regulation of mucous differentiation and mucin gene expression in the tracheobronchial epithelium
TRANSCRIPT

Toxicology 160 (2001) 35–46
Regulation of mucous differentiation and mucin geneexpression in the tracheobronchial epithelium
Thomas Gray *, Ja-Seok Koo, Paul NettesheimLaboratory of Pulmonary Pathobiology, National Institute of En6ironmental Health Sciences, Research Triangle Park,
NC 27709, USA
Abstract
The goal of our studies is to elucidate mechanisms that control and modulate mucous differentiation and mucingene expression in the conducting airways. We used cultures of normal human tracheobronchial epithelial (NHTBE)cells that were shown to secrete two major airway mucins, namely MUC5AC and MUC5B as well as several othersecretory products. Mucous differentiation and expression of MUC2, MUC5AC, MUC5B and MUC7, but notMUCi, MUC4, and MUC8 mucin genes, were shown to be retinoic acid- (RA) or retinol-dependent. We found thatRA control of mucin genes was mediated by the retinoid acid receptors RAR a and, to a lesser extent, by RAR g.Our studies also showed that other important bioregulators such as thyroid hormone (T3) and epidermal growthfactor (EGF) modulate basal expression of mucin genes, interacting with RA in a concentration-dependent manner.T3, which binds to thyroid receptors (TRs) belonging to the same superfamily of steroid hormone nuclear receptorsas the RARs, inhibits mucin gene expression, particularly MUC5AC. One possible mechanism of this T3 effect isdownregulation of RAR proteins, which are critical for mucin gene expression. However, we also found that T3
inhibits MUC5AC transcription.EGF, which had previously been shown to stimulate mucin expression and mucin secretion in cultured rat tracheal
epithelial (RTE) cells, inhibited mucin secretion in human bronchial epithelial cell cultures. This effect was EGFconcentration- and time-dependent and was progressively abolished by increasing the RA concentration. Subsequentstudies suggested that the inhibitory effects of high concentrations of EGF may result from selective reduction ofMUC5AC expression. These studies thus point to potentially important species differences in the mechanismsregulating mucous production, and they also confirm previous findings indicating differential regulation of MUC5ACand MUC5B gene expression. Published by Elsevier Science Ireland Ltd.
Keywords: Mucin gene expression; Mucous differentiation; Tracheobronchial epithelium
www.elsevier.com/locate/toxicol
Abbre6iations: EGF, epidermal growth factor; NHTBE, normal human tracheobronchial epithelial; RA, retinoic acid; RAR,retinoic acid receptor; RT-PCR, reverse transcriptase-polymerase chain reaction; T3, triiodothyronine.
* Corresponding author. Tel.: +1-919-5413449; fax +1-919-5414133.E-mail address: [email protected] (T. Gray).
0300-483X/01/$ - see front matter Published by Elsevier Science Ireland Ltd.
PII: S 0300 -483X(00 )00455 -8

T. Gray et al. / Toxicology 160 (2001) 35–4636
1. Introduction
The mucociliary epithelium of the conductingairways fulfills many functions that are essentialfor maintaining the health of the respiratory tract.One of the critical functions is to produce secre-tions such as mucins, which protect the airwaysagainst microbial, particulate and chemical toxinsthat contaminate the breathing air. Virtually allforms of airway inflammation are associated withmucus hypersecretion, which can lead to airwayobstruction. An important goal of respiratorymedicine is to elucidate the mechanisms involvedin the regulation of mucin synthesis and secretion.
To date, 12 mucin genes, designated MUCs1–4, 5AC, 5B, 7–8 (Rose and Gendler, 1997),MUC9 (Lagow et al., 1999) and MUCs11–12,(Williams et al., 1999) have been identified. Threeof these mucin genes, MUC2, MUC5AC andMUC5B, encode polymeric mucins that are ex-pressed by airway epithelium; however, so far,only two major components of airway mucushave been identified, namely MUC5AC andMUC5B (Hovenberg et al., 1996a,b; Thornton etal., 1997; Wickstrom et al., 1998).
Vitamin A and its derivatives, so-calledretinoids, have been shown to regulate growthand differentiation of tracheobronchial epithelialcells (Gray et al., 1996; Guzman et al., 1996; Kooet al., 1999, 2000; Yoon et al., 1997, 1999). Ourefforts have been focused on the elucidation ofmechanisms by which retinoic acid (RA), a proto-typical retinoid, regulates differentiation of airwayepithelium and controls mucin gene expression.
