reconstructions of late ordovician crinoids and bryozoans
TRANSCRIPT

Reconstructions of Late Ordovician crinoids and bryozoans from the Decorah Shale, Upper Mississippi Valley
Sibo Wang Senior Integrative Exercise
March 10, 2010
Submitted in partial fulfillment of the requirements for a Bachelor of Arts degree from Carleton College, Northfield, Minnesota

TABLE OF CONTENTS ABSTRACT INTRODUCTION ........................................................................................................ 01 GEOLOGIC SETTING ................................................................................................ 03 Late Ordovician world ................................................................................. 03
Southern Minnesota and the Decorah Shale ............................................... 03 Benthic community ....................................................................................... 05 Marine conditions ........................................................................................ 05
CRINOIDS ................................................................................................................. 06 General background and fossil record ........................................................ 06
Anatomy ....................................................................................................... 07 Decorah Shale crinoids ................................................................................10
BRYOZOANS ............................................................................................................. 10 General background and fossil record ........................................................ 10 Anatomy ....................................................................................................... 11 Decorah Shale bryozoans ............................................................................ 13
METHODS ................................................................................................................. 13 Research area .............................................................................................. 13 Processing fossils ......................................................................................... 14
Photographing ............................................................................................. 14 Reconstructions ............................................................................................ 14 RESULTS AND RECONSTRUCTIONS .......................................................................... 16
Crinoid fossils .............................................................................................. 16 Crinoid reconstructions ............................................................................... 18 Bryozoan fossils ........................................................................................... 18 Bryozoan reconstructions ............................................................................ 21
DISCUSSION .............................................................................................................. 21 Crinoids ....................................................................................................... 21 Bryozoans ..................................................................................................... 24
Further studies ............................................................................................. 25 CONCLUSION ............................................................................................................ 26 ACKNOWLEDGEMENTS ............................................................................................ 26 REFERENCES CITED ................................................................................................ 27 APPENDICES ............................................................................................................. 31

Reconstructions of Late Ordovician crinoids and bryozoans from the Decorah Shale,
Upper Mississippi Valley
Sibo Wang Carleton College
Senior Integrative Exercise March 10, 2010
Advisors: Clinton A. Cowan, Carleton College Department of Geology Cameron Davidson, Carleton College Department of Geology
ABSTRACT Questions are prevalent among introductory geology students about the appearances and forms of the animals whose fossils are found in local formations, one of which is the Upper Ordovician Decorah Shale. The Decorah is highly fossiliferous, with abundant remains of bryozoans, crinoids, brachiopods, and other various animal groups that are often loose within the formation and thus easily collectable. Fossils from outcrops at Wang’s Corner, MN; Massey Quarry near Rochester, MN; and Country Road 9 Quarry, Decorah, IA were collected with the intention of using them as the bases for reconstructions, primarily focusing on crinoids and bryozoans. The aims of this study are to reconstruct several crinoid species (Carabocrinus dicyclicus and Pycnocrinus sardesoni) as well as both a general bryozoan colony and the animals within the colony (Stictoporellina cribrosa) to visually aid students in understanding the anatomy and form of these organisms. General background and anatomical information is also presented for crinoids and bryozoans. Keywords: Decorah Shale, reconstruction, Carabocrinus, Pycnocrinus, Stictoporellina, Crinoidea, Bryozoa

INTRODUCTION
One of the most prevalent uncertainties paleontologists face is the nature and
appearance of the organisms they study, both extinct and extant. The interest is not solely
confined to scientists, however; understanding what extinct animals look like has
interested people of all ages and backgrounds. It is therefore no surprise that
reconstructions have been a familiar sight in museum paintings and dioramas and other
media intended for public consumption for more than a century (Isham et al., 1989; Paul
& Chase, 1989). In the past 20 years, demand for reconstructions within the scientific
community itself has increased as well (Paul & Chase, 1989). Reconstructions of
hominoids and large vertebrates such as dinosaurs and extinct mammals have dominated
the public’s interest; however, reconstructions of invertebrates, though perhaps less
lucrative, are nevertheless both scientifically and educationally valuable. Reconstructions
of invertebrates have been useful in gaining an understanding of paleoenvironmental
conditions—mostly marine—(Brower and Veinus, 1978) and can also be used to provide
a more accessible pathway for the public to learn about a sometimes enigmatic group of
animals.
Many invertebrates are preferentially preserved in the rock record because of their
mineralized skeletons and the marine environment in which they live, and consequently
there is an abundance of specimens from which to work (Moore et al., 1952; Müller,
1979). Two groups of animals that have been focused on are the class Crinoidea and the
phylum Bryozoa (Ectoprocta). Although there are extant species in both groups,
questions still remain about their predecessors, particularly Paleozoic (and specifically
Ordovician) species (Boardman et al., 1983). Soft part preservation is rare, and, as is the
1

case for crinoids, whole specimen preservation is uncommon because their calcareous
endoskeletons often disarticulate after death (Sprinkle and Guensburg, 2004).
Many crinoid reconstructions have been attempted (Jaekel, 1918; Pisera and Dzik,
1979; Haude et al., 1994), although there are only a few reconstructions specific to
Ordovician species (Ubaghs, 1969; Brower and Veinus, 1978). Crinoids found in
Lagerstätten have naturally been focused on (Liddell, 1975; Brett & Liddell, 1978;
Ausich, 1999) but reconstructions of crinoids from their disarticulated parts are rarely
tackled (Brower and Veinus, 1978). Brower and Veinus (1978) did attempt such
reconstructions of crinoids from the Twin Cities area, although they are schematic at best.
Information on Ordovician bryozoans is more lacking than on Ordovician crinoids
(Boardman et al., 1983). Despite the fact that bryozoan species were diverse and
abundant in the Ordovician (Moore et al., 1952), little research has been done on them in
comparison to other phyla of the time, in part because they are not particularly useful in
biostratigraphy (Taylor and Ernst, 2004). Identification of bryozoans beyond order-level
is also difficult because it requires thin sections taken at specific angles through the
skeleton (Moore et al., 1952; Tayler and Ernst, 2004). Reconstructions of the individual
animals within a bryozoan colony are often generalized or schematic (Boardman et al.,
1983; Utgaard, 1983) because several bryozoan orders have no extant species (Moore et
al., 1952).
In this paper, I am attempting to reconstruct crinoid and bryozoan species from
the Upper Ordovician Decorah Shale. Using illustrations, I am aiming to reconstruct
whole specimens with their soft parts and in life position. The bases for the
reconstructions are fossils that have been collected from the Decorah Shale in addition to
2

graphical and descriptive information from published literature. The reconstructions are
done with the intention of providing a visual aid to undergraduate introductory geology
students about the organisms that have existed locally. A general overview of crinoid and
bryozoan anatomy is also presented as a resource for students.
GEOLOGIC SETTING
Late Ordovician world
During the Late Ordovician, Earth is thought to have experienced two eustatic
highstands, the first of which occurred during the Caradoc and was the largest in the
entirety of the Phanerozoic (Hallam, 1992). A high greenhouse state and high carbon
dioxide levels led to a warm climate (Berner, 2001; Barnes, 2004a), although it is
theorized that the Earth plunged into an ice age during the terminal Ordovician which
correlated with the end Ordovician extinction event (Berry and Boucot, 1973; Jablonski,
1991; Brenchley, 2004). Increased volcanism, possibly in conjunction with a Mid
Ordovician mantle superplume, also contributed to shaping the climate (Barnes et al.,
1996; Qing et al., 1998; Barnes, 2004b).
Southern Minnesota and the Decorah Shale
Much of the Laurentian craton (North America) was submerged during the
extensive Caradoc transgression (Barnes, 2004a), and what is now southern Minnesota
was inundated by the Hollandale Embayment, part of a larger epeiric sea (Simo et al.,
2003; Fig. 1). The Hollandale Embayment was bounded by the Transcontinental Arch to
the north and west and the Wisconsin Dome to the east. The paleolatitude of the
embayment was approximately 15°S, and thus, the climate was likely tropical to
subtropical (Simo et al., 2003).
3

