regulatory strategies: atcase & haemoglobinwebspace.qmul.ac.uk/rwjanes/basic_13_16_web.pdf ·...
TRANSCRIPT

Regulatory Strategies: ATCase &
Haemoglobin
Allosteric Regulation
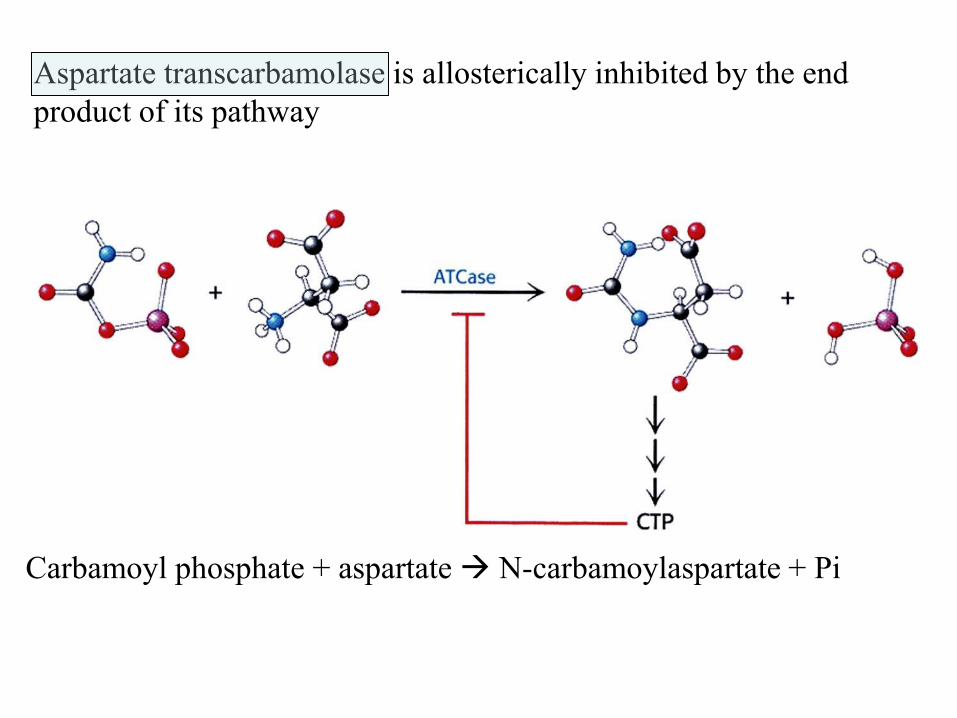
Aspartate transcarbamolase is allosterically inhibited by the end
product of its pathway
Carbamoyl phosphate + aspartate N-carbamoylaspartate + Pi

Aspartate transcarbamolase
• Catalyses the first step (the committed step)
in the biosynthesis of pyrimidines (thiamine
and cytosine), bases that are components of
nucleic acids

Condensation of aspartate and carbomyl phosphate to form N-
Carbamoylaspartate

• How is the enzyme regulated to generate
precisely the amount of CTP needed by the
cell?

CTP inhibits ATCase, despite
having little structural similarity to
reactants or products

CTP inhibits ATCase, despite
having little structural similarity to
reactants or products
FEEDBACK INHIBITION
Allosteric regulation

ATCase Structure

ATCase Consists of Separate Catalytic
and Regulatory Subunits
• Can be separated
into regulatory and
catalytic subunits
by treatment with
p-hydroxy-
mercuribenzoate
(PCMB), which
reacts with
sulfhydryl groups

2c3 + 3r2 c6r6
Ultracentrifugation Activity
PCMB treated
ACTase Native ACTase 11.6S
2.8S 5.8S
Mercurial dissociate ATCase into
two subunits

Subunit characteristics
• Regulatory subunit (r2)
– Two chains (17kd each)
– Binds CTP
– No enzyme activity
• Catalytic subunit (c3)
– Three chains
– Retains enzyme activity
– No response to CTP

Cysteine binds Zn – PCMB
displaces Zn and destabilizes
the domain
Structure of ATCase