In the studies reported here, we show that theeffects of RA are modified by thyroid hormone(T3). Both RA and T3 are ligands of closelyrelated nuclear receptors that act as transcrip-tional factors (Beato, 1991). We also examinedthe role of epidermal growth factor (EGF) in theregulation of mucin gene expression, becauseEGF has been shown in vivo, as well as in vitro,to effect mucous differentiation as well as mucinproduction (Stahlman et al., 1988; VanScott et al.,1988; St. George et al., 1991; Guzman et al., 1995;Takeyama et al., 1999).
2. Materials and methods
2.1. Air– liquid interface cultures
Passage-2, NHTBE cells (Clonetics Corp., LaJolla, CA) were seeded onto uncoated, semiper-meable Transwell clear membranes (CorningCostar, Cambridge, MA) in serum-free, hormoneand growth factor-supplemented medium (all me-dia supplements were purchased from Sigma, St.Louis, MO), containing all-trans retinoic acid(RA, Sigma). For the complete medium formula-tion, see Gray et al. (1996). Cultures were grownsubmerged for the first 7 days, at which time, theair–liquid interface was created. Cultures weremaintained at 37°C in a humidified atmosphere of5% CO2 in air.
The RAR-selective retinoid (Lehman et al.,1991), the RXR-selective retinoid (Lehman et al.,1992), and the RARg-selective retinoid (Martin et
Fig. 1. Morphology of NHTBE cultures. (A) Histological examination of NHTBE cultures grown in the presence of retinoic acidand stained with Alcian Blue (AB)-periodic acid shift (PAS) reveals a basal layer overlaid by a columnar epithelium containingseveral AB positive stained cells with typical goblet cell morphology (arrow); the apical surface of the epithelium is covered withcilia. (B) NHTBE cultures grown in the absence of RA reveal a stratified epithelium typical of a squamous pattern of differentiation.

T. Gray et al. / Toxicology 160 (2001) 35–46 37
Fig. 2. Time course of MUC2, MUC5AC and MUC5B mucin protein and mRNA levels. Total RNA, apical secretions and celllysates were collected from day 10, 14 and 21 day old cultures. The levels of secreted (A) and cell-associated (B) mucins weredetermined with mucin specific antibodies. The levels of (C) mucin gene mRNAs were determined by competitive RT-PCR. ForMUC 2, 10−4 amol of mimic was present in the PCR while for MUCSAC and MUCSB 10−1 amol of mimic was present.
al., 1992), were a generous gift from Dr. MarcialDawson (SRI International, Menlo Park, CA).The RARa-selective retinoid (Martin et al., 1992;Jetten et al., 1987) and the RARb selectiveretinoid (Geisen et al., 1997) were provided byHoffmann-La Roche, Ltd. (Nutley, NJ)
Immunodetection and quantitation of secretedtotal mucin, and specifically MUC5AC andMUC5B mucins. To monitor total secreted mucinproduction, apical secretions accumulating over a
24 h time period (triplicate samples per group)were collected and assayed by dot blotting (Grayet al., 1996) using the anti-mucin antibody H6C5(a generous gift from C.W. Davis, University ofNorth Carolina, Chapel Hill, NC). MUC5AC andMUCB levels were determined from intracellularand secreted material collected in 6 or 4 M guani-dinium hydrochloride, respectively. The levelswere determined by immunoreactivity withmucin-specific antisera (Thornton et al., 2000),

T. Gray et al. / Toxicology 160 (2001) 35–4638
and the signals were detected by chemilumines-cence (ECL kit, Amersham, Buckinghamshire,UK) and measured densitometrically. Statisticalcomparisons were made using Student’s t-test.
2.2. Competiti6e RT-PCR
Methods to detect and quantitate MUC5ACand MUC5B have been previously reported indetail (Guzman et al., 1996; Koo et al., 1999).Briefly, total RNA was collected from triplicateseparate cultures and was reverse-transcribed intocDNA. Oligonucleotide primers were designed ac-cording to published sequences (Guzman et al.,1996; Koo et al., 1999). PCR products were sepa-rated on 2% agarose gels (containing EtBr), andthe resulting bands were analyzed using a digitalimaging system (Alpha Innotech, San Eleandro,
CA). PCRs for MUC2, MUC5AC and MUC5Bwere performed in the presence of internal stan-dards so called MIMICs (Clontech PCR MIMICConstruction Kit, Palo Alto, CA), and the ratioof the signal intensity of the target cDNA to theMIMIC was determined.
