research article phlebotomines (diptera: psychodidae) in a...
TRANSCRIPT

Research ArticlePhlebotomines (Diptera Psychodidae) in a HydroelectricSystem Affected Area from Northern Amazonian BrazilFurther Insights into the Effects of Environmental Changeson Vector Ecology
Nercy Virginia Rabelo Furtado1 Allan Kardec Ribeiro Galardo1 Clicia Denis Galardo1
Viviane Caetano Firmino2 and Thiago Vasconcelos dos Santos3
1Laboratorio de Entomologia Medica Instituto de Pesquisas Cientıficas e Tecnologicas do Estado do Amapa Macapa AP Brazil2Universidade Federal do Amapa Campus Binacional Oiapoque AP Brazil3Instituto Evandro Chagas Secretaria de Vigilancia em Saude Ministerio da Saude Ananindeua PA Brazil
Correspondence should be addressed toThiago Vasconcelos dos Santos thiagovasconcelosiecpagovbr
Received 3 August 2016 Revised 6 October 2016 Accepted 12 October 2016
Academic Editor Aditya Prasad Dash
Copyright copy 2016 Nercy Virginia Rabelo Furtado et al This is an open access article distributed under the Creative CommonsAttribution License which permits unrestricted use distribution and reproduction in any medium provided the original work isproperly cited
During 2012ndash2015 an entomological survey was conducted as part of a phlebotomine (Diptera Psychodidae) monitoring programin an area influenced by the Santo Antonio do Jari hydroelectric system (Amapa State Brazil) The purpose was to study aspects ofAmazonGuiananAmerican cutaneous leishmaniasis (ACL) vectors subjected to stresses by anthropogenic environmental changesFor sampling CDC light traps were positioned 05 1 and 20m above ground at five capture locations along the Jari River BasinFluctuations in phlebotomine numbers were analyzed to determine any correlation with rainfall dam waterlogging andor ACLcases fromMay 2012 toMarch 2015 We captured 2800 individuals and among 45 species identified Bichromomyia flaviscutellataNyssomyia umbratilis and Psychodopygus squamiventris sl were determined to be the main putative vectors based on currentknowledge of the AmazonGuianan ACL scenario Rainfall but not complete flooding was relatively correlated with phlebotominefluctuation mainly observed for Ps squamiventris sl as were ACL cases with Ny umbratilis Behavioral changes were observed inthe unexpected high frequency of Bi flaviscutellata among CDC captures and the noncanopy dominance ofNy umbratilis possiblyattributable to environmental stress in the sampled ecotopes Continuous entomological surveillance is necessary to monitor theoutcomes of these findings
1 Introduction
Insects of Phlebotominae (Diptera Psychodidae) species areof great medical interest because of their implications inthe transmission of some vector-borne diseases in partic-ular leishmaniasis [1ndash3] American cutaneous leishmaniasis(ACL) is known to be endemic in Amazonian Brazil and isregarded as a serious risk to public health The high biodi-versity of the region induces a complex mosaic of ecologicalinteractions among reservoirs vectors and parasites andconsequently awide spectrumofACL clinicalmanifestationsincluding disfiguring and potentially life-threatening cases
[4] Moreover this diversity of transmission cycles becomeseven more complex to understand in the context of theinfluence of anthropic effects on natural ecological systemsThe consequences of anthropogenic environmental stress onthe expansion of the geographical distribution of ACL aretherefore difficult to predict with any certainty [5]
Over the past 30 years one of the main causes of humaninfluence on the environment in the Brazilian Amazon Basinis its exploitation for generating energy from hydropowerNegative effects of some of the resultant environmentalchanges have been observed an example is the disturbancecaused by the proliferation ofMansonia (Diptera Culicidae)
Hindawi Publishing CorporationJournal of Tropical MedicineVolume 2016 Article ID 9819723 12 pageshttpdxdoiorg10115520169819723
2 Journal of Tropical Medicine
in the dammed lake associatedwith theTucuruı hydroelectricsystem [6] Vector monitoring programs which are essentialto being granted a license to operate are included in environ-mental and public health planning in order to predict andminimize the effects of vector-borne diseases in hydroelectricsystems [7] This monitoring includes spatialtemporal stud-ies of phlebotomines and their potential implications for thetransmission dynamics of leishmaniasis agents [8]
The geographically closely related Brazilian states of Para(PA) and Amapa (AP) share a border through the JariRiver and cover a region historically associated with theexploitation and processing of cellulose Early informationabout leishmaniasis in the region first emerged in the 1970ndash1980s In this period studies conducted in ldquoMonte Douradordquo(Almeirim PA) provided substantial knowledge about ACLecoepidemiology with particular reference to the ldquopian-boisrdquo a form of cutaneous leishmaniasis caused by Leishma-nia (Viannia) guyanensis (formerly referred to as Leishmaniabraziliensis guyanensis) and its respective identity-obscuredvector Nyssomyia umbratilis [9ndash16]
Since those original studies were undertaken the regionalso now hosts the Santo Antonio do Jari hydroelectricsystem The present study therefore aimed to characterizephlebotomine fauna in the area in order to assess thepotential impact of the Santo Antonio do Jari hydroelectricsystem and in particular the spatiotemporal implications ofenvironmental changes on the ecology of ACL vectors in thatregion
2 Materials and Methods
21 Study Area The Santo Antonio do Jari hydroelectricsystem (0∘39101584002410158401015840S 52∘30101584056910158401015840W) is situated between themunicipalities of Almeirim (PA) and Laranjal do Jari (AP) aregion within the Guianan Ecoregion Complex The hydro-electric system comprises 3734 MW of installed potency anda reservoir area of 317 km2 Its construction began in 2011and the system was totally operative from 2014
The Amazon Region has a ldquohot-humidrdquo equatorial cli-mate however only minor differences are observed in thedistinct ecoregions In this study four Guianan EcoregionComplex seasons were characterized as follows a short rainy(wet) season from mid-November (ldquonovembre humiderdquo) tolate January a short dry season between early February andmid-March a long rainy season from late March to late Julyand a long dry season from late July to mid-November [17]
22 Sampled Locations Five locations each with the follow-ing characteristics were sampled up- and downstream of thecaisson (Figure 1)
The eastern bank of the Jari River (AP) covered by alluvialdense forest
(I) Porto Sabao (S 00∘37101584002210158401015840 W 052∘30101584042110158401015840) a for-ested area with only a few dwelling A small harboroperates from the opposite side of the river
(II) Iratapuru Village (S 00∘33101584050910158401015840 W 052∘34101584048510158401015840) acluster of dwellings forming a community of approx-imately 200 inhabitants which primarily acts as anextractive reserve
(III) SantoAntonioVillage (S 00∘38101584057410158401015840W052∘30101584027910158401015840)which is the only site downstreamof the caisson com-prising 27 agriculturist families inhabiting riverinewooden buildingsThe western bank of the Jari River (PA) covered bysubmontane (lowland) dense forest
(IV) Traıra (S 00∘37101584050710158401015840W052∘31101584048810158401015840) a locationwithno dwellings on the Traıra river
(V) the power Plant (S 00∘39101584007710158401015840 W 052∘31101584006410158401015840) themost affected area close to the construction site andwith very little vegetation
23 Sampling Phlebotomine captures were undertaken dur-ing 2012 to 2015 in fourteen quadrimonthly field expedi-tions which aimed to sample periods in the short dry(FebruaryMarch) rainy to dry (JuneJuly) and dry to rainy(OctoberNovember) AmazonGuianan seasons Eight expe-ditions were undertaken before and six after dammed lakelevels were replenished (in May 2014) In each of the abovedescribed five areas a randomly selected horizontal transectwas established in the direction of the primarily forested areacomprising a phlebotomine monitoring station (MS) sitedapproximately 200m distance from the forest edge wherehuman settlementswere present EachMS incorporated threeCDC miniature light traps (Sudia and Chamberlain model)installed between 1800 and 0600 for two consecutive nightsEach was placed according to the following different verticalstrata 05m from ground level 1m from ground level andat 20m in the tree canopy they were placed 10m apartfrom each other Trees used for canopy placement of CDCtraps were selected based on a minimum height of 20mregardless of speciesThe ground level surface was composedof humid decomposed organic material originating fromthe trees Phlebotomines were triaged in a stereomicroscopecleaned with neutral detergent [1 (Extran)] washed withNaCl 09 and then stored in 70 ethanol In the laboratoryspecimens were processed according to the methodologyof Ryan [18] mounted in Berlese fluid (GBI LaboratoriesManchester England) and identified according toGalati [19]
24 Data Analysis Sampling effort was calculated basedon number of expeditions versus the number of installedCDC traps versus hours of exposition Phlebotomine fre-quency was analyzed for correlation with climatic data(monthly average rainfall temperature and air relativehumidity) sourced from the Jari Meteorological Station (ldquoJariEnergiardquo database) and with epidemiology (ACL monthlyprevalence) obtained after interrogating the database of theldquoCoordenacao de Vigilancia em Sauderdquo (Coordination ofSurveillance in Health) of the ldquoSecretaria de Estado da Saudedo Amapa-SESArdquo (Amapa State Health Secretary) Data weretested by a single linear regression with Pearsonrsquos coefficient(r) using BioEstat 50 where the r-value represents a strongcorrelation if above 07 a moderate correlation if between03 and 07 a weak correlation if between 01 and 03and an insignificant correlation if below 01 The index ofspecies abundance (ISA) and the standard index of speciesabundance (SISA) were calculated using Microsoft Excel for
Journal of Tropical Medicine 3
Atlantic Ocean
Brazil
PacificOcean
Atlantic Ocean
Amapaacute State(Brazil)
French Guiana
Paraacute State(Brazil)
IratapuruVillage
Traiacutera River
Power Plant
PortoSabatildeo
Santo AntocircnioVillage
Jari River
Almeirim(Paraacute State)
Laranjal do Jari(Amapaacute State)
2000 km 200 km
2 km
N N
N
Figure 1 Study area Maps showing the five surveyed locations for phlebotomine captures (2012ndash2015) in Santo Antonio do Jari hydroelectricsystem impacted area between the states of Para and Amapa BrazilM locations
the five ecotopes surveyed as described by Roberts and Hsi[20] Species diversity (Shannonrsquos diversity index H1015840) andthe coefficient of variation (S1015840) between monthly records(climatic and epidemiological) were calculated using Pastsoftware [21] All comparisons were performed using a t-testto determine significance level (119901 value le 005)
3 Results
A total of 2800 individuals were captured (1894 femalesand 906 males) belonging to 45 species in approximately
2520 h of sampling effort Females of Trichophoromyia(exceptionally Th ubiquitalis) and Pressatia (not identifiableat species level) and some unidentified Evandromyia Nys-somyia and Psychodopygus species were allocated at genericlevel (Table 1)
New AP records includedTh castanheirai Th eurypygaEvandromyia begonae Ev bacula Pressatia equatorialis andPsathyromyia punctigeniculata
The five most frequent species captured were Sciopemyiasordellii (119899 = 316 SISA = 087) Ps davisi (119899 = 314 SISA =074) Bichromomyia flaviscutellata (119899 = 265 SISA= 095) Ev
4 Journal of Tropical MedicineTa
ble1Ph
ebotom
inespeciescompo
sitionofSantoAnton
iodo
Jarihydroelectric
syste
mim
pacted
area
(AmapaS
tateA
mazon
ianBrazil)
ontheb
asisoffivesurveyedlocatio
ns(2012ndash2015)
Species
PortoSabao
Traıra
River
Iratapuru
Santo
Power
Plant
ISA
SISA
Rank
Total
Village
Anton
ioVillage
CD
CD
CD
CD
CD
1Sciopemyiasordellii
149
6613
1015
442
16mdash
136
087
2nd
316
1129
2Psycho
dopygu
sdavisi
4637
9556
1510
3817
mdashmdash
64
074
3rd
314
1121
mdashTrich
ophoromyiaspp
177
mdash34
mdash40
mdash17
mdashmdash
mdashmdash
mdashmdash
268
957
3Bichromom
yiafla
viscutellata
7632
2414
2211
5231
12
2095
1st
265
946
4Evandrom
yiainfra
spinosa
4410
2518
32
3018
mdashmdash
9206
6th
150
536
5Ny
ssom
yiaum
bratilis
2612
3130
196
139
mdashmdash
86
063
5th
146
521
6Trich
ophoromyiacasta
nheir
aimdash
104
mdash6
mdash14
mdash11
mdashmdash
112
051
9th
135
482
mdashPressatia
spp
43mdash
1mdash
9mdash
78mdash
mdashmdash
mdashmdash
mdash131
468
mdashEvandrom
yiaspp
5210
4mdash
3911
12mdash
mdashmdash
mdashmdash
mdash128
457
7Viannamyiafurcata
444
115
111
305
mdashmdash
104
055
7th
111
396
8Psycho
dopygu
ssqu
amiventrissensulato
3614
155
10mdash
9mdash
mdashmdash
11052
8th
89318
9Trich
opygom
yiatrichopyga
1116
98
9mdash
2112
mdashmdash
7207
4th
86307
10Tricho
phorom
yiaub
iquitalis
216
58
148
139
2mdash
114
05
10th
86307
11Evandrom
yiasaulensis
409
5mdash
44
121
mdashmdash
128
043
11th
75268
12Psycho
dopygu
shirs
utus
hirsutus
13mdash
204
144
mdashmdash
mdashmdash
136
04
12th
5519
613
Trich
ophoromyiabrachypyga
mdash36
mdash12
mdash2
mdash2
mdashmdash
152
032
14th
5218
614
Pressatia
choti
mdash17
mdash3
mdash5
mdash25
mdashmdash
142
037
13th
5017
915
Nyssom
yiaan
duzei
65
2410
mdashmdash
mdashmdash
mdashmdash
17023
16th
4516
116
Lutzom
yiagomezi
2711
mdashmdash
mdashmdash
mdashmdash
mdashmdash
194
012
22th
3813
6mdash
Nysso
myiaspp
mdashmdash
mdash1
1mdash
1125
mdashmdash
mdashmdash
mdash38
136
17Ny
ssom
yiaan
tunesi
mdash2
9mdash
52
mdashmdash
1mdash
142
037
13th
19068
18Sciopemyiaflu
viatilis
21
3mdash
91
mdashmdash
mdashmdash
18019
18th
16057
19Micropygom
yiarorotaensis
4mdash
mdashmdash
8mdash
21
mdashmdash
178
02
17th
15054
20Psycho
dopygu
sparaensis
mdashmdash
mdashmdash
63
51
mdashmdash
176
02
17th
15054
21Psychodopygusclaustre
i7
1mdash
mdashmdash
mdash2
mdash3
1154
031
15th
14050
mdashPsychodopygusspp
4
mdash2
mdash6
mdash2
mdashmdash
mdashmdash
mdashmdash
14050
22Trich
ophoromyiaeurypyga
mdash8
mdashmdash
mdash3
mdash1
mdashmdash
182
018
19th
12043
23Psathyromyiaaragaoi
mdashmdash
52
mdashmdash
4mdash
mdashmdash
188
015
20th
11039
24Evandrom
yiabrachyphalla
43
21
1mdash
mdashmdash
mdashmdash
19014
21th
11039
25Psychodopyguscarreraicarrerai
11
mdash7
mdashmdash
mdashmdash
mdashmdash
202
008
26th
9032
26Evandrom
yiabegona
e2
2mdash
mdash4
mdashmdash
mdashmdash
mdash196
011
23th
8029
27Ny
ssom
yiawh
itman
imdash
mdashmdash
mdashmdash
mdash8
mdashmdash
mdash20
009
25th
8029
28Psathyromyiascaffi
32
mdashmdash
2mdash
mdashmdash
mdashmdash
198
01
24th
7025
29Pressatia
trispinosa
mdashmdash
mdash7
mdashmdash
mdashmdash
mdashmdash
204
007
27th
7025
30Micropygom
yiatrinidadensis
7mdash
mdashmdash
mdashmdash
mdashmdash
mdashmdash
208
005
29th
7025
31Viannamyiatuberculata
6mdash
mdashmdash
mdashmdash
mdashmdash
mdashmdash
21004
30th
6021
32Pintom
yiadamascenoi
mdashmdash
mdashmdash
mdashmdash
5mdash
mdashmdash
20009
25th
5018
33Evandrom
yiaevandroi
mdashmdash
mdashmdash
41
mdashmdash
mdashmdash
204
007
27th
5018
34Psathyromyialutziana
5mdash
mdashmdash
mdashmdash
mdashmdash
mdashmdash
212
003
31th
5018
35Brum
ptom
iatra
vasso
simdash
mdashmdash
mdashmdash
mdash3
1mdash
mdash196
011
23th
4014
36Evandrom
yiabacula
mdashmdash
mdashmdash
1mdash
3mdash
mdashmdash
202
008
26th
4014
37Evandrom
yiamonstruosa
mdashmdash
4mdash
mdashmdash
mdashmdash
mdashmdash
206
006
28th
4014
Journal of Tropical Medicine 5
Table1Con
tinued
Species
PortoSabao
Traıra
River
Iratapuru
Santo
Power
Plant
ISA
SISA
Rank
Total
Village
Anton
ioVillage
CD
CD
CD
CD
CD
38Psychodopygusa
mazonensis
mdashmdash
mdashmdash
22
mdashmdash
mdashmdash
21004
30th
4014
39Psychodopyguscorossoniensis
mdashmdash
mdashmdash
mdash2
mdashmdash
mdashmdash
206
006
28th
2007
40Pressatia
equatoria
lismdash
2mdash
mdashmdash
mdashmdash
mdashmdash
mdash21
004
30th
2007
41Psathyromyiainflata
mdashmdash
mdashmdash
mdashmdash
2mdash
mdashmdash
21004
30th
2007
42Micropygom
yiamicropyga
mdashmdash
mdashmdash
mdashmdash
11
mdashmdash
214
002
32th
2007
43Psychodopygusgenicu
latus
mdashmdash
mdash2
mdashmdash
mdashmdash
mdashmdash
218
033th
2007
44Lu
tzom
yiacarvalhoi
mdashmdash
1mdash
mdashmdash
mdashmdash
mdashmdash
208
005
29th
1004
45Psathyromyiapunctigenicu
lata
mdashmdash
mdashmdash
1mdash
mdashmdash
mdashmdash
216
001
32th
1004
Total
856
411
342
209
274
96415
186
74
2800
1000
1267
551
370
601
11Ea
chlocatio
ncomprise
dam
onito
ringstation(M
S)sampled
with
051and20
mCD
Ctrapsprovidingapproxim
ately
504h
MS
Cfem
aleDm
aleISAind
exof
speciesa
bund
anceSISAstand
ardindexof
speciesa
bund
ance
ldquothe
bold
italic
fontrdquoP
rovenandor
putativ
eACL
vectorsintheA
mazon
Region
based
inYo
ungandDun
can[1]Ra
ngelandLainson[4]Re
ady[2]andLainsonandShaw
[16]
6 Journal of Tropical Medicine
Table 2 Vertical stratification of Phebotomine species composition of Santo Antonio do Jari hydroelectric system impacted area (AmapaState Amazonian Brazil) on the basis of five surveyed locations (2012ndash2015)
Species 05m ground 1m ground 20m canopy TotalC D C D C D
1 Sciopemyia sordellii 117 71 91 23 11 3 3162 Psychodopygus davisi 48 27 44 39 102 54 314mdash Trichophoromyia spp 117 mdash 127 mdash 24 mdash 2683 Bichromomyia flaviscutellata 85 45 79 41 11 4 2654 Evandromyia infraspinosa 38 18 48 27 16 3 1505 Nyssomyia umbratilis 35 23 26 13 28 21 1466 Trichophoromyia castanheirai mdash 50 mdash 59 mdash 26 135mdash Pressatia spp 66 mdash 39 mdash 26 mdash 131mdash Evandromyia spp 55 12 41 9 11 mdash 1287 Vianamyia furcata 42 8 46 6 8 1 1118 Psychodopygus squamiventris sl 27 11 20 1 23 7 899 Trichopygomyia trichopyga 18 20 10 5 22 11 8610 Trichophoromyia ubiquitalis 28 15 16 13 9 5 86mdash Other species (11ndash45) 137 112 129 69 71 57 575
Total 813 412 716 305 362 192 2800Total (C + D) 1225 1021 554 mdash
Each stratum was sampled with approximately 840 h of CDC trapC female D male
infraspinosa (119899 = 150 SISA = 06) and Ny umbratilis (119899 =146 SISA = 063) Less frequently captured but nonethelessepidemiologically relevant species included Ps squamiventrissl (119899 = 89 SISA = 052) Th ubiquitalis (119899 = 86 SISA=05)Ps hirsutus hirsutus (119899 = 55 SISA = 04)Ny anduzei (119899 = 45SISA = 023) Lutzomyia gomezi (119899 = 38 SISA = 012) Nyantunesi (119899 = 19 SISA = 037) Ps paraensis (119899 = 15 SISA =02) and Ny whitmani (119899 = 8 SISA = 009)
Sampling effort for each area comprised approximately504 h of CDC trapping (14 expeditions 3 CDC traps and12-hour CDC exposition) Porto Sabao was the site of themost captured specimens (1267) followed by Santo AntonioVillage (601) The highest diversity was observed in capturesfrom Iratapuru Village (1198671015840 = 3027) All 1198671015840 value com-parisons between the different environments were significant(119901 le 005) except when comparing Traıra River and thePower Plant (119901 le 0099) (Figure 2)
Regarding vertical distribution sampling of the samecaptures described above accounted for approximately 840 hof CDC trapping by stratum (14 expeditions 5 ecotopes and12-hour CDC exposition) CDC traps placed 05m aboveground level accounted for a greater number of individuals(1225) than those placed 1m above ground level (1021) and20m above ground level in the canopy (554) (Table 2) WhilePs davisi was most likely to be found at canopy level (75individuals at 05m 83 at 1m and 156 at 20m) and Biflaviscutellata closer to ground level (130 at 05m 120 at 1mand 15 at 20m) there was little difference in numbers of Nyumbratilis individuals captured at the different levels of strata(58 at 05m 39 at 1m and 49 at 20m)
The monthly average rainfall was the only climatic datawith a representative coefficient of variance (1198781015840 = 7081)whentested against ACL occurrence and phlebotomine frequency
005
115
225
335
PortoSabatildeo
TraiacuteraRiver
IratapuruVillage
Santo AntocircnioVillage
PowerPlant
p = 0009p = 0099
p = 0007
p = 0012p = 0001
p = 0031
p = 0001p = 0004
p = 0001p = 00012855
26543027
2819
1468
Figure 2 Comparison of Shannon diversity indexes for CDC-captured phlebotomines from the five surveyed locations in SantoAntonio do Jari hydroelectric system impacted area between thestates of Para and Amapa Brazil (2012ndash2015)
(1198781015840 = 6595) in contrast to temperature (1198781015840 = 596)and relative air humidity (1198781015840 = 956) The available dataenabled correlations to be determined between the periodsof May 2012 to April 2015 a period within which nine of thefourteen capture efforts were undertaken Rainfall presenteda moderate correlation with overall phlebotomine frequency(119903 = 065 119901 = 000001) as also individually compared withPs squamiventris sl (119903 = 065 119901 = 000001) and a similarlymoderate but lower correlation with Bi flaviscutellata (119903 =048 119901 = 00021) and Ny umbratilis (119903 = 048 119901 = 0002)
Journal of Tropical Medicine 7
A decrease in the overall phlebotomine population wasobserved after March until November 2014 coinciding withthe transition between the rainy and dry periods and withflooding (Figure 3)
The number of ACL cases presented a moderate cor-relation with overall phlebotomine frequency (119903 = 044119901 = 00063) and also with Ps squamiventris (119903 = 032119901 = 00003) and an insignificant negative correlation with Biflaviscutellata (119903 = minus002 119901 = 00001) A similarly moderatebut higher correlation was however found withNy umbratilis(119903 = 048 119901 = 0002) (Figure 4)
4 Discussion
It is important to consider current investigations of phle-botomine fauna in the Jari River Basin in the context ofthe historical findings from the 1970s and 1980s Theseearlier studies represent comprehensive research conductedin Monte Dourado (northern PA) a region close to thearea investigated in the present study and on which thepresent epidemiological background to ACL transmissionin the associated region is based In brief the previousresearch includes a description of Ny umbratilis (formerlyldquoobscuredrdquo as Ny anduzei) [13] and its incrimination as themain vector of L (V) guyanensis [9ndash11] confirmed by strongecological associations [15] Just as important a parasitemore recently described as L (V) naiffi was isolated fromthe armadilloDasypus novemcinctus for which transmissionwas locally driven and attributed to Ps squamiventris sl andother Psychodopygus species [22] this comprised a primarilyenzootic ACL occasionally infecting humans [23] Finallythe isolation of L (L) amazonensis from rodents in an areaaffected by occurrence of the main vector Bi flaviscutellata[11] comprised the third known Jari ACL transmission cycleThese facts described above suggest that Ny umbratilis Pssquamiventris sl and Bi flaviscutellata are implicated in thetransmission scenario of the area covered by the present studyand thus merit particular attention
The present study incorporated a representative compo-sition of the known AP fauna Considering the substantialsampling effort (2520 h) the determined diversity showedagreement with the findings of other inventories from theBrazilian Amazon basin [1 24] Moreover all new records forAP have already been recorded for PA [25]
Apart from Ny umbratilis Ps squamiventris sl and Biflaviscutellata other species captured were noted as beingepidemiologically relevant to ACL in the Brazilian AmazonRegion Ps paraensis Ps davisi and Ps hirsutus hirsutuswhich are putative vectors of L (V) naiffi [26 27] Thubiquitalis a proven vector of L (V) lainsoni [28] Nyanduzei a secondary vector of L (V) guyanensis [4] Nywhitmani and Lu gomezi proven and putative vectors of L(V) shawi respectively [4 26] and lastly Ny antunesi asuspected vector of L (V) lindenbergi [4]
Among the above species Ps davisi is notable as thethird highest ranking according to SISA (074) and as havingthe highest presence in all vertical strata in particular atthe canopy level This species is generally anthropophilic iswidely distributed in the Amazon Basin [1 24] and is related
to Leishmania (Viannia) infections in the States of Rondonia(RO) [29] and PA [27 30] Interestingly this species remainsfrequent in recently surveyed areas affected by hydroelectricschemes in RO [8] and PA (TVS unpublished) Theseobservations add weight to the need for further investigationinto the putative epidemiological role of Ps davisi in Jari
The species most frequently captured Sc sordellii wouldappear not to be of epidemiological concern Firstly theapparent abundance of a given phlebotomine species is notsufficient of itself to incriminate it as a vector [31] particularlywhen it does not demonstrate high levels of anthropophilicbehavior Secondly flagellate infections isolated from thisspecies (formerly referred to as Lutzomyia nordestina) arehistorically recognized as distinct from Leishmania [22] orremain undetermined [32] and are more likely to be othertrypanosomatids [22] Furthermore despite recent reports ofmolecular Leishmania ascribed to this fly [33 34] currentlypolymerase chain reaction-based techniques do not allowldquotrue infectionrdquo to be distinguished from a ldquosimple and occa-sionally ingestedrdquo DNA fragment [3] Thus there remainsinsufficient evidence for a potential vector role of S sordelliiin Leishmania transmission
High frequencies of light-trapped Bi flaviscutellata havebeen documented [8 35] despite rodent-baited trappingbeing better recognized as amethod for attracting the speciesAdditionally the first SISA Rank (095) was indicative ofthis high presence in all five surveyed ecotopes this hasbeen observed in the Belem metropolitan area (PA) whereBi flaviscutellata was the only omnipresent species in sevenurban and ecologically isolated forest fragments surveyed[36] Furthermore this species has been reported as beingable to adapt to plantations of introduced tree species and inother nonclimax forests in Jari [14] suggesting it is capableof responding positively to environmental change such asthe vegetal suppression and flooding caused by hydroelectricoperations Nonetheless ecological niche model projectionspredict no change in spatial distribution for Bi flaviscutellatain the Jari area which is an environment that remains suitablefor the species [37]
In an area in Regina French Guiana ecologically dis-turbed due to agricultural practices flagellates infecting a Biflaviscutellata specimen have been determined to be L (V)guyanensis [38] in similar circumstances to the observed lowpresence of Ny umbratilis Although this possibly changingpattern in ACL transmission should be interpreted withcaution it raises an intriguing possibility about the permis-siveness of Bi flaviscutellata to L (V) guyanensis in theAmazonGuianan ACL scenario
In the present study a higher number of Bi flaviscutellataindividuals was captured compared to Ny umbratilis (265versus 146 resp) A lack of available evidence for naturalinfection in this unnatural vectorparasite system howeverprecludes speculation about the possible involvement of Biflaviscutellata even in the context of occasional transmissionof parasites distinct from L (L) amazonensis Neverthelessthe current high occurrence of Bi flaviscutellata in theAmazonGuianan ACL scenario may be of future concernunnatural parasitevectorreservoir systems when closelyrelated in an ecological context can on a century-level scale
8 Journal of Tropical Medicine
010203040506070
Bi flaviscutellata
01020304050607080
Ny umbratilis
0510152025303540
Ps squamiventris sl
0100200300400500600
050
100150200250300350400
2012 2013 2014 2015
Wat
erflo
odin
g
Ps s
quya
miv
entri
s sl
Bi fl
avisc
utell
ata
Ny u
mbr
atili
s
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
050
100150200250300350400
050
100150200250300350400
050
100150200250300350400
010203040506070
10000 20000 30000 40000000Rainfall (mm)
01020304050607080
10000 20000 30000 40000000Rainfall (mm)
Rainfall (mm)Overall
Rainfall (mm)
Rainfall (mm)
Rainfall (mm)
0100200300400500600
Ove
rall
10000 20000 30000 40000000Rainfall (mm)
05
10152025303540
10000 20000 30000 40000000Rainfall (mm)
r = 065 p = 000001
r = 048 p = 00021
r = 049 p = 00021
r = 065 p = 000001
Figure 3 Correlation between rainfall and phlebotomine putative vector species captured in Santo Antonio do Jari hydroelectric systemimpacted area between the states of Para and Amapa Brazil (2012ndash2015)
Journal of Tropical Medicine 9
2012 2013 2014 2015
Wat
erflo
odin
g
0100200300400500600
010203040506070
Bi flaviscutellata
01020304050607080
Ny umbratilis
0510152025303540
Ps squamiventris sl
Bi fl
avisc
utell
ata
05
1015202530
05
1015202530
05
1015202530
05
1015202530
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
0100200300400500600
Ove
rall
10 20 300ACL cases
010203040506070
10 20 300ACL cases
01020304050607080
Ny u
mbr
atili
s
10 20 300ACL cases
ACL casesOverall
ACL cases
ACL cases
ACL cases
05
10152025303540
Ps s
quam
iven
tris s
l
10 20 300ACL cases
r = 044 p = 00063
r = minus002 p = 00001
r = 048 p = 0002
r = 032 p = 00003
Figure 4 Correlation between ACL cases and phlebotomine putative vector species captured in Santo Antonio do Jari hydroelectric systemimpacted area between the states of Para and Amapa Brazil (2012ndash2015)
10 Journal of Tropical Medicine
undergo stress adaptation (evolutionary fitness) and formnew ecological systems driven by coevolution [39]
There is no doubt that Ny umbratilis is the main vectorof ACL in the Jari River Basin [15] Surprisingly the well-documented tree-dwelling behavior of this species [15 40]was not observed in CDC canopy captures in the presentstudy very little difference was found between the number ofindividuals captured in the three sampled strata (58 at 05m39 at 1m and 49 at 20m) Although Ny umbratilis is knownto climb into the canopy at night to feed on edentates (mainlysloths) [12 41] it is possible that an alternative terrestrialsource of blood such as rodents may be attracting femalestoward ground level Sloths are very slow moving and aretherefore commonly rescued for relocation during environ-mentalmonitoring operations their resulting rarenessmeansNy umbratilis has had to find other food sources Converselyblood meal analysis conducted during studies in a forestfragment of the Western Amazon has shown rodent bloodto be the predominant feeding source for Ny umbratilis [42]Wild and synanthropic rodents have been shown to surviveenvironmental disturbance in Jari and to proliferate in areasof introduced tree species [14] this indicated a plausiblereason not only for the putative adaptation of Ny umbratilisbut also for the increasing population levels of the rodentblood-feeder Bi flaviscutellata as already discussed
Given the clear importance of Psychodopygus of theChagasi series (formerly referred to as the Squamiventrisseries) in the context of ACL transmission [22] taxonomicstudies on identifying the cryptic females have been under-taken previously [43] and more recently with interestingresults (Rodrigo E Godoy unpublished) These techniqueshowever could not be employed in the field at the time Earlyconsideration of the assumed geographic distribution of thecomplex Ps squamiventris sl [1 44] shows that individualsfrom AP are likely to be predominantly Ps s maripaensisDetermining males of the species present could clarify thesetaxonomic uncertainties in the area covered by the presentstudy however doubts about identification of some malespecimens meant they were grouped as Ps squamiventris sl
Ryan et al [22] did not find Ps squamiventris sl to benaturally infected in the Jari area but proposed a link to L(V) naiffi along the same northern bank of the AmazonRiver through studies in Cachoeira Porteira (PA) Fouqueet al [38] came to the same conclusions in French GuianaInterestingly this fly species complex was found alongside L(V) braziliensis DNA in Suriname [45] an area where theknown Amazonian vectors of this parasite Ps complexus andPs wellcomei are unlikely to occur As previously discussedin the context of Bi flaviscutellata the changing transmissionpatterns of unnatural vectorparasite systems must be care-fully interpreted and only after strong ecological associationshave been determined Thus the putative vector importanceof Ps squamiventris sl in Jari can only be assumed in respectof L (V) naiffi
The five surveyed areas presented in general differentdiversity indexes showing variations in the phlebotominecomposition along the affected area This finding can beattributed not only to the distance between capture sitesbut also to forest composition fragmentation geographic
barriers and migration of mammal fauna between sites thislatter can occur naturally or be induced by rescue opera-tions involved in hydroelectric planning whereby animalsare moved to suitable ldquodry landrdquo environments safe frompotential flooding
A decrease in the phlebotomine population in 2014 inthe period after filling the reservoir could not be attributedexclusively either to flooding or to the transition from therainy to the dry season Both eventualities although oppositein effect are likely to lead to unsuitable conditions forphlebotomine reproduction However they did coincide inMay 2014 when rainfall decreased and the lake was formed
Conversely rainfall was a valid determinant of overallphlebotomine fluctuation Ps squamiventris sl was the mainputative vector directly correlated with this Other variablessuch as forest composition and altitudinal variation areknown to influence seasonal dynamics of species from thecomplex Ps squamiventris sl For example the dry periodfrom October to November in Serra dos Carajas (southernPA) can be unsuitable for Ps wellcomei which is adapted tohigh altitudes in contrast to Ps complexus which flourishesin lowland areas during the same months [46]
An insignificant negative to a moderate positive correla-tion was observed between cases of ACL and phlebotominefluctuations In the case of Ny umbratilis however theapparently higher correlation when compared with otherspecies is in agreement with its known vector importance inAmazonGuianan ACL A strong spatiotemporal associationbetween a given vector and ACL cases is considered essentialto maintenance of the Leishmania life cycle [2 14]
In summary the present study filled a temporal gapon phlebotomine data relating to the Jari area It providesa meaningful update on species composition and furtherinsights into seasonal dynamics of species under pressurescaused by environmental change The presence of ACLvectors in particular Ny umbratilis Bi flaviscutellata andPs squamiventris sl is noteworthy due to their directassociation with AmazonGuianan-ACL transmission ForNy umbratilis the lower frequency associatedwith the unpre-dicted lack of tree-dwelling behavior may be attributableto a reduction in numbers of canopy-dwelling mammalsdue to the environmental disturbance and relocation effortsthis could consequently have led to seeking for alternativeterrestrial blood sources at ground level The possible asso-ciation of Ps davisi with Leishmania transmission cycles inother Amazon ecoregions draws attention to its high rankposition in Jari Rainfall can be partly associated with localseasonal fluctuations in fly numbers mainly with respect toPs squamiventris sl Cases of ACL were also more highlycorrelated with Ny umbratilis dynamics Further attentionwas also given to results for Bi flaviscutellata which hasbeen apparently tolerant of anthropic activity continuedphlebotominemonitoring is proposed in order to evaluate theimplication for social welfare and health in the future
Competing Interests
The authors declare that they have no competing interests
Journal of Tropical Medicine 11
Acknowledgments
Theauthorswish to thank the ldquoInstituto de Pesquisas Cientıfi-cas e Tecnologicas do Estado do Amapa IEPArdquo entomologyteam for the technical assistance given to the field work inparticular to Aderbal Amanajas Santana Rosalvo Dias deCastro Noel Fernandes Santos Neto Osita Cleyriane Lobatodo Nascimento Jose Luiz dos Santos Chaves and Joao doSocorro Gomes Campos Logistical facilities were financiallysupported by the ldquoJari Energiardquo
References
[1] D G Young and M A Duncan ldquoGuide to the identificationand geographic distribution of Lutzomyia sand flies in Mexicothe West Indies Central and South America (Diptera Psycho-didae)rdquo inMemoirs of the American Entomological Institute pp1ndash881 Associated Publishers American Entomological InstituteGainesville Fla USA 1994
[2] P D Ready ldquoBiology of phlebotomine sand flies as vectors ofdisease agentsrdquo Annual Review of Entomology vol 58 pp 227ndash250 2013
[3] R P Brazil A A Fuzzari and J D Andrade Filho ldquoSandfly vectors of Leishmania in the Americasmdasha mini reviewrdquoEntomology Ornithology amp Herpetology vol 4 no 2 Article ID1000144 2015
[4] E F Rangel and R Lainson ldquoProven and putative vectorsof American cutaneous leishmaniasis in Brazil aspects oftheir biology and vectorial competencerdquoMemorias do InstitutoOswaldo Cruz vol 104 no 7 pp 937ndash954 2009
[5] E F Rangel S M Costa and B M Carvalho ldquoEnvironmentalchanges and the geographic spreading of American cutaneousleishmaniasis in Brazilrdquo in Leishmaniasis-Trends in Epidemiol-ogy D Claborn Ed Diagnosis and Treatment InTech RijekaCroatia 2014
[6] W P Taddei ldquoO genero Mansonia (Diptera Culicidae) e aproliferacao de mosquitos na usina hidreletrica de Tucuruırdquo inEnergia na Amazonia S B Magalhaes R C Brito and E RCastro Eds vol 1 pp 311ndash318 MPEGFPAUNAMAZ BelemBrazil 1996 (Portuguese)
[7] A I A Oliveira Observacoes Sobre a Resolucao CONAMA no23797 que Dispoe Sobre o Licenciamento Ambiental MimeoConselho Empresarial Brasileiro Para o Desenvolvimento Sus-tentavel Rio de Janeiro Brazil 1997 (Portuguese)
[8] A K R Galardo C D Galardo G A Silveira et al ldquoPhle-botominae sand flies (Diptera Psychodidae) potential vectorsof American cutaneous leishmaniasis agents in the area asso-ciated with the Santo Antonio hydroelectric system in WesternAmazonian Brazilrdquo Revista da Sociedade Brasileira deMedicinaTropical vol 48 no 3 pp 265ndash271 2015
[9] R Lainson R DWard and J J Shaw ldquoCutaneous leishmaniasisin North Brazil Lutzomyia anduzei as a major vectorrdquo Transac-tions of the Royal Society of Tropical Medicine and Hygiene vol70 no 2 pp 171ndash172 1976
[10] R Lainson J J Shaw R D Ward P D Ready and R DNaiff ldquoLeishmaniasis in Brazil XIII Isolation of Leishmaniafrom armadillos (Dasypus novemcinctus) and observations onthe epidemiology of cutaneous leishmaniasis in north ParaStaterdquo Transactions of the Royal Society of Tropical Medicine andHygiene vol 73 no 2 pp 239ndash242 1979
[11] R Lainson J J Shaw P D Ready M A Miles and M PovoaldquoLeishmaniasis in Brazil XVI Isolation and identification of
Leishmania species from sandflies wild mammals and manin north Para State with particular reference to L braziliensisguyanensis causative agent of ldquopian-boisrdquordquo Transactions of theRoyal Society of Tropical Medicine and Hygiene vol 75 no 4pp 530ndash536 1981
[12] R Lainson J J Shaw and M Povoa ldquoThe importance of eden-tates (sloths and anteaters) as primary reservoirs of Leishmaniabraziliensis guyanensis causative agent of lsquopian boisrsquo in northBrazilrdquo Transactions of the Royal Society of Tropical Medicineand Hygiene vol 75 no 4 pp 611ndash612 1981
[13] R DWard and H Fraiha ldquoLutzomyia umbratilis a new speciesof sand fly from Brazil (Diptera Psychodidae)rdquo Journal ofMedical Entomology vol 14 no 3 pp 313ndash317 1977
[14] P D Ready R Lainson and J J Shaw ldquoLeishmaniasis in BrazilXX Prevalence of lsquoenzootic rodent leishmaniasisrsquo (Leishmaniamexicana amazonensis) and apparent absence of lsquopian boisrsquo(Le braziliensis guyanensis) in plantations of introduced treespecies and in other non-climax forests in eastern AmazoniardquoTransactions of the Royal Society of Tropical Medicine andHygiene vol 77 no 6 pp 775ndash785 1983
[15] P D Ready R Lainson J J Shaw and R DWard ldquoThe ecologyof Lutzomyia umbratilisWard amp Fraiha (Diptera Psychodidae)the major vector to man of Leishmania braziliensis guyanensisin north-eastern Amazonian Brazilrdquo Bulletin of EntomologicalResearch vol 76 no 1 pp 21ndash40 1986
[16] R Lainson and J J Shaw ldquoLeishmania (Viannia) naiffi spn a parasite of the armadillo Dasypus novemcinctus (L)in Amazonian Brazilrdquo Annales de Parasitologie Humaine etComparee vol 64 no 1 pp 3ndash9 1989
[17] A Roger M Nacher M Hanf et al ldquoClimate and leishmaniasisin French guianardquo The American Journal of Tropical Medicineand Hygiene vol 89 no 3 pp 564ndash569 2013
[18] L Ryan ldquoFlebotomos do estado do Para Brasil (Diptera Psy-chodidae)rdquo Thecnical Document 1 Instituto Evandro ChagasBelem Brazil 1986 (Portuguese)
[19] E A B Galati ldquoMorfologia e Taxonomia Morfologia termi-nologia de adultos e identificacao dos taxons da Americardquoin Flebotomıneos do Brasil E F Rangel and R LainsonEds pp 53ndash75 Editora Fiocruz Rio de Janeiro Brazil 2003(Portuguese)
[20] D R Roberts and B P Hsi ldquoAn index of species abundance foruse with mosquito surveillance datardquo Environmental Entomol-ogy vol 8 no 6 pp 1007ndash1013 1979
[21] Oslash Hammer D A T Harper and P D Ryan ldquoPast paleontolog-ical statistics software package for education and data analysisrdquoPalaeontologia Electronica vol 4 no 1 pp 1ndash9 2001
[22] L Ryan R Lainson J J Shaw R R Braga and E A IshikawaldquoLeishmaniasis in Brazil XXV Sandfly vectors of Leishmania inPara State Brazilrdquo Medical and Veterinary Entomology vol 1no 4 pp 383ndash395 1987
[23] R Lainson J J Shaw F T Silveira R R Braga and E AIshikawa ldquoCutaneous leishmaniasis of man due to Leishmania(Viannia) naiffirdquo Annales de Parasitologie Humaine et Compa-ree vol 65 no 5-6 pp 282ndash284 1990