Transcontinental A
rch
Wisconsin
Dome
15˚S
N
WC
MQ
CRD
Hollandale Embayment
100 km
Figure 1. Geologic map, showing Transcontinental Arch, Wisconsin Dome, and Hollandale Embayment (modified from Witzke, 1980). Gray denotes Precambrian rock, but is not necessarily above sea-level during the Late Ordovician. Paleolatitude of 15°S from Simo et al. (2003). Note direction of paleo-north. Study locations are also shown: WC: Wang’s Corner, MN; MQ: Massey Quarry, near Rochester, MN; CRD: County Road 9 Quarry, Decorah, IA.
4

The Decorah Shale was deposited in the Hollandale Embayment, which
comprised a large stretch of what is now the Upper Mississippi Valley (Simo et al., 2003).
Abundant volcanism led to the deposition of several K-bentonite layers within the
Decorah Shale (Sloan, 1988), which were used to date the formation’s deposition to
around 454 Ma (Gradstein et al., 2004; Sadler and Cooper, 2004; Ogg, 2009). The
Decorah crops out in many locations in southern Minnesota and northern Iowa (Sloan et
al., 1988). Moving southeast across the formation, there is a decrease in shale content and
an increase in carbonates, and in southern Minnesota, most of the formation is composed
of fossiliferous shale with carbonate lenses and some beds of limestone (Mossler, 1987).
Benthic community
Brachiopods and bryozoans are abundant throughout the Decorah Shale, and it is
likely they were the primary constituents of the benthic fauna (Mossler, 1987). Crinoids
can be found in abundance in several layers, though usually in the form of disarticulated
ossicles (Brower and Veinus, 1978). Also present, but in less abundance, are gastropods,
pelecypods, cephalopods, rugose corals, receptaculities, and trilobites (Mossler and
Benson, 1999).
Marine conditions
The fossils found throughout the Decorah belong to the Heterozoan Association
proposed by James (1998). Although a Heterozoan assemblage indicates the possibility of
cool-water conditions, it does not exclusively signify such an environment (James, 1998).
There are a number of different variables that could have contributed to the found
assemblage; for instance, salinity, nutrient levels, and upwelling from colder polar waters
could all have affected the fauna, and it is uncertain how large a role each factor played
5

(James, 1998). Worldwide climate conditions and the paleolatitude of the area favor
warmer marine conditions, but neither are definitive indicators either (Simo et al., 2003).
Brower and Veinus (1978) were able to infer that the benthic fauna of the
Decorah Shale experienced strong but intermittent agitation from currents as well as
periods of quiet water. The substrate was likely unstable and not well-lithified, but also
rough and hummocky (Brower and Veinus, 1978). They also concluded that the
environment had normal marine salinity; however Barnes (2004a) argues instead for
raised salinity due to the warm climate and high evaporation rates to which fauna slowly
adapted.
CRINOIDS
General background and fossil record
Crinoids belong to the class Crinoidea of the phylum Echinodermata and, like
most other organisms within the phylum, exhibit some form of pentameral radial
symmetry when mature (Moore et al., 1952; Ubachs, 1967). They are entirely marine,
mostly sessile or slow-moving, and unlike other echinoderms, filter food from the water
column rather than from the substrate (Hess and Ausich, 1999). As larvae they are free-
floating and typically exhibit bilateral symmetry (Ubachs, 1967). Crinoid skeletons are
composed of high-Mg calcite (Sprinkle and Guensburg, 2004), and crinoids were
important reef-builders from the Ordovician to the Jurassic, with many of their skeletal
parts being found in limestones worldwide (Moore and Teichert, 1978).
Some argue that a basal crinoid existed in the Cambrian (Sprinkle and Moore,
1978); this is, however, repeatedly disputed by Ausich and Babcock (1998; 2000), who
believe the specimen is more probably an octocoral. The first unequivocal fossil crinoid
6

is from the middle Early Ordovician (Ausich and Babcock, 1998; Guensburg and
Sprinkle, 2001). Nevertheless, it is thought that crinoids first evolved during the Middle
to Late Cambrian, despite the lack of fossil evidence (Guensburg and Sprinkle, 2001).
Within the Ordovician, crinoids experienced the most species-level diversity during the
Mohawkian/Caradoc Age of the Late Ordovician, rising to become the most prominent
class of echinoderms during this time (Sprinkle and Guensburg, 2004).
Anatomy
Crinoids are composed of two basic sections: the pelma (the stem and holdfast, if
any), and the crown or corona (which includes the calyx and free arms, if any) (Ubaghs,
1978). The crinoid skeleton is made up of individual ossicles, which can refer to any
segment or plate of the skeleton (Moore et al., 1952). Ossicles in the stem are called
columnals, whereas ossicles in the arms are known as brachials (Moore et al., 1952).
Moore et al. (1952) has an excellent introduction to and explanation of basic anatomy and
anatomical terminology.
Stem: The stem serves a dual purpose of anchoring the crinoid to the substrate as
well as elevating the crinoid above the seafloor so that it has a better position to filter
food from the water (Ausich et al., 1999; Hess and Ausich, 1999). The stem is composed
of numerous columnals connected to each other with ligaments and other living tissue,
and can take on a wide range of forms, varying in shape, length, and diameter depending
on the environment to which the crinoid is adapted (Ausich et al., 1999). Some crinoid
species have specialized holdfasts (Ausich et al., 1999), whereas in other species, the
distal part of the stem serves as a makeshift holdfast, curling around other objects on the
seafloor or itself propping the crinoid upright (Liddell, 1975).
7