2.3. Preparation of nuclear extracts and Westernblotting
Nuclear extracts were prepared from NHTBEcultures, separated by sodium dodecylsulfate–polyacrylamide gel electrophoresis, transferred tonitrocellulose membranes, incubated with nuclearreceptor specific antibodies (Santa Cruz Biotech-nology Inc., Santa Cruz, CA) and the signalsdetected by chemiluminescence (ECL kit).
3. Results and discussion
3.1. De6elopment of the mucociliary phenotypeand dependence of the expression of eicosanoidand nitric oxide enzymes on the state ofdifferentiation
To investigate factors and mechanisms involvedin the regulation of epithelial differentiation, wedeveloped the NHTBE air–liquid interface cul-ture system using early passage, airway epithelialcells. Following cell seeding, there is a rapidgrowth phase in which the epithelium is composedentirely of poorly differentiated, non-polarizedcells. As the cell density increases and the culturesreach confluency (�day 10–day 14), the epithe-lium becomes visibly polarized with many cellscontaining AB-PAS positively stained granulesand apical secretion of mucus and other secretoryproducts occurs (Gray et al., 1996). In the lastphase of differentiation (�day 21–day 28), cili-ated cells appear in great numbers (Fig. 1A). Theserum-free culture media, in which the cells aregrown, contain growth factors and hormones aswell as retinoic acid (Gray et al., 1996). If theretinoid is deleted from the media, cultures un-dergo metaplastic, squamous differentiation (Fig.1B), express biochemical markers of squamousdifferentiation and do not produce mucin (Gray
Fig. 3. Mucin gene expression in RA-deficient and RA-suffi-cient cultures. Total RNA was collected from NHTBE cul-tures grown in the absence or presence of RA for 14 days.Human tracheal RNA was obtained from a commercialsource. The levels of mucin gene mRNA expression weredetermined by RT-PCR using primers based on publishedsequences for each gene.

T. Gray et al. / Toxicology 160 (2001) 35–46 39
Fig. 4. Effects of RAR and RXR-selective retinoids on mucin gene expression. (A) NHTBE cultures were treated with the indicatedconcentration of RAR or RXR selective retinoids and the mRNA levels of MUC2, MUC5AC and b2M were determined byRT-PCR. (B) Using the same experimental protocol, NHTBE cultures were treated with various concentrations of RARa-selective,RARb-selective or RARg-selective retinoids, and mucin gene and b2M mRNA (PCR control gene) levels were determined.
et al., 1996; Guzman et al., 1996; Yoon et al.,1997).
We found that the state of differentiation of thecultures also dramatically affects the expression ofeicosanoid enzymes and their products, importantbioregulators of inflammation. Undifferentiated
and rapidly growing cells strongly expressed theinducible cyclooxygenase (COX)-2 but did notexpress 15-lipoxygenase (15-LO). In contrast,fully differentiated cultures repressed the COX-2expression and instead strongly expressed 15-LO.We also found that the squamous epithelium of
T. Gray et al. / Toxicology 160 (2001) 35–4640
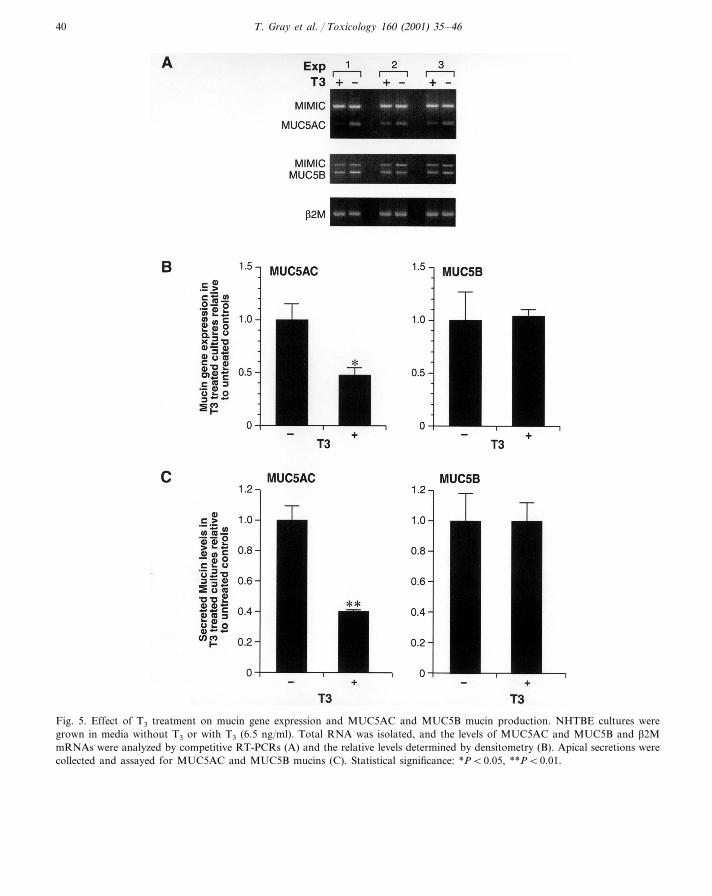
Fig. 5. Effect of T3 treatment on mucin gene expression and MUC5AC and MUC5B mucin production. NHTBE cultures weregrown in media without T3 or with T3 (6.5 ng/ml). Total RNA was isolated, and the levels of MUC5AC and MUC5B and b2MmRNAs were analyzed by competitive RT-PCRs (A) and the relative levels determined by densitometry (B). Apical secretions werecollected and assayed for MUC5AC and MUC5B mucins (C). Statistical significance: *PB0.05, **PB0.01.