[24] E G C Bermudez Lutzomyia Sand Flies in the BrazilianAmazon Basin (Diptera Psychodidae) INPA Manaus Brazil2009
[25] G M Aguiar and W M Medeiros ldquoDistribuicao regionale habitats das especies de flebotomıneos do Brasilrdquo in Fle-botomıneos do Brasil E F Rangel and R Lainson Eds pp 207ndash256 Fiocruz Rio de Janeiro Brazil 2003 (Portuguese)
12 Journal of Tropical Medicine
[26] J R Arias M A Miles R D Naiff et al ldquoFlagellate infectionsof Brazilian sand flies (Diptera Psychodidae) isolation invitro and biochemical identification of Endotrypanum andLeishmaniardquo The American Journal of Tropical Medicine andHygiene vol 34 no 6 pp 1098ndash1108 1985
[27] A A Souza T V Santos Y L Jennings et al ldquoNaturalLeishmania(Viannia) spp infections in phlebotomine sand flies(Diptera Psychodidae) from the Brazilian Amazon regionreveal new putative transmission cycles of American cutaneousleishmaniasisrdquo Parasite vol 23 article 22 2016
[28] F T Silveira A A Souza R Lainson J J Shaw R R Bragaand E A Ishikawa ldquoCutaneous leishmaniasis in the AmazonRegion natural infection of the sand fly Lutzomyia ubiquitalis(Psychodidae Phlebotominae) by Leishmania (Viannia) lain-soni in Para state BrazilrdquoMemorias do Instituto Oswaldo Cruzvol 86 no 1 pp 127ndash130 1991
[29] LH S Gil S A Basano A A Souza et al ldquoRecent observationson the sand fly (Diptera Psychodidae) fauna of the state ofRondonia Western Amazonia Brazil the importance of Psych-dopygus davisi as a vector of zoonotic cutaneous leishmaniasisrdquoMemorias do Instituto Oswaldo Cruz vol 98 no 6 pp 751ndash7552003
[30] A A A Souza F T Silveira R Lainson et al ldquoThe Phlebotom-inae fauna of Serra dos Carajas Para Brazil and its possibleimplication in the transmission of American tegumentaryleishmaniasisrdquo Revista Pan-Amazonica de Saude vol 1 no 1pp 45ndash51 2010
[31] R Killick-Kendrick ldquoPhlebotomine vectors of the leishmani-ases a reviewrdquoMedical and Veterinary Entomology vol 4 no 1pp 1ndash24 1990
[32] M E C Dorval G Cristaldo H C da Rocha et al ldquoPhle-botomine fauna (Diptera Psychodidae) of an American cuta-neous leishmaniasis endemic area in the state of Mato Grossodo Sul Brazilrdquo Memorias do Instituto Oswaldo Cruz vol 104no 5 pp 695ndash702 2009
[33] V C F V Guimaraes P L Costa F J da Silva et al ldquoMoleculardetection of Leishmania in phlebotomine sand flies in a cuta-neous and visceral leishmaniasis endemic area in northeasternBrazilrdquo Revista do Instituto de Medicina Tropical de Sao Paulovol 56 no 4 pp 357ndash360 2014
[34] R S Lana E M Michalsky C L Fortes-Dias et al ldquoPhle-botomine sand fly fauna and Leishmania infection in thevicinity of the Serra do Cipo National Park a natural Brazilianheritage siterdquo BioMed Research International vol 2015 ArticleID 385493 9 pages 2015
[35] V R Alves R A de Freitas F L Santos A F J de OliveiraT V Barrett and P H F Shimabukuro ldquoSand flies (DipteraPsychodidae Phlebotominae) from central amazonia and fournew records for the Amazonas State Brazilrdquo Revista Brasileirade Entomologia vol 56 no 2 pp 220ndash227 2012
[36] J V S Ferreira T V Santos E M Santos and I S GorayebldquoPhlebotomine sand flies (Diptera Psychodidae) in forestfragments of Belem metropolitan area Para State Brazil withconsiderations on vectors of American cutaneous leishmaniasisagentsrdquo Revista Pan-Amazonica de Saude vol 5 no 2 pp 29ndash35 2014
[37] B M Carvalho E F Rangel P D Ready and M M ValeldquoEcological niche modelling predicts southward expansion ofLutzomyia (Nyssomyia) flaviscutellata (Diptera PsychodidaePhlebotominae) vector of Leishmania (Leishmania) amazonen-sis in South America under climate changerdquo PLoS ONE vol 10no 11 Article ID e0143282 2015
[38] F Fouque P Gaborit J Issaly et al ldquoPhlebotomine sand flies(Diptera Psychodidae) associatedwith changing patterns in thetransmission of the human cutaneous leishmaniasis in FrenchGuianardquoMemorias do Instituto Oswaldo Cruz vol 102 no 1 pp35ndash40 2007
[39] R Killick-Kendrick ldquoSome epidemiological consequences ofthe evolutionary fit between Leishmaniae and their phle-botomine vectorsrdquo Bulletin de la Societe de Pathologie Exotiqueet de ses Filiales vol 78 no 5 pp 747ndash755 1985
[40] F X Pajot F Le Pont BGentile andR Besnard ldquoEpidemiologyof leishmaniasis in French Guianardquo Transactions of the RoyalSociety of Tropical Medicine and Hygiene vol 76 no 1 pp 112ndash113 1982
[41] H A Christensen J R Arias A M De Vasquez and R ADe Freitas ldquoHosts of sandfly vectors of Leishmania braziliensisguyanensis in the central Amazon of Brazilrdquo The AmericanJournal of Tropical Medicine and Hygiene vol 31 no 2 pp 239ndash242 1982
[42] L C R Nery E S Lorosa andAM R Franco ldquoFeeding prefer-ence of the sand flies Lutzomyia umbratilis and L spathotrichia(Diptera Psychodidae Phlebotominae) in an urban forest patchin the City ofManaus Amazonas BrazilrdquoMemorias do InstitutoOswaldo Cruz vol 99 no 6 pp 571ndash574 2004
[43] PD Ready R Lainson J J Shaw andAA Souza ldquoDNAprobesfor distinguishing Psychodopygus wellcomei from Psychodopy-gus complexus (Diptera Psychodidae)rdquo Memorias do InstitutoOswaldo Cruz vol 86 no 1 pp 41ndash49 1991
[44] P D Ready H Fraiha R P Lane J R Arias and F X PajotldquoOn distinguishing the female of Psychodopygus wellcomei avector of mucocutaneous leishmaniasis from other squamiven-tris series females I Characterization of Ps squamiventrissquamiventris and Ps s maripaensis stat nov (Diptera Psycho-didae)rdquo Annals of Tropical Medicine and Parasitology vol 76no 2 pp 201ndash214 1982
[45] A D Kent T V dos Santos A Gangadin A Samjhawan DR A Mans and H D F H Schallig ldquoStudies on the sandfly fauna (Diptera Psychodidae) in high-transmission areas ofcutaneous leishmaniasis in the Republic of Surinamerdquo Parasitesand Vectors vol 6 no 1 article 318 2013
[46] P D Ready R Lainson and J J Shaw ldquoHabitat and seasonalityof Psychodopygus wellcomei help incriminate it as a vector ofLeishmania braziliensis in Amazonia and Northeast BrazilrdquoTransactions of the Royal Society of Tropical Medicine andHygiene vol 78 no 4 pp 543ndash544 1984
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

2 Journal of Tropical Medicine
in the dammed lake associatedwith theTucuruı hydroelectricsystem [6] Vector monitoring programs which are essentialto being granted a license to operate are included in environ-mental and public health planning in order to predict andminimize the effects of vector-borne diseases in hydroelectricsystems [7] This monitoring includes spatialtemporal stud-ies of phlebotomines and their potential implications for thetransmission dynamics of leishmaniasis agents [8]
The geographically closely related Brazilian states of Para(PA) and Amapa (AP) share a border through the JariRiver and cover a region historically associated with theexploitation and processing of cellulose Early informationabout leishmaniasis in the region first emerged in the 1970ndash1980s In this period studies conducted in ldquoMonte Douradordquo(Almeirim PA) provided substantial knowledge about ACLecoepidemiology with particular reference to the ldquopian-boisrdquo a form of cutaneous leishmaniasis caused by Leishma-nia (Viannia) guyanensis (formerly referred to as Leishmaniabraziliensis guyanensis) and its respective identity-obscuredvector Nyssomyia umbratilis [9ndash16]
Since those original studies were undertaken the regionalso now hosts the Santo Antonio do Jari hydroelectricsystem The present study therefore aimed to characterizephlebotomine fauna in the area in order to assess thepotential impact of the Santo Antonio do Jari hydroelectricsystem and in particular the spatiotemporal implications ofenvironmental changes on the ecology of ACL vectors in thatregion
2 Materials and Methods
21 Study Area The Santo Antonio do Jari hydroelectricsystem (0∘39101584002410158401015840S 52∘30101584056910158401015840W) is situated between themunicipalities of Almeirim (PA) and Laranjal do Jari (AP) aregion within the Guianan Ecoregion Complex The hydro-electric system comprises 3734 MW of installed potency anda reservoir area of 317 km2 Its construction began in 2011and the system was totally operative from 2014
The Amazon Region has a ldquohot-humidrdquo equatorial cli-mate however only minor differences are observed in thedistinct ecoregions In this study four Guianan EcoregionComplex seasons were characterized as follows a short rainy(wet) season from mid-November (ldquonovembre humiderdquo) tolate January a short dry season between early February andmid-March a long rainy season from late March to late Julyand a long dry season from late July to mid-November [17]
22 Sampled Locations Five locations each with the follow-ing characteristics were sampled up- and downstream of thecaisson (Figure 1)
The eastern bank of the Jari River (AP) covered by alluvialdense forest
(I) Porto Sabao (S 00∘37101584002210158401015840 W 052∘30101584042110158401015840) a for-ested area with only a few dwelling A small harboroperates from the opposite side of the river
(II) Iratapuru Village (S 00∘33101584050910158401015840 W 052∘34101584048510158401015840) acluster of dwellings forming a community of approx-imately 200 inhabitants which primarily acts as anextractive reserve
(III) SantoAntonioVillage (S 00∘38101584057410158401015840W052∘30101584027910158401015840)which is the only site downstreamof the caisson com-prising 27 agriculturist families inhabiting riverinewooden buildingsThe western bank of the Jari River (PA) covered bysubmontane (lowland) dense forest
(IV) Traıra (S 00∘37101584050710158401015840W052∘31101584048810158401015840) a locationwithno dwellings on the Traıra river
(V) the power Plant (S 00∘39101584007710158401015840 W 052∘31101584006410158401015840) themost affected area close to the construction site andwith very little vegetation
23 Sampling Phlebotomine captures were undertaken dur-ing 2012 to 2015 in fourteen quadrimonthly field expedi-tions which aimed to sample periods in the short dry(FebruaryMarch) rainy to dry (JuneJuly) and dry to rainy(OctoberNovember) AmazonGuianan seasons Eight expe-ditions were undertaken before and six after dammed lakelevels were replenished (in May 2014) In each of the abovedescribed five areas a randomly selected horizontal transectwas established in the direction of the primarily forested areacomprising a phlebotomine monitoring station (MS) sitedapproximately 200m distance from the forest edge wherehuman settlementswere present EachMS incorporated threeCDC miniature light traps (Sudia and Chamberlain model)installed between 1800 and 0600 for two consecutive nightsEach was placed according to the following different verticalstrata 05m from ground level 1m from ground level andat 20m in the tree canopy they were placed 10m apartfrom each other Trees used for canopy placement of CDCtraps were selected based on a minimum height of 20mregardless of speciesThe ground level surface was composedof humid decomposed organic material originating fromthe trees Phlebotomines were triaged in a stereomicroscopecleaned with neutral detergent [1 (Extran)] washed withNaCl 09 and then stored in 70 ethanol In the laboratoryspecimens were processed according to the methodologyof Ryan [18] mounted in Berlese fluid (GBI LaboratoriesManchester England) and identified according toGalati [19]
24 Data Analysis Sampling effort was calculated basedon number of expeditions versus the number of installedCDC traps versus hours of exposition Phlebotomine fre-quency was analyzed for correlation with climatic data(monthly average rainfall temperature and air relativehumidity) sourced from the Jari Meteorological Station (ldquoJariEnergiardquo database) and with epidemiology (ACL monthlyprevalence) obtained after interrogating the database of theldquoCoordenacao de Vigilancia em Sauderdquo (Coordination ofSurveillance in Health) of the ldquoSecretaria de Estado da Saudedo Amapa-SESArdquo (Amapa State Health Secretary) Data weretested by a single linear regression with Pearsonrsquos coefficient(r) using BioEstat 50 where the r-value represents a strongcorrelation if above 07 a moderate correlation if between03 and 07 a weak correlation if between 01 and 03and an insignificant correlation if below 01 The index ofspecies abundance (ISA) and the standard index of speciesabundance (SISA) were calculated using Microsoft Excel for
Journal of Tropical Medicine 3
Atlantic Ocean
Brazil
PacificOcean
Atlantic Ocean
Amapaacute State(Brazil)
French Guiana
Paraacute State(Brazil)
IratapuruVillage
Traiacutera River
Power Plant
PortoSabatildeo
Santo AntocircnioVillage
Jari River
Almeirim(Paraacute State)
Laranjal do Jari(Amapaacute State)
2000 km 200 km
2 km
N N
N
Figure 1 Study area Maps showing the five surveyed locations for phlebotomine captures (2012ndash2015) in Santo Antonio do Jari hydroelectricsystem impacted area between the states of Para and Amapa BrazilM locations
the five ecotopes surveyed as described by Roberts and Hsi[20] Species diversity (Shannonrsquos diversity index H1015840) andthe coefficient of variation (S1015840) between monthly records(climatic and epidemiological) were calculated using Pastsoftware [21] All comparisons were performed using a t-testto determine significance level (119901 value le 005)
3 Results
A total of 2800 individuals were captured (1894 femalesand 906 males) belonging to 45 species in approximately
2520 h of sampling effort Females of Trichophoromyia(exceptionally Th ubiquitalis) and Pressatia (not identifiableat species level) and some unidentified Evandromyia Nys-somyia and Psychodopygus species were allocated at genericlevel (Table 1)
New AP records includedTh castanheirai Th eurypygaEvandromyia begonae Ev bacula Pressatia equatorialis andPsathyromyia punctigeniculata
The five most frequent species captured were Sciopemyiasordellii (119899 = 316 SISA = 087) Ps davisi (119899 = 314 SISA =074) Bichromomyia flaviscutellata (119899 = 265 SISA= 095) Ev
4 Journal of Tropical MedicineTa
ble1Ph
ebotom
inespeciescompo
sitionofSantoAnton
iodo
Jarihydroelectric
syste
mim
pacted
area
(AmapaS
tateA
mazon
ianBrazil)
ontheb
asisoffivesurveyedlocatio
ns(2012ndash2015)
Species
PortoSabao
Traıra
River
Iratapuru
Santo
Power
Plant
ISA
SISA
Rank
Total
Village
Anton
ioVillage
CD
CD
CD
CD
CD
1Sciopemyiasordellii
149
6613
1015
442
16mdash
136
087
2nd
316
1129
2Psycho
dopygu
sdavisi
4637
9556
1510
3817
mdashmdash
64
074
3rd
314
1121
mdashTrich
ophoromyiaspp
177
mdash34
mdash40
mdash17
mdashmdash
mdashmdash
mdashmdash
268
957
3Bichromom
yiafla
viscutellata
7632
2414
2211
5231
12
2095
1st
265
946
4Evandrom
yiainfra
spinosa
4410
2518
32
3018
mdashmdash
9206
6th
150
536
5Ny
ssom
yiaum
bratilis
2612
3130
196
139
mdashmdash
86
063
5th
146
521
6Trich
ophoromyiacasta
nheir
aimdash
104
mdash6
mdash14
mdash11
mdashmdash
112
051
9th
135
482
mdashPressatia
spp
43mdash
1mdash
9mdash
78mdash
mdashmdash
mdashmdash
mdash131
468
mdashEvandrom
yiaspp
5210
4mdash
3911
12mdash
mdashmdash
mdashmdash
mdash128
457
7Viannamyiafurcata
444
115
111
305
mdashmdash
104
055
7th
111
396
8Psycho
dopygu
ssqu
amiventrissensulato
3614
155
10mdash
9mdash
mdashmdash
11052
8th
89318
9Trich
opygom
yiatrichopyga
1116
98
9mdash
2112
mdashmdash
7207
4th
86307
10Tricho
phorom
yiaub
iquitalis
216
58
148
139
2mdash
114
05
10th
86307
11Evandrom
yiasaulensis
409
5mdash
44
121
mdashmdash
128
043
11th
75268
12Psycho
dopygu
shirs
utus
hirsutus
13mdash
204
144
mdashmdash
mdashmdash
136
04
12th
5519
613
Trich
ophoromyiabrachypyga
mdash36
mdash12
mdash2
mdash2
mdashmdash
152
032
14th
5218
614
Pressatia
choti
mdash17
mdash3
mdash5
mdash25
mdashmdash
142
037
13th
5017
915
Nyssom
yiaan
duzei
65
2410
mdashmdash
mdashmdash
mdashmdash
17023
16th
4516
116
Lutzom
yiagomezi
2711
mdashmdash
mdashmdash
mdashmdash
mdashmdash
194
012
22th
3813
6mdash
Nysso
myiaspp
mdashmdash
mdash1
1mdash
1125
mdashmdash
mdashmdash
mdash38
136
17Ny
ssom
yiaan
tunesi
mdash2
9mdash
52
mdashmdash
1mdash
142
037
13th
19068
18Sciopemyiaflu
viatilis
21
3mdash
91
mdashmdash
mdashmdash
18019
18th
16057
19Micropygom
yiarorotaensis
4mdash
mdashmdash
8mdash
21
mdashmdash
178
02
17th
15054
20Psycho
dopygu
sparaensis
mdashmdash
mdashmdash
63
51
mdashmdash
176
02
17th
15054
21Psychodopygusclaustre
i7
1mdash
mdashmdash
mdash2
mdash3
1154
031
15th
14050
mdashPsychodopygusspp
4
mdash2
mdash6
mdash2
mdashmdash
mdashmdash
mdashmdash
14050
22Trich
ophoromyiaeurypyga
mdash8
mdashmdash
mdash3
mdash1
mdashmdash
182
018
19th
12043
23Psathyromyiaaragaoi
mdashmdash
52
mdashmdash
4mdash
mdashmdash
188
015
20th
11039
24Evandrom
yiabrachyphalla
43
21
1mdash
mdashmdash
mdashmdash
19014
21th
11039
25Psychodopyguscarreraicarrerai
11
mdash7
mdashmdash
mdashmdash
mdashmdash
202
008
26th
9032
26Evandrom
yiabegona
e2
2mdash
mdash4
mdashmdash
mdashmdash
mdash196
011
23th
8029
27Ny
ssom
yiawh
itman
imdash
mdashmdash
mdashmdash
mdash8
mdashmdash
mdash20
009
25th
8029
28Psathyromyiascaffi
32
mdashmdash
2mdash
mdashmdash
mdashmdash
198
01
24th
7025
29Pressatia
trispinosa
mdashmdash
mdash7
mdashmdash
mdashmdash
mdashmdash
204
007
27th
7025
30Micropygom
yiatrinidadensis
7mdash
mdashmdash
mdashmdash
mdashmdash
mdashmdash
208
005
29th
7025
31Viannamyiatuberculata
6mdash
mdashmdash
mdashmdash
mdashmdash
mdashmdash
21004
30th
6021
32Pintom
yiadamascenoi
mdashmdash
mdashmdash
mdashmdash
5mdash
mdashmdash
20009
25th
5018
33Evandrom
yiaevandroi
mdashmdash
mdashmdash
41
mdashmdash
mdashmdash
204
007
27th
5018
34Psathyromyialutziana
5mdash
mdashmdash
mdashmdash
mdashmdash
mdashmdash
212
003
31th
5018
35Brum
ptom
iatra
vasso
simdash
mdashmdash
mdashmdash
mdash3
1mdash
mdash196
011
23th
4014
36Evandrom
yiabacula
mdashmdash
mdashmdash
1mdash
3mdash
mdashmdash
202
008
26th
4014
37Evandrom
yiamonstruosa
mdashmdash
4mdash
mdashmdash
mdashmdash
mdashmdash
206
006
28th
4014
Journal of Tropical Medicine 5
Table1Con
tinued
Species
PortoSabao
Traıra
River
Iratapuru
Santo
Power
Plant
ISA
SISA
Rank
Total
Village
Anton
ioVillage
CD
CD
CD
CD
CD
38Psychodopygusa
mazonensis
mdashmdash
mdashmdash
22
mdashmdash
mdashmdash
21004
30th
4014
39Psychodopyguscorossoniensis
mdashmdash
mdashmdash
mdash2
mdashmdash
mdashmdash
206
006
28th
2007
40Pressatia
equatoria
lismdash
2mdash
mdashmdash
mdashmdash
mdashmdash
mdash21
004
30th
2007
41Psathyromyiainflata
mdashmdash
mdashmdash
mdashmdash
2mdash
mdashmdash
21004
30th
2007
42Micropygom
yiamicropyga
mdashmdash
mdashmdash
mdashmdash
11
mdashmdash
214
002
32th
2007
43Psychodopygusgenicu
latus
mdashmdash
mdash2
mdashmdash
mdashmdash
mdashmdash
218
033th
2007
44Lu
tzom
yiacarvalhoi
mdashmdash
1mdash
mdashmdash
mdashmdash
mdashmdash
208
005
29th
1004
45Psathyromyiapunctigenicu
lata
mdashmdash
mdashmdash
1mdash
mdashmdash
mdashmdash
216
001
32th
1004
Total
856
411
342
209
274
96415
186
74
2800
1000
1267
551
370
601
11Ea
chlocatio
ncomprise
dam
onito
ringstation(M
S)sampled
with
051and20
mCD
Ctrapsprovidingapproxim
ately
504h
MS
Cfem
aleDm
aleISAind
exof
speciesa
bund
anceSISAstand
ardindexof
speciesa
bund
ance
ldquothe
bold
italic
fontrdquoP
rovenandor
putativ
eACL
vectorsintheA
mazon
Region
based
inYo
ungandDun
can[1]Ra
ngelandLainson[4]Re
ady[2]andLainsonandShaw
[16]
6 Journal of Tropical Medicine
Table 2 Vertical stratification of Phebotomine species composition of Santo Antonio do Jari hydroelectric system impacted area (AmapaState Amazonian Brazil) on the basis of five surveyed locations (2012ndash2015)
Species 05m ground 1m ground 20m canopy TotalC D C D C D
1 Sciopemyia sordellii 117 71 91 23 11 3 3162 Psychodopygus davisi 48 27 44 39 102 54 314mdash Trichophoromyia spp 117 mdash 127 mdash 24 mdash 2683 Bichromomyia flaviscutellata 85 45 79 41 11 4 2654 Evandromyia infraspinosa 38 18 48 27 16 3 1505 Nyssomyia umbratilis 35 23 26 13 28 21 1466 Trichophoromyia castanheirai mdash 50 mdash 59 mdash 26 135mdash Pressatia spp 66 mdash 39 mdash 26 mdash 131mdash Evandromyia spp 55 12 41 9 11 mdash 1287 Vianamyia furcata 42 8 46 6 8 1 1118 Psychodopygus squamiventris sl 27 11 20 1 23 7 899 Trichopygomyia trichopyga 18 20 10 5 22 11 8610 Trichophoromyia ubiquitalis 28 15 16 13 9 5 86mdash Other species (11ndash45) 137 112 129 69 71 57 575
Total 813 412 716 305 362 192 2800Total (C + D) 1225 1021 554 mdash
Each stratum was sampled with approximately 840 h of CDC trapC female D male
infraspinosa (119899 = 150 SISA = 06) and Ny umbratilis (119899 =146 SISA = 063) Less frequently captured but nonethelessepidemiologically relevant species included Ps squamiventrissl (119899 = 89 SISA = 052) Th ubiquitalis (119899 = 86 SISA=05)Ps hirsutus hirsutus (119899 = 55 SISA = 04)Ny anduzei (119899 = 45SISA = 023) Lutzomyia gomezi (119899 = 38 SISA = 012) Nyantunesi (119899 = 19 SISA = 037) Ps paraensis (119899 = 15 SISA =02) and Ny whitmani (119899 = 8 SISA = 009)
Sampling effort for each area comprised approximately504 h of CDC trapping (14 expeditions 3 CDC traps and12-hour CDC exposition) Porto Sabao was the site of themost captured specimens (1267) followed by Santo AntonioVillage (601) The highest diversity was observed in capturesfrom Iratapuru Village (1198671015840 = 3027) All 1198671015840 value com-parisons between the different environments were significant(119901 le 005) except when comparing Traıra River and thePower Plant (119901 le 0099) (Figure 2)
Regarding vertical distribution sampling of the samecaptures described above accounted for approximately 840 hof CDC trapping by stratum (14 expeditions 5 ecotopes and12-hour CDC exposition) CDC traps placed 05m aboveground level accounted for a greater number of individuals(1225) than those placed 1m above ground level (1021) and20m above ground level in the canopy (554) (Table 2) WhilePs davisi was most likely to be found at canopy level (75individuals at 05m 83 at 1m and 156 at 20m) and Biflaviscutellata closer to ground level (130 at 05m 120 at 1mand 15 at 20m) there was little difference in numbers of Nyumbratilis individuals captured at the different levels of strata(58 at 05m 39 at 1m and 49 at 20m)
The monthly average rainfall was the only climatic datawith a representative coefficient of variance (1198781015840 = 7081)whentested against ACL occurrence and phlebotomine frequency
005
115
225
335
PortoSabatildeo
TraiacuteraRiver
IratapuruVillage
Santo AntocircnioVillage
PowerPlant
p = 0009p = 0099
p = 0007
p = 0012p = 0001
p = 0031
p = 0001p = 0004
p = 0001p = 00012855
26543027
2819
1468
Figure 2 Comparison of Shannon diversity indexes for CDC-captured phlebotomines from the five surveyed locations in SantoAntonio do Jari hydroelectric system impacted area between thestates of Para and Amapa Brazil (2012ndash2015)
(1198781015840 = 6595) in contrast to temperature (1198781015840 = 596)and relative air humidity (1198781015840 = 956) The available dataenabled correlations to be determined between the periodsof May 2012 to April 2015 a period within which nine of thefourteen capture efforts were undertaken Rainfall presenteda moderate correlation with overall phlebotomine frequency(119903 = 065 119901 = 000001) as also individually compared withPs squamiventris sl (119903 = 065 119901 = 000001) and a similarlymoderate but lower correlation with Bi flaviscutellata (119903 =048 119901 = 00021) and Ny umbratilis (119903 = 048 119901 = 0002)
Journal of Tropical Medicine 7
A decrease in the overall phlebotomine population wasobserved after March until November 2014 coinciding withthe transition between the rainy and dry periods and withflooding (Figure 3)
The number of ACL cases presented a moderate cor-relation with overall phlebotomine frequency (119903 = 044119901 = 00063) and also with Ps squamiventris (119903 = 032119901 = 00003) and an insignificant negative correlation with Biflaviscutellata (119903 = minus002 119901 = 00001) A similarly moderatebut higher correlation was however found withNy umbratilis(119903 = 048 119901 = 0002) (Figure 4)
4 Discussion
It is important to consider current investigations of phle-botomine fauna in the Jari River Basin in the context ofthe historical findings from the 1970s and 1980s Theseearlier studies represent comprehensive research conductedin Monte Dourado (northern PA) a region close to thearea investigated in the present study and on which thepresent epidemiological background to ACL transmissionin the associated region is based In brief the previousresearch includes a description of Ny umbratilis (formerlyldquoobscuredrdquo as Ny anduzei) [13] and its incrimination as themain vector of L (V) guyanensis [9ndash11] confirmed by strongecological associations [15] Just as important a parasitemore recently described as L (V) naiffi was isolated fromthe armadilloDasypus novemcinctus for which transmissionwas locally driven and attributed to Ps squamiventris sl andother Psychodopygus species [22] this comprised a primarilyenzootic ACL occasionally infecting humans [23] Finallythe isolation of L (L) amazonensis from rodents in an areaaffected by occurrence of the main vector Bi flaviscutellata[11] comprised the third known Jari ACL transmission cycleThese facts described above suggest that Ny umbratilis Pssquamiventris sl and Bi flaviscutellata are implicated in thetransmission scenario of the area covered by the present studyand thus merit particular attention
The present study incorporated a representative compo-sition of the known AP fauna Considering the substantialsampling effort (2520 h) the determined diversity showedagreement with the findings of other inventories from theBrazilian Amazon basin [1 24] Moreover all new records forAP have already been recorded for PA [25]
Apart from Ny umbratilis Ps squamiventris sl and Biflaviscutellata other species captured were noted as beingepidemiologically relevant to ACL in the Brazilian AmazonRegion Ps paraensis Ps davisi and Ps hirsutus hirsutuswhich are putative vectors of L (V) naiffi [26 27] Thubiquitalis a proven vector of L (V) lainsoni [28] Nyanduzei a secondary vector of L (V) guyanensis [4] Nywhitmani and Lu gomezi proven and putative vectors of L(V) shawi respectively [4 26] and lastly Ny antunesi asuspected vector of L (V) lindenbergi [4]
Among the above species Ps davisi is notable as thethird highest ranking according to SISA (074) and as havingthe highest presence in all vertical strata in particular atthe canopy level This species is generally anthropophilic iswidely distributed in the Amazon Basin [1 24] and is related
to Leishmania (Viannia) infections in the States of Rondonia(RO) [29] and PA [27 30] Interestingly this species remainsfrequent in recently surveyed areas affected by hydroelectricschemes in RO [8] and PA (TVS unpublished) Theseobservations add weight to the need for further investigationinto the putative epidemiological role of Ps davisi in Jari
The species most frequently captured Sc sordellii wouldappear not to be of epidemiological concern Firstly theapparent abundance of a given phlebotomine species is notsufficient of itself to incriminate it as a vector [31] particularlywhen it does not demonstrate high levels of anthropophilicbehavior Secondly flagellate infections isolated from thisspecies (formerly referred to as Lutzomyia nordestina) arehistorically recognized as distinct from Leishmania [22] orremain undetermined [32] and are more likely to be othertrypanosomatids [22] Furthermore despite recent reports ofmolecular Leishmania ascribed to this fly [33 34] currentlypolymerase chain reaction-based techniques do not allowldquotrue infectionrdquo to be distinguished from a ldquosimple and occa-sionally ingestedrdquo DNA fragment [3] Thus there remainsinsufficient evidence for a potential vector role of S sordelliiin Leishmania transmission
High frequencies of light-trapped Bi flaviscutellata havebeen documented [8 35] despite rodent-baited trappingbeing better recognized as amethod for attracting the speciesAdditionally the first SISA Rank (095) was indicative ofthis high presence in all five surveyed ecotopes this hasbeen observed in the Belem metropolitan area (PA) whereBi flaviscutellata was the only omnipresent species in sevenurban and ecologically isolated forest fragments surveyed[36] Furthermore this species has been reported as beingable to adapt to plantations of introduced tree species and inother nonclimax forests in Jari [14] suggesting it is capableof responding positively to environmental change such asthe vegetal suppression and flooding caused by hydroelectricoperations Nonetheless ecological niche model projectionspredict no change in spatial distribution for Bi flaviscutellatain the Jari area which is an environment that remains suitablefor the species [37]
In an area in Regina French Guiana ecologically dis-turbed due to agricultural practices flagellates infecting a Biflaviscutellata specimen have been determined to be L (V)guyanensis [38] in similar circumstances to the observed lowpresence of Ny umbratilis Although this possibly changingpattern in ACL transmission should be interpreted withcaution it raises an intriguing possibility about the permis-siveness of Bi flaviscutellata to L (V) guyanensis in theAmazonGuianan ACL scenario
In the present study a higher number of Bi flaviscutellataindividuals was captured compared to Ny umbratilis (265versus 146 resp) A lack of available evidence for naturalinfection in this unnatural vectorparasite system howeverprecludes speculation about the possible involvement of Biflaviscutellata even in the context of occasional transmissionof parasites distinct from L (L) amazonensis Neverthelessthe current high occurrence of Bi flaviscutellata in theAmazonGuianan ACL scenario may be of future concernunnatural parasitevectorreservoir systems when closelyrelated in an ecological context can on a century-level scale
8 Journal of Tropical Medicine
010203040506070
Bi flaviscutellata
01020304050607080
Ny umbratilis
0510152025303540
Ps squamiventris sl
0100200300400500600
050
100150200250300350400
2012 2013 2014 2015
Wat
erflo
odin
g
Ps s
quya
miv
entri
s sl
Bi fl
avisc
utell
ata
Ny u
mbr
atili
s
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
050
100150200250300350400
050
100150200250300350400
050
100150200250300350400
010203040506070
10000 20000 30000 40000000Rainfall (mm)
01020304050607080
10000 20000 30000 40000000Rainfall (mm)
Rainfall (mm)Overall
Rainfall (mm)
Rainfall (mm)
Rainfall (mm)
0100200300400500600
Ove
rall
10000 20000 30000 40000000Rainfall (mm)
05
10152025303540
10000 20000 30000 40000000Rainfall (mm)
r = 065 p = 000001
r = 048 p = 00021
r = 049 p = 00021
r = 065 p = 000001
Figure 3 Correlation between rainfall and phlebotomine putative vector species captured in Santo Antonio do Jari hydroelectric systemimpacted area between the states of Para and Amapa Brazil (2012ndash2015)
Journal of Tropical Medicine 9
2012 2013 2014 2015
Wat
erflo
odin
g
0100200300400500600
010203040506070
Bi flaviscutellata
01020304050607080
Ny umbratilis
0510152025303540
Ps squamiventris sl
Bi fl
avisc
utell
ata
05
1015202530
05
1015202530
05
1015202530
05
1015202530
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
0100200300400500600
Ove
rall
10 20 300ACL cases
010203040506070
10 20 300ACL cases
01020304050607080
Ny u
mbr
atili
s
10 20 300ACL cases
ACL casesOverall
ACL cases
ACL cases
ACL cases
05
10152025303540
Ps s
quam
iven
tris s
l
10 20 300ACL cases
r = 044 p = 00063
r = minus002 p = 00001
r = 048 p = 0002
r = 032 p = 00003
Figure 4 Correlation between ACL cases and phlebotomine putative vector species captured in Santo Antonio do Jari hydroelectric systemimpacted area between the states of Para and Amapa Brazil (2012ndash2015)
10 Journal of Tropical Medicine
undergo stress adaptation (evolutionary fitness) and formnew ecological systems driven by coevolution [39]
There is no doubt that Ny umbratilis is the main vectorof ACL in the Jari River Basin [15] Surprisingly the well-documented tree-dwelling behavior of this species [15 40]was not observed in CDC canopy captures in the presentstudy very little difference was found between the number ofindividuals captured in the three sampled strata (58 at 05m39 at 1m and 49 at 20m) Although Ny umbratilis is knownto climb into the canopy at night to feed on edentates (mainlysloths) [12 41] it is possible that an alternative terrestrialsource of blood such as rodents may be attracting femalestoward ground level Sloths are very slow moving and aretherefore commonly rescued for relocation during environ-mentalmonitoring operations their resulting rarenessmeansNy umbratilis has had to find other food sources Converselyblood meal analysis conducted during studies in a forestfragment of the Western Amazon has shown rodent bloodto be the predominant feeding source for Ny umbratilis [42]Wild and synanthropic rodents have been shown to surviveenvironmental disturbance in Jari and to proliferate in areasof introduced tree species [14] this indicated a plausiblereason not only for the putative adaptation of Ny umbratilisbut also for the increasing population levels of the rodentblood-feeder Bi flaviscutellata as already discussed
Given the clear importance of Psychodopygus of theChagasi series (formerly referred to as the Squamiventrisseries) in the context of ACL transmission [22] taxonomicstudies on identifying the cryptic females have been under-taken previously [43] and more recently with interestingresults (Rodrigo E Godoy unpublished) These techniqueshowever could not be employed in the field at the time Earlyconsideration of the assumed geographic distribution of thecomplex Ps squamiventris sl [1 44] shows that individualsfrom AP are likely to be predominantly Ps s maripaensisDetermining males of the species present could clarify thesetaxonomic uncertainties in the area covered by the presentstudy however doubts about identification of some malespecimens meant they were grouped as Ps squamiventris sl
Ryan et al [22] did not find Ps squamiventris sl to benaturally infected in the Jari area but proposed a link to L(V) naiffi along the same northern bank of the AmazonRiver through studies in Cachoeira Porteira (PA) Fouqueet al [38] came to the same conclusions in French GuianaInterestingly this fly species complex was found alongside L(V) braziliensis DNA in Suriname [45] an area where theknown Amazonian vectors of this parasite Ps complexus andPs wellcomei are unlikely to occur As previously discussedin the context of Bi flaviscutellata the changing transmissionpatterns of unnatural vectorparasite systems must be care-fully interpreted and only after strong ecological associationshave been determined Thus the putative vector importanceof Ps squamiventris sl in Jari can only be assumed in respectof L (V) naiffi
The five surveyed areas presented in general differentdiversity indexes showing variations in the phlebotominecomposition along the affected area This finding can beattributed not only to the distance between capture sitesbut also to forest composition fragmentation geographic
barriers and migration of mammal fauna between sites thislatter can occur naturally or be induced by rescue opera-tions involved in hydroelectric planning whereby animalsare moved to suitable ldquodry landrdquo environments safe frompotential flooding
A decrease in the phlebotomine population in 2014 inthe period after filling the reservoir could not be attributedexclusively either to flooding or to the transition from therainy to the dry season Both eventualities although oppositein effect are likely to lead to unsuitable conditions forphlebotomine reproduction However they did coincide inMay 2014 when rainfall decreased and the lake was formed
Conversely rainfall was a valid determinant of overallphlebotomine fluctuation Ps squamiventris sl was the mainputative vector directly correlated with this Other variablessuch as forest composition and altitudinal variation areknown to influence seasonal dynamics of species from thecomplex Ps squamiventris sl For example the dry periodfrom October to November in Serra dos Carajas (southernPA) can be unsuitable for Ps wellcomei which is adapted tohigh altitudes in contrast to Ps complexus which flourishesin lowland areas during the same months [46]
An insignificant negative to a moderate positive correla-tion was observed between cases of ACL and phlebotominefluctuations In the case of Ny umbratilis however theapparently higher correlation when compared with otherspecies is in agreement with its known vector importance inAmazonGuianan ACL A strong spatiotemporal associationbetween a given vector and ACL cases is considered essentialto maintenance of the Leishmania life cycle [2 14]
In summary the present study filled a temporal gapon phlebotomine data relating to the Jari area It providesa meaningful update on species composition and furtherinsights into seasonal dynamics of species under pressurescaused by environmental change The presence of ACLvectors in particular Ny umbratilis Bi flaviscutellata andPs squamiventris sl is noteworthy due to their directassociation with AmazonGuianan-ACL transmission ForNy umbratilis the lower frequency associatedwith the unpre-dicted lack of tree-dwelling behavior may be attributableto a reduction in numbers of canopy-dwelling mammalsdue to the environmental disturbance and relocation effortsthis could consequently have led to seeking for alternativeterrestrial blood sources at ground level The possible asso-ciation of Ps davisi with Leishmania transmission cycles inother Amazon ecoregions draws attention to its high rankposition in Jari Rainfall can be partly associated with localseasonal fluctuations in fly numbers mainly with respect toPs squamiventris sl Cases of ACL were also more highlycorrelated with Ny umbratilis dynamics Further attentionwas also given to results for Bi flaviscutellata which hasbeen apparently tolerant of anthropic activity continuedphlebotominemonitoring is proposed in order to evaluate theimplication for social welfare and health in the future
Competing Interests
The authors declare that they have no competing interests
Journal of Tropical Medicine 11
Acknowledgments
Theauthorswish to thank the ldquoInstituto de Pesquisas Cientıfi-cas e Tecnologicas do Estado do Amapa IEPArdquo entomologyteam for the technical assistance given to the field work inparticular to Aderbal Amanajas Santana Rosalvo Dias deCastro Noel Fernandes Santos Neto Osita Cleyriane Lobatodo Nascimento Jose Luiz dos Santos Chaves and Joao doSocorro Gomes Campos Logistical facilities were financiallysupported by the ldquoJari Energiardquo
References
[1] D G Young and M A Duncan ldquoGuide to the identificationand geographic distribution of Lutzomyia sand flies in Mexicothe West Indies Central and South America (Diptera Psycho-didae)rdquo inMemoirs of the American Entomological Institute pp1ndash881 Associated Publishers American Entomological InstituteGainesville Fla USA 1994
[2] P D Ready ldquoBiology of phlebotomine sand flies as vectors ofdisease agentsrdquo Annual Review of Entomology vol 58 pp 227ndash250 2013
[3] R P Brazil A A Fuzzari and J D Andrade Filho ldquoSandfly vectors of Leishmania in the Americasmdasha mini reviewrdquoEntomology Ornithology amp Herpetology vol 4 no 2 Article ID1000144 2015
[4] E F Rangel and R Lainson ldquoProven and putative vectorsof American cutaneous leishmaniasis in Brazil aspects oftheir biology and vectorial competencerdquoMemorias do InstitutoOswaldo Cruz vol 104 no 7 pp 937ndash954 2009
[5] E F Rangel S M Costa and B M Carvalho ldquoEnvironmentalchanges and the geographic spreading of American cutaneousleishmaniasis in Brazilrdquo in Leishmaniasis-Trends in Epidemiol-ogy D Claborn Ed Diagnosis and Treatment InTech RijekaCroatia 2014
[6] W P Taddei ldquoO genero Mansonia (Diptera Culicidae) e aproliferacao de mosquitos na usina hidreletrica de Tucuruırdquo inEnergia na Amazonia S B Magalhaes R C Brito and E RCastro Eds vol 1 pp 311ndash318 MPEGFPAUNAMAZ BelemBrazil 1996 (Portuguese)
[7] A I A Oliveira Observacoes Sobre a Resolucao CONAMA no23797 que Dispoe Sobre o Licenciamento Ambiental MimeoConselho Empresarial Brasileiro Para o Desenvolvimento Sus-tentavel Rio de Janeiro Brazil 1997 (Portuguese)
[8] A K R Galardo C D Galardo G A Silveira et al ldquoPhle-botominae sand flies (Diptera Psychodidae) potential vectorsof American cutaneous leishmaniasis agents in the area asso-ciated with the Santo Antonio hydroelectric system in WesternAmazonian Brazilrdquo Revista da Sociedade Brasileira deMedicinaTropical vol 48 no 3 pp 265ndash271 2015
[9] R Lainson R DWard and J J Shaw ldquoCutaneous leishmaniasisin North Brazil Lutzomyia anduzei as a major vectorrdquo Transac-tions of the Royal Society of Tropical Medicine and Hygiene vol70 no 2 pp 171ndash172 1976
[10] R Lainson J J Shaw R D Ward P D Ready and R DNaiff ldquoLeishmaniasis in Brazil XIII Isolation of Leishmaniafrom armadillos (Dasypus novemcinctus) and observations onthe epidemiology of cutaneous leishmaniasis in north ParaStaterdquo Transactions of the Royal Society of Tropical Medicine andHygiene vol 73 no 2 pp 239ndash242 1979
[11] R Lainson J J Shaw P D Ready M A Miles and M PovoaldquoLeishmaniasis in Brazil XVI Isolation and identification of
Leishmania species from sandflies wild mammals and manin north Para State with particular reference to L braziliensisguyanensis causative agent of ldquopian-boisrdquordquo Transactions of theRoyal Society of Tropical Medicine and Hygiene vol 75 no 4pp 530ndash536 1981
[12] R Lainson J J Shaw and M Povoa ldquoThe importance of eden-tates (sloths and anteaters) as primary reservoirs of Leishmaniabraziliensis guyanensis causative agent of lsquopian boisrsquo in northBrazilrdquo Transactions of the Royal Society of Tropical Medicineand Hygiene vol 75 no 4 pp 611ndash612 1981