Potent
competitive
inhibitor
Carbamoyl phosphate
Aspartate
Use of PALA to locate active site
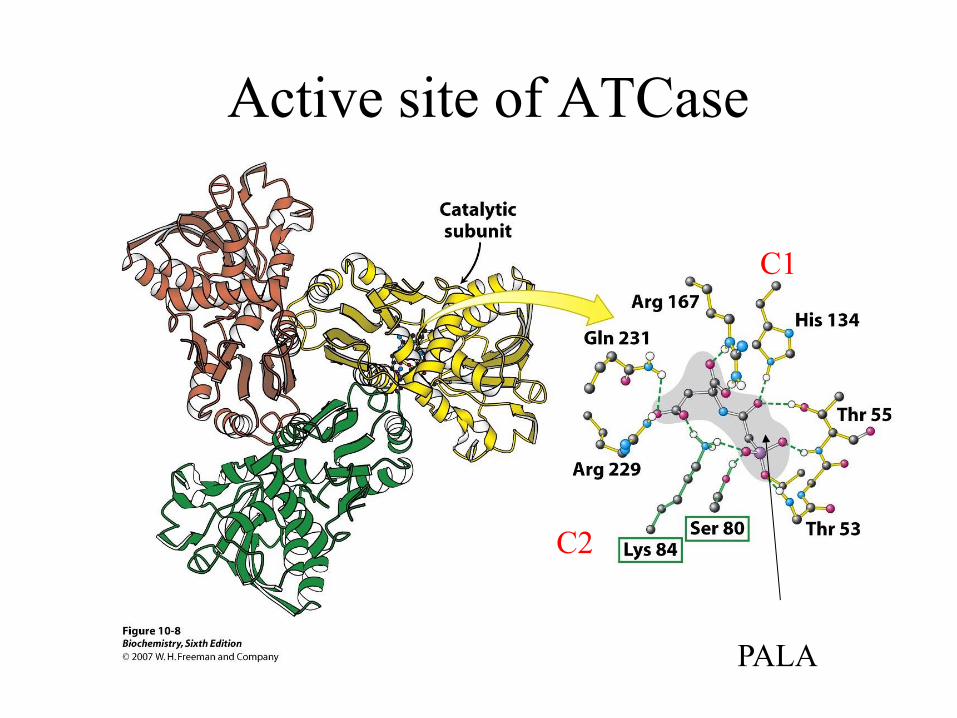
Active site of ATCase
PALA
C1
C2

The T-to-R state transition
Each catalytic trimer has 3 substrate binding sites
Enzyme has two quaternary forms.

CTP stabilises the T state
• T state when CTP bound
• Binding site for CTP
in each regulatory domain
• Binds 50Å from active site
– allosteric

R and T state are in equilibrium
Mechanism for CTP inhibition

ATCase displays sigmoidal kinetics
T>R
R>T
Cooperativity

Why does ATCase display
sigmoidal kinetics
• The importance of the changes in quaternary structure in determining the sigmoidal curve is illustrated by studies on the isolated catalytic trimer, freed by p-hydroxymercuribenzoate treatment.
• The catalytic subunit shows Michaelis-Menten kinetics with kinetic parameters indistinguishable from those deduced for the R-state.
• The term tense is apt – the regulatory dimers hold the two catalytic trimers close so key loops collide & interfere with the conformational adjustments necessary for high affinity binding & catalysis.

Basis for the sigmoidal curve
(mixture of two Michaelis Menten enzymes)
High KM
Low KM

Allosteric regulators modulate
the T-to-R equilibrium

CTP is an allosteric inhibitor
T>R

ATP is an allosteric activator
High purine
High Energy
mRNA synthesis ↑
R>T
ATP Competes for same binding site

Haemoglobin


Myoglobin
• Myoglobin is a single
polypeptide,
hemoglobin has four
polypeptide chains.
• Haemoglobin is a
much more efficient
oxygen-carrying
protein. Why?