T. Gray et al. / Toxicology 160 (2001) 35–46 41
retinoid-deficient cultures expressed neither COX-2 nor 15-LO (Hill et al., 1996, 1998; Nettesheim etal., 2000). Further studies showed that the expres-sion of nitric oxide synthase (NOS) is also differ-entiatiation- and RA-dependent (Norford et al.,1998). iNOS and eNOS were only expressed indifferentiated epithelium, and bNOS was onlyexpressed in undifferentiated cultures. These stud-ies showed the dependency of two unrelated en-zyme systems, namely eicosanoid and nitric oxidesynthases, on the state of differentiation and serveto remind us that many biochemical activities aredependent on cell- and tissue-specific differentia-tion, which in turn is modulated by a variety ofbioregulators.
3.2. Two major gel-forming mucins, MUC5ACand MUC5B, are produced by NHTBE cultures
To determine whether the major gel-formingmucins present in human airways (i.e., MUC5ACand MUC5B: Hovenberg et al., 1996a,b; Sheehanet al., 1999) are expressed during mucociliarydevelopment of cultured NHTBE cells, we col-lected apical washings and cell lysates from 10 to21 day cultures and determined the levels ofsecreted and intracellular MUC2, MUC5AC and
MUC5B mucins using specific antibodies. Asshown in Fig. 2, over the course of the experi-ment, there was a 10-fold increase in the amountof secreted (Fig. 2A) and intracellular (Fig. 2B)MUC5AC and MUC5B mucin. MUC2 mucinwas detected at low levels approaching back-ground. As shown in Fig. 2C, MUC5AC andMUC5B mRNA levels, determined by competi-tive RT-PCR, were low on day 10 but wereclearly increased on days 14 and 21. The levels ofMUC2 mRNA expressed throughout the courseof the study (the amount of MIMIC in thesereactions was 10−3 of that present for measuringMUC5AC and MUC5B) were much lower com-pared to those expressed for both MUC5AC andMUC5B. These studies showed that culturedNHTBE cells secrete MUC5AC and MUC5B andfurther showed that both at the mRNA andprotein level, the expression of these two mucinscorrelates with the morphological differentiationof the mucous phenotype (Thornton et al., 2000).
3.3. Regulation of mucin gene expression by RAand the role of RA receptors
Of the 12 mucin genes identified to date, onlyMUC1, MUC2, MUC4, MUC5AC, MUC5B,MUC7 and MUC8 have been shown to be ex-pressed in the respiratory tract (Rose andGendler, 1997). We have shown earlier thatMUC2, MUC5AC and MUC5B mRNA expres-sion is retinoid-dependent (Guzman et al., 1996;Yoon et al., 1997; Koo et al., 1999, 2000). Morerecently, we compared the mucin gene expressionprofile in RA-deficient and RA-sufficient cultures.The findings summarized in Fig. 3 confirmed thatexpression of MUC2, MUC5AC and MUC5B isentirely dependent on the presence of RA; incontrast, MUC1, MUC4, MUC7 and MUC8 areexpressed regardless of the presence or absence ofRA, although RA slightly increases MUC1 andMUC7 mRNA levels.
The effects of retinoids are mediated via twofamilies of retinoid receptors, the retinoic acidreceptors (RARs) and the retinoid X receptors(RXRs), which belong to the superfamily ofsteroid hormone nuclear receptors. Each of thetwo subfamilies of retinoid receptors consists of
Fig. 6. Effect of T3 on retinoic acid receptor expression. Fiftymicrograms of nuclear lysates were separated on SDS–PAGE,and the levels of each receptor protein were examined usingrelevant specific antibodies.

