revista antipoda no. 13
DESCRIPTION
Revista Antipoda No. 13TRANSCRIPT




ANTIPODAJULIO - DICIEMBRE 2011
REVISTA DEL DEPARTAMENTO DE ANTROPOLOGÍA DE LA FACULTAD DE CIENCIAS SOCIALES UNIVERSIDAD DE LOS ANDES
BOGOTÁ, COLOMBIA
ANTROPOLOGÍA BIOLÓGICA Y ZOOARQUEOLOGÍA
DESDIBUJANDO BARRERAS DISCIPLINARES

.U N I V E R S I D A D D E L O S A N D E SPablo Navas Sanz de Santamaría
REC TOR
Hugo Fazio VengoaDEC ANO
FACULTAD DE C IENC IAS SOCIALES
Carlos Alberto UribeDIREC TOR
DEPARTAMENTO DE ANTROPOLOGÍA
.C O M I T É E D I T O R I A L
Alcida Rita Ramos, Ph.D.Uni v er s idad de Br a s i l i a , Br a s i l
a l c ida . r amos@uo l . com.b r
Carlos Alberto Uribe, Ph.D.Uni v er s idad de los Ande s , Colomb i a
cu r ibe@uniandes .edu .co
Jean Jackson, Ph.D.Massachuse t t s Ins t i tu t e of Technology, M IT
E s tados Un idos j j a c k son@mi t .edu
María Clemencia Ramírez, Ph.D.I n s t i tu to Colomb i ano de Antropolog í a
e H i s tor i a , IC ANH , Colomb i a c lema15@yahoo .com
Margarita Serje, Dra.Uni v er s idad de los Ande s , Colomb i a
mser je@un iandes .edu .co
Myriam Jimeno Santoyo, Dra.Uni v er s idad Nac iona l de Colomb i a
mj imenos@una l .edu .co
Roberto Pineda Camacho, Dr.Uni v er s idad Nac iona l de Colomb i a
r p inedac@una l .edu .co
Roberto Suárez, Dr.Uni v er s idad de los Ande s , Colomb i a
r sua rez@uniandes .edu .co
Santiago Giraldo, Ph.D.Globa l Her i tage Fund, Colomb i a
y E s tados Un idos sg i r a ldo@g loba lhe r i t age fund .o rg
.C O M I T É C I E N T Í F I C OBeatriz Castro, Ph.D.
Uni v er s idad del Va l l e , Colomb i a Cris Shore, Ph.D.
The Un i v er s i t y of Auck l and, Nue va Zel andaElizabeth Fitting, Ph.D.
Dalhous i e Un i v er s i t y, C anadáGerardo Otero, Ph.D.
S imon Fr a ser Un i v er s i t y, C anadáLiu Xin, Ph.D.
Univer s i t y of C al i forn i a , Berkele y. E s tados UnidosMichel Taussig, Ph.D.
Columb i a Un i v er s i t y, E s tados Un idosSergio Eduardo Visacovsky, Ph.D.
I n s t i tu to de De s arrol lo Económico y Soc i a l- IDES , Argent ina
Stephen Hugh-Jones, Ph.D.C ambr idge Un i v er s i t y, R e ino Un ido
Zandra Pedraza, Dr. Phil.Uni v er s idad de los Ande s , Colomb i a
.E D I T O R E S I N V I T A D O SElizabeth Ramos Roca
Uni v er s idad de los Ande s , Colomb i aLuis Borrero
Imh ic ihu - con ice t y Un i v er s idad de Buenos A ir e s , Argent ina
.E Q U I P O A N T Í P O D A
Direc tor a Claudia Steiner Sampedro
Uni v er s idad de los Ande s , Colomb i a mste ine r@un iandes .edu .co
Ed i tor a Gener a l Nidia Vargas Medina
Uni v er s idad de los Ande s , Colomb i a nva rgas@uniandes .edu .co
A s i s t ent e Ed i tor i a l Marisela Quiroz Díaz
Uni v er s idad de los Ande s , Colomb i a mar i s - qu@uniandes .edu .co
.Diseño or ig ina l
Diego Amaral CeballosEd ic ión , a rmada e l ec trón ic a
.Puntoaparte EditoresImpre s ión
Editorial KimpresCorrecc ión de e s t i lo
Guillermo DíezFotogr af í a s
Margarita Monsalve y Juan Antonio Monsalve
fo tomonsa l ve@gmai l . com
.V E N T A S Y S U S C R I P C I O N E S
Librería Uniandes [email protected]
La librería de la Uwww.lalibreriadelau.com
Siglo del Hombre Editoresw w w.s ig lode lhombre .com
C A N J E SSistema de Bibliotecas Uniandes
sbadqu i s i c iones@uniandes .edu .co
.P R E C I O P O R E J E M P L A R :
$24.000 / US$15.00 Ti r a d a : 3 0 0 e j e m p l a r e s • 17 x 24 c m • 2 9 2 p p .
Antípoda. Revista de Antropología y Arqueolo-gía Nº 13 se terminó de imprimir en el mes de diciembre de 2011.
Las opiniones e ideas aquí consignadas son de responsabilidad exclusiva de los autores y no nece-sariamente reflejan la opinión del Departamento de Antropología de la Universidad de los Andes.
El material de esta revista puede ser repro-ducido sin autorización para uso personal o en el aula de clase, siempre y cuando se mencionen como fuente el artículo y su autor y a Antípo-da. Revista de Antropología y Arqueología del Departamento de Antropología de la Universidad de los Andes. Para reproducciones con cualquier otro fin es necesario solicitar la autorización del Comité Editorial de la revista.
ANTIPODARE V IS TA D E A N T RO P O LO GÍ A Y A RQUEO LO GÍ A
N º 13 , JUL IO - D IC I EMBRE 2011 I SSN 19 0 0 -5 4 07 · h t t p : / /an t i p o da .un iande s .e du .co
.PUBLICACIÓN SEMESTRAL DEL DEPARTAMENTO DE ANTROPOLOGÍA, FACULTAD DE CIENCIAS SOCIALES
UNIVERSIDAD DE LOS ANDESDi recc ión pos t a l : C a r re ra 1 N º 18A -10 · Ed i f i c io F ranco , P i so 5 · Bogo t á D. C . , Co lombia
Te lé fono: 57.1.339.4949, E x t . 3483 · Te le fa x : 57.1.332.4510 Pág ina web: h t t p: //an t ropo log ia .un iandes .edu .co

Í N D I C E S
• CABAbstracts(www.cabi.org,EstadosUnidos),2011• SciELO-ScientificElectronicLibraryOnline(Colombia),2010• LatAm–EstudiosLatrinoamericanos(InternationalInformationServices,EstadosUnidos),2009• HLAS-HandbookofLatinAmericanStudies(LibraryodCongress,EstadosUnidos),2009• LATINDEX-SistemaRegionaldeInformaciónenLíneaparaRevistasCientíficasdeAméricaLatina,elCaribe,
EspañayPortugal,2008• HAPI-HispanicAmericanPeriodicalsIndex(UCLA-LatinAmericanInstitute,EstadosUnidos),2008• IBSS-InternationalBibliographyoftheSocialScience(Proquest,EstadosUnidos),2008• PUBLINDEXCategoríaA2-ÍndiceNacionaldePublicaciones(Colciencias,Colombia),2008• SociologicalAbstractsyLinguisticsandLanguageBehaviorAbstracts(CSA-CambridgeScientificAbstracts,
Proquest,EstadosUnidos),2008• DOAJ-DirectoryofOpenAcessJournals(LundUniversityLibraries,Suecia),2007• CREDI-Centrode recursosdocumentales e informáticos (OEI-Organizaciónde Estados Iberoamericanos),
2008• CLASE-CitaLatinoamericanasdeCienciasSocialesyHumanidades(UNAM,México),2007• CIBERA-BibliotecaVirtualIberoamericana(GermanInstituteofGlobalandAreaStudies,Alemania),2007• RedLyC-ReddeRevistasCientíficasdeAméricaLatinayelCaribe,EspañayPortugal(CLACSO,UAEM,Méxi-
co),2007• InformeAcadémico,AcadémicoOnefile(GaeCengageLearning,EstadosUnidos),2007• GALE-CENGAGELEARNING,2007• DIALNET-DifusióndeAlertasenlaRed(UniversidaddelaRioja,España),2007• EPSmartlinkFulltext,FuenteAcadémica,CurrentAbstract,TOCPremier,AcadémicaResearchComplete(EBS-
COInformationServices,EstadosUnidos),2005• Ulrich’sPeriodicalDirectory(Proquest,EstadosUnidos),2005• PRISMA–Publicacionesyrevistassocialeshamanísticas(Proquest,EstadosUnidos),2005• AnthropologicalIndexOnline-RoyalAnthropologicalInstitute(ReinoUnido),2005• OCENET(EditorialOcéanoEspaña),2005• (EBSCO InformationServices,EstadosUnidos)EPSmartlinkFulltext,FuenteAcadémica,CurrentAbstract,
TOCPremier,AcadémicaSearchComplete,2005
P O R T A L E S W E B http://www.lablaa.org/listado_revistas.htm(BibliotecaLuisÁngelArango,Colombia)http://www.portalquorum.org(QuórumPortaldeRevistas,España)http://sala.clacso.org.ar/biblioteca/Members/lenlaces(ReddeBibliotecasVirtualesdeCLACSO,Argentina)

Antípoda. Revista de Antropología y Arqueología del Departamento de Antro-pología (Facultad de Ciencias Sociales) de la Universidad de los Andes (Bogotá, Colombia), llega semestralmente a sus lectores desde el año 2005.
Antípoda conforma un foro abierto y plural en donde se publican artículos y trabajos que permitan adelantar discusiones en la frontera del conocimiento antropológico y de otras disciplinas afines de las ciencias sociales. El criterio para la publicación de trabajos es el de su calidad y pertinencia intelectual, así como su contribución en la discusión y el debate en la comunidad académica nacional e internacional. Los responsables editoriales de Antípoda garantizan una evaluación seria y profesional de todos los materiales sometidos a su con-sideración por parte de pares de reconocida solvencia intelectual, académica y ética. Antípoda tiene un enfoque regional latinoamericano, con un interés espe-cial en difundir y compartir las experiencias del trabajo antropológico tanto de Colombia como de las antropologías del mundo.
A partir del nombre de Antípoda como una metáfora de la alteridad, la revista pretende presentar las diversas visiones de los temas sobre los cuales trata la disciplina. Las secciones se organizan desde las siguientes alegorías espaciales:
Meridianos: en esta sección se señala la orientación del número. Aquí se publican artículos originales, resultados de investigaciones que están directamente relacionadas con el tema central. Igualmente se presentan trabajos de investigado-res reconocidos en la disciplina, los cuales hacen referencia al tema central y que en algunos casos se traducen para hacerlos accesibles al público hispanoparlante.
Paralelos: tienen lugar en esta sección artículos que están relacionados con el tema central del número, no necesariamente de una manera directa pero sí a través de aportes tanto teóricos como empíricos.
Panorámicas: una sección amplia y abierta que recoge escritos con temas de actualidad y que no necesariamente se relacionan directamente con el tema central del número.
Reseñas: presenta reseñas bibliográficas de nuevas publicaciones u otros trabajos de interés para la revista y sus lectores.
Documentos: sección que ofrece a los lectores trabajos inéditos que pue-den ser escritos, fotografías u otros documentos de carácter histórico. .
ANTIPODA

Ana María Boada, Universit yofPit tsburgh,EstadosUnidos
Andrés Link Ospina, UniversidaddelosAndes,Colombia
Angélica Monserrat Tivoli, CentroAustraldeInvestigacionesCientíf icas–ConsejoNacionaldeInvestigacionesCientíf icasyTécnicasCADIC-CONICET,Argentina
Atilio Francisco Zangrado, CentroAustraldeInvestigacionesCientíf icas–ConsejoNacionaldeInvestigacionesCientíf icasyTécnicasCADIC-CONICET,Argentina
Carl Langebaek, UniversidaddelosAndes,Colombia
Carlos Alberto Uribe,UniversidaddelosAndes,Colombia
Carolyn Freiwald,Universit yofWisconsin–Madison,EstadosUnidos
Eduardo Corona M.,Insti tutoNacionaldeAntropologíaeHistoriaINAH,México
Elizabeth Ramos Roca,UniversidaddelosAndes.Bogotá,Colombia
Federico Luis Scartascini, ConsejoNacionaldeInvestigacionesCientíf icasyTécnicasCONICET;InstitutoMultidisciplinariodeHistoriayCienciasHumanasIMHICIHU,Argentina
Francisco J. Prevosti, MuseoArgentinodeCienciasNaturalesBernardinoRivadavia,Argentina
Gabriel Eduardo José López, ConsejoNacionaldeInvestigacionesCientíf icasyTécnicas–CONICET,Argentina
Gustavo Neme, MuseodeHistoriadeSanRafael,Argentina
Heather McKillop, LouisianaStateUniversit y,EstadosUnidos
Helena Groot,UniversidaddelosAndes,Bogotá,Colombia
Joaquín Arroyo-Cabrales, Insti tutoNacionaldeAntropologíaeHistoria,México
Lourdes Márquez Morfín, Insti tutoNacionaldeAntropologíaeHistoria/EscuelaNacionaldeAntropologíaeHistoria,México
Luciano De Santis, UniversidadNacionaldeLaPlata,Argentina
Luis Alberto Borrero, ConsejoNacionaldeInvestigacionesCientíf icasyTécnicasCADIC-CONICET;InstitutoMultidisciplinariodeHistoriayCienciasHumanasIMHICIHU,Argentina
Mariana Eleonor De Nigris, UniversidaddeBuenosAires,Argentina
Mark Rohrig Hubbe, Insti tutodeInvestigacionesArqueológicasyMuseoUniversidadCatólicadelNor te,SanPedrodeAtacama,Chile
Pablo Marcelo Fernández, CentroAustraldeInvestigacionesCientíf icas–ConsejoNacionaldeInvestigacionesCientíf icasyTécnicasCADIC-CONICET,Argentina
Roberto Suárez Montañez, UniversidaddelosAndes,Colombia
Wendy G. Teeter, Universit yofCalifornia-UCLA,EstadosUnidos.
Antípoda. Revista de Antropología y Arqueología agradece la colaboración de las siguientes personas como árbitros de este número
ANTIPODA

A N T Í P O D A N º 1 2 | E N E R O - J U N I O 2 0 1 1 A N T Í P O D A N º 1 1 | J U L I O - D I C I E M B R E 2 0 1 0ANTIPODAN o t a E d i t o r i a l . . . . . . . . . . . . . . . . . . . . . . . . . . 1 5P r e s e n t a c i ó n . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 7M e r i d i a n o s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 7
Los aportes de la bioantropología al conocimiento de la variabilidad biológica de los sudamerindiosDescripción y análisis
Franc isco Raúl Carnese · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · 29
Adeptos a la adaptaciónTres propuestas clásicas para la arqueología y una evaluación
Viv ian Scheinsohn · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · 55
Aportación arqueológica al conocimiento del proceso de descomposición del cuerpo humano en posición sentada/flexionada
Karina Gerdau Radonic · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · 75
P a r a l e l o s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9 7La biología de la conservación aplicada a la zooarqueología:la sostenibilidad de la cacería del venado cola blanca, Odocoileus virginianus (Artiodactyla, Cervidae) en Aguazuque
María Fernanda Mart ínez-Polanco · · · · · · · · · · · · · · · · · · · · · · · 99
La arqueofauna de Xcambó, Yucatán, MéxicoChristopher M Götz y Thelma N. S ierra Sosa · · · · · · · · · · · · · · · 1 19
Tafonomía de huesos de avesEstado de la cuestión y perspectivas desde el sur del Neotrópico
I sabel Cruz · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · 147
Primer registro de perro doméstico prehispánico (canis familiaris) entre los grupos cazadores recolectores del humedal de Paraná inferior (Argentina)
Alejandro Acosta , Daniel Loponte y César García Esponda · · · · · 175
Arqueología subacuática y tafonomía Recientes avances en el estudio de sitios finipleistocénicos sumergidos en la costa pacífica de Chile central.
I sabel Carta jena F. , Patr ic io López M. , Diego Carabias , Carla Morales y Gabriel Vargas · · · · · · · · · · · · · · · · · · · · · · · · 201
Í N D I C E

L A B I O L O G Í A D E L A C O N S E R V A C I Ó N | M A R Í A F E R N A N D A M A R T Í N E Z - P O L A N C O A N T Í P O D A N º 1 2 | E N E R O - J U N I O 2 0 1 1 A N T Í P O D A N º 1 1 | J U L I O - D I C I E M B R E 2 0 1 0ANTIPODAÍ N D I C EP a n o r á m i c a s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 2 7
Aproximaciones y escalas de análisis en la zooarqueología y tafonomía sudamericanas. Algunas reflexiones sobre su estado actual y perspectivas para su desarrollo
mariana Mondini y A . Sebast ián Muñoz · · · · · · · · · · · · · · · · · · · 229
D o c u m e n t o s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 5 3

ANTIPODAE d i t o r i a l N o t e . . . . . . . . . . . . . . . . . . . . . . . . . . 1 5P r e s e n t a t i o n . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 7M e r i d i a n s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 7
Bioanthropology contributions to the knowledge of biological variability among south american indians:Analysis and description
Franc isco Raúl Carnese · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · 29
Believing in adaptation:Three classical proposals for archeology and one evaluation
Viv ian Scheinsohn · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · 55
Archaeological insights into the disarticulation pattern of a human skeleton in a sitting/squatting position
Karina Gerdau Radonic · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · 75
P a r a l l e l s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9 7The biology of conservation applied to zooarcheology:Hunting sustainability of the white tail deer odocoileus virginianus (artiodactyla, cervidae) in Aguazuque
María Fernanda Mart ínez-Polanco · · · · · · · · · · · · · · · · · · · · · · · 99
Archeofauna of Xcambo, Yucatan, MexicoChristopher M Götz & Thelma N. S ierra Sosa · · · · · · · · · · · · · · · 1 19
Bird bones taphonomy.State of the problem and perspectives from the southern Neotropics
I sabel Cruz · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · 147
First registry of the pre-hispanic domestic dog (canis familiaris) among gatherer-hunter groups in lower Parana (Argentina)
Alejandro Acosta , Daniel Loponte & César García Esponda · · · · · 175
Sub-aquatic archeology and taphonomy recent advances in the study of submerged phinipleistocenics in the pacific coast of central Chile
I sabel Carta jena F. , Patr ic io López M. , Diego Carabias , Carla Morales & Gabriel Vargas · · · · · · · · · · · · · · · · · · · · · · · · 201
C O N T E N T S

ANTIPODAC O N T E N T SP a n o r a m i c s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 2 7
Approaches and scale analyses in south american zooarcheology and taphonomy: reflections regarding its current state and perspectives for its development
Mariana Mondini & A . Sebast ián Muñoz · · · · · · · · · · · · · · · · · · · 229
D o c u m e n t s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 5 3

A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1 A N T Í P O D A N º 1 2 | J U L I O - D I C I E M B R E 2 0 1 1
N o t a E d i t o r i a l . . . . . . . . . . . . . . . . . . . . . . . . . . 1 5A p r e s e n t a ç ã o . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 7M e r i d i a n o s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 7
Os aportes da bioantropologia ao conhecimento da variabilidade biologica dos sul-amerindios:descrição e análise
Franc isco Raúl Carnese · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · 29
Adeptos à adaptação:Três propostas clássicas para a arqueología e uma avaliação
Viv ian Scheinsohn · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · 55
Aportação arqueológica ao conhecimento do processo de decomposição do corpo humano em posição sentada/flexionada
Karina Gerdau Radonic · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · 75
P a r a l e l o s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9 7A biologia da conservação aplicada à zooarqueologia:A sustentabilidade da caçada do veado cauda branca odocoileus virginianus (artiodactyla, cervidae) em Aguazuque
María Fernanda Mart ínez-Polanco · · · · · · · · · · · · · · · · · · · · · · · 99
A arqueofauna de Xcambó, Iucatã, MéxicoChristopher M Götz , Thelma N. S ierra Sosa · · · · · · · · · · · · · · · · 1 19
Tafonomia de ossos de aves. estado da questão e perspectivas desde o sul do Neotrópico
I sabel Cruz · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · 147
Primeiro registro do cão domestico pré-hispânico (canis familiaris) entre os grupos caçadores recolhedores da zona úmida de Paraná inferior (Argentina)
Alejandro Acosta , Daniel Loponte , César García Esponda · · · · · · 175
Arqueologia subaquatica e tafonomiaRecentes avanços no estudo de lugares finipleistocênicos submersos na costa pacífico do Chile central
I sabel Carta jena F. , Patr ic io López M. , Diego Carabias , Carla Morales , Gabriel Vargas · · · · · · · · · · · · · · · · · · · · · · · · · 201
A N T Í P O D A N º 1 2 | E N E R O - J U N I O 2 0 1 1 A N T Í P O D A N º 1 1 | J U L I O - D I C I E M B R E 2 0 1 0ANTIPODAÍ N D I C E

A N T Í P O D A N º 1 2 | J U L I O - D I C I E M B R E 2 0 1 1 A N T Í P O D A N º 1 2 | E N E R O - J U N I O 2 0 1 1 A N T Í P O D A N º 1 1 | J U L I O - D I C I E M B R E 2 0 1 0ANTIPODAÍ N D I C EP a n o r â m i c a s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 2 7
Abordagens e escalas de análise na zooarqueologia e tafonomia sul-americanas:Algumas reflexões sobre seu estado atual e perspectivas para seu desenvolvimento
Mariana Mondini , A . Sebast ián Muñoz · · · · · · · · · · · · · · · · · · · · 229
D o c u m e n t o s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 5 3


1 5
N o t a E d i t o r i a l
A n t r o p o l o g í a b i o l ó g i c a y z o o a r q u e o l o g í a . D e s d i b u j a n d o b a r r e r a s d i s c i p l i n a r e s
En s u n ú m e r o 1 3 , Antípoda se propone ampliar la discusión sobre los diferentes campos de nuestra disciplina. El reconocimiento de los aportes de investigaciones no directamente relacionadas con la antropo-logía social y cultural, pero que de manera permanente la influyen, es el obje-tivo de este número. Entre estos aportes se destacan aquellos hechos por la antropología biológica y la zooarqueología, los cuales, tal y como lo señalan nuestros editores invitados, Elizabeth Ramos Roca y Luis Borrero, en su sig-nificativa presentación, al involucrar aspectos tanto sociales como biológicos, permiten avanzar en la comprensión de las sociedades humanas.
A partir de la realización del “Encuentro de Antropología Biológica en Colombia” y del “Primer Encuentro Latinoamericano de Zooarqueología”, lle-vado a cabo durante el 13 Congreso Colombiano de Antropología en Bogotá en 2009, se hizo notoria la importancia de establecer diálogos transdisciplina-rios que permitieran ampliar las discusiones entre los diferentes campos de la antropología. Los editores invitados hicieron una cuidadosa labor de selección de las ponencias que se presentaron, y que hoy publicamos en este número en forma de artículos, con la intención de iniciar este tipo de diálogo, necesario para la disciplina antropológica. Para Elizabeth Ramos y Luis Borrero, nuestros agradecimientos por su labor. En el presente número, los artículos sobre antro-pología biológica aparecen en la sección Meridianos, y los relacionados con zooarqueología se presentan en la sección Paralelos.
Para esta edición de Antípoda que pretende establecer un diálogo entre diversos campos de la antropología, el trabajo de los artistas Margarita Mon-salve y Juan Antonio Monsalve no podía ser más adecuado para ilustrar el tema. Su obra, que también refleja un tipo de mirada transdisciplinaria, al transformar imágenes fotográficas convencionales para crear imágenes que se
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.15-16

1 6
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
esconden una tras otra, nos lleva, como espectadores, a establecer una mirada distinta sobre la representación fotográfica. Una mirada que no deja de mara-villarse ante el surgimiento casi mágico de las nuevas imágenes que aparecen en diferentes dimensiones. A ellos, nuestros agradecimientos por permitirnos utilizar las fotograf ías que acompañan este número. .
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.15-16

1 7
P r e s e n t a c i ó n
D I S C U S I O N E S C O M U N E S Y R E L E VA N C I A D E L O S D I Á L O G O S
T R A N S D I S C I P L I N A R I O S E N A N T R O P O L O G Í A : A P O R T E S D E S D E
L A A N T R O P O L O G Í A B I O L Ó G I C A Y L A A R Q U E O Z O O L O G Í A
e l i z a b e T h r a m o s r o c a *
[email protected] de los Andes, Bogotá, Colombia
L u i s A L b e r t o b o r r e r o * *
[email protected] y Universidad de Buenos Aires, ArgentinaLas aproximaciones transdisciplinarias son,
sin lugar a dudas, uno de los terrenos más fértiles y uno de los mayores desaf íos de la investigación en el presente siglo, ya que, más que un trabajo “paralelo” o “simultáneo”, lo que implican es la identificación de fundamentos teóricos y metodológicos que puedan resultar verdaderamente transversales y que, en consecuencia, permitan integrar las discusiones de los distintos campos del conocimiento alrededor de propuestas teóricamente sólidas y metodológica-mente coherentes.
Por ya largo tiempo se ha debatido sobre el impacto que para la recons-trucción y el entendimiento de los procesos de adaptación biocultural humana ha tenido la imposibilidad de integrar de manera coherente el conocimiento que se produce desde las ciencias sociales y naturales (Schultz, 2009; Borofsky, 2002; Goodman y Leatherman, 1998; Calcagno, 2003; Puccirelli, 1994)1. En el campo de la Antropología, esto se evidencia en la cada vez mayor separación entre los intereses disciplinares de los antropólogos y en el escaso interés por
* Ph.D.enArqueología-UniversidaddePittsburgh,EstadosUnidos.**Doctor,UniversidaddeBuenosAires,Argentina.1 VéasetambiénSpecial Issue:BiologicalAnthropology:HistoricalPerspectivesonCurrentIssues,Disciplinary
Connections, and FutureDirections, JamesM. Calcagno, editor invitado,American Anthropologist 105 (1),marzode2003.
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.17-25

1 8
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
generar discusiones que integren los resultados de investigaciones de diferentes campos, cuestión que se ve reflejada en una producción académica fragmen-tada, en la desarticulación de los contenidos en los currículos educativos y en el contenido de los textos antropológicos (White et al., 2009), y en la falta de cla-ridad que hasta cierto punto persiste sobre los aportes que las investigaciones de los antropólogos biológicos y los arqueólogos pueden hacer a las discusiones de la Antropología social y cultural, especialmente en lo que atañe al estudio de las problemáticas contemporáneas2. Estas situaciones, creemos, vívidamente reflejan unas aristas del debate sobre el “holismo” como carácter esencial de la Antropología (Calcagno, 2003).
En este sentido, vemos cómo, en la medida en que avanzamos en el enten-dimiento de los niveles de complejidad de la especie humana, biológica y cultu-ralmente hablando, más evidente se hace la necesidad de ahondar en la discu-sión sobre la importancia de generar marcos conceptuales integradores entre las ciencias sociales y biológicas, y en el caso particular de la Antropología, entre sus distintos campos, los cuales, como es de suponer, involucran aspectos sociales y biológicos. Es claro que no ha sido tarea fácil producir arenas de dis-cusión comunes sobre la coevolución biológica y cultural. Recientemente, se ha señalado, por ejemplo, que uno de los marcos teóricos más promisorios en este sentido es el de la teoría evolutiva, ya que se argumenta que este marco no sólo permite la integración entre las ciencias sociales y biológicas sino, en general, entre las distintas ciencias humanas (Richerdson y Boyd, 2005), ofreciendo así posibilidades alentadoras para la gestación de teorías sintéticas sobre el com-portamiento humano (Martínez y Lanata, 2002; O’Brien y Lyman, 2002; Lyman y O’Brien, 2007; Muscio y López, 2009; Borrero, 2009; López y Cardillo, 2009).
En el contexto de la Antropología y la Arqueología contemporánea, dos campos que ilustran de manera apropiada esta situación son el de la Arqueo-zoología3 y el de la Antropología Biológica, ambos con historias particulares de larga trayectoria y ampliamente reconocidos en el presente, y entre los cuales, a pesar de las múltiples posibilidades de diálogo, son escasos los intentos por construir marcos de discusión donde se integren las investigaciones de ambos campos bajo una perspectiva transdisciplinaria.
En este sentido, y tomando como signo del desarrollo de cada una de estas dos disciplinas la diversidad de “áreas” o “subacampos” que cada una de éstas acredita hoy en día, vemos cómo la Arqueozoología –entendida de manera general como el estudio de la fauna de contextos arqueológicos– involucra
2 Comounejemplodetrabajosrecientesqueaportanenestesentido,véaseVéran(2011).3 UsadoaquícomosinónimodeZooarqueología.Paramayorclaridadentornoalusodelosdostérminos,véase
ReitzyWing(1999),capítulo1.
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.17-25

1 9
P R E S E N T A C I Ó N | E L I Z A B E T H R A M O S R O C A Y L U I S A L B E R T O B O R R E R O
diversos objetivos a su vez ligados a la multiplicidad de énfasis y enfoques con los que podemos aproximarnos al estudio de la fauna, como son la reconstruc-ción medioambiental, la dieta, la subsistencia, la biología de la conservación, los usos extradietarios y el cambio sociocultural, etcétera.
Desde la óptica de la Antropología Biológica –entendida como el estu-dio dentro de un marco evolutivo de la variación biológica de las poblaciones humanas a lo largo de toda su historia–, los subcampos que se pueden recono-cer son, entre otros, los de la biología de poblaciones humanas, la bioarqueolo-gía y la antropología forense.
A pesar de la aparente especificidad de cada una de estas áreas del cono-cimiento, resulta evidente que los temas alrededor de los cuales se insertan las discusiones más generales son compartidos. Así, identificar y definir “dónde” se plasman las arenas comunes y los marcos teóricos y aproximaciones metodoló-gicas que nos sirvan de referente para establecer discusiones transdisciplinarias es a lo que apunta el presente volumen; se busca contribuir a la tarea de lograr cada vez más y oportunos espacios en los que esta perspectiva, central como la entendemos, logre convertirse en una agenda puntual de la academia latinoa-mericana y, por supuesto, mundial en el campo de la Antropología Biológica y de la Arqueozoología, que a su vez logre trascender y aportar a las discusiones antropológicas más amplias.
Tal fue también el espíritu que guió el interés de organizar, en el marco del 13 Congreso de Antropología en Colombia (Bogotá, septiembre-octubre de 2009), la realización paralela del “Encuentro de Antropología Biológica en Colombia” y del “Primer Encuentro Latinoamericano de Zooarqueología”, eventos dentro de los cuales se presentaron la mayoría de los trabajos que aquí se compilan. Creemos que uno de los mecanismos para lograr estos diálogos comunes a los que nos referimos, es propiciar espacios de discusión y divulga-ción donde confluyan distintas áreas del conocimiento y diversas perspectivas, pero donde se establezcan e identifiquen temas comunes.
En los trabajos que aquí se incluyen se evidencian al menos dos de estos temas donde se percibe la perspectiva transdiciplinar; uno es el de la adaptación humana –ligado, por supuesto, a la teoría evolutiva–, abordado explícita o implí-citamente en todos los trabajos, y el otro, el de la tafonomía. La articulación entre estos dos temas resulta en nuestro concepto evidente, en la medida en que estudiar los fenómenos de adaptación humana en sus múltiples dimensiones requiere, por una parte, la discusión conceptual acerca de “adaptación”, y por otra, estudios puntuales en distintos contextos geográficos y temporales sobre las diversas líneas de evidencia relacionadas con estos procesos; líneas de evidencia que incluyen varios tipos de análisis pertinentes al campo de la Antropología Biológica y Arque-
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.17-25

2 0
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
zooqueología, como la determinación de la dieta, los patrones de subsistencia, el perfil paleodemográfico y paleopatológico, entre otros, y para los cuales el aporte de la tafonomía resulta, por decir lo menos, necesario. La perspectiva tafonómica no sólo es la que permite decidir cuáles son los tipos de investigaciones para las que son adecuados determinados conjuntos de materiales, sino también establecer cuándo existen condiciones de comparabilidad entre distintas muestras.
De acuerdo con la estructura de la revista, en la sección Meridianos con-tamos con tres trabajos que involucran explícita o implícitamente el tema de la adaptación humana y de la teoría de la evolución, en términos bioculturales –el último de ellos también ligado al tema de la tafonomía–, ensayos que nos permiten dimensionar los campos de la Antropología Biológica en una óptica regional como lo es la suramericana, y el de la Zooarqueología en dimensiones que van desde lo teórico-metodológico del concepto básico de “adaptación” hasta las precisiones propias de un análisis tafonómico detallado, en función de prácticas culturales humanas como las mortuorias.
En efecto, el texto de Francisco Raúl Carnese, “Los aportes de la bioan-tropología al conocimiento de la variabilidad biológica de los sudamerindios. Descripción y análisis”, además de trazar el derrotero de los estudios de genética de poblaciones desde el siglo XIX hasta el presente en el contexto sudamericano, destaca los aportes metodológicos y técnicos de la Antropología Biológica, enfa-tizando en la contribución que este tipo de investigaciones ha venido haciendo a las discusiones sobre el origen, la dispersión y la variabilidad biológica de las poblaciones humanas, destacándose, de manera particular, el potencial que para la interpretación de estos fenómenos tienen los estudios de ADN.
Por su parte, Vivian Scheinsohn, en su ensayo “Adeptos a la adapta-ción: tres propuestas clásicas para la arqueología y una evaluación”, presenta un estado del arte sobre las conceptualizaciones vigentes en el pensamiento arqueológico contemporáneo de uno de los conceptos más importantes en la perspectiva evolutiva –el de adaptación–, evaluando su utilización con base en una revisión exhaustiva en distintos campos y corrientes de la Arqueología, revisión que la lleva a concluir que, dentro de la gama de propuestas teóri-cas existentes, hay algunas que parecen adecuarse mejor a la interpretación arqueológica, como es el caso de la “Teoría del Nicho Cultural” y el de la “Teoría de la Herencia Dual”, y en el caso de esta última se requeriría una “decodifica-ción en cuanto registro arqueológico”, lo que para la autora se convierte en uno de nuestros mayores retos.
El texto de Karina Gerdau-Radonic, “Aportación arqueológica al cono-cimiento del proceso de descomposición del cuerpo humano en posición sentada/flexionada”, por su parte, además de documentar sobre el proceso
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.17-25

2 1
de descomposición del cuerpo humano, discute sobre la posibilidad de dis-tinguir entre manipulación intencional y eventos tafonómicos, realzando la importancia de estudiar detalladamente estos contextos con el fin de refinar las estrategias que nos permiten inferir de manera más precisa el comporta-miento humano en sus múltiples dimensiones.
En la sección Paralelos, se incluyen cinco ensayos que compilan resulta-dos de trabajos de investigación sobre temáticas o áreas de estudio particulares, pero que están ligados en todos los casos con el tema de la adaptación humana y la importancia de las investigaciones tafonómicas para abordar el estudio de estos procesos. En este sentido, el texto de María Fernanda Martínez-Polanco, “La biología de la conservación aplicada a la zooarqueología: la sostenibilidad de la cacería del venado cola blanca, Odocoileus virginianus (Artiodactila, Cer-vidae) en Aguazuque”, aporta al debate sobre la aplicación de la investigación zooarqueológica a las discusiones sobre conservación de especies en la actua-lidad, mediante el estudio de los patrones de cacería del venado cola blanca en Aguazuque, sitio de cazadores-recolectores tardíos de la sabana de Bogotá, y centrándose la discusión alrededor de conceptos como sostenibilidad, biología de la conservación e índices de diversidad.
El texto de Christopher M. Götz y Thelma Noemí Sierra Sosa, “La arqueofauna de Xcambó, Yucatán, México”, se aproxima al estudio de los proce-sos de adaptación de las poblaciones prehispánicas de la península de Yucatán, por medio del estudio zooarqueológico de los patrones de aprovechamiento de la fauna en el sitio prehispánico de Xcambó, contribuyendo así a la reconstruc-ción de las trayectorias bioculturales de las sociedades Mayas.
El texto de Isabel Cruz, “Tafonomía de huesos de aves. Estado de la cues-tión y perspectivas desde el sur del Neotrópico”, presenta, por su parte, una discusión basada en una serie de investigaciones en Patagonia, las cuales evi-dencian el impacto de los procesos tafonómicos sobre los restos de aves, que varían en relación con el tamaño corporal, y la morfoanatomía de las mismas, en función del contexto ecológico y ambiental. La autora señala la necesidad de aplicar lo anterior en estudios futuros que, desde la perspectiva tafonómica, estén dirigidos a evaluar la acción de tales procesos sobre restos de vertebrados, discusión muy pertinente sobre la calidad de las reconstrucciones de las dietas pasadas, tanto en términos nutricionales como de prácticas de cacería o pesca.
Por su parte, el artículo de Alejandro Acosta, Daniel Loponte y César Gar-cía Esponda, “Primer registro de perro doméstico prehispánico (Canis fami-liaris) entre los grupos cazadores recolectores del humedal de Paraná inferior (Argentina)”, trata sobre las implicaciones que para la discusión sobre el origen e interacción entre los perros prehispánicos y las poblaciones aborígenes tienen
P R E S E N T A C I Ó N | E L I Z A B E T H R A M O S R O C A Y L U I S A L B E R T O B O R R E R O
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.17-25

2 2
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
los hallazgos en esta zona, señalando que, dado que los resultados de estas inves-tigaciones dejan “más interrogantes que respuestas”, resulta perentorio abordar otras técnicas de investigación, como los son los estudios de ADN, que nos per-mitan documentar con mayor precisión las relaciones filogenéticas y las trayec-torias de esta especie en el continente.
El último texto de esta sección corresponde al ensayo de Isabel Cartajena F., Patricio López M., Diego Carabias, Carla Morales y Gabriel Vargas, “Arqueo-logía subacuática y tafonomía: recientes avances en el estudio de sitios finipleis-tocénicos sumergidos en la costa pacífica de Chile central”, el cual nos ofrece una discusión de los resultados del análisis tafonómico del sitio GNLQ1 –que corresponde a un contexto primario conformado por una serie de restos de fauna continental extinta del Pleistoceno final (familias Camelidae, Cervidae, Artiodactyla, Equidae, Mylodontidae, Canidae y Xenarthra), recuperados bajo las aguas del océano Pacífico–, enfatizando en la representación anatómica y las características superficiales de los materiales, con el fin de conocer las condiciones paleoecológicas y delimitar los factores tafonómicos en contextos terrestres y subacuáticos.
En la tercera sección de la revista, Panorámicas, el trabajo de Mariana Mondini y A. Sebastián Muñoz, titulado “Aproximaciones y escalas de análi-sis en la zooarqueología y tafonomía sudamericanas”, ofrece una panorámica sobre la importancia del vínculo entre la investigación zooarqueológica y tafo-nómica en Sudamérica y los retos del uso de escalas de análisis apropiadas para abordar estos fenómenos. Así, la discusión se organiza alrededor de algunos puntos “sensibles” en los cuales los autores consideraran que debemos centrar nuestra atención en el futuro próximo, como la integración entre la tafonomía, la zooarqueología y la investigación arqueológica, los estudios actualísticos, investigaciones que involucren distintas escalas de análisis y la necesidad de la interlocución continental para abordar estos temas. Para finalizar, señalan la importancia que para el futuro de estas disciplinas tiene el estímulo de una producción teórica, metodológica y epistemológica.
La última sección de la revista, Documentos, incluye el texto de Raúl Vala-dez Azúa y Gilberto Pérez Roldán, “La zooarqueología dentro de la Universidad Nacional Autónoma de México”, un estudio de caso que ilustra la importancia que tiene el trabajo interdisciplinario para la interpretación tanto zooarqueoló-gica como arqueológica en general, Así mismo, en esta sección, y como reflexión final del volumen, tenemos el texto de Luis Borrero, “La función transdiscipli-naria de la arqueozoología en el siglo XXI: restos animales y más allá”, docu-mento que recoge, usando el ejemplo de la Arqueozoología, la importancia y los principales retos del trabajo transdisciplinar, destacando cómo el trabajo
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.17-25

2 3
del arqueozoólogo exige un conocimiento de la variación exhibida, llamando la atención sobre la importancia de innovar y hacer propuestas metodológicas concordantes con las discusiones teóricas en América Latina, particularmente a partir de lo que el autor denomina “tafonomía irrestricta”.
Como editores invitados, queremos concluir esta presentación recono-ciendo a los autores y el equipo coordinador de la revista Antípoda su paciencia y compromiso, con la convicción de que este esfuerzo servirá para hacer reali-dad los diálogos verdaderamente transdisciplinares. .
P R E S E N T A C I Ó N | E L I Z A B E T H R A M O S R O C A Y L U I S A L B E R T O B O R R E R O
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.17-25

2 4
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
R e f e R e n c i a s
Borofsky, Robert 2002.“TheFourSubfields:AnthropologistsasMythmakers”,American Anthropologist104(2),pp.463-480.
Borrero, Luis Alberto 2009.“Arqueologíayevolución:comentariosydigresiones”en Arqueología y evolución. Teoría,
metodología y casos de estudio, eds.GabrielLópezyMarceloCardillo,pp.7-21.BuenosAires,EditorialSB,ColecciónComplejidadHumana.
Calcagno, James M. 2003.“KeepingBiologicalAnthropologyinAnthropology,andAnthropologyinBiology”,American
Anthropologist105(1),pp.6-15.
Goodman, Alan y Thomas Leatherman 1998.“TraversingtheChasmbetweenBiologyandCulture:AnIntroduction”,en Building a New
Biocultural Synthesis: Political-economic Perspectives on Human Biology,eds.AlanGoodmanyThomasLeatherman,pp.3-42.Michigan,UniversityofMichiganPress.
López, Gabriel y Marcelo Cardillo (eds.) 2009.Arqueología y evolución. Teoría, metodología y casos de estudio.BuenosAires,EditorialSB,
ColecciónComplejidadHumana.
Lyman, R. Lee y Michael O’Brien 2007.“Losobjetivosdelaarqueologíaevolucionista:suhistoriayexplicación”,en Clásicos de la teoría
arqueológica contemporánea,trad.LuisA.Orquera;comp.VictoriaD.Horwitz,pp.213-287.BuenosAires,PublicacionesdelaSociedadArgentinadeAntropología.
Martínez, Gustavo A. y José Luis Lanata (eds.) 2002. Perspectivas integradoras entre arqueología y evolución.Olavarría,SerieTeóricaNº1de
INCUAPA–InvestigacionesArqueológicasyPaleontológicasdelCuaternarioPampeano.
Muscio, Hernán Juan y Gabriel Eduardo José López (eds.) 2009.Theoretical and Methodological Issues in Evolutionary Archaeology Toward an Unified
Darwinian Paradigm. ProceedingsoftheXVWorldCongress(Lisboa,4-9deseptiembrede2006).Oxford,Vol.20BARInternationalSeries1915.
O’Brien, Michael J. y R. Lee Lyman 2002. Applying Evolutionary Archaeology: A Systematic Approach.NuevaYork,KluwerAcademic
Publishers.
Pucciarelli, H. M. 1994.“Relacionesconceptualesentreantropologíabiológicayarqueología”,enJornadas de
Arqueología e Interdisciplinas,pp.67-80.ProgramadeEstudiosPrehistóricos.BuenosAires.
Reitz, Elizabeth Jean y Elizabeth S. Wing 1999.Zooarchaeology.Cambridge,CambridgeUniversityPress.
Richerdson, Peter J. y Robert Boyd 2005. Not by Genes Alone. How Culture Transformed Human Evolution.ChicagoyLondres,The
UniversityofChicagoPress.
Schultz, Emily 2009.“ResolvingtheAnti-antievolutionismDilemma:ABriefforRelationalEvolutionaryThinkingin
Anthropology”,American Anthropologist111(2),pp.224-237.
Véran, Jean-François 2011.“OldBones,NewPowers”.Documentodigitaldisponibleen:http://www.jstor.org/
stable/10.1086/662384.s.f.
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.17-25

2 5
White, Jess, Craig D. Tollini, W. Aaron Collie, Meredith B. Strueber, Linda H. Strueber y Jonathan W. Ward
2009.“EvolutionandUniversity-levelAnthropologyTextbooks:The‘MissingLink’?”,Evo Edu Outreach 2,pp.722-737.
P R E S E N T A C I Ó N | E L I Z A B E T H R A M O S R O C A Y L U I S A L B E R T O B O R R E R O
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.17-25

A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1

MeridianosLOS APORTES DE LA BIOANTROPOLOGÍA AL CONOCIMIENTO DE LA VARIABILIDAD BIOLÓGICA DE LOS SUDAMERINDIOS. DESCRIPCIÓN Y ANÁLISIS
F r a n c i s c o r a ú l c a r n e s e 2 9
ADEPTOS A LA ADAPTACIÓN: TRES PROPUESTAS CLÁSICAS PARA LA ARQUEOLOGÍA Y UNA EVALUACIÓN
V i V i a n s c h e i n s o h n 5 5
APORTACIÓN ARQUEOLÓGICA AL CONOCIMIENTO DEL PROCESO DE DESCOMPOSICIÓN DEL CUERPO HUMANO EN POSICIÓN SENTADA/FLEXIONADA
K a r i n a G e r d a u r a d o n i c 7 5

2 8

2 9
LOS A PORT ES DE L A BIOA N TROPOLOGÍ A A L CONOCI M I EN TO DE L A VA R I A BI LI DA D BIOLÓGICA DE LOS SU DA M ER I N DIOS . DESCR IPCIÓN Y A NÁ LISIS
Fr ancisco r aúl carnese* [email protected] de Buenos Aires
R e s u m e n En este trabajo se describe y analiza la información
proporcionada por los estudios bioantropológicos en poblaciones
amerindias sudamericanas. Se evaluaron tres etapas del
desarrollo de la antropología biológica, desde el siglo XIX hasta
el presente, con la finalidad de analizar los aportes técnico-
metodológicos y teóricos predominantes en cada una de ellas.
Para alcanzar esos objetivos, se consideraron, principalmente,
las contribuciones de la genética de poblaciones que abrieron
nuevos rumbos para abordar y comprender, desde otra
perspectiva, la variabilidad biológica de esos grupos humanos.
P A L A B R A S C L A V E :
Bioantropología, poblaciones sudamerindias, diversidad genética,
descripción y análisis.
Artículorecibido:1dejuniode2011|aceptado:1denoviembrede2011|modificado:18denoviembrede2011
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,294pp.ISSN1900-5407,pp.29-54
* DoctorenCienciasNaturales,UniversidaddeBuenosAires,Argentina.

3 0
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.29-54
Bioanthropology ContriButions to the Knowledge of BiologiCal VariaBility among south ameriCan indians: analysis and desCription
os aportes da Bioantropologia ao ConheCimento da VariaBilidade BiologiCa dos sul-amerindios: desCrição e análise
ABSTR AC T This paper analyzes and
describes bioanthropological studies
on South American Amerindians. Three
periods in the development of biological
anthropology, from the 19th century
up to the present, were evaluated with
the intention to analyze the technical,
methodological and theoretical contributions
predominant in each of them. To achieve
this objective, contributions on population
genetics were considered, opening new
ways to approach and understand, from
a different perspective, the biological
variability of these human groups.
K E Y W O R D S :
Bioanthropology, Genetic Diversity, South
Amerindian Populations, Description and
Analysis.
RESUMO Neste trabalho descreve-se e
analisa-se a informação proporcionada pelos
estudos bioantropológicos em populações
ameríndias sul-americanas. Foram avaliadas
três etapas do desenvolvimento da
antropologia biológica, desde o século XIX
até o presente, com a finalidade de analisar
os aportes técnico-metodológicos e teóricos
predominantes em cada uma delas. Para
alcançar esses objetivos, foram consideradas,
principalmente, as contribuições da genética
de populações que abriram novos rumos
para abordar e compreender, desde outra
perspectiva, a variabilidade biológica desses
grupos humanos.
P A L A V R A S C H A V E :
Bioantropología, populações sul-ameríndias,
diversidade genética, descrição e análise.

3 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.29-54
LOS A PORT ES DE L A BIOA N TROPOLOGÍ A A L CONOCI M I EN TO DE L A VA R I A BI LI DA D BIOLÓGICA DE LOS SU DA M ER I N DIOS . DESCR IPCIÓN Y A NÁ LISIS
F r a n c i s c o r a ú l c a r n e s e
IntroducciónEn este trabajo se describen y analizan los avan-ces del conocimiento de la biología de las poblaciones indígenas sudamerica-nas, en las distintas etapas del desarrollo de la antropología biológica. Dada la vastedad de esta tarea, y debido a las limitaciones del espacio y a las propias del autor acerca del conocimiento de las distintas áreas de la bioantropología, el estudio se circunscribió a evaluar, principalmente, las contribuciones realiza-das por la genética de poblaciones, disciplina que abrió nuevos caminos para abordar y comprender, desde otra perspectiva, la variabilidad biológica de los indígenas sudamericanos. Dentro de ese marco, el análisis se limitó a conside-rar los aportes técnico-metodológicos y teóricos más significativos que apun-talaron el desarrollo de la disciplina. Teniendo en cuenta esas contribuciones, el estudio se dividió en tres períodos, con una extensión temporal que va desde los albores de la antropología a mediados del siglo XIX hasta la época actual.
Para desarrollar la primera de las tres etapas propuestas, se examinaron trabajos de autores de Argentina, Brasil, Chile, Uruguay y Venezuela, sobre la historia de la antropología biológica de sus respectivos países. Estos artículos fueron publicados en la Enciclopedia editada por Spencer (1997). En esta publi-cación no se registraron estudios realizados por autores de otros países sudame-ricanos. No obstante, se ha recabado información bibliográfica para el conjunto de los períodos analizados, a partir de otras fuentes no contenidas en esa Enci-clopedia. Los libros de Salzano y Callegari-Jacques (1988), Crawford (1992) y artículos originales fueron analizados para valorar el estado de avance de los estudios genético-serológicos en las poblaciones nativas de América, durante

3 2
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.29-54
el período que se extendió desde la década de 1920 hasta mediados de los años ochenta. Por último, se consultaron trabajos que evaluaron la variabilidad bio-lógica de esos grupos poblacionales, a partir de la incorporación en los estudios bioantropológicos de técnicas y métodos provenientes de la biología molecular.
Los primeros pasosPara la mayoría de los autores, los estudios bioantropológicos comienzan en los inicios o a mediados del siglo XIX. Este período se caracterizó por la pre-sencia en las instituciones académicas, museos y universidades de investiga-dores extranjeros provenientes, principalmente, de países europeos. Poste-riormente, muchos de ellos se establecieron en la región (Peter W. Lund, Paul Ehrenreich, Max Uhle, Henri Dumont, Adolf Bastian, Hermann Burmeister, Hermann ten Kate, Lehmann Nitsche, Martin Gusinde, Adolf Ernst, entre otros) y se relacionaron con investigadores nativos que, inicialmente, eran en su mayoría autodidactas.
Según Rothhammer y Aspillaga (1997), en todos los países hubo un período precientífico en la antropología f ísica. Ese período termina en Chile con la publi-cación del libro Los aborígenes de Chile, en 1882, de José T. Medina (1852-1930), quien resumió las contribuciones realizadas por otros autores, y también aportó concepciones modernas que aplicó al estudio de comunidades indígenas. En 1912, Max Uhle, arqueólogo alemán, organizó el Museo de Etnología y Antro-pología Chilena, en Santiago de Chile. Colaboraron con él Aureliano Oyarzún y Martin Gusinde (1886-1969). Este último autor realizó una exhaustiva des-cripción de los indios fueguinos que, posteriormente, fue publicada, en idioma español, por el Centro Argentino de Etnología Americana (1990).
En Brasil, João Baptista de Lacerda fue considerado el primer antropó-logo biólogo de ese país. Entre 1875 y 1893, realizó estudios morfológicos en poblaciones actuales y extintas de Brasil (ver Lacerda y Peixoto, 1876). El libro Rondonia, publicado en 1917 por Edgard Roquette Pinto (1884-1954), fue uno de los hechos más importantes para el desarrollo de la antropología brasileña (Salzano, 1997).
Los estudios realizados por Adolf Ernst (1832-1899) en el Museo de Cien-cias Naturales de Caracas fueron profundizados por Gaspar Marcano (1850-1910), nacido en Venezuela y formado en París, quien estudió las características craneométricas de los habitantes precolombinos de varias regiones de Vene-zuela (Castro de Guerra, 1997).
En Uruguay deben mencionarse los trabajos de José H. Figueira (1860-1946), quien realizó una descripción de las características f ísicas del material recuperado en enterratorios, ubicados en el este del país (Sans, 1997).

3 3
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.29-54
L O S A P O R T E S D E L A B I O A N T R O P O L O G Í A | F R A N C I S C O R A Ú L C A R N E S E
Un hecho trascendente en este período fue el descubrimiento de los restos de Lagoa Santa por Peter W. Lund (1801-1880). En la actualidad, este hallazgo es motivo de interesantes debates, debido a que, por su antigüedad y características morfológicas, estos restos son considerados como pertenecien-tes a un grupo poblacional que ingresó a América, previamente a la llegada de los amerindios (Neves y Pucciarelli, 1990, 1991; González et al., 2008; Hubbe et al., 2010). En esta época predominaron los estudios antropométricos (cra-neometría y osteología) y antroposcópicos de poblaciones indígenas actuales y extintas y de grupos neoamericanos. También se realizaron investigaciones sobre mezcla racial, endogamia y consanguinidad.
En general, los principales objetivos de la disciplina se centraron en las clasificaciones biotipológicas de las poblaciones indígenas. Las especulaciones acerca del origen del hombre y del hombre americano fueron también un tema de interés de los investigadores. En este sentido, es importante mencionar las obras de Florentino Ameghino (1854-1911) y sus elucubraciones en torno al origen del hombre en América, que, si bien fueron desvirtuadas por la reali-dad, tuvieron el mérito de haber despertado e impulsado el interés por estos estudios tanto en Argentina como en el mundo entero. Por ejemplo, Hrdlička, el padre de la bioantropología norteamericana, dedicó un libro entero a los hallazgos de Sudamérica, motivado por los trabajos de Ameghino (Hrdlička, 1912). (Ver Carnese et al., 1997; Carnese y Pucciarelli, 2007).
El darwinismo fue el marco teórico preponderante de la época, no sólo en antropología sino, también, en otras disciplinas científicas. Las concepciones evolucionistas tuvieron una influencia considerable en los ámbitos académicos e institucionales de casi todos los países sudamericanos (Glick, 1989).
Es recién en la década de 1920 cuando comienzan a realizarse los prime-ros estudios genéticos en poblacionales amerindias de Sudamérica.
Los aportes de la genética serológicaUna segunda etapa en el desarrollo de la disciplina se corresponde, princi-palmente, con la incorporación de técnicas y métodos provenientes de la genética-serológica. Mediante la determinación de marcadores genéticos eri-trocitarios y séricos se realizaron estudios en diversas poblaciones indígenas y neoamericanas.
Uno de los hechos más importante en Sudamérica fue marcado por Baci-galupo, quien difundió, por primera vez, los resultados de sus análisis serológi-cos en aborígenes argentinos, mediante el empleo de técnicas estadísticas apro-piadas (Bacigalupo, 1926, 1934). Casi al mismo tiempo, Mazza y Franke (1927, 1928) determinaron el grupo ABO en más de un centenar de aborígenes del

3 4
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.29-54
Chaco argentino, con el objetivo de establecer el origen de esas poblaciones. En esa época, Imbelloni, a pesar de ser un investigador proveniente de la escuela morfológica adscripta al hiperdifusionismo del historicismo cultural, adhirió a la inclusión de los grupos sanguíneos en los estudios bioantropológicos, por su extraordinaria eficacia heurística; porque se heredan independientemente de la edad y el sexo; no se modifican durante el desarrollo ontogénico, y es escasa o nula la influencia ambiental. Por estas razones, entendía que eran de suma utilidad para las clasificaciones raciales (Imbelloni, 1937).
Otro hecho de trascendencia fue producido por Layrisse et al. (1955), quienes descubrieron el factor Diego A (DI*A) en una familia indígena vene-zolana (Castro de Guerra, 1997). Este hallazgo tuvo repercusión en antropo-logía, porque reveló las relaciones genéticas entre las poblaciones amerin-dias y asiáticas.
A su vez, las tipificaciones de enzimas eritrocitarias y séricas, de las inmu-noglobulinas Gm/Km, del sistema HLA y del carácter secretor ABH, entre otras determinaciones, posibilitaron la obtención de un panorama más actualizado sobre la variabilidad biológica de las poblaciones indígenas sudamericanas.
Una de las observaciones más interesantes de estos estudios está rela-cionada con los grupos sanguíneos. Se constató que los amerindios tenían fre-cuencias cercanas al 100% para el alelo ABO*O, eran casi todos Rh positivos y, en promedio, un 10% presentaban el alelo Diego A (DI*A).
En relación con el sistema ABO, Crawford (1992: 32) señaló que antro-pólogos y genetistas sugirieron “que los amerindios se habían separado de las demás poblaciones humanas antes de la evolución de los antígenos A y B, y que ésa sería la causa de la ausencia de esos alelos en la mayoría de las poblaciones americanas”1. Por consiguiente, se dedujo que la presencia de esos marcadores en grupos amerindios sería consecuencia de mezcla génica con europeos.
Sin embargo, las poblaciones siberianas actuales son polimórficas para el sistema ABO, con una frecuencia del alelo ABO*A1 de casi un 20% en los Chukchi de la península de Chukotka y un 14% entre los esquimales siberianos, quienes, a su vez, presentan un 23% del alelo ABO*B (Crawford, 1992). Estas variantes polimórficas fueron observadas también en otros grupos siberianos. ¿Si esos antígenos estaban presentes en esas poblaciones, y si se admite que ellas fueron parte de los grupos fundadores de América, cómo se explica la ausencia de los mismos en las actuales poblaciones amerindias?
Se sugirieron al respecto varias hipótesis, donde la deriva génica, la selec-ción natural o la acción combinada de ambos mecanismos microevolutivos
1 LaexcepciónsonlosindígenasBlodfootsyBlood,quepresentabanelevadasfrecuenciasdelaleloA.

3 5
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.29-54
L O S A P O R T E S D E L A B I O A N T R O P O L O G Í A | F R A N C I S C O R A Ú L C A R N E S E
habrían sido las causas determinantes de la fijación del alelo ABO*O y la pér-dida de los alelos ABO*A y ABO*B.
Afortunadamente, debido a que estos antígenos están presentes en distintos tejidos y órganos del cuerpo humano, y que su capacidad antigénica se preserva del paso del tiempo, se comenzaron a desarrollar investigaciones para detectar esas sustancias en material óseo y/o momificado de épocas precolombinas. Con estos estudios se intentó dar respuesta a los interrogantes arriba formulados.
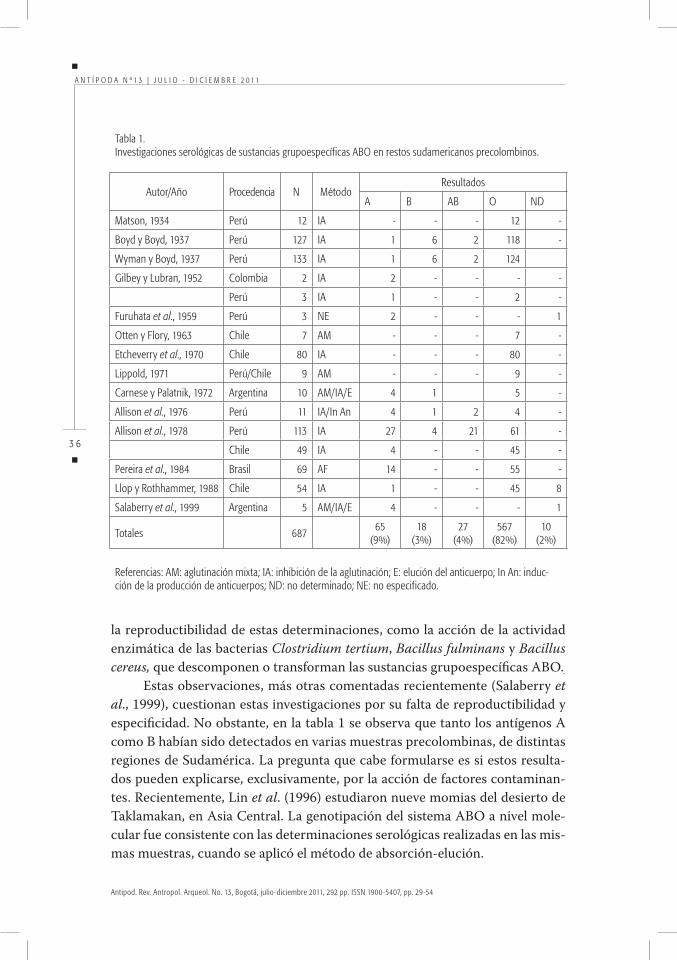
Los estudios paleoserológicosLa presencia de sustancias grupales sanguíneas, en su forma alcohol soluble, en tejidos humanos posibilitó su determinación mediante técnicas serológicas que habían sido empleadas en medicina legal.
En la tabla 1 se mencionan algunas de ellas, así como las determinaciones realizadas a partir de restos momificados de diferentes regiones de Sudamérica. Se puede observar que los antígenos A y B están presentes, aunque en bajas proporciones, en Colombia, Perú, Chile, Brasil y Argentina. La presencia de B en indios, precontacto con europeos, de la región andina de América del Sur es coincidente con lo remarcado para las poblaciones vivientes de esa misma área. A partir de estos datos se postuló que, si se descarta la acción de factores con-taminantes que interfieren en la tipificación de los especímenes, la existencia de los antígenos A y B en América precolombina no sería sólo consecuencia del mestizaje con europeos.
Se consideró, entonces, que la baja o nula frecuencia de esos antígenos en aborígenes actuales podría deberse a la acción de mecanismos microevolutivos como la deriva genética, la cual debió haber cumplido un importante rol en la diferenciación genética de esas poblaciones. A su vez, se sostuvo que un papel destacado pudo haber sido desempeñado por la selección natural, a través de mecanismos de susceptibilidad y resistencia a ciertas enfermedades y en fun-ción de determinados genes de grupos sanguíneos (Carnese y Palatnik, 1972).
Estas investigaciones habían proporcionado información de interés para aquellos que intentaban interpretar las relaciones y las variaciones entre las poblaciones pre y poscontacto con los europeos y para hipotetizar acerca de las probables causas que determinaron la composición genética de los indí-genas actuales.
Sin embargo, en su momento, estos estudios fueron criticados por Thieme y Otten (1957), quienes sostenían, entre otros argumentos, que la gran distri-bución en la naturaleza de sustancias semejantes a los antígenos de grupos san-guíneos puede actuar como contaminante, dando lugar a falsas reacciones de inhibición. Los mismos autores analizaron otros factores que influyen sobre
3 6
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.29-54
la reproductibilidad de estas determinaciones, como la acción de la actividad enzimática de las bacterias Clostridium tertium, Bacillus fulminans y Bacillus cereus, que descomponen o transforman las sustancias grupoespecíficas ABO.
Estas observaciones, más otras comentadas recientemente (Salaberry et al., 1999), cuestionan estas investigaciones por su falta de reproductibilidad y especificidad. No obstante, en la tabla 1 se observa que tanto los antígenos A como B habían sido detectados en varias muestras precolombinas, de distintas regiones de Sudamérica. La pregunta que cabe formularse es si estos resulta-dos pueden explicarse, exclusivamente, por la acción de factores contaminan-tes. Recientemente, Lin et al. (1996) estudiaron nueve momias del desierto de Taklamakan, en Asia Central. La genotipación del sistema ABO a nivel mole-cular fue consistente con las determinaciones serológicas realizadas en las mis-mas muestras, cuando se aplicó el método de absorción-elución.
Tabla1.InvestigacionesserológicasdesustanciasgrupoespecíficasABOenrestossudamericanosprecolombinos.
Autor/Año Procedencia N MétodoResultados
A B AB O ND
Matson,1934 Perú 12 IA - - - 12 -
BoydyBoyd,1937 Perú 127 IA 1 6 2 118 -
WymanyBoyd,1937 Perú 133 IA 1 6 2 124
GilbeyyLubran,1952 Colombia 2 IA 2 - - - -
Perú 3 IA 1 - - 2 -
Furuhataet al.,1959 Perú 3 NE 2 - - - 1
OttenyFlory,1963 Chile 7 AM - - - 7 -
Etcheverryet al.,1970 Chile 80 IA - - - 80 -
Lippold,1971 Perú/Chile 9 AM - - - 9 -
CarneseyPalatnik,1972 Argentina 10 AM/IA/E 4 1 5 -
Allisonet al.,1976 Perú 11 IA/InAn 4 1 2 4 -
Allisonet al.,1978 Perú 113 IA 27 4 21 61 -
Chile 49 IA 4 - - 45 -
Pereiraet al.,1984 Brasil 69 AF 14 - - 55 -
LlopyRothhammer,1988 Chile 54 IA 1 - - 45 8
Salaberryet al.,1999 Argentina 5 AM/IA/E 4 - - - 1
Totales 68765
(9%)18
(3%)27
(4%)567
(82%)10
(2%)
Referencias:AM:aglutinaciónmixta;IA:inhibicióndelaaglutinación;E:elucióndelanticuerpo;InAn:induc-cióndelaproduccióndeanticuerpos;ND:nodeterminado;NE:noespecificado.

3 7
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.29-54
L O S A P O R T E S D E L A B I O A N T R O P O L O G Í A | F R A N C I S C O R A Ú L C A R N E S E
En consecuencia, los intentos de explicar las causas determinantes de la distribución del sistema ABO en poblaciones amerindias no tienen, todavía, una respuesta satisfactoria. Seguramente, nuevas investigaciones a nivel mole-cular podrán, en un futuro próximo, aclarar esta cuestión.
En resumen, en este período, las investigaciones realizadas aportaron importante información sobre la biología de las poblaciones sudamerindias. Sin embargo, la incorporación de la genética serológica en los estudios bioan-tropológicos, si bien constituyó un cambio técnico y metodológico importante, no modificó la concepción básica de la variabilidad biológica de la época. Uno de los objetivos principales de la antropología biológica siguió siendo la carac-terización racial de los grupos humanos en entidades discretas, idea que pre-valeció hasta mediados del siglo XX. En un trabajo anterior se señaló que “El criterio tipológico de acuerdo a como fuera definido por Mayr (1968) y por Henning (1968) fue nodal en las explicaciones que se brindan en nuestra dis-ciplina. Aún con los trabajos serológicos de Boyd (1950) se llega a una concep-tualización raciológica similar, en lo esencial, a la de Blumenbach” (Pucciarelli, 1989: 28; ver Carnese et al., 1991-92: 44).
En esta etapa, el marco teórico predominante en la antropología biológica parece haber sido el darwinismo, por el enfoque dado a las investigaciones, aunque en Argentina se produjo un retroceso de las concepciones evolucionis-tas y un significativo avance de las ideas hiperdifusionistas de la Escuela Histó-rica-Cultural, asociada al método tipológico. El máximo representante de esa concepción fue José Imbelloni.
Las décadas de los años sesenta-ochenta Es a partir de la década de 1960 cuando comienza a producirse un paulatino reemplazo del método tipológico por métodos estadísticos uni y multiva-riado, que, junto con el desarrollo de la informática, se convirtieron en herra-mientas fundamentales para el procesamiento y análisis de una significativa cantidad de información. Estos aportes permitieron abrir nuevos caminos para una mejor interpretación de la variabilidad biológica humana. Se pro-fundiza el conocimiento de las estructuras genéticas de las poblaciones indí-genas, de los contactos interétnicos que establecieron y de la mezcla génica con grupos no amerindios. Entre otros significativos avances, las variaciones interpoblacionales comienzan a estimarse a partir de diversos coeficientes de distancias biológicas.
Además, surgieron nuevas hipótesis acerca del origen de los amerindios y de los grupos migratorios que ingresaron al continente americano. En ese sen-tido, el trabajo de Greenberg et al. (1986) fue una excelente contribución a toda

3 8
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.29-54
esa temática. Más adelante, una revisión de los estudios realizados en poblaciones indígenas sudamericanas fue publicada por Salzano y Callegari-Jacques (1988).
En este período, un hecho de trascendencia fue protagonizado por Lewon-tin (1972), quien demostró, mediante el análisis de diecisiete sistemas genéticos tradicionales, que de toda la variación genética humana, el 85% corresponde a variación entre individuos dentro de una nación o tribu, mientras que el resto se reparte entre naciones (8%) y las razas principales (6%). Posteriormente, los resultados obtenidos por Lewontin fueron confirmados mediante el empleo de marcadores genéticos moleculares (ver Barbujani, 2005). Al respecto, es intere-sante remarcar que los datos craneométricos son, también, coincidentes con las observaciones registradas a nivel serológico y molecular (ver Relethford, 1994). Estas observaciones originaron interesantes debates acerca de si el concepto de raza es reflejo de una realidad biológica o una construcción social.
La biología molecular Hacia mediados de la década de 1980 comienza a percibirse un importante cambio en las investigaciones bioantropológicas, con la incorporación de téc-nicas y métodos provenientes de la biología molecular2.
El primer trabajo en amerindios, que incluyó a treinta Warao de Venezuela, fue realizado por Johnson et al. (1983) mediante la técnica de longitud de frag-mentos polimórficos de restricción (restriction fragment length polymorphism, RFLP) empleando cinco enzimas de restricción. A partir de este estudio se inten-sificaron las investigaciones con el empleo de un número mayor de enzimas y la secuenciación de la región control del ADNmt (Wallace et al., 1985; Torroni et al., 1992; Ward et al., 1991). Con estas investigaciones se constató la presencia de cuatro haplogrupos mitocondriales de origen asiático: A, B, C y D, y, poste-riormente, sus variantes autóctonas americanas: A2, B2, C1 y D1. Más tarde, Brown et al. (1998) describen un quinto haplogrupo fundador denominado X, que está presente con bajas frecuencias, tanto en amerindios norteamericanos como en poblaciones europeas y siberianas actuales (Derenko et al., 2001), pero no se detectó en los grupos poblacionales que habitan actualmente en el este de Siberia, probablemente, por efecto de la deriva génica (Fagundes et al., 2008). En cambio, otros investigadores sugirieron la existencia de un número mayor de haplogrupos fundadores de América (Bailliet et al., 1994; Bianchi et al., 1995).
Según Salzano (2002), hasta 2001 se habían estudiado 90 muestras de nativos americanos, con un total de 3.809 individuos; 39 de esas muestras pro-
2 Paraeldesarrollode laprimerapartedeesteapartado se tuvoen consideración la revisiónefectuadaporSalzanoen2002.

3 9
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.29-54
L O S A P O R T E S D E L A B I O A N T R O P O L O G Í A | F R A N C I S C O R A Ú L C A R N E S E
venían de Sudamérica, e incluyen a 1.770 personas. Se observa que el haplo-grupo B era, en promedio, el más frecuente (0,318), seguido del C y el D, con valores similares, 0,234 y 0,233, respectivamente, y que el haplogrupo A (0,188) registraba la menor prevalencia. A partir de estos hallazgos, se concluyó que en la región sudamericana hay una distribución bastante uniforme de los cuatro haplogrupos, semejante a la de los amerindios de Norteamérica.
Las primeras investigaciones realizadas a partir del cromosoma Y per-mitieron indagar los linajes paternos y compararlos con los de origen materno derivados del ADN mitocondrial. Diversos estudios demostraron que su varia-bilidad era menor a la observada en el ADN mitocondrial.
Bianchi et al. (1997) sugirieron la existencia de un simple haplotipo fun-dador de América, mientras que otros autores plantearon que por lo menos dos serían los linajes paternos que han contribuido al acervo génico de las poblaciones americanas (Rodríguez-Delfin et al., 1997; Karafet et al., 1999; Ruiz-Linares et al., 1999).
En otras investigaciones se analizaron marcadores genéticos autosoma-les: sistema HLA, polimorfismos de inserción cortos o largos (Short or Large, LINE); longitud de fragmentos polimórficos de restricción (RFLP, en inglés); pequeñas secuencias de nucleótidos repetidos en tándem o microsatélites (Short tandem repeats, STRs); polimorfismos nucleotídicos simples de sustitu-ción (Single nucleotide polymorphisms, SNPS) y número variable de nucleóti-dos repetidos en tándem (Variable number of tandem repeats, VNTRs).
Por otra parte, hasta 2002 fueron escasos los trabajos en los que se utili-zaron marcadores genéticos del cromosoma X, para el análisis de poblaciones aborígenes sudamericanas (Salzano, 2002).
A partir de ese año, que se corresponde con la revisión realizada por Sal-zano, se produjeron significativos aportes técnico-metodológicos que permitie-ron analizar con mayor profundidad las estructuras y la diversidad genética de las poblaciones indígenas y plantear nuevas hipótesis acerca del poblamiento americano. Es profusa la bibliograf ía existente sobre estos temas, como para intentar realizar una revisión exhaustiva de la misma.
Por esa razón, se presenta un análisis de la variabilidad biológica de los sudamerindios, a partir de los datos disponibles hasta 2008, que fue comu-nicado ese mismo año en el Congreso de la Asociación Latinoamericana de Antropología Biológica (ALAB), realizado en la ciudad de La Plata, Argentina (Carnese et al., 2008).
En la tabla 2 se muestran las determinaciones genéticas realizadas en las poblaciones estudiadas, y en la figura 1, la distribución geográfica de las mismas. Como puede observarse, no fueron incluidos en el estudio todos los

4 0
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.29-54
Tabla2.Marcadoresgenéticosempleadosparaelanálisisdeladiversidadgenéticadelaspoblacionessudamerindias.
Marcadores Poblaciones
Proteicos(1)
ABO,MNSs,Rh,Fy,Di,JK,P,KELL,G6PD,HP,TF.
Mapuche(Argentina),Mapuche(Chile),Wichi,Tehuelche,Toba,Mocoví,Pilagá,Chorote,Chiriguano,Lengua,Xavante,Yanomami,Wapishama,Aymara,Macushi,Makiritare,Ayoreo,Guarani,Kaingang,Ticuna.
(N=19519)
Moleculares(2)
ADNmitocondrial Mapuche(Argentina),Mapuche(Chile),Wichi,Tehuelche,Toba,Mocoví,Pilagá,Chorote,Chiriguano,Lengua,Xavante,Yanomami,Wapishama,Aymara,Macushi,Makiritare,Ayoreo,Guarani,Kaingang,Ticuna.
(N=1518)
SistemaHLAClaseII: locusHLA-DRB1
Mapuche(Argentina),Mapuche(Chile),Chiri-guano,Wichi,Tehuelche,Toba,Pilagá,Lengua,Xavante,Guarani(Brasil),Kaingang,Ticuna.(N=1118)
CromosomaY.Loci:DYS19,DYS390,DYS391,DYS392,DYS393.
Mapuche(Argentina),Tehuelche,Wichí,Toba,Pilagá,Chorote,Lengua,Ayoreo,Guarani(Brasil),Ticuna.(N=211)
STRsautosómicosI.Loci:D3S1358,D5S818,D7S820,D8S1179,D13S317 Wichí,Toba,Pilagá,Xavante,Kaingang,Gua-
rani,Ayoreo.(N=309)
STRsautosómicosII.Loci:D7S820,D13S317,vWA,D16S539
Mapuche(Argentina),Wichi,Tehuelche,Pilagá,Toba,Xavante,Guarani(Brasil),Kaingang,Ayoreo.(N=411)
Referencias:(1)NagelyEtcheverry,1963;Matsonet al.,1967,1969;CabuttiyPalatnik,1975;PagesLarrayaet al.,1978;Haaset al.,1985;SalzanoyCallegari-Jacques,1988;Carnese,1995;Goicoecheaet al.,2001;Carneseet al.,2002.(2)Cernaet al.,1993;Gintheret al.,1993;Torroniet al.,1992-1994;Baillietet al.,1994;Merriwetheret al.,1995;Wardet al.,1996;Blagitkoet al.,1997;Bianchiet al.,1997-98;Salaet al.,1999;Pando,1998;Moragaet al.,2000;Demarchiet al.,2001;Hutzet al.,2002;Tsunetoet al.,2003;DemarchiyMichell,2004;Dornelleset al.,2004;Bravi,2005;Kohlrauschet al.,2005;Cabanaet al.,2006;Catanesiet al.,2006;Marreroet al.,2007;ParolínyCarnese,2009;Crossettiet al.,2008;Toscaniniet al.,2008.
grupos indígenas sudamericanos. Se seleccionaron algunos de ellos, que com-parten las mismas determinaciones genéticas y pertenecen a diversos troncos lingüísticos: Araucarian (mapuches); Aymará (Aymará); Patagon (tehuelches); Mataco (wichi, chorote); Guaycurú (toba, mocoví, pilaga); Mascoy (lengua); Zamuco (ayoreo); Kaingang (kaingang), Ge (xavante); Arawak (wapishana); Yanomami (yanomama); Karaib (macushi, makiritare); Tupí-Guaraní (guaraní, chiriguano); Ticuna (lengua independiente). Estas poblaciones fueron analiza-das para once marcadores proteicos y ADNmt. A su vez, sólo ha sido posible

4 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.29-54
L O S A P O R T E S D E L A B I O A N T R O P O L O G Í A | F R A N C I S C O R A Ú L C A R N E S E
emplear en el estudio al locus HLA*DRB1, porque fue el único estudiado en poblaciones tehuelches, lengua, y en cuatro grupos mapuches de la provincia de Río Negro, Argentina (Parolín y Carnese, 2009). En cambio, para el resto de los sistemas, y debido a que no todas las poblaciones fueron estudiadas para los mismos loci, se priorizó, en ciertos casos, el número de poblaciones, en detri-mento del número de tipificaciones genéticas, y en otros se procedió de manera inversa, como puede observarse, por ejemplo, en las determinaciones de los STRs autosomales. A pesar de estas limitaciones, se logró arribar a algunas conclusiones que coinciden, en general, con las formuladas por otros autores.
Se constata que la diversidad biológica varía según el sistema genético ana-lizado. En todos los casos, la variabilidad genética intrapoblacional explicó la mayor parte de la variación genética total, que va del 75% en el ADNmt al 93% en los sistemas proteicos. La elevada variación genética interpoblacional a nivel del ADNmt (25%) se explica porque al ser transmitido sólo por línea materna, su tamaño efectivo es ¼ respecto de los sistemas genéticos de transmisión bipa-rental. Un tamaño efectivo reducido acentúa las distancias genéticas entre las poblaciones, similar a la acción de la deriva genética, originando un aumento de la diferenciación genética interpoblacional. Asimismo, los STRs del cromosoma Y y el locus DRB1 del sistema HLA registraron, también, en términos relativos, valores elevados de variación genética interpoblacional, 19% y 12%, respectiva-mente. Estos valores disminuyeron al 9% con los marcadores STRs autosómicos en las dos muestras poblacionales, y al 6% para los sistemas proteicos.
Figura1.PoblacionesindígenasdeSudaméricaestudiadasapartirdediversosmarcadoresgenéticos.
Tehuelche
TobaWichiPilago
Guaraní
LenguaChiriguano
Chorote
Macushi
Wapishama
Makiritare
Mocoví
Ayoreo
Aymara
Xavante
TicunaYanomama
MapucheArgentina
MapucheChile
Kaingang

4 2
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.29-54
Estos altos porcentajes de diferenciación genética interpoblacional en sudamerindios son superiores a los observados entre poblaciones de otros con-tinentes (Barbujani, 2005) y se deberían, probablemente, al accionar de meca-nismos estocásticos que determinaron sus estructuras demográfico-genéticas. Esta característica había sido observada, previamente, por Salzano (2002) para el conjunto de los grupos amerindios.
Otro aspecto destacable de los amerindios es que el ADNmt presenta una variabilidad más restringida respecto de otras poblaciones mundiales. Hay que tener en cuenta que esos grupos sufrieron reducciones dramáticas de sus poblaciones como consecuencia de los enfrentamientos con los conquistadores europeos y los ejércitos nacionales y, adicionalmente, por las enfermedades y epidemias que asolaron gran parte de esas comunidades. Estos hechos podrían ser una de las causas determinantes de la variabilidad biológica observada en los indígenas actuales.
Para aclarar, en parte, esta problemática, las investigaciones del ADN antiguo se están convirtiendo en una herramienta útil que seguramente, en un futuro mediato, incrementará el conocimiento biológico de las poblacio-nes precolombinas, permitirá estimar las relaciones genéticas que se han esta-blecido entre ellas, calcular la diversidad genética precontacto con europeos y comparar ésta con la observada en sus descendientes actuales.
Con base en estas consideraciones, se presenta en el siguiente apartado una revisión de los estudios paleogenéticos realizados en grupos precolombi-nos e históricos de Sudamérica.
Los estudios paleogenéticosComo se mencionó en los párrafos anteriores, las investigaciones genéticas en poblaciones aborígenes sudamericanas actuales han tenido un significativo desarrollo y contribuyeron al conocimiento de la historia evolutiva de dichos grupos. Desde 1990, con el análisis del ADN antiguo en restos precolombinos se incorporó la dimensión temporal en las investigaciones bioantropológicas.
Con respecto a América del Sur, se realizaron estudios en diferentes sitios arqueológicos, desde el extremo norte del subcontinente hasta Tierra del Fuego (ver la tabla 3). Monsalve et al. (1996) analizaron ocho momias pre-colombinas de Colombia (1526-150 AP) y detectaron tres de los cuatro haplo-grupos mitocondriales amerindios (5A, 1B y 2C). Al mismo tiempo, Ribeiro dos Santos et al. (1996) amplificaron y secuenciaron la región control del ADN mitocondrial en dieciocho restos esqueletarios precolombinos (4.000-500 AP) de la cuenca amazónica. Diez de ellos presentaron las mutaciones típicas de los haplogrupos A (5), C (4) y D (1). Una muestra más antigua, 4.000 años AP,

4 3
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.29-54
L O S A P O R T E S D E L A B I O A N T R O P O L O G Í A | F R A N C I S C O R A Ú L C A R N E S E
fue asignada al haplogrupo B, que se distingue por la deleción de 9pb en la región V; las restantes muestras no pudieron ser asignadas a linajes mitocon-driales amerindios. Previamente, Horai et al. (1991) no observaron la deleción de 9pb en momias del norte de Chile.
Por otra parte, en el extremo sur del continente, en la región Fuego-Pata-gónica, Lalueza et al. (1993/4, 1997) no detectaron los haplogrupos A y B en 60 restos de aborígenes fueguinos de colecciones de Chile y Argentina (100-200 BP, incluidas dos muestras de restos esqueletales de 4.030 y 5.000 años). Además, mediante la secuenciación de 24 de esas muestras, García Bour et al. (2004) detectaron, en todos los casos, las mutaciones características de los
Tabla3.HaplogruposmitocondrialesengruposprecolombinosehistóricosdediferentesregionesdeSudamérica.
Sitio/población N Haplogrupos Autor
A B C D Otros
Colombia(152-150AP) 8 0,630 0,120 0,250 0,000 0,000 Monsalveet al.,1996
Amazonas(4.000-500AP) 18 0,280 0,050 0,220 0,050 0,390 RibeiroDosSantoset al.,1996
RegiónFuego-Patagónica (100-200AP,incluye1muestrade4.030APy1muestrade5.000AP)
60 0,000 0,000 0,380 0,600 0,020 Laluezaet al.,1997
Chile,MomiasdeAzapa (2.990-6.000AP)
32 0,312 0,219 0,312 0,031 0,125 Moragaet al.,2001
Tiwanaku 13 0,080 0,150 0,230 0,230 0,310 Rothhammeret al.,2003
ChileArcaicoTardío (6.000-3.900AP)
14 0,500 0,357 0,070 0,070 0,000 Moragaet al.,2005(*)
ChileHoriz.Medio (1.650-1.000AP)
19 0,316 0,421 0,263 0,000 0,000 Moragaet al.,2005
ChileIntermedioTardío (1.000-500AP)
15 0,200 0,533 0,200 0,067 0,000 Moragaet al.,2005
Perú(Paucarcancha,PatallactayHuata)
35 0,086 0,657 0,220 0,029 Shinodaet al.,2006
Argentina,PampaGrande (650AP)
19 0,110 0,470 0,000 0,420 0,000 Carneseet al.,2010
Referencias:RoganySalvo(1990),Horaiet al.(1991),Laluezaet al.(1993-94),Merriwetheret al.(1994)yDemarchiet al.(2001)nodetectanladelecióndelos9pbenmomiasdelnortedeChile,entejidoblandonidientesdelNorte,CentroySurdeArgentina.García-Bouret al.(2004)secuenciaron24restosdelos60estudiadosporLaluezaet al.(1997)yconfirmaronlaausenciadeloshaplogruposAyBenTierradelFuego-Patagonia.(*)LatotalidaddelosrestosdelosperíodosanalizadosporMoragaet al.,2005suman61individuos,deloscuales13(21%)nopertenecenaloshaplogruposclásicosamerindiosylosclasificancomo“otros”.

