL Pinkner J WinbergC Defusco A et al (2002) Structural basis of tropism ofEscherichia coli to the bladder during urinary tract infec-tion Mol Microbiol 44 903ndash915
Jones TA Zou JY Cowan SW and Kjeldgaard M(1991) Improved methods for building protein models inelectron density maps and the location of errors in thesemodels Acta Crystallogr A 47 110ndash119
Justice SS Hung C-S Theriot JA Fletcher DAAnderson GG Footer MJ and Hulgren SJ (2004)Differentiation and developmental pathways of uropatho-genic Escherichia coli in urinary tract pathogenesis ProcNatl Acad Sci USA 101 1333ndash1338
Kallenius G Mollby R Svenson SB Windberg J Lun-dblud A Svenson S and Cedergen B (1980) The Pk
antigen as receptor for the haemagglutinin of pyelonephri-togenic Escherichia coli FEMS Microbiol Lett 8 297ndash302
Knight SD Berglund J and Choudhury D (2000) Bacte-rial adhesins structural studies reveal chaperone functionand pilus biogenesis Curr Opin Chem Biol 4 653ndash660
Langermann S and Ballou WR (2003) Development of arecombinant FimCH vaccine for urinary tract infectionsAdv Exp Med Biol 539 635ndash653
Langermann S Palaszynski S Barnhart M Auguste GPinkner JS Burlein J et al (1997) Prevention ofmucosal Escherichia coli infection by FimH-adhesin-basedsystemic vaccination Science 276 607ndash611
Langermann S Mollby R Burlein JE Palaszynski SRAuguste CG Defusco A et al (2000) Vaccination withFimH adhesin protects cynomolgus monkeys from coloni-zation and infection by uropathogenic Escherichia coli JInfect Dis 181 774ndash778
Larsson A Ohlsson J Dodson KW Hultgren SJ Nils-son U and Kihlberg J (2003) Quantitative studies of thebinding of the class II PapG adhesin from uropathogenic
454 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
Escherichia coli to oligosaccharides Bioorg Med Chem 112255ndash2261
Leffler H and Svanborg-Eden C (1980) Chemical identifi-cation of a glycosphingolipid receptor for Escherichia coliattaching to human urinary tract epithelial cells and agglu-tinating human erythrocytes FEMS Microbiol Lett 8 127ndash134
Lindhorst TK Kieburg C and Krallmann-Wenzel U(1998) Inhibition of the type 1 fimbriae-mediated adhesionof Escherichia coli to erythrocytes by multiantennary alpha-mannosyl clusters the effect of multivalency Glycoconj J15 605ndash613
Loris R Tielker D Jaeger KE and Wyns L (2003a)Structural basis of carbohydrate recognition by the lectinLecB from Pseudomonas aeruginosa J Mol Biol 331 861ndash870
Loris R Imberty A Beeckmans S Van Driessche ERead JS Bouckaert J et al (2003b) Crystal structureof Pterocarpus angolensis lectin in complex with glucosesucrose and turanose J Biol Chem 278 16297ndash16303
Lund B Lindberg F Marklund BI and Normark S(1987) The PapG protein is the a-D-galactopyranosyl-(1-4)-b-D-galactopyranose-binding adhesin of uropathogenicEscherichia coli Proc Natl Acad Sci USA 84 5898ndash5902
Min G Stolz M Zhou G Liang F Sebbel P StofflerD et al (2002) Localization of uroplakin Ia the urothelialreceptor for bacterial adhesin FimH on the six innerdomains of the 16 nm urothelial plaque particle J Mol Biol317 697ndash706
Miroux B and Walker JE (1996) Over-production of pro-teins in Escherichia coli mutant hosts that allow synthesisof some membrane proteins and globular proteins at highlevels J Mol Biol 260 289ndash298
Morris GM Goodsell DS Halliday RS Huey R HartWE Belew RK and Olson AJ (1998) Automateddocking using a Lamarckian genetic algorithm and anempirical binding free energy function J Comput Chem 191639ndash1662
Mulvey MA (2002) Adhesion and entry of uropathogenicEscherichia coli Cell Microbiol 4 257ndash271
Mulvey MA Lopez-Boado YS Wilson CL Roth RParks WC Heuser J and Hultgren SJ (1998) Induc-tion and evasion of host defenses by type 1-piliated uro-pathogenic Escherichia coli Science 282 1494ndash1497
Murshudov GN Vagin AA and Dodson EJ (1997)Refinement of macromolecular structures by the maxi-mum-likelihood method Acta Crystallogr D Biol Crystallogr53 240ndash255
Nagahori N Lee RT Nishimura S Pageacute D Roy Rand Lee YC (2002) Inhibition of adhesion of type 1 fim-briated Escherichia coli to highly mannosylated ligandsChembiochem 3 836ndash844
Navaza J (2001) Implementation of molecular replacementin AMoRe Acta Crystallogr D Biol Crystallogr 57 1367ndash1372
Neeser JR Koellreutter B and Wuersch P (1986)Oligomannoside-type glycopeptides inhibiting adhesion ofEscherichia coli strains mediated by type 1 pili preparationof potent inhibitors from plant glycoproteins Infect Immun52 428ndash436
Old DC (1972) Inhibition of the interaction between fimbrial
hemagglutinatinins and erythrocytes by D-mannose andother carbohydrates J Gen Microbiol 71 149ndash157
Orndorff PE Devapali A Palestrant S Wyse A EverettML Bollinger RR and Parker W (2004) Immunoglob-ulin-mediated agglutination of and biofilm formation byEscherichia coli K-12 require the type 1 pilus fiber InfectImmun 72 1929ndash1938
Oscarson S and Tedebark U (1995) Syntheses of deoxyanalogues of methyl 36-di-O-a-D-mannopyranosyl-a-D-mannopyranoside for studies of the binding site of con-canavalin A Carbohydr Res 278 271ndash287
Oscarson S and Tiden AK (1993) Syntheses of the octyland tetradecyl glycosides of 36-di-O-alpha-deuterium-mannopyranosyl-alpha-deuterium-mannopyranose and of34-di-O-alpha-deuterium-mannopyranosyl-alpha-deute-rium-mannopyranose ndash a new way for 24-di-O-protectionof mannopyranosides Carbohydr Res 247 323ndash328
Otwinowski Z and Minor W (1997) Processing of X-raydiffraction data collected in oscillation mode MethodsEnzymol 276 307ndash326
Powell HR (1999) The Rossmann Fourier autoindexingalgorithm in MOSFLM Acta Crystallogr D Biol Crystallogr55 1690ndash1695
Pratt LA and Kolter R (1998) Genetic analysis of Escher-ichia coli biofilm formation roles of flagella motility chemo-taxis and type I pili Mol Microbiol 30 285ndash293
Remaut H and Waksman G (2004) Structural biology ofbacterial pathogenesis Curr Opin Struct Biol 14 161ndash170
Ronald AR Nicolle LE Stamm E Krieger J WarrenJ Schaeffer A et al (2001) Urinary tract infection inadults research priorities and strategies Int J AntimicrobAgents 17 343ndash348
Sanner MF Olson AJ and Spehner JC (1996)Reduced surface an efficient way to compute molecularsurfaces Biopolymers 38 305ndash320
Sauer FG Futterer K Pinkner JS Dodson KW Hult-gren SJ and Waksman G (1999) Structural basis ofchaperone function and pilus biogenesis Science 2851058ndash1061
Sauer FG Knight SD Waksman GW and HultgrenSJ (2000) PapD-like chaperones and pilus biogenesisSemin Cell Dev Biol 11 27ndash34
Sauer FG Pinkner JS Waksman G and Hultgren SJ(2002) Chaperone priming of pilus subunits facilitates atopological transition that drives fiber formation Cell 111543ndash551
Schembri MA Hasman H and Klemm P (2000)Expression and purification of the mannose recognitiondomain of the FimH adhesin FEMS Microbiol Lett 188147ndash151
Schembri MA Kjaergaard K and Klemm P (2003) Glo-bal gene expression in Escherichia coli biofilms Mol Micro-biol 48 253ndash267
Schilling JD Mulvey MA Vincent CD Lorenz RGand Hultgren SJ (2001) Bacterial invasion augments epi-thelial cytokine responses to Escherichia coli through alipopolysaccharide-dependent mechanism J Immunol166 1148ndash1155
Silhavy TJ Szmelcman S Boos W and Schwartz M(1975) On the significance of the retention of ligand byprotein Proc Natl Acad Sci USA 72 2120ndash2124
A novel class of FimH high-affinity ligands 455
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
Svanborg C and Godaly G (1997) Bacterial virulence inurinary tract infection Infect Dis Clin North Am 11 513ndash529
Thanassi DG Saulino ET and Hultgren SJ (1998) Thechaperoneusher pathway a major terminal branch of thegeneral secretory pathway Curr Opin Struct Biol 1 223ndash231
Thankavel K Madison B Ikeda T Malaviya R ShahAH Arumugen PM and Abraham SN (1997) Local-ization of a domain in the FimH adhesin of Escherichia colitype 1 fimbriae capable of receptor recognition and use ofa domain-specific antibody to confer protection againstexperimental urinary tract infection J Clin Invest 1001123ndash1126
Vagin A and Teplyakov A (1997) MOLREP an automatedprogram for molecular replacement J Appl Crystallogr 301022ndash1025
Wu XR Sun TT and Medina JJ (1996) In vitro bindingof type 1-fimbriated Escherichia coli to uroplakins 1a and
1b relation to urinary tract infections Proc Natl Acad SciUSA 93 9630ndash9635
Xia Y Gally D Forsman-Semb K and Uhlin BE (2000)Regulatory cross-talk between adhesin operons in Escher-ichia coli inhibition of type 1 fimbriae expression by thePapB protein EMBO J 19 1450ndash1457
Zafriri D Ofek I Adar R Pocino M and Sharon N(1989) Inhibitory activity of cranberry juice on adherenceof type-1 and type-P fimbriated Escherichia coli to eukary-otic cells Antimicrob Agents Chemother 33 92ndash98
Zavialov AV Berglund J Pudney AF Fooks LJ Ibra-him TM MacIntyre S and Knight SD (2003) Structureand biogenesis of the capsular F1 antigen from Yersiniapestis preserved folding energy drives fiber formation Cell113 587ndash596
Zhou G Mo WJ Sebbel P Min G Neubert TA Glock-shuber R et al (2001) Uroplakin Ia is the urothelial recep-tor for uropathogenic Escherichia coli evidence from invitro FimH binding J Cell Sci 114 4095ndash4103

A novel class of FimH high-affinity ligands 447
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
docks in the binding pocket in a conformation very closeto the crystallographically observed mannose conforma-tion (Fig 6) while the aromatic substituent interacts withthe tyrosine gate The different conformation of Tyr48 in
our two FimH truncate structures leads to different pre-dicted interactions between the aromatic aglycon andTyr48 and Tyr137 Docking to the FimHtr2 structureresulted in somewhat tighter interactions between thetyrosine gate and the aglycon and hence in somewhatlower docking energies Calculated FimHtr2 docking ener-gies of Edoc = -104 kcal mol-1 for pNPaMan (Kd = 46 nM)and Edoc = -109 kcal mol-1 for MeUmbaMan (Kd = 20 nM)are in good agreement with the experimental binding data(Table 1) The docked umbelliferyl ring system is inserteddeep into the tyrosine gate in the FimHtr2 conformationwith the carbonyl group within hydrogen bonding distanceto Thr51 at the back of the gate (Fig 6A) Docking to theFimHtr1 structure instead stacks the umbelliferyl ring face-to-face with the Tyr48 ring (not shown) The phenyl ring ofpNPaMan docks similarly to both our structures It doesnot quite reach in between the tyrosine side-chains butinstead hydrogen bonds with its nitro group to the hydroxyloxygen of Tyr137 (Fig 6B)
Discussion
Most lectins bind naturally occurring carbohydrates withlow affinities often in the millimolar range for monosac-charides and micromolar range for oligosaccharides Thiswas recently exemplified to also hold true for theadhesins of F17 fimbriae (Kd = 12 mM for the F17a-G-N-acetylglucosamine interaction Buts et al 2003) and Ppili (Kd = 84 mM for the PapGII-globotetraoside interactionLarsson et al 2003) The recent crystal structure of FimHin complex with mannose illustrated that FimH almostcompletely envelops the monosaccharide in a deeppocket and involves it in an intense hydrogen bondingnetwork suggestive of an unusual lectin exerting highaffinities (Hung et al 2002)
Fig 4 FimH binding experimentsA Surface plasmon resonance competition experiment of FimHtr1 binding to monoclonal antibody IC10 Fab fragments with butyl a-D-mannosideB Equilibrium binding of [3H]-mannose to FimHtr2C Equilibrium displacement measurements of butyl a-D-mannoside to the FimHtr2 [3H]-mannose complex
log[Butyl a-D-mannoside] (M)
cpm
cpm
Butyl a-D-mannoside (mM)
Mannose (mM)
A
B
C
ndash64
ndash66
ndash68
ndash70
ndash72
ndash74
ndash76
ndash78
ndash80
ndash82
ndash8 ndash7 ndash6 ndash5 ndash4 ndash3
log
[Fim
H]
M
Fig 5 Linear free energy relationship for the alkyl chain on mannose A linear fit was made using 1 up to 7 methyl groups with the program MICROCAL ORIGIN 50
Number of carbon atoms0 2 4 6 8
ndash75
ndash80
ndash90
ndash95
ndash100
ndash105
ndash110
ndash115
ndash85
D G
(kc
al m
olndash1
)
slopecorrelation
ndash064ndash099
+ndash 003+ndash 018
448 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
The measured affinity confirm the predictions that FimHbinds the monosacharide mannose exceptionally well(Kd = 23ndash41 mM) To investigate the contribution of thedifferent hydroxyl groups of mannose in the interactionwith FimH the binding of a series of hexoses to butylmannoside-free FimH was examined using SPR (Table 2)The effect of deoxygenation of the O2 hydroxyl group(methyl 2-deoxy-a-D-mannopyranoside) is clearly lessdestructive (100-fold reduction) than its equatorial positionas in glucose O2 is the only hydroxyl group of mannosethat interacts indirectly with the FimH pocket via a con-served water molecule (Hung et al 2002) Glucose onlydisplays a weak affinity for FimH about 4000 times lowerthan methyl a-D-mannoside Repositioning of both the O2and O4 hydroxyl groups as in galactose abolishes sugarbinding by FimH Similar to these results the inhibitoryconcentrations for bacterial adherence of methyl a-D-glucoside and galactose derivatives have been estimatedto be respectively 104-fold and 105-fold weaker than formethyl a-D-mannoside (Nagahori et al 2002)
In stark contrast with the other monosaccharides fruc-tose has a binding affinity only about 15-fold less thanmannose for FimH (Table 2) The decrease in affinity for
fructose relative to mannose agrees with studies of inhi-bition of adherence of type 1 piliated E coli by fructosein which fructose was found to be the active compound infruit juices (present at about 5) (Zafriri et al 1989)Fructose binding to FimH is stronger than the binding ofthe physiological globotetraoside receptor to PapGII theother well-characterized adhesin of UPEC strains (Lars-son et al 2003) The pyranose form of fructose is verywell superimposable onto mannose in the FimH bindingsite where similar interactions are established with FimHas for mannose except for the loss of the anomeric O1and an extra hydroxyl group (O2) on the C5 position ofmannose Probably however both changes of fructopyr-anoside compared with mannopyranoside could be easilyaccommodated by FimH The mannose O1 does notmake direct interactions within 45 Aring of the FimH bindingpocket (Fig 1B) The extra O2 group of fructose would notclash or interfere significantly with the binding pocket butit would make a 27 Aring van der Waals contact with the Ile52side-chain under the condition that the Ile52 side-chaindoes not move The crystal structure of the Pseudomonasaeruginosa lectin LecB also showed that fructopyranosebut not fructofuranose can substitute for mannose (Loris
Fig 6 Stereo diagrams of docked MeUm-baMan (A) and pNPaMan (B) in the FimH bind-ing site The crystallographically determined position of a-D-mannose is shown in orange for comparison
A novel class of FimH high-affinity ligands 449
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
et al 2003a) Hydrogen bonds of the more specificfucose complex are replaced by van der Waals interac-tions involving a hydroxyl group in the complexes with thelower-affinity binders mannose and fructopyranose as forthe proposed fructopyranoside binding to FimH
The fructose-containing disaccharides sucrose andturanose do not have an inhibitory effect and bind withaffinities similar to glucose Sucrose is a non-reducingsugar that has fructose locked in its furanose form Tura-nose on the other hand allows the interconversion of b-D-fructofuranose and b-D-fructopyranose eg both formsoccur in complex with a plant lectin from Pterocarpusangolensis (Loris et al 2003b) It is predictable howeverfrom the crystal structures that the glycosidic linkage ofglucose with C3 of fructose equivalent in position to C4of mannose would be disallowed in the tight FimH pocketSucrose and turanose thus most probably interact with theFimH mannose binding pocket via their glucose moietyresulting in similar low affinities
Our mass spectrometry and chemical analyses con-firms the crystallographic identification of the ligand boundto the FimH lectin domains as butyl a-D-mannoside(Figs 1B and 2) The mannose ring of the butyl a-D-mannoside coincides with the mannose bound to theFimCFimH complex (Hung et al 2002) and makes thesame interactions The four-carbon alkyl tail exits towardsthe back wall (loop b4ndashb5 and b10ndashb11) of the pocket(Fig 1A and B) making hydrophobic contacts to Tyr48Tyr137 and Ile52 The mass spectrometry data also showthat the bound butyl a-D-mannoside is present in LBmedium but absent from minimal medium Hence thesource of the alkyl mannoside is possibly the yeast extractused in preparation of LB medium Although we do notwant to speculate about the significance of this findingalkyl mannoside production by yeast has not beenreported and might warrant investigation
The butyl mannoside was retained by FimH despiteextensive dialysis The phenomenon of ligand retention byprotein was described as early as 1975 for periplasmicbinding proteins (Silhavy et al 1975) Applied to dialysisof butyl mannoside away from FimH grown in richmedium half-lives of up to 30 h can be estimated whichis far beyond the duration of a normal buffer exchangedialysis experiment In the present case the retainedligand does not appear to seriously affect [3H]-mannosedisplacement equilibrium binding experiments as wecould detect no significant differences in dissociation con-stants obtained using FimHtr2 produced in LB or minimalmedia (data not shown) In contrast proper SPR bindingcurves could not be obtained when butyl mannoside occu-pied the binding site Hence the displacement techniquemay be of more general use to obtain binding constantsfor proteins that tend to inadvertently retain ligands whichis a common trait of lectins
Surface plasmon resonance as well as equilibrium bind-ing and displacement measurements using [3H]-mannosewere carried out to obtain dissociation constants for a-D-mannose and a linear series of alkyl-substituted manno-sides to FimH (Table 1) Both equilibrium binding experi-ments show a very good agreement in their measurementof the carbohydrate affinity of the adhesin FimH bindsbutyl a-D-mannoside 15ndash30 times stronger than a-D-mannose The importance of the added hydrophobiccontacts is clearly demonstrated by the binding of alkylmannosides (Table 1) The best binding alkyl mannosidesbind a few hundred times stronger than mannose equiv-alent in affinity to the most tightly binding aromaticallysubstituted mannosides for FimH (Firon et al 1987) andof mannose dendrimers for type 1 piliated E coli (Fironet al 1984 1987 Nagahori et al 2002)
Interestingly the butyl moiety takes on significantlydifferent conformations in FimHtr1 and FimHtr2 (Fig 1C)indicating that intrinsic conformational flexibility in theaglycon is allowed Indeed in spite of the larger flexibilityexpected for longer alkyl mannosides the binding energydecreases linearly with increasing length of the alkyl chainbetween one and seven carbon atoms (Fig 5) The affinityreaches its maximum for mannose with a seven-carbonalkyl and SPR measurements suggest that the affinitydecreases from eight carbon atoms on Evidently theeighth carbon atom goes beyond the hydrophobic interac-tion surface and protein edge which is penalized with adecrease in affinity We propose that the affinity increaseresults from an increased interaction surface with thehydrophobic gateway Alkyl aglycons fit well in the tyrosinegate and as observed in the two crystal structures in atleast two different conformations that depend on theconformation of Tyr48 This flexibility may indicate thataglycons can interact rather non-specifically with thehydrophobic gateway nevertheless resulting in a 100- to1000-fold increase in affinity
FimH binding to ligands other than just mannose ispossibly of importance in terms of the different environ-ments that E coli will encounter ndash the urinary tract gas-trointestinal tract and in biofilms Alkyl mannosides havenot been previously recognized as strong binders to FimHAs they are easily synthesized and highly soluble in waterthey may be interesting as potential blocking agents forFimH-mediated adhesion Moreover they have beenshown to be good drug vehicles themselves acting aspermeability enhancers on tight junctions (Eley and Triu-malashetty 2001) As such they might be able to pene-trate into deeper cell layers of the epithelium to alsoprevent those bacteria that form a quiescent reservoirfrom causing recurrent infections (Mulvey et al 1998Justice et al 2004)
On the other hand the two aromatically substitutedmannosides pNPaMan and MeUmbaMan are known to
450 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
be particularly potent inhibitors of FimH-mediated bacte-rial adhesion (Firon et al 1987 Nagahori et al 2002)Based on adhesion inhibition studies using type 1 piliatedE coli and guinea pig ileal epithelial cells the relativeinhibitory potential of pNPaMan and MeUmbaMan havebeen estimated to be about 70 and 600 times strongerrespectively than that of methyl a-D-mannose (Fironet al 1987) Our measurements indicate an affinity ofFimH for pNPaMan and MeUmbaMan between 50 and90 times and between 110 and 200 times higher respec-tively than for methyl a-D-mannose in good agreementwith the adhesion inhibition results (Firon et al 1987)Dissociation constants predicted using AutoDock3 (Morriset al 1998) are similar to experimental values Thecomputational docking predicts that tight binding of thearomatically substituted mannosides pNPaMan andMeUmbaMan is accomplished similarly as for butyl a-D-mannoside with the aglycon interacting with the tyrosinegate
This is the first systematic study providing molecularbinding data for the FimH adhesin the prerequisite forcolonization by UPEC (Langermann and Ballou 2003)We also show crystallographic evidence that the lectinor receptor binding domain of FimH is a stable andfunctional entity Its structure is virtually identical to thesame domain in the FimCFimH complex indicating thata truncate corresponding to the lectin domain could pro-vide a good alternative for the development of a UTIvaccine (Langermann et al 1997 Langermann and Bal-lou 2003) However progress of this interesting UTI vac-cine candidate into clinical trials has not been reportedThe binding affinities of FimH at the molecular levelrelate convincingly well with the inhibitory concentrationsused in previous bacterial adhesion assays thus endors-ing the direct and conclusive correlation of FimH withbacterial adherence Alkyl mannosides have nanomolaraffinities for FimH which may overcome the significantobstacle of low affinity generally encountered in attemptsto develop carbohydrate-based drugs which interferewith microbial attachment to host cell surface glycocon-jugates New anti-adhesive drugs could be based onthese leads to allow specific interaction and targeting tothe urinary tract in order to prevent anticipated andrecurrent infections
Experimental procedures
Strains and constructs
FimH proteins from two different E coli strains the UPECstrain J96 (Hull et al 1981) and the E coli K-12 strainMG1655 (Blattner et al 1997) were used in this study TheFimH adhesins encoded by these two E coli strains areidentical FimHtr1 (residues 1ndash158 of UPEC J96 FimH) wasexpressed from plasmid pMMB91 transformed into E coli
C600 cells FimHtr2 (residues 1ndash158 of E coli MG1655FimH + carboxy-terminal 6-His tag) was expressed fromplasmid pPKL241 (Schembri et al 2000) transformed into Ecoli host strain HB101
Purification and crystallization
FimHtr1 Bacteria were grown in LB medium at 37infinC inducedat OD600 = 08 with 1 mM IPTG and harvested 2ndash3 h laterThe cells were resuspended in 20 mM Tris-HCl pH 80 5 mMEDTA and 30 sucrose and the periplasmic content wasextracted by osmotic shock upon 10-fold dilution with 20 mMTris-HCl pH 80 Purification of FimHtr1 was performed by ionexchange chromatography with MonoS at pH 45 followed byMonoQ (Amersham Biosciences) at pH 8 FimHtr1 was crys-tallized using vapour diffusion in hanging drops composed of3 ml of the precipitant solution (01 M Hepes pH 75 12PEG 6000 and 5 vv methyl-24-pentanediol) and 3 ml of12 mg ml-1 protein solution in 20 mM Tris-HCl pH 80 with150 mM NaCl
FimHtr2 Cells were grown in LB medium containing50 mg ml-1 ampicillin at 37infinC At OD600 = 06 the cells wereinduced with 5 mM IPTG Cells were harvested 2 h afterinduction and the periplasmic content was extracted Theprotein was purified on a Pharmacia HiTrap Chelating HP5 ml column (Pharmacia) loaded with Ni-chloride and elutedwith a sharp 0ndash500 mM imidazole gradient Fractions con-taining FimHtr2 were pooled dialysed against 50 mM sodiumacetate pH 525 and loaded onto a Mono S HR 8 ml columnThe protein was eluted with a 0ndash500 mM NaCl gradient anddialysed overnight against 20 mM MES pH 65 before crys-tallization Crystallization was performed at room tempera-ture by the vapour diffusion method with hanging dropsmade by mixing of 2 ml of 15 mg ml-1 protein solution with 2 mlof the precipitant solution (65 methyl-24-pentanediol100 mM cacodylate pH 65) Microseeding was employed tooptimize crystal quality and reproducibility Crystals suitablefor X-ray data collection generally appeared within 48 h aftermicroseeding
Structure determination
X-ray data for FimHtr1 were collected from a single crystalto 17 Aring resolution at the Advanced Photon SourceArgonne National Laboratory USA The crystal was flash-cooled to 100 K in the precipitant solution complementedwith 25 glycerol The data were processed and scaledusing the HKL2000 package (Otwinowski and Minor 1997)(Table S1 in Supplementary material) Data for FimHtr2
were collected from a single crystal under liquid-nitrogencryo conditions at 100 K at beamline ID29 ESRF Greno-ble France to 21 Aring Bragg spacing FimHtr2 data were pro-cessed and reduced using MOSFLM (Powell 1999) andscaled using SCALA (Collaborative computational project1994) (Table S1 in Supplementary material) The structuresof FimHtr1 and FimHtr2 were solved by molecular replace-ment using Amore (Navaza 2001) and MOLREP (Vaginand Teplyakov 1997) respectively with the FimH lectindomain from the FimCFimH complex (Choudhury et al
A novel class of FimH high-affinity ligands 451
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
1999) (PDB id 1QUN) as the search model Positional bulksolvent and isotropic temperature factor refinement wasperformed using CNS (Bruumlnger et al 1998) and REFMAC5(Murshudov et al 1997) respectively Model adjustmentand inspection were performed using Turbo-Frodo and O(Jones et al 1991) graphics programs respectively Refine-ment statistics are given in Table S1 Molecular graphicsillustrations were generated with Bobscript (Esnouf 1997)and Pymol 090 (DeLano Scientific LLC 1998ndash2003 httppymolsourceforgenet)
Mass spectrometry
The purified FimHtr1 protein (300 mg) that had been used forcrystallization was freeze-dried taken up in 100 ml of pyridineand 100 ml of acetic anhydride and incubated overnight atroom temperature in the dark for peracetylation The samplewas dried under nitrogen and taken up in chloroform (400 ml)The chloroform phase was extracted six times with waterdried and taken up in 200 ml of 50 aqueous methanol A1 ml aliquot was applied to a nano-scale RP column(75 mm yen 100 mm PepMap DionexLC Packings) using anUltimate nano-LC system (LC Packings) which was equili-brated with eluent A (H2Oacetonitrile 955 vv containing01 formic acid) at a flow rate of 100 nl min-1 After injectingthe sample the column was run isocratically for 5 min fol-lowed by a linear gradient to 100 eluent B (H2Oacetonitrile2080 vv containing 01 formic acid) in 20 min and a finalwash with 100 B for 5 min The system was directly coupledto an Esquire 3000 ESI-IT-MS (Bruker Daltonik) equippedwith an online nanospray source operating in the positive-ionmode For electrospray (900ndash1200 V) capillaries (360 mmOD 20 mm ID with 10 mm opening) from New Objective wereused The solvent was evaporated at 150infinC with a nitrogenstream of 6 l min-1
To verify the suspected source of the bound butyl manno-side mass spectrometry was performed on samples ofFimHtr2 grown in either LB or minimal M9 medium Proteinsamples were dialysed against water overnight before massspectrometry in order to reduce background from the Trisbuffer Samples were diluted to a concentration in the submi-cromolar range Electrospray mass spectra were recorded indirect inlet mode (50 aqueous methanol solutions) on aBruker Daltonics Esquire-LC (Billerica)