[13] R DWard and H Fraiha ldquoLutzomyia umbratilis a new speciesof sand fly from Brazil (Diptera Psychodidae)rdquo Journal ofMedical Entomology vol 14 no 3 pp 313ndash317 1977
[14] P D Ready R Lainson and J J Shaw ldquoLeishmaniasis in BrazilXX Prevalence of lsquoenzootic rodent leishmaniasisrsquo (Leishmaniamexicana amazonensis) and apparent absence of lsquopian boisrsquo(Le braziliensis guyanensis) in plantations of introduced treespecies and in other non-climax forests in eastern AmazoniardquoTransactions of the Royal Society of Tropical Medicine andHygiene vol 77 no 6 pp 775ndash785 1983
[15] P D Ready R Lainson J J Shaw and R DWard ldquoThe ecologyof Lutzomyia umbratilisWard amp Fraiha (Diptera Psychodidae)the major vector to man of Leishmania braziliensis guyanensisin north-eastern Amazonian Brazilrdquo Bulletin of EntomologicalResearch vol 76 no 1 pp 21ndash40 1986
[16] R Lainson and J J Shaw ldquoLeishmania (Viannia) naiffi spn a parasite of the armadillo Dasypus novemcinctus (L)in Amazonian Brazilrdquo Annales de Parasitologie Humaine etComparee vol 64 no 1 pp 3ndash9 1989
[17] A Roger M Nacher M Hanf et al ldquoClimate and leishmaniasisin French guianardquo The American Journal of Tropical Medicineand Hygiene vol 89 no 3 pp 564ndash569 2013
[18] L Ryan ldquoFlebotomos do estado do Para Brasil (Diptera Psy-chodidae)rdquo Thecnical Document 1 Instituto Evandro ChagasBelem Brazil 1986 (Portuguese)
[19] E A B Galati ldquoMorfologia e Taxonomia Morfologia termi-nologia de adultos e identificacao dos taxons da Americardquoin Flebotomıneos do Brasil E F Rangel and R LainsonEds pp 53ndash75 Editora Fiocruz Rio de Janeiro Brazil 2003(Portuguese)
[20] D R Roberts and B P Hsi ldquoAn index of species abundance foruse with mosquito surveillance datardquo Environmental Entomol-ogy vol 8 no 6 pp 1007ndash1013 1979
[21] Oslash Hammer D A T Harper and P D Ryan ldquoPast paleontolog-ical statistics software package for education and data analysisrdquoPalaeontologia Electronica vol 4 no 1 pp 1ndash9 2001
[22] L Ryan R Lainson J J Shaw R R Braga and E A IshikawaldquoLeishmaniasis in Brazil XXV Sandfly vectors of Leishmania inPara State Brazilrdquo Medical and Veterinary Entomology vol 1no 4 pp 383ndash395 1987
[23] R Lainson J J Shaw F T Silveira R R Braga and E AIshikawa ldquoCutaneous leishmaniasis of man due to Leishmania(Viannia) naiffirdquo Annales de Parasitologie Humaine et Compa-ree vol 65 no 5-6 pp 282ndash284 1990
[24] E G C Bermudez Lutzomyia Sand Flies in the BrazilianAmazon Basin (Diptera Psychodidae) INPA Manaus Brazil2009
[25] G M Aguiar and W M Medeiros ldquoDistribuicao regionale habitats das especies de flebotomıneos do Brasilrdquo in Fle-botomıneos do Brasil E F Rangel and R Lainson Eds pp 207ndash256 Fiocruz Rio de Janeiro Brazil 2003 (Portuguese)
12 Journal of Tropical Medicine
[26] J R Arias M A Miles R D Naiff et al ldquoFlagellate infectionsof Brazilian sand flies (Diptera Psychodidae) isolation invitro and biochemical identification of Endotrypanum andLeishmaniardquo The American Journal of Tropical Medicine andHygiene vol 34 no 6 pp 1098ndash1108 1985
[27] A A Souza T V Santos Y L Jennings et al ldquoNaturalLeishmania(Viannia) spp infections in phlebotomine sand flies(Diptera Psychodidae) from the Brazilian Amazon regionreveal new putative transmission cycles of American cutaneousleishmaniasisrdquo Parasite vol 23 article 22 2016
[28] F T Silveira A A Souza R Lainson J J Shaw R R Bragaand E A Ishikawa ldquoCutaneous leishmaniasis in the AmazonRegion natural infection of the sand fly Lutzomyia ubiquitalis(Psychodidae Phlebotominae) by Leishmania (Viannia) lain-soni in Para state BrazilrdquoMemorias do Instituto Oswaldo Cruzvol 86 no 1 pp 127ndash130 1991
[29] LH S Gil S A Basano A A Souza et al ldquoRecent observationson the sand fly (Diptera Psychodidae) fauna of the state ofRondonia Western Amazonia Brazil the importance of Psych-dopygus davisi as a vector of zoonotic cutaneous leishmaniasisrdquoMemorias do Instituto Oswaldo Cruz vol 98 no 6 pp 751ndash7552003
[30] A A A Souza F T Silveira R Lainson et al ldquoThe Phlebotom-inae fauna of Serra dos Carajas Para Brazil and its possibleimplication in the transmission of American tegumentaryleishmaniasisrdquo Revista Pan-Amazonica de Saude vol 1 no 1pp 45ndash51 2010
[31] R Killick-Kendrick ldquoPhlebotomine vectors of the leishmani-ases a reviewrdquoMedical and Veterinary Entomology vol 4 no 1pp 1ndash24 1990
[32] M E C Dorval G Cristaldo H C da Rocha et al ldquoPhle-botomine fauna (Diptera Psychodidae) of an American cuta-neous leishmaniasis endemic area in the state of Mato Grossodo Sul Brazilrdquo Memorias do Instituto Oswaldo Cruz vol 104no 5 pp 695ndash702 2009
[33] V C F V Guimaraes P L Costa F J da Silva et al ldquoMoleculardetection of Leishmania in phlebotomine sand flies in a cuta-neous and visceral leishmaniasis endemic area in northeasternBrazilrdquo Revista do Instituto de Medicina Tropical de Sao Paulovol 56 no 4 pp 357ndash360 2014
[34] R S Lana E M Michalsky C L Fortes-Dias et al ldquoPhle-botomine sand fly fauna and Leishmania infection in thevicinity of the Serra do Cipo National Park a natural Brazilianheritage siterdquo BioMed Research International vol 2015 ArticleID 385493 9 pages 2015
[35] V R Alves R A de Freitas F L Santos A F J de OliveiraT V Barrett and P H F Shimabukuro ldquoSand flies (DipteraPsychodidae Phlebotominae) from central amazonia and fournew records for the Amazonas State Brazilrdquo Revista Brasileirade Entomologia vol 56 no 2 pp 220ndash227 2012
[36] J V S Ferreira T V Santos E M Santos and I S GorayebldquoPhlebotomine sand flies (Diptera Psychodidae) in forestfragments of Belem metropolitan area Para State Brazil withconsiderations on vectors of American cutaneous leishmaniasisagentsrdquo Revista Pan-Amazonica de Saude vol 5 no 2 pp 29ndash35 2014
[37] B M Carvalho E F Rangel P D Ready and M M ValeldquoEcological niche modelling predicts southward expansion ofLutzomyia (Nyssomyia) flaviscutellata (Diptera PsychodidaePhlebotominae) vector of Leishmania (Leishmania) amazonen-sis in South America under climate changerdquo PLoS ONE vol 10no 11 Article ID e0143282 2015
[38] F Fouque P Gaborit J Issaly et al ldquoPhlebotomine sand flies(Diptera Psychodidae) associatedwith changing patterns in thetransmission of the human cutaneous leishmaniasis in FrenchGuianardquoMemorias do Instituto Oswaldo Cruz vol 102 no 1 pp35ndash40 2007
[39] R Killick-Kendrick ldquoSome epidemiological consequences ofthe evolutionary fit between Leishmaniae and their phle-botomine vectorsrdquo Bulletin de la Societe de Pathologie Exotiqueet de ses Filiales vol 78 no 5 pp 747ndash755 1985
[40] F X Pajot F Le Pont BGentile andR Besnard ldquoEpidemiologyof leishmaniasis in French Guianardquo Transactions of the RoyalSociety of Tropical Medicine and Hygiene vol 76 no 1 pp 112ndash113 1982
[41] H A Christensen J R Arias A M De Vasquez and R ADe Freitas ldquoHosts of sandfly vectors of Leishmania braziliensisguyanensis in the central Amazon of Brazilrdquo The AmericanJournal of Tropical Medicine and Hygiene vol 31 no 2 pp 239ndash242 1982
[42] L C R Nery E S Lorosa andAM R Franco ldquoFeeding prefer-ence of the sand flies Lutzomyia umbratilis and L spathotrichia(Diptera Psychodidae Phlebotominae) in an urban forest patchin the City ofManaus Amazonas BrazilrdquoMemorias do InstitutoOswaldo Cruz vol 99 no 6 pp 571ndash574 2004
[43] PD Ready R Lainson J J Shaw andAA Souza ldquoDNAprobesfor distinguishing Psychodopygus wellcomei from Psychodopy-gus complexus (Diptera Psychodidae)rdquo Memorias do InstitutoOswaldo Cruz vol 86 no 1 pp 41ndash49 1991
[44] P D Ready H Fraiha R P Lane J R Arias and F X PajotldquoOn distinguishing the female of Psychodopygus wellcomei avector of mucocutaneous leishmaniasis from other squamiven-tris series females I Characterization of Ps squamiventrissquamiventris and Ps s maripaensis stat nov (Diptera Psycho-didae)rdquo Annals of Tropical Medicine and Parasitology vol 76no 2 pp 201ndash214 1982
[45] A D Kent T V dos Santos A Gangadin A Samjhawan DR A Mans and H D F H Schallig ldquoStudies on the sandfly fauna (Diptera Psychodidae) in high-transmission areas ofcutaneous leishmaniasis in the Republic of Surinamerdquo Parasitesand Vectors vol 6 no 1 article 318 2013
[46] P D Ready R Lainson and J J Shaw ldquoHabitat and seasonalityof Psychodopygus wellcomei help incriminate it as a vector ofLeishmania braziliensis in Amazonia and Northeast BrazilrdquoTransactions of the Royal Society of Tropical Medicine andHygiene vol 78 no 4 pp 543ndash544 1984
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
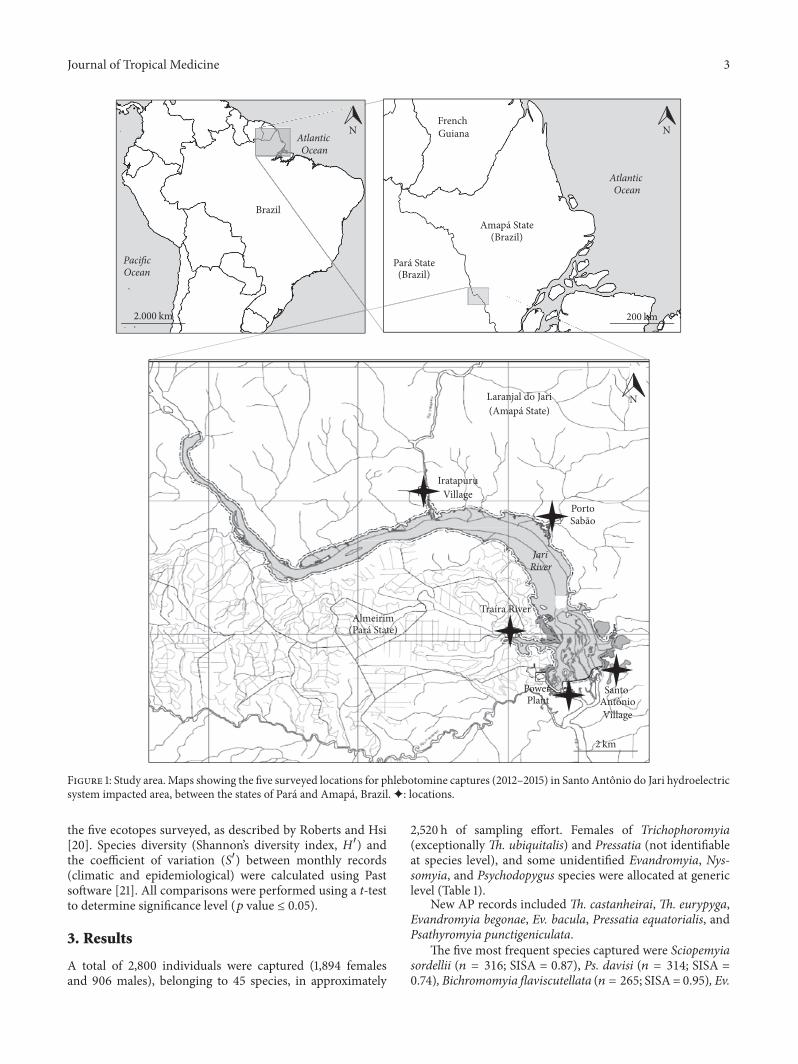
Journal of Tropical Medicine 3
Atlantic Ocean
Brazil
PacificOcean
Atlantic Ocean
Amapaacute State(Brazil)
French Guiana
Paraacute State(Brazil)
IratapuruVillage
Traiacutera River
Power Plant
PortoSabatildeo
Santo AntocircnioVillage
Jari River
Almeirim(Paraacute State)
Laranjal do Jari(Amapaacute State)
2000 km 200 km
2 km
N N
N
Figure 1 Study area Maps showing the five surveyed locations for phlebotomine captures (2012ndash2015) in Santo Antonio do Jari hydroelectricsystem impacted area between the states of Para and Amapa BrazilM locations
the five ecotopes surveyed as described by Roberts and Hsi[20] Species diversity (Shannonrsquos diversity index H1015840) andthe coefficient of variation (S1015840) between monthly records(climatic and epidemiological) were calculated using Pastsoftware [21] All comparisons were performed using a t-testto determine significance level (119901 value le 005)
3 Results
A total of 2800 individuals were captured (1894 femalesand 906 males) belonging to 45 species in approximately
2520 h of sampling effort Females of Trichophoromyia(exceptionally Th ubiquitalis) and Pressatia (not identifiableat species level) and some unidentified Evandromyia Nys-somyia and Psychodopygus species were allocated at genericlevel (Table 1)
New AP records includedTh castanheirai Th eurypygaEvandromyia begonae Ev bacula Pressatia equatorialis andPsathyromyia punctigeniculata
The five most frequent species captured were Sciopemyiasordellii (119899 = 316 SISA = 087) Ps davisi (119899 = 314 SISA =074) Bichromomyia flaviscutellata (119899 = 265 SISA= 095) Ev
4 Journal of Tropical MedicineTa
ble1Ph
ebotom
inespeciescompo
sitionofSantoAnton
iodo
Jarihydroelectric
syste
mim
pacted
area
(AmapaS
tateA
mazon
ianBrazil)
ontheb
asisoffivesurveyedlocatio
ns(2012ndash2015)
Species
PortoSabao
Traıra
River
Iratapuru
Santo
Power
Plant
ISA
SISA
Rank
Total
Village
Anton
ioVillage
CD
CD
CD
CD
CD
1Sciopemyiasordellii
149
6613
1015
442
16mdash
136
087
2nd
316
1129
2Psycho
dopygu
sdavisi
4637
9556
1510
3817
mdashmdash
64
074
3rd
314
1121
mdashTrich
ophoromyiaspp
177
mdash34
mdash40
mdash17
mdashmdash
mdashmdash
mdashmdash
268
957
3Bichromom
yiafla
viscutellata
7632
2414
2211
5231
12
2095
1st
265
946
4Evandrom
yiainfra
spinosa
4410
2518
32
3018
mdashmdash
9206
6th
150
536
5Ny
ssom
yiaum
bratilis
2612
3130
196
139
mdashmdash
86
063
5th
146
521
6Trich
ophoromyiacasta
nheir
aimdash
104
mdash6
mdash14
mdash11
mdashmdash
112
051
9th
135
482
mdashPressatia
spp
43mdash
1mdash
9mdash
78mdash
mdashmdash
mdashmdash
mdash131
468
mdashEvandrom
yiaspp
5210
4mdash
3911
12mdash
mdashmdash
mdashmdash
mdash128
457
7Viannamyiafurcata
444
115
111
305
mdashmdash
104
055
7th
111
396
8Psycho
dopygu
ssqu
amiventrissensulato
3614
155
10mdash
9mdash
mdashmdash
11052
8th
89318
9Trich
opygom
yiatrichopyga
1116
98
9mdash
2112
mdashmdash
7207
4th
86307
10Tricho
phorom
yiaub
iquitalis
216
58
148
139
2mdash
114
05
10th
86307
11Evandrom
yiasaulensis
409
5mdash
44
121
mdashmdash
128
043
11th
75268
12Psycho
dopygu
shirs
utus
hirsutus
13mdash
204
144
mdashmdash
mdashmdash
136
04
12th
5519
613
Trich
ophoromyiabrachypyga
mdash36
mdash12
mdash2
mdash2
mdashmdash
152
032
14th
5218
614
Pressatia
choti
mdash17
mdash3
mdash5
mdash25
mdashmdash
142
037
13th
5017
915
Nyssom
yiaan
duzei
65
2410
mdashmdash
mdashmdash
mdashmdash
17023
16th
4516
116
Lutzom
yiagomezi
2711
mdashmdash
mdashmdash
mdashmdash
mdashmdash
194
012
22th
3813
6mdash
Nysso
myiaspp
mdashmdash
mdash1
1mdash
1125
mdashmdash
mdashmdash
mdash38
136
17Ny
ssom
yiaan
tunesi
mdash2
9mdash
52
mdashmdash
1mdash
142
037
13th
19068
18Sciopemyiaflu
viatilis
21
3mdash
91
mdashmdash
mdashmdash
18019
18th
16057
19Micropygom
yiarorotaensis
4mdash
mdashmdash
8mdash
21
mdashmdash
178
02
17th
15054
20Psycho
dopygu
sparaensis
mdashmdash
mdashmdash
63
51
mdashmdash
176
02
17th
15054
21Psychodopygusclaustre
i7
1mdash
mdashmdash
mdash2
mdash3
1154
031
15th
14050
mdashPsychodopygusspp
4
mdash2
mdash6
mdash2
mdashmdash
mdashmdash
mdashmdash
14050
22Trich
ophoromyiaeurypyga
mdash8
mdashmdash
mdash3
mdash1
mdashmdash
182
018
19th
12043
23Psathyromyiaaragaoi
mdashmdash
52
mdashmdash
4mdash
mdashmdash
188
015
20th
11039
24Evandrom
yiabrachyphalla
43
21
1mdash
mdashmdash
mdashmdash
19014
21th
11039
25Psychodopyguscarreraicarrerai
11
mdash7
mdashmdash
mdashmdash
mdashmdash
202
008
26th
9032
26Evandrom
yiabegona
e2
2mdash
mdash4
mdashmdash
mdashmdash
mdash196
011
23th
8029
27Ny
ssom
yiawh
itman
imdash
mdashmdash
mdashmdash
mdash8
mdashmdash
mdash20
009
25th
8029
28Psathyromyiascaffi
32
mdashmdash
2mdash
mdashmdash
mdashmdash
198
01
24th
7025
29Pressatia
trispinosa
mdashmdash
mdash7
mdashmdash
mdashmdash
mdashmdash
204
007
27th
7025
30Micropygom
yiatrinidadensis
7mdash
mdashmdash
mdashmdash
mdashmdash
mdashmdash
208
005
29th
7025
31Viannamyiatuberculata
6mdash
mdashmdash
mdashmdash
mdashmdash
mdashmdash
21004
30th
6021
32Pintom
yiadamascenoi
mdashmdash
mdashmdash
mdashmdash
5mdash
mdashmdash
20009
25th
5018
33Evandrom
yiaevandroi
mdashmdash
mdashmdash
41
mdashmdash
mdashmdash
204
007
27th
5018
34Psathyromyialutziana
5mdash
mdashmdash
mdashmdash
mdashmdash
mdashmdash
212
003
31th
5018
35Brum
ptom
iatra
vasso
simdash
mdashmdash
mdashmdash
mdash3
1mdash
mdash196
011
23th
4014
36Evandrom
yiabacula
mdashmdash
mdashmdash
1mdash
3mdash
mdashmdash
202
008
26th
4014
37Evandrom
yiamonstruosa
mdashmdash
4mdash
mdashmdash
mdashmdash
mdashmdash
206
006
28th
4014
Journal of Tropical Medicine 5
Table1Con
tinued
Species
PortoSabao
Traıra
River
Iratapuru
Santo
Power
Plant
ISA
SISA
Rank
Total
Village
Anton
ioVillage
CD
CD
CD
CD
CD
38Psychodopygusa
mazonensis
mdashmdash
mdashmdash
22
mdashmdash
mdashmdash
21004
30th
4014
39Psychodopyguscorossoniensis
mdashmdash
mdashmdash
mdash2
mdashmdash
mdashmdash
206
006
28th
2007
40Pressatia
equatoria
lismdash
2mdash
mdashmdash
mdashmdash
mdashmdash
mdash21
004
30th
2007
41Psathyromyiainflata
mdashmdash
mdashmdash
mdashmdash
2mdash
mdashmdash
21004
30th
2007
42Micropygom
yiamicropyga
mdashmdash
mdashmdash
mdashmdash
11
mdashmdash
214
002
32th
2007
43Psychodopygusgenicu
latus
mdashmdash
mdash2
mdashmdash
mdashmdash
mdashmdash
218
033th
2007
44Lu
tzom
yiacarvalhoi
mdashmdash
1mdash
mdashmdash
mdashmdash
mdashmdash
208
005
29th
1004
45Psathyromyiapunctigenicu
lata
mdashmdash
mdashmdash
1mdash
mdashmdash
mdashmdash
216
001
32th
1004
Total
856
411
342
209
274
96415
186
74
2800
1000
1267
551
370
601
11Ea
chlocatio
ncomprise
dam
onito
ringstation(M
S)sampled
with
051and20
mCD
Ctrapsprovidingapproxim
ately
504h
MS
Cfem
aleDm
aleISAind
exof
speciesa
bund
anceSISAstand
ardindexof
speciesa
bund
ance
ldquothe
bold
italic
fontrdquoP
rovenandor
putativ
eACL
vectorsintheA
mazon
Region
based
inYo
ungandDun
can[1]Ra
ngelandLainson[4]Re
ady[2]andLainsonandShaw
[16]
6 Journal of Tropical Medicine
Table 2 Vertical stratification of Phebotomine species composition of Santo Antonio do Jari hydroelectric system impacted area (AmapaState Amazonian Brazil) on the basis of five surveyed locations (2012ndash2015)
Species 05m ground 1m ground 20m canopy TotalC D C D C D
1 Sciopemyia sordellii 117 71 91 23 11 3 3162 Psychodopygus davisi 48 27 44 39 102 54 314mdash Trichophoromyia spp 117 mdash 127 mdash 24 mdash 2683 Bichromomyia flaviscutellata 85 45 79 41 11 4 2654 Evandromyia infraspinosa 38 18 48 27 16 3 1505 Nyssomyia umbratilis 35 23 26 13 28 21 1466 Trichophoromyia castanheirai mdash 50 mdash 59 mdash 26 135mdash Pressatia spp 66 mdash 39 mdash 26 mdash 131mdash Evandromyia spp 55 12 41 9 11 mdash 1287 Vianamyia furcata 42 8 46 6 8 1 1118 Psychodopygus squamiventris sl 27 11 20 1 23 7 899 Trichopygomyia trichopyga 18 20 10 5 22 11 8610 Trichophoromyia ubiquitalis 28 15 16 13 9 5 86mdash Other species (11ndash45) 137 112 129 69 71 57 575
Total 813 412 716 305 362 192 2800Total (C + D) 1225 1021 554 mdash
Each stratum was sampled with approximately 840 h of CDC trapC female D male
infraspinosa (119899 = 150 SISA = 06) and Ny umbratilis (119899 =146 SISA = 063) Less frequently captured but nonethelessepidemiologically relevant species included Ps squamiventrissl (119899 = 89 SISA = 052) Th ubiquitalis (119899 = 86 SISA=05)Ps hirsutus hirsutus (119899 = 55 SISA = 04)Ny anduzei (119899 = 45SISA = 023) Lutzomyia gomezi (119899 = 38 SISA = 012) Nyantunesi (119899 = 19 SISA = 037) Ps paraensis (119899 = 15 SISA =02) and Ny whitmani (119899 = 8 SISA = 009)
Sampling effort for each area comprised approximately504 h of CDC trapping (14 expeditions 3 CDC traps and12-hour CDC exposition) Porto Sabao was the site of themost captured specimens (1267) followed by Santo AntonioVillage (601) The highest diversity was observed in capturesfrom Iratapuru Village (1198671015840 = 3027) All 1198671015840 value com-parisons between the different environments were significant(119901 le 005) except when comparing Traıra River and thePower Plant (119901 le 0099) (Figure 2)
Regarding vertical distribution sampling of the samecaptures described above accounted for approximately 840 hof CDC trapping by stratum (14 expeditions 5 ecotopes and12-hour CDC exposition) CDC traps placed 05m aboveground level accounted for a greater number of individuals(1225) than those placed 1m above ground level (1021) and20m above ground level in the canopy (554) (Table 2) WhilePs davisi was most likely to be found at canopy level (75individuals at 05m 83 at 1m and 156 at 20m) and Biflaviscutellata closer to ground level (130 at 05m 120 at 1mand 15 at 20m) there was little difference in numbers of Nyumbratilis individuals captured at the different levels of strata(58 at 05m 39 at 1m and 49 at 20m)
The monthly average rainfall was the only climatic datawith a representative coefficient of variance (1198781015840 = 7081)whentested against ACL occurrence and phlebotomine frequency
005
115
225
335
PortoSabatildeo
TraiacuteraRiver
IratapuruVillage
Santo AntocircnioVillage
PowerPlant
p = 0009p = 0099
p = 0007
p = 0012p = 0001
p = 0031
p = 0001p = 0004
p = 0001p = 00012855
26543027
2819
1468
Figure 2 Comparison of Shannon diversity indexes for CDC-captured phlebotomines from the five surveyed locations in SantoAntonio do Jari hydroelectric system impacted area between thestates of Para and Amapa Brazil (2012ndash2015)
(1198781015840 = 6595) in contrast to temperature (1198781015840 = 596)and relative air humidity (1198781015840 = 956) The available dataenabled correlations to be determined between the periodsof May 2012 to April 2015 a period within which nine of thefourteen capture efforts were undertaken Rainfall presenteda moderate correlation with overall phlebotomine frequency(119903 = 065 119901 = 000001) as also individually compared withPs squamiventris sl (119903 = 065 119901 = 000001) and a similarlymoderate but lower correlation with Bi flaviscutellata (119903 =048 119901 = 00021) and Ny umbratilis (119903 = 048 119901 = 0002)
Journal of Tropical Medicine 7
A decrease in the overall phlebotomine population wasobserved after March until November 2014 coinciding withthe transition between the rainy and dry periods and withflooding (Figure 3)
The number of ACL cases presented a moderate cor-relation with overall phlebotomine frequency (119903 = 044119901 = 00063) and also with Ps squamiventris (119903 = 032119901 = 00003) and an insignificant negative correlation with Biflaviscutellata (119903 = minus002 119901 = 00001) A similarly moderatebut higher correlation was however found withNy umbratilis(119903 = 048 119901 = 0002) (Figure 4)
4 Discussion
It is important to consider current investigations of phle-botomine fauna in the Jari River Basin in the context ofthe historical findings from the 1970s and 1980s Theseearlier studies represent comprehensive research conductedin Monte Dourado (northern PA) a region close to thearea investigated in the present study and on which thepresent epidemiological background to ACL transmissionin the associated region is based In brief the previousresearch includes a description of Ny umbratilis (formerlyldquoobscuredrdquo as Ny anduzei) [13] and its incrimination as themain vector of L (V) guyanensis [9ndash11] confirmed by strongecological associations [15] Just as important a parasitemore recently described as L (V) naiffi was isolated fromthe armadilloDasypus novemcinctus for which transmissionwas locally driven and attributed to Ps squamiventris sl andother Psychodopygus species [22] this comprised a primarilyenzootic ACL occasionally infecting humans [23] Finallythe isolation of L (L) amazonensis from rodents in an areaaffected by occurrence of the main vector Bi flaviscutellata[11] comprised the third known Jari ACL transmission cycleThese facts described above suggest that Ny umbratilis Pssquamiventris sl and Bi flaviscutellata are implicated in thetransmission scenario of the area covered by the present studyand thus merit particular attention
The present study incorporated a representative compo-sition of the known AP fauna Considering the substantialsampling effort (2520 h) the determined diversity showedagreement with the findings of other inventories from theBrazilian Amazon basin [1 24] Moreover all new records forAP have already been recorded for PA [25]
Apart from Ny umbratilis Ps squamiventris sl and Biflaviscutellata other species captured were noted as beingepidemiologically relevant to ACL in the Brazilian AmazonRegion Ps paraensis Ps davisi and Ps hirsutus hirsutuswhich are putative vectors of L (V) naiffi [26 27] Thubiquitalis a proven vector of L (V) lainsoni [28] Nyanduzei a secondary vector of L (V) guyanensis [4] Nywhitmani and Lu gomezi proven and putative vectors of L(V) shawi respectively [4 26] and lastly Ny antunesi asuspected vector of L (V) lindenbergi [4]
Among the above species Ps davisi is notable as thethird highest ranking according to SISA (074) and as havingthe highest presence in all vertical strata in particular atthe canopy level This species is generally anthropophilic iswidely distributed in the Amazon Basin [1 24] and is related
to Leishmania (Viannia) infections in the States of Rondonia(RO) [29] and PA [27 30] Interestingly this species remainsfrequent in recently surveyed areas affected by hydroelectricschemes in RO [8] and PA (TVS unpublished) Theseobservations add weight to the need for further investigationinto the putative epidemiological role of Ps davisi in Jari
The species most frequently captured Sc sordellii wouldappear not to be of epidemiological concern Firstly theapparent abundance of a given phlebotomine species is notsufficient of itself to incriminate it as a vector [31] particularlywhen it does not demonstrate high levels of anthropophilicbehavior Secondly flagellate infections isolated from thisspecies (formerly referred to as Lutzomyia nordestina) arehistorically recognized as distinct from Leishmania [22] orremain undetermined [32] and are more likely to be othertrypanosomatids [22] Furthermore despite recent reports ofmolecular Leishmania ascribed to this fly [33 34] currentlypolymerase chain reaction-based techniques do not allowldquotrue infectionrdquo to be distinguished from a ldquosimple and occa-sionally ingestedrdquo DNA fragment [3] Thus there remainsinsufficient evidence for a potential vector role of S sordelliiin Leishmania transmission
High frequencies of light-trapped Bi flaviscutellata havebeen documented [8 35] despite rodent-baited trappingbeing better recognized as amethod for attracting the speciesAdditionally the first SISA Rank (095) was indicative ofthis high presence in all five surveyed ecotopes this hasbeen observed in the Belem metropolitan area (PA) whereBi flaviscutellata was the only omnipresent species in sevenurban and ecologically isolated forest fragments surveyed[36] Furthermore this species has been reported as beingable to adapt to plantations of introduced tree species and inother nonclimax forests in Jari [14] suggesting it is capableof responding positively to environmental change such asthe vegetal suppression and flooding caused by hydroelectricoperations Nonetheless ecological niche model projectionspredict no change in spatial distribution for Bi flaviscutellatain the Jari area which is an environment that remains suitablefor the species [37]
In an area in Regina French Guiana ecologically dis-turbed due to agricultural practices flagellates infecting a Biflaviscutellata specimen have been determined to be L (V)guyanensis [38] in similar circumstances to the observed lowpresence of Ny umbratilis Although this possibly changingpattern in ACL transmission should be interpreted withcaution it raises an intriguing possibility about the permis-siveness of Bi flaviscutellata to L (V) guyanensis in theAmazonGuianan ACL scenario
In the present study a higher number of Bi flaviscutellataindividuals was captured compared to Ny umbratilis (265versus 146 resp) A lack of available evidence for naturalinfection in this unnatural vectorparasite system howeverprecludes speculation about the possible involvement of Biflaviscutellata even in the context of occasional transmissionof parasites distinct from L (L) amazonensis Neverthelessthe current high occurrence of Bi flaviscutellata in theAmazonGuianan ACL scenario may be of future concernunnatural parasitevectorreservoir systems when closelyrelated in an ecological context can on a century-level scale
8 Journal of Tropical Medicine
010203040506070
Bi flaviscutellata
01020304050607080
Ny umbratilis
0510152025303540
Ps squamiventris sl
0100200300400500600
050
100150200250300350400
2012 2013 2014 2015
Wat
erflo
odin
g
Ps s
quya
miv
entri
s sl
Bi fl
avisc
utell
ata
Ny u
mbr
atili
s
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
050
100150200250300350400
050
100150200250300350400
050
100150200250300350400
010203040506070
10000 20000 30000 40000000Rainfall (mm)
01020304050607080
10000 20000 30000 40000000Rainfall (mm)
Rainfall (mm)Overall
Rainfall (mm)
Rainfall (mm)
Rainfall (mm)
0100200300400500600
Ove
rall
10000 20000 30000 40000000Rainfall (mm)
05
10152025303540
10000 20000 30000 40000000Rainfall (mm)
r = 065 p = 000001
r = 048 p = 00021
r = 049 p = 00021
r = 065 p = 000001
Figure 3 Correlation between rainfall and phlebotomine putative vector species captured in Santo Antonio do Jari hydroelectric systemimpacted area between the states of Para and Amapa Brazil (2012ndash2015)
Journal of Tropical Medicine 9
2012 2013 2014 2015
Wat
erflo
odin
g
0100200300400500600
010203040506070
Bi flaviscutellata
01020304050607080
Ny umbratilis
0510152025303540
Ps squamiventris sl
Bi fl
avisc
utell
ata
05
1015202530
05
1015202530
05
1015202530
05
1015202530
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
0100200300400500600
Ove
rall
10 20 300ACL cases
010203040506070
10 20 300ACL cases
01020304050607080
Ny u
mbr
atili
s
10 20 300ACL cases
ACL casesOverall
ACL cases
ACL cases
ACL cases
05
10152025303540
Ps s
quam
iven
tris s
l
10 20 300ACL cases
r = 044 p = 00063
r = minus002 p = 00001
r = 048 p = 0002
r = 032 p = 00003
Figure 4 Correlation between ACL cases and phlebotomine putative vector species captured in Santo Antonio do Jari hydroelectric systemimpacted area between the states of Para and Amapa Brazil (2012ndash2015)
10 Journal of Tropical Medicine
undergo stress adaptation (evolutionary fitness) and formnew ecological systems driven by coevolution [39]
There is no doubt that Ny umbratilis is the main vectorof ACL in the Jari River Basin [15] Surprisingly the well-documented tree-dwelling behavior of this species [15 40]was not observed in CDC canopy captures in the presentstudy very little difference was found between the number ofindividuals captured in the three sampled strata (58 at 05m39 at 1m and 49 at 20m) Although Ny umbratilis is knownto climb into the canopy at night to feed on edentates (mainlysloths) [12 41] it is possible that an alternative terrestrialsource of blood such as rodents may be attracting femalestoward ground level Sloths are very slow moving and aretherefore commonly rescued for relocation during environ-mentalmonitoring operations their resulting rarenessmeansNy umbratilis has had to find other food sources Converselyblood meal analysis conducted during studies in a forestfragment of the Western Amazon has shown rodent bloodto be the predominant feeding source for Ny umbratilis [42]Wild and synanthropic rodents have been shown to surviveenvironmental disturbance in Jari and to proliferate in areasof introduced tree species [14] this indicated a plausiblereason not only for the putative adaptation of Ny umbratilisbut also for the increasing population levels of the rodentblood-feeder Bi flaviscutellata as already discussed
Given the clear importance of Psychodopygus of theChagasi series (formerly referred to as the Squamiventrisseries) in the context of ACL transmission [22] taxonomicstudies on identifying the cryptic females have been under-taken previously [43] and more recently with interestingresults (Rodrigo E Godoy unpublished) These techniqueshowever could not be employed in the field at the time Earlyconsideration of the assumed geographic distribution of thecomplex Ps squamiventris sl [1 44] shows that individualsfrom AP are likely to be predominantly Ps s maripaensisDetermining males of the species present could clarify thesetaxonomic uncertainties in the area covered by the presentstudy however doubts about identification of some malespecimens meant they were grouped as Ps squamiventris sl
Ryan et al [22] did not find Ps squamiventris sl to benaturally infected in the Jari area but proposed a link to L(V) naiffi along the same northern bank of the AmazonRiver through studies in Cachoeira Porteira (PA) Fouqueet al [38] came to the same conclusions in French GuianaInterestingly this fly species complex was found alongside L(V) braziliensis DNA in Suriname [45] an area where theknown Amazonian vectors of this parasite Ps complexus andPs wellcomei are unlikely to occur As previously discussedin the context of Bi flaviscutellata the changing transmissionpatterns of unnatural vectorparasite systems must be care-fully interpreted and only after strong ecological associationshave been determined Thus the putative vector importanceof Ps squamiventris sl in Jari can only be assumed in respectof L (V) naiffi
The five surveyed areas presented in general differentdiversity indexes showing variations in the phlebotominecomposition along the affected area This finding can beattributed not only to the distance between capture sitesbut also to forest composition fragmentation geographic
barriers and migration of mammal fauna between sites thislatter can occur naturally or be induced by rescue opera-tions involved in hydroelectric planning whereby animalsare moved to suitable ldquodry landrdquo environments safe frompotential flooding
A decrease in the phlebotomine population in 2014 inthe period after filling the reservoir could not be attributedexclusively either to flooding or to the transition from therainy to the dry season Both eventualities although oppositein effect are likely to lead to unsuitable conditions forphlebotomine reproduction However they did coincide inMay 2014 when rainfall decreased and the lake was formed
Conversely rainfall was a valid determinant of overallphlebotomine fluctuation Ps squamiventris sl was the mainputative vector directly correlated with this Other variablessuch as forest composition and altitudinal variation areknown to influence seasonal dynamics of species from thecomplex Ps squamiventris sl For example the dry periodfrom October to November in Serra dos Carajas (southernPA) can be unsuitable for Ps wellcomei which is adapted tohigh altitudes in contrast to Ps complexus which flourishesin lowland areas during the same months [46]
An insignificant negative to a moderate positive correla-tion was observed between cases of ACL and phlebotominefluctuations In the case of Ny umbratilis however theapparently higher correlation when compared with otherspecies is in agreement with its known vector importance inAmazonGuianan ACL A strong spatiotemporal associationbetween a given vector and ACL cases is considered essentialto maintenance of the Leishmania life cycle [2 14]
In summary the present study filled a temporal gapon phlebotomine data relating to the Jari area It providesa meaningful update on species composition and furtherinsights into seasonal dynamics of species under pressurescaused by environmental change The presence of ACLvectors in particular Ny umbratilis Bi flaviscutellata andPs squamiventris sl is noteworthy due to their directassociation with AmazonGuianan-ACL transmission ForNy umbratilis the lower frequency associatedwith the unpre-dicted lack of tree-dwelling behavior may be attributableto a reduction in numbers of canopy-dwelling mammalsdue to the environmental disturbance and relocation effortsthis could consequently have led to seeking for alternativeterrestrial blood sources at ground level The possible asso-ciation of Ps davisi with Leishmania transmission cycles inother Amazon ecoregions draws attention to its high rankposition in Jari Rainfall can be partly associated with localseasonal fluctuations in fly numbers mainly with respect toPs squamiventris sl Cases of ACL were also more highlycorrelated with Ny umbratilis dynamics Further attentionwas also given to results for Bi flaviscutellata which hasbeen apparently tolerant of anthropic activity continuedphlebotominemonitoring is proposed in order to evaluate theimplication for social welfare and health in the future
Competing Interests
The authors declare that they have no competing interests
Journal of Tropical Medicine 11
Acknowledgments
Theauthorswish to thank the ldquoInstituto de Pesquisas Cientıfi-cas e Tecnologicas do Estado do Amapa IEPArdquo entomologyteam for the technical assistance given to the field work inparticular to Aderbal Amanajas Santana Rosalvo Dias deCastro Noel Fernandes Santos Neto Osita Cleyriane Lobatodo Nascimento Jose Luiz dos Santos Chaves and Joao doSocorro Gomes Campos Logistical facilities were financiallysupported by the ldquoJari Energiardquo
References
[1] D G Young and M A Duncan ldquoGuide to the identificationand geographic distribution of Lutzomyia sand flies in Mexicothe West Indies Central and South America (Diptera Psycho-didae)rdquo inMemoirs of the American Entomological Institute pp1ndash881 Associated Publishers American Entomological InstituteGainesville Fla USA 1994
[2] P D Ready ldquoBiology of phlebotomine sand flies as vectors ofdisease agentsrdquo Annual Review of Entomology vol 58 pp 227ndash250 2013
[3] R P Brazil A A Fuzzari and J D Andrade Filho ldquoSandfly vectors of Leishmania in the Americasmdasha mini reviewrdquoEntomology Ornithology amp Herpetology vol 4 no 2 Article ID1000144 2015
[4] E F Rangel and R Lainson ldquoProven and putative vectorsof American cutaneous leishmaniasis in Brazil aspects oftheir biology and vectorial competencerdquoMemorias do InstitutoOswaldo Cruz vol 104 no 7 pp 937ndash954 2009
[5] E F Rangel S M Costa and B M Carvalho ldquoEnvironmentalchanges and the geographic spreading of American cutaneousleishmaniasis in Brazilrdquo in Leishmaniasis-Trends in Epidemiol-ogy D Claborn Ed Diagnosis and Treatment InTech RijekaCroatia 2014
[6] W P Taddei ldquoO genero Mansonia (Diptera Culicidae) e aproliferacao de mosquitos na usina hidreletrica de Tucuruırdquo inEnergia na Amazonia S B Magalhaes R C Brito and E RCastro Eds vol 1 pp 311ndash318 MPEGFPAUNAMAZ BelemBrazil 1996 (Portuguese)
[7] A I A Oliveira Observacoes Sobre a Resolucao CONAMA no23797 que Dispoe Sobre o Licenciamento Ambiental MimeoConselho Empresarial Brasileiro Para o Desenvolvimento Sus-tentavel Rio de Janeiro Brazil 1997 (Portuguese)
[8] A K R Galardo C D Galardo G A Silveira et al ldquoPhle-botominae sand flies (Diptera Psychodidae) potential vectorsof American cutaneous leishmaniasis agents in the area asso-ciated with the Santo Antonio hydroelectric system in WesternAmazonian Brazilrdquo Revista da Sociedade Brasileira deMedicinaTropical vol 48 no 3 pp 265ndash271 2015
[9] R Lainson R DWard and J J Shaw ldquoCutaneous leishmaniasisin North Brazil Lutzomyia anduzei as a major vectorrdquo Transac-tions of the Royal Society of Tropical Medicine and Hygiene vol70 no 2 pp 171ndash172 1976
[10] R Lainson J J Shaw R D Ward P D Ready and R DNaiff ldquoLeishmaniasis in Brazil XIII Isolation of Leishmaniafrom armadillos (Dasypus novemcinctus) and observations onthe epidemiology of cutaneous leishmaniasis in north ParaStaterdquo Transactions of the Royal Society of Tropical Medicine andHygiene vol 73 no 2 pp 239ndash242 1979
[11] R Lainson J J Shaw P D Ready M A Miles and M PovoaldquoLeishmaniasis in Brazil XVI Isolation and identification of
Leishmania species from sandflies wild mammals and manin north Para State with particular reference to L braziliensisguyanensis causative agent of ldquopian-boisrdquordquo Transactions of theRoyal Society of Tropical Medicine and Hygiene vol 75 no 4pp 530ndash536 1981
[12] R Lainson J J Shaw and M Povoa ldquoThe importance of eden-tates (sloths and anteaters) as primary reservoirs of Leishmaniabraziliensis guyanensis causative agent of lsquopian boisrsquo in northBrazilrdquo Transactions of the Royal Society of Tropical Medicineand Hygiene vol 75 no 4 pp 611ndash612 1981
[13] R DWard and H Fraiha ldquoLutzomyia umbratilis a new speciesof sand fly from Brazil (Diptera Psychodidae)rdquo Journal ofMedical Entomology vol 14 no 3 pp 313ndash317 1977
[14] P D Ready R Lainson and J J Shaw ldquoLeishmaniasis in BrazilXX Prevalence of lsquoenzootic rodent leishmaniasisrsquo (Leishmaniamexicana amazonensis) and apparent absence of lsquopian boisrsquo(Le braziliensis guyanensis) in plantations of introduced treespecies and in other non-climax forests in eastern AmazoniardquoTransactions of the Royal Society of Tropical Medicine andHygiene vol 77 no 6 pp 775ndash785 1983
[15] P D Ready R Lainson J J Shaw and R DWard ldquoThe ecologyof Lutzomyia umbratilisWard amp Fraiha (Diptera Psychodidae)the major vector to man of Leishmania braziliensis guyanensisin north-eastern Amazonian Brazilrdquo Bulletin of EntomologicalResearch vol 76 no 1 pp 21ndash40 1986
[16] R Lainson and J J Shaw ldquoLeishmania (Viannia) naiffi spn a parasite of the armadillo Dasypus novemcinctus (L)in Amazonian Brazilrdquo Annales de Parasitologie Humaine etComparee vol 64 no 1 pp 3ndash9 1989
[17] A Roger M Nacher M Hanf et al ldquoClimate and leishmaniasisin French guianardquo The American Journal of Tropical Medicineand Hygiene vol 89 no 3 pp 564ndash569 2013
[18] L Ryan ldquoFlebotomos do estado do Para Brasil (Diptera Psy-chodidae)rdquo Thecnical Document 1 Instituto Evandro ChagasBelem Brazil 1986 (Portuguese)
[19] E A B Galati ldquoMorfologia e Taxonomia Morfologia termi-nologia de adultos e identificacao dos taxons da Americardquoin Flebotomıneos do Brasil E F Rangel and R LainsonEds pp 53ndash75 Editora Fiocruz Rio de Janeiro Brazil 2003(Portuguese)
[20] D R Roberts and B P Hsi ldquoAn index of species abundance foruse with mosquito surveillance datardquo Environmental Entomol-ogy vol 8 no 6 pp 1007ndash1013 1979
[21] Oslash Hammer D A T Harper and P D Ryan ldquoPast paleontolog-ical statistics software package for education and data analysisrdquoPalaeontologia Electronica vol 4 no 1 pp 1ndash9 2001
[22] L Ryan R Lainson J J Shaw R R Braga and E A IshikawaldquoLeishmaniasis in Brazil XXV Sandfly vectors of Leishmania inPara State Brazilrdquo Medical and Veterinary Entomology vol 1no 4 pp 383ndash395 1987
[23] R Lainson J J Shaw F T Silveira R R Braga and E AIshikawa ldquoCutaneous leishmaniasis of man due to Leishmania(Viannia) naiffirdquo Annales de Parasitologie Humaine et Compa-ree vol 65 no 5-6 pp 282ndash284 1990
[24] E G C Bermudez Lutzomyia Sand Flies in the BrazilianAmazon Basin (Diptera Psychodidae) INPA Manaus Brazil2009
[25] G M Aguiar and W M Medeiros ldquoDistribuicao regionale habitats das especies de flebotomıneos do Brasilrdquo in Fle-botomıneos do Brasil E F Rangel and R Lainson Eds pp 207ndash256 Fiocruz Rio de Janeiro Brazil 2003 (Portuguese)
12 Journal of Tropical Medicine
[26] J R Arias M A Miles R D Naiff et al ldquoFlagellate infectionsof Brazilian sand flies (Diptera Psychodidae) isolation invitro and biochemical identification of Endotrypanum andLeishmaniardquo The American Journal of Tropical Medicine andHygiene vol 34 no 6 pp 1098ndash1108 1985
[27] A A Souza T V Santos Y L Jennings et al ldquoNaturalLeishmania(Viannia) spp infections in phlebotomine sand flies(Diptera Psychodidae) from the Brazilian Amazon regionreveal new putative transmission cycles of American cutaneousleishmaniasisrdquo Parasite vol 23 article 22 2016
[28] F T Silveira A A Souza R Lainson J J Shaw R R Bragaand E A Ishikawa ldquoCutaneous leishmaniasis in the AmazonRegion natural infection of the sand fly Lutzomyia ubiquitalis(Psychodidae Phlebotominae) by Leishmania (Viannia) lain-soni in Para state BrazilrdquoMemorias do Instituto Oswaldo Cruzvol 86 no 1 pp 127ndash130 1991
[29] LH S Gil S A Basano A A Souza et al ldquoRecent observationson the sand fly (Diptera Psychodidae) fauna of the state ofRondonia Western Amazonia Brazil the importance of Psych-dopygus davisi as a vector of zoonotic cutaneous leishmaniasisrdquoMemorias do Instituto Oswaldo Cruz vol 98 no 6 pp 751ndash7552003