4 4
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.29-54
haplogrupos C y D, coincidente con los resultados previos realizados por RFLP. A su vez, lograron tipificar varios microsatélites del cromosoma Y.
Cabe destacar que el haplogrupo B no había sido detectado en tejidos blandos, huesos y dientes en 56 restos prehispánicos del norte de Chile, y del norte, centro y sur de Argentina, y, particularmente, en aquellos grupos que habitaron el extremo sur del continente (Rogan y Salvo, 1990; Merriwether et al., 1994; Demarchi et al., 2001).
Sin embargo, Moraga et al. (2001), en momias provenientes del valle de Azapa (2.990-6.000 AP), y Rothhammer et al. (2003), en varios restos esque-letales provenientes de Tiwanaku, detectaron una frecuencia relativamente alta de B (15% y 22%, respectivamente). Más tarde, una alta prevalencia de B fue observada en grupos prehistóricos provenientes de los valles de Lluta, Azapa y Camarones, en el norte de Chile. En estas regiones, el haplogrupo B varía del 35% en el período Arcaico Tardío (6.000-3.900 BP) al 53% en el período Intermedio Tardío (1.000-500 BP) (Moraga et al., 2005). Además, Shinoda et al. (2006) también detectaron altas frecuencias del haplogrupo B (66%) en restos peruanos provenientes de Paucarcancha, Patallacta y Huata, que muestran grandes similitudes con las halladas en las actuales poblaciones Quechua y Aymará de Perú.
Recientemente, en Pampa Grande, provincia de Salta, Argentina, se estu-diaron diecinueve restos de una antigüedad de 1310 AP. En este estudio se detectaron elevadas frecuencias de B (47%) y D (42%), baja proporción de A (10%) y ausencia de C. La elevada presencia del haplogrupo B parece sugerir una estrecha relación genética con los antiguos habitantes de la región centro-sur andina. A su vez, en este trabajo se determinaron, también, nueve STRs autosomales y once STRs del cromosoma Y (Carnese et al., 2010).
En síntesis, de las series analizadas hasta ahora, se observa una relativa-mente elevada prevalencia de los haplogrupos A y baja frecuencia del haplo-grupo B en Colombia y en la región amazónica; frecuencias altas de B en el norte de Chile y en el noroeste de Argentina y en los antiguos habitantes de las altas tierras de Perú, y ausencia de A y B y altas frecuencias de C y D en fueguinos.
Por otra parte, de descartarse algún artefacto metodológico, los datos obtenidos por Ribeiro dos Santos et al. (1996) en la región amazónica, Moraga et al. (2001-2005) en el norte de Chile y Rothhammer et al. (2003) en Tiwanaku parecen indicar la existencia de otros haplogrupos fundadores de América, además de los clásicos A, B, C, D y X (ver la tabla 3). Estos resultados sugieren la existencia de una mayor variabilidad genética en las poblaciones extintas, respecto de los grupos actuales. Sin embargo, los STRs autosomales determina-dos en Pampa Grande presentan frecuencias génicas similares a las observadas

4 5
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.29-54
L O S A P O R T E S D E L A B I O A N T R O P O L O G Í A | F R A N C I S C O R A Ú L C A R N E S E
en poblaciones amerindias, que habitan actualmente en diversas regiones de Sudamérica (Carnese et al., 2010). Estos datos, por el contrario, nos estarían sugiriendo que no hubo variación en el tiempo, por lo menos en relación con ese sistema, de la diversidad genética de los sudamerindios.
Es evidente que más estudios serán necesarios para aclarar las causas determinantes de la variabilidad biológica de las poblaciones amerindias pre y poscontacto con europeos.
La actual superación de problemas técnico-metodológicos en los estudios de ADN antiguo posibilitará el desarrollo de nuevas investigaciones que contri-buirán, como ya señalamos, a analizar las relaciones entre los grupos extintos y actuales de los amerindios en cuanto a continuidad y/o reemplazo y a respon-der interrogantes aún no resueltos sobre el poblamiento americano.
Por último, del análisis efectuado en esta etapa del desarrollo de la antro-pología biológica, se puede observar una eclosión de datos acerca de la varia-bilidad biológica de los sudamerindios. Esta diversidad está siendo analizada desde distintos enfoques y con una magnífica batería de técnicas y métodos, cuyos resultados están aportando datos relevantes para una mejor compren-sión de las probables causas que la determinan. Otro aspecto de interés, que excede el análisis aquí realizado, es el surgimiento de nuevas y plausibles hipó-tesis acerca del origen de los amerindios y de los contactos y rutas migratorias que siguieron en el interior del continente.
Por otra parte, desde el punto de vista teórico, el paradigma predomi-nante en este período es el neodarwinismo, aunque algunos autores como Reig (1984: 583) ya habían sostenido, durante un homenaje a Darwin por los 125 años de la aparición de El origen de las especies, que posiblemente estemos transitando “los albores de una nueva síntesis o de un desarrollo expansivo de la síntesis moderna, que se perfila como una teoría jerárquica de la evolución que pretende superar las limitaciones reduccionistas del darwinismo original y de su versión de la síntesis moderna, admitiendo la existencia de distintos ámbitos y niveles de manifestación de los procesos evolutivos”.
SíntesisSe han descrito y analizado, durante las distintas etapas del desarrollo de la antropología biológica, algunas de las contribuciones que apuntalaron el cono-cimiento de la biología de los amerindios sudamericanos. El análisis efectuado presenta facetas muy interesantes que permiten valorar positivamente lo alcan-zado hasta el presente y vislumbrar un futuro promisorio, dado que, con el empleo de las excelentes herramientas técnico-metodológicas disponibles y dentro de un marco teórico apropiado, se podrá indagar y responder a muchos

4 6
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.29-54
de los interrogantes que, aún, nos formulamos sobre la biología de las poblacio-nes originarias de nuestra América.
Agradecimientos A Victoria Lois, por su colaboración en la preparación del presente manuscrito. A los revisores de este trabajo, quienes con sus sugerencias enriquecieron el contenido del artículo. Al Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) y a la Universidad de Buenos Aires (UBACyT), por su apoyo financiero. .

4 7
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.29-54
L O S A P O R T E S D E L A B I O A N T R O P O L O G Í A | F R A N C I S C O R A Ú L C A R N E S E
R e f e R e n c i a s
Allisson, Marvin, Ali Hossaini, Nora Castro, Juan Munizaga y Alejandro Pezzia1976.“ABOBloodGroupsinPeruvianMummies.I.AnEvaluationofTechniques”,American Journal of
Physical Anthropology 44,pp.55-61.
Allisson, Marvin, Ali Hossaini, Nora Castro, Juan Munizaga y Rosa Fung1978.“ABOBloodGroupsinChileanandPeruvianMummies.II.ResultsofAgglutination-inhibition
Technique”,American Journal of Physical Anthropology 49,pp.139-142.
Bacigalupo, Juan1926.“Estadísticassobreciencasosdegrupossanguíneos”,La Semana Médica 2, pp.734-736.1934. “Estadísticassobregrupossanguíneos”, en V Congreso Nacional de Medicina VII,pp.683-685.
Bailliet, Graciela, Francisco Rothhammer, Francisco R. Carnese, Claudio M. Bravi y Néstor O. Bianchi
1994.“FounderMitochondrialHaplotypesinAmerindianPopulation”,American Journal of Human Genetic54,pp.27-33.
Barbujani, Guido 2005.“HumanRaces:ClassifyingPeopleVs.UnderstandingDiversity”,Current Genomics6(4),
pp.215-226.
Bianchi, Néstor O., Graciela Bailliet y Claudio M. Bravi1995.“PeoplingoftheAmericasasInferredThroughtheAnalysisofMitochondrialDNA”,Brazilian
Journal of Genetics18,pp.661-668.
Bianchi, Néstor O., Graciela Bailliet, Claudio M. Bravi, Francisco R. Carnese, Francisco Rothhammer, Verónica Martinez-Marignac y Sergio D. J. Pena
1997.“OriginofAmerindianY-ChromosomesasInferredbytheAnalysisofSixPolymorphicMarkers”,American Journal of Physical Anthropology102,pp.79-89.
Bianchi, Néstor, Cecilia Catanesi, Graciela Bailliet, Verónica Martinez-Marignac, Claudio M. Bravi, Lidia Vidal-Rioja, René J. Herrera y Jorge Lopez-Camelo
1998.“CharacterizationofAncestralandDerivedY-chromosomeHaplotypesofNewWorldNativePopulations”,American Journal of Human Genetics63,pp.1862-71.
Blagitko, Nadezda, Colm O’Huigin, Felipe Figueroa, Satoshi Horai, Sunro Sonoda, Kazuo Tajima, David Watkins y Jan Klein
1997.“PolymorphismoftheHLA-DRB1LocusinColombian,Ecuadorian,andChileanAmerinds”,Human Immunology54,pp.74-81.
Boyd, William 1950.“BloodGroupsofSouthAmericanIndians”,Handbook of South American Indians, Vol.6. Bulletin
of the Bureau of American Ethnology143,pp.91-95.
Boyd, William C. y Lyle G. Boyd1937.“BloodGroupingTestson300Mummies,withNotesonthePrecipitin-test”,Journal of
Immunology32,pp.307-319.2005.“Análisisdelinajesmaternosenpoblacionesindígenasamericanas”.TesisDoctoral.Facultadde
CienciasNaturalesyMuseo,UniversidadNacionaldeLaPlata.
Brown, Michael D., Seyed H. Hosseini, Antonio Torroni, Hans-Jürgen Bandelt, Jon C. Allen, Theodore Schurr, Rosaria Scozzari, Fulvio Cruciani y Douglas C. Wallace
1998.“mtDNAHaplogroupX:AnAncientLinkbetweenEurope/WesternAsiaandNorthAmerica?”,American Journal of Human Genetics63,pp.1852-1861.
Cabana, Graciela, D. Andrew Merriwether, Keith Hunley y Darío Demarchi2006.“IstheGeneticStructureofGranChacoPopulationsUnique?InterregionalPerspectivesonNative

4 8
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.29-54
SouthAmericanMitochondrialDNAVariation”,American Journal of Physical Anthropology131,pp.108-119.
Cabutti, Nilda F. de y Marcos Palatnik1975.“Aspectosgenéticosmonofactorialeseritrocitarios”,enProgresos en biología,eds.F.Barbieri
yA.Legname,pp.425-434.Tucumán,FundaciónMiguelLillo.
Carnese, Francisco R.1995.“GeneticMarkersintheAboriginalPopulationsofArgentina”,Brazilian Journal of Genetics18,
pp.651-656.
Carnese, Francisco R. y Marcos Palatnik 1972.“Estudiospaleoserológicosderestosmomificadosdeaborígenesargentinos”,Sangre17,pp.201-210.
Carnese, Francisco R., Alberto J. Cocilovo y Alicia S. Goicoechea1991-1992.“HistoriaysituaciónactualdelaantropologíabiológicaenlaArgentina”,RUNA20,pp.35-67.1997.“Argentina”,enHistory of Physical Anthropology. An Encyclopedia, ed.FrankSpencer,pp.101-107.
NuevaYorkyLondres,GarlandPublishing,Inc.
Carnese, Francisco R., Alicia Caratini y Alicia S. Goicoechea2002.“InterethnicRelationsinNative-AmericanPopulationsofArgentinePatagonia:AGenetic
DemographicAnalysis”,enContemporary Perspectives on the Native Peoples of Pampa, Patagonia, and Tierra del Fuego, eds.ClaudiaBrionesy,JoséLuisLanata,pp.121-134Londres,Bergin&Garvey.pp121-134
Carnese, Francisco R. y Héctor M. Pucciarelli2007.“InvestigacionesantropobiológicasenArgentina,desdeladécadade1930hastalaactualidad”,
Relaciones de la Sociedad Argentina de AntropologíaXXXII,pp.243-280.
Carnese, Francisco R., Alicia S. Goicoechea, Sergio A. Avena, María Laura Parolín y Cristina B. Dejean
2008.“Diversidadgenéticaenindígenasargentinos”,XCongresodelaAsociaciónLatinoamericanadeAntropologíaBiológica,LaPlata,Argentina.20al23deoctubre.
Carnese, Francisco R., Fanny Mandisco, Christine Keyser, Cristina B. Dejean, Jean-Michel Dogoujon, Bertrand Ludes y Eric Crubézy
2010.“PaleogeneticalStudyofPre-ColumbianSamplesfromPampaGrande(Salta,Argentina)”,American Journal of Physical Anthropology141(3),pp.452-462.
Castro de Guerra, Dinorah 1997.“Venezuela”,enHistory of Physical Anthropology. An Encyclopedia, Ed.FrankSpencer,
pp.1087-1089.NuevaYorkyLondres,GarlandPublishing,Inc.
Catanesi, Cecilia, Noelia Tourret, Francisco R. Carnese y Lidia Vidal-Rioja2006.“SouthAmericanAmerindianDiversityatthevWASTRLocus”,Journal of Basic & Applied
Genetics 17(1),pp.35-40.
Cerna, Marie, Michela Falco, Horacio Friedman, Eduardo Raimondi, Armando Maccagno, Marcelo Fernández-Viña y Peter Stastny
1993.“DifferencesinHLAClassIIAllelesofIsolatedSouthAmericanIndianPopulationsfromBrazilandArgentina”,Human Immunology 37,pp.213-220.
Crawford, Michael H. 1992.Antropología biológica de los indios americanos.Madrid,Ed.Mapfre.
Crosset ti, Shaiane, Darío Demarchi, Paulo Raimann, Francisco M. Salzano, Mara H. Hutz y Sidia Callegari-Jacques
2008.“AutosomalSTRGeneticVariabilityintheGranChacoNativePopulation:HomogeneityorHeterogeneity?”,American Journal of Human Biology 20,pp.704-711.

4 9
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.29-54
L O S A P O R T E S D E L A B I O A N T R O P O L O G Í A | F R A N C I S C O R A Ú L C A R N E S E
Demarchi, Darío, Graciela Panzet ta-Dutari, Sonia Colantonio y Alberto Marcellino 2001.“Absenceofthe9-bpdeletionofMitochondrialDNAinPre-HispanicInhabitantsofArgentina”,
Human Biology73(4),pp.575-582.
Demarchi, Darío y R. John Mitchell2004.“GeneticStructureandGeneFlowinGranChacoPopulationsofArgentina:Evidencefrom
Y-chromosomeMarkers”,Human Biology76(3),pp.413-429.
Derenko, Miroslava V., Tomasz Grzybowski, Boris A. Malyarchuk, Jakub Czarny, Danuta Miscicka-Sliwka e Ilia A. Zakharov
2001.“ThePresenceofMitochondrialHaplogroupsXinAltaiansfromSouthSiberia”,American Journal of Human Genetics 69,pp.237-241.
Dornelles, Claudia H., Jacqueline Bat tilana, Nelson Fagundes, Loreta B. Freitas, Sandro L. Bonatto y Francisco M. Salzano
2004.“MitochondrialDNAandAluInsertionsinaGeneticallyPeculiarPopulation:TheAyoreoIndiansofBoliviaandParaguay”,American Journal of Human Biology 16,pp.479-488.
Etcheverry, Raúl, Carlos Guzmán, Carlos Regonesi, V. Tonkin, R. M. Urra y Norma Durán 1970.“GrupossanguíneosA-B-Oenlasmomiasprecolombinasdelosindígenasatacameños”,Revista
de Medicina de Chile 98,pp.277-282.
Fagundes, Nelson, Ricardo Kanitz, Roberta Eckert, Ana Valls, Mauricio Bogo, Francisco M. Salzano, David Smith, Wilson Silva Jr., Marco Zago, Andrea Ribeiro dos Santos, Sidney Santos, María Petzl-Erler y Sandro Bonatto
2008.“AReevaluationoftheNativeAmericanmtDNAGenomeDiversityandItsBearingoftheModelsofEarlyColonizationofBeringia”,PloS One 3(9),pp.e3157.
Furuhata, Tanemoto, Hachiro Nakajima, Eiichiro Ishid, Seiichi Izumi, Kazuo Terrada y Yoshitaro Amano
1959.“BloodGroupDeterminationsofPeruvianMummies”,Proceedings of Japan Academy35,pp.305-306.
García-Bour, Jaume, Alejandro Pérez-Pérez, Sara Álvarez, Eva Fernández, Ana M. López-Parra, Eduardo Arroyo-Pardo y Daniel Turbón
2004. “EarlyPopulationDifferentiationinExtinctAboriginesfromTierradelFuego-Patagonia:AncientmtDNASequencesandY-ChromosomeSTRCharacterization”,American Journal of Physical Anthropology123,pp.361-370.
Gilbrey, B. E. y M. Lubran 1952.“BloodGroupsofSouthAmericanIndianMummies”,Man 52,pp.115-117.
Ginther, Charles, Daniel Corach, R. Penacino, Jorge Rey, Francisco R. Carnese, Mara H. Hutz, A . Anderson, J. Just, Francisco M. Salzano y Marie C. King
1993.“GeneticVariationAmongtheMapucheIndiansfromthePatagonianRegionofArgentina:MitochondrialDNASequenceVariationandAlleleFrequenciesofSeveralNuclearGenes”,enDNA Finger-printing: State of the Science, eds.SergioPena,R.Chakraborty,J.EpplenyA.Jeffreys, pp.211-219.Basel,BirkhauserVerlag.
Glick, Thomas F. 1989.Darwin y el darwinismo en el Uruguay y en América Latina.Montevideo,Uruguay,Ed.UniversidaddelaRepública,FacultaddeHumanidadesyCiencias,DepartamentodePublicaciones.
Goicoechea, Alicia S., Francisco R. Carnese, Cristina B. Dejean, Sergio A. Avena, Tania A. Weimer, María H. L. P. Franco, Sidia M. Callegari-Jacques, Alberto Estalote, M. L. Simões, Marcos Palatnik y Francisco M. Salzano
2001.“GeneticRelationshipsbetweenAmerindianPopulationsofArgentina”,American Journal of Physical Anthropology115,pp.133-143.

5 0
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.29-54
González, José R., M. C. Bortolini, F. R. Santos y S. L. Bonatto 2008.“ThePeoplingofAmerica:CraniofacialShapeVariationonaContinentalScaleandItsInterpretation
fromanInterdisciplinaryView”,American Journal of Physical Anthropology 137,pp.175-187.
Greenberg, Joseph H., Christy Turner y Stephen L. Zegura 1986.“TheSettlementoftheAmericas:AComparisonoftheLinguistic,Dental,andGeneticEvidence”,
Current Anthropology 27,pp.477-497.
Gusinde, Martin1990.Los indios de Tierra del Fuego.BuenosAires,CentroArgentinodeEtnologíaAmericana.
Haas, Emilio, Francisco M. Salzano, H. Araujo, F. Grossman, A. Barbetti, Tania A. Weimer, María H. L. P. Franco, L. Verruno, O. Nasif, V. Morales y R. Arienti
1985.“HLAAntigensandotherGeneticMarkersintheMapucheIndiansofArgentina”,Human Heredity 35,pp.306-313.
Henning, Willi
1968. Elementos de una sistemática fi logenética. Buenos Aires, Editorial Universitaria de Buenos Aires (EUDEBA).
Horai, Satoshi, Rumi Kondo, Kumiko Murayama, Seiji Hayashi, Hiroko Koike, Nobuyuki Nakai, K. A . Joysey, G. A. Dover y Svante Päabo
1991.“PhylogeneticAffiliationofAncientandContemporaryHumansInferredfromMitochondrialDNA”,Philosophical Transactions of the Royal Society B: Biological Science333,pp.409-418.
Hrdlicka, Aleš 1912.Early Man in South America.WashingtonBureauofAmericanEthnologyBulletins.Reprint
ServicesCorporation.
Hubbe, Mark, Walter Neves y Katerina Harvati 2010.“TestingEvolutionaryandDispersionScenariosfortheSettlementoftheNewWorld”,PLoS ONE
5:e11105.doi:10.1371/journal.pone.0011105
Hutz, Mara H., Sidia M. Callegari-Jacques, Sabrina Almeida, Talita Armborst y Francisco M. Salzano
2002.“LowLevelsofSTRPVariabilityAreNotUniversalinAmericanIndians”,Human Biology 74, pp.791-806.
Imbelloni, José1937.“Razashumanasygrupossanguíneos”,Relaciones de la Sociedad Argentina de Antropología1,
pp.23-49.
Johnson, Michele J., Douglas C. Wallace, Stephen D. Ferris, Mario C. Rat tazzi y Luca Cavalli-Sforza
1983.“RadiationofHumanMitochondrialDNATypesAnalyzedbyRestrictionEndonucleaseCleavagePatterns”,Journal of Molecular Evolution19,pp.255-271.
Karafet, Tatiana M., Stephen L. Zegura, Olga Posukh, Ludmila Osipova, A. Bergen, J. Long, D. Goldman, W. Klitz, Shinji Harihara, Peter de Knijf f, V. Wiebe, Robert C. Grif f iths, Alan R. Templeton y Michael F. Hammer
1999.“AncestralAsianSource(S)oftheNewWorldY-chromosomeFounderHaplotypes”,American Journal of Human Genetics64,pp.817-831.
Kohlrausch, Fabiana, Sidia Callegari-Jacques, Luiza Tsuneto, María Petz-Erler, Kim Hill, A . Magdalena Hurtado, Francisco M. Salzano y Mara H. Hutz
2005.“GeographyInfluencesMicrosatellitePolymorphismDiversityinAmerindians”,American Journal of Physical Anthropology125,pp.463-470.
ˇ

5 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.29-54
L O S A P O R T E S D E L A B I O A N T R O P O L O G Í A | F R A N C I S C O R A Ú L C A R N E S E
Lacerda F. y R. Peixoto 1876.“ContribuiçõesparaoestudoantropológicodasraçasindígenasdoBrazil”,Archivos do Museu
Nacional do Rio de Janeiro1,pp.47-79.
Lalueza, Carles, Alejandro Pérez-Pérez, Eva Prats, Pons J. Moreno P. y Daniel Turbón1993-94.“Ausenciadeladeleciónde9bpCOII/tRNALysenaborígenesdeFuego-Patagoniamediante
análisisdeADNantiguo”,Anales del Instituto de la Patagonia22,pp.181-191.
Lalueza, Carles, Alejandro Pérez-Pérez, Eva Prats, Luis Cornudella y Daniel Turbón 1997.“LackofFoundingAmerindianMitochondrialDNALineagesinExtinctAboriginesfromTierradel
Fuego-Patagonia”,Human Molecular Genetics6,pp.41-46.
Layrisse, Miguel, Tulio Arends y Rafael Domínguez Sisco 1955.“Nuevogruposanguíneoencontradoendescendientesdeindios”,Acta Médica Venezolana 3,pp.132.
Lewontin, Richard C.1972.“TheApportionmentofHumanDiversity”,Evolutionary Biology6,pp.381-98.
Lin, Z., T. Kondo, T. Minamino, E. Sun, G. Liu y T. Ohshima 1996.“GenotypingofABOBloodGroupSystembyPCRandRFLPonMummiesDiscoveredat
TaklamakanDesertin1912”,Nihon Hoigaku Zasshi 50(5),pp.336-342.
Lippold, Lois1971.“TheMixedCellAglutinationMethodforTypingMummifiedHumanTissue”, American Journal of
Physical Anthropology 34,pp.377-383.
Llop, Elena y Francisco Rothhammer 1988.“ANoteonthePresenceofBloodGroupsAandBinPre-ColumbianSouthAmerica”,American
Journal of Physical Anthropology74(1),pp.107-111.
Marrero, Andrea, Wilson Silva-Junior, Claudio M. Bravi, Mara H. Hutz, María Petzl-Erler, Andrés Ruiz-Linares, Francisco M. Salzano y María C. Bortolini
2007.“DemographicandEvolutionaryTrajectoriesoftheGuaraniandKaingangNativesofBrazil”,American Journal of Physical Anthropology132,pp.301-310.
Matson, Albin G. 1934.“AProcedureforDeterminingDistributionofBloodGroupsinMummies”,Proceedings of the
Society for Experimental Biology and Medicine31,pp.964-968.
Matson, Albin G., Eldon Sutton, Raul Etcheverry, Jane Swanson y Abner Robinson1967.“DistributionofHereditaryBloodGroupsamongIndiansinSouthAmerica.IV.Chile”,American
Journal of Physical Anthropology 27,pp.157-193.
Matson, Albin G., Eldon Sutton, Jane Swanson y Abner Robinson1969.“DistributionofHereditaryBloodGroupsamongIndiansinSouthAmerica.VII.InArgentina”,
American Journal of Physical Anthropology 30,pp.61-83.
Mayr, Ernst1968.Especies animales y evolución.SantiagodeChile,Ed.UniversidaddeChile.
Mazza, Salvador e Isabel Franke1927.“Grupossanguíneosdeindiosydeautóctonosdelnorteargentino(Notapreliminar)”,Boletín del
Instituto de Clínica Quirúrgica3,pp.137-140.1928.“Grupossanguíneosdeindiosydeargentinos”,La Prensa Médica Argentina15,pp.607-610.
Merriwether, D. Andrew, Francisco Rothhammer y Robert Ferrell 1994.“GeneticVariationintheNewWorld:AncientTeeth,BoneandTissueasSourcesofDNA”,
Experientia50,pp.411-430.

5 2
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.29-54
1995.“DistributionoftheFourFoundingLineageHaplotypesinNativeAmericansSuggestsaSingleWaveofMigrationfortheNewWorld”,American Journal of Physical Anthropology98,pp.411-430.
Monsalve, María Victoria, F. Cárdenas, F. Guhl, A . D. Delaney y D. V. Devine 1996.“PhylogeneticAnalysisofmtDNALineagesinSouthAmericanmummies”,Annals of Human
Genetics60,pp.293-303.
Moraga, Mauricio, Paola Rocco, Juan F. Miquel, Flavio Nervi, Elena Llop, Ranajit Chakraborty, Francisco Rothhammer y Pilar Carvallo
2000.“MitochondrialDNAPolymorphismsinChileanAboriginalPopulations:ImplicationsforthePeoplingoftheSouthernConeoftheContinent”,American Journal of Physical Anthropology 113,pp.19-29.
Moraga, Mauricio, Eugenio Aspillaga, Calogero M. Santoro, Vivien G. Standen, Pilar Carvallo y Francisco Rothhammer
2001.“AnálisisdeADNmitocondrialenmomiasdelnortedeChileavalahipótesisdeorigenamazónicodepoblacionesandinas”,Revista Chilena de Historia Natural74,pp.719-726.
Moraga, Mauricio, Calogero M. Santoro, Vivien G. Standen, Pilar Carvallo y Francisco Rothhammer
2005.“MicroevolutioninPrehistoricAndeanPopulations:ChronologicmtDNAVariationintheDesertValleysofNorthernChile”,American Journal of Physical Anthropology127,pp.170-181.
Nagel, Ronald y Raúl Etcheverry1963.“TypesofHaptoglobinsinAraucanianIndiansofChile”,Nature197,pp.187-188.
Neves, Walter y Héctor Pucciarelli 1990.“TheOriginofFirstAmericans:AnAnalysisBasedontheCranialMorphologyofEarlySouth
AmericanHumanRemains”,American Journal of Physical Anthropology81,pp.274.1991.“MorphologicalAffinitiesoftheFirstAmericans:AnExploratoryAnalysisBasedonEarlySouth
AmericanHumanRemains”,Journal of Human Evolution 21,pp.261-273.
Otten, Charlot te y Lynn Flory 1963.“BloodTypingofChileanMummyTissue:ANewApproach”,American Journal of Physical
Anthropology 21,pp.283-285.
Pagés Larraya, Fernando, Norma Contardi y Elsa Servy1978.“MarcadoresgenéticosdelapoblaciónaborigendelChacoargentino”,Revista del Instituto de
Antropología 6,pp.217-241.
Pando, Marcelo1998.“EstudiomoleculardeladistribucióndealelosHLA-DR-DQenpoblacionesnormalesargentinas
yenpacientesconhepatitiscrónicaautoinmune”.TesisDoctoral,FacultaddeCienciesExactasyNaturales,UniversidaddeBuenosAires.
Parolín, Maria L. y Francisco R. Carnese 2009.“HLA-DRB1AllelesinfourAmerindianPopulationsfromArgentinaandParaguay”,Genetics and
Molecular Biology32(2),pp.212-219.
Pereira, M., J. A . Rohr, Y. Lengyel y O. Barreto1984.“OsgrupossanguíneosABOemesqueletospré-históricosdeaborígenesdaIlhaSantaCatarina,
Brazil”,Ciência e Cultura36,pp.1597-1599.
Pucciarelli, Héctor M.1989.“Contribuciónalconceptodeantropologíabiológica”,Revista de Antropología7,pp.27-31.
Reig, Osvaldo1984.“Lateoríadelaevoluciónaloscientoveinticincoañosdelaapariciónde‘Elorigendelas
especies’”,Boletín de la Academia Nacional de Medicina62,pp.545-586.

5 3
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.29-54
L O S A P O R T E S D E L A B I O A N T R O P O L O G Í A | F R A N C I S C O R A Ú L C A R N E S E
Relethford, John H. 1994.“CraniometricVariationamongModernHumanPopulations”,American Journal of Physical
Anthropology95(1),pp.53-62.
Ribeiro dos Santos, Andrea K., Sidney E. Santos, Ana Machado, Vera Guapindaia y Marco A. Zago
1996.“HeterogeneityofMitochondrialDNAHaplotypesinPre-ColumbianNativesoftheAmazonRegion”,American Journal of Physical Anthropology101,pp.29-37.
Rodríguez-Delfin, Luis, Sidney E. Santos y Marco A. Zago 1997.“DiversityoftheHumanYChromosomeofSouthAmericanAmerindians:AComparisonwith
Blacks,Whites,andJapanesefromBrazil”,Annals of Human Genetics61,pp.439-48.
Rogan, Peter K. y Joseph J. Salvo 1990.“MolecularGeneticsofPre-ColumbianSouthAmericanMummies”,UCLA Symposia on Molecular
and Cellular Biology 122,pp.223-234.
Rothhammer, Francisco y Eugenio Aspillaga 1997.“Chile”,enHistory of Physical Anthropology. An Encyclopedia, ed.FrankSpencer,pp.271-272.
NuevaYorkyLondres,GarlandPublishing,Inc.
Rothhammer, Francisco, Mauricio Moraga, Mario Rivera, Calogero M. Santoro, Vivien G. Standen, Federico García y Pilar Carvallo
2003.“AnálisisdeADNmtderestosesqueletalesdelsitioarqueológicodeTiwanakuysurelaciónconelorigendesusconstructores”,Chungara 35(2),pp.269-274.
Ruiz-Linares, Andrés, Daniel Ortiz-Barrientos, Mauricio Figueroa, Natalia Mesa, Juan G. Múnera, Gabriel Bedoya, Iván Vélez, Luis F. García, Anna Pérez-Lezaun, Jaume Bertranpetit , Marcus Feldman y David B. Goldstein
1999.“MicrosatellitesProvideEvidenceforYChromosomeDiversityamongtheFoundersoftheNewWorld”,Proceedings of the National Academy of Sciences of the United States of America96,pp.6312-6317.
Sala, Andrea, Gustavo Penacino, Francisco R. Carnese y Daniel Corach1999.“ReferenceDatabaseofHypervariableGeneticMarkersofArgentina:ApplicationforMolecular
AnthropologyandForensicCasework”,Electrophoresis20,pp.1733-1739.
Salaberry, María T., Alicia S. Goicoechea, Cristina. B. Dejean y Francisco R. Carnese 1999.“Grupossanguíneosenrestosmomificadosdeunapoblaciónprehispánica:LasPirguas(Salta,
Argentina)”,Revista Argentina de Antropología Biológica2(1),pp.267-280.
Salzano, Francisco M.1997.“Brazil”,enHistory of Physical Anthropology. An Encyclopedia,ed.FrankSpencer,pp.207-213.
NuevaYorkyLondres,GarlandPublishing,Inc.2002.“MolecularvariabilityinAmerindians:WidespreadbutUnevenInformation”,Anais da Academia
Brasileira de Ciências74(2),pp.223-263.
Salzano, Francisco M. y Sidia Callegari-Jacques 1988.South American Indians. A Case Study in Evolution.Oxford,ClarendonPress.
Sans, Mónica1997.“Venezuela”,enHistory of Physical Anthropology. An Encyclopedia, ed.FrankSpencer,
pp.1081-1083.NuevaYorkyLondres,GarlandPublishing,Inc.
Shinoda, Ken-ichi, Noboru Adachi, Sonia Guillén e Izumi Shimada 2006.“MitochondrialDNAAnalysisofAncientPeruvianHighlanders”,American Journal of Physical
Anthropology131,pp.98-107.

5 4
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.29-54
Spencer, Frank 1997.History of Physical Anthropology. An Enciclopedia.NuevaYorkyLondres,GarlandPublishing.
Thieme, Frederick P. y Charlot te M. Otten1957.“TheUnreliabilityofBloodTypingAgedBone”,American Journal of Physical Anthropology15,
pp.387-397.
Torroni, Antonio, Theodore G. Schurr, Chi-Chuang Yang, Emöke J. Szathmary, Robert Williams, Moses Schanfield, Gary Troup, William C. Knowler, Dale Lawrence, Kenneth M. Weiss y Douglas C. Wallace
1992.“NativeAmericanMitochondrialDNAAnalysisIndicatesthattheAmerindandtheNadenePopulationsWereFoundedbyTwoIndependentMigrations”,Genetics130(1),pp.153-162.
Torroni, Antonio, Theodore G. Schurr, M. Cabell, Michael Brown, James Neel, M Larsen, D. Smith, Carlos Vullo y Douglas Wallace
1994.“AsianAffinitiesandContinentalRadiationoftheFourFoundingNativeAmericanmtDNAs”,American Journal of Human Genetics53,pp.563-590.
Toscanini, Ulises, Leonor Gusmao, Gabriela Berardi, Antonio Amorim, Ángel Carracedo, Antonio Salas y Eduardo Raimondi
2008.“YChromosomeMicrosatelliteGeneticVariationinTwoNativeAmericanPopulationsfromArgentina”,Forensic Science international. Genetics2(4),pp.274-280.
Tsuneto, Luisa, Christian Probst, Mara H. Hutz, Francisco M. Salzano, Luis Rodríguez-Delfin, Marco Zago, Kim Hill, Magdalena A. Hurtado, Andrea Ribeiro dos Santos y María Petzl-Erler
2003.“HLAClassIIDiversityinSevenAmerindianPopulations.CluesabouttheOriginsoftheAché”,Tissue Antigens 62,pp.512-526.
Wallace, Douglas C., Katherine Garrison y William C. Knowler 1985.“DramaticFounderEffectsinAmerindianMitochondrialDNAs”,American Journal of Physical
Anthropology68,pp.149-155.
Ward, Ryk H., Barbara L. Frazier, Kerry Dew-Jager y Svante Pääbo 1991.“ExtensiveMitochondrialDiversitywithinaSingleAmerindianTribe”,Proceedings of the National
Academy of Sciences of the United States of America88,pp.8720-8724.
Ward, Ryk H., Francisco M. Salzano, Sandro Bonatto, Mara H. Hutz, C. Coimbra Jr. y Ricardo Santos
1996.“MitochondrialDNAPolymorphisminThreeBrazilianIndianTribes”,American Journal of Human Biology8,pp.317-323.
Wyman, Leland C. y William C. Boyd1937.“BloodGroupDeterminationsofPrehistoricAmericanIndians”,American Anthropologist39,
pp.583-592.

5 5
A DEPTOS A L A A DA PTACIÓN: TR ES PROPU ESTA S CL Á SICA S PA R A L A A RQU EOLOGÍ A Y U NA EVA LUACIÓN
ViVian scheinsohn*
[email protected] Nacional de Investigaciones Científicas y Técnicas (CONICET), Instituto Nacional de Antropología y Pensamiento Latinoamericano (INAPL), Argentina
R e s u m e n A más de treinta años del artículo fundacional
de Kirch (1980), este trabajo se propone investigar el papel
del concepto de adaptación en el pensamiento arqueológico
contemporáneo y evaluar su utilización. Para esto, se revisa cómo
fue utilizado este concepto en biología, cómo se dio su uso en
arqueología, enmarcado dentro del procesualismo, seleccionismo
y la teoría de la herencia dual, y se evalúa su papel actual.
P A L A B R A S C L A V E :
Adaptación, arqueología, procesualismo, seleccionismo, Teoría de la
Herencia Dual.
Artículorecibido:31deoctubrede2011|aceptado:10denoviembrede2011|modificado:20denoviembrede2011
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,294pp.ISSN1900-5407,pp.55-73
* DoctoraUniversidaddeBuenosAires-UBA,Argentina.

5 6
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.55-73
BelieVing in adaptation: three ClassiCal proposals for arCheology and one eValuation
ABSTR AC T Thirty years after Kirch
(1980) seminal paper, this work reviews
the role of adaptation in contemporary
archaeological thought and discusses its
use. In this process, the use of this concept
in biology will be examined as well as its
use in archaeology, as it is incorporated
in processualism, selectionism and Dual
Inheritance theories. The author concludes
with an evaluation of its current potential.
K E Y W O R D S :
Adaptation, Archaeology, Processualism,
Selectionism, Dual Inheritance Theory.
adeptos à adaptação: três propostas ClássiCas para a arqueología e uma aValiação
RESUMO A mais de trinta anos do artigo
fundacional de Kirch (1980), este trabalho
propõe-se pesquisar o papel do conceito
de adaptação no pensamento arqueológico
contemporâneo e avaliar sua utilização.
Para isto, é revisto como foi utilizado este
conceito em biologia, como se deu o seu
uso em arqueologia, moldurado dentro do
processualismo, selecionismo e a teoria da
herança dual, e avalia-se seu papel atual.
P A L A V R A S C H A V E :
Adaptação, arqueologia, processualismo,
selecionismo, Teoria da Dupla Herança.

5 7
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.55-73
A DEPTOS A L A A DA PTACIÓN: TR ES PROPU ESTA S CL Á SICA S PA R A L A A RQU EOLOGÍ A Y U NA EVA LUACIÓN
V i V i a n s c h e i n s o h n
IntroducciónAq u e l l o s s e r e s h u m a n o s que formamos parte de lo que se llama Occidente tenemos una forma particular de vernos. La “Gran cadena del ser” o Scala naturae, esa idea acuñada por los pensado-res cristianos que acomodaba toda la creación en distintos niveles jerárqui-cos (sobre esta idea y su historia, ver Lovejoy, 1960), sigue permeando nuestro pensamiento y nos sigue ubicando en un escalón superior al resto de los seres vivos. La tesis de la excepcionalidad humana (Schaeffer, 2009) postula que el ser humano es excepcional, en cuanto posee una dimensión ontológica que le per-mite trascender la realidad de otras formas de vida y su propia naturalidad. De manera concordante, y ya en un terreno científico, también nos consideramos sobresalientes en cuanto a nuestras posibilidades de adaptación. Ya sea que se hable de plasticidad fenotípica o construcción de nicho cultural, ya sea que se lo plantee en términos conductuales y genéticos o en términos estrictamente cul-turales, nadie discute la capacidad de adaptación de los seres humanos. Como lo ha dejado en claro Kirch, “Es casi pedante afirmar que la adaptación conduc-tual alcanzó un apogeo con el Homo sapiens” (1980: 105, traducción mía).
Sin embargo, y paradójicamente, el registro de fracasos y conductas maladaptativas en nuestra especie es extenso. En la vida cotidiana, nos preten-demos infalibles al partir de ilusiones que nos llevan, entre otras cosas, a cho-ques automovilísticos y equivocaciones groseras en los testimonios judiciales (ver Chabris y Simons, 2011). Con una profundidad temporal mayor, el registro arqueológico se ha encargado de demostrar muchas veces que esa maravillosa capacidad de adaptación no siempre ha sido puesta en práctica. Muchas veces los humanos han enfrentado la extinción de poblaciones antes de tener que cambiar conductas (ver algunos ejemplos en Diamond, 2004). Entonces, ¿cuál

5 8
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.55-73
es el papel del concepto de adaptación cuando se trata de estudiarnos? ¿Nece-sitamos ese concepto en arqueología? Este artículo pretende volver a esa pre-gunta, luego de transcurridos treinta años del artículo fundacional de Kirch (1980), que inauguró el uso científico del concepto de adaptación en arqueolo-gía. Para contestarla, empezaremos revisando cómo fue utilizado ese concepto en biología. Luego veremos cómo se dio su uso en arqueología, fundamental-mente a partir de tres propuestas correspondientes a sendos marcos teóricos, que consideramos clásicas. Nos limitaremos a estas propuestas en función de que: 1) en el marco del procesualismo y el seleccionismo, se publicaron traba-jos que se dedicaron específicamente al tema de la adaptación y 2) son estos marcos teóricos los que formularon conceptos de adaptación acordes con el evolucionismo darwiniano. Por ende, dejaremos de lado aquellas propuestas relacionadas con lo que se conoce como evolucionismo cultural. Finalmente, evaluaremos el potencial actual del concepto de adaptación.
El concepto de adaptación en biologíaNo es fácil definir qué es exactamente adaptación, a pesar de que es uno de los conceptos claves en teoría evolutiva (ver, entre otros, Ridley, 1997, y Hull y Ruse, 1998). En principio, es un concepto polisémico. En Origin of Species, Darwin describe la adaptación como un tipo de relación entre organismos:
[…] the structure of every organic being is related, in the most essential yet often hidden manner, to that of all other organic beings, with which it comes into competition for food or residence, or from which it has to escape, or on which it preys. (Darwin, 1859: 77, citado por Kampourakis, 2011)
Con posterioridad, la palabra adaptación fue usada para referirse tanto a un proceso como a un rasgo y a un estado (el de estar adaptado). West-Eberhard (1992) alude a esas primeras dos acepciones, en cuanto considera como adaptación
[…] a characteristic of an organism whose form is the result of selection in a particular functional context […]. Accordingly, the process of ‘adaptation’ is the evolutionary modification of a character under selection for efficient or advantageous (fitness-enhancing) functioning in a particular context or set of contexts. (West-Eberhard, 1992: 13)
En lo que respecta a la adaptación como rasgo, no es simplemente el hecho de que ese rasgo le otorgue alguna ventaja a su poseedor lo que lo consti-tuye como tal, ya que esa ventaja puede ser resultado del azar, sino que, debido a esa ventaja, sus ancestros deben haber sido seleccionados. Entonces, la selec-

5 9
A D E P T O S A L A A D A P T A C I Ó N | V I V I A N S C H E I N S O H N
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.55-73
ción natural de un rasgo para un rol es condición necesaria y suficiente para ser una adaptación, mientras que contribuir al aumento actual de la supervivencia y reproducción no es necesario ni suficiente. Esto es lo que se ha denominado definición histórica de la adaptación (Kampourakis, 2011). Las dificultades prácticas que surgieron al intentar aplicar esta definición llevaron a ciertos autores a favorecer una definición ahistórica, que considera como adaptación a un rasgo que mejora la supervivencia y reproducción de su poseedor sin impor-tar su origen. En este caso, no importa la historia, sólo importan el diseño y el éxito reproductivo. Así, una adaptación puede ser definida como un rasgo que puede ejecutar un papel o función biológica bajo circunstancias específi-cas o como una variante fenotípica que contribuye a la supervivencia y repro-ducción de sus poseedores entre un conjunto especificado de variantes en una ambiente dado (Kampourakis, 2011). En este caso, la contribución actual a la supervivencia y reproducción es una condición necesaria y suficiente para ser una adaptación, mientras que la selección de un rasgo para un papel o función no es ni necesaria ni suficiente (Kampourakis, 2011). Nótese también aquí una distinción importante, que es la diferencia entre adaptación (o adaptedness) y fitness. Los estados alternativos de una adaptación son la causa de los cambios evolutivos a través de sus diferencias en la interacción organismo-ambiente. Las diferencias adaptativas entre fenotipos alteran sus parámetros demográ-ficos. Estos parámetros son los componentes de lo que la genética de pobla-ciones denominó fitness, es decir, el éxito reproductivo de poblaciones enteras de genotipos. Adaptación y fitness1, entonces, son conceptos relacionados pero no son lo mismo, aunque Darwin y los primeros evolucionistas los utilizaban como sinónimos. El uso de fitness cambió a medida que se pudo modelar mate-máticamente el concepto de éxito reproductivo. Pero, en síntesis, hoy, adapta-ción es la causa y fitness es su resultado intrageneracional (Watt, 2001).
Gould y Lewontin (1979) fueron los primeros en plantear una serie de críticas a aquellos que le daban a la adaptación un peso excluyente en la his-toria evolutiva. Para ello elaboraron el concepto de programa adaptacionista o paradigma panglossiano, en alusión al doctor Pangloss, personaje de la novela Cándido de Voltaire, quien sostenía que vivimos en el mejor de los mundos posibles. En síntesis, la crítica al programa adaptacionista apunta a que bajo este programa 1) no se analiza a los organismos como un todo integrado y, por lo tanto, no se tienen en cuenta las constricciones internas dadas por la heren-cia filética y/o el desarrollo; 2) se relaciona la utilidad actual con la explicación
1 Comoesdifícildiferenciarestos términosenespañol,yaqueambospuedentraducirsecomoadaptaciónoajuste,utilizaréadaptaciónparalacausa,yparaeléxitoreproductivo,eltérminoinglésfitness.

6 0
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.55-73
del origen de cada rasgo; 3) no se consideran las alternativas a la selección natu-ral; 4) la sola plausibilidad es suficiente como para aceptar historias adaptativas (para detalles, ver Ginnobili y Blanco, 2007).
Unos años después, Gould y Vrba (1982) proponen una redefinición del concepto de adaptación. A partir de las dos definiciones que hemos mencionado (la histórica, que considera la historia del rasgo, y que por lo tanto lo vincula con la selección natural, y la ahistórica, que define adaptación en un sentido inme-diato como cualquier rasgo que mejora el éxito reproductivo de un organismo sin importar su origen histórico), estos autores consideran que existen rasgos que terminan siendo adaptativos, aunque no fueron diseñados en primera instan-cia para esa función, y que éstos no encajarían en ninguna de esas definiciones. A partir de la etimología de adaptación (del latín ad y aptus, que quiere decir “hacia un fin para un rol particular”), hablan de aptation (que podríamos traducir con un neologismo como “aptación”, o adecuación2), para considerar como tal al fenómeno general de ser adecuado (being fit) y diferencian entre adaptación (carácter diseñado para cumplir una función por selección natural) y exaptación (carácter diseñado para una determinada función que es cooptado para cumplir otra o que es no adaptativo por no ser originado por selección natural). Así, la selección natural no es el único proceso que lleva a la adecuación o “aptación”, definida como el fenómeno de estar adaptado, es decir, la definición ahistórica de adaptación, y que incluiría la adaptación y la exaptación (Gould y Vrba, 1982).
Existen tres maneras en las que puede ocurrir la adecuación (Ginnobili y Blanco, 2007): la genética (adaptación propiamente dicha); la fisiológica, debida a la plasticidad fenotípica que permite al organismo en su desarrollo ontogénico moldearse según su ambiente, y la cultural, que, según estos autores, tampoco surgiría por selección natural (aunque hay autores que sostienen otra posición; ver más adelante). Lo que es clave en este panorama es que no todas las adecua-ciones o “aptaciones” son óptimas.
El concepto de adaptación, la antropología y la arqueologíaEn lo que sigue de este artículo me concentraré en el concepto de adaptación tal como fue usado en la arqueología contemporánea. Sin embargo, me veré en la obligación de hacer referencias a cómo las ideas evolucionistas se trasladaron al campo antropológico, debido a que son estas ideas las que van a influenciar el desarrollo del evolucionismo en la arqueología.
2 Sensu Ginnobili (2009), aunque este autor considera diferencias respecto de la definición ahistórica deadaptación.

6 1
A D E P T O S A L A A D A P T A C I Ó N | V I V I A N S C H E I N S O H N
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.55-73
Se puede decir que incluso antes de Darwin ya había una idea de evolu-ción, al menos implícita, en el ámbito de la arqueología. Esta noción de evo-lución estaba más relacionada con las ideas propuestas por el inglés Herbert Spencer, y que son radicalmente diferentes a las de Darwin (ver Scheinsohn, 2001). Las diferencias entre el evolucionismo darwiniano y spenceriano que-dan claras en el siguiente párrafo:
Spencer’s doctrine, not having resulted from any kind of sustained empirical enquiry, was explicitly deductive in its structure, and rested on the metaphy-sical supposition that all evolutionary change was due to the persistence of an immanent “Power” that was […] both “unknown and unknowable.” In marked contrast, Darwin’s theory, as he published it in 1859, was authentically scien-tific, for, without recourse, for all practical purposes, to metaphysics or “final causes,” it postulated, on the basis of massive factual evidence, a non-teleo-logical mode of evolutionary change and incorporated a precise definition of the mechanism of natural selection which (as has since been conclusively demonstrated) does indeed result in the genetic evolution of populations of living organisms. (Freeman, 1974: 215)
En definitiva la evolución spenceriana es unilineal, direccional y progre-siva y opuesta a la darwiniana, en cuanto ésta es multilineal, no direccional y no progresiva.
Los trabajos del norteamericano Lewis Morgan y del inglés Edward Tylor se enmarcaron dentro de esta evolución spenceriana. Y los hallazgos arqueoló-gicos de la época, respecto de la antigüedad humana, parecían encajar en ella, por lo que la arqueología incorporó, amplió y defendió ese evolucionismo neo-lamarckiano (según la definición de Freeman, 1974), dando sustento a la idea de que el progreso humano era una ley de la naturaleza, y que todos los gru-pos humanos pasaban por estadios similares de desarrollo. El evolucionismo antropológico transfirió las desigualdades humanas del terreno de lo político al terreno de lo natural, interpretándolas como consecuencia de las diferencias biológicas y, finalmente, dando origen a las primeras doctrinas racistas de corte científico (Scheinsohn, 2001). Esta asociación entre evolucionismo y “darwi-nismo social” condujo finalmente al rechazo de los enfoques darwinistas dentro de las Ciencias Sociales y Humanas, durante la primera mitad del siglo. A fines del siglo XIX, además, la arqueología se vio asociada con el nacionalismo, al ser utilizada para reafirmar la historia nacional. Lentamente se fue dejando de lado el evolucionismo unilineal, y los cambios detectados en el registro arqueológico ya no eran atribuidos a la evolución de los pueblos, sino a la difusión de carac-teres culturales que se producían en ciertos centros de invención. Comenzaban a dominar el difusionismo y su corolario, el enfoque histórico-cultural. Así, el

6 2
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.55-73
supuesto de que grupos no relacionados que están al mismo nivel de desarrollo poseen similitudes culturales fue reemplazado por el de que, como las cultu-ras son estáticas, sólo la comparación con aquellas relacionadas históricamente permite interpretar el registro arqueológico. Sólo los trabajos del australiano Vere Gordon Childe propiciaron una perspectiva evolucionista multilineal, inmersa en su militancia socialista y pensamiento marxista. En la década del sesenta surge un grupo de antropólogos que moderniza esos primeros plan-teos evolucionistas, aunque siempre dentro de la misma matriz direccionista y progresiva de corte spenceriano. Los autores más representativos de esta ten-dencia son los norteamericanos Leslie White y Julián Steward. Ambos auto-res plantearon su perspectiva, en primer lugar, diferenciando el evolucionismo cultural del biológico y, en segundo lugar, apelando a conceptos procedentes de la Teoría de los Sistemas y de la Ecología (Scheinsohn, 2001). Se puede decir entonces que el concepto de adaptación entra al campo de la cultura a partir de Leslie White, al considerar que la cultura es el medio de adaptación extrasomá-tico humano (White, 1959). Para White la evolución cultural era unilineal, y la adaptación al ambiente se efectuaba al aumentar la eficiencia termodinámica. Steward (1955) aportó un concepto de adaptación cultural más moderno al sostener una evolución multilineal. La evolución era entonces concebida en términos ecológicos como la adaptación de una cultura a un ambiente especí-fico (Kirch, 1980). La convergencia de las perspectivas ecológicas y evolutivas dio lugar a un concepto de cultura distintivo: el de un sistema que relaciona las poblaciones humanas con su matriz ambiental, es decir, el concepto adaptativo de cultura (Kirch, 1980). Si bien, como señaló Kirch (1980), el marco de referen-cia adaptacionista despertó en antropólogos y arqueólogos un inmediato atrac-tivo, los principios fundamentales del proceso adaptativo no fueron definidos (Kirch, 1980). Es la Nueva Arqueología la que comienza a aplicar estos concep-tos en un marco arqueológico. Binford comenzó a formular una aproximación arqueológica al estudio de la adaptación cultural (Kirch, 1980), mientras que su propuesta programática de focalizarse en la variabilidad vino a representar en arqueología el equivalente del pensamiento poblacional en biología (Mayr, 1963), aunque con un cierto desfasaje temporal. A partir de las propuestas de Binford se empieza a hablar de un paradigma sistémico adaptativo, que fue considerado como equivalente del procesualismo.
La propuesta procesualEs Kirch (1980) quien definirá el uso del concepto de adaptación en ese marco, y por ello nos concentraremos en su trabajo. Para Kirch la adaptación cultural consiste en “the continuous modification of those behavioral patterns in res-

6 3
A D E P T O S A L A A D A P T A C I Ó N | V I V I A N S C H E I N S O H N
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.55-73
ponse to changing environments, by means of selective retention of behavior that confers increased adaptedness upon the members of that society” (Kirch, 1980: 110). Nótese que, al hablar de selección, Kirch no diferencia si se trata de selección natural o cultural. Esto posiblemente se deba a que apuesta al acopla-miento que habría entre ambas, lo que se revela en el siguiente párrafo:
In the short run, various criteria including efficiency of energy capture, and the satisfaction of perceived needs and wants may determine the selection and retention of certain behavior. In the longer term, however, only if that behavior contributes to the persistence of the group or population in terms of reproduc-tive continuity will it be truly retained. (Kirch, 1980: 122)
Kirch va a ser quien fije el concepto de “estrategia adaptativa”, central en el marco del procesualismo: “Adaptive strategy may be broadly defined as the set of culturally transmitted behaviors (extractive, exploitative, modifying, mani-pulative, competitive, mutualistic, and the like) with which a population inte-racts or interfaces with its natural and social environment” (Kirch, 1980: 129).
Los principios fundamentales que entonces propone Kirch para el estu-dio de la conducta humana desde una perspectiva adaptativa siguen los linea-mientos que corresponden a su estudio en biología (como cuando en su trabajo determina que la variación es la fuente última de la adaptación, que la selección natural es el mecanismo causante de discriminar entre las variantes conductua-les y que, en cuanto la persistencia de un grupo o población es determinada por el éxito reproductivo, el criterio último de la selección es el fitness reproduc-tivo), y que establece como aquellos propios de los humanos:
(6) In human behavior, the mechanism for selective retention and transmis-sion of adaptive behavior is cognitive and symbolic, with important conse-quences for the rate of adaptive response.
(7) Selection of behavior operates not only upon the individual, but also at the group level.
(8) All of human behavior cannot be reduced to function at the level of inte-raction with environment, and thus not all behavior is adaptive, or explicable in an evolutionary framework. (Kirch, 1980: 130-131)
Propone entonces, para un estudio arqueológico del proceso adaptativo, seguir el siguiente procedimiento: 1) identificar un marco temporal y espacial apropiado para la secuencia adaptativa en cuestión que contemple distintos niveles jerárquicos de unidades de adaptación, 2) buscar y analizar la variación conductual representada en el registro arqueológico e intentar identificar áreas probables de stress representadas por la variabilidad (por lo que interpreta la

6 4
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.55-73
variabilidad como un indicador indirecto de las presiones selectivas, y no como aleatoria), 3) identificar presiones selectivas probables causantes del stress en el ambiente local (que incluye las condiciones naturales y humanas), 4) analizar datos paleodemográficos para obtener información relacionada con la estabili-dad poblacional, el crecimiento o declinación como medida de la tasa e intensi-dad del proceso adaptativo en cuestión, 5) formular un modelo de las relaciones entre ambiente, estrategia adaptativa y demograf ía que permita realizar predic-ciones respecto de las respuestas conductuales a presiones selectivas ambienta-les específicas, 6) generar hipótesis alternativas que puedan someterse a prueba desde ese modelo, 7) a partir de los resultados, revisar el modelo y obtener más datos para repetir el proceso hasta que puedan eliminarse las hipótesis alterna-tivas (ver en Kirch, 1980).
Esta enumeración revela a las claras la raíz procesual y sistémica de la propuesta de Kirch. En principio, por un cierto optimismo que demuestra, en cuanto a la posibilidad de obtener los datos que requiere: identificación de las presiones selectivas, datos paleodemográficos (no siempre disponibles arqueológicamente) o un marco temporal y espacial apropiado (sin definir qué es apropiado en este contexto), y la idea de que las hipótesis alterna-tivas puedan descartarse sobre la base de la obtención de más datos. Pero también el peso que le otorga a la elaboración de modelos e hipótesis. Su postura podría ligarse a la ecología del comportamiento en ciertos aspectos (i.e., la idea de que la variación conductual permite identificar áreas de stress, es decir, que la variación es guiada); pero también puede verse esbozada una postura que después se elaborará en el marco de la Teoría del Nicho Cultu-ral (ver una reciente síntesis en Kendal et al., 2011), cuando reconoce que el ambiente incluye las poblaciones humanas y poblaciones humanas vecinas, o cuando afirma:
Man is the ultimate manipulator and modifier of environments, and any envi-ronmental analysis must reflect this fact of his behavior. Fortunately, some archaeologists have already paid serious attention to environmental selection pressures that are set up as a consequence of man’s own activity […]. Archaeo-logists occupy a rather privileged intellectual niche for the study of long-term consequences of ecosystem regression under the influence of human adaptive strategies. (Kirch, 1980: 136)
El problema que se plantea es que sus procedimientos no son estricta-mente arqueológicos. Su definición de estrategia adaptativa apunta al con-texto sistémico que arqueológicamente debería reflejarse en un “static set of patterned materials reflecting paleoenvironment, paleodemography, and past

6 5
A D E P T O S A L A A D A P T A C I Ó N | V I V I A N S C H E I N S O H N
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.55-73
human activity (i.e., archaeological data in the traditional sense)” (Kirch, 1980: 131). Cuando pasamos al procedimiento que establece en siete pasos, para un estudio arqueológico del proceso adaptativo, hay muy poca precisión respecto de lo que considera stress, presiones selectivas y paleodemograf ía en términos arqueológicos. Y los da como supuestos.
Se puede decir que el impacto más profundo del trabajo de Kirch se dio en cuanto a la definición de estrategia adaptativa, citada por muchos arqueó-logos. Pero muchas veces este término fue utilizado como mera etiqueta. A modo de ejemplo, puede argumentarse lo que sostiene Politis a propósito de la arqueología latinoamericana “[…] most of what is considered ‘processual archaeology’ by its practitioners is actually culture history with more sophis-ticated analytical methods, an emphasis on paleoenvironmental data and some fashionable themes (for example risk and uncertainty, adaptive strate-gies, technological efficiency, etc.) embedded in the discussion or sometimes only appended to introductions” (Politis, 2003: 248; las cursivas son mías). En definitiva, muchas veces el uso del término estrategia adaptativa sirvió para hablar de las culturas, que postuló la escuela histórico-cultural, y ponerle, al mismo perro, otro collar. Fueron pocos los que intentaron seguir los procedi-mientos de Kirch y encarar de manera rigurosa una visión adaptativa darwi-niana en arqueología.
La propuesta seleccionistaEsta propuesta de Kirch fue seguida en el tiempo por otra formulada dentro de un marco teórico ligado al procesualismo pero separado de éste. El selec-cionismo, tal como se lo define en arqueología, apunta a la aplicación directa de la teoría evolutiva al registro arqueológico. El registro arqueológico no es ni refleja la conducta humana ni las poblaciones. Precisamente, es la ausencia de conducta lo que permite identificarlo como registro arqueológico. Se pos-tula que el fenotipo humano tiene dos componentes: el individuo (componente material) y su comportamiento. Tanto la transmisión genética como la cultural son responsables del fenotipo, ya sea en humanos o en otros animales (ver deta-lles sobre el seleccionismo en Scheinsohn, 2001).
El seleccionismo se hizo cargo del concepto de adaptación con el artí-culo de O’Brien y Holland (1992). Comienzan diciendo que “[…] anthropo-logy in general, and archaeology in particular, are not even close to having a developed set of methods for the study of human adaptation” (O’Brien y Holland, 1992: 36).
Para estos autores, en cuanto seleccionistas, la selección natural es per-tinente respecto de los humanos. Esta postura viene a contrapesar las pro-

6 6
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.55-73
puestas de la ecología del comportamiento, para las que la selección natural sólo había intervenido en el pasado al conformar la plasticidad conductual humana: “human adaptedness and adaptation do not make us immune from natural selection and that despite historically based reasons for doing so, we cannot continue to equate the creative ability of humans with some biza-rre, extraordinary means of escaping selective environments” (O’Brien y Holland, 1992: 37).
Abogan entonces a favor de una visión darwinista del registro arqueoló-gico sin ahorrar críticas al procesualismo: “Marks and Staski (1988: 151) are correct in asserting that the ‘new archaeology’ called for evolutionary thought. It may have made such a call, but apparently no one familiar with Darwinism answered” (O’Brien y Holland, 1992: 37).
La premisa principal de la que parten es que el registro arqueológico se compone de partes del fenotipo humano del pasado. Los objetos que allí se encuentran no están relacionados con los fenotipos sino que son partes de él. Sin embargo, siguiendo a Leonard y Jones (1987), los objetos arqueológicos no son unidades de reproducción, por lo que deben ser considerados en cuanto al éxito replicativo y no respecto al éxito reproductivo: “[…] replicative success does not necessarily signal that a selective advantage has been passed to the possessors of the feature, nor does it imply that the feature was under selection” (O’Brien y Holland, 1992: 46).
El éxito replicativo está a cargo de la adaptedness, entendida como el estado de estar adaptado (y, por lo tanto, equivalente a la definición ahistórica de adaptación o a lo que Gould y Vrba, 1982, llaman aptation o adecuación). Así, sostienen que, si bien los arqueólogos no pueden determinar la adecuación que se dio efectivamente (realized adaptedness), sí pueden determinar la ade-cuación potencial de un rasgo.
O’Brien y Holland (1992) se concentran en las adaptaciones, en cuanto éstas son evidencia de la selección natural, y consideran que ése es el tipo de selección presente en el registro arqueológico, descartando la selección artifi-cial (o cultural o racional). Los autores argumentan:
Humans may be selective agents, but only if they affect the adaptedness either of themselves or of other organisms. For example, animal breeders are active selective agents. And, the seemingly capricious, but in reality patterned, choice by collectors of butterflies of one color or another is as potent an agent of selection as is the choice by any bird. In both cases the butterflies meet unhappy endings, and the gene pool of which they were a part is adjusted accordingly (O’Brien and Holland, 1990: 57). But this is very different from choosing one design element over another for vessel decoration.

6 7
A D E P T O S A L A A D A P T A C I Ó N | V I V I A N S C H E I N S O H N
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.55-73
Nunca dejan en claro por qué es diferente un caso del otro y por qué consideran que las selecciones realizadas por agentes humanos no pueden afectar su adecuación.
Si bien reconocen que la selección natural es sólo una causa de evolución, y no la única, consideran que la tarea esencial del arqueólogo es determinar adaptaciones, es decir, rastrear la selección natural en el registro arqueológico. Para ello, la clave es poder determinar, entonces, cuándo estamos tratando con una adaptación, y la escala de la misma.
Cuando se trata con el registro arqueológico, primero hay que separar rasgos que afectan la adecuación (adaptedeness), de aquellos que no la afectan:
Second, archaeologists are obliged to chart the evolutionary history of a par-ticular feature in order to decide whether it has been shaped by selection, since then and only then can a trait be considered an adaptation. Determining whether or not a trait was (is) adaptive is not always easy. For example, adap-tations may be nested in a number of hierarchical units, and it is necessary first to adjust the level of analysis to the proper unit. It then may be possible to determine the potential adaptedness of a trait (or suite of traits). Enginee-ring studies are a viable means of assessing fitness, and we suggest elsewhere (O’Brien and Holland 1991) that perhaps this is the most-useful role that the field of archaeometry can play. (O’Brien y Holland, 1992: 55)
Es de rigor entonces determinar si un rasgo es selectivamente neutral o no. Precisamente, consideran que este tema llevó a las malas interpretaciones relacionadas con la cuestión de estilo y función definidas por Dunnell (1978). Éste distingue entre función y estilo en cuanto a su fitness darwiniano y su valor selectivo. El problema surge cuando se equiparan adaptación y rasgos funcionales y no-adaptación y rasgos neutrales. El aumento en el fitness se usa como criterio para la inclusión en la categoría función, mientras que la ausencia de valor selectivo se usa como criterio para la inclusión en la cate-goría neutral (estilo).
But what do we do with features that increase the adaptedness of the possessor(s) but that are not products of selection? To bring the problem into focus and to avoid problems associated with the dichotomous terms “style” and “function,” we create three categories of traits: (1) traits that are under selective control and that increase adaptedness; (2) traits that are not under selective control and that increase adaptedness; and (3) traits that are not under selective control and that do not increase adaptedness. (O’Brien y Holland, 1992: 46-47)
Los rasgos 1) son definidos como adaptaciones. Sin embargo, y aunque no les dan un nombre a los otros dos casos, el caso 2 es una definición de ade-

6 8
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.55-73
cuación o “aptación” (que incluye exaptaciones), mientras que el caso 3 serían no-adaptaciones.
Diez años después Van Pool (2002), otro autor enrolado en el seleccio-nismo, publica un trabajo sobre el concepto de adaptación retomando las pro-puestas de O’Brien y Holland (1992).
Según Van Pool (2002), los arqueólogos que se enmarcan dentro de la evo-lución darwiniana concuerdan con la definición de adaptación y las implicancias de su uso tanto como un resultado y como un proceso, pero la usan en dos esca-las diferentes. Por un lado, está la posición de la ecología del comportamiento, que considera la conducta humana como una adaptación, ya que es producto de la selección natural que la modeló para facilitar la capacidad de modificar la conducta frente a circunstancias cambiantes y obtener respuestas conductuales consistentes ante estímulos ambientales (Boone y Smith, 1998). En cambio, los seleccionistas argumentan que la selección natural crea adaptaciones a la escala de artefactos específicos y patrones de conducta. Así, los artefactos y conduc-tas son adaptaciones, y no el resultado de adaptaciones. Consideran entonces la operación de la selección natural en rasgos conductuales y la posibilidad de adaptaciones al nivel de artefactos individuales. Aquellos rasgos culturales y artefactos que afectan la supervivencia y reproducción de los humanos se verán afectados por selección natural, y con el tiempo se convertirán en adaptaciones.
Thus by demonstrating that a trait affects the survival and reproduction of humans within a particular selective context and by identifying any changes in the trait that further increases the reproductive success of the associated indivi-duals, a particular trait can be identified as an adaptation. (Van Pool, 2002: 19)
Si bien reconoce las dificultades que esta identificación implica al nivel arqueológico, no se amilana:
Still the concept of adaptation is fundamental to all archaeologists emplo-ying Darwinian evolutionary perspective because it specifies the explanatory structure necessary to provide evolutionary explanations of some phenotypic changes […] Thus, differentiating between those traits that are adaptations and those that are not is the first step in creating evolutionary explanations. (Van Pool, 2002: 20)
Igual que en el caso de O’Brien y Holland (1992), considera el análisis fun-cional como una de las formas de determinar cuándo tratamos con una adap-tación: así, propone una relación entre una forma particular de un rasgo y la supervivencia y reproducción de los individuos. A manera de ejemplo, aporta el caso de las puntas de proyectil, en cuanto constituyen una adaptación para

6 9
A D E P T O S A L A A D A P T A C I Ó N | V I V I A N S C H E I N S O H N
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.55-73
la caza favorecida por la selección natural, por sus efectos beneficiosos en los seres humanos. El análisis del diseño de la punta de proyectil permitiría deter-minar si ésta afecta la eficiencia en la caza. Luego, hay que verificar que el desa-rrollo histórico del rasgo se correlaciona con el ambiente selectivo permitiendo la creación de explicaciones de adaptación, por ejemplo, si se correlaciona con cambios en tipo de caza y disponibilidad.
Los ejemplos a los que apelan O’Brien y Holland (1992) y Van Pool (2002) son sencillos y parecen convincentes. Pero son engañosos. Prueban en todo caso la existencia de adecuaciones (“aptaciones”) y exaptaciones, pero no adaptacio-nes. En ninguno de los ejemplos que mencionan puede argumentarse un éxito reproductivo concomitante de las poblaciones humanas portadoras de los ras-gos que, según estos autores, constituyen una adaptación. El problema es que las poblaciones de artefactos y las poblaciones humanas están desacopladas. Como señaló Cullen (2000), los artefactos muestran independencia genealógica de los fenotipos humanos en los que residen, y, por lo tanto, no puede establecerse una relación uno a uno entre esas poblaciones. Los humanos hacemos artefactos, los diseñamos y creamos pero al mismo tiempo los intercambiamos y copiamos. Por lo tanto, es dif ícil evaluar el éxito reproductivo de una población humana en virtud de sus artefactos, ya que es muy dif ícil delimitar qué poblaciones usan qué artefactos. Sólo puede evaluarse el éxito replicativo de los mismos.
La propuesta de la Teoría de la Herencia DualLuego del trabajo publicado por Van Pool, ya no hay trabajos dedicados espe-cíficamente al concepto de adaptación, aunque el tema no deja de mencionarse colateralmente en artículos que tratan sobre otros temas. Esta desaparición de la adaptación posiblemente se relaciona con las posturas que se dan en la Eco-logía del comportamiento humano y en la Teoría de la Herencia Dual (Boyd y Richerson, 1985 y 2005), en donde se argumenta que la selección natural es la causa última de la conducta humana, y, por lo tanto, su papel es indirecto. En el marco de la Teoría de la Herencia Dual (Boyd y Richerson, 1985 y 2005), la cultura es considerada como una adaptación: la cultura es la causa próxima de la conducta, mientras que la selección natural es su causa última. Para estos autores, el surgimiento de la cultura implicó un nueva disyuntiva (tradeoff) evolutiva. El aprendizaje social permitió a los humanos acumular reservorios de información adaptativa durante muchas generaciones. La cultura es útil y adaptativa porque las poblaciones de mentes humanas almacenan los esfuer-zos de las mentes de generaciones previas. Como este proceso es mucho más rápido que la evolución genética, las poblaciones humanas pueden generar adaptaciones culturales a ambientes locales, una capacidad importante ante

7 0
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.55-73
el caótico y cambiante mundo del Pleistoceno. Sin embargo, el mismo meca-nismo psicológico que creó este beneficio vino con un costo implicado. Para aprovechar los beneficios del aprendizaje social, los humanos tienen que acep-tar las maneras, los modos o las formas que observan en su sociedad. Esto da lugar a la dispersión de creencias maladaptativas: la conducta de otros indi-viduos es una fuente de información, ya que su imitación implica bajo costo, pero con un mayor riesgo de error (Boyd y Richerson, 2005). Así, la cultura puede ser adaptativa cuando el aprendizaje es dif ícil y el ambiente no es ni muy variable ni muy estable, pero ciertas conductas pueden ser maladapta-tivas desde el punto de vista genético. Boyd y Richerson consideran enton-ces que la cultura es información almacenada en los cerebros humanos, que se transmite por procesos de aprendizaje social, y que la cultura evoluciona (Boyd y Richerson, 2005). Esta evolución se da por medio de tres tipos de fuer-zas evolutivas: las random (que incluyen mutación y deriva), las que tienen que ver con la toma de decisiones (que incluyen la variación guiada y sesgada) y la selección natural, que tiene que ver con cambios en la composición cultural de una población causados por los efectos de adherir a una variante cultural antes que a otra, tanto grupal como individualmente.
ConclusionesDespués de haber revisado estas posturas, queda entonces hacer una evalua-ción del concepto de adaptación en arqueología.
Hemos señalado que la propuesta de Kirch (1980) adolece de una falta de precisión respecto de cómo operar al nivel del registro arqueológico y no reconoce las fuerzas evolutivas culturales (sensu Boyd y Richerson, 2005). Sin embargo, su consideración respecto del papel de la selección natural en cuanto causa última y su uso del concepto de nicho lo vinculan con las posibilidades que abre la Teoría del Nicho Cultural, de elaboración más reciente. Dentro de este marco muchas de las propuestas de Kirch son ahora más abordables. Por otro lado, la propuesta seleccionista de encontrar selección natural en los arte-factos no ofreció resultados alentadores. A pesar de que insisten en la nece-sidad de probar que se trata de adaptaciones, los ejemplos que ofrecen estos autores no garantizan que en realidad estén encontrado adecuaciones. Y esto ocurre porque se parte de la hipótesis nula de selección natural, cuando en rea-lidad la hipótesis nula debiera ser la neutralidad. Como se dijo más arriba, las poblaciones humanas están desacopladas de las poblaciones artefactuales, por lo que no hay forma de “medir” el éxito reproductivo de una población humana asociada a una población artefactual, como para determinar la intervención de la selección natural. Por otro lado, la selección natural es colocada como causa

7 1
A D E P T O S A L A A D A P T A C I Ó N | V I V I A N S C H E I N S O H N
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.55-73
primera, descartando también las fuerzas evolutivas culturales y considerando los artefactos o los rasgos de artefactos como adaptaciones a priori. Se supone que la adaptación es ubicua, y así se desanima la evaluación de hipótesis alter-nativas que consideren, por ejemplo, constricciones estructurales.
Hasta el momento las propuestas que mejor han funcionado son aque-llas que colocan a la selección natural como causa última, como es el caso de la Teoría del Nicho Cultural (Laland et al., 2000, entre otros) y la Teoría de la Herencia Dual, ya que éstas incluyen en el juego evolutivo otras fuerzas de carácter cultural. Estas posturas son hasta el momento las más parsimoniosas, en cuanto pueden explicar la paradoja señalada al principio de este trabajo respecto de la gran plasticidad conductual humana atada a una larga lista de fracasos. Específicamente, la explicación que ofrece la Teoría de la Herencia Dual, en cuanto la cultura implicó una nueva disyuntiva evolutiva (ver arriba), da cuenta de esta situación.
En una versión de lo que Gould y Lewontin (1979) plantearon para la biología, no se debe exagerar la importancia de las adaptaciones ni extender su uso de manera excesiva. Las mismas críticas que estos autores le hicieron al programa adaptacionista son las que se le pueden plantear al uso del concepto de adaptación en el marco del procesualismo y el seleccionismo: no se analiza a los organismos como un todo integrado y, por lo tanto, no se tienen en cuenta las constricciones internas (sobre esto, ver Scheinsohn, 2010b): se relaciona la utilidad actual con la explicación del origen de cada rasgo; no se consideran las alternativas a la selección natural (a este respecto, ver la importancia que les dan a las fuerzas culturales Boyd y Richerson, 2005), y la sola plausibilidad es suficiente como para aceptar historias adaptativas (ver los ejemplos en O’Brien y Holland, 1992, y Van Pool, 2002, a pesar de que eso es lo que critican).
En la evolución (cultural o no) hay más que adaptación y selección natural, y, después de un largo camino recorrido en la arqueología, estamos en condicio-nes de darles a las fuerzas evolutivas culturales la importancia que se merecen. Sin embargo, los planteos de Boyd y Richerson (1985 y 2005) se dan en un con-texto sistémico. Su decodificación en cuanto registro arqueológico es el siguiente desaf ío, lo que ha comenzado a encararse ya en algunos trabajos arqueológicos.
Teniendo en cuenta la imposibilidad de identificar la selección natural en el registro arqueológico, la identificación de adecuaciones o exaptaciones parece ser más realizable desde el punto de vista arqueológico. El concepto de adaptación debiera considerarse de manera indirecta y asociado a la causación última, en cuanto implica la acción directa de selección natural, quedando la búsqueda de adecuaciones o exaptaciones (ver, por ejemplo, Scheinsohn, 1997 y 2010a, y Borrero y Borella, 2010) como un terreno más fértil. .