Carbohydrate constituent analysis
Samples were hydrolysed in 100 ml of 4 M aqueous trifluoro-acetic acid (Sigma) at 100infinC for 4 h and dried under astream of nitrogen Monosaccharides were converted intotheir anthranilic acid derivatives by reductive aminationresolved by RP-HPLC and detected by fluorescence (Anu-mula 1994)
Carbohydrates for binding studies
[3H]-mannose was obtained from Amersham Methyl a-D-mannoside glucose galactose fructose sucrose turanosep-nitrophenyl a-D-mannopyranoside (pNPaMan) and 4-methylumbelliferyl a-D-mannopyranoside (MeUmbaMan)
were obtained from Sigma Monomannosylated bovineserum albumin (BSA-Man) was kindly provided by MedIm-mune We synthesized methyl 2-deoxy-a-D-mannopyrano-side (Oscarson and Tedebark 1995) Alkyl mannosides weresynthesized through silver triflate-promoted couplings of thecorresponding alcohol with 2346-tetra-O-benzoyl-a-D-mannopyranosyl bromide followed by Zempleacuten deacylationof the obtained protected alkyl mannosides according to theprocedure reported for the octyl and tetradecyl mannosides(Oscarson and Tiden 1993)
Binding studies
Competition experiments using SPR To obtain butyl man-noside-free FimHtr1 the FimH truncate was overproducedwith a carboxy-terminal 6-His tag after transformation into Ecoli C43 (DE3) (Miroux and Walker 1996) and the bacteriagrown in minA minimal medium supplemented with the 20amino acids (40 mg l-1) thiamin (2 mg l-1) and biotin (2 mgl-1) Purification of the His-tagged protein was performedusing Ni-NTA affinity chromatography (Qiagen) in 50 mM Tris-HCl pH 8 and 300 mM NaCl Elution was in the same solutioncomplemented with 500 mM imidazole Subsequently pooledand concentrated FimH fractions were gel filtrated over aSuperdex G75 column (Pharmacia) in 20 mM Tris-HCl pH 8
We used Fab fragments of a monoclonal antibody againstthe mannose-binding site of FimH to determine the solutionaffinity of FimHndashcarbohydrate interactions The monoclonalantibody IC10 was produced by a mouse hybridoma cell lineat Medimmune and its Fab fragments were prepared andpurified The Fabs were covalently immobilized via aminecoupling at 1000 Resonance Units (1000 pg ligand mm-2) inflowcell Fc2 on a CM5 biasensor chip (BIAapplications Hand-book Biacore AB Uppsala Sweden) Flowcell Fc1 was usedas the reference surface and was treated in the same wayas Fc2 except that it was blocked immediately after activationThe immobilization buffer was 100 mM sodium acetatepH 50 with 100 mM NaCl All SPR measurements were per-formed on a Biacore3000trade
The kinetic constants of binding of FimH to the immobilizedantibody were measured with different concentrations ofFimH (2000 nM 1000 nM 500 nM 250 nM 125 nM625 nM 312 nM 156 nM 78 nM) in running buffer [phos-phate-buffered saline (PBS) with 0005 surfactant P20 and3 mM EDTA] on both flowcells Fc1 and Fc2 simultaneouslyat a flow rate of 30 ml min-1 and at 298 K Complete dissoci-ation of FimH was performed with running buffer beforestarting a new binding cycle For all measurements the asso-ciation time was 3 min and the dissociation time was 30 minAll binding cycles were performed in duplicate including azero concentration cycle of FimH (injection of running buffer)All analyses were performed with the BIAeval software ALangmuir binding isotherm with a 11 stoichiometry was fittedto the data from both the association phase and the dissoci-ation phase after subtraction of the reference and zero con-centration data The kinetic constants ka and kd and themaximal binding Rmax were obtained and the dissociationconstant Kd was calculated
The equilibrium dissociation constants Kd of FimHndashsaccha-ride interactions were determined in a competition experi-ment using the same chip as for the previous experiment
452 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
As a control the Fabs of monoclonal antibody IA7 as wellas BSA-Man were also immobilized Samples containing afixed concentration of FimH were used (close to the Kd of theFimHndashantibody interaction) in combination with varying con-centrations of saccharide First 10-fold dilutions of the sac-charide solution were used to determine the concentrationrange for binding of the saccharide to FimH A Langmuirbinding isotherm with a 11 stoichiometry was fitted to thedata using the kinetic constants and Rmax from the foregoingexperiment to obtain the concentrations of FimH that werefree ([FimH]free) to bind the antibody immobilized on the chipSecond the concentration range of the saccharide wasextended and adapted to ensure accurate fitting and theequilibrium binding constant of the FimHndashsaccharide interac-tion was derived from the curve of [FimH]free against concen-tration of saccharide Every measurement was repeated atleast twice including testing different protein batches andwhenever possible also different saccharide stock solutions(typically 200 mM)
Equilibrium binding studies using radioactively labelledmannose Mannose-free FimHtr2 was produced inM9 minimal medium and purified as described above forFimHtr2 Alkyl mannosides were dissolved in double distilledwater pNPaMan and MeUmbaMan (6 mg each) were dis-solved in 20 ml of DMSO and diluted to 20 mM using doubledistilled water Binding experiments were performed at 310 Kusing six different concentrations of [3H]-a-D-mannose (finalconcentrations 435 mM 290 mM 193 mM 129 mM 86 mMand 57 mM) Protein (180 ml) at a concentration of about500 nM was mixed with 20 ml of the radioactive ligand [3H]-a-D-mannose and incubated at 37degC for 20 min To separatefree ligand from bound the mixture was rapidly filtratedthrough a Protran BA 85 cellulose nitrate filter (Schleicherand Schuell) and washed once with 1 ml of ice-cold 1yen PBSFilter-bound radioactivity was measured by scintillation spec-trometry within 24 h Displacement experiments were per-formed using six different concentrations of inhibitor (finalconcentrations in the range 00ndash435 mM) in the presence of435 mM [3H]-a-D-mannose Twenty microlitres of radioactiveligand 20 ml of inhibitor and 160 ml of protein (500 nM) weremixed and the experiments were performed as describedabove All experiments were performed in duplicates and thedata presented are mean values of at least two independentmeasurements For the determination of Kd for a-D-mannosean hyperbolic curve (y = P1x(P2 + x) where P1 is an arbitraryscale factor and P2 = Kd) was fitted to the data For the dis-placement experiments the curve y = P1(P2 + x) where P1
is an arbitrary scale factor and P2 is the concentration of theinhibitor displacing 50 of the labelled ligand [I]05 was usedinstead To calculate the inhibitor dissociation constant (KI)the Cheng and Prusoff equation [KI = [I]05[L]KL + 1) KL is theconstant of dissociation for the ligand] (Cheng and Prusoff1973) was used when both the concentration of the radio-active ligand (L) and the displacing agent (I) are in excessover the protein (LT gtgt PT IT gtgt PT T indicates total concen-tration) For the two strong inhibitors pNPaMan and MeUm-baMan where IT was not in excess over PT the equationof KI = IT[(1 - Y)Y(LTKL) - 1] - PTKLYLT where Y is thefraction of the ligand bound in presence of the inhibitor wasused instead (Horovitz and Levitzki 1987)
Computational docking
Computational docking studies were performed usingAutoDock3 (Morris et al 1998) In AutoDock3 the proteinis represented by a grid of affinity potentials Hence theprotein conformation is not allowed to change during thedocking procedure Flexibility of ligands is taken intoaccount by allowing rotation around flexible torsion anglesThe docked energies are calculated as the sum of the inter-molecular interaction energy and the internal energy of theligand The search for the best interaction energy was car-ried out by means of the Lamarckian genetic algorithmLigand structures were obtained from the Protein DataBank and prepared for docking by adding hydrogens andcharges using AutoDockTools (Sanner et al 1996) Eachsimulation consisted of 100 independent runs with a popu-lation size of 200 500 generations and a maximum of25 000 000 energy evaluations Solutions were rankedbased on their docking energies and similar solutions wereclustered (cluster cut-off rmsd lt 1 Aring) For both ligandsthe top solutions were evaluated using the graphics pro-gram O (Jones et al 1991)
Acknowledgements
We thank Andrej Zavialov and Maringhns Ehrenberg for help withequilibrium binding experiments and Thomas Norberg forassistance with mass spectrometry Anthony DeFusco fromMedimmune provided us with mannosylated bovine serumalbumin We are grateful to Fred Sauer for assistance withdata collection Thanks to Maor Bar-Peled of the CCC at theUniversity of Georgia Athens USA for his suggestion toexpress FimH in minimal medium We acknowledge the useof beamline 19BM of the Structural Biology Center at theAdvanced Photon Source Argonne National LaboratoryChicago IL USA and of beamline ID29 ESRF GrenobleFrance JB is a postdoctoral fellow of the Fonds voor Weten-schappelijk Onderzoek-Vlaanderen which also supportedthe BiaCore3000 instrument (FWONL35) This work was sup-ported by grants from the Swedish Research Council andfrom the Strategic Research Foundation (Structural BiologyNetwork) to SDK
Supplementary material
The following material is available from httpwwwblackwellpublishingcomproductsjournalssuppmatmmimmi4415mmi4415smhtmTable S1 Crystallographic data and refinement statistics
References
Anumula KR (1994) Quantitative determination ofmonosaccharides in glycoproteins by high-performanceliquid chromatography with highly sensitive fluorescencedetection Anal Biochem 220 275ndash283
Bahrani-Mougeot FK Buckles EL Lockatell CV HebelJR Johnson DE Tang CM and Donnenberg MS(2002) Type 1 fimbriae and extracellular polysaccharidesare preeminent uropathogenic Escherichia coli virulence
A novel class of FimH high-affinity ligands 453
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
determinants in the murine urinary tract Mol Microbiol 451079ndash1093
Berglund J and Knight SD (2003) Structural basis forbacterial adhesion in the urinary tract Glycobiol Med 53533ndash52
Blattner FR Plunkett G Bloch CA Perna NT BurlandV Riley M et al (1997) The complete genome sequenceof Escherichia coli K-12 Science 277 1453ndashbullbull
Bloch CA Stocker BA and Orndorff PE (1992) A keyrole for type 1 pili in enterobacterial communicability MolMicrobiol 6 697ndash701
Bork P Holm L and Sander C (1994) The immunoglob-ulin fold ndash structural classification sequence patterns andcommon core J Mol Biol 242 309ndash320
Brinton CC (1959) Non-flagellar appendages of bacteriaNature 183 782ndash786
Bruumlnger AT Adams PD Clore GM DeLano WLGros P Grosse-Kunstleve RW et al (1998) Crystallog-raphy and NMR system a new software suite for macro-molecular structure determination Acta Crystallogr D BiolCrystallogr 54 905ndash921
Buchanan K Falkow S Hull RA and Hull SI (1985)Frequency among Enterobacteriaceae of the DNA-sequences encoding type-1 pili J Bacteriol 162 799ndash803
Buts L Bouckaert J De Genst E Loris R Oscarson SLahmann M et al (2003) The fimbrial adhesin F17-G ofenterotoxigenic Escherichia coli has an immunoglobulin-like lectin domain that binds N-acetylglucosamine MolMicrobiol 49 705ndash715
Cheng Y and Prusoff WH (1973) Relationship betweeninhibition constant (K1) and concentration of inhibitor whichcauses 50 per cent inhibition (I50) of an enzymatic reac-tion Biochem Pharmacol 22 3099ndash3108
Choudhury D Thompson A Stojanoff V LangermannS Pinkner J Hultgren SJ and Knight SD (1999) X-ray structure of the FimCndashFimH chaperonendashadhesin com-plex from uropathogenic Escherichia coli Science 2851061ndash1066
Collaborative computational project n4 (1994) The CCP4suite programs for crystallography Acta Crystallogr D BiolCrystallogr 50 760ndash763
Dodson KW Pinkner JS Rose T Magnusson G Hult-gren SJ and Waksman G (2001) Structural basis of theinteraction of the pyelonephritic Escherichia coli adhesin toits human kidney receptor Cell 105 733ndash743
Eley JG and Triumalashetty P (2001) In vitro assessmentof alkylglycosides as permeability enhancers AAPSPharmSciTech 2 (3) Article 19
Esnouf RM (1997) An extensively modified version of Mol-script that includes greatly enhanced coloring capabilitiesJ Mol Graph Model 15 132ndash134
Firon N Ofek I and Sharon N (1982) Interaction ofmannose-containing oligosaccharides with the fimbriallectin of Escherichia coli Biochem Biophys Res Commun105 1426ndash1432
Firon N Ofek I and Sharon N (1983) Carbohydrate spec-ificity of the surface lectins of Escherichia coli Klebsiellapneumoniae and Salmonella typhimurium Carbohydr Res120 235ndash249
Firon N Ofek I and Sharon N (1984) Carbohydrate-
binding sites of the mannose-specific fimbrial lectins ofEnterobacteria Infect Immun 43 1088ndash1090
Firon N Ashkenazis S Mirelman D Ofek I and SharonN (1987) Aromatic a-glycosides of mannose are powerfulinhibitors of the adherence of type 1 fimbriated Escherichiacoli to yeast and intestinal cells Infect Immun 55 472ndash476
Fukiya S Mizoguchi H Tobe T and Mori H (2004)Extensive genomic diversity in pathogenic Escherichia coliand Shigella strains revealed by comparative genomichybridization microarray J Bacteriol 186 3911ndash3921
Hooton TM and Stamm WE (1997) Diagnosis and treat-ment of uncomplicated urinary tract infection Infect DisClin North Am 11 551ndash581
Horovitz A and Levitzki A (1987) An accurate method fordetermination of receptor ligand and enzyme-inhibitor dis-sociation-constants from displacement curves Proc NatlAcad Sci USA 84 6654ndash6658
Hull RA Gill RE Hsu P Minshaw BH and Falkow S(1981) Construction and expression of recombinant plas-mids encoding type 1 and D-mannose-resistant pili from aurinary tract infection Escherichia coli isolate Infect Immun33 933ndash938
Hultgren SJ Porter TN Schaeffer AJ and Duncan JL(1985) Role of type 1 pili and effects of phase variation onlower urinary tract infections produced by Escherichia coliInfect Immun 50 370ndash377
Hung C-S Bouckaert J Hung DL Pinkner J WinbergC Defusco A et al (2002) Structural basis of tropism ofEscherichia coli to the bladder during urinary tract infec-tion Mol Microbiol 44 903ndash915
Jones TA Zou JY Cowan SW and Kjeldgaard M(1991) Improved methods for building protein models inelectron density maps and the location of errors in thesemodels Acta Crystallogr A 47 110ndash119
Justice SS Hung C-S Theriot JA Fletcher DAAnderson GG Footer MJ and Hulgren SJ (2004)Differentiation and developmental pathways of uropatho-genic Escherichia coli in urinary tract pathogenesis ProcNatl Acad Sci USA 101 1333ndash1338
Kallenius G Mollby R Svenson SB Windberg J Lun-dblud A Svenson S and Cedergen B (1980) The Pk
antigen as receptor for the haemagglutinin of pyelonephri-togenic Escherichia coli FEMS Microbiol Lett 8 297ndash302
Knight SD Berglund J and Choudhury D (2000) Bacte-rial adhesins structural studies reveal chaperone functionand pilus biogenesis Curr Opin Chem Biol 4 653ndash660
Langermann S and Ballou WR (2003) Development of arecombinant FimCH vaccine for urinary tract infectionsAdv Exp Med Biol 539 635ndash653
Langermann S Palaszynski S Barnhart M Auguste GPinkner JS Burlein J et al (1997) Prevention ofmucosal Escherichia coli infection by FimH-adhesin-basedsystemic vaccination Science 276 607ndash611
Langermann S Mollby R Burlein JE Palaszynski SRAuguste CG Defusco A et al (2000) Vaccination withFimH adhesin protects cynomolgus monkeys from coloni-zation and infection by uropathogenic Escherichia coli JInfect Dis 181 774ndash778
Larsson A Ohlsson J Dodson KW Hultgren SJ Nils-son U and Kihlberg J (2003) Quantitative studies of thebinding of the class II PapG adhesin from uropathogenic
454 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
Escherichia coli to oligosaccharides Bioorg Med Chem 112255ndash2261
Leffler H and Svanborg-Eden C (1980) Chemical identifi-cation of a glycosphingolipid receptor for Escherichia coliattaching to human urinary tract epithelial cells and agglu-tinating human erythrocytes FEMS Microbiol Lett 8 127ndash134
Lindhorst TK Kieburg C and Krallmann-Wenzel U(1998) Inhibition of the type 1 fimbriae-mediated adhesionof Escherichia coli to erythrocytes by multiantennary alpha-mannosyl clusters the effect of multivalency Glycoconj J15 605ndash613
Loris R Tielker D Jaeger KE and Wyns L (2003a)Structural basis of carbohydrate recognition by the lectinLecB from Pseudomonas aeruginosa J Mol Biol 331 861ndash870
Loris R Imberty A Beeckmans S Van Driessche ERead JS Bouckaert J et al (2003b) Crystal structureof Pterocarpus angolensis lectin in complex with glucosesucrose and turanose J Biol Chem 278 16297ndash16303
Lund B Lindberg F Marklund BI and Normark S(1987) The PapG protein is the a-D-galactopyranosyl-(1-4)-b-D-galactopyranose-binding adhesin of uropathogenicEscherichia coli Proc Natl Acad Sci USA 84 5898ndash5902
Min G Stolz M Zhou G Liang F Sebbel P StofflerD et al (2002) Localization of uroplakin Ia the urothelialreceptor for bacterial adhesin FimH on the six innerdomains of the 16 nm urothelial plaque particle J Mol Biol317 697ndash706
Miroux B and Walker JE (1996) Over-production of pro-teins in Escherichia coli mutant hosts that allow synthesisof some membrane proteins and globular proteins at highlevels J Mol Biol 260 289ndash298
Morris GM Goodsell DS Halliday RS Huey R HartWE Belew RK and Olson AJ (1998) Automateddocking using a Lamarckian genetic algorithm and anempirical binding free energy function J Comput Chem 191639ndash1662
Mulvey MA (2002) Adhesion and entry of uropathogenicEscherichia coli Cell Microbiol 4 257ndash271
Mulvey MA Lopez-Boado YS Wilson CL Roth RParks WC Heuser J and Hultgren SJ (1998) Induc-tion and evasion of host defenses by type 1-piliated uro-pathogenic Escherichia coli Science 282 1494ndash1497
Murshudov GN Vagin AA and Dodson EJ (1997)Refinement of macromolecular structures by the maxi-mum-likelihood method Acta Crystallogr D Biol Crystallogr53 240ndash255
Nagahori N Lee RT Nishimura S Pageacute D Roy Rand Lee YC (2002) Inhibition of adhesion of type 1 fim-briated Escherichia coli to highly mannosylated ligandsChembiochem 3 836ndash844
Navaza J (2001) Implementation of molecular replacementin AMoRe Acta Crystallogr D Biol Crystallogr 57 1367ndash1372
Neeser JR Koellreutter B and Wuersch P (1986)Oligomannoside-type glycopeptides inhibiting adhesion ofEscherichia coli strains mediated by type 1 pili preparationof potent inhibitors from plant glycoproteins Infect Immun52 428ndash436
Old DC (1972) Inhibition of the interaction between fimbrial
hemagglutinatinins and erythrocytes by D-mannose andother carbohydrates J Gen Microbiol 71 149ndash157
Orndorff PE Devapali A Palestrant S Wyse A EverettML Bollinger RR and Parker W (2004) Immunoglob-ulin-mediated agglutination of and biofilm formation byEscherichia coli K-12 require the type 1 pilus fiber InfectImmun 72 1929ndash1938
Oscarson S and Tedebark U (1995) Syntheses of deoxyanalogues of methyl 36-di-O-a-D-mannopyranosyl-a-D-mannopyranoside for studies of the binding site of con-canavalin A Carbohydr Res 278 271ndash287
Oscarson S and Tiden AK (1993) Syntheses of the octyland tetradecyl glycosides of 36-di-O-alpha-deuterium-mannopyranosyl-alpha-deuterium-mannopyranose and of34-di-O-alpha-deuterium-mannopyranosyl-alpha-deute-rium-mannopyranose ndash a new way for 24-di-O-protectionof mannopyranosides Carbohydr Res 247 323ndash328
Otwinowski Z and Minor W (1997) Processing of X-raydiffraction data collected in oscillation mode MethodsEnzymol 276 307ndash326
Powell HR (1999) The Rossmann Fourier autoindexingalgorithm in MOSFLM Acta Crystallogr D Biol Crystallogr55 1690ndash1695
Pratt LA and Kolter R (1998) Genetic analysis of Escher-ichia coli biofilm formation roles of flagella motility chemo-taxis and type I pili Mol Microbiol 30 285ndash293
Remaut H and Waksman G (2004) Structural biology ofbacterial pathogenesis Curr Opin Struct Biol 14 161ndash170
Ronald AR Nicolle LE Stamm E Krieger J WarrenJ Schaeffer A et al (2001) Urinary tract infection inadults research priorities and strategies Int J AntimicrobAgents 17 343ndash348
Sanner MF Olson AJ and Spehner JC (1996)Reduced surface an efficient way to compute molecularsurfaces Biopolymers 38 305ndash320
Sauer FG Futterer K Pinkner JS Dodson KW Hult-gren SJ and Waksman G (1999) Structural basis ofchaperone function and pilus biogenesis Science 2851058ndash1061
Sauer FG Knight SD Waksman GW and HultgrenSJ (2000) PapD-like chaperones and pilus biogenesisSemin Cell Dev Biol 11 27ndash34
Sauer FG Pinkner JS Waksman G and Hultgren SJ(2002) Chaperone priming of pilus subunits facilitates atopological transition that drives fiber formation Cell 111543ndash551
Schembri MA Hasman H and Klemm P (2000)Expression and purification of the mannose recognitiondomain of the FimH adhesin FEMS Microbiol Lett 188147ndash151
Schembri MA Kjaergaard K and Klemm P (2003) Glo-bal gene expression in Escherichia coli biofilms Mol Micro-biol 48 253ndash267
Schilling JD Mulvey MA Vincent CD Lorenz RGand Hultgren SJ (2001) Bacterial invasion augments epi-thelial cytokine responses to Escherichia coli through alipopolysaccharide-dependent mechanism J Immunol166 1148ndash1155
Silhavy TJ Szmelcman S Boos W and Schwartz M(1975) On the significance of the retention of ligand byprotein Proc Natl Acad Sci USA 72 2120ndash2124
A novel class of FimH high-affinity ligands 455
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
Svanborg C and Godaly G (1997) Bacterial virulence inurinary tract infection Infect Dis Clin North Am 11 513ndash529
Thanassi DG Saulino ET and Hultgren SJ (1998) Thechaperoneusher pathway a major terminal branch of thegeneral secretory pathway Curr Opin Struct Biol 1 223ndash231
Thankavel K Madison B Ikeda T Malaviya R ShahAH Arumugen PM and Abraham SN (1997) Local-ization of a domain in the FimH adhesin of Escherichia colitype 1 fimbriae capable of receptor recognition and use ofa domain-specific antibody to confer protection againstexperimental urinary tract infection J Clin Invest 1001123ndash1126
Vagin A and Teplyakov A (1997) MOLREP an automatedprogram for molecular replacement J Appl Crystallogr 301022ndash1025
Wu XR Sun TT and Medina JJ (1996) In vitro bindingof type 1-fimbriated Escherichia coli to uroplakins 1a and
1b relation to urinary tract infections Proc Natl Acad SciUSA 93 9630ndash9635
Xia Y Gally D Forsman-Semb K and Uhlin BE (2000)Regulatory cross-talk between adhesin operons in Escher-ichia coli inhibition of type 1 fimbriae expression by thePapB protein EMBO J 19 1450ndash1457
Zafriri D Ofek I Adar R Pocino M and Sharon N(1989) Inhibitory activity of cranberry juice on adherenceof type-1 and type-P fimbriated Escherichia coli to eukary-otic cells Antimicrob Agents Chemother 33 92ndash98
Zavialov AV Berglund J Pudney AF Fooks LJ Ibra-him TM MacIntyre S and Knight SD (2003) Structureand biogenesis of the capsular F1 antigen from Yersiniapestis preserved folding energy drives fiber formation Cell113 587ndash596
Zhou G Mo WJ Sebbel P Min G Neubert TA Glock-shuber R et al (2001) Uroplakin Ia is the urothelial recep-tor for uropathogenic Escherichia coli evidence from invitro FimH binding J Cell Sci 114 4095ndash4103

448 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
The measured affinity confirm the predictions that FimHbinds the monosacharide mannose exceptionally well(Kd = 23ndash41 mM) To investigate the contribution of thedifferent hydroxyl groups of mannose in the interactionwith FimH the binding of a series of hexoses to butylmannoside-free FimH was examined using SPR (Table 2)The effect of deoxygenation of the O2 hydroxyl group(methyl 2-deoxy-a-D-mannopyranoside) is clearly lessdestructive (100-fold reduction) than its equatorial positionas in glucose O2 is the only hydroxyl group of mannosethat interacts indirectly with the FimH pocket via a con-served water molecule (Hung et al 2002) Glucose onlydisplays a weak affinity for FimH about 4000 times lowerthan methyl a-D-mannoside Repositioning of both the O2and O4 hydroxyl groups as in galactose abolishes sugarbinding by FimH Similar to these results the inhibitoryconcentrations for bacterial adherence of methyl a-D-glucoside and galactose derivatives have been estimatedto be respectively 104-fold and 105-fold weaker than formethyl a-D-mannoside (Nagahori et al 2002)
In stark contrast with the other monosaccharides fruc-tose has a binding affinity only about 15-fold less thanmannose for FimH (Table 2) The decrease in affinity for
fructose relative to mannose agrees with studies of inhi-bition of adherence of type 1 piliated E coli by fructosein which fructose was found to be the active compound infruit juices (present at about 5) (Zafriri et al 1989)Fructose binding to FimH is stronger than the binding ofthe physiological globotetraoside receptor to PapGII theother well-characterized adhesin of UPEC strains (Lars-son et al 2003) The pyranose form of fructose is verywell superimposable onto mannose in the FimH bindingsite where similar interactions are established with FimHas for mannose except for the loss of the anomeric O1and an extra hydroxyl group (O2) on the C5 position ofmannose Probably however both changes of fructopyr-anoside compared with mannopyranoside could be easilyaccommodated by FimH The mannose O1 does notmake direct interactions within 45 Aring of the FimH bindingpocket (Fig 1B) The extra O2 group of fructose would notclash or interfere significantly with the binding pocket butit would make a 27 Aring van der Waals contact with the Ile52side-chain under the condition that the Ile52 side-chaindoes not move The crystal structure of the Pseudomonasaeruginosa lectin LecB also showed that fructopyranosebut not fructofuranose can substitute for mannose (Loris
Fig 6 Stereo diagrams of docked MeUm-baMan (A) and pNPaMan (B) in the FimH bind-ing site The crystallographically determined position of a-D-mannose is shown in orange for comparison
A novel class of FimH high-affinity ligands 449
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
et al 2003a) Hydrogen bonds of the more specificfucose complex are replaced by van der Waals interac-tions involving a hydroxyl group in the complexes with thelower-affinity binders mannose and fructopyranose as forthe proposed fructopyranoside binding to FimH
The fructose-containing disaccharides sucrose andturanose do not have an inhibitory effect and bind withaffinities similar to glucose Sucrose is a non-reducingsugar that has fructose locked in its furanose form Tura-nose on the other hand allows the interconversion of b-D-fructofuranose and b-D-fructopyranose eg both formsoccur in complex with a plant lectin from Pterocarpusangolensis (Loris et al 2003b) It is predictable howeverfrom the crystal structures that the glycosidic linkage ofglucose with C3 of fructose equivalent in position to C4of mannose would be disallowed in the tight FimH pocketSucrose and turanose thus most probably interact with theFimH mannose binding pocket via their glucose moietyresulting in similar low affinities
Our mass spectrometry and chemical analyses con-firms the crystallographic identification of the ligand boundto the FimH lectin domains as butyl a-D-mannoside(Figs 1B and 2) The mannose ring of the butyl a-D-mannoside coincides with the mannose bound to theFimCFimH complex (Hung et al 2002) and makes thesame interactions The four-carbon alkyl tail exits towardsthe back wall (loop b4ndashb5 and b10ndashb11) of the pocket(Fig 1A and B) making hydrophobic contacts to Tyr48Tyr137 and Ile52 The mass spectrometry data also showthat the bound butyl a-D-mannoside is present in LBmedium but absent from minimal medium Hence thesource of the alkyl mannoside is possibly the yeast extractused in preparation of LB medium Although we do notwant to speculate about the significance of this findingalkyl mannoside production by yeast has not beenreported and might warrant investigation