Myoglobin and Haemoglobin
bind oxygen at iron atoms in
heme
1 2
3 4
Fe2+
Fe has 6 co-ordination sites
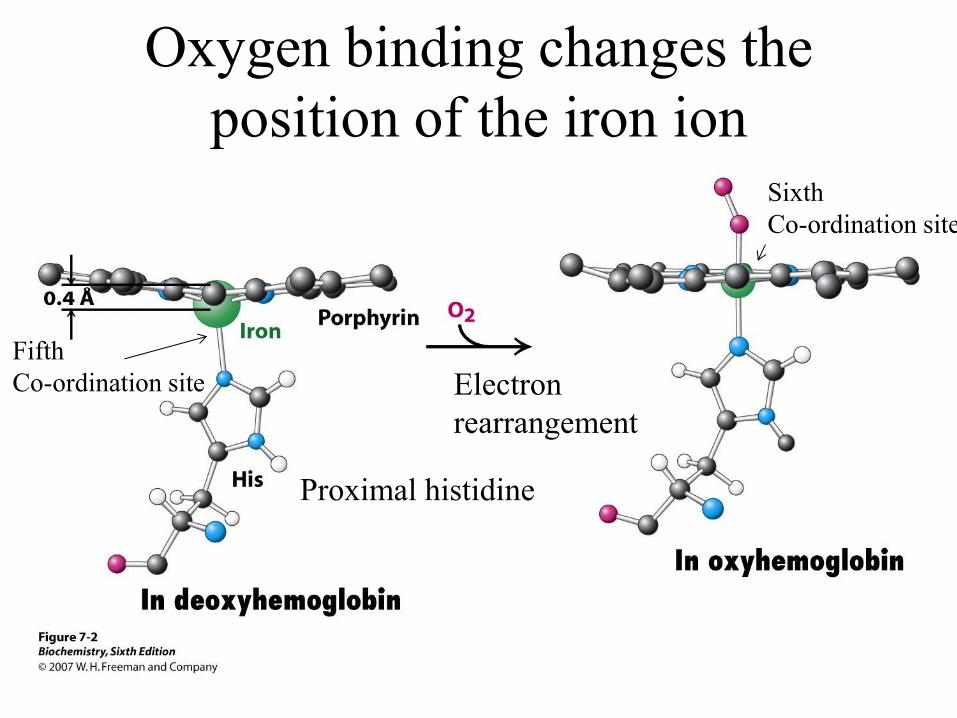
Proximal histidine
Sixth
Co-ordination site
Oxygen binding changes the
position of the iron ion
Fifth
Co-ordination site Electron
rearrangement

Myoglobin – stabilising bound
oxygen
Stronger binding

Why is haemoglobin more
efficient at binding oxygen?

a1b1 and a2b2 dimers
Quaternary structure of
deoxyhemoglobin - HbA

Oxygen binding to myoglobin
Simple equilibrium.

Haemoglobin as an allosteric protein
• Haemoglobin consists of 2a and 2b chains
• Each chain has an oxygen binding site,
therefore haemoglobin can bind 4 molecules
of oxygen in total
• The oxygen-binding characteristics of
haemoglobin show it to be allosteric

Oxygen binding to haemoglobin
in rbc
Cooperativity

Physiological significance of
allosteric regulation

Cooperative unloading of oxygen
enhances oxygen delivery

Haemoglobin
• Two principal models have been developed
to explain how allosteric interactions give
rise to sigmoidal binding curves
• The concerted model
• The sequential model