7 2
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.55-73
R e f e R e n c i a s
Boone, James y Eric A. Smith 1998.“IsItEvolutionYet?ACritiqueofEvolutionaryArchaeology”,Current Anthropology 39
(supplement),pp.141-173.
Borrero, Luis y Florencia Borella2010.“HarpoonsandTravellers:FuegianEthnographicCollectionsandtheRecentArchaeological
Record”,Before Farmingvol.2010p.1-2010/1article3Verenhttp://www.waspress.co.uk/journals/beforefarming/journal_20101/abstracts/index.php
Boyd, Robert y Peter Richerson1985.Culture and the Evolutionary Process.Chicago,UniversityofChicagoPress.2005. Not By Genes Alone: How Culture Transformed Human Evolution.Chicago,UniversityofChicagoPress.
Chabris, Christopher y Daniel Simons 2011. El gorila invisible y otras maneras en las que nuestra intuición nos engaña.BuenosAires,SigloXXI.
Cullen, Ben2000. Contagious Ideas: On Evolution, Culture, Archaeology, and Cultural Virus Theory.Londres,
OxbowBooks.
Diamond, Jared2004. Collapse: How Societies Choose to Fail or Succeed.NuevaYork,Viking.
Dunnell, Robert 1978.“StyleandFunction:AFundamentalDichotomy”,American Antiquity 43,pp.192-202.
Freeman, Derek.1974.“TheEvolutionaryTheoriesofCharlesDarwinandHerbertSpencer”,Current Anthropology15(3),
pp.211-237.
Ginnobili, Santiago2009.“Adaptaciónyfunción.Elpapeldelosconceptosfuncionalesenlateoríadelaselecciónnatural
darwiniana”,Ludus Vitalis XVII(31),pp.3-24.
Ginnobili, Santiago y Daniel Blanco 2007.“GouldyLewontincontraelprogramaadaptacionista:elucidacióndecríticas”,Scientiae Studia 5
(1),pp.35-48.
Gould, Stephen J. y Robert C. Lewontin 1979.“TheSpandrelsofSanMarcoandthePanglossianParadigm:ACritiqueoftheAdaptationist
Programme”,Proceedings of the Royal Society of London 205,pp.581-98.
Gould, Stephen J. y Elizabeth S. Vrba 1982.“Exaptation–aMissingTermintheScienceofForm”,Paleobiology 8(1),pp.4-15.
Hull, David L. y Michael Ruse (eds.) 1998.The Philosophy of Biology.Oxford,OxfordUniversityPress.
Kampourakis, Kostas 2011.“TeachingAboutAdaptation:WhyEvolutionaryHistoryMatters”,Science & EducationOnlineFirst,
7demayo.
Kendal, Jeremy, Jamshid Tehrani y John Odling-Smee2011.“HumanNicheConstructioninInterdisciplinaryFocus”,Philosophical Transactions of the Royal
Society Biological Sciences366,pp.785-792.
Kirch, Patrick1980.“TheArchaeologicalStudyofAdaptation:TheoreticalandMethodologicalIssues”,en Advances in
Archaeological Method and Theory, ed.M.Schiffer,Vol.3,pp.101-156.NuevaYork,AcademicPress.

7 3
A D E P T O S A L A A D A P T A C I Ó N | V I V I A N S C H E I N S O H N
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.55-73
Laland, Kevin, John Odling-Smee y Marcus Feldman2000.“NicheConstruction,BiologicalEvolution,andCulturalChange”,Behavioral and Brain Sciences
23,pp.131-175.
Leonard, Robert y George Jones 1987.“ElementsofanInclusiveEvolutionaryModelforArchaeology”,Journal of Anthropological
Archaeology6(3),pp.199-219.
Lovejoy, Arthur. 1960.The Great Chain of Being.NuevaYork,Harper.
Mayr, Ernest1963.Animal Species and Evolution.Cambridge,HarvardUniversityPress.
O’Brien, Michael y Thomas Holland 1992.“TheRoleofAdaptationinArchaeologicalExplanation”,American Antiquity57,pp.36-59.
Politis, Gustavo 2003.“TheTheoreticalLandscapeandtheMethodologicalDevelopmentofArchaeologyinLatin
America”American Antiquity68(2),pp.245-272.
Ridley, Mark 1997.Evolution.Oxford,OxfordUniversityPress.
Schaeffer, Jean M. 2009.El fin de la excepción humana.BuenosAires,FondodeCulturaEconómica.
Scheinsohn, Vivian1997.“ExplotacióndemateriasprimasóseasenlaIslaGrandedeTierradelFuego”.TesisdeDoctorado,
FacultaddeFilosofíayLetras,UBA.Ms.2001.“Elevolucionismoenarqueología”,en La evolución y las ciencias,comp.V.Scheinsohn,
pp.87-105.BuenosAires,Emecé.2010a.Hearts and Bones. Bone Raw Material Exploitation in Tierra del Fuego.Oxford,BARInternational
Series.2010b.“TheGood,theBadandtheUgly:HarpoonHeadsofBeagleChannel,IslaGrandedeTierradel
Fuego(Patagonia,Argentina)”,en Ancient and Modern Bone Artefacts from America to Russia. Cultural, Technological and Functional Signature, eds.Alexandra.Legrand-Pineau,Isabelle.Sidera,Natacha.Buc,Eva.DavidyVivian.Scheinsohn,pp.295-302.Oxford,BarInternationalSeries.
Steward, Julian1955.Theory of Culture Change: The Methodology of Multilinear Evolution.Urbana,Universityof
IllinoisPress.
Van Pool, Todd 2002.“Adaptation”,enDarwin and Archaeology: A Handbook of Key Concepts,eds.John.P.Harty
John.E.Terrell,pp.15-28.Westport,Bergin&Garvey.
Watt, Ward B. 2001.“Adaptation,Fitness,andEvolution”,en International Encyclopedia of the Social & Behavioral
Sciences,eds.N.SmelseryP.Baltes,Vol.1,pp.66-72.NuevaYork.Elsevier.
West-Eberhard, Mary 1992.“Adaptation:CurrentUses”,en Keywords in Evolutionary Biology,eds.E.FoxKelleryE.Lloyd,
pp.12-18.Cambridge,HarvardUniversityPress.
White, Leslie 1959.The Evolution of Culture: The Development of Civilization to the Fall of Rome.NuevaYork,
McGraw-Hill.

7 4

7 5
A PORTACIÓN A RQU EOLÓGICA A L CONOCI M I EN TO DEL PROC ESO DE DESCOM POSICIÓN DEL C U ER PO H U M A NO EN POSICIÓN SEN TA DA/F LEX IONA DA
K arina Gerdau r adonic*[email protected] University, Reino Unido
R e s u m e n La excavación arqueológica de restos humanos
permite recabar datos sobre el proceso de descomposición.
Basándose en estos conocimientos, se puede establecer la
disposición original de los restos, distinguir entre manipulación
intencional y eventos tafonómicos, así como revelar la presencia
de material orgánico desaparecido. A partir de observaciones
en contexto arqueológico, se presentan las modalidades de
descomposición en posición sentada/flexionada de un fardo
funerario y se ilustra cómo los huesos se desplazan naturalmente
durante el proceso.
P A L A B R A S C L A V E :
Descomposición, tafonomía, antropología forense, arqueología,
fardo funerario.
Artículorecibido:1dejuniode2011|aceptado:14deoctubrede2011|modificado:23denoviembrede2011
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,294pp.ISSN1900-5407,pp.75-94
* Ph.D.,DoctoratenAnthropologieBiologique,UniversitéBodeauxI,Talence,Francia.

7 6
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.75-94
arChaeologiCal insights into the disartiCulation pattern of a human sKeleton in a sitting/squatting position
ABSTR AC T Archaeological excavation of
human remains can provide information
on decomposition. From these data it is
possible to establish the original lay out
of the remains, to distinguish between
taphonomic processes and intentional
human intervention, and to reveal
the presence of decomposed organic
materials. From observations made in an
archaeological context, the author describes
the decomposition of a funerary bundle in a
sitting/squatting position and explains how
bones can be naturally displaced during
the process.
K E Y W O R D S :
Decomposition, Taphonomy, Forensic
Anthropology, Archaeology, Funerary Bundle.
aportação arqueológiCa ao ConheCimento do proCesso de deComposição do Corpo humano em posição sentada/flexionada
RESUMO A escavação arqueológica de
restos humanos permite recavar dados sobre
o processo de decomposição. Baseando-se
nestes conhecimentos, pode-se estabelecer
a disposição original dos restos, distinguir
entre manipulação intencional e eventos
tafonômicos, assim como revelar a presença
de material orgânico desaparecido. A partir
das observações em contexto arqueológico,
apresentam-se as modalidades de
decomposição em posição sentada/
flexionada de um fardo funerário e ilustra-se
como os ossos deslocam-se naturalmente
durante o processo.
P A L A V R A S C H A V E :
Decomposição, tafonomia, antropologia
forense, arqueologia, fardo funerário.

7 7
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.75-94
A PORTACIÓN A RQU EOLÓGICA A L CONOCI M I EN TO DEL PROC ESO DE DESCOM POSICIÓN DEL C U ER PO H U M A NO EN POSICIÓN SEN TA DA/F LEX IONA DA
K a r i n a G e r d a u r a d o n i c
La excavación arqueológica de restos humanos per-mite recabar datos sobre el proceso de descomposición, información útil no sólo en arqueología funeraria, sino también en antropología forense. Enten-der dicho proceso, cómo se desarticulan y desplazan los huesos del esqueleto al desaparecer el tejido blando que los une, es necesario para reconstruir los hechos y eventos desde la muerte, pasando por el sepelio o abandono del cadá-ver, hasta el momento en el que se hallan sus restos. El tratamiento post mórtem del cuerpo, su posición original al ser depositado y los eventos posdeposicio-nales influyen sobre el proceso de descomposición, el grado de desarticulación del esqueleto y, por último, su disposición final. Por ende, es posible recorrer el camino inverso, partiendo de los despojos hallados, a menudo desarticulados, y llegar a establecer los hechos en torno al tratamiento post mórtem, e incluso la muerte (véanse, entre otros, Andrews y Bello, 2006; Boddington, 1987; Brothwell, 1987; Duday, 1986, 1987, 2005, 2006; Duday et al., 1990; Duday y Guillon, 2006; Haglund y Sorg, 1997, 2002).
En cuestión de tratamientos mortuorios y eventos tafonómicos, los con-textos arqueológicos son una fuente de información inigualable y permiten enriquecer nuestro conocimiento sobre la tafonomía del cadáver en diversas situaciones, ya sean culturales o ambientales. La manera en la que se depo-sita el cuerpo y el momento en el que se le da sepultura varían en función de las creencias y preferencias del difunto y sus allegados. Así mismo, situaciones particulares, el entorno geográfico y el clima, entre otros factores, también pue-den influir en decisiones sobre el cuándo y cómo del sepelio.
Este trabajo busca ilustrar la tafonomía del cadáver de un fardo funera-rio que contiene un individuo adulto en posición sentada-flexionada, mediante

7 8
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.75-94
evidencias arqueológicas provenientes del yacimiento Tablada de Lurín (depar-tamento de Lima), en la costa central peruana. Además, también busca enri-quecer el conocimiento sobre conexiones anatómicas lábiles y persistentes en el proceso de descomposición del cuerpo humano.
Las tumbas colectivas de Tablada de LurínEl yacimiento arqueológico de Tablada de Lurín se encuentra en el valle de Lurín, en la costa central de Perú. La ocupación principal fue una gran necrópo-lis de la primera mitad del Intermedio Temprano (siglos II a. C.-II d. C.), la cual fue el objeto de dos fases funerarias consecutivas, aunque distintas: la primera caracterizada por entierros en pozo, mayormente individuales, y la segunda por tumbas colectivas en estructuras semisubterráneas en piedra, llamadas cistas (para una descripción más detallada de estas ocupaciones, véanse Gerdau y Makowski, 2011; Makowski, 2002, 2009a, 2009b).
Las observaciones que se presentan a continuación fueron recabadas en el marco del Proyecto Arqueológico-Taller de Campo “Lomas de Lurín” (PATL), dirigido por Krzysztof Makowski, de la Pontificia Universidad Católica del Perú. Los datos tafonómicos fueron recopilados por la autora durante la excavación de una tumba colectiva (EF 4 {NE-B}) y el análisis osteológico del material humano proveniente de ésta y otras cuatro tumbas –EFs 1, 2 y 3 (NE-B) y EF 6 (NE-A)–, excavadas con antelación por diversos miembros del pro-yecto PATL (véanse Gerdau, 2007; Gerdau y Makowski, 2011). Los informes, trabajos y registros gráficos y escritos de campo de estas otras cuatro tumbas brindaron información complementaria (Atalaya, ms.; Balbuena, 1996; García, ms.; Makowski, 2002; Makowski et al., 1996).
Las cinco cistas excavadas por el equipo de Makowski contenían entre 20 y 55 individuos, mayormente adultos, en entierros primarios o secundarios –algunos de estos últimos parciales–, y habían sido objeto de diversas remocio-nes, con el fin de ordenar el interior de las cámaras funerarias (figura 1; Gerdau, 2007; Gerdau y Makowski, 2011). Aun así, ciertos individuos se habían mante-nido intactos o casi, y en su ubicación original, facilitando información sobre el tratamiento mortuorio, la posición original en las que se les dio sepultura y, finalmente, el orden de desarticulación de las diferentes conexiones anatómicas del cuerpo humano.
Con base en los trabajos de Balbuena (1996), de la autora (Gerdau, 2007; Gerdau y Makowski, 2011) y de Makowski (2002), se ha llegado a la conclu-sión que, en un principio, los difuntos eran enfardelados de manera simple, sin momificación. Se les sentaba o acuclillaba sobre un cesto, con las extremidades superiores flexionadas y cruzadas delante del pecho, las extremidades inferio-

7 9
A P O R T A C I Ó N A R Q U E O L Ó G I C A A L C O N O C I M I E N T O | K A R I N A G E R D A U R A D O N I C
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.75-94
res flexionadas, las rodillas frente al mentón, y los pies delante o bajo la zona púbica. A continuación se les envolvía en uno o más mantos, a fin de constituir un fardo pequeño y simple. Este tipo de fardo se puede apreciar, por ejemplo, entre las imágenes fotográficas y radiográficas de las momias sudamericanas albergadas en el Museum für Völkerkunde de Berlín (ahora el Ethnologisches Museum; Herrmann y Meyer, 1993).
Los restos óseos de Tablada no presentan evidencia alguna de momifi-cación, ya sean tejidos anatómicos desecados o huellas de desarticulación en orden paradójico. En situaciones naturales donde el cadáver no ha sido momi-ficado, las articulaciones lábiles (labiles, en francés) son aquellas que se desha-cen primero (como las de las manos y los pies), y las persistentes (persistentes, en francés) perduran más tiempo (como la de la rodilla) (véanse Duday, 2005, y Duday et al., 1990, para una discusión más amplia; términos traducidos por la autora). Por lo tanto, es posible, en el caso de entierros secundarios, y a pesar del desplazamiento de los despojos, encontrar la rodilla aún articulada y los huesos de la mano completamente removidos y fuera de su lugar. Por el contrario, en casos de momificación que con el paso del tiempo terminan por descomponerse, Maureille y Sellier (1996) describen un proceso de desarti-culación invertido, en donde las conexiones anatómicas lábiles perduran (v.g., la mano) y las persistentes se deshacen primero. Es por eso que individuos inicialmente momificados (natural o artificialmente) que han sido desplaza-dos pueden llegar a presentar un orden de desarticulación llamado paradójico (Duday et al., 1990: 33).
Materiales y métodosEl patrón de desarticulación descrito a continuación fue observado durante excavaciones llevadas a cabo por el Proyecto Arqueológico-Taller de Campo “Lomas de Lurín”, proyecto de investigación dirigido por K. Makowski, de la Pontificia Universidad Católica del Perú. Aunque el proyecto estudió ambas ocupaciones funerarias, la autora del presente trabajo sólo participó en la investigación de la segunda fase de ocupación del cementerio. Por lo tanto, como ya se mencionó, las observaciones presentadas provienen de la excava-ción de una estructura funeraria (EF 4 {NE-B}) y de información procedente de informes, tesis y registros gráficos y escritos de las temporadas de exca-vación anteriores (Atalaya, ms.; Balbuena, 1996; García, ms.; Makowski et al., 1996). Se observaron asociaciones osteológicas de primer grado, según la definición de Duday (2006: 50), directamente en campo, y también se les buscó en los registros gráficos de excavación generados por otros miembros del equipo de investigación.

8 0
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.75-94
Excavación y registro del yacimiento
Como se describió más arriba, el equipo de Makowski excavó cinco estructu-ras funerarias, las cuales contenían entierros colectivos secuenciales (Gerdau, 2007; Gerdau y Makowski, 2011). Las técnicas de excavación y de registro de campo siguieron las recomendaciones de la escuela francesa de Archéothanato-logie, la cual especifica que observaciones antropológicas detalladas deben ser recabadas en el campo, en conjunto con un registro gráfico y escrito detallado del contexto. Se ruega al lector referirse a los trabajos de Duday y sus colegas para más amplia información sobre los detalles que deben observarse (v.g., aso-ciaciones osteológicas, articulaciones lábiles o persistentes, entre otros) y sobre las interpretaciones que ciertas observaciones conllevan (v.g., espacio vacío o rellenado, efecto de pared, inter alia), puesto que la metodología y sus aplica-ciones son discutidas ampliamente en estos trabajos (Duday, 1986, 1987, 2005, 2006; Duday y Guillon, 2006; Duday et al., 1990).
Dada la densidad de material contenido dentro de cada estructura, des-pués de desarmar el techo de éstas para facilitar el acceso, la excavación pro-gresó levantando una capa de material a la vez. Es necesario precisar que estas capas no representan diferentes eventos o momentos en el uso de cada estruc-tura. Por el contrario, las capas eran delimitadas de manera artificial según la cantidad de elementos que podían ser identificados, registrados y levantados en una sola vista fotográfica en planta de toda el área interior de cada estructura (ver la figura 1). Por ejemplo, en la estructura EF 4 (NE-B), se requirieron un total de 64 vistas en planta para registrar todos los especímenes osteológicos u otros dentro de dicha estructura. Por ende, el primer levantamiento representa la capa superior, y el último representa la capa de restos arqueológicos que yacían directamente sobre el piso de la estructura. Los dibujos en planta de cada estructura están basados en estas imágenes fotográficas.
Cada espécimen osteológico (según la definición de Grayson [1984, en Lyman, 1994: 100]) levantado fue registrado en estas fotograf ías de vista en planta. Además, la profundidad a la cual yacía cada espécimen fue registrada a partir de un punto fijo datum. Para la excavación de EF 4 (NE-B), la autora registró la profundidad de cada hueso largo a ambos extremos de éste (proximal y distal), a fin de identificar si éstos yacían de manera plana sobre la superficie, o de manera inclinada. Este tipo de registro permite ubicar cada espécimen en el espacio tridimensional, facilitando así el estudio de las asociaciones espaciales de todos los especímenes recuperados.
Durante la excavación de EF 4 (NE-B), la autora identificó cada espéci-men de más de 1 cm2 de forma detallada (elemento anatómico o parte de éste y lateralidad); sin embargo, elementos anatómicos menores que ese tamaño,

8 1
A P O R T A C I Ó N A R Q U E O L Ó G I C A A L C O N O C I M I E N T O | K A R I N A G E R D A U R A D O N I C
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.75-94
como pueden ser las falanginas del pie, también fueron identificados y regis-trados in situ. Los registros de las otras cuatro estructuras por otras personas del equipo de investigación procedieron de manera similar pero con menor detalle en cuanto a los especímenes de menor tamaño. Aun así, para fines de este estudio fue posible establecer la ubicación de la mayoría de especímenes y evaluar las asociaciones espaciales, así como el grado de desarticulación de ciertos individuos y de partes anatómicas aisladas como pies, extremidades u otras. Asociaciones osteológicas de primera grado habían sido registradas en campo por todos los miembros del equipo, y a pesar de las remociones y el con-glomerado óseo dentro de todas las estructuras, fue posible identificar ciertos individuos o partes de éstos dentro de cada estructura. El material asociado también fue registrado en el espacio tridimensional.
Tamaño de la muestraEl número mínimo de individuos (NMI) es diferente según la estructura (ver el cuadro 1). Como se mencionó más arriba, dentro de cada estructura, durante las excavaciones se pudieron identificar ciertos individuos completos o par-ciales. Como se puede apreciar en el cuadro 1, dentro de las cinco estructuras
Figura1.EF4(NE-B),Levantamiento23,vistaenplanta.

8 2
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.75-94
sólo cinco de éstos estaban completos y aparecían como depósitos mortuorios primarios intactos. Sin embargo, un número más grande (39) estaban incom-pletos y hasta la fecha es dif ícil concluir si se trata de depósitos primarios dis-turbados o de depósitos secundarios, debido a que algunos de ellos habían sido removidos y desplazados dentro de la misma estructura múltiples veces (véanse Gerdau, 2007; Gerdau y Makowski, 2011).
Estos pocos individuos completos son los que han provisto la mayoría de la información sobre el rito mortuorio, incluidas la preparación post mórtem del cuerpo con fines de entierro, la posición original de los restos y, finalmente, la secuencia de desarticulación esquelética dentro de un fardo funerario. El tamaño de la muestra es limitado, puesto que consiste en cuatro individuos adultos y un juvenil identificados como depósitos funerarios primarios (cuadro 1). De los cuatro adultos no todos mostraban el mismo patrón, pero se pueden distinguir rasgos generales, que se describen a continuación. Más aún, los indi-viduos incompletos sirvieron para corroborar ciertas observaciones. El cuadro 2 presenta las articulaciones que se pudieron observar, y a aquellos individuos que las presentaban.
Por último, es importante enfatizar que la secuencia de desarticulación que se detalla en este trabajo sólo se aplica a individuos adultos que fueron enterrados enfardelados, con las extremidades inferiores y superiores flexio-nadas, y que fueron depositados dentro de un espacio vacío (no en una tumba rellenada con tierra). Los juveniles, quienes aún no tienen las epífisis consoli-dadas, pueden presentar un patrón distinto, pero se necesitan más investiga-ciones al respecto.
Cuadro1.
NMI(Gerdau,2007)
Individuosidentificadoscomodepósitosprimarios
Otrosindividuos(adultosyjuveniles) identificados(puedenserdepósitos primariosdisturbadososecundarios)
EF1(NE-B) 36 0 11
EF2(NE-B) 38 0 2
EF3(NE-B) 29 1adulto 8
EF4(NE-B) 55 3adultos(y1juvenil) 15
EF6(NE-A) 20 0 3
NúmeroMínimodeIndividuos(NMI),númerodeindividuosidentificadoscomodepósitosprimariosynúmerodeotrosindividuos(disturbados)dentrodecadaestructuraexcavadaporelProyectoArqueológico-TallerdeCampo“LomasdeLurín”.CaberesaltarqueladiscrepanciaesgrandeentreelNMIyelnúmerototaldeindividuosidenti-ficados.Estosedebeaquesóloalgunosdeéstospudieronseridentificadoscomotalesencadaestructura;todoslosdemásindividuosestabanconglomeradosynofueposibledistinguiraunosdeotros(segúnAtalaya,1996;Balbuena,1996;García,ms.;Gerdau,ms.

8 3
A P O R T A C I Ó N A R Q U E O L Ó G I C A A L C O N O C I M I E N T O | K A R I N A G E R D A U R A D O N I C
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.75-94
Tafonomía del esqueleto de un individuo sentado/flexionado y enfardelado
Como se describió más arriba, la posición original del cuerpo era sentado o acuclillado, con las extremidades superiores e inferiores flexionadas delante del pecho. Los individuos habían sido colocados sobre cestos y envueltos en un manto, al menos. Los textiles y otro material orgánico no se preservan bien en Tablada. Mientras que estos materiales se descomponían fueron progresi-vamente reemplazados por la tierra utilizada para rellenar las estructuras en el momento de su clausura, o por sedimento que se infiltraba desde el exterior de
Cuadro2.
CabezacaídasobreeltorsoEF3(NE-B):inds.IEF4(NE-B):inds.VI,X,XIII(juvenil)
Persistenciadelaarticulacióndelhombrocon dislocaciónescápulo-torácicayclavículo-esternal
EF4(NE-B):inds.X
ColumnaarticuladahastavértebrascervicalesEF3(NE-B):inds.IEF4(NE-B):inds.VI;X,XIII(juvenil)
PersistenciadelaarticulacióndelcodoEF3(NE-B):inds.I,IIIEF4(NE-B):inds.I,V,VI,VIII,X
Dislocacióndelamuñecaydelasarticulaciones delamano
EF3(NE-B):inds.I,IV,VEF4(NE-B):inds.I,II,VI,VIII,X,XIII(juvenil)
Persistenciadelacolumnalumbar
EF1(NE-B):ind.IEF2(NE-B):inds.I,IIEF3(NE-B):inds.IIIEF4(NE-B):inds.I;II,XIV(juvenil),XVI,XVII
Persistenciadelapelvis
EF2(NE-B):inds.I,IIEF3(NE-B):inds.I,IIIEF4(NE-B):conjuntodepelvisyunpardepies,inds.I;II,V,VI(dislocacióndelsacro),VIII,X,XIII(juvenil),XIV(juvenil),XVI,XVII
Persistenciadelpie
EF1(NE-B):ind.IEF2(NE-B):inds.I,IIEF3(NE-B):inds.I,III,IV,VEF4(NE-B):conjuntodepelvisyunpardepies; inds.VI,X,XIII(juvenil),XVII
PersistenciadelarodillaEF1(NE-B):ind.IEF3(NE-B):inds.III,IV,VEF4(NE-B):inds.V;VI,VIII,X,XIII(juvenil),XVII
Desplazamientodelaextremidadinferiorsin dislocacióndeloshuesosdeltarso
EF3(NE-B):inds.IEF4(NE-B):inds.VI,X,XIII(juvenil)
Individuossobreloscualessepudieronobservarlasarticulacionescitadasenelcuadro.VéaseGerdau(2007)paraunadescripcióndetalladadecadaindividuo.

8 4
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.75-94
las estructuras con el paso del tiempo (Balbuena, 1996; Gerdau, 2007; Gerdau y Makowski, 2011). Son la posición de los restos descritos más abajo, así como restos de fibra orgánica hallados entre ellos, los que sugieren la presencia de un cesto y textiles.
Esqueleto axialDada la posición vertical del cuerpo, es natural que la mitad superior de éste, así como los huesos largos de las extremidades inferiores, se hallasen en des-equilibrio a medida que avanzaba el proceso de descomposición. Al desapare-cer los órganos internos y generar un vacío en las cavidades torácica y pélvica, la mitad superior de la columna vertebral y de las costillas cayó dentro de estas cavidades perdiendo su orden anatómico y la mayoría de sus conexiones. El esqueleto cefálico se asentó sobre los huesos del tórax, en buena parte desarti-culados, guardando su orientación original. Se ubicó el atlas inmediatamente por debajo de la base del cráneo o en proximidad de ésta.
En algunos individuos, el fardo facilitó la conservación de las articulacio-nes de la columna vertebral en su casi totalidad (ver las figuras 3 a 6), proporcio-nando un soporte sobre el cual podían descansar las vértebras sin deshacerse las conexiones. Con el paso del tiempo, el tejido del fardo fue reemplazado por sedimento que se infiltró y por los despojos de individuos contiguos, que pro-porcionaban un apoyo adicional.
En la mayoría de los casos observados en Tablada, fue únicamente la mitad inferior de la columna vertebral la que quedó articulada, apoyándose contra la parte posterior del cesto dentro del cual se había sentado al individuo, y contra el manto que lo envolvía. Al transcurrir el tiempo, el sedimento fue sustituyendo la cesta y el manto.
La mitad superior de las costillas cayó en desorden dentro de las cavidades torácica y pélvica. Sin embargo, la mitad inferior de las costillas cayó de parte y otra de las vértebras, conservando más o menos su secuencia anatómica (figu-ras 3 a 6). Si el individuo yacía en un principio levemente inclinado hacia atrás, las costillas se asentaban detrás de la parte superior del hueso coxal, a menos profundidad que éste. Si el individuo yacía con la espalda recta o ligeramente inclinada hacia adelante, las costillas inferiores se asentaban sobre los iliones.
Cintura escapular y extremidades superioresEl movimiento de las extremidades superiores y de los huesos de la cintura escapular dependía mucho del espacio disponible en torno al cuerpo. Si el individuo se encontraba con la espalda recta y tenía algún soporte posterior, una vez liberada de sus conexiones anatómicas, la escápula se deslizaba hacia

8 5
A P O R T A C I Ó N A R Q U E O L Ó G I C A A L C O N O C I M I E N T O | K A R I N A G E R D A U R A D O N I C
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.75-94
la zona pélvica y aparecía en su orientación anatómica natural, es decir, con la parte superior hacia arriba detrás del hueso coxal de la misma lateralidad (figuras 5 y 6). Si el individuo se hallaba inclinado hacia adelante, la escápula aparecía en vista posterior, por encima de las costillas y la pelvis (figuras 5 y 6).
Con frecuencia, la cabeza humeral y la clavícula seguían en su caída a la escápula, hallándose a menudo en proximidad de ésta. Al pasar el tiempo, y según los movimientos de los demás huesos de la mitad superior del cuerpo, la articulación del codo se dislocaba levemente, pero los extremos proximales del radio y de la ulna (cúbito) se hallaban cerca del extremo distal del húmero (figu-ras 3 a 6). Era fácil reconocer la posición cruzada de las extremidades superiores.
La posición original de las manos fue más dif ícil de establecer con clari-dad. Puesto que los antebrazos estaban cruzados delante del pecho, es posible
Figura2.Dibujoesquemáticodelos19individuosidentificadosdentrodeEF4(NE-B)ydesucronologíarelativa.Lasflechasindicanelordendellegadadelosindividuos(delmásrecientealmásantiguo),cuandoéstasepudoestablecer.Porejemplo,elindividuoXVIIfueposiblementeelprimeroenllegar;losindividuosIyIIestuvieronentrelosúltimos.

8 6
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.75-94
Figura6.EF4(NE-B),IndividuoX, vistaenplanta,mira10cm.
Figura3.EF4(NE-B),IndividuoVI, vistaenplanta(dibujo,KarlaGerdau).
Figura4.EF4(NE-B),IndividuoVI, vistaenplanta(dibujo,KarlaGerdau).
Figura5.EF4(NE-B),IndividuoX, vistaenplanta(dibujo,KarlaGerdau).

8 7
A P O R T A C I Ó N A R Q U E O L Ó G I C A A L C O N O C I M I E N T O | K A R I N A G E R D A U R A D O N I C
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.75-94
que las manos se hallasen en una de cuatro posiciones: delante de la faz, a la altura del mentón, sobre el pecho o sobre los hombros. Estas posiciones han sido registradas en momias prehispánicas peruanas (véase Herrmann y Meyer, 1993). Cualquiera de estas posiciones es inestable para los huesos de la mano, una vez que desaparecen los tejidos blandos que los mantienen unidos. Por lo tanto, los huesos de la mano se hallaban desarticulados entre los huesos de las cavidades torácica y pélvica.
Cintura pélvica y extremidades inferioresLas extremidades inferiores caían hacia un lado u otro, en función de la posi-ción original del cuerpo. La cadera y el tobillo se desarticulaban tan sólo leve-mente (figuras 3 a 6). En varias ocasiones la rodilla se mantuvo en conexión, con la patela (rótula) aún in situ, a pesar de la caída de los huesos largos hacia un lado. Es muy probable que el manto del fardo mantuviese la patela en su articulación con el fémur y la tibia. El sedimento hubiera reemplazado progre-sivamente al textil.
Gracias a su posición dentro de un cesto, los huesos de la pelvis y del pie se mantenían en conexión anatómica. La forma de la pelvis dictaría que una vez sus conexiones anatómicas se desintegraran, ésta se abriría, los huesos coxales girarían lateral y posteriormente, y el sacro caería ventral o dorsalmente (Duday, 2005: 170; 2006: 35). Sin embargo, las paredes del cesto sobre el cual estaba sentado el individuo proveían un soporte contra el cual se apoyaban los huesos coxales sin perder su precario equilibrio. Así mismo, al poner los pies a plano sobre el fondo de la canastilla, los huesos del pie lograban mantenerse en equilibrio y en casi perfecta conexión, a pesar de la descomposición de los tejidos blandos.
En un caso en particular (Individuo XVII, EF 4 {NE-B}), los dedos del pie se hallaron dorsi-flexionados, aparentemente apoyados contra la pared anterior del cesto en el cual se sentó al individuo. Las falanges se encontraron, por ende, en posición vertical, distal hacia arriba.
Por último, la presencia de un envoltorio (manto) y de un soporte (cesto) permite desplazar conjuntamente dentro de un mismo espacio o de un lugar a otro los despojos de un solo individuo, pese a su grado de descomposición avanzando. Así, se hallaron en conjunto restos fuertemente desarticulados que pertenecían a un mismo individuo, y otros despojos parcialmente articulados, en especial la pelvis y los pies, con evidencias de haber sido empujados hacia un lado (ver las figuras 7 a 10).

8 8
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.75-94
DiscusiónLa muestra de individuos completos dentro de cada estructura es limitada (cua-dro 1). Aun así, se puede discernir un patrón general, aunque se necesita reca-bar más observaciones. En Tablada, la posición vertical del cuerpo del individuo sentado o acuclillado aumentó el grado de desarticulación de la mitad superior del cuerpo, puesto que una vez el estado de descomposición es avanzado, ya no queda tejido blando que mantenga en su sitio los diferentes elementos óseos. Por lo tanto, éstos caen naturalmente. El manto-envoltorio y el soporte-conte-nedor inferior (cesto) estabilizan la mitad inferior del tórax, la pelvis y los pies, manteniéndolos en articulación completa o parcial.
Cuando se halla un individuo en esta posición pero con la caja torá-cica, la columna y la pelvis aún en conexión anatómica, esto puede deberse a dos posibilidades:
Figura10.EF4(NE-B),IndividuoIII,vistaenplanta(dibujo,KarlaGerdau).
Figura7.EF4(NE-B),IndividuoI, vistaenplanta(dibujo,KarlaGerdau).
Figura8.EF4(NE-B),IndividuoI, vistaenplanta,mira10cm,nortehaciaarriba.
Figura9.EF4(NE-B),IndividuoXVI, vistaenplanta.

8 9
A P O R T A C I Ó N A R Q U E O L Ó G I C A A L C O N O C I M I E N T O | K A R I N A G E R D A U R A D O N I C
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.75-94
1) el reemplazo progresivo de los tejidos blandos descompuestos por sedi-mento que se infiltra dentro del contexto (en el caso de entierros en sedimento fluido, como es la arena (Duday, 2005: 185-186; 2006: 41); o,
2) porque los elementos óseos fueron sujetos por algún soporte exterior, en el caso de depósitos en espacio vacío, como lo son los entierros en las cistas de Tablada. En dos individuos dentro de la estructura EF 4 (NE-B), la columna se mantuvo casi intacta por el soporte que ofrecían el envoltorio textil y los restos adyacentes (figuras 3 a 6).
Articulaciones lábiles y persistentes
El cuelloAlgunas observaciones hechas en Tablada corroboran aquellas de otros estu-dios sobre las articulaciones lábiles y persistentes (ver el cuadro 3). Por ejem-plo, el atlas se halló bajo la base del cráneo del individuo X (EF 4 {NE-B}). Duday y sus colegas (1990: 31) afirman que la articulación atlanto-occipi-tal es más persistente que otras articulaciones intervertebrales cervicales. Duday (2006: 35) escribe que la desarticulación inicial en el cuello ocurre al nivel de la articulación atlas-axis o axis-cervical 3. Andrews y Bello (2006: 17) también afirman que el cuello es una articulación lábil. Es posible que algunas articulaciones cervicales (atlas-axis, axis-cervical 3) sean más lábiles que otras (atlas-occipital). Más aún, dentro de los fardos las vértebras cer-vicales se encontraban en desorden dentro de las cavidades torácica y abdo-minal, con la excepción del atlas, que podía hallarse bajo la base del cráneo, indicación de que cuando el cráneo se asentaba sobre el torso, el atlas aún mantenía su conexión.
Cintura escapularEn Tablada, el hombro presenta evidencias de ser una conexión más persistente que la articulación clavículo-esternal o la conexión escápulo-torácica. Cuando la escápula se desarticula de la caja torácica, la clavícula y el húmero la siguen en su caída. Por otra parte, la articulación entre el esternón y la clavícula se deshace. Duday (2005: 167; 2006: 33; Duday et al., 1990: 31) también presenta la conexión escápulo-torácica como lábil.
La muñeca y las articulaciones de la manoLas articulaciones de la mano, incluida la muñeca, son citadas como lábiles (Andrews y Bello, 2006: 17; Duday, 2005: 167; 2006: 33; Duday et al., 1990: 31). Entre los restos humanos de Tablada, estas articulaciones también aparecen

9 0
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.75-94
como lábiles. Más aún, la muñeca debe descomponerse después de las articu-laciones entre los carpos, los metacarpos y las falanges. En Tablada no hay evi-dencias de que las manos cayesen enteras de la muñeca. Más bien, los huesos de la mano parecen haber caído progresivamente de la mano hacia la cavidad abdominal o pélvica.
Cuadro3.Listadearticulacioneslábilesypersistentes,segúnlaliteratura.
Lábil Persistente Referencia Tablada*
Articulacionesentrevértebrascervicales
Dudayet al.,1990;Duday,2005,2006;AndrewsyBello,2006
X
Articulacionesdentro delamano
Dudayet al.,1990;Duday,2005,2006;AndrewsyBello,2006
X
Muñecamenoslábilquearticulacionesdentrodelamano
X
Articulacionesdistalesdelpie
Dudayet al.,1990;Duday,2005,2006;AndrewsyBello,2006
Articulacionescosto-esternales
Dudayet al.,1990;Duday,2005,2006;AndrewsyBello,2006
Conexión escápulo-torácica
Dudayet al.,1990;Duday,2005,2006;AndrewsyBello,2006
X
Articulación clavículo-esternal
Dudayet al.,1990;Duday,2005,2006;AndrewsyBello,2006
X
Cadera Duday,2005,2006
Articulaciónatlanto-occipital
Dudayet al.,1990;Duday,2005,2006
X
HombroDudayet al.,1990;Duday,2005,2006
X
Columnalumbar,articulaciónsacro-lumbar,articula-ciónsacro-iliaca
Dudayet al.,1990;Duday,2005,2006
X
Rodilla,tobillo,huesosdeltarso
Dudayet al.,1990;Duday,2005,2006
X
Rodilla,máspersistentequetobillo
Pereira,1996X(tambiénmáspersistentequecadera)
*LacolumnadeTabladasehamarcadoconunaXsilaevidenciatafonómicaindicaquelaarticulaciónoconexiónanatómicaeslábilopersistente;noindicasilasarticulacionessepodíanobservar(véaseelcuadro2).

9 1
A P O R T A C I Ó N A R Q U E O L Ó G I C A A L C O N O C I M I E N T O | K A R I N A G E R D A U R A D O N I C
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.75-94
Columna lumbar y pelvisEn Tablada, la pelvis y la columna lumbar aparecen como articulaciones persis-tentes. Esto confirma otras observaciones (Duday, 2005: 167; 2006: 33; Duday et al., 1990: 31). Sin embargo, cabe resaltar que esta observación puede ser un producto del patrón funerario, ya que la columna lumbar y la pelvis fueron mantenidas en articulación con ayuda del cesto y el envoltorio del fardo.
Cadera, rodilla y tobilloEl grado de persistencia de la articulación de la rodilla parece superior al de la cadera y los tobillos. Dentro de los fardos de Tablada, una vez descompuesta la articulación de la cadera, el fémur no podía mantenerse en su posición vertical y levemente inclinada. Por lo tanto, el muslo, la patela y los huesos de la pierna caían hacia un lado. Estos huesos caían conjuntamente y la rodilla se mantenía articulada. La tibia y la f íbula (peroné) se desarticulaban del talo (astrágalo), pero no jalaban ninguno de los huesos del tarso en la caída. Los pies se man-tenían articulados. Estas observaciones sobre la rodilla y el tobillo confirman aquellas hechas por Grégory Pereira (1996) en el yacimiento arqueológico de Potrero de Guadalupe (Michoacán, México). Duday (2005: 167; 2006: 33; Duday et al., 1990: 31) también afirma que la rodilla y el tobillo son articulacio-nes persistentes. Tablada y Michoacán no contradicen esto. Más bien, indican que la rodilla es más persistente que el tobillo.
ConclusiónLa excavación y las observaciones detalladas de la disposición de restos óseos humanos ofrecen información sobre la secuencia de desarticulación de ciertas conexiones anatómicas del cuerpo humano. En los casos presentados aquí, la posición vertical del individuo acuclillado-sentado incrementó el grado de des-articulación de la mitad superior del esqueleto, ya que la desaparición del tejido blando que mantenía unidos los huesos generó una pérdida de equilibrio en esta posición. La presencia de un envoltorio (manto) y de un soporte inferior (cesto), en el caso de los fardos funerarios, proporcionó estabilidad a la mitad inferior del tórax, a la zona pélvica y a los pies, ayudando a mantenerlos en conexión anatómica parcial o completa. Así mismo, el cesto y el manto faci-litaron el transporte de los restos de un lugar a otro dentro de la estructura, minimizando la pérdida de elementos óseos de los individuos.
Por último, las observaciones hechas en Tablada tienden a demostrar que la articulación del hombro (escápula, húmero y clavícula) es más persis-tente que las conexiones clavículo-esternal y escápulo-torácica, puesto que al caer el hombro, los tres huesos que lo constituyen se mantienen en proximi-

9 2
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.75-94
dad; sin embargo, la clavícula se desarticula del esternón de manera marcada, así como la escápula de la caja torácica. Así mismo, el grado de “persistencia” de la articulación de la rodilla es más fuerte que el de la cadera y el tobillo. Una vez deshecha la articulación de la cadera, el muslo, la patela, y la pierna caen conjuntamente y de manera articulada. Al caer, la tibia se desarticula del talo sin que la caída afecte las conexiones anatómicas de los huesos del tarso. Estas observaciones sobre la rodilla y el tobillo confirman aquellas hechas por Pereira (1996) en el yacimiento arqueológico de Potrero de Guadalupe (Michoacán, México).
Información sobre articulaciones lábiles y persistentes puede ayudar a la interpretación de depósitos primarios y secundarios, así como, posiblemente, a revelar la presencia de restos momificados descompuestos. Además, la obser-vación detallada de los restos y su entorno puede revelar la presencia de envol-torios, soportes u otro material asociado que podrían haberse descompuesto y desaparecido con el paso del tiempo (véanse Boddington, 1987; Duday, 1986, 1987, 2005, 2006; Duday y Guillon, 2006; Duday et al., 1990).
AgradecimientosQuisiera agradecer a H. Duday, K. Makowski, a todos los miembros presentes y pasados del Laboratoire d’anthropologie des populations du passé (UMR 5199 – PACEA, U. Bordeaux I) y del Proyecto Arqueológico Taller de Campo “Lomas de Lurín” (Departamento de Humanidades, PUCP), así como a O. Dutour, M. Ziolkowski, P. Murail y D. Castex. Se agradecen de igual manera los comenta-rios y sugerencias de los evaluadores anónimos.
Igualmente, agradezco a mis familiares y amigos por su paciencia y apoyo, en especial a Alex, Tadeo, Silvia, mis padres, Kati, Karla, y Greg.
Este trabajo se llevó a cabo gracias al generoso apoyo financiero del Minis-tère de l’Enseignement Supérieur et de la Recherche (Francia), Cementos Lima, S. A. (Perú) y PATL-PUCP (Perú). .

9 3
A P O R T A C I Ó N A R Q U E O L Ó G I C A A L C O N O C I M I E N T O | K A R I N A G E R D A U R A D O N I C
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.75-94
R e f e R e n c i a s
Andrews, Peter y Silvia Bello2006.“PatterninHumanBurialPractice”,enSocial Archaeology of Funerary Remains, Rebecca
GowlandyChristopherKnüsel,eds.,pp.14-29.Oxford,OxbowBooks.
Atalaya Huerta, Elmerms.Informe. Sector Noreste. Unidad de exposición A. Unidad de excavación N 123-126, E 127-131.
1996.InformeinternodelProyectoArqueológicoTabladadeLurín,Lima,PontificiaUniversidadCatólicadelPerú.
Balbuena Cotlear, Lucía1996.“EntierrosmúltiplesenTabladadeLurín:unaaproximaciónalritualfunerario”.Tesisde
Licenciatura,Lima,PontificiaUniversidadCatólicadelPerú.
Boddington, Andrew1987.“Chaos,DisturbanceandDecayinanAnglo-SaxonCemetery”,enDeath, Decay, and
Reconstruction: Approaches to Archaeology and Forensic Science, AndrewBoddington,A.NeilGarland,yRobC.Janaway, eds.,pp.27-42.Manchester,ManchesterUniversityPress.
Brothwell, Don R. 1987.“DecayandDisorderintheYorkJewburySkeletons”,enDeath, Decay, and Reconstruction:
Approaches to Archaeology and Forensic Science, AndrewBoddington,A.NeilGarland,yRobC.Janaway,eds.,pp.22-26.Manchester,ManchesterUniversityPress.
Duday, Henri1986.“Contributiondesobservationsostéologiquesàlachronologieinternedessépulturescollectives”,
en Anthropologie physique et Archéologie. Méthodes d’étude des sépultures: actes du colloque de Toulouse, 4, 5 et 6 novembre 1982, HenriDudayyClaudeMasset,eds.,pp.51-59.París,CNRS.
Duday, Henri1987.“Laquantificationdesresteshumains.Applicationàl’étudedessépulturesàincinérationsoudes
differentielsautresquelaconservation”,en Méthodes d’études des sépultures, compte-rendu de la 2e table ronde de la R.C.P. 742 du CNRS, St. Germain-en-Laye (16 et 17 mai 1987),pp.17-22.París,R.C.P.742duCNRS,
Duday, Henri2005.“L’archéothanatologie”enObjets et méthodes en paléoanthropologie, OlivierDutour,Jean-
JacquesHublinyBernardVandermeersch,eds.,pp.153-215.París,Comitédestravauxhistoriquesetscientifiques.
Duday, Henri2006.“ArchaeothanatologyortheArchaeologyofDeath”,en Social Archaeology of Funerary Remains,
RebeccaGowlandyChristopherKnüsel,eds.,pp.30-56.Oxford,OxbowBooks.
Duday, Henri, Patrice Courtaud, Éric Crubézy, Pascal Sellier y Anne-Marie Tillier1990.“L’anthropologiedeterrain”,Bulletins et mémoires de la Société d’Anthropologie de Paris,N.S.,
T.2,No.3-4,pp.29-50.
Duday, Henri y Marc Guillon 2006.“UnderstandingtheCircumstancesofDepositionwhentheBodyisSkeletonized”,enForensic
Anthropology and Forensic Medicine: Complementary Sciences From Recovery to Cause of Death, AuroreSchmitt,EugéniaCunhayJoãoPinheiro,eds.,pp.117-158.Totowa,NewJersey,HumanaPress.
García Malpartida, Ricardoms.Informe de la segunda práctica pre-profesional: la estructura funeraria 2 en Tablada de Lurín.
1994.InformeinternodelProyectoArqueológicoTabladadeLurín,Lima,PontificiaUniversidadCatólicadelPerú.

9 4
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.75-94
Gerdau Radonic, Karina2007.“LestombescollectivesdeTabladadeLurín(ValléedeLurín,Pérou;Ier-IIIes.ap.J.C.)”.Tesisde
Doctorado,Burdeos,Francia,UniversitéBordeauxI.
Gerdau Radonic, Karina y Krzysztof Makowski 2011.“LassepulturascolectivasdeTabladadeLurín:unaperspectivadesdelaantropologíabiológica”,en
Arqueología peruana: homenaje a Mercedes Cárdenas, LuisaVetter,SandraTellezyRafaelVega-Centeno,eds.,pp.145-176.Lima,InstitutoRiva-Agüero,PontificiaUniversidadCatólicadelPerú.
Haglund, William D. y Marcella H. Sorg (eds.)1997.Forensic Taphonomy: The Postmortem Fate of Human Remains.BocaRaton,FL,CRCPress.2002. Advances in Forensic Taphonomy. Method, Theory and Archaeological Perspectives. BocaRaton,
FL,CRCPress.
Herrmann, Bernd y Roelf D. Meyer1993.Südamerikanische Mumien aus vorspanischer Zeit: eine radiologische Untersuchung.Berlín,
StaatlicheMuseenzuBerlin–PreussischerKulturbesitz.
Lyman, R. Lee1994. Vertebrate Taphonomy.Cambridge,CambridgeUniversityPress.
Makowski, Krzysztof2002.“PowerandSocialRankingattheEndoftheFormativePeriod.TheLowerLurinValley
Cemeteries”,enAndean Archaeology I: Variations in Sociopolitical Organization, WilliamH.IsbellyHelaineSilverman,eds.,pp.89-120.NuevaYork,KluwerAcademic/PlenumPublishers.
2009a.“PoderyestatussocialafinesdelPeriodoFormativo:loscementeriosdelvallebajodeLurín”,enArqueología del período formativo en la cuenca baja de Lurín,RichardL.BurgeryKrzysztofMakowski,eds.,pp.209-236.Lima,FondoEditorialPUCP.
2009b.“TabladadeLurín:aspectoscronológicosdelaocupacióndelomascosterasdeAtocongo”,enArqueología del período formativo en la cuenca baja de Lurín, RichardL.BurgeryKrzysztofMakowski,eds.,pp.237-282.Lima,FondoEditorialPUCP.
Makowski, Krzysztof, Rafael Vega-Centeno, Iván Amaro, Hernán Carillo, Elmo León, Cecilia Rodríguez de March, Pamela Castro de la Mata, Jahl Dulanto, Elsa Tomasto, Lucía Balbuena, Elmer Atalaya, Manuel Gorriti, Iván Santos, Luis Salcedo, Glenda Escajadillo y Emma Arana
1996.Proyecto Arqueológico-Taller de Campo PUCP “Tablada de Lurín”: informe de las temporadas 1993/1994 y 1994/1995.Lima,InstitutoNacionaldeCultura.
Maureille, Bruno y Pascal Sellier1996.“Dislocationenordreparadoxal,mommificationetdecomposition:observationsethypothèses”,
Bulletinset Mémoires de la Société d’Anthropologie de Paris,n.s.,8(3-4),pp.313-327.
Pereira, Grégory1996.“PotrerodeGuadalupe:anthropologiefunéraired’unecommunautépré-tarasquedunorddu
Michoacán,Mexique”.TesisdeDoctorado,París,Francia,UniversitéParis1–laSorbonne.

9 5


ParalelosLA BIOLOGÍA DE LA CONSERVACIÓN APLICADA A LA ZOOARQUEOLOGÍA. LA SOSTENIBILIDAD DE LA CACERÍA DEL VENADO COLA BLANCA, ODOCOILEUS VIRGINIANUS (ARTIODACTYLA, CERVIDAE) EN AGUAZUQUE
m a r í a F e r n a n d a m a r T í n e z - p o l a n c o 9 9
LA ARQUEOFAUNA DE XCAMBÓ, YUCATÁN, MÉXICOc h r i s T o p h e r m G ö T z y Th e l m a n . s i e r r a s o s a 119
TAFONOMÍA DE HUESOS DE AVES. ESTADO DE LA CUESTIÓN Y PERSPECTIVAS DESDE EL SUR DEL NEOTRÓPICO
i s a b e l c r u z 147
PRIMER REGISTRO DE PERRO DOMÉSTICO PREHISPÁNICO (CANIS FAMILIARIS) ENTRE LOS GRUPOS CAZADORES RECOLECTORES DEL HUMEDAL DE PARANÁ INFERIOR (ARGENTINA)
a l e j a n d r o a c o s T a , d a n i e l l o p o n T e y c é s a r G a r c í a e s p o n d a 175
ARQUEOLOGÍA SUBACUÁTICA Y TAFONOMÍA: RECIENTES AVANCES EN EL ESTUDIO DE SITIOS FINIPLEISTOCÉNICOS SUMERGIDOS EN LA COSTA PACÍFICA DE CHILE CENTRAL
i s a b e l c a r T a j e n a F. , pa T r i c i o l ó p e z m . , d i e G o c a r a b i a s , c a r l a m o r a l e s y G a b r i e l Va r G a s 201

9 8

9 9
L A BIOLOGÍ A DE L A CONSERVACIÓN A PLICA DA A L A ZOOA RQU EOLOGÍ A: L A SOST EN IBI LI DA D DE L A CAC ER Í A DEL V ENA DO COL A BL A NCA, ODOCOI L EUS V I RGI N I A N US (A RTIODACT Y L A, C ERV I DA E), EN AGUA Z UQU E
maría Fernanda marTínez-polanco*
[email protected] de Ciencias Naturales, Universidad Nacional de Colombia
R e s u m e n El objetivo de este artículo es evaluar la cacería de
O. virginianus en Aguazuque, un sitio de cazadores-recolectores
tardíos de la sabana de Bogotá, a partir de elementos teóricos y
metodológicos de la biología de la conservación, como lo es el
concepto de sostenibilidad. Se utilizaron índices de diversidad
comúnmente utilizados en ecología, además de índices
normalmente empleados en zooarqueología como NISP y MNI.
En el caso de la cacería del venado en Aguazuque, se encontró
que ésta fue sostenible.
P A L A B R A S C L A V E :
Biología de la conservación, zooarqueología, sostenibilidad,
O. virginianus, sabana de Bogotá, cazadores recolectores tardíos.
Artículorecibido:1dejuniode2011|aceptado:14deoctubrede2011|modificado:23deoctubrede2011
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,294pp.ISSN1900-5407,pp.99-118
* MSc.,Antropología,UniversidadNacionaldeColombia.

1 0 0
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.99-118
the Biology of ConserVation applied to ZooarCheology: hunting sustainaBility of the white tail deer odoCoileus Virginianus (artiodaCtyla, CerVidae) in aguaZuque
ABSTR AC T The aim of this article is
to evaluate O. Virginianus hunting in
Aguazuque, a late hunter-gatherer site at the
Sabana de Bogotá by using theoretical and
methodological elements of conservation
biology, such as sustainability. In this study,
diversity indexes which are commonly
utilized in ecology were used as well as
in zooarchaeology with indexes like NISP
and MNI. In the case of the O. virginianus
hunting at Aguazuque the result is that it
was a sustainable activity.
K E Y W O R D S :
Conservation Biology, Zooarchaeology,
Sustainability, O. virginianus, Bogotá High
Plane, Late Hunters Gatherers.
a Biologia da ConserVação apliCada à Zooarqueologia: a sustentaBilidade da Caçada do Veado Cauda BranCa odoCoileus Virginianus (artiodaCtyla, CerVidae) em aguaZuque
RESUMO O objetivo deste artigo é avaliar
a caçada do O. virginianus em Aguazuque,
um lugar de caçadores-recolhedores
tardios na Sabana de Bogotá, a partir de
elementos teóricos e metodológicos da
biologia da conservação, como é o conceito
de sustentabilidade. Utilizaram-se índices
de diversidade comumente utilizados em
ecologia além de índices normalmente
empregados em zooarqueologia como NISP
e MNI. No caso da caçada do veado em
Aguazuque encontrou-se que foi sustentável.
P A L A V R A S C H A V E :
Biologia da conservação, zooarqueologia,
sustentabilidade, O. virginianus, sabana de
Bogotá, caçadores recolhedores tardios.

1 0 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.99-118
L A BIOLOGÍ A DE L A CONSERVACIÓN A PLICA DA A L A ZOOA RQU EOLOGÍ A: L A SOST EN IBI LI DA D DE L A CAC ER Í A DEL V ENA DO COL A BL A NCA, ODOCOI L EUS V I RGI N I A N US (A RTIODACT Y L A, C ERV I DA E), EN AGUA Z UQU E
m a r í a F e r n a n d a m a r T í n e z - p o l a n c o
El punto de inicio: conceptos claves de la biología de la conservación La biología de la conservación es una ciencia
multidisciplinaria que surge como respuesta a la crisis que enfrenta la diversi-dad biológica en la actualidad. Esta ciencia tiene tres propósitos: 1) Investigar y describir la diversidad que hay en el mundo; 2) Entender los efectos de las actividades humanas sobre las especies, las comunidades y los ecosistemas y 3) Desarrollar enfoques interdisciplinarios que tengan como objetivo proteger y restaurar la diversidad biológica (Primack, 2000).
La biología de la conservación es considerada como una disciplina de cri-sis que debe responder rápidamente a diferentes problemas actuales (Groom et al., 2006). Para lograr este objetivo se debe conocer con detalle el problema por resolver y poner en práctica conceptos de biología y ecología, además de incluir información proveniente de otras áreas de conocimiento que puedan brindar más elementos para tomar decisiones acertadas de manejo y conserva-ción (Primack, 2000).
Las mayores causas de la pérdida de la diversidad biológica son: 1) Des-trucción, fragmentación y degradación de hábitats, 2) Contaminación, 3) Sobreexplotación, 4) Introducción de especies exóticas y 5) Enfermedades. Estas causas son producto de la actividad humana, el incremento de la pobla-ción mundial, que ha promovido el excesivo uso de los recursos naturales, ade-más de los procesos de industrialización que se han dado en los últimos siglos (Primack, 2000). La mayor causa de pérdida de diversidad biológica es, justa-

1 0 2
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.99-118
mente, la pérdida de hábitat, que ha reducido en gran medida las zonas bosco-sas aptas para la conservación de la biodiversidad (IAvH, 1997).
La contaminación es un factor indirecto de pérdida de biodiversidad; aunque su impacto puede no ser evidente, sí podría estar afectando la supervi-vencia de una especie (Primack, 2000). De la misma forma, las enfermedades pueden tener una incidencia indirecta en la conservación de las especies; usual-mente, éstas son un efecto colateral de la introducción de especies exóticas, las cuales, además de competir con los organismos locales, pueden traer consigo enfermedades que pueden afectarlos (IAvH, 1997).
La sobreexplotación es la segunda causa de pérdida de la diversidad bio-lógica (Reynolds y Peres, 2006). Sin embargo, durante mucho tiempo los gru-pos humanos han convivido y explotado muchas especies tanto animales como vegetales sin llevarlas a la extinción (Ojasti, 2000). Aunque, en otros casos, la fuerte sobreexplotación que han hecho los humanos de una especie la ha lle-vado a la extinción, como es el caso de la paloma migratoria (Ectopistes migra-torius), que se extinguió en el siglo pasado (Reynolds y Peres, 2006).
Por otro lado, en la medida en que la tecnología se ha hecho más efi-ciente, la cacería se ha convertido en una seria amenaza para muchas especies. Además, la introducción de la cacería de subsistencia dentro de la economía de mercado ha hecho que muchas especies de fauna silvestre se encuentren en peligro (Primack, 2000). En algunos lugares del mundo la sobreexplotación ha sido tan dramática que ha dejado bosques vacíos (Empty forest), en donde los mamíferos grandes son los que más se han visto afectados, dado que son las presas favoritas de los cazadores (Redford, 1992).
Sin embargo, por definición, las poblaciones biológicas pueden soportar ciertos grados de explotación humana, debido a que las poblaciones son reno-vables; esto quiere decir que pueden compensar los efectos de la mortalidad con la natalidad y mantener constante el tamaño poblacional dentro de cier-tos límites (Reynolds y Peres, 2006). Esta característica de las poblaciones se denomina denso-dependencia (Caughley y Sinclair, 1994). La teoría biológica de la sostenibilidad se basa en este principio (Primack, 2000; Reynolds y Peres, 2006). La máxima cosecha sostenible (Maximum Sustainable Yield [MSY]) es la cantidad máxima de un recurso que puede ser cosechada cada año depen-diendo de la capacidad que tenga la población de renovarse por el crecimiento poblacional natural (Begon, 2002; Bodmer et al., 1997).
Se puede decir, entonces, a partir de la teoría biológica de la sostenibilidad, que una cosecha es insostenible cuando los niveles de extracción son mayores que los niveles de producción o reemplazo de la población, lo que puede llevar a que una especie se agote hasta su extinción (Robinson y Redford, 1991; Robin-

1 0 3
L A B I O L O G Í A D E L A C O N S E R V A C I Ó N | M A R Í A F E R N A N D A M A R T Í N E Z - P O L A N C O
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.99-118
son y Bennett, 1999). Una población expuesta a la sobreexplotación presenta una o varias de las siguientes características: 1) densidades bajas (Alvard, 1999, Hart, 1999; Peres, 1999), 2) reducción del tamaño promedio del cuerpo de los individuos, 3) reducción del promedio de la edad de la primera reproducción (Hart, 1999), 4) reducción en el número de animales de determinada clase de edad (Altrichter, 2005; Leeuwenberg y Robinson, 1999; Maffei, 2001 y 2003), 5) potencial disminución de la población (Robinson y Redford, 1991), 6) inminente extinción de poblaciones de especies vulnerables (Alvard et al., 1997; Alvard, 1999; Bennett et al., 1999). Además, la sobreexplotación puede tener un efecto en la producción, estructura y composición de la comunidad biológica de la cual la población explotada forma parte, debido a que, al disminuir o desaparecer la taxa de más alto rendimiento, se empiezan a cazar otras especies de menor pro-ductividad (Hart, 1999). Para lograr que la cacería sea sostenible a largo plazo, un grupo humano puede manejar una población silvestre con el fin de explotarla sin agotarla (Caughley y Sinclair, 1994; Sinclair et al., 2006).
Para evaluar la sostenibilidad de la cacería en la actualidad, se utilizan diferentes métodos; por una parte, se realiza una evaluación al nivel de comu-nidad analizando qué especies son cazadas, a partir de entrevistas y visitas a los cazadores, donde se pesan y miden los animales cazados para calcular su aporte proteico (Townsend, 1999; Fitzgibbon et al., 1999; Hill y Padwe, 1999; Mena et al., 1999). También se comparan las densidades de poblaciones animales en lugares con diferentes intensidades de cacería (Alvard, 1999; Fitzgibbon et al., 1999; Hill y Padwe, 1999; Mena et al., 1999). El otro nivel de análisis utilizado es el de poblaciones; en este caso, se analizan aspectos como el sexo, la edad y el tamaño de los animales cazados de cierta especie (Altrichter, 2005; Hart, 1999; Leeuwenberg y Robinson, 1999; Maffei, 2001 y 2003).
La sostenibilidad y la zooarqueologíaA pesar de que el estado actual de las poblaciones animales es producto de una dinámica histórica, son muy pocos los casos en los que se incluye una perspectiva temporal en los estudios de manejo y conservación de fauna sil-vestre. Recientemente se ha venido dando una nueva tendencia dentro de la zooarqueología, interesada en aportar elementos prácticos útiles para los pla-nes de manejo de ciertas especies (Lyman, 1996; Lyman y Cannon, 2004). La zooarqueología puede brindar elementos de análisis a la sostenibilidad, ya que puede indagar sobre el manejo de la fauna por parte de diversos grupos huma-nos en diferentes momentos y lugares del planeta (Frazier, 2007). La sosteni-bilidad se puede estudiar a partir de muestras faunísticas analizando aspectos tales como 1) Cambios en la composición de la caza a través del tiempo (Bovy,

1 0 4
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.99-118
2007; Carder et al., 2007; Emery, 2007; McKechnie, 2007; Morwood et al., 2008), 2) Aumento en el número de individuos cazados de una especie deter-minada (McNiven y Bedingfield, 2008; Sapir-Hen et al., 2009), 3) Selección de animales más jóvenes de cierta especie después de un período determinado (Lyman, 1987; Etnier, 2004 y 2007; Walker, 2000), 4) Disminución de la talla promedio de los animales cazados (Blick, 2007; Carder et al., 2007; Hill Jr. et al., 2008; McKechnie, 2007; Whitaker, 2008).
La biología de la conservación y la zooarqueología pueden aportarse entre sí elementos tanto teóricos como metodológicos que permitan entender el pasado a la luz del presente, así como comprender la relación entre gru-pos humanos y animales que se dieron en el pasado, y pueden dar claves para lograr un mejor manejo, conservación y protección de aquellas especies que se encuentran en peligro en la actualidad (Lyman y Cannon, 2004).
En zooarqueología se utiliza el modelo de la presa de la teoría del forrajeo óptimo, para evaluar qué presas son utilizadas por el grupo humano estudiado y qué partes de las presas se están transportando hasta el lugar de asentamiento (Byers et al., 2005; Byers y Ugan, 2005; Reitz y Wing, 2008; Ugan, 2005). Para determinar qué presas son utilizadas se analizan las abundancias relativas de los diferentes taxa representados (Reitz y Wing, 2008). Para conocer las partes de las presas que son utilizadas se llevan a cabo análisis de la distribución dife-rencial (Reitz y Wing, 2008; Byers et al., 2005; Byers y Ugan, 2005; Ugan, 2005).
Grayson y Delpech (1998) proponen que para entender modelos como la amplitud de la dieta (Diet breadth), que también se desprende de la teoría del forrajeo óptimo, se deben emplear análisis combinados que incluyan el número de taxas, el número de especímenes, la fragmentación y la distribución de las partes del esqueleto de los animales presentes en la muestra, para poder evi-denciar claramente si ocurrieron cambios en la dieta de un grupo humano a lo largo del tiempo.
Usualmente, el modelo de la presa es utilizado para probar hipótesis de sobreexplotación en el pasado de megafauna (Byers y Ugan, 2005), de mamí-feros grandes (Broughton, 2002; Broughton y Bayham, 2003; Byers et al., 2005; Emery, 2007), entre otros. A la luz de este modelo, se espera que los cazadores siempre elijan aquellas presas que les generen mayores beneficios. Esto puede ser un riesgo, porque si se caza siempre la misma especie, se puede generar una extinción, lo que le significaría más costos a los cazadores (Blick, 2007). En este caso, tendrían que empezar a recorrer distancias mayores para cazar la misma especie o cambiar de presa; generalmente, se inicia la cacería de animales más pequeños, que representan menos ganancias (Ugan, 2005; Byers et al., 2005; Byers y Ugan, 2005).

1 0 5
L A B I O L O G Í A D E L A C O N S E R V A C I Ó N | M A R Í A F E R N A N D A M A R T Í N E Z - P O L A N C O
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.99-118
Sin embargo, los cazadores pueden elegir otro tipo de estrategias que impliquen el manejo de las especies; estas medidas podrían incluir una cacería selectiva de animales adultos que hayan tenido la oportunidad de reproducirse (Martínez-Polanco, 2008a), o cazar más animales de un sexo o de otro (Etnier, 2007). Otra estrategia de manejo puede ser cazar varias especies a la vez; de esta forma, no se está agotando una sola especie (Emery, 2007). También puede existir otra clase de estrategias que impliquen la movilidad, de tal modo que no siempre se esté cazando en el mismo lugar (Whitaker, 2008).
Un punto en común: la sostenibilidad de la cacería de O. virginianus en la sabana de BogotáEsta investigación surge de un problema de manejo y conservación actual: la extinción local de O. virginianus de la sabana de Bogotá, de la cual no se conocen sus orígenes (Martínez-Polanco, 2008b). Se sabe que O. virginianus fue la especie más importante para los humanos que habitaron la sabana de Bogotá, desde los grupos de cazadores-recolectores hasta los grupos alfareros Herrera y Muisca, porque era la especie de mayor tamaño que se encontraba en la zona y la que mayores beneficios representaba. O. virginianus no sólo fue utilizado como alimento; también sus pieles pudieron haber sido empleadas como abrigo, al igual que sus huesos y astas como herramientas (Peña y Pinto, 1996). Posteriormente, en la época de la Colonia, los cronistas hacen referencia a la abundancia de esta especie en el área (Fernández Piedrahíta, 1987 [1688]; Rodríguez Freyle, 1985 [1636]; Simón, 1981 [1625]).
El O. virginianus es una especie de amplia distribución que se expande desde Norteamérica hasta el norte de Suramérica (Einsberg, 1989; Emmons, 1999; Smith, 1991). En Colombia, O. virginianus se distribuye en la Orinoquia y la Amazonia, en la planicie del Caribe y en las cordilleras Central, Oriental, y en Nariño (López-Arévalo y González-Hernández, 2006).
Esta especie se caracteriza por poseer un marcado dimorfismo sexual propio de los cérvidos. Los machos son más grandes que las hembras, pesan unos 50 kg y poseen astas ramificadas; las hembras pesan unos 30 kg y no pre-sentan astas. Las hembras viven en grupos junto con los cervatillos; los machos tienen hábitos solitarios. Son animales muy adaptables, incluso en su alimen-tación, lo que les permite vivir en diferentes tipos de hábitats (Smith, 1991; Emmons, 1999; Teer, 1994) (ver la figura 1).

1 0 6
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.99-118
a. b.
Figura1.a.HembraO. virginianus;b.MachoO. virginianusPuertoGaitán,Meta.Foto:ArchivopersonalMartínez-Polanco.
Teniendo en cuenta que la sobreexplotación es la segunda causa de pérdida de diversidad, en el caso de O. virginianus en la sabana de Bogotá es importante realizar estudios de largo alcance temporal acerca del impacto que pudieron causar los grupos humanos sobre las poblaciones de esta especie.
El período precerámico de la sabana de Bogotá se encuentra bien documen-tado porque se han excavado varios yacimientos como El Abra (Hurt et al., 1976), Tequendama I (Correal y Van der Hammen, 1977), Nemocón y Sueva (Correal, 1979), Tibitó (Correal, 1981), Chía (Ardila, 1984), Vistahermosa (Correal, 1987), Aguazuque (Correal, 1990), Neusa (Rivera, 1992), Checua (Groot, 1992) y Galindo (Pinto, 2003). En todos estos sitios se han encontrado abundantes evi-dencias de O. virginianus; sin embargo, no se conoce el impacto de la cacería en este período desde una perspectiva regional (Martínez-Polanco, 2008a).
El yacimiento arqueológico de Aguazuque fue excavado a finales de la década de los ochenta por Gonzalo Correal (Correal, 1990). En este sitio se encon-traron cinco ocupaciones precerámicas datadas entre 2725 AP y 5025 AP y una cerámica de dif ícil interpretación (Correal, 1990). Todas las ocupaciones prece-rámicas tienen en común la presencia de instrumentos líticos de tipo abriense, elementos utilizados para el manejo de vegetales (molinos y yunques) y nume-rosos restos de O. virginianus (Correal, 1990). Éste es un sitio muy interesante, debido a que se encuentra en un área abierta, a diferencia de la mayoría de sitios de cazadores-recolectores, ubicados cerca de abrigos rocosos (Correal, 1990).
Partiendo de los conceptos expuestos anteriormente, el propósito de este artículo es evaluar la sostenibilidad de la cacería del O. virginianus utilizando la muestra arqueofaunística proveniente del yacimiento arqueológico de Agua-zuque. Para tal fin, se determinará si existieron cambios en la diversidad de

1 0 7
L A B I O L O G Í A D E L A C O N S E R V A C I Ó N | M A R Í A F E R N A N D A M A R T Í N E Z - P O L A N C O
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.99-118
especies cazadas a lo largo del tiempo, aplicando los índices de diversidad de Shannon-Weber (H) y el índice de uniformidad (E); además, se determinarán cambios en la representatividad del O. virginianus a lo largo de la ocupación utilizando el Número Mínimo de Individuos (NMI).
Evaluación de la sostenibilidad de la cacería de O. virginianus en Aguazuque
Área de estudioEl yacimiento arqueológico Aguazuque se encuentra ubicado en el predio de la hacienda del mismo nombre, en el municipio de Soacha, Cundina-marca (ver la figura 2). En este sitio se realizaron dos cortes estratigráficos. El corte 1 abarcó un área de 12 m2, y el corte 2, un área de 64 m2. Ambos cortes fueron excavados siguiendo niveles estratigráficos naturales y se utili-zaron cernidores y tamices para recuperar evidencias de pequeños animales y vegetales (Correal, 1990).
El área de Aguazuque en la actualidad se caracteriza por tener una vege-tación de alta montaña; sin embargo, cuando estuvo ocupada se encontraba próxima a los dilatados pantanos remanentes del gran lago pleistocénico que cubría la sabana de Bogotá (Correal, 1990). Como se mencionó anteriormente, se encontraron seis ocupaciones; sin embargo, la sexta se encontró muy alte-rada, y se excluyó del presente análisis. Estas ocupaciones se prolongaron
Figura2.LocalizacióngeográficadeAguazque.(Fuente:imagensatelitalGoogleEarth,consultadaenagostode2007).

1 0 8
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.99-118
durante períodos de cierta amplitud y fueron ejercidas por grupos humanos más o menos densos (ver la tabla 1) (Correal, 1990).
Tabla1.FechasdelasocupacionesencontradasenAguazuque(Correal,1990).
Ocupación Fecha
1 5025±40 AP
2 4030±35 AP
3 3850±35AP
4 3400-2800AP
5 2725±35AP
Una característica muy interesante de Aguazuque es su ubicación geográ-fica, que les permitió a los grupos humanos que lo ocuparon acceder a recur-sos, no sólo del área de la sabana, sino también de otros provenientes del valle del río Magdalena (Correal, 1990).
Materiales Para realizar este trabajo se utilizó la muestra arqueofaunística de O. virgi-nianus proveniente del sitio arqueológico Aguazuque. El material se encuentra depositado en la Colección de Arqueología del Instituto de Ciencias Naturales de la Universidad Nacional de Colombia, sede Bogotá. Se trabajó con los mate-riales catalogados como ICN 661 hasta ICN 740. Fueron analizados los mate-riales provenientes del corte 2 de Aguazuque; en dicho corte se identificaron 3.524 elementos óseos (Martínez-Polanco, 2008a).
MétodosMedidas de diversidad: para calcular los índices de diversidad, se determinó el Número de Restos (NR) o Number of Identified Specimens (NISP) de la muestra de O. virginianus de Aguazuque (Martínez-Polanco, 2008a); además, se utiliza-ron los datos publicados sobre el número de restos óseos de fauna colectados en Aguazuque (Correal, 1990) y el número de restos del género Cavia analiza-dos por Pinto et al. (2002) para la revisión de este género en el país.
Índice de diversidad de Shannon-Weaver (H): este índice tiene en cuenta el número de taxa y la abundancia relativa de cada taxón, para evaluar la hete-rogeneidad de la muestra (Krebs, 1985; Reitz y Wing, 2008). El índice se calcula de la siguiente forma:
H=_Σs
i=1(pi) (Log pi)

1 0 9
L A B I O L O G Í A D E L A C O N S E R V A C I Ó N | M A R Í A F E R N A N D A M A R T Í N E Z - P O L A N C O
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.99-118
donde:pi = es la abundancia relativa de cada taxa en la muestra.S = número de taxa.Con este índice, las muestras con una distribución uniforme entre los
taxa tienen un valor mayor de H que aquellas muestras con el mismo número de taxa pero sin distribución uniforme (Grayson, 1984; Reitz y Wing, 2008).
Índice de uniformidad (Evenness index –E–): este índice mide la abundan- este índice mide la abundan-cia proporcional de cada taxa en la muestra. Los valores más altos indican una distribución uniforme en la muestra, es decir, una representación equitativa de cada taxa en la misma. Valores pequeños de este índice indican el predominio de una o pocas especies en la muestra (Grayson, 1984; Magurran, 1988; Reitz y Wing, 2008), y se calcula de la siguiente manera:
E = H/Log Sdonde:H = índice de diversidad de Shannon-WeaverS = número de taxaNúmero Mínimo de Individuos (NMI): el Número Mínimo de Individuos
(NMI) se puede definir como el menor número posible de individuos de una especie que pueden existir en una muestra zooarqueológica (Reitz y Wing, 2008). Para determinar el NMI se utilizaron las tibias, que fueron el elemento más frecuente de la muestra arqueofaunística de O. virginianus de Aguazuque (n = 220). Se tuvieron en cuenta la lateralidad, la edad (grado de fusión de las epífisis), el sexo y la talla, para determinar el NMI (Reitz y Wing, 2008).
ResultadosMedidas de diversidad: el número de especies cazadas en Aguazuque varió entre 12 y 16. Estas especies comprenden a peces como el capitán (Eremo-philus mustisii), la guapucha (Grundulus bogotensis) y el capitán pequeño (Pygidium bogotensis); reptiles como tortugas (Kinosternon leucostomum) y cocodrilos (Crocodylia); aves como patos (Anatidae), pollas de agua (Ralli-dae) y pavas (Cracidae) (Correal, 1990). La muestra de mamíferos está com-puesta por armadillos (Dasypus novemcinctus), ocelotes (Leopardos par-dalis), zorros (Cerdocyon thous), venados de cola blanca (Odocoileus virgi-nianus), venados soche (Mazama sp.), pecarís (Tayassu pecari), curíes (Cavia sp.) y pacas (Cuniculus paca) (Correal, 1990). Los valores del índice de diver-sidad de Shannon variaron entre 1,41 y 0,83 entre las diferentes ocupacio-nes, encontrándose el mayor valor en la primera ocupación, y el menor, en la cuarta (ver la tabla 2).