[30] A A A Souza F T Silveira R Lainson et al ldquoThe Phlebotom-inae fauna of Serra dos Carajas Para Brazil and its possibleimplication in the transmission of American tegumentaryleishmaniasisrdquo Revista Pan-Amazonica de Saude vol 1 no 1pp 45ndash51 2010
[31] R Killick-Kendrick ldquoPhlebotomine vectors of the leishmani-ases a reviewrdquoMedical and Veterinary Entomology vol 4 no 1pp 1ndash24 1990
[32] M E C Dorval G Cristaldo H C da Rocha et al ldquoPhle-botomine fauna (Diptera Psychodidae) of an American cuta-neous leishmaniasis endemic area in the state of Mato Grossodo Sul Brazilrdquo Memorias do Instituto Oswaldo Cruz vol 104no 5 pp 695ndash702 2009
[33] V C F V Guimaraes P L Costa F J da Silva et al ldquoMoleculardetection of Leishmania in phlebotomine sand flies in a cuta-neous and visceral leishmaniasis endemic area in northeasternBrazilrdquo Revista do Instituto de Medicina Tropical de Sao Paulovol 56 no 4 pp 357ndash360 2014
[34] R S Lana E M Michalsky C L Fortes-Dias et al ldquoPhle-botomine sand fly fauna and Leishmania infection in thevicinity of the Serra do Cipo National Park a natural Brazilianheritage siterdquo BioMed Research International vol 2015 ArticleID 385493 9 pages 2015
[35] V R Alves R A de Freitas F L Santos A F J de OliveiraT V Barrett and P H F Shimabukuro ldquoSand flies (DipteraPsychodidae Phlebotominae) from central amazonia and fournew records for the Amazonas State Brazilrdquo Revista Brasileirade Entomologia vol 56 no 2 pp 220ndash227 2012
[36] J V S Ferreira T V Santos E M Santos and I S GorayebldquoPhlebotomine sand flies (Diptera Psychodidae) in forestfragments of Belem metropolitan area Para State Brazil withconsiderations on vectors of American cutaneous leishmaniasisagentsrdquo Revista Pan-Amazonica de Saude vol 5 no 2 pp 29ndash35 2014
[37] B M Carvalho E F Rangel P D Ready and M M ValeldquoEcological niche modelling predicts southward expansion ofLutzomyia (Nyssomyia) flaviscutellata (Diptera PsychodidaePhlebotominae) vector of Leishmania (Leishmania) amazonen-sis in South America under climate changerdquo PLoS ONE vol 10no 11 Article ID e0143282 2015
[38] F Fouque P Gaborit J Issaly et al ldquoPhlebotomine sand flies(Diptera Psychodidae) associatedwith changing patterns in thetransmission of the human cutaneous leishmaniasis in FrenchGuianardquoMemorias do Instituto Oswaldo Cruz vol 102 no 1 pp35ndash40 2007
[39] R Killick-Kendrick ldquoSome epidemiological consequences ofthe evolutionary fit between Leishmaniae and their phle-botomine vectorsrdquo Bulletin de la Societe de Pathologie Exotiqueet de ses Filiales vol 78 no 5 pp 747ndash755 1985
[40] F X Pajot F Le Pont BGentile andR Besnard ldquoEpidemiologyof leishmaniasis in French Guianardquo Transactions of the RoyalSociety of Tropical Medicine and Hygiene vol 76 no 1 pp 112ndash113 1982
[41] H A Christensen J R Arias A M De Vasquez and R ADe Freitas ldquoHosts of sandfly vectors of Leishmania braziliensisguyanensis in the central Amazon of Brazilrdquo The AmericanJournal of Tropical Medicine and Hygiene vol 31 no 2 pp 239ndash242 1982
[42] L C R Nery E S Lorosa andAM R Franco ldquoFeeding prefer-ence of the sand flies Lutzomyia umbratilis and L spathotrichia(Diptera Psychodidae Phlebotominae) in an urban forest patchin the City ofManaus Amazonas BrazilrdquoMemorias do InstitutoOswaldo Cruz vol 99 no 6 pp 571ndash574 2004
[43] PD Ready R Lainson J J Shaw andAA Souza ldquoDNAprobesfor distinguishing Psychodopygus wellcomei from Psychodopy-gus complexus (Diptera Psychodidae)rdquo Memorias do InstitutoOswaldo Cruz vol 86 no 1 pp 41ndash49 1991
[44] P D Ready H Fraiha R P Lane J R Arias and F X PajotldquoOn distinguishing the female of Psychodopygus wellcomei avector of mucocutaneous leishmaniasis from other squamiven-tris series females I Characterization of Ps squamiventrissquamiventris and Ps s maripaensis stat nov (Diptera Psycho-didae)rdquo Annals of Tropical Medicine and Parasitology vol 76no 2 pp 201ndash214 1982
[45] A D Kent T V dos Santos A Gangadin A Samjhawan DR A Mans and H D F H Schallig ldquoStudies on the sandfly fauna (Diptera Psychodidae) in high-transmission areas ofcutaneous leishmaniasis in the Republic of Surinamerdquo Parasitesand Vectors vol 6 no 1 article 318 2013
[46] P D Ready R Lainson and J J Shaw ldquoHabitat and seasonalityof Psychodopygus wellcomei help incriminate it as a vector ofLeishmania braziliensis in Amazonia and Northeast BrazilrdquoTransactions of the Royal Society of Tropical Medicine andHygiene vol 78 no 4 pp 543ndash544 1984
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

4 Journal of Tropical MedicineTa
ble1Ph
ebotom
inespeciescompo
sitionofSantoAnton
iodo
Jarihydroelectric
syste
mim
pacted
area
(AmapaS
tateA
mazon
ianBrazil)
ontheb
asisoffivesurveyedlocatio
ns(2012ndash2015)
Species
PortoSabao
Traıra
River
Iratapuru
Santo
Power
Plant
ISA
SISA
Rank
Total
Village
Anton
ioVillage
CD
CD
CD
CD
CD
1Sciopemyiasordellii
149
6613
1015
442
16mdash
136
087
2nd
316
1129
2Psycho
dopygu
sdavisi
4637
9556
1510
3817
mdashmdash
64
074
3rd
314
1121
mdashTrich
ophoromyiaspp
177
mdash34
mdash40
mdash17
mdashmdash
mdashmdash
mdashmdash
268
957
3Bichromom
yiafla
viscutellata
7632
2414
2211
5231
12
2095
1st
265
946
4Evandrom
yiainfra
spinosa
4410
2518
32
3018
mdashmdash
9206
6th
150
536
5Ny
ssom
yiaum
bratilis
2612
3130
196
139
mdashmdash
86
063
5th
146
521
6Trich
ophoromyiacasta
nheir
aimdash
104
mdash6
mdash14
mdash11
mdashmdash
112
051
9th
135
482
mdashPressatia
spp
43mdash
1mdash
9mdash
78mdash
mdashmdash
mdashmdash
mdash131
468
mdashEvandrom
yiaspp
5210
4mdash
3911
12mdash
mdashmdash
mdashmdash
mdash128
457
7Viannamyiafurcata
444
115
111
305
mdashmdash
104
055
7th
111
396
8Psycho
dopygu
ssqu
amiventrissensulato
3614
155
10mdash
9mdash
mdashmdash
11052
8th
89318
9Trich
opygom
yiatrichopyga
1116
98
9mdash
2112
mdashmdash
7207
4th
86307
10Tricho
phorom
yiaub
iquitalis
216
58
148
139
2mdash
114
05
10th
86307
11Evandrom
yiasaulensis
409
5mdash
44
121
mdashmdash
128
043
11th
75268
12Psycho
dopygu
shirs
utus
hirsutus
13mdash
204
144
mdashmdash
mdashmdash
136
04
12th
5519
613
Trich
ophoromyiabrachypyga
mdash36
mdash12
mdash2
mdash2
mdashmdash
152
032
14th
5218
614
Pressatia
choti
mdash17
mdash3
mdash5
mdash25
mdashmdash
142
037
13th
5017
915
Nyssom
yiaan
duzei
65
2410
mdashmdash
mdashmdash
mdashmdash
17023
16th
4516
116
Lutzom
yiagomezi
2711
mdashmdash
mdashmdash
mdashmdash
mdashmdash
194
012
22th
3813
6mdash
Nysso
myiaspp
mdashmdash
mdash1
1mdash
1125
mdashmdash
mdashmdash
mdash38
136
17Ny
ssom
yiaan
tunesi
mdash2
9mdash
52
mdashmdash
1mdash
142
037
13th
19068
18Sciopemyiaflu
viatilis
21
3mdash
91
mdashmdash
mdashmdash
18019
18th
16057
19Micropygom
yiarorotaensis
4mdash
mdashmdash
8mdash
21
mdashmdash
178
02
17th
15054
20Psycho
dopygu
sparaensis
mdashmdash
mdashmdash
63
51
mdashmdash
176
02
17th
15054
21Psychodopygusclaustre
i7
1mdash
mdashmdash
mdash2
mdash3
1154
031
15th
14050
mdashPsychodopygusspp
4
mdash2
mdash6
mdash2
mdashmdash
mdashmdash
mdashmdash
14050
22Trich
ophoromyiaeurypyga
mdash8
mdashmdash
mdash3
mdash1
mdashmdash
182
018
19th
12043
23Psathyromyiaaragaoi
mdashmdash
52
mdashmdash
4mdash
mdashmdash
188
015
20th
11039
24Evandrom
yiabrachyphalla
43
21
1mdash
mdashmdash
mdashmdash
19014
21th
11039
25Psychodopyguscarreraicarrerai
11
mdash7
mdashmdash
mdashmdash
mdashmdash
202
008
26th
9032
26Evandrom
yiabegona
e2
2mdash
mdash4
mdashmdash
mdashmdash
mdash196
011
23th
8029
27Ny
ssom
yiawh
itman
imdash
mdashmdash
mdashmdash
mdash8
mdashmdash
mdash20
009
25th
8029
28Psathyromyiascaffi
32
mdashmdash
2mdash
mdashmdash
mdashmdash
198
01
24th
7025
29Pressatia
trispinosa
mdashmdash
mdash7
mdashmdash
mdashmdash
mdashmdash
204
007
27th
7025
30Micropygom
yiatrinidadensis
7mdash
mdashmdash
mdashmdash
mdashmdash
mdashmdash
208
005
29th
7025
31Viannamyiatuberculata
6mdash
mdashmdash
mdashmdash
mdashmdash
mdashmdash
21004
30th
6021
32Pintom
yiadamascenoi
mdashmdash
mdashmdash
mdashmdash
5mdash
mdashmdash
20009
25th
5018
33Evandrom
yiaevandroi
mdashmdash
mdashmdash
41
mdashmdash
mdashmdash
204
007
27th
5018
34Psathyromyialutziana
5mdash
mdashmdash
mdashmdash
mdashmdash
mdashmdash
212
003
31th
5018
35Brum
ptom
iatra
vasso
simdash
mdashmdash
mdashmdash
mdash3
1mdash
mdash196
011
23th
4014
36Evandrom
yiabacula
mdashmdash
mdashmdash
1mdash
3mdash
mdashmdash
202
008
26th
4014
37Evandrom
yiamonstruosa
mdashmdash
4mdash
mdashmdash
mdashmdash
mdashmdash
206
006
28th
4014
Journal of Tropical Medicine 5
Table1Con
tinued
Species
PortoSabao
Traıra
River
Iratapuru
Santo
Power
Plant
ISA
SISA
Rank
Total
Village
Anton
ioVillage
CD
CD
CD
CD
CD
38Psychodopygusa
mazonensis
mdashmdash
mdashmdash
22
mdashmdash
mdashmdash
21004
30th
4014
39Psychodopyguscorossoniensis
mdashmdash
mdashmdash
mdash2
mdashmdash
mdashmdash
206
006
28th
2007
40Pressatia
equatoria
lismdash
2mdash
mdashmdash
mdashmdash
mdashmdash
mdash21
004
30th
2007
41Psathyromyiainflata
mdashmdash
mdashmdash
mdashmdash
2mdash
mdashmdash
21004
30th
2007
42Micropygom
yiamicropyga
mdashmdash
mdashmdash
mdashmdash
11
mdashmdash
214
002
32th
2007
43Psychodopygusgenicu
latus
mdashmdash
mdash2
mdashmdash
mdashmdash
mdashmdash
218
033th
2007
44Lu
tzom
yiacarvalhoi
mdashmdash
1mdash
mdashmdash
mdashmdash
mdashmdash
208
005
29th
1004
45Psathyromyiapunctigenicu
lata
mdashmdash
mdashmdash
1mdash
mdashmdash
mdashmdash
216
001
32th
1004
Total
856
411
342
209
274
96415
186
74
2800
1000
1267
551
370
601
11Ea
chlocatio
ncomprise
dam
onito
ringstation(M
S)sampled
with
051and20
mCD
Ctrapsprovidingapproxim
ately
504h
MS
Cfem
aleDm
aleISAind
exof
speciesa
bund
anceSISAstand
ardindexof
speciesa
bund
ance
ldquothe
bold
italic
fontrdquoP
rovenandor
putativ
eACL
vectorsintheA
mazon
Region
based
inYo
ungandDun
can[1]Ra
ngelandLainson[4]Re
ady[2]andLainsonandShaw
[16]
6 Journal of Tropical Medicine
Table 2 Vertical stratification of Phebotomine species composition of Santo Antonio do Jari hydroelectric system impacted area (AmapaState Amazonian Brazil) on the basis of five surveyed locations (2012ndash2015)
Species 05m ground 1m ground 20m canopy TotalC D C D C D
1 Sciopemyia sordellii 117 71 91 23 11 3 3162 Psychodopygus davisi 48 27 44 39 102 54 314mdash Trichophoromyia spp 117 mdash 127 mdash 24 mdash 2683 Bichromomyia flaviscutellata 85 45 79 41 11 4 2654 Evandromyia infraspinosa 38 18 48 27 16 3 1505 Nyssomyia umbratilis 35 23 26 13 28 21 1466 Trichophoromyia castanheirai mdash 50 mdash 59 mdash 26 135mdash Pressatia spp 66 mdash 39 mdash 26 mdash 131mdash Evandromyia spp 55 12 41 9 11 mdash 1287 Vianamyia furcata 42 8 46 6 8 1 1118 Psychodopygus squamiventris sl 27 11 20 1 23 7 899 Trichopygomyia trichopyga 18 20 10 5 22 11 8610 Trichophoromyia ubiquitalis 28 15 16 13 9 5 86mdash Other species (11ndash45) 137 112 129 69 71 57 575
Total 813 412 716 305 362 192 2800Total (C + D) 1225 1021 554 mdash
Each stratum was sampled with approximately 840 h of CDC trapC female D male
infraspinosa (119899 = 150 SISA = 06) and Ny umbratilis (119899 =146 SISA = 063) Less frequently captured but nonethelessepidemiologically relevant species included Ps squamiventrissl (119899 = 89 SISA = 052) Th ubiquitalis (119899 = 86 SISA=05)Ps hirsutus hirsutus (119899 = 55 SISA = 04)Ny anduzei (119899 = 45SISA = 023) Lutzomyia gomezi (119899 = 38 SISA = 012) Nyantunesi (119899 = 19 SISA = 037) Ps paraensis (119899 = 15 SISA =02) and Ny whitmani (119899 = 8 SISA = 009)
Sampling effort for each area comprised approximately504 h of CDC trapping (14 expeditions 3 CDC traps and12-hour CDC exposition) Porto Sabao was the site of themost captured specimens (1267) followed by Santo AntonioVillage (601) The highest diversity was observed in capturesfrom Iratapuru Village (1198671015840 = 3027) All 1198671015840 value com-parisons between the different environments were significant(119901 le 005) except when comparing Traıra River and thePower Plant (119901 le 0099) (Figure 2)
Regarding vertical distribution sampling of the samecaptures described above accounted for approximately 840 hof CDC trapping by stratum (14 expeditions 5 ecotopes and12-hour CDC exposition) CDC traps placed 05m aboveground level accounted for a greater number of individuals(1225) than those placed 1m above ground level (1021) and20m above ground level in the canopy (554) (Table 2) WhilePs davisi was most likely to be found at canopy level (75individuals at 05m 83 at 1m and 156 at 20m) and Biflaviscutellata closer to ground level (130 at 05m 120 at 1mand 15 at 20m) there was little difference in numbers of Nyumbratilis individuals captured at the different levels of strata(58 at 05m 39 at 1m and 49 at 20m)
The monthly average rainfall was the only climatic datawith a representative coefficient of variance (1198781015840 = 7081)whentested against ACL occurrence and phlebotomine frequency
005
115
225
335
PortoSabatildeo
TraiacuteraRiver
IratapuruVillage
Santo AntocircnioVillage
PowerPlant
p = 0009p = 0099
p = 0007
p = 0012p = 0001
p = 0031
p = 0001p = 0004
p = 0001p = 00012855
26543027
2819
1468
Figure 2 Comparison of Shannon diversity indexes for CDC-captured phlebotomines from the five surveyed locations in SantoAntonio do Jari hydroelectric system impacted area between thestates of Para and Amapa Brazil (2012ndash2015)
(1198781015840 = 6595) in contrast to temperature (1198781015840 = 596)and relative air humidity (1198781015840 = 956) The available dataenabled correlations to be determined between the periodsof May 2012 to April 2015 a period within which nine of thefourteen capture efforts were undertaken Rainfall presenteda moderate correlation with overall phlebotomine frequency(119903 = 065 119901 = 000001) as also individually compared withPs squamiventris sl (119903 = 065 119901 = 000001) and a similarlymoderate but lower correlation with Bi flaviscutellata (119903 =048 119901 = 00021) and Ny umbratilis (119903 = 048 119901 = 0002)
Journal of Tropical Medicine 7
A decrease in the overall phlebotomine population wasobserved after March until November 2014 coinciding withthe transition between the rainy and dry periods and withflooding (Figure 3)
The number of ACL cases presented a moderate cor-relation with overall phlebotomine frequency (119903 = 044119901 = 00063) and also with Ps squamiventris (119903 = 032119901 = 00003) and an insignificant negative correlation with Biflaviscutellata (119903 = minus002 119901 = 00001) A similarly moderatebut higher correlation was however found withNy umbratilis(119903 = 048 119901 = 0002) (Figure 4)
4 Discussion
It is important to consider current investigations of phle-botomine fauna in the Jari River Basin in the context ofthe historical findings from the 1970s and 1980s Theseearlier studies represent comprehensive research conductedin Monte Dourado (northern PA) a region close to thearea investigated in the present study and on which thepresent epidemiological background to ACL transmissionin the associated region is based In brief the previousresearch includes a description of Ny umbratilis (formerlyldquoobscuredrdquo as Ny anduzei) [13] and its incrimination as themain vector of L (V) guyanensis [9ndash11] confirmed by strongecological associations [15] Just as important a parasitemore recently described as L (V) naiffi was isolated fromthe armadilloDasypus novemcinctus for which transmissionwas locally driven and attributed to Ps squamiventris sl andother Psychodopygus species [22] this comprised a primarilyenzootic ACL occasionally infecting humans [23] Finallythe isolation of L (L) amazonensis from rodents in an areaaffected by occurrence of the main vector Bi flaviscutellata[11] comprised the third known Jari ACL transmission cycleThese facts described above suggest that Ny umbratilis Pssquamiventris sl and Bi flaviscutellata are implicated in thetransmission scenario of the area covered by the present studyand thus merit particular attention
The present study incorporated a representative compo-sition of the known AP fauna Considering the substantialsampling effort (2520 h) the determined diversity showedagreement with the findings of other inventories from theBrazilian Amazon basin [1 24] Moreover all new records forAP have already been recorded for PA [25]
Apart from Ny umbratilis Ps squamiventris sl and Biflaviscutellata other species captured were noted as beingepidemiologically relevant to ACL in the Brazilian AmazonRegion Ps paraensis Ps davisi and Ps hirsutus hirsutuswhich are putative vectors of L (V) naiffi [26 27] Thubiquitalis a proven vector of L (V) lainsoni [28] Nyanduzei a secondary vector of L (V) guyanensis [4] Nywhitmani and Lu gomezi proven and putative vectors of L(V) shawi respectively [4 26] and lastly Ny antunesi asuspected vector of L (V) lindenbergi [4]
Among the above species Ps davisi is notable as thethird highest ranking according to SISA (074) and as havingthe highest presence in all vertical strata in particular atthe canopy level This species is generally anthropophilic iswidely distributed in the Amazon Basin [1 24] and is related
to Leishmania (Viannia) infections in the States of Rondonia(RO) [29] and PA [27 30] Interestingly this species remainsfrequent in recently surveyed areas affected by hydroelectricschemes in RO [8] and PA (TVS unpublished) Theseobservations add weight to the need for further investigationinto the putative epidemiological role of Ps davisi in Jari
The species most frequently captured Sc sordellii wouldappear not to be of epidemiological concern Firstly theapparent abundance of a given phlebotomine species is notsufficient of itself to incriminate it as a vector [31] particularlywhen it does not demonstrate high levels of anthropophilicbehavior Secondly flagellate infections isolated from thisspecies (formerly referred to as Lutzomyia nordestina) arehistorically recognized as distinct from Leishmania [22] orremain undetermined [32] and are more likely to be othertrypanosomatids [22] Furthermore despite recent reports ofmolecular Leishmania ascribed to this fly [33 34] currentlypolymerase chain reaction-based techniques do not allowldquotrue infectionrdquo to be distinguished from a ldquosimple and occa-sionally ingestedrdquo DNA fragment [3] Thus there remainsinsufficient evidence for a potential vector role of S sordelliiin Leishmania transmission
High frequencies of light-trapped Bi flaviscutellata havebeen documented [8 35] despite rodent-baited trappingbeing better recognized as amethod for attracting the speciesAdditionally the first SISA Rank (095) was indicative ofthis high presence in all five surveyed ecotopes this hasbeen observed in the Belem metropolitan area (PA) whereBi flaviscutellata was the only omnipresent species in sevenurban and ecologically isolated forest fragments surveyed[36] Furthermore this species has been reported as beingable to adapt to plantations of introduced tree species and inother nonclimax forests in Jari [14] suggesting it is capableof responding positively to environmental change such asthe vegetal suppression and flooding caused by hydroelectricoperations Nonetheless ecological niche model projectionspredict no change in spatial distribution for Bi flaviscutellatain the Jari area which is an environment that remains suitablefor the species [37]
In an area in Regina French Guiana ecologically dis-turbed due to agricultural practices flagellates infecting a Biflaviscutellata specimen have been determined to be L (V)guyanensis [38] in similar circumstances to the observed lowpresence of Ny umbratilis Although this possibly changingpattern in ACL transmission should be interpreted withcaution it raises an intriguing possibility about the permis-siveness of Bi flaviscutellata to L (V) guyanensis in theAmazonGuianan ACL scenario
In the present study a higher number of Bi flaviscutellataindividuals was captured compared to Ny umbratilis (265versus 146 resp) A lack of available evidence for naturalinfection in this unnatural vectorparasite system howeverprecludes speculation about the possible involvement of Biflaviscutellata even in the context of occasional transmissionof parasites distinct from L (L) amazonensis Neverthelessthe current high occurrence of Bi flaviscutellata in theAmazonGuianan ACL scenario may be of future concernunnatural parasitevectorreservoir systems when closelyrelated in an ecological context can on a century-level scale
8 Journal of Tropical Medicine
010203040506070
Bi flaviscutellata
01020304050607080
Ny umbratilis
0510152025303540
Ps squamiventris sl
0100200300400500600
050
100150200250300350400
2012 2013 2014 2015
Wat
erflo
odin
g
Ps s
quya
miv
entri
s sl
Bi fl
avisc
utell
ata
Ny u
mbr
atili
s
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
050
100150200250300350400
050
100150200250300350400
050
100150200250300350400
010203040506070
10000 20000 30000 40000000Rainfall (mm)
01020304050607080
10000 20000 30000 40000000Rainfall (mm)
Rainfall (mm)Overall
Rainfall (mm)
Rainfall (mm)
Rainfall (mm)
0100200300400500600
Ove
rall
10000 20000 30000 40000000Rainfall (mm)
05
10152025303540
10000 20000 30000 40000000Rainfall (mm)
r = 065 p = 000001
r = 048 p = 00021
r = 049 p = 00021
r = 065 p = 000001
Figure 3 Correlation between rainfall and phlebotomine putative vector species captured in Santo Antonio do Jari hydroelectric systemimpacted area between the states of Para and Amapa Brazil (2012ndash2015)
Journal of Tropical Medicine 9
2012 2013 2014 2015
Wat
erflo
odin
g
0100200300400500600
010203040506070
Bi flaviscutellata
01020304050607080
Ny umbratilis
0510152025303540
Ps squamiventris sl
Bi fl
avisc
utell
ata
05
1015202530
05
1015202530
05
1015202530
05
1015202530
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
0100200300400500600
Ove
rall
10 20 300ACL cases
010203040506070
10 20 300ACL cases
01020304050607080
Ny u
mbr
atili
s
10 20 300ACL cases
ACL casesOverall
ACL cases
ACL cases
ACL cases
05
10152025303540
Ps s
quam
iven
tris s
l
10 20 300ACL cases
r = 044 p = 00063
r = minus002 p = 00001
r = 048 p = 0002
r = 032 p = 00003
Figure 4 Correlation between ACL cases and phlebotomine putative vector species captured in Santo Antonio do Jari hydroelectric systemimpacted area between the states of Para and Amapa Brazil (2012ndash2015)
10 Journal of Tropical Medicine
undergo stress adaptation (evolutionary fitness) and formnew ecological systems driven by coevolution [39]
There is no doubt that Ny umbratilis is the main vectorof ACL in the Jari River Basin [15] Surprisingly the well-documented tree-dwelling behavior of this species [15 40]was not observed in CDC canopy captures in the presentstudy very little difference was found between the number ofindividuals captured in the three sampled strata (58 at 05m39 at 1m and 49 at 20m) Although Ny umbratilis is knownto climb into the canopy at night to feed on edentates (mainlysloths) [12 41] it is possible that an alternative terrestrialsource of blood such as rodents may be attracting femalestoward ground level Sloths are very slow moving and aretherefore commonly rescued for relocation during environ-mentalmonitoring operations their resulting rarenessmeansNy umbratilis has had to find other food sources Converselyblood meal analysis conducted during studies in a forestfragment of the Western Amazon has shown rodent bloodto be the predominant feeding source for Ny umbratilis [42]Wild and synanthropic rodents have been shown to surviveenvironmental disturbance in Jari and to proliferate in areasof introduced tree species [14] this indicated a plausiblereason not only for the putative adaptation of Ny umbratilisbut also for the increasing population levels of the rodentblood-feeder Bi flaviscutellata as already discussed
Given the clear importance of Psychodopygus of theChagasi series (formerly referred to as the Squamiventrisseries) in the context of ACL transmission [22] taxonomicstudies on identifying the cryptic females have been under-taken previously [43] and more recently with interestingresults (Rodrigo E Godoy unpublished) These techniqueshowever could not be employed in the field at the time Earlyconsideration of the assumed geographic distribution of thecomplex Ps squamiventris sl [1 44] shows that individualsfrom AP are likely to be predominantly Ps s maripaensisDetermining males of the species present could clarify thesetaxonomic uncertainties in the area covered by the presentstudy however doubts about identification of some malespecimens meant they were grouped as Ps squamiventris sl
Ryan et al [22] did not find Ps squamiventris sl to benaturally infected in the Jari area but proposed a link to L(V) naiffi along the same northern bank of the AmazonRiver through studies in Cachoeira Porteira (PA) Fouqueet al [38] came to the same conclusions in French GuianaInterestingly this fly species complex was found alongside L(V) braziliensis DNA in Suriname [45] an area where theknown Amazonian vectors of this parasite Ps complexus andPs wellcomei are unlikely to occur As previously discussedin the context of Bi flaviscutellata the changing transmissionpatterns of unnatural vectorparasite systems must be care-fully interpreted and only after strong ecological associationshave been determined Thus the putative vector importanceof Ps squamiventris sl in Jari can only be assumed in respectof L (V) naiffi
The five surveyed areas presented in general differentdiversity indexes showing variations in the phlebotominecomposition along the affected area This finding can beattributed not only to the distance between capture sitesbut also to forest composition fragmentation geographic
barriers and migration of mammal fauna between sites thislatter can occur naturally or be induced by rescue opera-tions involved in hydroelectric planning whereby animalsare moved to suitable ldquodry landrdquo environments safe frompotential flooding
A decrease in the phlebotomine population in 2014 inthe period after filling the reservoir could not be attributedexclusively either to flooding or to the transition from therainy to the dry season Both eventualities although oppositein effect are likely to lead to unsuitable conditions forphlebotomine reproduction However they did coincide inMay 2014 when rainfall decreased and the lake was formed
Conversely rainfall was a valid determinant of overallphlebotomine fluctuation Ps squamiventris sl was the mainputative vector directly correlated with this Other variablessuch as forest composition and altitudinal variation areknown to influence seasonal dynamics of species from thecomplex Ps squamiventris sl For example the dry periodfrom October to November in Serra dos Carajas (southernPA) can be unsuitable for Ps wellcomei which is adapted tohigh altitudes in contrast to Ps complexus which flourishesin lowland areas during the same months [46]
An insignificant negative to a moderate positive correla-tion was observed between cases of ACL and phlebotominefluctuations In the case of Ny umbratilis however theapparently higher correlation when compared with otherspecies is in agreement with its known vector importance inAmazonGuianan ACL A strong spatiotemporal associationbetween a given vector and ACL cases is considered essentialto maintenance of the Leishmania life cycle [2 14]
In summary the present study filled a temporal gapon phlebotomine data relating to the Jari area It providesa meaningful update on species composition and furtherinsights into seasonal dynamics of species under pressurescaused by environmental change The presence of ACLvectors in particular Ny umbratilis Bi flaviscutellata andPs squamiventris sl is noteworthy due to their directassociation with AmazonGuianan-ACL transmission ForNy umbratilis the lower frequency associatedwith the unpre-dicted lack of tree-dwelling behavior may be attributableto a reduction in numbers of canopy-dwelling mammalsdue to the environmental disturbance and relocation effortsthis could consequently have led to seeking for alternativeterrestrial blood sources at ground level The possible asso-ciation of Ps davisi with Leishmania transmission cycles inother Amazon ecoregions draws attention to its high rankposition in Jari Rainfall can be partly associated with localseasonal fluctuations in fly numbers mainly with respect toPs squamiventris sl Cases of ACL were also more highlycorrelated with Ny umbratilis dynamics Further attentionwas also given to results for Bi flaviscutellata which hasbeen apparently tolerant of anthropic activity continuedphlebotominemonitoring is proposed in order to evaluate theimplication for social welfare and health in the future
Competing Interests
The authors declare that they have no competing interests
Journal of Tropical Medicine 11
Acknowledgments
Theauthorswish to thank the ldquoInstituto de Pesquisas Cientıfi-cas e Tecnologicas do Estado do Amapa IEPArdquo entomologyteam for the technical assistance given to the field work inparticular to Aderbal Amanajas Santana Rosalvo Dias deCastro Noel Fernandes Santos Neto Osita Cleyriane Lobatodo Nascimento Jose Luiz dos Santos Chaves and Joao doSocorro Gomes Campos Logistical facilities were financiallysupported by the ldquoJari Energiardquo
References
[1] D G Young and M A Duncan ldquoGuide to the identificationand geographic distribution of Lutzomyia sand flies in Mexicothe West Indies Central and South America (Diptera Psycho-didae)rdquo inMemoirs of the American Entomological Institute pp1ndash881 Associated Publishers American Entomological InstituteGainesville Fla USA 1994
[2] P D Ready ldquoBiology of phlebotomine sand flies as vectors ofdisease agentsrdquo Annual Review of Entomology vol 58 pp 227ndash250 2013
[3] R P Brazil A A Fuzzari and J D Andrade Filho ldquoSandfly vectors of Leishmania in the Americasmdasha mini reviewrdquoEntomology Ornithology amp Herpetology vol 4 no 2 Article ID1000144 2015
[4] E F Rangel and R Lainson ldquoProven and putative vectorsof American cutaneous leishmaniasis in Brazil aspects oftheir biology and vectorial competencerdquoMemorias do InstitutoOswaldo Cruz vol 104 no 7 pp 937ndash954 2009
[5] E F Rangel S M Costa and B M Carvalho ldquoEnvironmentalchanges and the geographic spreading of American cutaneousleishmaniasis in Brazilrdquo in Leishmaniasis-Trends in Epidemiol-ogy D Claborn Ed Diagnosis and Treatment InTech RijekaCroatia 2014
[6] W P Taddei ldquoO genero Mansonia (Diptera Culicidae) e aproliferacao de mosquitos na usina hidreletrica de Tucuruırdquo inEnergia na Amazonia S B Magalhaes R C Brito and E RCastro Eds vol 1 pp 311ndash318 MPEGFPAUNAMAZ BelemBrazil 1996 (Portuguese)
[7] A I A Oliveira Observacoes Sobre a Resolucao CONAMA no23797 que Dispoe Sobre o Licenciamento Ambiental MimeoConselho Empresarial Brasileiro Para o Desenvolvimento Sus-tentavel Rio de Janeiro Brazil 1997 (Portuguese)
[8] A K R Galardo C D Galardo G A Silveira et al ldquoPhle-botominae sand flies (Diptera Psychodidae) potential vectorsof American cutaneous leishmaniasis agents in the area asso-ciated with the Santo Antonio hydroelectric system in WesternAmazonian Brazilrdquo Revista da Sociedade Brasileira deMedicinaTropical vol 48 no 3 pp 265ndash271 2015
[9] R Lainson R DWard and J J Shaw ldquoCutaneous leishmaniasisin North Brazil Lutzomyia anduzei as a major vectorrdquo Transac-tions of the Royal Society of Tropical Medicine and Hygiene vol70 no 2 pp 171ndash172 1976
[10] R Lainson J J Shaw R D Ward P D Ready and R DNaiff ldquoLeishmaniasis in Brazil XIII Isolation of Leishmaniafrom armadillos (Dasypus novemcinctus) and observations onthe epidemiology of cutaneous leishmaniasis in north ParaStaterdquo Transactions of the Royal Society of Tropical Medicine andHygiene vol 73 no 2 pp 239ndash242 1979
[11] R Lainson J J Shaw P D Ready M A Miles and M PovoaldquoLeishmaniasis in Brazil XVI Isolation and identification of
Leishmania species from sandflies wild mammals and manin north Para State with particular reference to L braziliensisguyanensis causative agent of ldquopian-boisrdquordquo Transactions of theRoyal Society of Tropical Medicine and Hygiene vol 75 no 4pp 530ndash536 1981
[12] R Lainson J J Shaw and M Povoa ldquoThe importance of eden-tates (sloths and anteaters) as primary reservoirs of Leishmaniabraziliensis guyanensis causative agent of lsquopian boisrsquo in northBrazilrdquo Transactions of the Royal Society of Tropical Medicineand Hygiene vol 75 no 4 pp 611ndash612 1981
[13] R DWard and H Fraiha ldquoLutzomyia umbratilis a new speciesof sand fly from Brazil (Diptera Psychodidae)rdquo Journal ofMedical Entomology vol 14 no 3 pp 313ndash317 1977
[14] P D Ready R Lainson and J J Shaw ldquoLeishmaniasis in BrazilXX Prevalence of lsquoenzootic rodent leishmaniasisrsquo (Leishmaniamexicana amazonensis) and apparent absence of lsquopian boisrsquo(Le braziliensis guyanensis) in plantations of introduced treespecies and in other non-climax forests in eastern AmazoniardquoTransactions of the Royal Society of Tropical Medicine andHygiene vol 77 no 6 pp 775ndash785 1983
[15] P D Ready R Lainson J J Shaw and R DWard ldquoThe ecologyof Lutzomyia umbratilisWard amp Fraiha (Diptera Psychodidae)the major vector to man of Leishmania braziliensis guyanensisin north-eastern Amazonian Brazilrdquo Bulletin of EntomologicalResearch vol 76 no 1 pp 21ndash40 1986
[16] R Lainson and J J Shaw ldquoLeishmania (Viannia) naiffi spn a parasite of the armadillo Dasypus novemcinctus (L)in Amazonian Brazilrdquo Annales de Parasitologie Humaine etComparee vol 64 no 1 pp 3ndash9 1989
[17] A Roger M Nacher M Hanf et al ldquoClimate and leishmaniasisin French guianardquo The American Journal of Tropical Medicineand Hygiene vol 89 no 3 pp 564ndash569 2013
[18] L Ryan ldquoFlebotomos do estado do Para Brasil (Diptera Psy-chodidae)rdquo Thecnical Document 1 Instituto Evandro ChagasBelem Brazil 1986 (Portuguese)
[19] E A B Galati ldquoMorfologia e Taxonomia Morfologia termi-nologia de adultos e identificacao dos taxons da Americardquoin Flebotomıneos do Brasil E F Rangel and R LainsonEds pp 53ndash75 Editora Fiocruz Rio de Janeiro Brazil 2003(Portuguese)
[20] D R Roberts and B P Hsi ldquoAn index of species abundance foruse with mosquito surveillance datardquo Environmental Entomol-ogy vol 8 no 6 pp 1007ndash1013 1979
[21] Oslash Hammer D A T Harper and P D Ryan ldquoPast paleontolog-ical statistics software package for education and data analysisrdquoPalaeontologia Electronica vol 4 no 1 pp 1ndash9 2001
[22] L Ryan R Lainson J J Shaw R R Braga and E A IshikawaldquoLeishmaniasis in Brazil XXV Sandfly vectors of Leishmania inPara State Brazilrdquo Medical and Veterinary Entomology vol 1no 4 pp 383ndash395 1987
[23] R Lainson J J Shaw F T Silveira R R Braga and E AIshikawa ldquoCutaneous leishmaniasis of man due to Leishmania(Viannia) naiffirdquo Annales de Parasitologie Humaine et Compa-ree vol 65 no 5-6 pp 282ndash284 1990
[24] E G C Bermudez Lutzomyia Sand Flies in the BrazilianAmazon Basin (Diptera Psychodidae) INPA Manaus Brazil2009
[25] G M Aguiar and W M Medeiros ldquoDistribuicao regionale habitats das especies de flebotomıneos do Brasilrdquo in Fle-botomıneos do Brasil E F Rangel and R Lainson Eds pp 207ndash256 Fiocruz Rio de Janeiro Brazil 2003 (Portuguese)
12 Journal of Tropical Medicine
[26] J R Arias M A Miles R D Naiff et al ldquoFlagellate infectionsof Brazilian sand flies (Diptera Psychodidae) isolation invitro and biochemical identification of Endotrypanum andLeishmaniardquo The American Journal of Tropical Medicine andHygiene vol 34 no 6 pp 1098ndash1108 1985
[27] A A Souza T V Santos Y L Jennings et al ldquoNaturalLeishmania(Viannia) spp infections in phlebotomine sand flies(Diptera Psychodidae) from the Brazilian Amazon regionreveal new putative transmission cycles of American cutaneousleishmaniasisrdquo Parasite vol 23 article 22 2016
[28] F T Silveira A A Souza R Lainson J J Shaw R R Bragaand E A Ishikawa ldquoCutaneous leishmaniasis in the AmazonRegion natural infection of the sand fly Lutzomyia ubiquitalis(Psychodidae Phlebotominae) by Leishmania (Viannia) lain-soni in Para state BrazilrdquoMemorias do Instituto Oswaldo Cruzvol 86 no 1 pp 127ndash130 1991
[29] LH S Gil S A Basano A A Souza et al ldquoRecent observationson the sand fly (Diptera Psychodidae) fauna of the state ofRondonia Western Amazonia Brazil the importance of Psych-dopygus davisi as a vector of zoonotic cutaneous leishmaniasisrdquoMemorias do Instituto Oswaldo Cruz vol 98 no 6 pp 751ndash7552003
[30] A A A Souza F T Silveira R Lainson et al ldquoThe Phlebotom-inae fauna of Serra dos Carajas Para Brazil and its possibleimplication in the transmission of American tegumentaryleishmaniasisrdquo Revista Pan-Amazonica de Saude vol 1 no 1pp 45ndash51 2010
[31] R Killick-Kendrick ldquoPhlebotomine vectors of the leishmani-ases a reviewrdquoMedical and Veterinary Entomology vol 4 no 1pp 1ndash24 1990
[32] M E C Dorval G Cristaldo H C da Rocha et al ldquoPhle-botomine fauna (Diptera Psychodidae) of an American cuta-neous leishmaniasis endemic area in the state of Mato Grossodo Sul Brazilrdquo Memorias do Instituto Oswaldo Cruz vol 104no 5 pp 695ndash702 2009
[33] V C F V Guimaraes P L Costa F J da Silva et al ldquoMoleculardetection of Leishmania in phlebotomine sand flies in a cuta-neous and visceral leishmaniasis endemic area in northeasternBrazilrdquo Revista do Instituto de Medicina Tropical de Sao Paulovol 56 no 4 pp 357ndash360 2014
[34] R S Lana E M Michalsky C L Fortes-Dias et al ldquoPhle-botomine sand fly fauna and Leishmania infection in thevicinity of the Serra do Cipo National Park a natural Brazilianheritage siterdquo BioMed Research International vol 2015 ArticleID 385493 9 pages 2015
[35] V R Alves R A de Freitas F L Santos A F J de OliveiraT V Barrett and P H F Shimabukuro ldquoSand flies (DipteraPsychodidae Phlebotominae) from central amazonia and fournew records for the Amazonas State Brazilrdquo Revista Brasileirade Entomologia vol 56 no 2 pp 220ndash227 2012
[36] J V S Ferreira T V Santos E M Santos and I S GorayebldquoPhlebotomine sand flies (Diptera Psychodidae) in forestfragments of Belem metropolitan area Para State Brazil withconsiderations on vectors of American cutaneous leishmaniasisagentsrdquo Revista Pan-Amazonica de Saude vol 5 no 2 pp 29ndash35 2014
[37] B M Carvalho E F Rangel P D Ready and M M ValeldquoEcological niche modelling predicts southward expansion ofLutzomyia (Nyssomyia) flaviscutellata (Diptera PsychodidaePhlebotominae) vector of Leishmania (Leishmania) amazonen-sis in South America under climate changerdquo PLoS ONE vol 10no 11 Article ID e0143282 2015
[38] F Fouque P Gaborit J Issaly et al ldquoPhlebotomine sand flies(Diptera Psychodidae) associatedwith changing patterns in thetransmission of the human cutaneous leishmaniasis in FrenchGuianardquoMemorias do Instituto Oswaldo Cruz vol 102 no 1 pp35ndash40 2007
[39] R Killick-Kendrick ldquoSome epidemiological consequences ofthe evolutionary fit between Leishmaniae and their phle-botomine vectorsrdquo Bulletin de la Societe de Pathologie Exotiqueet de ses Filiales vol 78 no 5 pp 747ndash755 1985
[40] F X Pajot F Le Pont BGentile andR Besnard ldquoEpidemiologyof leishmaniasis in French Guianardquo Transactions of the RoyalSociety of Tropical Medicine and Hygiene vol 76 no 1 pp 112ndash113 1982
[41] H A Christensen J R Arias A M De Vasquez and R ADe Freitas ldquoHosts of sandfly vectors of Leishmania braziliensisguyanensis in the central Amazon of Brazilrdquo The AmericanJournal of Tropical Medicine and Hygiene vol 31 no 2 pp 239ndash242 1982
[42] L C R Nery E S Lorosa andAM R Franco ldquoFeeding prefer-ence of the sand flies Lutzomyia umbratilis and L spathotrichia(Diptera Psychodidae Phlebotominae) in an urban forest patchin the City ofManaus Amazonas BrazilrdquoMemorias do InstitutoOswaldo Cruz vol 99 no 6 pp 571ndash574 2004
[43] PD Ready R Lainson J J Shaw andAA Souza ldquoDNAprobesfor distinguishing Psychodopygus wellcomei from Psychodopy-gus complexus (Diptera Psychodidae)rdquo Memorias do InstitutoOswaldo Cruz vol 86 no 1 pp 41ndash49 1991
[44] P D Ready H Fraiha R P Lane J R Arias and F X PajotldquoOn distinguishing the female of Psychodopygus wellcomei avector of mucocutaneous leishmaniasis from other squamiven-tris series females I Characterization of Ps squamiventrissquamiventris and Ps s maripaensis stat nov (Diptera Psycho-didae)rdquo Annals of Tropical Medicine and Parasitology vol 76no 2 pp 201ndash214 1982
[45] A D Kent T V dos Santos A Gangadin A Samjhawan DR A Mans and H D F H Schallig ldquoStudies on the sandfly fauna (Diptera Psychodidae) in high-transmission areas ofcutaneous leishmaniasis in the Republic of Surinamerdquo Parasitesand Vectors vol 6 no 1 article 318 2013
[46] P D Ready R Lainson and J J Shaw ldquoHabitat and seasonalityof Psychodopygus wellcomei help incriminate it as a vector ofLeishmania braziliensis in Amazonia and Northeast BrazilrdquoTransactions of the Royal Society of Tropical Medicine andHygiene vol 78 no 4 pp 543ndash544 1984
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
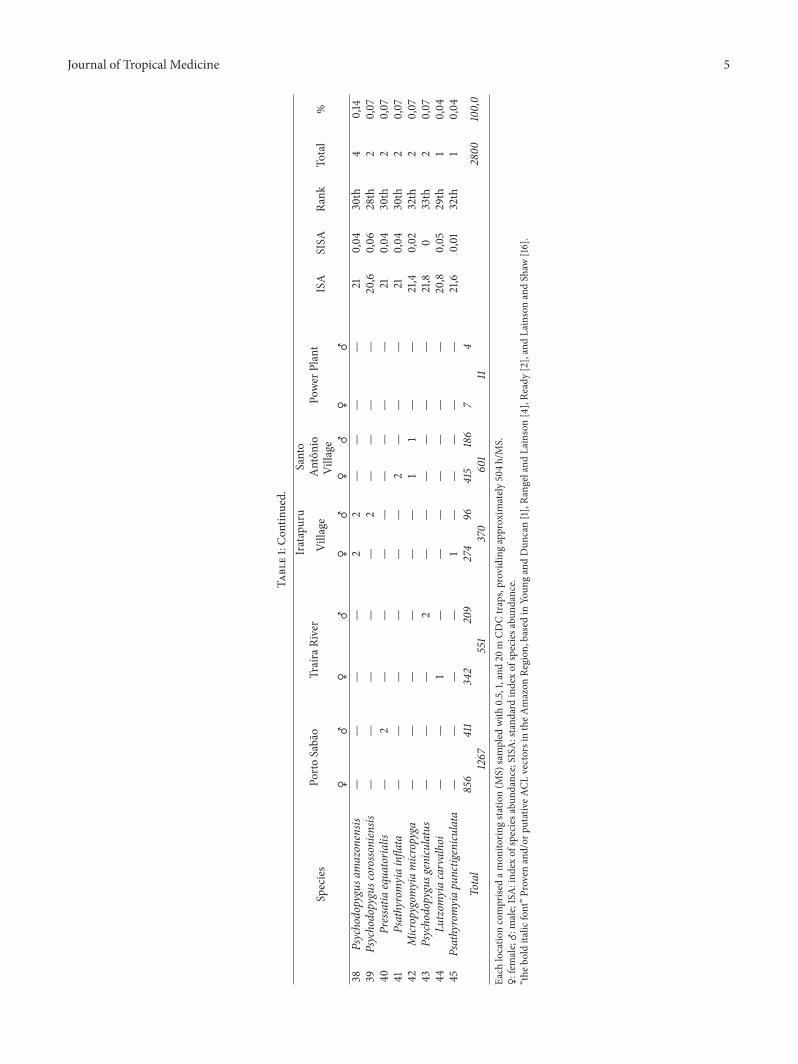
Journal of Tropical Medicine 5
Table1Con
tinued
Species
PortoSabao
Traıra
River
Iratapuru
Santo
Power
Plant
ISA
SISA
Rank
Total
Village
Anton
ioVillage
CD
CD
CD
CD
CD
38Psychodopygusa
mazonensis
mdashmdash
mdashmdash
22
mdashmdash
mdashmdash
21004
30th
4014
39Psychodopyguscorossoniensis
mdashmdash
mdashmdash
mdash2
mdashmdash
mdashmdash
206
006
28th
2007
40Pressatia
equatoria
lismdash
2mdash
mdashmdash
mdashmdash
mdashmdash
mdash21
004
30th
2007
41Psathyromyiainflata
mdashmdash
mdashmdash
mdashmdash
2mdash
mdashmdash
21004
30th
2007
42Micropygom
yiamicropyga
mdashmdash
mdashmdash
mdashmdash
11
mdashmdash
214
002
32th
2007
43Psychodopygusgenicu
latus
mdashmdash
mdash2
mdashmdash
mdashmdash
mdashmdash
218
033th
2007