Concerted model
• Oxygen can bind to either conformation, but
as the number of sites with oxygen bound
increases, so the equilibrium becomes
biased towards one conformation (in the
case of increasing oxygen bound, the R
conformation)
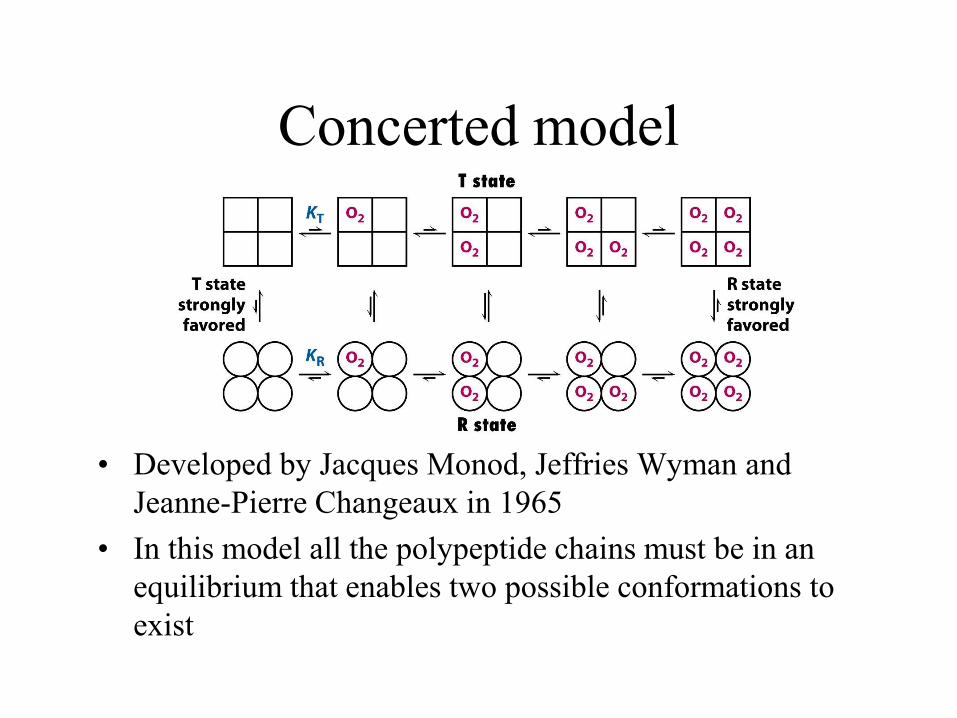
Concerted model
• Developed by Jacques Monod, Jeffries Wyman and
Jeanne-Pierre Changeaux in 1965
• In this model all the polypeptide chains must be in an
equilibrium that enables two possible conformations to
exist

Concerted model
• The concerted model assumes:
1. The protein interconverts between the two conformation T and R but all subunits must be in the same conformation
2. Ligands bind with low affinity to the T state and high affinity to the R state
3. Binding of each ligand increases the probability that all subunits in that protein molecule will be in the R state

Sequential model
• Assumes
1. Each polypeptide chain can only adopt one of two conformations T and R.
2. Binding of ligand switches the conformation of only the subunit bound.
3. Conformational change in this subunit alters the binding affinity of a neighbouring subunit i.e. a T subunit in a TR pair has higher affinity that in a TT pair because the TR subunit interface is different from the TT subunit interface.

Sequential model
• Devised by Dan Koshland in the 1950s
• Substrate binds to one site and causes the polypeptide to
change conformation
• Substrate binding to the first site affects the binding of a
second substrate to an adjoining site
• And so on for other binding sites …

How does oxygen binding induce
change from T to R state
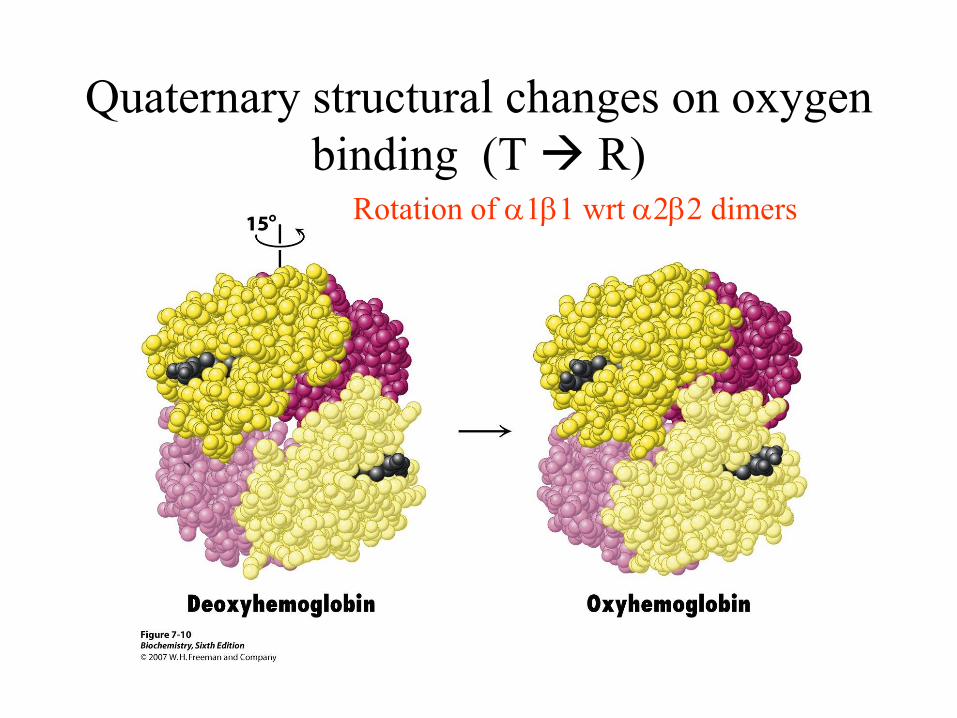
Quaternary structural changes on oxygen
binding (T R) Rotation of a1b1 wrt a2b2 dimers