1 1 0
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.99-118
Tabla2.MedidasdediversidaddelamuestraarqueofaunísticadeAguazuque.
OcupaciónS
Númerodesp.cazadasH
ÍndicedediversidadE
Índicedeequitabilidad
1 16 1,41 0,50
2 13 1,14 0,44
3 13 1,21 0,47
4 12 0,83 0,33
5 13 0,86 0,33
Los valores del índice de uniformidad oscilan entre 0,3 y 0,5 a lo largo de las cinco ocupaciones de Aguazuque. Hay una tendencia a la reducción tanto en el índice de diversidad H como en el de uniformidad E, lo que evidencia una composición de la caza dominada por pocas especies (ver la tabla 2).
Número Mínimo de Individuos (NMI): en total, en Aguazuque se iden-tificaron 105 individuos de O. virginianus. El NMI muestra una estabilidad en las tres primeras ocupaciones, con valores que varían entre 9 y 12 individuos. Posteriormente, se evidencia una tendencia al aumento de individuos cazados de O. virginianus desde la ocupación 3 hasta la 5, donde se observa el mayor NMI de toda la ocupación, con un total de 58 individuos, que representan el 55% de la muestra analizada (ver la figura 2 y la tabla 3).
Tabla3.NúmeroderestosyNúmeroMínimodeIndividuosdevenado colablanca,O. virginianus,cazadosenAguazuque.
Ocupación NISP NISP% NMI NMI%
1 214 8,04 10 9,52
2 267 10,03 9 8,57
3 248 9,31 12 11,43
4 536 20,13 16 15,24
5 1398 52,50 58 55,24
Total 2663 100,00 105 100,00
Síntesis y conclusionesEn este trabajo se abordó el tema de la sostenibilidad de la cacería del O. vir-ginianus en Aguazuque examinando varios aspectos de la caza. El primero de ellos fue diversidad de especies cazadas en la secuencia de ocupación, donde

1 1 1
L A B I O L O G Í A D E L A C O N S E R V A C I Ó N | M A R Í A F E R N A N D A M A R T Í N E Z - P O L A N C O
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.99-118
se esperaba encontrar un aumento en la diversidad de especies, generado por el agotamiento de O. virginianus, como lo predice el modelo de la presa de la teoría del forrajeo óptimo (Broughton, 2002; Emery, 2007). No obstante, el resultado de los índices de diversidad de Shannon y de equitabilidad no refleja esta tendencia. Por el contrario, el valor del índice de diversidad tiene una ten-dencia a disminuir con el paso del tiempo, lo cual puede reflejar más bien una tendencia a la especialización (Betts y Friesen, 2004) o intensificación (Earle, 1980) en la cacería del O. virginianus.
Es posible que la densidad poblacional humana no fuera tan alta y que los niveles de extracción no alcanzaran a agotar el recurso, como sí ha ocurrido en otros sitios arqueológicos. En el área maya, por ejemplo, la representatividad del O. virginianus disminuyó con el tiempo de ocupación, dado el aumento en la densidad humana. Esta situación provocó que los cazadores explotaran una mayor diversidad de especies pequeñas (Emery, 2007).
El segundo aspecto analizado para evaluar la sostenibilidad de la cace-ría fueron los cambios en la representatividad de esta especie a lo largo de la secuencia de ocupación de Aguazuque. Para tal fin, se determinó el número mínimo de individuos entre los diferentes estratos. Este análisis mostró que el aprovechamiento del O. virginianus fue estable en las cuatro primeras ocu-paciones, con una tendencia al aumento en las últimas, especialmente en la quinta. El aumento en el NMI puede ser un indicativo de cambios significativos en el aprovechamiento de este recurso.
En resumen, los análisis presentados en este trabajo evidencian que la cacería de O. virginianus fue una actividad sostenible en Aguazuque a lo largo de la secuencia de ocupación. Este resultado está de acuerdo con los hallaz-gos encontrados en otras investigaciones recientes que evidencian aprovecha-mientos sostenibles de recursos faunísticos en diferentes lugares del mundo y en diversas épocas (Carder et al., 2007; Etnier, 2004; Etnier, 2007; McKechnie, 2007; Whitaker, 2008).
Las características de historia de vida del O. virginianus (Smith, 1991), su capacidad de adaptarse con facilidad a diferentes ambientes (Smith, 1991; Emmons, 1999), sus comportamientos alimenticios (Teer, 1994; Galindo-Leal y Weber, 1998) y sus comportamientos sociales (Rodríguez y Vaughan, 1994) posibilitan que las poblaciones de O. virginianus tengan la capacidad, hasta cierto punto, de soportar extracciones de individuos sin que esto ponga en peli-gro a toda la población o la especie. Es posible que estas características del O. virginianus hubiesen favorecido su permanencia en Aguazuque, a pesar de la continua explotación que hicieron los cazadores-recolectores a lo largo de la secuencia de ocupación del sitio.

1 1 2
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.99-118
En la última ocupación de Aguazuque ya empiezan a observarse diferen-cias bastante marcadas respecto a las otras ocupaciones, ya que se aprovechó O. virginianus de una forma más intensiva, lo que evidencia otro tipo de apro-vechamiento del recurso. Este nuevo manejo de O. virginianus podría estar demostrando cambios en la organización social de los grupos humanos que habitaron Aguazuque, relacionados con un aumento poblacional y/o con un uso más permanente del sitio (Martínez-Polanco, 2008a). Del mismo modo que la población humana aumentó, creció la demanda de O. virginianus en Agua-zuque; sin embargo, esta hipótesis debe ser explorada más a fondo.
Es posible que la extinción de O. virginianus en el área de la sabana de Bogotá haya sido un evento relativamente reciente, probablemente más rela-cionado con la fragmentación y destrucción del hábitat que con la sobrecaza. Por tal razón, es conveniente seguir haciendo investigaciones tanto históricas como arqueológicas que den luces sobre el proceso del agotamiento del O. vir-ginianus en la sabana y que sean una herramienta a la hora de proponer estra-tegias de manejo y conservación de la especie, no sólo en la sabana de Bogotá, sino en todo el país. .

1 1 3
L A B I O L O G Í A D E L A C O N S E R V A C I Ó N | M A R Í A F E R N A N D A M A R T Í N E Z - P O L A N C O
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.99-118
R e f e R e n c i a s
Altrichter, Mariana2005.“TheSustainabilityofSubsistenceHuntingofPeccariesintheArgentineChaco”,Biological
Conservation(126),pp.351-362.
Alvard, Michael1999.“TheImpactofTraditionalSubsistenceHuntingandTrappingonPreyPopulations:Datafrom
WanaHorticulturalistofUplandCentralSulawesi,Indonesia”,enHunting for Sustainability in Tropical Forest,JohnRobinsonyElizabethBennett,eds.,pp.214-232.NuevaYork,ColumbiaUniversityPress.
2000.“ThePotentialforSustainableHarvestbyTraditionalWanaHuntersinMorowaliNatureReserve,CentralSulawesi,Indonesia”,Human Organization59(4),pp.428-440.
Alvard, Michael, John Robinson, Kent Redford y Hillard Kaplan 1997.“TheSustainabilityofSubsistenceHuntingintheNeotropics”,Conservation Biology11(4),
pp.977-982.
Ardila, Gerardo1984.Chía: un sitio precerámico en la sabana de Bogotá.Bogotá,FIANBancodelaRepública.
Begon, Michael, Martin Mortimer y David Thompson2002. Population Ecology.Oxford,BlackwellScience.
Bennett, Elizabeth, Adrian Nyaoi y Jephte Sompud1999.“SavingBorneo’sBacon:TheSustainabilityofHuntinginSarawakandSabah”,enHunting for
Sustainability in Tropical Forest,JohnRobinsonyElizabethBennett,eds.,pp.305-324.NuevaYork,ColumbiaUniversityPress.
Betts, Matthew y T. Max Friesen2004.“QuantifyingHunter-gathererIntensification:AZooarchaeologicalCaseStudyfromArticCanada”,
Journal of Anthropological Archaeology23,pp.357-384.
Blick, Jef frey2007.“Pre-ColumbianImpactinTerrestrial,Intertidal,andMarineResources,SanSalvador,Bahamas
(A.D.950-1500)”,Journal of Nature Conservation 15,pp.174-183.
Bodmer, Richard, John Einsberg y Kent Redford1997.“HuntingandLikelihoodofExtinctionofAmazonianMammals”,Conservation Biology11,
pp.460-466.
Bovy, Kristine2007.“GlobalHumanImpactsorClimateChange?:ExplainingtheSootyShearwaterDeclineatthe
MinardSite,WashingtonState,USA”,Journal of Archaeological Science34,pp.1087-1097.
Broughton, Jack2002.“PreySpatialStructureandBehaviorAffectArchaeologicalTestofOptimalForagingModels:Examples
fromtheEmeryvilleShellmoundVertebrateFauna”,World Archaeology34(1),pp.60-83.
Broughton, Jack y Frank Bayham2003.“ShowingOff,ForagingModels,andtheAscendanceofLarge-gameHuntingintheCalifornia
MiddleArchaic”,American Antiquity68(4),pp.783-789.
Byers, David, Craig Smith y Jack Broughton 2005.“HoloceneArtiodactylsPopulationHistoriesandLargeGameHuntingintheWyomingBasin,
USA”,Journal of Archaeological Science32,pp.125-142.

1 1 4
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.99-118
Byers David y Andrew Ugan2005.“ShouldWeExpectedLargeGameSpecializationintheLatePleistocene?AnOptimalForaging
PerspectiveonEarlyPaleoindianPreyChoice”,Journal of Archaeological Science32, pp.1624-1640.
Carder, Nanny, Elizabeth Reitz y John Crock2007.“FishCommunitiesandPopulationsDuringthePost-SaloidPeriod(AD600/800-1500),Anguilla,
LesserAntillas”,Journal of Archaeological Science34,pp.588-599.
Caughley, Graeme y Anthony R. E. Sinclair 1994.Wildlife Ecology and Management.Cambridge,BlackwellScience.
Correal, Gonzalo1979.Investigaciones arqueológicas en los abrigos rocosos de Nemocón y Sueva.Bogotá,FIANBanco
delaRepública.1981.Evidencias culturales y megafauna pleistocénica en Colombia.Bogotá,FIANBancodelaRepública.1987.“ExcavacionesarqueológicasenMosquera”,Revista de Estudiantes de Antropología,Universidad
NacionaldeColombia3(1),pp.13-17.1990.Aguazuque, evidencias de cazadores recolectores y plantadores en la altiplanicie de la cordillera
Oriental.Bogotá,FIANBancodelaRepública.
Correal, Gonzalo y Thomas Van der Hammen1977.Investigaciones arqueológicas en los abrigos rocosos del Tequendama. 11.000 años de
prehistoria en la sabana de Bogotá.Bogotá,BancoPopular.
Earle, Timothy1980.“AModelofSubsistenceChange”,enModeling Change in Prehistoric Subsistence Economies,
TimothyEarleyA.Christenson,eds.,pp.1-29.NuevaYork,AcademicPress.
Einsberg, John1989.Mammals of the Neotropics1.Chicago,ChicagoPress.
Emery, Kit ty2007.“AssessingtheImpactofAncientMayaAnimalUse”,Journal for Nature Conservation15,
pp.184-195.
Emmons, Louis1999.Mamíferos de los bosques húmedos de América tropical.SantaCruz,EditoralFAN.
Etnier, Michael2004.“ThePotentialofArchaeologicalDatatoGuidePinnipedManagementDecisionsintheEastern
NorthPacific”,enZooarchaeology and Conservation Biology,LeeLymanyKennethCannon,eds.,pp.88-102.SaltLakeCity,TheUniversityofUtahPress.
2007.“DefiningandIdentifyingSustainableHarvestofResources:ArchaeologicalExamplesofPinnipedHarvestintheEasternNorthPacific”,Journal for Nature Conservation15,pp.196-207.
Fernández Piedrahíta, Lucas/1688/1987.Historia general de la conquista del Nuevo Reino de Granada.SantanderdeQuilichao,
Carvajal.
Fitzgibbon, Clare, Hezron Mogaka y John Fanshawe1999.“ThreatenedMammals,SubsistenceHarvesting,andHighHumanPopulationDensities:ARecipe
forDisaster?”,enHunting for Sustainability in Tropical Forest, JohnRobinsonyElizabethBennett,eds.,pp.154-167.NuevaYork,ColumbiaUniversityPress.
Frazier, J.2007.“SustainableUseofWildlife:TheViewfromArchaeozoology”,Journal for Nature Conservation15,
pp.163-173.

1 1 5
L A B I O L O G Í A D E L A C O N S E R V A C I Ó N | M A R Í A F E R N A N D A M A R T Í N E Z - P O L A N C O
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.99-118
Galindo-Leal, Carlos y Manuel Weber1998.El venado de la Sierra Madre Occidental.México,Edicusa-Conabio.
Grayson, Donald1984.Quantitative Zooarchaeology.Washington,AcademicPress.
Grayson, Donald y Françoise Delpech1998.“ChangingDietBreadthintheEarlyUpperPalaeolithicofSouthwesternFrance”,Journal of
Archaeological Science25,pp.1119-1129.
Groom, Martha, Gary Meffe y Ronald Carrol 2006.Principles of Conservation Biology.Sunderland,M.ASinauer.
Groot, Ana María1992.Checua. Una secuencia cultural entre 8500 y 3000 años antes del presente. Bogotá,FIANBanco
delaRepública.
Hart, John1999.“ImpactandSustainabilityofIndigenousHuntingintheIturiForest,Congo-Zaire:AComparison
ofUnhuntedandHuntedDuikerPopulations”,enHunting for Sustainability in Tropical Forest,JohnRobinsonyElizabethBennett,eds.,pp,106-153.NuevaYork,ColumbiaUniversityPress.
Hill, Kim y Jonathan Padwe1999.“SustainabilityofAchéHuntingintheMbaracayuReserve”,enHunting for Sustainability in Tropical
Forest,JohnRobinsonyElizabethBennett,eds.,pp.79-195.NuevaYork,ColumbiaUniversityPress.
Hill Jr., Matthew, Matthew Hill y Christopher Widga2008.“LateQuaternaryBisonDiminutionontheGreatPlainsofNorthAmerica:EvaluatingtheRoleof
HumanHuntingVersusClimateChange”,Quaternary Science Reviews27,pp.1752-1771.
Hurt, Wesley, Thomas Van der Hammen y Gonzalo Correal1976.The El Abra Rockshelters, Sabana de Bogotá, Colombia, South America.Indiana,Indiana
UniversityMuseum,IndianaUniversityPress.
IAvH1997.Informenacionalsobreelestadodelabiodiversidad.Tomo2,Causas de pérdida de la
biodiversidad.Bogotá,IAvH.
Krebs, Charles1985.Ecología: estudio de la distribución y la abundancia.México,Harla.
Leeuwenberg, Frans y John Robinson1999.“TraditionalManagementofHuntingbyaXavanteCommunityinCentralBrazil:TheSearchfor
Sustainability”,enHunting for Sustainability in Tropical Forest,JohnRobinsonyElizabethBennett,eds.,pp.375-394.NuevaYork,ColumbiaUniversityPress.
López-Arévalo, Hugo y Andrés González-Hernández2006.“VenadosabaneroOdocoileus virginianus”,enLibro rojo de los mamíferos de Colombia,José
Vicente,Rodríguez-Mahecha;MichaelAlberico,FernandoTrujilloyJeffJogenson,eds., pp.114-120.Bogotá,SerieLibrosRojosdeEspeciesAmenazadasdeColombia,ConservaciónInternacionalyMinisteriodeAmbienteyDesarrolloTerritorial.
Lyman, Lee1987.“OntheAnalysisofVertebrateMortalityProfiles:SampleSize,MortalityTypeandHunting
Pressure”,American Antiquity52(1),pp.125-142.1996.“AppliedZooarchaeology:TheRelevanceofFaunalAnalysistoWildlifeManagement”,World
Archaeology28(1),pp.110-125.
Lyman, Lee y Kenneth Cannon 2004. Zooarchaeology and Conservation Biology.SaltLakeCity,TheUniversityofUtahPress.

11 6
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.99-118
Maffei, Leonardo2001.“Estructuradeedadesdelaurina(Mazama gouazoubira)enelChacoboliviano”,Mastozoología
Neotropical8(2),pp.149-155.2003.“TheAgeStructureofTapirs(Tapirus terrestris)intheChaco”,Tapir Conservation Newsletter of
the IUCN/SSC Tapir Specialist Group12(2).
Magurran, Ann1988.Ecological Diversity and Its Measurement.Princeton,Princeton,UniversityPress.
Martínez-Polanco, María Fernanda2008a.“LacaceríadelvenadocolablancaOdocoileus virginianusporcazadores-recolectorestardíos:
elcasodeAguazuque”.TesisinéditaMagisterenCiencias-Biología,UniversidadNacionaldeColombia.
2008b.“Delpasadoalpresente:breveanálisisdelestadodecincoespeciesdemamíferossilvestresenColombia”,Canto Rodado3,pp.95-112.
McKechnie, Iain2007.“InvestigatingtheComplexitiesofSustainableFishingataPrehistoricVillageonWestern
VancouverIsland,BritishColumbia,Canada”,Journal of Nature Conservation15,pp.208-222.
McNiven, Ian y Alice Bedingfield 2008.“PastandPresentMarineMammalHuntingRatesandAbundances:Dugong(Dugong dugon)
EvidencefromDabangaiBoneMound,TorresStrait”,Journal of Archaeological Science35(2), pp.505-515.
Mena, Patricio, Jody Stallings, Jhanira Regalado y Rubén Cueva1999“TheSustainabilityofCurrentHuntingPracticesbytheHuaorani”,Hunting for Sustainability in
Tropical Forest,enJohnRobinsonyElizabethBennett,eds.,pp.57-78.NuevaYork,ColumbiaUniversityPress.
Morwood, M. J., T. Sutikna, E.W. Saptomo, K.E. Westaway, Jatmiko R. Awe Due; M.W. Moore, Dwi Yani Yuniawati, P. Haidi, J-x. Zhao, C.S.M. Turney, K. Fif ield, H. Allen y R. P. Soejono
2008.“Climate,PeopleandFaunalSuccessiononJava,Indonesia:EvidencefromSongGupuh”,Journal of Archaeological Science35,pp.1776-1789.
Ojasti, Juhani2000. Manejo de fauna silvestre neotropical. Washington,SmithsonianInstitution/MABProgram.
Peña, Germán y María Pinto 1996.Mamíferos más comunes en sitios precerámicos de la sabana de Bogotá: guía ilustrada para
arqueólogos.Bogotá,AcademiaColombianadeCienciasExactas,FísicasyNaturales,ColecciónJulioCarrizosaValenzuela:6.
Peres, Carlos1999.“EvaluatingtheImpactandSustainabilityofSubsistenceHuntingatMultipleAmazonianForest
Sites”,enHunting for sustainability in Tropical Forest,JohnRobinsonyElizabethBennett,eds., pp.57-78.NuevaYork,ColumbiaUniversityPress.
Pinto, María2003. Galindo, un sitio a cielo abierto de cazadores-recolectores en la sabana de Bogotá (Colombia).
Bogotá,FIANBancodelaRepública.
Pinto, María, Herly Zúñiga y Olga Torres2002. Estudio sistemático del género Cavia, Pallas, 1766 (Rodentia: Caviidae) en Colombia. Revisión
del registro arqueológico colombiano.Bogotá,AcademiaColombianadeCienciasExactas,FísicasyNaturales.

11 7
L A B I O L O G Í A D E L A C O N S E R V A C I Ó N | M A R Í A F E R N A N D A M A R T Í N E Z - P O L A N C O
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.99-118
Primack, Richard2000. A Primer of Conservation Biology.Sunderland,SinauerAssociates,IncPublishers.
Redford, Kent1992.“TheEmptyForest”,BioScience42,pp.412-422.
Reitz, Elizabeth y Elizabeth Wing2008.Zooarchaeology. Cambridge Manuals in Archaeology,segundaedición.Cambridge,Cambridge
UniversityPress.
Reynolds, John y Carlos Peres2006.“Oveexploitation”,enPrinciples of Conservation Biology,MarthaGroomGaryMeffeyRonald
Carrol,eds.,pp.254-291Sunderland,M.A..Sinauer.
Rivera, Sergio1992. Neusa: 9.000 años de presencia humana en el páramo.Bogotá,FIANBancodelaRepública.
Robinson, John y Kent Redford1991.“SustainableHarvestofNeotropicalForestMammals”,enNeotropical Wildlife Use and
Conservation,JohnRobinsonyKentRedford,eds.,pp.415-429.Chicago,ChicagoPress.
Robinson, John y Elizabeth Bennett 1999.“HuntingforSustainability:TheStartofaSynthesis”,enHunting for Sustainability in Tropical For-
est,JohnRobinsonyElizabethBennett,eds.,pp.499-519.NuevaYork,ColumbiaUniversityPress.
Rodríguez, Manuel y Chistopher Vaughan1994.“TamañoycomposicióndelosgrupossocialesdelvenadocolablancaenlaislaSanLucas”,en
ChistopherVaughanyManuelRodríguezeds.,Ecología y manejo del venado cola blanca en México y Costa Rica,pp.131-162.Heredia,EUNA.
Rodríguez Freyle, Juan[1636]1985.El Carnero.Medellín,Bedout.
Sapir-Hen, Lidar, Guy Bar-Oz, Hamoudy Khalaily y Tamar Dayan2009.“GazelleExploitationintheEarlyNeolithicSiteofMotza,Israel:TheLastoftheGazelleHuntersin
theSouthernLevant”,Journal of Archaeological Science36,pp.1538-1546.
Simón, fray Pedro[1625]1981.Noticias historiales de las conquistas de tierra firme en las indias occidentales.Bogotá,
BibliotecaBancoPopular.
Sinclair, Anthony, John Fryxell y Graeme Caughley2006.Wildlife Ecology, Conservation and Management,segundaedición.Victoria,Australia,Blackwell
Publishing.
Smith, Winston1991.“Odocoileus virginianus”, Mammalian Species388(6).
Teer, James1994.“Elvenadocolablanca:historianaturalyprincipiosdemanejo”,enEcología y manejo del venado
cola blanca en México y Costa Rica,ChistopherVaughanyManuelRodríguez,eds.,pp.32-48.Heredia,EUNA.
Townsend, Wendy1999.“TheSustainabilityofSubsistenceHuntingbytheSirionoIndiansofBolivia”,enHunting for
Sustainability in Tropical Forest,JohnRobinsonyElizabethBennett,eds.,pp.267-281.NuevaYork,ColumbiaUniversityPress.
Ugan, Andrew2005.“Climate,BoneDensity,andResourceDepression:WhatIsDrivingVariationinLargeandSmall
GameinFremontArchaeofaunas”,Journal of Anthropological Archaeology24,pp.227-251.

11 8
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.99-118
Walker, Renne2000.“White-tailedDeer(Odocoileus virginianus)MortalityProfiles:ExamplesfromtheSoutheastern
UnitedStatus”,ArchaeozoologiaXI,pp.175-186.
Whitaker, Adrian2008.“IncipientAquacultureInPrehistoricCalifornia?Long-termProductivityandSustainabilityVs.
ImmediateReturnsfortheHarvestofMarineInvertebrates”,Journal of Archaeological Science 35 (4),pp.1114-1123.

11 9
L A A RQU EOFAU NA DE XCA M BÓ, Y UCATÁ N, M ÉX ICO
chrisTopher m. GöTz*[email protected] de Ciencias Antropológicas, Universidad Autónoma de Yucatán, México
theLmA Noemí sierr A sosA**[email protected] Regional Yucatán del Instituto Nacional de Antropología e Historia (INAH), México
R e s u m e n Este artículo discute el patrón de aprovechamiento
seguido por parte de los mayas prehispánicos del litoral yucateco
(México), basándose en una muestra zooarqueológica del sitio de
Xcambó. La discusión acerca del aprovechamiento gira en torno
al amplio y diverso espectro faunístico del sitio, haciendo énfasis
en el hallazgo de restos de foca tropical (Monachus tropicalis),
una especie extinta hoy día. Especímenes de foca tropical son
extremadamente raros en Mesoamérica, pero demuestran
que estos animales fueron ocasionalmente utilizados por los
pobladores precolombinos.
P A L A B R A S C L A V E :
Arqueología maya, zooarqueología, subsistencia, pesca
prehispánica, Monachus tropicalis.
Artículorecibido:1dejuniode2011|aceptado:1denoviembrede2011|modificado:16denoviembrede2011
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,294pp.ISSN1900-5407,pp.119-145
* Doctor,UniversidaddeHamburgo,Alemania.MaestroenCienciasyArtes(MagisterArtium)delaUniversidadRhinenseFederico-GuillermodeBonn,Alemania.
**Doctoraenestudiosmesoamericanos,UniversidadNacionalAutónomadeMéxico,México.

1 2 0
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.119-145
arCheofauna of xCamBo, yuCatan, mexiCo
ABSTR AC T This article discusses the faunal
use pattern of the pre-Hispanic Mayas of
the Yucatan coast (Mexico), based on a
zooarchaeological sample from the site of
Xcambo. The discussion addresses the use
of a wide and diverse spectrum of fauna,
emphasizing the discovery of remains of the
tropical seal (Monachus tropicalis) a species
currently extinct. Tropical seal specimens
are extremely rare in Mesoamerica, but they
show that these animals were occasionally
used by pre-Columbian inhabitants.
a arqueofauna de xCamBó, iuCatã, méxiCo
RESUMO Este artigo discute o padrão de
aproveitamento seguido por parte dos Maias
pré-hispânicos do litoral iucateco (México),
baseando-se numa amostra zooarqueológica
do lugar de Xcambó. A discussão sobre o
aproveitamento gira em torno ao amplo
e diverso espectro faunístico do lugar,
fazendo ênfase no descobrimento de restos
de foca tropical (Monachus tropicalis), uma
espécie extinta hoje em dia. Espécimes de
foca tropical são extremadamente raros em
Mesoamérica, mas demonstram que estes
animais foram ocasionalmente usados pelos
povoadores pré-colombianos.
P A L A V R A S C H A V E :
Arqueologia maya, zooarqueologia,
subsistência, pesca pré-hispânica, Monachus
tropicalis.
K E Y W O R D S :
Mayan Archaeology, Zooarchaeology,
Subsistence, Pre-hispanic Fishing, Monachus
Tropicalis.

1 2 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.119-145
L A A RQU EOFAU NA DE XCA M BÓ, Y UCATÁ N, M ÉX ICO
c h r i s T o p h e r m . G ö T z
t h e L m A N o e m í s i e r r A s o s A
IntroducciónLos recientes estudios zooarqueológicos en las tierras bajas maya del norte han arrojado datos acerca de la dieta alimenticia, el uso ritual de animales, patrones de caza, pesca y domesticación por parte del pueblo maya prehispánico (véase, por ejemplo, Götz, 2008a y b; Márquez Morf ín, 1991; Masson y Peraza, 2008). En este artículo queremos presentar al conjunto zooar-queológico del sitio arqueológico de Xcambó, ubicado en la costa noroeste de la península yucateca (ver la figura 1), que fue excavado y analizado recientemente. Se discutirá el patrón de aprovechamiento de animales vertebrados e invertebra-dos1 que se seguía por parte de la población xcamboense durante los períodos de su apogeo (períodos Clásico Temprano [250 a 550 d. C.] y Clásico Tardío [550 a 700 d. C.]). Se efectúa una reconstrucción de las áreas de obtención de los recursos faunísticos explotados por los habitantes costeros prehispánicos, estudiando si el material faunístico del asentamiento que arqueológicamente ha sido reconocido como puerto comercial permite identificar también el traslado de animales o partes de éstos, desde o hacia regiones distantes.
Una parte del artículo se adscribe a estudiar la interacción entre los antiguos habitantes del lugar y la foca tropical (Monachus tropicalis), cuyos restos se han encontrado en Xcambó y un sitio vecinos. Los fragmentos esqueléticos de foca tropical son de particular interés para el conocimiento zooarqueológico de Meso-américa, porque hace más de veinte años se había postulado el aprovechamiento
1 Lamuestra arqueofaunística de Xcambó consistió primordialmente en animales vertebrados. Los animalesinvertebradosserefierenúnicamenteacrustáceos,yaquelosmoluscosformanpartedeotroestudio.

1 2 2
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.119-145
de focas por parte de los mayas prehispánicos (Andrews, 1984), pero entonces no hubo pruebas f ísicas al respecto. El hallazgo arqueológico reciente de restos de esta especie permite confirmar que sí hubo aprovechamiento, a pequeña escala, de esta especie por parte de los pobladores costeros prehispánicos de la región.
1. Descripción del área de estudio La costa norte del estado mexicano de Yucatán, donde se encuentra el sitio de Xcambó, está conformada por sedimentos generados en el ambiente marino (Bautista-Zúñiga et al., 2003: 25-26). Entre dunas costeras y la llanura cárstica se encuentra una amplia depresión que acumula aguas saladas y semisaladas entre petenes2, donde las aguas dulces de los mantos freáticos únicamente aflo-ran en pocas fuentes (Bautista-Zúñiga et al., 2003: 27; Contreras Espinosa, 1993: 179) y la agricultura no puede practicarse, debido a las inundaciones marinas del litoral (Bautista-Zúñiga et al., 2003: 32). Los fondos marinos someros del norte peninsular consisten en arena y corales, bajo aguas de tipo tropical con una salinidad entre 32‰ y 37,5‰, en los que se observa una circulación vertical
2 Eltérminomayapetén sedefine,segúnBastarracheayCanto(2004),como“islaoacamposllanosjuntosalmaramaneradeisletas,dondehaylagartos,tortugasyalgunospeces”.
Figura1.Mapadeláreadeestudio,queindicalossitiosyregionesmencionadoseneltexto.EnelrecuadrosemuestraelmapadelsitioprehispánicodeXcambó,Yucatán,México.MapageneraldibujadoporC.Götz,mapadeXcambótomadodeSierraSosa,1997,modificadoporC.Götz.

1 2 3
L A A R Q U E O F A U N A D E X C A M B Ó , Y U C A T Á N | C H R I S T O P H E R M . G Ö T Z Y T H E L M A N O E M Í S I E R R A
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.119-145
intensa y un enriquecimiento de nutrientes que aumentan la productividad del ecosistema (Chávez, 1994: 3).
El Proyecto Arqueológico Xcambó del Centro INAH Yucatán, dirigido por Thelma Sierra Sosa, se llevó a cabo entre 1996 y 2000. El sitio, una comuni-dad que administraba y controlaba un puerto comercial y las salinas cercanas, está ubicado en la carretera costera Progreso-Telchac, al costado sur de la cié-naga. Xcambó fue construido modificando arquitectónicamente un pequeño petén y exhibe una infraestructura que concilia intereses económicos, políti-cos y religiosos con diseños y arreglos de construcciones y terreno. Los lími-tes naturales del petén fueron modificados mediante la nivelación externa y la construcción de elevados basamentos, impidiendo daños por inundaciones. Alrededor de las plazas se construyeron residencias muy cerca unas de otras. Al oeste de la Plaza Principal se levantan las residencias mejor elaboradas, con áreas de actividad doméstica de los señores principales y de la élite.
En el sector este se construyeron amplios basamentos bajos que soporta-ban viviendas más modestas. Los amplios espacios sin techar en este sector sir-vieron probablemente para el trabajo al aire libre. De la pequeña plaza pública parten dos andadores o sak beo’ob3; uno se dirige a una unidad residencial loca-lizada en el extremo este del sitio y el otro se interna tierra adentro, rumbo al moderno poblado de Dzemul, correspondiente a un antiguo sitio prehispánico. Xcambó mantuvo una especie de atracadero que se localiza al norte de la plaza principal, en la orilla de la ciénaga.
En un primer período de ocupación (Clásico Temprano, 250 a 550 d. C.), el sitio fue planeado arquitectónicamente como un centro integrado por un grupo de edificios modestos de tipo administrativo, religioso, cívico y residen-cial, alrededor de los cuales se instalaron amplios basamentos con viviendas y numerosos depósitos de almacenamiento. El puerto comercial tuvo una posi-ción económica y política importante, y suponemos que Xcambó controlaba económicamente las salinas costeras, así como los sitios aledaños costeros que producían sal y obtenían mariscos y pescados (Jiménez Álvarez, 2002; Sierra Sosa, 1997: 351; Sierra Sosa, 1999: 42ss.; Sierra Sosa y Martínez, 2001: 7; Sie-rra Sosa, 2004). El sitio tierra adentro de Izamal controlaba probablemente los recursos costeros, y Xcambó pudo haber sido un sitio de comercio intermedia-rio, a través del cual se aseguraban las transacciones.
La transformación de Xcambó durante el Clásico Tardío (550 a 700 d. C.) fue importante, y se modificaron la fisonomía del sitio y la ocupación de los
3 Unsak be(pl.sak beo’ob)consisteenunacalzadainternaoexternadelossitiosprecolombinosmayas,edifi-cadademamposteríaycubiertadeestuco(véaseGarzayKurjack1980:52-65).

1 2 4
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.119-145
habitantes. Se cubrieron las construcciones y áreas de almacenamiento ante-riores, para sustituirlas por residencias tanto de la élite como de la población de menor posición social, además de las construcciones de otra índole, como talleres, almacenes, áreas de actividad doméstica, etc. (véase Sierra Sosa y Mar-tínez, 2001). Para el Clásico Tardío se incrementó la explotación de las salinas, pero parece que la población del sitio sobre todo administraba dicho trabajo (Wanner et al., 2007). Xcambó tuvo relaciones con los sitios de las costas aleda-ñas de Campeche, y del occidente de Yucatán, con el centro y noroccidente de Yucatán, con el noroeste peninsular, con el Petén campechano-guatemalteco, con algunos sitios de la cuenca del río Grijalva-Mezcalapa de Tabasco y la sierra Zoque de Chiapas, con algunos lugares de la cuenca media y baja del río Usuma-cinta, de Tabasco y Chiapas, con la zona de la Laguna de Términos y la región de Atasta, y con algunos sitios de la región central veracruzana (Sierra Sosa, 2004).
2. Materiales y métodos La muestra de material arqueofaunístico de Xcambó considerada en este texto suma 1.539 especímenes y procede de la porción oeste del sitio (plaza oeste y estructuras NE2, NE14, NE19, NO1, NO2, NO4 y NO9), un área dominado por estructuras habitacionales de rango elevado. Los especímenes faunísticos están mayormente bien preservados, aunque restos microfaunísticos se encuentran poco. A pesar de haber una fuerte fragmentación del material esquelético, la mayoría de las piezas aún posee características diagnósticas para una identifi-cación taxonómica y osteológica.
Los materiales de Xcambó se obtuvieron durante la excavación de pozos estratigráficos y la liberación de los edificios, donde formaron parte de relle-nos constructivos o se hallaban encima de antiguas superficies de uso. Muchos desechos alimenticios y de utensilios, entre otros, fueron usados en rellenos constructivos, de tal manera que Xcambó manifiesta, como también lo men-cionan Cobos et al. (2007: 7) respecto a otro sitio costero maya, una virtual ausencia de basureros formales4, dificultando y muchas veces imposibilitando un fechamiento de los ecofactos con base en la estratigraf ía. Los pozos fueron excavados en el interior de las estructuras, en patios y plazas, con dimensiones de 2 x 2 m. La excavación siguió capas constructivas y se utilizaron cernidores de aperturas de 5 mm, para recuperar los materiales pequeños. En todos los casos, la excavación y recolección estuvieron en manos de trabajadores bajo la dirección y vigilancia de arqueólogos.
4 Esterasgopresentaunagrandiferenciarespectoaasentamientosmayastierraadentro,dondedichosrasgossonfrecuentes(véaseChase,ChaseyTeeter2004).

1 2 5
L A A R Q U E O F A U N A D E X C A M B Ó , Y U C A T Á N | C H R I S T O P H E R M . G Ö T Z Y T H E L M A N O E M Í S I E R R A
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.119-145
El análisis zooarqueológico se llevó a cabo en el Taller de Zooarqueolo-gía de la Facultad de Ciencias Antropológicas de la Universidad Autónoma de Yucatán. Durante el estudio zooarqueológico se realizó la identificación taxo-nómica y osteológica del material arqueofaunístico5, y se cuantificó mediante el conteo simple (NISP) y porcentajes de éste (véanse O’Connor, 2003; Reitz y Wing, 2008). Para las identificaciones de los especímenes, se utilizaron una colección de referencia osteológica, así como varios manuales de osteología de vertebrados (EA.FLMNH, 2003; Gilbert, 2003; Gilbert, Martin y Savage, 1996; Olsen, 1964, 1968 y 1982). La terminología taxonómica empleada en este trabajo se basa en estudios zoológicos recientes (Adam y García, 2003; Adam, 2004; Howell y Webb, 2004; Lee, 2000; Mojetta, 2005; Robins, Ray y Douglass, 1986; Reid, 1997), pero fue verificada en el Integrated Taxonomic Information System (ITIS, véase referencias). Los huesos de foca fueron identificados por medio de fotograf ías de especímenes modernos, facilitadas a los autores. Para las mediciones osteomorfométricas de los huesos de foca, se empleó el manual de Ericson y Storå (1999).
3. Resultados
3.1. Panorama general del aprovechamiento de animales por los mayas prehispánicos de Xcambó, Yucatán En la muestra de 1.539 fragmentos esqueléticos faunísticos se encontraron especímenes de crustáceos (7,7% del NISP total), peces (actinopterígios y con-dríctios juntos) (38,8% del NISP total), reptiles (34% del NISP total), aves (3,7% del NISP total) y mamíferos (15,8% del NISP total) (ver la tabla 1).
De los crustáceos se encontraron 87,3% del NISP pertenecientes a can-grejos (brachyura). Un alto porcentaje de los brachyura (expresado en relación con el NISP total de cada respectiva muestra) corresponde al cangrejo de pie-dra (Menippe mercenaria), mientras que las demás especies de cangrejos, como la jaiba (Calinectes sapidus), el juey común (Cardisoma guanhuni), el cangrejo señalador (Uca rapax), el cangrejo araña (Libinia emarginata) y la cucaracha del mar (Limulus polyphemus) son poco representados (ver la tabla 2).
5 Unestudiotafonómicodeperfilesdepreservaciónydegeneraciónderestosfaunísticosenlacostaestádesa-rrollándoseenelmomentoderedaccióndeestemanuscrito(sept.2011).

1 2 6
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.119-145
Tabla1.TablaqueindicalosporcentajesdelNISP(%NISP)delosgrupostaxonómicosmásrelevantesdelasmuestras.
ORDEN XCAMBÓ%NISP(N=1539) XCAMBÓNISP
CRUSTÁCEOS 7,7 118
PECES 38,8 597
REPTILES 34 524
AVES 3,7 57
MAMÍFEROS 15,8 243
CRUSTÁCEOS XCAMBÓ%NISP(N=118) XCAMBÓNISP(N=118)
Cangrejos 87,3 103
PECES XCAMBÓ%NISP(N=597) XCAMBÓNISP(N=597)
Tiburonesypezsierra 39,7 237
Rayas 2,3 14
Actinopterígios 58 346
REPTILES XCAMBÓ%NISP(N=524) XCAMBÓNISP(N=524)
Crocodilos 1,3 7
Tortugasdemar 65,5 343
Tortugasdetierra/aguadulce 14,9 78
Iguanas 18,3 96
AVES XCAMBÓ%NISP(N=57) XCAMBÓNISP(N=57)
Avesdemar 22,8 13
Buitres 0 0
Pavosyespeciesafines 36,8 21
Avespequeñas 3,5 2
MAMÍFEROS XCAMBÓ%NISP(N=243) XCAMBÓNISP(N=243)
Zarigüeyas 0,4 1
Armadillos 0,4 1
Roedoresyconejos 3,3 8
Carnívorosmenores 12,3 30
Felinos 0,4 1
Manatíes 1,2 3
Perisodáctilos 9,9 24
Artiodáctilos 50,2 122
Mamíferosmarinos 4,5 11

1 2 7
L A A R Q U E O F A U N A D E X C A M B Ó , Y U C A T Á N | C H R I S T O P H E R M . G Ö T Z Y T H E L M A N O E M Í S I E R R A
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.119-145
Tabla2. Tablaqueindicalosperfilestaxonómicosdetalladosdelosdosconjuntoscotejados,mostrandoelNISP yelporcentajedelNISP(%NISP).
TaxónXCAMBÓ(N=1539)
NISP %NISP
Noidentificado
CRUSTÁCEOS
Brachyuranoidentificado 15 1
Cangrejos
Callinectessapidus 1 0,1
Menippemercenaria 100 6,5
Cardisomaguanhuni
Ucarapax 1 0,1
Libiniaemarginata
Limuluspolyphemus 1 0,1
PECES
Condrictionoidentificado
Tiburones ypezsierra
Ginglymostomacirratum 12 0,7
Carcharhinusspp. 188 12,2
Galeocerdocuvieri 12 0,8
Mustelusnorrisi
Rhizoprionodonterraenovae 22 1,4
Sphyrnaspp. 3 0,2
Pristispectinata
Rayas
Rajiformenoidentificado 8 0,5
Dasyatiscf.americana 1 0,1
Aeobatusnarinari 5 0,3
Actinopterígionoidentificado 41 2,7
Actinopterigios
Megalopsatlanticus
Cynoscioncf.nebulosus
Ariopsisfelis 68 4,4
Opsanusspp. 4 0,3
Centropomusspp. 174 11,3
Epinephelusspp. 15 1
Caranxspp.
Lutjanusspp. 7 0,5
Heamulonplumieri
Archosargusspp. 1 0,1
Calamusleucosteus 4 0,3

1 2 8
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.119-145
TaxónXCAMBÓ(N=1539)
NISP %NISP
Peces Actinopterígios
Micropogoniasundulatus 32 2,1
Pomacanthidae
Sphoeroidesspp.
REPTILES
Crocodilos Crocodyluscf.moreletii 7 0,5
Tortugasdemar
Cheloniidaenoidentificado 247 16
Carettacaretta 49 3,2
Cheloniamydas 31 2
Eretmochelysimbricata 16 1
EretmochelysoChelonia
Tortugasdetierra/aguadulce
Dermatemysmawii 1 0,1
Kinosternonspp. 62 4
Emydidaenoidentificado
Terrapenespp. 7 0,5
TerrapeneoKinosternon 2 0,1
Trachemysspp. 6 0,4
Iguanas Ctenosauracf.similis 96 6,2
AVES
Avenoidentificada 21 1,4
Avesdemar
Sulaspp. 1 0,1
Pelecanusspp. 6 0,4
Phalacrocoraxauritus
Fregatamagnificens
Ardeaherodias 1 0,1
Egrettaspp. 5 0,3
Egrettaspp.óArdeaherodias
Phoenicopterusruber
Buitres Coragypsatratus
Pavos yespeciesafines
Cracidae 1 0,1
Galliformes 2 0,1
Meleagrisspp. 13 0,8
Meleagrisocellata 4 0,2
Puffinusspp.oGallinulachloropus
1 0,1
Avespequeñas
Bartramialongicauda 1 0,1
Columbidae
Aratingaspp.oAmazonaspp. 1 0,1
Cyanocoraxyucatanicus
(Cont.)

1 2 9
L A A R Q U E O F A U N A D E X C A M B Ó , Y U C A T Á N | C H R I S T O P H E R M . G Ö T Z Y T H E L M A N O E M Í S I E R R A
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.119-145
Los peces, subdivididos en tiburones y peces sierra, rayas y actinopterí-gios, muestran una distribución heterogénea: el 39,7% del NISP total de peces de Xcambó corresponde a tiburones y peces sierra, mientras que las rayas no superan el 2,3%. Los actinopterígios fueron representados por un 58% (tabla 1). Entre la fauna íctia se realzan porcentualmente diversos géneros de tibu-rones (Ginglymostoma, Carcharhinus, Galeocerdo, Rhizoprionodon, Sphyrna), al igual que varios especies y géneros de actinopterígios, como bagre (Ariop-sis felis), mero (Epinephelus sp.) y gurrubata (Micropogonias sp.). El pez más numeroso y presumiblemente importante en Xcambó fue el robalo (Centropo-mus sp.), con un 11,3% del NISP de la muestra total del sitio (tabla 2).
TaxónXCAMBÓ(N=1539)
NISP %NISP
MAMÍFEROS
Mamíferonoidentificado 42 2,7
ZarigüeyasDidelphisspp.
Didelphidaecf.PhilanderopossumoMarmosaspp.
1 0,1
Armadillos Dasypusnovemcinctus 1 0,1
Roedoresyconejos
Dasyproctapunctata
DasyproctidaeoAgoutidae
Agoutipaca 4 0,3
Sylvilagusspp. 4 0,3
Carnívorosmenores
Canislupusfamiliaris 18 1,2
Urocyoncinereoargenteus 9 0,6
Procyonlotor 3 0,2
FelinosFelidaecf.Pumaconcolor 1 0,1
Felidaecf.Pantheraonca
Manatíes Trichechusmanatus 3 0,2
Perisodáctilos
Equuscaballus 7 0,5
EquidaeoBovidae 4 0,3
Tapirusbairdi 13 0,8
Artiodáctilos
Pecaritajacu 9 0,6
Bostaurus 2 0,1
Cervidae
Odocioleusvirginianus 106 6,9
Mazamaspp. 5 0,3
Mamíferosmarinos
Delphinidae 2 0,1
Monachustropicalis 9 0,6
(Cont.)

1 3 0
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.119-145
Entre los reptiles se distinguen cocodrilos, tortugas de mar, tortugas de tierra/agua dulce e iguanas. Los cocodrilos (Crocodylus moreletii) son poco frecuentes, con 1,3% de la muestra total de reptiles, mientras que las tortugas de mar alcanzan 65,5% del NISP total de reptiles, las iguanas 18,3% y las tor-tugas de tierra y agua dulce 14,9% (tabla 1). En el sitio se encontraron espe-címenes de la tortuga caguama (Caretta caretta), tortuga verde (Chelonia mydas) y tortuga carey (Eretmochelys imbricata), mientras que las tortugas de agua dulce comprenden a la tortuga de río (Dermatemys mawii), tortuga casquito (Kinosternon sp.), tortuga ‘japonesa’ (Trachemys sp.) y de caja (Terra-pene sp.). En cambio, la única iguana identificada fue la Ctenosaura similis (ver la tabla 2).
La colección arqueofaunística de aves se compone de aves de mar (22,8% del NISP total de aves) y pavos y especies afines (36,8%) (tabla 1). Entre los taxones marinos de aves hubo restos de pelícano (Pelecanus sp.), gallareta (Sula sp.), fregata (Fregata magnificens), garza (Egretta sp.), mientras que las aves de tierra comprenden pavos ocelados (Meleagris ocellata), gallinola (Gallinula chloropus) y perico (Arathinga o Amazona sp.) (ver la tabla 2).
Con respecto a los mamíferos, se observan zarigüeyas, con 0,4% del NISP total de mamíferos; armadillos, con 0,4%; roedores y conejos, con 3,3%; felinos, con 0,4%, y mamíferos marinos (exceptuando el manatí), con 4,5%; los artiodáctilos son el grupo más numeroso en la muestra mamífera, y alcan-zaron 50,2% (ver la tabla 1). El mamífero más frecuente en el conjunto es el venado cola blanca (Odocoileus virginianus), pero se encontraron también restos de conejo (Sylvilagus sp.), agutí (Agouti paca), pecarí (Pecari tajacu) y venado temazate (Mazama sp.)6. Semejante a otros sitios prehispánicos costeros, se hallaron numerosos restos de perro (Canis lupus familiaris) en Xcambó, que sumaron 12,3%. Hay pocas evidencias de felinos grandes (Puma concolor), y los restos de la foca tropical alcanzan un 0,6 % del NISP en Xcambó (ver la tabla 2).
3.2 Los hallazgos de foca De la foca tropical, foca monje o West Indian Seal (Hairr, 2010), se hallaron diez especímenes esqueléticos de las extremidades y del cráneo entre la muestra arqueofaunística de Xcambó. Los diez especímenes esqueléticos se encontra-ron incrustados en rellenos constructivos de nivelaciones y plataformas de la parte oeste del sitio y corresponden a un fémur izquierdo (ver la figura 2a), un
6 Los restosdebóvidosyéquidos,aunqueconsideradosen la tabla,correspondena intrusionesmodernasyfueronhalladossinexcepciónensuperficie.

1 3 1
L A A R Q U E O F A U N A D E X C A M B Ó , Y U C A T Á N | C H R I S T O P H E R M . G Ö T Z Y T H E L M A N O E M Í S I E R R A
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.119-145
radio proximal derecho (ver la figura 2b), un fragmento anterior de un maxilar derecho con el tercer premolar in situ (ver la figura 2c), así como un canino superior derecho (correspondiente al maxilar anteriormente descrito) (ver la figura 2d). Además de esto, se encontraron en Xcambó un canino inferior y tres primeros premolares de foca, modificados como artefactos (véase Canto Mén-dez, 2009), ya que recibieron una perforación a nivel de raíz, probablemente para ser usados como colgantes. Casi todos los especímenes óseos de foca encontrados en Xcambó pertenecieron a individuos adultos; el único hueso de un animal juvenil corresponde a un radio proximal (ver la figura 2e), hallado en conjunto con los demás huesos.
El fémur izquierdo, encontrado en el relleno constructivo de la estruc-tura NE2, está muy bien preservado. Posee un largo total, con base en Eric-son y Storå (1999), del cóndilo medial hasta el extremo distal del trocánter mayor, de 108,5 mm. Tomando los dos cóndilos en un plano, se alcanza un largo total de 98 mm, correspondiente a la medida 2 del fémur, según Eric-son y Storå (1999). El ancho máximo del extremo proximal, medido desde la porción medial de la cabeza femoral hasta el lado lateral del trocánter
Figura2.ImágenesdelosespecímenesóseoscaracterísticosdefocatropicalhalladosenXcambó,Yucatán.FotografíasydibujodeC.Götz.

1 3 2
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.119-145
mayor, es de 65 mm (medida 3 del fémur del manual arriba mencionado). La distancia entre la depresión entre la cabeza femoral y el trocánter mayor y la depresión intercondilar distal alcanza 93 mm (medida 9 del fémur, según el manual arriba mencionado).
Del radio, excavado en la plaza oeste del sitio, falta una parte de la diá-fisis distal, por lo que no se pudo establecer con exactitud la edad biológica del mismo. Las inserciones musculares y la epífisis proximal están claramente definidas, y es muy probable que el animal haya sido de edad adulta. La longitud total del espécimen hasta la fractura es de 101,5 mm, y el ancho más próximo a la fractura, de 43,4 mm.
Muy cerca de la fractura y de la inserción principal muscular de la línea áspera del radio se encontraron cinco ligeras raspaduras encima del hueso córtico, perpendiculares al axis longitudinal del hueso y paralelas entre sí. La revisión de las raspaduras bajo estereoscopio a 40 x reveló que presentan poca profundidad en un corte transversal en forma de U. Junto con el extremo den-ticulado de la fractura del radio, puede suponerse que fueron efectuadas por dientes de algún carnívoro, muy probablemente un perro pequeño, que mas-ticó el hueso en la parte proximal, destruyendo una parte y raspando la otra entre el cuarto premolar y primer molar.
El fragmento de maxilar, del lado derecho, posee aún in situ el tercer premolar, además del alvéolo del tercer y segundo premolar y de la mitad del alvéolo del canino, junto con el diente correspondiente, y fue hallado en el relleno constructivo de la plaza oeste de Xcambó. La edad biológica del maxi-lar es adulta, ya que se aprecia no solamente la erupción total de las piezas dentales, sino también un aumentado desgaste del premolar y una retracción alveolar7. La distancia entre el borde posterior del alvéolo del cuarto premolar y del borde anterior del canino mide 45,2 mm.
El canino superior, perteneciente al maxilar y encontrado en asocia-ción estratigráfica con éste, muestra daños en la porción lingual de la corona, habiéndose quebrado, al parecer en un momento post mórtem por falta de redondeo de los bordes de la fractura, un tercio de ésta. La raíz, sin embargo, está completa y el espécimen mide en total 46 mm. El primer premolar, que se encontró suelto en la estructura NE10, presenta únicamente el típico desgaste de la porción distal de la corona y mide 31 mm.
7 Apesardequeeldesgastedentalylaretracciónalveolarsonsobretodoenesqueletoshumanossignospatoló-gicosodeunaedadavanzada,parecequeenlasfocasdelaespecieMonachus tropicalis constituyenunrasgocomún,yaqueinvestigadoresquevieronindividuosdeestaespecieaúnvivosenlosprimerosañosdelsigloXXafirmanquetodoslosindividuos,exceptuandolosmuyjóvenesyjóvenes,padecierondeunaumentadodesgastedental,posiblementerelacionadoconunadietabasadaenpecesycrustáceos(véaseNelson1900,citadoenAdamyGarcía,2003:302,305).

1 3 3
L A A R Q U E O F A U N A D E X C A M B Ó , Y U C A T Á N | C H R I S T O P H E R M . G Ö T Z Y T H E L M A N O E M Í S I E R R A
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.119-145
Los especímenes de foca evidentemente procesados por seres humanos corresponden a cuatro dientes sueltos, asociados a las estructuras NO2, NE3 y NE7A (Canto Méndez, 2009). Estos dientes consisten en un canino inferior y tres primeros premolares, y concuerdan en tamaño con los dientes descri-tos anteriormente.
4. Discusión
4.1 Variabilidad y continuidad cronológica del aprovechamiento de especies acuáticas en la costa yucatecaEl perfil faunístico de Xcambó (ver la figura 3) permite esbozar el panorama de aprovechamiento que se seguía por parte de los habitantes del sitio entre 250 y 700 d. C.8. No fue posible separar cronológicamente con mayor detalle los espe-címenes faunísticos, porque la estratigraf ía de Xcambó, como ya se mencionó, se vio alterada por las frecuentes modificaciones y reutilizaciones de material de relleno, como lo demostró también el estudio de la cerámica por Jiménez Álvarez (2002). Wanner et al. (2007) y Cucina et al. (2011) demostraron que las actividades de la población xcamboense cambiaron entre el Clásico Temprano y el Clásico Tardío, pero no fue posible captar estos cambios en el material arqueofaunístico, hasta este momento.
8 LosdatosdeIslaCerritosprocedendeHerreraFlores(2011).
Figura3.GráficoqueindicalariquezadeespeciesdeXcambó(laalturadelascolumnasindicalacantidaddeespeciesidentificadasporgrupotaxonómico.
Cang
rejo
s
Tibu
rone
syp
ezsi
erra
Raya
s
Actin
opte
rigio
s
Tortu
gasd
em
ar
Tortu
gasd
etie
rra
/agu
adu
lce
Aves
de
mar
Pavo
sye
spec
iesa
fines
Aves
peq
ueña
s
Zaru
güey
as
Roed
ores
ycon
ejos
Carn
ívor
osm
enor
es
Felin
os
Peris
odác
tilos
Artio
dáct
ilos
Mam
ífero
smar
inos
Xcambó

1 3 4
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.119-145
Con base en la reconstrucción de las áreas de obtención podemos cons-tatar, tal como se había postulado en publicaciones anteriores (Götz, 2008b), que los antiguos moradores de la península obtenían animales silvestres mayo-ritariamente en las zonas cercanas y circundantes a los sitios prehispánicos. El sitio de Xcambó no es la excepción de este patrón, y la fauna consumida venía ya sea de las aguas poco profundas del litoral yucateco incluido el manglar, de las playas del litoral, o bien, de los bosques cercanos a la zona intermedia entre el litoral y la región tierra adentro (Götz, 2008b: 165).
Los perfiles taxonómicos de la fauna íctica indican que en tiempos pre-hispánicos se aprovechaban principalmente de recursos bentónicos, de aguas menos profundas de lo que se pesca hoy día. No hay evidencia de que se navegaba, con fines de la pesca, en alta mar, ya que ninguno de los taxones encontrados ocurre exclusivamente en aguas pelágicas, aunque algunas especies encontradas en las muestras ocasionalmente habitan o viajan a través de aguas profundas (véase Froese y Pauly, 2009). Xcambó manifiesta una predominancia de especies que habitan en fondos lodosos cercanos a la costa y aguas costeras y estuarios, como bagre marino (Ariopsis felis) y robalo (Centropomus spp.). Actualmente, estas dos especies ya no son de importancia para la pesca, yaciendo el énfasis de la pesca yucateca moderna en meros (Epinephelus spp.) y camarones (Penaeus spp.), especies más alejadas de la zona costera inmediata (Chávez, 1994).
La información ecológica de los crustáceos y de la herpetofauna marina apunta igualmente hacia un aprovechamiento de las zonas lacustres en tiem-pos prehispánicos, e indica que todas las especies de cangrejos tuvieron que obtenerse en la playa, los manglares o las aguas poco profundas costeras (véase Kaplan, 1988). En cuanto a las tortugas marinas, cuyos restos fueron recupera-dos en gran cantidad en la muestra, se observa que la mayoría de los individuos era de edad adulta (Cárdenas, 2011). Es posible que estos individuos hayan sido hembras adultas, capturadas, probablemente como aún sucedía hace algunas décadas en esta zona, en el momento de desovar en las playas, entre primavera y verano (Lee, 2000: 135-142; Marmolejo Monsiváis, 2000: 89-90). Otra alter-nativa de la pesca de tortugas de mar pudiera haber sido cazarlas durante la cópula o mientras descansaban en el agua (Carr, 1987: 8).
Entre las aves marinas, llaman la atención los restos de la garza ceniza (Ardea herodias) y pelícano (Pelecanus spp.), ya que entre éstas se han obser-vado movimientos migratorios para la temporada de invierno, que vienen del norte, lo que podría indicar que estos individuos fueron capturados en los meses de invierno (véase Howell y Webb, 2004).
En relación con el comercio, actividad aparentemente de gran impor-tancia para el asentamiento, pudo evidenciarse que en la muestra no se

1 3 5
L A A R Q U E O F A U N A D E X C A M B Ó , Y U C A T Á N | C H R I S T O P H E R M . G Ö T Z Y T H E L M A N O E M Í S I E R R A
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.119-145
encuentra realmente ningún taxón claramente identificable como procedente de regiones muy alejadas del sitio, tales como el sur y centro de Veracruz, Guerrero, la región del Chontalpa y Grijalva de Tabasco y Chiapas, así como las tierras altas de Guatemala, áreas con las que Xcambó mantuvo contacto comercial. Sin embargo, existe la posibilidad de que Xcambó tuviera una rela-ción de intercambio con asentamientos tierra adentro cercanos. Los indicios de un intercambio de recursos faunísticos apuntan hacia un transporte de animales a menor distancia. Un gran porcentaje de los restos arqueofaunís-ticos de Xcambó procede de las inmediatas zonas costeras, refiriéndose en principio a animales marinos. Sin embargo, entre la fauna hubo también res-tos de animales que al menos en la actualidad no se encuentran en las inme-diaciones de los sitios, como los pavos (Meleagrididae) y especies afines, así como particularmente los cérvidos (O. virginianus y Mazama spp.). Estos dos grupos de animales constituyen la fauna de aprovechamiento más importante y frecuente en sitios tierra adentro en las tierras bajas mayas del norte (Götz, 2008a, 2008b; Masson y Peraza, 2008; Pollock y Ray, 1957), mientras que no forman parte frecuente de la fauna natural en la costa yucateca (véase Howell y Webb, 2004: 225,226; Reid, 1997: 283).
Los restos de pavo ocelado (Meleagris ocellata) podrían indicar que hubo un intercambio directo y estrecho con asentamientos tierra adentro, quizás para intercambiar sal por carne. La probable conexión entre el sitio de Xcambó y el sitio de Izamal podría haber servido para tal traslado.
También el venado cola blanca, como varias otras especies de vertebrados generalistas que se encontraron en la muestra, habita tanto en la zona norte de la península de Yucatán como en las mencionadas áreas con las que Xcambó mantuvo contacto (véase Reid, 1997), pero no hay indicios certeros de que estos animales se hubieran transportado desde allí. Como mamífero más represen-tado, el venado cola blanca (Odocoileus virginianus) pudo haber sido sujeto al intercambio entre habitantes costeros y de tierra adentro. Las partes esqueléti-cas de estos cérvidos, que representan todo el esqueleto, poseen una gran can-tidad de huellas de hervido y de fuego directo, de cortes y huellas de impacto (Götz, 2008a). Esto concuerda con la gran mayoría de los sitios mayas de las Tierras Bajas, en donde el venado cola blanca fue el mamífero más aprovechado para el consumo en tiempos prehispánicos, y rebasa en sitios tierra adentro, en cantidad, a todos los demás taxones (véanse por ejemplo, Pohl, 1990; Götz, 2008b). Mediante el análisis de las marcas isotópicas en el colágeno de los hue-sos arqueológicos podría evidenciarse un traslado de animales desde áreas no locales (véanse, por ejemplo, Bentley, 2006; Emery y Thornton, 2008), pero estos estudios aún quedan pendientes para la arqueofauna de Xcambó.

1 3 6
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.119-145
La abundancia de restos de pavos y cérvidos en sitios tierra adentro, la falta natural de éstos en la costa hoy día y la presumida existencia de una red de comercio dentro de la península (Andrews, 1998) permiten proponer que los restos de estos animales terrestres representan bienes de intercambio que se recibieron en la costa a cambio de productos marinos como sal, moluscos y peces. Esta hipótesis se ve apoyada por el hecho de que en Xcambó, una gran cantidad (n = 106, no considerados en el desglose numérico de la tabla 2) de restos óseos de cérvidos corresponde a metapodios, afilados en la parte distal en forma de punta y probablemente utilizados para la fabricación y reparación de redes (Canto Méndez, 2009: 93ss.), mientras que no se hallaron los restos postcraneales y craneales correspondientes en cantidad a estos especímenes en el sitio. Según Carr (1987: 15), estos artefactos probablemente llegaron ya fabricados a los sitios mayas costeros, lo que subraya entonces los argumentos al respecto de la propuesta red de intercambio.
Hay que mencionar que a poca distancia de la costa inician en la actua-lidad zonas de bosques bajos donde sí se encuentran pavos, venados y otros animales terrestres (Anthony Andrews, comunicación personal, 2011). Estas zonas también pudieron haber existido en tiempos prehispánicos, abriendo la posibilidad de que los pobladores de los asentamientos hubiesen aprovechado este recurso, obteniendo la fauna alimenticia de asentamientos cercanos o cazándolos ellos mismos. Wanner et al. (2007) y Cucina et al. (2011) proponen que la población del Clásico Tardío del sitio se empeñaba especialmente en la administración del comercio, argumento a favor de que los animales terrestres hayan sido obtenidos por medio del intercambio.
4.2 La foca tropical y el ser humano en las costas yucatecas prehispánicasLa foca tropical (Orden: Carnivora; Familia: Phocidae; Subfamilia: Monachi-nae; Género: Monachus; Especie: tropicalis [Gray, 1850]) (Davies, 2008; Adam, 2004) es una de tres especies de la familia de las “focas verdaderas” (Mangel, 2003: 3), especializadas en clima cálido o tropical. El nombre de la foca en maya fue tsulá (Andrews, 1984: 4); alcanzaba un peso promedio de 135 kg (Davies, 2008), y los individuos adultos tuvieron un color café a gris oscuro (Adam, 2004: 1). Las focas tropicales se alimentaban probablemente de peces y crustáceos (Adam y García, 2003: 305; Adam, 2004: 3) y moraban predominantemente en los arrecifes e islas lejanas de la costa (Adam y García, 2003: 305).
Se conocen tres especies emparentadas del género Monachus, distribuidas en las aguas cálidas del planeta. La especie Monachus monachus es moradora del Mediterráneo, mientras que la especie Monachus schauinslandi habita las costas de Hawái (Adam, 2004: 1; Varona, 1980: 42 y 43). Se presume que las tres espe-

1 3 7
L A A R Q U E O F A U N A D E X C A M B Ó , Y U C A T Á N | C H R I S T O P H E R M . G Ö T Z Y T H E L M A N O E M Í S I E R R A
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.119-145
cies se distribuyeron mediante la corriente ecuatorial en la era terciaria (Baker y Johanos, 2004: 108; Mangel, 2003: 3; Varona, 1980: 43).
Monachus tropicalis se extinguó entre 1950 y 1970, debido a la sobreex-plotación y destrucción del hábitat por el humano (Andrews, 1984; Mangel, 2003; Mignucci-Giannoni y Haddow, 2001; Adam y García, 2003; Davies, 2008; Adam, 2004). Reportes de recientes avistamientos de focas en el área cen-troamericana se atribuyen a otras especies (Mignucci-Giannoni y Haddow, 2001). Hay reportes de pescadores e investigadores que afirman haber visto y, en ocasiones, cazado animales de esta especie, principalmente en el siglo XIX y en la primera mitad del siglo XX, respecto a la costa de Veracruz, las aguas cerca de la ciudad de Campeche, del banco de Campeche y del arrecife Trián-gulos del estado mexicano de Campeche, así como al arrecife Alacrán y a la isla Anina del estado mexicano de Yucatán, pero también en el arrecife Chinchorro del estado mexicano de Quintana Roo y la bahía de Amatique, Belice (Adam y García, 2003: 302-3, 311, Fig. 2; Hairr, 2010). La mayoría de los reportes históri-cos de avistamientos de focas se refieren a las islas caribeñas de Cuba, República Dominicana, Bahamas y Jamaica (Adam y García, 2003: 302-3, 311, Fig. 2). En tiempos históricos ha habido gran abundancia de focas; en la Historia Natural de Jamaica, publicada en 1707 por H. Sloane, se menciona que las Bahamas no solamente estaban llenas de focas, sino que los pescadores capturaban a veces hasta cien en una noche, para luego freírlas y usar el aceite para las lámparas (Andrews, 1984: 6; Varona, 1980: 45).
Fuera de las islas en el golfo de México y el Caribe, hay numerosos hallaz-gos arqueológicos de especímenes de la foca Monachus tropicalis, desde las costas este de Texas, EE. UU., sobre el litoral tanto del golfo de México como del Atlántico de Florida, EE. UU., hasta Puerto Rico, las Antillas Holandesas, Saint Kitts y Nevis y las British Virgin Islands (Adam y García, 2003; Adam, 2004; Walker, 2005). A nivel de Mesoamérica solamente existen los huesos aquí descritos de Xcambó, además de un espécimen arqueológico descubierto en Isla Cerritos, también en Yucatán, como testimonio zooarqueológico del apro-vechamiento de esta especie por los mayas prehispánicos.
El hallazgo de especímenes esqueléticos de foca en dos sitios costeros de la península yucateca, Xcambó e Isla Cerritos permite esbozar una interacción entre el ser humano y estos animales por un largo tiempo, correspondiendo Xcambó cronológicamente a los siglos III a VII, e Isla Cerritos, a los siglos IX a XI. Los datos taxonómicos y osteológicos de los especímenes de foca se ven enriquecidos por las informaciones contextual y tafonómico-antrópica encon-tradas en algunos especímenes. La frecuencia esquelética de los restos de foca en ambos sitios es demasiado baja para hacer inferencias estadísticas en el sen-

1 3 8
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.119-145
tido estricto. Sin embargo, se pueden notar algunas particularidades respecto de los hallazgos.
No podemos asegurar que los animales hayan sido encontrados ya muer-tos en la playa, tal vez ahogados en una tormenta, como se supone para la foca de Pineland (Walker, 2005; véase también Gifford-González et al., 2005: 26), o si fueron cazados, porque los huesos carecieron de huellas antrópicas relacio-nadas con la caza y/o el procesamiento. Parece seguro, sin embargo, que hubo un aprovechamiento ocasional, lo que correspondería a una caza oportunista de estos animales, de manera semejante a lo que se ha reportado en cuanto al aprovechamiento de otras especies de focas por diferentes pueblos prehis-tóricos (véase Smith, 2005: 10). La foca tropical hawaiana busca alimento en incursiones que pueden durar varias semanas, durante el día y la noche, cerca de fondos marinos de poco relieve (Baker y Johanos, 2004: 107; Mangel, 2003: 3), por lo que existe la posibilidad de que los individuos de las muestras hayan sido capturados al buscar alimento cerca de la costa yucateca.
Debido al hecho de que uno de los especímenes de foca de Xcambó corres-ponde a un individuo juvenil, podría argumentarse que su existencia en este sitio, junto a los huesos de uno o varios individuos adultos, significó que estos animales fueron capturados, ¿hembra y cría?, juntos durante el comienzo de la primavera, ya que Andrews (1984: 7) reporta, con base en informes biológicos del siglo XIX, que la época de procreación de las focas tropicales se llevaba a cabo en invierno. No sabemos si existieron áreas específicas de anidación/crianza de focas caribeñas en tierra firme, y los únicos avistamientos de hembras preñadas se hicieron en el arrecife Triángulo (Adam, 2004: 4; Adam y García, 2003).
Si existieron áreas de anidación en la costa de la tierra firme peninsu-lar, éstas tuvieron que haberse ubicado en algún sitio más alejado del asenta-miento de Xcambó, ya que Mangel (2003: 7) reporta, nuevamente de la especie emparentada M. schauinslandi, que aun mínimas perturbaciones del hábitat, como la ocasional presencia del ser humano, causan que las focas abandonen las zonas de visita en las playas (véanse también Baker y Johanos, 2004: 106, y Gifford-González et al., 2005: 26). Otra razón por la que las áreas de anidación, si bien existieron en tierra firme, tuvieron que haber estado lejos de los asenta-mientos se basa en la frecuente presencia de perros, que al menos hoy día tran-sitan aún fuera de los límites de los sitios, habiendo representado posiblemente un depredador de potencial peligro para crías de focas.
La presencia de partes del cráneo y de los huesos grandes apendiculares, así como probablemente de la porción axial, podría sugerir que ciertas par-tes de los animales fueron, al capturarse o hallarse en la playa, separadas del cuerpo, llevándose únicamente las partes de provecho alimenticio o artefac-

1 3 9
L A A R Q U E O F A U N A D E X C A M B Ó , Y U C A T Á N | C H R I S T O P H E R M . G Ö T Z Y T H E L M A N O E M Í S I E R R A
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.119-145
tual. De manera específica, llama la atención que no se hallaron restos de falan-ges, metapodios y podios, lo que podría significar, semejantemente a un caso encontrado en Pineland, Florida (Walker, 2005), que en este caso aplique el llamado efecto schlepp (Perkins y Daly, 1968), con el que el despiece de animales grandes, si se considera que focas tropicales alcanzaban un peso aproximado de 160 kg (Adam, 2004: 2) y hasta 200 kg (Davies, 2008), se llevaba a cabo en el sitio de caza o hallazgo. El cráneo, partes del esqueleto axial y algunas porcio-nes de las extremidades se transportaron posiblemente a los sitios.
No hay evidencia directa de que se haya consumido la carne de la foca, pero la ubicación de sus restos en los basureros secundarios de Xcambó podría apun-tar hacia este uso. Esto resulta interesante, ya que los especímenes se asocian en Xcambó a una parte del asentamiento, donde se presume habitaba la gente socioeconómicamente importante. La rareza de focas cerca de tierra firme, y el hecho de que los mayas prehispánicos parecen no haber podido navegar a los arrecifes alejados de la tierra firme para cazarlos (véase también Gifford-Gonzá-lez et al., 2005: 26), pudieran haber ocasionado que su consumo haya sido restrin-gido a personas de la élite, tal como se menciona, en casos históricos, para grupos indígenas de Florida (Adam, 2004: 4, según Larson, 1980; Andrews, 1984: 5). La utilización de los dientes de la foca para fines ornamentales pudo también haber obedecido a la rareza del animal en el ambiente costero prehispánico.
Por otro lado, tanto la foca caribeña (Adam, 2004) como la foca hawaiana visitaban las playas para descansar, mudar el pelaje, dar a luz o escapar de depre-dadores acuáticos (Mangel, 2003: 5). Para dar a luz y cuidar las crías, las focas hawaianas frecuentan playas con aguas poco profundas, alejadas de depreda-dores (Mangel, 2003: 5, según Westlake y Gilmartin, 1990).
Las focas nunca cumplieron un papel importante en la dieta de los pobladores prehispánicos, ni en Mesoamérica ni en el sur de EE. UU. El hábi-tat de estos animales, alejado del alcance prehispánico, principalmente en islas y arrecifes lejanos de tierra firme (véase arriba, Adam y García, 2003; Adam, 2004; Andrews, 1984: 4-5), fue aparentemente protección suficiente contra una depredación severa (véase también Smith, 2005), en tiempos en los que la navegación aún no permitía moverse en alta mar. El cambio drástico en cuanto al aprovechamiento de éstas y otras especies de focas en diferentes partes del mundo tuvo lugar a partir de la colonización europea (Smith, 2005: 11). El hecho de que especímenes arqueológicos de esta especie se asocian a restos materiales de personas de estatus elevado es posiblemente una respuesta a la rareza de estos animales, en donde el consumo, así como el uso de trofeos y ornamentos, estaban restringidos a los que hubieran tenido el poder de reclamarlos.

1 4 0
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.119-145
Conclusiones
La interacción entre los habitantes humanos del sitio costero de Xcambó y su entorno natural, vista a través de los materiales arqueofaunísticos, muestra que los ocupantes del asentamiento se alimentaban de una amplia gama de anima-les, que incluye miembros de todos los órdenes de animales vertebrados, como peces, reptiles, aves y mamíferos (Jiménez Cano, 2009). La obtención de esta gran variedad de animales vertebrados e invertebrados, al parecer, exclusiva-mente de aguas cercanas al litoral yucateco, fue posible gracias a la riqueza y gran biodiversidad del ambiente natural mencionado, y es de suponer que dicha riqueza en recursos naturales haya posibilitado el asentamiento en la zona costera y el sustento obtenido en áreas cercanas.
Sin embargo, se pudo notar también que la riqueza de recursos faunís-ticos marinos aparentemente no reemplazó por completo las fuentes cárnicas de animales de tierra adentro, que se manifiestan, si bien en pequeña escala, en las muestras costeras de Xcambó, que pueden representar bienes de inter-cambio traídos desde tierra adentro. Un estudio de la ocurrencia de caries entre los antiguos habitantes de Xcambó (Cucina et al., 2011) mostró que los habitantes tanto del Clásico Temprano como del Clásico Tardío padecieron de esta patología, evidenciándose el consumo de maíz, junto con otros alimentos ricos en carbohidratos (cacao, miel, etc.), que también debían traerse de zonas tierra adentro.
Un animal poco aprovechado, pero de gran importancia para la inves-tigación de las zonas litorales prehispánicas, es la foca tropical, de la que se encontraron restos tanto en Isla Cerritos como en Xcambó, y se evidenció que los mayas prehispánicos utilizaron, cuando pudieron conseguirlos, a estos ani-males para fines artefactuales y, muy probablemente, también culinarios.
Las investigaciones zooarqueológicas en sitios prehispánicos costeros han enriquecido nuestra visión acerca del desenvolvimiento de la cultura maya prehispánica en su entorno natural. Nuevas y detalladas excavaciones en otros sitios costeros generarán un mayor conocimiento del aprovechamiento de la fauna y del impacto que produjeron los asentamientos en tiempos prehispáni-cos en el ambiente costero. Futuros proyectos, tanto arqueológicos como pro-piamente zooarqueológicos, pueden intentar acercarse a estas problemáticas, buscando soluciones de las interrogantes.
AgradecimientosQueremos agradecer a la doctora Elizabeth Ramos Roca por la invitación a esta publicación. La elaboración de este artículo no hubiera sido posible sin el apoyo de David A. Herrera Flores y Nayeli G. Jiménez Cano, quienes estudia-

1 4 1
L A A R Q U E O F A U N A D E X C A M B Ó , Y U C A T Á N | C H R I S T O P H E R M . G Ö T Z Y T H E L M A N O E M Í S I E R R A
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.119-145
ron partes importantes de las muestras esqueléticas de Xcambó e Isla Cerri-tos. Fotograf ías de especímenes esqueléticos de foca tropical, usados para la identificación zooarqueológica, fueron facilitadas por Jim Thomason, National Marine Mammal Laboratory, Alaska Fisheries Science Center, NMFS y Carol E. Colaninno, Department of Anthropology, University of Georgia. Peter Adam, Department of Biology, North West Missouri State University, Maryville, Mis-souri, ayudó a la identificación del radio y maxilar de la foca, al igual que a la descripción de rasgos diagnósticos de especímenes de esta especie, realizada en 2006. Cualquier error contenido en estas líneas, sin embargo, será asumido por los autores de este artículo. .

1 4 2
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.119-145
R e f e R e n c i a s
Adam, Peter 2004.“Monachustropicalis”,Mammalian Species747,pp.1-9.
Adam, Peter y Gabriela G. García2003.“NewInformationontheNaturalHistory,Distribution,andSkullSizeoftheExtinct(?)WestIndian
MonkSeal,Monachus tropicalis”,Marine Mammal Science 19(2),pp.297-317.
Andrews, Anthony P.1984.“Laextincióndelafoca(Monachus tropicalis)enYucatán”,Boletín E.C.A.U.D.Y.,12(68),pp.3-12.1998.“ElcomerciomarítimodelosmayasdelPostclásico”,Arqueología Mexicana VI(33),pp.16-23.
Baker, Jason D. y Thea C. Johanos2004.“AbundanceoftheHawaiianMonkSealintheMainHawaiianIslands”,Biological Conservation
116,pp.103-110.
Bastarrachea Manzano, Juan Ramón y Jorge Canto (eds.)2004. Diccionario maya popular.Mérida,Méx.,AcademiadelaLenguaMayaA.C.
Bautista Zúñiga, Francisco, Eduardo Batllori Sampedro, Mario Arturo Ortiz Pérez, Gerardo Palacio Aponte y Miguel Castillo González
2003.“Geoformas,aguaysueloenlapenínsuladeYucatán”,enNaturaleza y sociedad en el área maya: pasado, presente y futuro,eds.PatriciaColunga-MartínyAlfonsoLarqué-Saavedra,pp.21-36.Mérida,Méx.,AcademiaMexicanadeCienciasyCentrodeInvestigaciónCientíficaenYucatán.
Bentley, R. Alexander 2006.“StrontiumIsotopesfromtheEarthtotheArchaeologicalSkeleton:AReview”,Journal of
Archaeological Method and Theory13(3),pp.135-187.
Canto Méndez, Pilar Margarita2009.“Análisistipológicoyfuncionaldelosartefactosdehueso,astaydientedeXcambó”,Yucatán.
TesisdeLicenciaturanopublicada,presentadaenlaFacultaddeCienciasAntropológicas,UniversidadAutónomadeYucatán.Mérida,Méx.
Cárdenas Cervantes, José Miguel2011.“Lautilizacióndelastortugasmarinas(Caretta caretta,Chelonia mydasyEretmochelys imbricata)
enlasociedadmayaprehispánicadeXcambó”.TesisdeLicenciaturanopublicada,presentadaenlaFacultaddeCienciasAntropológicas,UniversidadAutónomadeYucatán.Mérida,Méx.
Carr, Helen Sorayya1987.“PreliminaryAnalysisofNonmolluscanFaunalRemainsfromIslaCerritos,Yucatán”.Informeno
publicado.
Chase, Arlen F., Diane Z. Chase y Wendy G. Teeter2004.“Archaeology,FaunalAnalysis,andInterpretation:LessonsfromMayaStudies”,Archaeofauna13,
pp.11-18.
Chávez, Ernesto A.1994.“LosrecursosmarinosdelapenínsuladeYucatán”,en Recursos faunísticos del litoral de la
península de Yucatán, ed. AlejandroYáñez-Arancibia,pp.1-12.Campeche,Méx.,EPOMEXSerieCientífica;UniversidadAutónomadeCampeche.
Cobos, Rafael, Lilia Fernández Souza, Rodolfo Canto Carrillo, Vera Tiesler Blos, Andrea Cucina, Socorro del Pilar Jiménez Álvarez, Christopher Götz, Guillermo de Anda Alanis, Nancy Peniche May
2007.“ProyectoArqueológicoIslaCerritos:EstudiodeunaComunidadMayaCosteradelClásicoTerminal”.Informedelatemporadadecampo2006,entregadoalConsejoNacionaldeArqueología,InstitutoNacionaldeAntropologíaeHistoria(inédito).

1 4 3
L A A R Q U E O F A U N A D E X C A M B Ó , Y U C A T Á N | C H R I S T O P H E R M . G Ö T Z Y T H E L M A N O E M Í S I E R R A
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.119-145
Contreras Espinosa, Francisco1993.Ecosistemas costeros mexicanos.México:ComisiónNacionalparaelConocimientoyUsodela
Biodiversidad;UniversidadAutónomaMetropolitana,UnidadIztapalapa.
Cucina, Andrea, Cristina Perera Cantillo, Thelma Sierra Sosa y Vera Tiesler Blos2011.“CariousLesionsandMaizeConsumptionamongthePrehispanicMaya:AnAnalysisofaCoastal
CommunityinNorthernYucatan”.American Journal of Physical Anthropology145(4),pp.560-567.
Davies, Laura2008.“Monachustropicalis”(On-line),AnimalDiversityWeb.Consultadoel28demayode2009en
http://animaldiversity.ummz.umich.edu/site/accounts/information/Monachus_tropicalis.html.
EA.FLMNH2003.“Colecciónfotográficadigitaldeelementosesqueléticosde243especiesfaunísticastropicales,
subtropicalesyneárticas”(tomoII),elaboradaporChristopherM.GötzenelFloridaMuseumofNaturalHistory,Gainesville(inédito).
Emery, Kit ty F. y Erin Kennedy Thornton2008.“ARegionalPerspectiveonBioticChangeDuringtheClassicMayaOccupationUsing
ZooarchaeologicalIsotopeChemistry”,Quaternary International191,pp.131-143.
Ericson, Per G. y Jan Storå1999.A Manual to the Skeletal Measurements of the Seal Genera Halichoerus and Phoca (Mammalia:
Pinnipedia).Estocolmo,DepartmentofVertebrateZoology,SwedishMuseumofNaturalHistory.
Froese, Rainer, y Daniel Pauly (eds.)2009.Fishbase.WorldWideWebelectronicpublication.www.fishbase.org,version(03/2009),fechadel
últimoacceso,19demayode2009.
Garza Tarazona, Silvia y Edward B. Kurjack1980.Atlas arqueológico del estado de Yucatán,Mérida,Méx.SEP,INAH,CentroRegionaldelSureste.
Gif ford-González, Diane, Seth D. Newsome, Paul L. Koch, Tom P. Guilderson, Josh J. Snodgrass y R. K. Burton
2005.“ArchaeofaunalInsightsonPinniped-HumanInteractionsontheNortheasternPacific”,en The Exploitation and Cultural Importance of Sea Mammals, ed. GregoryG.Monks,pp.19-38.Oxford,OxbowBooks.
Gilbert, B. Miles, Larry Martin y Howard Savage1996.Avian Osteology.Colombia,Missouri,MissouriArchaeologicalSociety.
Gilbert, B. Miles2003. Mammalian Osteology.Colombia,Missouri,MissouriArchaeologicalSociety.
Götz, Christopher M.2008a.Die Verwendung von Wirbeltieren durch die Maya des nördlichen Tieflandes während der
Klassik und Postklassik (600 - 1500 n.Chr.).InternationaleArchäologie,tomo106.Rahden,Alemania,VerlagMarieLeidorf.
2008b.“CoastalandInlandPatternsofFaunalExploitationinthePrehispanicNorthernMayaLowlands”,Quaternary International191,pp.154-169.
Hairr, John2010. Caribbean Monk Seals: Lost Seals of the Gulf of Mexico and Caribbean Sea.Landisville,
CoachwhipPublications.
Herrera Flores, David2011.“AspectosculinariosrelacionadosalaidentidaddelosantiguoshabitantesdeIslaCerritos,
Yucatán:unestudiozooarqueológico”.TesisdeLicenciaturanopublicada,porpresentarseenlaFacultaddeCienciasAntropológicasdelaUniversidadAutónomadeYucatán.