T. Gray et al. / Toxicology 160 (2001) 35–4642
Fig. 7. Effect of EGF on MUC5AC and MUC5B mRNA expression. Completely differentiated NHTBE cultures were treated withvarious concentrations of EGF, and the mRNA levels of MUC5AC (solid bars) and MUC5B (hatched bars) were determined (A).(B) Similar NHTBE cultures were treated for indicated times with either control levels of EGF (0.5 ng/ml) or 25 ng/ml. Total RNAwas collected and assayed for MUC5A and MUC5B gene expression. The data are reported as levels of expression in EGF-treatedcultures compared to control.
three receptor isoforms designated RARa, b, g,for which RA is the major ligand, and RXRa, b,g, for which 9-cis RA is the major ligand. Specificreceptor selective agonists for the RAR isoformshave been synthesized (Majewski et al., 1995). Togain some insight into the retinoid signaling path-ways involved in the regulation of mucin geneexpression, we used several retinoid receptor-se-
lective agonists (Koo et al., 2000). As shown inFig. 4A, RA and RAR-selective retinoids wereboth effective in inducing mucin gene expression.In contrast, the RXR-selective agonist did not byitself induce mucin gene expression or mucousdifferentiation. However, the RXR-selective ago-nist enhanced the induction of mucin gene expres-sion in cultures treated with sub-optimal
T. Gray et al. / Toxicology 160 (2001) 35–46 43
concentrations of the RAR-selective agonist (datanot shown). We further found, as shown in Fig.4B, that mucin gene expression can be induced byboth RARa and RARg-selective agonists but notby the RARb-selective agonist, and the RARa-se-lective antagonist (R041-5253) completely blockedmucin gene expresssion induced by RA (Koo etal., 2000). These studies revealed that RARa isthe major retinoid receptor mediating the induc-tion of mucin gene expression by RA.
3.4. Differential regulation of MUC5AC andMUC5B by T3
The effects of the hormone T3 on various cellsand tissues are mediated by the thyroid hormonereceptors (TR) that belong to the same nuclearreceptor family as the retinoid receptors. It is wellknown that both the RARs and the TRs form
heterodimers with RXRs, so T3 might modulateRA effects on mucin gene expression. As shownin Fig. 5A and B, the addition of T3 caused a 60%decrease in the levels of steady-state MUC5ACmRNA, but MUC5B, and b2M, RNA used as acontrol gene, were unaffected by T3. SecretedMUC5AC mucin level also was reduced 50–60%in T3-treated cultures (Fig. 5C, left panel), butMUC5B mucin was not affected (Fig. 5C, rightpanel). These data indicate that MUC5AC andMUC5B are differentially regulated by T3 (Grayet al., in press).
T3 has been shown to downregulate a numberof different proteins (Samuels et al., 1988), so weconsidered the possibility that it downregulatesnuclear receptors. We therefore prepared nuclearextracts from NHTBE cultures treated with T3.The receptors were identified by Western blottingusing antibodies directed against specific receptor
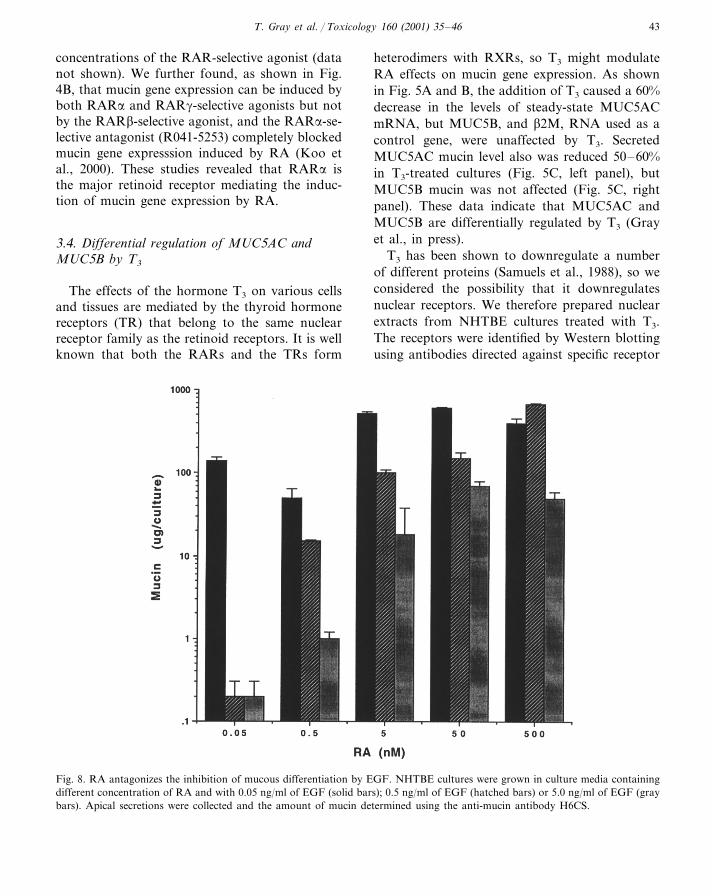
Fig. 8. RA antagonizes the inhibition of mucous differentiation by EGF. NHTBE cultures were grown in culture media containingdifferent concentration of RA and with 0.05 ng/ml of EGF (solid bars); 0.5 ng/ml of EGF (hatched bars) or 5.0 ng/ml of EGF (graybars). Apical secretions were collected and the amount of mucin determined using the anti-mucin antibody H6CS.