Conformational change in haemoglobin
T → R
N

The role of 2,3
bisphosphoglycerate in red blood
cells

Haemoglobin must remain in T
state in absence of oxygen
T – state is extremely
unstable
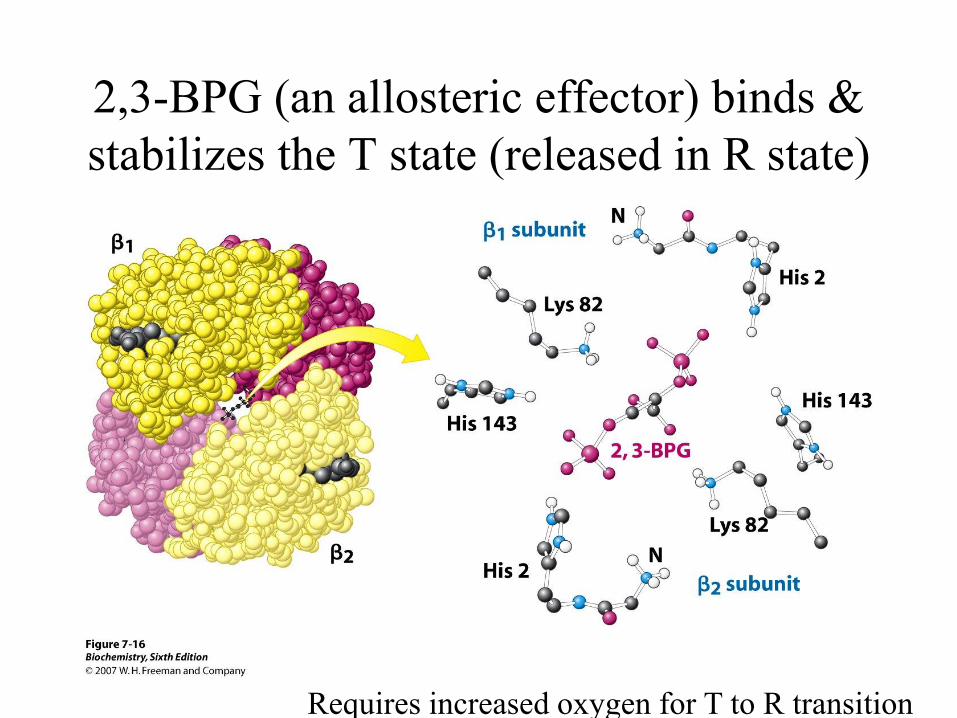
2,3-BPG (an allosteric effector) binds &
stabilizes the T state (released in R state)
Requires increased oxygen for T to R transition
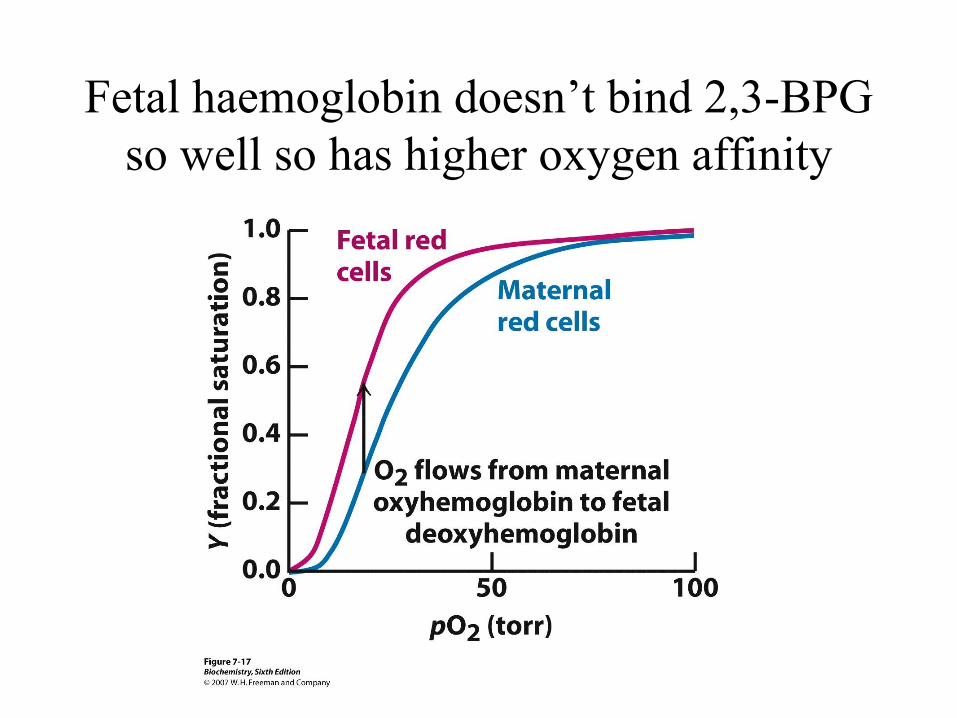
Fetal haemoglobin doesn’t bind 2,3-BPG
so well so has higher oxygen affinity