Calyx: The calyx (or cup) is generally bulb-shaped and is composed of plates that
sit atop the stem and house the crinoid’s internal organs (Hess and Ausich, 1999).
Calyxes can have several different plate configurations depending on order, family, etc.,
and the treatment of specific configurations is beyond the scope of this paper (see Moore
et al., 1952; Moore and Teichert, 1978; Simms, 1999).
In some species, the cup comprises the calyx and the entirety of the arms, as
plates connect all the appendages together; in other species, the cup is composed solely of
the calyx and the entire extent of the arms are free-moving (Ausich et al., 1999).
Arms: Crinoid arms comprise brachials that are connected by ligaments and other
tissue (Ausich et al., 1999). The primary function of the arms is to filter out food from the
water column (Ausich et al., 1999). Each arm ray can branch in a variety of ways (see Fig.
2). Arm sections between branch nodes are known as brachs (Moore et al., 1952). Arms
may or may not possess pinnules, which are small branches on the arms intended to
improve filtration (Ausich et al., 1999).
Soft Parts: Crinoids are composed mostly of skeleton, with little living tissue in
comparison (Ausich et al., 1999). The stem is not covered by skin; however the
columnals have numerous microscopic holes that are filled with tissue that excrete the
calcium carbonate skeleton. The tegmen that covers the calyx is composed of leathery,
tough skin, small calcareous plates, or a mixture of both (Moore et al., 1952). Towards
the center of the tegmen is the mouth opening, which is connected to a digestive tract
comprising a feeding tube, stomach, intestines and anus (Moore et al., 1952). The anal
opening is usually located on the side of the tegmen, sometimes on a very large anal tube
(Moore et al., 1952). A water-vascular system runs through the arms and ends in the tube
8

Figure 2. Two possible arm branching forms. Sections between branching points are known as brachs. The entire arm with branches is known as an arm ray. Modified from Moore et al., 1952.
9

feet that are located in grooves within the arm brachials (Moore et al., 1952; Ausich et al.,
1999). The tube feet possess cilia, which are used both to move food down to the mouth
as well as diffuse oxygen into the crinoid (Ausich et al., 1999). Crinoids possess an
extensive nervous system that also runs throughout the animal, though the nerves are
concentrated in the arms and calyx (Moore et al., 1952; Ausich et al., 1999).
Decorah Shale crinoids
Crinoid fossils are moderately abundant throughout the Decorah Shale, which
contains fourteen known species and six types of holdfasts in the Twin Cities area
(Brower and Veinus, 1978; Appendix 1). Information about crinoids within the Decorah
in areas other than the Twin Cities is relatively unknown. In the Twin Cities area, two
subclasses are represented within the Decorah: the Camerata and the Inadunata (Brower
and Veinus, 1978). Disarticulated columnals, short stem fragments, and plate fragments
are the most commonly crinoid fossils found (Brower and Veinus, 1978). Intact calyxes
and arm segments are rare (Brower and Veinus, 1978).
BRYOZOANS
General background and fossil record
In this paper, bryozoans will refer to Ectoprocta only, and Entoprocta will not be
classified within the phylum (cf. Tudge, 2000). Bryozoans have been an enigmatic
phylum throughout history, and it was only recently confirmed using nucleotide data that
the phylum was truly monophyletic (Fuchs et al., 2009). Although bryozoans appear in
the fossil record during the Early Ordovician, it is probable that, like crinoids, more
primitive and basal forms evolved prior to the Ordovician (Fuchs et al., 2009). Bryozoans
10

are unique in that they are the only phylum in which all species are colonial organisms
(Boardman et al., 1983).
Bryozoans reproduce both sexually and asexually (Moore et al., 1952). New
colonies are formed from the sexually-produced larva of old colonies, and when a larva
settles onto a hard surface, it matures into the ancestrula, or the first adult individual of a
colony (Moore et al., 1952; Boardman et al., 1983). All subsequent members of that
colony are clones of the ancestrula and created through asexual budding (Moore et al,
1952; Boardman et al., 1983). Unlike crinoids, bryozoan species have an exoskeleton,
though some species do not excrete a mineralized skeleton (Boardman et al., 1983).
Bryozoans are also filter feeders, and use a lophophore to sift food particles from the
water (Moore et al., 1952).
Anatomy
Zooid: The individual animal within a bryozoan colony is called the zooid.
Zooids are not usually larger than 1 mm in length, and are often smaller (Boardman et al.,
1983; McKinney and Jackson, 1989). Basic zooids are called autozooids, and perform
feeding and sometimes reproductive functions (McKinney and Jackson, 1989). The
simplified anatomy of autozooids comprises a lophophore that is composed of tentacles
and encircles a mouth, an esophagus, stomach and anus (Moore et al., 1952; Boardman et
al., 1983; McKinney and Jackson, 1989). The digestive tract is recurved, with the anus
located by the mouth (Moore et al., 1952; Boardman et al., 1983). Zooids can become
highly specialized forms, known as heterozooids, and can function in several different
roles (e.g. reproduction, cleaning, protection) (Boardman et al., 1983; McKinney and
Jackson, 1989). In species that secrete calcium carbonate, zooids are enclosed by an
11
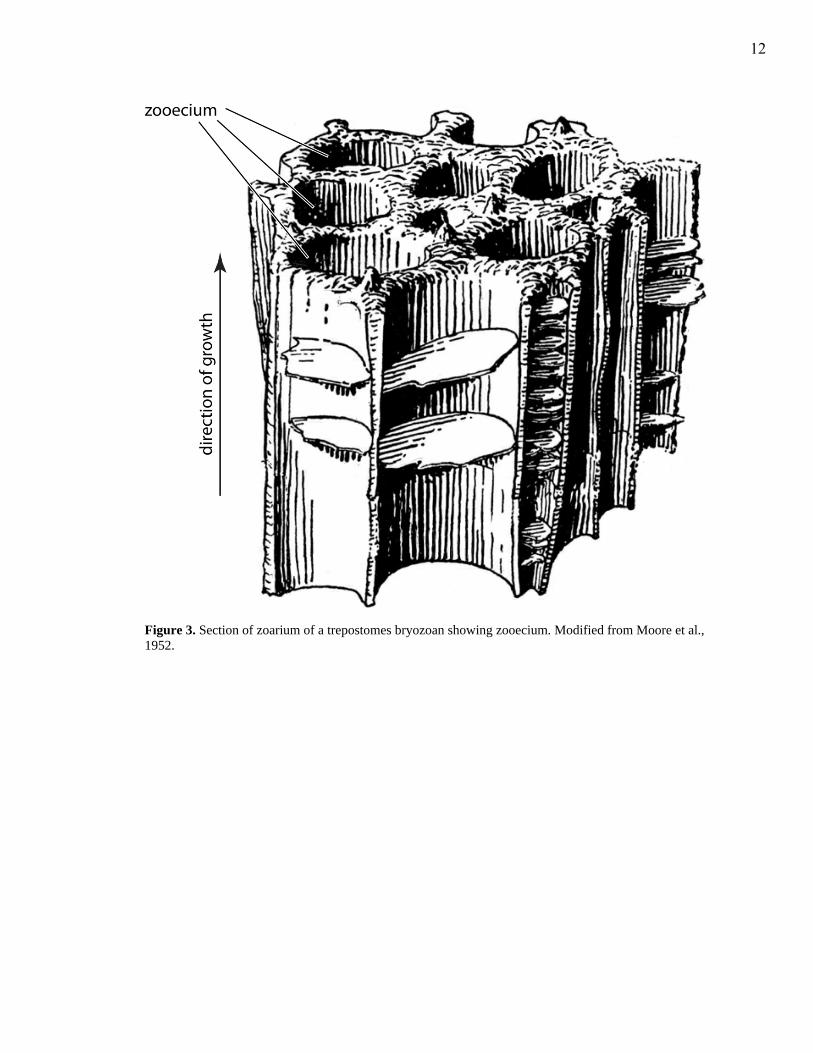
Figure 3. Section of zoarium of a trepostomes bryozoan showing zooecium. Modified from Moore et al., 1952.
12