The butyl mannoside was retained by FimH despiteextensive dialysis The phenomenon of ligand retention byprotein was described as early as 1975 for periplasmicbinding proteins (Silhavy et al 1975) Applied to dialysisof butyl mannoside away from FimH grown in richmedium half-lives of up to 30 h can be estimated whichis far beyond the duration of a normal buffer exchangedialysis experiment In the present case the retainedligand does not appear to seriously affect [3H]-mannosedisplacement equilibrium binding experiments as wecould detect no significant differences in dissociation con-stants obtained using FimHtr2 produced in LB or minimalmedia (data not shown) In contrast proper SPR bindingcurves could not be obtained when butyl mannoside occu-pied the binding site Hence the displacement techniquemay be of more general use to obtain binding constantsfor proteins that tend to inadvertently retain ligands whichis a common trait of lectins
Surface plasmon resonance as well as equilibrium bind-ing and displacement measurements using [3H]-mannosewere carried out to obtain dissociation constants for a-D-mannose and a linear series of alkyl-substituted manno-sides to FimH (Table 1) Both equilibrium binding experi-ments show a very good agreement in their measurementof the carbohydrate affinity of the adhesin FimH bindsbutyl a-D-mannoside 15ndash30 times stronger than a-D-mannose The importance of the added hydrophobiccontacts is clearly demonstrated by the binding of alkylmannosides (Table 1) The best binding alkyl mannosidesbind a few hundred times stronger than mannose equiv-alent in affinity to the most tightly binding aromaticallysubstituted mannosides for FimH (Firon et al 1987) andof mannose dendrimers for type 1 piliated E coli (Fironet al 1984 1987 Nagahori et al 2002)
Interestingly the butyl moiety takes on significantlydifferent conformations in FimHtr1 and FimHtr2 (Fig 1C)indicating that intrinsic conformational flexibility in theaglycon is allowed Indeed in spite of the larger flexibilityexpected for longer alkyl mannosides the binding energydecreases linearly with increasing length of the alkyl chainbetween one and seven carbon atoms (Fig 5) The affinityreaches its maximum for mannose with a seven-carbonalkyl and SPR measurements suggest that the affinitydecreases from eight carbon atoms on Evidently theeighth carbon atom goes beyond the hydrophobic interac-tion surface and protein edge which is penalized with adecrease in affinity We propose that the affinity increaseresults from an increased interaction surface with thehydrophobic gateway Alkyl aglycons fit well in the tyrosinegate and as observed in the two crystal structures in atleast two different conformations that depend on theconformation of Tyr48 This flexibility may indicate thataglycons can interact rather non-specifically with thehydrophobic gateway nevertheless resulting in a 100- to1000-fold increase in affinity
FimH binding to ligands other than just mannose ispossibly of importance in terms of the different environ-ments that E coli will encounter ndash the urinary tract gas-trointestinal tract and in biofilms Alkyl mannosides havenot been previously recognized as strong binders to FimHAs they are easily synthesized and highly soluble in waterthey may be interesting as potential blocking agents forFimH-mediated adhesion Moreover they have beenshown to be good drug vehicles themselves acting aspermeability enhancers on tight junctions (Eley and Triu-malashetty 2001) As such they might be able to pene-trate into deeper cell layers of the epithelium to alsoprevent those bacteria that form a quiescent reservoirfrom causing recurrent infections (Mulvey et al 1998Justice et al 2004)
On the other hand the two aromatically substitutedmannosides pNPaMan and MeUmbaMan are known to
450 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
be particularly potent inhibitors of FimH-mediated bacte-rial adhesion (Firon et al 1987 Nagahori et al 2002)Based on adhesion inhibition studies using type 1 piliatedE coli and guinea pig ileal epithelial cells the relativeinhibitory potential of pNPaMan and MeUmbaMan havebeen estimated to be about 70 and 600 times strongerrespectively than that of methyl a-D-mannose (Fironet al 1987) Our measurements indicate an affinity ofFimH for pNPaMan and MeUmbaMan between 50 and90 times and between 110 and 200 times higher respec-tively than for methyl a-D-mannose in good agreementwith the adhesion inhibition results (Firon et al 1987)Dissociation constants predicted using AutoDock3 (Morriset al 1998) are similar to experimental values Thecomputational docking predicts that tight binding of thearomatically substituted mannosides pNPaMan andMeUmbaMan is accomplished similarly as for butyl a-D-mannoside with the aglycon interacting with the tyrosinegate
This is the first systematic study providing molecularbinding data for the FimH adhesin the prerequisite forcolonization by UPEC (Langermann and Ballou 2003)We also show crystallographic evidence that the lectinor receptor binding domain of FimH is a stable andfunctional entity Its structure is virtually identical to thesame domain in the FimCFimH complex indicating thata truncate corresponding to the lectin domain could pro-vide a good alternative for the development of a UTIvaccine (Langermann et al 1997 Langermann and Bal-lou 2003) However progress of this interesting UTI vac-cine candidate into clinical trials has not been reportedThe binding affinities of FimH at the molecular levelrelate convincingly well with the inhibitory concentrationsused in previous bacterial adhesion assays thus endors-ing the direct and conclusive correlation of FimH withbacterial adherence Alkyl mannosides have nanomolaraffinities for FimH which may overcome the significantobstacle of low affinity generally encountered in attemptsto develop carbohydrate-based drugs which interferewith microbial attachment to host cell surface glycocon-jugates New anti-adhesive drugs could be based onthese leads to allow specific interaction and targeting tothe urinary tract in order to prevent anticipated andrecurrent infections
Experimental procedures
Strains and constructs
FimH proteins from two different E coli strains the UPECstrain J96 (Hull et al 1981) and the E coli K-12 strainMG1655 (Blattner et al 1997) were used in this study TheFimH adhesins encoded by these two E coli strains areidentical FimHtr1 (residues 1ndash158 of UPEC J96 FimH) wasexpressed from plasmid pMMB91 transformed into E coli
C600 cells FimHtr2 (residues 1ndash158 of E coli MG1655FimH + carboxy-terminal 6-His tag) was expressed fromplasmid pPKL241 (Schembri et al 2000) transformed into Ecoli host strain HB101
Purification and crystallization
FimHtr1 Bacteria were grown in LB medium at 37infinC inducedat OD600 = 08 with 1 mM IPTG and harvested 2ndash3 h laterThe cells were resuspended in 20 mM Tris-HCl pH 80 5 mMEDTA and 30 sucrose and the periplasmic content wasextracted by osmotic shock upon 10-fold dilution with 20 mMTris-HCl pH 80 Purification of FimHtr1 was performed by ionexchange chromatography with MonoS at pH 45 followed byMonoQ (Amersham Biosciences) at pH 8 FimHtr1 was crys-tallized using vapour diffusion in hanging drops composed of3 ml of the precipitant solution (01 M Hepes pH 75 12PEG 6000 and 5 vv methyl-24-pentanediol) and 3 ml of12 mg ml-1 protein solution in 20 mM Tris-HCl pH 80 with150 mM NaCl
FimHtr2 Cells were grown in LB medium containing50 mg ml-1 ampicillin at 37infinC At OD600 = 06 the cells wereinduced with 5 mM IPTG Cells were harvested 2 h afterinduction and the periplasmic content was extracted Theprotein was purified on a Pharmacia HiTrap Chelating HP5 ml column (Pharmacia) loaded with Ni-chloride and elutedwith a sharp 0ndash500 mM imidazole gradient Fractions con-taining FimHtr2 were pooled dialysed against 50 mM sodiumacetate pH 525 and loaded onto a Mono S HR 8 ml columnThe protein was eluted with a 0ndash500 mM NaCl gradient anddialysed overnight against 20 mM MES pH 65 before crys-tallization Crystallization was performed at room tempera-ture by the vapour diffusion method with hanging dropsmade by mixing of 2 ml of 15 mg ml-1 protein solution with 2 mlof the precipitant solution (65 methyl-24-pentanediol100 mM cacodylate pH 65) Microseeding was employed tooptimize crystal quality and reproducibility Crystals suitablefor X-ray data collection generally appeared within 48 h aftermicroseeding
Structure determination
X-ray data for FimHtr1 were collected from a single crystalto 17 Aring resolution at the Advanced Photon SourceArgonne National Laboratory USA The crystal was flash-cooled to 100 K in the precipitant solution complementedwith 25 glycerol The data were processed and scaledusing the HKL2000 package (Otwinowski and Minor 1997)(Table S1 in Supplementary material) Data for FimHtr2
were collected from a single crystal under liquid-nitrogencryo conditions at 100 K at beamline ID29 ESRF Greno-ble France to 21 Aring Bragg spacing FimHtr2 data were pro-cessed and reduced using MOSFLM (Powell 1999) andscaled using SCALA (Collaborative computational project1994) (Table S1 in Supplementary material) The structuresof FimHtr1 and FimHtr2 were solved by molecular replace-ment using Amore (Navaza 2001) and MOLREP (Vaginand Teplyakov 1997) respectively with the FimH lectindomain from the FimCFimH complex (Choudhury et al
A novel class of FimH high-affinity ligands 451
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
1999) (PDB id 1QUN) as the search model Positional bulksolvent and isotropic temperature factor refinement wasperformed using CNS (Bruumlnger et al 1998) and REFMAC5(Murshudov et al 1997) respectively Model adjustmentand inspection were performed using Turbo-Frodo and O(Jones et al 1991) graphics programs respectively Refine-ment statistics are given in Table S1 Molecular graphicsillustrations were generated with Bobscript (Esnouf 1997)and Pymol 090 (DeLano Scientific LLC 1998ndash2003 httppymolsourceforgenet)
Mass spectrometry
The purified FimHtr1 protein (300 mg) that had been used forcrystallization was freeze-dried taken up in 100 ml of pyridineand 100 ml of acetic anhydride and incubated overnight atroom temperature in the dark for peracetylation The samplewas dried under nitrogen and taken up in chloroform (400 ml)The chloroform phase was extracted six times with waterdried and taken up in 200 ml of 50 aqueous methanol A1 ml aliquot was applied to a nano-scale RP column(75 mm yen 100 mm PepMap DionexLC Packings) using anUltimate nano-LC system (LC Packings) which was equili-brated with eluent A (H2Oacetonitrile 955 vv containing01 formic acid) at a flow rate of 100 nl min-1 After injectingthe sample the column was run isocratically for 5 min fol-lowed by a linear gradient to 100 eluent B (H2Oacetonitrile2080 vv containing 01 formic acid) in 20 min and a finalwash with 100 B for 5 min The system was directly coupledto an Esquire 3000 ESI-IT-MS (Bruker Daltonik) equippedwith an online nanospray source operating in the positive-ionmode For electrospray (900ndash1200 V) capillaries (360 mmOD 20 mm ID with 10 mm opening) from New Objective wereused The solvent was evaporated at 150infinC with a nitrogenstream of 6 l min-1
To verify the suspected source of the bound butyl manno-side mass spectrometry was performed on samples ofFimHtr2 grown in either LB or minimal M9 medium Proteinsamples were dialysed against water overnight before massspectrometry in order to reduce background from the Trisbuffer Samples were diluted to a concentration in the submi-cromolar range Electrospray mass spectra were recorded indirect inlet mode (50 aqueous methanol solutions) on aBruker Daltonics Esquire-LC (Billerica)
Carbohydrate constituent analysis
Samples were hydrolysed in 100 ml of 4 M aqueous trifluoro-acetic acid (Sigma) at 100infinC for 4 h and dried under astream of nitrogen Monosaccharides were converted intotheir anthranilic acid derivatives by reductive aminationresolved by RP-HPLC and detected by fluorescence (Anu-mula 1994)
Carbohydrates for binding studies
[3H]-mannose was obtained from Amersham Methyl a-D-mannoside glucose galactose fructose sucrose turanosep-nitrophenyl a-D-mannopyranoside (pNPaMan) and 4-methylumbelliferyl a-D-mannopyranoside (MeUmbaMan)
were obtained from Sigma Monomannosylated bovineserum albumin (BSA-Man) was kindly provided by MedIm-mune We synthesized methyl 2-deoxy-a-D-mannopyrano-side (Oscarson and Tedebark 1995) Alkyl mannosides weresynthesized through silver triflate-promoted couplings of thecorresponding alcohol with 2346-tetra-O-benzoyl-a-D-mannopyranosyl bromide followed by Zempleacuten deacylationof the obtained protected alkyl mannosides according to theprocedure reported for the octyl and tetradecyl mannosides(Oscarson and Tiden 1993)
Binding studies
Competition experiments using SPR To obtain butyl man-noside-free FimHtr1 the FimH truncate was overproducedwith a carboxy-terminal 6-His tag after transformation into Ecoli C43 (DE3) (Miroux and Walker 1996) and the bacteriagrown in minA minimal medium supplemented with the 20amino acids (40 mg l-1) thiamin (2 mg l-1) and biotin (2 mgl-1) Purification of the His-tagged protein was performedusing Ni-NTA affinity chromatography (Qiagen) in 50 mM Tris-HCl pH 8 and 300 mM NaCl Elution was in the same solutioncomplemented with 500 mM imidazole Subsequently pooledand concentrated FimH fractions were gel filtrated over aSuperdex G75 column (Pharmacia) in 20 mM Tris-HCl pH 8
We used Fab fragments of a monoclonal antibody againstthe mannose-binding site of FimH to determine the solutionaffinity of FimHndashcarbohydrate interactions The monoclonalantibody IC10 was produced by a mouse hybridoma cell lineat Medimmune and its Fab fragments were prepared andpurified The Fabs were covalently immobilized via aminecoupling at 1000 Resonance Units (1000 pg ligand mm-2) inflowcell Fc2 on a CM5 biasensor chip (BIAapplications Hand-book Biacore AB Uppsala Sweden) Flowcell Fc1 was usedas the reference surface and was treated in the same wayas Fc2 except that it was blocked immediately after activationThe immobilization buffer was 100 mM sodium acetatepH 50 with 100 mM NaCl All SPR measurements were per-formed on a Biacore3000trade
The kinetic constants of binding of FimH to the immobilizedantibody were measured with different concentrations ofFimH (2000 nM 1000 nM 500 nM 250 nM 125 nM625 nM 312 nM 156 nM 78 nM) in running buffer [phos-phate-buffered saline (PBS) with 0005 surfactant P20 and3 mM EDTA] on both flowcells Fc1 and Fc2 simultaneouslyat a flow rate of 30 ml min-1 and at 298 K Complete dissoci-ation of FimH was performed with running buffer beforestarting a new binding cycle For all measurements the asso-ciation time was 3 min and the dissociation time was 30 minAll binding cycles were performed in duplicate including azero concentration cycle of FimH (injection of running buffer)All analyses were performed with the BIAeval software ALangmuir binding isotherm with a 11 stoichiometry was fittedto the data from both the association phase and the dissoci-ation phase after subtraction of the reference and zero con-centration data The kinetic constants ka and kd and themaximal binding Rmax were obtained and the dissociationconstant Kd was calculated
The equilibrium dissociation constants Kd of FimHndashsaccha-ride interactions were determined in a competition experi-ment using the same chip as for the previous experiment
452 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
As a control the Fabs of monoclonal antibody IA7 as wellas BSA-Man were also immobilized Samples containing afixed concentration of FimH were used (close to the Kd of theFimHndashantibody interaction) in combination with varying con-centrations of saccharide First 10-fold dilutions of the sac-charide solution were used to determine the concentrationrange for binding of the saccharide to FimH A Langmuirbinding isotherm with a 11 stoichiometry was fitted to thedata using the kinetic constants and Rmax from the foregoingexperiment to obtain the concentrations of FimH that werefree ([FimH]free) to bind the antibody immobilized on the chipSecond the concentration range of the saccharide wasextended and adapted to ensure accurate fitting and theequilibrium binding constant of the FimHndashsaccharide interac-tion was derived from the curve of [FimH]free against concen-tration of saccharide Every measurement was repeated atleast twice including testing different protein batches andwhenever possible also different saccharide stock solutions(typically 200 mM)
Equilibrium binding studies using radioactively labelledmannose Mannose-free FimHtr2 was produced inM9 minimal medium and purified as described above forFimHtr2 Alkyl mannosides were dissolved in double distilledwater pNPaMan and MeUmbaMan (6 mg each) were dis-solved in 20 ml of DMSO and diluted to 20 mM using doubledistilled water Binding experiments were performed at 310 Kusing six different concentrations of [3H]-a-D-mannose (finalconcentrations 435 mM 290 mM 193 mM 129 mM 86 mMand 57 mM) Protein (180 ml) at a concentration of about500 nM was mixed with 20 ml of the radioactive ligand [3H]-a-D-mannose and incubated at 37degC for 20 min To separatefree ligand from bound the mixture was rapidly filtratedthrough a Protran BA 85 cellulose nitrate filter (Schleicherand Schuell) and washed once with 1 ml of ice-cold 1yen PBSFilter-bound radioactivity was measured by scintillation spec-trometry within 24 h Displacement experiments were per-formed using six different concentrations of inhibitor (finalconcentrations in the range 00ndash435 mM) in the presence of435 mM [3H]-a-D-mannose Twenty microlitres of radioactiveligand 20 ml of inhibitor and 160 ml of protein (500 nM) weremixed and the experiments were performed as describedabove All experiments were performed in duplicates and thedata presented are mean values of at least two independentmeasurements For the determination of Kd for a-D-mannosean hyperbolic curve (y = P1x(P2 + x) where P1 is an arbitraryscale factor and P2 = Kd) was fitted to the data For the dis-placement experiments the curve y = P1(P2 + x) where P1
is an arbitrary scale factor and P2 is the concentration of theinhibitor displacing 50 of the labelled ligand [I]05 was usedinstead To calculate the inhibitor dissociation constant (KI)the Cheng and Prusoff equation [KI = [I]05[L]KL + 1) KL is theconstant of dissociation for the ligand] (Cheng and Prusoff1973) was used when both the concentration of the radio-active ligand (L) and the displacing agent (I) are in excessover the protein (LT gtgt PT IT gtgt PT T indicates total concen-tration) For the two strong inhibitors pNPaMan and MeUm-baMan where IT was not in excess over PT the equationof KI = IT[(1 - Y)Y(LTKL) - 1] - PTKLYLT where Y is thefraction of the ligand bound in presence of the inhibitor wasused instead (Horovitz and Levitzki 1987)
Computational docking
Computational docking studies were performed usingAutoDock3 (Morris et al 1998) In AutoDock3 the proteinis represented by a grid of affinity potentials Hence theprotein conformation is not allowed to change during thedocking procedure Flexibility of ligands is taken intoaccount by allowing rotation around flexible torsion anglesThe docked energies are calculated as the sum of the inter-molecular interaction energy and the internal energy of theligand The search for the best interaction energy was car-ried out by means of the Lamarckian genetic algorithmLigand structures were obtained from the Protein DataBank and prepared for docking by adding hydrogens andcharges using AutoDockTools (Sanner et al 1996) Eachsimulation consisted of 100 independent runs with a popu-lation size of 200 500 generations and a maximum of25 000 000 energy evaluations Solutions were rankedbased on their docking energies and similar solutions wereclustered (cluster cut-off rmsd lt 1 Aring) For both ligandsthe top solutions were evaluated using the graphics pro-gram O (Jones et al 1991)
Acknowledgements
We thank Andrej Zavialov and Maringhns Ehrenberg for help withequilibrium binding experiments and Thomas Norberg forassistance with mass spectrometry Anthony DeFusco fromMedimmune provided us with mannosylated bovine serumalbumin We are grateful to Fred Sauer for assistance withdata collection Thanks to Maor Bar-Peled of the CCC at theUniversity of Georgia Athens USA for his suggestion toexpress FimH in minimal medium We acknowledge the useof beamline 19BM of the Structural Biology Center at theAdvanced Photon Source Argonne National LaboratoryChicago IL USA and of beamline ID29 ESRF GrenobleFrance JB is a postdoctoral fellow of the Fonds voor Weten-schappelijk Onderzoek-Vlaanderen which also supportedthe BiaCore3000 instrument (FWONL35) This work was sup-ported by grants from the Swedish Research Council andfrom the Strategic Research Foundation (Structural BiologyNetwork) to SDK
Supplementary material
The following material is available from httpwwwblackwellpublishingcomproductsjournalssuppmatmmimmi4415mmi4415smhtmTable S1 Crystallographic data and refinement statistics
References
Anumula KR (1994) Quantitative determination ofmonosaccharides in glycoproteins by high-performanceliquid chromatography with highly sensitive fluorescencedetection Anal Biochem 220 275ndash283
Bahrani-Mougeot FK Buckles EL Lockatell CV HebelJR Johnson DE Tang CM and Donnenberg MS(2002) Type 1 fimbriae and extracellular polysaccharidesare preeminent uropathogenic Escherichia coli virulence
A novel class of FimH high-affinity ligands 453
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
determinants in the murine urinary tract Mol Microbiol 451079ndash1093
Berglund J and Knight SD (2003) Structural basis forbacterial adhesion in the urinary tract Glycobiol Med 53533ndash52
Blattner FR Plunkett G Bloch CA Perna NT BurlandV Riley M et al (1997) The complete genome sequenceof Escherichia coli K-12 Science 277 1453ndashbullbull
Bloch CA Stocker BA and Orndorff PE (1992) A keyrole for type 1 pili in enterobacterial communicability MolMicrobiol 6 697ndash701
Bork P Holm L and Sander C (1994) The immunoglob-ulin fold ndash structural classification sequence patterns andcommon core J Mol Biol 242 309ndash320
Brinton CC (1959) Non-flagellar appendages of bacteriaNature 183 782ndash786
Bruumlnger AT Adams PD Clore GM DeLano WLGros P Grosse-Kunstleve RW et al (1998) Crystallog-raphy and NMR system a new software suite for macro-molecular structure determination Acta Crystallogr D BiolCrystallogr 54 905ndash921
Buchanan K Falkow S Hull RA and Hull SI (1985)Frequency among Enterobacteriaceae of the DNA-sequences encoding type-1 pili J Bacteriol 162 799ndash803
Buts L Bouckaert J De Genst E Loris R Oscarson SLahmann M et al (2003) The fimbrial adhesin F17-G ofenterotoxigenic Escherichia coli has an immunoglobulin-like lectin domain that binds N-acetylglucosamine MolMicrobiol 49 705ndash715
Cheng Y and Prusoff WH (1973) Relationship betweeninhibition constant (K1) and concentration of inhibitor whichcauses 50 per cent inhibition (I50) of an enzymatic reac-tion Biochem Pharmacol 22 3099ndash3108
Choudhury D Thompson A Stojanoff V LangermannS Pinkner J Hultgren SJ and Knight SD (1999) X-ray structure of the FimCndashFimH chaperonendashadhesin com-plex from uropathogenic Escherichia coli Science 2851061ndash1066
Collaborative computational project n4 (1994) The CCP4suite programs for crystallography Acta Crystallogr D BiolCrystallogr 50 760ndash763
Dodson KW Pinkner JS Rose T Magnusson G Hult-gren SJ and Waksman G (2001) Structural basis of theinteraction of the pyelonephritic Escherichia coli adhesin toits human kidney receptor Cell 105 733ndash743
Eley JG and Triumalashetty P (2001) In vitro assessmentof alkylglycosides as permeability enhancers AAPSPharmSciTech 2 (3) Article 19
Esnouf RM (1997) An extensively modified version of Mol-script that includes greatly enhanced coloring capabilitiesJ Mol Graph Model 15 132ndash134
Firon N Ofek I and Sharon N (1982) Interaction ofmannose-containing oligosaccharides with the fimbriallectin of Escherichia coli Biochem Biophys Res Commun105 1426ndash1432
Firon N Ofek I and Sharon N (1983) Carbohydrate spec-ificity of the surface lectins of Escherichia coli Klebsiellapneumoniae and Salmonella typhimurium Carbohydr Res120 235ndash249
Firon N Ofek I and Sharon N (1984) Carbohydrate-
binding sites of the mannose-specific fimbrial lectins ofEnterobacteria Infect Immun 43 1088ndash1090
Firon N Ashkenazis S Mirelman D Ofek I and SharonN (1987) Aromatic a-glycosides of mannose are powerfulinhibitors of the adherence of type 1 fimbriated Escherichiacoli to yeast and intestinal cells Infect Immun 55 472ndash476
Fukiya S Mizoguchi H Tobe T and Mori H (2004)Extensive genomic diversity in pathogenic Escherichia coliand Shigella strains revealed by comparative genomichybridization microarray J Bacteriol 186 3911ndash3921
Hooton TM and Stamm WE (1997) Diagnosis and treat-ment of uncomplicated urinary tract infection Infect DisClin North Am 11 551ndash581
Horovitz A and Levitzki A (1987) An accurate method fordetermination of receptor ligand and enzyme-inhibitor dis-sociation-constants from displacement curves Proc NatlAcad Sci USA 84 6654ndash6658
Hull RA Gill RE Hsu P Minshaw BH and Falkow S(1981) Construction and expression of recombinant plas-mids encoding type 1 and D-mannose-resistant pili from aurinary tract infection Escherichia coli isolate Infect Immun33 933ndash938
Hultgren SJ Porter TN Schaeffer AJ and Duncan JL(1985) Role of type 1 pili and effects of phase variation onlower urinary tract infections produced by Escherichia coliInfect Immun 50 370ndash377
Hung C-S Bouckaert J Hung DL Pinkner J WinbergC Defusco A et al (2002) Structural basis of tropism ofEscherichia coli to the bladder during urinary tract infec-tion Mol Microbiol 44 903ndash915
Jones TA Zou JY Cowan SW and Kjeldgaard M(1991) Improved methods for building protein models inelectron density maps and the location of errors in thesemodels Acta Crystallogr A 47 110ndash119
Justice SS Hung C-S Theriot JA Fletcher DAAnderson GG Footer MJ and Hulgren SJ (2004)Differentiation and developmental pathways of uropatho-genic Escherichia coli in urinary tract pathogenesis ProcNatl Acad Sci USA 101 1333ndash1338
Kallenius G Mollby R Svenson SB Windberg J Lun-dblud A Svenson S and Cedergen B (1980) The Pk
antigen as receptor for the haemagglutinin of pyelonephri-togenic Escherichia coli FEMS Microbiol Lett 8 297ndash302
Knight SD Berglund J and Choudhury D (2000) Bacte-rial adhesins structural studies reveal chaperone functionand pilus biogenesis Curr Opin Chem Biol 4 653ndash660
Langermann S and Ballou WR (2003) Development of arecombinant FimCH vaccine for urinary tract infectionsAdv Exp Med Biol 539 635ndash653
Langermann S Palaszynski S Barnhart M Auguste GPinkner JS Burlein J et al (1997) Prevention ofmucosal Escherichia coli infection by FimH-adhesin-basedsystemic vaccination Science 276 607ndash611
Langermann S Mollby R Burlein JE Palaszynski SRAuguste CG Defusco A et al (2000) Vaccination withFimH adhesin protects cynomolgus monkeys from coloni-zation and infection by uropathogenic Escherichia coli JInfect Dis 181 774ndash778
Larsson A Ohlsson J Dodson KW Hultgren SJ Nils-son U and Kihlberg J (2003) Quantitative studies of thebinding of the class II PapG adhesin from uropathogenic
454 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
Escherichia coli to oligosaccharides Bioorg Med Chem 112255ndash2261
Leffler H and Svanborg-Eden C (1980) Chemical identifi-cation of a glycosphingolipid receptor for Escherichia coliattaching to human urinary tract epithelial cells and agglu-tinating human erythrocytes FEMS Microbiol Lett 8 127ndash134
Lindhorst TK Kieburg C and Krallmann-Wenzel U(1998) Inhibition of the type 1 fimbriae-mediated adhesionof Escherichia coli to erythrocytes by multiantennary alpha-mannosyl clusters the effect of multivalency Glycoconj J15 605ndash613
Loris R Tielker D Jaeger KE and Wyns L (2003a)Structural basis of carbohydrate recognition by the lectinLecB from Pseudomonas aeruginosa J Mol Biol 331 861ndash870
Loris R Imberty A Beeckmans S Van Driessche ERead JS Bouckaert J et al (2003b) Crystal structureof Pterocarpus angolensis lectin in complex with glucosesucrose and turanose J Biol Chem 278 16297ndash16303
Lund B Lindberg F Marklund BI and Normark S(1987) The PapG protein is the a-D-galactopyranosyl-(1-4)-b-D-galactopyranose-binding adhesin of uropathogenicEscherichia coli Proc Natl Acad Sci USA 84 5898ndash5902
Min G Stolz M Zhou G Liang F Sebbel P StofflerD et al (2002) Localization of uroplakin Ia the urothelialreceptor for bacterial adhesin FimH on the six innerdomains of the 16 nm urothelial plaque particle J Mol Biol317 697ndash706