T. Gray et al. / Toxicology 160 (2001) 35–4644
isotypes. As shown in Fig. 6, the levels of RARa,b and g and RXRa and g proteins were reducedfollowing T3 treatment. In contrast, expression ofTRb and b-actin protein was only marginally, ifat all, affected (no TRa was detected; data notshown). Since the expression of MUC5AC is de-pendent on RA, the effect of which is mediated byretinoid receptors, it is likely that the reason forthe inhibition of mucin gene expression is thereduction of retinoid receptor proteins caused byT3. However, we have recently also found evi-dence for transcriptional inhibition of MUC5ACby T3 (Gray et al., in press).
3.5. EGF suppresses MUC5AC but not MUC5Bexpression
We previously reported that NHTBE cell cul-tures require low levels of EGF for growth andmucous differentiation (Gray et al., 1996). How-ever, we found that treatment of fully differenti-ated cultures with increasing concentrations ofEGF suppressed MUC5AC mRNA levels (Fig.7A). Reduced MUC5AC levels were first notice-able at 48 h after EGF treatment (Fig. 7B). Incontrast, MUC5B mRNA levels were not affectedby EGF.
We hypothesized that EGF might be suppress-ing the mucous phenotype and that the degree ofsuppression might depend on the balance betweenEGF and RA. To test this hypothesis, NHTBEcells were grown in media containing 0.05 to 500nM RA (the standard concentration used is 50nM) and the EGF concentration was either 0.05,0.5 or 5.0 ng/ml. As shown in Fig. 8, culturesgrown in low concentrations of EGF requiredlittle RA to produce large amounts of mucin, butunder these conditions, mucin secretion was dra-matically inhibited when the EGF concentrationwas increased to 0.5 or 5.0 ng/ml. Increasing theRA concentration in cultures containing higherlevels of EGF completely restored mucin secre-tion. We concluded that in the absence of RA, orat low concentrations of RA, EGF suppressed themucous phenotype, and higher concentrations ofRA rescued mucous differentiation from the sup-pressive effects of EGF.
The results reported by various investigators onthe effect of EGF and other EGF ligands on themucous phenotype appear to be contradictory. Insome studies, EGF seems to support or enhanceexpression of the mucous phenotype; in others, itseems to suppress it (Stahlman et al., 1988;VanScott et al., 1988; St. George et al., 1991;Takeyama et al., 1999). In earlier studies withcultured rat tracheal epithelial cells, EGF en-hanced mucous cell differentiation and mucingene expression (Guzman et al., 1995). It appearsthat important species differences, as well as dif-ferences of experimental design such as RA andEGF concentrations, may account for some of thediscrepancies. The mechanisms of the RA–EGFinteractions demonstrated here are presently notunderstood. Recent studies have demonstratedthat retinoic acid receptor complexes inhibit fos/jun APi mediated transcription of a number ofgenes including genes induced by EGF (Nicholsonet al., 1990). In addition, RA has been shown toalter transcription, translation as well as phospho-rylation of the EGF-receptor (Raymond et al.,1990; Kim et al., 1992; Zheng et al., 1992).Whether such mechanisms are involved in theEGF–RA interaction in the regulation ofMUC5AC expression remains to be determined.
In summary, our studies show that MUC5ACand MUC5B are differentially regulated by RA,T3 and EGF. MUC5AC responds much moredramatically and rapidly to external signals thanMUC5B. We have shown that mucous differentia-tion in cultured NHTBE cells is mediated throughRAR signaling pathways. These complex signal-ing pathways are modulated by thyroid hormone,which suppresses retinoic acid receptor expressionand MUC5AC transcription. The mechanism bywhich EGF interacts with the RA signaling path-way in the regulation of MUC5AC mucin geneexpression is currently unknown.