44Lu
tzom
yiacarvalhoi
mdashmdash
1mdash
mdashmdash
mdashmdash
mdashmdash
208
005
29th
1004
45Psathyromyiapunctigenicu
lata
mdashmdash
mdashmdash
1mdash
mdashmdash
mdashmdash
216
001
32th
1004
Total
856
411
342
209
274
96415
186
74
2800
1000
1267
551
370
601
11Ea
chlocatio
ncomprise
dam
onito
ringstation(M
S)sampled
with
051and20
mCD
Ctrapsprovidingapproxim
ately
504h
MS
Cfem
aleDm
aleISAind
exof
speciesa
bund
anceSISAstand
ardindexof
speciesa
bund
ance
ldquothe
bold
italic
fontrdquoP
rovenandor
putativ
eACL
vectorsintheA
mazon
Region
based
inYo
ungandDun
can[1]Ra
ngelandLainson[4]Re
ady[2]andLainsonandShaw
[16]
6 Journal of Tropical Medicine
Table 2 Vertical stratification of Phebotomine species composition of Santo Antonio do Jari hydroelectric system impacted area (AmapaState Amazonian Brazil) on the basis of five surveyed locations (2012ndash2015)
Species 05m ground 1m ground 20m canopy TotalC D C D C D
1 Sciopemyia sordellii 117 71 91 23 11 3 3162 Psychodopygus davisi 48 27 44 39 102 54 314mdash Trichophoromyia spp 117 mdash 127 mdash 24 mdash 2683 Bichromomyia flaviscutellata 85 45 79 41 11 4 2654 Evandromyia infraspinosa 38 18 48 27 16 3 1505 Nyssomyia umbratilis 35 23 26 13 28 21 1466 Trichophoromyia castanheirai mdash 50 mdash 59 mdash 26 135mdash Pressatia spp 66 mdash 39 mdash 26 mdash 131mdash Evandromyia spp 55 12 41 9 11 mdash 1287 Vianamyia furcata 42 8 46 6 8 1 1118 Psychodopygus squamiventris sl 27 11 20 1 23 7 899 Trichopygomyia trichopyga 18 20 10 5 22 11 8610 Trichophoromyia ubiquitalis 28 15 16 13 9 5 86mdash Other species (11ndash45) 137 112 129 69 71 57 575
Total 813 412 716 305 362 192 2800Total (C + D) 1225 1021 554 mdash
Each stratum was sampled with approximately 840 h of CDC trapC female D male
infraspinosa (119899 = 150 SISA = 06) and Ny umbratilis (119899 =146 SISA = 063) Less frequently captured but nonethelessepidemiologically relevant species included Ps squamiventrissl (119899 = 89 SISA = 052) Th ubiquitalis (119899 = 86 SISA=05)Ps hirsutus hirsutus (119899 = 55 SISA = 04)Ny anduzei (119899 = 45SISA = 023) Lutzomyia gomezi (119899 = 38 SISA = 012) Nyantunesi (119899 = 19 SISA = 037) Ps paraensis (119899 = 15 SISA =02) and Ny whitmani (119899 = 8 SISA = 009)
Sampling effort for each area comprised approximately504 h of CDC trapping (14 expeditions 3 CDC traps and12-hour CDC exposition) Porto Sabao was the site of themost captured specimens (1267) followed by Santo AntonioVillage (601) The highest diversity was observed in capturesfrom Iratapuru Village (1198671015840 = 3027) All 1198671015840 value com-parisons between the different environments were significant(119901 le 005) except when comparing Traıra River and thePower Plant (119901 le 0099) (Figure 2)
Regarding vertical distribution sampling of the samecaptures described above accounted for approximately 840 hof CDC trapping by stratum (14 expeditions 5 ecotopes and12-hour CDC exposition) CDC traps placed 05m aboveground level accounted for a greater number of individuals(1225) than those placed 1m above ground level (1021) and20m above ground level in the canopy (554) (Table 2) WhilePs davisi was most likely to be found at canopy level (75individuals at 05m 83 at 1m and 156 at 20m) and Biflaviscutellata closer to ground level (130 at 05m 120 at 1mand 15 at 20m) there was little difference in numbers of Nyumbratilis individuals captured at the different levels of strata(58 at 05m 39 at 1m and 49 at 20m)
The monthly average rainfall was the only climatic datawith a representative coefficient of variance (1198781015840 = 7081)whentested against ACL occurrence and phlebotomine frequency
005
115
225
335
PortoSabatildeo
TraiacuteraRiver
IratapuruVillage
Santo AntocircnioVillage
PowerPlant
p = 0009p = 0099
p = 0007
p = 0012p = 0001
p = 0031
p = 0001p = 0004
p = 0001p = 00012855
26543027
2819
1468
Figure 2 Comparison of Shannon diversity indexes for CDC-captured phlebotomines from the five surveyed locations in SantoAntonio do Jari hydroelectric system impacted area between thestates of Para and Amapa Brazil (2012ndash2015)
(1198781015840 = 6595) in contrast to temperature (1198781015840 = 596)and relative air humidity (1198781015840 = 956) The available dataenabled correlations to be determined between the periodsof May 2012 to April 2015 a period within which nine of thefourteen capture efforts were undertaken Rainfall presenteda moderate correlation with overall phlebotomine frequency(119903 = 065 119901 = 000001) as also individually compared withPs squamiventris sl (119903 = 065 119901 = 000001) and a similarlymoderate but lower correlation with Bi flaviscutellata (119903 =048 119901 = 00021) and Ny umbratilis (119903 = 048 119901 = 0002)
Journal of Tropical Medicine 7
A decrease in the overall phlebotomine population wasobserved after March until November 2014 coinciding withthe transition between the rainy and dry periods and withflooding (Figure 3)
The number of ACL cases presented a moderate cor-relation with overall phlebotomine frequency (119903 = 044119901 = 00063) and also with Ps squamiventris (119903 = 032119901 = 00003) and an insignificant negative correlation with Biflaviscutellata (119903 = minus002 119901 = 00001) A similarly moderatebut higher correlation was however found withNy umbratilis(119903 = 048 119901 = 0002) (Figure 4)
4 Discussion
It is important to consider current investigations of phle-botomine fauna in the Jari River Basin in the context ofthe historical findings from the 1970s and 1980s Theseearlier studies represent comprehensive research conductedin Monte Dourado (northern PA) a region close to thearea investigated in the present study and on which thepresent epidemiological background to ACL transmissionin the associated region is based In brief the previousresearch includes a description of Ny umbratilis (formerlyldquoobscuredrdquo as Ny anduzei) [13] and its incrimination as themain vector of L (V) guyanensis [9ndash11] confirmed by strongecological associations [15] Just as important a parasitemore recently described as L (V) naiffi was isolated fromthe armadilloDasypus novemcinctus for which transmissionwas locally driven and attributed to Ps squamiventris sl andother Psychodopygus species [22] this comprised a primarilyenzootic ACL occasionally infecting humans [23] Finallythe isolation of L (L) amazonensis from rodents in an areaaffected by occurrence of the main vector Bi flaviscutellata[11] comprised the third known Jari ACL transmission cycleThese facts described above suggest that Ny umbratilis Pssquamiventris sl and Bi flaviscutellata are implicated in thetransmission scenario of the area covered by the present studyand thus merit particular attention
The present study incorporated a representative compo-sition of the known AP fauna Considering the substantialsampling effort (2520 h) the determined diversity showedagreement with the findings of other inventories from theBrazilian Amazon basin [1 24] Moreover all new records forAP have already been recorded for PA [25]
Apart from Ny umbratilis Ps squamiventris sl and Biflaviscutellata other species captured were noted as beingepidemiologically relevant to ACL in the Brazilian AmazonRegion Ps paraensis Ps davisi and Ps hirsutus hirsutuswhich are putative vectors of L (V) naiffi [26 27] Thubiquitalis a proven vector of L (V) lainsoni [28] Nyanduzei a secondary vector of L (V) guyanensis [4] Nywhitmani and Lu gomezi proven and putative vectors of L(V) shawi respectively [4 26] and lastly Ny antunesi asuspected vector of L (V) lindenbergi [4]
Among the above species Ps davisi is notable as thethird highest ranking according to SISA (074) and as havingthe highest presence in all vertical strata in particular atthe canopy level This species is generally anthropophilic iswidely distributed in the Amazon Basin [1 24] and is related
to Leishmania (Viannia) infections in the States of Rondonia(RO) [29] and PA [27 30] Interestingly this species remainsfrequent in recently surveyed areas affected by hydroelectricschemes in RO [8] and PA (TVS unpublished) Theseobservations add weight to the need for further investigationinto the putative epidemiological role of Ps davisi in Jari
The species most frequently captured Sc sordellii wouldappear not to be of epidemiological concern Firstly theapparent abundance of a given phlebotomine species is notsufficient of itself to incriminate it as a vector [31] particularlywhen it does not demonstrate high levels of anthropophilicbehavior Secondly flagellate infections isolated from thisspecies (formerly referred to as Lutzomyia nordestina) arehistorically recognized as distinct from Leishmania [22] orremain undetermined [32] and are more likely to be othertrypanosomatids [22] Furthermore despite recent reports ofmolecular Leishmania ascribed to this fly [33 34] currentlypolymerase chain reaction-based techniques do not allowldquotrue infectionrdquo to be distinguished from a ldquosimple and occa-sionally ingestedrdquo DNA fragment [3] Thus there remainsinsufficient evidence for a potential vector role of S sordelliiin Leishmania transmission
High frequencies of light-trapped Bi flaviscutellata havebeen documented [8 35] despite rodent-baited trappingbeing better recognized as amethod for attracting the speciesAdditionally the first SISA Rank (095) was indicative ofthis high presence in all five surveyed ecotopes this hasbeen observed in the Belem metropolitan area (PA) whereBi flaviscutellata was the only omnipresent species in sevenurban and ecologically isolated forest fragments surveyed[36] Furthermore this species has been reported as beingable to adapt to plantations of introduced tree species and inother nonclimax forests in Jari [14] suggesting it is capableof responding positively to environmental change such asthe vegetal suppression and flooding caused by hydroelectricoperations Nonetheless ecological niche model projectionspredict no change in spatial distribution for Bi flaviscutellatain the Jari area which is an environment that remains suitablefor the species [37]
In an area in Regina French Guiana ecologically dis-turbed due to agricultural practices flagellates infecting a Biflaviscutellata specimen have been determined to be L (V)guyanensis [38] in similar circumstances to the observed lowpresence of Ny umbratilis Although this possibly changingpattern in ACL transmission should be interpreted withcaution it raises an intriguing possibility about the permis-siveness of Bi flaviscutellata to L (V) guyanensis in theAmazonGuianan ACL scenario
In the present study a higher number of Bi flaviscutellataindividuals was captured compared to Ny umbratilis (265versus 146 resp) A lack of available evidence for naturalinfection in this unnatural vectorparasite system howeverprecludes speculation about the possible involvement of Biflaviscutellata even in the context of occasional transmissionof parasites distinct from L (L) amazonensis Neverthelessthe current high occurrence of Bi flaviscutellata in theAmazonGuianan ACL scenario may be of future concernunnatural parasitevectorreservoir systems when closelyrelated in an ecological context can on a century-level scale
8 Journal of Tropical Medicine
010203040506070
Bi flaviscutellata
01020304050607080
Ny umbratilis
0510152025303540
Ps squamiventris sl
0100200300400500600
050
100150200250300350400
2012 2013 2014 2015
Wat
erflo
odin
g
Ps s
quya
miv
entri
s sl
Bi fl
avisc
utell
ata
Ny u
mbr
atili
s
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
050
100150200250300350400
050
100150200250300350400
050
100150200250300350400
010203040506070
10000 20000 30000 40000000Rainfall (mm)
01020304050607080
10000 20000 30000 40000000Rainfall (mm)
Rainfall (mm)Overall
Rainfall (mm)
Rainfall (mm)
Rainfall (mm)
0100200300400500600
Ove
rall
10000 20000 30000 40000000Rainfall (mm)
05
10152025303540
10000 20000 30000 40000000Rainfall (mm)
r = 065 p = 000001
r = 048 p = 00021
r = 049 p = 00021
r = 065 p = 000001
Figure 3 Correlation between rainfall and phlebotomine putative vector species captured in Santo Antonio do Jari hydroelectric systemimpacted area between the states of Para and Amapa Brazil (2012ndash2015)
Journal of Tropical Medicine 9
2012 2013 2014 2015
Wat
erflo
odin
g
0100200300400500600
010203040506070
Bi flaviscutellata
01020304050607080
Ny umbratilis
0510152025303540
Ps squamiventris sl
Bi fl
avisc
utell
ata
05
1015202530
05
1015202530
05
1015202530
05
1015202530
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
0100200300400500600
Ove
rall
10 20 300ACL cases
010203040506070
10 20 300ACL cases
01020304050607080
Ny u
mbr
atili
s
10 20 300ACL cases
ACL casesOverall
ACL cases
ACL cases
ACL cases
05
10152025303540
Ps s
quam
iven
tris s
l
10 20 300ACL cases
r = 044 p = 00063
r = minus002 p = 00001
r = 048 p = 0002
r = 032 p = 00003
Figure 4 Correlation between ACL cases and phlebotomine putative vector species captured in Santo Antonio do Jari hydroelectric systemimpacted area between the states of Para and Amapa Brazil (2012ndash2015)
10 Journal of Tropical Medicine
undergo stress adaptation (evolutionary fitness) and formnew ecological systems driven by coevolution [39]
There is no doubt that Ny umbratilis is the main vectorof ACL in the Jari River Basin [15] Surprisingly the well-documented tree-dwelling behavior of this species [15 40]was not observed in CDC canopy captures in the presentstudy very little difference was found between the number ofindividuals captured in the three sampled strata (58 at 05m39 at 1m and 49 at 20m) Although Ny umbratilis is knownto climb into the canopy at night to feed on edentates (mainlysloths) [12 41] it is possible that an alternative terrestrialsource of blood such as rodents may be attracting femalestoward ground level Sloths are very slow moving and aretherefore commonly rescued for relocation during environ-mentalmonitoring operations their resulting rarenessmeansNy umbratilis has had to find other food sources Converselyblood meal analysis conducted during studies in a forestfragment of the Western Amazon has shown rodent bloodto be the predominant feeding source for Ny umbratilis [42]Wild and synanthropic rodents have been shown to surviveenvironmental disturbance in Jari and to proliferate in areasof introduced tree species [14] this indicated a plausiblereason not only for the putative adaptation of Ny umbratilisbut also for the increasing population levels of the rodentblood-feeder Bi flaviscutellata as already discussed
Given the clear importance of Psychodopygus of theChagasi series (formerly referred to as the Squamiventrisseries) in the context of ACL transmission [22] taxonomicstudies on identifying the cryptic females have been under-taken previously [43] and more recently with interestingresults (Rodrigo E Godoy unpublished) These techniqueshowever could not be employed in the field at the time Earlyconsideration of the assumed geographic distribution of thecomplex Ps squamiventris sl [1 44] shows that individualsfrom AP are likely to be predominantly Ps s maripaensisDetermining males of the species present could clarify thesetaxonomic uncertainties in the area covered by the presentstudy however doubts about identification of some malespecimens meant they were grouped as Ps squamiventris sl
Ryan et al [22] did not find Ps squamiventris sl to benaturally infected in the Jari area but proposed a link to L(V) naiffi along the same northern bank of the AmazonRiver through studies in Cachoeira Porteira (PA) Fouqueet al [38] came to the same conclusions in French GuianaInterestingly this fly species complex was found alongside L(V) braziliensis DNA in Suriname [45] an area where theknown Amazonian vectors of this parasite Ps complexus andPs wellcomei are unlikely to occur As previously discussedin the context of Bi flaviscutellata the changing transmissionpatterns of unnatural vectorparasite systems must be care-fully interpreted and only after strong ecological associationshave been determined Thus the putative vector importanceof Ps squamiventris sl in Jari can only be assumed in respectof L (V) naiffi
The five surveyed areas presented in general differentdiversity indexes showing variations in the phlebotominecomposition along the affected area This finding can beattributed not only to the distance between capture sitesbut also to forest composition fragmentation geographic
barriers and migration of mammal fauna between sites thislatter can occur naturally or be induced by rescue opera-tions involved in hydroelectric planning whereby animalsare moved to suitable ldquodry landrdquo environments safe frompotential flooding
A decrease in the phlebotomine population in 2014 inthe period after filling the reservoir could not be attributedexclusively either to flooding or to the transition from therainy to the dry season Both eventualities although oppositein effect are likely to lead to unsuitable conditions forphlebotomine reproduction However they did coincide inMay 2014 when rainfall decreased and the lake was formed
Conversely rainfall was a valid determinant of overallphlebotomine fluctuation Ps squamiventris sl was the mainputative vector directly correlated with this Other variablessuch as forest composition and altitudinal variation areknown to influence seasonal dynamics of species from thecomplex Ps squamiventris sl For example the dry periodfrom October to November in Serra dos Carajas (southernPA) can be unsuitable for Ps wellcomei which is adapted tohigh altitudes in contrast to Ps complexus which flourishesin lowland areas during the same months [46]
An insignificant negative to a moderate positive correla-tion was observed between cases of ACL and phlebotominefluctuations In the case of Ny umbratilis however theapparently higher correlation when compared with otherspecies is in agreement with its known vector importance inAmazonGuianan ACL A strong spatiotemporal associationbetween a given vector and ACL cases is considered essentialto maintenance of the Leishmania life cycle [2 14]
In summary the present study filled a temporal gapon phlebotomine data relating to the Jari area It providesa meaningful update on species composition and furtherinsights into seasonal dynamics of species under pressurescaused by environmental change The presence of ACLvectors in particular Ny umbratilis Bi flaviscutellata andPs squamiventris sl is noteworthy due to their directassociation with AmazonGuianan-ACL transmission ForNy umbratilis the lower frequency associatedwith the unpre-dicted lack of tree-dwelling behavior may be attributableto a reduction in numbers of canopy-dwelling mammalsdue to the environmental disturbance and relocation effortsthis could consequently have led to seeking for alternativeterrestrial blood sources at ground level The possible asso-ciation of Ps davisi with Leishmania transmission cycles inother Amazon ecoregions draws attention to its high rankposition in Jari Rainfall can be partly associated with localseasonal fluctuations in fly numbers mainly with respect toPs squamiventris sl Cases of ACL were also more highlycorrelated with Ny umbratilis dynamics Further attentionwas also given to results for Bi flaviscutellata which hasbeen apparently tolerant of anthropic activity continuedphlebotominemonitoring is proposed in order to evaluate theimplication for social welfare and health in the future
Competing Interests
The authors declare that they have no competing interests
Journal of Tropical Medicine 11
Acknowledgments
Theauthorswish to thank the ldquoInstituto de Pesquisas Cientıfi-cas e Tecnologicas do Estado do Amapa IEPArdquo entomologyteam for the technical assistance given to the field work inparticular to Aderbal Amanajas Santana Rosalvo Dias deCastro Noel Fernandes Santos Neto Osita Cleyriane Lobatodo Nascimento Jose Luiz dos Santos Chaves and Joao doSocorro Gomes Campos Logistical facilities were financiallysupported by the ldquoJari Energiardquo
References
[1] D G Young and M A Duncan ldquoGuide to the identificationand geographic distribution of Lutzomyia sand flies in Mexicothe West Indies Central and South America (Diptera Psycho-didae)rdquo inMemoirs of the American Entomological Institute pp1ndash881 Associated Publishers American Entomological InstituteGainesville Fla USA 1994
[2] P D Ready ldquoBiology of phlebotomine sand flies as vectors ofdisease agentsrdquo Annual Review of Entomology vol 58 pp 227ndash250 2013
[3] R P Brazil A A Fuzzari and J D Andrade Filho ldquoSandfly vectors of Leishmania in the Americasmdasha mini reviewrdquoEntomology Ornithology amp Herpetology vol 4 no 2 Article ID1000144 2015
[4] E F Rangel and R Lainson ldquoProven and putative vectorsof American cutaneous leishmaniasis in Brazil aspects oftheir biology and vectorial competencerdquoMemorias do InstitutoOswaldo Cruz vol 104 no 7 pp 937ndash954 2009
[5] E F Rangel S M Costa and B M Carvalho ldquoEnvironmentalchanges and the geographic spreading of American cutaneousleishmaniasis in Brazilrdquo in Leishmaniasis-Trends in Epidemiol-ogy D Claborn Ed Diagnosis and Treatment InTech RijekaCroatia 2014
[6] W P Taddei ldquoO genero Mansonia (Diptera Culicidae) e aproliferacao de mosquitos na usina hidreletrica de Tucuruırdquo inEnergia na Amazonia S B Magalhaes R C Brito and E RCastro Eds vol 1 pp 311ndash318 MPEGFPAUNAMAZ BelemBrazil 1996 (Portuguese)
[7] A I A Oliveira Observacoes Sobre a Resolucao CONAMA no23797 que Dispoe Sobre o Licenciamento Ambiental MimeoConselho Empresarial Brasileiro Para o Desenvolvimento Sus-tentavel Rio de Janeiro Brazil 1997 (Portuguese)
[8] A K R Galardo C D Galardo G A Silveira et al ldquoPhle-botominae sand flies (Diptera Psychodidae) potential vectorsof American cutaneous leishmaniasis agents in the area asso-ciated with the Santo Antonio hydroelectric system in WesternAmazonian Brazilrdquo Revista da Sociedade Brasileira deMedicinaTropical vol 48 no 3 pp 265ndash271 2015
[9] R Lainson R DWard and J J Shaw ldquoCutaneous leishmaniasisin North Brazil Lutzomyia anduzei as a major vectorrdquo Transac-tions of the Royal Society of Tropical Medicine and Hygiene vol70 no 2 pp 171ndash172 1976
[10] R Lainson J J Shaw R D Ward P D Ready and R DNaiff ldquoLeishmaniasis in Brazil XIII Isolation of Leishmaniafrom armadillos (Dasypus novemcinctus) and observations onthe epidemiology of cutaneous leishmaniasis in north ParaStaterdquo Transactions of the Royal Society of Tropical Medicine andHygiene vol 73 no 2 pp 239ndash242 1979
[11] R Lainson J J Shaw P D Ready M A Miles and M PovoaldquoLeishmaniasis in Brazil XVI Isolation and identification of
Leishmania species from sandflies wild mammals and manin north Para State with particular reference to L braziliensisguyanensis causative agent of ldquopian-boisrdquordquo Transactions of theRoyal Society of Tropical Medicine and Hygiene vol 75 no 4pp 530ndash536 1981
[12] R Lainson J J Shaw and M Povoa ldquoThe importance of eden-tates (sloths and anteaters) as primary reservoirs of Leishmaniabraziliensis guyanensis causative agent of lsquopian boisrsquo in northBrazilrdquo Transactions of the Royal Society of Tropical Medicineand Hygiene vol 75 no 4 pp 611ndash612 1981
[13] R DWard and H Fraiha ldquoLutzomyia umbratilis a new speciesof sand fly from Brazil (Diptera Psychodidae)rdquo Journal ofMedical Entomology vol 14 no 3 pp 313ndash317 1977
[14] P D Ready R Lainson and J J Shaw ldquoLeishmaniasis in BrazilXX Prevalence of lsquoenzootic rodent leishmaniasisrsquo (Leishmaniamexicana amazonensis) and apparent absence of lsquopian boisrsquo(Le braziliensis guyanensis) in plantations of introduced treespecies and in other non-climax forests in eastern AmazoniardquoTransactions of the Royal Society of Tropical Medicine andHygiene vol 77 no 6 pp 775ndash785 1983
[15] P D Ready R Lainson J J Shaw and R DWard ldquoThe ecologyof Lutzomyia umbratilisWard amp Fraiha (Diptera Psychodidae)the major vector to man of Leishmania braziliensis guyanensisin north-eastern Amazonian Brazilrdquo Bulletin of EntomologicalResearch vol 76 no 1 pp 21ndash40 1986
[16] R Lainson and J J Shaw ldquoLeishmania (Viannia) naiffi spn a parasite of the armadillo Dasypus novemcinctus (L)in Amazonian Brazilrdquo Annales de Parasitologie Humaine etComparee vol 64 no 1 pp 3ndash9 1989
[17] A Roger M Nacher M Hanf et al ldquoClimate and leishmaniasisin French guianardquo The American Journal of Tropical Medicineand Hygiene vol 89 no 3 pp 564ndash569 2013
[18] L Ryan ldquoFlebotomos do estado do Para Brasil (Diptera Psy-chodidae)rdquo Thecnical Document 1 Instituto Evandro ChagasBelem Brazil 1986 (Portuguese)
[19] E A B Galati ldquoMorfologia e Taxonomia Morfologia termi-nologia de adultos e identificacao dos taxons da Americardquoin Flebotomıneos do Brasil E F Rangel and R LainsonEds pp 53ndash75 Editora Fiocruz Rio de Janeiro Brazil 2003(Portuguese)
[20] D R Roberts and B P Hsi ldquoAn index of species abundance foruse with mosquito surveillance datardquo Environmental Entomol-ogy vol 8 no 6 pp 1007ndash1013 1979
[21] Oslash Hammer D A T Harper and P D Ryan ldquoPast paleontolog-ical statistics software package for education and data analysisrdquoPalaeontologia Electronica vol 4 no 1 pp 1ndash9 2001
[22] L Ryan R Lainson J J Shaw R R Braga and E A IshikawaldquoLeishmaniasis in Brazil XXV Sandfly vectors of Leishmania inPara State Brazilrdquo Medical and Veterinary Entomology vol 1no 4 pp 383ndash395 1987
[23] R Lainson J J Shaw F T Silveira R R Braga and E AIshikawa ldquoCutaneous leishmaniasis of man due to Leishmania(Viannia) naiffirdquo Annales de Parasitologie Humaine et Compa-ree vol 65 no 5-6 pp 282ndash284 1990
[24] E G C Bermudez Lutzomyia Sand Flies in the BrazilianAmazon Basin (Diptera Psychodidae) INPA Manaus Brazil2009
[25] G M Aguiar and W M Medeiros ldquoDistribuicao regionale habitats das especies de flebotomıneos do Brasilrdquo in Fle-botomıneos do Brasil E F Rangel and R Lainson Eds pp 207ndash256 Fiocruz Rio de Janeiro Brazil 2003 (Portuguese)
12 Journal of Tropical Medicine
[26] J R Arias M A Miles R D Naiff et al ldquoFlagellate infectionsof Brazilian sand flies (Diptera Psychodidae) isolation invitro and biochemical identification of Endotrypanum andLeishmaniardquo The American Journal of Tropical Medicine andHygiene vol 34 no 6 pp 1098ndash1108 1985
[27] A A Souza T V Santos Y L Jennings et al ldquoNaturalLeishmania(Viannia) spp infections in phlebotomine sand flies(Diptera Psychodidae) from the Brazilian Amazon regionreveal new putative transmission cycles of American cutaneousleishmaniasisrdquo Parasite vol 23 article 22 2016
[28] F T Silveira A A Souza R Lainson J J Shaw R R Bragaand E A Ishikawa ldquoCutaneous leishmaniasis in the AmazonRegion natural infection of the sand fly Lutzomyia ubiquitalis(Psychodidae Phlebotominae) by Leishmania (Viannia) lain-soni in Para state BrazilrdquoMemorias do Instituto Oswaldo Cruzvol 86 no 1 pp 127ndash130 1991
[29] LH S Gil S A Basano A A Souza et al ldquoRecent observationson the sand fly (Diptera Psychodidae) fauna of the state ofRondonia Western Amazonia Brazil the importance of Psych-dopygus davisi as a vector of zoonotic cutaneous leishmaniasisrdquoMemorias do Instituto Oswaldo Cruz vol 98 no 6 pp 751ndash7552003
[30] A A A Souza F T Silveira R Lainson et al ldquoThe Phlebotom-inae fauna of Serra dos Carajas Para Brazil and its possibleimplication in the transmission of American tegumentaryleishmaniasisrdquo Revista Pan-Amazonica de Saude vol 1 no 1pp 45ndash51 2010
[31] R Killick-Kendrick ldquoPhlebotomine vectors of the leishmani-ases a reviewrdquoMedical and Veterinary Entomology vol 4 no 1pp 1ndash24 1990
[32] M E C Dorval G Cristaldo H C da Rocha et al ldquoPhle-botomine fauna (Diptera Psychodidae) of an American cuta-neous leishmaniasis endemic area in the state of Mato Grossodo Sul Brazilrdquo Memorias do Instituto Oswaldo Cruz vol 104no 5 pp 695ndash702 2009
[33] V C F V Guimaraes P L Costa F J da Silva et al ldquoMoleculardetection of Leishmania in phlebotomine sand flies in a cuta-neous and visceral leishmaniasis endemic area in northeasternBrazilrdquo Revista do Instituto de Medicina Tropical de Sao Paulovol 56 no 4 pp 357ndash360 2014
[34] R S Lana E M Michalsky C L Fortes-Dias et al ldquoPhle-botomine sand fly fauna and Leishmania infection in thevicinity of the Serra do Cipo National Park a natural Brazilianheritage siterdquo BioMed Research International vol 2015 ArticleID 385493 9 pages 2015
[35] V R Alves R A de Freitas F L Santos A F J de OliveiraT V Barrett and P H F Shimabukuro ldquoSand flies (DipteraPsychodidae Phlebotominae) from central amazonia and fournew records for the Amazonas State Brazilrdquo Revista Brasileirade Entomologia vol 56 no 2 pp 220ndash227 2012
[36] J V S Ferreira T V Santos E M Santos and I S GorayebldquoPhlebotomine sand flies (Diptera Psychodidae) in forestfragments of Belem metropolitan area Para State Brazil withconsiderations on vectors of American cutaneous leishmaniasisagentsrdquo Revista Pan-Amazonica de Saude vol 5 no 2 pp 29ndash35 2014
[37] B M Carvalho E F Rangel P D Ready and M M ValeldquoEcological niche modelling predicts southward expansion ofLutzomyia (Nyssomyia) flaviscutellata (Diptera PsychodidaePhlebotominae) vector of Leishmania (Leishmania) amazonen-sis in South America under climate changerdquo PLoS ONE vol 10no 11 Article ID e0143282 2015
[38] F Fouque P Gaborit J Issaly et al ldquoPhlebotomine sand flies(Diptera Psychodidae) associatedwith changing patterns in thetransmission of the human cutaneous leishmaniasis in FrenchGuianardquoMemorias do Instituto Oswaldo Cruz vol 102 no 1 pp35ndash40 2007
[39] R Killick-Kendrick ldquoSome epidemiological consequences ofthe evolutionary fit between Leishmaniae and their phle-botomine vectorsrdquo Bulletin de la Societe de Pathologie Exotiqueet de ses Filiales vol 78 no 5 pp 747ndash755 1985
[40] F X Pajot F Le Pont BGentile andR Besnard ldquoEpidemiologyof leishmaniasis in French Guianardquo Transactions of the RoyalSociety of Tropical Medicine and Hygiene vol 76 no 1 pp 112ndash113 1982
[41] H A Christensen J R Arias A M De Vasquez and R ADe Freitas ldquoHosts of sandfly vectors of Leishmania braziliensisguyanensis in the central Amazon of Brazilrdquo The AmericanJournal of Tropical Medicine and Hygiene vol 31 no 2 pp 239ndash242 1982
[42] L C R Nery E S Lorosa andAM R Franco ldquoFeeding prefer-ence of the sand flies Lutzomyia umbratilis and L spathotrichia(Diptera Psychodidae Phlebotominae) in an urban forest patchin the City ofManaus Amazonas BrazilrdquoMemorias do InstitutoOswaldo Cruz vol 99 no 6 pp 571ndash574 2004
[43] PD Ready R Lainson J J Shaw andAA Souza ldquoDNAprobesfor distinguishing Psychodopygus wellcomei from Psychodopy-gus complexus (Diptera Psychodidae)rdquo Memorias do InstitutoOswaldo Cruz vol 86 no 1 pp 41ndash49 1991
[44] P D Ready H Fraiha R P Lane J R Arias and F X PajotldquoOn distinguishing the female of Psychodopygus wellcomei avector of mucocutaneous leishmaniasis from other squamiven-tris series females I Characterization of Ps squamiventrissquamiventris and Ps s maripaensis stat nov (Diptera Psycho-didae)rdquo Annals of Tropical Medicine and Parasitology vol 76no 2 pp 201ndash214 1982
[45] A D Kent T V dos Santos A Gangadin A Samjhawan DR A Mans and H D F H Schallig ldquoStudies on the sandfly fauna (Diptera Psychodidae) in high-transmission areas ofcutaneous leishmaniasis in the Republic of Surinamerdquo Parasitesand Vectors vol 6 no 1 article 318 2013
[46] P D Ready R Lainson and J J Shaw ldquoHabitat and seasonalityof Psychodopygus wellcomei help incriminate it as a vector ofLeishmania braziliensis in Amazonia and Northeast BrazilrdquoTransactions of the Royal Society of Tropical Medicine andHygiene vol 78 no 4 pp 543ndash544 1984
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

6 Journal of Tropical Medicine
Table 2 Vertical stratification of Phebotomine species composition of Santo Antonio do Jari hydroelectric system impacted area (AmapaState Amazonian Brazil) on the basis of five surveyed locations (2012ndash2015)
Species 05m ground 1m ground 20m canopy TotalC D C D C D
1 Sciopemyia sordellii 117 71 91 23 11 3 3162 Psychodopygus davisi 48 27 44 39 102 54 314mdash Trichophoromyia spp 117 mdash 127 mdash 24 mdash 2683 Bichromomyia flaviscutellata 85 45 79 41 11 4 2654 Evandromyia infraspinosa 38 18 48 27 16 3 1505 Nyssomyia umbratilis 35 23 26 13 28 21 1466 Trichophoromyia castanheirai mdash 50 mdash 59 mdash 26 135mdash Pressatia spp 66 mdash 39 mdash 26 mdash 131mdash Evandromyia spp 55 12 41 9 11 mdash 1287 Vianamyia furcata 42 8 46 6 8 1 1118 Psychodopygus squamiventris sl 27 11 20 1 23 7 899 Trichopygomyia trichopyga 18 20 10 5 22 11 8610 Trichophoromyia ubiquitalis 28 15 16 13 9 5 86mdash Other species (11ndash45) 137 112 129 69 71 57 575
Total 813 412 716 305 362 192 2800Total (C + D) 1225 1021 554 mdash
Each stratum was sampled with approximately 840 h of CDC trapC female D male
infraspinosa (119899 = 150 SISA = 06) and Ny umbratilis (119899 =146 SISA = 063) Less frequently captured but nonethelessepidemiologically relevant species included Ps squamiventrissl (119899 = 89 SISA = 052) Th ubiquitalis (119899 = 86 SISA=05)Ps hirsutus hirsutus (119899 = 55 SISA = 04)Ny anduzei (119899 = 45SISA = 023) Lutzomyia gomezi (119899 = 38 SISA = 012) Nyantunesi (119899 = 19 SISA = 037) Ps paraensis (119899 = 15 SISA =02) and Ny whitmani (119899 = 8 SISA = 009)
Sampling effort for each area comprised approximately504 h of CDC trapping (14 expeditions 3 CDC traps and12-hour CDC exposition) Porto Sabao was the site of themost captured specimens (1267) followed by Santo AntonioVillage (601) The highest diversity was observed in capturesfrom Iratapuru Village (1198671015840 = 3027) All 1198671015840 value com-parisons between the different environments were significant(119901 le 005) except when comparing Traıra River and thePower Plant (119901 le 0099) (Figure 2)
Regarding vertical distribution sampling of the samecaptures described above accounted for approximately 840 hof CDC trapping by stratum (14 expeditions 5 ecotopes and12-hour CDC exposition) CDC traps placed 05m aboveground level accounted for a greater number of individuals(1225) than those placed 1m above ground level (1021) and20m above ground level in the canopy (554) (Table 2) WhilePs davisi was most likely to be found at canopy level (75individuals at 05m 83 at 1m and 156 at 20m) and Biflaviscutellata closer to ground level (130 at 05m 120 at 1mand 15 at 20m) there was little difference in numbers of Nyumbratilis individuals captured at the different levels of strata(58 at 05m 39 at 1m and 49 at 20m)
The monthly average rainfall was the only climatic datawith a representative coefficient of variance (1198781015840 = 7081)whentested against ACL occurrence and phlebotomine frequency
005
115
225
335
PortoSabatildeo
TraiacuteraRiver
IratapuruVillage
Santo AntocircnioVillage
PowerPlant
p = 0009p = 0099
p = 0007
p = 0012p = 0001
p = 0031
p = 0001p = 0004
p = 0001p = 00012855
26543027
2819
1468
Figure 2 Comparison of Shannon diversity indexes for CDC-captured phlebotomines from the five surveyed locations in SantoAntonio do Jari hydroelectric system impacted area between thestates of Para and Amapa Brazil (2012ndash2015)
(1198781015840 = 6595) in contrast to temperature (1198781015840 = 596)and relative air humidity (1198781015840 = 956) The available dataenabled correlations to be determined between the periodsof May 2012 to April 2015 a period within which nine of thefourteen capture efforts were undertaken Rainfall presenteda moderate correlation with overall phlebotomine frequency(119903 = 065 119901 = 000001) as also individually compared withPs squamiventris sl (119903 = 065 119901 = 000001) and a similarlymoderate but lower correlation with Bi flaviscutellata (119903 =048 119901 = 00021) and Ny umbratilis (119903 = 048 119901 = 0002)
Journal of Tropical Medicine 7
A decrease in the overall phlebotomine population wasobserved after March until November 2014 coinciding withthe transition between the rainy and dry periods and withflooding (Figure 3)
The number of ACL cases presented a moderate cor-relation with overall phlebotomine frequency (119903 = 044119901 = 00063) and also with Ps squamiventris (119903 = 032119901 = 00003) and an insignificant negative correlation with Biflaviscutellata (119903 = minus002 119901 = 00001) A similarly moderatebut higher correlation was however found withNy umbratilis(119903 = 048 119901 = 0002) (Figure 4)
4 Discussion
It is important to consider current investigations of phle-botomine fauna in the Jari River Basin in the context ofthe historical findings from the 1970s and 1980s Theseearlier studies represent comprehensive research conductedin Monte Dourado (northern PA) a region close to thearea investigated in the present study and on which thepresent epidemiological background to ACL transmissionin the associated region is based In brief the previousresearch includes a description of Ny umbratilis (formerlyldquoobscuredrdquo as Ny anduzei) [13] and its incrimination as themain vector of L (V) guyanensis [9ndash11] confirmed by strongecological associations [15] Just as important a parasitemore recently described as L (V) naiffi was isolated fromthe armadilloDasypus novemcinctus for which transmissionwas locally driven and attributed to Ps squamiventris sl andother Psychodopygus species [22] this comprised a primarilyenzootic ACL occasionally infecting humans [23] Finallythe isolation of L (L) amazonensis from rodents in an areaaffected by occurrence of the main vector Bi flaviscutellata[11] comprised the third known Jari ACL transmission cycleThese facts described above suggest that Ny umbratilis Pssquamiventris sl and Bi flaviscutellata are implicated in thetransmission scenario of the area covered by the present studyand thus merit particular attention
The present study incorporated a representative compo-sition of the known AP fauna Considering the substantialsampling effort (2520 h) the determined diversity showedagreement with the findings of other inventories from theBrazilian Amazon basin [1 24] Moreover all new records forAP have already been recorded for PA [25]
Apart from Ny umbratilis Ps squamiventris sl and Biflaviscutellata other species captured were noted as beingepidemiologically relevant to ACL in the Brazilian AmazonRegion Ps paraensis Ps davisi and Ps hirsutus hirsutuswhich are putative vectors of L (V) naiffi [26 27] Thubiquitalis a proven vector of L (V) lainsoni [28] Nyanduzei a secondary vector of L (V) guyanensis [4] Nywhitmani and Lu gomezi proven and putative vectors of L(V) shawi respectively [4 26] and lastly Ny antunesi asuspected vector of L (V) lindenbergi [4]
Among the above species Ps davisi is notable as thethird highest ranking according to SISA (074) and as havingthe highest presence in all vertical strata in particular atthe canopy level This species is generally anthropophilic iswidely distributed in the Amazon Basin [1 24] and is related
to Leishmania (Viannia) infections in the States of Rondonia(RO) [29] and PA [27 30] Interestingly this species remainsfrequent in recently surveyed areas affected by hydroelectricschemes in RO [8] and PA (TVS unpublished) Theseobservations add weight to the need for further investigationinto the putative epidemiological role of Ps davisi in Jari
The species most frequently captured Sc sordellii wouldappear not to be of epidemiological concern Firstly theapparent abundance of a given phlebotomine species is notsufficient of itself to incriminate it as a vector [31] particularlywhen it does not demonstrate high levels of anthropophilicbehavior Secondly flagellate infections isolated from thisspecies (formerly referred to as Lutzomyia nordestina) arehistorically recognized as distinct from Leishmania [22] orremain undetermined [32] and are more likely to be othertrypanosomatids [22] Furthermore despite recent reports ofmolecular Leishmania ascribed to this fly [33 34] currentlypolymerase chain reaction-based techniques do not allowldquotrue infectionrdquo to be distinguished from a ldquosimple and occa-sionally ingestedrdquo DNA fragment [3] Thus there remainsinsufficient evidence for a potential vector role of S sordelliiin Leishmania transmission
High frequencies of light-trapped Bi flaviscutellata havebeen documented [8 35] despite rodent-baited trappingbeing better recognized as amethod for attracting the speciesAdditionally the first SISA Rank (095) was indicative ofthis high presence in all five surveyed ecotopes this hasbeen observed in the Belem metropolitan area (PA) whereBi flaviscutellata was the only omnipresent species in sevenurban and ecologically isolated forest fragments surveyed[36] Furthermore this species has been reported as beingable to adapt to plantations of introduced tree species and inother nonclimax forests in Jari [14] suggesting it is capableof responding positively to environmental change such asthe vegetal suppression and flooding caused by hydroelectricoperations Nonetheless ecological niche model projectionspredict no change in spatial distribution for Bi flaviscutellatain the Jari area which is an environment that remains suitablefor the species [37]
In an area in Regina French Guiana ecologically dis-turbed due to agricultural practices flagellates infecting a Biflaviscutellata specimen have been determined to be L (V)guyanensis [38] in similar circumstances to the observed lowpresence of Ny umbratilis Although this possibly changingpattern in ACL transmission should be interpreted withcaution it raises an intriguing possibility about the permis-siveness of Bi flaviscutellata to L (V) guyanensis in theAmazonGuianan ACL scenario