individual skeleton, called the zooecium (Fig. 3; McKinney and Jackson, 1989).
Colony: The skeleton of the entire colony is known as the zoarium (Moore et al,
1952; Boardman et al., 1983). Colonies can be encrusting, erect, free-living, or rooted,
and several forms have been preferentially evolved throughout geologic time by different
species in what is known as iterative evolution (McKinney and Jackson, 1989).
McKinney and Jackson (1989) determined five basic growth forms that most colonies
take on. Colony forms can be determined by species, environment, or a combination of
both (Boardman et al., 1983).
Decorah Shale Bryozoans
Bryozoan fossils are abundant throughout the Decorah Shale (Karklins, 1988).
The bryozoan fossils found in the Decorah are primarily of the orders Trepostomata (32
species) and Cryptostomata (18 species); bryozoan fossils of the order Cystoporata are
rare and represented by one species, Bythotrypa laxata (Karklins, 1988; Appendix 2).
Within the trepostomes, genera Homotrypa, Heterotrypa, Batostoma, Parvohallopora,
and Eridotrypa are the most prevalent; within the cryptostomes, the genera that are most
common are Stictopora, Trigonodictya, and Pseudostictoporella (Karklins, 1988).
METHODS
Research area
Fossils were collected in the fall of 2009 during several field trips for the
Sedimentology and Stratigraphy course at Carleton College, as well as on independent
return outings to Wang’s Corner, MN; Massey Quarry near Rochester, MN; and County
Road 9 Quarry in Decorah, IA (Fig. 1). In general, fossils were removed from 1)
weathered shale, 2) poorly-lithified shale, or 3) surface to subsurface soil, depending on
13

location; no large scale excavations were attempted and no formal sampling methods
were used.
Processing fossils
Crinoid and bryozoan fossils were separated from other irrelevant fossils and
organized by location. The fossils were initially cleaned with water spray to remove large
amounts of shaley material, soil, and other debris. They were then more thoroughly
cleaned with a Bransonic UL-452E ultrasonic cleaner. Depending on their size, fossils
were placed into either a 50 ml or 250 ml Pyrex beaker with soap and tap water and then
placed in the cleaner between 30 and 60 seconds. The cleaner itself was filled
approximately three-quarters full with water, although the water level was adjusted
depending on the size of the beaker being used. Fossils were completely immersed in
solution if possible. Some small scale iron oxidation occurred after cleaning, but did not
significantly obscure important anatomical details. Although not used in this study,
cleaning with distilled water is recommended.
Photographing
Representative fossils were photographed to show the assemblages present at each
location using a Panasonic Lumix DMC-LZ7 camera. A Nikon SMZ-10 stereozoom
microscope was used to magnify the details on fossils to aid in the reconstructions. A
Nikon Coolpix 990 digital camera was used in conjunction with the microscope in order
to further zoom in on fossil details for the reconstructions.
Reconstructions
Reconstructions were created using Adobe Photoshop CS4 Extended with a
Wacom Intuos2 6x8 graphics tablet. Illustrations, photographs, and reconstructions from
14

Figure 4. Crinoid fossil assemblages. A. Assemblage of Wang’s Corner, MN; 1-7, 9-11, Pycnocrinus sp.?; 8, stem of unknown crinoid; 12, 13, Isotomocrinus tenuis? Cincinnaticrinus sp.?; 14-29, 31-33, individual columnals of several Pycnocrinus sp.? 30, Iocrinus sp.? B. Assemblage of Massey Quarry, near Rochester, MN; 1-5, Pycnocrinus sp.; 6, 8, Pycnocrinus sp.?; 7, 9, 10, Carabocrinus sp.?
15

the literature were used in addition to the fossils collected to create the most accurate
reconstructions possible.
RESULTS AND RECONSTRUCTIONS
Crinoid fossils
No whole specimens were found in the field; most crinoid fossils were in the form
of fossil debris (Fig. 4). Additionally, no recognizable crinoid calyxes or arm brachials
were found. However, numerous columnals were collected, as well as a few stem
fragments that had not disarticulated.
Crinoid fossils found at Wang’s Corner were mostly in the form of disarticulated
columnals or short (<2 cm) stem sections (Fig. 4A). The largest individual columnal had
a diameter of ~11 mm, while the smallest columnal section had a diameter of ~2 mm.
Most of the crinoid fossils that were found likely comprised the genus Pycnocrinus. Two
stem fragments appeared similar to an Isotomocrinus tenuis identified by Brower and
Veinus (1978), although they admitted that the identification was tentative, at best.
However, the stem fragments as well as the specimen identified by Brower and Veinus
(1978) bear a striking resemblance to a Cincinnaticrinus sp. found in an approximately
coeval formation from New York (Brower, 2005). An individual columnal (30) was also
found that is tentatively being identified here as Iocrinus sp. (Fig. 4A; Meyer et al., 2002);
its occurrence in the Decorah has never previously been mentioned.
Much longer columnal sections were found at Massey Quarry, with the longest
being about 4.1 cm (Fig. 4B). The crinoid fossils found likely all belonged to either the
genus Pycnocrinus or the genus Carabocrinus. No crinoid fossils were found at the
County Road 9 Quarry; this was, however, the least surveyed location, and thus, lack of
16

Figure 5. Reconstruction of the crinoid Carobocrinus dicyclicus. Note lack of pinnules and short stem. Calyx is approximately 4 cm in height.
17