Miroux B and Walker JE (1996) Over-production of pro-teins in Escherichia coli mutant hosts that allow synthesisof some membrane proteins and globular proteins at highlevels J Mol Biol 260 289ndash298
Morris GM Goodsell DS Halliday RS Huey R HartWE Belew RK and Olson AJ (1998) Automateddocking using a Lamarckian genetic algorithm and anempirical binding free energy function J Comput Chem 191639ndash1662
Mulvey MA (2002) Adhesion and entry of uropathogenicEscherichia coli Cell Microbiol 4 257ndash271
Mulvey MA Lopez-Boado YS Wilson CL Roth RParks WC Heuser J and Hultgren SJ (1998) Induc-tion and evasion of host defenses by type 1-piliated uro-pathogenic Escherichia coli Science 282 1494ndash1497
Murshudov GN Vagin AA and Dodson EJ (1997)Refinement of macromolecular structures by the maxi-mum-likelihood method Acta Crystallogr D Biol Crystallogr53 240ndash255
Nagahori N Lee RT Nishimura S Pageacute D Roy Rand Lee YC (2002) Inhibition of adhesion of type 1 fim-briated Escherichia coli to highly mannosylated ligandsChembiochem 3 836ndash844
Navaza J (2001) Implementation of molecular replacementin AMoRe Acta Crystallogr D Biol Crystallogr 57 1367ndash1372
Neeser JR Koellreutter B and Wuersch P (1986)Oligomannoside-type glycopeptides inhibiting adhesion ofEscherichia coli strains mediated by type 1 pili preparationof potent inhibitors from plant glycoproteins Infect Immun52 428ndash436
Old DC (1972) Inhibition of the interaction between fimbrial
hemagglutinatinins and erythrocytes by D-mannose andother carbohydrates J Gen Microbiol 71 149ndash157
Orndorff PE Devapali A Palestrant S Wyse A EverettML Bollinger RR and Parker W (2004) Immunoglob-ulin-mediated agglutination of and biofilm formation byEscherichia coli K-12 require the type 1 pilus fiber InfectImmun 72 1929ndash1938
Oscarson S and Tedebark U (1995) Syntheses of deoxyanalogues of methyl 36-di-O-a-D-mannopyranosyl-a-D-mannopyranoside for studies of the binding site of con-canavalin A Carbohydr Res 278 271ndash287
Oscarson S and Tiden AK (1993) Syntheses of the octyland tetradecyl glycosides of 36-di-O-alpha-deuterium-mannopyranosyl-alpha-deuterium-mannopyranose and of34-di-O-alpha-deuterium-mannopyranosyl-alpha-deute-rium-mannopyranose ndash a new way for 24-di-O-protectionof mannopyranosides Carbohydr Res 247 323ndash328
Otwinowski Z and Minor W (1997) Processing of X-raydiffraction data collected in oscillation mode MethodsEnzymol 276 307ndash326
Powell HR (1999) The Rossmann Fourier autoindexingalgorithm in MOSFLM Acta Crystallogr D Biol Crystallogr55 1690ndash1695
Pratt LA and Kolter R (1998) Genetic analysis of Escher-ichia coli biofilm formation roles of flagella motility chemo-taxis and type I pili Mol Microbiol 30 285ndash293
Remaut H and Waksman G (2004) Structural biology ofbacterial pathogenesis Curr Opin Struct Biol 14 161ndash170
Ronald AR Nicolle LE Stamm E Krieger J WarrenJ Schaeffer A et al (2001) Urinary tract infection inadults research priorities and strategies Int J AntimicrobAgents 17 343ndash348
Sanner MF Olson AJ and Spehner JC (1996)Reduced surface an efficient way to compute molecularsurfaces Biopolymers 38 305ndash320
Sauer FG Futterer K Pinkner JS Dodson KW Hult-gren SJ and Waksman G (1999) Structural basis ofchaperone function and pilus biogenesis Science 2851058ndash1061
Sauer FG Knight SD Waksman GW and HultgrenSJ (2000) PapD-like chaperones and pilus biogenesisSemin Cell Dev Biol 11 27ndash34
Sauer FG Pinkner JS Waksman G and Hultgren SJ(2002) Chaperone priming of pilus subunits facilitates atopological transition that drives fiber formation Cell 111543ndash551
Schembri MA Hasman H and Klemm P (2000)Expression and purification of the mannose recognitiondomain of the FimH adhesin FEMS Microbiol Lett 188147ndash151
Schembri MA Kjaergaard K and Klemm P (2003) Glo-bal gene expression in Escherichia coli biofilms Mol Micro-biol 48 253ndash267
Schilling JD Mulvey MA Vincent CD Lorenz RGand Hultgren SJ (2001) Bacterial invasion augments epi-thelial cytokine responses to Escherichia coli through alipopolysaccharide-dependent mechanism J Immunol166 1148ndash1155
Silhavy TJ Szmelcman S Boos W and Schwartz M(1975) On the significance of the retention of ligand byprotein Proc Natl Acad Sci USA 72 2120ndash2124
A novel class of FimH high-affinity ligands 455
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
Svanborg C and Godaly G (1997) Bacterial virulence inurinary tract infection Infect Dis Clin North Am 11 513ndash529
Thanassi DG Saulino ET and Hultgren SJ (1998) Thechaperoneusher pathway a major terminal branch of thegeneral secretory pathway Curr Opin Struct Biol 1 223ndash231
Thankavel K Madison B Ikeda T Malaviya R ShahAH Arumugen PM and Abraham SN (1997) Local-ization of a domain in the FimH adhesin of Escherichia colitype 1 fimbriae capable of receptor recognition and use ofa domain-specific antibody to confer protection againstexperimental urinary tract infection J Clin Invest 1001123ndash1126
Vagin A and Teplyakov A (1997) MOLREP an automatedprogram for molecular replacement J Appl Crystallogr 301022ndash1025
Wu XR Sun TT and Medina JJ (1996) In vitro bindingof type 1-fimbriated Escherichia coli to uroplakins 1a and
1b relation to urinary tract infections Proc Natl Acad SciUSA 93 9630ndash9635
Xia Y Gally D Forsman-Semb K and Uhlin BE (2000)Regulatory cross-talk between adhesin operons in Escher-ichia coli inhibition of type 1 fimbriae expression by thePapB protein EMBO J 19 1450ndash1457
Zafriri D Ofek I Adar R Pocino M and Sharon N(1989) Inhibitory activity of cranberry juice on adherenceof type-1 and type-P fimbriated Escherichia coli to eukary-otic cells Antimicrob Agents Chemother 33 92ndash98
Zavialov AV Berglund J Pudney AF Fooks LJ Ibra-him TM MacIntyre S and Knight SD (2003) Structureand biogenesis of the capsular F1 antigen from Yersiniapestis preserved folding energy drives fiber formation Cell113 587ndash596
Zhou G Mo WJ Sebbel P Min G Neubert TA Glock-shuber R et al (2001) Uroplakin Ia is the urothelial recep-tor for uropathogenic Escherichia coli evidence from invitro FimH binding J Cell Sci 114 4095ndash4103

A novel class of FimH high-affinity ligands 449
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
et al 2003a) Hydrogen bonds of the more specificfucose complex are replaced by van der Waals interac-tions involving a hydroxyl group in the complexes with thelower-affinity binders mannose and fructopyranose as forthe proposed fructopyranoside binding to FimH
The fructose-containing disaccharides sucrose andturanose do not have an inhibitory effect and bind withaffinities similar to glucose Sucrose is a non-reducingsugar that has fructose locked in its furanose form Tura-nose on the other hand allows the interconversion of b-D-fructofuranose and b-D-fructopyranose eg both formsoccur in complex with a plant lectin from Pterocarpusangolensis (Loris et al 2003b) It is predictable howeverfrom the crystal structures that the glycosidic linkage ofglucose with C3 of fructose equivalent in position to C4of mannose would be disallowed in the tight FimH pocketSucrose and turanose thus most probably interact with theFimH mannose binding pocket via their glucose moietyresulting in similar low affinities
Our mass spectrometry and chemical analyses con-firms the crystallographic identification of the ligand boundto the FimH lectin domains as butyl a-D-mannoside(Figs 1B and 2) The mannose ring of the butyl a-D-mannoside coincides with the mannose bound to theFimCFimH complex (Hung et al 2002) and makes thesame interactions The four-carbon alkyl tail exits towardsthe back wall (loop b4ndashb5 and b10ndashb11) of the pocket(Fig 1A and B) making hydrophobic contacts to Tyr48Tyr137 and Ile52 The mass spectrometry data also showthat the bound butyl a-D-mannoside is present in LBmedium but absent from minimal medium Hence thesource of the alkyl mannoside is possibly the yeast extractused in preparation of LB medium Although we do notwant to speculate about the significance of this findingalkyl mannoside production by yeast has not beenreported and might warrant investigation
The butyl mannoside was retained by FimH despiteextensive dialysis The phenomenon of ligand retention byprotein was described as early as 1975 for periplasmicbinding proteins (Silhavy et al 1975) Applied to dialysisof butyl mannoside away from FimH grown in richmedium half-lives of up to 30 h can be estimated whichis far beyond the duration of a normal buffer exchangedialysis experiment In the present case the retainedligand does not appear to seriously affect [3H]-mannosedisplacement equilibrium binding experiments as wecould detect no significant differences in dissociation con-stants obtained using FimHtr2 produced in LB or minimalmedia (data not shown) In contrast proper SPR bindingcurves could not be obtained when butyl mannoside occu-pied the binding site Hence the displacement techniquemay be of more general use to obtain binding constantsfor proteins that tend to inadvertently retain ligands whichis a common trait of lectins
Surface plasmon resonance as well as equilibrium bind-ing and displacement measurements using [3H]-mannosewere carried out to obtain dissociation constants for a-D-mannose and a linear series of alkyl-substituted manno-sides to FimH (Table 1) Both equilibrium binding experi-ments show a very good agreement in their measurementof the carbohydrate affinity of the adhesin FimH bindsbutyl a-D-mannoside 15ndash30 times stronger than a-D-mannose The importance of the added hydrophobiccontacts is clearly demonstrated by the binding of alkylmannosides (Table 1) The best binding alkyl mannosidesbind a few hundred times stronger than mannose equiv-alent in affinity to the most tightly binding aromaticallysubstituted mannosides for FimH (Firon et al 1987) andof mannose dendrimers for type 1 piliated E coli (Fironet al 1984 1987 Nagahori et al 2002)
Interestingly the butyl moiety takes on significantlydifferent conformations in FimHtr1 and FimHtr2 (Fig 1C)indicating that intrinsic conformational flexibility in theaglycon is allowed Indeed in spite of the larger flexibilityexpected for longer alkyl mannosides the binding energydecreases linearly with increasing length of the alkyl chainbetween one and seven carbon atoms (Fig 5) The affinityreaches its maximum for mannose with a seven-carbonalkyl and SPR measurements suggest that the affinitydecreases from eight carbon atoms on Evidently theeighth carbon atom goes beyond the hydrophobic interac-tion surface and protein edge which is penalized with adecrease in affinity We propose that the affinity increaseresults from an increased interaction surface with thehydrophobic gateway Alkyl aglycons fit well in the tyrosinegate and as observed in the two crystal structures in atleast two different conformations that depend on theconformation of Tyr48 This flexibility may indicate thataglycons can interact rather non-specifically with thehydrophobic gateway nevertheless resulting in a 100- to1000-fold increase in affinity
FimH binding to ligands other than just mannose ispossibly of importance in terms of the different environ-ments that E coli will encounter ndash the urinary tract gas-trointestinal tract and in biofilms Alkyl mannosides havenot been previously recognized as strong binders to FimHAs they are easily synthesized and highly soluble in waterthey may be interesting as potential blocking agents forFimH-mediated adhesion Moreover they have beenshown to be good drug vehicles themselves acting aspermeability enhancers on tight junctions (Eley and Triu-malashetty 2001) As such they might be able to pene-trate into deeper cell layers of the epithelium to alsoprevent those bacteria that form a quiescent reservoirfrom causing recurrent infections (Mulvey et al 1998Justice et al 2004)
On the other hand the two aromatically substitutedmannosides pNPaMan and MeUmbaMan are known to
450 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
be particularly potent inhibitors of FimH-mediated bacte-rial adhesion (Firon et al 1987 Nagahori et al 2002)Based on adhesion inhibition studies using type 1 piliatedE coli and guinea pig ileal epithelial cells the relativeinhibitory potential of pNPaMan and MeUmbaMan havebeen estimated to be about 70 and 600 times strongerrespectively than that of methyl a-D-mannose (Fironet al 1987) Our measurements indicate an affinity ofFimH for pNPaMan and MeUmbaMan between 50 and90 times and between 110 and 200 times higher respec-tively than for methyl a-D-mannose in good agreementwith the adhesion inhibition results (Firon et al 1987)Dissociation constants predicted using AutoDock3 (Morriset al 1998) are similar to experimental values Thecomputational docking predicts that tight binding of thearomatically substituted mannosides pNPaMan andMeUmbaMan is accomplished similarly as for butyl a-D-mannoside with the aglycon interacting with the tyrosinegate
This is the first systematic study providing molecularbinding data for the FimH adhesin the prerequisite forcolonization by UPEC (Langermann and Ballou 2003)We also show crystallographic evidence that the lectinor receptor binding domain of FimH is a stable andfunctional entity Its structure is virtually identical to thesame domain in the FimCFimH complex indicating thata truncate corresponding to the lectin domain could pro-vide a good alternative for the development of a UTIvaccine (Langermann et al 1997 Langermann and Bal-lou 2003) However progress of this interesting UTI vac-cine candidate into clinical trials has not been reportedThe binding affinities of FimH at the molecular levelrelate convincingly well with the inhibitory concentrationsused in previous bacterial adhesion assays thus endors-ing the direct and conclusive correlation of FimH withbacterial adherence Alkyl mannosides have nanomolaraffinities for FimH which may overcome the significantobstacle of low affinity generally encountered in attemptsto develop carbohydrate-based drugs which interferewith microbial attachment to host cell surface glycocon-jugates New anti-adhesive drugs could be based onthese leads to allow specific interaction and targeting tothe urinary tract in order to prevent anticipated andrecurrent infections
Experimental procedures
Strains and constructs
FimH proteins from two different E coli strains the UPECstrain J96 (Hull et al 1981) and the E coli K-12 strainMG1655 (Blattner et al 1997) were used in this study TheFimH adhesins encoded by these two E coli strains areidentical FimHtr1 (residues 1ndash158 of UPEC J96 FimH) wasexpressed from plasmid pMMB91 transformed into E coli
C600 cells FimHtr2 (residues 1ndash158 of E coli MG1655FimH + carboxy-terminal 6-His tag) was expressed fromplasmid pPKL241 (Schembri et al 2000) transformed into Ecoli host strain HB101
Purification and crystallization
FimHtr1 Bacteria were grown in LB medium at 37infinC inducedat OD600 = 08 with 1 mM IPTG and harvested 2ndash3 h laterThe cells were resuspended in 20 mM Tris-HCl pH 80 5 mMEDTA and 30 sucrose and the periplasmic content wasextracted by osmotic shock upon 10-fold dilution with 20 mMTris-HCl pH 80 Purification of FimHtr1 was performed by ionexchange chromatography with MonoS at pH 45 followed byMonoQ (Amersham Biosciences) at pH 8 FimHtr1 was crys-tallized using vapour diffusion in hanging drops composed of3 ml of the precipitant solution (01 M Hepes pH 75 12PEG 6000 and 5 vv methyl-24-pentanediol) and 3 ml of12 mg ml-1 protein solution in 20 mM Tris-HCl pH 80 with150 mM NaCl
FimHtr2 Cells were grown in LB medium containing50 mg ml-1 ampicillin at 37infinC At OD600 = 06 the cells wereinduced with 5 mM IPTG Cells were harvested 2 h afterinduction and the periplasmic content was extracted Theprotein was purified on a Pharmacia HiTrap Chelating HP5 ml column (Pharmacia) loaded with Ni-chloride and elutedwith a sharp 0ndash500 mM imidazole gradient Fractions con-taining FimHtr2 were pooled dialysed against 50 mM sodiumacetate pH 525 and loaded onto a Mono S HR 8 ml columnThe protein was eluted with a 0ndash500 mM NaCl gradient anddialysed overnight against 20 mM MES pH 65 before crys-tallization Crystallization was performed at room tempera-ture by the vapour diffusion method with hanging dropsmade by mixing of 2 ml of 15 mg ml-1 protein solution with 2 mlof the precipitant solution (65 methyl-24-pentanediol100 mM cacodylate pH 65) Microseeding was employed tooptimize crystal quality and reproducibility Crystals suitablefor X-ray data collection generally appeared within 48 h aftermicroseeding
Structure determination
X-ray data for FimHtr1 were collected from a single crystalto 17 Aring resolution at the Advanced Photon SourceArgonne National Laboratory USA The crystal was flash-cooled to 100 K in the precipitant solution complementedwith 25 glycerol The data were processed and scaledusing the HKL2000 package (Otwinowski and Minor 1997)(Table S1 in Supplementary material) Data for FimHtr2
were collected from a single crystal under liquid-nitrogencryo conditions at 100 K at beamline ID29 ESRF Greno-ble France to 21 Aring Bragg spacing FimHtr2 data were pro-cessed and reduced using MOSFLM (Powell 1999) andscaled using SCALA (Collaborative computational project1994) (Table S1 in Supplementary material) The structuresof FimHtr1 and FimHtr2 were solved by molecular replace-ment using Amore (Navaza 2001) and MOLREP (Vaginand Teplyakov 1997) respectively with the FimH lectindomain from the FimCFimH complex (Choudhury et al
A novel class of FimH high-affinity ligands 451
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
1999) (PDB id 1QUN) as the search model Positional bulksolvent and isotropic temperature factor refinement wasperformed using CNS (Bruumlnger et al 1998) and REFMAC5(Murshudov et al 1997) respectively Model adjustmentand inspection were performed using Turbo-Frodo and O(Jones et al 1991) graphics programs respectively Refine-ment statistics are given in Table S1 Molecular graphicsillustrations were generated with Bobscript (Esnouf 1997)and Pymol 090 (DeLano Scientific LLC 1998ndash2003 httppymolsourceforgenet)
Mass spectrometry
The purified FimHtr1 protein (300 mg) that had been used forcrystallization was freeze-dried taken up in 100 ml of pyridineand 100 ml of acetic anhydride and incubated overnight atroom temperature in the dark for peracetylation The samplewas dried under nitrogen and taken up in chloroform (400 ml)The chloroform phase was extracted six times with waterdried and taken up in 200 ml of 50 aqueous methanol A1 ml aliquot was applied to a nano-scale RP column(75 mm yen 100 mm PepMap DionexLC Packings) using anUltimate nano-LC system (LC Packings) which was equili-brated with eluent A (H2Oacetonitrile 955 vv containing01 formic acid) at a flow rate of 100 nl min-1 After injectingthe sample the column was run isocratically for 5 min fol-lowed by a linear gradient to 100 eluent B (H2Oacetonitrile2080 vv containing 01 formic acid) in 20 min and a finalwash with 100 B for 5 min The system was directly coupledto an Esquire 3000 ESI-IT-MS (Bruker Daltonik) equippedwith an online nanospray source operating in the positive-ionmode For electrospray (900ndash1200 V) capillaries (360 mmOD 20 mm ID with 10 mm opening) from New Objective wereused The solvent was evaporated at 150infinC with a nitrogenstream of 6 l min-1
To verify the suspected source of the bound butyl manno-side mass spectrometry was performed on samples ofFimHtr2 grown in either LB or minimal M9 medium Proteinsamples were dialysed against water overnight before massspectrometry in order to reduce background from the Trisbuffer Samples were diluted to a concentration in the submi-cromolar range Electrospray mass spectra were recorded indirect inlet mode (50 aqueous methanol solutions) on aBruker Daltonics Esquire-LC (Billerica)
Carbohydrate constituent analysis
Samples were hydrolysed in 100 ml of 4 M aqueous trifluoro-acetic acid (Sigma) at 100infinC for 4 h and dried under astream of nitrogen Monosaccharides were converted intotheir anthranilic acid derivatives by reductive aminationresolved by RP-HPLC and detected by fluorescence (Anu-mula 1994)
Carbohydrates for binding studies
[3H]-mannose was obtained from Amersham Methyl a-D-mannoside glucose galactose fructose sucrose turanosep-nitrophenyl a-D-mannopyranoside (pNPaMan) and 4-methylumbelliferyl a-D-mannopyranoside (MeUmbaMan)
were obtained from Sigma Monomannosylated bovineserum albumin (BSA-Man) was kindly provided by MedIm-mune We synthesized methyl 2-deoxy-a-D-mannopyrano-side (Oscarson and Tedebark 1995) Alkyl mannosides weresynthesized through silver triflate-promoted couplings of thecorresponding alcohol with 2346-tetra-O-benzoyl-a-D-mannopyranosyl bromide followed by Zempleacuten deacylationof the obtained protected alkyl mannosides according to theprocedure reported for the octyl and tetradecyl mannosides(Oscarson and Tiden 1993)
Binding studies
Competition experiments using SPR To obtain butyl man-noside-free FimHtr1 the FimH truncate was overproducedwith a carboxy-terminal 6-His tag after transformation into Ecoli C43 (DE3) (Miroux and Walker 1996) and the bacteriagrown in minA minimal medium supplemented with the 20amino acids (40 mg l-1) thiamin (2 mg l-1) and biotin (2 mgl-1) Purification of the His-tagged protein was performedusing Ni-NTA affinity chromatography (Qiagen) in 50 mM Tris-HCl pH 8 and 300 mM NaCl Elution was in the same solutioncomplemented with 500 mM imidazole Subsequently pooledand concentrated FimH fractions were gel filtrated over aSuperdex G75 column (Pharmacia) in 20 mM Tris-HCl pH 8
We used Fab fragments of a monoclonal antibody againstthe mannose-binding site of FimH to determine the solutionaffinity of FimHndashcarbohydrate interactions The monoclonalantibody IC10 was produced by a mouse hybridoma cell lineat Medimmune and its Fab fragments were prepared andpurified The Fabs were covalently immobilized via aminecoupling at 1000 Resonance Units (1000 pg ligand mm-2) inflowcell Fc2 on a CM5 biasensor chip (BIAapplications Hand-book Biacore AB Uppsala Sweden) Flowcell Fc1 was usedas the reference surface and was treated in the same wayas Fc2 except that it was blocked immediately after activationThe immobilization buffer was 100 mM sodium acetatepH 50 with 100 mM NaCl All SPR measurements were per-formed on a Biacore3000trade
The kinetic constants of binding of FimH to the immobilizedantibody were measured with different concentrations ofFimH (2000 nM 1000 nM 500 nM 250 nM 125 nM625 nM 312 nM 156 nM 78 nM) in running buffer [phos-phate-buffered saline (PBS) with 0005 surfactant P20 and3 mM EDTA] on both flowcells Fc1 and Fc2 simultaneouslyat a flow rate of 30 ml min-1 and at 298 K Complete dissoci-ation of FimH was performed with running buffer beforestarting a new binding cycle For all measurements the asso-ciation time was 3 min and the dissociation time was 30 minAll binding cycles were performed in duplicate including azero concentration cycle of FimH (injection of running buffer)All analyses were performed with the BIAeval software ALangmuir binding isotherm with a 11 stoichiometry was fittedto the data from both the association phase and the dissoci-ation phase after subtraction of the reference and zero con-centration data The kinetic constants ka and kd and themaximal binding Rmax were obtained and the dissociationconstant Kd was calculated
The equilibrium dissociation constants Kd of FimHndashsaccha-ride interactions were determined in a competition experi-ment using the same chip as for the previous experiment
452 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
As a control the Fabs of monoclonal antibody IA7 as wellas BSA-Man were also immobilized Samples containing afixed concentration of FimH were used (close to the Kd of theFimHndashantibody interaction) in combination with varying con-centrations of saccharide First 10-fold dilutions of the sac-charide solution were used to determine the concentrationrange for binding of the saccharide to FimH A Langmuirbinding isotherm with a 11 stoichiometry was fitted to thedata using the kinetic constants and Rmax from the foregoingexperiment to obtain the concentrations of FimH that werefree ([FimH]free) to bind the antibody immobilized on the chipSecond the concentration range of the saccharide wasextended and adapted to ensure accurate fitting and theequilibrium binding constant of the FimHndashsaccharide interac-tion was derived from the curve of [FimH]free against concen-tration of saccharide Every measurement was repeated atleast twice including testing different protein batches andwhenever possible also different saccharide stock solutions(typically 200 mM)
Equilibrium binding studies using radioactively labelledmannose Mannose-free FimHtr2 was produced inM9 minimal medium and purified as described above forFimHtr2 Alkyl mannosides were dissolved in double distilledwater pNPaMan and MeUmbaMan (6 mg each) were dis-solved in 20 ml of DMSO and diluted to 20 mM using doubledistilled water Binding experiments were performed at 310 Kusing six different concentrations of [3H]-a-D-mannose (finalconcentrations 435 mM 290 mM 193 mM 129 mM 86 mMand 57 mM) Protein (180 ml) at a concentration of about500 nM was mixed with 20 ml of the radioactive ligand [3H]-a-D-mannose and incubated at 37degC for 20 min To separatefree ligand from bound the mixture was rapidly filtratedthrough a Protran BA 85 cellulose nitrate filter (Schleicherand Schuell) and washed once with 1 ml of ice-cold 1yen PBSFilter-bound radioactivity was measured by scintillation spec-trometry within 24 h Displacement experiments were per-formed using six different concentrations of inhibitor (finalconcentrations in the range 00ndash435 mM) in the presence of435 mM [3H]-a-D-mannose Twenty microlitres of radioactiveligand 20 ml of inhibitor and 160 ml of protein (500 nM) weremixed and the experiments were performed as describedabove All experiments were performed in duplicates and thedata presented are mean values of at least two independentmeasurements For the determination of Kd for a-D-mannosean hyperbolic curve (y = P1x(P2 + x) where P1 is an arbitraryscale factor and P2 = Kd) was fitted to the data For the dis-placement experiments the curve y = P1(P2 + x) where P1
is an arbitrary scale factor and P2 is the concentration of theinhibitor displacing 50 of the labelled ligand [I]05 was usedinstead To calculate the inhibitor dissociation constant (KI)the Cheng and Prusoff equation [KI = [I]05[L]KL + 1) KL is theconstant of dissociation for the ligand] (Cheng and Prusoff1973) was used when both the concentration of the radio-active ligand (L) and the displacing agent (I) are in excessover the protein (LT gtgt PT IT gtgt PT T indicates total concen-tration) For the two strong inhibitors pNPaMan and MeUm-baMan where IT was not in excess over PT the equationof KI = IT[(1 - Y)Y(LTKL) - 1] - PTKLYLT where Y is thefraction of the ligand bound in presence of the inhibitor wasused instead (Horovitz and Levitzki 1987)
Computational docking
Computational docking studies were performed usingAutoDock3 (Morris et al 1998) In AutoDock3 the proteinis represented by a grid of affinity potentials Hence theprotein conformation is not allowed to change during thedocking procedure Flexibility of ligands is taken intoaccount by allowing rotation around flexible torsion anglesThe docked energies are calculated as the sum of the inter-molecular interaction energy and the internal energy of theligand The search for the best interaction energy was car-ried out by means of the Lamarckian genetic algorithmLigand structures were obtained from the Protein DataBank and prepared for docking by adding hydrogens andcharges using AutoDockTools (Sanner et al 1996) Eachsimulation consisted of 100 independent runs with a popu-lation size of 200 500 generations and a maximum of25 000 000 energy evaluations Solutions were rankedbased on their docking energies and similar solutions wereclustered (cluster cut-off rmsd lt 1 Aring) For both ligandsthe top solutions were evaluated using the graphics pro-gram O (Jones et al 1991)
Acknowledgements