Bohr effect (protons are also allosteric
effectors) T-state stabilized by
salt bridges
Thus oxygen is
released
Salt bridges

Carbonic anhydrase
Also … CO2 forms carbamate (R-NH-CO2) with N-ter – at
interface between αβ dimers favours release of O2 by
favouring the T state

Carbon dioxide promotes the release of
oxygen

Sickle cell anaemia
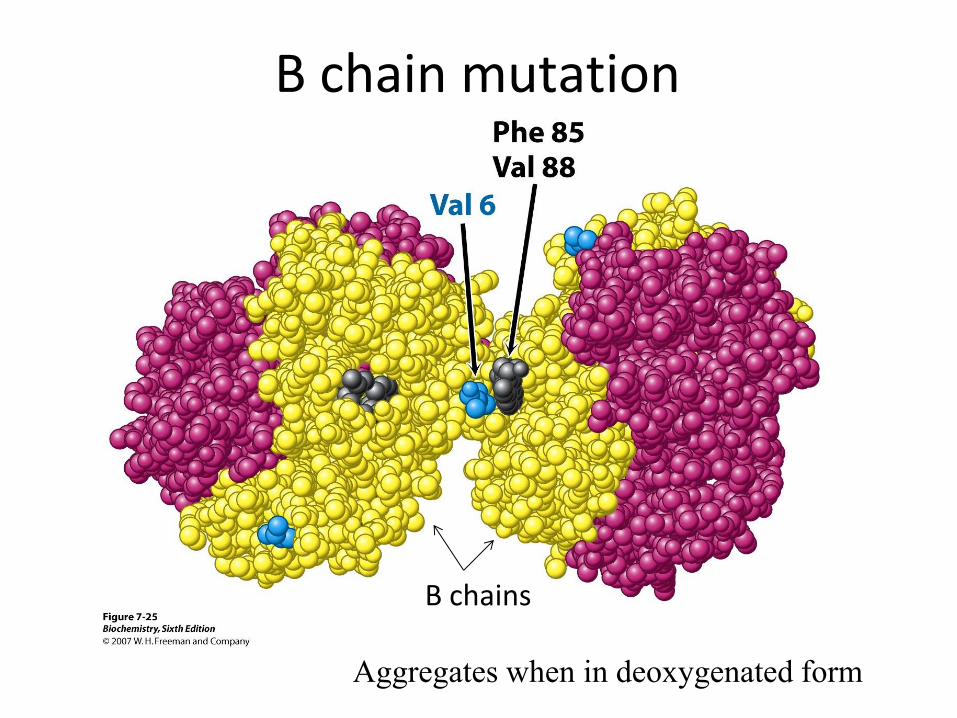
Aggregates when in deoxygenated form
Β chains
Β chain mutation

Plasmodium falciparum
Why is HbS so prevalent in
Africa
• Sickle cell trait (one allele mutation)
resistant to malaria