recovered crinoid fossils should not be an indication that there were no crinoids present.
Crinoid reconstructions
Carabocrinus dicyclicus was reconstructed primarily using photographs by
Brower and Veinus (1978; Fig. 5). The crinoid is illustrated as having three primibrachs;
arms branch a maximum of five times, both isotomously and heterotomously (Fig. 2).
Tube feet, or the feeding apparatuses, are shown on the most distal arm from the
foreground in orange. The crinoid is shown cemented directly to the substrate with a
basal plate.
Pycnocrinus sardesoni was also reconstructed primarily using photographs by
Brower and Veinus (1978; Fig. 6) as well as using a Pycnocrinus dyeri reconstruction by
Ausich (1999). The distal stem and holdfast are excluded.
Bryozoan fossils
Many bryozoans of the orders Trepostomata and Cryptostomata were collected
from each field site (Fig. 7). At Wang’s Corner, two Prasopora insularis were found
cemented onto on mass composed of shale and fossil debris, with one colony growing on
top of the other (Fig. 7A). Two other Prasopora fossils were found, one being identified
as Prasopora simulatrix, and the other too small to identify the species. Several bryozoan
fossils were likely Batostoma sp., and one (6) was Stictopora sp.
Fossils recovered from Massey Quarry were largely of the genus Batostoma, with
two Stictoporellina cribosa fossils, two Stictopora sp. fossils, and a Hallopora
multitabulata that was still cemented to other fossil debris (Fig. 7B). Several fossils were
unable to be identified, largely due to their small size and incomplete nature. Bryozoans
collected from the County Road 9 Quarry were mostly from the genus Prasopora, with
18
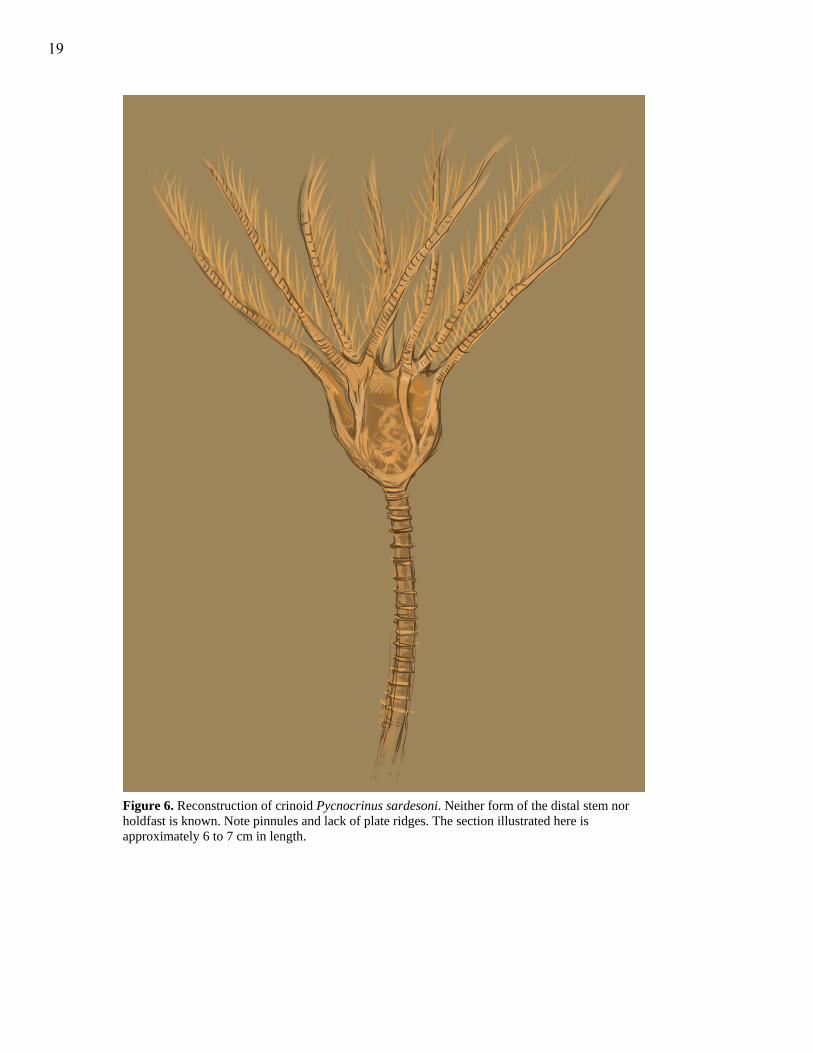
Figure 6. Reconstruction of crinoid Pycnocrinus sardesoni. Neither form of the distal stem nor holdfast is known. Note pinnules and lack of plate ridges. The section illustrated here is approximately 6 to 7 cm in length.
19

Figu
re 7
. Bry
ozoa
n fo
ssil
asse
mbl
ages
. A. A
ssem
blag
e of
Wan
g’s C
orne
r, M
N; 1
, Pra
sopo
ra in
sula
ris;
2-5
, 8, B
atos
tom
a sp
.?; 6
, Stic
topo
ra sp
.; 7,
uni
dent
ified
br
yozo
an; 9
, Pra
sopo
ra si
mul
atri
x; 1
0, P
raso
pora
sp. B
. Ass
embl
age
of M
asse
y Q
uarr
y, n
ear R
oche
ster
, MN
; 1, H
allo
pora
(Tar
phop
hrag
ma)
mul
titab
ulat
a; 2
-9,
Bat
osto
ma
sp.;
10-1
3, u
nide
ntifi
ed b
ryoz
oan;
15,
Stic
topo
ra sp
.; 14
,16,
Stic
topo
relli
na c
ribr
osa.
C. A
ssem
blag
e of
Cou
nty
Roa
d 9
Qua
rry,
Dec
orah
, IA
; 1-1
1,
13-1
5, P
raso
pora
sp.;
12, B
atos
tom
a sp
.? 2
-5a,
9-1
1a, t
op v
iew
of b
ryoz
oans
; 2-5
b, 9
-11b
, bot
tom
vie
w o
f bry
ozoa
ns.
20

one fossil tentatively placed in the genus Batostoma (Fig. 7C).
Bryozoan reconstructions
A Stictoporellina cribrosa colony was reconstructed (Fig. 8) based on the fossil
fragments found at Massey Quarry in addition to illustrations by Sardeson (1901) and
models of erect fan growth (McKinney and Jackson, 1989).
Stictoporellina cribrosa autozooids were illustrated within the living chambers of
the zoarium (Fig. 9). Autozooid forms are schematic. Zooecium forms were based off of
thin section photographs (Karklins, 1983) and an illustration by Sardeson (1901).
Schematic autozooid form based off of general zooid illustrations (Boardman et al., 1983;
Utgaard, 1983). Autozooid anatomical features that are illustrated are the tentacles of the
lophophore, mouth, cardia, caecum, pylorus, intestine, and anus.
DISCUSSION
It should be noted that the colors used in the reconstructions are pure conjecture,
and used only to enhance a more realistic view of specimens. Because the species being
reconstructed are extinct, it is inevitable that more imagination and speculation are
involved than is optimal.
Crinoids
A full crinoid reconstruction was difficult without an entire specimen; calyx and
arm configuration and appearance were essential to understanding the basic form, and
because no calyxes or arm segments were recovered, crinoid species were chosen based
on the amount of graphical information present in the literature. Brower and Veinus
(1978) had the most comprehensive information on Decorah Shale crinoids, and thus, the
species best represented in their compilation were chosen for reconstruction (i.e.,
21

Figu
re 8
. Rec
onst
ruct
ion
of th
e br
yozo
an S
ticto
pore
llina
cri
bros
a. C
olon
y is
app
roxi
mat
ely
15 c
m w
ide.
22

Figure 9. Generalized reconstruction of Stictoporellina cribrosa autozooids w
ithin the living chambers of
the zoarium. C
ross-section view of colony perpendicular to plane of zoarium
. Zooids are approximately 1
mm
long.
23