We thank Andrej Zavialov and Maringhns Ehrenberg for help withequilibrium binding experiments and Thomas Norberg forassistance with mass spectrometry Anthony DeFusco fromMedimmune provided us with mannosylated bovine serumalbumin We are grateful to Fred Sauer for assistance withdata collection Thanks to Maor Bar-Peled of the CCC at theUniversity of Georgia Athens USA for his suggestion toexpress FimH in minimal medium We acknowledge the useof beamline 19BM of the Structural Biology Center at theAdvanced Photon Source Argonne National LaboratoryChicago IL USA and of beamline ID29 ESRF GrenobleFrance JB is a postdoctoral fellow of the Fonds voor Weten-schappelijk Onderzoek-Vlaanderen which also supportedthe BiaCore3000 instrument (FWONL35) This work was sup-ported by grants from the Swedish Research Council andfrom the Strategic Research Foundation (Structural BiologyNetwork) to SDK
Supplementary material
The following material is available from httpwwwblackwellpublishingcomproductsjournalssuppmatmmimmi4415mmi4415smhtmTable S1 Crystallographic data and refinement statistics
References
Anumula KR (1994) Quantitative determination ofmonosaccharides in glycoproteins by high-performanceliquid chromatography with highly sensitive fluorescencedetection Anal Biochem 220 275ndash283
Bahrani-Mougeot FK Buckles EL Lockatell CV HebelJR Johnson DE Tang CM and Donnenberg MS(2002) Type 1 fimbriae and extracellular polysaccharidesare preeminent uropathogenic Escherichia coli virulence
A novel class of FimH high-affinity ligands 453
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
determinants in the murine urinary tract Mol Microbiol 451079ndash1093
Berglund J and Knight SD (2003) Structural basis forbacterial adhesion in the urinary tract Glycobiol Med 53533ndash52
Blattner FR Plunkett G Bloch CA Perna NT BurlandV Riley M et al (1997) The complete genome sequenceof Escherichia coli K-12 Science 277 1453ndashbullbull
Bloch CA Stocker BA and Orndorff PE (1992) A keyrole for type 1 pili in enterobacterial communicability MolMicrobiol 6 697ndash701
Bork P Holm L and Sander C (1994) The immunoglob-ulin fold ndash structural classification sequence patterns andcommon core J Mol Biol 242 309ndash320
Brinton CC (1959) Non-flagellar appendages of bacteriaNature 183 782ndash786
Bruumlnger AT Adams PD Clore GM DeLano WLGros P Grosse-Kunstleve RW et al (1998) Crystallog-raphy and NMR system a new software suite for macro-molecular structure determination Acta Crystallogr D BiolCrystallogr 54 905ndash921
Buchanan K Falkow S Hull RA and Hull SI (1985)Frequency among Enterobacteriaceae of the DNA-sequences encoding type-1 pili J Bacteriol 162 799ndash803
Buts L Bouckaert J De Genst E Loris R Oscarson SLahmann M et al (2003) The fimbrial adhesin F17-G ofenterotoxigenic Escherichia coli has an immunoglobulin-like lectin domain that binds N-acetylglucosamine MolMicrobiol 49 705ndash715
Cheng Y and Prusoff WH (1973) Relationship betweeninhibition constant (K1) and concentration of inhibitor whichcauses 50 per cent inhibition (I50) of an enzymatic reac-tion Biochem Pharmacol 22 3099ndash3108
Choudhury D Thompson A Stojanoff V LangermannS Pinkner J Hultgren SJ and Knight SD (1999) X-ray structure of the FimCndashFimH chaperonendashadhesin com-plex from uropathogenic Escherichia coli Science 2851061ndash1066
Collaborative computational project n4 (1994) The CCP4suite programs for crystallography Acta Crystallogr D BiolCrystallogr 50 760ndash763
Dodson KW Pinkner JS Rose T Magnusson G Hult-gren SJ and Waksman G (2001) Structural basis of theinteraction of the pyelonephritic Escherichia coli adhesin toits human kidney receptor Cell 105 733ndash743
Eley JG and Triumalashetty P (2001) In vitro assessmentof alkylglycosides as permeability enhancers AAPSPharmSciTech 2 (3) Article 19
Esnouf RM (1997) An extensively modified version of Mol-script that includes greatly enhanced coloring capabilitiesJ Mol Graph Model 15 132ndash134
Firon N Ofek I and Sharon N (1982) Interaction ofmannose-containing oligosaccharides with the fimbriallectin of Escherichia coli Biochem Biophys Res Commun105 1426ndash1432
Firon N Ofek I and Sharon N (1983) Carbohydrate spec-ificity of the surface lectins of Escherichia coli Klebsiellapneumoniae and Salmonella typhimurium Carbohydr Res120 235ndash249
Firon N Ofek I and Sharon N (1984) Carbohydrate-
binding sites of the mannose-specific fimbrial lectins ofEnterobacteria Infect Immun 43 1088ndash1090
Firon N Ashkenazis S Mirelman D Ofek I and SharonN (1987) Aromatic a-glycosides of mannose are powerfulinhibitors of the adherence of type 1 fimbriated Escherichiacoli to yeast and intestinal cells Infect Immun 55 472ndash476
Fukiya S Mizoguchi H Tobe T and Mori H (2004)Extensive genomic diversity in pathogenic Escherichia coliand Shigella strains revealed by comparative genomichybridization microarray J Bacteriol 186 3911ndash3921
Hooton TM and Stamm WE (1997) Diagnosis and treat-ment of uncomplicated urinary tract infection Infect DisClin North Am 11 551ndash581
Horovitz A and Levitzki A (1987) An accurate method fordetermination of receptor ligand and enzyme-inhibitor dis-sociation-constants from displacement curves Proc NatlAcad Sci USA 84 6654ndash6658
Hull RA Gill RE Hsu P Minshaw BH and Falkow S(1981) Construction and expression of recombinant plas-mids encoding type 1 and D-mannose-resistant pili from aurinary tract infection Escherichia coli isolate Infect Immun33 933ndash938
Hultgren SJ Porter TN Schaeffer AJ and Duncan JL(1985) Role of type 1 pili and effects of phase variation onlower urinary tract infections produced by Escherichia coliInfect Immun 50 370ndash377
Hung C-S Bouckaert J Hung DL Pinkner J WinbergC Defusco A et al (2002) Structural basis of tropism ofEscherichia coli to the bladder during urinary tract infec-tion Mol Microbiol 44 903ndash915
Jones TA Zou JY Cowan SW and Kjeldgaard M(1991) Improved methods for building protein models inelectron density maps and the location of errors in thesemodels Acta Crystallogr A 47 110ndash119
Justice SS Hung C-S Theriot JA Fletcher DAAnderson GG Footer MJ and Hulgren SJ (2004)Differentiation and developmental pathways of uropatho-genic Escherichia coli in urinary tract pathogenesis ProcNatl Acad Sci USA 101 1333ndash1338
Kallenius G Mollby R Svenson SB Windberg J Lun-dblud A Svenson S and Cedergen B (1980) The Pk
antigen as receptor for the haemagglutinin of pyelonephri-togenic Escherichia coli FEMS Microbiol Lett 8 297ndash302
Knight SD Berglund J and Choudhury D (2000) Bacte-rial adhesins structural studies reveal chaperone functionand pilus biogenesis Curr Opin Chem Biol 4 653ndash660
Langermann S and Ballou WR (2003) Development of arecombinant FimCH vaccine for urinary tract infectionsAdv Exp Med Biol 539 635ndash653
Langermann S Palaszynski S Barnhart M Auguste GPinkner JS Burlein J et al (1997) Prevention ofmucosal Escherichia coli infection by FimH-adhesin-basedsystemic vaccination Science 276 607ndash611
Langermann S Mollby R Burlein JE Palaszynski SRAuguste CG Defusco A et al (2000) Vaccination withFimH adhesin protects cynomolgus monkeys from coloni-zation and infection by uropathogenic Escherichia coli JInfect Dis 181 774ndash778
Larsson A Ohlsson J Dodson KW Hultgren SJ Nils-son U and Kihlberg J (2003) Quantitative studies of thebinding of the class II PapG adhesin from uropathogenic
454 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
Escherichia coli to oligosaccharides Bioorg Med Chem 112255ndash2261
Leffler H and Svanborg-Eden C (1980) Chemical identifi-cation of a glycosphingolipid receptor for Escherichia coliattaching to human urinary tract epithelial cells and agglu-tinating human erythrocytes FEMS Microbiol Lett 8 127ndash134
Lindhorst TK Kieburg C and Krallmann-Wenzel U(1998) Inhibition of the type 1 fimbriae-mediated adhesionof Escherichia coli to erythrocytes by multiantennary alpha-mannosyl clusters the effect of multivalency Glycoconj J15 605ndash613
Loris R Tielker D Jaeger KE and Wyns L (2003a)Structural basis of carbohydrate recognition by the lectinLecB from Pseudomonas aeruginosa J Mol Biol 331 861ndash870
Loris R Imberty A Beeckmans S Van Driessche ERead JS Bouckaert J et al (2003b) Crystal structureof Pterocarpus angolensis lectin in complex with glucosesucrose and turanose J Biol Chem 278 16297ndash16303
Lund B Lindberg F Marklund BI and Normark S(1987) The PapG protein is the a-D-galactopyranosyl-(1-4)-b-D-galactopyranose-binding adhesin of uropathogenicEscherichia coli Proc Natl Acad Sci USA 84 5898ndash5902
Min G Stolz M Zhou G Liang F Sebbel P StofflerD et al (2002) Localization of uroplakin Ia the urothelialreceptor for bacterial adhesin FimH on the six innerdomains of the 16 nm urothelial plaque particle J Mol Biol317 697ndash706
Miroux B and Walker JE (1996) Over-production of pro-teins in Escherichia coli mutant hosts that allow synthesisof some membrane proteins and globular proteins at highlevels J Mol Biol 260 289ndash298
Morris GM Goodsell DS Halliday RS Huey R HartWE Belew RK and Olson AJ (1998) Automateddocking using a Lamarckian genetic algorithm and anempirical binding free energy function J Comput Chem 191639ndash1662
Mulvey MA (2002) Adhesion and entry of uropathogenicEscherichia coli Cell Microbiol 4 257ndash271
Mulvey MA Lopez-Boado YS Wilson CL Roth RParks WC Heuser J and Hultgren SJ (1998) Induc-tion and evasion of host defenses by type 1-piliated uro-pathogenic Escherichia coli Science 282 1494ndash1497
Murshudov GN Vagin AA and Dodson EJ (1997)Refinement of macromolecular structures by the maxi-mum-likelihood method Acta Crystallogr D Biol Crystallogr53 240ndash255
Nagahori N Lee RT Nishimura S Pageacute D Roy Rand Lee YC (2002) Inhibition of adhesion of type 1 fim-briated Escherichia coli to highly mannosylated ligandsChembiochem 3 836ndash844
Navaza J (2001) Implementation of molecular replacementin AMoRe Acta Crystallogr D Biol Crystallogr 57 1367ndash1372
Neeser JR Koellreutter B and Wuersch P (1986)Oligomannoside-type glycopeptides inhibiting adhesion ofEscherichia coli strains mediated by type 1 pili preparationof potent inhibitors from plant glycoproteins Infect Immun52 428ndash436
Old DC (1972) Inhibition of the interaction between fimbrial
hemagglutinatinins and erythrocytes by D-mannose andother carbohydrates J Gen Microbiol 71 149ndash157
Orndorff PE Devapali A Palestrant S Wyse A EverettML Bollinger RR and Parker W (2004) Immunoglob-ulin-mediated agglutination of and biofilm formation byEscherichia coli K-12 require the type 1 pilus fiber InfectImmun 72 1929ndash1938
Oscarson S and Tedebark U (1995) Syntheses of deoxyanalogues of methyl 36-di-O-a-D-mannopyranosyl-a-D-mannopyranoside for studies of the binding site of con-canavalin A Carbohydr Res 278 271ndash287
Oscarson S and Tiden AK (1993) Syntheses of the octyland tetradecyl glycosides of 36-di-O-alpha-deuterium-mannopyranosyl-alpha-deuterium-mannopyranose and of34-di-O-alpha-deuterium-mannopyranosyl-alpha-deute-rium-mannopyranose ndash a new way for 24-di-O-protectionof mannopyranosides Carbohydr Res 247 323ndash328
Otwinowski Z and Minor W (1997) Processing of X-raydiffraction data collected in oscillation mode MethodsEnzymol 276 307ndash326
Powell HR (1999) The Rossmann Fourier autoindexingalgorithm in MOSFLM Acta Crystallogr D Biol Crystallogr55 1690ndash1695
Pratt LA and Kolter R (1998) Genetic analysis of Escher-ichia coli biofilm formation roles of flagella motility chemo-taxis and type I pili Mol Microbiol 30 285ndash293
Remaut H and Waksman G (2004) Structural biology ofbacterial pathogenesis Curr Opin Struct Biol 14 161ndash170
Ronald AR Nicolle LE Stamm E Krieger J WarrenJ Schaeffer A et al (2001) Urinary tract infection inadults research priorities and strategies Int J AntimicrobAgents 17 343ndash348
Sanner MF Olson AJ and Spehner JC (1996)Reduced surface an efficient way to compute molecularsurfaces Biopolymers 38 305ndash320
Sauer FG Futterer K Pinkner JS Dodson KW Hult-gren SJ and Waksman G (1999) Structural basis ofchaperone function and pilus biogenesis Science 2851058ndash1061
Sauer FG Knight SD Waksman GW and HultgrenSJ (2000) PapD-like chaperones and pilus biogenesisSemin Cell Dev Biol 11 27ndash34
Sauer FG Pinkner JS Waksman G and Hultgren SJ(2002) Chaperone priming of pilus subunits facilitates atopological transition that drives fiber formation Cell 111543ndash551
Schembri MA Hasman H and Klemm P (2000)Expression and purification of the mannose recognitiondomain of the FimH adhesin FEMS Microbiol Lett 188147ndash151
Schembri MA Kjaergaard K and Klemm P (2003) Glo-bal gene expression in Escherichia coli biofilms Mol Micro-biol 48 253ndash267
Schilling JD Mulvey MA Vincent CD Lorenz RGand Hultgren SJ (2001) Bacterial invasion augments epi-thelial cytokine responses to Escherichia coli through alipopolysaccharide-dependent mechanism J Immunol166 1148ndash1155
Silhavy TJ Szmelcman S Boos W and Schwartz M(1975) On the significance of the retention of ligand byprotein Proc Natl Acad Sci USA 72 2120ndash2124
A novel class of FimH high-affinity ligands 455
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
Svanborg C and Godaly G (1997) Bacterial virulence inurinary tract infection Infect Dis Clin North Am 11 513ndash529
Thanassi DG Saulino ET and Hultgren SJ (1998) Thechaperoneusher pathway a major terminal branch of thegeneral secretory pathway Curr Opin Struct Biol 1 223ndash231
Thankavel K Madison B Ikeda T Malaviya R ShahAH Arumugen PM and Abraham SN (1997) Local-ization of a domain in the FimH adhesin of Escherichia colitype 1 fimbriae capable of receptor recognition and use ofa domain-specific antibody to confer protection againstexperimental urinary tract infection J Clin Invest 1001123ndash1126
Vagin A and Teplyakov A (1997) MOLREP an automatedprogram for molecular replacement J Appl Crystallogr 301022ndash1025
Wu XR Sun TT and Medina JJ (1996) In vitro bindingof type 1-fimbriated Escherichia coli to uroplakins 1a and
1b relation to urinary tract infections Proc Natl Acad SciUSA 93 9630ndash9635
Xia Y Gally D Forsman-Semb K and Uhlin BE (2000)Regulatory cross-talk between adhesin operons in Escher-ichia coli inhibition of type 1 fimbriae expression by thePapB protein EMBO J 19 1450ndash1457
Zafriri D Ofek I Adar R Pocino M and Sharon N(1989) Inhibitory activity of cranberry juice on adherenceof type-1 and type-P fimbriated Escherichia coli to eukary-otic cells Antimicrob Agents Chemother 33 92ndash98
Zavialov AV Berglund J Pudney AF Fooks LJ Ibra-him TM MacIntyre S and Knight SD (2003) Structureand biogenesis of the capsular F1 antigen from Yersiniapestis preserved folding energy drives fiber formation Cell113 587ndash596
Zhou G Mo WJ Sebbel P Min G Neubert TA Glock-shuber R et al (2001) Uroplakin Ia is the urothelial recep-tor for uropathogenic Escherichia coli evidence from invitro FimH binding J Cell Sci 114 4095ndash4103

450 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
be particularly potent inhibitors of FimH-mediated bacte-rial adhesion (Firon et al 1987 Nagahori et al 2002)Based on adhesion inhibition studies using type 1 piliatedE coli and guinea pig ileal epithelial cells the relativeinhibitory potential of pNPaMan and MeUmbaMan havebeen estimated to be about 70 and 600 times strongerrespectively than that of methyl a-D-mannose (Fironet al 1987) Our measurements indicate an affinity ofFimH for pNPaMan and MeUmbaMan between 50 and90 times and between 110 and 200 times higher respec-tively than for methyl a-D-mannose in good agreementwith the adhesion inhibition results (Firon et al 1987)Dissociation constants predicted using AutoDock3 (Morriset al 1998) are similar to experimental values Thecomputational docking predicts that tight binding of thearomatically substituted mannosides pNPaMan andMeUmbaMan is accomplished similarly as for butyl a-D-mannoside with the aglycon interacting with the tyrosinegate
This is the first systematic study providing molecularbinding data for the FimH adhesin the prerequisite forcolonization by UPEC (Langermann and Ballou 2003)We also show crystallographic evidence that the lectinor receptor binding domain of FimH is a stable andfunctional entity Its structure is virtually identical to thesame domain in the FimCFimH complex indicating thata truncate corresponding to the lectin domain could pro-vide a good alternative for the development of a UTIvaccine (Langermann et al 1997 Langermann and Bal-lou 2003) However progress of this interesting UTI vac-cine candidate into clinical trials has not been reportedThe binding affinities of FimH at the molecular levelrelate convincingly well with the inhibitory concentrationsused in previous bacterial adhesion assays thus endors-ing the direct and conclusive correlation of FimH withbacterial adherence Alkyl mannosides have nanomolaraffinities for FimH which may overcome the significantobstacle of low affinity generally encountered in attemptsto develop carbohydrate-based drugs which interferewith microbial attachment to host cell surface glycocon-jugates New anti-adhesive drugs could be based onthese leads to allow specific interaction and targeting tothe urinary tract in order to prevent anticipated andrecurrent infections
Experimental procedures
Strains and constructs
FimH proteins from two different E coli strains the UPECstrain J96 (Hull et al 1981) and the E coli K-12 strainMG1655 (Blattner et al 1997) were used in this study TheFimH adhesins encoded by these two E coli strains areidentical FimHtr1 (residues 1ndash158 of UPEC J96 FimH) wasexpressed from plasmid pMMB91 transformed into E coli
C600 cells FimHtr2 (residues 1ndash158 of E coli MG1655FimH + carboxy-terminal 6-His tag) was expressed fromplasmid pPKL241 (Schembri et al 2000) transformed into Ecoli host strain HB101
Purification and crystallization
FimHtr1 Bacteria were grown in LB medium at 37infinC inducedat OD600 = 08 with 1 mM IPTG and harvested 2ndash3 h laterThe cells were resuspended in 20 mM Tris-HCl pH 80 5 mMEDTA and 30 sucrose and the periplasmic content wasextracted by osmotic shock upon 10-fold dilution with 20 mMTris-HCl pH 80 Purification of FimHtr1 was performed by ionexchange chromatography with MonoS at pH 45 followed byMonoQ (Amersham Biosciences) at pH 8 FimHtr1 was crys-tallized using vapour diffusion in hanging drops composed of3 ml of the precipitant solution (01 M Hepes pH 75 12PEG 6000 and 5 vv methyl-24-pentanediol) and 3 ml of12 mg ml-1 protein solution in 20 mM Tris-HCl pH 80 with150 mM NaCl
FimHtr2 Cells were grown in LB medium containing50 mg ml-1 ampicillin at 37infinC At OD600 = 06 the cells wereinduced with 5 mM IPTG Cells were harvested 2 h afterinduction and the periplasmic content was extracted Theprotein was purified on a Pharmacia HiTrap Chelating HP5 ml column (Pharmacia) loaded with Ni-chloride and elutedwith a sharp 0ndash500 mM imidazole gradient Fractions con-taining FimHtr2 were pooled dialysed against 50 mM sodiumacetate pH 525 and loaded onto a Mono S HR 8 ml columnThe protein was eluted with a 0ndash500 mM NaCl gradient anddialysed overnight against 20 mM MES pH 65 before crys-tallization Crystallization was performed at room tempera-ture by the vapour diffusion method with hanging dropsmade by mixing of 2 ml of 15 mg ml-1 protein solution with 2 mlof the precipitant solution (65 methyl-24-pentanediol100 mM cacodylate pH 65) Microseeding was employed tooptimize crystal quality and reproducibility Crystals suitablefor X-ray data collection generally appeared within 48 h aftermicroseeding
Structure determination
X-ray data for FimHtr1 were collected from a single crystalto 17 Aring resolution at the Advanced Photon SourceArgonne National Laboratory USA The crystal was flash-cooled to 100 K in the precipitant solution complementedwith 25 glycerol The data were processed and scaledusing the HKL2000 package (Otwinowski and Minor 1997)(Table S1 in Supplementary material) Data for FimHtr2
were collected from a single crystal under liquid-nitrogencryo conditions at 100 K at beamline ID29 ESRF Greno-ble France to 21 Aring Bragg spacing FimHtr2 data were pro-cessed and reduced using MOSFLM (Powell 1999) andscaled using SCALA (Collaborative computational project1994) (Table S1 in Supplementary material) The structuresof FimHtr1 and FimHtr2 were solved by molecular replace-ment using Amore (Navaza 2001) and MOLREP (Vaginand Teplyakov 1997) respectively with the FimH lectindomain from the FimCFimH complex (Choudhury et al
A novel class of FimH high-affinity ligands 451
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
1999) (PDB id 1QUN) as the search model Positional bulksolvent and isotropic temperature factor refinement wasperformed using CNS (Bruumlnger et al 1998) and REFMAC5(Murshudov et al 1997) respectively Model adjustmentand inspection were performed using Turbo-Frodo and O(Jones et al 1991) graphics programs respectively Refine-ment statistics are given in Table S1 Molecular graphicsillustrations were generated with Bobscript (Esnouf 1997)and Pymol 090 (DeLano Scientific LLC 1998ndash2003 httppymolsourceforgenet)
Mass spectrometry
The purified FimHtr1 protein (300 mg) that had been used forcrystallization was freeze-dried taken up in 100 ml of pyridineand 100 ml of acetic anhydride and incubated overnight atroom temperature in the dark for peracetylation The samplewas dried under nitrogen and taken up in chloroform (400 ml)The chloroform phase was extracted six times with waterdried and taken up in 200 ml of 50 aqueous methanol A1 ml aliquot was applied to a nano-scale RP column(75 mm yen 100 mm PepMap DionexLC Packings) using anUltimate nano-LC system (LC Packings) which was equili-brated with eluent A (H2Oacetonitrile 955 vv containing01 formic acid) at a flow rate of 100 nl min-1 After injectingthe sample the column was run isocratically for 5 min fol-lowed by a linear gradient to 100 eluent B (H2Oacetonitrile2080 vv containing 01 formic acid) in 20 min and a finalwash with 100 B for 5 min The system was directly coupledto an Esquire 3000 ESI-IT-MS (Bruker Daltonik) equippedwith an online nanospray source operating in the positive-ionmode For electrospray (900ndash1200 V) capillaries (360 mmOD 20 mm ID with 10 mm opening) from New Objective wereused The solvent was evaporated at 150infinC with a nitrogenstream of 6 l min-1
To verify the suspected source of the bound butyl manno-side mass spectrometry was performed on samples ofFimHtr2 grown in either LB or minimal M9 medium Proteinsamples were dialysed against water overnight before massspectrometry in order to reduce background from the Trisbuffer Samples were diluted to a concentration in the submi-cromolar range Electrospray mass spectra were recorded indirect inlet mode (50 aqueous methanol solutions) on aBruker Daltonics Esquire-LC (Billerica)
Carbohydrate constituent analysis
Samples were hydrolysed in 100 ml of 4 M aqueous trifluoro-acetic acid (Sigma) at 100infinC for 4 h and dried under astream of nitrogen Monosaccharides were converted intotheir anthranilic acid derivatives by reductive aminationresolved by RP-HPLC and detected by fluorescence (Anu-mula 1994)
Carbohydrates for binding studies
[3H]-mannose was obtained from Amersham Methyl a-D-mannoside glucose galactose fructose sucrose turanosep-nitrophenyl a-D-mannopyranoside (pNPaMan) and 4-methylumbelliferyl a-D-mannopyranoside (MeUmbaMan)
were obtained from Sigma Monomannosylated bovineserum albumin (BSA-Man) was kindly provided by MedIm-mune We synthesized methyl 2-deoxy-a-D-mannopyrano-side (Oscarson and Tedebark 1995) Alkyl mannosides weresynthesized through silver triflate-promoted couplings of thecorresponding alcohol with 2346-tetra-O-benzoyl-a-D-mannopyranosyl bromide followed by Zempleacuten deacylationof the obtained protected alkyl mannosides according to theprocedure reported for the octyl and tetradecyl mannosides(Oscarson and Tiden 1993)
Binding studies
Competition experiments using SPR To obtain butyl man-noside-free FimHtr1 the FimH truncate was overproducedwith a carboxy-terminal 6-His tag after transformation into Ecoli C43 (DE3) (Miroux and Walker 1996) and the bacteriagrown in minA minimal medium supplemented with the 20amino acids (40 mg l-1) thiamin (2 mg l-1) and biotin (2 mgl-1) Purification of the His-tagged protein was performedusing Ni-NTA affinity chromatography (Qiagen) in 50 mM Tris-HCl pH 8 and 300 mM NaCl Elution was in the same solutioncomplemented with 500 mM imidazole Subsequently pooledand concentrated FimH fractions were gel filtrated over aSuperdex G75 column (Pharmacia) in 20 mM Tris-HCl pH 8
We used Fab fragments of a monoclonal antibody againstthe mannose-binding site of FimH to determine the solutionaffinity of FimHndashcarbohydrate interactions The monoclonalantibody IC10 was produced by a mouse hybridoma cell lineat Medimmune and its Fab fragments were prepared andpurified The Fabs were covalently immobilized via aminecoupling at 1000 Resonance Units (1000 pg ligand mm-2) inflowcell Fc2 on a CM5 biasensor chip (BIAapplications Hand-book Biacore AB Uppsala Sweden) Flowcell Fc1 was usedas the reference surface and was treated in the same wayas Fc2 except that it was blocked immediately after activationThe immobilization buffer was 100 mM sodium acetatepH 50 with 100 mM NaCl All SPR measurements were per-formed on a Biacore3000trade
The kinetic constants of binding of FimH to the immobilizedantibody were measured with different concentrations ofFimH (2000 nM 1000 nM 500 nM 250 nM 125 nM625 nM 312 nM 156 nM 78 nM) in running buffer [phos-phate-buffered saline (PBS) with 0005 surfactant P20 and3 mM EDTA] on both flowcells Fc1 and Fc2 simultaneouslyat a flow rate of 30 ml min-1 and at 298 K Complete dissoci-ation of FimH was performed with running buffer beforestarting a new binding cycle For all measurements the asso-ciation time was 3 min and the dissociation time was 30 minAll binding cycles were performed in duplicate including azero concentration cycle of FimH (injection of running buffer)All analyses were performed with the BIAeval software ALangmuir binding isotherm with a 11 stoichiometry was fittedto the data from both the association phase and the dissoci-ation phase after subtraction of the reference and zero con-centration data The kinetic constants ka and kd and themaximal binding Rmax were obtained and the dissociationconstant Kd was calculated
The equilibrium dissociation constants Kd of FimHndashsaccha-ride interactions were determined in a competition experi-ment using the same chip as for the previous experiment
452 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
As a control the Fabs of monoclonal antibody IA7 as wellas BSA-Man were also immobilized Samples containing afixed concentration of FimH were used (close to the Kd of theFimHndashantibody interaction) in combination with varying con-centrations of saccharide First 10-fold dilutions of the sac-charide solution were used to determine the concentrationrange for binding of the saccharide to FimH A Langmuirbinding isotherm with a 11 stoichiometry was fitted to thedata using the kinetic constants and Rmax from the foregoingexperiment to obtain the concentrations of FimH that werefree ([FimH]free) to bind the antibody immobilized on the chipSecond the concentration range of the saccharide wasextended and adapted to ensure accurate fitting and theequilibrium binding constant of the FimHndashsaccharide interac-tion was derived from the curve of [FimH]free against concen-tration of saccharide Every measurement was repeated atleast twice including testing different protein batches andwhenever possible also different saccharide stock solutions(typically 200 mM)
Equilibrium binding studies using radioactively labelledmannose Mannose-free FimHtr2 was produced inM9 minimal medium and purified as described above forFimHtr2 Alkyl mannosides were dissolved in double distilledwater pNPaMan and MeUmbaMan (6 mg each) were dis-solved in 20 ml of DMSO and diluted to 20 mM using doubledistilled water Binding experiments were performed at 310 Kusing six different concentrations of [3H]-a-D-mannose (finalconcentrations 435 mM 290 mM 193 mM 129 mM 86 mMand 57 mM) Protein (180 ml) at a concentration of about500 nM was mixed with 20 ml of the radioactive ligand [3H]-a-D-mannose and incubated at 37degC for 20 min To separatefree ligand from bound the mixture was rapidly filtratedthrough a Protran BA 85 cellulose nitrate filter (Schleicherand Schuell) and washed once with 1 ml of ice-cold 1yen PBSFilter-bound radioactivity was measured by scintillation spec-trometry within 24 h Displacement experiments were per-formed using six different concentrations of inhibitor (finalconcentrations in the range 00ndash435 mM) in the presence of435 mM [3H]-a-D-mannose Twenty microlitres of radioactiveligand 20 ml of inhibitor and 160 ml of protein (500 nM) weremixed and the experiments were performed as describedabove All experiments were performed in duplicates and thedata presented are mean values of at least two independentmeasurements For the determination of Kd for a-D-mannosean hyperbolic curve (y = P1x(P2 + x) where P1 is an arbitraryscale factor and P2 = Kd) was fitted to the data For the dis-placement experiments the curve y = P1(P2 + x) where P1