References
Beato, M., 1991. Transcriptional control by nuclear receptors.FASEB J. 3, 2044–2051.
Geisen, C., Denk, C., Gremm, B., Braust, C., Karger, A.,Bollag, W., Schwarz, E., 1997. Cancer Res. 57, 1460–1467.

T. Gray et al. / Toxicology 160 (2001) 35–46 45
Gray, T., Davis, C.W., Abdullah, L.H., Nettesheim, P.,1996. Mucociliary differentiation of serially passaged nor-mal human tracheobronchial epithelial (NHTBE) cells.Am. J. Respir. Cell Mol. Biol. 14, 104–112.
Gray, T., Nettesheim, P., Basbaum, C., Koo, J.-S., in press.Regulation of mucin gene expression in human tra-cheobronchial epithelial cells by thyroid hormone.
Guzman, K., Gray, T., Yoon, J.-H., Nettesheim, P., 1996.Quantitation of mucin RNA by PCR reveals inductionof both MUC2 and MUCS mRNA levels by retinoids.Am. J. Physiol. 271, L1023–L1028.
Guzman, K., Randell, S., Nettesheim, P., 1995. Epidermalgrowth factor regulates expression of mucous phenotypeof rat tracheal epithelial cells. Biochem. Biophys. Res.Commun. 217, 412–418.
Hill, E., Bader, T., Eling, T, Nettesheim, P., 1996. Retinoid-induced differentiation regulates prostaglandin-H syn-thase and cPLA2 expression in tracheal epithelium. Am.J. Physiol. 270, L852–L854.
Hill, E., Eling, T., Nettesheim, P., 1998. Changes in expres-sion of 15-lipoxygenase and prostaglandin-H synthaseduring differentiation of human tracheobronchial epithe-lial cells. Am. J. Respir. Cell Mol. Biol. 18, 662–669.
Hovenberg, H., Davies, J., Carlstedt, I., 1996a. Differentmucins are produced by surface epithelium and the sub-mucosa of human trachea: Identification of MUC5AC asa major mucin from goblet cells. Biochem. J. 318, 319–324.
Hovenberg, H., Davies, J., Herrmann, A., Linden, C., Carl-stedt, I., 1996b. MUC5AC, but not MUC2, is a promi-nent mucin in respiratory secretions. Glycoconj. J. 13,839–847.
Jetten, A., Anderson, K., Deas, M., Kagechika, H., Lotan,R., Rearick, J., Shudo, K., 1987. Cancer Res. 47, 3523–3527.
Kim, J., Steck, P., Calleck, G., Lee, J., Blick, M, Hong, W,Lotan, R., 1992. Suppression by retinoic acid of epider-mal growth factor receptor autophosphorylation and gly-cosylation in cultured human head and neck squamouscarcinoma cells. J. Natl. Cancer Inst. Monogr. 13, 101–111.
Koo, J.-S., Jetten, A., Belloni, P., Yoon, J.-H., Kim, Y.D.,Nettesheim, P., 2000. Role of retinoid receptors in theregulation of mucin gene expression by retinoic acid inhuman tracheobronchial epithelial cells. Biochem. J. 338,351–357.
Koo, J.-S., Yoon, J.-H., Gray, T., Norford, D., Jetten, A.,Nettesheim, P., 1999. Restoration of the mucous pheno-type by retinoic acid in retinoid-deficient humanbronchial cell cultures: Changes in mucin gene expres-sion. Am. J. Respir. Cell Mol. Biol. 20, 43–52.
Lagow, E., DeSouza, M., Carson, D., 1999. Mammalianreproductive tract mucins. Hum. Reprod. Update 5,280–290.
Lehman, J., Dawson, M., Hobbs, P., Hussman, M., Pfahl,M, 1991. Cancer Res. 51, 4804–4809.
Lehman, J., Jong, L., Fanjul, A., Cameron, J., Lu, X.,Haefner, P., Dawson, M., Pfahl, M., 1992. Science 258,1944–1946.
Majewski, S., Marczak, M., Szmurlo, A., Jablonska, S., Bol-lag, W., 1995. Cancer Lett. 89, 117–124.
Martin, B., Bernardon, J., Cavey, M., Bernard, B., Carla-van, I., Carpentier, B., Pilgrim, W., Shroot, B., Reichert,U., 1992. Skin Pharmacol. 5, 57–65.