Carabocrinus dicyclicus and Pycnocrinus sardesoni).
Carabocrinus dicyclicus (Fig. 5) is illustrated as having a very short stem due to
the large bulk of the calyx and the fact that strong agitation from currents would more
easily detach the calyx from a longer stem (Brower and Veinus, 1978). The tegmen is
only partially known, and thus not very well detailed in the reconstruction (Brower and
Veinus, 1978). The species only has two to three primibrachs and relatively short arms
(Brower and Veinus, 1978). Total arm length is likely a little too long in the
reconstruction; in adults, the calyx is approximately 4 cm tall, while the arms are
approximately 5 cm in length (Brower and Veinus, 1978). The holdfast is disk-shaped,
but its structure is hard to make out in the photographs of the fossil counterparts by
Brower and Veinus (1978), and so the holdfast is not very well articulated in the
reconstruction.
Pycnocrinus sardesoni (Fig. 6) is depicted without the distal stem segment or the
holdfast because the forms are unknown (Brower and Veinus, 1978). However, it is likely
that the species did not have a holdfast, instead using the distal stem to curl around
objects, such as branching bryozoan species, to anchor itself to the substrate, much like
Pycnocrinus dyeri (Ausich, 1999). Arms are pinnulated, and the plates of the calyx lack
plate margins (Brower and Veinus, 1978).
Bryozoans
When possible, I tried to identify the bryozoan fossils at the genus or species level.
Zoarium forms were relatively species-specific, although generalized forms are relatively
known at the genus level. The zoarium of many species is more or less preserved, while
the forms of the zooids are unknown, and thus, zooid reconstructions are more
24

scientifically interesting and useful.
Stictoporellina cribrosa was chosen for reconstruction because of the relative
straightforwardness of the zoarium form. Only small sections of the zoarium were found
in the field (Fig. 7B), and so the fossils were used to extrapolate the form of the entire
zoarium.
The reconstruction of the zooid of Stictoporellina cribrosa (Fig. 9) was difficult
because the forms of Ordovician zooids are almost completely unknown. Twitchell (1934)
even argues that the Trepostomata, an extinct order of Bryozoa, should be reassigned to
the Entoprocta, and though today it is widely accepted that trepostomes are Ectoprocta
(Boardman et al.), it illustrates the uncertainty of soft part anatomy of extinct bryozoan
orders. Therefore, a generalized and schematic reconstruction was attempted, more to
show zooid placement within the zoarium than for anatomical accuracy. Thin sections of
longitudinal sections of the zoarium were used as a guide for zooecium form (Karklins,
1983), as well as modeled growth forms of fan-shaped bryozoans (McKinney and
Jackson, 1989).
Further studies
In order to aid in more accurate reconstructions, detailed observations of pertinent
crinoid and bryozoan collections at museums, universities, and colleges should be
completed. More thorough and systematic collections of fossils from the field should be
also implemented, especially at different times during a week or month, as weather had a
significant effect on fossil-collecting conditions. Reconstructions of other bryozoan and
crinoid species of the Decorah Shale should also be illustrated because there was a
relatively high diversity of species within the Decorah, and knowing complete forms
25

would be educationally useful. A reconstruction of the benthic community would also be
valuable, especially to illustrate how the benthic fauna interacted with one another.
CONCLUSIONS
Undergraduate introductory geology students often come away from fossiliferous
outcrop excursions with questions about the form and living appearance of the fossilized
animals that they discover. This study was completed with the intention of providing a
basic introduction into two groups of animals that are found in the Upper Ordovician
Decorah Shale, the Crinoidea and the Bryozoa. A general overview of crinoid and
bryozoan anatomy is discussed with the hopes of providing a useful introductory resource
for students. Fossils were collected from three outcrops of the Decorah and used in
conjunction with published literature to reconstruct the likely forms of 1) the crinoids
Carabocrinus sardesoni and Pycnocrinus dicyclicus and 2) the bryozoan Stictoporellina
cribrosa colony and zooids as they appeared when living. There was some difficulty
encountered while doing the reconstructions, and collected fossils from the field were
less useful than previously anticipated. Although it is inevitable that certain aspects of the
reconstructions are the product of speculation and imagination, the intended goal in
creating the reconstructions is to provide a visual aid to students to further their
understanding, and I hope that this has been accomplished.
ACKNOWLEDGEMENTS
I would like to thank my advisors Clint Cowan and Cam Davidson—Clint, for
inspiring my interest in paleobiology, providing me with ideas for this paper, and
supplying invaluable resources; and Cam, who gave incredibly useful advice, critical
commentary, and encouragement. I would also like to extend my gratitude to the Carleton
26

College Geology Department for use of their resources; to Stuart Sweeney Smith for
doing several edits, offering general advice, and providing emotional support; and to all
my fellow geology senior majors for their encouragement and support throughout the
process.
REFERENCES CITED Ausich, W.I., 1999, Upper Ordovician of the Cincinnati, Ohio, area, USA, in Hess, H.,
ed., Fossil crinoids: New York, New York, Cambridge University Press, p. 75–80. Ausich, W.I., and Babcock, L.E., 1998, The phylogenetic position of Echmatocrinus
brachiatus, a probable octocoral from the Burgess Shale: Palaeontology, v. 41, p. 193–202.
Ausich, W.I., and Babcock, L.E., 2000, Echmatocrinus, a Burgess Shale animal reconsidered: Lethaia, v. 33, p. 92–94.
Ausich, W.I., and Kammer, T.W., 2001, The study of crinoids during the 20th century and the challenges of the 21st century: Journal of Paleontology, v. 75, no. 6, p. 1161–1173.
Ausich, W.I., Brett, C.E., Hess, H., and Simms, M.J., 1999, Crinoid form and function, in Hess, H., ed., Fossil crinoids: New York, New York, Cambridge University Press, p. 3–30.
Barnes, C.R., 2004a, Ordovician oceans and climate, in Webby, B.D., Paris, F., Droser, M.L., and Percival, I.G., eds., The Great Ordovician Biodiversification Event: New York, New York, Columbia University Press, p.72–76.
Barnes, C.R., 2004b, Was there an Ordovician superplume event?, in Webby, B.D., Paris, F., Droser, M.L., and Percival, I.G., eds., The Great Ordovician Biodiversification Event: New York, New York, Columbia University Press, p. 77–80.
Barnes, C.R, Fortey, R.A., and Williams, S.H., 1996, The pattern of global bio-events during the Ordovician Period, in Walliser, O.H., ed., Global events and event stratigraphy in the Phanerozoic: New York, New York, Springer-Verlag, p. 139–172.
Berry, W.B.N., and Boucot, A.J., 1973, Glacio-eustatic control of late Ordovician-early Silurian platform sedimentation and faunal changes: Geological Society of America Bulletin, v. 84, p. 275–284.
Boardman, R.S., Cheetham, A.H., and Cook, P.L., Introduction to the Bryozoa, in Robison, R.A., ed., Treatise on invertebrate paleontology, Part G, Bryozoa, revised, Volume 1: Boulder, Colorado, Geological Society of America, and Lawrence, University of Kansas, p. 3–48.
Brenchley, P.J., 2004, End Ordovician glaciation, in Webby, B.D., Paris, F., Droser, M.L., and Percival, I.G., eds., The Great Ordovician Biodiversification Event: New York, New York, Columbia University Press, p. 81–83.
Brett, C.E., and Liddell, W.D., 1978, Preservation and paleoecology of a Middle Ordovician hardground community: Paleobiology, v. 4, p. 329–348.
27