1 4 4
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.119-145
Howell, Steve N.G. y Sophie Webb2004. A Guide to the Birds of Mexico and Northern Central America.Londres,OxfordUniversityPress.
ITIS(IntegratedTaxonomicInformationSystem;www.itis.gov),consultadoporúltimavezel23demayode2009.
Jiménez Álvarez, Socorro del Pilar2002.“LacronologíacerámicadelpuertomayadeXcambó,costanortedeYucatán:complejocerámico
XcambóyCayalac”.TesisdeLicenciaturanopublicada,presentadaenlaFacultaddeCienciasAntropológicas,UniversidadAutónomadeYucatán.Mérida,Méx.
Jiménez Cano, Nayeli Guadalupe2009.“Ecologíaysubsistenciaenlacostapeninsularyucateca,250-1150d.C.,unacercamiento
paleoecológicoatravésdelaarqueofaunacostera”.TesisdeLicenciaturanopublicada,presentadaenlaFacultaddeCienciasAntropológicas,UniversidadAutónomadeYucatán.Mérida,Méx.
Kaplan, Eugene H.1988.A Field Guide to Coral Reefs: Caribbean and Florida. Peterson Field Guide.NuevaYork,Houghton
andMifflinComp.
Larson, L. H.1980.Aboriginal Subsistence Technology on the Southeastern Coastal Plain During the Late Prehistoric
Period.Gainesville,Florida,UniversityPress.
Lee, Julian G.2000. A Field Guide to the Amphibians and Reptiles of the Maya World.Cornell,CornellUniversityPress.
Mangel, Jef frey C.2003.“SpatialandTemporalHauloutPatternsoftheHawaiianMonkSeal(Monachus schauinslandi)at
MidwayAtoll,1997-2000”.TesisdeMaestríapresentadaenlaNicholasSchoolofEnvironmentalandEarthSciencesdelaDukeUniversity.
Marmolejo Monsiváis, Miguel Ángel2000. Fauna alimentaria de la península de Yucatán.México,InstitutoNacionalIndigenistayPrograma
delasNacionesUnidasparaelDesarrollo.
Márquez Morfín, Lourdes1991.“Ladietamayaprehispánicadelacostayucateca”,Estudios de la Cultura MayaXVIII,pp.359-394.
Masson, Marilyn y Carlos Peraza2008.“AnimalUseatthePostclassicMayaCenterofMayapán,Quaternary International191,pp.170-183.
Mignucci-Giannoni, Antonio y Peter Haddow2001.“CaribbeanMonkSealsorHoodedSeals?”,The Monachus Guardian 4(2),consultado
electrónicamentebajoladireccióndehttp://www.monachusguardian.org/mguard08/08newcar.htm,el23demayode2009.
Mojet ta, Angelo2005. Tiburones. Guía del mundo submarino.Madrid,EditorialDiana
O’Connor, Terry2003. The Analysis of Animal Bone Assemblages: A Handbook for Archaeologists. The Archaeology of
York, Principles and Methods 19/2.York,YorkArchaeologicalTrustyCouncilforBritishArchaeology.
Olsen, Stanley J.1964.Mammal Remains from Archaeological Sites. Papers of the Peabody Museum of Archaeology
and Ethnology56(1).Cambridge,Massachusetts,HarvardUniversityPress.1968.Fish, Amphibian and Reptile Remains from Archaeological Sites. Papers of the Peabody Museum
of Archaeology and Ethnology56(2).Cambridge,Massachusetts,HarvardUniversityPress.

1 4 5
L A A R Q U E O F A U N A D E X C A M B Ó , Y U C A T Á N | C H R I S T O P H E R M . G Ö T Z Y T H E L M A N O E M Í S I E R R A
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.119-145
1982.An Osteology of some Maya Mammals. Papers of the Peabody Museum of Archaeology and Ethnology,vol.73.Cambridge,Massachusetts,HarvardUniversityPress.
Perkins, Dexter Jr. y Patricia Daly1968.“AHunter’SVillageinNeolithicTurkey”,Scientific American219(5),pp.96-106.
Pohl, Mary Ellen Deland1990.“TheEthnozoologyoftheMaya”,en Excavations at Seibal, ed.GordonR.Willey,pp.142-174.
Massachusetts,HarvardUniversityPress.Cambridge.
Pollock, Harry D. E. y Clayton E. Ray1957.Notes on Vertebrate Animal Remains from Mayapan.CurrentReportsNo.41.Washington,
CarnegieInstitutionofWashington,DepartmentofArchaeology.
Reid, Fiona A.1997.A Field Guide to the Mammals of Central America and Southeastern Mexico.NuevaYork,Oxford
UniversityPress.
Reitz, Elizabeth J. y Elizabeth S. Wing2008.Zooarchaeology(segundaedición).CambridgeManualsinArchaeology.Cambridge,Cambridge
UniversityPress.
Robins, Richard, Charleton Ray y John Douglass1986.A Field Guide to Atlantic Coast Fishes. North America.BostonyNuevaYork,HoughtonMiffinCompany.
Sierra Sosa, Thelma N.1997.“ProyectoarqueológicoXcambó.Informetécnicodelasegundatemporada”.CRY-Instituto
NacionaldeAntropologíaeHistoria.Mérida(inédito).1999.“Xcambó.CodiciadoenclaveeconómicodelClásicomaya”,Arqueología Mexicana VII(37),
pp.40-47.2004.“LaarqueologíadeXcambó,Yucatán,centroadministrativosalineroypuertocomercialde
importanciaregionalduranteelClásico”.Tesisdoctoral.UniversidadNacionalAutónomadeMéxico,EstudiosMesoamericanos.México,DF,Méx.
Sierra Sosa, Thelma N. y Ángel Martínez2001.“LosentierrosdeXcambóysusimplicacionessociales”,I’Inaj. Semilla de Maíz. Revista de
divulgación del patrimonio cultural del Yucatán12,pp.6-12.
Smith, Ian2005.“RetreatandResilience:FurSealsandHumanSettlementsinNewZealand”,enThe Exploitation
and Cultural Importance of Sea Mammals, ed.GregoryGMonks,pp.6-18.Oxford,OxbowBooks.
Varona, Luis S.1980.Mamíferos de Cuba.LaHabana,EditorialGenteNueva.
Walker, Karen2005.“PinelandMysteryBoneIsfromMonkSeal–Now–ExtinctAnimalWasPineland1650YearsAgo“,
Friends of the Randall Research Center 4(1).
Wanner, Isabel Sora, Thelma Sierra Sosa, Kurt Alt y Vera Tiesler2007.Lifestyle,Occupation,andWholeBoneMorphologyofthePre-HispanicMayaCoastalPopulation
fromXcambó,Yucatán,Mexico,International Journal of Osteoarchaeology17(3),pp.253-268.
Westlake, R. y W. Gilmartin1990.“HawaiianMonkSealPuppingLocationsintheNorthwestHawaiianIslands”,Pacific Science 44
(4),pp.366-383.

1 4 6

1 4 7
TA FONOM Í A DE H U ESOS DE AV ES . ESTA DO DE L A C U ESTIÓN Y PER SPECTI VA S DESDE EL SU R DEL N EOTRÓPICO
isabel cruz *
[email protected] Nacional de la Patagonia Austral Santa Cruz, Argentina
R e s u m e n Las investigaciones en el sur del Neotrópico
señalan que el impacto de los procesos tafonómicos sobre los
restos de aves varía en relación con el tamaño corporal y la
morfoanatomía de las mismas, así como en función del contexto
ecológico y ambiental considerado. Los estudios futuros deberán
enmarcarse en análisis comparativos para evaluar la acción de
diversos procesos sobre los restos de diferentes vertebrados en
un mismo contexto ambiental, y también deberán desarrollarse
observaciones controladas para profundizar algunos temas.
P A L A B R A S C L A V E :
Tafonomía de vertebrados, aves, meteorización, representación
anatómica, modificaciones por carnívoros.
Artículorecibido:1dejuniode2011|aceptado:10denoviembrede2011|modificado:21denoviembrede2011
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,294pp.ISSN1900-5407,pp.147-174
* DoctoraenArqueología,UniversidaddeBuenosAires,Argentina.

A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
1 4 8
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.147-174
Bird Bones taphonomy. state of the proBlem and perspeCtiVes from the southern neotropiCs
ABSTR AC T Current studies on bird bones
conducted at the Neotropics show that
differences in the impact of taphonomic
processes are mainly due to body size
and anatomy, as well as to the role of
environmental and ecological contexts.
Future research must include comparative
studies and controlled observations to
generate specific models that will help
to understand taphonomic histories of
zooarchaeological remains in the region.
K E Y W O R D S :
Vertebrate Taphonomy, Birds, Weathering,
Anatomic Representation, Carnivore’s
Modifications.
tafonomia de ossos de aVes. estado da questão e perspeCtiVas desde o sul do neotrópiCo
RESUMO As pesquisas no sul do Neotrópico
sinalizam que o impacto dos processos
tafonômicos sobre os restos de aves varia
em relação ao tamanho corporal e a
morfoanatomia das mesmas, assim como em
função do contexto ecológico e ambiental
considerado. Os estudos futuros deveriam
moldar-se em análises comparativos para
avaliar a ação de diversos processos sobre
os restos de diferentes vertebrados num
mesmo contexto ambiental e também
deveriam desenvolver-se observações
controladas para aprofundar alguns temas.
P A L A V R A S C H A V E :
Tafonomia de vertebrados, aves,
meteorização, representação anatômica,
modificações por carnívoros.

1 4 9
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.147-174
TA FONOM Í A DE H U ESOS DE AV ES . ESTA DO DE L A C U ESTIÓN Y PER SPECTI VA S DESDE EL SU R DEL N EOTRÓPICO
i s a b e l c r u z
IntroducciónLas aves presentan características que las convierten en instrumentos interesantes para analizar varios aspectos de la ecología humana. En primer lugar, constituyen un recurso que puede ser uti-lizado integralmente: su carne y sus huevos son un alimento importante, que contiene gran cantidad de proteínas y ácidos grasos. Además, sus plumas y huesos pueden ser utilizados como materia prima para la confección de diver-sos artefactos. En segundo lugar, las aves pueden ser importantes proxys en la determinación de variables paleoecológicas (Emslie, 1995; Morales Muñiz, 1993; Savanti, 1994, entre otros). Muchas aves están estrechamente asociadas a una comunidad vegetal o a un hábitat específico. La presencia de sus huesos en un depósito arqueológico puede ser una línea más en el análisis paleoambien-tal, que necesariamente debe integrar información de varios campos. Debido a que no todos los huesos de aves presentes en estos depósitos son producto de la predación humana, también permiten abordar aspectos relacionados con la formación del registro arqueológico, así como la distribución de especies pre-sentes en un ambiente y su uso por los predadores no humanos.
Durante los últimos años ha aumentado el interés en desarrollar este potencial a partir del estudio de los restos de aves provenientes de depósitos zooarqueológicos en diversos lugares de la región Neotropical (Corona Mar-tínez, 2001, 2002; Cruz, 2007a; deFrance, 2005; Higgins, 1999; Lefèvre, 1989, 1992, 1997; Mameli, 2002; Martínez et al., 2009; Savanti, 1994; Tivoli, 2010, entre otros). A pesar de ello, todavía es poco lo que conocemos acerca de las

A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
1 5 0
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.147-174
aves mediante el registro arqueológico en la región. Uno de los factores que han impedido lograr avances es que sus huesos pueden ser muy escasos en gran parte de los sitios arqueológicos. En virtud de que es poco lo que sabemos acerca de la forma en que se preservan sus restos, es dif ícil dilucidar si esta escasez se debe a que eran poco cazadas o a que sus huesos fueron destruidos por procesos naturales antes de poder incorporarse a los registros arqueológicos.
La disciplina científica que se dedica a comprender cómo se conservan o destruyen los restos orgánicos es la tafonomía. Los estudios tafonómicos cons-tituyen un paso previo al análisis de los restos zooarqueológicos, ya que brin-dan las herramientas interpretativas adecuadas para que las inferencias acerca del pasado sean correctas. El objetivo de este trabajo, por lo tanto, es sintetizar las investigaciones tafonómicas de restos de aves efectuadas hasta el momento en Patagonia (ver la figura 1), con el fin de evaluar el grado de conocimiento alcanzado y establecer las bases para las investigaciones futuras. La Patago-nia se ubica en el sur del Neotrópico (región biogeográfica que abarca desde México hasta Tierra del Fuego), y muchas de las consideraciones presentadas aquí son de utilidad para toda la región. Sin embargo, tal como se enfatizará al final del trabajo, será necesario que en el futuro se desarrollen investigaciones específicas en distintos ambientes de esta región, que completen los estudios presentados aquí y muestren la diversidad ecológica que presentan los proce-sos tafonómicos en diferentes localidades de la región. Esto permitirá contar con un conjunto amplio de principios tafonómicos que podrá ser utilizado por todos aquellos que quieran abordar el estudio de las aves a partir de registros óseos (tanto arqueológicos como paleontológicos).
Zooarqueología y tafonomíaDesde sus inicios como disciplina científica, la zooarqueología ha estado ínti-mamente ligada al desarrollo de los estudios tafonómicos (Andrews, 1990; Binford, 1981; Brain, 1981; Blumenschine, 1986; Gifford, 1981; Lyman, 1994; Pickering et al., 2007, entre otros). La tafonomía es el estudio de los procesos de preservación y de la forma en que éstos afectan a la información que puede obtenerse a partir del registro fósil, tanto paleontológico como arqueológico (Behrensmeyer et al., 2000).
Como el registro arqueológico es el parámetro a partir del cual se deter-mina la relevancia de las explicaciones acerca del pasado humano (Binford, 1981), gran parte de la tarea de los arqueólogos consiste en establecer la histo-ria de formación de los depósitos. Una vez conocida esta historia –o, al menos, la parte de ella accesible a los investigadores–, entonces se puede proceder a evaluar la forma en que el registro en cuestión permite avalar o descartar las

1 5 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.147-174
T A F O N O M Í A D E H U E S O S D E A V E S | I S A B E L C R U Z
hipótesis acerca de la conducta humana. Desde esta perspectiva, el análisis zooarqueológico no sólo implica estudiar los depósitos en cuanto a los actores humanos, sino que también es necesario entender cuál pudo ser la contribu-ción de otros actores y procesos en su formación. No hacerlo implica correr el riesgo de asignar contenido cultural a características de los restos óseos que son producto de procesos naturales.
En el marco de la zooarqueología, uno de los objetivos de los estudios tafonómicos es generar instrumentos interpretativos que permitan garanti-zar las inferencias sobre la conducta humana que se basan en los restos de animales recuperados en contextos arqueológicos. Las observaciones actuales son un medio fructífero para comprender la manera en que diversos actores y procesos afectan a los restos orgánicos, ya que permiten establecer cuáles son las relaciones que existen entre procesos geobiológicos específicos y los conjuntos de huesos que son contemporáneos a los mismos (Binford, 1981; Gifford, 1981, entre otros).
A lo largo de las últimas décadas se han multiplicado las observaciones tafonómicas en el sur del Neotrópico. Sin embargo, ni en esta región ni en otros lugares del mundo los estudios destinados a los restos de aves han sido tan exhaustivos como los dedicados a los mamíferos (Behrensmeyer et al., 2003; Bickart, 1984; Cruz, 2007b; Davis y Briggs, 1998; Livingston, 1989; Muñoz y
Figura1.UbicacióndePatagoniacontinentalyTierradelFuegoenlaregiónNeotropical.

A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
1 5 2
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.147-174
Savanti, 1998; Oliver y Graham, 1994, entre otros). Las investigaciones tafonó-micas sobre huesos de aves en el sur del Neotrópico comenzaron a mediados de la década de 1990 y se desarrollaron fundamentalmente en ambientes de la Patagonia. Aunque todavía escasas, estas investigaciones han permitido cono-cer la forma en que diversos procesos afectan a los restos de estos vertebrados, así como algunos de sus efectos en la formación de los conjuntos óseos recupe-rados de depósitos arqueológicos.
El esqueleto de las aves y los procesos tafonómicosLa interacción entre huesos y procesos tafonómicos está mediada por varios factores, entre los cuales se destacan las propiedades específicas de los hue-sos de cada grupo de organismos. El esqueleto de las aves difiere del de otros vertebrados en varios aspectos, especialmente los relacionados con la princi-pal adaptación locomotora del taxón: el vuelo. El modo de locomoción plantea constricciones anatómicas y, aunque no todas las aves son voladoras, muchos aspectos corporales de las aves reflejan los especiales requerimientos que demanda el vuelo (Livingston, 1989).
Los huesos de cualquier vertebrado tienen una función, que se relaciona con su “fuerza tafonómica” (Lyman, 1994). La función anatómica de un hueso determina sus propiedades mecánicas, las que a su vez median sobre los efectos que los procesos tafonómicos tendrán sobre ese hueso. Muchas de estas pro-piedades de los huesos pueden ser tafonómicamente significativas: el diseño y el tamaño corporal, así como la microestructura del tejido óseo, son facto-res que influirán en la destrucción o preservación de los restos de cada taxón (Andrews, 1990; Behrensmeyer, 1991; Blumenschine, 1987; Kidwell y Behrens-meyer, 1988; Lyman, 1994, entre otros). En el caso de las aves, entre los reque-rimientos para el vuelo que tienen correlatos importantes en el esqueleto se cuentan una reducción general del peso, la obtención de mayor rigidez corporal y una mayor resistencia para sobrellevar el stress que implica el vuelo (Bellairs y Jenkin, 1960; Feduccia, 1992; Gilbert et al., 1981). Debido a ello, varios inves-tigadores han planteado la necesidad de efectuar estudios tafonómicos especí-ficos para sus restos (Cruz, 2000a, 2007b; Davis y Briggs, 1998; Higgins, 1999; Lyman, 1994; Trapani, 1998, entre otros).
Como grupo, las aves exhiben pocas diferencias osteológicas entre sí (Bellairs y Jenkin, 1960), pero presentan gran variabilidad con respecto al tamaño. Éste es un factor que debe ser considerado al analizar el impacto de los procesos tafonómicos sobre sus huesos, porque se ha demostrado que procesos tafonómicos como la meteorización, la acción de carnívoros o el pisoteo varían

1 5 3
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.147-174
T A F O N O M Í A D E H U E S O S D E A V E S | I S A B E L C R U Z
según el tamaño corporal de los animales implicados (Andrews, 1990; Behrens-meyer, 1978; Behrensmeyer y Dechant-Boaz, 1980; Behrensmeyer et al., 2003; Blumenschine, 1987).
Por otro lado, en relación con el vuelo, muchos de los huesos del cráneo y del esqueleto poscraneano de las aves son huecos y contienen sacos de aire (Bellairs y Jenkin, 1960; Higgins, 1999). El grado de neumatización varía entre taxones: es mínimo en las aves más pequeñas, mientras que involucra la mayo-ría de los huesos de las aves grandes y activamente voladoras, como los albatros y las águilas. También entre los Ratites, aves no voladoras, los huesos presentan un alto grado de neumatización, característica posiblemente derivada de sus ancestros adaptados al vuelo. En cambio, los huesos de las aves buceadoras están muy poco neumatizados y sus esqueletos reflejan el equilibrio entre los requerimientos del vuelo y del buceo (Gilbert et al., 1981).
En síntesis, en función de las necesidades que plantea el vuelo, las aves han desarrollado un esqueleto más liviano y rígido, con menor cantidad de ele-mentos individuales que el de otros vertebrados. Además, también la microes-tructura y la morfología de los huesos han sufrido modificaciones. Todos estos aspectos tienen particulares implicaciones para la tafonomía de sus huesos y permiten predecir que los procesos tafonómicos no los afectarán de la misma manera que a los de otros vertebrados.
Aspectos metodológicos Todos los estudios tafonómicos dedicados a los restos de aves efectuados en Patagonia se desarrollaron en el marco de lo que Marean (1995) denominó tafonomía naturalista. A diferencia de la tafonomía experimental, el enfoque naturalista permite conocer las condiciones variables en las que se conforman los registros óseos derivados de comunidades específicas, en ambientes con-cretos. Para ello, las investigaciones no se realizan en laboratorios o ambien-tes controlados sino que se basan en la observación de los restos depositados naturalmente. De esta manera, es posible establecer nexos entre los procesos tafonómicos y sus trazas en los restos.
Tal como lo efectuaron otros investigadores en diferentes ecosistemas de África (Behrensmeyer, 1993; Behrensmeyer y Dechant-Boaz, 1980; Behrens-meyer et al., 2003; Blumenschine, 1989; Bunn et al., 1991; Sept, 1994; Tappen, 1995, entre otros), la metodología utilizada en Patagonia consistió en el registro y posterior análisis de todas las características de los restos óseos actuales a lo largo de unidades espaciales amplias, por medio de la implementación de tran-sectas en diferentes sectores del paisaje (Belardi, 1999; Cruz, 1999, 2000b, 2004, 2007b; Muñoz y Savanti, 1998, entre otros).

A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
1 5 4
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.147-174
Las observaciones se centraron en aquellas aves de tamaño mediano a grande, que son las de posible utilidad económica para las poblaciones huma-nas del pasado (ver la figura 2). Con excepciones que luego se detallan, la mayor parte de estas aves están incluidas en lo que Behrensmeyer (1991) con-sidera como microvertebrados, es decir, con un peso menor a 5 kg. Dentro del marco de este rango de tamaño corporal, las observaciones diferenciaron por lo menos tres grupos de aves en función de su morfoanatomía. El fundamento de esta diferenciación es que las características anatómicas tienen correlatos en la constitución del esqueleto y, por lo tanto, pueden ocasionar diferencias en el potencial de preservación de huesos correspondientes a distintas arqui-tecturas corporales.
Figura2.Algunasdelasavespatagónicasmáscomunes: a)cauquéncomún(Chloephaga picta),b)pingüinodeMagallanes(Speniscus magellanicus),c)ñandúpetiso(Rhea pennata)yd)martineta(Eudromia elegans).
En función de esta premisa, durante las observaciones efectuadas en Patagonia se consideraron las características que presentaban los restos de aves voladoras, de ñandúes (Rheidae) –aves no voladoras del grupo de las Rati-tes– y de pingüinos (Spheniscidae) –aves marinas no voladoras–. Las anato-mías de estas aves presentan marcadas diferencias entre sí y sus pesos varían entre los 27 kg de los ñandúes, los 3,5-4 kg de los pingüinos y la diversidad de pesos (entre unos pocos cientos de gramos hasta los 4 kg) de las aves voladoras consideradas en la región.
a) b)
c) d)

1 5 5
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.147-174
T A F O N O M Í A D E H U E S O S D E A V E S | I S A B E L C R U Z
Los resultados obtenidos
Las investigaciones desarrolladas tanto en el sur del continente (Belardi, 1999; Cruz, 1999, 2000b, 2007b, entre otros) como en la Isla Grande de Tierra del Fuego (Muñoz y Savanti, 1998) permitieron conocer diversos aspectos relacio-nados con la depositación, modificación y destrucción de huesos de aves por los procesos tafonómicos en diferentes ambientes (la costa marina, la estepa y el bosque andino-patagónico). Muchos de estos resultados son concordantes con los obtenidos en otros lugares del mundo (Behrensmeyer et al., 2003; Bic-kart, 1984; Davis y Briggs, 1998; Oliver y Graham, 1994) pero también mostra-ron diferencias en algunos aspectos (Cruz, 2007b, 2008).
Como en otros estudios actualísticos, las investigaciones patagónicas se centraron en dos aspectos relacionados con la preservación de restos orgá-nicos: la depositación y las modificaciones que se producen antes de que los mismos sean cubiertos por sedimentos (sensu Behrensmeyer et al., 2000). Esto tiene sentido si se considera que muchos de los factores que determinan las condiciones bajo las que se preservarán algunos organismos –o partes de los mismos– se definen en el momento de la muerte y/o en un corto lapso des-pués de ella (Binford, 1981; Andrews, 1990; Behrensmeyer, 1991; Lyman, 1994, entre otros). Conocer la dinámica relativa a estos dos aspectos bajo condiciones ambientales particulares es un medio para establecer cuántos y cuáles podrán ser los restos que se preservarán o los que, por lo menos, tendrán posibilidades de ser cubiertos por los sedimentos.
Aunque los temas abordados por estos estudios han sido muchos, aquí se sintetizan los resultados alcanzados en relación con algunos de ellos: la depo-sitación de restos por causas naturales y su posible impacto sobre los registros arqueológicos de la región, la secuencia de desarticulación y la representación anatómica de aves, la meteorización y la acción de los carnívoros. Todos estos temas se consideran particularmente relevantes para una adecuada interpreta-ción de los restos de aves contenidos en depósitos arqueológicos, para enten-der los contextos en los que los mismos se produjeron y como un instrumento más entre los varios necesarios para generar expectativas acerca de los registros óseos derivados de la actividad humana.
La depositación de huesos por causas naturalesUna de las formas más evidentes en que los procesos naturales pueden afectar a los registros arqueológicos es mediante la acumulación de restos en lugares recurrentemente utilizados por los humanos, que forman palimpsestos en los que se mezclan huesos depositados por distintos procesos y actores.

A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
1 5 6
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.147-174
Binford (1981) define “integridad” como la homogeneidad relativa de los agentes causantes de la acumulación de los materiales presentes en un depó-sito arqueológico. Una herramienta útil para evaluar la integridad de los regis-tros arqueofaunísticos en diferentes puntos del espacio y establecer las posi-bilidades de “contaminación” con huesos depositados por causas no humanas es conocer la dinámica que determina la depositación natural de restos en un ambiente dado. En un mismo paisaje existen variaciones en la depositación de restos (Behrensmeyer y Dechant-Boaz, 1980; Cruz, 2004, 2009a; Sept, 1994; Tappen, 1995, entre otros). La posibilidad de que exista superposición entre artefactos u otros restos arqueológicos y huesos derivados de muertes naturales variará en función de las características del registro arqueológico y de la forma que asuma la depositación natural de restos (Borrero, 2001).
En Patagonia, la mayor diferencia en este sentido se encuentra entre los hábitats costeros y los del interior (tanto sectores de estepa como de bosque y humedales). En los hábitats costeros se producen grandes concentraciones de restos durante el período de nidificación y cría de las aves marinas, cuando la mortalidad es generalmente muy alta (Cruz, 1999, 2007b, 2009a). Dadas las características que presenta el registro arqueológico en muchos sectores coste-ros de la región, las probabilidades de “contaminación” de depósitos arqueoló-gicos por la muerte natural de aves son muy altas. Como se ha planteado para varios casos en Patagonia continental, es posible que todos o gran parte de los restos de aves relacionados con un registro arqueológico particular sean pro-ducto de la depositación natural, debido a la dinámica de las colonias de nidifi-cación (Cruz, 1999, 2004, 2006, 2009a).
En cambio, en la estepa y el bosque característicos del sur de Patagonia las posibilidades de superposición o mezcla con restos arqueológicos son muy bajas, ya que, si se compara con lo que ocurre en la costa, son muy pocos los huesos de aves que se depositan, y, además, en virtud de la inten-sidad de los procesos tafonómicos, es factible que gran parte de ellos sean destruidos antes de ser cubiertos por sedimentos (Cruz, 2000b, 2007c, 2008, 2009a). La excepción detectada la constituyen los abrigos rocosos, que son sectores de gran acumulación natural por la acción de los carnívoros y por muertes naturales (Cruz, 2000a, 2000b, 2007a, 2009a). Los huesos de aves voladoras son frágiles y su destrucción puede ser rápida si no son cubiertos por sedimentos poco tiempo después de la depositación. Los abrigos rocosos constituyen puntos en el paisaje que generalmente presentan altas tasas de sedimentación o condiciones que atemperan la acción de los procesos des-tructivos, lo que facilita la conservación de huesos de aves. Como muchos de los depósitos arqueológicos en Patagonia continental se ubican en abrigos

1 5 7
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.147-174
T A F O N O M Í A D E H U E S O S D E A V E S | I S A B E L C R U Z
rocosos, el potencial para la mezcla con restos derivados de muertes natura-les es muy alto en estos loci.
En síntesis, conocer la forma general que adopta la depositación natural de restos en un ambiente dado puede ser una herramienta útil para modelar la integridad de los depósitos arqueológicos y para generar los medios que permi-tan evaluarla en cada caso.
Secuencia de desarticulaciónOtro de los temas al que se le ha prestado especial atención en la tafonomía de aves es la secuencia de desarticulación. Este interés ha sido importante en los estudios tafonómicos de otros vertebrados y se vincula con el hecho de que la representación de partes esqueletarias, utilizada frecuentemente para efectuar inferencias acerca del comportamiento humano, puede estar relacionada con esta secuencia. Si bien la desarticulación en sí misma no implica la destrucción o desaparición de los elementos, aumenta la vulnerabilidad de los huesos frente a la acción de otros procesos.
La secuencia de desarticulación de aves voladoras sigue un patrón que, en líneas generales, ha sido registrado mediante observaciones actuales en diferen-tes lugares del mundo (ver la tabla 1). Entre los aspectos más recurrentes, se ha observado que las extremidades posteriores se desarticulan de la carcasa antes que las anteriores. Asimismo, los elementos de las extremidades posteriores se desarticulan entre sí más rápidamente que los de las extremidades anterio-res. Estas últimas permanecen más tiempo como una unidad, frecuentemente junto a coracoides, fúrcula, escápula y, en algunos casos, esternón. En ambas extremidades, las porciones proximales se desarticulan antes que las distales. Estos aspectos de la secuencia planteada para carcasas de aves voladoras son coincidentes con la detectada en Patagonia, y también se cumplen en los restos de pingüinos (Cruz, 2007b).
En las carcasas de ñandúes, en cambio, los primeros elementos en des-articularse son el cráneo, la cintura escapular, el esternón y las extremidades anteriores, que son rápidamente destruidos por los procesos tafonómicos. Vér-tebras y costillas pueden permanecer articuladas como una unidad en conjunto con la pelvis, pero se desarticulan y destruyen antes que las extremidades pos-teriores. Estas extremidades, especialmente la porción distal (tibiatarso, meta-tarso y dígitos), pueden permanecer articuladas por largo tiempo y sus elemen-tos son los más resistentes a los procesos destructivos (Belardi, 1999; Cruz, 2007c; Cruz y Elkin, 2003; Cruz y Fernández, 2004).
Estas diferencias en las secuencias de desarticulación pueden correlacio-narse con diferencias en la preservación de partes esqueletarias de las distintas

A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
1 5 8
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.147-174
aves patagónicas. En efecto, aquellos elementos que se desarticulan primero son los que posiblemente se destruyan primero. Los elementos correspondien-tes a las extremidades anteriores de las aves voladoras, que permanecen arti-culados por más tiempo, son los que posiblemente se conserven mejor (ver la figura 3) en la superficie terrestre. En el caso de los ñandúes, los elementos que perdurarán más tiempo articulados son los de las extremidades posteriores, por lo que serán los últimos en destruirse y los que, por lo tanto, tendrán más posibilidades de preservarse tanto en contextos naturales como arqueológicos.
Tabla1.Secuenciasdedesarticulacióndecarcasasdeaves (tomadodeBickart,1984;OliveryGraham;1994,yDavisyBriggs,1998).
Bickart(1984) OliveryGraham(1994) DavisyBriggs(1998)
Lascostillassedesarticulandelesternón
Launidadcabeza-vértebrascervicalessedesarticuladelasvértebrastorácicas
Elcráneoylasvértebrascervicalescomounidadseseparandeltórax
Lasextremidades posterioressedesarticulandeltronco
Elcráneoseseparadelascervicales Elfémurseseparadelsinsacro
Desarticulacióndelasvértebras
Elesternónylasalascomounidadseseparandelrestodelacarcasa
Lacinturaescapularylas extremidadesanteriorescomounidadseseparandeltórax
Desarticulacióndelacinturaescapular
Seseparanelesternónyelcoracoides Lasvértebrastorácicasse desarticulan,causandoque eltóraxseseparedelsinsacro
Laporciónproximaldelasalassedesarticulaantesqueladistal
Lasextremidadesposterioresseseparandeltronco
Lascostillasseseparandelasvértebrastorácicas,lasqueasuvezseseparanentresí
Lasvértebrastorácicasseseparandelsinsacro
Sedesarticulanlasextremidadesposteriores
Elcoracoidesseseparadelhúmero Loselementosdelasextremidadesanterioresylacinturaescapularsedesarticulanentresí
Launidadtarsometatarso-falangesseseparadelaporciónproximaldelaextremidadposterior
Lasfalangessedesarticulandeltarso-metatarso
Eltibiatarsoyelfémursedesarticulan
Elhúmerosedesarticuladelaporcióndistaldelasalas
Radioyulnasedesarticulandelcarpo-metacarpoylasfalanges

1 5 9
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.147-174
T A F O N O M Í A D E H U E S O S D E A V E S | I S A B E L C R U Z
Representación de partes esqueletariasLa frecuencia de partes esqueletarias es una de las variables que permiten dis-cutir la historia tafonómica de un conjunto óseo (Behrensmeyer, 1991) y es uno de los medios más utilizados por los arqueólogos para abordar diversos aspec-tos de la subsistencia humana. Debido a ello, muchos de los estudios tafonómi-cos se han dedicado muy especialmente a establecer las diferencias que existen entre el patrón natural de representación anatómica y el derivado de la acción humana (Cruz, 2005; Ericsson, 1987; Livingston, 1989, entre otros).
Las investigaciones efectuadas dejaron en claro que la representación de partes esqueletarias de aves es el resultado de un conjunto complejo de procesos, que varía en función de gran número de variables (tamaño de las presas, tipo de predador implicado, si constituyen restos de comida o son restos regurgitados, etc.). Incluso, tal como ha planteado Bovy (2002) al res-pecto, la variabilidad puede extenderse a los conjuntos generados por acti-vidades humanas, ya que pueden existir diferencias en función del tipo de aprovechamiento efectuado.
Entre los logros más importantes de las investigaciones en Patagonia, se cuenta el haber definido patrones naturales de representación anatómica diferentes para cada uno de los tipos de aves considerados. Entre los restos de aves voladoras predominan los elementos correspondientes a las extremidades anteriores (especialmente húmeros, radios y ulnas), las vértebras, así como los tibiatarsos (Cruz, 2005; Muñoz y Savanti, 1998). En el caso de los ñandúes, los elementos más representados son los de las extremidades posteriores (tibiatar-sos y tarsometatarsos) y las vértebras (Belardi, 1999; Cruz, 2000, 2005; Cruz y Elkin, 2003; Cruz y Fernández, 2004). Los restos de pingüinos más abundantes son los correspondientes a ambas extremidades (especialmente húmero, fémur y tibiatarso) y el coracoides (Cruz, 1999, 2005, 2007b; Emslie, 1995). La cintura pélvica es la porción anatómica menos representada en todos los casos.
Figura3.Restosarticuladosdeunjuvenildegaviotacocinera(Larus dominicanus).

A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
1 6 0
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.147-174
En síntesis, la representación de partes esqueletarias en conjuntos natu-rales no es azarosa, sino que presenta patrones consistentes. Las diferencias detectadas no pueden atribuirse a las respectivas historias tafonómicas: en todos los casos estos patrones son el resultado de la acción conjunta de varios procesos tafonómicos (principalmente, meteorización y acción de carnívoros) que facilitan la destrucción diferencial de elementos. Tampoco reflejan un pro-blema de muestreo, ya que se puso especial cuidado en relevar hábitats especí-ficos de cada una de las aves consideradas. Contrariamente a lo planteado por algunos investigadores, las investigaciones patagónicas mostraron que estos patrones son un correlato de la morfoanatomía de cada una de las aves y no se relacionan con el agente causal de la depositación (Cruz, 2005).
El caso de los ñandúes constituye un ejemplo de aplicación de estos resul-tados: el patrón natural observado en los estudios tafonómicos es coincidente con la abundancia anatómica detectada en varios sitios arqueológicos de Pata-gonia y de todo el sur de Sudamérica en general. Las primeras interpretaciones de esta representación de partes esqueletarias proponían que la misma era el resultado del descarte de las extremidades posteriores en los sitios de caza, debido a que sólo se transportaban hasta los campamentos aquellas porcio-nes con mayor contenido cárnico, que es variable entre elementos esqueleta-rios (Giardina, 2006). Sin embargo, esta representación anatómica también es característica de los conjuntos naturales y puede explicarse como el resultado de la destrucción mediada por la densidad (Cruz y Elkin, 2003; Fernández et al., 2001, entre otros). Por lo tanto, la abundancia anatómica de los conjun-tos arqueológicos no debería atribuirse directamente a un comportamiento humano específico (el transporte de partes de una presa y el descarte de otras), ya que también puede ser el resultado de un gran número de procesos, en los que la destrucción mediada por la densidad mineral tiene el papel preponde-rante. Si bien las investigaciones acerca del aprovechamiento del ñandú por las poblaciones prehistóricas aún deben profundizarse, los estudios tafonómicos tuvieron un rol central para descartar una hipótesis y señalar nuevas líneas de investigación, las cuales deberán incluir explicaciones más complejas acerca de la representación de partes esqueletarias que presentan los restos de estas aves en los conjuntos arqueológicos de la región.
MeteorizaciónLa meteorización es uno de los principales procesos que causan la destrucción de restos óseos en la superficie terrestre (Behrensmeyer, 1978; Lyman, 1994). Los huesos de distintos animales presentan diferentes tasas de meteorización en un mismo ambiente (Gifford, 1981), y el tamaño corporal es uno de los

1 6 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.147-174
T A F O N O M Í A D E H U E S O S D E A V E S | I S A B E L C R U Z
factores determinantes para que esto ocurra (Behrensmeyer, 1978). Debido a las características de los huesos de aves y a su tamaño corporal generalmente pequeño, el impacto de la meteorización sobre ellos es muy diferente al que sufren los huesos de otros animales, especialmente los mamíferos. Por otro lado, como la meteorización es un proceso ecológicamente mediado, son nece-sarias investigaciones específicas para comprender la forma en que puede afec-tar los depósitos arqueológicos en un ambiente particular.
Las primeras investigaciones en Patagonia dedicadas a los restos de aves fueron las de Muñoz y Savanti (1998) en la Isla Grande de Tierra del Fuego. A partir de sus observaciones, estos investigadores formularon varios estadios de meteorización para los huesos de aves que, en líneas generales, seguían los linea-mientos propuestos por Behrensmeyer (1978) para los huesos de mamíferos de más de 5 kg. Estudios posteriores efectuados en Amboseli (África) corroboraron que los estadios inicialmente propuestos para los mamíferos también son útiles para evaluar la meteorización en huesos de aves (Behrensmeyer et al., 2003).
A partir del perfil de meteorización de huesos de aves derivado de los resultados de sus observaciones, Muñoz y Savanti (1998) plantearon que el pre-dominio de los estadios más bajos se debe a que las propiedades mecánicas de estos huesos facilitan su destrucción antes de alcanzar una meteorización avanzada. Estos resultados fueron confirmados por los estudios en el sector continental de Patagonia (Cruz, 2000a, 2007b, 2008, entre otros) y son acordes con los derivados de las investigaciones en Amboseli (Beherensmeyer et al., 2003). En todos los casos, la acción de la meteorización provocó que los huesos de aves desaparecieran más rápidamente que los de mamíferos, especialmente de ungulados. Existe un aspecto, sin embargo, en el que los resultados de Pata-gonia y Amboseli difieren: la destrucción de los huesos de aves producida por la meteorización es más rápida en Patagonia (Cruz, 2008). Esto es acorde con lo registrado para los huesos de ungulados en la región (Borrero, 2007), por lo que debe considerarse una consecuencia de las condiciones ambientales preva-lecientes en el sur del continente.
Un último punto por destacar se relaciona con las diferencias registradas entre los huesos de los distintos grupos de aves patagónicas con respecto a la meteorización. La comparación entre los perfiles de meteorización de huesos de aves voladoras y pingüinos en Patagonia permite afirmar que las diferencias que pueden existir en la estructura de sus huesos no son importantes frente a la acción de este proceso tafonómico (Cruz, 2007b). En cambio, se registraron diferencias al respecto en el caso de los ñandúes, ya que todas las observaciones destacan que los elementos correspondientes a las extremidades posteriores de estas aves tienen mayor resistencia a los procesos tafonómicos, incluida la

A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
1 6 2
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.147-174
meteorización (Belardi, 1999; Cruz, 2007a; Cruz y Fernández, 2004). Esta resis-tencia puede atribuirse a dos factores: 1) los ñandúes son las aves de mayor tamaño corporal en Patagonia, y 2) estas aves están adaptadas para correr a grandes velocidades, por lo que los huesos de sus extremidades posteriores –especialmente tibiatarso, tarsometatarso y dígitos– poseen paredes gruesas y altos valores de densidad mineral (Cruz y Elkin, 2003). Como consecuencia, parte del esqueleto del ñandú (especialmente las extremidades anteriores, el cráneo y el esternón) comparte con el de las aves voladoras una rápida des-trucción por la meteorización, mientras que las extremidades posteriores se asemejan a los huesos de ungulados, que poseen una mayor resistencia a este proceso (Cruz, 2007c).
Modificaciones y transporte por carnívorosA diferencia de lo registrado en otros ecosistemas (Binford, 1981; Blumens-chine, 1989; Tappen, 1995, entre otros), los carnívoros patagónicos no tienen gran potencial para destruir huesos y carcasas de ungulados (Borrero, 1990, 2000; Cruz, 2008, entre otros). Sin embargo, en función de su pequeño tamaño corporal y su particular estructura ósea, las aves constituyen presas cuyos hue-sos y carcasas pueden ser profundamente afectados por la acción de predado-res y carroñeros (Bickart, 1984; Oliver y Graham, 1994).
Las observaciones efectuadas en Patagonia permitieron establecer que el impacto de la acción de los carnívoros varía según las características de las comunidades de aves y las condiciones ecológicas imperantes en distintos ambientes de la región. Esto implica que para una adecuada evaluación del impacto que puede tener este proceso sobre los restos de aves hay que conside-rar los contextos en que se producen la predación y el carroñeo. La costa marina presenta marcadas diferencias con los otros ambientes en lo que respecta a sus comunidades de aves, lo cual determina el grado en el que las modificaciones producidas por los carnívoros influyen en la conformación del registro (Cruz, 2004, 2007b, 2008; Muñoz y Savanti, 1998). En la costa, la gran mortalidad de aves concentrada espacial y temporalmente se expresa en una gran deposita-ción de restos, que constituyen una importante fuente de alimento disponible para los carnívoros locales. Entonces, si bien podría esperarse una gran des-trucción inicial en función del tamaño corporal de las aves, las condiciones de abundancia de carcasas permiten la preservación de gran parte de los restos. En la estepa y el bosque, en cambio, las aves son menos abundantes y las carcasas disponibles son escasas y están muy dispersas, por lo que los carnívoros las aprovechan más intensamente. Como parte de este aprovechamiento integral, se produce una mayor destrucción de las carcasas, y, además, el transporte de

1 6 3
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.147-174
T A F O N O M Í A D E H U E S O S D E A V E S | I S A B E L C R U Z
algunas partes anatómicas por los predadores afecta la localización de los hue-sos de aves (Cruz, 2000b, 2007c, 2008).
Los autores que han efectuado observaciones sobre la forma en que los carnívoros modifican los huesos y carcasas de aves concuerdan en que exis-ten diferencias según se trate de predadores mamíferos o aves (Davis y Briggs, 1998; Oliver y Graham, 1994). Los mamíferos generalmente producen muchas modificaciones y destrucción y pueden transportar los restos fuera del sitio de muerte. En cambio, en general las aves carnívoras tienen una menor capacidad de modificación de los huesos y, si ocurre transporte, el mismo es de carne, vísceras o partes anatómicas segmentadas.
En Patagonia, aves y mamíferos también tienen diferentes roles en la modificación de huesos y carcasas (Cruz, 1999, 2004, 2007b). En la costa marina se ha registrado poca acción de los mamíferos terrestres sobre las carcasas de aves; en cambio, es importante la acción de las aves, que generalmente se cen-tra en los tejidos blandos. Tal cual se observó en otros lugares del mundo, su papel como actores tafonómicos consiste fundamentalmente en contribuir a la desarticulación de las carcasas. Sin embargo, en un área de nidificación de cor-moranes emplazada en una isla en la que no habitan mamíferos se registraron restos de aves con modificaciones en el esternón (Cruz, 2009b). Estas modifi-caciones, similares a los punctures producidos por mamíferos y descritos por Binford (1981), fueron efectuadas por gaviotas (Larus dominicanus) o eskúas (Catharacta skua), e indican que en algunos casos las aves pueden producir efectos sobre los huesos.
En la estepa patagónica, en cambio, son los mamíferos quienes suelen destruir, modificar y transportar carcasas y huesos de aves (Belardi, 1999; Cruz, 2000a; Muñoz y Savanti, 1998). El transporte se produce principalmente a sus madrigueras de cría, que en este ambiente suelen ubicarse en abrigos rocosos (Cruz, 2000a; Fernández et al., 2010; Martín, 1998). No todos los huesos de aves en estas madrigueras presentan modificaciones claramente asignables a carnívoros, por lo que, si en el abrigo existen depósitos arqueológicos, estos restos pueden incorporarse a los mismos sin trazas que permitan establecer su procedencia (Fernández y Cruz, 2010).
Como en el caso de la meteorización, existen diferencias entre las distin-tas aves en lo que respecta a las modificaciones por estos actores tafonómicos. Los huesos de los ñandúes, en función de su tamaño y estructura, son más resistentes a la acción de los carnívoros. Los daños registrados no afectan a todo el esqueleto, sino que se ubican principalmente en las epífisis de huesos largos, el esternón y cintura pélvica (Belardi, 1999) (ver la figura 4). La destruc-ción de las carcasas de aves voladoras y pingüinos, en cambio, puede ser más

A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
1 6 4
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.147-174
completa, y los huesos con más probabilidades de sobrevivir son los corres-pondientes a las extremidades anteriores y la porción distal de las posteriores (Cruz, 2000a, 2005, 2007b).
Aplicación a casos zooarqueológicosRobert Blumenschine (1998) planteó que la habilidad de los arqueólogos para “leer” el registro arqueofaunístico de manera confiable y verificable no está a la altura de la creatividad alcanzada por las explicaciones acerca del pasado. Para este investigador, el desarrollo desigual de estos dos aspectos de las inves-tigaciones zooarqueológicas no se debe a la carencia de principios y modelos tafonómicos, ya que afortunadamente los mismos son muchos y variados. La causa de esta inexplicable falta de profundización de los aspectos tafonómicos involucrados en la interpretación arqueológica es que muchos zooarqueólogos todavía se niegan a aplicar los modelos de esta disciplina en sus interpretacio-nes de los conjuntos que describen (Blumenschine, 1998). En los comienzos de la segunda década del siglo XXI, esto continúa ocurriendo en gran parte de las investigaciones en la región Neotropical. Por lo tanto, además del ejem-plo mencionado en relación con el aprovechamiento de Rheidos en el sur de Sudamérica (ver “Representación de partes esqueletarias”), aquí presentaremos brevemente dos casos que permiten apreciar el potencial de los modelos tafo-nómicos para una adecuada interpretación zooarqueológica.
El primer caso se vincula con la necesidad de evaluar si los restos de aves recuperados en varios sitios arqueológicos (CCP 5, CCP 7, ADG y AGV) en abrigos rocosos ubicados en el sector cordillerano del sur de Patagonia continental en Argentina derivan de la actividad humana o son el producto de procesos tafonómicos, como la acción de los carnívoros (Cruz, 2000a, 2007a). Teniendo en cuenta las altas posibilidades de superposición de huesos deri-
Figura4.Sinsacrodeñandúpetiso(Rhea pennata)conmodificacionesefectuadasporcánidos.Elcírculoseñalalazonaafectada.

1 6 5
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.147-174
T A F O N O M Í A D E H U E S O S D E A V E S | I S A B E L C R U Z
vados de muertes naturales y restos arqueológicos en los abrigos rocosos (ver “La depositación de huesos por causas naturales” y “Modificaciones y trans-porte por carnívoros”), se planteó la necesidad de efectuar un análisis tafo-nómico que incluyese diversas líneas de evidencia para evaluar las hipótesis planteadas acerca de la explotación de aves en este sector cordillerano. El énfasis de este análisis estuvo puesto en la caracterización de la acción de los carnívoros en el área (Cruz, 2000a) y en las propiedades del registro óseo de los sitios arqueológicos de la localidad (Cruz, 2000b, 2007a). Por ejemplo, en varios de los conjuntos zooarqueológicos se registraron huesos de carní-voros sin modificaciones antrópicas, por lo que es posible pensar que estos animales murieron naturalmente en el abrigo y que debieron tener un papel en la acumulación y modificación de los restos recuperados (Cruz, 2000b). En el sitio CCP 5, se determinaron huesos de un cánido –posiblemente zorro extinto (Dusicyon avus)– y de puma, además de la presencia de marcas atri-buibles a carnívoros en 3,4-6,6% de los especímenes de artiodáctilos identi-ficados. En CCP 7, ADG y AGV también se registraron restos de carnívoros entre los materiales en estratigraf ía. Estos restos incluyen tanto huesos como excrementos, y posiblemente corresponden a cánidos (Cruz, 2007a). Ade-más, en la actualidad, las aves rapaces ocupan varios de estos abrigos en los que se emplazan sitios arqueológicos (CCP 5, CCP 7 y ADG), y su presencia en las secuencias arqueológicas ha sido demostrada a partir de egagrópilas y plumas, de restos de sus nidos y de sus propios huesos. En síntesis, la evi-dencia indica que los carnívoros (tanto mamíferos como aves) utilizaron los abrigos rocosos durante el período de formación de los depósitos arqueoló-gicos. El análisis de las escasas modificaciones culturales en los restos de aves y la información contextual de los mismos en los conjuntos arqueológicos permitieron determinar que los dos actores más importantes en la conforma-ción del conjunto avifaunístico son los carnívoros y los humanos. Tanto unos como otros contribuyeron a la formación del mismo, aportando y modifi-cando los huesos de aves, lo cual fue tenido en cuenta en las interpretaciones arqueológicas. En síntesis, este caso mostró la necesidad de evaluar todos los posibles actores tafonómicos involucrados en la formación de los conjuntos óseos, para ajustar las interpretaciones y corroborar adecuadamente las hipó-tesis acerca de la forma en que los humanos predaron las aves en este sector de la cordillera de los Andes (Cruz, 2007a).
El segundo caso muestra los beneficios de comparar la representación natural de partes esqueletarias observada mediante estudios tafonómicos de huesos actuales de cormoranes (Phalacrocorax sp.) con la registrada en depó-sitos arqueológicos que incluyen restos de estas aves (Cruz, 2009b). A partir

A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
1 6 6
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.147-174
de las observaciones actuales, se determinó que el perfil de representación anatómica de los cormoranes se destaca por el predominio de elementos de las extremidades posteriores (mayoritariamente, fémures y tibiatarsos) y ante-riores (especialmente, húmeros). Esta diferencia con el perfil anatómico de aves voladoras (ver “Representación de partes esqueletarias”) se explica por la carencia de neumatización en muchos de los huesos de cormoranes y el papel de las extremidades posteriores de estas aves en la locomoción bajo el agua, por lo que sus huesos son más resistentes a los procesos tafonómicos que los correspondientes a los de otras aves voladoras (Cruz, 2009b). El análisis tafo-nómico estuvo destinado, entre otros temas, a comparar esta representación natural con la determinada en varios conjuntos zooarqueológicos que tienen una alta representación de los cormoranes. Tres de estos conjuntos provie-nen de Patagonia: Cabo Blanco 1 (Moreno et al., 1998), Punta Baja (Lefèvre, 1989) y Túnel VII (Mameli, 2002); el restante, de la costa peruana: Quebrada Tacahuay (de France, 2005). La única similitud detectada en la representación anatómica de todos los conjuntos (los arqueológicos y el natural) fue que la cintura pélvica es la unidad menos representada. El perfil de Punta Baja se asemeja en líneas generales al natural, pero con una alta representación del esqueleto axial que no posee el conjunto natural (Cruz, 2009a). Los otros con-juntos tienen representaciones anatómicas muy diferentes a las registradas a partir de los estudios tafonómicos. Lo más interesante de esta comparación es que no fue posible aislar un patrón común a todos los depósitos arqueológicos. Lo mismo sucede con otros conjuntos de Patagonia, como los analizados por Lefèvre (1992, 1997) y Savanti (1994). Esto no implica que el patrón detectado en cada uno de los depósitos no sea el resultado de la acción humana, lo cual debería determinarse en cada caso mediante un análisis tafonómico exhaus-tivo. Lo que estas diferencias señalan es que es posible una gran variabilidad en la representación de partes esqueletarias de cormoranes en los conjuntos zooarqueológicos, relacionada con la diversidad de actividades humanas que pueden involucrar a estas aves. Muchas de estas actividades no dejan una firma específica en los registros óseos y pueden asemejarse al perfil natural observado (tal como sucede con el conjunto de Punta Baja). Dada esta ambi-güedad, el análisis tafonómico permitió concluir que los patrones de repre-sentación de partes esqueletarias no pueden considerarse un indicador claro de los actores y procesos involucrados, por lo que es poco aconsejable que se efectúen afirmaciones sobre la conducta humana utilizando exclusivamente estos perfiles anatómicos. Sin embargo, la representación de elementos cons-tituye una de las variables que permiten discutir la historia de un conjunto óseo (Behrensmeyer, 1991), y su análisis en el marco de las investigaciones

1 6 7
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.147-174
T A F O N O M Í A D E H U E S O S D E A V E S | I S A B E L C R U Z
zooarqueológicas permitirá efectuar estudios tafonómicos más precisos en cuanto a los procesos involucrados.
Las investigaciones futurasLas investigaciones tafonómicas efectuadas hasta el momento son un primer paso para comprender las particularidades del proceso de reciclado de huesos de aves en el sur del Neotrópico. Son varias las líneas de investigación sobre tafonomía de los restos de aves que quedan abiertas y que deberán desarrollarse para completar los resultados que presentamos aquí.
En primer lugar, es necesario profundizar las investigaciones acerca de la relación entre morfología esqueletaria y resistencia a los procesos tafonómicos, especialmente entre diferentes especies de aves voladoras. Esta categoría abarca un amplio rango de tamaños corporales y de variación anatómica, dentro del marco general del esqueleto básico de las aves. La anatomía de muchas de estas aves está también influenciada por aspectos independientes de la locomoción, e incluso por modos de locomoción alternativos. En Patagonia se ha avanzado en este aspecto, iniciándose investigaciones en colonias de nidificación de cor-moranes (Phalacrocoracidae). Como se mencionó anteriormente, a partir de estos estudios, se pudo establecer que la representación de partes esqueletarias de estas aves, que vuelan y bucean, se asemeja más a la de los Spheniscidae que a la de otras aves voladoras (Cruz, 2009b). En el futuro, las investigaciones deberían explorar éste y otros aspectos en los restos de aves con diferencias anatómicas y tamaños corporales marcados.
En segundo lugar, los estudios tafonómicos de aves deben enmarcarse en investigaciones más amplias, destinadas a establecer el impacto que tienen diversos actores y procesos tafonómicos sobre los restos de distintos vertebrados en un mismo contexto ambiental. Esto permitirá evaluar el rol de las diferencias anatómicas más amplias en la preservación de restos de distintos organismos y puede constituir un instrumento adecuado para el análisis de conjuntos zooar-queológicos, generalmente conformados por restos de diferentes taxones.
Además, algunos aspectos requieren ser tratados mediante observacio-nes controladas. Entre ellos, se destacan la secuencia de desarticulación y el impacto de la meteorización. Estos temas han sido tratados por medio de las observaciones efectuadas, pero la rápida destrucción de los restos de aves en contextos naturales hace necesario que se implementen metodologías especí-ficas que permitan un control más estricto de las variables que harán posible comprenderlos más acabadamente.
Por último, sería interesante contrastar los principios derivados de estas investigaciones tafonómicas en una escala más amplia, que permita aumentar

A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
1 6 8
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.147-174
los contextos ambientales en los que se depositan y modifican los restos de aves. La dinámica tafonómica de ambientes más cálidos, con otras relacio-nes ecológicas o con mayor diversidad de aves, seguramente es diferente a la registrada en el extremo sur del Neotrópico. Esta comparación más amplia permitirá una comprensión más acabada del comportamiento tafonómico de los restos de aves y será de utilidad para interpretar depósitos en ésta y otras regiones biogeográficas.
Consideraciones finalesTodo el conocimiento acerca del pasado está fuertemente mediado por la comprensión de los factores tafonómicos que han actuado para conformar los registros fósiles que son la base para las interpretaciones científicas (Behrens-meyer et al., 2000). El entendimiento amplio de la forma en que operan los mecanismos que promueven la destrucción y/o preservación de restos orgáni-cos permite generar los modelos e hipótesis que constituyen el corpus teórico a partir del cual se pueden interpretar los casos específicos.
El principal aporte de las observaciones tafonómicas en Patagonia, o en cualquier otra región del Neotrópico, radica en que pueden dar cuenta de con-diciones diferentes a las consideradas por los modelos y principios que actual-mente constituyen la base de la disciplina, formulados a partir de observacio-nes en otros lugares del mundo. Cuanto mayor sea la cantidad de ecosistemas y ambientes en los que se evalúen los modelos tafonómicos, mayor podrá ser su grado de generalidad, debido a que habrán sido puestos a prueba bajo gran variedad de condiciones. Al mismo tiempo, las investigaciones en diferentes regiones del mundo permitirán desarrollar modelos más específicos, en virtud de que los ya existentes podrán ser ajustados para dar cuenta de situaciones particulares. En este sentido, las observaciones patagónicas permitieron, por un lado, mostrar la persistencia de los patrones regulares propios de los proce-sos considerados, que se expresan independientemente de las distintas condi-ciones ambientales. Por otro, posibilitaron establecer cuáles son las particula-ridades propias de la región y, de esta manera, comprender cuáles son algunos de los aspectos que pueden incidir en las variaciones.
Como señaló Binford (1981), conocer la dinámica de las poblaciones humanas del pasado es imposible sin comprender el registro arqueológico. La tafonomía, como parte de las investigaciones arqueológicas, brinda la posi-bilidad de interpretar adecuadamente una porción del mismo –el registro arqueofaunístico–, al tiempo que permite abordar aspectos de los contextos en los cuales se generaron los depósitos. Dados la madurez de la disciplina y el desarrollo que la misma ha alcanzado en todo el mundo, esperamos que en los

1 6 9
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.147-174
T A F O N O M Í A D E H U E S O S D E A V E S | I S A B E L C R U Z
próximos años la tafonomía pueda incorporarse definitivamente en las investi-gaciones zooarqueológicas del Neotrópico.
AgradecimientosEste trabajo es un resultado de los proyectos UNPA 29A/260-1 y PIP/CONI-CET 112-200801-00996. .

A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
1 7 0
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.147-174
R e f e R e n c i a s
Andrews, Peter 1990.Owls, Caves and Fossils.Chicago,TheUniversityofChicagoPress.
Behrensmeyer, Anna K. 1978.“TaphonomicandEcologicalInformationfromBoneWeathering”,Paleobiology4,pp.150-162.1991.“TerrestrialVertebrateAccumulations”,enTaphonomy: Releasing the Data Locked in the Fossil
Record,eds.P.A.AllisonyD.E.G.Briggs,pp.291-335.NuevaYork,PlenumPress.1993“TheBonesofAmboseli.TheTaphonomicRecordofEcologicalChangeinAmboseliPark,Kenya”,
enNational Geographic Research & Exploration9,pp.402-421.
Behrensmeyer, Anna K. y Dorothy E. Dechant-Boaz 1980.“TheRecentBonesofAmboseliPark,Kenya,inRelationtoEastAfricanPaleoecology”,enFossils
in the Making. Vertebrate Taphonomy and Paleoecology,eds.AnnaK.BehrensmeyeryAndrewP.Hill,pp.72-92.Chicago,UniversityofChicagoPress.
Behrensmeyer, Anna K., Susan M. Kidwell y Roger Gastaldo 2000.“TaphonomyandPaleobiology”,enDeep Time. Paleobiology’s Perspective,Suplementode
Paleobiology,eds.DouglasH.ErwinyScottL.Wing, 26,pp.103-147.
Behrensmeyer, Anna K., C. Tristan Stayton y Ralph E. Chapman 2003.“TaphonomyandEcologyofModernAvifaunalRemainsfromAmboseliPark,Kenya”,
Paleobiology29,pp.52-70.
Belardi, Juan Bautista1999“Haychoiquesenlaterraza.Informacióntafonómicayprimerasimplicacionesarqueofaunísticas
paraPatagonia”,enArqueología. Revista de la Sección Prehistoria 9,pp.163-185.
Bellairs, A . D’A. y C. R. Jenkin 1960.“TheSkeletonofBirds”,en Biology and Comparative Physiology of Birds,ed.A.J.Marshall,Vol.1,
pp.241-300.NuevaYork,J.Wiley&Sons.
Bickart, Karl J. 1984.“AFieldExperimentinAvianTaphonomy”,Journal of Vertebrate Paleontology4,pp.525-535.
Binford, Lewis R. 1981.Bones. Ancient Men and Modern Myths.NuevaYork,AcademicPress.
Blumenschine, Robert J. 1986.Early Hominid Scavenging Opportunities. Implications of Carcass Availability in the Serengeti
and Ngorongoro Ecosystems.Oxford,BARInternationalSeries283.1987.“CharacteristicsofanEarlyHominidScavengingNiche”,Current Anthropology28,pp.383-407.1989.“ALandscapeTaphonomicModeloftheScaleofPrehistoricScavengingOpportunities”,Journal of
Human Evolution18,pp.345-371.1998.“CommentstoMusterianLarge-MammalRemainsfromKobehCave:BehavioralImplications(C.
MareanyS.Y.Kim)”,Current Anthropology39(Suplemento),pp.79-113.
Borrero, Luis A.1990.“TaphonomyofGuanacoBonesinTierradelFuego”,Quaternary Research34,pp.361-371.2000.“TenYearsAfter:esquemaparaunatafonomíaregionaldelaPatagoniaMeridionalynortede
TierradelFuego”,enDesde el país de los gigantes. Perspectivas arqueológicas en Patagonia,tomoI,pp.183-193.RíoGallegos,UniversidadNacionaldelaPatagoniaAustral.
2001.“RegionalTaphonomy:BackgroundNoiseandtheIntegrityoftheArchaeologicalRecord”,enEthnoarchaeology of Andean South America. Contributions to Archaeological Method and Theory,ed.LawrenceA.Kuznar,pp.243-254.AnnArbor,Michigan.InternationalMonographsinPrehistory.EthnoarchaeologicalSeries4.

1 7 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.147-174
T A F O N O M Í A D E H U E S O S D E A V E S | I S A B E L C R U Z
2007.“LongitudinalTaphonomicStudiesinTierradelFuego,Argentina”,enTaphonomy and Archaeozoology in Argentina,eds.MaríaA.Gutiérrez,GustavoBarrientos,MónicaSalemme,LauraMiottiyGuillermoL.MengoniGoñalons,pp.219-233.Oxford,BAR(BritishArchaeologicalReports)InternationalSeriesS1601.
Bovy, Kristine M. 2002.“DifferentialAvianSkeletalPartDistribution:ExplainingtheAbundanceofWings”,Journal of
Archaeological Science29,pp.965-978.
Brain, Charles Kimberlin 1981.The Hunters or the Hunted? An Introduction to African Cave Taphonomy.Chicago,TheUniversity
ofChicagoPress.
Bunn, Henry T., Ellen M. Kroll y Lawrence E. Bartram 1991.“BoneDistributiononaModernEastAfricanLandscapeanditsArchaeologicalImplications”,
enCultural Beginnings. Approaches to Understanding Early Hominid Life-ways in the African Savanna,pp.33-54.Bonn,Dr.RudolfHabeltGMBA.
Corona Martínez, Eduardo2001.“Larelaciónhombre-aveenelsigloXVI.CrónicasyarqueozoologíaenelcentrodeMéxico”,
Arqueología25,pp.109-120.2002. Las aves en la historia natural novohispana.México,InstitutoNacionaldeAntropología
eHistoria.
Cruz, Isabel 1999.“PingüinosdeCaboVírgenes(SantaCruz).Aspectostafonómicoseimplicacionesarqueológicas”,
enActas del XIII Congreso Nacional de Arqueología Argentina,tomo4,pp.95-108.Córdoba,UniversidadNacionaldeCórdoba.
2000a.“Líneastafonómicasyecológicasparaevaluarlaexplotaciónprehistóricadeavesacuáticasenlazonacordillerana(Prov.deSantaCruz)”,en Desde el país de los gigantes. Perspectivas arqueológicas en Patagonia,tomoI,pp.202-217.RíoGallegos,UniversidadNacionaldelaPatagoniaAustral.
2000b.“LosrestosdeavesdelossitiosarqueológicosdelParqueNacionalPeritoMoreno(SantaCruz,Argentina)”,en Anales del Instituto de la Patagonia,SerieCienciasSociales(Chile),28,pp.305-313.
2004.“TafonomíadehuesosdeavesenPuntaMedanosa(Depto.PuertoDeseado,SantaCruz,Argentina)”,enContra viento y marea. Arqueología de la Patagonia,eds.MaríaTeresaCivalero,PabloMarceloFernándezyAnaGabrielaGuraieb,pp.455-468.BuenosAires,InstitutoNacionaldeAntropologíayPensamientoLatinoamericanoySociedadArgentinadeAntropología.
2005.“Larepresentacióndepartesesqueléticasdeaves.Patronesnaturaleseinterpretaciónarqueológica”,Archaeofauna14,pp.69-81.
2006.“Loshuesosdepingüinos(Spheniscidae)delossitiosdeCaboBlanco(SantaCruz,PatagoniaArgentina).Análisistafonómicoyperspectivasarqueológicas”,Intersecciones en Antropología7,pp.15-26.
2007a.“LaexplotacióndelasavesenlazonacordilleranadelsurdePatagonia(SantaCruz,Argentina)”,Werken 11,pp.107-126.
2007b.“AvianTaphonomy:ObservationsatTwoMagellanicPenguin(Spheniscus magellanicus)BreedingColoniesandTheirImplicationsfortheFossilRecord”,Journal of Archaeological Science 34,pp.1252-1261.
2007c.“TheRecentBonesoftheRíoGallegosBasin(SantaCruz,Argentina)andTheirPreservationPotential”,enTaphonomy and Archaeozoology in Argentina,eds.MaríaA.Gutiérrez,LauraMiotti,GustavoBarrientos,GuillermoL.MengoniGoñalonsyMónicaSalemme,pp.161-170.Oxford,BAR(BritishArchaeologicalReports)InternationalSeriesS1601.
2008.“AvianandMammalianBoneTaphonomyinSouthernContinentalPatagonia.AComparativeApproach”,Quaternary International180,pp.30-37.

A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
1 7 2
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.147-174
2009a.“Tafonomíaenescalasespacialesamplias:elregistroóseodelasavesenelsurdePatagonia”,enTemas de arqueología 2: estudios tafonómicos y zooarqueológicos,eds.AlejandroAcosta,DanielLoponteyLeonardoMucciolo,pp.15-34.BuenosAires,InstitutoNacionaldeAntropologíayPensamientoLatinoamericano.
2009b.“Tafonomíadehuesosdecormoranesenlacostapatagónica.Primerosresultados”,enArqueología de la Patagonia. Una mirada desde el último confín,pp.729-741.Ushuaia(Argentina),EditorialUtopía.
Cruz, Isabel y Dolores C. Elkin2003.“StructuralBoneDensityoftheLesserRhea(Pterocnemia pennata)(Aves:Rheidae).Taphonomic
andArchaeologicalImplications”,Journal of Archaeological Science 30,pp.37-44.
Cruz, Isabel y Pablo M. Fernández2004.“Comprendiendoelpasado:avancesentafonomíadeRheidos”,enActas del 1º Congreso
Latinoamericano sobre Conservación y Cría Comercial de Ñandúes,CD-ROM.BuenosAires,INTA-Rheacultura.com.ar-SecretaríadeAgricultura,Ganadería,PescayAlimentacióndelaRepúblicaArgentina.
Davis, Paul G. y Derek E. G. Briggs 1998.“TheImpactofDecayandDisarticulationonthePreservationofFossilBirds”,Palaios13,pp.3-13.
deFrance, Susan2005.“LatePleistoceneMarineBirdsfromSouthernPeru:DistinguishingHumanCapturefromElNiño-
inducedWindfall”,Journal of Archaeological Science32,pp.1131-1146.
Emslie, Steven D. 1995.“AgeandTaphonomyofAbandonedPenguinRookeriesintheAntarcticPeninsulaRegion”,Polar
Record31,pp.409-418.
Ericsson, Per G.1987.“InterpretationsofArchaeologicalBirdRemains:ATaphonomicApproach”,Journal of
Archaeological Science14,pp.65-75.
Feduccia, Alan1992“Osteologíadelasaves”,enAnatomía de los animales domésticos,eds.SamuelSisson,J.B.
GrossmanyRobertGetti,tomoI,pp.1960-1972,México,EditorialSalvat.
Fernández, Pablo Marcelo, Isabel Cruz y Dolores C. Elkin2001“DensidadmineralóseadePterocnemia pennata(Aves:Rheidae).Unaherramientaparaevaluar
frecuenciasanatómicasensitiosarqueológicos”,Relaciones, Revista de la Sociedad Argentina de Antropología 26,pp.243-260.
Fernández, Pablo M., Isabel Cruz y Ana Forlano2010.“Sitio37:unamadrigueradecarnívoroenelNortedelaPatagoniaAndina(Cholila,Provinciade
Chubut,Argentina)”,enZooarqueología a principios del siglo XXI: aportes teóricos, metodológicos y casos de estudio,eds.MaríaA.Gutiérrez,MarianaDeNigris,PabloM.Fernández,MiguelGiardina,AdolfoF.Gil,AndrésIzeta,GustavoNemeyHugoD.Yacobaccio,pp.409-417.BuenosAires,EdicionesdelEspinillo.
Fernández, Pablo M. e Isabel Cruz2010.“TheFox,theHare,andtheSheep.BoneModificationsinaModernCarnivoreDenatthe
DeciduousAndeanForest(Cholila,Argentina)”,pósterpresentadoenlaReuniónInternacionaldelICAZ(InternationalCouncilofArchaeozoology).París(Francia).
Giardina, Miguel A.2006.“AnatomíaeconómicadeRheidae”,Intersecciones en Antropología7,pp.263-276.

1 7 3
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.147-174
T A F O N O M Í A D E H U E S O S D E A V E S | I S A B E L C R U Z
Gif ford, Dianne P. 1981.“TaphonomyandPaleoecology:ACriticalReviewofArchaeology’sSisterDisciplines”,enAdvances
in Archaeological Method and Theory,ed.MichaelB.Schiffer,4,pp.365-438.NuevaYork,AcademicPress.
Gilbert, B. Miles., Larry D. Martin y Howard G. Savage 1981.Avian Osteology.Laramie,ModernPrintingCo.
Higgins, Jude 1999.“Túnel:ACaseStudyofAvianZooarchaeologyandTaphonomy”,Journal of Archaeological
Science26,pp.1449-1447.
Kidwell, Susan y Anna K. Behrensmeyer1988“Overview:EcologicalandEvolutionaryImplicationsofTaphonomicProcesses”,en
Palaeogeography, Palaeoclimatology and Palaeobiology 63, pp. 1-13.
Lefèvre, Christine 1989.“Lesoiseaux”,enEthno-archéologie dans les archipels de Patagonie: les nomades marins de
Punta Baja,ed.DominiqueLegoupil,pp.99-113.París,EditionsRecherchesurlesCivilisations,MémoireNo.84.
1992.“PuntaMaría2.Losrestosdeaves”,Palimpsesto, Revista de Arqueología2,pp.71-98.1997.“SeaBirdFowlinginSouthernPatagonia:AContributiontoUnderstandingtheNomadicRoundof
theCanoerosIndians”,International Journal of Osteoarchaeology7,pp.260-270.
Livingston, Stephanie D. 1989.“TheTaphonomicInterpretationofAvianSkeletalPartFrecuencies”,Journal of Archaeological
Science16,pp.537-547.
Lyman, Richard L. 1994.Vertebrate Taphonomy.Cambridge,CambridgeUniverstiyPress.
Mameli, Laura2002.“BirdManagementinAmerica’sExtremeSouthduringthe19thCentury”,enActa zoologica
cracoviensia45,pp.151-165.
Marean, Curtis W.1995.“OfTaphonomyandZooarchaeology”,Evolutionary Anthropology4,pp.64-72.
Martín, Fabiana M. 1998.“Madrigueras,dormiderosyletrinas:aproximaciónalatafonomíadezorros”,enArqueología
de la Patagonia Meridional (Proyecto Magallania),comp.LuisAlbertoBorrero,pp.73-96.ConcepcióndelUruguay,EdicionesBúsquedadeAyllu.
Martínez, Ismael, Isabel Cartajena y Patricio López M.2009.“PresenceofLate-PleistoceneRheidaeintheOasisofCalama(SecondRegion,Chile)”,Current
Research in the Pleistocene27,pp.33-35.
Morales Muñiz, Arturo 1993.“Ornithoarchaeology:TheVariousAspectsoftheClassificationofBirdRemainsfrom
ArchaeologicalSites”,Archaeofauna2,pp.1-13.
Moreno, Eduardo, Alicia Castro, Karina Martinelli y Andrés Abello 1998.“ElmaterialfaunísticodeCaboBlanco1”,Anales del Instituto de la Patagonia,26,pp.169-180.
Muñoz, A. Sebastián y Florencia Savanti1998“ObservacionestafonómicassobrerestosavifaunísticosdelacostanorestedeTierradelFuego”,
enActas y Memorias del XI Congreso Nacional de Arqueología Argentina (8ª Parte), Revista del Museo de Historia Natural de San RafaelTomoXX1/2:107-121.

A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
1 7 4
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.147-174
Oliver, James S. y Russell W. Graham 1994.“ACatastrophicKillofIce-trapedCoots:Time-AveragedVersusScavenger-specificDisarticulation
Patterns”,Paleobiology20,pp.229-244.
Pickering, Travis, Kathy Schick y Nicholas Toth (eds.)2007.Breathing Life into Fossils: Taphonomic Studies in Honor of C. K. (Bob) Brain.Gosport,StoneAge
InstitutePublicationSeriesNumber2.
Savanti, Florencia1994.Las aves en la dieta de los cazadores-recolectores terrestres de la costa fueguina.Temasde
Arqueología.BuenosAires,CONICET/PREP.
Sept, Jean M. 1994.“BoneDistributioninaSemi-aridRiverineHabitatinEasternZaire:Implicationsforthe
InterpretationofFaunalAssemblagesatEarlyArchaeologicalSites”,Journal of Archaeological Science21,pp.217-235.
Tappen, Martha 1995.“SavannaEcologyandNaturalBoneDeposition.ImplicationsforEarlyHominidSiteFormation,
Hunting,andScavenging”,Current Anthropology 36,pp.223-260.
Tívoli, Angélica M. 2010.“ExploitationofBirdResourcesamongPrehistoricSea-nomadSocietiesoftheBeagleChannel
Region,SouthernSouthAmerica”,Before Farming2,artículo3.
Trapani, Joshua 1998.“HydrodynamicSortingofAvianSkeletalRemains”,Journal of Archaeological Science25,
pp.477-487.

1 7 5
PR I M ER R EGISTRO DE PER RO DOM ÉSTICO PR EH ISPÁ N ICO (C A N IS FA M I L I A R IS) EN TR E LOS GRU POS CA Z A DOR ES R ECOLECTOR ES DEL H U M EDA L DE PA R A NÁ I N F ER IOR (A RGEN TI NA)
ale jandro acosTa*
[email protected] Instituto Nacional de Antropología y Pensamiento Latinoamericano, Argentina
DANieL LopoNte**
[email protected] Instituto Nacional de Antropología y Pensamiento Latinoamericano, Argentina
césar García esponda***
[email protected] de Ciencias Naturales y Museo, Universidad Nacional de La Plata, Argentina
R e s u m e n Se analiza el primer registro confiable de un
perro prehispánico (Canis familiaris) recuperado en un sitio
de cazadores recolectores del humedal del Paraná inferior
(Argentina). Se discuten distintos aspectos relacionados con su
origen, antigüedad y tipo de interacción que habrían establecido
con las poblaciones humanas en el marco de la escasa
información de la que actualmente disponemos acerca de su
presencia entre los grupos cazadores-recolectores del extremo
sur de Sudamérica.
P A L A B R A S C L A V E :
Perro prehispánico, cazadores-recolectores, humedal del Paraná
inferior.
Artículorecibido:1dejuniode2011|aceptado:12deoctubrede2011|modificado:5denoviembrede2011
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,294pp.ISSN1900-5407,pp.175-199
* DoctorenCienciasNaturales,FacultaddeCienciasNaturalesyMuseo,UniversidadNacionaldeLaPlata,Argentina.**DoctorenCienciasNaturales,FacultaddeCienciasNaturalesyMuseo,UniversidadNacionaldeLaPlata,Argentina.***Doctor en CienciasNaturales, Facultad de CienciasNaturales yMuseo,UniversidadNacional de La Plata,
Argentina.

1 7 6
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.175-199
first registry of the pre-hispaniC domestiC dog (Canis familiaris) among gatherer-hunter groups in lower parana (argentina)
ABSTR AC T The first reliable record of the
dog (Canis familiaris) recovered in a hunter
gatherer site from the lower Paraná wetlands
(Argentina) is analyzed in this paper. We
discuss several aspects related to the
origin, chronology and types of interactions
established between dogs and aboriginal
groups in the context of scarce information
currently available about their presence
among hunter gatherer groups of Southern
South America.
primeiro registro do Cão domestiCo pré-hispâniCo (Canis familiaris) entre os grupos Caçadores reColhedores da Zona úmida de paraná inferior (argentina)
RESUMO Analisa-se o primeiro registro
confiável de um cão pré-hispânico (Canis
familiaris) recuperado num lugar de
caçadores recolhedores da zona úmida do
Paraná inferior (Argentina). Discutem-se
distintos aspectos relacionados com sua
origem, antiguidade e tipo de interação
que teriam estabelecido com as populações
humanas na moldura da escassa informação
que temos atualmente sobre sua presença
entre os grupos caçadores–recolhedores do
extremo sul de América do Sul.
P A L A V R A S C H A V E :
Cão pré-hispânico, caçadores-recolhedores,
zona úmida do Paraná inferior.
K E Y W O R D S :
Prehispanic Dog, Hunter Gatherer, Lower
Paraná Wetland.