is an arbitrary scale factor and P2 is the concentration of theinhibitor displacing 50 of the labelled ligand [I]05 was usedinstead To calculate the inhibitor dissociation constant (KI)the Cheng and Prusoff equation [KI = [I]05[L]KL + 1) KL is theconstant of dissociation for the ligand] (Cheng and Prusoff1973) was used when both the concentration of the radio-active ligand (L) and the displacing agent (I) are in excessover the protein (LT gtgt PT IT gtgt PT T indicates total concen-tration) For the two strong inhibitors pNPaMan and MeUm-baMan where IT was not in excess over PT the equationof KI = IT[(1 - Y)Y(LTKL) - 1] - PTKLYLT where Y is thefraction of the ligand bound in presence of the inhibitor wasused instead (Horovitz and Levitzki 1987)
Computational docking
Computational docking studies were performed usingAutoDock3 (Morris et al 1998) In AutoDock3 the proteinis represented by a grid of affinity potentials Hence theprotein conformation is not allowed to change during thedocking procedure Flexibility of ligands is taken intoaccount by allowing rotation around flexible torsion anglesThe docked energies are calculated as the sum of the inter-molecular interaction energy and the internal energy of theligand The search for the best interaction energy was car-ried out by means of the Lamarckian genetic algorithmLigand structures were obtained from the Protein DataBank and prepared for docking by adding hydrogens andcharges using AutoDockTools (Sanner et al 1996) Eachsimulation consisted of 100 independent runs with a popu-lation size of 200 500 generations and a maximum of25 000 000 energy evaluations Solutions were rankedbased on their docking energies and similar solutions wereclustered (cluster cut-off rmsd lt 1 Aring) For both ligandsthe top solutions were evaluated using the graphics pro-gram O (Jones et al 1991)
Acknowledgements
We thank Andrej Zavialov and Maringhns Ehrenberg for help withequilibrium binding experiments and Thomas Norberg forassistance with mass spectrometry Anthony DeFusco fromMedimmune provided us with mannosylated bovine serumalbumin We are grateful to Fred Sauer for assistance withdata collection Thanks to Maor Bar-Peled of the CCC at theUniversity of Georgia Athens USA for his suggestion toexpress FimH in minimal medium We acknowledge the useof beamline 19BM of the Structural Biology Center at theAdvanced Photon Source Argonne National LaboratoryChicago IL USA and of beamline ID29 ESRF GrenobleFrance JB is a postdoctoral fellow of the Fonds voor Weten-schappelijk Onderzoek-Vlaanderen which also supportedthe BiaCore3000 instrument (FWONL35) This work was sup-ported by grants from the Swedish Research Council andfrom the Strategic Research Foundation (Structural BiologyNetwork) to SDK
Supplementary material
The following material is available from httpwwwblackwellpublishingcomproductsjournalssuppmatmmimmi4415mmi4415smhtmTable S1 Crystallographic data and refinement statistics
References
Anumula KR (1994) Quantitative determination ofmonosaccharides in glycoproteins by high-performanceliquid chromatography with highly sensitive fluorescencedetection Anal Biochem 220 275ndash283
Bahrani-Mougeot FK Buckles EL Lockatell CV HebelJR Johnson DE Tang CM and Donnenberg MS(2002) Type 1 fimbriae and extracellular polysaccharidesare preeminent uropathogenic Escherichia coli virulence
A novel class of FimH high-affinity ligands 453
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
determinants in the murine urinary tract Mol Microbiol 451079ndash1093
Berglund J and Knight SD (2003) Structural basis forbacterial adhesion in the urinary tract Glycobiol Med 53533ndash52
Blattner FR Plunkett G Bloch CA Perna NT BurlandV Riley M et al (1997) The complete genome sequenceof Escherichia coli K-12 Science 277 1453ndashbullbull
Bloch CA Stocker BA and Orndorff PE (1992) A keyrole for type 1 pili in enterobacterial communicability MolMicrobiol 6 697ndash701
Bork P Holm L and Sander C (1994) The immunoglob-ulin fold ndash structural classification sequence patterns andcommon core J Mol Biol 242 309ndash320
Brinton CC (1959) Non-flagellar appendages of bacteriaNature 183 782ndash786
Bruumlnger AT Adams PD Clore GM DeLano WLGros P Grosse-Kunstleve RW et al (1998) Crystallog-raphy and NMR system a new software suite for macro-molecular structure determination Acta Crystallogr D BiolCrystallogr 54 905ndash921
Buchanan K Falkow S Hull RA and Hull SI (1985)Frequency among Enterobacteriaceae of the DNA-sequences encoding type-1 pili J Bacteriol 162 799ndash803
Buts L Bouckaert J De Genst E Loris R Oscarson SLahmann M et al (2003) The fimbrial adhesin F17-G ofenterotoxigenic Escherichia coli has an immunoglobulin-like lectin domain that binds N-acetylglucosamine MolMicrobiol 49 705ndash715
Cheng Y and Prusoff WH (1973) Relationship betweeninhibition constant (K1) and concentration of inhibitor whichcauses 50 per cent inhibition (I50) of an enzymatic reac-tion Biochem Pharmacol 22 3099ndash3108
Choudhury D Thompson A Stojanoff V LangermannS Pinkner J Hultgren SJ and Knight SD (1999) X-ray structure of the FimCndashFimH chaperonendashadhesin com-plex from uropathogenic Escherichia coli Science 2851061ndash1066
Collaborative computational project n4 (1994) The CCP4suite programs for crystallography Acta Crystallogr D BiolCrystallogr 50 760ndash763
Dodson KW Pinkner JS Rose T Magnusson G Hult-gren SJ and Waksman G (2001) Structural basis of theinteraction of the pyelonephritic Escherichia coli adhesin toits human kidney receptor Cell 105 733ndash743
Eley JG and Triumalashetty P (2001) In vitro assessmentof alkylglycosides as permeability enhancers AAPSPharmSciTech 2 (3) Article 19
Esnouf RM (1997) An extensively modified version of Mol-script that includes greatly enhanced coloring capabilitiesJ Mol Graph Model 15 132ndash134
Firon N Ofek I and Sharon N (1982) Interaction ofmannose-containing oligosaccharides with the fimbriallectin of Escherichia coli Biochem Biophys Res Commun105 1426ndash1432
Firon N Ofek I and Sharon N (1983) Carbohydrate spec-ificity of the surface lectins of Escherichia coli Klebsiellapneumoniae and Salmonella typhimurium Carbohydr Res120 235ndash249
Firon N Ofek I and Sharon N (1984) Carbohydrate-
binding sites of the mannose-specific fimbrial lectins ofEnterobacteria Infect Immun 43 1088ndash1090
Firon N Ashkenazis S Mirelman D Ofek I and SharonN (1987) Aromatic a-glycosides of mannose are powerfulinhibitors of the adherence of type 1 fimbriated Escherichiacoli to yeast and intestinal cells Infect Immun 55 472ndash476
Fukiya S Mizoguchi H Tobe T and Mori H (2004)Extensive genomic diversity in pathogenic Escherichia coliand Shigella strains revealed by comparative genomichybridization microarray J Bacteriol 186 3911ndash3921
Hooton TM and Stamm WE (1997) Diagnosis and treat-ment of uncomplicated urinary tract infection Infect DisClin North Am 11 551ndash581
Horovitz A and Levitzki A (1987) An accurate method fordetermination of receptor ligand and enzyme-inhibitor dis-sociation-constants from displacement curves Proc NatlAcad Sci USA 84 6654ndash6658
Hull RA Gill RE Hsu P Minshaw BH and Falkow S(1981) Construction and expression of recombinant plas-mids encoding type 1 and D-mannose-resistant pili from aurinary tract infection Escherichia coli isolate Infect Immun33 933ndash938
Hultgren SJ Porter TN Schaeffer AJ and Duncan JL(1985) Role of type 1 pili and effects of phase variation onlower urinary tract infections produced by Escherichia coliInfect Immun 50 370ndash377
Hung C-S Bouckaert J Hung DL Pinkner J WinbergC Defusco A et al (2002) Structural basis of tropism ofEscherichia coli to the bladder during urinary tract infec-tion Mol Microbiol 44 903ndash915
Jones TA Zou JY Cowan SW and Kjeldgaard M(1991) Improved methods for building protein models inelectron density maps and the location of errors in thesemodels Acta Crystallogr A 47 110ndash119
Justice SS Hung C-S Theriot JA Fletcher DAAnderson GG Footer MJ and Hulgren SJ (2004)Differentiation and developmental pathways of uropatho-genic Escherichia coli in urinary tract pathogenesis ProcNatl Acad Sci USA 101 1333ndash1338
Kallenius G Mollby R Svenson SB Windberg J Lun-dblud A Svenson S and Cedergen B (1980) The Pk
antigen as receptor for the haemagglutinin of pyelonephri-togenic Escherichia coli FEMS Microbiol Lett 8 297ndash302
Knight SD Berglund J and Choudhury D (2000) Bacte-rial adhesins structural studies reveal chaperone functionand pilus biogenesis Curr Opin Chem Biol 4 653ndash660
Langermann S and Ballou WR (2003) Development of arecombinant FimCH vaccine for urinary tract infectionsAdv Exp Med Biol 539 635ndash653
Langermann S Palaszynski S Barnhart M Auguste GPinkner JS Burlein J et al (1997) Prevention ofmucosal Escherichia coli infection by FimH-adhesin-basedsystemic vaccination Science 276 607ndash611
Langermann S Mollby R Burlein JE Palaszynski SRAuguste CG Defusco A et al (2000) Vaccination withFimH adhesin protects cynomolgus monkeys from coloni-zation and infection by uropathogenic Escherichia coli JInfect Dis 181 774ndash778
Larsson A Ohlsson J Dodson KW Hultgren SJ Nils-son U and Kihlberg J (2003) Quantitative studies of thebinding of the class II PapG adhesin from uropathogenic
454 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
Escherichia coli to oligosaccharides Bioorg Med Chem 112255ndash2261
Leffler H and Svanborg-Eden C (1980) Chemical identifi-cation of a glycosphingolipid receptor for Escherichia coliattaching to human urinary tract epithelial cells and agglu-tinating human erythrocytes FEMS Microbiol Lett 8 127ndash134
Lindhorst TK Kieburg C and Krallmann-Wenzel U(1998) Inhibition of the type 1 fimbriae-mediated adhesionof Escherichia coli to erythrocytes by multiantennary alpha-mannosyl clusters the effect of multivalency Glycoconj J15 605ndash613
Loris R Tielker D Jaeger KE and Wyns L (2003a)Structural basis of carbohydrate recognition by the lectinLecB from Pseudomonas aeruginosa J Mol Biol 331 861ndash870
Loris R Imberty A Beeckmans S Van Driessche ERead JS Bouckaert J et al (2003b) Crystal structureof Pterocarpus angolensis lectin in complex with glucosesucrose and turanose J Biol Chem 278 16297ndash16303
Lund B Lindberg F Marklund BI and Normark S(1987) The PapG protein is the a-D-galactopyranosyl-(1-4)-b-D-galactopyranose-binding adhesin of uropathogenicEscherichia coli Proc Natl Acad Sci USA 84 5898ndash5902
Min G Stolz M Zhou G Liang F Sebbel P StofflerD et al (2002) Localization of uroplakin Ia the urothelialreceptor for bacterial adhesin FimH on the six innerdomains of the 16 nm urothelial plaque particle J Mol Biol317 697ndash706
Miroux B and Walker JE (1996) Over-production of pro-teins in Escherichia coli mutant hosts that allow synthesisof some membrane proteins and globular proteins at highlevels J Mol Biol 260 289ndash298
Morris GM Goodsell DS Halliday RS Huey R HartWE Belew RK and Olson AJ (1998) Automateddocking using a Lamarckian genetic algorithm and anempirical binding free energy function J Comput Chem 191639ndash1662
Mulvey MA (2002) Adhesion and entry of uropathogenicEscherichia coli Cell Microbiol 4 257ndash271
Mulvey MA Lopez-Boado YS Wilson CL Roth RParks WC Heuser J and Hultgren SJ (1998) Induc-tion and evasion of host defenses by type 1-piliated uro-pathogenic Escherichia coli Science 282 1494ndash1497
Murshudov GN Vagin AA and Dodson EJ (1997)Refinement of macromolecular structures by the maxi-mum-likelihood method Acta Crystallogr D Biol Crystallogr53 240ndash255
Nagahori N Lee RT Nishimura S Pageacute D Roy Rand Lee YC (2002) Inhibition of adhesion of type 1 fim-briated Escherichia coli to highly mannosylated ligandsChembiochem 3 836ndash844
Navaza J (2001) Implementation of molecular replacementin AMoRe Acta Crystallogr D Biol Crystallogr 57 1367ndash1372
Neeser JR Koellreutter B and Wuersch P (1986)Oligomannoside-type glycopeptides inhibiting adhesion ofEscherichia coli strains mediated by type 1 pili preparationof potent inhibitors from plant glycoproteins Infect Immun52 428ndash436
Old DC (1972) Inhibition of the interaction between fimbrial
hemagglutinatinins and erythrocytes by D-mannose andother carbohydrates J Gen Microbiol 71 149ndash157
Orndorff PE Devapali A Palestrant S Wyse A EverettML Bollinger RR and Parker W (2004) Immunoglob-ulin-mediated agglutination of and biofilm formation byEscherichia coli K-12 require the type 1 pilus fiber InfectImmun 72 1929ndash1938
Oscarson S and Tedebark U (1995) Syntheses of deoxyanalogues of methyl 36-di-O-a-D-mannopyranosyl-a-D-mannopyranoside for studies of the binding site of con-canavalin A Carbohydr Res 278 271ndash287
Oscarson S and Tiden AK (1993) Syntheses of the octyland tetradecyl glycosides of 36-di-O-alpha-deuterium-mannopyranosyl-alpha-deuterium-mannopyranose and of34-di-O-alpha-deuterium-mannopyranosyl-alpha-deute-rium-mannopyranose ndash a new way for 24-di-O-protectionof mannopyranosides Carbohydr Res 247 323ndash328
Otwinowski Z and Minor W (1997) Processing of X-raydiffraction data collected in oscillation mode MethodsEnzymol 276 307ndash326
Powell HR (1999) The Rossmann Fourier autoindexingalgorithm in MOSFLM Acta Crystallogr D Biol Crystallogr55 1690ndash1695
Pratt LA and Kolter R (1998) Genetic analysis of Escher-ichia coli biofilm formation roles of flagella motility chemo-taxis and type I pili Mol Microbiol 30 285ndash293
Remaut H and Waksman G (2004) Structural biology ofbacterial pathogenesis Curr Opin Struct Biol 14 161ndash170
Ronald AR Nicolle LE Stamm E Krieger J WarrenJ Schaeffer A et al (2001) Urinary tract infection inadults research priorities and strategies Int J AntimicrobAgents 17 343ndash348
Sanner MF Olson AJ and Spehner JC (1996)Reduced surface an efficient way to compute molecularsurfaces Biopolymers 38 305ndash320
Sauer FG Futterer K Pinkner JS Dodson KW Hult-gren SJ and Waksman G (1999) Structural basis ofchaperone function and pilus biogenesis Science 2851058ndash1061
Sauer FG Knight SD Waksman GW and HultgrenSJ (2000) PapD-like chaperones and pilus biogenesisSemin Cell Dev Biol 11 27ndash34
Sauer FG Pinkner JS Waksman G and Hultgren SJ(2002) Chaperone priming of pilus subunits facilitates atopological transition that drives fiber formation Cell 111543ndash551
Schembri MA Hasman H and Klemm P (2000)Expression and purification of the mannose recognitiondomain of the FimH adhesin FEMS Microbiol Lett 188147ndash151
Schembri MA Kjaergaard K and Klemm P (2003) Glo-bal gene expression in Escherichia coli biofilms Mol Micro-biol 48 253ndash267
Schilling JD Mulvey MA Vincent CD Lorenz RGand Hultgren SJ (2001) Bacterial invasion augments epi-thelial cytokine responses to Escherichia coli through alipopolysaccharide-dependent mechanism J Immunol166 1148ndash1155
Silhavy TJ Szmelcman S Boos W and Schwartz M(1975) On the significance of the retention of ligand byprotein Proc Natl Acad Sci USA 72 2120ndash2124
A novel class of FimH high-affinity ligands 455
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
Svanborg C and Godaly G (1997) Bacterial virulence inurinary tract infection Infect Dis Clin North Am 11 513ndash529
Thanassi DG Saulino ET and Hultgren SJ (1998) Thechaperoneusher pathway a major terminal branch of thegeneral secretory pathway Curr Opin Struct Biol 1 223ndash231
Thankavel K Madison B Ikeda T Malaviya R ShahAH Arumugen PM and Abraham SN (1997) Local-ization of a domain in the FimH adhesin of Escherichia colitype 1 fimbriae capable of receptor recognition and use ofa domain-specific antibody to confer protection againstexperimental urinary tract infection J Clin Invest 1001123ndash1126
Vagin A and Teplyakov A (1997) MOLREP an automatedprogram for molecular replacement J Appl Crystallogr 301022ndash1025
Wu XR Sun TT and Medina JJ (1996) In vitro bindingof type 1-fimbriated Escherichia coli to uroplakins 1a and
1b relation to urinary tract infections Proc Natl Acad SciUSA 93 9630ndash9635
Xia Y Gally D Forsman-Semb K and Uhlin BE (2000)Regulatory cross-talk between adhesin operons in Escher-ichia coli inhibition of type 1 fimbriae expression by thePapB protein EMBO J 19 1450ndash1457
Zafriri D Ofek I Adar R Pocino M and Sharon N(1989) Inhibitory activity of cranberry juice on adherenceof type-1 and type-P fimbriated Escherichia coli to eukary-otic cells Antimicrob Agents Chemother 33 92ndash98
Zavialov AV Berglund J Pudney AF Fooks LJ Ibra-him TM MacIntyre S and Knight SD (2003) Structureand biogenesis of the capsular F1 antigen from Yersiniapestis preserved folding energy drives fiber formation Cell113 587ndash596
Zhou G Mo WJ Sebbel P Min G Neubert TA Glock-shuber R et al (2001) Uroplakin Ia is the urothelial recep-tor for uropathogenic Escherichia coli evidence from invitro FimH binding J Cell Sci 114 4095ndash4103

A novel class of FimH high-affinity ligands 451
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
1999) (PDB id 1QUN) as the search model Positional bulksolvent and isotropic temperature factor refinement wasperformed using CNS (Bruumlnger et al 1998) and REFMAC5(Murshudov et al 1997) respectively Model adjustmentand inspection were performed using Turbo-Frodo and O(Jones et al 1991) graphics programs respectively Refine-ment statistics are given in Table S1 Molecular graphicsillustrations were generated with Bobscript (Esnouf 1997)and Pymol 090 (DeLano Scientific LLC 1998ndash2003 httppymolsourceforgenet)
Mass spectrometry
The purified FimHtr1 protein (300 mg) that had been used forcrystallization was freeze-dried taken up in 100 ml of pyridineand 100 ml of acetic anhydride and incubated overnight atroom temperature in the dark for peracetylation The samplewas dried under nitrogen and taken up in chloroform (400 ml)The chloroform phase was extracted six times with waterdried and taken up in 200 ml of 50 aqueous methanol A1 ml aliquot was applied to a nano-scale RP column(75 mm yen 100 mm PepMap DionexLC Packings) using anUltimate nano-LC system (LC Packings) which was equili-brated with eluent A (H2Oacetonitrile 955 vv containing01 formic acid) at a flow rate of 100 nl min-1 After injectingthe sample the column was run isocratically for 5 min fol-lowed by a linear gradient to 100 eluent B (H2Oacetonitrile2080 vv containing 01 formic acid) in 20 min and a finalwash with 100 B for 5 min The system was directly coupledto an Esquire 3000 ESI-IT-MS (Bruker Daltonik) equippedwith an online nanospray source operating in the positive-ionmode For electrospray (900ndash1200 V) capillaries (360 mmOD 20 mm ID with 10 mm opening) from New Objective wereused The solvent was evaporated at 150infinC with a nitrogenstream of 6 l min-1
To verify the suspected source of the bound butyl manno-side mass spectrometry was performed on samples ofFimHtr2 grown in either LB or minimal M9 medium Proteinsamples were dialysed against water overnight before massspectrometry in order to reduce background from the Trisbuffer Samples were diluted to a concentration in the submi-cromolar range Electrospray mass spectra were recorded indirect inlet mode (50 aqueous methanol solutions) on aBruker Daltonics Esquire-LC (Billerica)
Carbohydrate constituent analysis
Samples were hydrolysed in 100 ml of 4 M aqueous trifluoro-acetic acid (Sigma) at 100infinC for 4 h and dried under astream of nitrogen Monosaccharides were converted intotheir anthranilic acid derivatives by reductive aminationresolved by RP-HPLC and detected by fluorescence (Anu-mula 1994)
Carbohydrates for binding studies
[3H]-mannose was obtained from Amersham Methyl a-D-mannoside glucose galactose fructose sucrose turanosep-nitrophenyl a-D-mannopyranoside (pNPaMan) and 4-methylumbelliferyl a-D-mannopyranoside (MeUmbaMan)
were obtained from Sigma Monomannosylated bovineserum albumin (BSA-Man) was kindly provided by MedIm-mune We synthesized methyl 2-deoxy-a-D-mannopyrano-side (Oscarson and Tedebark 1995) Alkyl mannosides weresynthesized through silver triflate-promoted couplings of thecorresponding alcohol with 2346-tetra-O-benzoyl-a-D-mannopyranosyl bromide followed by Zempleacuten deacylationof the obtained protected alkyl mannosides according to theprocedure reported for the octyl and tetradecyl mannosides(Oscarson and Tiden 1993)
Binding studies
Competition experiments using SPR To obtain butyl man-noside-free FimHtr1 the FimH truncate was overproducedwith a carboxy-terminal 6-His tag after transformation into Ecoli C43 (DE3) (Miroux and Walker 1996) and the bacteriagrown in minA minimal medium supplemented with the 20amino acids (40 mg l-1) thiamin (2 mg l-1) and biotin (2 mgl-1) Purification of the His-tagged protein was performedusing Ni-NTA affinity chromatography (Qiagen) in 50 mM Tris-HCl pH 8 and 300 mM NaCl Elution was in the same solutioncomplemented with 500 mM imidazole Subsequently pooledand concentrated FimH fractions were gel filtrated over aSuperdex G75 column (Pharmacia) in 20 mM Tris-HCl pH 8
We used Fab fragments of a monoclonal antibody againstthe mannose-binding site of FimH to determine the solutionaffinity of FimHndashcarbohydrate interactions The monoclonalantibody IC10 was produced by a mouse hybridoma cell lineat Medimmune and its Fab fragments were prepared andpurified The Fabs were covalently immobilized via aminecoupling at 1000 Resonance Units (1000 pg ligand mm-2) inflowcell Fc2 on a CM5 biasensor chip (BIAapplications Hand-book Biacore AB Uppsala Sweden) Flowcell Fc1 was usedas the reference surface and was treated in the same wayas Fc2 except that it was blocked immediately after activationThe immobilization buffer was 100 mM sodium acetatepH 50 with 100 mM NaCl All SPR measurements were per-formed on a Biacore3000trade
The kinetic constants of binding of FimH to the immobilizedantibody were measured with different concentrations ofFimH (2000 nM 1000 nM 500 nM 250 nM 125 nM625 nM 312 nM 156 nM 78 nM) in running buffer [phos-phate-buffered saline (PBS) with 0005 surfactant P20 and3 mM EDTA] on both flowcells Fc1 and Fc2 simultaneouslyat a flow rate of 30 ml min-1 and at 298 K Complete dissoci-ation of FimH was performed with running buffer beforestarting a new binding cycle For all measurements the asso-ciation time was 3 min and the dissociation time was 30 minAll binding cycles were performed in duplicate including azero concentration cycle of FimH (injection of running buffer)All analyses were performed with the BIAeval software ALangmuir binding isotherm with a 11 stoichiometry was fittedto the data from both the association phase and the dissoci-ation phase after subtraction of the reference and zero con-centration data The kinetic constants ka and kd and themaximal binding Rmax were obtained and the dissociationconstant Kd was calculated
The equilibrium dissociation constants Kd of FimHndashsaccha-ride interactions were determined in a competition experi-ment using the same chip as for the previous experiment
452 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
As a control the Fabs of monoclonal antibody IA7 as wellas BSA-Man were also immobilized Samples containing afixed concentration of FimH were used (close to the Kd of theFimHndashantibody interaction) in combination with varying con-centrations of saccharide First 10-fold dilutions of the sac-charide solution were used to determine the concentrationrange for binding of the saccharide to FimH A Langmuirbinding isotherm with a 11 stoichiometry was fitted to thedata using the kinetic constants and Rmax from the foregoingexperiment to obtain the concentrations of FimH that werefree ([FimH]free) to bind the antibody immobilized on the chipSecond the concentration range of the saccharide wasextended and adapted to ensure accurate fitting and theequilibrium binding constant of the FimHndashsaccharide interac-tion was derived from the curve of [FimH]free against concen-tration of saccharide Every measurement was repeated atleast twice including testing different protein batches andwhenever possible also different saccharide stock solutions(typically 200 mM)
Equilibrium binding studies using radioactively labelledmannose Mannose-free FimHtr2 was produced inM9 minimal medium and purified as described above forFimHtr2 Alkyl mannosides were dissolved in double distilledwater pNPaMan and MeUmbaMan (6 mg each) were dis-solved in 20 ml of DMSO and diluted to 20 mM using doubledistilled water Binding experiments were performed at 310 Kusing six different concentrations of [3H]-a-D-mannose (finalconcentrations 435 mM 290 mM 193 mM 129 mM 86 mMand 57 mM) Protein (180 ml) at a concentration of about500 nM was mixed with 20 ml of the radioactive ligand [3H]-a-D-mannose and incubated at 37degC for 20 min To separatefree ligand from bound the mixture was rapidly filtratedthrough a Protran BA 85 cellulose nitrate filter (Schleicherand Schuell) and washed once with 1 ml of ice-cold 1yen PBSFilter-bound radioactivity was measured by scintillation spec-trometry within 24 h Displacement experiments were per-formed using six different concentrations of inhibitor (finalconcentrations in the range 00ndash435 mM) in the presence of435 mM [3H]-a-D-mannose Twenty microlitres of radioactiveligand 20 ml of inhibitor and 160 ml of protein (500 nM) weremixed and the experiments were performed as describedabove All experiments were performed in duplicates and thedata presented are mean values of at least two independentmeasurements For the determination of Kd for a-D-mannosean hyperbolic curve (y = P1x(P2 + x) where P1 is an arbitraryscale factor and P2 = Kd) was fitted to the data For the dis-placement experiments the curve y = P1(P2 + x) where P1
is an arbitrary scale factor and P2 is the concentration of theinhibitor displacing 50 of the labelled ligand [I]05 was usedinstead To calculate the inhibitor dissociation constant (KI)the Cheng and Prusoff equation [KI = [I]05[L]KL + 1) KL is theconstant of dissociation for the ligand] (Cheng and Prusoff1973) was used when both the concentration of the radio-active ligand (L) and the displacing agent (I) are in excessover the protein (LT gtgt PT IT gtgt PT T indicates total concen-tration) For the two strong inhibitors pNPaMan and MeUm-baMan where IT was not in excess over PT the equationof KI = IT[(1 - Y)Y(LTKL) - 1] - PTKLYLT where Y is thefraction of the ligand bound in presence of the inhibitor wasused instead (Horovitz and Levitzki 1987)
Computational docking
Computational docking studies were performed usingAutoDock3 (Morris et al 1998) In AutoDock3 the proteinis represented by a grid of affinity potentials Hence theprotein conformation is not allowed to change during thedocking procedure Flexibility of ligands is taken intoaccount by allowing rotation around flexible torsion anglesThe docked energies are calculated as the sum of the inter-molecular interaction energy and the internal energy of theligand The search for the best interaction energy was car-ried out by means of the Lamarckian genetic algorithmLigand structures were obtained from the Protein DataBank and prepared for docking by adding hydrogens andcharges using AutoDockTools (Sanner et al 1996) Eachsimulation consisted of 100 independent runs with a popu-lation size of 200 500 generations and a maximum of25 000 000 energy evaluations Solutions were rankedbased on their docking energies and similar solutions wereclustered (cluster cut-off rmsd lt 1 Aring) For both ligandsthe top solutions were evaluated using the graphics pro-gram O (Jones et al 1991)
Acknowledgements