Nettesheim, P., Koo, J.S., Gray, T., in press. Regulation ofdifferentiation of tracheobronchial epithelium. J. AerosolMed.
Nicholson, R., Mader, S., Nagpal, S., Leid, M., Rochette-Egly, C., Chambon, P., 1990. Negative regulation of ratstromelysin gene promoter by retinoic acid is mediatedby an APi binding site. EMBO J. 9, 4443–4454.
Norford, D., Koo, J.-S., Gray, T., Adler, K., Nettesheim,P., 1998. Expression of nitric oxide synthase isoforms innormal human tracheobronchial epithelial cells in vitro:Dependence on retinoic acid and state of differentiation.Exp. Lung Res. 24, 355–366.
Raymond, V., Grisham, L., Earp, S., 1990. Characterizationof epidermal growth factor receptor induction by retinoicacid in a chemically transformed rat liver cell line. CellGrowth Diff. 1, 393–399.
Rose, M., Gendler, S., 1997. Airway mucin genes and geneproducts. In: Rogers, D.F., Lethem, M.I. (Eds.), AirwayMucus: Basic Mechanisms and Clinical Perspectives.Birkhauser Publishing Limited, Basel, pp. 41–66.
St. George, J., Read, L., Crauz, D., Tarantal, A., George-Nascimento, G., Plopper, C., 1991. Effect of epidermalgrowth factor on the fetal development of tra-cheobronchial secretory apparatus in rhesus monkeys.Am. J. Respir. Cell Mol. Biol. 4, 95–101.
Samuels, H., Forman, B., Horowitz, Z., Ye, Z., 1988. Regu-lation of gene expression by thyroid hormone. J. Clin.Invest. 81, 957–967.
Sheehan, J., Howard, M., Richardson, P., Longwill, T.,Thornton, D., 1999. Physical characterization of a low-charge glycoform of MUC5B mucin comprising the gel-phase of an asthmatic respiratory mucous plug. Biochem.J. 338, 507–513.
Stahlman, M., Gray, M., Chytil, F., Sundell, H., 1988. Ef-fect of retinol on fetal lamb tracheal epithelium with andwithout epidermal growth factor. Lab. Invest. 59, 25–35.
Takeyama, K., Dabbagh, K., Lee, H.-M., Agusti, C.,Lausier, J., Ueki, I., Grattan, K., Nadel, J., 1999. Epi-dermal growth factor system regulates mucin productionin airways. Proc. Natl. Acad. Sci. 96, 3081–3086.
Thornton, D., Gray, T., Nettesheim, P., Howard, M., Koo,J.-S., Sheehan, J., 2000. Characterization of mucins fromcultured normal tracheobronchial epithelial cells. Am. J.Physiol. 278, L1118–L1128.
Thornton, D., Howard, M., Khan, N., Sheehan, J., 1997.Identification of two glycoforms of the MUC5B mucin inhuman respiratory mucus. Evidence for a cysteine-richsequence repeated within the molecule. J. Biol. Chem.272, 9561–9566.

T. Gray et al. / Toxicology 160 (2001) 35–4646
VanScott, M., Lee, N., Yankaskas, J., Boucher, R.,1988. Effect of hormones on growth and function ofcultured canine epithelial cells. Am J. Physiol. 24, C237–C245.
Wickstrom, C., Davies, J., Eriksen, G., Veerman, E., Carlst-edt, I., 1998. MUC5B is a major gel-forming, oligomericmucin from salivary gland, respiratory tract and endo-cervix. Biochem. J. 334, 685–693.
Williams, S., McGluckin, M., Gotley, D., Eyre, H., Suther-land, G., Antal is, T., 1999. Two novel mucin genesdown-regulated in colorectal cancer identified by differen-tial display. Cancer Res. 59, 4083–4089.
Yoon, J.-H., Gray, T, Guzman, K, Nettesheim, P., 1997.Regulation of secretory phenotype of human airway ep-ithelium by retinoic acid, triiodothyronine and extracellu-lar matrix. Am. J. Respir. Cell Mol. Biol. 16, 724–731.
Yoon, J.-H., Koo, J.-S., Norford, D., Guzman, K., Gray,T., Nettesheim, P., 1999. Lysozyme expression duringmetaplastic squamous differentiation of retinoic acid-defi-cient human tracheobronchial epithelial cells. Am. J.Respir. Cell Mol. Biol. 20, 573–581.
Zheng, Z-Sh., Polakowska, R., Johnson, A., Goldsmith, L.,1992. Transcriptional control of epidermal growth factorreceptor by retinoic acid. Cell Growth Diff. 3, 225–230.
.