1 7 7
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.175-199
PR I M ER R EGISTRO DE PER RO DOM ÉSTICO PR EH ISPÁ N ICO (C A N IS FA M I L I A R IS) EN TR E LOS GRU POS CA Z A DOR ES R ECOLECTOR ES DEL H U M EDA L DE PA R A NÁ I N F ER IOR (A RGEN TI NA)
a l e j a n d r o a c o s T a
D A N i e L L o p o N t e
c é s a r G a r c í a e s p o n d a
IntroducciónEn este trabajo presentamos y discutimos el primer registro de perro (Canis familiaris) prehispánico recuperado en un contexto arqueológico de cazadores-recolectores que habitaron el humedal del Paraná inferior (Argentina) durante el Holoceno tardío. Para ello se contextualiza la problemática abordada en función de la información de la que actualmente se dispone sobre la evolución y domesticación del perro en Euroasia y América. Posteriormente, se resumen los principales antecedentes y el estado actual del conocimiento sobre la presencia y origen de C. familiaris en el extremo sur de Sudamérica. Finalmente, analizamos y evaluamos nuestro caso de estudio a partir de otras evidencias disponibles, con el fin de discutir diferen-tes aspectos relacionados con el origen del perro, y las posibles interacciones que se habrían establecido entre estos últimos y las poblaciones cazadoras-recolectoras tanto en nuestra área de estudio como en el extremo sur de Sud-américa en general.
El estudio sobre el origen y evolución del perro ha sido un tema que ha despertado un gran interés entre los especialistas de diferentes ramas de la ciencia. Hoy sabemos que esta especie fue una de las primeras en ser domes-ticadas por los seres humanos y que se utilizó con fines diversos (Schwartz, 1997; Morey, 2006). Durante mucho tiempo se discutió acerca de sus posibles centros de domesticación y cuál había sido su antepasado más directo, situa-ción que implicaba establecer su potencial relación evolutiva con los cánidos salvajes tanto del Viejo como del Nuevo Mundo, tales como el coyote, el chacal,

1 7 8
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.175-199
distintas variedades de lobos, e incluso con algunos cánidos extintos de Suda-mérica (e.g., Allen, 1920; Cabrera, 1934). Si bien gran parte de estos esfuerzos se basaron en estudios comparativos relacionados con el análisis morfométrico de las distintas especies implicadas (fósiles y modernas; ver ejemplos en Morey, 1992), las investigaciones genéticas realizadas en los últimos quince años han aportado pruebas de sustancial importancia para comprender este fenómeno (ver Okumura et al., 1996; Tsuda et al., 1997; Vilá et al., 1997; Leonard et al., 2002; Ostrander y Wayne, 2005; entre otros). En la actualidad, casi todos los investigadores aceptan que el perro habría derivado de la domesticación de Canis lupus (lobo gris) (ver, sin embargo, Koler-Matznick, 2002).
Vilá et al. (1997) realizaron uno de los primeros estudios en donde se analizó y comparó el ADN mitocondrial de distintas poblaciones de perros (N = 67) y de lobos (N = 162) procedentes de Europa, Asia y América del Norte. A través de este trabajo se confirmó la estrecha afinidad genética que existe entre los perros y los lobos grises (euroasiáticos), siendo tan sólo del 2 % la diferencia estimada entre ambas especies. Asimismo, el análisis filogenético efectuado a partir de las secuencias de ADN mitocondrial permitió reconocer cuatro gru-pos o clados (I a IV), hecho que indicaría que los lobos fueron domesticados en distintas regiones y/o momentos (sin embargo, ver también Savolainen et al., 2002); es muy probable que luego de su domesticación se hayan producido diversos episodios de cruzamiento o procesos de hibridización entre perros y lobos (Andersone et al., 2002; Wayne et al., 2006).
Las variaciones de ADN mitocondrial identificadas dentro del clado I indican que la divergencia genética en este clado es mucho mayor que en los otros tres. Al respecto, Vilá et al. (1997) calcularon que la divergencia entre los perros y los lobos grises habría acontecido alrededor de 135.000 años AP, mien-tras que Okumura et al. (1996), también sobre la base de información molecu-lar, sugirieron una antigüedad de 76.000 a 121.000 años AP (para una discu-sión sobre la divergencia de ambas especies, ver Wayne y Vilá, 2003). De todos modos, los cambios morfológicos que diferencian a C. familiaris de C. lupus recién se presentan en los últimos 11.000 a 17.000 años AP, rango cronológico dentro del que se encuentran los primeros perros domésticos hallados en dis-tintos sitios arqueológicos del continente euroasiático (Olsen, 1985; Sablin y Khlopachev, 2002; Morey, 2006; sin embargo, véase Germonpré et al., 2009, quienes reportan un ejemplar de perro paleolítico proveniente de la cueva de Goyet, en Bélgica, con una antigüedad de 31.700 años AP). En relación con este aspecto, se ha considerado que la selección fenotípica de determinados rasgos morfológicos no tendría una relación causal directa con los cambios observa-dos en las secuencias del ADN mitocondrial (Vilá et al., 1997; Wayne et al.,

1 7 9
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.175-199
P R I M E R R E G I S T R O D E P E R R O D O M É S T I C O | A L E J A N D R O A C O S T A , D A N I E L L O P O N T E Y C É S A R G A R C Í A
2006). Es probable que, durante un largo tiempo, las primeras poblaciones de C. familiaris (o protoperros) no presentaran significativas diferencias morfoló-gicas respecto de sus ancestros. Distintos autores han argumentado que ciertos rasgos (fenotípicos y/o conductuales) que diferencian a los perros de los lobos aparecieron más tardíamente, debido a las presiones selectivas ejercidas por los grupos humanos, en la medida en que se intensificó la interacción entre las poblaciones de H. sapiens y C. familiaris. Esta situación estaría estrechamente relacionada con la reducción de la movilidad y con la estabilidad de las ocu-paciones de las sociedades humanas que habitaron Euroasia hace unos 15.000 años AP (Wayne et al., 2006; Kubinyi et al., 2007).
En cuanto al Nuevo Mundo, los perros domésticos más antiguos provie-nen de América del Norte y poseen una antigüedad de entre 10.000 y 8.500 años AP, existiendo numerosos registros a partir de los 7.000 años AP en ade-lante (Grayson, 1988; Morey, 1992; Morey y Wiant, 1992). En el centro y sur del continente los registros tempranos han sido, en general, más escasos y/o controvertidos, y aunque existen evidencias entre los 7.500 y 4.500 años AP, la mayor parte de los hallazgos se ubican dentro de los 3.500 a 1.000 años AP (Wing, 1989; Schwartz, 1997; Valadez et al., 2003). Uno de los principales pro-blemas que ha surgido en torno a los perros americanos ha sido el posible ori-gen independiente de algunas razas a partir de las poblaciones de lobos origina-rias del norte del continente (ver, por ejemplo, Koop et al., 2000). Leonard et al. (2002) analizaron el ADN mitocondrial de perros prehispánicos en 37 muestras arqueológicas procedentes de México, Perú y Bolivia, más 11 de Alaska. Los resultados obtenidos fueron comparados con las secuencias genéticas de otros perros y de poblaciones de lobos (N = 259) de distintas regiones del mundo. A través de este estudio los autores lograron establecer que los haplotipos de los perros americanos se agrupaban en dos de los clados (I y IV) identificados por Vilá et al. (1997) y que ninguno de ellos tenía relación directa con los de los lobos (americanos), situación que indujo a plantear que los perros (ameri-canos) serían de origen euroasiático. A su vez, en función de las variaciones registradas dentro de los clados (I y IV), y a partir del reconocimiento de una serie de nuevos haplotipos denominados como “grupo a”, sostuvieron que al menos cinco líneas de perros debieron ingresar al continente americano, las cuales habrían estado expuestas a diferentes grados de aislamiento (ver detalles en Leonard et al., 2002; Valadez et al., 2003).
Dado que el ejemplar de C. familiaris que aquí se estudia proviene del sur de Sudamérica, es conveniente realizar algunas apreciaciones sobre la presen-cia y distribución de los perros prehispánicos en el subcontinente. En términos generales, el registro de perros es cuantitativamente menor en relación con el que

1 8 0
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.175-199
se dispone para Mesoamérica y para América del Norte. Casi todos los hallaz-gos que dan cuenta de su existencia se distribuyen a lo largo del eje cordillerano de la región andina, habiéndose registrado en Venezuela, Ecuador, Perú, Bolivia, norte de Chile y noroeste de Argentina (N.O.A.). Es importante destacar que la mayoría de estas evidencias se encuentran asociadas a ocupaciones de socieda-des agroalfareras con diferentes y complejos niveles de organización socioeconó-mica (Cabrera, 1934; Gallardo, 1964-65; Zetti, 1973; Wing, 1989; Schwartz, 1997; Mendoza España y Valadez, 2006; entre otros). En cuanto a los grupos cazadores-recolectores del extremo sur de Sudamérica, la antigüedad e identificación de perros domésticos antes del contacto europeo han sido más bien controvertidas. En general, son muy pocos los casos que confirman su presencia prehispánica dentro de la vasta región que se extiende desde el paralelo 32º hacia el sur, que-dando involucrados prácticamente más del 50% de los territorios que correspon-den a Chile Argentina y Uruguay. Dentro de esta macrorregión, las determina-ciones más confiables de C. familiaris que hasta ahora se conocen provienen de Uruguay y Argentina, habiéndose identificado su presencia en tan sólo siete sitios arqueológicos, todos ellos cronológicamente situados en el Holoceno tardío (ver más abajo), incluido el ejemplar que aquí se estudia.
La existencia de perros prehispánicos entre los grupos cazadores-reco-lectores del extremo sur de Sudamérica ha sido recientemente confirmada. De todos modos, aún son escasos los registros confiables que se conocen y las regiones en donde se los ha identificado. Esto indicaría, aparentemente, que habrían existido en proporciones muy bajas, cuyas posibles causas más ade-lante trataremos de dilucidar. Más allá de su reciente confirmación, práctica-mente desde principios del siglo XX, ya sea mediante evidencias arqueofaunís-ticas y/o a partir de fuentes históricas, distintos autores han aportado diver-sas pruebas para evaluar si habían existido o no perros con anterioridad a la conquista europea. La mayor parte de esta discusión se centró en la región de Pampa y Patagonia (Roth, 1902; Cabrera, 1932 y 1934; Kriscautzky, 1975b, cit. en Tonni y Politis, 1981; Cardich et al., 1977; Clutton-Brock, 1977 y 1978; Caviglia, 1980 y 1985-86; Tonni y Politis, 1981).
La mayoría de los restos de cánidos que fueron determinados como C. familiaris provienen de la Patagonia meridional, más precisamente de ocupa-ciones arqueológicas datadas entre 10.000 y 7.500 años AP, aproximadamente. Éste es el caso de las cuevas Fell y Pali Aike (Clutton-Brock, 1977 y 1978; Bird, 1988), cueva Eberhard (Roth, 1902) (Magallanes, Chile) y la cueva 3 de Los Toldos (Santa Cruz, Argentina) (Cardich et al., 1977). Sobre la base de estos hallazgos y considerando su antigüedad, algunos autores llegaron a proponer como hipótesis, tanto para el área patagónica como para la pampeana, la pre-

1 8 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.175-199
P R I M E R R E G I S T R O D E P E R R O D O M É S T I C O | A L E J A N D R O A C O S T A , D A N I E L L O P O N T E Y C É S A R G A R C Í A
sencia y/o el mantenimiento del proceso de domesticación de C. familiaris por parte de los aborígenes prehispánicos (ver Tonni y Politis, 1981: 264). Cabe aclarar que, para la región pampeana, el único registro que se conocía era el cráneo de un cánido asociado a un entierro humano recuperado en un sitio del Holoceno tardío denominado Río Luján I (Partido de Campana, Buenos Aires). Dicho cráneo, en una primera instancia, fue asignado por Kriscautzky (1975) a Chrysocyon brachyurus (aguará guazú), aunque muy poco tiempo des-pués lo reasignó a Canis familiaris (Kriscautzky, 1975b, cit. en Tonni y Politis, 1981). De todas maneras, en un reciente trabajo se informó que el cráneo no correspondería a C. familiaris, ni a C. brachyurus, sino a Dusicyon avus (zorro extinto) (ver Prevosti et al., 2004: 33).
Una posición diferente a las mencionadas anteriormente fue la asumida por Caviglia (1980, 1985-86), quien cuestionó la existencia de perros en la Patagonia antes de la conquista europea. A partir de un análisis realizado con los restos de cánidos recuperados en la cueva de Las Buitreras (Santa Cruz, Argentina) y en otros sitios de la Patagonia meridional y de Tierra del Fuego, sostuvo que los especímenes que habían sido atribuidos a cánidos domésticos tempranos corresponderían en realidad a D. avus, incluidos los recuperados en la cueva 3 de Los Toldos y que fueran referidos a C. familiaris por Cardich et al. (1977). Esto último fue puesto en duda por Tonni y Politis (1981), quienes sostuvieron que su asignación a C. familiaris era la correcta (ver Tonni y Poli-tis, 1981: 262-263). Unos años después Caviglia (1985-86) planteó nuevamente la ausencia de perros prehispánicos en Patagonia y Tierra del Fuego. En este trabajo, realizó una revisión de los materiales de la cueva Fell e incorporó al análisis nuevos registros de cánidos, tanto tempranos como tardíos. A partir de una base mucho más sólida de datos efectuó un análisis cuantitativo de sus atri-butos (cráneo-dentarios) y logró establecer cuatro grupos de valores, los cuales se correspondían con las distintas especies de zorros (actuales y extinguidas) conocidas para la región de Fuego-Patagonia. También comparó el material estudiado con la información publicada sobre perros tempranos y otros cáni-dos del Cuaternario provenientes de América del Norte, estableciendo que no existían relaciones entre estos últimos y los zorros de Fuego-Patagonia. Ade-más, tuvo en cuenta algunas fuentes etnohistóricas de los siglos XVI y XVII, que también sugieren el origen poshispánico de C. familiaris en la región. Es importante mencionar que, en un reciente trabajo, Amorosi y Prevosti (2008) reexaminaron los restos provenientes de las cuevas Fell y Pali Aike que Clutton-Brock (1978; ver también Bird, 1988) asignara a C. familiaris, y determinaron que debían ser referidos a D. avus, tal como había originalmente propuesto Caviglia (1980 y 1985-86).

1 8 2
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.175-199
Desde mediados de los ochenta hasta la actualidad, a pesar del conside-rable aumento de las investigaciones efectuadas en los sitios del Pleistoceno-Holoceno inicial de las regiones Pampeana y Patagónica (ver Borrero, 2009), no se recuperaron restos que hayan sido de manera precisa o potencialmente refe-ridos a C. familiaris. Esta misma situación puede plantearse para el Holoceno medio. En cuanto al Holoceno tardío, en la Patagonia meridional y Tierra del Fuego, hasta ahora no se conocen evidencias que avalen claramente la presen-cia de perros prehispánicos, hecho que sustentaría las ideas de Caviglia (1985-86). En cambio, en la Patagonia septentrional y en el límite sur de la Región Pampeana fueron recientemente identificados en dos contextos arqueológicos de cazadores recolectores (Prates et al., 2010a y 2010b). Los restos provienen de los sitios Angostura 1 y Chenque 1. El primero se encuentra localizado en la provincia de Río Negro, en la costa norte cercana a la desembocadura del río homónimo. Los materiales referidos a C. familiaris son un canino y un molar inferior. El contexto de procedencia de estos restos fue datado en 938 ± 45 C14
años AP. El segundo es un cementerio aborigen ubicado en el suroeste de la provincia de La Pampa, en el Parque Nacional Lihué Calel. En este sitio se recu-peró un esqueleto completo (C. familiaris) enterrado deliberadamente junto a uno humano correspondiente a un individuo subadulto. Una fecha taxón rea-lizada sobre un fragmento de costilla del perro arrojó una antigüedad de 930 ± 30 años C14 AP (ver Prates et al., 2010a y 2010b).
Finalmente, en Uruguay desde los años noventa se ha mencionado la pre-sencia de perros asociados a enterratorios humanos en al menos cuatro estruc-turas tumulares situadas en el departamento de Rocha (Uruguay) (González, 1999). Los depósitos fueron generados por cazadores-recolectores del Holo-ceno tardío, conocidos en la literatura como “constructores de cerritos” (Pintos y Bracco, 1999; López Mazz, 2001). Cabe destacar que en su mayoría se trata de esqueletos que poseen un alto grado de completitud anatómica. Las deter-minaciones efectuadas por González (1999) incluyen distintos análisis morfo-métricos, además de comparaciones con otras razas de perros y de cánidos salvajes propios de la región. Lamentablemente, no se cuenta con fechas taxón. Sin embargo, en virtud de las cronologías radiocarbónicas obtenidas para los sitios, y en función de su procedencia estratigráfica y otros rasgos asociados, se estimó que los ejemplares, en su conjunto, tendrían algo más de 1.000 años AP (González, 1999: 67). Las edades radiocarbónicas obtenidas en Angostura 1 y Chenque 1, y probablemente las de Uruguay, se encuentran dentro de un rango temporal muy cercano al que posee el ejemplar recuperado en el sitio Cerro Lutz, y que seguidamente describiremos.

1 8 3
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.175-199
P R I M E R R E G I S T R O D E P E R R O D O M É S T I C O | A L E J A N D R O A C O S T A , D A N I E L L O P O N T E Y C É S A R G A R C Í A
El registro prehispánico de Canis familiaris en el sitio Cerro Lutz
El sitio Cerro Lutz se encuentra en el sector centro-oriental de la Región Pam-peana, en el departamento de Gualeguaychú (provincia de Entre Ríos); 33º 38’ 47" LS y 58º 36’ 20” LO (ver la figura 1), localizado en términos ecológi-cos dentro del humedal del Paraná inferior (HPI), correspondiente a la Eco-región denominada “Delta e islas del Paraná” (Burkart et al., 1999). El depósito arqueológico, generado durante el Holoceno tardío por grupos de cazadores-recolectores, se ubica sobre un albardón situado en la margen izquierda del arroyo Martínez, cercano a su intersección con el arroyo Sagastume Chico. La superficie de la ocupación arqueológica ha sido estimada en unos 24.000 m² aproximadamente. El total de superficie excavada hasta el momento es de 19 m², producto de cuatro Unidades de Excavación (UE) ubicadas en los sectores Norte y Sur del sitio. Los muestreos realizados han posibilitado obtener una abundante colección faunística y artefactual, además de una importante can-tidad de enterratorios humanos (primarios y secundarios), cuya distribución y disposición indican una variada y compleja gama de prácticas mortuorias. Los materiales arqueológicos se distribuyen dentro del Horizonte A, habién-dose detectado una mayor redundancia de ocupaciones en el sector Norte, en donde la capa arqueológica alcanza una potencia de entre 65 y 95 cm, situación que pudo verificarse en las UE 2 y 4, respectivamente. Los perfiles expuestos en estas dos unidades de excavación presentan una secuencia estra-tigráfica compleja, observándose una superposición de actividades separadas entre sí por pequeños hiatos sedimentarios y por concentraciones de valvas de Diplodon sp., características que implican la existencia de diferentes eventos de descarte. Los fechados radiocarbónicos efectuados (ver la tabla 1) indican que los sucesivos episodios de ocupación registrados en Cerro Lutz se habrían producido, en principio, dentro de un rango temporal que se ubica entre los 976 ± 42 y los 730 ± 70 años C14 AP.
El hallazgo de C. familiaris proviene de la UE-4 (sector Norte) ubicada a unos 10 metros hacia el norte de la UE-2. La UE-4 posee tan sólo 0,70 m² de superficie y presenta una potencia arqueológica de 95 cm. En esta UE se detectaron por lo menos tres estructuras de combustión separadas por densas y compactas capas de valvas de Diplodon sp. Se pudo observar que la distri-bución de estas acumulaciones en los perfiles era relativamente homogénea, aunque presentaban una concentración ligeramente mayor por debajo de las estructuras de combustión. Si bien en esta UE no se recuperaron restos huma-nos, se obtuvieron abundantes restos faunísticos y artefactuales. El ejemplar de C. familiaris corresponde a un individuo cuyo esqueleto se halla casi completo

1 8 4
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.175-199
y anatómicamente articulado. La mayoría de sus huesos (incluido el cráneo) se encuentran en un excelente estado de conservación, que no supera el estadio 1 de meteorización (sensu Behrensmeyer, 1978). Tampoco se observaron marcas naturales que sugieran que el cadáver sufrió algún tipo de alteración tafonómica significativa luego de su depositación. El esqueleto fue recuperado en el perfil Este (UE-4), en donde afloraba gran parte de la cintura pélvica y de los miem-bros posteriores, a los 93 cm de profundidad, entre la base del horizonte A y el techo del nivel A/C, hallándose literalmente sellado por una compacta capa de valvas de Diplodon sp. (ver la figura 2) (Acosta et al., 2010). La fecha taxón obtenida de un fragmento de costilla arrojó una edad de 916 ± 42 años C14 AP, cronología que resultó ser penecontemporánea con la datación efectuada sobre los restos humanos provenientes de la UE-3, cuya antigüedad fue fijada en 976
Figura1.UbicacióndelsitioCerroLutz.
Tabla1.FechadosradiocarbónicosobtenidosparaelsitioCerroLutz.
CerroLutz
Unidaddeexcavación
Materialdatado Laboratorio C13 CronologíaenañosC14AP
UE-2 Hueso H. sapiens LP-1711 -20.0±2‰ 730±70 665±66añoscal.
UE-2 Hueso H. sapiens AA77311 -19.7±2‰ 796±42 724±30añoscal.
UE-3 Hueso H. sapiens AA77310 -20.3±2‰ 976±42 877±53añoscal.
UE-4 Hueso C. familiaris AA77312 -22.0±2‰ 916±42 844±54añoscal.

1 8 5
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.175-199
P R I M E R R E G I S T R O D E P E R R O D O M É S T I C O | A L E J A N D R O A C O S T A , D A N I E L L O P O N T E Y C É S A R G A R C Í A
± 42 años C14 AP (ver la tabla 1). Esto último sugiere una estrecha asociación entre la ocupación arqueológica y C. familiaris. En tal sentido, el alto grado de articulación anatómica que presentaba el esqueleto y su alta integridad tafonó-mica permiten plantear que el perro habría sido intencionalmente depositado y enterrado por los grupos humanos que ocuparon Cerro Lutz, aspecto que luego retomaremos.
Figura2.Perfilestratigráfico(E)delaUnidaddeExcavación4,endondeserecuperóelejemplardeC. familiaris(INAPL/CL1-UE-3).
Descripción y determinación taxonómica
El ejemplar recuperado en Cerro Lutz (INAPL/CL1-UE-3) fue determinado como C. familiaris, principalmente sobre la base del análisis de sus caracteres cráneo-dentarios. El mismo se comparó con otros ejemplares de perros y de cánidos sudamericanos como Lycalopex culpeus (zorro colorado), L. gymno-cercus (zorro de las pampas), L. griseus (zorro gris), Cerdocyon thous (zorro de monte) y Ch. brachyurus (aguará guazú), depositados en las colecciones del Museo de La Plata y Museo Argentino de Ciencias Naturales “Bernardino Rivadavia” (Argentina).
Con el fin de determinar el tamaño del ejemplar, se consideraron la altura
Unidadexcavación4
Valvas (Diplodon sp.)

1 8 6
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.175-199
en la cruz y el peso del mismo. La estimación de la altura se realizó mediante las fórmulas propuestas por Harcourt (1974), las cuales utilizan la longitud total de los huesos largos de los miembros. El peso se calculó a partir de las circunfe-rencias del húmero y del fémur, mediante las fórmulas propuestas por Anyonge (1993), y a partir de la altura de la rama horizontal de la mandíbula, utilizando la fórmula de Wing (1978).
Para evaluar la forma general del cráneo, se calcularon los índices facial (IF = ancho bicigomático x 100/longitud nasion-prostion), de la caja craneana (ICC = ancho interparietal x 100/longitud inion-nasion) y craneal (IC = ancho bicigomático x 100/longitud inion-prostion). Estos índices fueron tomados de Evans (1993) y comparados con los allí publicados para cráneos dolicocéfalos, mesocéfalos y braquicéfalos. Todas las medidas lineales fueron tomadas con un calibre digital con una precisión de 0,01 mm. Para determinar el sexo se observó la morfología del hueso basioccipital y se calculó un índice que tiene en cuenta la relación longitud-ancho del mismo (Trouth et al., 1977).
Análisis morfológicoEl cráneo (arco cigomático derecho incompleto, proceso paraoccipital izquierdo roto en su base), al igual que la dentición, presentan un alto grado de integridad (ver la figura 3). El esqueleto postcraneano se encuentra representado por las vértebras cervicales, doce vértebras torácicas, cinco vértebras lumbares, nume-rosas costillas, restos de las dos escápulas, húmeros, radios, ambos cúbitos sin el extremo distal, metacarpales II-V, huesos coxales, fémures, tibias, extremo distal de la f íbula izquierda; calcáneos, astrágalo izquierdo y metatarsales II-V (ver algunos ejemplos en la figura 4).
Entre los caracteres cráneo-dentarios que permitieron asignar clara-mente el material al género Canis y diferenciarlo de los zorros sudamericanos se pueden mencionar los siguientes (sensu Tedford et al., 1995): el gran desa-rrollo de los senos frontales, los cuales penetran en los procesos postorbitarios y se extienden caudalmente ensanchando la constricción postorbitaria y otor-gando un perfil convexo al cráneo en la región dorsal a las órbitas; el desarrollo aguzado del inion, que, en vista lateral, claramente sobrepasa caudalmente a los cóndilos occipitales; el considerable desarrollo del proceso angular de la mandíbula, debido a la gran extensión de la fosa para la inserción de la rama dorsal del m. pterygoideus medialis; el gran desarrollo de los I3, los cuales pre-sentan un cíngulo posteromedial bien definido; el mayor desarrollo del para-cono en relación con el metacono en los M 1-2. Asimismo, la relación entre la longitud del radio y la longitud de la tibia es del 90% (en los zorros sudameri-canos es menor que ese valor, excepto en el aguará guazú). Otros caracteres

1 8 7
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.175-199
P R I M E R R E G I S T R O D E P E R R O D O M É S T I C O | A L E J A N D R O A C O S T A , D A N I E L L O P O N T E Y C É S A R G A R C Í A
cráneo-dentarios, que habitualmente son utilizados para diferenciar a C. fami-liaris de C. lupus, también permitieron diferenciar al ejemplar de Cerro Lutz de los zorros sudamericanos. Generalmente, se interpreta que estos caracteres estarían relacionados con el proceso de domesticación del perro (e.g., Clutton-Brock et al., 1976; Olsen, 1985; Koler-Matznick, 2002, entre otros); entre ellos se pueden enumerar: el mayor ancho relativo de la región facial del cráneo; el desarrollo de un perfil cóncavo en la región frontal, por delante de la órbita; el desarrollo de una mayor convexidad en el borde ventral de la mandíbula; una mayor curvatura caudal del proceso coronoides y el desarrollo más cóncavo de su borde caudal; un menor desarrollo de la bulla timpánica, la cual es rela-tivamente más pequeña y aplanada; la disposición apiñada de los premolares y, en algunos casos, su orientación oblicua respecto de la serie dentaria; un menor desarrollo de los dientes que componen la cuchilla carnicera (PM4-m1), los cuales son relativamente más cortos. Se observaron, además, algunas anomalías dentarias, tales como ausencia del pm1 izquierdo y presencia de incisivos supernumerarios. Estos últimos están representados por un pequeño incisivo ubicado entre los I1-I2 derechos y por un pequeño alvéolo ubicado entre los mismos dientes del lado izquierdo. Los valores estimados mediante las mediciones cráneo-dentarias, al igual que los obtenidos para los huesos largos, pueden verse en la tabla 2.
Figura3.Vistaslateral,dorsalyventraldelcráneodeC. familiaris(INAPL/CL1-UE-3)recuperadoenelsitioCerroLutz.Escala:5cm.

1 8 8
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.175-199
Los restos recuperados corresponden a un individuo adulto (epífi-sis soldadas, dentición definitiva totalmente erupcionada y con poco des-gaste, sutura basioccipital-basiesfenoides fusionada), cuyo sexo es probable que corresponda a una hembra (índice sexual = 146,4; hembra si es mayor que 136, macho si es menor que 123). La altura en la cruz estimada es de unos 47 cm (media = 46,6 cm; mínimo = 44,6, promedio a partir de las tibias; máximo = 47,7, promedio a partir de los fémures). El peso promedio estimado es de 15,9 kg (mínimo = 14,1 kg, a partir de la altura de la rama horizontal de la mandíbula; máximo = 17,3, a partir del promedio de la circunferencia de los húmeros). Los índices del cráneo (IF = 117,2; ICC = 53,1; IC = 57,2) corres-ponden a los de uno del tipo mesocéfalo. Los resultados obtenidos indican que el ejemplar (INAPL/CL1-UE-3) habría sido de un tamaño equivalente al de las razas medianas de perros actuales.
Discusión y conclusionesLa presencia de perros prehispánicos en el humedal del Paraná inferior (HPI), y en el extremo sur de Sudamérica en general, genera nuevos interrogantes arqueológicos tanto en lo que se refiere a su origen como al grado y tipo de interacción que habrían existido entre éstos y las poblaciones aborígenes.
Figura4.ElementosdelesqueletoapendiculardeC. familiaris(INAPL/CL1-UE-3)recuperadoenelsitioCerroLutz.Arriba,deizquierdaaderecha:húmerosderechoeizquierdo(vistaposterior);radiosycúbitosderechoseizquierdos(vistamedial);fémuresderechoeizquierdo(vistaanterior);tibiasderechaeizquierda(vistaanterior).Abajo,dederechaaizquierda:fragmentodistaldelaescápulaizquierda(vistalateral),huesoscoxalesderechoeizquierdo(vistalateral).Escala:5cm.

1 8 9
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.175-199
P R I M E R R E G I S T R O D E P E R R O D O M É S T I C O | A L E J A N D R O A C O S T A , D A N I E L L O P O N T E Y C É S A R G A R C Í A
Tabla2.Medidas(enmm)cráneo-dentariasydeloshuesoslargosdeC. familiaris(INAPL/CL1-UE-3), procedentedeCerroLutz.
Longituddelcráneo Akrokranion-prosthion 174,6*
Longituddelacajacraneana Akrokranion-nasion 95,4*
Longitudcondilobasal Margenposteriorcóndilosoccipitales-prosthion 167,5
Longitudfacial Nasion-prosthion 85,2*
Longitudnasal Nasion-rhinion 53,0
Longitudpalatal Staphylion-prosthion 87,5
Anchobimastoideo Otion-Otion 60,7
Anchobicigomático Zygion-zygion 99,9*
Anchointerparietal Euryon-euryon 51,0*
Anchopalatal TomadaanivelPM4-M1 65,4
Anchointerorbitario Entorbitale-entorbitale 32,0
Anchopostorbitario Ectorbitale-ectorbitale 46,6
Constricciónpostorbitaria Menoranchodelaconstricciónpostorbitaria 33,5
Longituddelabulla Longitudmáxima(sinelprocesoestiloides) 20,1
LongitudPM4 Tomadaaniveldelcíngulo 18,2
AnchoPM4 Tomadaaniveldelcíngulo 10,1
LongitudM1 Tomadaaniveldelcíngulo 12,2
AnchoM1 Tomadaaniveldelcíngulo 16,0
Longituddelamandíbula Infradentale-processusangularis 131,43
Alturadelamandíbula TomadaaniveldelmargenalveolarposteriordelM1 22,0*
LongituddelM1 Tomadaaniveldelcíngulo 20,0
AnchodelM1 Tomadaaniveldelcíngulo 8,7
Longituddelhúmero Longitudmáximadelhúmero 146,1*
CircunferenciadelhúmeroTomadaa35%delalongitudtotal,desdeelextremodistal
40*
Longituddelradio Longitudmáximadelradio 141,0*
Longituddelfémur Longitudmáximadelfémur 156,2*
Circunferenciadelfémur Tomadaenlamitaddeladiáfisis 40*
Longituddelatibia Longitudmáximadelatibia 156,2*
*Medidasutilizadasparaelcálculodeíndicescraneanosoestimacionesdelpesoyalzada(véaseeltexto)

1 9 0
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.175-199
Una primera pregunta surge, justamente, a raíz de su reciente identificación, un hecho realmente notable es su escasa presencia arqueológica, a pesar de los numerosos sitios que se han excavado, particularmente en las regiones Pam-peana y Patagónica en los últimos veinte años. Esto no implica descartar que existan defectos relacionados con el muestreo arqueológico. Tampoco creemos que su baja frecuencia pueda ser simplemente atribuida a que fueron introdu-cidos tardíamente (ver más abajo), tal como lo indican los pocos hallazgos y fechados radiocarbónicos que hasta ahora se conocen.
Por otro lado, debe notarse la escasa a nula existencia de fuentes etnohis-tóricas del siglo XVI (e incluso del XVII) en donde se mencione la presencia de perros entre los grupos aborígenes de Pampa-Patagonia, hecho que contrasta con la información de la que se dispone para el noroeste argentino y la región andina en general (ver algunos ejemplos en Gallardo, 1964-65, Raffino et al., 1977). En relación con el HPI, uno de los pocos datos del siglo XVI que se cono-cen proviene de Gonzalo Fernández de Oviedo y Valdés (1944), quien señaló que los grupos Timbúes que habitaban los territorios adyacentes al curso medio e inferior del río Paraná tenían “[…] muchos perros como los nuestros grandes y pequeños, que ellos estiman mucho, los cuales allá no avía, y se han hecho de la casta que quedó de quando Sebastián Gaboto y el capitan Jhoan del Junco anduvieron por aquella tierra” (Fernández de Oviedo y Valdés, 1944 V, XI: 155). En la misma página, se refiere a otros grupos indígenas que habitaban un sector más septentrional de la cuenca del Plata, y dice que también tenían “[…] unos perrillos que crían en sus casas, mudos, que no ladran y los tienen por buen manjar, y los comen cuando quieren” (Fernández de Oviedo y Valdés, 1944 V, XI: 155). Aunque en esta referencia no menciona el origen de dichos perros, es posible que en este caso se tratara de una raza nativa.
Dentro del HPI, lamentablemente, aún no se conocen sitios arqueológi-cos que correspondan al siglo XVI, en donde, de acuerdo con la crónica de Fernández de Oviedo y Valdés, se pueda eventualmente constatar la presencia de perros domésticos. En cambio, son numerosos los depósitos arqueológicos que se han investigado para momentos previos a la conquista europea, los cua-les en su mayoría se ubican dentro de los últimos 1.000 años C14 AP (Caggiano, 1984; Loponte, 2008), rango temporal que involucra a los perros recuperados tanto en Cerro Lutz como en los sitios Angostura 1 y Chenque 1 y, posible-mente, los hallados en Uruguay. En este sentido, y retomando el tema de su escasa representación en tiempos prehispánicos, es necesario realizar algunas consideraciones. Prates et al. (2010a y 2010b) sostienen que el origen tardío de los perros en el extremo sur de Sudamérica podría deberse a los complejos procesos sociales que experimentaron las poblaciones cazadoras-recolectoras

1 9 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.175-199
P R I M E R R E G I S T R O D E P E R R O D O M É S T I C O | A L E J A N D R O A C O S T A , D A N I E L L O P O N T E Y C É S A R G A R C Í A
de Pampa-Patagonia durante la fase final del Holoceno tardío, momento en el que se habrían incrementado la interacción y la circulación de los individuos, bienes e ideas a lo largo de grandes distancias. De ser así, es probable que la incorporación de los perros dentro de las poblaciones aborígenes haya sido un fenómeno más bien circunstancial que sistemático. De otro modo, a no ser que existan importantes deficiencias de muestreo, es muy dif ícil explicar su baja identificabilidad arqueológica entre los grupos cazadores-recolectores antes del contacto europeo. Por lo tanto, en el caso de que los perros hayan cons-tituido bienes (individuales) de intercambio provenientes de regiones distan-tes, es factible que la frecuencia de este tipo de eventos haya sido significativa-mente baja y ocasional. Una situación similar puede plantearse en relación con el intercambio de ciertos objetos y/o materias primas alóctonos (e.g., metales) que también suelen tener una muy baja frecuencia, llegando a constituir en algunos casos hallazgos excepcionales; un claro ejemplo puede verse, para el norte de la Patagonia, en Gómez Otero y Dahinten (1999).
De haberse producido el ingreso de C. familiaris bajo las condiciones antes señaladas, es posible que sus poblaciones en el extremo sur de Sudamérica no hayan alcanzado a tener el mismo grado de proliferación como el que habrían tenido en la región andina. Esto sería concordante con la poca información etnohistórica que se conoce sobre la presencia de perros entre los aborígenes de Pampa y Patagonia durante los primeros siglos de la conquista europea. Si esto fue así, puede pensarse en que debió ser extremadamente baja la cantidad de perros que ingresaron como bienes de intercambio. De haber sido mayor, es probable que existiesen muchos más casos arqueológicos de los que hasta hoy se registraron. Aquí debe reiterarse que los hallazgos realizados en Cerro Lutz, Angostura 1 y Chenque 1 presentan una antigüedad circa a los 1.000 años C14 AP. Por lo tanto, y de haber existido un mayor número de perros circulando entre los grupos aborígenes, puede considerarse que el tiempo transcurrido entre la fecha aludida y el siglo XVI habría sido más que suficiente para que se produjera un significativo aumento de su población. Esto parece ser lo que ocurrió en la Región Pampeana con los perros europeos introducidos a partir del siglo XVI, en donde en el transcurso de unos doscientos años habrían dado origen a las grandes y densas jaurías de perros cimarrones que fueron histórica-mente registradas unos años antes de mediados del siglo XVIII (Cabrera, 1932). Dada la gran magnitud que tuvo dicho fenómeno, y de ser cierto que existió un reducido número de perros prehispánicos, es posible que estos últimos hayan sido rápidamente asimilados por las razas introducidas por los europeos, tal como parece haber sucedido con las poblaciones nativas de perros de otras regiones de América (ver Scott, 1968: 264).

1 9 2
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.175-199
Volviendo a nuestro caso de estudio, la estructura arqueológica regional del HPI y los distintos rasgos que han sido identificados en el nivel arqueoló-gico, y/o a partir de diversas crónicas del siglo XVI, sugieren que durante el Holoceno reciente los grupos cazadores-recolectores habían generado adapta-ciones denso-dependientes y se hallaban en un creciente proceso de comple-jidad social (ver detalles en Acosta et al., 2010; Loponte et al., 2005; Loponte, 2008). Bajo estas condiciones, y debido al crecimiento de la población, pudie-ron incrementarse la interacción entre los grupos, la generación de conflictos, la diferenciación entre los individuos y la adquisición de prestigio mediante distintas conductas (cf. Hayden y Gargett, 1990; Clark y Blake, 1992), dentro de las que puede incluirse la obtención de bienes suntuarios o exóticos mediante el intercambio (cf. Ames, 2005), situación bajo la cual pudieron ser adquiridos los perros. Para el HPI, Loponte (2008), en función de la información arqueo-lógica y etnohistórica disponible, ha propuesto cuatro posibles y grandes vec-tores relacionados con el intercambio y el flujo de bienes y de información que habrían existido entre los grupos humanos del HPI y los que habitaban otras regiones. Estos procesos habrían presentado diferentes trayectorias en cuanto a las distancias y al tipo de elementos implicados en las actividades de intercam-bio. Los principales vectores o macroespacios involucrados tanto en la circu-lación interna como externa de los bienes habrían sido: a) el corredor fluvial y áreas adyacentes del Paraná medio e inferior y del estuario superior del Río de La Plata, b) las sierras septentrionales bonaerenses, c) las sierras centrales de la provincia de Córdoba y d) el área Chaco-santiagueña y el N.O.A. Presumi-blemente de alguna de estas dos últimas regiones pudieron provenir los perros (prehispánicos), al igual que otros objetos tales como los metales y los textiles de los que existen evidencias arqueológicas e históricas (para diferentes ejem-plos acerca de esta dinámica, ver Loponte, 2008).
Un comentario aparte merecen los perros que fueron recuperados en Uruguay. Aunque es prematuro saber si en esta región también pudieron haber constituido o no bienes de intercambio, es posible plantear algunas ideas al respecto. Es interesante notar que durante el Holoceno tardío los denominados “constructores de cerritos” habrían desarrollado complejos sistemas socioeco-nómicos (ver Pintos, 1999), por lo que también sería esperable que hayan gene-rado distintas redes y formas de intercambio. Además, debe considerarse que hasta ahora en Uruguay, al igual que en otras regiones del extremo sur de Sud-américa, no se han recuperado restos de perros que puedan remitirse al límite Pleistoceno-Holoceno, ni al Holoceno medio, hecho que también permite pre-suponer su aparente origen tardío. Más allá de los mecanismos culturales que pudieron estar implicados, para el Holoceno tardío, existen dos posibles vías

1 9 3
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.175-199
P R I M E R R E G I S T R O D E P E R R O D O M É S T I C O | A L E J A N D R O A C O S T A , D A N I E L L O P O N T E Y C É S A R G A R C Í A
de ingreso de los perros al territorio uruguayo: sur de Brasil, o bien desde el Litoral Argentino, a través del río Uruguay, siendo este último un curso fluvial fácilmente vadeable, particularmente cuando se producen las grandes bajantes. Esta segunda ruta de acceso sería la más probable, dado que en Brasil no existe hasta el momento información que avale la presencia de perros prehispánicos.
Por otra parte, sabemos que los perros, ya sea por sus fines prácticos y/o simbólicos, fueron bienes muy valorados por numerosas sociedades humanas a través del tiempo. Entre sus principales aspectos utilitarios figuran su apro-vechamiento como alimento y como complemento en las actividades relacio-nadas con la caza (cf. Schwartz, 1997). A juzgar por la documentación histó-rica, recién en los siglos XVIII y XIX los perros parecen haber sido activamente incorporados a los sistemas de caza por los aborígenes que habitaban distintas regiones de la Pampa y Fuego-Patagonia, e incluso del Chaco. En el HPI, ya hacia fines del siglo XVII y principios del XVIII, las poblaciones indígenas se encontraban en un franco proceso de extinción. Por lo tanto, y dada la escasa representación que habrían tenido los perros en momentos prehispánicos, es sumamente dif ícil evaluar si tuvieron algún grado de importancia cinegética dentro de las estrategias de caza, siendo más dificultoso aún establecer si cons-tituyeron parte de la alimentación. Esto mismo se plantea en cuanto a su posi-ble intervención como agente tafonómico.
Un aspecto que sí puede considerarse a pesar de su baja frecuencia, es su evidente relación con la funebría. El entierro de perros, ya sean solos y/o asociados a las inhumaciones humanas, es una conducta que se ha registrado en diferentes lugares del mundo desde hace por lo menos 14.000 años AP, hecho que pone de manifiesto el estrecho vínculo y la importancia que tuvie-ron los perros para las grupos humanos luego de que fueran domesticados (una síntesis puede verse en Morey, 2006). No es casual que de los pocos casos registrados entre los cazadores-recolectores del extremo sur de Sudamérica, con excepción de Angostura 1, en el resto de los sitios se haya identificado este tipo de prácticas mortuorias. Tal es el caso de Chenque 1 (Prates et al., 2010a y 2010b) y el de los perros hallados en los cerritos de Uruguay, ya sea porque estaban en asociación directa con los entierros humanos o porque se encontraban muy cercanos a ellos (González, 1999). En Cerro Lutz es real-mente llamativo que, en la unidad de excavación (UE-4) en donde se recuperó el esqueleto, no se hayan identificado restos humanos (ver más arriba). De confirmarse esta situación en futuras excavaciones, y considerando las carac-terísticas arriba mencionadas, estaríamos ante uno de los primeros entierros individuales de C. familiaris que se conocen para los grupos cazadores-reco-lectores del extremo sur del subcontinente.

1 9 4
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.175-199
La reciente identificación de perros prehispánicos en el extremo sur de Sudamérica abre nuevas e interesantes líneas de investigación. Es evidente que varios de los aspectos aquí planteados han generado más interrogantes que respuestas, tanto en lo que se refiere a su origen como al tipo y grado de interacción que mantuvieron con las poblaciones humanas. Un aspecto que aún debe explorarse en profundidad es la relación filogenética que poseen los ejemplares aquí mencionados con otros perros prehispánicos americanos y de otras regiones del mundo. Los resultados que se obtengan del C. familiaris recuperado en Cerro Lutz, a partir del análisis de ADN en curso, permitirán conocer un poco más sobre la trayectoria histórica que tuvieron los perros en nuestro subcontinente. .

1 9 5
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.175-199
P R I M E R R E G I S T R O D E P E R R O D O M É S T I C O | A L E J A N D R O A C O S T A , D A N I E L L O P O N T E Y C É S A R G A R C Í A
R e f e R e n c i a s
Acosta, Alejandro, Daniel Loponte y Pablo Tchilinguirian 2010. “Análisiscomparativosobrelaestructuraylosprocesosdeformacióndelosdepósitos
arqueológicosenelhumedaldelParanáinferior”,enArqueología de cazadores recolectores de la cuenca del Plata,G.CoccoyM.R.FeuilletTerzaghi,comps.,pp.191-208.SantaFe,CentrodeEstudiosHispanoamericanos.
Allen, Glover W.1920.“DogsoftheAmericanAborigines”,Bulletin of the Museum of Comparative Zoology at Harvard
College63,pp.431-517.
Ames, Kenneth M. 2005.“IntensificationofFoodProductionontheNorthwestCoastandElsewhere”,enThe Northwest
Coast, Foragers or Farmers?,D.DueryN.Turner,eds.,pp.64-94.Seattle,UniversityofWashingtonPress.
Amorosi, Thomas y Francisco J. Prevosti 2008.“APreliminaryReviewoftheCanidRemainsfromJuniusBird’sExcavationsatFell’sandPaliAike
Caves,Magallanes,Chile”, Current Research in the Pleistocene, Archaeology: Latin America25, pp.25-27.
Andersone, Žanete, Vit torio Lucchini, Et tore Randi y J. Ozoliš2002.“HybridizationbetweenWolvesandDogsinLatviaasDocumentedUsingMitochondrialand
MicrosatelliteDNAMarkers”,Mammalian Biology 67,pp.79-90.
Anyonge, William1993.“BodyMassinLargeExtantandExtinctCarnivores”,Journal of Zoology231,pp.339-350.
Behrensmeyer, Anna Kay 1978.“TaphonomicandEcologicInformationfromBoneWeathering”,Paleobiology4,pp.150-162.
Bird, Junius B.1988.Travels and Archaeology in South Chile.IowaCity,UniversityofIowaPress.
Borrero, Luis A.2009.“TheElusiveEvidence:TheArcheologicalRecordofTheSouthAmericanExtinctMegafauna”,
enAmerican Megafaunal Extinctions at the End of the Pleistocene”,G.Haynes,pp.145-168, ed.,Dordrecht,SpringerPublishers.
Burkart, Rodolfo, Néstor Bárbaro, Roberto O. Sánchez y Daniel A . Gómez1999.Ecorregiones de la Argentina.BuenosAires,AdministracióndeParquesNacionales.
Cabrera, Ángel1932.“ElperrocimarróndelaPampaArgentina”,Publicaciones del Museo Antropológico y Etnográfico
de la Facultad de Filosofía y Letras,Serie A,2,pp.7-36.1934.“Losperrosdomésticosdelosindígenasdelterritorioargentino”, 25º Congreso Internacional
Americanistas 1,pp.83-93.
Caggiano, María Amanda1984.“PrehistoriadelNordesteArgentino,susvinculacionesconlaRepúblicaOrientaldelUruguayySur
deBrasil”,Pesquisas Antropología38,pp.1-109.
Cardich, Augusto, Eduardo P. Tonni y Néstor Kriscautzky1977.“PresenciadeCanis familiarisenrestosarqueológicosdeLosToldos(ProvinciadeSantaCruz,
Argentina)”,Relaciones de la Sociedad Argentina de Antropología11,pp.115-119.

1 9 6
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.175-199
Caviglia, Sergio1980. “LapresenciadeDusicyon avus(Burmeister),1864,enlacapaVIIIdelacueva‘LasBuitreras’
(Patagonia,Argentina):surelaciónconotroshallazgosenPatagoniaMeridional”,Runa13, pp.31-33.
1985-86.“NuevosrestosdecánidostempranosensitiosarqueológicosdeFuego-Patagonia”,Anales del Instituto de la Patagonia16,pp.85-93.
Clark, John E. y Michael Blake1992.“ThePowerofPrestige:CompetitiveGenerosityandtheEmergenceofRankSocietiesinLowland
Mesoamerica”,enFactional Competition and Political Development in the New World, E. M. BrumfielyJ.W.Fox,eds.,pp.17-30.Cambridge,CambridgeUniversityPress.
Clut ton-Brock, Juliet1977.“Man-madeDogs”, Science197,pp.1340-1342.1978.“TheCarnivoreRemainsfromtheExcavationsatFell’sCave,ChileCarriedoutbyDr.JuniusBirdin
1970”,pp.1-6.Ms.
Clut ton-Brock, Juliet, Gordon B. Corbert y Michael Hills1976.“AReviewoftheFamilyCanidae,withaClassificationbyNumericalMethods”,Bulletin of the
British Museum (Natural History), Zoology 29,pp.117-199.
Evans, Howard E.1993.Miller´s Anatomy of the Dog.Filadelfia,W.B.SaundersCompany.
Fernández de Oviedo y Valdés, Gonzalo1944.Historia General y Natural de las Indias. Islas y Tierra-Firme del Mar Océano.Sietevol.Asunción
delParaguay,EditorialGuarania.
Gallardo, Guillermo1964-1965.“Perrosamericanosprecolombinos”,Cuadernos del Instituto Nacional de Antropología5,
pp.31-68.
Germonpré, Mietje; Mikhail V. Sablin, Rhiannon E. Stevens, Robert E. M. Hedges, Michael Hofreiter, Mathias Stiller y Viviane R. Després
2009.“FossilDogsandWolvesfromPalaeolithicSitesinBelgium,theUkraineandRussia:Osteometry,AncientDNAandStableIsotopes”,Journal of Archaeological Science 36,pp.473-490.
Gómez Otero, Julieta y Silvia Dahinten1999.“EvidenciasdecontactosinterétnicosenelsigloXVIenPatagonia:informepreliminarsobreel
sitioenterratorioRawson(Chubut)”.Actas XII Congreso Nacional de Arqueología Argentina,TomoIII,pp.44-53.LaPlata.
González, Roberto1999.Canis familiaris y constructores de cerritos: una perspectiva funeraria.TallerIIdeArqueología,
FHCE.Montevideo.Uruguay.Ms.
Grayson, Donald K.1988.“DangerCave,LastSupperCave,andHangingRockShelter:Thefaunas”.Anthropological Papers
of the American Museum of Natural History66,pp.1-130.
Harcourt, Robert A.1974.“TheDoginPrehistoricandEarlyHistoricBritain”,Journal of Archaeological Science1,
pp.151-175.
Hayden, Brian y Rob Garget t1990.“BigMan,BigHeart?AMesoamericanViewofTheEmergenceofComplexSocieties”,Ancient
Mesoamerica1,pp.3-20.

1 9 7
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.175-199
P R I M E R R E G I S T R O D E P E R R O D O M É S T I C O | A L E J A N D R O A C O S T A , D A N I E L L O P O N T E Y C É S A R G A R C Í A
Koler-Matznick, Janice2002.“TheOriginoftheDogRevisited”,Anthrozoös15,pp.98-118.
Koop Ben F., Maryann Burbidge, Ashley Byun, Ute Rink y Susan. J. Crockford2000.“AncientDNAEvidenceofaSeparateOriginforNorthAmericanIndigenousDogs”,enDogs
Through Time: An Archaeological Perspective,S.J.Crockford,ed.,pp.271-286.Oxford,Archaeopress,B.A.R.
Kriscautzky, Néstor1975.“Nuevosaportesparaelconocimientodelosmaterialesarqueológicosobtenidosenlaexcavación
delsitioRíoLujánenlallanuraaluvialdedichocurso”,Arqueología1,pp.9-13.
Kubinyi, Eniko, Zsófia Virányi y Adám Miklósi2007.“ComparativeSocialCognition:FromWolfandDogtoHumans”,Comparative Cognition &
Behavior Reviews 2,pp.26-46.
Leonard, Jennifer A., Robert K. Wayne, Jane Wheeler, Raúl Valadez, Sonia Guillen y Carles Vilá2002.“AncientDNAEvidenceforOldWorldOriginofNewWorldDogs”,Science298,pp.1613-1616.
Lopéz Mazz, José M.2001.“Lasestructurastumulares(cerritos)dellitoralatlánticouruguayo”,Latin American Antiquity12,
pp.231-255.
Loponte, Daniel2008.Arqueología del Humedal del Paraná Inferior (Bajíos Ribereños meridionales).BuenosAires,
InstitutoNacionaldeAntropologíayPensamientoLatinoamericano,SecretaríadeCulturadelaNación.
Loponte, Daniel, Alejandro Acosta y Javier Musali2005.“ComplexityamongHunter-gatherersfromthePampeanRegion,SouthAmerica”,enBeyond
Affluent Foragers, C.Grier,J.KimyJ.Uchiyama,eds.,pp.106-125.Oxford,OxbowBooks.
Mendoza España, Velia y Raúl Valadez2006.“Elperroprehispánicoandino:funciónytiposapartirdelanálisisarqueozoológico”,Anales de la
XIX Reunión Anual de Etnología,pp.31-38.LaPaz,MuseoNacionaldeEtnografíayFolklore.
Morey, Darcy. F.1992.“Size,ShapeandDevelopmentintheEvolutionoftheDomesticDog”,Journal of Archaeological
Science19,pp.181-204.2006.“BuryingKeyEvidence:theSocialBondBetweenDogsandPeople”,Journal of Archaeological
Science33,pp. 158-175.
Morey, Darcy F. y Michael D. Wiant1992.“EarlyHoloceneDomesticDogBurialsfromtheNorthAmericanMidwest”,Current Anthropology33,
pp.224-229.
Okumura, Naohito, Naotakana Ishiguro, Masuo Nakano, Akira Matsui y Makoto Sahara1996.“Intra-andInterbreedGeneticVariationsOfMitochondrialDNAMajorNoncodingRegionsin
JapaneseNativeDogBreeds(Canis familiaris),Animal Genetics27,pp.397-405.
Olsen, Stanley J.1985.Origins of the Domestic Dog: The Fossil Record.Tucson,TheUniversityofArizonaPress.
Ostrander, Elaine. A. y Robert K. Wayne2005.“TheCanineGenome”,Genome Research15,pp.1706-1716.
Pintos, Sebastián1999.“Túmulos,caciquesyotrashistoriascazadoresrecolectorescomplejosenlacuencadelalaguna
deCastillos,Uruguay”,Complutum10,pp.213-226.

1 9 8
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.175-199
Pintos, Sebastián y Roberto Bracco1999.“Modalidadesdeenterramientoyhuellasdeorigenantrópicoenespecímenesóseoshumanos.
TierrasBajasdelestedelUruguay(R.O.U.)”,enArqueología y Bioantropología de las Tierras Bajas,J.M.LópezyM.Sans,comps.,pp.81-106.Montevideo,FacultaddeHumanidadesyCienciasdelaEducación,UniversidaddelaRepública.
Prates, Luciano, Francisco J. Prevosti y Mónica Berón 2010a.“FirstRecordsofPrehispanicDogsinSouthernSouthAmerica(Pampa-Patagonia,Argentina)”,
Current Anthropology51,pp.273-280.
Prates, Luciano, Mónica Berón y Francisco J. Prevosti2010b.“LosperrosprehispánicosdelConoSur.Tendenciasynuevosregistros”,Mamül Mapu: pasado
y presente desde la arqueología pampeana,enM.Berón,L.Luna,M.Bonomo,C.Montalvo,C.ArandayM.CarreraAizpitarte,eds.,pp.215-228.Ayacucho,EditorialLibrosdelEspinillo.
Prevosti Francisco J, Mariano Bonomo y Eduardo P. Tonni2004.“LadistribucióndeChrysocyon brachyurus(Illiger,1811)(Mammalia:Carnivora:Canidae)duranteel
HolocenoenlaArgentina:implicanciaspaleoambientales”, Mastozoología Neotropical 11,pp.27-43.
Raffino, Rodolfo Eduardo Tonni y Alberto Cione1977.“RecursosalimentariosyeconomíaenlaQuebradadelToro”,Relaciones de la Sociedad
Argentina de Antropología11,pp.9-29.
Roth, Santiago1902.“NuevosrestosdemamíferosdelacavernaEberhardtenÚltimaEsperanza”,Revista del Museo
de La Plata11,pp.37–54.
Sablin, Mikhail V. y Gennady A. Khlopachev2002.“TheEarliestIceAgedogs:EvidencefromEliseevichi”,Current Anthropology 43,pp.795-799.
Savolainen, Peter, Ya-ping Zhang, Jing Lou, Joakim Lundeberg y Thomas Leitner 2002.“GeneticEvidenceforanEastAsianOriginofDomesticDogs”,Science298,pp.1610-1613.
Scott, John Paul1968.“EvolutionandDomesticationoftheDog”,Evolutionary Biology2,pp.243-275.
Schwartz, Marion1997.A History of Dogs in the Early Americas.NewHaven,YaleUniversityPress.
Tedford, Richard H., Beryl. E. Taylor y Xiaomong Wang1995.“PhylogenyoftheCaninae(Carnivora:Canidae):theLivingTaxa”,American Museum Novitates
3146,pp.1-37.
Tonni, Eduardo P. y Gustavo Politis1981.“UngrancánidodelHolocenodelaProvinciadeBuenosAiresyelregistroprehispánicodeCanis
(Canis) familiarisenlasáreasPampeanayPatagónica”,Ameghiniana18,pp.251-265.
Trouth, Ovid., Susan Winter, Karen C., Gupta, Richard M. Millis y James A. Holloway1977.“AnalysisoftheSexualDimorphismintheBasioccipitalPortionoftheDog´sSkull”,Acta
Anatomica98,pp.469-473.
Tsuda, Kaoru, Yoshiaki Kikkawa, Hiromichi Yonekawa y Yuichi Tanabe1997.“ExtensiveInterbreedingOccurredamongMultipleMatriarchalAncestorsDuringthe
DomesticationofDogs:EvidencefromInter-andIntraspeciesPolymorphismsintheD-loopRegionofMitochondrialDNAbetweenDogsandWolves”,Genes and Genetic Systems72, pp.229-238.

1 9 9
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.175-199
P R I M E R R E G I S T R O D E P E R R O D O M É S T I C O | A L E J A N D R O A C O S T A , D A N I E L L O P O N T E Y C É S A R G A R C Í A
Valadez, Raúl, Jennifer Leonard y Carles Vilá2003.“Elorigendelperroamericanovistoatravésdelabiologíamolecular”,Asociación Mexicana de
Médicos Veterinarios Especialistas en Pequeñas Especies14,pp.73-82.
Vilá, Carles, Peter Savolainen, Jesús Maldonado, Isabel R. Amorim, John E. Rice, Rodney L. Honeycutt, Keith A. Crandall, Joakim Lundeberg, y Robert K. Wayne
1997.“MultipleandAncientOriginsoftheDomesticDog”,Science276,pp.1687-1689.
Wayne, Robert K. y Carles Vilá2003.“MolecularGeneticStudiesofWolves”,enWolves Behavior, Ecology, and Conservation, L. D.
MechyL.Boitani,eds.,pp.218-238.Chicago,UniversityofChicagoPress.
Wayne, Robert K., Jennifer A. Leonard y Carles Vilá2006.“GeneticAnalysisofdogDomestication”,enDocumenting Domestication. New Genetic and Ar-
chaeological Paradigms,M.AZeder,D.G.Bradley,E.EmshwilleryB.D.Smith,eds.,pp.279-293. Berkeley,LosÁngelesyLondres,UniversityCaliforniaPress.
Wing, Elizabeth1978.“UseofDogsasFood:AnAdaptationtotheCoastalEnvironment”,enPrehistoric Coastal
Adaptations: The Economy and Ecology of Maritime Middle America, B.L.StarkyB.Voorhies,eds.,pp.29-41.NuevaYork,AcademicPress.
1989.“HumanUseofCanidsintheCentralAndes”,enAdvances in Neotropical Mammalogy, K. RedfordyJ.Eisenberg,eds.,pp.265-278.Gainesville,TheSandhill CranePress.
Zetti, Jorge1973.“PaleoetnozoologíadelTastil”,enTastil, una ciudad preincaica argentina,E.M.Cigliano,comp.,
pp.565-578.BuenosAires,Ed.Cabargon.

2 0 0

2 0 1
A RQU EOLOGÍ A SU BACUÁTICA Y TAFONOMÍ A: R ECIENTES AVA NCES EN EL ESTU DIO DE SITIOS FINIPLEISTOCÉNICOS SUM ERGIDOS EN LA COSTA PACÍFICA DE CHILE CENTR A L
isabel carTa jena F.*
[email protected] de Antropología, Universidad de Chile, ChilepaTricio lópez m.**
[email protected] Católica del Norte, IIAM, ChiledieGo car abias a .***
[email protected] Consultores S. A., Chilecarla mor ales****
[email protected] Consultores S. A., ChileGAbrieL VArGAs*****
[email protected] de Geología, Universidad de Chile, Chile
Resumen Se presentan los resultados del análisis tafonómico
del sitio GNLQ1 (32º S – 71º O, Chile) descubierto a 13 m de
profundidad bajo el mar. El sitio corresponde a un contexto
primario conformado por una serie de restos de fauna continental
extinta (familias Camelidae, Cervidae, Cricetidae, Artiodactyla,
Equidae, Mylodontidae, Canidae, así como del Orden Xenarthra)
recuperados en concentraciones dispuestas sobre una unidad
estratigráfica asignada al Pleistoceno final. En este trabajo se
abordan la determinación taxonómica y representación anatómica,
además de las características superficiales del material, con el fin
de conocer las condiciones paleoecológicas y delimitar los factores
tafonómicos en contextos terrestres y subacuáticos.
P A L A B R A S C L A V E :
Arqueología subacuática, tafonomía, fauna extinta, Pleistoceno
final, Chile central.
Artículorecibido:1dejuniode2011|aceptado:11denoviembrede2011|modificado:22denoviembrede2011
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,294pp.ISSN1900-5407,pp.201-225
* DoctoraenArqueologíaAmericana(Altamerikanistik,FreieUniversität,Berlín).**DoctorenAntropología.ProgramadeDoctoradoenAntropología,UniversidadCatólicadelNorte,IIAM,Chile.***Arqueólogo,UniversidaddeChile,Chile.****Doctora,HistoriadelArte,RestauraciónyConservación,UniversidadInternacionalS.E.K.,Chile.*****DoctorenOceanografíayPaleoceanografía,UniversidaddeBordeauxI,Francia.

2 0 2
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.201-225
suB-aquatiC arCheology and taphonomy: reCent adVanCes in the study of suBmerged phinipleistoCeniCs in the paCifiC Coast of Central Chile
ABSTR AC T This paper presents the results
of a taphonomic analysis of the site GNLQ1
(32 º S – 71º W Chile) discovered at 13
m of depth below sea level. The site can
be characterized as a primary context,
conformed by diverse remains of extinct
continental fauna (families Camelidae,
Cervidae, Cricetidae, Artiodactyla, Equidae,
Mylodontidae, Canidae and the Xenarthra
Order) recovered in concentrations arranged
on a stratigraphic unit also assigned
to the Late Pleistocene. We deal with
taxonomic determination, skeletal units’
representations, and surface modifications
of the material. We aim to understand
paleo-ecological conditions and identify
taphonomic factors in terrestrial and sub
aquatic contexts.
K E Y W O R D S :
Underwater Archaeology, Taphonomy, Extinct
Fauna, Late Pleistocene, Central Chile.
arqueologia suBaquatiCa e tafonomia: reCentes aVanços no estudo de lugares finipleistoCêni-Cos suBmersos na Costa paCífiCo do Chile Central
RESUMO Apresentam-se os resultados
da análise tafonômica do lugar GNLQ1
(32º S – 71º O, Chile) descoberto a
13 cm de profundidade baixo o mar.
O lugar corresponde a um contexto
primário conformado por uma série
de restos de fauna continental extinta
(famílias Camelidae, Cervidae, Cricetidae,
Artiodactyla, Equidae, Mylodontidae,
Canidae e assim como da ordem Xenarthra)
recuperados em concentrações dispostas
sob uma unidade estratigráfica atribuída ao
pleistoceno final. Neste trabalho abordam-se
a determinação taxonômica e representação
anatômica, além das características
superficiais do material com o fim de
conhecer as condições paleoecológicas
e delimitar os fatores tafonômicos em
contextos terrestres e subaquáticos.
P A L A V R A S C H A V E :
Arqueologia subaquática, tafonomia, fauna
extinta, pleistoceno final, Chile central.

2 0 3
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.201-225
A RQU EOLOGÍ A SU BACUÁTICA Y TAFONOMÍ A: R ECIENTES AVA NCES EN EL ESTU DIO DE SITIOS FINIPLEISTOCÉNICOS SUM ERGIDOS EN LA COSTA PACÍFICA DE CHILE CENTR A L
i s a b e l c a r T a j e n a F .
p a T r i c i o l ó p e z m .
d i e G o c a r a b i a s
c a r l a m o r a l e s
G A b r i e L V A r G A s
IntroducciónLa arqueología subacuática es una disciplina que ha comenzado a ocupar su propio espacio dentro de los estudios arqueológi-cos en Chile, ya sea mediante el desarrollo de proyectos de investigación con objetivos particulares y claramente definidos, o bien, en el marco de estudios de impacto ambiental (Carabias, 2001, 2005). Las dificultades de trabajar en un medio diametralmente distinto al terrestre, unidas a procesos de forma-ción disímiles a los tradicionalmente estudiados, han llevado a la arqueología subacuática a formular sus propios marcos teóricos y metodológicos y adaptar otras técnicas a partir de los referentes clásicos de la arqueología de campo. No obstante, la excavación en un contexto subacuático no difiere en demasía de la de sitios sobre tierra, siendo similares en el registro visual, las técnicas de exca-vación, la recolecta de muestras y el registro planimétrico, aunque cambian las condiciones y los tiempos de trabajo, debido a las limitantes del medio.
Los avances de la arqueología subacuática en las últimas décadas han posibilitado que esta disciplina haya ido consolidando metodologías propias en el estudio de restos de relevancia arqueológica que actualmente se encuentran sumergidos tanto en ambientes marinos como de agua dulce (Dean et al., 2000; Petriaggi y Davidde, 2007; Masters y Flemming, 1983). Si bien por tradición la arqueología subacuática y marítima ha enfatizado el estudio de contextos his-tóricos, los trabajos en sitios prehistóricos son cada vez más frecuentes y se han enmarcado en problemáticas relacionadas con el uso de rutas costeras dentro

2 0 4
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.201-225
de procesos de poblamiento inicial de una zona, el uso de antiguas embarca-ciones, la aplicación de técnicas de pesca y recolección de recursos marinos y sus correlatos materiales (Blanton, 1996; Clausen et al., 1979; Dunbar et al., 1992; Faught, 1996, 2004a, 2004b; Faught y Donoghue, 1997; Flemming, 2004; Galili et al., 1993; Gusick y Faught, 2011). Así mismo, el estudio de contex-tos paleontológicos y arqueológicos prehistóricos ha permitido recolectar una gran cantidad de información geoarqueológica relacionada con el aumento del nivel del mar, o bien, la inundación de sectores de gran potencial para estudios paleoambientales y geomorfológicos (Dunbar et al., 1992; Faught, 1996; Faught y Donoghue, 1997; Faught, 2004a, 2004b; Flemming, 2004).
Dentro de la arqueología americana, las investigaciones subacuáticas han adquirido cierta relevancia en el estudio del poblamiento inicial del continente, debido al posible uso de la ruta costera (Dillehay, 2009; Goebel et al., 2008), cuyos sitios se encontrarían actualmente sumergidos, producto del avance marino durante el Pleistoceno final, y que suponen un poblamiento previo al denominado “Clovis First” (Bradley y Stanford, 2004; Dillehay, 2009; Faught, 1996, 2004a, 2004b; Goebel et al., 2008; Hoffecker et al., 1993). No obstante, la hipótesis de entrada por el Pacífico (Pacific Rim o Coastal-entry) es un modelo complejo que involucra diversas propuestas. Si bien la ruta por el Pacífico es lógicamente atractiva, la evidencia es escasa. Desde el Último Glacial Máximo, el nivel del mar ha subido hasta 120 m, inundando paisajes costeros y estuarios ribereños donde se podrían haber localizado los sitios. A excepción de Josen-hans et al. (1997), que han logrado localizar sitios en la costa de Queen Char-lotte Islands, se han realizado escasos esfuerzos de búsqueda a lo largo de las líneas costeras sumergidas (Bonnichsen et al., 2005: 321). Si bien la evidencia arqueológica señala la presencia de adaptaciones costeras, también cabe espe-rar la presencia de cazadores terrestres, no en el sentido planteado por Bon-nichsen et al. (2005) en cuanto a la reubicación de asentamientos y un acerca-miento a los recursos costeros ante un escenario cambiante, sino en el marco de la utilización de quebradas y estuarios por parte de grupos interiores, cuyas evidencias quedaron cubiertas posteriormente por el mar.
El reconocimiento de yacimientos en este contexto no involucra única-mente la recolección de datos paleoambientales, geomorfológicos o eviden-cias artefactuales y/o ecofactuales, sino que además, como todo sitio arqueo-lógico temprano, requiere una contrastación rigurosa de la antigüedad real de las ocupaciones humanas, así como de la integridad de las asociaciones esperadas para estos eventos ocupacionales (Stewart, 1999; Waters, 1992). Al respecto, uno de los problemas recurrentes a lo largo de las investigaciones de sitios del Pleistoceno final se centra en una evidencia poco diagnóstica para

2 0 5
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.201-225
A R Q U E O L O G Í A S U B A C U Á T I C A | I . C A R T A J E N A , P . L Ó P E Z , D . C A R A B I A S , C . M O R A L E S Y G . V A R G A S
discriminar eventos naturales y/o culturales en la generación de un yacimiento determinado, de manera específica, entre las evidencias osteofaunísticas de mamíferos actualmente extintos producidas por agentes naturales versus la acción humana. Este problema ha sido abordado con profusión, y la tafonomía constituye un referente metodológico y teórico clave para el entendimiento e interpretación de sitios donde los agentes naturales y culturales se mezclan, o son dif íciles de separar (Bonnichsen, 1982; Borrero, 2003; Frison, 1982; Hill y Behrensmeyer, 1985).
Los proyectos orientados al estudio de sitios tempranos se caracterizan por el trabajo conjunto de numerosos especialistas, con el fin de entender y comprender los conjuntos recuperados. Disciplinas como la geología, geoar-queología, botánica, palinología, botánica y zooarqueología se congregan comúnmente para el estudio de dichos yacimientos. Mediante este trabajo con-junto, no sólo se busca la colaboración y compartir experiencias sino abrir nue-vas perspectivas de análisis.
Reflexiones en torno a una tafonomía en contextos subacuáticosDentro de los estudios tafonómicos, los depósitos de mamíferos de talla mediana a grande han sido abordados en detalle, tanto en el marco de trabajos experimentales (Binford, 1981; Blumenschine, 1995), etnoarqueológicos (Bunn et al., 1988; Bunn, 1993; O’Connell et al., 1990), como mediante observacio-nes actualísticas en ensambles faunísticos modernos (Beherensmeyer, 1978; Borrero y Martin, 1996; De Ruiter y Berger, 2000; Domínguez-Rodrigo, 1999; Haynes, 1983a, 1983b). No obstante, los agentes y procesos tafonómicos estu-diados se enmarcan en contextos terrestres, y escasamente han sido pensados para sitios sumergidos (Stewart, 1999; Stright, 1995; Waters, 1992). Dentro de estas reflexiones destacan las de Dunbar y colaboradores (1989), que proponen, a partir del caso del sitio de Aucilla River (Florida, EE. UU.), al menos cinco tipos de modificaciones que deben ser consideradas en contextos finipleistocénicos sumergidos con restos de fauna, a saber: 1) modificaciones culturales en huesos frescos sincrónicas a la muerte del animal (por ejemplo, huellas de corte), 2) modificaciones naturales previas a la depositación de los huesos (por ejemplo, meteorización, marcas de carnívoros), 3) modificaciones naturales posdeposita-cionales (mineralización, degradación por acción del suelo), 4) modificaciones naturales dentro de la interfaz sedimento-agua (por ejemplo, oxidación, abra-sión) y 5) alteraciones posexcavación producto del cambio de ambiente de los huesos. Esta alteración, sin embargo, es controlable a partir de un debido trabajo de conservación y restauración de las osamentas (Bouzas y Laborde, 2003).

2 0 6
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.201-225
A estas alteraciones agregamos aquellas que se producen en sitios con alta frecuencia de tránsito de barcos de distintos calado como muelles y puer-tos, además de zonas aledañas a pueblos y ciudades costeras. En el primer caso, los muelles y puertos, como la bahía de Quintero, constantemente movilizan y arrastran anclas, cadenas, o “muertos” que corresponden a bloques de concreto que alteran subsuperficialmente el fondo marino. En el caso de las poblaciones humanas asentadas en las cercanías de la costa, éstas producen alteraciones, debido a la incorporación de basura y tubos de desagüe.
En el caso del sitio a estudiar, GNLQ1, ubicado en localidad de Quintero (Latitud: 32º 46’ 59” S; Longitud: 71º 31’ 59” O), en el litoral de Chile central, se encuentra aproximadamente a 13 m de profundidad bajo el mar y presenta características únicas en la costa del Pacífico sur. El sitio GNLQ1 corresponde a un contexto primario conformado por una serie de restos de fauna con-tinental extinta del Pleistoceno final, depositados en sedimentos terrígenos actualmente sumergidos. Dentro de un área de 1 m2 de excavación, se regis-traron diversos restos del esqueleto apendicular y axial de especies extintas pertenecientes a las familias Camelidae, Cervidae, Cricetidae, Artiodactyla, Equidae, Mylodontidae, Canidae y restos del Orden Xenarthra, que fueron recuperados en concentraciones discretas dispuestas sobre una unidad estra-tigráfica caracterizada por una grava arcillosa en una matriz de arena fina a limo y arcillas con presencia de lentes de carbón. Los antecedentes faunísti-cos, geomorfológicos y sedimentológicos sugieren que el contexto durante el Pleistoceno final correspondía a un ambiente tipo estuario-lagunar, con un período de somerización-disecación, a uno de tipo de anteplaya superior, inundado por el mar.
Las características del emplazamiento permiten, de acuerdo con Bishop (1980), adscribirlo a aquellos paleoambientes fosilíferos en regímenes conti-nentales, que permiten la fosilización y conservación. Entre ellos se encuentran las aguas poco profundas de estuario, los deltas y depósitos lagunares asociados a las transgresiones marinas (Bishop, 1980: 23). Además, en un eje temporal, el contexto terrestre representa una escala temporal corta, en la cual es posible observar cambios rápidos en el ambiente, que ocurren y se perciben en el nivel sedimentario. Por otra parte, en su paso a un contexto subacuático, es posible adscribirlo a un ambiente instantáneo (“Instant environment” sensu Bishop, 1980: 22), definido como eventos a corto plazo, que permiten un conocimiento acabado de un evento catastrófico de diversa magnitud. En este medio acuoso, por su parte, el conjunto no presenta las particularidades de restos transporta-dos o expuestos al movimiento de las aguas (Hanson, 1980).

2 0 7
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.201-225
A R Q U E O L O G Í A S U B A C U Á T I C A | I . C A R T A J E N A , P . L Ó P E Z , D . C A R A B I A S , C . M O R A L E S Y G . V A R G A S
Figura1.a)UbicaciónnacionaldelsitioGNLQ1yb)regional.c)UbicacióndelsitioenlabahíadeQuintero.Lospuntosindicanelcontextogeomorfológicodelasmuestras,mientrasquelaestrellaseñalaeltestigodesedi-mentoobtenidodelabahíadeQuintero.Laslíneasgrisesindicanlasisóbatas.Laflechademayortamañoindicaladireccióndepropagacióndelfrentedeolasdesdeelsuroeste;mientrasquelasflechasdelgadasindicansudireccióndepropagaciónluegodeproducirseladifraccióndelfrentedeolasporefectodelapuntadeQuintero(figuratomadaymodificadadeVargasyOrtega,2008).
A través del presente trabajo se busca caracterizar el conjunto faunístico recuperado del sitio GNLQ1, tanto al nivel de la representación de unidades anatómicas como de la diversidad taxonómica, poniendo especial hincapié en el análisis taxonómico, para lo cual se consideraron las características de las superficies, integridad y articulación de unidades. Si bien este análisis es pre-liminar, mediante los resultados obtenidos se busca conocer las condiciones paleoecológicas y delimitar los factores tafonómicos en contextos terrestres y subacuáticos (Behrensmeyer y Hill, 1980). Así mismo, a partir de este análisis se busca generar expectativas para el desarrollo de los análisis zooarqueológicos y tafonómicos en estrecha relación con programas de investigación que intenten formalizar la búsqueda de yacimientos arqueológicos prehispánicos sumergi-dos y su potencial para comprender y caracterizar de mejor manera procesos de poblamiento humano hacia el Pleistoceno final.