In the present study a higher number of Bi flaviscutellataindividuals was captured compared to Ny umbratilis (265versus 146 resp) A lack of available evidence for naturalinfection in this unnatural vectorparasite system howeverprecludes speculation about the possible involvement of Biflaviscutellata even in the context of occasional transmissionof parasites distinct from L (L) amazonensis Neverthelessthe current high occurrence of Bi flaviscutellata in theAmazonGuianan ACL scenario may be of future concernunnatural parasitevectorreservoir systems when closelyrelated in an ecological context can on a century-level scale
8 Journal of Tropical Medicine
010203040506070
Bi flaviscutellata
01020304050607080
Ny umbratilis
0510152025303540
Ps squamiventris sl
0100200300400500600
050
100150200250300350400
2012 2013 2014 2015
Wat
erflo
odin
g
Ps s
quya
miv
entri
s sl
Bi fl
avisc
utell
ata
Ny u
mbr
atili
s
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
050
100150200250300350400
050
100150200250300350400
050
100150200250300350400
010203040506070
10000 20000 30000 40000000Rainfall (mm)
01020304050607080
10000 20000 30000 40000000Rainfall (mm)
Rainfall (mm)Overall
Rainfall (mm)
Rainfall (mm)
Rainfall (mm)
0100200300400500600
Ove
rall
10000 20000 30000 40000000Rainfall (mm)
05
10152025303540
10000 20000 30000 40000000Rainfall (mm)
r = 065 p = 000001
r = 048 p = 00021
r = 049 p = 00021
r = 065 p = 000001
Figure 3 Correlation between rainfall and phlebotomine putative vector species captured in Santo Antonio do Jari hydroelectric systemimpacted area between the states of Para and Amapa Brazil (2012ndash2015)
Journal of Tropical Medicine 9
2012 2013 2014 2015
Wat
erflo
odin
g
0100200300400500600
010203040506070
Bi flaviscutellata
01020304050607080
Ny umbratilis
0510152025303540
Ps squamiventris sl
Bi fl
avisc
utell
ata
05
1015202530
05
1015202530
05
1015202530
05
1015202530
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
0100200300400500600
Ove
rall
10 20 300ACL cases
010203040506070
10 20 300ACL cases
01020304050607080
Ny u
mbr
atili
s
10 20 300ACL cases
ACL casesOverall
ACL cases
ACL cases
ACL cases
05
10152025303540
Ps s
quam
iven
tris s
l
10 20 300ACL cases
r = 044 p = 00063
r = minus002 p = 00001
r = 048 p = 0002
r = 032 p = 00003
Figure 4 Correlation between ACL cases and phlebotomine putative vector species captured in Santo Antonio do Jari hydroelectric systemimpacted area between the states of Para and Amapa Brazil (2012ndash2015)
10 Journal of Tropical Medicine
undergo stress adaptation (evolutionary fitness) and formnew ecological systems driven by coevolution [39]
There is no doubt that Ny umbratilis is the main vectorof ACL in the Jari River Basin [15] Surprisingly the well-documented tree-dwelling behavior of this species [15 40]was not observed in CDC canopy captures in the presentstudy very little difference was found between the number ofindividuals captured in the three sampled strata (58 at 05m39 at 1m and 49 at 20m) Although Ny umbratilis is knownto climb into the canopy at night to feed on edentates (mainlysloths) [12 41] it is possible that an alternative terrestrialsource of blood such as rodents may be attracting femalestoward ground level Sloths are very slow moving and aretherefore commonly rescued for relocation during environ-mentalmonitoring operations their resulting rarenessmeansNy umbratilis has had to find other food sources Converselyblood meal analysis conducted during studies in a forestfragment of the Western Amazon has shown rodent bloodto be the predominant feeding source for Ny umbratilis [42]Wild and synanthropic rodents have been shown to surviveenvironmental disturbance in Jari and to proliferate in areasof introduced tree species [14] this indicated a plausiblereason not only for the putative adaptation of Ny umbratilisbut also for the increasing population levels of the rodentblood-feeder Bi flaviscutellata as already discussed
Given the clear importance of Psychodopygus of theChagasi series (formerly referred to as the Squamiventrisseries) in the context of ACL transmission [22] taxonomicstudies on identifying the cryptic females have been under-taken previously [43] and more recently with interestingresults (Rodrigo E Godoy unpublished) These techniqueshowever could not be employed in the field at the time Earlyconsideration of the assumed geographic distribution of thecomplex Ps squamiventris sl [1 44] shows that individualsfrom AP are likely to be predominantly Ps s maripaensisDetermining males of the species present could clarify thesetaxonomic uncertainties in the area covered by the presentstudy however doubts about identification of some malespecimens meant they were grouped as Ps squamiventris sl
Ryan et al [22] did not find Ps squamiventris sl to benaturally infected in the Jari area but proposed a link to L(V) naiffi along the same northern bank of the AmazonRiver through studies in Cachoeira Porteira (PA) Fouqueet al [38] came to the same conclusions in French GuianaInterestingly this fly species complex was found alongside L(V) braziliensis DNA in Suriname [45] an area where theknown Amazonian vectors of this parasite Ps complexus andPs wellcomei are unlikely to occur As previously discussedin the context of Bi flaviscutellata the changing transmissionpatterns of unnatural vectorparasite systems must be care-fully interpreted and only after strong ecological associationshave been determined Thus the putative vector importanceof Ps squamiventris sl in Jari can only be assumed in respectof L (V) naiffi
The five surveyed areas presented in general differentdiversity indexes showing variations in the phlebotominecomposition along the affected area This finding can beattributed not only to the distance between capture sitesbut also to forest composition fragmentation geographic
barriers and migration of mammal fauna between sites thislatter can occur naturally or be induced by rescue opera-tions involved in hydroelectric planning whereby animalsare moved to suitable ldquodry landrdquo environments safe frompotential flooding
A decrease in the phlebotomine population in 2014 inthe period after filling the reservoir could not be attributedexclusively either to flooding or to the transition from therainy to the dry season Both eventualities although oppositein effect are likely to lead to unsuitable conditions forphlebotomine reproduction However they did coincide inMay 2014 when rainfall decreased and the lake was formed
Conversely rainfall was a valid determinant of overallphlebotomine fluctuation Ps squamiventris sl was the mainputative vector directly correlated with this Other variablessuch as forest composition and altitudinal variation areknown to influence seasonal dynamics of species from thecomplex Ps squamiventris sl For example the dry periodfrom October to November in Serra dos Carajas (southernPA) can be unsuitable for Ps wellcomei which is adapted tohigh altitudes in contrast to Ps complexus which flourishesin lowland areas during the same months [46]
An insignificant negative to a moderate positive correla-tion was observed between cases of ACL and phlebotominefluctuations In the case of Ny umbratilis however theapparently higher correlation when compared with otherspecies is in agreement with its known vector importance inAmazonGuianan ACL A strong spatiotemporal associationbetween a given vector and ACL cases is considered essentialto maintenance of the Leishmania life cycle [2 14]
In summary the present study filled a temporal gapon phlebotomine data relating to the Jari area It providesa meaningful update on species composition and furtherinsights into seasonal dynamics of species under pressurescaused by environmental change The presence of ACLvectors in particular Ny umbratilis Bi flaviscutellata andPs squamiventris sl is noteworthy due to their directassociation with AmazonGuianan-ACL transmission ForNy umbratilis the lower frequency associatedwith the unpre-dicted lack of tree-dwelling behavior may be attributableto a reduction in numbers of canopy-dwelling mammalsdue to the environmental disturbance and relocation effortsthis could consequently have led to seeking for alternativeterrestrial blood sources at ground level The possible asso-ciation of Ps davisi with Leishmania transmission cycles inother Amazon ecoregions draws attention to its high rankposition in Jari Rainfall can be partly associated with localseasonal fluctuations in fly numbers mainly with respect toPs squamiventris sl Cases of ACL were also more highlycorrelated with Ny umbratilis dynamics Further attentionwas also given to results for Bi flaviscutellata which hasbeen apparently tolerant of anthropic activity continuedphlebotominemonitoring is proposed in order to evaluate theimplication for social welfare and health in the future
Competing Interests
The authors declare that they have no competing interests
Journal of Tropical Medicine 11
Acknowledgments
Theauthorswish to thank the ldquoInstituto de Pesquisas Cientıfi-cas e Tecnologicas do Estado do Amapa IEPArdquo entomologyteam for the technical assistance given to the field work inparticular to Aderbal Amanajas Santana Rosalvo Dias deCastro Noel Fernandes Santos Neto Osita Cleyriane Lobatodo Nascimento Jose Luiz dos Santos Chaves and Joao doSocorro Gomes Campos Logistical facilities were financiallysupported by the ldquoJari Energiardquo
References
[1] D G Young and M A Duncan ldquoGuide to the identificationand geographic distribution of Lutzomyia sand flies in Mexicothe West Indies Central and South America (Diptera Psycho-didae)rdquo inMemoirs of the American Entomological Institute pp1ndash881 Associated Publishers American Entomological InstituteGainesville Fla USA 1994
[2] P D Ready ldquoBiology of phlebotomine sand flies as vectors ofdisease agentsrdquo Annual Review of Entomology vol 58 pp 227ndash250 2013
[3] R P Brazil A A Fuzzari and J D Andrade Filho ldquoSandfly vectors of Leishmania in the Americasmdasha mini reviewrdquoEntomology Ornithology amp Herpetology vol 4 no 2 Article ID1000144 2015
[4] E F Rangel and R Lainson ldquoProven and putative vectorsof American cutaneous leishmaniasis in Brazil aspects oftheir biology and vectorial competencerdquoMemorias do InstitutoOswaldo Cruz vol 104 no 7 pp 937ndash954 2009
[5] E F Rangel S M Costa and B M Carvalho ldquoEnvironmentalchanges and the geographic spreading of American cutaneousleishmaniasis in Brazilrdquo in Leishmaniasis-Trends in Epidemiol-ogy D Claborn Ed Diagnosis and Treatment InTech RijekaCroatia 2014
[6] W P Taddei ldquoO genero Mansonia (Diptera Culicidae) e aproliferacao de mosquitos na usina hidreletrica de Tucuruırdquo inEnergia na Amazonia S B Magalhaes R C Brito and E RCastro Eds vol 1 pp 311ndash318 MPEGFPAUNAMAZ BelemBrazil 1996 (Portuguese)
[7] A I A Oliveira Observacoes Sobre a Resolucao CONAMA no23797 que Dispoe Sobre o Licenciamento Ambiental MimeoConselho Empresarial Brasileiro Para o Desenvolvimento Sus-tentavel Rio de Janeiro Brazil 1997 (Portuguese)
[8] A K R Galardo C D Galardo G A Silveira et al ldquoPhle-botominae sand flies (Diptera Psychodidae) potential vectorsof American cutaneous leishmaniasis agents in the area asso-ciated with the Santo Antonio hydroelectric system in WesternAmazonian Brazilrdquo Revista da Sociedade Brasileira deMedicinaTropical vol 48 no 3 pp 265ndash271 2015
[9] R Lainson R DWard and J J Shaw ldquoCutaneous leishmaniasisin North Brazil Lutzomyia anduzei as a major vectorrdquo Transac-tions of the Royal Society of Tropical Medicine and Hygiene vol70 no 2 pp 171ndash172 1976
[10] R Lainson J J Shaw R D Ward P D Ready and R DNaiff ldquoLeishmaniasis in Brazil XIII Isolation of Leishmaniafrom armadillos (Dasypus novemcinctus) and observations onthe epidemiology of cutaneous leishmaniasis in north ParaStaterdquo Transactions of the Royal Society of Tropical Medicine andHygiene vol 73 no 2 pp 239ndash242 1979
[11] R Lainson J J Shaw P D Ready M A Miles and M PovoaldquoLeishmaniasis in Brazil XVI Isolation and identification of
Leishmania species from sandflies wild mammals and manin north Para State with particular reference to L braziliensisguyanensis causative agent of ldquopian-boisrdquordquo Transactions of theRoyal Society of Tropical Medicine and Hygiene vol 75 no 4pp 530ndash536 1981
[12] R Lainson J J Shaw and M Povoa ldquoThe importance of eden-tates (sloths and anteaters) as primary reservoirs of Leishmaniabraziliensis guyanensis causative agent of lsquopian boisrsquo in northBrazilrdquo Transactions of the Royal Society of Tropical Medicineand Hygiene vol 75 no 4 pp 611ndash612 1981
[13] R DWard and H Fraiha ldquoLutzomyia umbratilis a new speciesof sand fly from Brazil (Diptera Psychodidae)rdquo Journal ofMedical Entomology vol 14 no 3 pp 313ndash317 1977
[14] P D Ready R Lainson and J J Shaw ldquoLeishmaniasis in BrazilXX Prevalence of lsquoenzootic rodent leishmaniasisrsquo (Leishmaniamexicana amazonensis) and apparent absence of lsquopian boisrsquo(Le braziliensis guyanensis) in plantations of introduced treespecies and in other non-climax forests in eastern AmazoniardquoTransactions of the Royal Society of Tropical Medicine andHygiene vol 77 no 6 pp 775ndash785 1983
[15] P D Ready R Lainson J J Shaw and R DWard ldquoThe ecologyof Lutzomyia umbratilisWard amp Fraiha (Diptera Psychodidae)the major vector to man of Leishmania braziliensis guyanensisin north-eastern Amazonian Brazilrdquo Bulletin of EntomologicalResearch vol 76 no 1 pp 21ndash40 1986
[16] R Lainson and J J Shaw ldquoLeishmania (Viannia) naiffi spn a parasite of the armadillo Dasypus novemcinctus (L)in Amazonian Brazilrdquo Annales de Parasitologie Humaine etComparee vol 64 no 1 pp 3ndash9 1989
[17] A Roger M Nacher M Hanf et al ldquoClimate and leishmaniasisin French guianardquo The American Journal of Tropical Medicineand Hygiene vol 89 no 3 pp 564ndash569 2013
[18] L Ryan ldquoFlebotomos do estado do Para Brasil (Diptera Psy-chodidae)rdquo Thecnical Document 1 Instituto Evandro ChagasBelem Brazil 1986 (Portuguese)
[19] E A B Galati ldquoMorfologia e Taxonomia Morfologia termi-nologia de adultos e identificacao dos taxons da Americardquoin Flebotomıneos do Brasil E F Rangel and R LainsonEds pp 53ndash75 Editora Fiocruz Rio de Janeiro Brazil 2003(Portuguese)
[20] D R Roberts and B P Hsi ldquoAn index of species abundance foruse with mosquito surveillance datardquo Environmental Entomol-ogy vol 8 no 6 pp 1007ndash1013 1979
[21] Oslash Hammer D A T Harper and P D Ryan ldquoPast paleontolog-ical statistics software package for education and data analysisrdquoPalaeontologia Electronica vol 4 no 1 pp 1ndash9 2001
[22] L Ryan R Lainson J J Shaw R R Braga and E A IshikawaldquoLeishmaniasis in Brazil XXV Sandfly vectors of Leishmania inPara State Brazilrdquo Medical and Veterinary Entomology vol 1no 4 pp 383ndash395 1987
[23] R Lainson J J Shaw F T Silveira R R Braga and E AIshikawa ldquoCutaneous leishmaniasis of man due to Leishmania(Viannia) naiffirdquo Annales de Parasitologie Humaine et Compa-ree vol 65 no 5-6 pp 282ndash284 1990
[24] E G C Bermudez Lutzomyia Sand Flies in the BrazilianAmazon Basin (Diptera Psychodidae) INPA Manaus Brazil2009
[25] G M Aguiar and W M Medeiros ldquoDistribuicao regionale habitats das especies de flebotomıneos do Brasilrdquo in Fle-botomıneos do Brasil E F Rangel and R Lainson Eds pp 207ndash256 Fiocruz Rio de Janeiro Brazil 2003 (Portuguese)
12 Journal of Tropical Medicine
[26] J R Arias M A Miles R D Naiff et al ldquoFlagellate infectionsof Brazilian sand flies (Diptera Psychodidae) isolation invitro and biochemical identification of Endotrypanum andLeishmaniardquo The American Journal of Tropical Medicine andHygiene vol 34 no 6 pp 1098ndash1108 1985
[27] A A Souza T V Santos Y L Jennings et al ldquoNaturalLeishmania(Viannia) spp infections in phlebotomine sand flies(Diptera Psychodidae) from the Brazilian Amazon regionreveal new putative transmission cycles of American cutaneousleishmaniasisrdquo Parasite vol 23 article 22 2016
[28] F T Silveira A A Souza R Lainson J J Shaw R R Bragaand E A Ishikawa ldquoCutaneous leishmaniasis in the AmazonRegion natural infection of the sand fly Lutzomyia ubiquitalis(Psychodidae Phlebotominae) by Leishmania (Viannia) lain-soni in Para state BrazilrdquoMemorias do Instituto Oswaldo Cruzvol 86 no 1 pp 127ndash130 1991
[29] LH S Gil S A Basano A A Souza et al ldquoRecent observationson the sand fly (Diptera Psychodidae) fauna of the state ofRondonia Western Amazonia Brazil the importance of Psych-dopygus davisi as a vector of zoonotic cutaneous leishmaniasisrdquoMemorias do Instituto Oswaldo Cruz vol 98 no 6 pp 751ndash7552003
[30] A A A Souza F T Silveira R Lainson et al ldquoThe Phlebotom-inae fauna of Serra dos Carajas Para Brazil and its possibleimplication in the transmission of American tegumentaryleishmaniasisrdquo Revista Pan-Amazonica de Saude vol 1 no 1pp 45ndash51 2010
[31] R Killick-Kendrick ldquoPhlebotomine vectors of the leishmani-ases a reviewrdquoMedical and Veterinary Entomology vol 4 no 1pp 1ndash24 1990
[32] M E C Dorval G Cristaldo H C da Rocha et al ldquoPhle-botomine fauna (Diptera Psychodidae) of an American cuta-neous leishmaniasis endemic area in the state of Mato Grossodo Sul Brazilrdquo Memorias do Instituto Oswaldo Cruz vol 104no 5 pp 695ndash702 2009
[33] V C F V Guimaraes P L Costa F J da Silva et al ldquoMoleculardetection of Leishmania in phlebotomine sand flies in a cuta-neous and visceral leishmaniasis endemic area in northeasternBrazilrdquo Revista do Instituto de Medicina Tropical de Sao Paulovol 56 no 4 pp 357ndash360 2014
[34] R S Lana E M Michalsky C L Fortes-Dias et al ldquoPhle-botomine sand fly fauna and Leishmania infection in thevicinity of the Serra do Cipo National Park a natural Brazilianheritage siterdquo BioMed Research International vol 2015 ArticleID 385493 9 pages 2015
[35] V R Alves R A de Freitas F L Santos A F J de OliveiraT V Barrett and P H F Shimabukuro ldquoSand flies (DipteraPsychodidae Phlebotominae) from central amazonia and fournew records for the Amazonas State Brazilrdquo Revista Brasileirade Entomologia vol 56 no 2 pp 220ndash227 2012
[36] J V S Ferreira T V Santos E M Santos and I S GorayebldquoPhlebotomine sand flies (Diptera Psychodidae) in forestfragments of Belem metropolitan area Para State Brazil withconsiderations on vectors of American cutaneous leishmaniasisagentsrdquo Revista Pan-Amazonica de Saude vol 5 no 2 pp 29ndash35 2014
[37] B M Carvalho E F Rangel P D Ready and M M ValeldquoEcological niche modelling predicts southward expansion ofLutzomyia (Nyssomyia) flaviscutellata (Diptera PsychodidaePhlebotominae) vector of Leishmania (Leishmania) amazonen-sis in South America under climate changerdquo PLoS ONE vol 10no 11 Article ID e0143282 2015
[38] F Fouque P Gaborit J Issaly et al ldquoPhlebotomine sand flies(Diptera Psychodidae) associatedwith changing patterns in thetransmission of the human cutaneous leishmaniasis in FrenchGuianardquoMemorias do Instituto Oswaldo Cruz vol 102 no 1 pp35ndash40 2007
[39] R Killick-Kendrick ldquoSome epidemiological consequences ofthe evolutionary fit between Leishmaniae and their phle-botomine vectorsrdquo Bulletin de la Societe de Pathologie Exotiqueet de ses Filiales vol 78 no 5 pp 747ndash755 1985
[40] F X Pajot F Le Pont BGentile andR Besnard ldquoEpidemiologyof leishmaniasis in French Guianardquo Transactions of the RoyalSociety of Tropical Medicine and Hygiene vol 76 no 1 pp 112ndash113 1982
[41] H A Christensen J R Arias A M De Vasquez and R ADe Freitas ldquoHosts of sandfly vectors of Leishmania braziliensisguyanensis in the central Amazon of Brazilrdquo The AmericanJournal of Tropical Medicine and Hygiene vol 31 no 2 pp 239ndash242 1982
[42] L C R Nery E S Lorosa andAM R Franco ldquoFeeding prefer-ence of the sand flies Lutzomyia umbratilis and L spathotrichia(Diptera Psychodidae Phlebotominae) in an urban forest patchin the City ofManaus Amazonas BrazilrdquoMemorias do InstitutoOswaldo Cruz vol 99 no 6 pp 571ndash574 2004
[43] PD Ready R Lainson J J Shaw andAA Souza ldquoDNAprobesfor distinguishing Psychodopygus wellcomei from Psychodopy-gus complexus (Diptera Psychodidae)rdquo Memorias do InstitutoOswaldo Cruz vol 86 no 1 pp 41ndash49 1991
[44] P D Ready H Fraiha R P Lane J R Arias and F X PajotldquoOn distinguishing the female of Psychodopygus wellcomei avector of mucocutaneous leishmaniasis from other squamiven-tris series females I Characterization of Ps squamiventrissquamiventris and Ps s maripaensis stat nov (Diptera Psycho-didae)rdquo Annals of Tropical Medicine and Parasitology vol 76no 2 pp 201ndash214 1982
[45] A D Kent T V dos Santos A Gangadin A Samjhawan DR A Mans and H D F H Schallig ldquoStudies on the sandfly fauna (Diptera Psychodidae) in high-transmission areas ofcutaneous leishmaniasis in the Republic of Surinamerdquo Parasitesand Vectors vol 6 no 1 article 318 2013
[46] P D Ready R Lainson and J J Shaw ldquoHabitat and seasonalityof Psychodopygus wellcomei help incriminate it as a vector ofLeishmania braziliensis in Amazonia and Northeast BrazilrdquoTransactions of the Royal Society of Tropical Medicine andHygiene vol 78 no 4 pp 543ndash544 1984
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Journal of Tropical Medicine 7
A decrease in the overall phlebotomine population wasobserved after March until November 2014 coinciding withthe transition between the rainy and dry periods and withflooding (Figure 3)
The number of ACL cases presented a moderate cor-relation with overall phlebotomine frequency (119903 = 044119901 = 00063) and also with Ps squamiventris (119903 = 032119901 = 00003) and an insignificant negative correlation with Biflaviscutellata (119903 = minus002 119901 = 00001) A similarly moderatebut higher correlation was however found withNy umbratilis(119903 = 048 119901 = 0002) (Figure 4)
4 Discussion
It is important to consider current investigations of phle-botomine fauna in the Jari River Basin in the context ofthe historical findings from the 1970s and 1980s Theseearlier studies represent comprehensive research conductedin Monte Dourado (northern PA) a region close to thearea investigated in the present study and on which thepresent epidemiological background to ACL transmissionin the associated region is based In brief the previousresearch includes a description of Ny umbratilis (formerlyldquoobscuredrdquo as Ny anduzei) [13] and its incrimination as themain vector of L (V) guyanensis [9ndash11] confirmed by strongecological associations [15] Just as important a parasitemore recently described as L (V) naiffi was isolated fromthe armadilloDasypus novemcinctus for which transmissionwas locally driven and attributed to Ps squamiventris sl andother Psychodopygus species [22] this comprised a primarilyenzootic ACL occasionally infecting humans [23] Finallythe isolation of L (L) amazonensis from rodents in an areaaffected by occurrence of the main vector Bi flaviscutellata[11] comprised the third known Jari ACL transmission cycleThese facts described above suggest that Ny umbratilis Pssquamiventris sl and Bi flaviscutellata are implicated in thetransmission scenario of the area covered by the present studyand thus merit particular attention
The present study incorporated a representative compo-sition of the known AP fauna Considering the substantialsampling effort (2520 h) the determined diversity showedagreement with the findings of other inventories from theBrazilian Amazon basin [1 24] Moreover all new records forAP have already been recorded for PA [25]
Apart from Ny umbratilis Ps squamiventris sl and Biflaviscutellata other species captured were noted as beingepidemiologically relevant to ACL in the Brazilian AmazonRegion Ps paraensis Ps davisi and Ps hirsutus hirsutuswhich are putative vectors of L (V) naiffi [26 27] Thubiquitalis a proven vector of L (V) lainsoni [28] Nyanduzei a secondary vector of L (V) guyanensis [4] Nywhitmani and Lu gomezi proven and putative vectors of L(V) shawi respectively [4 26] and lastly Ny antunesi asuspected vector of L (V) lindenbergi [4]
Among the above species Ps davisi is notable as thethird highest ranking according to SISA (074) and as havingthe highest presence in all vertical strata in particular atthe canopy level This species is generally anthropophilic iswidely distributed in the Amazon Basin [1 24] and is related
to Leishmania (Viannia) infections in the States of Rondonia(RO) [29] and PA [27 30] Interestingly this species remainsfrequent in recently surveyed areas affected by hydroelectricschemes in RO [8] and PA (TVS unpublished) Theseobservations add weight to the need for further investigationinto the putative epidemiological role of Ps davisi in Jari
The species most frequently captured Sc sordellii wouldappear not to be of epidemiological concern Firstly theapparent abundance of a given phlebotomine species is notsufficient of itself to incriminate it as a vector [31] particularlywhen it does not demonstrate high levels of anthropophilicbehavior Secondly flagellate infections isolated from thisspecies (formerly referred to as Lutzomyia nordestina) arehistorically recognized as distinct from Leishmania [22] orremain undetermined [32] and are more likely to be othertrypanosomatids [22] Furthermore despite recent reports ofmolecular Leishmania ascribed to this fly [33 34] currentlypolymerase chain reaction-based techniques do not allowldquotrue infectionrdquo to be distinguished from a ldquosimple and occa-sionally ingestedrdquo DNA fragment [3] Thus there remainsinsufficient evidence for a potential vector role of S sordelliiin Leishmania transmission
High frequencies of light-trapped Bi flaviscutellata havebeen documented [8 35] despite rodent-baited trappingbeing better recognized as amethod for attracting the speciesAdditionally the first SISA Rank (095) was indicative ofthis high presence in all five surveyed ecotopes this hasbeen observed in the Belem metropolitan area (PA) whereBi flaviscutellata was the only omnipresent species in sevenurban and ecologically isolated forest fragments surveyed[36] Furthermore this species has been reported as beingable to adapt to plantations of introduced tree species and inother nonclimax forests in Jari [14] suggesting it is capableof responding positively to environmental change such asthe vegetal suppression and flooding caused by hydroelectricoperations Nonetheless ecological niche model projectionspredict no change in spatial distribution for Bi flaviscutellatain the Jari area which is an environment that remains suitablefor the species [37]
In an area in Regina French Guiana ecologically dis-turbed due to agricultural practices flagellates infecting a Biflaviscutellata specimen have been determined to be L (V)guyanensis [38] in similar circumstances to the observed lowpresence of Ny umbratilis Although this possibly changingpattern in ACL transmission should be interpreted withcaution it raises an intriguing possibility about the permis-siveness of Bi flaviscutellata to L (V) guyanensis in theAmazonGuianan ACL scenario
In the present study a higher number of Bi flaviscutellataindividuals was captured compared to Ny umbratilis (265versus 146 resp) A lack of available evidence for naturalinfection in this unnatural vectorparasite system howeverprecludes speculation about the possible involvement of Biflaviscutellata even in the context of occasional transmissionof parasites distinct from L (L) amazonensis Neverthelessthe current high occurrence of Bi flaviscutellata in theAmazonGuianan ACL scenario may be of future concernunnatural parasitevectorreservoir systems when closelyrelated in an ecological context can on a century-level scale
8 Journal of Tropical Medicine
010203040506070
Bi flaviscutellata
01020304050607080
Ny umbratilis
0510152025303540
Ps squamiventris sl
0100200300400500600
050
100150200250300350400
2012 2013 2014 2015
Wat
erflo
odin
g
Ps s
quya
miv
entri
s sl
Bi fl
avisc
utell
ata
Ny u
mbr
atili
s
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
050
100150200250300350400
050
100150200250300350400
050
100150200250300350400
010203040506070
10000 20000 30000 40000000Rainfall (mm)
01020304050607080
10000 20000 30000 40000000Rainfall (mm)
Rainfall (mm)Overall
Rainfall (mm)
Rainfall (mm)
Rainfall (mm)
0100200300400500600
Ove
rall
10000 20000 30000 40000000Rainfall (mm)
05
10152025303540
10000 20000 30000 40000000Rainfall (mm)
r = 065 p = 000001
r = 048 p = 00021
r = 049 p = 00021
r = 065 p = 000001
Figure 3 Correlation between rainfall and phlebotomine putative vector species captured in Santo Antonio do Jari hydroelectric systemimpacted area between the states of Para and Amapa Brazil (2012ndash2015)
Journal of Tropical Medicine 9
2012 2013 2014 2015
Wat
erflo
odin
g
0100200300400500600
010203040506070
Bi flaviscutellata
01020304050607080
Ny umbratilis
0510152025303540
Ps squamiventris sl
Bi fl
avisc
utell
ata
05
1015202530
05
1015202530
05
1015202530
05
1015202530
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
0100200300400500600
Ove
rall
10 20 300ACL cases
010203040506070
10 20 300ACL cases
01020304050607080
Ny u
mbr
atili
s
10 20 300ACL cases
ACL casesOverall
ACL cases
ACL cases
ACL cases
05
10152025303540
Ps s
quam
iven
tris s
l
10 20 300ACL cases
r = 044 p = 00063
r = minus002 p = 00001
r = 048 p = 0002
r = 032 p = 00003
Figure 4 Correlation between ACL cases and phlebotomine putative vector species captured in Santo Antonio do Jari hydroelectric systemimpacted area between the states of Para and Amapa Brazil (2012ndash2015)
10 Journal of Tropical Medicine
undergo stress adaptation (evolutionary fitness) and formnew ecological systems driven by coevolution [39]
There is no doubt that Ny umbratilis is the main vectorof ACL in the Jari River Basin [15] Surprisingly the well-documented tree-dwelling behavior of this species [15 40]was not observed in CDC canopy captures in the presentstudy very little difference was found between the number ofindividuals captured in the three sampled strata (58 at 05m39 at 1m and 49 at 20m) Although Ny umbratilis is knownto climb into the canopy at night to feed on edentates (mainlysloths) [12 41] it is possible that an alternative terrestrialsource of blood such as rodents may be attracting femalestoward ground level Sloths are very slow moving and aretherefore commonly rescued for relocation during environ-mentalmonitoring operations their resulting rarenessmeansNy umbratilis has had to find other food sources Converselyblood meal analysis conducted during studies in a forestfragment of the Western Amazon has shown rodent bloodto be the predominant feeding source for Ny umbratilis [42]Wild and synanthropic rodents have been shown to surviveenvironmental disturbance in Jari and to proliferate in areasof introduced tree species [14] this indicated a plausiblereason not only for the putative adaptation of Ny umbratilisbut also for the increasing population levels of the rodentblood-feeder Bi flaviscutellata as already discussed
Given the clear importance of Psychodopygus of theChagasi series (formerly referred to as the Squamiventrisseries) in the context of ACL transmission [22] taxonomicstudies on identifying the cryptic females have been under-taken previously [43] and more recently with interestingresults (Rodrigo E Godoy unpublished) These techniqueshowever could not be employed in the field at the time Earlyconsideration of the assumed geographic distribution of thecomplex Ps squamiventris sl [1 44] shows that individualsfrom AP are likely to be predominantly Ps s maripaensisDetermining males of the species present could clarify thesetaxonomic uncertainties in the area covered by the presentstudy however doubts about identification of some malespecimens meant they were grouped as Ps squamiventris sl
Ryan et al [22] did not find Ps squamiventris sl to benaturally infected in the Jari area but proposed a link to L(V) naiffi along the same northern bank of the AmazonRiver through studies in Cachoeira Porteira (PA) Fouqueet al [38] came to the same conclusions in French GuianaInterestingly this fly species complex was found alongside L(V) braziliensis DNA in Suriname [45] an area where theknown Amazonian vectors of this parasite Ps complexus andPs wellcomei are unlikely to occur As previously discussedin the context of Bi flaviscutellata the changing transmissionpatterns of unnatural vectorparasite systems must be care-fully interpreted and only after strong ecological associationshave been determined Thus the putative vector importanceof Ps squamiventris sl in Jari can only be assumed in respectof L (V) naiffi
The five surveyed areas presented in general differentdiversity indexes showing variations in the phlebotominecomposition along the affected area This finding can beattributed not only to the distance between capture sitesbut also to forest composition fragmentation geographic
barriers and migration of mammal fauna between sites thislatter can occur naturally or be induced by rescue opera-tions involved in hydroelectric planning whereby animalsare moved to suitable ldquodry landrdquo environments safe frompotential flooding
A decrease in the phlebotomine population in 2014 inthe period after filling the reservoir could not be attributedexclusively either to flooding or to the transition from therainy to the dry season Both eventualities although oppositein effect are likely to lead to unsuitable conditions forphlebotomine reproduction However they did coincide inMay 2014 when rainfall decreased and the lake was formed
Conversely rainfall was a valid determinant of overallphlebotomine fluctuation Ps squamiventris sl was the mainputative vector directly correlated with this Other variablessuch as forest composition and altitudinal variation areknown to influence seasonal dynamics of species from thecomplex Ps squamiventris sl For example the dry periodfrom October to November in Serra dos Carajas (southernPA) can be unsuitable for Ps wellcomei which is adapted tohigh altitudes in contrast to Ps complexus which flourishesin lowland areas during the same months [46]
An insignificant negative to a moderate positive correla-tion was observed between cases of ACL and phlebotominefluctuations In the case of Ny umbratilis however theapparently higher correlation when compared with otherspecies is in agreement with its known vector importance inAmazonGuianan ACL A strong spatiotemporal associationbetween a given vector and ACL cases is considered essentialto maintenance of the Leishmania life cycle [2 14]
In summary the present study filled a temporal gapon phlebotomine data relating to the Jari area It providesa meaningful update on species composition and furtherinsights into seasonal dynamics of species under pressurescaused by environmental change The presence of ACLvectors in particular Ny umbratilis Bi flaviscutellata andPs squamiventris sl is noteworthy due to their directassociation with AmazonGuianan-ACL transmission ForNy umbratilis the lower frequency associatedwith the unpre-dicted lack of tree-dwelling behavior may be attributableto a reduction in numbers of canopy-dwelling mammalsdue to the environmental disturbance and relocation effortsthis could consequently have led to seeking for alternativeterrestrial blood sources at ground level The possible asso-ciation of Ps davisi with Leishmania transmission cycles inother Amazon ecoregions draws attention to its high rankposition in Jari Rainfall can be partly associated with localseasonal fluctuations in fly numbers mainly with respect toPs squamiventris sl Cases of ACL were also more highlycorrelated with Ny umbratilis dynamics Further attentionwas also given to results for Bi flaviscutellata which hasbeen apparently tolerant of anthropic activity continuedphlebotominemonitoring is proposed in order to evaluate theimplication for social welfare and health in the future
Competing Interests
The authors declare that they have no competing interests
Journal of Tropical Medicine 11
Acknowledgments
Theauthorswish to thank the ldquoInstituto de Pesquisas Cientıfi-cas e Tecnologicas do Estado do Amapa IEPArdquo entomologyteam for the technical assistance given to the field work inparticular to Aderbal Amanajas Santana Rosalvo Dias deCastro Noel Fernandes Santos Neto Osita Cleyriane Lobatodo Nascimento Jose Luiz dos Santos Chaves and Joao doSocorro Gomes Campos Logistical facilities were financiallysupported by the ldquoJari Energiardquo
References
[1] D G Young and M A Duncan ldquoGuide to the identificationand geographic distribution of Lutzomyia sand flies in Mexicothe West Indies Central and South America (Diptera Psycho-didae)rdquo inMemoirs of the American Entomological Institute pp1ndash881 Associated Publishers American Entomological InstituteGainesville Fla USA 1994
[2] P D Ready ldquoBiology of phlebotomine sand flies as vectors ofdisease agentsrdquo Annual Review of Entomology vol 58 pp 227ndash250 2013
[3] R P Brazil A A Fuzzari and J D Andrade Filho ldquoSandfly vectors of Leishmania in the Americasmdasha mini reviewrdquoEntomology Ornithology amp Herpetology vol 4 no 2 Article ID1000144 2015
[4] E F Rangel and R Lainson ldquoProven and putative vectorsof American cutaneous leishmaniasis in Brazil aspects oftheir biology and vectorial competencerdquoMemorias do InstitutoOswaldo Cruz vol 104 no 7 pp 937ndash954 2009
[5] E F Rangel S M Costa and B M Carvalho ldquoEnvironmentalchanges and the geographic spreading of American cutaneousleishmaniasis in Brazilrdquo in Leishmaniasis-Trends in Epidemiol-ogy D Claborn Ed Diagnosis and Treatment InTech RijekaCroatia 2014
[6] W P Taddei ldquoO genero Mansonia (Diptera Culicidae) e aproliferacao de mosquitos na usina hidreletrica de Tucuruırdquo inEnergia na Amazonia S B Magalhaes R C Brito and E RCastro Eds vol 1 pp 311ndash318 MPEGFPAUNAMAZ BelemBrazil 1996 (Portuguese)
[7] A I A Oliveira Observacoes Sobre a Resolucao CONAMA no23797 que Dispoe Sobre o Licenciamento Ambiental MimeoConselho Empresarial Brasileiro Para o Desenvolvimento Sus-tentavel Rio de Janeiro Brazil 1997 (Portuguese)
[8] A K R Galardo C D Galardo G A Silveira et al ldquoPhle-botominae sand flies (Diptera Psychodidae) potential vectorsof American cutaneous leishmaniasis agents in the area asso-ciated with the Santo Antonio hydroelectric system in WesternAmazonian Brazilrdquo Revista da Sociedade Brasileira deMedicinaTropical vol 48 no 3 pp 265ndash271 2015
[9] R Lainson R DWard and J J Shaw ldquoCutaneous leishmaniasisin North Brazil Lutzomyia anduzei as a major vectorrdquo Transac-tions of the Royal Society of Tropical Medicine and Hygiene vol70 no 2 pp 171ndash172 1976
[10] R Lainson J J Shaw R D Ward P D Ready and R DNaiff ldquoLeishmaniasis in Brazil XIII Isolation of Leishmaniafrom armadillos (Dasypus novemcinctus) and observations onthe epidemiology of cutaneous leishmaniasis in north ParaStaterdquo Transactions of the Royal Society of Tropical Medicine andHygiene vol 73 no 2 pp 239ndash242 1979
[11] R Lainson J J Shaw P D Ready M A Miles and M PovoaldquoLeishmaniasis in Brazil XVI Isolation and identification of
Leishmania species from sandflies wild mammals and manin north Para State with particular reference to L braziliensisguyanensis causative agent of ldquopian-boisrdquordquo Transactions of theRoyal Society of Tropical Medicine and Hygiene vol 75 no 4pp 530ndash536 1981
[12] R Lainson J J Shaw and M Povoa ldquoThe importance of eden-tates (sloths and anteaters) as primary reservoirs of Leishmaniabraziliensis guyanensis causative agent of lsquopian boisrsquo in northBrazilrdquo Transactions of the Royal Society of Tropical Medicineand Hygiene vol 75 no 4 pp 611ndash612 1981
[13] R DWard and H Fraiha ldquoLutzomyia umbratilis a new speciesof sand fly from Brazil (Diptera Psychodidae)rdquo Journal ofMedical Entomology vol 14 no 3 pp 313ndash317 1977
[14] P D Ready R Lainson and J J Shaw ldquoLeishmaniasis in BrazilXX Prevalence of lsquoenzootic rodent leishmaniasisrsquo (Leishmaniamexicana amazonensis) and apparent absence of lsquopian boisrsquo(Le braziliensis guyanensis) in plantations of introduced treespecies and in other non-climax forests in eastern AmazoniardquoTransactions of the Royal Society of Tropical Medicine andHygiene vol 77 no 6 pp 775ndash785 1983
[15] P D Ready R Lainson J J Shaw and R DWard ldquoThe ecologyof Lutzomyia umbratilisWard amp Fraiha (Diptera Psychodidae)the major vector to man of Leishmania braziliensis guyanensisin north-eastern Amazonian Brazilrdquo Bulletin of EntomologicalResearch vol 76 no 1 pp 21ndash40 1986
[16] R Lainson and J J Shaw ldquoLeishmania (Viannia) naiffi spn a parasite of the armadillo Dasypus novemcinctus (L)in Amazonian Brazilrdquo Annales de Parasitologie Humaine etComparee vol 64 no 1 pp 3ndash9 1989
[17] A Roger M Nacher M Hanf et al ldquoClimate and leishmaniasisin French guianardquo The American Journal of Tropical Medicineand Hygiene vol 89 no 3 pp 564ndash569 2013
[18] L Ryan ldquoFlebotomos do estado do Para Brasil (Diptera Psy-chodidae)rdquo Thecnical Document 1 Instituto Evandro ChagasBelem Brazil 1986 (Portuguese)
[19] E A B Galati ldquoMorfologia e Taxonomia Morfologia termi-nologia de adultos e identificacao dos taxons da Americardquoin Flebotomıneos do Brasil E F Rangel and R LainsonEds pp 53ndash75 Editora Fiocruz Rio de Janeiro Brazil 2003(Portuguese)
[20] D R Roberts and B P Hsi ldquoAn index of species abundance foruse with mosquito surveillance datardquo Environmental Entomol-ogy vol 8 no 6 pp 1007ndash1013 1979
[21] Oslash Hammer D A T Harper and P D Ryan ldquoPast paleontolog-ical statistics software package for education and data analysisrdquoPalaeontologia Electronica vol 4 no 1 pp 1ndash9 2001
[22] L Ryan R Lainson J J Shaw R R Braga and E A IshikawaldquoLeishmaniasis in Brazil XXV Sandfly vectors of Leishmania inPara State Brazilrdquo Medical and Veterinary Entomology vol 1no 4 pp 383ndash395 1987