Brower, J.C., 2005, The paleobiology and ontogeny of Cincinnaticrinus varibrachialus Warn and Strimple, 1977 from the Middle Ordovician (Shermanian) Walcott-Rust Quarry of New York: Journal of Paleontology, v. 79, no. 1, p. 152–174.
Brower, J.C., and Veinus, J., 1978, Middle Ordovician crinoids from the Twin Cities area of Minnesota: Bulletins of American Paleontology, v.74, no. 304, p. 372–506.
Fuchs, J., Obst, M., and Sundberg, P., 2009, The first comprehensive molecular phylogeny of Bryozoa (Ectoprocta) based on combined analyses of nuclear and mitochondrial genes: Molecular Phylogenetics and Evolution, v. 52, p. 225–233.
Gradstein, F.M., Ogg, J.G., and Smith, A.G., 2004, A Geologic Time Scale 2004: New York, New York, Cambridge University Press, 589 p.
Guensburg, T.E., and Sprinkle, J., 2001, Earliest crinoids: New evidence for the origin of the dominant Paleozoic echinoderms: Geology, v. 29, no. 2, p. 131–134.
Hallam, A., 1992, Phanerozoic sea-level changes: New York, New York, Columbia University Press, 266 p.
Haude, R., Jahnke, H., and Walliser, O.H., 1994, Scyphocrinoïden an der Wende Silur/Devon: Aufschluss, v. 45, no. 2, p. 49–55.
Hess, H., and Ausich, W.I., 1999, Introduction, in Hess, H., ed., Fossil crinoids: New York, New York, Cambridge University Press, p. xiii–xv.
Isham, L.B., Klitz, K., and Hueber, F.M., 1989, Illustrating fossils, in Hodges, E.R.S., ed., The Guild Handbook of Scientific Illustration: New York, New York, Van Nostrand Reinhold, p. 221–223.
Jablonski, D., 1991, Extinctions: A paleontological perspective: Science, v. 253, no. 5021, p. 754–757.
Jaekel, O., 1918, Phylogenie und System der Pelmatozoen: Paläontologische Zeitschrift, v. 3, p. 1–128.
James, N.P., 1997, The cool-water carbonate depositional realm, in James, N.P., and Clarke, J.A.D., eds., Cool-Water Carbonates: Society for Sedimentary Geology Special Publication no. 56: Tulsa, Oklahoma, Society for Sedimentary Geology, p. 1–20.
Karklins, O.L., 1983, Systematic descriptions for the suborder Ptilodictyina, in Robison, R.A., ed., Treatise on invertebrate paleontology, Part G, Bryozoa, revised, Volume 1: Boulder, Colorado, Geological Society of America, and Lawrence, University of Kansas, p. 489–529.
Karklins, O.L., 1988, Bryozoa from Rocklandian (Middle Ordovician) rocks of the Upper Mississippi Valley Region, in Middle and Late Ordovician lithostratigraphy and biostratigraphy of the Upper Mississippi Valley: Minnesota Geological Survey Report of investigations, p. 173–176.
Liddell, W.D., 1975, Ecology and biostratinomy of a Middle Ordovician echinoderm assemblage from Kirkfield, Ontario: unpublished M.A. thesis, University of Michigan, Ann Arbor, 44 p.
McKinney, F.K., and Jackson, J.B.C., 1989, Bryozoan Evolution: Boston, Massachusetts, Unwin Hyman, 238 p.
Meyer, D.L., Miller, A.I., Holland, S.M., and Dattilo, B.F., 2002, Crinoid distribution and feeding morphology through a depositional sequence: Kope and Fairview Formations, Upper Ordovician, Cincinnati Arch Region: Journal of Paleontology, v. 74, no. 4, p. 725–732.
28

Moore, R.C., Lalicker, C.G., and Fischer, A.G., 1952, Invertebrate Fossils: New York, New York, McGraw-Hill Book Company, 766 p.
Moore, R.C., and Teichert, C., 1978, Introduction, in Moore, R.C., and Teichert, C., eds., Treatise on invertebrate paleontology, Part T, Echinodermata 2, Volume 1: Boulder, Colorado, Geological Society of America, and Lawrence, University of Kansas, p. T7–T9.
Moore, R.C., and Teichert, C., 1978, Treatise on invertebrate paleontology, Part T, Echinodermata: Boulder, Colorado, Geological Society of America, and Lawrence, University of Kansas, 1027 p.
Mossler, J.H., 1987, Paleozoic lithostratigraphic nomenclature for Minnesota: Minnesota Geological Survey Report of Investigations 36, 36 p.
Mossler, J.H., and Benson, S., 1999, Fossil collecting in the Twin Cities area: Minnesota Geology Survey Minnesota at a Glance, 4 p.
Müller, A.H., 1979, Fossilization (taphonomy), in Robison, R.A., and Teichert, C., eds., Treatise on invertebrate paleontology, Part A, Introduction: Boulder, Colorado, Geological Society of America, and Lawrence, University of Kansas, p. A2–A78.
Ogg, J.G., 2009, International Stratigraphic Chart, International Commission on Stratigraphy, retrieved from http://www.stratigraphy.org/upload/ISChart2009.pdf, accessed on 19 Feb 2010.
Paul, G.S., and Chase, T.L., 1989, Reconstructing extinct vertebrates, in Hodges, E.R.S., ed., The guild handbook of scientific illustration: New York, New York, Van Nostrand Reinhold, p. 239–256.
Pisera, A., and Dzik, J., 1979, Tithonian crinoids from Rogoźnik (Pieniny Klippen Belt, Poland) and their evolutionary relationships: Eclogae geologicae Helvetia, v. 72, p. 805–849.
Qing, H., Barnes, C.R., Buhl, D., Veizer, J., 1998, The strontium isotopic composition of Ordovician and Silurian brachiopods and conodonts: Relationships to geological events and implications for coeval seawater: Geochimica et Cosmochimica Acta, v. 62, p. 1721–1733.
Sadler, P.M., and Cooper, R.A., 2004, Calibration of the Ordovician Timescale, in Webby, B.D., Paris, F., Droser, M.L., and Percival, I.G., eds., The Great Ordovician Biodiversification Event: New York, New York, Columbia University Press, p. 48–51.
Sardeson, F.W., 1901, Problem of the Monticuliporoidea. II: The Journal of Geology, v. 9, no. 2, p. 149–173.
Simo, J.A., Emerson, N.R., Byers, C.W., and Ludvigson, G.A., 2003, Anatomy of an embayment in an Ordovician epeiric sea, Upper Mississippi Valley, USA: Geology, v. 31, no. 6, p. 545–548.
Simms, M.J., 1999, Systematics, phylogeny and evolution history, in Hess, H., ed., Fossil crinoids: New York, New York, Cambridge University Press, p. 31–40.
Sloan, R.E., 1988, Tectonics, biostratigraphy and lithostratigraphy of the Middle and Late Ordovician of the upper Mississippi Valley, in Middle and Late Ordovician lithostratigraphy and biostratigraphy of the Upper Mississippi Valley: Minnesota Geological Survey Report of investigations, p. 7–20.
Sloan, R.E., Kolata, D.R., Witzke, B.J., and Ludvigson, G.A., 1988, Descriptions of major outcrops in Minnesota and Iowa, in Middle and Late Ordovician
29