2 0 8
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.201-225
El sitio GNLQ1
Como se mencionó anteriormente, el sitio GNLQ1 se ubica en la bahía de Quin-tero, emplazada al norte de Valparaíso, V Región de Chile (ver las figuras 1a, 1b y 1c). Ésta corresponde a una bahía somera, con una profundidad máxima de 55 a 60 m, abierta hacia el noroeste. En ella descargan algunas quebradas con escorrentía de permanente a perenne, que generan lagunas y/o humedales entrampados por barras de arena asociadas a la acción de la hidrodinámica lito-ral (Vargas y Ortega, 2008). El perfil batimétrico muestra una mayor pendiente en los primeros ca. 250 m desde la línea de costa y una menor pendiente hasta los 1.600 m alejados de ésta. El sitio fue descubierto durante las labores de pros-pección y excavación arqueológica, en el marco de un proyecto relacionado con una terminal de gas.
En este contexto se realizó una serie de sondeos subsuperficiales, en puntos distribuidos a lo largo de una transecta trazada dentro de un área de 40 x 25 m (Carabias, 2008). A partir de estas evidencias, se excavó una unidad de 1 x 1 m (Unidad K8) delimitada sobre una concentración de huesos que asomaban del fondo marino (erosionado por las actividades realizadas en el muelle cercano) y que presentaban potencial estratigráfico, de acuerdo con los sondeos realizados previamente. En el caso de la excavación del sitio GNLQ1, se procedió mediante niveles artificiales de 10 cm, registrando todo tipo de evidencias en planta, extra-yendo el sedimento mediante dragado para ser harneado.
Figura2.VistadelsitioGNLQ1.a)Procesodedecapadodeunidadde1x1mconmaterialinsitu.b)Fósilesinsituenconcentraciónerosionadaporactividadesdesarrolladasenelmuelle.
A partir del análisis del testigo se determinaron tres unidades estratigráficas con características sedimentológicas distintas, que por orden de antigüedad son: Unidad 1, Unidad 2 y Unidad 3. La Unidad 1 está constituida por arena fina de buena selección, con un sedimento de color café. La Unidad 2 está constituida por grava arcillosa clasto-soportada de aglomerados de arena fina y arcilla, y eventual-

2 0 9
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.201-225
A R Q U E O L O G Í A S U B A C U Á T I C A | I . C A R T A J E N A , P . L Ó P E Z , D . C A R A B I A S , C . M O R A L E S Y G . V A R G A S
mente calcedonia, de alta dureza, de color naranja y gris, algunas con pátinas de carbón, en una matriz de arena fina a limo y arcillas. En la parte superior e inferior de la unidad se intercalan lentes milimétricos de carbón. La Unidad 3 está cons-tituida por arena fina arcillosa de color café anaranjado (Vargas y Ortega, 2008).
Las evidencias óseas fueron encontradas en la Unidad 2, distribuidas hori-zontalmente sobre una extensa área, directamente asociadas a lentes de carbón presentes en los primeros 5 cm de la Unidad 2 (Vargas y Ortega, 2008; ver las figuras 2a y 2b). Las muestras, luego de extraídas, fueron desalinizadas mediante lavados progresivos en agua desmineralizada. Al respecto, durante la excavación de los sondeos del sitio GNLQ1, los materiales recuperados fueron extraídos manteniendo gran parte del sedimento o matriz que los rodeaba. Este levan-tamiento “en bloque” permitió rigidez y soporte al hueso y sus fragmentos, y minimizó la pérdida de información y deterioro f ísico que pudiera ocasionar la extracción. Considerando la fragilidad de algunos de los restos, se puso especial cuidado en no afectarlos mecánicamente durante la intervención arqueológica. El primer daño que pueden sufrir los materiales orgánicos, cualquiera que éstos sean, es la pérdida de humedad. Para evitarla, se dispuso de recipientes indi-viduales con agua salada para ir introduciendo los objetos a medida que son extraídos del fondo, impidiendo que se produzca la deshidratación de éstos, y así no romper el equilibrio que han logrado estando en el fondo marino. Los materiales óseos, así como la mayor parte de los materiales recuperados de sitios arqueológicos en tierra o bajo el agua, son vulnerables también a los cambios bruscos de temperatura. Para controlar ese tipo de daño los huesos se fueron almacenando, según se iban levantando, en una zona protegida de los rayos sola-res, en tanques de almacenamiento realizados especialmente para este proyecto.
Una vez despejado, el material óseo se depositó en contenedores indivi-duales con agua salada proveniente del mismo sitio. Allí se dio inicio al segundo proceso, quizás el más importante, del que dependen el éxito de la estabiliza-ción de los materiales y su posterior conservación. Este proceso consistió en bajar los niveles de sal que contienen, primero cambiando de manera gradual el agua que los contenía por agua libre de minerales. Este reemplazo se realizó periódicamente, con cambios del porcentaje de agua salada versus agua desti-lada una vez cada diez días, hasta que el 100% fuera agua dulce.
Después de 50 días comenzó el tratamiento de rinsing de los huesos. Éste consistió en un lavado delicado con agua corriente utilizando una escobilla suave para acelerar la limpieza, teniendo cuidado de no rayar la superficie de la pieza. Mientras se realizaban estos lavados, los materiales fueron deposita-dos en contenedores con agua desmineralizada renovada periódicamente en un lugar oscuro y fresco para evitar el desarrollo de microorganismos, como hon-

2 1 0
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.201-225
gos o bacterias. Al igual que en el proceso anterior, se buscó liberar al material óseo de la sal que tenía en su interior; en este caso en particular, los lavados se realizaron una vez por semana durante cuatro meses, hasta que los controles de acidez fueran bajos y pudiera iniciarse el último de los tratamientos, el secado. Estos controles se realizaron midiendo el pH durante cada cambio de agua, y correspondieron a uno de los procesos más importantes durante el tratamiento de conservación (Carabias, 2008).
Ahora bien: para determinar la antigüedad del sitio fueron enviadas dos muestras de hueso de paleocamélidos para su datación por C14 (AMS), pero no presentaban colágeno suficiente. De acuerdo con el ambiente de sedimentación de las Unidades 3 y 2, el sitio GNLQ1 habría sido un estuario (Vargas y Ortega, 2008), y teniendo en cuenta la variación global del nivel del mar, junto a un despreciable alzamiento tectónico para la zona en los últimos miles de años, se podría tentativamente sugerir una edad superior a los 7.000 años AP, para la formación de la Unidad 2, puesto que el nivel del mar se encontraba alrededor de 10-20 m más bajo que su posición actual, entre los 7.000-9.000 años AP, respectivamente (Lambeck et al., 2002). Lo anterior ha quedado ratificado a partir de una muestra de sedimentos de la parte inferior de la matriz en donde se encontraban los restos (Unidad 2) bajo los restos fósiles, la cual aportó una fecha por C14 de 13.640±40 años AP sin calibrar (UGAMS#9194).
Resultados
Características del conjunto óseoSe analizó un total de 224 especímenes óseos (ver la tabla 1), que actualmente se encuentran depositados en Departamento de Antropología de la Univer-sidad de Chile. Los restos provienen mayoritariamente de la cuadrícula K8-2 (Nivel 1, Concentraciones 1, 3 y 6), y, como se mencionó anteriormente, previo a cualquier análisis fueron sometidos a largos procesos de estabilización y con-servación producto del cambio brusco de ambiente (Carabias, 2008).
El conjunto óseo se encuentra en muy buenas condiciones de conserva-ción. No obstante, algunos especímenes evidencian estados iniciales de meteo-rización como agrietamientos longitudinales en fragmentos de diáfisis, lo que sugiere una exposición a condiciones subáreas previa a su enterramiento final. Esta meteorización podría haber favorecido la fragmentación del conjunto, en especial astillas (27% de la muestra total) y fragmentos de huesos planos (17% de la muestra total). Pese a esto, un porcentaje no menor pudo ser remontado, pese a la erosión de los bordes de los huesos producto de la constante acción del agua y sedimentos abrasivos propios del ambiente de depositación del sitio.

2 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.201-225
A R Q U E O L O G Í A S U B A C U Á T I C A | I . C A R T A J E N A , P . L Ó P E Z , D . C A R A B I A S , C . M O R A L E S Y G . V A R G A S
Determinación taxonómicaPara la determinación taxonómica de los restos se utilizaron colecciones de refe-rencia albergadas en el Departamento de Antropología de la Universidad de Chile correspondientes a équidos, camélidos y cérvidos actuales. Así mismo, se utilizó la colección paleontológica del sitio Kamac Mayu (II Región, Chile) depositada en el Laboratorio de Estudios de Arqueología y Patrimonio (LEAP) de la Univer-sidad Internacional SEK-Chile. Debido a las cercanías y similitud faunística con el sitio GNLQ1, se utilizaron las colecciones del sitio Quereo (IV Región), que están actualmente depositadas en el Museo Arqueológico de La Serena.
Para los restos de Canidae fueron utilizadas colecciones particulares de Pseudalopex griseus y Lycalopex culpaeus recolectadas en las cercanías al área de estudio, y que tenían por objeto caracterizar las diferencias de tamaño y morfología. Finalmente, en el caso de los restos de xenartros, el material paleon-tológico disponible en Chile para fines comparativos es escaso; sin embargo, existe en el Museo Nacional de Historia Natural de Santiago material bastante completo de Megatherium medinae y Glossotherium, que fue utilizado para la identificación anatómica de los restos. Debido a la relevancia del sitio, la fase de asignación taxonómica del material de GNLQ1 aún se encuentra en proceso, por lo que en el presente trabajo se discutirán y presentarán datos generales relacionados con este tema.
Clase MammaliaEsta categoría taxonómica es la que presenta la mayor representación de res-tos dentro del conjunto (ver la figura 4). La mayor parte de los especímenes corresponden a astillas, fragmentos de huesos planos (muchos de ellos restos de costillas), fragmentos mínimos y algunas unidades anatómicas identificables como fragmentos de vértebras y epífisis humeral y femoral (tabla 1). Los restos
Tabla1.RestosasignadoscomoMammaliadelsitioGNLQ1.
Mammaliaindeterminados NISP %NISP
Astillas 61 41,2
Fragmentosdehuesosplanos 40 27,0
Fragmentosmínimos 31 20,9
Fragmentosdehuesoslargos 2 1,4
Identificables 14 9,5
Total 148 100,0

2 1 2
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.201-225
corresponderían a grandes mamíferos, encontrándose tanto fragmentos perte-necientes a individuos adultos como juveniles (no fusionados).
Clase avesEstos restos fueron registrados en una cuadrícula diferente (K7), en la cual la presencia de huesos era mínima. Se recuperaron un fémur y una tibia de ave correspondientes a un individuo joven, lo que dificulta su asignación taxonó-mica. Sin embargo, la morfología y tamaño es compatible con especímenes de la familia Phasanidae.
Orden ArtiodactylaSe recobró un premolar deciduo de tipo selenodonte propio de un artiodáctilo. De igual forma, se registró un fragmento de cráneo (meato auditivo). Ambos restos, ante la ausencia de otros rasgos diagnósticos, se asignaron a esta cate-goría taxonómica.
Figura3.RestosóseosrecuperadosdelsitioGNLQ1:a)fragmentodemandíbuladeCervidae;b)calcáneodecánido;c)caninodecánido;d)falangeunguealdexenanrtro;e)molardeéquidojuvenil;f)húmeroyradioulnaarticuladosdecamélido;yg)húmerodecamélidoconflechaquedenotalaescotadurayfracturaenladiáfisis.

2 1 3
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.201-225
A R Q U E O L O G Í A S U B A C U Á T I C A | I . C A R T A J E N A , P . L Ó P E Z , D . C A R A B I A S , C . M O R A L E S Y G . V A R G A S
Familia Camelidaecf. Palaeolama sp.El total de restos recuperados es de 23 especímenes. Éstos corresponden prin-cipalmente a la extremidad anterior superior (húmero distal y radioulna), un cuarto carpiano y fragmentos de tibia (ver las figuras 3f, 3g y 5). La conservación de la epífisis permitió la obtención de medidas, que, comparadas con especíme-nes de guanaco (Lama guanicoe) y Palaeolama sp. provenientes de depósitos del Pleistoceno final de la zona aledaña de Los Vilos (31º S), denotan un camélido de gran tamaño, comparable con los géneros Palaeolama y Hemiauchenia.
En el sitio GNLQ1 no fueron recuperados materiales que permitieran una asignación certera al género Palaeolama. Los restos recuperados (húmero distal y radioulna) son similares en tamaño y morfología al material recobrado del sitio de Quereo (López et al., 2004). De acuerdo con esto, se ha clasificado el material de Quintero como cf. Palaeolama, en espera de un registro más completo.
Familia CervidaeEl material analizado corresponde a fragmentos de la mandíbula izquierda con la serie dental completa, es decir, Pm2, Pm3, Pm4, M1, M2 y M3, además del I3, Pm2, Pm3, Pm4 derechos de un mismo individuo (ver la figura 3a). En comparación con Hippocamelus antisensis (taruca del norte o huemul), los restos de Quintero
Figura4.HistogramaconlaabundanciataxonómicadelsitioGNLQ1,expresadaen%NISP.

2 1 4
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.201-225
son de mayor tamaño, acercándose a géneros como Antifer y Blastocerus. Ahora bien, dados los escasos antecedentes sobre la sistemática de cérvidos extintos en territorio chileno (Casamiquela, 1984, 1999; Labarca y Alcaraz, 2011), se ha optado por asignar el material en estudio como Cervidae gen. et sp. indet.
Familia Equidae El material estudiado corresponde a un P2 y un P3, ambos derechos, de un ani-mal juvenil de aproximadamente dos años (figura 3e). La superficie oclusal, en el caso del P3, prácticamente no presenta desgaste y comienza a denotar el pro-tocono, aunque con un pliegue caballuno casi inexistente. Si bien la forma del protocono tiende a ser ovalada, el escaso desarrollo de diente impide asignarlo con certeza al género Hippidion. Debido a esto, han sido asignadas ambas pie-zas como Equidae gen. et sp. indet.
Orden XenarthraEl registro fósil de xenartros recobrados del sitio GNLQ1 corresponde a un osteodermo, la secuencia completa de los molariformes de la mandíbula dere-cha aparentemente de un mismo individuo y una falange ungueal (figura 3d).
En el caso de los molariformes, las superficies oclusales por forma son afines a especies de la Subfamilia Scelidotheriinae. Por su parte, la ausencia de
Figura5.Histogramaconlarepresentacióndeelementosóseos(MNE),comparadosconlosespecímenesóseos(NISP)decf.Palaeolamasp.

2 1 5
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.201-225
A R Q U E O L O G Í A S U B A C U Á T I C A | I . C A R T A J E N A , P . L Ó P E Z , D . C A R A B I A S , C . M O R A L E S Y G . V A R G A S
rasgos diagnósticos de la falange ungueal impide por el momento asignarlo a una Familia específica. Por su parte, los osteodermos están presentes tanto en especies de la Familia Mylodontidae como en algunos ejemplares de Glyptodon (Rinderknecht, 2000). Este material se encuentra en proceso de estudio, y, de acuerdo con resultados preliminares, la totalidad del registro ha sido clasificada como Xenarthra gen. et sp. indet., en espera de mayores comparaciones.
Familia Canidae El material de Quintero corresponde a un calcáneo y a un canino cuya morfo-logía y cuyo tamaño son similares al actual Lycalopex culpaeus. No obstante, debido a lo escaso del material, se ha asignado el registro del sitio GNLQ1 como Canidae gen. et sp. indet. (figuras 3b y 3c).
Familia CricetidaeSe recuperó un incisivo de un roedor de un Cricetidae de baja diagnosis taxonómica.
Tafonomía
Alteraciones posdepositacionales del sitioTal como se mencionó en párrafos anteriores, el sitio GNLQ1 se encuentra emplazado en una bahía somera que ha funcionado a lo largo de su historia como puerto natural para embarcaciones de distinto calado, y en la actualidad es un muelle artificial usado como terminal para el ingreso de gas licuado al país. A causa de ello, recalan en las cercanías del sitio embarcaciones de gran tamaño, lo que ha motivado a definir un área de exclusión alrededor del sitio para su debida protección (Carabias, 2008). Sin embargo, en el momento de descubrirse el yacimiento GNLQ1 se observó una serie de alteraciones meno-res, que afectaron mediante erosión los sedimentos superiores que sellaban el sitio y, por tanto, han acelerado la exposición superficial de algunas concen-traciones de restos fósiles. Esta erosión se ha producido por la acción mecá-nica sobre el fondo marino de trabajos submarinos desarrollados en el marco de la construcción del muelle producto del traslado de “muertos”, bloques de cemento y cadenas que son llevados en remolques mediante embarcaciones en superficies (Carabias, 2008). Otro proceso de alteración que tiene una larga data en la bahía es el fondeo de anclas que constantemente remueven el fondo del muelle. Si bien estas alteraciones son continuas y han afectado mediante la erosión ciertos sectores del sitio, no han movilizado mayormente restos óseos, por cuanto se ha observado en dichas concentraciones cierta integridad anató-

2 1 6
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.201-225
mica, debido a que unidades de un mismo individuo que se articulan entre sí se disponen espacialmente en un mismo espacio acotado.
Otras alteraciones posdepositacionales corresponden a la incorporación de basura actual sobre la capa superior del sitio. Estas incorporaciones han sido discriminadas sin problemas, debido a que corresponden a desechos históricos absolutamente diacrónicos con el depósito pleistocénico.
Alteraciones tafonómicas en el registro óseo
AbrasiónEntre las alteraciones tafonómicas comunes para la totalidad de la muestra, o buena parte de ésta, se observa un leve grado de abrasión de la superficie de los huesos. Esta abrasión es homogénea y se manifiesta como un pulimento parejo tanto en los bordes como en las superficies planas de las osamentas. Esta alte-ración es esperable, debido a las condiciones de enterramiento de los huesos en un ambiente acuoso y de sedimentos abrasivos, aunque no se afectaron de forma muy agresiva, puesto que es posible distinguir rasgos como marcas pro-ducidas por distintos agentes biológicos (carnívoros y roedores), y aún pueden remontarse sin problemas astillas pertenecientes a un mismo elemento óseo, tal como se señaló anteriormente.
Alteraciones químicasUna segunda alteración común a todos los restos es la presencia de manchas de oxidación producto de la interacción de los huesos con una interfaz de sedi-mentos de grava-arcillosa y agua (ver alteración química en la tabla 2). Este tipo de alteración es frecuente en ambientes similares a los del sitio GNLQ1 (Dunbar et al., 1989; Noakes et al., 2009), por lo que se la debe considerar a futuro como una expectativa dentro de las alteraciones tafonómicas. Por su parte, la presencia de sal en el agua marina y, por ende, en los huesos fue con-trolada mediante lavados continuos con agua desmineralizada, como se men-cionó anteriormente.
La meteorización afectó al 87,5% de la muestra (ver la tabla 2). Esta altera-ción es leve y se concentra en los estadios 1 a 2, según la escala de Behrensme-yer (1978). Así mismo, indica que los restos quedaron expuestos a condiciones subaéreas, previo a su enterramiento y posterior inundación. Cabe destacar que los restos de cf. Palaeolama presentan un estadio de meteorización más avan-zado que el resto de la muestra (Estadio 2), que no sólo indica tiempos más prolongados de exposición, sino que también sugiere que en el sitio GNLQ1 se formó a partir de distintos eventos de enterramiento.

2 1 7
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.201-225
A R Q U E O L O G Í A S U B A C U Á T I C A | I . C A R T A J E N A , P . L Ó P E Z , D . C A R A B I A S , C . M O R A L E S Y G . V A R G A S
RoedoresLas únicas marcas producidas por roedores se concentran en el calcáneo de cánido (ver la figura 6a). Esta evidencia presenta características propias del paso de incisivos de roedores dentro de una zona acotada en la diáfisis del cal-cáneo, a modo de líneas delgadas, cortas y paralelas.
Tabla2.AlteracionestaxonómicasobservadasenelconjuntoóseodelsitioGNLQ1expresadasenNISP.
NISP Meteorización Abrasión Alt.química Carnívoros Roedores Radículas
Total 224 196 222 222 13 1 220
RadículasLa mayor parte del registro óseo presenta en su superficie evidencias de impron-tas de raíces (ver la tabla 2 y la figura 6b). La presencia de estas improntas en los restos no sólo indica el crecimiento de una cubierta vegetacional sobre los depósitos fósiles previa a la transgresión marina, sino que además es un buen indicador que sugiere cierta estabilidad del sitio. Al respecto, las improntas se distribuyen marcadamente sobre la superficie superior de las osamentas y decrecen en la superficie inferior, e indican que los huesos no se movilizaron dentro del estrato portador, previa a la inundación del sitio.
CarnívorosLa acción de carnívoros se observó en el 5,8% de la muestra total. Pese a lo bajo de la incidencia de carnívoros en la alteración del conjunto óseo, estos agen-tes son significativos, ya que no sólo aportan información referente a los pro-cesos de formación del sitio sino que además sirven para una reconstrucción paleoecológica del ensamble faunístico.
Ahora bien, de la totalidad del conjunto, los especímenes de cf. Palaeo-lama sp. presentan la mayor cantidad de marcas producidas por carnívoros. Por un lado, la epífisis distal de húmero presenta dos perforaciones circula-res, que podrían ser atribuidas a la acción de un carnívoro de gran tamaño, y que muestran características similares a las observadas en restos de mamí-feros extintos de diversos sitios registrados en la costa de la zona aledaña de Los Vilos (López, 2007). En la diáfisis de este hueso se observa una fractura transversal, que guardaría relación con la acción del carnívoro, puesto que en la superficie dorsal de la diáfisis y en el borde de la fractura se observan dos escotaduras cuyo agente podría ser el mismo carnívoro de gran tamaño, ya

2 1 8
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.201-225
que presentan una distancia (ca. 35 mm) similar a las perforaciones registra-das en la epífisis.
Tal como se señaló en el párrafo anterior, este tipo de marcas presen-tes en especies de similar tamaño y peso que Palaeolama en distintos sitios paleontológicos y arqueológicos de la zona de Los Vilos fueron atribuidas por López (2007) a un carnívoro de gran tamaño, específicamente a Panthera onca mesembrina. En el caso del sitio GNLQ1, no se ha identificado con claridad el agente específico de las alteraciones producidas en los restos de cf. Palaeolama, aunque parecen indicar un predador cuya talla supera a la de los cánidos, y se relacionan más bien, por morfología y tamaño, con las de un gran carnívoro (Borrero y Martin, 1996). Estas marcas fueron observadas también en la epífisis distal del radio de cf. Palaeolama, que por su tamaño y proximidad espacial parece corresponder al mismo individuo al que pertenece el resto del material asignado a este camélido extinto.
Dentro de los daños producidos por carnívoros se identificaron en frag-mentos óseos y cuerpos vertebrales no identificados marcas de menor tamaño comparadas con las descritas para los restos de cf. Palaeolama sp., y que dan cuenta de la acción de pequeños carnívoros. Éstas son compatibles con la mor-fología y características de las modificaciones de cánidos, cuyos restos están presentes en el sitio GNLQ1 (ver la figura 6c).
Figura6.MarcastafonómicasenrestosóseosdelsitioGNLQ1:a)marcaderoedoresenhuesodeCanidae;b)improntasderadículasenhúmerodecf.Palaeolama;c)marcadecarnívoroencostillademamíferoindeterminado.

2 1 9
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.201-225
A R Q U E O L O G Í A S U B A C U Á T I C A | I . C A R T A J E N A , P . L Ó P E Z , D . C A R A B I A S , C . M O R A L E S Y G . V A R G A S
Evidencias culturalesEn los restos de cf. Palaeolama sp., el húmero distal presenta una fractura sobre hueso fresco. No obstante, esta fractura se asocia con perforaciones pro-ducidas por la acción de un gran carnívoro, por lo que es probable que este agente haya sido el causante de los daños observados en el húmero (Borrero y Martin, 1996; Haynes, 1983a). Si bien, este tipo de fracturas fue considerado propio de las actividades humanas, diversos análisis determinaron el grado de ambigüedad de este rasgo, ya que también aparecen en contextos formados por agentes naturales (Haynes, 1983a, 1983b; Myers et al., 1980). Cabe desta-car, no obstante, el notable parecido de este espécimen con un húmero distal proveniente del cercano sitio de Quereo, donde se observa una fractura muy similar y que ha sido interpretada como cultural (López et al., 2004). Pese a esto, las diferencias observadas en el caso de la muestra de Quereo tienen relación con la morfología y los rasgos asociados al borde de fractura, puesto que se observaron escotaduras y negativos de impacto producidos por el golpe directo sobre la diáfisis del fósil de Quereo, que en el húmero del sitio GNLQ1 no se observan con claridad y además se asocian a las marcas de carnívoros descritas anteriormente.
La totalidad de la evidencia ósea no permite atestiguar de manera clara la acción antrópica en el sitio GNLQ1. No obstante, el análisis sedimentológico de la Unidad 2, es decir, la capa portadora del registro fósil, evidenció lentes milimétricos de carbón cuyo origen podría deberse eventualmente a combus-tiones locales producidas en una ambiente de sedimentación de tipo somero a estuarial-lagunar (Vargas y Ortega, 2008).
Ahora bien: en las superficies óseas se observaron manchas producidas por la oxidación propia de la interfaz del sedimento con el agua, aunque en algunos ejemplares estas manchas parecían mezclarse en algunos huesos con pátinas carbonosas dif íciles de separar macroscópicamente. Análisis mediante Microscopio Electrónico de Barrido (MEB) y Espectroscopía de Energía Dis-persiva (EDS) indican que la totalidad de estas manchas son producto de alte-raciones diagenéticas, debido a la presencia de óxidos de hierro.
Discusión y conclusionesLa información geomorfológica, sedimentaria y paleontológica del sitio GNLQ1 sugiere la presencia de un contexto primario con escasas alteraciones posteriores al depósito de los restos óseos. La unidad estratigráfica portadora de los restos fósiles, denominada Unidad 2, sugiere un ambiente estuarial-lagunar en proceso de desecación, el cual dio paso a un ambiente de anteplaya superior inundada por el mar. En relación con las dataciones del evento de enterramiento del con-

2 2 0
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.201-225
junto fósil, si se consideran la variación global del nivel del mar y el alzamiento tectónico de la zona en los últimos miles de años, se sugiere una edad superior a los 7.000 años AP para la formación de la Unidad 2, puesto que el nivel del mar se encontraba alrededor de 10-20 m más bajo que su posición actual, entre los 7.000-9.000 años AP, respectivamente (Vargas y Ortega, 2008). Esta observación es consistente con el fechado de C14 obtenido de una muestra de la matriz sedi-mentaria sobre la cual se encuentra depositado el conjunto óseo.
Del análisis tafonómico se desprenden diversos agentes que alteraron el registro previo, durante y posterior a su ingreso a la litósfera. Al respecto, se identificó la acción de al menos dos tipos de carnívoros sobre los restos, deno-tados por el tamaño y distancia de las punturas presentes. Uno de los agentes modificadores correspondería a un carnívoro de gran tamaño, como lo indi-can las marcas dejadas en huesos largos de cf. Palaeolama sp. Por otra parte, se observaron punturas más pequeñas compatibles con una arcada similar a la dejada por cánidos. En menor medida, la acción de roedores se manifiesta claramente en las marcas observables en un calcáneo perteneciente a un carní-voro. Por su parte, la presencia de improntas de raicillas indica el crecimiento de una cubierta vegetal, lo que denota que los restos, una vez expuestos y antes de la transgresión marina, habrían sido cubiertos por sedimentos, lo que favo-reció la preservación e integridad del registro.
Dentro de las alteraciones producidas después del ingreso de los hue-sos en los sedimentos, la más elocuente corresponde a las manchas oxidación producto de la interacción de los huesos con una interfaz de sedimentos de grava-arcillosa y agua, que pueden ser consideradas como propias de ambien-tes depositacionales similares a los del sitio GNLQ1.
Por último, si bien no se identificaron restos culturales artefactuales ni en los sondeos ni en superficie, ni claros rastros de consumo humano de la fauna estudiada, no debe descartarse la acción del hombre en parte de la conforma-ción del sitio. En relación con esta hipótesis, las investigaciones desarrolladas en la costa del norte semiárido chileno señalan una alta densidad de sitios con mamíferos extintos distribuidos en ambientes lagunares y ecosistemas estua-rinos que se formaron a partir de eventos de mortandad natural de especies actualmente extintas, ya sea mediante la incorporación de carcasas y huesos dejados por carroñeros y carnívoros, o bien, por la acción predadora de las pri-meras poblaciones humanas asentadas en la zona (Jackson et al., 2005; López et al., 2004; López, 2007). El resultado de la intervención de estos tres agentes se encuentra muchas veces conjuntamente en un mismo sitio, como en el caso del sitio de Quereo y en otros sitios subsuperficiales del semiárido, observán-dose incluso posibles evidencias de carroñeo humano sobre restos dejados por

2 2 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.201-225
A R Q U E O L O G Í A S U B A C U Á T I C A | I . C A R T A J E N A , P . L Ó P E Z , D . C A R A B I A S , C . M O R A L E S Y G . V A R G A S
grandes carnívoros (López, 2007; Núñez et al., 1994). A esto se suma que la evidencia artefactual es mínima y se complementa o prioriza el uso de huesos como artefactos expeditivos para el procesamiento de las carcasas (Jackson et al., 2007; López, 2007; Núñez et al., 1994).
Estos antecedentes proponen un escenario que debe evaluarse en futuras investigaciones y sugieren que las evidencias del sitio GNLQ1 no son exclu-yentes de la posible acción antrópica dentro del conjunto óseo, aspecto que dependerá de un mayor cúmulo de evidencias.
Sin lugar a dudas, este hallazgo es único por su naturaleza y estado de conservación; sin embargo, su similitud con otros yacimientos hace necesario ahondar los análisis, puesto que habría una serie de indicadores como la diver-sidad taxonómica, la concentración de los restos, la presencia de fracturas en hueso frescos, entre otros, que nos llevan a indagar su posible asociación con ocupaciones humanas.
AgradecimientosEl presente trabajo fue financiado por GNL Quintero S. A. y Arka Consulto-res. Los autores desean agradecer al arqueólogo Joaquín Vega por la informa-ción y ayuda aportada. Especial agradecimiento a los organizadores del Primer Encuentro Latinoamericano de Zooarqueología, en particular a la doctora Eli-zabeth Ramos, por la ayuda y facilidades brindadas. Por último, agradecemos a los evaluadores anónimos, que ayudaron a mejorar el presenta manuscrito. Los errores de este trabajo son de exclusiva responsabilidad de los autores. .

2 2 2
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.201-225
R e f e R e n c i a s
Behrensmeyer, Anna Kay 1978.“TaphonomicandEcologicInformationfromBoneWeathering”,Paleobiology4(2),pp.150-62.
Behrensmeyer, Anna Kay y Andrew Hill1980.Fossils in the Making: Vertebrate Taphonomy and Paleoecology.Chicago,Universityof
ChicagoPress.Binford, Lewis1981.Bones: Ancient Men and Modern Myths.NuevaYork,AcademicPress.
Bishop, Walter W.1980.“PaleogeomorphologyandContinentalTaphonomy”,en Fossils in the Making,eds.AnnaK.
BehrensmeyeryAndrewP.Hill,pp.20-40.Chicago,PrehistoricArchaeologyandEcologySeries,UniversityofChicagoPress.
Blanton, Dennis 1996.“AccountingforSubmergedMid-HoloceneArchaeologicalSitesintheSoutheast.ACaseStudy
fromtheChesapeakeBayEstuary,Virginia”,en Archaeology of the Mid-Holocene Southeast, eds.KennethE.SassamanyDavidG.Anderson,pp.200-217.Gainesville,UniversityPressofFlorida.
Blumenschine, Robert J. 1995.“PercussionMarks,ToothMarks,andExperimentalDeterminationsoftheTimingofHominid
andCarnivoreAccesstoLongBonesatFLKZinjanthropus,OlduvaiGorge,Tanzania”,Journal of Human Evolution 29,pp.21-51.
Bonnichsen, Robson 1982.“BoneTechnologyasaTaphonomicFactor:AnIntroductoryStatement”,Canadian Journal of
Anthropology2(2),pp.137-144.
Bonnichsen, Robson, Bradley Lepper, Dennis Stanford y Michael Waters2005. Paleoamerican Origins: Beyond Clovis.Orono,CenterfortheStudyoftheFirstAmericans,Texas
UniversityPress.
Borrero, Luis A.2003.“TaphonomyoftheTresArroyosRockshelter”,Quaternary International109-110,pp.87-93.
Borrero, Luis A. y Fabiana Martin1996.“Tafonomíadecarnívoros:unenfoqueregional”,en Arqueología Solo Patagonia. Segundas
Jornadas de Arqueología de la Patagonia,ed.JulietaGómezOtero,pp.189-198.PuertoMadryn,CentroNacionalPatagónico.
Bouzas, Ana y Ana Laborde2003.“Ladegradacióndelhueso”,Monte Buciero 9, pp.267-276.
Bradley, Bruce y Dennis Stanford2004.“ThenorthAtlanticIce-edgeCorridor:APossiblePaleolithicRoutetotheNewWorld”,World
Archaeology36(4),pp.459-478.
Bunn, Henry T. 1993.“BoneAssemblagesatBaseCamps:AFurtherConsiderationofCarcassTransportandBone
DestructionbytheHadza”,en From bones to behavior: Etnoarchaeological and Experimental Contributions to the Interpretation of Faunal Remains,ed.JeanHudson,pp.156-68.Illinois,SouthernIllinoisUniversityatCarbondale.
Bunn, Henry. T., Laurence Bartram y Ellen Kroll1988.“VariabilityinBoneAssemblageFormationfromHadzaHunting,Scavenging,andCarcass
Processing”,Journal of Anthropological Archaeology 7,pp.412-457.

2 2 3
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.201-225
A R Q U E O L O G Í A S U B A C U Á T I C A | I . C A R T A J E N A , P . L Ó P E Z , D . C A R A B I A S , C . M O R A L E S Y G . V A R G A S
Carabias, Diego2001.“PrimerasaproximacionesatravésdelaarqueologíasubacuáticaalpasadoprehistóricodelNorte
chico:PuntaÑagué”,Boletín de la Sociedad Chilena de Arqueología 31,pp.39-41.2005.“PatrimonioculturalsubacuáticodeChile.ElrescatedeunlegadosumergidoenChile”,enPaís
oceánico,pp.124-131.Santiago,OchoLibrosEditores.
Carabias, Diego2008.“Informedeevaluaciónarqueológicasubacuáticaeimplementacióndeáreadeexclusión
preventivaentornoalsitioGNLQ1”,enProyecto Terminal de Gas Natural Licuadi (GNL) en Quintero, V Región, Comuna de Quintero.Valparaíso.
Casamiquela, Rodolfo1984.“CriticalCatalogueofSomeChileanFossilVertebratesI.TheDeers:Complementary
ConsiderationsonAntifer (Antifer niemeyeri n.sp.),thePleistoceneGiantDeer”,Quaternary of South America & Antarctic Peninsula 2,pp.41-50.
1999.“ThePleistoceneVertebrateRecordofChile”,Quaternary of South America & Antarctic Peninsula 7,pp.91-107.
Clausen, Carl J., Arthur Cohen, Cesare Emiliani, Alan Holman y Jerry Stipp1979.“LittleSaltSpringFlorida:AUniqueUnderwaterSite”,Science 203(4381),pp.609-614.
De Ruiter, Darryl y Lee Berger2000.“LeopardsasTaphonomicAgentsinDolomiticCaves.ImplicationsforBoneAccumulationsinthe
Hominid-bearingDepositsofSouthAfrica”,Journal of Archaeological Science 27,pp.665-684.
Dean, Martin, Ben Ferrari, Ian Oxley, Mark Redknap y Kit tie Watson2000. Archaeology Underwater. The NAS Guide to Principles and Practice.Londres.EditorialArchetype
PublicationsLtd.
Dillehay, Tom 2009.“ProbingDeeperintoFirstAmericanStudies”,Proceedings of the National Academy of Sciences
106(4),pp.971-978.
Domínguez-Rodrigo, Manuel 1999.“FleshAvailabilityandBoneModificationsinCarcassesConsumedbyLions”,Palaeogeography,
Palaeoclimatology, Palaeoecology149,pp.373-388.
Dunbar, James, David Webb y Dan Cring1989.“CulturallyandNaturallyModifiedBonesfromaPaleoindianSiteintheAucillaRiver,North
Florida”,en Bone Modifications,eds.RobsonBonnichsenyMarcellaSorg,pp.473-497.Orono,CenterfortheStudyoftheFirstAmericans.
Dunbar, James, David Webb y Michael Faught 1992.“InundatedPrehistoricSitesinApalacheeBay,Florida,andtheSearchfortheClovisShoreline”,
en Paleoshorelines and Prehistory: An Investigation of Method,eds.LucilleJohnsonyMelanieStright,pp.117-148.AnnArbor,CRCPress.
Faught, Michael 1996.“ClovisOriginsandUnderwterPrehistoricArchaeologyinNorthwesternFlorida”.Ph.D.
dissertation,UniversityofArizona,Tucson.UniversityMicrofilms,AnnArbor.2004a.“TheUnderwaterArchaeologyofPaleolandscapes,ApalacheeBay,Florida”,American Antiquity
69(2),pp.275-289.2004b.“SubmergedPaleoindianandArchaicSitesoftheBigBend,Florida”,Journal of Field
Archaeology 29(3-4),pp.273-290.

2 2 4
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.201-225
Faught, Michael y Joseph Donoghue 1997.“MarineInundatedArchaeologicalSitesandPaleofluvialSystems:ExamplesfromaKarst-
controlledContinentalShelfSettinginApalacheeBay,NortheasternGulfofMexico”,Geoarchaeology 12(5),pp.417-458.
Flemming, Nic C. 2004.“SubmarinePrehistoricArchaeologyoftheIndianContinentalShelf:APotentialResource”,
Current Science 86(9),pp.1225-1230.
Frison, George 1982.“BoneButcheringToolsinArchaeologicalSites”,Canadian Journal of Anthropology 2(2),
pp.159-167.
Galili, Ehud, Mina Weinstein-Evron, Israel Hershkovitz, Avi Gopher, Kislev Mordecai, Omri Lernau, Liora Kolska-Horwitz y Hanan Lernau
1993.“Atlit-Yam:APrehistoricSiteontheSeaFloorOfftheIsraeliCoast”,Journal of Field Archaeology 20(2),pp.133-157.
Goebel, Ted, Michael Waters y Dennis O’Rourke2008.“TheLatePleistoceneDispersalofModernHumansintheAmericas”,Science 319,pp.1497-1502.
Gusick, Amy y Michael Faught2011.“PrehistoricArchaeologyUnderwater:ANascentSubdisciplineCriticaltoUnderstandingEarly
CoastalOccupationsandMigrationRoutes”,enTrekking the Shore, Changing coastlines and the antiquity of coastal settlements,eds.NunoF.Bicho,JonathanA.HawsyLorenG.Davis,pp.27-50.NuevaYork,Springer.
Hanson, C. Bruce1980.“FluvialTaphonomicProcesses:ModelsandExperiments”,enFossils in the Making, eds.AnnaK.
BehrensmeyeryAndrewP.Hill,pp.156-181.PrehistoricArchaeologyandEcologySeries.Chicago,UniversityofChicagoPress.
Haynes, Gary 1983a.“AGuideforDifferentiatingMammalianCarnivoresTaxaResponsableforGnawDamageto
HerbivoreLimbBones”,Paleobiology9(2),pp.164-172.1983b.“FrequenciesofSpiralandGreenboneFracturesonUngulateLimbBonesinModernSurface
Assemblages”,American Antiquity 48,pp.102-114.
Hoffecker, John, Roger Powers y Ted Goebel1993.“TheColonizationofBeringiaandthePeoplingoftheNewWorld”,Science 259,pp.46-53.
Hill, Andrew y Anna Kay Behrensmeyer 1985.“NaturalDisarticulationandBisonButchery”,American Antiquity 50(1),pp.141-145.
Jackson, Donald, César Méndez, Patricio López, Douglas Jackson y Roxana Seguel2005.“EvaluacióndeunasentamientoarqueológicoenelsemiáridodeChile:procesosdeformación,
faunaextintaycomponentesculturales”,Intersecciones en Antropología6,pp.139-152.
Jackson, Donald, César Méndez, Roxana Seguel, Antonio Maldonado y Gabriel Vargas2007.“InitialOccupationofthePacificCoastofChileduringLatePleistoceneTimes”,Current
Anthropology48(5),pp.725-731.
Josenhans, Heiner, Daryl Fedje, Reinhard Pienitz y John Southon1997.“EarlyHumansandRapidlyChangingHoloceneSeaLevelsintheQueenCharlotteIslands-Hecate
StraitBritishColumbia,Canada”,Science 277,pp.71-74.
Labarca, Rafael y M. Alejandra Alcaraz2011.“PresenciadeAntiferultraAmeghino(=AntiferniemeyeriCasamiquela)(Artiodactyla,Cervidae)enel
Pleistocenotardío-HolocenoTempranodeChileCentral(30-35°S)”,Andean Geology 38,pp.156-170.

2 2 5
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.201-225
A R Q U E O L O G Í A S U B A C U Á T I C A | I . C A R T A J E N A , P . L Ó P E Z , D . C A R A B I A S , C . M O R A L E S Y G . V A R G A S
Lambeck, Kurt, Emma-Kate Potter y Tezer Esat2002.“LinksbetweenClimateandSeaLevelsforthePastThreeMillionYears”,Nature 419,pp.199-206.
López, Patricio2007.“TafonomíadelosmamíferosextintosdelPleistocenoTardíodelacostameridionaldelsemiárido
deChile(IVRegión-32°latitud):alcancesculturalesypaleoecológicos”,Chungara39(1), pp.69-86.
López, Patricio, Rafael Labarca y Lautaro Núñez2004.“NivelQuereoI:unadiscusiónacercadelpoblamientotempranoenlaProvinciadelChoapa”,
Revista Werkén 5,pp.15-20.
Masters, Patricia M. y Nic C. Flemming 1983.Quaternary Coastlines and Marine Archaeology. Towards the Prehistory of Land Bridges and
Continental Shelves.NuevaYork,AcademicPress.
Myers, Thomas, Michael Voorhies y Georde Corner1980.“SpiralFracturesandBonePseudotoolsatPaleontologicalSites”,American Antiquity 45(3),
pp.483-490.
Noakes, Scot t, Ervan Garrison y Greg McFall 2009.“UnderwaterPaleontology:RecoveryofaPrehistoricWhaleMandibleOffshoreGeorgia”,en
Proceedings of the American Academy of Underwater Sciences 28th Symposium, ed.N.W.Pollock,pp.245-251.Atlanta,AmericanAcademyofUnderwaterSciences.
Núñez, Lautaro, Juan Varela, Rodolfo Casamiquela y Carolina Villagrán1994.“ReconstrucciónmultidisciplinariadelaocupaciónprehistóricadeQuereo,CentrodeChile”,Latin
American Antiquity 5(2),pp.99-118.
O’Connell, James, Kristen Hawkes y Nicholas Blurton Jones 1990.“ReanalysisofLargeMammalBodyPartTransportamongtheHadza”,Journal of Anthropological
Research 17,pp.31-316.
Petriaggi, Roberto y Barbara Davidde2007.Archeologia sott’acqua. Teoria y practica.Pisa,AccademiaEditoriale.
Rinderknecht, Andrés2000.“LapresenciadeosteodermosenlasextremidadesposterioresdeGlyptodon clavipes Owen,
1839(Mammalia:Cingulata)”,Ameghiniana 37(3),pp.369-373.
Stewart, David1999.“FormationProcessesAffectingSubmergedArchaeologicalSites:AnOverview”en
Geoarchaeology 14(6),pp.565-587.
Stright, Melanie 1995.“ArchaicPeriodSitesontheContinentalShelfofNorthAmerica:TheEffectofRelativeSea-level
ChangesonArchaeologicalSiteLocationsandPreservation”,enArchaeological Geology of the Archaic Period in North America,ed.ElmerA.Bettis,pp.131-147.GeologicalSocietyofAmericaSpecialPaper297.
Vargas, Gabriel y Cristina Ortega2008.“Contextogeomorfológicoyanálisissedimentológicopreliminardemuestrasdelabahía
Quintero,VRegión”,enProyecto Terminal de Gas Natural Licuado (GNL) en Quintero, V Región, Comuna de Quintero.Valparaíso.
Waters, Michael 1992.Principles of Geoarchaeology: A North American Perspective.Tucson,TheUniversityofArizonaPress.

2 2 6
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1

APROXIMACIONES Y ESCALAS DE ANÁLISIS EN LA ZOOARQUEOLOGÍA Y TAFONOMÍA SUDAMERICANAS. ALGUNAS REFLEXIONES SOBRE SU ESTADO ACTUAL Y PERSPECTIVAS PARA SU DESARROLLO
m a r i a n a m o n d i n i y a . s e b a s T i á n m u ñ o z 2 2 9
Panorámicas

2 2 8

2 2 9
A PROX I M ACION ES Y ESCA L A S DE A NÁ LISIS EN L A ZOOA RQU EOLOGÍ A Y TA FONOM Í A SU DA M ER ICA NA S . A LGU NA S R EF LEX ION ES SOBR E SU ESTA DO ACT UA L Y PER SPECTI VA S PA R A SU DESA R ROLLO
mAriANA moNDiNi*[email protected], IDACOR-CONICET, Universidad de Buenos Aires, Argentina
a. sebasTián muñoz**[email protected], IDACOR-CONICET, Universidad Nacional de Córdoba, Argentina
Resumen En este artículo se discuten algunos aspectos teóricos y
metodológicos de la zooarqueología y la tafonomía en relación con
el desarrollo de estas disciplinas en Sudamérica. Particularmente,
se tratan cuestiones relacionadas con las escalas de análisis,
las aproximaciones teórico-metodológicas sobre el papel de
los procesos tafonómicos y el comportamiento humano, y la
interacción entre tafonomía y zooarqueología. Estos temas son
abordados en el marco del estado actual de estas disciplinas y en
el contexto de las particularidades de las faunas Neotropicales.
P A L A B R A S C L A V E :
Zooarqueología, tafonomía, escalas, actores, Sudamérica.
Artículorecibido:1dejuniode2011|aceptado:21deoctubrede2011|modificado:23denoviembrede2011
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,294pp.ISSN1900-5407,pp.229-250
* DoctoraenArqueología,UniversidaddeBuenosAires,Argentina.**DoctorenArqueología,UniversidaddeBuenosAires,Argentina.

2 3 0
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.229-250
approaChes and sCale analyses in south ameriCan ZooarCheology and taphonomy: refleCtions regarding its Cur-rent state and perspeCtiVes for its deVelopment
ABSTR AC T Theoretical and methodological
aspects of zooarchaeology and taphonomy
are discussed in relation to the development
of these disciplines in South America.
In particular, issues regarding scales of
analysis, theoretical-methodological
approaches to the role of taphonomic
processes and human behavior, and the
interaction between taphonomy and
zooarchaeology. Current issues in these
disciplines and the particularities of
Neotropical faunas are considered.
K E Y W O R D S :
Zooarchaeology, Taphonomy, Scales, Actors,
South America.
aBordagens e esCalas de análise na Zooarqueologia e tafonomia sul-ameriCanas: algumas reflexões soBre seu estado atual e perspeCtiVas para seu desenVolVimento
RESUMO Neste artigo discutem-se alguns
aspectos teóricos e metodológicos da
zooarqueología e a tafonomia em relação
ao desenvolvimento de estas disciplinas
em América do Sul. Particularmente
tratam-se questões relacionadas com
as escalas de análise, as abordagens
teórico-metodológicas sobre o papel dos
processos tafonômicos e o comportamento
humano, e a interação entre tafonomia
e zooarqueologia. Estes temas são
abordados na moldura do estado atual
destas disciplinas e no contexto das
particularidades das faunas neotropicais.
P A L A V R A S C H A V E :
Zooarqueologia, tafonomia, escalas, atores,
América do Sul.

2 3 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.229-250
A PROX I M ACION ES Y ESCA L A S DE A NÁ LISIS EN L A ZOOA RQU EOLOGÍ A Y TA FONOM Í A SU DA M ER ICA NA S . A LGU NA S R EF LEX ION ES SOBR E SU ESTA DO ACT UA L Y PER SPECTI VA S PA R A SU DESA R ROLLO
m A r i A N A m o N D i N i
a . s e b a s T i á n m u ñ o z
IntroducciónEl desarrollo de la tafonomía y la zooarqueología en la historia de la arqueología sudamericana está estrechamente vinculado a la introducción de una perspectiva teórica procesual en el tratamiento del regis-tro arqueológico (ver Borrero, 1988a; Mondini y Muñoz, 1996; Politis, 2003; Mengoni Goñalons, 2007). Este desarrollo no ha sido homogéneo a lo largo del continente, aunque puede plantearse que en general siguió distintas etapas vinculadas a la evolución que tuvieron ambas disciplinas en el resto del mundo, muchas veces con cierta dilación y en otros casos con desarrollos pioneros (Politis, 2003; Mengoni Goñalons, 2004; Mengoni Goñalons et al., 2010).
Durante las últimas décadas del siglo XX, las investigaciones tafonómicas sobre vertebrados en los países centrales estuvieron muy vinculadas al estudio de la información que ofrece el registro arqueofaunístico. En una primera etapa su lugar fue el de una instancia preliminar en la investigación arqueológica, destinada a “despojar” al registro arqueológico de los “sesgos” introducidos por agentes y procesos “naturales”, generalmente concebidos meramente como posdepositacionales. Posteriormente se dio importancia a la incorporación de la información paleoecológicamente relevante que brindan las trazas de estos procesos y agentes, lo que involucra no sólo evaluar la resolución y compara-bilidad de los conjuntos analizados sino también estudiar la variabilidad de los mismos (Gifford, 1981). Estos enfoques sobre la variabilidad derivaron en un mayor conocimiento y comprensión de la complejidad de los procesos tafonó-micos, lo que llevó paulatinamente a la formulación de marcos interpretativos

2 3 2
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.229-250
mejorados (Behrensmeyer, 1993). Más recientemente, las investigaciones tafo-nómicas vinculadas a la zooarqueología han incorporado un importante tra-bajo en la investigación de los procesos diagenéticos, tornándose asimismo más interdisciplinarias (O’Connor, 2005; ver también Behrensmeyer et al., 2000).
La zooarqueología, por su parte, ha pasado también por distintas etapas. De manera muy sintética, puede decirse que un período inicial, en los años setenta, se caracterizó por la discusión de unidades de análisis y de cuanti-ficación. A finales de esta década y en los años ochenta, muchos trabajos se orientaron hacia el reconocimiento de patrones y la búsqueda de situaciones típicas para abordar la variabilidad del registro zooarqueológico (ver discusión en Binford, 1978; Gifford, 1981; O’Connell et al., 1988; Schiffer, 1996 [1987]), buscando desarrollar, de este modo, un marco teórico y conceptual que, puede decirse, culmina con la publicación del modelo epistemológico propuesto en 1991 por Gifford-Gonzalez. En la década de los noventa el espacio destinado a la discusión del papel que los restos faunísticos ocupan en las explicaciones arqueológicas ha sido importante, tratándose de diversas maneras la comple-jidad existente entre la evidencia y los distintos procesos que la originan (por ejemplo, Stiner, 1993a y b, 1994; Lyman, 1994; Marean, 1995; Marciniak, 1999). Sin embargo, esta discusión no llevó a cambios sustanciales en las metodologías implementadas (Reitz y Wing, 2008). Para este momento comenzó una tenden-cia saludable que se continúa hasta hoy, que consiste en la publicación, cada vez más frecuente, de volúmenes dedicados a la zooarqueología y, en menor medida, a la tafonomía, acompañada de la publicación de numerosos artículos y síntesis en obras generales y revistas, así como la multiplicación de reuniones académicas en torno a estas disciplinas.
En una evaluación reciente del estado actual de la zooarqueología, Reitz y Wing (2008) encuentran que los cambios ocurridos en los últimos diez años no han sido tanto en las metodologías más tradicionales sino más bien en el desarrollo de aplicaciones específicas, tales como los estudios genéticos y los isotópicos. Es decir, no se trataría de cambios importantes en la teoría y meto-dología zooarqueológicas sino más bien en la ampliación del espectro de téc-nicas utilizadas y en el énfasis dado a líneas de trabajo que antes no tenían una representación importante. Estas autoras señalan asimismo que una consecuen-cia de la importancia relativa que adquirieron las explicaciones geoquímicas se hace manifiesta en la integración de distintas perspectivas en el estudio de las relaciones entre los humanos y el ambiente, que diluyó en parte las distinciones entre profesionales de distintas disciplinas que hasta ese momento eran pro-blemáticas (Reitz y Wing, 2008). Esta mayor amplitud de actores y perspectivas en el estudio de las relaciones humano-animales se hace evidente también en

2 3 3
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.229-250
A P R O X I M A C I O N E S Y E S C A L A S D E A N Á L I S I S | M A R I A N A M O N D I N I Y A . S E B A S T I Á N M U Ñ O Z
la aparición de volúmenes que integran perspectivas metodológicas (por ejem-plo, Maltby, 2006; Lyman, 2008), que desarrollan aspectos de la zooarqueología aplicada (por ejemplo, Lauwerier y Plug, 2004; Lyman y Cannon, 2004) o que integran diferentes disciplinas (por ejemplo, De Renzi et al., 2002; Lyman, 2008).
En síntesis, puede decirse que la situación actual de la tafonomía y la zooarqueología en el mundo es de una alta productividad y de un refinamiento en el abanico de técnicas disponibles para inferir información sobre el papel de los animales en el pasado y la historia de sus restos. Sin embargo, esta situación sigue siendo de alguna manera transicional, ya que el problema de fondo yace, como señala James O’Connell (1993), en los marcos de referencia explicativos a los que recurrimos para dar cuenta de la variabilidad en el registro arqueoló-gico, ya sea que se relacionen con los contextos ecológicos en que los registros fueron generados (por ejemplo, Gifford, 1981; Behrensmeyer, 1993; Hudson, 1993; O’Connell, 1993) o, más recientemente, con contextos sociales, religio-sos, de género y otros (por ejemplo, Gifford-Gonzalez, 1993, 2007; O’Day et al., 2004; Orton, 2010). En buena medida, se trata de un problema relacionado con el modo en que es utilizada la información sobre el mundo contemporáneo. Asimismo, se relaciona también con una visión paradigmática de la naturaleza de las inferencias zooarqueológicas y tafonómicas.
La tafonomía y la zooarquelogía en SudaméricaLa tafonomía y la zooarqueología en Sudamérica han seguido en rasgos genera-les los mismos caminos que en el resto del mundo, aunque con particularidades que expresan una diversidad de situaciones en las comunidades académicas de cada país (ver Politis, 2003; Mengoni Goñalons, 2004, 2007; Mengoni Goñalons et al., 2010; Borrero, 2008). El recuento que presentamos a continuación es limitado, ya que está principalmente basado en el desarrollo que estas disci-plinas han tenido en Argentina, no sólo porque como miembros de esa comu-nidad académica es el caso que más conocemos, sino además porque en este país la zooarqueología y tafonomía han tenido un desarrollo muy importante (Politis, 2003; Mengoni Goñalons, 2009). Consideramos que, pese a esta parti-cularidad, se trata de un ejemplo interesante para analizar la agenda actual de las disciplinas en el continente.
Un primer aspecto por señalar es que en Sudamérica los problemas de investigación abordados desde la zooarqueología y la tafonomía resultan tanto de la historia propia de estas disciplinas en la región (Mengoni Goñalons, 2004, 2009; Gutiérrez, 2009) como de su inserción en el escenario global (Reitz y Wing, 2008). En buena medida, los temas definidos han seguido lineamien-

2 3 4
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.229-250
tos similares a los del resto del mundo (ver, por ejemplo, Mengoni Goñalons, 2010), entre los que se destacan la explotación de megafauna y la domesticación de camélidos, agregándose más recientemente preguntas de carácter social. En este sentido, se destacan asimismo una preocupación metodológica desde el comienzo de las investigaciones sistemáticas (Mengoni Goñalons, 1988, 2010) y la incorporación de innovaciones técnico-metodológicas en concordancia con los desarrollos introducidos en países centrales (por ejemplo, Borrero, 1990, Barberena et al., 2009). Más aún, como mencionamos antes, algunos desarrollos teóricos y metodológicos han sido originales en la región, como es el caso del planteo de una explotación oportunista de la megafauna por parte de las primeras poblaciones humanas (Borrero, 2009, entre otros), a diferencia de los planteos dominantes en América del Norte, y el desarrollo de modelos de densidad ósea de camélidos basados en nuevas metodologías (Elkin, 1995; Stahl, 1999). Sin embargo, este desarrollo de la zooarqueología y la tafonomía en América del Sur no siempre se traduce en una incorporación de sus resul-tados fuera de la zona de producción en la que fueron generados (Politis, 2003; Mengoni Goñalons, 2007).
En la actualidad puede decirse que, más allá del grado de originalidad o desarrollo que hayan alcanzado desde lo metodológico o teórico, la zooarquelo-gía y, en menor medida, la tafonomía son campos de investigación reconocidos como importantes en la arqueología de la región (ver Nogueira de Queiroz, 1999; Politis, 2003; Mengoni Goñalons, 2007; Muñoz y Mondini, 2008a; este volumen). Más aún, las dos disciplinas han alcanzado en el continente altos estándares de calidad. En ambas existen, sin embargo, aspectos que aún no se han resuelto o que están siendo explorados, y sobre ellos se focaliza el presente trabajo.
Entre los problemas mencionados en la literatura se destaca reiterada-mente la falta de integración entre la investigación tafonómica y la arqueoló-gica (Mondini y Muñoz, 1996; Borella y Cruz, 2000; Borrero, 2007a; Mengoni Goñalons, 2007). Esto hace que, a pesar de tener un importante desarrollo en determinadas comunidades académicas, la tafonomía no sea considerada un campo de investigación completamente establecido en la actualidad. Éste es el caso de Argentina (Borrero, 2007a), una de las comunidades académicas que más aportes ha realizado en este sentido en Sudamérica (Politis, 2003).
Como señala Borrero (2007a), entre los aspectos positivos se destaca que en los últimos años se ampliaron los estudios tafonómicos planteados en escalas regionales (por ejemplo, Borrero, 1988b, 2001; Cruz, 2000), aunque su representación todavía es escasa. Esto se relaciona a su vez con la escala tem-poral de los estudios actualísticos, ya que son necesarios más estudios de largo plazo (por ejemplo, Borrero, 2007b; Cruz y Muñoz, 2010), capaces de reflejar la

2 3 5
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.229-250
A P R O X I M A C I O N E S Y E S C A L A S D E A N Á L I S I S | M A R I A N A M O N D I N I Y A . S E B A S T I Á N M U Ñ O Z
variabilidad de los procesos estudiados a lo largo del tiempo (Borrero, 2007a). Este autor también señala la necesidad de incorporar una perspectiva tafonó-mica en las investigaciones que abordan preguntas relacionadas con el género, poder, y aspectos simbólicos.
En este sentido, puede decirse además que las observaciones tafonómicas son relativamente abundantes pero los estudios sistemáticos no tanto, lo que sugiere que aún no se ha alcanzado un nivel de madurez en el reconocimiento de actores y procesos específicos. Como se señaló, esta situación en parte deriva de que las investigaciones tafonómicas no suelen formar parte integral y relevante de las investigaciones arqueológicas más generales. Incluso para los actores tafonómicos más estudiados localmente carecemos de un conoci-miento exhaustivo de su variabilidad y de sus efectos. Esta situación se rela-ciona también con que en muchos casos se aborda la relación entre trazas y actores determinados con base en información de otras regiones del mundo, que no siempre resultan relevantes para las condiciones locales.
La zooarqueología, como la tafonomía, ha logrado producir información con estándares internacionales de calidad, aunque la dependencia teórica y la tendencia a la replicación de los paradigmas hegemónicos son un problema por abordar en la agenda actual y futura de la disciplina en la región (Mengoni Goñalons, 2007). También la zooarqueología sudamericana ha apelado muchas veces a modelos generados en otras regiones, no siempre relevantes para las condiciones locales.
En lo metodológico, aunque se han hecho importantes trabajos experi-mentales (por ejemplo, Miotti, 1998; De Nigris, 2004; Gutiérrez y Kaufmann, 2007), se trata de un campo que aún puede desarrollarse más con resultados productivos (Mengoni Goñalons, 2010). Las investigaciones etnoarqueológi-cas también tienen un gran potencial para la zooarqueología, que apenas ha comenzado a explorarse (por ejemplo, Caracotche, 2001; Yacobaccio, 2007; Martínez, 2009).
En proporción, la mayor parte de las investigaciones zooarqueológicas han tratado diferentes aspectos de la subsistencia de sociedades cazadoras-recolectoras, y en menor medida, de los primeros productores de alimentos y de momentos históricos, por lo que es necesario un mayor énfasis en otras formas de organización social. También está pendiente un mayor trabajo en los aspectos políticos, sociales y simbólicos que se pueden abordar a partir del registro arqueológico (Mengoni Goñalons, 2010), que, como se señaló arriba, requieren además una aproximación tafonómica (Borrero, 2007a). De todos modos, estos temas comienzan a hacerse más comunes en los últimos años (ver, por ejemplo, deFrance, 2009).

2 3 6
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.229-250
La zooarqueología ha estado mucho más integrada que la tafonomía a las investigaciones arqueológicas más generales, pasándose rápidamente de apén-dices con listados taxonómicos a estudios plenamente integrados, que incluso proponían la agenda de investigaciones futuras. Sin embargo, esta integración podría ser todavía más productiva, de profundizarse en el futuro.
De una manera muy sintética, y con el riesgo de obviar matices y comple-jidades que ello implica, puede decirse que la arqueología de cazadores-reco-lectores en Sudamérica ha incorporado más activamente los estudios zooar-queológicos y los tafonómicos, incluso desde la concepción más productiva que plantea que las improntas tafonómicas no sólo tienen consecuencias nega-tivas para nuestras interpretaciones zooarqueológicas sino que también nos informan sobre las condiciones paleoecológicas en que vivieron estas socie-dades (ver Behrensmeyer y Kidwell, 1985; Behrensmeyer et al., 2000; Muñoz y Mondini; 2010). En la arqueología de sociedades complejas, en cambio, la zooarqueología no ha sido un motor tan importante de modelos, y la tafonomía ha ocupado un lugar más secundario, incluso muchas veces limitado a eva-luar posibles sesgos postdepositacionales del registro arqueológico. Puede asi-mismo plantearse que los modelos zooarqueológicos desarrollados en el marco de la arqueología de cazadores-recolectores tienden a tener niveles más altos de generalidad que los de sociedades complejas (pero ver deFrance, 2009).
Apuntes para la agenda de la zooarquelogía y tafonomía de los próximos años en SudaméricaUna tendencia saludable de la zooarqueología y la tafonomía sudamericanas que creemos puede profundizarse es la generación de modelos localmente rele-vantes que den cuenta de las interacciones entre las sociedades humanas y la fauna regional, así como de la acción de los procesos y agentes tafonómicos. Generar estos modelos, o bien apelar a las particularidades de los ambientes y faunas de la región al aplicar modelos desarrollados en otros ámbitos, son estrategias fundamentales, ya que estas propiedades dictarán la relevancia o no de los modelos o aspectos de los mismos, más allá de cuán corrientes sean en otras regiones del mundo.
La historia de las poblaciones humanas y su relación con la fauna en Sudamérica ha ocurrido bajo contextos que están profundamente condicio-nados por las propiedades f ísicas y la historia biótica del continente, y difieren de los de otras regiones en una serie de aspectos significativos. Por ejemplo, las interacciones entre predadores en la subregión Andino-Patagónica de los Neotrópicos, que abarca buena parte del continente, no implicarían los altos niveles de competencia involucrados en muchos modelos sobre tafonomía

2 3 7
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.229-250
A P R O X I M A C I O N E S Y E S C A L A S D E A N Á L I S I S | M A R I A N A M O N D I N I Y A . S E B A S T I Á N M U Ñ O Z
de carnívoros derivados de otros contextos (Muñoz y Mondini, 2007, 2008b). En los últimos años han comenzado a diversificarse los estudios destinados a abordar regionalmente este tipo de problemas (por ejemplo, Morales et al., 2009), que generan así marcos de referencia relevantes para la historia de las poblaciones humanas en América del Sur y para la formación del registro arqueofaunístico en la región.
Un problema relacionado que tenemos que resolver para avanzar en esta dirección es la dificultad que surge de no contar con buena información de base biológica y ecológica. Desconocemos, por ejemplo, en muchos casos la diversidad de especies actual y pasada y los cambios en las distribuciones de las mismas. Es decir, un mismo registro arqueológico o paleontológico muchas veces es objeto de estudio y referencia, ya que resulta dif ícil disponer de información independiente contra la que discutir nuestras ideas. Incluso procesos de gran impacto, como el resultante de la introducción de nuevas especies a partir de la ocupación europea del continente, recién empiezan a conocerse sistemáticamente. Esta falta de información se hace evidente también en lo limitado de los repositorios y las colecciones de museos, lo cual dificulta la contextualización y el estudio de la información pasada. Un mayor compromiso en la creación y sistematización de colecciones públicas es entonces un esfuerzo necesario.
Profundizar la tendencia a producir modelos regionalmente relevantes implica considerar las interacciones entre las poblaciones humanas y ani-males en América del Sur como instancias del amplio rango de variabilidad en las mismas a lo largo de nuestra trayectoria como especie, instancias que implicaron una serie de particularidades. Las mismas, por ende, suelen no estar representadas en otros continentes, y sin embargo forman una parte significativa de nuestra historia. Ello se relaciona a su vez con la relativa-mente baja inserción de los estudios sudamericanos en el escenario global antes mencionada, una tendencia que desde los años noventa ha comenzado a revertirse saludablemente.
Esto último es una excelente señal de la maduración relativa de la zooar-queología y de la tafonomía en nuestra región y de la divulgación de sus resul-tados en la comunidad académica más general. Sin embargo, dentro del con-tinente y en el resto de Latinoamérica están pendientes todavía un desarrollo más homogéneo de las disciplinas a lo largo de las distintas regiones y los diver-sos países, así como una mayor integración de las investigaciones realizadas y de los propios investigadores.
En un plano más general, uno de los principales desaf íos que a nuestro entender enfrentan la zooarqueología y la tafonomía consiste en evaluar qué

2 3 8
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.229-250
tan generales han de ser nuestras inferencias, más allá del marco teórico abor-dado. Esto se relaciona con nuestra concepción de la clase de datos que la evi-dencia arqueofaunística potencialmente ofrece y con los límites y la resolución de los mismos. Éste es un asunto que merece más atención aquí y en otras regiones del mundo, si bien en Sudamérica se hace particularmente relevante, dada la historia más joven de ambas disciplinas.
Una serie de preguntas surgen en torno a este tema, tales como: ¿ha habido avances en lo que respecta a cómo acceder a niveles más altos de gene-ralidad en las inferencias zooarqueológicas y tafonómicas, o los enfoques que aplicamos están todavía muy vinculados a explicaciones del tipo de las “his-torias de precaución” y las relaciones típicas entre actores y trazas? En otras palabras, ¿hemos avanzado en la construcción de narrativas históricas que vinculen la formación de registros faunísticos y la producción de trazas por humanos y otros agentes con contextos más generales? Más aún, cabe pre-guntarse si ésta es todavía una aspiración en la comunidad académica, o si la misma está actualmente más orientada a estudios particulares. También cabe preguntarse si las aproximaciones teóricas más recientemente incorporadas buscan la causalidad en principios generales o más bien en historias particula-res. Sobre este punto, pensamos que en la arqueología sudamericana pueden verse actualmente ambas tendencias.
Aun en los casos en que se busca elucidar historias más o menos particu-lares, puede pensarse que para dar cuenta de la formación del registro arqueo-lógico es necesario remitirse en última instancia a marcos de referencia más o menos generales. Incluso en el caso de los registros que presentan una resolu-ción suficientemente fina como para inferir actividades cotidianas, biograf ías de objetos, etc., su integridad debe demostrarse en cada caso (Borrero, 2007a).
Todo esto nos plantea que es necesario rediscutir, en esta nueva etapa de la zooarqueología y la tafonomía, el rol de la analogía y el uniformismo (ver Gifford, 1981; Gifford-Gonzalez, 1991). Un punto de partida para ello es reto-mar el modelo propuesto por Gifford-Gonzalez (1991), en el que se planteaba explícitamente la integración de niveles más amplios de causalidad en la zooar-queología y la tafonomía. La pregunta que así surge es cómo vemos en la actua-lidad la aplicación de un modelo epistemológico vinculado a las relaciones de vida y las narrativas históricas, cuyas consecuencias han sido exploradas pero no profundizadas todavía, tanto en la arqueología sudamericana como en la de otras regiones del mundo (ver, por ejemplo, Marean, 1995; Gifford-Gonzalez, 1999; Marciniak 1999; Muñoz, 2007).
Más allá de la valoración que se haga de estos intentos y de las alternati-vas que se propusieron, pensamos que el modelo de Gifford-Gonzalez (1991)

2 3 9
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.229-250
A P R O X I M A C I O N E S Y E S C A L A S D E A N Á L I S I S | M A R I A N A M O N D I N I Y A . S E B A S T I Á N M U Ñ O Z
sigue siendo una guía programática con plena vigencia en la actualidad, ya que sugiere maneras específicas y epistemológicamente explícitas de explotar el máximo potencial de la zooarqueología y la tafonomía. Más aún, el mismo puede adaptarse a marcos teóricos diferentes de aquel dentro del que fuera originalmente elaborado (ver, por ejemplo, Gifford-Gonzalez, 2007). Se trata, asimismo, de un modelo interesante para estudiar el papel de los humanos en la comunidad faunística sudamericana, si tenemos en cuenta la historia de la biota del subcontinente, caracterizada por períodos de conexión y aislamiento con Norteamérica y por extinciones catastróficas en el Pleistoceno, entre otros procesos relevantes.
Uno de los problemas que enfrentamos al plantearnos este tipo de pro-gramas tiene que ver con el modo de trabajar la variabilidad de la evidencia y las distintas escalas de análisis involucradas (Gifford-Gonzalez, 1991; Speth, 1991; O’Connell, 1993, 1995, entre otros). Como señala Gifford-Gonzalez (1991), el asignar significados más complejos y generales a los agregados de trazas y res-tos en los conjuntos zooarqueológicos plantea nuevas fuentes de incertidum-bre inferencial, tanto metodológica (por ejemplo, respecto de la equifinalidad) como teórica (por ejemplo, respecto de la causalidad en los sistemas biológi-cos). Para ello, la autora propuso, entre otras cosas, una investigación actua-lística más compleja y epistemológicamente explícita. Como se analizó más arriba, si bien se han producido notables avances, esto requiere aún de mucho desarrollo y de un fuerte trabajo tafonómico y geoarqueológico.
Los estudios actualísticos de escalas regionales y de largo plazo son un camino productivo en este sentido que es necesario seguir profundizando. A pesar de algunas opiniones en detrimento de los estudios actualísticos natu-ralísticos y los estudios comparativos sobre registros fósiles modernos (ver Marean, 1995), pensamos que cuando son bien controlados, éstos tienen un potencial aún mayor que los experimentales, ya que, si bien se pierde certi-dumbre respecto de la relación entre actores y trazas, se gana mucha informa-ción contextual que contribuye de manera fundamental a dar sentido a nues-tras observaciones. Lo que contribuye a garantizar nuestras inferencias es más bien un análisis profundo de cuáles propiedades pueden considerarse unifor-mes y cuáles no, y bajo qué circunstancias pueden variar. De todos modos, si bien esto último dif ícilmente pueda derivarse de los estudios experimentales, éstos son necesarios para ahondar en las relaciones entre trazas y los niveles de causalidad más próximos.
En general, entonces, es fundamental desarrollar aún más los estudios tafonómicos actualísticos y los etnoarqueológicos en Sudamérica. Una vía con mucho potencial son asimismo la búsqueda de patrones y el análisis de proce-

2 4 0
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.229-250
sos a escala suprarregional, una perspectiva más cercana a la biogeograf ía y la macroecología, estrategia que aún no ha tenido tanto desarrollo mundial.
Gifford-Gonzalez planteaba en 1991 que, habiéndose estudiado muchos actores y procesos tafonómicos en un nivel básico, podía proseguirse con el estudio de los contextos en los cuales varían. En la situación actual de la tafo-nomía y la zooarqueología en nuestro continente, consideramos necesario avanzar con ambas clases de objetivos paralelamente: ampliar nuestro cono-cimiento sobre los actores y procesos de menor nivel, especialmente en lo que incumbe a su variabilidad, a la vez que indagamos sobre las condiciones bajo las cuales varían sus acciones y los efectos de las mismas. Éste puede ser un punto de partida para acotar el problema derivado de la mayor ambigüedad propia de niveles de análisis en los que los resultados son probabilísticos y las propiedades emergentes algo esperado, en un contexto disciplinar en donde las sucesivas rondas de investigación sobre agentes y procesos particulares no han alcanzado aún niveles de redundancia. Una estrategia de investigación pro-ductiva sería una que apunte simultáneamente tanto a reconocer la variabilidad potencial y a reducir la ambigüedad de trazas y patrones en el registro faunís-tico como a explorar desde sus objetivos mismos las condiciones bajo las cuales se produce tal variabilidad y, por ende, su significado en cuanto al comporta-miento humano en el pasado y el contexto en que tuvo lugar.
Un tema pendiente en relación con estos planteos tiene que ver con la cuestión de si hemos avanzado en el tratamiento de la equifinalidad o nos hemos vuelto menos exigentes en este aspecto. En la medida en que se destaque una visión típica del comportamiento de los agentes tafonómi-cos, incluso del humano, y demos mayor importancia a los sesgos que a la información paleoecológica, que las trazas tafonómicas brindan, será dif ícil articular la idea de rangos de efectos producto de un agente que es varia-ble contextualmente y de efectos similares producto de distintos agentes. En esto es importante la integración de los modelos tafonómicos a la par de los zooarqueológicos, de manera que la tafonomía no ocupe un lugar de com-promiso, para garantizar que no haya fuertes sesgos postdepositacionales, sino que dote de significado ecológico y material a la interpretación de la evidencia arqueológica.
En este punto nos preguntamos en qué medida han sido reconocidas las “contribuciones positivas” de la tafonomía (Behrensmeyer y Kidwell, 1985). Una estrategia mixta como la aquí propuesta, orientada a recono-cer la variabilidad tafonómica al mismo tiempo que las condiciones bajo las cuales se genera, implica asimismo considerar tanto las contribuciones “negativas” (sesgos) como las “positivas” (paleoecología) de la tafonomía.

2 4 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.229-250
A P R O X I M A C I O N E S Y E S C A L A S D E A N Á L I S I S | M A R I A N A M O N D I N I Y A . S E B A S T I Á N M U Ñ O Z
Este aspecto de las investigaciones tafonómicas sigue siendo central en la agenda pendiente de la arqueología de Sudamérica, ya que ahondar en la información paleoecológica que ofrecen las trazas y restos de organismos en el registro arqueológico permitirá a su vez contextualizar los comporta-mientos humanos del pasado.
Esto se relaciona con la necesidad de modelos localmente relevantes comentada arriba. La incorporación de principios generales, en conjunto con las trayectorias históricas que moldearon la biota sudamericana, aplicados al estudio de las poblaciones humanas y su registro arqueológico, son una herramienta con mucho potencial para construir narrativas históricas sobre la trayectoria de estas poblaciones en el continente (por ejemplo, Stahl, 1996; Muñoz y Mondini, 2008b).
Discusión y conclusiones Como se señaló, la tendencia que muestra la zooarqueología en la actualidad puede sintetizarse como la de una disciplina orientada a la incorporación de nuevos objetivos de investigación y marcos de referencia (simbólicos y socia-les, por ejemplo), y sobre todo de nuevas técnicas de análisis para resolver los viejos y nuevos problemas, pero dentro de un universo de preguntas que no ha implicado en todos los casos un cambio sustancial en los niveles de causalidad (Reitz y Wing, 2008). La tafonomía vinculada a la zooarqueología, por su parte, ha avanzado sobre la generación de información vinculada a la variabilidad de agentes y procesos pero aún no se encuentra en una etapa de síntesis y de hipó-tesis generales (Behrensmeyer, 1993).
En Sudamérica, la zooarqueología y tafonomía han alcanzado un impor-tante grado de desarrollo, si bien éste es heterogéneo a través de la gama de investigaciones arqueológicas que se llevan a cabo en el continente y de las dife-rentes regiones y países dentro del mismo. Del diagnóstico actual se desprende que al menos los siguientes puntos son sensibles y aún requieren trabajo:• laintegraciónentretafonomíayzooarqueología,yentreambasyelrestode
las investigaciones arqueológicas;• el desarrollo de estudios actualísticos: estudios tafonómicos en escalas
espaciales y temporales suficientemente amplias como para abordar la variabilidad de los distintos actores y procesos, y estudios experimentales y etnoarqueológicos que complementen los marcos de referencia generados por otras vías;
• eldesarrollodemásestudioszooarquelógicossobreaspectossociales,reli-giosos, simbólicos, de género y otros más recientemente introducidos en la arqueología de la región, y la incorporación activa de la tafonomía en

2 4 2
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.229-250
los mismos, así como más investigaciones zooarqueológicas y tafonómicas sobre sociedades complejas;
• eldesarrollodemodelos ymétodos zooarqueológicos y tafonómicosqueden cuenta de los procesos particulares que incumben a la historia de las poblaciones humanas y los ambientes en Sudamérica, y que no dependan necesariamente de los paradigmas propuestos en otras regiones;
• eltratamientodelanaturalezayvariabilidaddelaevidenciaylasdistintasesca-las de análisis involucradas en una estrategia de investigación de este tipo, y
• la superaciónde lasdiferencias señaladas respectodelestadodecono-cimiento disponible en las distintas regiones y comunidades académicas del continente.
El desarrollo futuro de las disciplinas en la región se verá beneficiado por la ampliación de los espacios de intercambio y discusión, así como por genera-ción de modelos relevantes para las condiciones locales y, sobre todo, la reflexión sobre la generalidad de nuestras inferencias y los modos de obtenerlas.
Creemos que pasar del reconocimiento de patrones y explicaciones próxi-mas a explicaciones más generales y niveles más altos de causalidad implica necesariamente apelar a escalas más amplias, tanto temporales como espacia-les. También destacamos la necesidad de continuar el desarrollo de estudios actualísticos, y especialmente la utilidad de una estrategia mixta de investiga-ción, que busque reconocer la variabilidad de los procesos y agentes tafonómi-cos al mismo tiempo que se exploran las condiciones bajo las cuales esa varia-bilidad se genera, y, por ende, los contextos en que vivieron las poblaciones humanas en el pasado. Una propuesta programática como la de Gifford-Gon-zalez (1991) puede ser más productiva en Sudamérica si se generan investiga-ciones en ambos niveles simultáneamente, y se integran desde el comienzo de las investigaciones arqueológicas, teniendo en cuenta el contexto específico del continente y de cada región en particular.
Cabe preguntarse de todos modos qué tanto interés ha suscitado entre los investigadores de la región la aspiración a niveles de causalidad más gene-rales. En este sentido, debemos tener en cuenta, además de la joven historia de las disciplinas en Sudamérica, algunas cuestiones paradigmáticas. En cual-quier caso, como señalan muchos autores, una mayor integración de tafonomía y zooarqueología es una pieza clave tanto de un programa como el señalado como de otras propuestas, ya que sólo los arqueólogos buscamos responder a las preguntas sobre la formación del registro arqueológico, y ninguna otra dis-ciplina ha generado la información necesaria para responderlas (Binford, 1981; Gifford-Gonzalez, 1991, entre otros).

2 4 3
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.229-250
A P R O X I M A C I O N E S Y E S C A L A S D E A N Á L I S I S | M A R I A N A M O N D I N I Y A . S E B A S T I Á N M U Ñ O Z
Como se señaló, esto no niega que el desarrollo de la zooarqueología y la tafonomía sudamericanas ha sido muy destacable en general, tanto en canti-dad como en calidad. Así lo evidencia, por ejemplo, el grado de avance del que disponemos hoy sobre algunos temas planteados como importantes hace más de veinte años, tales como la tafonomía de la acción de carnívoros sudameri-canos (Borrero, 1988b) y los aspectos vinculados a la economía prehistórica y a los estudios paleoambientales abordados desde el registro zooarqueológico (Haber y Ratto, 1988).
Un repaso de la literatura de síntesis (ver, por ejemplo, Mengoni Goña-lons et al., 2010) deja en claro que en la última década la zooarqueología y tafo-nomía sudamericanas han dado en efecto un gran paso, expresado asimismo en el valor que se le asigna a la comunicación entre profesionales y la circulación de resultados originales. Esto es evidente si uno releva las herramientas que para ello se han propuesto, tales como la Red Iberoamericana de Arqueozoolo-gía propuesta en el congreso del ICAZ de 2002, la Red de Laboratorios Latinoa-mericanos de Zooarqueología (http://redlabz.uniandes.edu.co/) y otras redes de investigación con miembros de diferentes países, así como la publicación de volúmenes específicos para dar cuenta del estado actual de la disciplina en la región (Mengoni Goñalons, 2004, 2009; Gutiérrez et al., 2007, 2010; Muñoz y Mondini, 2008a; López et al., 2009; Mengoni Goñalons et al., 2010; Álvarez et al., 2011), el surgimiento de espacios propios en reuniones internacionales (como el Plenario sobre Zooarqueología Latinoamericana en el congreso del ICAZ de 2006 y los simposios de zooarqueología y tafonomía Neotropicales en los dos últimos congresos del ICAZ), e incluso reuniones científicas específicas (como las reuniones nacionales de zooarqueología llevadas a cabo en Chile en 2006 y Argentina en 2008 y 2011, los Encuentros de Zooarqueología Latinoa-mericana en 2009, y próximamente en 2012; el congreso “Culturas Americanas y su Ambiente: Perspectivas desde la Zooarqueología, la Paleoetnobotánica y la Etnobiología”, realizado en México en 2010, y el recientemente creado Grupo de Trabajo de Zooarqueología Neotropical del ICAZ y su primer reunión aca-démica en 2012, además de los congresos del ICAZ realizados en la región: en 2006 en México y próximamente, en 2014, en Argentina). Estos espacios ponen en evidencia que existe una necesidad de generar y compartir conocimiento en lo que podemos definir como comunidad académica sudamericana, y que los desarrollos que se requieren no necesariamente van a venir a futuro de la mano de la replicación de modelos generados en otras regiones o en el marco de problemas de investigación diferentes (ver discusión en Politis, 2003; Men-goni Goñalons, 2007; Muñoz y Mondini, 2008a). Para ello es necesario enton-ces profundizar estos espacios académicos compartidos.