We thank Andrej Zavialov and Maringhns Ehrenberg for help withequilibrium binding experiments and Thomas Norberg forassistance with mass spectrometry Anthony DeFusco fromMedimmune provided us with mannosylated bovine serumalbumin We are grateful to Fred Sauer for assistance withdata collection Thanks to Maor Bar-Peled of the CCC at theUniversity of Georgia Athens USA for his suggestion toexpress FimH in minimal medium We acknowledge the useof beamline 19BM of the Structural Biology Center at theAdvanced Photon Source Argonne National LaboratoryChicago IL USA and of beamline ID29 ESRF GrenobleFrance JB is a postdoctoral fellow of the Fonds voor Weten-schappelijk Onderzoek-Vlaanderen which also supportedthe BiaCore3000 instrument (FWONL35) This work was sup-ported by grants from the Swedish Research Council andfrom the Strategic Research Foundation (Structural BiologyNetwork) to SDK
Supplementary material
The following material is available from httpwwwblackwellpublishingcomproductsjournalssuppmatmmimmi4415mmi4415smhtmTable S1 Crystallographic data and refinement statistics
References
Anumula KR (1994) Quantitative determination ofmonosaccharides in glycoproteins by high-performanceliquid chromatography with highly sensitive fluorescencedetection Anal Biochem 220 275ndash283
Bahrani-Mougeot FK Buckles EL Lockatell CV HebelJR Johnson DE Tang CM and Donnenberg MS(2002) Type 1 fimbriae and extracellular polysaccharidesare preeminent uropathogenic Escherichia coli virulence
A novel class of FimH high-affinity ligands 453
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
determinants in the murine urinary tract Mol Microbiol 451079ndash1093
Berglund J and Knight SD (2003) Structural basis forbacterial adhesion in the urinary tract Glycobiol Med 53533ndash52
Blattner FR Plunkett G Bloch CA Perna NT BurlandV Riley M et al (1997) The complete genome sequenceof Escherichia coli K-12 Science 277 1453ndashbullbull
Bloch CA Stocker BA and Orndorff PE (1992) A keyrole for type 1 pili in enterobacterial communicability MolMicrobiol 6 697ndash701
Bork P Holm L and Sander C (1994) The immunoglob-ulin fold ndash structural classification sequence patterns andcommon core J Mol Biol 242 309ndash320
Brinton CC (1959) Non-flagellar appendages of bacteriaNature 183 782ndash786
Bruumlnger AT Adams PD Clore GM DeLano WLGros P Grosse-Kunstleve RW et al (1998) Crystallog-raphy and NMR system a new software suite for macro-molecular structure determination Acta Crystallogr D BiolCrystallogr 54 905ndash921
Buchanan K Falkow S Hull RA and Hull SI (1985)Frequency among Enterobacteriaceae of the DNA-sequences encoding type-1 pili J Bacteriol 162 799ndash803
Buts L Bouckaert J De Genst E Loris R Oscarson SLahmann M et al (2003) The fimbrial adhesin F17-G ofenterotoxigenic Escherichia coli has an immunoglobulin-like lectin domain that binds N-acetylglucosamine MolMicrobiol 49 705ndash715
Cheng Y and Prusoff WH (1973) Relationship betweeninhibition constant (K1) and concentration of inhibitor whichcauses 50 per cent inhibition (I50) of an enzymatic reac-tion Biochem Pharmacol 22 3099ndash3108
Choudhury D Thompson A Stojanoff V LangermannS Pinkner J Hultgren SJ and Knight SD (1999) X-ray structure of the FimCndashFimH chaperonendashadhesin com-plex from uropathogenic Escherichia coli Science 2851061ndash1066
Collaborative computational project n4 (1994) The CCP4suite programs for crystallography Acta Crystallogr D BiolCrystallogr 50 760ndash763
Dodson KW Pinkner JS Rose T Magnusson G Hult-gren SJ and Waksman G (2001) Structural basis of theinteraction of the pyelonephritic Escherichia coli adhesin toits human kidney receptor Cell 105 733ndash743
Eley JG and Triumalashetty P (2001) In vitro assessmentof alkylglycosides as permeability enhancers AAPSPharmSciTech 2 (3) Article 19
Esnouf RM (1997) An extensively modified version of Mol-script that includes greatly enhanced coloring capabilitiesJ Mol Graph Model 15 132ndash134
Firon N Ofek I and Sharon N (1982) Interaction ofmannose-containing oligosaccharides with the fimbriallectin of Escherichia coli Biochem Biophys Res Commun105 1426ndash1432
Firon N Ofek I and Sharon N (1983) Carbohydrate spec-ificity of the surface lectins of Escherichia coli Klebsiellapneumoniae and Salmonella typhimurium Carbohydr Res120 235ndash249
Firon N Ofek I and Sharon N (1984) Carbohydrate-
binding sites of the mannose-specific fimbrial lectins ofEnterobacteria Infect Immun 43 1088ndash1090
Firon N Ashkenazis S Mirelman D Ofek I and SharonN (1987) Aromatic a-glycosides of mannose are powerfulinhibitors of the adherence of type 1 fimbriated Escherichiacoli to yeast and intestinal cells Infect Immun 55 472ndash476
Fukiya S Mizoguchi H Tobe T and Mori H (2004)Extensive genomic diversity in pathogenic Escherichia coliand Shigella strains revealed by comparative genomichybridization microarray J Bacteriol 186 3911ndash3921
Hooton TM and Stamm WE (1997) Diagnosis and treat-ment of uncomplicated urinary tract infection Infect DisClin North Am 11 551ndash581
Horovitz A and Levitzki A (1987) An accurate method fordetermination of receptor ligand and enzyme-inhibitor dis-sociation-constants from displacement curves Proc NatlAcad Sci USA 84 6654ndash6658
Hull RA Gill RE Hsu P Minshaw BH and Falkow S(1981) Construction and expression of recombinant plas-mids encoding type 1 and D-mannose-resistant pili from aurinary tract infection Escherichia coli isolate Infect Immun33 933ndash938
Hultgren SJ Porter TN Schaeffer AJ and Duncan JL(1985) Role of type 1 pili and effects of phase variation onlower urinary tract infections produced by Escherichia coliInfect Immun 50 370ndash377
Hung C-S Bouckaert J Hung DL Pinkner J WinbergC Defusco A et al (2002) Structural basis of tropism ofEscherichia coli to the bladder during urinary tract infec-tion Mol Microbiol 44 903ndash915
Jones TA Zou JY Cowan SW and Kjeldgaard M(1991) Improved methods for building protein models inelectron density maps and the location of errors in thesemodels Acta Crystallogr A 47 110ndash119
Justice SS Hung C-S Theriot JA Fletcher DAAnderson GG Footer MJ and Hulgren SJ (2004)Differentiation and developmental pathways of uropatho-genic Escherichia coli in urinary tract pathogenesis ProcNatl Acad Sci USA 101 1333ndash1338
Kallenius G Mollby R Svenson SB Windberg J Lun-dblud A Svenson S and Cedergen B (1980) The Pk
antigen as receptor for the haemagglutinin of pyelonephri-togenic Escherichia coli FEMS Microbiol Lett 8 297ndash302
Knight SD Berglund J and Choudhury D (2000) Bacte-rial adhesins structural studies reveal chaperone functionand pilus biogenesis Curr Opin Chem Biol 4 653ndash660
Langermann S and Ballou WR (2003) Development of arecombinant FimCH vaccine for urinary tract infectionsAdv Exp Med Biol 539 635ndash653
Langermann S Palaszynski S Barnhart M Auguste GPinkner JS Burlein J et al (1997) Prevention ofmucosal Escherichia coli infection by FimH-adhesin-basedsystemic vaccination Science 276 607ndash611
Langermann S Mollby R Burlein JE Palaszynski SRAuguste CG Defusco A et al (2000) Vaccination withFimH adhesin protects cynomolgus monkeys from coloni-zation and infection by uropathogenic Escherichia coli JInfect Dis 181 774ndash778
Larsson A Ohlsson J Dodson KW Hultgren SJ Nils-son U and Kihlberg J (2003) Quantitative studies of thebinding of the class II PapG adhesin from uropathogenic
454 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
Escherichia coli to oligosaccharides Bioorg Med Chem 112255ndash2261
Leffler H and Svanborg-Eden C (1980) Chemical identifi-cation of a glycosphingolipid receptor for Escherichia coliattaching to human urinary tract epithelial cells and agglu-tinating human erythrocytes FEMS Microbiol Lett 8 127ndash134
Lindhorst TK Kieburg C and Krallmann-Wenzel U(1998) Inhibition of the type 1 fimbriae-mediated adhesionof Escherichia coli to erythrocytes by multiantennary alpha-mannosyl clusters the effect of multivalency Glycoconj J15 605ndash613
Loris R Tielker D Jaeger KE and Wyns L (2003a)Structural basis of carbohydrate recognition by the lectinLecB from Pseudomonas aeruginosa J Mol Biol 331 861ndash870
Loris R Imberty A Beeckmans S Van Driessche ERead JS Bouckaert J et al (2003b) Crystal structureof Pterocarpus angolensis lectin in complex with glucosesucrose and turanose J Biol Chem 278 16297ndash16303
Lund B Lindberg F Marklund BI and Normark S(1987) The PapG protein is the a-D-galactopyranosyl-(1-4)-b-D-galactopyranose-binding adhesin of uropathogenicEscherichia coli Proc Natl Acad Sci USA 84 5898ndash5902
Min G Stolz M Zhou G Liang F Sebbel P StofflerD et al (2002) Localization of uroplakin Ia the urothelialreceptor for bacterial adhesin FimH on the six innerdomains of the 16 nm urothelial plaque particle J Mol Biol317 697ndash706
Miroux B and Walker JE (1996) Over-production of pro-teins in Escherichia coli mutant hosts that allow synthesisof some membrane proteins and globular proteins at highlevels J Mol Biol 260 289ndash298
Morris GM Goodsell DS Halliday RS Huey R HartWE Belew RK and Olson AJ (1998) Automateddocking using a Lamarckian genetic algorithm and anempirical binding free energy function J Comput Chem 191639ndash1662
Mulvey MA (2002) Adhesion and entry of uropathogenicEscherichia coli Cell Microbiol 4 257ndash271
Mulvey MA Lopez-Boado YS Wilson CL Roth RParks WC Heuser J and Hultgren SJ (1998) Induc-tion and evasion of host defenses by type 1-piliated uro-pathogenic Escherichia coli Science 282 1494ndash1497
Murshudov GN Vagin AA and Dodson EJ (1997)Refinement of macromolecular structures by the maxi-mum-likelihood method Acta Crystallogr D Biol Crystallogr53 240ndash255
Nagahori N Lee RT Nishimura S Pageacute D Roy Rand Lee YC (2002) Inhibition of adhesion of type 1 fim-briated Escherichia coli to highly mannosylated ligandsChembiochem 3 836ndash844
Navaza J (2001) Implementation of molecular replacementin AMoRe Acta Crystallogr D Biol Crystallogr 57 1367ndash1372
Neeser JR Koellreutter B and Wuersch P (1986)Oligomannoside-type glycopeptides inhibiting adhesion ofEscherichia coli strains mediated by type 1 pili preparationof potent inhibitors from plant glycoproteins Infect Immun52 428ndash436
Old DC (1972) Inhibition of the interaction between fimbrial
hemagglutinatinins and erythrocytes by D-mannose andother carbohydrates J Gen Microbiol 71 149ndash157
Orndorff PE Devapali A Palestrant S Wyse A EverettML Bollinger RR and Parker W (2004) Immunoglob-ulin-mediated agglutination of and biofilm formation byEscherichia coli K-12 require the type 1 pilus fiber InfectImmun 72 1929ndash1938
Oscarson S and Tedebark U (1995) Syntheses of deoxyanalogues of methyl 36-di-O-a-D-mannopyranosyl-a-D-mannopyranoside for studies of the binding site of con-canavalin A Carbohydr Res 278 271ndash287
Oscarson S and Tiden AK (1993) Syntheses of the octyland tetradecyl glycosides of 36-di-O-alpha-deuterium-mannopyranosyl-alpha-deuterium-mannopyranose and of34-di-O-alpha-deuterium-mannopyranosyl-alpha-deute-rium-mannopyranose ndash a new way for 24-di-O-protectionof mannopyranosides Carbohydr Res 247 323ndash328
Otwinowski Z and Minor W (1997) Processing of X-raydiffraction data collected in oscillation mode MethodsEnzymol 276 307ndash326
Powell HR (1999) The Rossmann Fourier autoindexingalgorithm in MOSFLM Acta Crystallogr D Biol Crystallogr55 1690ndash1695
Pratt LA and Kolter R (1998) Genetic analysis of Escher-ichia coli biofilm formation roles of flagella motility chemo-taxis and type I pili Mol Microbiol 30 285ndash293
Remaut H and Waksman G (2004) Structural biology ofbacterial pathogenesis Curr Opin Struct Biol 14 161ndash170
Ronald AR Nicolle LE Stamm E Krieger J WarrenJ Schaeffer A et al (2001) Urinary tract infection inadults research priorities and strategies Int J AntimicrobAgents 17 343ndash348
Sanner MF Olson AJ and Spehner JC (1996)Reduced surface an efficient way to compute molecularsurfaces Biopolymers 38 305ndash320
Sauer FG Futterer K Pinkner JS Dodson KW Hult-gren SJ and Waksman G (1999) Structural basis ofchaperone function and pilus biogenesis Science 2851058ndash1061
Sauer FG Knight SD Waksman GW and HultgrenSJ (2000) PapD-like chaperones and pilus biogenesisSemin Cell Dev Biol 11 27ndash34
Sauer FG Pinkner JS Waksman G and Hultgren SJ(2002) Chaperone priming of pilus subunits facilitates atopological transition that drives fiber formation Cell 111543ndash551
Schembri MA Hasman H and Klemm P (2000)Expression and purification of the mannose recognitiondomain of the FimH adhesin FEMS Microbiol Lett 188147ndash151
Schembri MA Kjaergaard K and Klemm P (2003) Glo-bal gene expression in Escherichia coli biofilms Mol Micro-biol 48 253ndash267
Schilling JD Mulvey MA Vincent CD Lorenz RGand Hultgren SJ (2001) Bacterial invasion augments epi-thelial cytokine responses to Escherichia coli through alipopolysaccharide-dependent mechanism J Immunol166 1148ndash1155
Silhavy TJ Szmelcman S Boos W and Schwartz M(1975) On the significance of the retention of ligand byprotein Proc Natl Acad Sci USA 72 2120ndash2124
A novel class of FimH high-affinity ligands 455
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
Svanborg C and Godaly G (1997) Bacterial virulence inurinary tract infection Infect Dis Clin North Am 11 513ndash529
Thanassi DG Saulino ET and Hultgren SJ (1998) Thechaperoneusher pathway a major terminal branch of thegeneral secretory pathway Curr Opin Struct Biol 1 223ndash231
Thankavel K Madison B Ikeda T Malaviya R ShahAH Arumugen PM and Abraham SN (1997) Local-ization of a domain in the FimH adhesin of Escherichia colitype 1 fimbriae capable of receptor recognition and use ofa domain-specific antibody to confer protection againstexperimental urinary tract infection J Clin Invest 1001123ndash1126
Vagin A and Teplyakov A (1997) MOLREP an automatedprogram for molecular replacement J Appl Crystallogr 301022ndash1025
Wu XR Sun TT and Medina JJ (1996) In vitro bindingof type 1-fimbriated Escherichia coli to uroplakins 1a and
1b relation to urinary tract infections Proc Natl Acad SciUSA 93 9630ndash9635
Xia Y Gally D Forsman-Semb K and Uhlin BE (2000)Regulatory cross-talk between adhesin operons in Escher-ichia coli inhibition of type 1 fimbriae expression by thePapB protein EMBO J 19 1450ndash1457
Zafriri D Ofek I Adar R Pocino M and Sharon N(1989) Inhibitory activity of cranberry juice on adherenceof type-1 and type-P fimbriated Escherichia coli to eukary-otic cells Antimicrob Agents Chemother 33 92ndash98
Zavialov AV Berglund J Pudney AF Fooks LJ Ibra-him TM MacIntyre S and Knight SD (2003) Structureand biogenesis of the capsular F1 antigen from Yersiniapestis preserved folding energy drives fiber formation Cell113 587ndash596
Zhou G Mo WJ Sebbel P Min G Neubert TA Glock-shuber R et al (2001) Uroplakin Ia is the urothelial recep-tor for uropathogenic Escherichia coli evidence from invitro FimH binding J Cell Sci 114 4095ndash4103

452 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
As a control the Fabs of monoclonal antibody IA7 as wellas BSA-Man were also immobilized Samples containing afixed concentration of FimH were used (close to the Kd of theFimHndashantibody interaction) in combination with varying con-centrations of saccharide First 10-fold dilutions of the sac-charide solution were used to determine the concentrationrange for binding of the saccharide to FimH A Langmuirbinding isotherm with a 11 stoichiometry was fitted to thedata using the kinetic constants and Rmax from the foregoingexperiment to obtain the concentrations of FimH that werefree ([FimH]free) to bind the antibody immobilized on the chipSecond the concentration range of the saccharide wasextended and adapted to ensure accurate fitting and theequilibrium binding constant of the FimHndashsaccharide interac-tion was derived from the curve of [FimH]free against concen-tration of saccharide Every measurement was repeated atleast twice including testing different protein batches andwhenever possible also different saccharide stock solutions(typically 200 mM)
Equilibrium binding studies using radioactively labelledmannose Mannose-free FimHtr2 was produced inM9 minimal medium and purified as described above forFimHtr2 Alkyl mannosides were dissolved in double distilledwater pNPaMan and MeUmbaMan (6 mg each) were dis-solved in 20 ml of DMSO and diluted to 20 mM using doubledistilled water Binding experiments were performed at 310 Kusing six different concentrations of [3H]-a-D-mannose (finalconcentrations 435 mM 290 mM 193 mM 129 mM 86 mMand 57 mM) Protein (180 ml) at a concentration of about500 nM was mixed with 20 ml of the radioactive ligand [3H]-a-D-mannose and incubated at 37degC for 20 min To separatefree ligand from bound the mixture was rapidly filtratedthrough a Protran BA 85 cellulose nitrate filter (Schleicherand Schuell) and washed once with 1 ml of ice-cold 1yen PBSFilter-bound radioactivity was measured by scintillation spec-trometry within 24 h Displacement experiments were per-formed using six different concentrations of inhibitor (finalconcentrations in the range 00ndash435 mM) in the presence of435 mM [3H]-a-D-mannose Twenty microlitres of radioactiveligand 20 ml of inhibitor and 160 ml of protein (500 nM) weremixed and the experiments were performed as describedabove All experiments were performed in duplicates and thedata presented are mean values of at least two independentmeasurements For the determination of Kd for a-D-mannosean hyperbolic curve (y = P1x(P2 + x) where P1 is an arbitraryscale factor and P2 = Kd) was fitted to the data For the dis-placement experiments the curve y = P1(P2 + x) where P1
is an arbitrary scale factor and P2 is the concentration of theinhibitor displacing 50 of the labelled ligand [I]05 was usedinstead To calculate the inhibitor dissociation constant (KI)the Cheng and Prusoff equation [KI = [I]05[L]KL + 1) KL is theconstant of dissociation for the ligand] (Cheng and Prusoff1973) was used when both the concentration of the radio-active ligand (L) and the displacing agent (I) are in excessover the protein (LT gtgt PT IT gtgt PT T indicates total concen-tration) For the two strong inhibitors pNPaMan and MeUm-baMan where IT was not in excess over PT the equationof KI = IT[(1 - Y)Y(LTKL) - 1] - PTKLYLT where Y is thefraction of the ligand bound in presence of the inhibitor wasused instead (Horovitz and Levitzki 1987)
Computational docking
Computational docking studies were performed usingAutoDock3 (Morris et al 1998) In AutoDock3 the proteinis represented by a grid of affinity potentials Hence theprotein conformation is not allowed to change during thedocking procedure Flexibility of ligands is taken intoaccount by allowing rotation around flexible torsion anglesThe docked energies are calculated as the sum of the inter-molecular interaction energy and the internal energy of theligand The search for the best interaction energy was car-ried out by means of the Lamarckian genetic algorithmLigand structures were obtained from the Protein DataBank and prepared for docking by adding hydrogens andcharges using AutoDockTools (Sanner et al 1996) Eachsimulation consisted of 100 independent runs with a popu-lation size of 200 500 generations and a maximum of25 000 000 energy evaluations Solutions were rankedbased on their docking energies and similar solutions wereclustered (cluster cut-off rmsd lt 1 Aring) For both ligandsthe top solutions were evaluated using the graphics pro-gram O (Jones et al 1991)
Acknowledgements
We thank Andrej Zavialov and Maringhns Ehrenberg for help withequilibrium binding experiments and Thomas Norberg forassistance with mass spectrometry Anthony DeFusco fromMedimmune provided us with mannosylated bovine serumalbumin We are grateful to Fred Sauer for assistance withdata collection Thanks to Maor Bar-Peled of the CCC at theUniversity of Georgia Athens USA for his suggestion toexpress FimH in minimal medium We acknowledge the useof beamline 19BM of the Structural Biology Center at theAdvanced Photon Source Argonne National LaboratoryChicago IL USA and of beamline ID29 ESRF GrenobleFrance JB is a postdoctoral fellow of the Fonds voor Weten-schappelijk Onderzoek-Vlaanderen which also supportedthe BiaCore3000 instrument (FWONL35) This work was sup-ported by grants from the Swedish Research Council andfrom the Strategic Research Foundation (Structural BiologyNetwork) to SDK
Supplementary material
The following material is available from httpwwwblackwellpublishingcomproductsjournalssuppmatmmimmi4415mmi4415smhtmTable S1 Crystallographic data and refinement statistics
References
Anumula KR (1994) Quantitative determination ofmonosaccharides in glycoproteins by high-performanceliquid chromatography with highly sensitive fluorescencedetection Anal Biochem 220 275ndash283
Bahrani-Mougeot FK Buckles EL Lockatell CV HebelJR Johnson DE Tang CM and Donnenberg MS(2002) Type 1 fimbriae and extracellular polysaccharidesare preeminent uropathogenic Escherichia coli virulence
A novel class of FimH high-affinity ligands 453
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
determinants in the murine urinary tract Mol Microbiol 451079ndash1093
Berglund J and Knight SD (2003) Structural basis forbacterial adhesion in the urinary tract Glycobiol Med 53533ndash52
Blattner FR Plunkett G Bloch CA Perna NT BurlandV Riley M et al (1997) The complete genome sequenceof Escherichia coli K-12 Science 277 1453ndashbullbull
Bloch CA Stocker BA and Orndorff PE (1992) A keyrole for type 1 pili in enterobacterial communicability MolMicrobiol 6 697ndash701
Bork P Holm L and Sander C (1994) The immunoglob-ulin fold ndash structural classification sequence patterns andcommon core J Mol Biol 242 309ndash320
Brinton CC (1959) Non-flagellar appendages of bacteriaNature 183 782ndash786
Bruumlnger AT Adams PD Clore GM DeLano WLGros P Grosse-Kunstleve RW et al (1998) Crystallog-raphy and NMR system a new software suite for macro-molecular structure determination Acta Crystallogr D BiolCrystallogr 54 905ndash921
Buchanan K Falkow S Hull RA and Hull SI (1985)Frequency among Enterobacteriaceae of the DNA-sequences encoding type-1 pili J Bacteriol 162 799ndash803
Buts L Bouckaert J De Genst E Loris R Oscarson SLahmann M et al (2003) The fimbrial adhesin F17-G ofenterotoxigenic Escherichia coli has an immunoglobulin-like lectin domain that binds N-acetylglucosamine MolMicrobiol 49 705ndash715
Cheng Y and Prusoff WH (1973) Relationship betweeninhibition constant (K1) and concentration of inhibitor whichcauses 50 per cent inhibition (I50) of an enzymatic reac-tion Biochem Pharmacol 22 3099ndash3108
Choudhury D Thompson A Stojanoff V LangermannS Pinkner J Hultgren SJ and Knight SD (1999) X-ray structure of the FimCndashFimH chaperonendashadhesin com-plex from uropathogenic Escherichia coli Science 2851061ndash1066
Collaborative computational project n4 (1994) The CCP4suite programs for crystallography Acta Crystallogr D BiolCrystallogr 50 760ndash763
Dodson KW Pinkner JS Rose T Magnusson G Hult-gren SJ and Waksman G (2001) Structural basis of theinteraction of the pyelonephritic Escherichia coli adhesin toits human kidney receptor Cell 105 733ndash743
Eley JG and Triumalashetty P (2001) In vitro assessmentof alkylglycosides as permeability enhancers AAPSPharmSciTech 2 (3) Article 19
Esnouf RM (1997) An extensively modified version of Mol-script that includes greatly enhanced coloring capabilitiesJ Mol Graph Model 15 132ndash134
Firon N Ofek I and Sharon N (1982) Interaction ofmannose-containing oligosaccharides with the fimbriallectin of Escherichia coli Biochem Biophys Res Commun105 1426ndash1432
Firon N Ofek I and Sharon N (1983) Carbohydrate spec-ificity of the surface lectins of Escherichia coli Klebsiellapneumoniae and Salmonella typhimurium Carbohydr Res120 235ndash249
Firon N Ofek I and Sharon N (1984) Carbohydrate-
binding sites of the mannose-specific fimbrial lectins ofEnterobacteria Infect Immun 43 1088ndash1090
Firon N Ashkenazis S Mirelman D Ofek I and SharonN (1987) Aromatic a-glycosides of mannose are powerfulinhibitors of the adherence of type 1 fimbriated Escherichiacoli to yeast and intestinal cells Infect Immun 55 472ndash476
Fukiya S Mizoguchi H Tobe T and Mori H (2004)Extensive genomic diversity in pathogenic Escherichia coliand Shigella strains revealed by comparative genomichybridization microarray J Bacteriol 186 3911ndash3921
Hooton TM and Stamm WE (1997) Diagnosis and treat-ment of uncomplicated urinary tract infection Infect DisClin North Am 11 551ndash581
Horovitz A and Levitzki A (1987) An accurate method fordetermination of receptor ligand and enzyme-inhibitor dis-sociation-constants from displacement curves Proc NatlAcad Sci USA 84 6654ndash6658
Hull RA Gill RE Hsu P Minshaw BH and Falkow S(1981) Construction and expression of recombinant plas-mids encoding type 1 and D-mannose-resistant pili from aurinary tract infection Escherichia coli isolate Infect Immun33 933ndash938
Hultgren SJ Porter TN Schaeffer AJ and Duncan JL(1985) Role of type 1 pili and effects of phase variation onlower urinary tract infections produced by Escherichia coliInfect Immun 50 370ndash377
Hung C-S Bouckaert J Hung DL Pinkner J WinbergC Defusco A et al (2002) Structural basis of tropism ofEscherichia coli to the bladder during urinary tract infec-tion Mol Microbiol 44 903ndash915
Jones TA Zou JY Cowan SW and Kjeldgaard M(1991) Improved methods for building protein models inelectron density maps and the location of errors in thesemodels Acta Crystallogr A 47 110ndash119
Justice SS Hung C-S Theriot JA Fletcher DAAnderson GG Footer MJ and Hulgren SJ (2004)Differentiation and developmental pathways of uropatho-genic Escherichia coli in urinary tract pathogenesis ProcNatl Acad Sci USA 101 1333ndash1338
Kallenius G Mollby R Svenson SB Windberg J Lun-dblud A Svenson S and Cedergen B (1980) The Pk
antigen as receptor for the haemagglutinin of pyelonephri-togenic Escherichia coli FEMS Microbiol Lett 8 297ndash302
Knight SD Berglund J and Choudhury D (2000) Bacte-rial adhesins structural studies reveal chaperone functionand pilus biogenesis Curr Opin Chem Biol 4 653ndash660
Langermann S and Ballou WR (2003) Development of arecombinant FimCH vaccine for urinary tract infectionsAdv Exp Med Biol 539 635ndash653
Langermann S Palaszynski S Barnhart M Auguste GPinkner JS Burlein J et al (1997) Prevention ofmucosal Escherichia coli infection by FimH-adhesin-basedsystemic vaccination Science 276 607ndash611
Langermann S Mollby R Burlein JE Palaszynski SRAuguste CG Defusco A et al (2000) Vaccination withFimH adhesin protects cynomolgus monkeys from coloni-zation and infection by uropathogenic Escherichia coli JInfect Dis 181 774ndash778
Larsson A Ohlsson J Dodson KW Hultgren SJ Nils-son U and Kihlberg J (2003) Quantitative studies of thebinding of the class II PapG adhesin from uropathogenic
454 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
Escherichia coli to oligosaccharides Bioorg Med Chem 112255ndash2261
Leffler H and Svanborg-Eden C (1980) Chemical identifi-cation of a glycosphingolipid receptor for Escherichia coliattaching to human urinary tract epithelial cells and agglu-tinating human erythrocytes FEMS Microbiol Lett 8 127ndash134
Lindhorst TK Kieburg C and Krallmann-Wenzel U(1998) Inhibition of the type 1 fimbriae-mediated adhesionof Escherichia coli to erythrocytes by multiantennary alpha-mannosyl clusters the effect of multivalency Glycoconj J15 605ndash613
Loris R Tielker D Jaeger KE and Wyns L (2003a)Structural basis of carbohydrate recognition by the lectinLecB from Pseudomonas aeruginosa J Mol Biol 331 861ndash870
Loris R Imberty A Beeckmans S Van Driessche ERead JS Bouckaert J et al (2003b) Crystal structureof Pterocarpus angolensis lectin in complex with glucosesucrose and turanose J Biol Chem 278 16297ndash16303
Lund B Lindberg F Marklund BI and Normark S(1987) The PapG protein is the a-D-galactopyranosyl-(1-4)-b-D-galactopyranose-binding adhesin of uropathogenicEscherichia coli Proc Natl Acad Sci USA 84 5898ndash5902
Min G Stolz M Zhou G Liang F Sebbel P StofflerD et al (2002) Localization of uroplakin Ia the urothelialreceptor for bacterial adhesin FimH on the six innerdomains of the 16 nm urothelial plaque particle J Mol Biol317 697ndash706
Miroux B and Walker JE (1996) Over-production of pro-teins in Escherichia coli mutant hosts that allow synthesisof some membrane proteins and globular proteins at highlevels J Mol Biol 260 289ndash298
Morris GM Goodsell DS Halliday RS Huey R HartWE Belew RK and Olson AJ (1998) Automateddocking using a Lamarckian genetic algorithm and anempirical binding free energy function J Comput Chem 191639ndash1662
Mulvey MA (2002) Adhesion and entry of uropathogenicEscherichia coli Cell Microbiol 4 257ndash271