[23] R Lainson J J Shaw F T Silveira R R Braga and E AIshikawa ldquoCutaneous leishmaniasis of man due to Leishmania(Viannia) naiffirdquo Annales de Parasitologie Humaine et Compa-ree vol 65 no 5-6 pp 282ndash284 1990
[24] E G C Bermudez Lutzomyia Sand Flies in the BrazilianAmazon Basin (Diptera Psychodidae) INPA Manaus Brazil2009
[25] G M Aguiar and W M Medeiros ldquoDistribuicao regionale habitats das especies de flebotomıneos do Brasilrdquo in Fle-botomıneos do Brasil E F Rangel and R Lainson Eds pp 207ndash256 Fiocruz Rio de Janeiro Brazil 2003 (Portuguese)
12 Journal of Tropical Medicine
[26] J R Arias M A Miles R D Naiff et al ldquoFlagellate infectionsof Brazilian sand flies (Diptera Psychodidae) isolation invitro and biochemical identification of Endotrypanum andLeishmaniardquo The American Journal of Tropical Medicine andHygiene vol 34 no 6 pp 1098ndash1108 1985
[27] A A Souza T V Santos Y L Jennings et al ldquoNaturalLeishmania(Viannia) spp infections in phlebotomine sand flies(Diptera Psychodidae) from the Brazilian Amazon regionreveal new putative transmission cycles of American cutaneousleishmaniasisrdquo Parasite vol 23 article 22 2016
[28] F T Silveira A A Souza R Lainson J J Shaw R R Bragaand E A Ishikawa ldquoCutaneous leishmaniasis in the AmazonRegion natural infection of the sand fly Lutzomyia ubiquitalis(Psychodidae Phlebotominae) by Leishmania (Viannia) lain-soni in Para state BrazilrdquoMemorias do Instituto Oswaldo Cruzvol 86 no 1 pp 127ndash130 1991
[29] LH S Gil S A Basano A A Souza et al ldquoRecent observationson the sand fly (Diptera Psychodidae) fauna of the state ofRondonia Western Amazonia Brazil the importance of Psych-dopygus davisi as a vector of zoonotic cutaneous leishmaniasisrdquoMemorias do Instituto Oswaldo Cruz vol 98 no 6 pp 751ndash7552003
[30] A A A Souza F T Silveira R Lainson et al ldquoThe Phlebotom-inae fauna of Serra dos Carajas Para Brazil and its possibleimplication in the transmission of American tegumentaryleishmaniasisrdquo Revista Pan-Amazonica de Saude vol 1 no 1pp 45ndash51 2010
[31] R Killick-Kendrick ldquoPhlebotomine vectors of the leishmani-ases a reviewrdquoMedical and Veterinary Entomology vol 4 no 1pp 1ndash24 1990
[32] M E C Dorval G Cristaldo H C da Rocha et al ldquoPhle-botomine fauna (Diptera Psychodidae) of an American cuta-neous leishmaniasis endemic area in the state of Mato Grossodo Sul Brazilrdquo Memorias do Instituto Oswaldo Cruz vol 104no 5 pp 695ndash702 2009
[33] V C F V Guimaraes P L Costa F J da Silva et al ldquoMoleculardetection of Leishmania in phlebotomine sand flies in a cuta-neous and visceral leishmaniasis endemic area in northeasternBrazilrdquo Revista do Instituto de Medicina Tropical de Sao Paulovol 56 no 4 pp 357ndash360 2014
[34] R S Lana E M Michalsky C L Fortes-Dias et al ldquoPhle-botomine sand fly fauna and Leishmania infection in thevicinity of the Serra do Cipo National Park a natural Brazilianheritage siterdquo BioMed Research International vol 2015 ArticleID 385493 9 pages 2015
[35] V R Alves R A de Freitas F L Santos A F J de OliveiraT V Barrett and P H F Shimabukuro ldquoSand flies (DipteraPsychodidae Phlebotominae) from central amazonia and fournew records for the Amazonas State Brazilrdquo Revista Brasileirade Entomologia vol 56 no 2 pp 220ndash227 2012
[36] J V S Ferreira T V Santos E M Santos and I S GorayebldquoPhlebotomine sand flies (Diptera Psychodidae) in forestfragments of Belem metropolitan area Para State Brazil withconsiderations on vectors of American cutaneous leishmaniasisagentsrdquo Revista Pan-Amazonica de Saude vol 5 no 2 pp 29ndash35 2014
[37] B M Carvalho E F Rangel P D Ready and M M ValeldquoEcological niche modelling predicts southward expansion ofLutzomyia (Nyssomyia) flaviscutellata (Diptera PsychodidaePhlebotominae) vector of Leishmania (Leishmania) amazonen-sis in South America under climate changerdquo PLoS ONE vol 10no 11 Article ID e0143282 2015
[38] F Fouque P Gaborit J Issaly et al ldquoPhlebotomine sand flies(Diptera Psychodidae) associatedwith changing patterns in thetransmission of the human cutaneous leishmaniasis in FrenchGuianardquoMemorias do Instituto Oswaldo Cruz vol 102 no 1 pp35ndash40 2007
[39] R Killick-Kendrick ldquoSome epidemiological consequences ofthe evolutionary fit between Leishmaniae and their phle-botomine vectorsrdquo Bulletin de la Societe de Pathologie Exotiqueet de ses Filiales vol 78 no 5 pp 747ndash755 1985
[40] F X Pajot F Le Pont BGentile andR Besnard ldquoEpidemiologyof leishmaniasis in French Guianardquo Transactions of the RoyalSociety of Tropical Medicine and Hygiene vol 76 no 1 pp 112ndash113 1982
[41] H A Christensen J R Arias A M De Vasquez and R ADe Freitas ldquoHosts of sandfly vectors of Leishmania braziliensisguyanensis in the central Amazon of Brazilrdquo The AmericanJournal of Tropical Medicine and Hygiene vol 31 no 2 pp 239ndash242 1982
[42] L C R Nery E S Lorosa andAM R Franco ldquoFeeding prefer-ence of the sand flies Lutzomyia umbratilis and L spathotrichia(Diptera Psychodidae Phlebotominae) in an urban forest patchin the City ofManaus Amazonas BrazilrdquoMemorias do InstitutoOswaldo Cruz vol 99 no 6 pp 571ndash574 2004
[43] PD Ready R Lainson J J Shaw andAA Souza ldquoDNAprobesfor distinguishing Psychodopygus wellcomei from Psychodopy-gus complexus (Diptera Psychodidae)rdquo Memorias do InstitutoOswaldo Cruz vol 86 no 1 pp 41ndash49 1991
[44] P D Ready H Fraiha R P Lane J R Arias and F X PajotldquoOn distinguishing the female of Psychodopygus wellcomei avector of mucocutaneous leishmaniasis from other squamiven-tris series females I Characterization of Ps squamiventrissquamiventris and Ps s maripaensis stat nov (Diptera Psycho-didae)rdquo Annals of Tropical Medicine and Parasitology vol 76no 2 pp 201ndash214 1982
[45] A D Kent T V dos Santos A Gangadin A Samjhawan DR A Mans and H D F H Schallig ldquoStudies on the sandfly fauna (Diptera Psychodidae) in high-transmission areas ofcutaneous leishmaniasis in the Republic of Surinamerdquo Parasitesand Vectors vol 6 no 1 article 318 2013
[46] P D Ready R Lainson and J J Shaw ldquoHabitat and seasonalityof Psychodopygus wellcomei help incriminate it as a vector ofLeishmania braziliensis in Amazonia and Northeast BrazilrdquoTransactions of the Royal Society of Tropical Medicine andHygiene vol 78 no 4 pp 543ndash544 1984
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

8 Journal of Tropical Medicine
010203040506070
Bi flaviscutellata
01020304050607080
Ny umbratilis
0510152025303540
Ps squamiventris sl
0100200300400500600
050
100150200250300350400
2012 2013 2014 2015
Wat
erflo
odin
g
Ps s
quya
miv
entri
s sl
Bi fl
avisc
utell
ata
Ny u
mbr
atili
s
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
050
100150200250300350400
050
100150200250300350400
050
100150200250300350400
010203040506070
10000 20000 30000 40000000Rainfall (mm)
01020304050607080
10000 20000 30000 40000000Rainfall (mm)
Rainfall (mm)Overall
Rainfall (mm)
Rainfall (mm)
Rainfall (mm)
0100200300400500600
Ove
rall
10000 20000 30000 40000000Rainfall (mm)
05
10152025303540
10000 20000 30000 40000000Rainfall (mm)
r = 065 p = 000001
r = 048 p = 00021
r = 049 p = 00021
r = 065 p = 000001
Figure 3 Correlation between rainfall and phlebotomine putative vector species captured in Santo Antonio do Jari hydroelectric systemimpacted area between the states of Para and Amapa Brazil (2012ndash2015)
Journal of Tropical Medicine 9
2012 2013 2014 2015
Wat
erflo
odin
g
0100200300400500600
010203040506070
Bi flaviscutellata
01020304050607080
Ny umbratilis
0510152025303540
Ps squamiventris sl
Bi fl
avisc
utell
ata
05
1015202530
05
1015202530
05
1015202530
05
1015202530
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
0100200300400500600
Ove
rall
10 20 300ACL cases
010203040506070
10 20 300ACL cases
01020304050607080
Ny u
mbr
atili
s
10 20 300ACL cases
ACL casesOverall
ACL cases
ACL cases
ACL cases
05
10152025303540
Ps s
quam
iven
tris s
l
10 20 300ACL cases
r = 044 p = 00063
r = minus002 p = 00001
r = 048 p = 0002
r = 032 p = 00003
Figure 4 Correlation between ACL cases and phlebotomine putative vector species captured in Santo Antonio do Jari hydroelectric systemimpacted area between the states of Para and Amapa Brazil (2012ndash2015)
10 Journal of Tropical Medicine
undergo stress adaptation (evolutionary fitness) and formnew ecological systems driven by coevolution [39]
There is no doubt that Ny umbratilis is the main vectorof ACL in the Jari River Basin [15] Surprisingly the well-documented tree-dwelling behavior of this species [15 40]was not observed in CDC canopy captures in the presentstudy very little difference was found between the number ofindividuals captured in the three sampled strata (58 at 05m39 at 1m and 49 at 20m) Although Ny umbratilis is knownto climb into the canopy at night to feed on edentates (mainlysloths) [12 41] it is possible that an alternative terrestrialsource of blood such as rodents may be attracting femalestoward ground level Sloths are very slow moving and aretherefore commonly rescued for relocation during environ-mentalmonitoring operations their resulting rarenessmeansNy umbratilis has had to find other food sources Converselyblood meal analysis conducted during studies in a forestfragment of the Western Amazon has shown rodent bloodto be the predominant feeding source for Ny umbratilis [42]Wild and synanthropic rodents have been shown to surviveenvironmental disturbance in Jari and to proliferate in areasof introduced tree species [14] this indicated a plausiblereason not only for the putative adaptation of Ny umbratilisbut also for the increasing population levels of the rodentblood-feeder Bi flaviscutellata as already discussed
Given the clear importance of Psychodopygus of theChagasi series (formerly referred to as the Squamiventrisseries) in the context of ACL transmission [22] taxonomicstudies on identifying the cryptic females have been under-taken previously [43] and more recently with interestingresults (Rodrigo E Godoy unpublished) These techniqueshowever could not be employed in the field at the time Earlyconsideration of the assumed geographic distribution of thecomplex Ps squamiventris sl [1 44] shows that individualsfrom AP are likely to be predominantly Ps s maripaensisDetermining males of the species present could clarify thesetaxonomic uncertainties in the area covered by the presentstudy however doubts about identification of some malespecimens meant they were grouped as Ps squamiventris sl
Ryan et al [22] did not find Ps squamiventris sl to benaturally infected in the Jari area but proposed a link to L(V) naiffi along the same northern bank of the AmazonRiver through studies in Cachoeira Porteira (PA) Fouqueet al [38] came to the same conclusions in French GuianaInterestingly this fly species complex was found alongside L(V) braziliensis DNA in Suriname [45] an area where theknown Amazonian vectors of this parasite Ps complexus andPs wellcomei are unlikely to occur As previously discussedin the context of Bi flaviscutellata the changing transmissionpatterns of unnatural vectorparasite systems must be care-fully interpreted and only after strong ecological associationshave been determined Thus the putative vector importanceof Ps squamiventris sl in Jari can only be assumed in respectof L (V) naiffi
The five surveyed areas presented in general differentdiversity indexes showing variations in the phlebotominecomposition along the affected area This finding can beattributed not only to the distance between capture sitesbut also to forest composition fragmentation geographic
barriers and migration of mammal fauna between sites thislatter can occur naturally or be induced by rescue opera-tions involved in hydroelectric planning whereby animalsare moved to suitable ldquodry landrdquo environments safe frompotential flooding
A decrease in the phlebotomine population in 2014 inthe period after filling the reservoir could not be attributedexclusively either to flooding or to the transition from therainy to the dry season Both eventualities although oppositein effect are likely to lead to unsuitable conditions forphlebotomine reproduction However they did coincide inMay 2014 when rainfall decreased and the lake was formed
Conversely rainfall was a valid determinant of overallphlebotomine fluctuation Ps squamiventris sl was the mainputative vector directly correlated with this Other variablessuch as forest composition and altitudinal variation areknown to influence seasonal dynamics of species from thecomplex Ps squamiventris sl For example the dry periodfrom October to November in Serra dos Carajas (southernPA) can be unsuitable for Ps wellcomei which is adapted tohigh altitudes in contrast to Ps complexus which flourishesin lowland areas during the same months [46]
An insignificant negative to a moderate positive correla-tion was observed between cases of ACL and phlebotominefluctuations In the case of Ny umbratilis however theapparently higher correlation when compared with otherspecies is in agreement with its known vector importance inAmazonGuianan ACL A strong spatiotemporal associationbetween a given vector and ACL cases is considered essentialto maintenance of the Leishmania life cycle [2 14]
In summary the present study filled a temporal gapon phlebotomine data relating to the Jari area It providesa meaningful update on species composition and furtherinsights into seasonal dynamics of species under pressurescaused by environmental change The presence of ACLvectors in particular Ny umbratilis Bi flaviscutellata andPs squamiventris sl is noteworthy due to their directassociation with AmazonGuianan-ACL transmission ForNy umbratilis the lower frequency associatedwith the unpre-dicted lack of tree-dwelling behavior may be attributableto a reduction in numbers of canopy-dwelling mammalsdue to the environmental disturbance and relocation effortsthis could consequently have led to seeking for alternativeterrestrial blood sources at ground level The possible asso-ciation of Ps davisi with Leishmania transmission cycles inother Amazon ecoregions draws attention to its high rankposition in Jari Rainfall can be partly associated with localseasonal fluctuations in fly numbers mainly with respect toPs squamiventris sl Cases of ACL were also more highlycorrelated with Ny umbratilis dynamics Further attentionwas also given to results for Bi flaviscutellata which hasbeen apparently tolerant of anthropic activity continuedphlebotominemonitoring is proposed in order to evaluate theimplication for social welfare and health in the future
Competing Interests
The authors declare that they have no competing interests
Journal of Tropical Medicine 11
Acknowledgments
Theauthorswish to thank the ldquoInstituto de Pesquisas Cientıfi-cas e Tecnologicas do Estado do Amapa IEPArdquo entomologyteam for the technical assistance given to the field work inparticular to Aderbal Amanajas Santana Rosalvo Dias deCastro Noel Fernandes Santos Neto Osita Cleyriane Lobatodo Nascimento Jose Luiz dos Santos Chaves and Joao doSocorro Gomes Campos Logistical facilities were financiallysupported by the ldquoJari Energiardquo
References
[1] D G Young and M A Duncan ldquoGuide to the identificationand geographic distribution of Lutzomyia sand flies in Mexicothe West Indies Central and South America (Diptera Psycho-didae)rdquo inMemoirs of the American Entomological Institute pp1ndash881 Associated Publishers American Entomological InstituteGainesville Fla USA 1994
[2] P D Ready ldquoBiology of phlebotomine sand flies as vectors ofdisease agentsrdquo Annual Review of Entomology vol 58 pp 227ndash250 2013
[3] R P Brazil A A Fuzzari and J D Andrade Filho ldquoSandfly vectors of Leishmania in the Americasmdasha mini reviewrdquoEntomology Ornithology amp Herpetology vol 4 no 2 Article ID1000144 2015
[4] E F Rangel and R Lainson ldquoProven and putative vectorsof American cutaneous leishmaniasis in Brazil aspects oftheir biology and vectorial competencerdquoMemorias do InstitutoOswaldo Cruz vol 104 no 7 pp 937ndash954 2009
[5] E F Rangel S M Costa and B M Carvalho ldquoEnvironmentalchanges and the geographic spreading of American cutaneousleishmaniasis in Brazilrdquo in Leishmaniasis-Trends in Epidemiol-ogy D Claborn Ed Diagnosis and Treatment InTech RijekaCroatia 2014
[6] W P Taddei ldquoO genero Mansonia (Diptera Culicidae) e aproliferacao de mosquitos na usina hidreletrica de Tucuruırdquo inEnergia na Amazonia S B Magalhaes R C Brito and E RCastro Eds vol 1 pp 311ndash318 MPEGFPAUNAMAZ BelemBrazil 1996 (Portuguese)
[7] A I A Oliveira Observacoes Sobre a Resolucao CONAMA no23797 que Dispoe Sobre o Licenciamento Ambiental MimeoConselho Empresarial Brasileiro Para o Desenvolvimento Sus-tentavel Rio de Janeiro Brazil 1997 (Portuguese)
[8] A K R Galardo C D Galardo G A Silveira et al ldquoPhle-botominae sand flies (Diptera Psychodidae) potential vectorsof American cutaneous leishmaniasis agents in the area asso-ciated with the Santo Antonio hydroelectric system in WesternAmazonian Brazilrdquo Revista da Sociedade Brasileira deMedicinaTropical vol 48 no 3 pp 265ndash271 2015
[9] R Lainson R DWard and J J Shaw ldquoCutaneous leishmaniasisin North Brazil Lutzomyia anduzei as a major vectorrdquo Transac-tions of the Royal Society of Tropical Medicine and Hygiene vol70 no 2 pp 171ndash172 1976
[10] R Lainson J J Shaw R D Ward P D Ready and R DNaiff ldquoLeishmaniasis in Brazil XIII Isolation of Leishmaniafrom armadillos (Dasypus novemcinctus) and observations onthe epidemiology of cutaneous leishmaniasis in north ParaStaterdquo Transactions of the Royal Society of Tropical Medicine andHygiene vol 73 no 2 pp 239ndash242 1979
[11] R Lainson J J Shaw P D Ready M A Miles and M PovoaldquoLeishmaniasis in Brazil XVI Isolation and identification of
Leishmania species from sandflies wild mammals and manin north Para State with particular reference to L braziliensisguyanensis causative agent of ldquopian-boisrdquordquo Transactions of theRoyal Society of Tropical Medicine and Hygiene vol 75 no 4pp 530ndash536 1981
[12] R Lainson J J Shaw and M Povoa ldquoThe importance of eden-tates (sloths and anteaters) as primary reservoirs of Leishmaniabraziliensis guyanensis causative agent of lsquopian boisrsquo in northBrazilrdquo Transactions of the Royal Society of Tropical Medicineand Hygiene vol 75 no 4 pp 611ndash612 1981
[13] R DWard and H Fraiha ldquoLutzomyia umbratilis a new speciesof sand fly from Brazil (Diptera Psychodidae)rdquo Journal ofMedical Entomology vol 14 no 3 pp 313ndash317 1977
[14] P D Ready R Lainson and J J Shaw ldquoLeishmaniasis in BrazilXX Prevalence of lsquoenzootic rodent leishmaniasisrsquo (Leishmaniamexicana amazonensis) and apparent absence of lsquopian boisrsquo(Le braziliensis guyanensis) in plantations of introduced treespecies and in other non-climax forests in eastern AmazoniardquoTransactions of the Royal Society of Tropical Medicine andHygiene vol 77 no 6 pp 775ndash785 1983
[15] P D Ready R Lainson J J Shaw and R DWard ldquoThe ecologyof Lutzomyia umbratilisWard amp Fraiha (Diptera Psychodidae)the major vector to man of Leishmania braziliensis guyanensisin north-eastern Amazonian Brazilrdquo Bulletin of EntomologicalResearch vol 76 no 1 pp 21ndash40 1986
[16] R Lainson and J J Shaw ldquoLeishmania (Viannia) naiffi spn a parasite of the armadillo Dasypus novemcinctus (L)in Amazonian Brazilrdquo Annales de Parasitologie Humaine etComparee vol 64 no 1 pp 3ndash9 1989
[17] A Roger M Nacher M Hanf et al ldquoClimate and leishmaniasisin French guianardquo The American Journal of Tropical Medicineand Hygiene vol 89 no 3 pp 564ndash569 2013
[18] L Ryan ldquoFlebotomos do estado do Para Brasil (Diptera Psy-chodidae)rdquo Thecnical Document 1 Instituto Evandro ChagasBelem Brazil 1986 (Portuguese)
[19] E A B Galati ldquoMorfologia e Taxonomia Morfologia termi-nologia de adultos e identificacao dos taxons da Americardquoin Flebotomıneos do Brasil E F Rangel and R LainsonEds pp 53ndash75 Editora Fiocruz Rio de Janeiro Brazil 2003(Portuguese)
[20] D R Roberts and B P Hsi ldquoAn index of species abundance foruse with mosquito surveillance datardquo Environmental Entomol-ogy vol 8 no 6 pp 1007ndash1013 1979
[21] Oslash Hammer D A T Harper and P D Ryan ldquoPast paleontolog-ical statistics software package for education and data analysisrdquoPalaeontologia Electronica vol 4 no 1 pp 1ndash9 2001
[22] L Ryan R Lainson J J Shaw R R Braga and E A IshikawaldquoLeishmaniasis in Brazil XXV Sandfly vectors of Leishmania inPara State Brazilrdquo Medical and Veterinary Entomology vol 1no 4 pp 383ndash395 1987
[23] R Lainson J J Shaw F T Silveira R R Braga and E AIshikawa ldquoCutaneous leishmaniasis of man due to Leishmania(Viannia) naiffirdquo Annales de Parasitologie Humaine et Compa-ree vol 65 no 5-6 pp 282ndash284 1990
[24] E G C Bermudez Lutzomyia Sand Flies in the BrazilianAmazon Basin (Diptera Psychodidae) INPA Manaus Brazil2009
[25] G M Aguiar and W M Medeiros ldquoDistribuicao regionale habitats das especies de flebotomıneos do Brasilrdquo in Fle-botomıneos do Brasil E F Rangel and R Lainson Eds pp 207ndash256 Fiocruz Rio de Janeiro Brazil 2003 (Portuguese)
12 Journal of Tropical Medicine
[26] J R Arias M A Miles R D Naiff et al ldquoFlagellate infectionsof Brazilian sand flies (Diptera Psychodidae) isolation invitro and biochemical identification of Endotrypanum andLeishmaniardquo The American Journal of Tropical Medicine andHygiene vol 34 no 6 pp 1098ndash1108 1985
[27] A A Souza T V Santos Y L Jennings et al ldquoNaturalLeishmania(Viannia) spp infections in phlebotomine sand flies(Diptera Psychodidae) from the Brazilian Amazon regionreveal new putative transmission cycles of American cutaneousleishmaniasisrdquo Parasite vol 23 article 22 2016
[28] F T Silveira A A Souza R Lainson J J Shaw R R Bragaand E A Ishikawa ldquoCutaneous leishmaniasis in the AmazonRegion natural infection of the sand fly Lutzomyia ubiquitalis(Psychodidae Phlebotominae) by Leishmania (Viannia) lain-soni in Para state BrazilrdquoMemorias do Instituto Oswaldo Cruzvol 86 no 1 pp 127ndash130 1991
[29] LH S Gil S A Basano A A Souza et al ldquoRecent observationson the sand fly (Diptera Psychodidae) fauna of the state ofRondonia Western Amazonia Brazil the importance of Psych-dopygus davisi as a vector of zoonotic cutaneous leishmaniasisrdquoMemorias do Instituto Oswaldo Cruz vol 98 no 6 pp 751ndash7552003
[30] A A A Souza F T Silveira R Lainson et al ldquoThe Phlebotom-inae fauna of Serra dos Carajas Para Brazil and its possibleimplication in the transmission of American tegumentaryleishmaniasisrdquo Revista Pan-Amazonica de Saude vol 1 no 1pp 45ndash51 2010
[31] R Killick-Kendrick ldquoPhlebotomine vectors of the leishmani-ases a reviewrdquoMedical and Veterinary Entomology vol 4 no 1pp 1ndash24 1990
[32] M E C Dorval G Cristaldo H C da Rocha et al ldquoPhle-botomine fauna (Diptera Psychodidae) of an American cuta-neous leishmaniasis endemic area in the state of Mato Grossodo Sul Brazilrdquo Memorias do Instituto Oswaldo Cruz vol 104no 5 pp 695ndash702 2009
[33] V C F V Guimaraes P L Costa F J da Silva et al ldquoMoleculardetection of Leishmania in phlebotomine sand flies in a cuta-neous and visceral leishmaniasis endemic area in northeasternBrazilrdquo Revista do Instituto de Medicina Tropical de Sao Paulovol 56 no 4 pp 357ndash360 2014
[34] R S Lana E M Michalsky C L Fortes-Dias et al ldquoPhle-botomine sand fly fauna and Leishmania infection in thevicinity of the Serra do Cipo National Park a natural Brazilianheritage siterdquo BioMed Research International vol 2015 ArticleID 385493 9 pages 2015
[35] V R Alves R A de Freitas F L Santos A F J de OliveiraT V Barrett and P H F Shimabukuro ldquoSand flies (DipteraPsychodidae Phlebotominae) from central amazonia and fournew records for the Amazonas State Brazilrdquo Revista Brasileirade Entomologia vol 56 no 2 pp 220ndash227 2012
[36] J V S Ferreira T V Santos E M Santos and I S GorayebldquoPhlebotomine sand flies (Diptera Psychodidae) in forestfragments of Belem metropolitan area Para State Brazil withconsiderations on vectors of American cutaneous leishmaniasisagentsrdquo Revista Pan-Amazonica de Saude vol 5 no 2 pp 29ndash35 2014
[37] B M Carvalho E F Rangel P D Ready and M M ValeldquoEcological niche modelling predicts southward expansion ofLutzomyia (Nyssomyia) flaviscutellata (Diptera PsychodidaePhlebotominae) vector of Leishmania (Leishmania) amazonen-sis in South America under climate changerdquo PLoS ONE vol 10no 11 Article ID e0143282 2015
[38] F Fouque P Gaborit J Issaly et al ldquoPhlebotomine sand flies(Diptera Psychodidae) associatedwith changing patterns in thetransmission of the human cutaneous leishmaniasis in FrenchGuianardquoMemorias do Instituto Oswaldo Cruz vol 102 no 1 pp35ndash40 2007
[39] R Killick-Kendrick ldquoSome epidemiological consequences ofthe evolutionary fit between Leishmaniae and their phle-botomine vectorsrdquo Bulletin de la Societe de Pathologie Exotiqueet de ses Filiales vol 78 no 5 pp 747ndash755 1985
[40] F X Pajot F Le Pont BGentile andR Besnard ldquoEpidemiologyof leishmaniasis in French Guianardquo Transactions of the RoyalSociety of Tropical Medicine and Hygiene vol 76 no 1 pp 112ndash113 1982
[41] H A Christensen J R Arias A M De Vasquez and R ADe Freitas ldquoHosts of sandfly vectors of Leishmania braziliensisguyanensis in the central Amazon of Brazilrdquo The AmericanJournal of Tropical Medicine and Hygiene vol 31 no 2 pp 239ndash242 1982
[42] L C R Nery E S Lorosa andAM R Franco ldquoFeeding prefer-ence of the sand flies Lutzomyia umbratilis and L spathotrichia(Diptera Psychodidae Phlebotominae) in an urban forest patchin the City ofManaus Amazonas BrazilrdquoMemorias do InstitutoOswaldo Cruz vol 99 no 6 pp 571ndash574 2004
[43] PD Ready R Lainson J J Shaw andAA Souza ldquoDNAprobesfor distinguishing Psychodopygus wellcomei from Psychodopy-gus complexus (Diptera Psychodidae)rdquo Memorias do InstitutoOswaldo Cruz vol 86 no 1 pp 41ndash49 1991
[44] P D Ready H Fraiha R P Lane J R Arias and F X PajotldquoOn distinguishing the female of Psychodopygus wellcomei avector of mucocutaneous leishmaniasis from other squamiven-tris series females I Characterization of Ps squamiventrissquamiventris and Ps s maripaensis stat nov (Diptera Psycho-didae)rdquo Annals of Tropical Medicine and Parasitology vol 76no 2 pp 201ndash214 1982
[45] A D Kent T V dos Santos A Gangadin A Samjhawan DR A Mans and H D F H Schallig ldquoStudies on the sandfly fauna (Diptera Psychodidae) in high-transmission areas ofcutaneous leishmaniasis in the Republic of Surinamerdquo Parasitesand Vectors vol 6 no 1 article 318 2013
[46] P D Ready R Lainson and J J Shaw ldquoHabitat and seasonalityof Psychodopygus wellcomei help incriminate it as a vector ofLeishmania braziliensis in Amazonia and Northeast BrazilrdquoTransactions of the Royal Society of Tropical Medicine andHygiene vol 78 no 4 pp 543ndash544 1984
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Journal of Tropical Medicine 9
2012 2013 2014 2015
Wat
erflo
odin
g
0100200300400500600
010203040506070
Bi flaviscutellata
01020304050607080
Ny umbratilis
0510152025303540
Ps squamiventris sl
Bi fl
avisc
utell
ata
05
1015202530
05
1015202530
05
1015202530
05
1015202530
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
May Ju
lSe
pN
ov Jan
Mar
0100200300400500600
Ove
rall
10 20 300ACL cases
010203040506070
10 20 300ACL cases
01020304050607080
Ny u
mbr
atili
s
10 20 300ACL cases
ACL casesOverall
ACL cases
ACL cases
ACL cases
05
10152025303540
Ps s
quam
iven
tris s
l
10 20 300ACL cases
r = 044 p = 00063
r = minus002 p = 00001
r = 048 p = 0002
r = 032 p = 00003
Figure 4 Correlation between ACL cases and phlebotomine putative vector species captured in Santo Antonio do Jari hydroelectric systemimpacted area between the states of Para and Amapa Brazil (2012ndash2015)
10 Journal of Tropical Medicine
undergo stress adaptation (evolutionary fitness) and formnew ecological systems driven by coevolution [39]
There is no doubt that Ny umbratilis is the main vectorof ACL in the Jari River Basin [15] Surprisingly the well-documented tree-dwelling behavior of this species [15 40]was not observed in CDC canopy captures in the presentstudy very little difference was found between the number ofindividuals captured in the three sampled strata (58 at 05m39 at 1m and 49 at 20m) Although Ny umbratilis is knownto climb into the canopy at night to feed on edentates (mainlysloths) [12 41] it is possible that an alternative terrestrialsource of blood such as rodents may be attracting femalestoward ground level Sloths are very slow moving and aretherefore commonly rescued for relocation during environ-mentalmonitoring operations their resulting rarenessmeansNy umbratilis has had to find other food sources Converselyblood meal analysis conducted during studies in a forestfragment of the Western Amazon has shown rodent bloodto be the predominant feeding source for Ny umbratilis [42]Wild and synanthropic rodents have been shown to surviveenvironmental disturbance in Jari and to proliferate in areasof introduced tree species [14] this indicated a plausiblereason not only for the putative adaptation of Ny umbratilisbut also for the increasing population levels of the rodentblood-feeder Bi flaviscutellata as already discussed
Given the clear importance of Psychodopygus of theChagasi series (formerly referred to as the Squamiventrisseries) in the context of ACL transmission [22] taxonomicstudies on identifying the cryptic females have been under-taken previously [43] and more recently with interestingresults (Rodrigo E Godoy unpublished) These techniqueshowever could not be employed in the field at the time Earlyconsideration of the assumed geographic distribution of thecomplex Ps squamiventris sl [1 44] shows that individualsfrom AP are likely to be predominantly Ps s maripaensisDetermining males of the species present could clarify thesetaxonomic uncertainties in the area covered by the presentstudy however doubts about identification of some malespecimens meant they were grouped as Ps squamiventris sl
Ryan et al [22] did not find Ps squamiventris sl to benaturally infected in the Jari area but proposed a link to L(V) naiffi along the same northern bank of the AmazonRiver through studies in Cachoeira Porteira (PA) Fouqueet al [38] came to the same conclusions in French GuianaInterestingly this fly species complex was found alongside L(V) braziliensis DNA in Suriname [45] an area where theknown Amazonian vectors of this parasite Ps complexus andPs wellcomei are unlikely to occur As previously discussedin the context of Bi flaviscutellata the changing transmissionpatterns of unnatural vectorparasite systems must be care-fully interpreted and only after strong ecological associationshave been determined Thus the putative vector importanceof Ps squamiventris sl in Jari can only be assumed in respectof L (V) naiffi
The five surveyed areas presented in general differentdiversity indexes showing variations in the phlebotominecomposition along the affected area This finding can beattributed not only to the distance between capture sitesbut also to forest composition fragmentation geographic
barriers and migration of mammal fauna between sites thislatter can occur naturally or be induced by rescue opera-tions involved in hydroelectric planning whereby animalsare moved to suitable ldquodry landrdquo environments safe frompotential flooding
A decrease in the phlebotomine population in 2014 inthe period after filling the reservoir could not be attributedexclusively either to flooding or to the transition from therainy to the dry season Both eventualities although oppositein effect are likely to lead to unsuitable conditions forphlebotomine reproduction However they did coincide inMay 2014 when rainfall decreased and the lake was formed
Conversely rainfall was a valid determinant of overallphlebotomine fluctuation Ps squamiventris sl was the mainputative vector directly correlated with this Other variablessuch as forest composition and altitudinal variation areknown to influence seasonal dynamics of species from thecomplex Ps squamiventris sl For example the dry periodfrom October to November in Serra dos Carajas (southernPA) can be unsuitable for Ps wellcomei which is adapted tohigh altitudes in contrast to Ps complexus which flourishesin lowland areas during the same months [46]
An insignificant negative to a moderate positive correla-tion was observed between cases of ACL and phlebotominefluctuations In the case of Ny umbratilis however theapparently higher correlation when compared with otherspecies is in agreement with its known vector importance inAmazonGuianan ACL A strong spatiotemporal associationbetween a given vector and ACL cases is considered essentialto maintenance of the Leishmania life cycle [2 14]
In summary the present study filled a temporal gapon phlebotomine data relating to the Jari area It providesa meaningful update on species composition and furtherinsights into seasonal dynamics of species under pressurescaused by environmental change The presence of ACLvectors in particular Ny umbratilis Bi flaviscutellata andPs squamiventris sl is noteworthy due to their directassociation with AmazonGuianan-ACL transmission ForNy umbratilis the lower frequency associatedwith the unpre-dicted lack of tree-dwelling behavior may be attributableto a reduction in numbers of canopy-dwelling mammalsdue to the environmental disturbance and relocation effortsthis could consequently have led to seeking for alternativeterrestrial blood sources at ground level The possible asso-ciation of Ps davisi with Leishmania transmission cycles inother Amazon ecoregions draws attention to its high rankposition in Jari Rainfall can be partly associated with localseasonal fluctuations in fly numbers mainly with respect toPs squamiventris sl Cases of ACL were also more highlycorrelated with Ny umbratilis dynamics Further attentionwas also given to results for Bi flaviscutellata which hasbeen apparently tolerant of anthropic activity continuedphlebotominemonitoring is proposed in order to evaluate theimplication for social welfare and health in the future
Competing Interests
The authors declare that they have no competing interests
Journal of Tropical Medicine 11
Acknowledgments
Theauthorswish to thank the ldquoInstituto de Pesquisas Cientıfi-cas e Tecnologicas do Estado do Amapa IEPArdquo entomologyteam for the technical assistance given to the field work inparticular to Aderbal Amanajas Santana Rosalvo Dias deCastro Noel Fernandes Santos Neto Osita Cleyriane Lobatodo Nascimento Jose Luiz dos Santos Chaves and Joao doSocorro Gomes Campos Logistical facilities were financiallysupported by the ldquoJari Energiardquo
References
[1] D G Young and M A Duncan ldquoGuide to the identificationand geographic distribution of Lutzomyia sand flies in Mexicothe West Indies Central and South America (Diptera Psycho-didae)rdquo inMemoirs of the American Entomological Institute pp1ndash881 Associated Publishers American Entomological InstituteGainesville Fla USA 1994
[2] P D Ready ldquoBiology of phlebotomine sand flies as vectors ofdisease agentsrdquo Annual Review of Entomology vol 58 pp 227ndash250 2013
[3] R P Brazil A A Fuzzari and J D Andrade Filho ldquoSandfly vectors of Leishmania in the Americasmdasha mini reviewrdquoEntomology Ornithology amp Herpetology vol 4 no 2 Article ID1000144 2015
[4] E F Rangel and R Lainson ldquoProven and putative vectorsof American cutaneous leishmaniasis in Brazil aspects oftheir biology and vectorial competencerdquoMemorias do InstitutoOswaldo Cruz vol 104 no 7 pp 937ndash954 2009
[5] E F Rangel S M Costa and B M Carvalho ldquoEnvironmentalchanges and the geographic spreading of American cutaneousleishmaniasis in Brazilrdquo in Leishmaniasis-Trends in Epidemiol-ogy D Claborn Ed Diagnosis and Treatment InTech RijekaCroatia 2014
[6] W P Taddei ldquoO genero Mansonia (Diptera Culicidae) e aproliferacao de mosquitos na usina hidreletrica de Tucuruırdquo inEnergia na Amazonia S B Magalhaes R C Brito and E RCastro Eds vol 1 pp 311ndash318 MPEGFPAUNAMAZ BelemBrazil 1996 (Portuguese)
[7] A I A Oliveira Observacoes Sobre a Resolucao CONAMA no23797 que Dispoe Sobre o Licenciamento Ambiental MimeoConselho Empresarial Brasileiro Para o Desenvolvimento Sus-tentavel Rio de Janeiro Brazil 1997 (Portuguese)
[8] A K R Galardo C D Galardo G A Silveira et al ldquoPhle-botominae sand flies (Diptera Psychodidae) potential vectorsof American cutaneous leishmaniasis agents in the area asso-ciated with the Santo Antonio hydroelectric system in WesternAmazonian Brazilrdquo Revista da Sociedade Brasileira deMedicinaTropical vol 48 no 3 pp 265ndash271 2015
[9] R Lainson R DWard and J J Shaw ldquoCutaneous leishmaniasisin North Brazil Lutzomyia anduzei as a major vectorrdquo Transac-tions of the Royal Society of Tropical Medicine and Hygiene vol70 no 2 pp 171ndash172 1976
[10] R Lainson J J Shaw R D Ward P D Ready and R DNaiff ldquoLeishmaniasis in Brazil XIII Isolation of Leishmaniafrom armadillos (Dasypus novemcinctus) and observations onthe epidemiology of cutaneous leishmaniasis in north ParaStaterdquo Transactions of the Royal Society of Tropical Medicine andHygiene vol 73 no 2 pp 239ndash242 1979
[11] R Lainson J J Shaw P D Ready M A Miles and M PovoaldquoLeishmaniasis in Brazil XVI Isolation and identification of
Leishmania species from sandflies wild mammals and manin north Para State with particular reference to L braziliensisguyanensis causative agent of ldquopian-boisrdquordquo Transactions of theRoyal Society of Tropical Medicine and Hygiene vol 75 no 4pp 530ndash536 1981
[12] R Lainson J J Shaw and M Povoa ldquoThe importance of eden-tates (sloths and anteaters) as primary reservoirs of Leishmaniabraziliensis guyanensis causative agent of lsquopian boisrsquo in northBrazilrdquo Transactions of the Royal Society of Tropical Medicineand Hygiene vol 75 no 4 pp 611ndash612 1981
[13] R DWard and H Fraiha ldquoLutzomyia umbratilis a new speciesof sand fly from Brazil (Diptera Psychodidae)rdquo Journal ofMedical Entomology vol 14 no 3 pp 313ndash317 1977
[14] P D Ready R Lainson and J J Shaw ldquoLeishmaniasis in BrazilXX Prevalence of lsquoenzootic rodent leishmaniasisrsquo (Leishmaniamexicana amazonensis) and apparent absence of lsquopian boisrsquo(Le braziliensis guyanensis) in plantations of introduced treespecies and in other non-climax forests in eastern AmazoniardquoTransactions of the Royal Society of Tropical Medicine andHygiene vol 77 no 6 pp 775ndash785 1983
[15] P D Ready R Lainson J J Shaw and R DWard ldquoThe ecologyof Lutzomyia umbratilisWard amp Fraiha (Diptera Psychodidae)the major vector to man of Leishmania braziliensis guyanensisin north-eastern Amazonian Brazilrdquo Bulletin of EntomologicalResearch vol 76 no 1 pp 21ndash40 1986
[16] R Lainson and J J Shaw ldquoLeishmania (Viannia) naiffi spn a parasite of the armadillo Dasypus novemcinctus (L)in Amazonian Brazilrdquo Annales de Parasitologie Humaine etComparee vol 64 no 1 pp 3ndash9 1989
[17] A Roger M Nacher M Hanf et al ldquoClimate and leishmaniasisin French guianardquo The American Journal of Tropical Medicineand Hygiene vol 89 no 3 pp 564ndash569 2013
[18] L Ryan ldquoFlebotomos do estado do Para Brasil (Diptera Psy-chodidae)rdquo Thecnical Document 1 Instituto Evandro ChagasBelem Brazil 1986 (Portuguese)
[19] E A B Galati ldquoMorfologia e Taxonomia Morfologia termi-nologia de adultos e identificacao dos taxons da Americardquoin Flebotomıneos do Brasil E F Rangel and R LainsonEds pp 53ndash75 Editora Fiocruz Rio de Janeiro Brazil 2003(Portuguese)
[20] D R Roberts and B P Hsi ldquoAn index of species abundance foruse with mosquito surveillance datardquo Environmental Entomol-ogy vol 8 no 6 pp 1007ndash1013 1979
[21] Oslash Hammer D A T Harper and P D Ryan ldquoPast paleontolog-ical statistics software package for education and data analysisrdquoPalaeontologia Electronica vol 4 no 1 pp 1ndash9 2001
[22] L Ryan R Lainson J J Shaw R R Braga and E A IshikawaldquoLeishmaniasis in Brazil XXV Sandfly vectors of Leishmania inPara State Brazilrdquo Medical and Veterinary Entomology vol 1no 4 pp 383ndash395 1987
[23] R Lainson J J Shaw F T Silveira R R Braga and E AIshikawa ldquoCutaneous leishmaniasis of man due to Leishmania(Viannia) naiffirdquo Annales de Parasitologie Humaine et Compa-ree vol 65 no 5-6 pp 282ndash284 1990
[24] E G C Bermudez Lutzomyia Sand Flies in the BrazilianAmazon Basin (Diptera Psychodidae) INPA Manaus Brazil2009
[25] G M Aguiar and W M Medeiros ldquoDistribuicao regionale habitats das especies de flebotomıneos do Brasilrdquo in Fle-botomıneos do Brasil E F Rangel and R Lainson Eds pp 207ndash256 Fiocruz Rio de Janeiro Brazil 2003 (Portuguese)
12 Journal of Tropical Medicine
[26] J R Arias M A Miles R D Naiff et al ldquoFlagellate infectionsof Brazilian sand flies (Diptera Psychodidae) isolation invitro and biochemical identification of Endotrypanum andLeishmaniardquo The American Journal of Tropical Medicine andHygiene vol 34 no 6 pp 1098ndash1108 1985
[27] A A Souza T V Santos Y L Jennings et al ldquoNaturalLeishmania(Viannia) spp infections in phlebotomine sand flies(Diptera Psychodidae) from the Brazilian Amazon regionreveal new putative transmission cycles of American cutaneousleishmaniasisrdquo Parasite vol 23 article 22 2016
[28] F T Silveira A A Souza R Lainson J J Shaw R R Bragaand E A Ishikawa ldquoCutaneous leishmaniasis in the AmazonRegion natural infection of the sand fly Lutzomyia ubiquitalis(Psychodidae Phlebotominae) by Leishmania (Viannia) lain-soni in Para state BrazilrdquoMemorias do Instituto Oswaldo Cruzvol 86 no 1 pp 127ndash130 1991