2 4 4
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.229-250
Afortunadamente, como señalan Reitz y Wing (2008), la zooarqueología es un campo disciplinar internacional llevado adelante por una comunidad inclusiva y en crecimiento, por lo que seguramente el futuro mostrará más desarrollos teóricos, síntesis y contribuciones originales, como ya comienza a ser el caso.
En suma, tanto aquí como en otras regiones del mundo, el futuro más productivo de la zooarqueología y la tafonomía está ligado a nuestra capacidad de discutir teoría, metodología y epistemología desde estas disciplinas. Pensa-mos que ambas disciplinas se encuentran ya en ese camino en la región.
AgradecimientosExpresamos nuestro agradecimiento a Guillermo Mengoni Goñalons, quien nos facilitó bibliograf ía; a los organizadores del Primer Encuentro Latino-americano de Zooarqueología; a dos evaluadores anónimos y a los editores de Antípoda. .

2 4 5
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.229-250
A P R O X I M A C I O N E S Y E S C A L A S D E A N Á L I S I S | M A R I A N A M O N D I N I Y A . S E B A S T I Á N M U Ñ O Z
R e f e R e n c i a s
Álvarez, M. Clara, Agustina Massigoge, Andrés D. Izeta, Mariela González y Daniel Rafuse (comp.)2011.“LibrodeResúmenes”,IICongresoNacionaldeZooarqueologíaArgentina,Olavarría,Fac.Cs.
Sociales,UniversidadNacionaldelCentrodelaPcia.deBuenosAires.
Barberena, Ramiro, Adolfo Gil, Gustavo A. Neme y Robert H. Tykot2009.“StableIsotopesandArchaeologyinSouthernSouthAmerica.Hunter-Gatherers,Pastoralismand
Agriculture:AnIntroduction”,International Journal of Osteoarchaeology 9,pp.127-129.
Behrensmeyer, Anna K.1993.“Discussion:NonculturalProcesses”,enFrom Bones to Behavior, Ethnoarchaeological and
Experimental Contributions to the Interpretation of Faunal Remains,ed.JeanHudson,pp.342-348.Carbondale,CenterforArchaeologicalInvestigations,SouthernIllinoisUniversityatCarbondale.
Behrensmeyer Anna K. y Susan M. Kidwell1985.“Taphonomy’sContributionstoPaleobiology”,Paleobiology 11(1),pp.105-119.
Behrensmeyer, Anna. K., Susan M. Kidwell y Robert A. Gastaldo2000.“TaphonomyandPaleobiology”,Deep Time: Paleobiology’s Perspective, Paleobiology,26
(SupplementtoNo.4),pp.103-147.
Binford, Lewis R. 1978.Nunamiut Ethnoarchaeology.NuevaYork,AcademicPress.1981. Bones, Ancient Men and Modern Myths.NuevaYork,AcademicPress.
Borella, Florencia e Isabel Cruz2000“Simposio:diferentesenfoquesparaelestudiodelosprocesosnaturalesdeformacióndelregistro
arqueológico”,enDesde el país de los gigantes. Perspectivas arqueológicas en Patagonia, pp.179-181.RíoGallegos,UniversidadNacionaldelaPatagoniaAustral.
Borrero, Luis Alberto 1988a.“EstudiostafonómicosenTierradelFuego:surelevanciaparaentenderprocesosdeformación
delregistroarqueológico”,enArqueología contemporánea argentina. Actualidad y perspectivas,ed.HugoDanielYacobaccio,pp.13-32.BuenosAires,Búsqueda.
1988b.“Tafonomíaregional”,enDe procesos, contextos y otros huesos,eds.NormaR.RattoyAlejandroF.Haber,pp.9-15.BuenosAires,ICA(SecciónPrehistoria),FFyL,UBA.
1990.“Fuego-PatagonianBoneAssemblagesandtheProblemofCommunalGuanacoHunting”,enHunters of the Recent Past,eds.LeslieB.DavisyBrianO.K.Reeves,pp.373-299.Londres,UnwinHyman.
2001.“RegionalTaphonomy:BackgroundNoiseandtheIntegrityoftheArchaeologicalRecord”,enEthnoarchaeology of Andean South America,ed.LawrenceA.Kuznar,pp.243-254.AnnArbor,InternationalMonographsinPrehistory,EthnoarchaeologicalSeries4.
2007a.“NoDirectionHome:VertebrateTaphonomyinArgentina”,enTaphonomy and Zooarchaeology in Argentina,eds.MaríaGutierrez,LauraMiotti,GustavoBarrientos,GuillermoMengoniGoñalonsyMónicaSalemme,pp.9-12.Oxford,Archaeopress,BritishArchaeologicalReports,InternationalSeries1601.
2007b.“LongitudinalTaphonomicStudiesinTierradelFuego,Argentina”,enTaphonomy and Zooarchaeology in Argentina,eds.MaríaA.Gutierrez,,LauraMiotti,GustavoBarrientos,GuillermoMengoniGoñalonsyMónicaSalemme,pp.219-233.Oxford,Archaeopress,BritishArchaeologicalReports,InternationalSeries1601.
2008.“TheArchaeologyoftheNeotropics”,Neotropical Zooarchaeology and Taphonomy, Quaternary International180(1),pp.152-157.

2 4 6
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.229-250
2009.“TheElusiveEvidence:TheArcheologicalRecordoftheSouthAmericanExtinctMegafauna”,enAmerican Megafaunal Extinctions at the End of the Pleistocene,ed.GaryHaynes,pp.145-168.Dordrecht,Springer.
Caracotche, Soledad 2001.“TheInvisibilityofTime:AnEthnoarchaeologicalStudyoftheTemporarySitesofHerdersofthe
SouthernPuna”,enEthnoarchaeology of Andean South America,ed.LawrenceA.Kuznar, pp.97-115.AnnArbor,InternationalMonographsinPrehistory,EthnoarchaeologicalSeries4.
Cruz, Isabel 2000.“Líneastafonómicasyecológicasparaevaluarlaexplotaciónprehistóricadeavesacuáticas
enlazonacordillerana(Prov.deSantaCruz)”,enDesde el país de los gigantes. Perspectivas arqueológicas en Patagonia.TomoI,pp.202-217.RíoGallegos,UniversidadNacionaldelaPatagoniaAustral.
Cruz, Isabel y A. Sebastián Muñoz2010.“Tafonomíacomparativa:seguimientodecarcasasdemamíferosenPuntaEntrada(SantaCruz,
Argentina)”,enZooarqueología a principios del siglo XXI. Aportes teóricos, metodológicos y casos de estudio,eds.MaríaA.Gutiérrez,MarianaDeNigris,PabloM.Fernández,MiguelGiardina,AdolfoGil,AndrésIzeta,GustavoNemeyHugoYacobaccio,pp.387-396.BuenosAires,EdicionesdelEspinillo.
deFrance, Susan D.2009.“ZooarchaeologyinComplexSocieties:PoliticalEconomy,Status,andIdeology”,Journal of
Archaeological Research 17(2),pp.105-168.
De Nigris, Mariana2004. El consumo en grupos cazadores recolectores. Un ejemplo zooarqueológico de Patagonia
meridional.BuenosAires,SociedadArgentinadeAntropología.
De Renzi, Miquel, Miguel Pardo Alonso, Margarita Belinchón, Enrique Peñalver Moyá, Plinio Montoya, Ana Márquez-Aliaga y Valentín Villaverde (eds.)
2002. Current Topics on Taphonomy and Fossilization.Valencia,ColleccióEncontres5,Ayuntamiento deValencia.
Elkin, Dolores1995.“StructuralDensityofSouthAmericanCamelidSkeletalParts”,International Journal of
Osteoarchaeology 5,pp.29-37.
Gif ford, Diane P. 1981.“TaphonomyandPaleoecology:ACriticalReviewofArchaeology’sSisterDisciplines”,enAdvances
in Archaeological Method and Theory,ed.MichaelSchiffer,Vol.4,pp.365-438.NuevaYork,AcademicPress.
Gif ford-Gonzalez, Diane P. 1991.“BonesArenotEnough:Analogues,Knowledge,andInterpretiveStrategiesinZooarchaeology”,
enJournal of Anthropological Archaeology10,pp.215-254.1993.“GapsinZooarchaeologicalAnalysesofButchery.IsGenderAnIssue?”,enFrom Bones to
Behavior. Ethnoarchaeological and Experimental Contributions to the Interpretation of Faunal Remains,ed.JeanHudson,pp.181-199.Carbondale,CenterforArchaeologicalInvestigations,SouthernIllinoisUniversityatCarbondale.
1999.“ZooarchaeologyinAfrica:TheFirstTwoMillionYears”,ArchaeozoologiaX,pp.55-82.2007.“ToughtsonaMethodforZooarchaeologcalStudyofQuotidianLife”,enInterpreting Household
Practices. Treballs d’Arqueologia13,pp.5-27.
Gutiérrez, María A.2009.“Tafonomía:¿tiraníaomultivocalidad?”,enPerspectivas actuales en arqueología argentina,eds.
RamiroBarberena,KarenBorrazzoyLuisBorrero,pp.57-87.BuenosAires,CONICET-IMHICIHU.

2 4 7
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.229-250
A P R O X I M A C I O N E S Y E S C A L A S D E A N Á L I S I S | M A R I A N A M O N D I N I Y A . S E B A S T I Á N M U Ñ O Z
Gutiérrez, María y Cristian Kaufmann2007.“MethodologicalCriteriafortheIdentificationofFormationProcessesinGuanaco(Lama
guanicoe)BoneAssemblagesinFluvial-LacustrineEnvironments”,Journal of Taphonomy5(4),pp.151-176.
Gutiérrez, María A., Laura Miot ti, Gustavo Barrientos, Guillermo Mengoni Goñalons y Mónica Salemme (eds.)
2007.Taphonomy and Zooarchaeology in Argentina.Oxford,Archaeopress,BritishArchaeologicalReports,InternationalSeries1601.
Gutiérrez, María, Mariana De Nigris, Pablo M. Fernández, Miguel Giardina, Adolfo Gil, Andrés Izeta, Gustavo Neme y Hugo Yacobaccio (eds.)
2010. Zooarqueología a principios del siglo XXI. Aportes teóricos, metodológicos y casos de estudio. BuenosAires,EdicionesdelEspinillo.
Haber, Alejandro y Norma Ratto1988.“Deprocesos,contextosyotroshuesos:presentación”,enDe proce sos, contextos y otros huesos,
eds.AlejandroHaberyNormaRatto,pp.1-7.BuenosAires,ICA(SecciónPrehistoria),FFyL,UBA.
Hudson, Jean (ed.)1993.From Bones to Behavior, Ethnoarchaeological and Experimental Contributions to the
Interpretation of Faunal Remains.Carbondale,CenterforArchaeologicalInvestigations,SouthernIllinoisUniversityatCarbondale.
Lauwerier, Roel C. G. M. e Ina Plug (eds.)2004. Future from the Past: Archaeozoology in Wildlife Conservation and Heritage Management.
Oxford,OxbowBooks.
López, Patricio, Isabel Cartajena, Christian García y Francisco Mena (eds.)2009.Zooarqueología y tafonomía en el confín del mundo.SantiagodeChile,SeriedeMonografías
Arqueológicas,FacultaddeEstudiosdelPatrimonioCulturaldelaUniversidadInternacionalSEK-Chile.
Lyman, R. Lee 1994.Vertebrate Taphonomy (Cambridge Manuals in Archaeology).Cambridge,CambridgeUniversity
Press.2008.Quantitative Paleozoology (Cambridge Manuals in Archaeology).Cambridge,Cambridge
UniversityPress.
Lyman, R. Lee y Kenneth P. Cannon 2004. Zooarchaeology & Conservation Biology. SaltLakeCity,UniversityofUtahPress.Maltby, J. Mark (ed.)2006.Integrating Zooarchaeology.Oxford,OxbowBooks.
Marciniak, Arkadiusz 1999.“FaunalMaterialsandInterpretativeArchaeology-epistemologyReconsidered”,Journal of
Archaeological Method and Theory6(4),pp.293-320.
Marean, Curtis W.1995.“OfTaphonomyandZooarchaeology”,Evolutionary Anthropology4,pp.64-72.
Martínez, Gustavo2009.“HumanChewingBoneSurfaceModificationandProcessingofSmallandMediumPreyAmongst
theNukak(ForagersoftheColombianAmazon)”,Journal of Taphonomy7(1),pp.1-19.
Mengoni Goñalons, Guillermo L.1988.“Análisisdematerialesfaunísticosensitiosarqueológicos”,Xama1,pp.71-120.2004. Zooarchaeology of South America.Oxford,BritishArchaeologicalReports,InternationalSeries1298.

2 4 8
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.229-250
2007.“ArchaeofaunalStudiesinArgentina:AHistoricalOverview”,enTaphonomy and Zooarchaeology in Argentina,eds.MaríaGutiérrez,LauraMiotti,GustavoBarrientos,GuillermoMengoniGoñalonsyMónicaSalemme,pp.13-34.Oxford,Archaeopress,BritishArchaeologicalReports.
2009.“LaZooarqueologíadeSudamérica:sulugarenelmundo”,enZooarqueología y tafonomía en el confín del mundo,eds.PatricioLópez,IsabelCartajena,ChristianGarcíayFranciscoMena, pp.9-13.SantiagodeChile,SeriedeMonografíasArqueológicas,FacultaddeEstudiosdelPatrimonioCulturaldelaUniversidadInternacionalSEK-Chile.
2010.“AdvancesinAnimalBoneArchaeologyinArgentina:GeneralTrendsandSomeProspectsfortheFuture”,enArqueozoología latinoamericana: origenes y desarrollo,eds.GuillermoMengoniGoñalons,JoaquínArroyo-Cabrales,OscarJ.PolacoyFelisaJ.Aguilar,pp.15-24.México,InstitutoNacionaldeAntropologíaeHistoriaparalaCienciaylaTecnología,ConsejoNacionalparalaCienciaylaTecnologíaeInternationalCouncilforArchaeozoology.
Mengoni Goñalons, Guillermo, Joaquín Arroyo-Cabrales, Oscar J. Polaco y Felisa J. Aguilar (eds.)
2010. Arqueozoología latinoamericana: orígenes y desarrollo.México,InstitutoNacionaldeAntropologíaeHistoria,ConsejoNacionalparalaCienciaylaTecnología,InternationalCouncilforArchaeozoology,yUniversidaddeBuenosAires.
Miotti, Laura1998.“ZooarqueologíadelamesetacentralycostadeSantaCruz.Unenfoquedelasestrategias
adaptativasaborígenesylospaleoambientes”,Revista del Museo de Historia Natural de San Rafael10(1/4),pp.3-306.
Mondini, Mariana y A. Sebastián Muñoz1996.“Eldesarrollodelatafonomíaenlaarqueologíaargentina.Estadoactualyperspectivas”,enII
Reunión de Tafonomía y Fosilización,eds.M.FernandaBlascoSanchoeIsabelPérezUrresti, pp.255-258.Zaragoza,InstituciónFernandoelCatólico.
Morales, Marcelo, Ramiro Barberena, Juan B. Belardi, Luis Borrero, Valeria Cortegoso, Victor Durán, A. Guerci, Rafael Goñi, Adolfo Gil, Gustavo Neme, Hugo Yacobaccio y Marcelo Zárate
2009.“ReviewingHuman-environmentInteractionsinAridRegionsofSouthernSouthAmericaduringthePast3000Years”,Palaeogeography, Palaeoclimatology, Palaeoecology281,pp.283-295.
Muñoz, A. Sebastián2007.“Lacomplejidaddelossistemasecológicosenlaexplicacióndelregistroarqueofaunísticodelos
cazadoresrecolectoresdelaIslaGrandedeTierradelFuego”,enSurvey on Human and Fauna Relationships, Human and Faunal Relationships Reviewed: An Archaeozoological Approach,eds.EduardoCorona-M.yJoaquínArroyo-Cabrales,pp.35-42.Oxford,BritishArchaeologicalReports,InternationalSeries,S1627.
Muñoz, A. Sebastián y Mariana Mondini2007.“HumansinSouthAmericanFaunalCommunities.InteractionswithPreyandPredatorsin
theSouthernCone”,enLate Neogene and Quaternary Biodiversity and Evolution: Regional Developments and Interregional Correlations,eds.Ralf-DietrichKahlke,LutzChristianMaulyPaulP. Mazza,Courier Forschungsinstitut Senckenberg,258,pp.205-211.
2008a.“NeotropicalZooarchaeologyandTaphonomy”,Quaternary International 180(1),pp.1-4.2008b.“LongTermHuman/AnimalInteractionsandTheirImplicationsforHunter-gathererArchaeology
inSouthAmerica”,enTime and Change: Archaeological and Anthropological Perspectives on the Long Term,eds.DimitraPapagianni,RobertLaytonyHerbertD.G.Maschner,pp.55-76.Oxford,OxbowBooks.

2 4 9
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.229-250
A P R O X I M A C I O N E S Y E S C A L A S D E A N Á L I S I S | M A R I A N A M O N D I N I Y A . S E B A S T I Á N M U Ñ O Z
2010.“Taphonomy”,enEncyclopedia of Geography,ed.BarneyWarf,Vol.6,pp.2775-2776.ThousandOaks,SAGE.
Nogueira de Queiroz, Alberico 1999.“Panoramadel’archéozoologieauBrésil”,enL’Amérique du Sud: Des chasseurs-cueilleurs à
l’Empire Inca (Actes des journées d’étude d’archéologie précolombienne, Genève, 10-11 octobre 1997),eds.A.Chevalier,L.VelardeeI.Velarde,pp.23-29.Oxford,BritishArchaeologicalReports,InternationalSeries746.
O’Connell, James1993.“Discussion:SubsistenceandSettlementInterpretation”,enFrom Bones to Behavior,
Ethnoarchaeological and Experimental Contributions to the Interpretation of Faunal Remains,ed.J.Hudson,pp.169-176.Carbondale,CenterforArchaeologicalInvestigations,SouthernIllinoisUniversityatCarbondale.
1995.“EthnoarchacologyNeedsaGeneralTheoryofBehavior”,Journal of Archaelogical Research 1, pp.205-255.
O’Connell, James, Kristen Hawkes y Nicholas G. Blurton-Jones1988“HazdaHunting,andBoneTransportandTheirArchaeologicalImplications”,Journal of Anthro-
pological Research 44,pp.113-161.
O’Connor, Terry (ed.)2005. Biosphere to Lithosphere: New Studies in Vertebrate Taphonomy.Oxford,OxbowBooks.
O’Day, Jones Sharyn, Wim Van Neer y Anton Envynck (eds.)2004. Behaviour Behind Bones: The Zooarchaeology of Ritual, Religion, Status and Identity.Oxford,
OxbowBooks.
Orton, David C. 2010.“TaphonomyandInterpretation:AnAnalyticalFrameworkforSocialZooarchaeology”,
International Journal of Osteoarchaeology.Artículopublicadooriginalmenteonline:26deseptiembrede2010|DOI:10.1002/oa.1212.
Politis, Gustavo G.2003.“TheTheoreticalLandscapeandtheMethologicalDevelpmentofArchaeologyinLatinAmerica”,
American Antiquity68(2),pp.248-272.
Reitz, Elizabeth J. y Elizabeth S. Wing2008.Zooarchaeology (Cambridge Manuals in Archaeology),segundaedición.Cambridge,Cambridge
UniversityPress.
Schif fer, Michael B.1996[1987].Formation Processes of the Archaeological Record.SaltLakeCity,UniversityofUtahPress.
Speth, John D.1991.“TaphonomyandEarlyHominidBehavior.ProblemsinDistinguishingCulturalandNon-cultural
Agents”,enHuman Predators and Prey Mortality,ed.MaryStiner,pp.187-200.Boulder,WestwiewPress,WestviewSpecialStudiesinArchaeologicalResearch.
Stahl, Peter W.1996.“HoloceneBiodiversity:AnArchaeologicalPerspectivefromtheAmericas”,Annual Review of
Anthropology 25,pp.105-126.1999.“StructuralDensityofDomesticatedSouthAmericanCamelidSkeletalElementsandthe
ArchaeologicalInvestigationofPrehistoricAndeanCh’arki”,Journal of Archaeological Science26,pp.1347-1368.

2 5 0
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.229-250
Stiner, Mary C.1993a.“ModernHumanOrigins-faunalPerspectives”,Annual Review of Anthropology22,pp.55-82.1993b.“ThePlaceofHominidsAmongPredators:InterspecificComparisonsofFoodProcurementand
Transport”,enFrom Bones to Behavior. Ethnoarchaeological and Experimental Contributions to the Interpretation of Faunal Remains,ed.JeanHudson,pp.38-61.Carbondale,CenterforArchaeologicalInvestigations,SouthernIllinoisUniversityatCarbondale.
1994.Honor among Thieves. A Zooar chaeological Study of Neandertal Ecology.Princeton,PrincetonUniversityPress.
Yacobaccio, Hugo D. 2007.“AndeanCamelidHerdingintheSouthAndes:EthnoarchaeologicalModelsforArchaeozoological
Reseach”,enHerding Practices and Techniques: Models and Methodological Tools for Archaeozoological Analysis.ActsoftheICAZsession(Mexico,August2006),Anthropozoologica 42(2),pp.143-154.

2 5 1


Documentos

2 5 4

2 5 5
L A ZOOA RQU EOLOGÍ A DEN TRO DE L A U N I V ER SI DA D NACIONA L AU TÓNOM A DE M ÉX ICO
r aúl Valadez azúa*
[email protected] de Paleozoología, Instituto de Investigaciones Antropológicas de la Universidad Nacional Autónoma de México, México
GilberTo pérez roldán**
[email protected] de Paleozoología, Instituto de Investigaciones Antropológicas de la Universidad Nacional Autónoma de México, México
IntroducciónEn México la zooarqueología es una disciplina de fuertes contrastes. Por un lado, sabemos que desde finales del siglo XVIII se reportaron hallazgos de restos animales asociados a estructuras religiosas de tiempos prehispánicos (Bernal, 1992), pero aún en la actualidad muchos arqueólogos dudan de si vale la pena rescatar la arqueofauna descubierta por-que no saben si los resultados podrán aportar información relevante para su investigación; por otro lado, desde el siglo XX, a partir de que la arqueología mexicana se constituyó como disciplina con un método de trabajo estructu-rado, siempre el arqueólogo ha vivido la disyuntiva de impulsar el estudio de la colección zooarqueológica obtenida, o mejor buscar información general en fuentes escritas del siglo XVI sobre la fauna que utilizaban los pueblos indíge-nas antes de la Conquista.
Parte de este esquema tiene su origen en que la zooarqueología apare-ció en México, en el Instituto Nacional de Antropología e Historia (INAH)
* DoctorenCienciasBiológicas,UniversidadNacionalAutónomadeMéxico,México.**MaestríaenAntropología,Arqueología,UniversidadNacionalAutónomadeMéxico,México.

2 5 6
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.255-265
(Pérez, 2009), medio siglo después de los primeros estudios arqueológicos formales, pero también existe la circunstancia de que gran parte de los tra-bajos zooarqueológicos se han realizado bajo un esquema conservador, es decir, limitando la investigación a la identificación de las especies presentes, que concluye con una sencilla lista de restos estudiados bajo la forma de un informe técnico, sin que exista en ningún momento la comunicación necesa-ria entre zooarqueólogo y arqueólogo, de forma que impulse la investigación hacia la comprensión de cómo el recurso animal fue concebido y empleado por la comunidad ahí establecida. Resultado inevitable de este patrón es que ambas partes (el arqueólogo y el zooarqueólogo) aceptan sin problemas los limitados productos bajo la certeza de que la ausencia de información (poten-cialmente existente pero no extraída) podrá ser sustituida por datos prove-nientes de fuentes históricas.
Origen de la zooarqueología en la Universidad Nacional Autónoma de MéxicoLa más importante institución académica mexicana, la Universidad Nacional Autónoma de México (UNAM), cuenta con un laboratorio de zooarqueolo-gía adscrito al Instituto de Investigaciones Antropológicas (IIA) desde 1986, cuando Raúl Valadez Azúa fue contratado para desarrollarlo.
Aunque aproximadamente un 25% de los académicos del IIA son arqueó-logos, cuando el Laboratorio de Paleozoología fue creado no se sugirieron esquemas de trabajo más allá de que su principal responsabilidad fuera el apoyo a los proyectos arqueológicos; por otro lado, el responsable, siendo bió-logo formado en la UNAM, no tenía un antecedente de cómo era el trabajo zooarqueológico “normal” en entidades como el INAH, lo que permitió que la propuesta de cómo funcionaría el laboratorio se estructurara al margen de esquemas tradicionales y con las sugerencias de algunos antropólogos que con el paso de los años se han convertido en importantes figuras de la investigación antropológica mexicana, por ejemplo, los doctores Linda Manzanilla, Luis A. Vargas Guadarrama y Carlos Serrano Sánchez.
Uno de los grandes beneficios de la UNAM es la libertad de cátedra, la cual, aunque no siempre puede rebasar los conflictos personales o las limita-ciones de recursos, generalmente se mantiene como un enorme paraguas que protege a la academia de las tormentas económicas o políticas que se viven en el país. Dicha libertad, si es convertida en productos concretos de investigación –publicaciones, participación en congresos, cursos, conferencias, etc.–, dejará abierta la posibilidad de que el académico involucrado pueda desarrollar su línea de investigación con libertad.

2 5 7
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.255-265
L A Z O O A R Q U E O L O G Í A D E N T R O D E L A U N A M | R A Ú L V A L A D E Z A Z Ú A Y G I L B E R T O P É R E Z R O L D Á N
Este conjunto de condiciones permitieron que desde el principio los pro-ductos zooarqueológicos derivados del laboratorio rebasaran el esquema tra-dicional y buscaran dar opciones acerca de la forma como se había dado la relación hombre-fauna en el sitio de estudio (Serra y Valadez, 1985), aspecto que, a partir de ese momento, se convirtió en la meta mínima por cubrir dentro de cualquier estudio zooarqueológico que se vaya a realizar.
Impulso interdisciplinarioPara mediados de los ochenta, el tema de la interdisciplina era algo poco manejado en México, pero en el laboratorio de Paleozoología este esquema de pensamiento se dio de manera natural como consecuencia de que el acadé-mico responsable tenía formación biológica y fuertemente encaminada hacia la investigación científica formal, pero dentro de un ámbito laboral completa-mente antropológico. Esto permitió que los problemas antropológicos fueran emprendidos bajo otros esquemas de pensamiento y que el resultado tuviera una alta aceptación académica (Barba et al., 1987; Serra y Valadez, 1986; Vala-dez, 1988a, 1988b y 1989), por lo que, cuando surgen las primeras demandas de estudios por realizar bajo una concepción interdisciplinaria, esto fue abordado sin problemas (Valadez, 1992, 1993, 1994, 1995 y 1996). Es necesario recalcar que dentro de este proceso fue nuevamente factor clave la libertad de cátedra de la UNAM, pues permitió que los productos obtenidos justificaran el esquema de pensamiento sin necesidad de explicarle a nadie. Por otro lado, debido a que en el campus de la ciudad universitaria (donde se encuentra el IIA) es rela-tivamente fácil encontrar especialistas de diversas áreas que acepten apoyar el trabajo académico de colegas (zoólogos en general, médicos, nutriólogos, paleontólogos, médicos veterinarios, etc.), el ambiente mismo propicia la bús-queda de soluciones a problemas antropológicos a partir de otras disciplinas.
Ruta de la investigación zooarqueológica en el Laboratorio de PaleozoologíaA partir de los noventa, ya con lo interdisciplinario como esquema normal de trabajo, el reto académico principal ha sido incorporar todo tipo de estu-dio y visión innovadora dentro de la investigación zooarqueológica. Esto, sin embargo, ha requerido una planeación más cuidadosa de la ruta de estudio que sigue cada elemento óseo, a fin de saber qué cantidad y tipo de datos podrán obtenerse.
La investigación zooarqueológica del Laboratorio de Paleozoología posee tres niveles (ver la figura 1). El primero incluye el estudio del elemento óseo, el hueso como tal, el cual se analiza para saber si requiere consolidación, y

2 5 8
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.255-265
después, para ubicarlo anatómica, taxonómica y ontogénicamente. Si en este primer nivel se reconocen detalles que involucren la necesidad o el interés de seguir estudiándolo con otros fines (búsqueda de evidencias de manipulación del hueso, ser parte de un esqueleto identificado, posibles malformaciones del individuo, detalles que sugieran manejo de organismos vivos o uso de los espe-címenes para estudios químicos o de otro tipo), entonces entrará a alguna de las dos categorías de estudio especializado.
El segundo nivel involucra la colección arqueozoológica, ya como un conjunto de individuos reconocidos. El acervo de información derivado del anterior nivel se estudia bajo una perspectiva arqueológica, ecológica, zoogeo-gráfica e histórica, para determinar el Mínimo Número de Individuos (MNI),
Fasesdetrabajo ysecuencia
Nivelysecuencia delosestudios
Disciplinasinvolucradas
I.Individual (elelementoóseocomo entidaddeestudio)
BásicoZoologíaAnatomíacomparadaTaxonomíaOntogeniaRestauraciónyconservación
EspecializadoIAnálisisdematerialesArqueologíaexperimentalVeterinariaZootecnia
EspecializadoIIQuímicaBiologíamolecularGenética
II.Colectivo (lacolecciónarqueozoológica comoentidaddeestudio)
Estratigráficocontextual Arqueología
Poblacional EcologíaGenéticadepoblaciones
Paleoambiental EcologíaZoogeografíaGeologíahistórica
III.Integral
EtnobiológicoHistoriaEtnohistoriaArqueologíaLingüísticaPaleobotánicaEtnobiología
Figura1.EsquemadetrabajozooarqueológicoqueserealizadentrodellaboratoriodePaleozoologíadelInsti-tutodeInvestigacionesAntropológicasdelaUniversidadNacionalAutónomadeMéxico.

2 5 9
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.255-265
L A Z O O A R Q U E O L O G Í A D E N T R O D E L A U N A M | R A Ú L V A L A D E Z A Z Ú A Y G I L B E R T O P É R E Z R O L D Á N
reconstruir los esquema de uso, los cambios naturales y antropogénicos que se dieron para con la fauna al paso del tiempo y la reconstrucción de las condicio-nes ambientales que existieron durante el período estudiado.
Por último, en el tercer nivel se involucran diversas disciplinas antropo-lógicas (y otras afines) para colocar la información dentro de un marco etno-biológico y, así, tanto enriquecer los resultados obtenidos como aportar nuevo conocimiento a la etnobiología.
El cuadro mostrado, sin embargo, requiere especialistas de diversas áreas, incluso dentro del mismo grupo de trabajo, pues es imposible que una sola persona pueda realizar toda la línea de estudio. En nuestro caso, tenemos un equipo constituido por dos arqueólogos (uno especializado en la manufactura del hueso), un biólogo con estudios de posgrado en antropología y un bió-logo, además de químicos, f ísicos, médicos veterinarios, odontólogos, zoólo-gos, paleoetnobotánicos, antropólogos f ísicos, lingüistas y etnólogos, y otros arqueozoólogos con quienes se ha colaborado.
Pero dif ícilmente podría llegarse a este nivel de esfuerzo académico si no existiera siempre el propósito de involucrar al arqueólogo responsable del pro-yecto dentro del estudio zooarqueológico, para que se integre en la medida de lo posible al espíritu de la investigación y proporcione la información necesaria para avanzar en los estudios. Ciertamente, no todos aceptan en principio la idea, pero conforme se han ampliado el banco de publicaciones hechas por el labora-torio y el alto nivel de beneficios académicos, se hace más fácil la labor de con-vencimiento e, incluso, puede darse la situación de que conozca el tipo de estudio que se realiza y busque precisamente este tipo de trabajos y resultados.
Algunos casos ilustrativosA lo largo de tres décadas han existido tanto proyectos de investigación con todo el acervo necesario para estudiar al máximo cada elemento óseo como aquellos con los cuales los resultados han quedado estacionados en cierto nivel, por la ausencia de información o por el pequeño tamaño de la colección. Por otro lado, se han llevado a cabo líneas de investigación dentro del laboratorio, independientes de proyectos arqueológicos, cuyo fin es el desarrollo de méto-dos de estudio del material zooarqueológico.
El estudio de los cánidos mesoamericanos. Perteneciente a estas líneas de investigación tenemos, como primer caso, el estudio de los cánidos arqueo-zoológicos, el cual se inició desde 1989, junto con la Maestra en Antropología Alicia Blanco Padilla, de la Dirección de Salvamento Arqueológico del INAH. Su origen se relaciona con la enorme cantidad de restos del género Canis en contextos mesoamericanos, los cuales, sin embargo, nunca habían sido objeto

2 6 0
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.255-265
de estudio detallado. En el presente poseemos el esquema metodológico nece-sario para reconocer especie, edad, sexo, raza (en el caso del perro), dimensio-nes (alzada, longitud, peso, patrones alimentarios, padecimientos congénitos, relación cánido-hombre, valor simbólico, posible uso post mórtem, y si se trató de ejemplares que al morir tenían menos de un año de edad, época en que se dio el evento) (Blanco, Valadez y Rodríguez, 2009) (ver la figura 2).
Fasesdetrabajo ysecuencia
Nivelysecuenciadelosestudios
I.Individual
Consolidación,siserequiere,determinaciónanatómica,tomademedidas,identificacióndeespecie,reconocimientodeedad,huellasdemanipulación,aspectostafonómicos,sisetratadepartedeunconjuntodehuesosoesunapiezaaislada
Estudiodehuellasdemanipulación,determinaciónderaza,desexo,deanomalías,deposiblesusosdelhuesoydelcánido
Limpiezadelamuestra,elaboracióndecolecciones
demuestras,realizacióndeestudios
II.Colectivo
Contextualizacióndelosmateriales,determinacióndelMNI,debiotipos,reconstruccióndelaspoblacionesdecánidos,determinacióndeactividadesinvolucradasconcadatipodecánidos
Cánidospresentesenelsitio,condicionesdevidacondicionesambientalesyhumanasquedeterminaron
supresencia,esquemasdeinteracciónconelhombreenespacioytiempo,determinacióndelaépocademuertedeindividuosnoadultos
Empleodelosdatosdeloscánidossilvestresparareconocercondicionesambientales
III.IntegralComparacióndelainformaciónobtenidarespectoaloscánidosconlaprovenientedeotrossitiosuotrasfuentesdeinformación. Reconocimientodelarelaciónhombre-cánidocomofenómenocultural
Figura2.ProcesodeestudiodecánidoszooarqueológicosdentrodellaboratoriodePaleozoologíadelaUNAM.

2 6 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.255-265
L A Z O O A R Q U E O L O G Í A D E N T R O D E L A U N A M | R A Ú L V A L A D E Z A Z Ú A Y G I L B E R T O P É R E Z R O L D Á N
Elementos traza y colecciones zooarqueológicas. Otra línea de investi-gación, aún en proceso de desarrollo, es el estudio de elementos traza, para determinar patrones alimentarios (en humanos y en fauna doméstica), pero el esquema metodológico requiere el empleo de la fauna silvestre del sitio estu-diado para estructurar un marco de referencia que permita convertir los valores obtenidos en tendencias alimentarias (Valadez et al., 2005; Valadez y Tejeda, 2005) (ver la figura 3).
Figura3.Ejemplodeusodelaarqueofaunademamíferosdeunsitioarqueológicoparareconocerpatronesalimentariosenhumanos.Losnivelesdeestronciosondirectamenteproporcionalesalasdietasricasenvegetales,ylosdezinc,alascarnívoras.AlobtenerdatosderelaciónSr-Znenfaunasilvestrecreamosunmarcodereferenciaquepermiteubicarlastendenciashumanas.Losresultados,deacuerdoconnuestracondiciónnaturalomnívoraysuposiciónenlagráfica,nospresentanalosindividuosestudiadosconunadietaintermediaentreardillasyzarigüeyas,indicativodeunaalimentaciónmuydiversa,peroconpredominanciadealimentosdeorigenvegetal(Valadezet al.,2005;ValadezyTejeda,2005).
Fauna del proyecto “Túneles y Cuevas” de Teotihuacán. Respecto de pro-yectos arqueológicos que nos ilustran sobre el esquema de investigación tene-mos, en primer lugar, “Estudio de Túneles y Cuevas en Teotihuacan”, del cual se derivó una colección zooarqueológica de poco más de 3.000 individuos per-tenecientes a 275 taxa, distribuidos a lo largo de trece siglos (siglo VII-VIII d. C. hasta el presente). Su estudio permitió proponer secuencias ocupacionales del sitio, uso de la fauna en diversas épocas, evolución paleoambiental de la región, el hallazgo de una nueva forma de cánido doméstico, determinación de patrones alimentarios en humanos y cánidos domésticos y la elaboración de propuestas sobre el uso de los perros para realizar fechamientos relativos mediante el colágeno residual (Valadez, en prensa).
Fauna del “Entierro 6” de la Pirámide de la Luna. En este momento se están concluyendo los estudios de la fauna descubierta en el “Entierro 6” de la segunda estructura piramidal más importante de la ciudad arqueológica
Ominívoros o carnívorosno especializados

2 6 2
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.255-265
de Teotihuacán, la “Pirámide de la Luna” (Sugiyama y López, 2006). La investi-gación zooarqueológica se inició en 2004 con el rescate de cuarenta organismos descubiertos, seguido de su consolidación, estudio y armado. En este momento se ha determinado el esquema de manejo que se dio con cada uno de ellos, que en algunos casos incluyó su captura y posterior cautividad, el reconocimiento de lesiones, forma de muerte y esquema de traslado a la pirámide (Blanco et al., en prensa) (ver la figura 4).
Conclusiones Aunque la investigación zooarqueológica que se realiza en el Laboratorio de Paleozoología del IIA de la UNAM tuvo un inicio lleno de situaciones fortuitas, con el paso de los años ha tenido la oportunidad de consolidar un esquema de trabajo interdisciplinario de amplio beneficio académico, como lo demuestran las aproximadamente 200 publicaciones que se han derivado al paso de 23 años.
Entre los factores que han tenido un papel crucial para el desarrollo de la zooarqueología bajo el esquema mostrado se encuentran, en primer lugar, la libertad de cátedra; en segundo término, el involucramiento del arqueólogo con los estudios por realizar, y, finalmente, la posibilidad de constituir grupos de trabajo donde las humanidades y las ciencias experimentales tienen un peso similar y el mismo objetivo, llevando así a una propuesta de investigación muy detallada en cuanto a la forma como el material faunístico es estudiado, y ambi-ciosa, en cuanto a la información que se busca obtener; afortunadamente, los resultados logrados demuestran, sin lugar a dudas, que este esquema de investi-gación es una realidad en todo sentido y un referente de la forma como se puede conducir la investigación zooarqueológica en América Latina. .

2 6 3
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.255-265
L A Z O O A R Q U E O L O G Í A D E N T R O D E L A U N A M | R A Ú L V A L A D E Z A Z Ú A Y G I L B E R T O P É R E Z R O L D Á N
Fasesdetrabajoysecuencia
Nivelysecuenciadelosestudios
I.Individual
Remineralizaciónyconsolidacióndeloshuesos,reconstruccióndelosmismos, determinaciónanatómica,tomademedidas,identificacióndeespecie,deedad, búsquedadehuellasdemanipulaciónydeotrotipodealteracionestafonómicas
Estudiodehuellasdemanipulación,reconstruccióndecadaejemplar,determinacióndelsexoydesubiotipo,estudioderestosóseosdecavidadgástricaydelesionesidentificadas
Separacióndemuestras,limpieza,elaboracióndecoleccionesde
muestras,realización deestudiosdeelementostraza
II.Colectivo
Búsquedadepatronesdeacomododeindividuos,determinacióndelosesquemasdemanejodecadaunoyposiblesasociacionessimbólicas
Reconocimientodeindividuoscompletosoquellegaronbajolaformadepieles,reconstruccióndelmanejoquesedioconcadaunoprevioalevento,estudiode
condicionesdecautividadymanejodeejemplaresvivos,determinacióndelposiblemomentodemuertedelosanimalescautivosyformaenqueserealizó
Determinacióndelugaresdeprocedenciadelosejemplares,estudiodelas circunstanciasbiológicasquefavorecieronlacautividaddealgunos
III.Integral
Comparacióndelainformaciónobtenidaconladeotrosentierrosdescubiertos enlaPirámidedelaLuna,estudiodeotroseventosdecautividaddeanimalesen
Mesoamérica,reconocimientodelatradicióndecapturarcríasymantenercautivosaciertosanimalescomoesquemaculturalmesoamericano
Figura4.Secuenciadetrabajozooarqueológicorealizadoconlosejemplaresdescubiertosenel“Entierro6”delaPirámidedelaLuna(Blancoet al.,enprensa).

2 6 4
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.255-265
R e f e R e n c i a s
Barba, Luis, Beatriz Ludlow, Linda Manzanilla, Emily McClung y Raúl Valadez1987.“LavidadomésticaenTeotihuacn.Unestudiointerdisciplinario”,Ciencia y DesarrolloXIII(77),
pp.21-32.
Bernal, Ignacio1992.Historia de la arqueología mexicana.México:Porrúa.
Blanco, Alicia, Raúl Valadez y Bernardo Rodríguez2009.El estudio de los cánidos arqueológicos del México prehispánico.México:Institutode
InvestigacionesAntropológicas-InstitutoNacionaldeAntropologíaeHistoria.
Blanco, Alicia, Gilberto Pérez, Bernardo Rodríguez, Nawa Sugiyama, Fabiola Torres y Raúl ValadezEnprensa.“FaunalRemainsfromBurial6attheMoonPyramid”,enThe Burials of the Moon Pyramid,
SaburoSugiyamayDavidCarballo,coords.,ArizonaStateUniversity.
Pérez, Gilberto2009.“Laarqueozoología:presenteyfuturo”,TesisdeMaestría,FacultaddeFilosofíayLetras.México,
UniversidadNacionalAutónomadeMéxico.
Serra. Mari Carmen y Raúl Valadez1985.“FaunadeTerremote-Tlatenco,D.F.”,Anales de AntropologíaXXII,pp.159-213.1986.“LosrecursoslacustresdelaCuencadeMéxico:LosPatos”,Anales de AntropologíaXXIII,
pp.120-148.
Sugiyama, Saburo y Leonardo López (eds.)2006.Sacrificios de consagración en la Pirámide de la Luna.México:InstitutoNacionaldeAntropología
eHistoria-ConsejoNacionalparalaCulturaylasArtes-MuseodelTemploMayor-ArizonaStateUniversity.
Valadez, Raúl1988a.“RestosdeOvis canadiensisenelcentrodeMéxico”,Ciencia y DesarrolloXIV(81),pp.28-39.1988b.“Algunosaspectossobreaprovechamientodelosrecursossilvestres,alimentaciónymodelos
deestudioencomunidadesprehispánicasdelaCuencadeMéxico”,enEtnoarqueología,MariCarmenSerrayYokoSugiura,eds.,PrimerColoquioPedroBoschGimpera,pp.283-304.México:InstitutodeInvestigacionesAntropológicas.
1989.“Restosanimales”,enLas máscaras de la cueva de Santa Ana Teloxtoc,ErnestoVargas,ed., pp.142-150.México:UniversidadNacionalAutónomadeMéxico.
1992.“Impactodelrecursofaunísticoenlasociedadteotihuacana”,TesisdeDoctoradoenCienciasBiológicas,FacultaddeCiencias,UniversidadNacionalAutónomadeMéxico,México.
1993.“Macrofósilesfaunísticos”,enAnatomía de un conjunto residencial teotihuacano en Oztoyohualco,LindaManzanilla,ed.,II,pp.729-813.México:UniversidadNacionalAutónomadeMéxico.
1994.“¿CuántasrazasdeperrosexistieronenelMéxicoprehispánico?”,Veterinaria México25,(1),pp.1-11.1995.El perro mexicano.México:InstitutodeInvestigacionesAntropológicas.1996.La domesticación animal.México:PlazayValdez-InstitutodeInvestigacionesAntropológicas.
Valadez, Raúl (coord.)Enprensa.El ambiente y el hombre: Arqueofauna de los túneles de Teotihuacan: Estudios
Interdisciplinarios,vol.II,El inframundo de Teotihuacán: ocupaciones post-teotihuacanas en los túneles al este de la Pirámide del Sol, ed.general,LindaManzanilla,México:ElColegioNacional.

2 6 5
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.255-265
L A Z O O A R Q U E O L O G Í A D E N T R O D E L A U N A M | R A Ú L V A L A D E Z A Z Ú A Y G I L B E R T O P É R E Z R O L D Á N
Valadez, Raúl, Samuel Tejeda, Graciela Zarazúa, Leticia Carapias y Mario Casas2005.“Elestudiodeelementostrazaenrestosarqueozoológicosysuempleoenlareconstrucciónde
paleodietas”,Estudios de Antropología BiológicaXII(2),pp.945-970.
Valadez, Raúl y Samuel Tejeda2005.“Modeloparaladeterminacióndepatronesalimentariosenpoblacionesprehispánicasdelcentro
deMéxicoatravésdelacuantificacióndeelementostraza”,enPrimer Congreso Argentino de Arqueometría,AdríanPifferettiyRaúlBolmaro,eds.,pp.290-299.HumanidadesyArtesEdiciones.

2 6 6

2 6 7
L A F U NCIÓN TR A NSDISCIPLI NA R I A DE L A A RQU EOZOOLOGÍ A EN EL SIGLO X X I: R ESTOS A N I M A LES Y M Á S A LL Á
Luis ALberto borrero*[email protected] y Universidad de Buenos Aires CONICET-IMHICIHU y Universidad de Buenos Aires, Argentina
A la memoria de Oscar J. Polaco
La arqueozoologíaLa arqueozoología se ha asentado sólida-mente como uno de los pilares de la arqueología en el siglo XXI. Su objetivo gira alrededor del estudio de los huesos, pero esto no debe confundirse con una invitación a la hiperespecialización, que no es la mejor vía para lograr estudios integrados y comprensivos. He dedicado este trabajo a la memoria de Oscar J. Polaco, para mí un modelo en el tipo de búsqueda que defiendo, ya que en su obra abarcó desde el Pleistoceno hasta los períodos históricos (hablo exclusi-vamente de sus contribuciones arqueozoológicas), incluyendo una variedad de problemas desde muy distintos ángulos, generando puentes entre disciplinas (ver Polaco et al., 2001; Arroyo-Cabrales et al., 2005).
Recientemente se ha destacado que dentro de este campo disciplinario existen variantes tales como la arqueozoología propiamente dicha –centrali-zada en la relevancia cultural de los restos faunísticos– o la “arqueología bioló-gica” –centralizada en comparaciones entre distribuciones pasadas y presentes de faunas– (Mengoni Goñalons, 2007: 14), a las que se pueden agregar estudios de arqueozoología aplicada (Lyman, 2006), quizá incluidos los de transforma-ciones ambientales (Odling-Smee et al., 2003; Stahl, 2008). Como ha enfatizado Mengoni Goñalons, estas perspectivas se complementan y son integrables.
* Doctor,UniversidaddeBuenosAires,Argentina.

2 6 8
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.267-274
Durante mucho tiempo la aspiración de la arqueozología fue multidisciplina-ria, en el sentido de tratar su objeto desde ángulos muy diferentes, creando las sólidas bases observables hoy en distintos países (Mengoni Goñalons, 2004; Quiroz, 2009). La tafonomía asumió un papel crucial en el logro de todos estos fines, dada su capacidad para apuntalar la construcción de conocimiento. Más allá de esto, por definición operativa la tafonomía se constituye en parte inte-gral de esta disciplina. Lo que hace no tanto tiempo llegó en algunos ámbitos a ser considerado como una moda, hoy se asienta en el mismo nivel de nece-sidad que tienen el informe estratigráfico o la taxonomía. De todas maneras, claramente “zooarchaeology is not just taphonomy, it goes far beyond” (Men-goni Goñalons, 2007: 22). Diré incluso que los objetivos de la arqueozoología trascienden el análisis de las faunas, pues debe presentar información creíble, operativa y –aquí está el desaf ío principal– transdisciplinaria. En otras palabras, debe servir a fines que no están exclusivamente centralizados en el mundo ani-mal. Una de las formas es la consideración de las prácticas alimenticias utili-zando distintos tipos de fuentes desde las arqueofaunas, los textos o los arte-factos, como lo ha hecho María Marschoff al analizar el caso de Floridablanca, el asentamiento español del siglo XVIII (Marschoff, 2007). Otra forma es tratar de discutir el problema en diferentes escalas espaciales o temporales, utilizando el mismo tipo de materiales (Mengoni Goñalons et al., 2000). Cualquiera de las soluciones obliga a una interacción fuerte con los materiales no faunísticos.
Por otra parte, no creo que se requiera un acercamiento teórico especial ni una arqueología exclusiva de América Latina (Borrero, 2004), opinión que extiendo a la arqueozoología. Entiendo las formas de practicar arqueozoología como universales, con matices que se seleccionan de acuerdo con una enorme variedad de circunstancias –entre las cuales se cuentan, por cierto, las geo-gráficas o las políticas–, pero para mí la forma de la distribución resultante no tiene un eje geográfico.
Existen varios temas comunes que parecen específicos para América Latina, como el estudio de los camélidos (Mengoni Goñalons et al., 2000; Yaco-baccio, 2007) o su particular biodiversidad (Mengoni Goñalons, 2004). Sin embargo, los análisis realizados sobre los restos de camélidos no difieren de los que se eligen para estudiar otros animales. Los enfoques pueden ser morfomé-tricos (L’Heureux, 2005), etnoarqueológicos (Martinez, 2007; Yacobaccio, 2007; Beretta, 2008), moleculares (Orlando et al., 2009), etc., ninguno de los cuales es exclusivo para los camélidos. Entonces, la existencia de problemas arqueozooló-gicos específicos de América Latina es innegable, tan sólo que éstos no pueden constituirse en el criterio demarcador de las metodologías de investigación.
Vivian Scheinsohn (2009) analizó el panorama teórico relacionado con

2 6 9
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.267-274
L A F U N C I Ó N T R A N S D I S C I P L I N A R I A D E L A A R Q U E O Z O O L O G Í A E N E L S I G L O X X I | L U I S A L B E R T O B O R R E R O
la construcción de una arqueología darwiniana en la periferia de los grandes centros productores de conocimiento, en particular en Argentina. Uno de sus resultados es que en Argentina operó una mezcla de esquemas. Scheinsohn indica que muchas de las aplicaciones teóricas realizadas en la periferia exce-den o contrarían el préstamo, para constituirse en elaboración teórica (Schein-sohn, 2009: 82). Éstas son algunas de las condiciones desde las que se pueden realizar aportes metodológicos y teóricos desde América Latina, teniendo en claro que éstos también servirán en otros lugares del mundo. Por otra parte, el momento por el que pasa la arqueozoología en general requiere una postura metodológica. Lo avalan los avances producidos, entre otros, por Elkin (1995), Scheinsohn y Ferretti (1995), Stahl (1999), Gutiérrez y Kaufmann (2007), Bore-lla et al. (2007), San Román (2009) o Kaufmann (2009). Estos aportes, a la vez, apuntalan la arqueozoología de América Latina y prestan un servicio que tras-ciende sus fronteras. No es casual que estos ejemplos provengan mayoritaria-mente del campo metodológico, crucial para la construcción arqueofaunística. Los estudios de anatomía económica, tafonómicos o ingenieriles mencionados plantean claros puentes hacia discusiones más amplias. En suma, hay varias razones por las que importa avanzar en la incorporación activa de estudios arqueozoológicos en general y metodológicos en particular.
En síntesis, vale la pena alentar la exploración metodológica y explotar la capacidad de innovación que ofrece el caldo de cultivo creado por las excursio-nes teóricas muchas veces eclécticas de la arqueozoología en América Latina. Parte de esta búsqueda incluye lo que denomino “tafonomía irrestricta”.
Tafonomía irrestrictaAlgunas recientes revisiones de la historia de la arqueología o la arqueozología en América Latina han destacado la importancia de la tafonomía (Politis, 2003; Mengoni Goñalons, 2007; De Queiroz y De Carvalho, 2008; Quiroz, 2009). Muy adecuadamente la tafonomía se ha concentrado en el estudio de huesos, dado que éstos se adaptan más al análisis de procesos en escalas cortas. En otras palabras, su estudio se desarrolla en condiciones bajo las cuales el trabajo actualístico es más sencillo. Dado que el tiempo de reciclado óseo se mide en décadas, la vida nos alcanza para observar cambios significativos. Además, informan muy sensiblemente sobre su historia de vida y, cuando se destruyen, lo hacen rápidamente. Así, los estudios tafonómicos óseos nos han permitido sensibilizarnos con los ritmos de depositación, preservación y destrucción ósea. Más allá de esto, se espera que su uso sirva también para la comprensión de las historias de otros materiales. Éste es el gambito que permite proyectar la aventura ósea tafonómica a otros planos y otros tipos de restos. La tafonomía

2 7 0
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.267-274
ósea sirve como control, por ejemplo, de la historia de exposición subaérea de conjuntos arqueológicos que incluyen restos no óseos (Martin et al., 2004; Borrazzo, en preparación).
Entonces, habiendo sentado una sólida base en tafonomía ósea, el próximo desarrollo está en no restringir la tafonomía a los huesos –los estudios de tejidos blandos (Finucane, 2007) o moleculares (Willerslev y Cooper, 2005; Orlando et al., 2009) ya hace bastante tiempo que constituyen una parte integral– o no pen-sar la arqueozoología sólo en función de “animales”.
La tafonomía estudia la tensión constante entre medios preservaciona-les y medios destructivos1. Al ocuparse de sentar las bases para discusiones arqueológicas posteriores, no se interesa sólo en la fauna. La mera inclusión de conceptos tafonómicos en una discusión arqueozoológica usualmente deberá llevarnos a trascender el mundo de los huesos, o el de los materiales orgánicos en general. Las funciones de una perspectiva tafonómica no sólo incorporan criterios formacionales, sino que llevan toda la discusión a un estado de siner-gia en el que todas las clases de materiales –sedimentos, plástico, restos líticos o cerámicos, concreto, huesos, etc.– informan acerca de todo lo demás. No se trata meramente de desarrollar, por ejemplo, una tafonomía lítica (Hiscock, 1985; Borrazzo, 2006), cerámica (Ozán, 2009) o de fitolitos (Piperno, 1985) –todas incorporaciones positivas– sino de utilizar interactivamente los dis-tintos marcadores. La fidelidad a esta posición permitirá incluir un espectro mucho más amplio de objetivos y expandirá los resultados. Si bien existen ante-cedentes para la mayoría de estos desarrollos, en América Latina adquieren un buen potencial de integración, cosa aún no totalmente formalizada en otros lugares. En este sentido, puede ser una ventaja la actual falta de sistematización de los componentes tafonómicos de una investigación que es observable en este sector del mundo, ya que facilitará esa expansión.
He dicho, con otras palabras, que la tafonomía está detrás de toda empresa arqueológica. Simplemente no podemos creerle al registro en la forma en que se nos presenta sin una preocupación acerca de su historia formacional y pre-servacional (Gutiérrez, 2009). Siendo estrictos, se pueden concebir e identifi-car marcadores tafonómicos que abarquen toda la gama de procesos de inte-rés arqueológico, desde los de mezcla hasta los indicadores de “depositación no funcionalmente significativa” –conjuntos de bifaces sin uso y/o quebrados (Carr y Boszhardt, 2003), taxa representados diferencialmente (Corona, 2007), segmentos de animales que no responden a marcos de referencia económicos
1 Nomepreocupoaquíporcuestionesnominales, talescomoelexcesivorespetoaunadefiniciónclásicade“tafonomía”.Entiendoquelostrabajosclásicosdelinearonuncaminoque,pornecesidad,deberáestarsignadoporlasadicionesytransformaciones.

2 7 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.267-274
L A F U N C I Ó N T R A N S D I S C I P L I N A R I A D E L A A R Q U E O Z O O L O G Í A E N E L S I G L O X X I | L U I S A L B E R T O B O R R E R O
(Stahl, 2004; Politis, 2007), etc.–. El punto es que toda la búsqueda y discusión interpretativa estarán asentadas sobre una base sólida.
Dicho esto, hay que enfatizar que el trabajo tafonómico clásico, caracteri-zado por el enfoque naturalista (cf. Marean, 1995), aún no ha dejado de prestar servicios. Tal vez se ha pensado que esa forma de adquisición de conocimiento ya cumplió su tarea, pero en realidad apenas ha comenzado a desarrollarla. Por ejemplo, los poderosos resultados de la tafonomía ósea naturalista aún deben ser explorados en su dimensión geográfica. ¿Cuáles son los valores de las varia-bles en distintos ambientes, latitudes, alturas, etcétera? A esto se suman las apenas iniciadas series de observaciones sobre otros tipos de materiales, tales como concreto o plástico (ver Weisman, 2007). No hay nada que sugiera que esta tarea deba esperar por la existencia de un interés masivo por una arqueo-logía que se ocupe de los mismos (ver Gould y Schiffer, 1981). Éste es un campo en el que es posible realizar las observaciones actualísticas antes de la inicia-ción de los programas intensivos de investigación, una situación que permitirá seleccionar las variables adecuadas de manera informada.
En otras palabras, aún nos falta conocer ritmos básicos de la naturaleza en relación con nuestros problemas de investigación. El trabajo del arqueozoólogo, como se sigue de las bases epistemológicas de la teoría de la evolución, míni-mamente exige un conocimiento de la variación exhibida por un fenómeno. Esto significa, sencillamente, que es mucho más lo que queda por conocer que lo ya hecho. No conozco proposición más alentadora que ésa. .

2 7 2
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.267-274
R e f e R e n c i a s
Araujo, Adauto, Karl Reinhard y Luis Fernando Ferreira 2008.“ParasiteFindingsinArchaeologicalRemains:DiagnosisandInterpretation”,Neotropical
Zooarchaeology and Taphonomy(eds.S.MuñozyM.Mondini)Quaternary International180(1),pp.17-21.
Arroyo-Cabrales, Joaquín, Oscar J. Polaco, César Laurito, Eileen Johnson, María Teresa Alberdi y Ana Lucía Valerio Zamora
2005.“TheProboscideans(Mammalia)fromMesoamerica”,Quaternary International169-170,pp.17-23.
Beret ta, J.M. 2008.“Laetnoarqueologíaylasedentarizaciónobligadadelosmby’adelvalledeCuñapirú”.Relaciones
de la Sociedad Argentina de Antropología33,pp.85-112.
Borella, F., M. Gutiérrez, H.R. Foderé y J. F. Merlo 2007.“Estudiodedensidadmineralóseaparadosespeciesdeotáridosfrecuentesenelregistro
arqueofaunísticopatagónico(Otaria flavescensyArctocephalus australis)”,pp.421-426.
Borrazzo, Karen 2006.“Tafonomíalíticaendunas:unapropuestaparaelanálisisdelosartefactoslíticos”,Revista
Intersecciones en Antropología 7,pp.247-261.Enpreparación.“Arqueologíadelosestepariosfueguinos”.TesisdeDoctorado,UniversidaddeBuenos
Aires.
Borrero, Luis Alberto 2004.“ArqueologíaenAméricadelSur.¿Serequiereunacercamientoespecial?”,enTeoría
arqueológica en América del Sur, eds.G.PolitisyR.D.Peretti,pp.71-83.Olavarría,UNCPBA.
Carr, D. y R. Boszhardt 2003.“TheKrieselCache:ALatePaleoindianCachefromWesternWisconsin”,Plains Anthropologist48,
pp.225-235.
Corona Martinez, E. 2007.“ZoogeogeographicalAffinitiesandtheUseofVertebralesinXochicalco”,Quaternary
International180(1),pp.145-151.2008.Las aves en el Cenozoico tardío de México.Madrid,UniversidadAutónomadeMadrid.
Nogueira De Queiroz, Albérico N. y Olivia A. De Carvalho 2008.“ProblemsintheInterpretationofBrazilianArchaeofaunas:DifferentContextsandtheImportant
RoleofTaphonomy”,Neotropical Zooarchaeology and Taphonomy(eds.S.MuñozyM.Mondini) Quaternary Internacional180(1),pp.75-89.
Elkin, Dolores 1995.“VolumeDensityofSouthAmericanCamelidsSkeletalParts”,International Journal of
Osteoarchaeology5,pp.29-37.
Finucane, Brian C. 2007.“Mummies,MaizeandManure:Muti-tissueStableIsotopeAnalysisofLatePrehistoricHuman
RemainsfromtheAyacuchoValley,Perú”,Journal of Archaeological Science 34,pp.2115-2124.
Gould, Richard y Michael Schif fer 1981.Modern Material Culture: The Archaeology of US.NuevaYork,AcademicPress.
Gutiérrez, María 2009.“Tafonomía:¿tiraníaomultivocalidad?”,enPerspectivas actuales en Arqueología Argentina,eds.
R.Barberena,K.BorrazzoyL.A.Borrero,pp.57-87.BuenosAires,CONICET-IMHICIHU.

2 7 3
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.267-274
L A F U N C I Ó N T R A N S D I S C I P L I N A R I A D E L A A R Q U E O Z O O L O G Í A E N E L S I G L O X X I | L U I S A L B E R T O B O R R E R O
Gutiérrez, M. y C. Kaufmann 2007.“CriteriafortheIdentificationoftheFormationProcessesinGuanaco(Lama guanicoe)Bone
AssemblagesinFluvial-LacustrineEnvironments”,Journal of Taphonomy5(4),pp.151-176.
Hiscock, Peter 1985.“TheNeedforaTaphonomicPerspectiveinStoneArtefactAnalysis”,Queensland Archaeological
Research 2,pp.82-95.
Kaufmann, Cristian 2009.Estudio de edad y sexo en guanaco. Estudio actualístico y arqueológico en Pampa y Patagonia.
BuenosAires,SociedadArgentinadeAntropología.
L’Heureux, Gabriela L. 2005.“VariaciónmorfométricaenrestosóseosdeguanacodesitiosarqueológicosdePatagoniaaustral
continentalydelaIslaGrandedeTierradelFuego”,Magallania3(1),pp.81-94.
Lyman, R. Lee2006.“PaleozoologyintheServiceofConservationBiology”,Evolutionary Anthropology15,pp.11-19.
Marean, C. 1995.“OfTaphonomyandZooarchaeology”,Evolutionary Anthropology4,pp.64-72.
Marschoff, M. 2007.Gato por liebre. Prácticas alimenticias en Floridablanca.BuenosAires,Teseo.
Martin, F.M., R. Barberena y R. A. Guichón 2004.“Erosiónyhuesoshumanos.ElcasodelalocalidadChorrillos,TierradelFuego”,Magallania32,
pp.125-142.
Martinez, G. 2007.“PatternsofBoneRepresentedandSufaceBoneModificationCausedbyNukakPreyAcquisition.
Appendix2”,en Ethnoarchaeology of an Amazonian People,ed.G.Politis,pp.357-376.WalnutCreek,LeftCoastPress.
Mengoni Goñalons, G. L. 2004.“Introduction:AnOverviewofSouthAmericanZooarchaeology”,enZooarchaeology of South
America,ed.G.L.MengoniGoñalons,pp.1-9.Oxford,BARInternationalSeries1298.2007.“ArchaeofaunalStudiesinArgentina:AHistoricalOverview”,enTaphonomy and Zooarchaeology
in Argentina,eds.M.Gutiérrez,L.Miotti,G.Barrientos,G.L.MengoniGoñalonsyM.Salemme,pp.13-34.Oxford,BARInternationalSeries1601.
Mengoni Goñalons, G. L., D.Olivera y H. Yacobaccio 2000. El uso de los camélidos a través del tiempo.BuenosAires,GZC-ICAZ.
Orlando, L., J. L. Metcalf, M. T. Alberdi, M. Telles-Antunes, D. Bonjean, M. Otte, F. Martin, V. Eisenmann, M. Mashkouri, F. Morello, J. L. Prado, R. Salas-Gismondi, B. J. Shockey, P. J. Wrinn, S. K. Vasil’ev, N. D. Ovodov, M. I. Cherry, B. Hopwood, D. Male, J. J. Austin, C. Hanni, y A. Cooper
2009.Revising the recent Evolutionary History of Equids Using Ancient DNA.ProceedingsNationalAcademyofSciences,EarlyEdition,www.pnas.org_cgi_doi_10.1073_pnas.0903672106
Ozán, I. L. 2009.“Procesosdeformacióndelregistrocerámicoencazadoresrecolectoresdelcentro-estey
sudoestedelaProvinciadeLaPampa”.TesisdeLicenciatura,UniversidaddeBuenosAires.
Piperno, Dolores1985.“PhytolithTaphonomyandDistributionsinArchaeologicalSedimentsfromPanama”,Journal of
Archaeological Science12,pp.247-267.

2 7 4
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antipod.Rev.Antropol.Arqueol.No.13,Bogotá,julio-diciembre2011,292pp.ISSN1900-5407,pp.267-274
Polaco, O. J., Joaquín Arroyo-Cabrales, Eduardo Corona-M., J. G. Oliva-López 2001.“TheAmericanMastodonMammutamericanuminMéxico”,enLa Terra degli Elefante,eds.,
G.Cavarretta,P.Gioia,M.MussiyM.R.Palombo,pp.237-242:Roma,ConsiglioNazionaledelleRicerche.
Politis, Gustavo 2003.“TheTheoreticalLandscapeandtheMethodologicalDevelopmentofArchaeologyinLatin
America”,American Antiquity68(2),pp.245-272.2007.Ethnoarchaeology of an Amazonian People.WalnutCreek,LeftCoastPress.
Quiroz, Daniel 2009.“ZooarqueologíaenChile:historia,problemas,perspectivas”,enZooarqueología y tafonomía
en el confín del mundo,eds.P.López,I.Cartajena,C.GarcíayF.Mena,pp.15-26.Santiago,UniversidadInternacionalSEK.
San Román, M. 2009.“AnatomíaeconómicadeOtaria flavescens[Shaw,1800]”,enZooarqueología y tafonomía
en el confín del mundo,eds.P.López,I.Cartajena,C.GarcíayF.Mena,pp.169-180.Santiago,UniversidadInternacionalSEK.
Scheinsohn, Vivian 2009.“Evoluciónenlaperiferia.ElcasodelaarqueologíaevolutivaenlaArgentina”,enArqueología y
evolución. Teoría, metodología y casos de estudio,eds.G.LópezyM.Cardillo,pp.73-86.BuenosAires,EditorialSB.
Scheinsohn, Vivian y J. L. Ferret ti 1995.“TheMechanicalPropertiesofBoneMaterialsinRelationtotheDesignandFunctionofPrehistoric
ToolsfromTierradelFuego(Argentina)”,Journal of Archaeological Science22,pp.711-717.
Stahl, Peter 1999.“StructuralDensityofDomesticatedSouthAmericanCameliaSkeletalElementsandthe
ArchaeologicalInvestigationofPrehistoricAndeanCh’arki”,Journal of Archaeological Science26,pp.1347-1368.
2004.“NeotropicalZooarchaeologyinEcuador”,enZooarchaeology of South America,ed.G.L.MengoniGoñalons,pp.203-220.Oxford,BARInternationalSeries1298.
2008.“TheContributionofZooarchaeologytoHistoricalEcologyintheNeotropics”,Neotropical Zooarchaeology and Taphonomy (eds.S.MuñozyM.Mondini).
Weisman, Alan 2007.The World without Us.NuevaYork-Londres,Picador.
Willerslev, E. y Alan Cooper 2005.“AncientDNA”.ReviewPaper,Proceedings Royal Society of LondonB.272,pp.3-16.
Yacobaccio, Hugo 2007.“AndeanCamelidHerdingintheSouthAndes:EthnoarchaeologicalModelsforArchaeozoological
Research”,Anthropozoologica42(2),pp.143-154.

2 7 5
C r é d i t o s i m á g e n e s A n t í p o d a 1 3 MargaritaMonsalveyJuanAntonioMonsalve©
P o R t a d a Diente de león2004.
G u a R d a i n i c i a l
Estructuras2004.
i m a G e n P á G i n a 2 6 :PatinadoresNuevaYork,2009.
i m a G e n P á G i n a 9 6 :Parque de diversionesMexicoDF,2002.
i m a G e n P á G i n a 2 2 6 :Cony islandNuevaYork,2009.
i m a G e n P á G i n a 2 5 2 :Pérgolas Barcelona,2004.
G u a R d a f i n a l :Tensores(det)2004.

2 7 6

Normas

2 7 8
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Antípoda. Revista de Antropología y Arqueología, de la Universidad de los Andes es una publicación semestral que se proyecta como un espacio de encuentro y discusión sobre temas de interés para la disciplina antropológica. Su objetivo es incentivar el intercambio académico entre la Antropología y la Arqueología con otras disciplinas sociales y humanas. La revista tiene un interés especial en desarrollar y profundizar en los análisis de dichas áreas en torno a problemáti-cas actuales y regionales. Dado nuestro interés por crear lazos entre pensamientos académicos de otros contextos nacionales y continentales para discutir temas hispanoame-ricanos, la revista Antípoda publica textos inéditos en español, inglés y por-tugués. En volúmenes especiales, y de acuerdo con la decisión del Comité Editorial, se pueden publicar artículos en otros idiomas con su respectiva traducción al español.
Proceso editorial• Cadanúmerodelarevistacontieneundossier especial en donde se presen-
tan artículos relacionados con el tema principal de la publicación. Este tema es sugerido por el Comité Editorial o a partir de las propuestas recibidas por académicos interesados en participar como editores invitados.
• Unavezseleccionadoeltemadelnúmero,larevistalanzaconseismesesdeanticipación una convocatoria por un período determinado a través de dis-tintas redes académicas. De forma regular, se reciben artículos que cumplan con el objetivo general de la convocatoria.
• Sesugiereconsultarconlaeditorageneralantesdeenviartrabajosenidio-mas distintos al castellano. [email protected]
• Losartículospresentadosnodebenestarenprocesodeevaluaciónnitenercompromisos editoriales con ninguna otra publicación.
• Los autores son responsablesde conseguir lospermisos correspondientespara la reproducción de imágenes, ilustraciones, figuras y citas extensas. La revista puede apoyar para solicitar dichos permisos. Una vez se cierra la con-vocatoria, orientada y coordinada por el (los) editor(es) invitados, el Comité Editorial en pleno hace una revisión cuidadosa de los artículos presentados. Allí se evalúa si el escrito cumple con los requisitos básicos exigidos por la revista y su pertinencia para figurar en cada número temático de Antípoda.
• Una vez se cierra la convocatoria, orientada y coordinada por el (los)editore(es) invitados, el Comité Editorial en pleno hace una revisión cui-
N o r m a s a c t u a l i z a d a s p a r a p u b l i c a r

2 7 9
dadosa de los artículos presentados. Allí se evalúa si el escrito cumple con los requisitos básicos exigidos por la revista y su pertinencia para figurar en cada número temático de Antípoda.
• Posteriormente,losartículosseleccionadossonsometidosaevaluaciónporparte de dos árbitros anónimos, sugeridos por el (los) editor(es) invitado(s) y avalados por el Comité Editorial. La decisión final se informará a los auto-res en un plazo máximo de seis meses.
• Elequipoeditorialseencargarádeenviarunconceptoinicialacadaautor(aprobado, aprobado con cambios o rechazado). Igualmente, se inicia un proceso de seguimiento a los artículos aceptados, que consta de las siguien-tes etapas: evaluación, corrección por autores, revisión editorial y correc-ción de estilo. Las modificaciones y correcciones del artículo deberán ser realizadas por el autor en el plazo que le será indicado por el equipo edito-rial de la Revista.
• Alfinalizarelprocesodeevaluación,el(los)editor(es)invitado(s),juntoconel equipo editorial, deberá(n) revisar los conceptos recibidos y determinar cuáles serán los artículos publicados. Igualmente, definirán en qué sección serán publicados y qué cambios requieren. Esta decisión será comunicada al autor tan pronto se tenga toda la documentación que le es solicitada en el plazo indicado.
• Losautoresdelostextosaceptadosautorizan,mediantelafirmadel"docu-mento de autorización" de uso de derechos de propiedad intelectual, la uti-lización de los derechos patrimoniales de autor (reproducción, comunica-ción pública, transformación y distribución) a la Universidad de los Andes, Departamento de Antropología, para incluir el texto en Antípoda. Revista de Antropología y Arqueología (versión impresa y versión electrónica). En este mismo documento los autores confirman que el texto es de su autoría y que en el mismo se respetan los derechos de propiedad intelectual de terceros.
• Alfinalizaresteproceso,lostextosnoserándevueltos.• Losautoresrecibirándosejemplaresdelnúmeroenelqueparticiparon.• Encasodequeunartículoquisieraincluirseposteriormenteenotrapubli-
cación, deberán señalarse claramente los datos de la publicación original en Antípoda. Revista de Antropología y Arqueología, previa autorización soli-citada a la dirección de la revista.
Requisitos para la presentación de artículosAntípoda recibe textos en español máximo de veinticinco (25) páginas (8.500 palabras Aprox.). En otros idiomas, no podrán sobrepasar las veinte (20) pági-nas (7.000 palabras Aprox.), (incluídos pies de página y bibliograf ía).

2 8 0
A N T Í P O D A N º 1 3 | J U L I O - D I C I E M B R E 2 0 1 1
Al presentar un artículo, se deben cumplir las siguientes especificaciones:• Los trabajos deberán presentarse en el siguiente formato: tamaño carta,
márgenes de 2,54 cm, doble espacio, letra Times New Roman 12 puntos, numeración de página desde 1 hasta n, en el margen superior derecho, con un uso mínimo de estilos –negrita sólo para títulos y subtítulos; cursiva para énfasis dentro del texto–.
• Cada texto debe incluir el título del artículo y un resumen en español einglés que no supere las 80 palabras, y 5 palabras clave en español e inglés.
• Debe incluirse en pie de página un breve perfil del autor o autores queincluya su actual vinculación institucional, el título académico más alto con la institución que lo otorgó, y su correo electrónico.
• Todoslosdiagramasyfotograf íasserándenominados“Figuras”,lascualesdeben ser identificadas en el texto mediante un pie de foto.
• Todas las imágenesdebensernumeradasconsecutivamente.Deberánsereviandas en formato .tiff de alta resolución, es decir de 300 pixeles por pul-gada (ppp).
• La formadecitacióndebecorrespondera lasnormasChicagoadaptadaspor Antípoda.
• Lasreferenciasbibliográficadebenestarorganizadasenestrictoordenalfa-bético. Ver modelos de presentación de los datos bibliográficos que apare-cen en los siguientes ejemplos:
Libros:Apellido, NombreAño Título. Ciudad, Editorial.
Behrensmeyer, Anna Kay y Andrew Hill1980 Fossils in the Making: Vertebrate Taphonomy and Paleoecology. Chicago, University of Chicago Press.
Capítulo de libro o artículo de contribución:Uribe, Carlos Alberto1992 Aculturación. En Palabras para desarmar, eds. Margarita Serje, María Cristina Suaza y Roberto Pineda, pp.25-37. Bogotá, ICANH.
Artículos en revistas:Bradley, Bruce y Dennis Standford 2004 The North Atlantic Ice-edge Corridor: A Possible Paleolithic Rout to the New World. World Archaeology 36 (4), pp. 459-478.

2 8 1
Se recomienda a los convicados tener en cuenta estas normas actualizadas a partir de Antípoda número 14.
Como revista académica, Antípoda respeta y está abierta a diferentes líneas de pensamiento. Sin embargo, Antípoda no se hace responsable de las opiniones y conceptos de los autores que aparecen en cada número.




2 8 5




2 8 9
C O N V O C A T O R I A
ANTROPOLOGÍA Y LITERATURA
Desde Herodoto hasta Frazer las relaciones entre la antropología y la lite-ratura han sido estrechas. No pocas veces los cuadros etnográficos han estado acompañados de relatos ficticios en los cuales el pensamiento antropológico ha sido plasmado en textos de carácter literario que, en ocasiones, han tenido impacto mayor que el de la producción propiamente académica. A su vez, los antropólogos —como lo señalaron Clifford y Marcus en su ya clásico Writing Culture (1986)—, al hilar sus representaciones culturales en los textos etnográ-ficos, han construido no sólo políticas de la diferencia, sino verdaderas geopoé-ticas en la cuales quedan involucrados como personajes o narradores literarios.
Para su edición número 15, Antípoda, con el fin de aportar en este campo y de hacer una aproximación sistemática a la compleja relación entre antropo-logía y literatura, está convocando contribuciones que propongan reflexiones sobre los espacios de intersección entre éstas poniendo en evidencia tanto la forma en que la literatura ha alimentado el pensamiento antropológico como la manera en que la antropología y, en particular, la etnograf ía se han constituido como prácticas narrativas.
Editores invitados: Margarita Serje, profesora Departamento de Antro-pología Universidad de los Andes, Colombia; Juan Carlos Orrego, profesor Departamento de Antropología Universidad de Antioquia, Colombia.Fecha límite para la recepción de artículos: 12 de marzo de 2012Los artículos pueden ser enviados a:antí[email protected]@[email protected]
Más información en: http://antipoda.uniandes.edu.co/page.php?c=Convocatorias

2 9 0
C A L L O F P A P E R S
ANTHROPOLOGY AND LITERATURE
From Herodotus to Frazer relations between anthropology and literature have been close. Not infrequently were the ethnographic pictures accompanied by fictitious stories in which anthropological thought has been expressed in literary texts that, at times, have had greater impact than the actual academic production. In turn, anthropologists like Clifford and Marcus said in their clas-sic Writing Culture (1986) – by spinning cultural representations in ethnogra-phic texts they have built not only policies of difference, but real geo-poetics in which they are involved in as literary characters and narrators.
For its 15th edition, Antípoda, in order to contribute to this field and make a systematic approach to the complex relationship between anthropology and literature, it’s calling for contributions to propose reflections on the inter-section between these, highlighting both how literature has fueled anthropo-logical thought and the way anthropology and ethnography in particular, have become narrative practices.
Guest Editors: Margarita Serje, Professor at the Department of Anthro-pology of the University of the Andes (Universidad de los Andes), Colombia Juan Carlos Orrego, Professor at the Department of Anthropology of the University of Antioquia (Universidad de Antioquia), Colombia
Deadline for receipt of articles: March 12, 2012
The articles can be submitted to:antí[email protected] [email protected] [email protected]
For more information go to: http://antipoda.uniandes.edu.co/page.php?c=Convocatorias

2 9 1
C H A M A D A
ANTROPOLOGIA E LITERATURA
Desde Herodoto até Frazer as relações entre a antropologia e a literatura sempre foram muito próximas. Não poucas vezes, os quadros etnográficos vieram acompanhados de relatos fictícios em que o se colocava o pensamento antropológico em textos de caráter literário que, ocasionalmente, tiveram maior impacto comparado com aquele da produção acadêmica própria. Por sua vez, os antropólogos, como já apontaram Clifford e Marcus no clássico Wri-ting Culture (1986)— enquanto lavram as representações culturais nos textos etnográficos, construíram não apenas políticas da diferença, mas verdadeiras geo-poéticas em que personagens ou narradores literários ficam envolvidos.
Para a edição número 15, Antípoda, no intuito de contribuir neste campo e de se aproximar sistematicamente à complexa relação entre antropologia e literatura, pretende reunir contribuições que levantem reflexões sobre os espaços de encontro entre elas, evidenciando tanto a forma em que a literatura tem alimentado o pensamento antropológico quanto a forma em que a antro-pologia, e nomeadamente, a etnografia, tornaram-se práticas narrativas.
Editores invitados: Margarita Serje, professora Departamento de Antropo-logia Universidade de los Andes, Colômbia; Juan Carlos Orrego, professor Departamento de Antropologia Universidade de Antioquia, Colômbia.
Data limite para recebimento de artigos 12 de março de 2012.
Os artigos podem ser enviados a:antí[email protected] [email protected] [email protected]
Mais informações: http://antipoda.uniandes.edu.co/page.php?c=Convocatorias