lithostratigraphy and biostratigraphy of the Upper Mississippi Valley: Minnesota Geological Survey Report of Investigations 35, p. 197–223.
Sprinkle, J., and Guensburg, T.E., 2004, Crinozoan, blastozoan, echinozoan, asterozoan, and homalozoan echinoderms, in Webby, B.D., Paris, F., Droser, M.L., and Percival, I.G., eds., The Great Ordovician Biodiversification Event: New York, New York, Columbia University Press, p. 266–280.
Sprinkle, J., and Moore, R.C., 1978, Echmatocrinea, in Moore, R.C., and Teichert, C., eds., Treatise on invertebrate paleontology, Part T, Echinodermata 2, Volume 2: Boulder, Colorado, Geological Society of America, and Lawrence, University of Kansas, p. T405–T407.
Taylor, P.D., and Ernst, A., 2004, Bryozoans, in Webby, B.D., Paris, F., Droser, M.L., and Percival, I.G., eds., The Great Ordovician Biodiversification Event: New York, New York, Columbia University Press, p. 147–156.
Tudge, Colin, 2000, The variety of life: Oxford, United Kingdom, Oxford University Press, 684 p.
Twitchell, G.B., 1934, Urnatella gracilis leidy, a living trepostomatous bryozoan: The American Midland Naturalist, v. 15, no. 6, p. 629–661.
Ubaghs, G., 1967, General characters of Echinodermata, in Moore, R.C., ed., Treatise on invertebrate paleontology, Part S, Echinodermata 1, Volume 1: Boulder, Colorado, Geological Society of America, and Lawrence, University of Kansas, p. S3–S60.
Ubaghs, G., 1969, Aethocrinus moorei Ubaghs, n. gen., n. sp., le plus ancien crinoïde dicyclique connu, University of Kansas Paleontological Contributions Paper 38, 26 p.
Ubaghs, G., 1978, Skeletal morphology of fossil crinoids, in Moore, R.C., and Teichert, C., eds., Treatise on invertebrate paleontology, Part T, Echinodermata 2, Volume 1: Boulder, Colorado, Geological Society of America, and Lawrence, University of Kansas, p. T58–T216.
Utgaard, J., 1983, Paleobiology and taxonomy of the order Cystoporata, in Robison, R.A., ed., Treatise on invertebrate paleontology, Part G, Bryozoa, revised, Volume 1: Boulder, Colorado, Geological Society of America, and Lawrence, University of Kansas, p. 327–357.
Witzke, B.J., 1980, Middle and Upper Ordovician Paleogeography of the region bordering the Transcontinental Arch, in Fouch, T.D., and Magathan, E.R., eds., Paleozoic paleogeography of the west-central United States: Rocky Mountain section, Society of Economic Paleontologists and Mineralogists, p.1–18.
30

APPENDICES APPENDIX 1. CRINOID FOSSILS OF THE DECORAH SHALE (FROM BROWER AND VEINUS, 1978). Subclass Camerata Wachsmuth and Springer, 1885 Order Diplobathrida (Moore and Laudon, 1943) Family Archaeocrinidae Archaeocrinus sp. Order Monobathrida Suborder Glyptocrinina Family Glyptocrinidae
Glyptocrinus tridactylus (Brower and Veinus, 1978) Pycnocrinus multibrachialis (Brower and Veinus, 1978) Pycnocrinus sardesoni (Brower and Veinus, 1978) Periglyptocrinus spinuliferus (Brower and Veinus, 1978)
Subclass Inadunata Order Cladida Suborder Dendrocrinina Family Cupulocrinidae
Cupulocrinus jewetti (E. Billings) Cupulocrinus canaliculatus (Brower and Veinus, 1978)
Family Dendrocrinidae Grenprisia billingsi (Springer)
Suborder Cyathocrinina Family Carabocrinidae
Carabocrinus dicyclicus (Sardeson) Carabocrinus magnificus Sardeson
Family Palaeocrinidae Palaeocrinus angulatus (E. Billings)
Family Porocrinidae Porocrinus pentagonius Meek and Worthen Order Disparida Superfamily Cincinnaticrinacea Family Cincinnaticrinidae
Isotomocrinus tenuis (E. Billings) Superfamily Homocrinicae Family Calceocrinidae
Cremacrinus punctatus Ulrich Attachment devices Lichenocrinid holdfasts Lobate and digitate holdfasts cemented to bryozoans Tree stump-like cirrus root Massive conical attachment-disk
31

APPENDIX 2. BRYOZOAN FOSSILS OF THE DECORAH SHALE (FROM KARKLINS, 1988). Order Trepostomata Family Amplexoporidae Amplexopora minnesotensis Amplexopora winchelli Family Trematoporidae Batostoma humile Batostoma increbescens Batostoma varium
Family Atactotoechidae Cyphotrypa cf. C. pachymuralis Cyphotrypa acervulosa Family Diplotrypidae Diplotrypa sp. Family Aisenvergiidae
Eridotrypa minor Family Dittoporidae
Hemiphragma irrasum Family Heterotrypidae
Heterotrypa* pauca Heterotrypa praenuntia echinata Heterotrypa praenuntia simplex Heterotrypa trentonensis Stigmatella crenulata Stigmatella spinosa
Family Mesotrypidae Homotrypa cf. H. curvata Homotrypa dickeyvillensis Homotrypa exilis
Homotrypa similis Homotrypa pauperata Homotrypa subramosa Prasopora insularis Prasopora simulatrix
Family Monticuliporidae Monticulipora grandis
Family Halloporidae Parvohallopora arguta Parvohallopora cf. P. tolli Parvohallopora incontroversa Tarphophragma ampla Tarphophragma multitabulata = Hallopora multitabulata
Unassigned Trepostome genera Nicholsonella ponderosa Nicholsonella sp.
32

Order Cryptostomata Suborder Ptilodictyina
Family Rhinidictyidae Athrophragma foliata Stictopora cf. S. neglecta Stictopora exigua Stictopora lita Stictopora minima Stictopora mutabilis Stictopora paupera Trigonodictya acuta Trigonodictya conciliatrix Trigonodictya elegans Trigonodictya fimbriata Trigonodictya pumila
Family Stictoporellidae Pseudostictoporella angularis Pseudostictoporella dumosa Pseudostictoporella frondifera
Stictoporellina cribrosa Family Escharoporidae
Escharopora subrecta Graptodictya simplex = Arthropora simplex?
Order Cystoporata Suborder Fistuliporina Family Anolotichiidae
Bythotrypa laxata *Heterotrypa = Dekayella
33