Mulvey MA Lopez-Boado YS Wilson CL Roth RParks WC Heuser J and Hultgren SJ (1998) Induc-tion and evasion of host defenses by type 1-piliated uro-pathogenic Escherichia coli Science 282 1494ndash1497
Murshudov GN Vagin AA and Dodson EJ (1997)Refinement of macromolecular structures by the maxi-mum-likelihood method Acta Crystallogr D Biol Crystallogr53 240ndash255
Nagahori N Lee RT Nishimura S Pageacute D Roy Rand Lee YC (2002) Inhibition of adhesion of type 1 fim-briated Escherichia coli to highly mannosylated ligandsChembiochem 3 836ndash844
Navaza J (2001) Implementation of molecular replacementin AMoRe Acta Crystallogr D Biol Crystallogr 57 1367ndash1372
Neeser JR Koellreutter B and Wuersch P (1986)Oligomannoside-type glycopeptides inhibiting adhesion ofEscherichia coli strains mediated by type 1 pili preparationof potent inhibitors from plant glycoproteins Infect Immun52 428ndash436
Old DC (1972) Inhibition of the interaction between fimbrial
hemagglutinatinins and erythrocytes by D-mannose andother carbohydrates J Gen Microbiol 71 149ndash157
Orndorff PE Devapali A Palestrant S Wyse A EverettML Bollinger RR and Parker W (2004) Immunoglob-ulin-mediated agglutination of and biofilm formation byEscherichia coli K-12 require the type 1 pilus fiber InfectImmun 72 1929ndash1938
Oscarson S and Tedebark U (1995) Syntheses of deoxyanalogues of methyl 36-di-O-a-D-mannopyranosyl-a-D-mannopyranoside for studies of the binding site of con-canavalin A Carbohydr Res 278 271ndash287
Oscarson S and Tiden AK (1993) Syntheses of the octyland tetradecyl glycosides of 36-di-O-alpha-deuterium-mannopyranosyl-alpha-deuterium-mannopyranose and of34-di-O-alpha-deuterium-mannopyranosyl-alpha-deute-rium-mannopyranose ndash a new way for 24-di-O-protectionof mannopyranosides Carbohydr Res 247 323ndash328
Otwinowski Z and Minor W (1997) Processing of X-raydiffraction data collected in oscillation mode MethodsEnzymol 276 307ndash326
Powell HR (1999) The Rossmann Fourier autoindexingalgorithm in MOSFLM Acta Crystallogr D Biol Crystallogr55 1690ndash1695
Pratt LA and Kolter R (1998) Genetic analysis of Escher-ichia coli biofilm formation roles of flagella motility chemo-taxis and type I pili Mol Microbiol 30 285ndash293
Remaut H and Waksman G (2004) Structural biology ofbacterial pathogenesis Curr Opin Struct Biol 14 161ndash170
Ronald AR Nicolle LE Stamm E Krieger J WarrenJ Schaeffer A et al (2001) Urinary tract infection inadults research priorities and strategies Int J AntimicrobAgents 17 343ndash348
Sanner MF Olson AJ and Spehner JC (1996)Reduced surface an efficient way to compute molecularsurfaces Biopolymers 38 305ndash320
Sauer FG Futterer K Pinkner JS Dodson KW Hult-gren SJ and Waksman G (1999) Structural basis ofchaperone function and pilus biogenesis Science 2851058ndash1061
Sauer FG Knight SD Waksman GW and HultgrenSJ (2000) PapD-like chaperones and pilus biogenesisSemin Cell Dev Biol 11 27ndash34
Sauer FG Pinkner JS Waksman G and Hultgren SJ(2002) Chaperone priming of pilus subunits facilitates atopological transition that drives fiber formation Cell 111543ndash551
Schembri MA Hasman H and Klemm P (2000)Expression and purification of the mannose recognitiondomain of the FimH adhesin FEMS Microbiol Lett 188147ndash151
Schembri MA Kjaergaard K and Klemm P (2003) Glo-bal gene expression in Escherichia coli biofilms Mol Micro-biol 48 253ndash267
Schilling JD Mulvey MA Vincent CD Lorenz RGand Hultgren SJ (2001) Bacterial invasion augments epi-thelial cytokine responses to Escherichia coli through alipopolysaccharide-dependent mechanism J Immunol166 1148ndash1155
Silhavy TJ Szmelcman S Boos W and Schwartz M(1975) On the significance of the retention of ligand byprotein Proc Natl Acad Sci USA 72 2120ndash2124
A novel class of FimH high-affinity ligands 455
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
Svanborg C and Godaly G (1997) Bacterial virulence inurinary tract infection Infect Dis Clin North Am 11 513ndash529
Thanassi DG Saulino ET and Hultgren SJ (1998) Thechaperoneusher pathway a major terminal branch of thegeneral secretory pathway Curr Opin Struct Biol 1 223ndash231
Thankavel K Madison B Ikeda T Malaviya R ShahAH Arumugen PM and Abraham SN (1997) Local-ization of a domain in the FimH adhesin of Escherichia colitype 1 fimbriae capable of receptor recognition and use ofa domain-specific antibody to confer protection againstexperimental urinary tract infection J Clin Invest 1001123ndash1126
Vagin A and Teplyakov A (1997) MOLREP an automatedprogram for molecular replacement J Appl Crystallogr 301022ndash1025
Wu XR Sun TT and Medina JJ (1996) In vitro bindingof type 1-fimbriated Escherichia coli to uroplakins 1a and
1b relation to urinary tract infections Proc Natl Acad SciUSA 93 9630ndash9635
Xia Y Gally D Forsman-Semb K and Uhlin BE (2000)Regulatory cross-talk between adhesin operons in Escher-ichia coli inhibition of type 1 fimbriae expression by thePapB protein EMBO J 19 1450ndash1457
Zafriri D Ofek I Adar R Pocino M and Sharon N(1989) Inhibitory activity of cranberry juice on adherenceof type-1 and type-P fimbriated Escherichia coli to eukary-otic cells Antimicrob Agents Chemother 33 92ndash98
Zavialov AV Berglund J Pudney AF Fooks LJ Ibra-him TM MacIntyre S and Knight SD (2003) Structureand biogenesis of the capsular F1 antigen from Yersiniapestis preserved folding energy drives fiber formation Cell113 587ndash596
Zhou G Mo WJ Sebbel P Min G Neubert TA Glock-shuber R et al (2001) Uroplakin Ia is the urothelial recep-tor for uropathogenic Escherichia coli evidence from invitro FimH binding J Cell Sci 114 4095ndash4103

A novel class of FimH high-affinity ligands 453
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
determinants in the murine urinary tract Mol Microbiol 451079ndash1093
Berglund J and Knight SD (2003) Structural basis forbacterial adhesion in the urinary tract Glycobiol Med 53533ndash52
Blattner FR Plunkett G Bloch CA Perna NT BurlandV Riley M et al (1997) The complete genome sequenceof Escherichia coli K-12 Science 277 1453ndashbullbull
Bloch CA Stocker BA and Orndorff PE (1992) A keyrole for type 1 pili in enterobacterial communicability MolMicrobiol 6 697ndash701
Bork P Holm L and Sander C (1994) The immunoglob-ulin fold ndash structural classification sequence patterns andcommon core J Mol Biol 242 309ndash320
Brinton CC (1959) Non-flagellar appendages of bacteriaNature 183 782ndash786
Bruumlnger AT Adams PD Clore GM DeLano WLGros P Grosse-Kunstleve RW et al (1998) Crystallog-raphy and NMR system a new software suite for macro-molecular structure determination Acta Crystallogr D BiolCrystallogr 54 905ndash921
Buchanan K Falkow S Hull RA and Hull SI (1985)Frequency among Enterobacteriaceae of the DNA-sequences encoding type-1 pili J Bacteriol 162 799ndash803
Buts L Bouckaert J De Genst E Loris R Oscarson SLahmann M et al (2003) The fimbrial adhesin F17-G ofenterotoxigenic Escherichia coli has an immunoglobulin-like lectin domain that binds N-acetylglucosamine MolMicrobiol 49 705ndash715
Cheng Y and Prusoff WH (1973) Relationship betweeninhibition constant (K1) and concentration of inhibitor whichcauses 50 per cent inhibition (I50) of an enzymatic reac-tion Biochem Pharmacol 22 3099ndash3108
Choudhury D Thompson A Stojanoff V LangermannS Pinkner J Hultgren SJ and Knight SD (1999) X-ray structure of the FimCndashFimH chaperonendashadhesin com-plex from uropathogenic Escherichia coli Science 2851061ndash1066
Collaborative computational project n4 (1994) The CCP4suite programs for crystallography Acta Crystallogr D BiolCrystallogr 50 760ndash763
Dodson KW Pinkner JS Rose T Magnusson G Hult-gren SJ and Waksman G (2001) Structural basis of theinteraction of the pyelonephritic Escherichia coli adhesin toits human kidney receptor Cell 105 733ndash743
Eley JG and Triumalashetty P (2001) In vitro assessmentof alkylglycosides as permeability enhancers AAPSPharmSciTech 2 (3) Article 19
Esnouf RM (1997) An extensively modified version of Mol-script that includes greatly enhanced coloring capabilitiesJ Mol Graph Model 15 132ndash134
Firon N Ofek I and Sharon N (1982) Interaction ofmannose-containing oligosaccharides with the fimbriallectin of Escherichia coli Biochem Biophys Res Commun105 1426ndash1432
Firon N Ofek I and Sharon N (1983) Carbohydrate spec-ificity of the surface lectins of Escherichia coli Klebsiellapneumoniae and Salmonella typhimurium Carbohydr Res120 235ndash249
Firon N Ofek I and Sharon N (1984) Carbohydrate-
binding sites of the mannose-specific fimbrial lectins ofEnterobacteria Infect Immun 43 1088ndash1090
Firon N Ashkenazis S Mirelman D Ofek I and SharonN (1987) Aromatic a-glycosides of mannose are powerfulinhibitors of the adherence of type 1 fimbriated Escherichiacoli to yeast and intestinal cells Infect Immun 55 472ndash476
Fukiya S Mizoguchi H Tobe T and Mori H (2004)Extensive genomic diversity in pathogenic Escherichia coliand Shigella strains revealed by comparative genomichybridization microarray J Bacteriol 186 3911ndash3921
Hooton TM and Stamm WE (1997) Diagnosis and treat-ment of uncomplicated urinary tract infection Infect DisClin North Am 11 551ndash581
Horovitz A and Levitzki A (1987) An accurate method fordetermination of receptor ligand and enzyme-inhibitor dis-sociation-constants from displacement curves Proc NatlAcad Sci USA 84 6654ndash6658
Hull RA Gill RE Hsu P Minshaw BH and Falkow S(1981) Construction and expression of recombinant plas-mids encoding type 1 and D-mannose-resistant pili from aurinary tract infection Escherichia coli isolate Infect Immun33 933ndash938
Hultgren SJ Porter TN Schaeffer AJ and Duncan JL(1985) Role of type 1 pili and effects of phase variation onlower urinary tract infections produced by Escherichia coliInfect Immun 50 370ndash377
Hung C-S Bouckaert J Hung DL Pinkner J WinbergC Defusco A et al (2002) Structural basis of tropism ofEscherichia coli to the bladder during urinary tract infec-tion Mol Microbiol 44 903ndash915
Jones TA Zou JY Cowan SW and Kjeldgaard M(1991) Improved methods for building protein models inelectron density maps and the location of errors in thesemodels Acta Crystallogr A 47 110ndash119
Justice SS Hung C-S Theriot JA Fletcher DAAnderson GG Footer MJ and Hulgren SJ (2004)Differentiation and developmental pathways of uropatho-genic Escherichia coli in urinary tract pathogenesis ProcNatl Acad Sci USA 101 1333ndash1338
Kallenius G Mollby R Svenson SB Windberg J Lun-dblud A Svenson S and Cedergen B (1980) The Pk
antigen as receptor for the haemagglutinin of pyelonephri-togenic Escherichia coli FEMS Microbiol Lett 8 297ndash302
Knight SD Berglund J and Choudhury D (2000) Bacte-rial adhesins structural studies reveal chaperone functionand pilus biogenesis Curr Opin Chem Biol 4 653ndash660
Langermann S and Ballou WR (2003) Development of arecombinant FimCH vaccine for urinary tract infectionsAdv Exp Med Biol 539 635ndash653
Langermann S Palaszynski S Barnhart M Auguste GPinkner JS Burlein J et al (1997) Prevention ofmucosal Escherichia coli infection by FimH-adhesin-basedsystemic vaccination Science 276 607ndash611
Langermann S Mollby R Burlein JE Palaszynski SRAuguste CG Defusco A et al (2000) Vaccination withFimH adhesin protects cynomolgus monkeys from coloni-zation and infection by uropathogenic Escherichia coli JInfect Dis 181 774ndash778
Larsson A Ohlsson J Dodson KW Hultgren SJ Nils-son U and Kihlberg J (2003) Quantitative studies of thebinding of the class II PapG adhesin from uropathogenic
454 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
Escherichia coli to oligosaccharides Bioorg Med Chem 112255ndash2261
Leffler H and Svanborg-Eden C (1980) Chemical identifi-cation of a glycosphingolipid receptor for Escherichia coliattaching to human urinary tract epithelial cells and agglu-tinating human erythrocytes FEMS Microbiol Lett 8 127ndash134
Lindhorst TK Kieburg C and Krallmann-Wenzel U(1998) Inhibition of the type 1 fimbriae-mediated adhesionof Escherichia coli to erythrocytes by multiantennary alpha-mannosyl clusters the effect of multivalency Glycoconj J15 605ndash613
Loris R Tielker D Jaeger KE and Wyns L (2003a)Structural basis of carbohydrate recognition by the lectinLecB from Pseudomonas aeruginosa J Mol Biol 331 861ndash870
Loris R Imberty A Beeckmans S Van Driessche ERead JS Bouckaert J et al (2003b) Crystal structureof Pterocarpus angolensis lectin in complex with glucosesucrose and turanose J Biol Chem 278 16297ndash16303
Lund B Lindberg F Marklund BI and Normark S(1987) The PapG protein is the a-D-galactopyranosyl-(1-4)-b-D-galactopyranose-binding adhesin of uropathogenicEscherichia coli Proc Natl Acad Sci USA 84 5898ndash5902
Min G Stolz M Zhou G Liang F Sebbel P StofflerD et al (2002) Localization of uroplakin Ia the urothelialreceptor for bacterial adhesin FimH on the six innerdomains of the 16 nm urothelial plaque particle J Mol Biol317 697ndash706
Miroux B and Walker JE (1996) Over-production of pro-teins in Escherichia coli mutant hosts that allow synthesisof some membrane proteins and globular proteins at highlevels J Mol Biol 260 289ndash298
Morris GM Goodsell DS Halliday RS Huey R HartWE Belew RK and Olson AJ (1998) Automateddocking using a Lamarckian genetic algorithm and anempirical binding free energy function J Comput Chem 191639ndash1662
Mulvey MA (2002) Adhesion and entry of uropathogenicEscherichia coli Cell Microbiol 4 257ndash271
Mulvey MA Lopez-Boado YS Wilson CL Roth RParks WC Heuser J and Hultgren SJ (1998) Induc-tion and evasion of host defenses by type 1-piliated uro-pathogenic Escherichia coli Science 282 1494ndash1497
Murshudov GN Vagin AA and Dodson EJ (1997)Refinement of macromolecular structures by the maxi-mum-likelihood method Acta Crystallogr D Biol Crystallogr53 240ndash255
Nagahori N Lee RT Nishimura S Pageacute D Roy Rand Lee YC (2002) Inhibition of adhesion of type 1 fim-briated Escherichia coli to highly mannosylated ligandsChembiochem 3 836ndash844
Navaza J (2001) Implementation of molecular replacementin AMoRe Acta Crystallogr D Biol Crystallogr 57 1367ndash1372
Neeser JR Koellreutter B and Wuersch P (1986)Oligomannoside-type glycopeptides inhibiting adhesion ofEscherichia coli strains mediated by type 1 pili preparationof potent inhibitors from plant glycoproteins Infect Immun52 428ndash436
Old DC (1972) Inhibition of the interaction between fimbrial
hemagglutinatinins and erythrocytes by D-mannose andother carbohydrates J Gen Microbiol 71 149ndash157
Orndorff PE Devapali A Palestrant S Wyse A EverettML Bollinger RR and Parker W (2004) Immunoglob-ulin-mediated agglutination of and biofilm formation byEscherichia coli K-12 require the type 1 pilus fiber InfectImmun 72 1929ndash1938
Oscarson S and Tedebark U (1995) Syntheses of deoxyanalogues of methyl 36-di-O-a-D-mannopyranosyl-a-D-mannopyranoside for studies of the binding site of con-canavalin A Carbohydr Res 278 271ndash287
Oscarson S and Tiden AK (1993) Syntheses of the octyland tetradecyl glycosides of 36-di-O-alpha-deuterium-mannopyranosyl-alpha-deuterium-mannopyranose and of34-di-O-alpha-deuterium-mannopyranosyl-alpha-deute-rium-mannopyranose ndash a new way for 24-di-O-protectionof mannopyranosides Carbohydr Res 247 323ndash328
Otwinowski Z and Minor W (1997) Processing of X-raydiffraction data collected in oscillation mode MethodsEnzymol 276 307ndash326
Powell HR (1999) The Rossmann Fourier autoindexingalgorithm in MOSFLM Acta Crystallogr D Biol Crystallogr55 1690ndash1695
Pratt LA and Kolter R (1998) Genetic analysis of Escher-ichia coli biofilm formation roles of flagella motility chemo-taxis and type I pili Mol Microbiol 30 285ndash293
Remaut H and Waksman G (2004) Structural biology ofbacterial pathogenesis Curr Opin Struct Biol 14 161ndash170
Ronald AR Nicolle LE Stamm E Krieger J WarrenJ Schaeffer A et al (2001) Urinary tract infection inadults research priorities and strategies Int J AntimicrobAgents 17 343ndash348
Sanner MF Olson AJ and Spehner JC (1996)Reduced surface an efficient way to compute molecularsurfaces Biopolymers 38 305ndash320
Sauer FG Futterer K Pinkner JS Dodson KW Hult-gren SJ and Waksman G (1999) Structural basis ofchaperone function and pilus biogenesis Science 2851058ndash1061
Sauer FG Knight SD Waksman GW and HultgrenSJ (2000) PapD-like chaperones and pilus biogenesisSemin Cell Dev Biol 11 27ndash34
Sauer FG Pinkner JS Waksman G and Hultgren SJ(2002) Chaperone priming of pilus subunits facilitates atopological transition that drives fiber formation Cell 111543ndash551
Schembri MA Hasman H and Klemm P (2000)Expression and purification of the mannose recognitiondomain of the FimH adhesin FEMS Microbiol Lett 188147ndash151
Schembri MA Kjaergaard K and Klemm P (2003) Glo-bal gene expression in Escherichia coli biofilms Mol Micro-biol 48 253ndash267
Schilling JD Mulvey MA Vincent CD Lorenz RGand Hultgren SJ (2001) Bacterial invasion augments epi-thelial cytokine responses to Escherichia coli through alipopolysaccharide-dependent mechanism J Immunol166 1148ndash1155
Silhavy TJ Szmelcman S Boos W and Schwartz M(1975) On the significance of the retention of ligand byprotein Proc Natl Acad Sci USA 72 2120ndash2124
A novel class of FimH high-affinity ligands 455
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
Svanborg C and Godaly G (1997) Bacterial virulence inurinary tract infection Infect Dis Clin North Am 11 513ndash529
Thanassi DG Saulino ET and Hultgren SJ (1998) Thechaperoneusher pathway a major terminal branch of thegeneral secretory pathway Curr Opin Struct Biol 1 223ndash231
Thankavel K Madison B Ikeda T Malaviya R ShahAH Arumugen PM and Abraham SN (1997) Local-ization of a domain in the FimH adhesin of Escherichia colitype 1 fimbriae capable of receptor recognition and use ofa domain-specific antibody to confer protection againstexperimental urinary tract infection J Clin Invest 1001123ndash1126
Vagin A and Teplyakov A (1997) MOLREP an automatedprogram for molecular replacement J Appl Crystallogr 301022ndash1025
Wu XR Sun TT and Medina JJ (1996) In vitro bindingof type 1-fimbriated Escherichia coli to uroplakins 1a and
1b relation to urinary tract infections Proc Natl Acad SciUSA 93 9630ndash9635
Xia Y Gally D Forsman-Semb K and Uhlin BE (2000)Regulatory cross-talk between adhesin operons in Escher-ichia coli inhibition of type 1 fimbriae expression by thePapB protein EMBO J 19 1450ndash1457
Zafriri D Ofek I Adar R Pocino M and Sharon N(1989) Inhibitory activity of cranberry juice on adherenceof type-1 and type-P fimbriated Escherichia coli to eukary-otic cells Antimicrob Agents Chemother 33 92ndash98
Zavialov AV Berglund J Pudney AF Fooks LJ Ibra-him TM MacIntyre S and Knight SD (2003) Structureand biogenesis of the capsular F1 antigen from Yersiniapestis preserved folding energy drives fiber formation Cell113 587ndash596
Zhou G Mo WJ Sebbel P Min G Neubert TA Glock-shuber R et al (2001) Uroplakin Ia is the urothelial recep-tor for uropathogenic Escherichia coli evidence from invitro FimH binding J Cell Sci 114 4095ndash4103

454 J Bouckaert et al
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
Escherichia coli to oligosaccharides Bioorg Med Chem 112255ndash2261
Leffler H and Svanborg-Eden C (1980) Chemical identifi-cation of a glycosphingolipid receptor for Escherichia coliattaching to human urinary tract epithelial cells and agglu-tinating human erythrocytes FEMS Microbiol Lett 8 127ndash134
Lindhorst TK Kieburg C and Krallmann-Wenzel U(1998) Inhibition of the type 1 fimbriae-mediated adhesionof Escherichia coli to erythrocytes by multiantennary alpha-mannosyl clusters the effect of multivalency Glycoconj J15 605ndash613
Loris R Tielker D Jaeger KE and Wyns L (2003a)Structural basis of carbohydrate recognition by the lectinLecB from Pseudomonas aeruginosa J Mol Biol 331 861ndash870
Loris R Imberty A Beeckmans S Van Driessche ERead JS Bouckaert J et al (2003b) Crystal structureof Pterocarpus angolensis lectin in complex with glucosesucrose and turanose J Biol Chem 278 16297ndash16303
Lund B Lindberg F Marklund BI and Normark S(1987) The PapG protein is the a-D-galactopyranosyl-(1-4)-b-D-galactopyranose-binding adhesin of uropathogenicEscherichia coli Proc Natl Acad Sci USA 84 5898ndash5902
Min G Stolz M Zhou G Liang F Sebbel P StofflerD et al (2002) Localization of uroplakin Ia the urothelialreceptor for bacterial adhesin FimH on the six innerdomains of the 16 nm urothelial plaque particle J Mol Biol317 697ndash706
Miroux B and Walker JE (1996) Over-production of pro-teins in Escherichia coli mutant hosts that allow synthesisof some membrane proteins and globular proteins at highlevels J Mol Biol 260 289ndash298
Morris GM Goodsell DS Halliday RS Huey R HartWE Belew RK and Olson AJ (1998) Automateddocking using a Lamarckian genetic algorithm and anempirical binding free energy function J Comput Chem 191639ndash1662
Mulvey MA (2002) Adhesion and entry of uropathogenicEscherichia coli Cell Microbiol 4 257ndash271
Mulvey MA Lopez-Boado YS Wilson CL Roth RParks WC Heuser J and Hultgren SJ (1998) Induc-tion and evasion of host defenses by type 1-piliated uro-pathogenic Escherichia coli Science 282 1494ndash1497
Murshudov GN Vagin AA and Dodson EJ (1997)Refinement of macromolecular structures by the maxi-mum-likelihood method Acta Crystallogr D Biol Crystallogr53 240ndash255
Nagahori N Lee RT Nishimura S Pageacute D Roy Rand Lee YC (2002) Inhibition of adhesion of type 1 fim-briated Escherichia coli to highly mannosylated ligandsChembiochem 3 836ndash844
Navaza J (2001) Implementation of molecular replacementin AMoRe Acta Crystallogr D Biol Crystallogr 57 1367ndash1372
Neeser JR Koellreutter B and Wuersch P (1986)Oligomannoside-type glycopeptides inhibiting adhesion ofEscherichia coli strains mediated by type 1 pili preparationof potent inhibitors from plant glycoproteins Infect Immun52 428ndash436
Old DC (1972) Inhibition of the interaction between fimbrial
hemagglutinatinins and erythrocytes by D-mannose andother carbohydrates J Gen Microbiol 71 149ndash157
Orndorff PE Devapali A Palestrant S Wyse A EverettML Bollinger RR and Parker W (2004) Immunoglob-ulin-mediated agglutination of and biofilm formation byEscherichia coli K-12 require the type 1 pilus fiber InfectImmun 72 1929ndash1938
Oscarson S and Tedebark U (1995) Syntheses of deoxyanalogues of methyl 36-di-O-a-D-mannopyranosyl-a-D-mannopyranoside for studies of the binding site of con-canavalin A Carbohydr Res 278 271ndash287
Oscarson S and Tiden AK (1993) Syntheses of the octyland tetradecyl glycosides of 36-di-O-alpha-deuterium-mannopyranosyl-alpha-deuterium-mannopyranose and of34-di-O-alpha-deuterium-mannopyranosyl-alpha-deute-rium-mannopyranose ndash a new way for 24-di-O-protectionof mannopyranosides Carbohydr Res 247 323ndash328
Otwinowski Z and Minor W (1997) Processing of X-raydiffraction data collected in oscillation mode MethodsEnzymol 276 307ndash326
Powell HR (1999) The Rossmann Fourier autoindexingalgorithm in MOSFLM Acta Crystallogr D Biol Crystallogr55 1690ndash1695
Pratt LA and Kolter R (1998) Genetic analysis of Escher-ichia coli biofilm formation roles of flagella motility chemo-taxis and type I pili Mol Microbiol 30 285ndash293
Remaut H and Waksman G (2004) Structural biology ofbacterial pathogenesis Curr Opin Struct Biol 14 161ndash170
Ronald AR Nicolle LE Stamm E Krieger J WarrenJ Schaeffer A et al (2001) Urinary tract infection inadults research priorities and strategies Int J AntimicrobAgents 17 343ndash348
Sanner MF Olson AJ and Spehner JC (1996)Reduced surface an efficient way to compute molecularsurfaces Biopolymers 38 305ndash320
Sauer FG Futterer K Pinkner JS Dodson KW Hult-gren SJ and Waksman G (1999) Structural basis ofchaperone function and pilus biogenesis Science 2851058ndash1061
Sauer FG Knight SD Waksman GW and HultgrenSJ (2000) PapD-like chaperones and pilus biogenesisSemin Cell Dev Biol 11 27ndash34
Sauer FG Pinkner JS Waksman G and Hultgren SJ(2002) Chaperone priming of pilus subunits facilitates atopological transition that drives fiber formation Cell 111543ndash551
Schembri MA Hasman H and Klemm P (2000)Expression and purification of the mannose recognitiondomain of the FimH adhesin FEMS Microbiol Lett 188147ndash151
Schembri MA Kjaergaard K and Klemm P (2003) Glo-bal gene expression in Escherichia coli biofilms Mol Micro-biol 48 253ndash267
Schilling JD Mulvey MA Vincent CD Lorenz RGand Hultgren SJ (2001) Bacterial invasion augments epi-thelial cytokine responses to Escherichia coli through alipopolysaccharide-dependent mechanism J Immunol166 1148ndash1155
Silhavy TJ Szmelcman S Boos W and Schwartz M(1975) On the significance of the retention of ligand byprotein Proc Natl Acad Sci USA 72 2120ndash2124
A novel class of FimH high-affinity ligands 455
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
Svanborg C and Godaly G (1997) Bacterial virulence inurinary tract infection Infect Dis Clin North Am 11 513ndash529
Thanassi DG Saulino ET and Hultgren SJ (1998) Thechaperoneusher pathway a major terminal branch of thegeneral secretory pathway Curr Opin Struct Biol 1 223ndash231
Thankavel K Madison B Ikeda T Malaviya R ShahAH Arumugen PM and Abraham SN (1997) Local-ization of a domain in the FimH adhesin of Escherichia colitype 1 fimbriae capable of receptor recognition and use ofa domain-specific antibody to confer protection againstexperimental urinary tract infection J Clin Invest 1001123ndash1126
Vagin A and Teplyakov A (1997) MOLREP an automatedprogram for molecular replacement J Appl Crystallogr 301022ndash1025
Wu XR Sun TT and Medina JJ (1996) In vitro bindingof type 1-fimbriated Escherichia coli to uroplakins 1a and
1b relation to urinary tract infections Proc Natl Acad SciUSA 93 9630ndash9635
Xia Y Gally D Forsman-Semb K and Uhlin BE (2000)Regulatory cross-talk between adhesin operons in Escher-ichia coli inhibition of type 1 fimbriae expression by thePapB protein EMBO J 19 1450ndash1457
Zafriri D Ofek I Adar R Pocino M and Sharon N(1989) Inhibitory activity of cranberry juice on adherenceof type-1 and type-P fimbriated Escherichia coli to eukary-otic cells Antimicrob Agents Chemother 33 92ndash98
Zavialov AV Berglund J Pudney AF Fooks LJ Ibra-him TM MacIntyre S and Knight SD (2003) Structureand biogenesis of the capsular F1 antigen from Yersiniapestis preserved folding energy drives fiber formation Cell113 587ndash596
Zhou G Mo WJ Sebbel P Min G Neubert TA Glock-shuber R et al (2001) Uroplakin Ia is the urothelial recep-tor for uropathogenic Escherichia coli evidence from invitro FimH binding J Cell Sci 114 4095ndash4103

A novel class of FimH high-affinity ligands 455
copy 2004 Blackwell Publishing Ltd Molecular Microbiology 55 441ndash455
Svanborg C and Godaly G (1997) Bacterial virulence inurinary tract infection Infect Dis Clin North Am 11 513ndash529
Thanassi DG Saulino ET and Hultgren SJ (1998) Thechaperoneusher pathway a major terminal branch of thegeneral secretory pathway Curr Opin Struct Biol 1 223ndash231
Thankavel K Madison B Ikeda T Malaviya R ShahAH Arumugen PM and Abraham SN (1997) Local-ization of a domain in the FimH adhesin of Escherichia colitype 1 fimbriae capable of receptor recognition and use ofa domain-specific antibody to confer protection againstexperimental urinary tract infection J Clin Invest 1001123ndash1126
Vagin A and Teplyakov A (1997) MOLREP an automatedprogram for molecular replacement J Appl Crystallogr 301022ndash1025
Wu XR Sun TT and Medina JJ (1996) In vitro bindingof type 1-fimbriated Escherichia coli to uroplakins 1a and
1b relation to urinary tract infections Proc Natl Acad SciUSA 93 9630ndash9635
Xia Y Gally D Forsman-Semb K and Uhlin BE (2000)Regulatory cross-talk between adhesin operons in Escher-ichia coli inhibition of type 1 fimbriae expression by thePapB protein EMBO J 19 1450ndash1457
Zafriri D Ofek I Adar R Pocino M and Sharon N(1989) Inhibitory activity of cranberry juice on adherenceof type-1 and type-P fimbriated Escherichia coli to eukary-otic cells Antimicrob Agents Chemother 33 92ndash98
Zavialov AV Berglund J Pudney AF Fooks LJ Ibra-him TM MacIntyre S and Knight SD (2003) Structureand biogenesis of the capsular F1 antigen from Yersiniapestis preserved folding energy drives fiber formation Cell113 587ndash596
Zhou G Mo WJ Sebbel P Min G Neubert TA Glock-shuber R et al (2001) Uroplakin Ia is the urothelial recep-tor for uropathogenic Escherichia coli evidence from invitro FimH binding J Cell Sci 114 4095ndash4103