[29] LH S Gil S A Basano A A Souza et al ldquoRecent observationson the sand fly (Diptera Psychodidae) fauna of the state ofRondonia Western Amazonia Brazil the importance of Psych-dopygus davisi as a vector of zoonotic cutaneous leishmaniasisrdquoMemorias do Instituto Oswaldo Cruz vol 98 no 6 pp 751ndash7552003
[30] A A A Souza F T Silveira R Lainson et al ldquoThe Phlebotom-inae fauna of Serra dos Carajas Para Brazil and its possibleimplication in the transmission of American tegumentaryleishmaniasisrdquo Revista Pan-Amazonica de Saude vol 1 no 1pp 45ndash51 2010
[31] R Killick-Kendrick ldquoPhlebotomine vectors of the leishmani-ases a reviewrdquoMedical and Veterinary Entomology vol 4 no 1pp 1ndash24 1990
[32] M E C Dorval G Cristaldo H C da Rocha et al ldquoPhle-botomine fauna (Diptera Psychodidae) of an American cuta-neous leishmaniasis endemic area in the state of Mato Grossodo Sul Brazilrdquo Memorias do Instituto Oswaldo Cruz vol 104no 5 pp 695ndash702 2009
[33] V C F V Guimaraes P L Costa F J da Silva et al ldquoMoleculardetection of Leishmania in phlebotomine sand flies in a cuta-neous and visceral leishmaniasis endemic area in northeasternBrazilrdquo Revista do Instituto de Medicina Tropical de Sao Paulovol 56 no 4 pp 357ndash360 2014
[34] R S Lana E M Michalsky C L Fortes-Dias et al ldquoPhle-botomine sand fly fauna and Leishmania infection in thevicinity of the Serra do Cipo National Park a natural Brazilianheritage siterdquo BioMed Research International vol 2015 ArticleID 385493 9 pages 2015
[35] V R Alves R A de Freitas F L Santos A F J de OliveiraT V Barrett and P H F Shimabukuro ldquoSand flies (DipteraPsychodidae Phlebotominae) from central amazonia and fournew records for the Amazonas State Brazilrdquo Revista Brasileirade Entomologia vol 56 no 2 pp 220ndash227 2012
[36] J V S Ferreira T V Santos E M Santos and I S GorayebldquoPhlebotomine sand flies (Diptera Psychodidae) in forestfragments of Belem metropolitan area Para State Brazil withconsiderations on vectors of American cutaneous leishmaniasisagentsrdquo Revista Pan-Amazonica de Saude vol 5 no 2 pp 29ndash35 2014
[37] B M Carvalho E F Rangel P D Ready and M M ValeldquoEcological niche modelling predicts southward expansion ofLutzomyia (Nyssomyia) flaviscutellata (Diptera PsychodidaePhlebotominae) vector of Leishmania (Leishmania) amazonen-sis in South America under climate changerdquo PLoS ONE vol 10no 11 Article ID e0143282 2015
[38] F Fouque P Gaborit J Issaly et al ldquoPhlebotomine sand flies(Diptera Psychodidae) associatedwith changing patterns in thetransmission of the human cutaneous leishmaniasis in FrenchGuianardquoMemorias do Instituto Oswaldo Cruz vol 102 no 1 pp35ndash40 2007
[39] R Killick-Kendrick ldquoSome epidemiological consequences ofthe evolutionary fit between Leishmaniae and their phle-botomine vectorsrdquo Bulletin de la Societe de Pathologie Exotiqueet de ses Filiales vol 78 no 5 pp 747ndash755 1985
[40] F X Pajot F Le Pont BGentile andR Besnard ldquoEpidemiologyof leishmaniasis in French Guianardquo Transactions of the RoyalSociety of Tropical Medicine and Hygiene vol 76 no 1 pp 112ndash113 1982
[41] H A Christensen J R Arias A M De Vasquez and R ADe Freitas ldquoHosts of sandfly vectors of Leishmania braziliensisguyanensis in the central Amazon of Brazilrdquo The AmericanJournal of Tropical Medicine and Hygiene vol 31 no 2 pp 239ndash242 1982
[42] L C R Nery E S Lorosa andAM R Franco ldquoFeeding prefer-ence of the sand flies Lutzomyia umbratilis and L spathotrichia(Diptera Psychodidae Phlebotominae) in an urban forest patchin the City ofManaus Amazonas BrazilrdquoMemorias do InstitutoOswaldo Cruz vol 99 no 6 pp 571ndash574 2004
[43] PD Ready R Lainson J J Shaw andAA Souza ldquoDNAprobesfor distinguishing Psychodopygus wellcomei from Psychodopy-gus complexus (Diptera Psychodidae)rdquo Memorias do InstitutoOswaldo Cruz vol 86 no 1 pp 41ndash49 1991
[44] P D Ready H Fraiha R P Lane J R Arias and F X PajotldquoOn distinguishing the female of Psychodopygus wellcomei avector of mucocutaneous leishmaniasis from other squamiven-tris series females I Characterization of Ps squamiventrissquamiventris and Ps s maripaensis stat nov (Diptera Psycho-didae)rdquo Annals of Tropical Medicine and Parasitology vol 76no 2 pp 201ndash214 1982
[45] A D Kent T V dos Santos A Gangadin A Samjhawan DR A Mans and H D F H Schallig ldquoStudies on the sandfly fauna (Diptera Psychodidae) in high-transmission areas ofcutaneous leishmaniasis in the Republic of Surinamerdquo Parasitesand Vectors vol 6 no 1 article 318 2013
[46] P D Ready R Lainson and J J Shaw ldquoHabitat and seasonalityof Psychodopygus wellcomei help incriminate it as a vector ofLeishmania braziliensis in Amazonia and Northeast BrazilrdquoTransactions of the Royal Society of Tropical Medicine andHygiene vol 78 no 4 pp 543ndash544 1984
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

10 Journal of Tropical Medicine
undergo stress adaptation (evolutionary fitness) and formnew ecological systems driven by coevolution [39]
There is no doubt that Ny umbratilis is the main vectorof ACL in the Jari River Basin [15] Surprisingly the well-documented tree-dwelling behavior of this species [15 40]was not observed in CDC canopy captures in the presentstudy very little difference was found between the number ofindividuals captured in the three sampled strata (58 at 05m39 at 1m and 49 at 20m) Although Ny umbratilis is knownto climb into the canopy at night to feed on edentates (mainlysloths) [12 41] it is possible that an alternative terrestrialsource of blood such as rodents may be attracting femalestoward ground level Sloths are very slow moving and aretherefore commonly rescued for relocation during environ-mentalmonitoring operations their resulting rarenessmeansNy umbratilis has had to find other food sources Converselyblood meal analysis conducted during studies in a forestfragment of the Western Amazon has shown rodent bloodto be the predominant feeding source for Ny umbratilis [42]Wild and synanthropic rodents have been shown to surviveenvironmental disturbance in Jari and to proliferate in areasof introduced tree species [14] this indicated a plausiblereason not only for the putative adaptation of Ny umbratilisbut also for the increasing population levels of the rodentblood-feeder Bi flaviscutellata as already discussed
Given the clear importance of Psychodopygus of theChagasi series (formerly referred to as the Squamiventrisseries) in the context of ACL transmission [22] taxonomicstudies on identifying the cryptic females have been under-taken previously [43] and more recently with interestingresults (Rodrigo E Godoy unpublished) These techniqueshowever could not be employed in the field at the time Earlyconsideration of the assumed geographic distribution of thecomplex Ps squamiventris sl [1 44] shows that individualsfrom AP are likely to be predominantly Ps s maripaensisDetermining males of the species present could clarify thesetaxonomic uncertainties in the area covered by the presentstudy however doubts about identification of some malespecimens meant they were grouped as Ps squamiventris sl
Ryan et al [22] did not find Ps squamiventris sl to benaturally infected in the Jari area but proposed a link to L(V) naiffi along the same northern bank of the AmazonRiver through studies in Cachoeira Porteira (PA) Fouqueet al [38] came to the same conclusions in French GuianaInterestingly this fly species complex was found alongside L(V) braziliensis DNA in Suriname [45] an area where theknown Amazonian vectors of this parasite Ps complexus andPs wellcomei are unlikely to occur As previously discussedin the context of Bi flaviscutellata the changing transmissionpatterns of unnatural vectorparasite systems must be care-fully interpreted and only after strong ecological associationshave been determined Thus the putative vector importanceof Ps squamiventris sl in Jari can only be assumed in respectof L (V) naiffi
The five surveyed areas presented in general differentdiversity indexes showing variations in the phlebotominecomposition along the affected area This finding can beattributed not only to the distance between capture sitesbut also to forest composition fragmentation geographic
barriers and migration of mammal fauna between sites thislatter can occur naturally or be induced by rescue opera-tions involved in hydroelectric planning whereby animalsare moved to suitable ldquodry landrdquo environments safe frompotential flooding
A decrease in the phlebotomine population in 2014 inthe period after filling the reservoir could not be attributedexclusively either to flooding or to the transition from therainy to the dry season Both eventualities although oppositein effect are likely to lead to unsuitable conditions forphlebotomine reproduction However they did coincide inMay 2014 when rainfall decreased and the lake was formed
Conversely rainfall was a valid determinant of overallphlebotomine fluctuation Ps squamiventris sl was the mainputative vector directly correlated with this Other variablessuch as forest composition and altitudinal variation areknown to influence seasonal dynamics of species from thecomplex Ps squamiventris sl For example the dry periodfrom October to November in Serra dos Carajas (southernPA) can be unsuitable for Ps wellcomei which is adapted tohigh altitudes in contrast to Ps complexus which flourishesin lowland areas during the same months [46]
An insignificant negative to a moderate positive correla-tion was observed between cases of ACL and phlebotominefluctuations In the case of Ny umbratilis however theapparently higher correlation when compared with otherspecies is in agreement with its known vector importance inAmazonGuianan ACL A strong spatiotemporal associationbetween a given vector and ACL cases is considered essentialto maintenance of the Leishmania life cycle [2 14]
In summary the present study filled a temporal gapon phlebotomine data relating to the Jari area It providesa meaningful update on species composition and furtherinsights into seasonal dynamics of species under pressurescaused by environmental change The presence of ACLvectors in particular Ny umbratilis Bi flaviscutellata andPs squamiventris sl is noteworthy due to their directassociation with AmazonGuianan-ACL transmission ForNy umbratilis the lower frequency associatedwith the unpre-dicted lack of tree-dwelling behavior may be attributableto a reduction in numbers of canopy-dwelling mammalsdue to the environmental disturbance and relocation effortsthis could consequently have led to seeking for alternativeterrestrial blood sources at ground level The possible asso-ciation of Ps davisi with Leishmania transmission cycles inother Amazon ecoregions draws attention to its high rankposition in Jari Rainfall can be partly associated with localseasonal fluctuations in fly numbers mainly with respect toPs squamiventris sl Cases of ACL were also more highlycorrelated with Ny umbratilis dynamics Further attentionwas also given to results for Bi flaviscutellata which hasbeen apparently tolerant of anthropic activity continuedphlebotominemonitoring is proposed in order to evaluate theimplication for social welfare and health in the future
Competing Interests
The authors declare that they have no competing interests
Journal of Tropical Medicine 11
Acknowledgments
Theauthorswish to thank the ldquoInstituto de Pesquisas Cientıfi-cas e Tecnologicas do Estado do Amapa IEPArdquo entomologyteam for the technical assistance given to the field work inparticular to Aderbal Amanajas Santana Rosalvo Dias deCastro Noel Fernandes Santos Neto Osita Cleyriane Lobatodo Nascimento Jose Luiz dos Santos Chaves and Joao doSocorro Gomes Campos Logistical facilities were financiallysupported by the ldquoJari Energiardquo
References
[1] D G Young and M A Duncan ldquoGuide to the identificationand geographic distribution of Lutzomyia sand flies in Mexicothe West Indies Central and South America (Diptera Psycho-didae)rdquo inMemoirs of the American Entomological Institute pp1ndash881 Associated Publishers American Entomological InstituteGainesville Fla USA 1994
[2] P D Ready ldquoBiology of phlebotomine sand flies as vectors ofdisease agentsrdquo Annual Review of Entomology vol 58 pp 227ndash250 2013
[3] R P Brazil A A Fuzzari and J D Andrade Filho ldquoSandfly vectors of Leishmania in the Americasmdasha mini reviewrdquoEntomology Ornithology amp Herpetology vol 4 no 2 Article ID1000144 2015
[4] E F Rangel and R Lainson ldquoProven and putative vectorsof American cutaneous leishmaniasis in Brazil aspects oftheir biology and vectorial competencerdquoMemorias do InstitutoOswaldo Cruz vol 104 no 7 pp 937ndash954 2009
[5] E F Rangel S M Costa and B M Carvalho ldquoEnvironmentalchanges and the geographic spreading of American cutaneousleishmaniasis in Brazilrdquo in Leishmaniasis-Trends in Epidemiol-ogy D Claborn Ed Diagnosis and Treatment InTech RijekaCroatia 2014
[6] W P Taddei ldquoO genero Mansonia (Diptera Culicidae) e aproliferacao de mosquitos na usina hidreletrica de Tucuruırdquo inEnergia na Amazonia S B Magalhaes R C Brito and E RCastro Eds vol 1 pp 311ndash318 MPEGFPAUNAMAZ BelemBrazil 1996 (Portuguese)
[7] A I A Oliveira Observacoes Sobre a Resolucao CONAMA no23797 que Dispoe Sobre o Licenciamento Ambiental MimeoConselho Empresarial Brasileiro Para o Desenvolvimento Sus-tentavel Rio de Janeiro Brazil 1997 (Portuguese)
[8] A K R Galardo C D Galardo G A Silveira et al ldquoPhle-botominae sand flies (Diptera Psychodidae) potential vectorsof American cutaneous leishmaniasis agents in the area asso-ciated with the Santo Antonio hydroelectric system in WesternAmazonian Brazilrdquo Revista da Sociedade Brasileira deMedicinaTropical vol 48 no 3 pp 265ndash271 2015
[9] R Lainson R DWard and J J Shaw ldquoCutaneous leishmaniasisin North Brazil Lutzomyia anduzei as a major vectorrdquo Transac-tions of the Royal Society of Tropical Medicine and Hygiene vol70 no 2 pp 171ndash172 1976
[10] R Lainson J J Shaw R D Ward P D Ready and R DNaiff ldquoLeishmaniasis in Brazil XIII Isolation of Leishmaniafrom armadillos (Dasypus novemcinctus) and observations onthe epidemiology of cutaneous leishmaniasis in north ParaStaterdquo Transactions of the Royal Society of Tropical Medicine andHygiene vol 73 no 2 pp 239ndash242 1979
[11] R Lainson J J Shaw P D Ready M A Miles and M PovoaldquoLeishmaniasis in Brazil XVI Isolation and identification of
Leishmania species from sandflies wild mammals and manin north Para State with particular reference to L braziliensisguyanensis causative agent of ldquopian-boisrdquordquo Transactions of theRoyal Society of Tropical Medicine and Hygiene vol 75 no 4pp 530ndash536 1981
[12] R Lainson J J Shaw and M Povoa ldquoThe importance of eden-tates (sloths and anteaters) as primary reservoirs of Leishmaniabraziliensis guyanensis causative agent of lsquopian boisrsquo in northBrazilrdquo Transactions of the Royal Society of Tropical Medicineand Hygiene vol 75 no 4 pp 611ndash612 1981
[13] R DWard and H Fraiha ldquoLutzomyia umbratilis a new speciesof sand fly from Brazil (Diptera Psychodidae)rdquo Journal ofMedical Entomology vol 14 no 3 pp 313ndash317 1977
[14] P D Ready R Lainson and J J Shaw ldquoLeishmaniasis in BrazilXX Prevalence of lsquoenzootic rodent leishmaniasisrsquo (Leishmaniamexicana amazonensis) and apparent absence of lsquopian boisrsquo(Le braziliensis guyanensis) in plantations of introduced treespecies and in other non-climax forests in eastern AmazoniardquoTransactions of the Royal Society of Tropical Medicine andHygiene vol 77 no 6 pp 775ndash785 1983
[15] P D Ready R Lainson J J Shaw and R DWard ldquoThe ecologyof Lutzomyia umbratilisWard amp Fraiha (Diptera Psychodidae)the major vector to man of Leishmania braziliensis guyanensisin north-eastern Amazonian Brazilrdquo Bulletin of EntomologicalResearch vol 76 no 1 pp 21ndash40 1986
[16] R Lainson and J J Shaw ldquoLeishmania (Viannia) naiffi spn a parasite of the armadillo Dasypus novemcinctus (L)in Amazonian Brazilrdquo Annales de Parasitologie Humaine etComparee vol 64 no 1 pp 3ndash9 1989
[17] A Roger M Nacher M Hanf et al ldquoClimate and leishmaniasisin French guianardquo The American Journal of Tropical Medicineand Hygiene vol 89 no 3 pp 564ndash569 2013
[18] L Ryan ldquoFlebotomos do estado do Para Brasil (Diptera Psy-chodidae)rdquo Thecnical Document 1 Instituto Evandro ChagasBelem Brazil 1986 (Portuguese)
[19] E A B Galati ldquoMorfologia e Taxonomia Morfologia termi-nologia de adultos e identificacao dos taxons da Americardquoin Flebotomıneos do Brasil E F Rangel and R LainsonEds pp 53ndash75 Editora Fiocruz Rio de Janeiro Brazil 2003(Portuguese)
[20] D R Roberts and B P Hsi ldquoAn index of species abundance foruse with mosquito surveillance datardquo Environmental Entomol-ogy vol 8 no 6 pp 1007ndash1013 1979
[21] Oslash Hammer D A T Harper and P D Ryan ldquoPast paleontolog-ical statistics software package for education and data analysisrdquoPalaeontologia Electronica vol 4 no 1 pp 1ndash9 2001
[22] L Ryan R Lainson J J Shaw R R Braga and E A IshikawaldquoLeishmaniasis in Brazil XXV Sandfly vectors of Leishmania inPara State Brazilrdquo Medical and Veterinary Entomology vol 1no 4 pp 383ndash395 1987
[23] R Lainson J J Shaw F T Silveira R R Braga and E AIshikawa ldquoCutaneous leishmaniasis of man due to Leishmania(Viannia) naiffirdquo Annales de Parasitologie Humaine et Compa-ree vol 65 no 5-6 pp 282ndash284 1990
[24] E G C Bermudez Lutzomyia Sand Flies in the BrazilianAmazon Basin (Diptera Psychodidae) INPA Manaus Brazil2009
[25] G M Aguiar and W M Medeiros ldquoDistribuicao regionale habitats das especies de flebotomıneos do Brasilrdquo in Fle-botomıneos do Brasil E F Rangel and R Lainson Eds pp 207ndash256 Fiocruz Rio de Janeiro Brazil 2003 (Portuguese)
12 Journal of Tropical Medicine
[26] J R Arias M A Miles R D Naiff et al ldquoFlagellate infectionsof Brazilian sand flies (Diptera Psychodidae) isolation invitro and biochemical identification of Endotrypanum andLeishmaniardquo The American Journal of Tropical Medicine andHygiene vol 34 no 6 pp 1098ndash1108 1985
[27] A A Souza T V Santos Y L Jennings et al ldquoNaturalLeishmania(Viannia) spp infections in phlebotomine sand flies(Diptera Psychodidae) from the Brazilian Amazon regionreveal new putative transmission cycles of American cutaneousleishmaniasisrdquo Parasite vol 23 article 22 2016
[28] F T Silveira A A Souza R Lainson J J Shaw R R Bragaand E A Ishikawa ldquoCutaneous leishmaniasis in the AmazonRegion natural infection of the sand fly Lutzomyia ubiquitalis(Psychodidae Phlebotominae) by Leishmania (Viannia) lain-soni in Para state BrazilrdquoMemorias do Instituto Oswaldo Cruzvol 86 no 1 pp 127ndash130 1991
[29] LH S Gil S A Basano A A Souza et al ldquoRecent observationson the sand fly (Diptera Psychodidae) fauna of the state ofRondonia Western Amazonia Brazil the importance of Psych-dopygus davisi as a vector of zoonotic cutaneous leishmaniasisrdquoMemorias do Instituto Oswaldo Cruz vol 98 no 6 pp 751ndash7552003
[30] A A A Souza F T Silveira R Lainson et al ldquoThe Phlebotom-inae fauna of Serra dos Carajas Para Brazil and its possibleimplication in the transmission of American tegumentaryleishmaniasisrdquo Revista Pan-Amazonica de Saude vol 1 no 1pp 45ndash51 2010
[31] R Killick-Kendrick ldquoPhlebotomine vectors of the leishmani-ases a reviewrdquoMedical and Veterinary Entomology vol 4 no 1pp 1ndash24 1990
[32] M E C Dorval G Cristaldo H C da Rocha et al ldquoPhle-botomine fauna (Diptera Psychodidae) of an American cuta-neous leishmaniasis endemic area in the state of Mato Grossodo Sul Brazilrdquo Memorias do Instituto Oswaldo Cruz vol 104no 5 pp 695ndash702 2009
[33] V C F V Guimaraes P L Costa F J da Silva et al ldquoMoleculardetection of Leishmania in phlebotomine sand flies in a cuta-neous and visceral leishmaniasis endemic area in northeasternBrazilrdquo Revista do Instituto de Medicina Tropical de Sao Paulovol 56 no 4 pp 357ndash360 2014
[34] R S Lana E M Michalsky C L Fortes-Dias et al ldquoPhle-botomine sand fly fauna and Leishmania infection in thevicinity of the Serra do Cipo National Park a natural Brazilianheritage siterdquo BioMed Research International vol 2015 ArticleID 385493 9 pages 2015
[35] V R Alves R A de Freitas F L Santos A F J de OliveiraT V Barrett and P H F Shimabukuro ldquoSand flies (DipteraPsychodidae Phlebotominae) from central amazonia and fournew records for the Amazonas State Brazilrdquo Revista Brasileirade Entomologia vol 56 no 2 pp 220ndash227 2012
[36] J V S Ferreira T V Santos E M Santos and I S GorayebldquoPhlebotomine sand flies (Diptera Psychodidae) in forestfragments of Belem metropolitan area Para State Brazil withconsiderations on vectors of American cutaneous leishmaniasisagentsrdquo Revista Pan-Amazonica de Saude vol 5 no 2 pp 29ndash35 2014
[37] B M Carvalho E F Rangel P D Ready and M M ValeldquoEcological niche modelling predicts southward expansion ofLutzomyia (Nyssomyia) flaviscutellata (Diptera PsychodidaePhlebotominae) vector of Leishmania (Leishmania) amazonen-sis in South America under climate changerdquo PLoS ONE vol 10no 11 Article ID e0143282 2015
[38] F Fouque P Gaborit J Issaly et al ldquoPhlebotomine sand flies(Diptera Psychodidae) associatedwith changing patterns in thetransmission of the human cutaneous leishmaniasis in FrenchGuianardquoMemorias do Instituto Oswaldo Cruz vol 102 no 1 pp35ndash40 2007
[39] R Killick-Kendrick ldquoSome epidemiological consequences ofthe evolutionary fit between Leishmaniae and their phle-botomine vectorsrdquo Bulletin de la Societe de Pathologie Exotiqueet de ses Filiales vol 78 no 5 pp 747ndash755 1985
[40] F X Pajot F Le Pont BGentile andR Besnard ldquoEpidemiologyof leishmaniasis in French Guianardquo Transactions of the RoyalSociety of Tropical Medicine and Hygiene vol 76 no 1 pp 112ndash113 1982
[41] H A Christensen J R Arias A M De Vasquez and R ADe Freitas ldquoHosts of sandfly vectors of Leishmania braziliensisguyanensis in the central Amazon of Brazilrdquo The AmericanJournal of Tropical Medicine and Hygiene vol 31 no 2 pp 239ndash242 1982
[42] L C R Nery E S Lorosa andAM R Franco ldquoFeeding prefer-ence of the sand flies Lutzomyia umbratilis and L spathotrichia(Diptera Psychodidae Phlebotominae) in an urban forest patchin the City ofManaus Amazonas BrazilrdquoMemorias do InstitutoOswaldo Cruz vol 99 no 6 pp 571ndash574 2004
[43] PD Ready R Lainson J J Shaw andAA Souza ldquoDNAprobesfor distinguishing Psychodopygus wellcomei from Psychodopy-gus complexus (Diptera Psychodidae)rdquo Memorias do InstitutoOswaldo Cruz vol 86 no 1 pp 41ndash49 1991
[44] P D Ready H Fraiha R P Lane J R Arias and F X PajotldquoOn distinguishing the female of Psychodopygus wellcomei avector of mucocutaneous leishmaniasis from other squamiven-tris series females I Characterization of Ps squamiventrissquamiventris and Ps s maripaensis stat nov (Diptera Psycho-didae)rdquo Annals of Tropical Medicine and Parasitology vol 76no 2 pp 201ndash214 1982
[45] A D Kent T V dos Santos A Gangadin A Samjhawan DR A Mans and H D F H Schallig ldquoStudies on the sandfly fauna (Diptera Psychodidae) in high-transmission areas ofcutaneous leishmaniasis in the Republic of Surinamerdquo Parasitesand Vectors vol 6 no 1 article 318 2013
[46] P D Ready R Lainson and J J Shaw ldquoHabitat and seasonalityof Psychodopygus wellcomei help incriminate it as a vector ofLeishmania braziliensis in Amazonia and Northeast BrazilrdquoTransactions of the Royal Society of Tropical Medicine andHygiene vol 78 no 4 pp 543ndash544 1984
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Journal of Tropical Medicine 11
Acknowledgments
Theauthorswish to thank the ldquoInstituto de Pesquisas Cientıfi-cas e Tecnologicas do Estado do Amapa IEPArdquo entomologyteam for the technical assistance given to the field work inparticular to Aderbal Amanajas Santana Rosalvo Dias deCastro Noel Fernandes Santos Neto Osita Cleyriane Lobatodo Nascimento Jose Luiz dos Santos Chaves and Joao doSocorro Gomes Campos Logistical facilities were financiallysupported by the ldquoJari Energiardquo
References
[1] D G Young and M A Duncan ldquoGuide to the identificationand geographic distribution of Lutzomyia sand flies in Mexicothe West Indies Central and South America (Diptera Psycho-didae)rdquo inMemoirs of the American Entomological Institute pp1ndash881 Associated Publishers American Entomological InstituteGainesville Fla USA 1994
[2] P D Ready ldquoBiology of phlebotomine sand flies as vectors ofdisease agentsrdquo Annual Review of Entomology vol 58 pp 227ndash250 2013
[3] R P Brazil A A Fuzzari and J D Andrade Filho ldquoSandfly vectors of Leishmania in the Americasmdasha mini reviewrdquoEntomology Ornithology amp Herpetology vol 4 no 2 Article ID1000144 2015
[4] E F Rangel and R Lainson ldquoProven and putative vectorsof American cutaneous leishmaniasis in Brazil aspects oftheir biology and vectorial competencerdquoMemorias do InstitutoOswaldo Cruz vol 104 no 7 pp 937ndash954 2009
[5] E F Rangel S M Costa and B M Carvalho ldquoEnvironmentalchanges and the geographic spreading of American cutaneousleishmaniasis in Brazilrdquo in Leishmaniasis-Trends in Epidemiol-ogy D Claborn Ed Diagnosis and Treatment InTech RijekaCroatia 2014
[6] W P Taddei ldquoO genero Mansonia (Diptera Culicidae) e aproliferacao de mosquitos na usina hidreletrica de Tucuruırdquo inEnergia na Amazonia S B Magalhaes R C Brito and E RCastro Eds vol 1 pp 311ndash318 MPEGFPAUNAMAZ BelemBrazil 1996 (Portuguese)
[7] A I A Oliveira Observacoes Sobre a Resolucao CONAMA no23797 que Dispoe Sobre o Licenciamento Ambiental MimeoConselho Empresarial Brasileiro Para o Desenvolvimento Sus-tentavel Rio de Janeiro Brazil 1997 (Portuguese)
[8] A K R Galardo C D Galardo G A Silveira et al ldquoPhle-botominae sand flies (Diptera Psychodidae) potential vectorsof American cutaneous leishmaniasis agents in the area asso-ciated with the Santo Antonio hydroelectric system in WesternAmazonian Brazilrdquo Revista da Sociedade Brasileira deMedicinaTropical vol 48 no 3 pp 265ndash271 2015
[9] R Lainson R DWard and J J Shaw ldquoCutaneous leishmaniasisin North Brazil Lutzomyia anduzei as a major vectorrdquo Transac-tions of the Royal Society of Tropical Medicine and Hygiene vol70 no 2 pp 171ndash172 1976
[10] R Lainson J J Shaw R D Ward P D Ready and R DNaiff ldquoLeishmaniasis in Brazil XIII Isolation of Leishmaniafrom armadillos (Dasypus novemcinctus) and observations onthe epidemiology of cutaneous leishmaniasis in north ParaStaterdquo Transactions of the Royal Society of Tropical Medicine andHygiene vol 73 no 2 pp 239ndash242 1979
[11] R Lainson J J Shaw P D Ready M A Miles and M PovoaldquoLeishmaniasis in Brazil XVI Isolation and identification of
Leishmania species from sandflies wild mammals and manin north Para State with particular reference to L braziliensisguyanensis causative agent of ldquopian-boisrdquordquo Transactions of theRoyal Society of Tropical Medicine and Hygiene vol 75 no 4pp 530ndash536 1981
[12] R Lainson J J Shaw and M Povoa ldquoThe importance of eden-tates (sloths and anteaters) as primary reservoirs of Leishmaniabraziliensis guyanensis causative agent of lsquopian boisrsquo in northBrazilrdquo Transactions of the Royal Society of Tropical Medicineand Hygiene vol 75 no 4 pp 611ndash612 1981
[13] R DWard and H Fraiha ldquoLutzomyia umbratilis a new speciesof sand fly from Brazil (Diptera Psychodidae)rdquo Journal ofMedical Entomology vol 14 no 3 pp 313ndash317 1977
[14] P D Ready R Lainson and J J Shaw ldquoLeishmaniasis in BrazilXX Prevalence of lsquoenzootic rodent leishmaniasisrsquo (Leishmaniamexicana amazonensis) and apparent absence of lsquopian boisrsquo(Le braziliensis guyanensis) in plantations of introduced treespecies and in other non-climax forests in eastern AmazoniardquoTransactions of the Royal Society of Tropical Medicine andHygiene vol 77 no 6 pp 775ndash785 1983
[15] P D Ready R Lainson J J Shaw and R DWard ldquoThe ecologyof Lutzomyia umbratilisWard amp Fraiha (Diptera Psychodidae)the major vector to man of Leishmania braziliensis guyanensisin north-eastern Amazonian Brazilrdquo Bulletin of EntomologicalResearch vol 76 no 1 pp 21ndash40 1986
[16] R Lainson and J J Shaw ldquoLeishmania (Viannia) naiffi spn a parasite of the armadillo Dasypus novemcinctus (L)in Amazonian Brazilrdquo Annales de Parasitologie Humaine etComparee vol 64 no 1 pp 3ndash9 1989
[17] A Roger M Nacher M Hanf et al ldquoClimate and leishmaniasisin French guianardquo The American Journal of Tropical Medicineand Hygiene vol 89 no 3 pp 564ndash569 2013
[18] L Ryan ldquoFlebotomos do estado do Para Brasil (Diptera Psy-chodidae)rdquo Thecnical Document 1 Instituto Evandro ChagasBelem Brazil 1986 (Portuguese)
[19] E A B Galati ldquoMorfologia e Taxonomia Morfologia termi-nologia de adultos e identificacao dos taxons da Americardquoin Flebotomıneos do Brasil E F Rangel and R LainsonEds pp 53ndash75 Editora Fiocruz Rio de Janeiro Brazil 2003(Portuguese)
[20] D R Roberts and B P Hsi ldquoAn index of species abundance foruse with mosquito surveillance datardquo Environmental Entomol-ogy vol 8 no 6 pp 1007ndash1013 1979
[21] Oslash Hammer D A T Harper and P D Ryan ldquoPast paleontolog-ical statistics software package for education and data analysisrdquoPalaeontologia Electronica vol 4 no 1 pp 1ndash9 2001
[22] L Ryan R Lainson J J Shaw R R Braga and E A IshikawaldquoLeishmaniasis in Brazil XXV Sandfly vectors of Leishmania inPara State Brazilrdquo Medical and Veterinary Entomology vol 1no 4 pp 383ndash395 1987
[23] R Lainson J J Shaw F T Silveira R R Braga and E AIshikawa ldquoCutaneous leishmaniasis of man due to Leishmania(Viannia) naiffirdquo Annales de Parasitologie Humaine et Compa-ree vol 65 no 5-6 pp 282ndash284 1990
[24] E G C Bermudez Lutzomyia Sand Flies in the BrazilianAmazon Basin (Diptera Psychodidae) INPA Manaus Brazil2009
[25] G M Aguiar and W M Medeiros ldquoDistribuicao regionale habitats das especies de flebotomıneos do Brasilrdquo in Fle-botomıneos do Brasil E F Rangel and R Lainson Eds pp 207ndash256 Fiocruz Rio de Janeiro Brazil 2003 (Portuguese)
12 Journal of Tropical Medicine
[26] J R Arias M A Miles R D Naiff et al ldquoFlagellate infectionsof Brazilian sand flies (Diptera Psychodidae) isolation invitro and biochemical identification of Endotrypanum andLeishmaniardquo The American Journal of Tropical Medicine andHygiene vol 34 no 6 pp 1098ndash1108 1985
[27] A A Souza T V Santos Y L Jennings et al ldquoNaturalLeishmania(Viannia) spp infections in phlebotomine sand flies(Diptera Psychodidae) from the Brazilian Amazon regionreveal new putative transmission cycles of American cutaneousleishmaniasisrdquo Parasite vol 23 article 22 2016
[28] F T Silveira A A Souza R Lainson J J Shaw R R Bragaand E A Ishikawa ldquoCutaneous leishmaniasis in the AmazonRegion natural infection of the sand fly Lutzomyia ubiquitalis(Psychodidae Phlebotominae) by Leishmania (Viannia) lain-soni in Para state BrazilrdquoMemorias do Instituto Oswaldo Cruzvol 86 no 1 pp 127ndash130 1991
[29] LH S Gil S A Basano A A Souza et al ldquoRecent observationson the sand fly (Diptera Psychodidae) fauna of the state ofRondonia Western Amazonia Brazil the importance of Psych-dopygus davisi as a vector of zoonotic cutaneous leishmaniasisrdquoMemorias do Instituto Oswaldo Cruz vol 98 no 6 pp 751ndash7552003
[30] A A A Souza F T Silveira R Lainson et al ldquoThe Phlebotom-inae fauna of Serra dos Carajas Para Brazil and its possibleimplication in the transmission of American tegumentaryleishmaniasisrdquo Revista Pan-Amazonica de Saude vol 1 no 1pp 45ndash51 2010
[31] R Killick-Kendrick ldquoPhlebotomine vectors of the leishmani-ases a reviewrdquoMedical and Veterinary Entomology vol 4 no 1pp 1ndash24 1990
[32] M E C Dorval G Cristaldo H C da Rocha et al ldquoPhle-botomine fauna (Diptera Psychodidae) of an American cuta-neous leishmaniasis endemic area in the state of Mato Grossodo Sul Brazilrdquo Memorias do Instituto Oswaldo Cruz vol 104no 5 pp 695ndash702 2009
[33] V C F V Guimaraes P L Costa F J da Silva et al ldquoMoleculardetection of Leishmania in phlebotomine sand flies in a cuta-neous and visceral leishmaniasis endemic area in northeasternBrazilrdquo Revista do Instituto de Medicina Tropical de Sao Paulovol 56 no 4 pp 357ndash360 2014
[34] R S Lana E M Michalsky C L Fortes-Dias et al ldquoPhle-botomine sand fly fauna and Leishmania infection in thevicinity of the Serra do Cipo National Park a natural Brazilianheritage siterdquo BioMed Research International vol 2015 ArticleID 385493 9 pages 2015
[35] V R Alves R A de Freitas F L Santos A F J de OliveiraT V Barrett and P H F Shimabukuro ldquoSand flies (DipteraPsychodidae Phlebotominae) from central amazonia and fournew records for the Amazonas State Brazilrdquo Revista Brasileirade Entomologia vol 56 no 2 pp 220ndash227 2012
[36] J V S Ferreira T V Santos E M Santos and I S GorayebldquoPhlebotomine sand flies (Diptera Psychodidae) in forestfragments of Belem metropolitan area Para State Brazil withconsiderations on vectors of American cutaneous leishmaniasisagentsrdquo Revista Pan-Amazonica de Saude vol 5 no 2 pp 29ndash35 2014
[37] B M Carvalho E F Rangel P D Ready and M M ValeldquoEcological niche modelling predicts southward expansion ofLutzomyia (Nyssomyia) flaviscutellata (Diptera PsychodidaePhlebotominae) vector of Leishmania (Leishmania) amazonen-sis in South America under climate changerdquo PLoS ONE vol 10no 11 Article ID e0143282 2015
[38] F Fouque P Gaborit J Issaly et al ldquoPhlebotomine sand flies(Diptera Psychodidae) associatedwith changing patterns in thetransmission of the human cutaneous leishmaniasis in FrenchGuianardquoMemorias do Instituto Oswaldo Cruz vol 102 no 1 pp35ndash40 2007
[39] R Killick-Kendrick ldquoSome epidemiological consequences ofthe evolutionary fit between Leishmaniae and their phle-botomine vectorsrdquo Bulletin de la Societe de Pathologie Exotiqueet de ses Filiales vol 78 no 5 pp 747ndash755 1985
[40] F X Pajot F Le Pont BGentile andR Besnard ldquoEpidemiologyof leishmaniasis in French Guianardquo Transactions of the RoyalSociety of Tropical Medicine and Hygiene vol 76 no 1 pp 112ndash113 1982
[41] H A Christensen J R Arias A M De Vasquez and R ADe Freitas ldquoHosts of sandfly vectors of Leishmania braziliensisguyanensis in the central Amazon of Brazilrdquo The AmericanJournal of Tropical Medicine and Hygiene vol 31 no 2 pp 239ndash242 1982
[42] L C R Nery E S Lorosa andAM R Franco ldquoFeeding prefer-ence of the sand flies Lutzomyia umbratilis and L spathotrichia(Diptera Psychodidae Phlebotominae) in an urban forest patchin the City ofManaus Amazonas BrazilrdquoMemorias do InstitutoOswaldo Cruz vol 99 no 6 pp 571ndash574 2004
[43] PD Ready R Lainson J J Shaw andAA Souza ldquoDNAprobesfor distinguishing Psychodopygus wellcomei from Psychodopy-gus complexus (Diptera Psychodidae)rdquo Memorias do InstitutoOswaldo Cruz vol 86 no 1 pp 41ndash49 1991
[44] P D Ready H Fraiha R P Lane J R Arias and F X PajotldquoOn distinguishing the female of Psychodopygus wellcomei avector of mucocutaneous leishmaniasis from other squamiven-tris series females I Characterization of Ps squamiventrissquamiventris and Ps s maripaensis stat nov (Diptera Psycho-didae)rdquo Annals of Tropical Medicine and Parasitology vol 76no 2 pp 201ndash214 1982
[45] A D Kent T V dos Santos A Gangadin A Samjhawan DR A Mans and H D F H Schallig ldquoStudies on the sandfly fauna (Diptera Psychodidae) in high-transmission areas ofcutaneous leishmaniasis in the Republic of Surinamerdquo Parasitesand Vectors vol 6 no 1 article 318 2013
[46] P D Ready R Lainson and J J Shaw ldquoHabitat and seasonalityof Psychodopygus wellcomei help incriminate it as a vector ofLeishmania braziliensis in Amazonia and Northeast BrazilrdquoTransactions of the Royal Society of Tropical Medicine andHygiene vol 78 no 4 pp 543ndash544 1984
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

12 Journal of Tropical Medicine
[26] J R Arias M A Miles R D Naiff et al ldquoFlagellate infectionsof Brazilian sand flies (Diptera Psychodidae) isolation invitro and biochemical identification of Endotrypanum andLeishmaniardquo The American Journal of Tropical Medicine andHygiene vol 34 no 6 pp 1098ndash1108 1985
[27] A A Souza T V Santos Y L Jennings et al ldquoNaturalLeishmania(Viannia) spp infections in phlebotomine sand flies(Diptera Psychodidae) from the Brazilian Amazon regionreveal new putative transmission cycles of American cutaneousleishmaniasisrdquo Parasite vol 23 article 22 2016
[28] F T Silveira A A Souza R Lainson J J Shaw R R Bragaand E A Ishikawa ldquoCutaneous leishmaniasis in the AmazonRegion natural infection of the sand fly Lutzomyia ubiquitalis(Psychodidae Phlebotominae) by Leishmania (Viannia) lain-soni in Para state BrazilrdquoMemorias do Instituto Oswaldo Cruzvol 86 no 1 pp 127ndash130 1991
[29] LH S Gil S A Basano A A Souza et al ldquoRecent observationson the sand fly (Diptera Psychodidae) fauna of the state ofRondonia Western Amazonia Brazil the importance of Psych-dopygus davisi as a vector of zoonotic cutaneous leishmaniasisrdquoMemorias do Instituto Oswaldo Cruz vol 98 no 6 pp 751ndash7552003
[30] A A A Souza F T Silveira R Lainson et al ldquoThe Phlebotom-inae fauna of Serra dos Carajas Para Brazil and its possibleimplication in the transmission of American tegumentaryleishmaniasisrdquo Revista Pan-Amazonica de Saude vol 1 no 1pp 45ndash51 2010
[31] R Killick-Kendrick ldquoPhlebotomine vectors of the leishmani-ases a reviewrdquoMedical and Veterinary Entomology vol 4 no 1pp 1ndash24 1990
[32] M E C Dorval G Cristaldo H C da Rocha et al ldquoPhle-botomine fauna (Diptera Psychodidae) of an American cuta-neous leishmaniasis endemic area in the state of Mato Grossodo Sul Brazilrdquo Memorias do Instituto Oswaldo Cruz vol 104no 5 pp 695ndash702 2009
[33] V C F V Guimaraes P L Costa F J da Silva et al ldquoMoleculardetection of Leishmania in phlebotomine sand flies in a cuta-neous and visceral leishmaniasis endemic area in northeasternBrazilrdquo Revista do Instituto de Medicina Tropical de Sao Paulovol 56 no 4 pp 357ndash360 2014
[34] R S Lana E M Michalsky C L Fortes-Dias et al ldquoPhle-botomine sand fly fauna and Leishmania infection in thevicinity of the Serra do Cipo National Park a natural Brazilianheritage siterdquo BioMed Research International vol 2015 ArticleID 385493 9 pages 2015
[35] V R Alves R A de Freitas F L Santos A F J de OliveiraT V Barrett and P H F Shimabukuro ldquoSand flies (DipteraPsychodidae Phlebotominae) from central amazonia and fournew records for the Amazonas State Brazilrdquo Revista Brasileirade Entomologia vol 56 no 2 pp 220ndash227 2012
[36] J V S Ferreira T V Santos E M Santos and I S GorayebldquoPhlebotomine sand flies (Diptera Psychodidae) in forestfragments of Belem metropolitan area Para State Brazil withconsiderations on vectors of American cutaneous leishmaniasisagentsrdquo Revista Pan-Amazonica de Saude vol 5 no 2 pp 29ndash35 2014
[37] B M Carvalho E F Rangel P D Ready and M M ValeldquoEcological niche modelling predicts southward expansion ofLutzomyia (Nyssomyia) flaviscutellata (Diptera PsychodidaePhlebotominae) vector of Leishmania (Leishmania) amazonen-sis in South America under climate changerdquo PLoS ONE vol 10no 11 Article ID e0143282 2015
[38] F Fouque P Gaborit J Issaly et al ldquoPhlebotomine sand flies(Diptera Psychodidae) associatedwith changing patterns in thetransmission of the human cutaneous leishmaniasis in FrenchGuianardquoMemorias do Instituto Oswaldo Cruz vol 102 no 1 pp35ndash40 2007
[39] R Killick-Kendrick ldquoSome epidemiological consequences ofthe evolutionary fit between Leishmaniae and their phle-botomine vectorsrdquo Bulletin de la Societe de Pathologie Exotiqueet de ses Filiales vol 78 no 5 pp 747ndash755 1985
[40] F X Pajot F Le Pont BGentile andR Besnard ldquoEpidemiologyof leishmaniasis in French Guianardquo Transactions of the RoyalSociety of Tropical Medicine and Hygiene vol 76 no 1 pp 112ndash113 1982
[41] H A Christensen J R Arias A M De Vasquez and R ADe Freitas ldquoHosts of sandfly vectors of Leishmania braziliensisguyanensis in the central Amazon of Brazilrdquo The AmericanJournal of Tropical Medicine and Hygiene vol 31 no 2 pp 239ndash242 1982
[42] L C R Nery E S Lorosa andAM R Franco ldquoFeeding prefer-ence of the sand flies Lutzomyia umbratilis and L spathotrichia(Diptera Psychodidae Phlebotominae) in an urban forest patchin the City ofManaus Amazonas BrazilrdquoMemorias do InstitutoOswaldo Cruz vol 99 no 6 pp 571ndash574 2004
[43] PD Ready R Lainson J J Shaw andAA Souza ldquoDNAprobesfor distinguishing Psychodopygus wellcomei from Psychodopy-gus complexus (Diptera Psychodidae)rdquo Memorias do InstitutoOswaldo Cruz vol 86 no 1 pp 41ndash49 1991
[44] P D Ready H Fraiha R P Lane J R Arias and F X PajotldquoOn distinguishing the female of Psychodopygus wellcomei avector of mucocutaneous leishmaniasis from other squamiven-tris series females I Characterization of Ps squamiventrissquamiventris and Ps s maripaensis stat nov (Diptera Psycho-didae)rdquo Annals of Tropical Medicine and Parasitology vol 76no 2 pp 201ndash214 1982
[45] A D Kent T V dos Santos A Gangadin A Samjhawan DR A Mans and H D F H Schallig ldquoStudies on the sandfly fauna (Diptera Psychodidae) in high-transmission areas ofcutaneous leishmaniasis in the Republic of Surinamerdquo Parasitesand Vectors vol 6 no 1 article 318 2013
[46] P D Ready R Lainson and J J Shaw ldquoHabitat and seasonalityof Psychodopygus wellcomei help incriminate it as a vector ofLeishmania braziliensis in Amazonia and Northeast BrazilrdquoTransactions of the Royal Society of Tropical Medicine andHygiene vol 78 no 4 pp 543ndash544 1984
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom