rodrigo josé custodio - livros grátislivros01.livrosgratis.com.br/cp142429.pdf · preparada pela...
TRANSCRIPT

Rodrigo José Custodio
Hipóxia sanguínea relacionada a doenças respiratórias agudas e a regulação do
sistema IGF em crianças.
Tese de doutorado apresentada à
Faculdade de Medicina de Ribeirão
Preto da Universidade de São Paulo
para obtenção do título de Doutor em
Saúde da Criança e do Adolescente –
Opção: Investigação em Pediatria.
Orientador: Prof. Dr. Carlos Eduardo
Martinelli Júnior
Ribeirão Preto
2010

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

Autorizo a reprodução e divulgação total ou parcial deste trabalho, por qualquer meio
convencional ou eletrônico, para fins de estudo e pesquisa, desde que citada a fonte.
Catalogação da Publicação
Preparada pela Biblioteca Central do Campus da Universidade de São Paulo de
Ribeirão Preto
Faculdade de Medicina de Ribeirão Preto da Universidade de São Paulo
FICHA CATALOGRÁFICA
Custodio, Rodrigo José
Hipóxia sanguínea relacionada a doenças respiratórias agudas
e a regulação do sistema IGF em crianças. Ribeirão Preto, 2010.
129 p.: il. ; 30cm
Tese de Doutorado, apresentada à Faculdade de Medicina de
Ribeirão Preto/USP. Área de concentração: Saúde da Criança e do
Adolescente – Opção: Investigação em Pediatria.
Orientador: Martinelli Júnior, Carlos Eduardo.
1. IGF. 2. IGFBPs. 3. IGF1R. 4. Hipóxia. 5. Criança.

Custodio RJ. Hipóxia sanguínea relacionada a doenças respiratórias agudas e a
regulação do sistema IGF em crianças.
Tese de doutorado apresentada à Faculdade de Medicina de Ribeirão Preto da
Universidade de São Paulo para obtenção do título de Doutor em Saúde da Criança e do
Adolescente – Opção: Investigação em Pediatria.
Aprovado em:
Banca Examinadora
Prof. Dr.________________________________ Instituição:_____________________
Julgamento:______________________________ Assinatura:_____________________
Prof. Dr.________________________________ Instituição:_____________________
Julgamento:______________________________ Assinatura:_____________________
Prof. Dr.________________________________ Instituição:_____________________
Julgamento:______________________________ Assinatura:_____________________
Prof. Dr.________________________________ Instituição:_____________________
Julgamento:______________________________ Assinatura:_____________________
Prof. Dr.________________________________ Instituição:_____________________
Julgamento:______________________________ Assinatura:_____________________

Dedicatória
Aos meus inesquecíveis e amados pais, Vanderlei (in memorian) e Neli,
paradigmas de retidão e honestidade que alicerçaram no esforço contínuo todo o meu
progresso;
Aos meus irmãos, Fabrício, Letícia e Ludmila: a palavra irmão encerra
tudo;
Aos meus cunhados Carla, Júnior e Rogério por completarem a grande
família;
Aos meus sobrinhos Giovana, Enzo, Laura e Isadora pela imensa alegria
que nos proporcionam;
Aos meus sogros, Antônio e Maria, colaboradores sempre presentes e
zelosos, impossível imaginar a conclusão desse trabalho sem o enorme auxílio prestado;
À Viviane. Minha eterna colega de sala de aula, namorada, noiva, esposa,
colega de profissão, mãe, crítica, pesquisadora, entusiasta Viviane. Pessoa, que viveu, e
vive profundamente todos os meus mais importantes momentos e que realmente sabe
como os sinto;
Ao Plínio e ao Henrique, que trouxeram consigo uma nova perspectiva
para a minha vida, e que, nesse pouco tempo de convívio, já mudaram meus passos
diversas vezes, ensinando-me a enxergar muito além dos meus prévios e parcos limites.

Agradecimentos
Ao Prof. Dr. Carlos Eduardo Martinelli Júnior pela orientação segura,
tranquila e sempre otimista durante a condução desse estudo; pela minha formação em
endocrinologia pediátrica e muito além dela; pela convivência deveras agradável como
colega na endocrinologia; pela amizade honesta e cuidadosa que surgiu durante todos
esses anos; enfim, por ter apontado direções na minha vida que jamais imaginei nem
sequer entrever, obrigado.
Ao Prof. Dr. Carlos Alberto Scrideli, além de proporcionar toda
disponibilidade do Laboratório de Pediatria do HCFMRPUSP, colaborou com sugestões
imprescindíveis quanto às técnicas laboratoriais e análise da expressão do gene IGF1R;
as observações, sempre pertinentes, foram fundamentais para o avanço desse estudo.
À Profa. Dra. Palmira Cupo e à Profa. Dra. Sylvia Evelyn Hering pelos
exemplos que são como professoras e pediatras, e por toda a colaboração constante na
minha formação como médico. Pela oportunidade de trabalhar na Enfermaria de
Pediatria da Unidade de Emergência do HCFMRPUSP onde pude aprender muito e em
diversas dimensões, principalmente na maneira como atua um médico; enfim, pela
colaboração no recrutamento de indivíduos participantes desse estudo oriundos da
Enfermaria pela qual são responsáveis. Agradeço também, especialmente à Profa. Dra.
Palmira, pelas valiosas sugestões na elaboração desse estudo.
À Profa. Dra. Maria Célia Cervi e à Dra. Márcia de Lima Isaac pelo
auxílio em efetuar as observações em pacientes internados sob seus cuidados na
Enfermaria de Moléstias Infecciosas da Unidade de Emergência do HCFMRPUSP.

À Profa Dra. Enilza Espreáfico que, nesse momento importante, jamais
poderia me esquecer de tudo o que me proporcionou ainda como aluno de iniciação
científica. Foram marcas indeléveis em meu conhecimento como pesquisador que
facilitaram substancialmente o meu progresso acadêmico. Agradeço imensamente.
À Profa. Dra. Alcione Artioli Machado, ao Prof. Dr. João Carlos da
Costa e à Profa. Dra. Maria Inez Machado Fernandes por continuarem o trabalho
cuidadoso da Profa. Enilza, guiando-me pelo método científico, porém naquela ocasião
com enfoque médico.
À equipe de Enfermagem da Pediatria da Unidade de Emergência do
HCFMRPUSP, especialmente, às enfermeiras Suzete, Estela, Ana Cláudia, Valéria,
Márcia, Jaqueline, Sandra e Marilurdes pelo auxílio na coleta das amostras; e a todas
Auxiliares de Enfermagem, em especial à Lindamir, Rita, Maria Antônia, enfim a todas,
pela paciência com que atenderam os meus telefones em momentos mais improváveis.
Aos colegas Regina e Wagner pela convivência e a compreensão durante
a condução desse estudo.
À responsável pelo Laboratório de Toxicologia da Unidade de
Emergência do HCFMRPUSP Maria José Leite e a Consuelo, Juliana, e especialmente à
Marisa Elias Amêndola por ter colaborado de forma constante com o adequado
acondicionamento das amostras, sempre prestativa.
A todos os médicos residentes do Departamento de Puericultura e
Pediatria da FMRPUSP que cumpriram seus programas entre os anos de 2006 e 2009,
pela tolerância com a minha insistência em incluir novos indivíduos no estudo e pela
efetiva inclusão desses. Sem a vigilância desse imenso grupo, esse estudo seria inviável.
A todos os técnicos e pós-graduandos que frequentam e trabalham no
Laboratório de Pediatria do HCFMRPUSP. Não poderia deixar de citar, dentre eles, a

Dra. Rosane Queiroz, Maria Angélica Abdalla de Freitas Cortez, Vanessa da Silva
Silveira, Priscila Leite e Daniel Antunes Moreno, por tudo, enfim, por tudo que me
ensinaram com a mais profunda paciência e rara harmonia vista nesse tipo de ambiente
de trabalho.
Aos pós-graduandos do Departamento de Puericultura e Pediatria,
especialmente àqueles da Divisão de Endocrinologia Pediátrica: Soraya Sader Milani,
Fábio Rosa, Ana Cláudia, Sabrina e Marcelo Ruiz pela convivência, compreensão e
sugestões durante a coleta e análises laboratoriais.
Às secretárias do Departamento de Puericultura e Pediatria da
FMRPUSP, Dulcides, Vera e Sandra pela amizade e colaboração durante toda a pós-
graduação.
Às famílias das crianças que permitiram a inclusão de seus filhos nesse
estudo: agradeço profundamente.
À Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) e
ao Conselho Nacional de Pesquisa (CNPq) pelo auxílio financeiro.

“Sonhar é acordar-se para dentro”.
Mário Quintana
“...Ah, eu quero te dizer que o instante de te ver custou tanto penar
Não vou me arrepender,
Só vim te convencer que eu vim prá não morrer
De tanto te esperar,
Eu quero te contar das chuvas que apanhei
Das noites que varei
No escuro a te buscar
Eu quero te mostrar
As marcas que ganhei
Nas lutas contra o rei
Nas discussões com Deus,
E agora que cheguei
Eu quero a recompensa
Eu quero a prenda imensa
Dos carinhos teus...”
Chico Buarque de Hollanda.

Resumo
CUSTODIO RJ. Hipóxia sanguínea relacionada a doenças respiratórias agudas e a
regulação do sistema IGF em crianças [tese]. Ribeirão Preto: Universidade de São
Paulo, Faculdade de Medicina de Ribeirão Preto, 2010. 129 f.
A hipóxia, relacionada à restrição do crescimento intra-uterino em
animais, foi capaz de promover aumento na concentração sérica da IGFBP-1 sem
alterações nas concentrações séricas de IGF-I. Além disso, a hipóxia grave provoca
aumento na expressão do IGF1R de cones neuronais de crescimento dos fetos de
ovelhas. Entretanto, em situações clínicas graves, houve redução das concentrações
séricas do IGF-I e aumento da IGFBP-1 sérica. Porém, há pouca informação sobre a
regulação do sistema IGF exercida pela hipóxia aguda em humanos. O objetivo desse
estudo foi avaliar o sistema IGF e sua relação com a hipóxia aguda em crianças
previamente normais. Para tanto foram dosadas as concentrações séricas de IGF-I,
IGFBP-1, IGFBP-3 e insulina e quantificada a expressão do IGF1R em linfócitos de 27
crianças (14 meninos e 13 meninas) de 15 dias a 9,5 anos de idade com hipóxia aguda
devida a problemas respiratórios e após sua recuperação, na mesma criança cujos
valores são mostrados em médias (±DP) e medianas (P25 – P75). As concentrações de
IGF-I, IGFBP-1, IGFBP-3 e insulina foram determinadas por ELISA específico. Os
linfócitos periféricos foram isolados de outras células utilizando Fycoll-Hypaque® e,
após, o RNA foi extraído. A expressão do mRNA do IGF1R foi obtida mediante
realização de RT-PCR em tempo real. As análises estatísticas foram realizadas por
pareamento. A saturação de oxigênio foi 87,8 (±3,5)% durante a hipóxia e 96,3
(±1,2)% após recuperação (p<0.0001). Houve redução das concentrações séricas de
IGF-I durante a hipóxia aguda: 21,3 (±37,8) e 60,2 (±58,6); e medianas de 12,3 (6,24 –
20,14) e 54,8 (24,51 – 72) ng/ml (p < 0,0001); assim como das concentrações séricas da

IGFBP-3: 1,53 (±0,8) e 2,34 (±0,7); e medianas de 1,32 (0,86 – 2,17) e 2,25 (1,9 – 2,78)
mg/l (p = 0,0002). As concentrações séricas da IGFBP-1 aumentaram na hipóxia: 135,3
(±79,3) e 75,2 (±55,5); e medianas de 115,5 (77,4 – 171,6) e 68,5 (28,25 – 106,8) ng/ml
(p = 0,0014). As concentrações séricas de insulina permaneceram inalteradas: 11,7 (±7)
e 14,5 (±9,6) e medianas de 9,1 (8,48 – 11,1) e 9,8 (8,79 – 18,6) µUI/ml, nas situações
de hipóxia e sem hipóxia (p = 0,132), respectivamente. Houve aumento da expressão do
IGF1R nos linfócitos onde as médias e medianas durante a hipóxia e sem hipóxia foram
de 1,61 (±1,26) e 1,21 (±1,1); e de 1,28 (0,8 – 2,11) e 0,93 (0,41 – 1,7),
respectivamente. Após pareamento, houve diferença significativa na expressão do
IGF1R entre as duas situações (p = 0,0373). Os dados apresentados, analisados em
conjunto, sugerem que a hipóxia aguda, provavelmente, associada a fatores infecciosos,
foi capaz de estimular alterações do sistema IGF com a redução do estímulo anabólico
provocado pelo IGF-I e o aumento na expressão do gene codificador do IGF1R sugere
um mecanismo compensatório de ativação tecidual.
Palavras-chave: sistema IGF, hipóxia, crianças.

Abstract - Summary
CUSTODIO RJ. Blood hypoxia related to acute respiratory diseases and the
regulation of the IGF system in children [thesis]. Ribeirão Preto: Universidade de São
Paulo, Faculdade de Medicina de Ribeirão Preto, 2010. 129 f.
Hypoxia related to intrauterine growth restriction promotes an increase in
serum IGFBP-1 concentrations in animals, with no changes in serum IGF-I
concentrations. In addition, severe hypoxia provokes an increase in IGF1R expression
in the neuronal growth cones of sheep fetuses. However, in serious clinical situations
there is a reduction of serum IGF-1 and an increase of serum IGFBP-1. Little
information is available about the regulation of the IGF system by acute hypoxia in
humans. The objective of the present study was to assess the IGF system and its relation
to acute hypoxia in previously normal children. To this end, the serum concentrations of
IGF-I, IGFBP-1, IGFBP-3 and insulin were determined and the expression of IGF1R
was quantitated in the lymphocytes of 27 children (14 boys and 13 girls) aged 15 days
to 9.5 years with acute hypoxia due to respiratory problems, during the crisis and after
their recovery. Data are reported as means ± SD and medians (P25 – P75). IGF-I, IGFBP-
1, IGFBP-3 and insulin concentrations were determined by specific ELISA. Peripheral
lymphocytes were isolated from other cells using Fycoll-Hypaque® and RNA was then
extracted. The expression of IGF1R mRNA was obtained by real time RT-PCR.
Statistical analyses were performed by pairing. Oxygen saturation was 87.8 ± 3.5%
during hypoxia and 96.3 ± 1.2% after recovery (p<0.0001). Mean serum IGF-I
concentrations were reduced during acute hypoxia: 213 ± 37.8 and 60.2 ± 58.6, with
medians of 12.3 (6.24 – 20.14) and 54.8 (24.51 – 72) ng/ml (p < 0.0001); as also were
mean serum IGFBP-3 concentrations: 1.53 ± 0.8 and 2.34 ± 0.7, with medians of 1.32

(0.86 – 2.17) and 2.25 (1.9 – 2.78) mg/l (p = 0.0002). Mean serum IGFBP-1
concentrations increased during hypoxia: 135.3 ± 79.3 and 75.2 ± 55.5, with medians of
115.5 (77.4 – 171.6) and 68.5 (28.25 – 106.8) ng/ml (p = 0.0014). Mean serum insulin
concentrations were unchanged: 11.7 ± 7 and 14.5 ± 9.6, with medians of 9.1 (8.48 –
11.1) and 9.8 (8.79 – 18.6) µIU/ml, in the presence and absence of hypoxia (p = 0.132),
respectively. There was an increase in IGF1R expression in lymphocytes, with means
and medians in the presence and absence of hypoxia of 1.61 ± 1.26 and 1.21 ±1.1, and
1.28 (0.8 – 2.11) and 0.93 (0.41 – 1.7), respectively. After pairing, a significant
difference in IGF1R expression was observed between the two situations (p = 0.0373).
Taken as a whole, these data suggest that acute hypoxia, probably associated with
infectious factors, was able to stimulate changes in the IGF system leading to the
reduction of the anabolic stimulus provoked by IGF-I and the increased expression of
the gene coding for IGF1R suggests a compensatory tissue-activating mechanism.
Key-words: IGF system, hypoxia, children.

Lista de abreviaturas
ADAM-9: a disintegrin and metallo-protease-9;
ALS: subunidade protéica ácido-lábil;
Bcl2 e BclX: nomenclatura derivada de B cell lymphoma;
cDNA: DNA complementar;
CRE: elementos regulatórios responsivos ao AMPc;
DEPC: dietilpirocarbonato;
DNA: ácido desoxirribonucléico;
dNTPs: desoxirribonucleotídeos fosfatados;
DP: desvio-padrão;
EDP: escore de desvio-padrão;
EDTA: etil-diamino-tetracético;
Foxo1, -o3: fator de transcrição forkhead box O1, -O3
GH: growth hormone ou hormônio de crescimento;
HIF-1 α e β: hypoxia-inducible factor-1 α e β;
HOMA-IR: homeostasis model assessment of insulin resistance;
HRE: elementos de resposta à hipóxia;
HRP: peroxidase de rábano;
IGF-I, -II: insulin-like growth factor -I, –II, ou fator de crescimento
insulina símile I, -II;
IGF1R: receptor tipo 1 de IGFs;
IGF2R: receptor tipo 2 de IGFs;
IGFBPs: Insulin-like growth factor binding proteins ou proteínas ligantes
de IGFs;

IGFBP-rPs: IGFBP-related proteins ou proteínas relacionadas às
IGFBPs;
IRS-1 e -2: substrato do receptor de insulina-1, -2;
Kb: quilobases;
Kd: dissociation constant ou constante de dissociação
kDa: quilodaltons;
MAPK: mitogen-activated protein kinase ou proteína cinase mitogênica
ativada;
MPC: morte celular programada;
mRNA: RNA mensageiro;
mTOR: mammalian target of rapamycin;
NSILA: nonsuppressible insulin-like activity ou atividade insulina-simile
não suprimível;
PAPP-A: pregnancy-associated plasma protein-A;
pb: pares de bases;
PBS: phosphate-buffered saline;
PDK-1: phosphoinositide-dependent kinase-1 ou fosfoinositol-
dependente cinase-1;
PI3K: phosphoinositide 3-kinase ou fosfatidil-inusitol-3-cinase;
PIP3: fosfatidilinositol 3,4,5-fosfato;
PKA: proteína cinase A;
PKB: Akt/proteína cinase B;
proMBP: pró-forma da proteína básica maior de eosinófilos;
QR: Quantificação relativa;
RCIU: restrição do crescimento intra-uterino;

RN: recém-nascido;
RNA: ácido ribonucléico;
RT-PCR: reverse transcription polymerase chain reaction ou transcrição
reversa - reação em cadeia da polimerase;
qPCR: PCR em tempo real;
RNs: Recém-nascidos;
RPM: rotações por minuto;
SH2: Src homology 2;
TGF- β: transforming growth factor-β;
TMB: tetrametilbenzidina.

SUMÁRIO
I - Introdução.................................................................................................................17
I.1 – O sistema IGF..........................................................................................................18
I.2 – Os receptores de IGFs.............................................................................................21
I.3 – As proteínas transportadoras dos IGFs....................................................................26
I.4 - O sistema IGF e sua relação com situações de hipóxia...........................................38
II – Objetivos..................................................................................................................44
III – Casuística e Métodos.............................................................................................46
III.1 – Crianças participantes...........................................................................................47
III.2 - Critérios de Exclusão............................................................................................50
III.3 - Desenho Experimental..........................................................................................51
III.4 - Dosagem Laboratorial...........................................................................................53
III.4.1. - Ensaio imunoenzimático do IGF-I....................................................................53
III.4.2 - Ensaio imunoenzimático da IGFBP-1................................................................54
III.4.3 - Ensaio imunoenzimático da IGFBP-3................................................................56
III.4.4 - Ensaio imunoenzimático de insulina..................................................................57
III.5 - Avaliação da expressão do IGF1R........................................................................59
III.5.1 – Separação de linfócitos......................................................................................59
III.5.2 – Extração do RNA...............................................................................................60
III.5.3 – Quantificação do RNA......................................................................................61
III.5.4 – Síntese da primeira fita de cDNA......................................................................61
III.5.5 – Controles endógenos.........................................................................................62

III.5.6 – RT-PCR.............................................................................................................62
III.5.7 – qPCR..................................................................................................................65
III.5.8 – Quantificação da expressão gênica....................................................................69
III.6 - Análise estatística..................................................................................................70
IV – Resultados..............................................................................................................71
IV.1 – Concentrações séricas de IGF-I...........................................................................72
IV.2 – Concentrações séricas de IGFBP-1.....................................................................75
IV.3 – Concentrações séricas de IGFBP-3.....................................................................78
IV.4 – Concentrações séricas de Insulina.......................................................................81
IV.5 – Expressão do IGF1R...........................................................................................84
IV.6 – Síntese dos resultados...........................................................................................88
V – Discussão..................................................................................................................90
VI – Conclusões............................................................................................................107
VII – Referências bibliográficas.................................................................................109
VIII – Apêndices e Anexos..........................................................................................126

17
I – Introdução

18
I.1 - O sistema IGF
O crescimento e o desenvolvimento do corpo humano são regulados por uma
rede complexa de fatores de ação local e sistêmica, dentre esses podem ser destacados os
integrantes do denominado sistema IGF.
Historicamente, verificou-se que os efeitos promotores do crescimento
provocados pelo hormônio de crescimento ou GH eram mediados por um fator hepático,
inicialmente chamado de “fator de sulfatação”, que tinha sua síntese estimulada pelo
próprio GH (SALMON; DAUGHDAY, 1957; HALL, 1972). Simultaneamente, outro
grupo de pesquisadores identificou uma fração, de atividade semelhante à insulina, presente
no soro, que não era suprimível quando adicionados anticorpos anti-insulina, chamada de
NSILA (FROESCH et al., 1963; ZAPF et al., 1978). Posteriormente, observou-se que esses
dois fatores eram idênticos, sendo unificada a nomenclatura para “somatomedinas”, termo
que permaneceu até o momento no qual foram sequenciados e, a partir de então, chamados
de IGFs.
A família dos IGFs é composta por 2 fatores de crescimento peptídicos: o
IGF-I e o IGF-II. Ambos apresentam alto grau de homologia com a pró-insulina, pois suas
estruturas são compostas por 4 domínios (A, B, C e D) sendo os 3 primeiros similares ao
pró-hormônio e 50% das sequências de aminoácidos dos IGFs são semelhantes à molécula
de insulina (JUUL, 2003). As moléculas do IGF-I e do IGF-II são compostas por
sequências únicas de 70 e 67 aminoácidos, respectivamente. Apresentam estruturas
constituídas por 3 pontes dissulfídricas internas sendo que o peso molecular do IGF-I é de

19
7649 dáltons e do IGF-II é de 7471 dáltons. Na figura 1 estão esquematizadas as moléculas
de IGF-I e IGF-II.
Figura 1 – Esquemas representativos das moléculas de IGF-I (1a) e IGF-II (1b)
reproduzido de MILANI, 2009 (com autorização). Os círculos brancos representam
aminoácidos distintos entre as moléculas; os verdes aminoácidos comuns e os marrons são
os aminoácidos suprimidos da molécula do IGF-I para a composição do IGF-II.
O gene codificador do IGF-I localiza-se no braço longo do cromossomo 12,
estende-se por 73 kb sendo constituído por 6 exons e 4 promoters. O IGF-II tem o seu gene

20
composto por 9 exons e 4 promoters no braço curto do cromossomo 11 e estende-se por 40
kb (SARA; HALL, 1990; SCHOFIELD, 1991; SUSSENBACH et al., 1991).
Os IGFs são produzidos na maioria dos tecidos e órgãos, entretanto o fígado
é a fonte responsável pela maior parte das concentrações plasmáticas circulantes desses
fatores, o que aponta para as ações tanto endócrinas como parácrinas dos IGFs. Não há
órgão onde ocorra armazenamento e, da mesma forma, não há órgão-alvo específico
(THISSEN; KETELSLEGERS; UNDERWOOD, 1994). Os IGFs apresentam diversas
ações biológicas sendo consideradas principais aquelas relacionadas à proliferação e
diferenciação celulares. IGF-I e IGF-II promovem tanto o aumento na síntese de DNA
quanto a divisão celular, portanto são considerados importantes estímulos mitogênicos
(JONES; CLEMMONS, 1995). São descritos inúmeros tipos celulares, tais como,
osteoblastos, adipócitos e oligodendrócitos nos quais os IGFs atuam como indutores da
diferenciação celular (SARA; HALL, 1990). Ainda no ciclo celular, os IGFs são capazes de
influenciar na MCP, processo no qual ocorre o desmantelamento e o acondicionamento do
material celular. Vários tipos celulares tiveram esse processo inibido pela resultante de
ações promovida pelo sistema IGF. Tal efeito inibidor da MCP foi descrito em
osteoblastos, células de melanoma, mioblastos cardíacos, células neuronais e epiteliais
(VINCENT; FELDMAN, 2002).
Além das ações que influenciam o ciclo celular em geral, os IGFs também
são capazes de mimetizar a ação hipoglicemiante da insulina quando administrados em
bolus. Esse efeito é conseguido mediante o aumento na captação periférica e redução da
produção hepática de glicose, além do aumento da sensibilidade à insulina (JONES;
CLEMMONS, 1995). A diminuição da proteólise e o aumento da síntese protéica são
outros exemplos de ações insulina-símile do IGF-I (MAURAS; BEAUFRERE, 1995).

21
A regulação da síntese dos IGFs é complexa e envolve diversos fatores,
sendo o GH considerado um dos principais promotores da síntese do IGF-I. Em crianças e
adultos normais as concentrações de IGF-I relacionam-se com a secreção espontânea de
GH (RUDMAN et al., 1981; BLUM et al., 1993). Por outro lado, deve ser lembrado que
baixas concentrações séricas de IGF-I são relacionadas à baixa ingestão protéica em
crianças, o que demonstra a influência exercida por outros fatores, como os nutricionais
(BOUHADDIOUI et al., 1989).
I.2 - Os receptores de IGFs
Os IGFs atuam mediante interação com dois receptores diferentes chamados
de IGF1R e IGF2R.
O IGF1R apresenta grande semelhança estrutural com receptor da insulina
sendo ambos compostos por 2 subunidades α e 2 subunidades β de 135 kDa e 90 kDa,
respectivamente (RECHLER; NISSLEY, 1985). Cada subunidade α é ligada a uma
subunidade β através de uma ponte dissulfídrica e as duas subunidades α são interligadas
através de 2 pontes dissulfídricas formando assim um heterotetrâmero β-α-α-β (LEROITH
et al., 1995).
Um resíduo de metionina inicia a estrutura primária do precursor do IGF1R
e é seguido por uma sequência sinalizadora de 30 aminoácidos rica em treonina e serina
precedendo a extremidade N-terminal do receptor. Após o peptídeo sinalizador, um resíduo
de ácido glutâmico inicia a subunidade α que contém uma região rica em cisteína (24
aminoácidos) e 11 sítios potenciais de glicosilação sendo essa subunidade localizada
totalmente na região extracelular. O sítio de clivagem existente entre as subunidades α e β

22
localiza-se entre os aminoácidos 706 e 711, em uma sequência composta por arginina,
lisina, arginina e arginina. Portanto, a cadeia β inicia-se na posição 711 com uma
asparagina. A subunidade β apresenta, em sua porção extracelular, 5 potenciais sítios de
glicosilação que são seguidos por uma única sequência de aminoácidos hidrofóbicos
composta por 24 aminoácidos localizados entre as posições 906 e 929 que perfaz a porção
transmembrânica do IGF1R. Além disso, essa região é flanqueada, em sua porção C-
terminal, por aminoácidos básicos que facilitam a fixação do receptor à membrana. Os
domínios intra-citoplasmáticos das subunidades β que apresentam atividade tirosina-cinase
são localizados numa região composta por 257 aminoácidos entre as posições 973 e 1229.
Esses sítios são capazes de promover a sinalização interna a partir da ligação, em regiões
ricas em cisteína nas subunidades α, com os IGFs (ULLRICH et al., 1986; VINCENT;
FELDMAN, 2002). A estrutura cristalizada de 3 primeiros domínios da subunidade α foi
descrita em 1998 por Garrett et al. Essa região é responsável pela interação desse receptor
com seus ligantes: IGF-I, IGF-II e insulina. Esse fragmento é compostos por 462
aminoácidos distribuídos em 3 domínios: L1, rico em cisteína e L2. Os domínios L1 e 2 são
formados por β hélice única torcida enquanto que o domínio rico em cisteína contém 8
módulos de pontes dissulfídricas e intercala os outros 2. Esses domínios, nessa sequência,
delimitam um espaço central na molécula capaz de comportar moléculas ligantes
(GARRETT et al., 1998). Sob a forma cristalizada, a distância entre os domínios L-1 e -2,
imposta pelo domínio rico em cisteína, parece ser grande, entretanto, em solução,
provavelmente, uma rotação de até 25º do domínio L-2 pode ser capaz de facilitar o contato
de regiões dos domínios L com os ligantes. Com isso, apesar do domínio rico em cisteína
da subunidade α parecer ser o responsável pela alta afinidade do IGF1R aos IGFs, os outros
domínios também colaboram para essa interação, haja vista que mutações nos domínios L

23
provocam alterações nas afinidades com os IGFs (GARRETT et al., 1998; DUPONT et al.,
2003).
O IGF1R tem alta afinidade pelos IGFs e, principalmente, para o IGF-I. A
afinidade para o IGF-II, embora alta, é menor quando ambas são comparadas. Dada a
homologia existente entre o IGF1R e os receptores de insulina, os IGFs podem interagir
com os receptores de insulina, embora com afinidade menor, e, da mesma forma, à insulina
pode ligar-se ao IGF1R com afinidade também menor que os IGFs (NEELY et al., 1991;
JONES; CLEMMONS, 1995).
Após a ligação dos IGFs às cadeias α do IGF1R, ocorre a internalização do
sinal através dos domínios transmembrânicos e as cadeias β respondem com alterações de
conformação que estimulam a atividade tirosina-cinase dessas cadeias em seus respectivos
domínios onde ocorre a autofosforilação nos resíduos de tirosina (O‟CONNOR et al.,
1997).
O IGF1R atua mediante ativação de múltiplas vias de sinalização, dentre elas a
via da PI3K e da MAPK. Como exemplo pode ser citada a expressão do fator de crescimento
vascular endotelial que é induzida via ativação da MAPK em células que expressam
excessivamente o IGF1R (MIELE et al., 2000).
Dentre diversos mecanismos que são ativados após o processo de
autofosforilação nas cadeias β, destacam-se aqueles relacionados com a MCP. Tal
fenômeno é geneticamente programado e facilita a remoção celular pela fagocitose.
Também foi observado que a expressão do IGF1R é relacionada ao grau de indução da
MCP, sendo que a expressão desse receptor diminui a suscetibilidade a esse processo
(RESNICOFF et al., 1995). A regulação da MCP é exemplo de ação mediada pelo IGF1R
onde parte dos mecanismos conhecidos utilizados após a ligação dos IGFs nesse receptor

24
são conhecidos. Nesse processo, uma vez iniciada a autofosforilação, vários substratos
protéicos associados são fosforilados e diversas vias são ativadas sendo que muitas delas
são envolvidas no impedimento da MCP (VINCENT; FELDMAN, 2002). Após a
autofosforilação do IGF1R, a fosforilação do IRS-1 promove a ativação da PI3K, sendo
essa via melhor conhecida nesse efeito da ativação do IGF1R. Essa ativação é obtida
através da ligação entre o domínio SH2 da PI3K ao IRS-1 levando ao aumento do PIP3. Em
concentrações altas o PIP3 liga-se à PKB e à PDK-1. Após essa ligação, a PDK-1 promove
a fosforilação do Akt/PKB, proteína ribossomal p70 cinase S6, proteína cinase C e PKA,
sendo essas 3 últimas consideradas partes de um mecanismo que também protege a célula
da MCP. A ativação do Akt/PKB também é considerada fundamental no impedimento da
MCP, pois aumenta a atividade de fatores anti-apoptóticos tais como Bcl2 e BclX. Por
outro lado, o Akt/PKB ativado inibe fatores que contribuem na MCP (VINCENT;
FELDMAN, 2002).
A análise da expressão através da técnica de cDNA microarray possibilitou a
identificação dos genes que tem sua expressão estimulada mediante ativação do IGF1R.
Tais genes habitualmente são envolvidos em várias funções celulares incluindo a
proliferação, diferenciação e MCP, porém vale destacar que mais da metade deles são
associados à mitogênese e à diferenciação. Como exemplos, podem ser citados os
receptores 3 e 4 de interleucinas (envolvidos no crescimento celular) e o fator neurotrófico
derivado da linhagem celular da glia (importante no desenvolvimento e manutenção de
neurônios), que têm sua expressão gênica estimulada após a ativação do IGF1R (KEEGAN
et al., 1994; AIRAKSINEN; SAARMA, 2002). Além desses exemplos relacionados ao
desenvolvimento celular, deve ser destacado o aumento da expressão do gene Twist
(DUPONT et al., 2003). Esse gene pertence à família de fatores de transcrição básicos em

25
forma de hélice-alça-hélice, os quais desempenham papel fundamental na diferenciação
celular de vertebrados e invertebrados (OLSON; KLEIN, 1994). O tratamento de células
NWTb3 (linhagem celular desenvolvida a partir de fibroblastos de camundongos e que
apresenta expressão aumentada de IGF1R) com IGF-1 foi capaz de aumentar o mRNA do
gene Twist. Esse aumento ocorre de forma dependente da ativação do IGF1R conforme
verificado em estudos que utilizaram culturas celulares, além disso, o mesmo efeito é
observado em músculo, quando injeções de IGF-1 são realizadas na veia cava. A via de
sinalização que ativa a MAPK parece ser crucial para que o aumento no mRNA do gene
Twist ocorra (DUPONT et al., 2003).
O gene codificador do IGF1R localiza-se no braço longo do cromossomo 15
(15q26.3), estima-se que possua mais que 100 kb e é composto por 21 exons e 20 íntrons. A
subunidade α é codificada pelos exons 1 até o 11, já a β pelos exons 12 ao 21. O primeiro
exon contém uma extensa sequência 5‟ não traduzida, definindo um sítio de início de
transcrição e, dessa forma, codifica somente os 2 primeiros aminoácidos da subunidade α.
Os exons 2 a 6 são responsáveis pelos domínios de ligação aos IGFs presentes nas
subunidades α do receptor. Cinco exons (16-20) são responsáveis pelo domínio tirosina-
cinase presente na subunidade β e o exon 14 pelo domínio transmembrânico. O exon
terminal do gene que codifica o IGF1R também contém uma ampla sequência 3‟ não
traduzida. A homologia organizacional desse gene quando comparado ao gene codificador
do receptor de insulina é notável. Dentre os 21 exons, 12 apresentam tamanho idêntico ao
gene homólogo do receptor de insulina e, no restante de exons, com exceção do primeiro e
do último, a diferença de tamanho não ultrapassa 15 nucleotídeos. Essa similaridade
observada na estrutura gênica é mantida mesmo nas regiões dos receptores onde a
homologia não é grande (ABBOTT et al., 1992; ABUZZAHAB et al., 2003).

26
O IGF2R é estruturalmente diferente do IGF1R e do receptor de insulina,
sendo idêntico ao receptor de manose-6-fosfato e não apresenta atividade tirosina-cinase
presente no IGF1R. O IGF2R apresenta peso molecular de 220-250 kDa e alta afinidade
pelo IGF-II sendo os sítios de ligação do IGF-II e da manose-6-fosfato provavelmente
diferentes (NEELY et al., 1991; JONES; CLEMMONS, 1995). Nota-se que, uma vez
ligados, IGF2R e IGF-II, o complexo formado é internalizado e degradado (NIELSEN,
1992). O papel fisiológico desse receptor ainda não é claro, embora existam indícios de que
participe do processo de clearance do IGF-II.
Vale salientar que, além do IGF1R e IGF2R, existem receptores híbridos
constituídos por dímeros α-β do IGF1R e dímeros α-β do receptor de insulina que
apresentam afinidade pelos IGFs semelhante àquela apresentada pelo IGF1R e menor pela
insulina (SOOS; SIDDLE, 1989; SOOS et al., 1993).
I.3 - As proteínas transportadoras dos IGFs
A regulação das ações promovidas pelos IGFs é influenciada pela presença
das proteínas transportadoras de IGFs ou IGFBPs. Até o momento, foram bem
caracterizadas 6 proteínas de mamíferos dessa família, chamadas de IGFBP-1, -2, -3, -4, -5
e -6. Nos fluidos biológicos, grande parte dos IGFs é encontrada em associação com as
IGFBPs que apresentam grande afinidade pelos mesmos (Kd de 10-10
M). Tal afinidade é
maior que a apresentada pelo próprio IGF1R a esses fatores de crescimento (Kd de 10-8
- 10-
9 M).
Atualmente, esse grupo de proteínas é classificado como uma família. Essa
classificação é baseada na grande homologia existente nas estruturas primárias sendo de até

27
50% entre as 6 proteínas e de 80% quando comparadas diversas espécies de mamíferos
(RECHLER, 1993; KELLEY et al., 1996). Além disso, e principalmente, as IGFBPs são
proteínas ricas em resíduos de cisteína localizados na porção N-terminal e C-terminal e
apresentam afinidade alta pelos IGFs, provavelmente proporcionada pela conformação
terciária desses dois domínios. É importante salientar que é justamente a alta afinidade
pelos IGFs a principal característica que define essas proteínas como sendo ligantes dos
IGFs e componentes de uma família (HWA; YOUNGMAN; ROSENFELD, 1999).
No entanto, na década passada, foram descritas diversas proteínas que
também apresentavam regiões ricas em cisteína e porções N-terminais semelhantes àquelas
presentes nas IGFBPs e que, funcionalmente, também ligavam-se aos IGFs com grande
afinidade, porém menor que as proteínas originalmente ligantes. Além disso, apesar de
apresentar um domínio N-terminal semelhante ao das IGFBPs, o restante da estrutura
dessas proteínas era diferente das IGFBPs. Em função dessas peculiaridades e semelhanças,
essas moléculas foram chamadas de IGFBP-rPs e, no seu conjunto, IGFBPs e IGFBP-rPs
puderam ser agrupadas em uma superfamília de proteínas (BAXTER et al., 1998). Existem
aproximadamente 9 proteínas com características que permitem agrupá-las nessa
classificação, assim denominadas: IGFBP-rP1, -rP2, -rP3, -rP4, -rP5, -rP6, -rP7, -rP8 e -
rP9. Observa-se grande semelhança na porção N-terminal entre esse grupo e as IGFBPs,
variando entre 40% a 57% (HWA; YOUNGMAN; ROSENFELD, 1999). A afinidade da
IGFBP-rP1, -rP2 e –rP3 pelos IGFs foi mensurada, sendo sempre menor que aquelas
verificadas em relação às IGFBPs. Provavelmente, a ausência do domínio C-terminal e,
portanto, também da configuração terciária contribuem para essa afinidade reduzida. Por
outro lado, observou-se que a IGFBP-rP1 e –rP3 são capazes de se ligar à insulina com
afinidade semelhante aos IGFs (AKAOGI et al., 1996; BURREN et al., 1999).

28
As IGFBPs apresentam elevado grau de especificidade e afinidade pelos
IGFs e, além de aumentar a meia vida desses fatores de crescimento, tais proteínas também
podem facilitar ou inibir as suas ações biológicas. Deve ser ressaltado que, além das ações
dependentes dos IGFs, também foram descritas ações dessas proteínas transportadoras que
são independentes dos IGFs (MOHAN; BAYLING, 2002). Habitualmente, são produzidas
diversas IGFBPs num mesmo tecido do organismo. No entanto, a produção de uma, ou no
máximo duas, dessas proteínas é maior que as demais, tornando a concentração de
determinadas IGFBPs maior em alguns tecidos específicos (RECHLER, 1993; KELLEY et
al., 1996; BAXTER, 2000; CLEMMONS, 2001).
Em geral, as estruturas primárias das IGFBPs dos mamíferos têm, em
comum, 3 domínios distintos de tamanhos equivalentes: o conservado domínio N-terminal,
a região mediana (extremamente variável) e o domínio C-terminal também conservado
(HWA; YOUNGMAN; ROSENFELD, 1999).
O domínio N-terminal contém entre 80 e 93 aminoácidos na molécula
madura. Dentre os 16 a 20 resíduos de cisteína encontrados no pré-peptídeo, 10 (na IGFBP-
6) a 12 (no restante das IGFBPs) estão nesse domínio. O grande número de cisteína
encontrado nesse domínio sustenta a hipótese de que essa região seja altamente conservada.
Além disso, existem evidências de que pontes dissulfídricas sejam formadas dentro desse
mesmo domínio, portanto, sem formações dessa natureza com o domínio C-terminal
(FORBES et al., 1998). Essas pontes dissulfidricas internas, presentes no domínio N-
terminal, formam 2 subdomínios: um composto por 6 cisteínas e outro por 4 na IGFBP-6
(NEUMANN; MARINARO; BACH, 1998). Além desses subdomínios, existe no domínio
N-terminal dentre as IGFBPs um motif composto por GCGCCxxC bem conservado entre as
IGFBPs (exceto na IGFBP-6, onde o número de cisteínas é menor). É postulado que esse

29
motif seja importante nas interações entre IGFs e IGFBPs uma vez que também é
encontrado nas IGFBP-rPs, porém a importância atribuída a esse motif pode ser sutil, uma
vez que a conservação dessa região não é completa na IGFBP-6 (HWA; YOUNGMAN;
ROSENFELD, 1999).
A região mediana das IGFBPs humanas separa os domínios N- e C-terminal
estendendo-se do aminoácido 55 ao 95. É uma região extremamente variável e parece não
haver semelhanças entre as diversas IGFBPs, dado que a similaridade entre elas é menor
que 15%.
Após a tradução, as IGFBPs podem sofrer diversas modificações, dentre
elas, destacam-se a glicosilação, a proteólise, a fosforilação e a ligação à matriz
extracelular, que são responsáveis por mudanças na afinidade das IGFBPs pelos IGFs. Com
isso, essas modificações sofridas pelas IGFBPs são capazes de promover uma regulação
diferenciada das ações desses fatores de crescimento. Além desses processos, as IGFBPs
também podem ser alvo de glicosilação (MOHAN; BAYLING, 2002).
Apesar da variabilidade da região mediana, provavelmente, é nessa
localização que ocorrem algumas das modificações pós-traducionais, dentre elas a
glicosilação e a fosforilação. A N-glicosilação ocorre em resíduos de asparagina localizados
em sequências consensuais (Asn-X-Ser/Thr), que são em número de 3, identificadas na
IGFBP-3 (ZAPF et al., 1988). A capacidade de ligação com alta afinidade com os IGFs não
parece ser influenciada pela glicosilação, embora possa conferir algum grau de resistência à
proteólise das IGFBPs (NEUMANN; MARINARO; BACH, 1998).
Foi descrito que, dentre as 6 IGFBPs, ao menos 3 (IGFBP-1, -3 e -5) são
modificadas após sua tradução pela fosforilação (HWA; YOUNGMAN; ROSENFELD,
1999; MOHAN; BAYLING, 2002). Nessas 3 proteínas, esse processo ocorre em resíduos

30
de serina localizados na região mediana. Exemplo disso é verificado na IGFBP-1 onde 2
resíduos de serina da região mediana são fosforilados e somente 1 resíduo da porção C-
terminal também é fosforilado (JONES et al., 1993c). A IGFBP-3 humana pode ser
fosforilada nas serinas que ocupam a posição 111 e 113. O processo de fosforilação dessa
IGFBP parece ser regulado pelos IGFs mediante interação com o IGF1R (COVERLEY;
BAXTER, 1997).
A região C-terminal é altamente conservada sendo a semelhança
compartilhada entre as 6 proteínas humanas de aproximadamente 34%. Nessa região, ao
menos 6 cisteínas são envolvidas na formação de ligações dissulfídricas dentro desse
domínio (FORBES et al., 1998). A estrutura primária da região próxima às últimas 5
cisteínas é semelhante (40%). Dado esse fato, a estrutura terciária dessa região
provavelmente é idêntica. Outro dado interessante observado sobre essa região é a
semelhança de 37% na sequência de aminoácidos presentes entre as cisteínas com outros
domínios de outras proteínas tais como o domínio tipo I da tireoglobulina (MALTHIERY;
LISSITZKY, 1987). A função desse domínio ainda não é conhecida, porém poderia estar
associada à interação com os IGFs, matriz extracelular e superfície celular (HWA; OH;
ROSENFELD, 1999).
É notório que a disposição dessas regiões que compõem as IGFBPs está
relacionada às suas diversas funções dependentes ou independentes dos IGFs. Diferentes
estudos demonstram uma participação importante da região N-terminal na interação com os
IGFs. Substituições de aminoácidos em determinadas posições da região N-terminal
promove reduções importantes da afinidade das IGFBPs pelos IGFs. Com precisão maior,
os 2 subdomínios propostos por Neumann, Marinaro e Bach (1998), localizados nessa
região, apresentam papel de destaque na afinidade com os IGFs. Entretanto, ambas, a

31
região C-terminal e a região mediana, também parecem ter importância nessa interação
IGFs-IGFBPs. A região mediana parece não ligar-se diretamente aos IGFs, porém colabora
com a alta afinidade da IGFBPs, garantindo uma estrutura ternária da molécula capaz de
interagir adequadamente com os IGFs (HWA; OH; ROSENFELD, 1999).
A estrutura gênica codificadora das IGFBPs também apresenta grande
semelhança entre si. Todas essas proteínas são codificadas por uma estrutura gênica comum
composta por 4 exons, dentre as quais, a única exceção é a IGFBP-3 que é codificada por 5
exons sendo que o quinto não é traduzido. O exon 1 corresponde à porção N-terminal em
todas as IGFBPs, o exon 2 corresponde à região mediana e, ambos, 3 e o 4 à C-terminal,
refletindo, dessa forma, a sua estrutura primária.
A IGFBP-1 foi a primeira IGFBP identificada e sequenciada; é constituída
por 234 aminoácidos e tem peso molecular de 30 kDa. Inicialmente isolada no líquido
amniótico, onde sua concentração é aproximadamente 1000 vezes maior que no soro,
apresenta, dentre as ações dependentes de IGFs, a capacidade de inibir essas ações
demonstrada em diversos estudos in vitro e in vivo (POVOA et al., 1984; LEE et al., 1997;
MOHAN; BAYLING, 2002). Entretanto, a transfecção da IGFBP-1 promoveu um estímulo
na migração de células ovarianas de hamster chinês. Vale ressaltar que essas células não
produzem IGFs e não respondem quando a eles são expostas, demonstrando-se, dessa
forma, uma ação da IGFBP-1, que independe dos IGFs (JONES et al., 1993b). Após a
expressão dessa proteína, a afinidade da IGFBP-1 pelos IGFs também pode ser modulada
pela fosforilação nos resíduos de serina localizados nas posições 101, 119 e 169. A forma
fosforilada tem sua afinidade pelo IGF-I aumentada em até 6 vezes (JONES et al., 1991).
As concentrações séricas de IGFBP-1 apresentam variação circadiana
evidente e sua expressão pode ser suprimida após refeições, coincidindo, inversamente,

32
com a secreção de insulina. A produção hepática também é inversamente regulada pela
insulinemia portal, sendo observado que nos pacientes diabéticos as concentrações séricas
de IGFBP-1 estão elevadas (COTERRIL et al., 1988; HOLLY et al., 1990; BRISMAR et
al., 1994). Portanto, a insulina constitui-se, assim, no principal regulador da produção de
IGFBP-1. Outros hormônios, dentre eles GH, cortisol e glucagon, também colaboram nessa
regulação. Em situações experimentais de hipoinsulinemia, houve aumento das
concentrações séricas de IGFBP-1 após a administração de cortisol e glucagon, entretanto o
GH apresentou ação inibidora nessa situação (CONOVER; DIVERTIE; LEE, 1993;
HILDING et al., 1993). Após a ligação com a insulina, ocorre a autofosforilação do
receptor de insulina e a fosforilação de substratos específicos (IRS-1 e -2). As fosfotirosinas
ligam-se a uma subunidade reguladora da PI3K chamada p85. Consequentemente, ocorre a
ativação de outras proteínas, dentre elas a proteína cinase B/Akt. A insulina regula a
expressão do gene da IGFBP-1, inibindo-a, através desse mecanismo que utiliza a PI3K
(BAND; POSNER, 1997). Entretanto, a partir da PI3K duas vias são potencialmente usadas
para que a inibição da expressão da IGFBP-1 ocorra. Uma delas envolve a ativação da Akt
que acaba por afetar a transcrição de fatores, tais como o Foxo1, Foxa2 e Foxo3. Após a
exclusão nuclear desses fatores, há inibição da ligação do Foxo1, –a2 e -o3 ao elemento
responsivo à insulina no promotor do gene IGFBP-1 (MOUNIER; DUMAS; POSNER,
2006). Noutra via, há ativação da mTOR, uma proteína com domínio catalítico semelhante
à PI3K, que é ativada downstream PI3K. A partir da ativação da mTOR há diminuição da
expressão do mRNA da IGFBP-1 independente da modulação do Foxo1 (PATEL et al.,
2002). Entretanto, recentemente, um estudo observou a importância de uma via mTOR-
dependente, que pode ser ativada independentemente da ativação da PI3K, e que pode
regular a expressão da IGFBP-1 na ausência de fosforilação do Foxo1 e -3. A fosforilação

33
do Foxo1 e -o3 não foi necessária nem suficiente para o efeito da insulina na transcrição do
gene da IGFBP-1, entretanto pareceu ter o papel de aumentar e consolidar o efeito
inibitório da insulina (MOUNIER; DUMAS; POSNER, 2006).
O gene humano codificador da IGFBP-1 é localizado no braço curto do
cromossomo 7 (7p14), separado somente por 20 kb região do gene codificador da IGFBP-3.
Estende-se por 5,2 kb e possui 4 exons que geram um mRNA de 1,6 kb. De forma
semelhante ao que ocorre nos genes das outras IGFBPs, o gene codificador da IGFBP-1
tem seu exon 1 composto por menos que 600 pb, correspondente à região N-terminal, já os
exons 2 e 3 são compostos por menos de 230 pb e correspondem a regiões variáveis da
molécula, que ainda tem na sua composição o sítio conservado C-terminal, correspondente
ao exon 4 (HWA; OH; ROSENFELD, 1999).
Aproximadamente 75% dos IGFs circulam sob a forma de um complexo
formado pela IGFBP-3 em associação com a ALS. Esse complexo ternário tem peso
molecular de 150 kDa e, portanto, não atravessa a barreira endotelial o que aumenta a vida
média desses fatores de 10 minutos, quando na forma livre, para 15 horas sob a forma de
complexos (ZAPF, 1995; BAXTER; MARTIN, 1989; BAXTER, 2000). A IGFBP-5
também apresenta a capacidade de formar complexos ternários com os IGFs e ALS, porém
a importância, em comparação com a IGFBP-3, parece ser menor (TWIGG et al., 1998;
BAXTER et al., 2002). A IGFBP-3 também pode ser fosforilada, porém esse processo
parece não alterar a sua afinidade pelos IGFs, diferentemente do que ocorre com a IGFBP-
1, além disso, quando ligada a fibroblastos, tem sua afinidade pelos IGFs diminuída em 40
vezes, ao ser comparada com a afinidade da IGFBP-3 em solução (CONOVER; POWELL,
1991; COVERLEY; BAXTER, 1997). Foram observadas algumas ações da IGFBP-3 que
são independentes dos IGFs em fibroblastos de camundongos e linhagens de células

34
humanas de câncer de mama (VILLAUDY et al., 1991; OH et al., 1993). A ALS é um
membro da família de proteínas “ricas em repetição” de leucina, sendo composta por quase
25% desse aminoácido; tem essa denominação devido ao fato de que se torna
irreversivelmente desnaturada quando submetida à acidificação do meio até um pH de 4 ou
menor. Assim como a IGFBP-3, a secreção da ALS é regulada principalmente pelo GH
(BAXTER; DAI, 1994). A presença dessa subunidade é fundamental no aumento
observado na meia-vida proporcionado pelo complexo ternário. Tanto em modelos animais,
quanto em humanos, as concentrações séricas totais de IGF-1 são reduzidas na deficiência
da ALS (BOISCLAIR et al., 2001; HWA et al., 2006).
A segunda proteína ligante de IGFs mais abundante no plasma é a IGFBP-2
sendo composta por 289 aminoácidos e possui um peso molecular de 31,3 kDa
(DAUGHADAY, 1995). Na vida intra-uterina, é a IGFBP que predomina na circulação e
tem no fígado a sua principal fonte de expressão. É encontrada em altas concentrações no
líquor e sêmen (ROSENFELD et al., 1994). O gene que codifica a IGFBP-2 localiza-se no
braço longo do cromossomo 2. Possui 4 exons e sua extensão é de 32 kb (HWA;
YOUNGMAN; ROSENFELD, 1999). Essa proteína liga-se com maior afinidade ao IGF-II
que ao IGF-I e é capaz de carrear os IGFs, através da transposição da barreira endotelial,
para os diversos tecidos. Concentrações elevadas dessa proteína são observadas em
pacientes com diabetes, hipopituitarismo, nanismo de Laron e em vários tipos de tumores,
embora os mecanismos envolvidos na sua regulação ainda não sejam completamente
conhecidos (CLEMMONS et al., 1991; CAMACHO-HÜBNER et al., 1991; COTTERILL
et al., 1992; JUUL, 2003). Em uma deficiência genética e isolada do GH, por uma mutação
no receptor do GHRH, não houve diferença entre as concentrações de IGFBP-2 e IGFBP-1
entre os indivíduos afetados e os controles em todas as faixas etárias (AGUIAR-

35
OLIVEIRA et al., 1999). Entretanto, situações de restrição protéica provocam aumento nas
concentrações séricas de IGFBP-2 em adultos e crianças (SMITH; UNDERWOOD;
CLEMMONS, 1995).
A IGFBP-4 apresenta uma massa molecular de 26 kDa sendo composta por
237 aminoácidos. O gene codificador dessa proteína localiza-se no braço longo do
cromossomo 17, mede aproximadamente 15,3 kb e sua estrutura também é formada por 4
exons (HWA; YOUNGMAN; ROSENFELD, 1999). Essa proteína apresenta alta afinidade
pelo IGF-I e IGF-II e também atravessa o endotélio juntamente com os fatores de
crescimento (COHICK; CLEMMONS, 1993). IGF-I e IGF-II promovem aumento nas
concentrações da IGFBP-4 em células de carcinoma mamário e fibroblastos fetais em
cultura, porém sem aumento concomitante da expressão do mRNA correspondente. Tal fato
indica um provável mecanismo regulatório pós-tradução (CAMACHO-HÜBNER et al.,
1992). Dentre esses mecanismos, destaca-se a proteólise de que essa IGFBP possa ser
substrato. Provavelmente, a protease envolvida nessa reação é a chamada PAPP-A que
promove a proteólise da IGFBP-4 com relativa especificidade. A PAPP-A é produzida por
uma grande variedade de células dentre as quais podem ser citados os fibroblastos, as
células da granulosa e os osteoblastos. Além disso, é presente no soro em indivíduos
normais e em mulheres grávidas (LAWRENCE et al., 1999; BYUN et al., 2001;
CONOVER et al., 2001; MOHAN; BAYLINK, 2002). Os IGFs parecem exercer papel
importante nessa proteólise uma vez que, quando ligados à IGFBP-4, expõem o sítio de
clivagem dessa proteína ligante à PAPP-A (QIN et al., 2000). Por outro lado, a proMBP
liga-se à PAPP-A e bloqueia a proteólise da IGFBP-4 (GIUDICE et al., 2002).
Outros hormônios parecem estar ligados ao controle da expressão da IGFBP-
4. O tratamento de células de osteossarcoma TE89 com GH produziu aumento e diminuição

36
das concentrações da IGFBP-4 em culturas de alta e baixa densidade celulares
respectivamente (MOHAN et al., 1992). O paratormônio produziu aumento da expressão
dessa proteína em culturas de osteoblastos (LATOUR et al., 1990). Outros hormônios
sistêmicos podem ser citados como reguladores da IGFBP-4 em osteoblastos tais como a
1,25 diidroxi-vitamina D3 e os glicocorticóides. Além dos IGFs, outros fatores de
crescimento locais também colaboram nessa regulação dentre eles destacando-se as
proteínas morfogenéticas ósseas, TGF- β e interleucinas (MOHAN; BAYLINK, 1999). A
IGFBP-4 apresentou efeitos inibitórios sobre a bioatividade dos IGFs em diversos estudos,
além disso, os fragmentos resultantes da proteólise dessa IGFBP também parecem não
interferir na ação dos IGFs (JONES; CLEMMONS, 1995).
A IGFBP-5 é composta por 252 aminoácidos apresentando peso molecular
de 31 kDa (DAUGHDAY, 1995). O gene codificador dessa proteína, com tamanho de 33
kb, localiza-se no braço longo do cromossomo 2 apresentando 4 exons o que gera um
mRNA de 6 kb (HWA; YOUNGMAN; ROSENFELD, 1999). É a IGFBP mais abundante
no tecido ósseo (CLAYTON; GILL, 2001). Vale lembrar que, da mesma forma que a
IGFBP-3, a IGFBP-5 também pode formar complexos ternários com os IGFs e a ALS,
porém, quando comparado com a IGFBP-3, a importância desses complexos parece ser
menor (TWIGG et al., 1998; BAXTER et al., 2002). Observou-se que, in vitro, o IGF-I é
capaz de aumentar a concentração de IGFBP-5, porém sem aumento do seu mRNA. Essa
observação, em conjunto com o fato de que a insulina foi incapaz, quando estimulou o
IGF1R, de aumentar as concentrações da IGFBP-5, ou seja, de forma independente do
receptor, levou a elaboração da hipótese de que os IGFs poderiam proteger a IGFBP-5 da
ação de proteases específicas (CAMACHO-HÜBNER et al., 1992; CONOVER; KIEFER,
1993). Vale lembrar que, mais recentemente, algumas dessas proteases foram identificadas.

37
Dentre elas, o complemento C1s produzido por células de músculo liso (BUSBY et al.,
2000) e a ADAM-9 (MOHAN et al., 2002), produzida por osteoblastos humanos, são
relativamente específicas para essa IGFBP. Os glicocorticóides produziram redução do
mRNA da IGFBP-5 em células semelhantes aos osteoblastos em cultura (CONOVER;
KIEFER, 1993). De forma semelhante ao que ocorre com a IGFBP-4, a IGFBP-5 parece ser
regulada também por outros hormônios, tais como, o GH, o paratormônio e a 1,25-diidroxi-
vitamina D3, além de fatores locais, como as proteínas morfogenéticas ósseas, TGF- β e
interleucinas (MOHAN; BAYLINK, 1999).
O estímulo às ações dos IGFs na presença da IGFBP-5 é verificado em
diversos estudos. Essa proteína, quando ligada à matriz extracelular em cultura de
fibroblasto, potencializa a ação do IGF-I aumentando a síntese de DNA e o crescimento
celular mediante a redução da afinidade da IGFBP-5 pelo IGF-I (JONES et al., 1993a).
Além disso, fragmentos dessa IGFBP parecem ser capazes de potencializar as ações dos
IGFs em osteoblastos (ANDRESS et al., 1993).
A IGFBP-6 é formada por 216 aminoácidos, apresentando peso molecular de
22,8 kDa na sua forma não glicosilada. O gene que codifica essa proteína está localizado no
cromossomo 12 e, também, é composto por 4 exons, gerando um mRNA de 1,1 kb (HWA;
YOUNGMAN; ROSENFELD, 1999). É a menor dentre as IGFBPs sendo, ao lado da
IGFBP-2, a IGFBP mais abundante no líquor (ROGHANI et al., 1991).
As IGFBPs, portanto, são importantes colaboradoras no sistema IGF, pois,
além de aumentar a vida média dos IGFs, modulam suas ações endócrinas e parácrinas
através do controle de suas concentrações nos tecidos e células e da interação com os seus
receptores, estão sujeitas a outros diversos mecanismos regulatórios importantes para o
crescimento e desenvolvimento celulares (JONES; CLEMMONS, 1995).

38
I.4 - O sistema IGF e sua relação com situações de hipóxia
Diversos estudos analisaram o sistema IGF perante situações de hipóxia,
utilizando como modelo experimental o crescimento intra-uterino, que depende
fundamentalmente da função placentária adequada e da oferta de substratos e oxigênio ao
feto e é regulado por complexos mecanismos, incluindo fatores genéticos e ambientais.
Devido à potente ação mitogênica e à colaboração na diferenciação celular,
IGF-I e II são considerados importantes reguladores do crescimento fetal (McLELLAN et
al., 1992). Camundongos que não apresentaram genes para IGF-I e II nasceram pequenos e
muitos morreram precocemente (POWELL-BRAXTON et al., 1993; LIU et al., 1998). Em
seres humanos, as concentrações séricas de IGF-I, pesquisadas em amostras obtidas de
cordocentese, apresentam-se crescentes em fetos normais entre 18 e 40 semanas de
gestação (LANGFORD; NICOLAIDES; MIELL, 1998). Ademais, RNs considerados
pequenos para a idade gestacional tiveram concentrações séricas menores de IGF-I no
sangue de cordão umbilical (GIUDICE et al., 1995). Há somente a descrição de um
paciente com deleção do gene codificador do IGF-I que, além do prejuízo neurológico,
apresentava grave restrição do crescimento pré-natal (WOODS et al., 1996).
A causa mais comum de RCIU é a insuficiência placentária caracterizada
pela oferta inadequada de nutrientes e oxigênio para a placenta e, portanto, para o feto
(POPOVICI et al., 2001). Sob esse aspecto, a hipóxia, como ferramenta de experimentação
in vivo, têm sido amplamente utilizada na compreensão do papel dos IGFs na RCIU. Nesse
sentido, fetos ovinos submetidos à hipóxia por redução do fluxo sanguíneo uterino
apresentaram aumento nas concentrações séricas de IGFBP-1 e redução da IGFBP-2, sendo

39
que, nessa análise, não houve variações significativas das concentrações séricas de IGFBP-
3 e IGFBP-4 (McLELLAN et al., 1992). Da mesma forma, filhotes de ratas submetidas à
hipóxia apresentaram, além da RCIU, concentrações séricas de IGFBP-1 e IGFBP-2 1,75
vezes maiores que os controles e concentrações séricas de IGFBP-4 2,5 vezes também
maiores. As concentrações séricas de IGF-II tenderam a ser maiores alcançando exatamente
o nível de significância (p = 0,05), entretanto as concentrações séricas de IGF-I foram
semelhantes àquelas observadas nos controles. A expressão do mRNA de IGFBP-1 e
IGFBP-2 nos fígados fetais submetidos à hipóxia foram 4 e 6 vezes maiores que os
controles, respectivamente (TAPANAINEN et al., 1994).
Em hepatócitos fetais humanos, obtidos de fetos entre 18 e 22 semanas de
gestação, e, posteriormente, postos em cultura, a hipóxia também provocou aumento de 3
vezes no mRNA da IGFBP-1. As concentrações dessa proteína no meio de cultura
aumentaram 3 vezes quando comparadas com situação de normoxia. Ainda nesse estudo, as
concentrações de IGFBP-3 aumentaram significativamente, IGFBP-2 e IGFBP-4 também
aumentaram, porém não significativamente. A produção da IGFBP-1 nesses hepatócitos
fetais submetidos à hipóxia, reforçou a hipótese de que o fígado poderia ser a fonte primária
da produção dessa proteína em fetos com RCIU (POPOVICI et al., 2001).
Corroborando a relação entre o crescimento intra-uterino e a IGFBP-1,
recentemente, a expressão de pequenas quantidades de IGFBP-1 foi verificada em tecidos
de peixe Danio rerio (popularmente conhecido como paulistinha) em diversos estágios do
desenvolvimento embrionário. A hipóxia, em embriões desse peixe, também provocou
aumento na transcrição hepática da IGFBP-1 e, em adultos, houve aumento no seu mRNA
(MAURES; DUAN, 2002).

40
Alguns estudos procuraram relacionar a presença de fetos com RCIU e as
concentrações de IGF-I e IGFBP-1 no líquido amniótico e no plasma materno, no entanto,
não se evidenciou correlação positiva com as concentrações desses fatores (BAHTIA et al.,
2002). Recentemente, através de amniocentese, as concentrações de IGF-I, IGFBP-1 e
leptina foram avaliadas com a mesma finalidade, no entanto, a correlação com RCIU não
foi observada (ROIG et al., 2005). Percebe-se, portanto, que a hipóxia é capaz de estimular
um conjunto de alterações metabólicas que visam proteger o indivíduo através da inibição
de processos que requeiram grandes aportes de energia.
A análise da expressão através da técnica de cDNA microarray também
possibilitou a identificação dos genes que tem sua expressão induzida mediante ao estímulo
do IGF-I. Esses genes responsivos ao IGF-I podem ser divididos em diversas categorias:
relacionados ao ciclo celular, fatores de transcrição, relacionados ao metabolismo, etc.
Porém vale ressaltar que, além de um grande número deles estar relacionado à angiogênese,
vários deles também foram responsivos ao HIF-1α. De fato, o próprio IGF-I também foi
capaz de induzir a translocação nuclear do HIF-1α (DUPONT et al., 2003).
A importância da integridade funcional do IGF1R e do IGF2R no
desenvolvimento humano é demonstrada quando são descritos indivíduos que apresentam
alterações no seu funcionamento. Várias mutações no gene que codifica o IGF1R foram
observadas em diversos sítios do receptor podendo resultar, inclusive, em alterações do
desenvolvimento dos indivíduos portadores dessas alterações. No momento, diversos
estudos foram realizados sendo observadas relações importantes entre o desenvolvimento
intra-uterino humano e o IGF1R. Exemplo dessa relação é encontrado na mutação em
heterozigose que provocou a substituição de uma arginina por um ácido glutâmico no
códon 709. Descrita em dois indivíduos (mãe e filha) com RCIU sem causa aparente, tal

41
mutação, localizada no exon 11, provocou a perda do sítio de clivagem do pró-receptor tipo
1 de IGF impedindo a formação das cadeias α e β resultando em retardo do crescimento pré
e pós-natal e retardo mental em um desses indivíduos (filha) (KAWASHIMA et al., 2005).
Aproximadamente 10% das crianças nascidas com RCIU permanecem com baixa estatura.
Nesse aspecto, alguns estudos procuraram demonstrar relação entre essa
característica clínica e alterações do IGF1R. Em 2003, 2 grupos de crianças com baixa
estatura foram avaliados, sendo que em um deles havia RCIU sem causa aparente (42
crianças) e o outro era composto por crianças somente com baixa estatura. No primeiro
grupo (RCIU associada à baixa estatura), havia um indivíduo com mutação composta em
heterozigose que alterava a sequência de aminoácidos de arginina para glutamina na
posição 108 num alelo e lisina para asparagina na posição 115 no outro alelo, ambas no
exon 2 do gene IGF1R. Nesse caso, além dos problemas com o crescimento pré e pós-natal,
houve discreto atraso da aquisição motora sendo que as concentrações de IGF-I sérico e
ALS estavam acentuadamente acima do normal e a IGFBP-3 no limite superior da
normalidade de acordo com os parâmetros do método aplicado pelos autores. Entretanto, a
paciente não apresentou redução do seu quociente de inteligência e apresentou puberdade
com início e progressão normais. A ligação entre o IGF1R e o IGF-1 esteve reduzida e
houve necessidade de concentrações maiores de IGF-I para que a atividade de fosforilação
no receptor fosse igual aos controles em culturas de fibroblastos. Nota-se, portanto, que
esse indivíduo apresentou uma mutação missense em heterozigose composta numa região
conservada da molécula do IGF1R associada a características de resistência ao IGF-1. A
mudança nas cargas dos aminoácidos envolvidos (de básica para neutra) provavelmente
alterou o sítio de ligação do receptor proporcionando a redução na ligação com o IGF-1
(ABUZZAHAB et al., 2003).

42
Além desse exemplo, também pode ser citada a mutação em heterozigose no
exon 2 (cadeia α) resultando na substituição de uma arginina por um códon de terminação
no resíduo 59 em outros 2 indivíduos (irmãos) com RCIU e discreto atraso do
desenvolvimento neuropsicomotor associados à microcefalia. Essa mutação provocou
resistência à ação do IGF-1 sendo necessária dose 10 vezes maior desse fator para se obter
fosforilação semelhante aos controles (RAILE et al., 2006).
Apesar das diversas relações observadas entre o IGF1R e RCIU associada à
baixa estatura, existem poucas análises do comportamento da expressão desse receptor nos
diversos tecidos em situações de hipóxia, tanto em fetos quanto em indivíduos adultos. Um
estudo, no qual ovelhas foram submetidas à hipóxia aguda, induzida pela substituição de
oxigênio por nitrogênio após intubação durante a gestação, verificou que fetos ovinos
apresentaram aumento na presença e na expressão do IGF1R nos cones de crescimento
neuronais do cérebro e cerebelo (MORGAN; CHAO, 2004).
Por outro lado, em indivíduos fora do período neonatal, as concentrações
reduzidas de IGF-I sérico foram observadas em crianças em bom estado nutricional com
doenças cardíacas congênitas cianosantes (DÜNDAR; AKÇORAL; SAYLAM et al., 2000).
As concentrações séricas de IGF-I também foram menores em crianças que desenvolveram
bronquiolite por vírus sincicial respiratório quando comparadas com as concentrações
nessas mesmas crianças após 3 meses do evento respiratório agudo (TASKER et al., 2004).
Concentrações séricas aumentadas de IGFBP-1 foram observadas em crianças nascidas
com acidose mista (componente respiratório e metabólico presentes simultaneamente)
(TAZUKE et al., 1998).
Entretanto, existem poucas informações sobre a relação entre a oxigenação
sanguínea e o sistema IGF, particularmente a respeito da IGFBP-1, IGF1R e seus

43
reguladores, nos seres humanos. Portanto, novas abordagens são necessárias para melhor
compreensão do comportamento desses fatores nessas condições, a fim de se obter melhor
entendimento desses mecanismos adaptativos.

44
II – Objetivos

45
II.1 – Objetivos
O objetivo geral desse estudo foi avaliar os diferentes componentes do
sistema IGF e sua relação com a oxigenação sanguínea associada a doenças respiratórias
agudas em crianças.
II.1.1 - Objetivos específicos:
Comparar as concentrações séricas de IGF-I, IGFBP-1, IGFBP-3 e insulina
em crianças nas situações de hipóxia aguda, associada a infecções respiratórias, e após a
sua resolução;
Comparar a expressão do mRNA do IGF1R na situação de hipóxia aguda
associada a doenças respiratórias agudas em crianças.

46
III - Casuística e Métodos

47
III.1 – Crianças participantes
Foram selecionadas 27 crianças dentre as quais 14 eram do sexo masculino e
13 do sexo feminino, com idades que variaram entre 15 dias e 9 anos e 6 meses. O paciente
com maior idade era do sexo masculino e, no momento da sua participação do estudo, não
apresentava sinais de puberdade. O tempo de internação hospitalar foi em média de 8 (±3,3)
e mediana de 7 dias. Os dados gerais e antropométricos de todos os indivíduos são
apresentados na tabela 1.
Todos os indivíduos apresentaram desconforto respiratório devido a diversas
situações clínicas (7 pacientes com sibilância, 4 com sibilância associada à pneumonia, 6
com bronquiolite, 2 com bronquiolite associada à pneumonia, 2 com pneumonia, 6 com
pneumonia associada a derrame e/ou espessamento pleural). Vale ressaltar que os
diagnósticos foram baseados em dados clínicos dos indivíduos sendo as etiologias diversas.
Durante as suas evoluções, todas essas crianças apresentaram saturações de oxigênio,
verificada em oximetria de pulso, menores que 95% e necessitaram de oferta suplementar
de oxigênio de qualquer natureza (cateter nasal, capela ou máscara não-reinalante).
Nenhuma criança necessitou de cuidados intensivos ou de respiração auxiliada por
aparelhos e cada uma participou somente uma vez do estudo.
Vinte crianças apresentaram história de febre que antecedeu a admissão
hospitalar e somente 7 estiveram sem febre no momento da sua inclusão no estudo; 13
pacientes não utilizaram antibióticos durante a sua evolução sendo que os 14 restantes
usaram esquemas terapêuticos diversos (3 utilizaram penicilina cristalina endovenosa
seguida de amoxicilina oral e 11 utilizaram ampicilina endovenosa seguida de amoxicilina).

48
Vale ressaltar que nenhuma das 7 crianças afebris utilizaram antibióticos durante sua
evolução clínica. Na tabela 2 são mostrados as crianças de acordo com o diagnóstico
clínico, oximetria de pulso na admissão hospitalar e o tempo de terapia com oxigênio que
precedeu a primeira coleta de amostras.
O projeto de pesquisa foi aprovado pelo Comitê de Ética em Pesquisa do
Hospital das Clínicas da Faculdade de Medicina de Ribeirão Preto – USP (HCFMRPUSP)
em 22 de maio de 2006 de acordo com o Processo HCRP no. 5231/2006 (Anexo A).
O contato com os responsáveis pelas crianças foi realizado no momento do
atendimento na Unidade de Emergência do HCFMRPUSP. Após apresentação da pesquisa,
esclarecimento de possíveis dúvidas e concordância da participação das crianças no estudo,
o termo de consentimento livre e esclarecido foi assinado por um responsável.
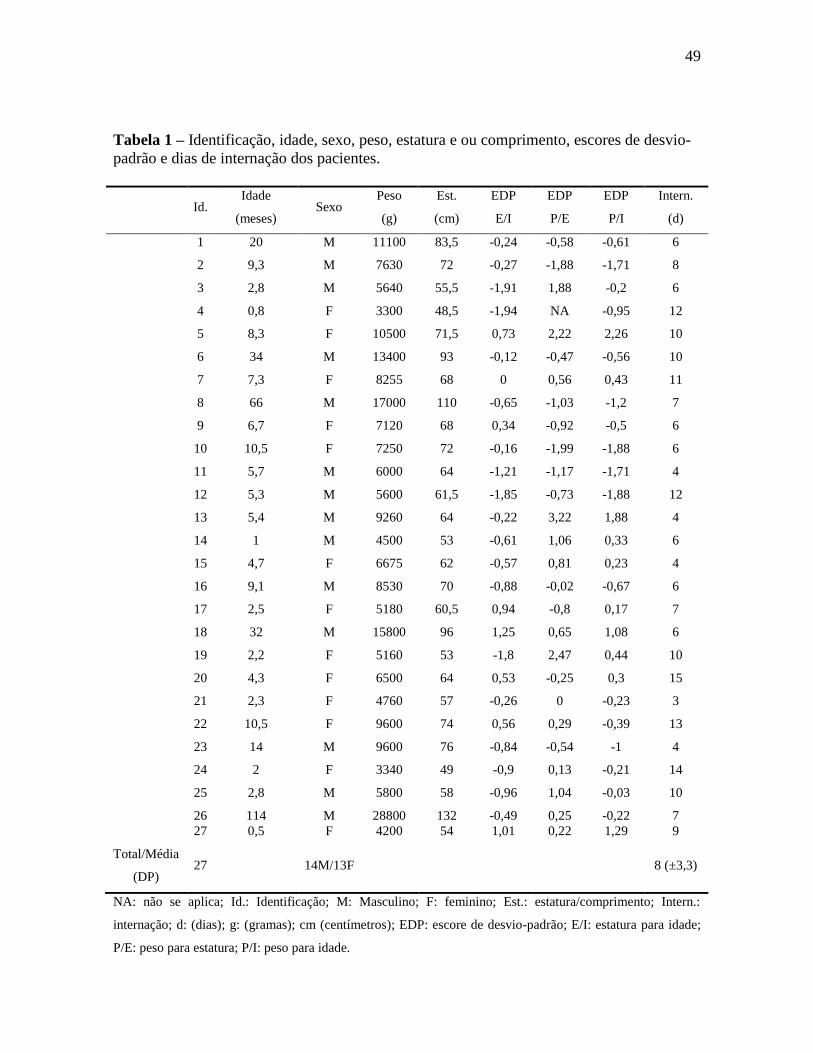
49
Tabela 1 – Identificação, idade, sexo, peso, estatura e ou comprimento, escores de desvio-
padrão e dias de internação dos pacientes.
Id. Idade
(meses) Sexo
Peso
(g)
Est.
(cm)
EDP
E/I
EDP
P/E
EDP
P/I
Intern.
(d)
1 20 M 11100 83,5 -0,24 -0,58 -0,61 6
2 9,3 M 7630 72 -0,27 -1,88 -1,71 8
3 2,8 M 5640 55,5 -1,91 1,88 -0,2 6
4 0,8 F 3300 48,5 -1,94 NA -0,95 12
5 8,3 F 10500 71,5 0,73 2,22 2,26 10
6 34 M 13400 93 -0,12 -0,47 -0,56 10
7 7,3 F 8255 68 0 0,56 0,43 11
8 66 M 17000 110 -0,65 -1,03 -1,2 7
9 6,7 F 7120 68 0,34 -0,92 -0,5 6
10 10,5 F 7250 72 -0,16 -1,99 -1,88 6
11 5,7 M 6000 64 -1,21 -1,17 -1,71 4
12 5,3 M 5600 61,5 -1,85 -0,73 -1,88 12
13 5,4 M 9260 64 -0,22 3,22 1,88 4
14 1 M 4500 53 -0,61 1,06 0,33 6
15 4,7 F 6675 62 -0,57 0,81 0,23 4
16 9,1 M 8530 70 -0,88 -0,02 -0,67 6
17 2,5 F 5180 60,5 0,94 -0,8 0,17 7
18 32 M 15800 96 1,25 0,65 1,08 6
19 2,2 F 5160 53 -1,8 2,47 0,44 10
20 4,3 F 6500 64 0,53 -0,25 0,3 15
21 2,3 F 4760 57 -0,26 0 -0,23 3
22 10,5 F 9600 74 0,56 0,29 -0,39 13
23 14 M 9600 76 -0,84 -0,54 -1 4
24 2 F 3340 49 -0,9 0,13 -0,21 14
25 2,8 M 5800 58 -0,96 1,04 -0,03 10
26 114 M 28800 132 -0,49 0,25 -0,22 7
27 0,5 F 4200 54 1,01 0,22 1,29 9
Total/Média
(DP) 27 14M/13F 8 (±3,3)
NA: não se aplica; Id.: Identificação; M: Masculino; F: feminino; Est.: estatura/comprimento; Intern.:
internação; d: (dias); g: (gramas); cm (centímetros); EDP: escore de desvio-padrão; E/I: estatura para idade;
P/E: peso para estatura; P/I: peso para idade.

50
Tabela 2 – Pacientes de acordo com o diagnóstico clínico, oximetria de pulso na admissão
hospitalar e tempo de terapia com oxigênio antes da primeira coleta de amostras.
Pacientes Diagnóstico
clínico
Oximetria de pulso na
admissão hospitalar (%)
Tempo de terapia com
oxigênio antes da primeira
coleta (min)
1 pn+dp 86 30
2 sbl+pn 90 90
3 sbl+pn 87 20
4 brl+pn 88 Não usou
5 pn+dp 81 17
6 pn 84 50
7 pn+dp 87 30
8 sbl 88 Não usou
9 sbl+pn 90 Não usou
10 sbl+pn 90 20
11 sbl 89 Não usou
12 brl 91 Não usou
13 sbl 89 60
14 brl 80 38
15 brl 92 Não usou
16 pn+dp 95 Não usou
17 pn 89 30
18 sbl 87 90
19 sbl 90 Não usou
20 brl+pn 89 20
21 sbl 89 Não usou
22 pn+dp 87 3
23 sbl 91 Não usou
24 brl 84 5
25 brl 90 60
26 pn+dp 80 15*
27 brl 88 30 pn+dp: pneumonia com derrame pleural; sbl+pn: sibilância com pneumonia; pn: pneumonia; sbl: sibilância;
brl+pn: bronquiolite com pneumonia; brl: bronquiolite; *tempo estimado.
III.2 - Critérios de Exclusão
Foram excluídas do estudo as crianças consideradas desnutridas, para tanto
foram calculados escores Z (desvio-padrão) das relações estatura por idade (E/I), peso por
estatura (P/E) e peso por idade (P/I) utilizando-se como padrão de referência os dados do

51
National Center for Health Statistics (CDC, 2000), sendo considerados indivíduos com
stunting aqueles que apresentaram o indicador E/I abaixo de -2 escores z dos valores de
referência; wasting quando o indicador P/E foi menor que -2 escores z; e, underweight
aqueles com o indicador P/I menor que -2 escores z (WORLD HEALTH
ORGANIZATION, 1995). Os dados de todas as crianças que participaram do estudo são
apresentados na tabela 1.
Também foram excluídas as crianças que em qualquer momento da
avaliação utilizaram corticóides ou que apresentavam síndromes genéticas, malformações
de sistema nervoso, infecções congênitas e endocrinopatias.
III.3 - Desenho Experimental
A coleta do material de todos os pacientes foi realizada no momento da sua
admissão no Pronto Atendimento de Pediatria em conjunto com a coleta de exames
pertinentes à investigação clínica e, preferencialmente, antes do início da oxigenioterapia.
Foram incluídos somente indivíduos que apresentaram saturações de oxigênio, verificadas
mediante oximetria de pulso, menores que 95% (momento de hipóxia). Amostras com
volume total de 2 ml a 5 ml de sangue foram obtidas em frascos com gel de separação para
a avaliação das concentrações de IGF-I, IGFBP-1, IGFBP-3, insulina; e, em frascos
contendo EDTA, foi colhido sangue total para a obtenção de linfócitos dos quais foi
extraído o RNA para a verificação da expressão do IGF1R. As amostras foram
armazenadas a 4oC por curto período de tempo e, assim que possível, foram
definitivamente armazenadas a -70oC. Da mesma forma, no dia da alta hospitalar, ou seja,

52
quando o paciente não mais precisava de oferta suplementar de oxigênio novas amostras
foram coletadas, obtendo, assim, o pareamento das amostras em 2 momentos: na situação
de hipóxia e fora dela, na mesma criança. A coleta das amostras fora da hipóxia foi
realizada no dia da alta hospitalar em todos os indivíduos. Nos casos em que a coleta de
gasometria foi desnecessária, a oximetria de pulso foi o critério adotado como parâmetro de
oxigenação sanguínea tanto durante a hipóxia quanto fora dela.
Na primeira coleta, ou seja, durante o momento de hipóxia, 10 crianças
estavam sem oxigênio e, dentre as 17 que estavam usando oxigênio naquele momento, o
período de uso foi em média de 37,1 (±26,4) minutos com mediana de 30 minutos.
O período de jejum médio antes da primeira coleta do material foi de 3,3
(±2,2) horas com mediana de 2,75. Na segunda coleta, o jejum médio foi de 3,2 (±0,9)
horas com mediana de 3. A oximetria de pulso foi realizada em todos os indivíduos nos
momentos de hipoxemia e fora dela (quando a oferta suplementar de oxigênio não foi
necessária). Essa medida foi realizada com oxímetro de pulso modelo DX 2405 – Oxímetro
de Pulso – Oxyplenthy Super Bright (Dixtal – Brasil). No momento da hipoxemia, em
média a oximetria de pulso foi de 87,8 (±3,5) % e mediana de 88 (86,5 – 90); enquanto que
no momento de ausência de hipoxemia essa medida foi em média de 96,3 (±1,3) % e
mediana de 96 (95,5 – 97).

53
III.4 - Dosagem Laboratorial
III.4.1. - Ensaio imunoenzimático do IGF-I
As concentrações séricas de IGF-I foram determinadas no Laboratório de
Pediatria do HCFMRP-USP utilizando o kit DSL-10-5600 Active® IGF-I ELISA da
Diagnostic Systems Laboratories (DSL – Webster, Texas, EUA). Esse kit é composto por
um imunoensaio enzimático tratando-se, portanto, de um ELISA (enzyme linked immuno
sorbent assay) de “um passo” do tipo sanduíche.
Nesse ensaio, houve uma etapa na qual o IGF-I foi separado de suas
proteínas de ligação no soro. Essa extração, que ocorreu previamente ao ensaio
propriamente dito, foi essencial para a precisa determinação das concentrações de IGF-I.
No ensaio, padrões, amostras e controles em duplicata foram encubados com
anticorpo anti-IGF-I marcado com enzima HRP em cavidades de microtitulação revestidas
com outro anticorpo anti-IGF-I durante 2 horas em temperatura ambiente (~25oC) agitados
a 500-700 rpm em agitadora orbital The Jitterbug™ model 130000 (Boekel Scientific,
Pennsylvania, EUA). Após a encubação, as cavidades foram lavadas 5 vezes em lavadora
automática Immunowash Model 1575 (Bio-Rad Laboratories, EUA) utilizando-se o tampão
de lavagem que acompanhou o kit para a dosagem do IGF-I. Após a lavagem, as cavidades
foram encubadas com o substrato TMB por 10 minutos. Uma solução ácida de interrupção
foi adicionada e o grau de reposição enzimática foi determinado por medida de absorbância
em duplo comprimento de onda a 450 e 620 nm na leitora iMarkTM
Microplate Absorbance

54
Reader (Bio-Rad Laboratories, EUA). Após a leitura, os dados obtidos foram
encaminhados para análise no software Microplate Manager® (Bio-Rad Laboratories,
EUA) conectado à leitora.
Nesse tipo de ensaio, a absorbância medida é diretamente proporcional à
concentração de IGF-I presente na amostra. Um conjunto de padrões de IGF-I foi utilizado
para plotar a absorbância da curva padrão versus a concentração de IGF-I, e, a partir dessa
curva, as concentrações nas amostras desconhecidas foram calculadas. A curva padrão foi
construída a partir das seguintes concentrações: 11, 43, 147, 273 e 600 ng/ml.
Todas as concentrações séricas de um mesmo indivíduo com e sem hipóxia
foram obtidas no mesmo ensaio em duplicatas e os resultados expressos em ng/ml. Os
coeficientes de variação intra- e inter-ensaios foram, respectivamente, 6,05% e 10,8%. A
sensibilidade do ensaio foi de 0,03 ng/ml.
III.4.2 - Ensaio imunoenzimático da IGFBP-1
As concentrações séricas de IGFBP-1 foram determinadas no Laboratório de
Pediatria do HCFMRP-USP utilizando o kit DSL-10-78001 Active® Total IGFBP-1
ELISA da Diagnostic Systems Laboratories (DSL – Webster, Texas, EUA). Esse kit é
composto por um imunoensaio enzimático tratando-se, portanto, de um ELISA (enzyme
linked immuno sorbent assay) de “dois passos” do tipo sanduíche.
No ensaio, padrões, amostras e controles em duplicata foram encubados em
cavidades de microtitulação os quais foram revestidos com anticorpos anti-IGFBP-1
durante 1 hora em temperatura ambiente (~25°C) agitados a 500-700 rpm em agitadora

55
orbital The Jitterbug™ model 130000 (Boekel Scientific, Pennsylvania, EUA). Após essa
encubação, as cavidades foram lavadas 5 vezes em lavadora automática Immunowash
Model 1575 (Bio-Rad Laboratories, EUA) utilizando-se o tampão de lavagem que
acompanhou o kit para a dosagem do IGFBP-1. Houve, então, nova encubação onde as
cavidades foram tratadas com outro anticorpo anti-IGFBP-1 marcado com enzima HRP
durante 30 minutos em temperatura ambiente (~25°C) agitados a 500-700 rpm em agitadora
orbital. Após essa segunda encubação e lavagem idêntica ao primeiro procedimento, as
cavidades foram encubadas com o substrato TMB por 10 minutos. Uma solução ácida foi
adicionada e o grau de reposição enzimática foi determinado por medida de absorbância em
duplo comprimento de onda a 450 e 620 nm na leitora iMarkTM
Microplate Absorbance
Reader (Bio-Rad Laboratories, EUA). Após a leitura, os dados obtidos foram
encaminhados para análise no software Microplate Manager® (Bio-Rad Laboratories,
EUA) conectado à leitora.
Nesse tipo de ensaio, a absorbância medida é diretamente proporcional à
concentração de IGFBP-1 presente na amostra. Um conjunto de padrões de IGFBP-1 foi
utilizado para plotar a absorbância da curva padrão versus a concentração de IGFBP-1, e, a
partir dessa curva, as concentrações nas amostras desconhecidas foram calculadas. A curva
padrão foi construída a partir das seguintes concentrações: 0,6; 3; 12; 37; 78 e 155 ng/ml.
Todas as concentrações séricas de um mesmo indivíduo com e sem hipóxia
foram obtidas no mesmo ensaio em duplicatas e os resultados expressos em ng/ml. Os
coeficientes de variação intra- e inter-ensaios foram, respectivamente, 3,3% e 8,8%. A
sensibilidade do ensaio foi 0,025 ng/ml.

56
III.4.3 - Ensaio imunoenzimático da IGFBP-3
As concentrações séricas de IGFBP-3 foram determinadas no Laboratório de
Pediatria do HCFMRP-USP utilizando o kit DSL-10-6600 Active® Total IGFBP-3 ELISA
da Diagnostic Systems Laboratories (DSL – Webster, Texas, EUA). Esse kit é composto
por um imunoensaio enzimático tratando-se, portanto, de um ELISA (enzyme linked
immuno sorbent assay) de “dois passos” do tipo sanduíche.
Nesse ensaio, houve uma etapa na qual as amostras foram diluídas em 1:100
utilizando-se diluente próprio componente do kit.
No ensaio, padrões, amostras e controles em duplicata foram encubados em
cavidades de microtitulação os quais foram revestidos com anticorpo policlonal anti-
IGFBP-3 durante 2 horas em temperatura ambiente (~25°C) agitados a 500-700 rpm em
agitadora orbital The Jitterbug™ model 130000 (Boekel Scientific, Pennsylvania, EUA).
Após essa encubação, as cavidades foram lavadas 5 vezes em lavadora automática
Immunowash Model 1575 (Bio-Rad Laboratories, EUA) utilizando-se o tampão de lavagem
que acompanhou o kit para a dosagem do IGFBP-3. Houve, então, nova encubação onde as
cavidades foram tratadas com outro anticorpo policlonal anti-IGFBP-3 marcado com
enzima HRP durante 1 hora em temperatura ambiente (~25°C) agitados a 500-700 rpm em
agitadora orbital. Após essa segunda encubação e lavagem idêntica ao primeiro
procedimento, as cavidades foram encubadas com o substrato TMB por 10 minutos. Uma
solução ácida foi adicionada e o grau de reposição enzimática foi determinado por medida
de absorbância em duplo comprimento de onda a 450 e 620 nm na leitora iMarkTM
Microplate Absorbance Reader (Bio-Rad Laboratories, EUA). Após a leitura, os dados

57
obtidos foram encaminhados para análise no software Microplate Manager® (Bio-Rad
Laboratories, EUA) conectado à leitora.
Nesse tipo de ensaio, a absorbância medida é diretamente proporcional à
concentração de IGFBP-3 presente na amostra. Um conjunto de padrões de IGFBP-3 foi
utilizado para plotar a absorbância da curva padrão versus a concentração de IGFBP-3, e, a
partir dessa curva, as concentrações nas amostras desconhecidas foram calculadas. A curva
padrão foi construída a partir das seguintes concentrações: 1,2; 4; 15; 39 e 74 ng/ml.
Todas as concentrações séricas de um mesmo indivíduo com e sem hipóxia
foram obtidas no mesmo ensaio em duplicatas e os resultados obtidos em ng/ml. Os
coeficientes de variação intra- e inter-ensaios foram, respectivamente, 4,5% e 10,4%. A
sensibilidade do ensaio foi de 0,04 ng/ml. As concentrações obtidas em ng/ml foram
corrigidas pelo fator de diluição e, após, transformadas em mg/l, sendo que foi nessa
unidade que foram reportadas nos resultados.
III.4.4 - Ensaio imunoenzimático de insulina
As concentrações séricas de insulina foram determinadas no Laboratório de
Pediatria do HCFMRP-USP utilizando o kit DSL-10-1600 Active® Insulina ELISA da
Diagnostic Systems Laboratories (DSL – Webster, Texas, EUA). Esse kit é composto por
um imunoensaio enzimático tratando-se, portanto, de um ELISA (enzyme linked immuno
sorbent assay) de “um passo” do tipo sanduíche.
No ensaio, padrões, amostras e controles em duplicata foram encubados com
anticorpo anti-insulina marcado com enzima HRP em cavidades de microtitulação

58
revestidas com outro anticorpo anti-insulina durante 1 hora em temperatura ambiente
(~25oC) agitados a 500-700 rpm em agitadora orbital The Jitterbug™ model 130000
(Boekel Scientific, Pennsylvania, EUA). Após a encubação, as cavidades foram lavadas 5
vezes em lavadora automática Immunowash Model 1575 (Bio-Rad Laboratories, EUA)
utilizando-se o tampão de lavagem que acompanhou o kit para a dosagem da insulina. Após
a lavagem, as cavidades foram encubadas com o substrato TMB por 10 minutos. Uma
solução ácida de interrupção foi adicionada e o grau de reposição enzimática foi
determinado por medida de absorbância em duplo comprimento de onda a 450 e 620 nm na
leitora iMarkTM
Microplate Absorbance Reader (Bio-Rad Laboratories, EUA). Após a
leitura, os dados obtidos foram encaminhados para análise no software Microplate
Manager® (Bio-Rad Laboratories, EUA) conectado à leitora.
Nesse tipo de ensaio, a absorbância medida é diretamente proporcional à
concentração de insulina presente na amostra. Um conjunto de padrões de insulina foi
utilizado para plotar a absorbância da curva padrão versus a concentração de insulina, e, a
partir dessa curva, as concentrações nas amostras desconhecidas foram calculadas. A curva
padrão foi construída a partir das seguintes concentrações: 3,5; 8,5; 25 e 84 µUI/ml.
Todas as concentrações séricas de um mesmo indivíduo com e sem hipóxia
foram obtidas no mesmo ensaio em duplicatas e os resultados expressos em µUI/ml. Os
coeficientes de variação intra- e inter-ensaios foram, respectivamente, 10,9% e 16%. A
sensibilidade do ensaio foi de 0,26 µUI/ml.

59
III.5 - Avaliação da expressão do IGF1R
III.5.1 – Separação de linfócitos
Amostras de sangue foram colhidas em frascos roxos contendo EDTA com
volumes que variaram entre 0,5 e 1 ml. Foi adicionada a esse material quantidade suficiente
de tampão PBS para obtenção do volume final de 5 ml. Essa solução foi posta sobre a
mesma quantidade de Ficoll-Hypaque® (Histopaque, Sigma Chemical, EUA) que
proporciona um gradiente de densidade para separação de linfócitos de outros elementos do
sangue total, e centrifugada a 1500 rpm durante 30 minutos. Obtiveram-se quatro camadas
distintas em ordem do sobrenadante para o precipitado: soro, células mononucleares que
compõem a camada opaca, Ficoll-Hypaque® e hemácias. Após desprezar as hemácias por
inversão, foram procedidas 3 lavagens acrescentando-se tampão PBS em quantidade
suficiente para 20 ml e centrifugando-se a 2500 rpm durante 10 minutos. Ao final, as
células foram ressuspensas com 500 µl de PBS, produzindo-se, dessa forma, 2 alíquotas de
250 µl às quais foram adicionados 750 µl de TRIzol LS Reagent® (Invitrogen, EUA) cada
uma, numa proporção de 3:1, sendo, então, homogeneizadas e imediatamente armazenadas
em congelador a -80oC até o momento da extração do RNA total dos linfócitos assim
separados.

60
III.5.2 – Extração do RNA
Depois de descongeladas, foram adicionados 200 µl de clorofórmio às
amostras armazenadas em TRIzol LS Reagent® (Invitrogen, EUA) sendo procedida, em
seguida, agitação vigorosa durante 15 segundos. As amostras permaneceram em repouso
por 3 minutos em temperatura ambiente e, após, centrifugadas a 13200 rpm por 15 minutos
a 4ºC. Obtiveram-se 3 fases assim descritas no sentido do sobrenadante para o precipitado:
fase superior (incolor) onde o RNA total está dissolvido, interfase de coloração branca onde
está dissolvido o DNA e proteínas; e a fase inferior de coloração rosa onde está o fenol e o
clorofórmio. A fase aquosa foi transferida para um novo tubo identificado onde foram
adicionados 500 µl de álcool isopropílico a 100% gelado e, imediatamente, postas no gelo.
As amostras, então, permaneceram a -20ºC por pelo menos 12 horas (overnight).
No dia seguinte, foram centrifugadas a 13200 rpm durante 20 minutos a 4ºC,
com isso, foi formado um pellet contendo o RNA total, sendo desprezado o sobrenadante.
Às amostras, foram acrescentados 1000 µl de etanol a 75% seguido novamente de
centrifugação a 13200 rpm por 5 minutos a 4ºC. Desprezou-se a fase superior e o pellet
seco foi dissolvido em 50 µl de água tratada com DEPC, sendo, após identificação correta,
estocado a -80oC até o momento da obtenção do cDNA.

61
III.5.3 – Quantificação do RNA
A quantificação do RNA foi obtida mediante espectrofotometria de onda em
aparelho Gene Quant Pro Spec® (Amersham Bioscience). Nesse método, a obtenção da
concentração de RNA baseia-se na relação entre 1 unidade em A260 ser igual a 40 µg/ml de
RNA dissolvido em água.
III.5.4 – Síntese da primeira fita de cDNA
Após a quantificação de RNA total das amostras armazenadas em freezer a -
80oC, o cDNA foi obtido a partir de aproximadamente 1000 µg de RNA. Foi utilizado o kit
High Capacity® (Applied Biosystems, EUA) onde, para o volume calculado da amostra de
RNA a ser reversamente transcrito, foram adicionados 1 µl de c/NTP, 2,5 µl de random P,
1,25 µl de Multiscribe, 0,63 µl de inibidor de RNase (RNase-Out, Invitrogen, EUA), 2,5 µl
de tampão e 15,12 µl de água tratada com DEPC. Essa mistura foi posta a 25ºC durante 10
minutos e, a seguir, a 37oC durante 2 horas sendo, então, estocada a -20ºC até o momento
da qPCR.
Naquelas amostras onde a quantificação de RNA não foi possível, utilizou-se
a quantidade de RNA de acordo com as recomendações do fabricante do kit High
Capacity® (Applied Biosystems, EUA).

62
III.5.5 – Controles endógenos
Os genes de controle endógeno (housekeeping) são utilizados para a
normalização dos níveis da expressão dos genes de estudo (alvo). Nesse estudo, foram
escolhidos dois genes para controle endógeno: B2M (β2-microglobulina) e o GUSB (β-
glicuronidase).
O B2M localiza-se no braço longo do cromossomo 15 (15q21-q22.2) e
codifica uma proteína relacionada ao complexo de histocompatibilidade maior de classe 1
chamada β-2 microglobulina. O gene GUSB localiza-se no braço longo do cromossomo 7
(7q21.11) e codifica a proteína β-glicuronidase presente em todas as células nucleadas,
sendo que sua expressão é mantida constante em diversos tipos celulares e diferentes
situações do ciclo celular mantendo sua expressão estável. Tais genes, em estudos
anteriores, apresentaram melhor desempenho durante a qPCR quando a expressão gênica
foi analisada em linfócitos humanos (BEILLARD et al., 2003).
Além de controle endógeno, o gene GUSB também foi empregado para
testar a integridade das amostras de cDNA em RT-PCR convencional (conforme descrição
a seguir).
III.5.6 – RT-PCR
Todas as amostras de cDNA obtidas após a sua síntese foram submetidas à
transcrição reversa convencional utilizando primers para amplificação do gene utilizado
para controle endógeno GUSB. Nessa reação, 1,0 µl do cDNA de cada amostra foi

63
adicionado a uma solução contendo 18,8 µl de água, 2,5 µl de solução tampão 5x
(Invitrogen, Brasil), 1,0 µl de cloreto de magnésio 50 nM (Invitrogen, Brasil), 0,5 µl de
solução 10 mM de cada dNTP, 15 pM de cada primer (sense e antisense), 0,2 µl de Taq-
polimerase 5 U/µl (Platinum Taq DNA polymerase, Invitrogen, Brasil), perfazendo um
volume total de 25 µl. As melhores condições de desnaturação, pareamento e amplificação
já haviam sido padronizadas previamente. As amostras foram, então, postas no
termociclador PTC-200 (MJ Research, Inc, EUA) sob as seguintes condições: desnaturação
inicial a 94ºC durante 4 minutos e pareamento a 60ºC por 2 minutos, seguidos de 38 ciclos
compostos por desnaturação a 94ºC, pareamento a 60ºC e extensão a 72ºC durante 1 minuto
cada, seguidos por um ciclo de extensão final a 72ºC durante 10 minutos.
Após a RT-PCR, os seus produtos foram submetidos à eletroforese em gel de
agarose a 2% com brometo de etídeo (descrito no apêndice A). Com isso, demonstrou-se a
integridade do cDNA e a sua capacidade de amplificação. A figura 2 mostra a visualização
do gel de eletroforese do produto de RT-PCR para o gene GUSB.

64
Figura 2 – Imagens obtidas dos géis de agarose a 2% com os produtos da amplificação do
gene GUSB após uma RT-PCR de todos os cDNAs das amostras coletadas.

65
III.5.7 – qPCR
O mRNA total foi extraído a partir de linfócitos periféricos (BERNABEI et
al, 2003) e, após a sua quantificação e obtenção do cDNA, esse foi submetido à técnica de
qPCR que permite a quantificação da expressão de um gene alvo. Tal procedimento permite
que no mesmo aparelho, a reação de transcrição reversa, amplificação do cDNA e
quantificação específica da expressão de um gene, no caso desse estudo, o gene IGF1R,
seja realizada. Para tanto, é necessária a presença de um marcador fluorescente que emite
um sinal quando excitado, por exemplo, por um laser. Esse processo é totalmente
automatizado sendo que no momento em que um feixe de laser é direcionado através dos
tubos ópticos utilizados durante as reações, os dados da fluorescência emitida pelas
amostras são coletados e analisados por um programa de computador possibilitando, dessa
forma, a quantificação (BUSTIN, 2000).
Habitualmente, dois sistemas de sondas fluorescentes têm sido utilizados:
aqueles nos quais as sondas tipo TaqMan® (Applied Biosystems, EUA) são usadas e
aqueles onde o marcador é do tipo SYBR Green® (Applied Biosystems, EUA). Ambos os
sistemas apresentam eficiências semelhantes, porém, quando comparados, apresentam
algumas diferenças em função do princípio básico das reações. Deve ser ressaltado que os
sistemas que utilizam sondas TaqMan® (Applied Biosystems, EUA) apresenta melhor
especificidade dado o uso de sondas específicas para o gene alvo. Esses sistemas são
compostos por 2 primers específicos e uma sonda com 2 fluorocromos diferentes. Um dos
fluorocromos é chamado de reporter e localiza-se na extremidade 5‟ da sonda, o outro
localizado na extremidade 3‟ da sonda é chamado de quencher (repressor). A proximidade
de ambos fluorocromos proporciona uma inibição da fluorescência do reporter através de

66
um fenômeno de transferência de energia conhecido como FRET (fluorescence ressonance
energy transference). A sonda, durante a qPCR, sofre hibridização com uma sequência
específica de interesse. No momento em que ocorre cada ciclo de extensão, a Taq-
polimerase cliva o reporter da sonda, separando-o do quencher o que proporciona a
emissão da fluorescência característica do reporter que é captada e analisada pelo aparelho.
Essa técnica, portanto, é capaz de medir a formação de produtos da amplificação dos
fragmentos de cDNA de forma reprodutível e precisa. É importante observar que a análise
deve ser realizada na fase exponencial da qPCR e que a fluorescência detectada é
proporcional à quantidade de sonda degradada e, portanto, ao produto da RT-PCR
acumulado (SCRIDELI; TONE, 2007).
A leitura obtida pelo aparelho de qPCR foi expressa na forma de número de
ciclos de PCR necessários para que um determinado nível de fluorescência fosse
significativamente detectável assim chamado de ciclo threshold ou Ct. Tal resultado
possibilitou o cálculo do ΔCt onde houve a normalização dos dados oriundos das amostras
em relação aos genes de controle endógeno. Além disso, também foram normalizados os
resultados obtidos das amostras em relação ao ΔCt de um calibrador, e a partir de então, foi
calculado o ΔΔCt das amostras.
Nesse estudo, a expressão do gene IGF1R (gene alvo) foi quantificada
utilizando-se sondas específicas TaqMan® (Applied Biosystems, EUA) durante a
realização da qRT-PCR sendo que todas as reações foram realizadas no aparelho 7500
Real-time PCR System® (Applied Biosystems, EUA) disponibilizado no Laboratório de
Pediatria do HCFMRPUSP.
Todas as reações foram preparadas em duplicatas imersas em gelo e sob
proteção de luz em placas de polipropileno contendo 96 poços próprios para esse fim

67
(UltraAmp 96-well Semi-Skirt PCR plates, Sorenson BioScience Inc, EUA). Todos os
genes do estudo (controles endógenos, GUSB e B2M, e o alvo, IGF1R) foram amplificados
nas amostras do estudo e também nas amostras do calibrador (nesse estudo, o cDNA foi
obtido de um indivíduo normal). Em paralelo, foram acrescentadas reações que não
continham cDNA (controles brancos) e, caso houvesse algum tipo de contaminação nessas
amostras, uma nova reação era realizada.
Nos poços foram colocados 10 µl de TaqMan® Universal Master Mix
(Applied Biosystems, EUA); 1 µl de sonda TaqMan® (Applied Biosystems, EUA)
específica para o gene (GUSB ou B2M ou IGF1R) e 9 µl do cDNA oriundo das amostras e
diluído na proporção de 1:50 em água, totalizando um volume final da reação de 20 µl. A
seguir, a placa foi coberta com adesivo plástico para microplacas resistente a álcool e a
altas temperaturas (Adhesive PCR film, ABgene) e seu conteúdo foi centrifugado durante 3
minutos a 3000 rpm. Os reagentes, dessa forma acondicionados, foram, então, submetidos
às condições para a realização da qPCR no aparelho 7500 Real-time PCR System®
(Applied Biosystems, EUA). Tais condições de temperatura foram as seguintes: pré-
aquecimento até 50ºC durante 2 minutos seguidos de desnaturação realizada a 95ºC durante
10 minutos e, após, ciclos de amplificação e quantificação nos quais as amostras foram
submetidas a 95ºC por 15 segundos e 60ºC por 60 segundos nessa ordem por 40 vezes. A
figura 3 exemplifica um gráfico de amplificação da qPCR.
68
a)
b)
Figura 3 – Imagens obtidas pelo software do aparelho 7500 Real-time PCR Systems® que
representam as curvas de amplificação do gene IGF1R em duas amostras do indivíduo 17.
Na primeira amostra (a), a amplificação na situação de hipóxia aguda; na segunda amostra
(b), a amplificação no momento sem hipóxia. A linha verde representa o threshold (0,01).

69
III.5.8 – Quantificação da expressão gênica
Conforme supracitado, cada amostra foi amplificada em duplicata e, ao
mesmo tempo, para cada gene a ser verificado, foram preparadas, na mesma placa, reações
contendo cDNA obtido de linfócitos extraídos de sangue periférico de um indivíduo
normal. Essa amostra, denominada calibrador, foi utilizada como referência na comparação
entre as amostras na análise da expressão gênica em todas as reações. O calibrador não foi
considerado como referência normal e, sim, somente como parâmetro da expressão gênica
dos genes abordados nesse estudo.
Foram incluídas na análise somente as amostras cujas diferenças de
amplificação entre as duplicatas foi de, no máximo, 1,5 ciclos. As amostras que excederam
tal limite foram repetidas a fim de se obter resultados confiáveis.
O momento em que todas as amostras apresentaram a mesma intensidade de
fluorescência (threshold) foi determinado manualmente sendo fixado em 0,1 para todos os
genes e utilizado em todas as reações de qPCR na sua fase exponencial. Dado o threshold,
o software do aparelho 7500 Real-time PCR Systems® forneceu os valores de Ct de cada
amostra testada. A média aritmética dos valores de Ct das duplicatas foi utilizada para o
cálculo do ΔΔCt.
Nesse estudo, a quantificação da expressão gênica realizada foi a relativa.
Essa modalidade de quantificação determina a mudança na expressão de uma sequência de
ácido nucléico (alvo) em relação a uma mesma sequência presente numa amostra de
calibrador. Uma vez que 2 amostras de um mesmo indivíduo foram obtidas em situações
diferentes, esse tipo de quantificação pode ser usada na comparação entre as expressões

70
verificadas na situação inicial (hipóxia aguda associada a infecções respiratórias) e na
situação final (sem hipóxia).
III.6 - Análise estatística
Todas as variáveis tiveram as médias, desvios-padrão, medianas e análises
interquartílicas calculados. Os resultados foram mostrados na forma de média (±DP) e
medianas (P25 – P75)
O teste de Wilcoxon foi aplicado respectivamente na análise das variáveis
com ou sem distribuição normal, antes e após o tratamento da hipoxemia.
O teste D‟Agostino e Pearson foi utilizado para verificar a normalidade das
variáveis.
Significância estatística será assumida para P 0,05.

71
IV – Resultados

72
IV.1 – Concentrações séricas de IGF-I
As concentrações séricas de IGF-I foram obtidas nas amostras colhidas
durante o momento de hipóxia aguda associada a infecções respiratórias e após a
recuperação da mesma em cada um dos 24 indivíduos, sendo que dentre os 27 indivíduos
inicialmente selecionados, em 3 deles as amostras durante a hipóxia mostraram-se com
volumes insuficientes para a dosagem, fato que ocasionou a exclusão desses sujeitos nessa
análise.
Na tabela 3 estão os resultados obtidos das concentrações séricas de IGF-I
nas crianças durante a hipóxia aguda associada a infecções respiratórias e quando a
normalidade foi restabelecida. Os resultados observados em conjunto, de acordo com a
situação hipóxia ou ausência dela, apresentaram médias de 21,3 (±37,8) e 60,2 (±58,6)
ng/ml, respectivamente. As medianas, sob esse mesmo aspecto, foram 12,3 (6,24 – 24,14)
em hipóxia e 54,8 (24,51 – 72) ng/ml na ausência dela.
A figura 4 ilustra o comportamento das concentrações séricas de IGF-I de
cada indivíduo, inicialmente, na hipóxia aguda associada a infecções respiratórias e, após,
na ausência dela.
Cada indivíduo avaliado foi representado por um símbolo gráfico e uma cor
específicos, sendo que nas figuras 3, 4, 5, 6 e 7 essa representação gráfica se repete e que
cada indivíduo, portanto, conserva a mesma representação em todas as figuras.
Na ausência de padrão de normalidade para as concentrações séricas de IGF-
I em crianças com idade inferior a 2 anos, 23 indivíduos tiveram suas concentrações
comparadas com a referência apresentada nas instruções do fabricante. A mediana

73
apontada, para essa faixa etária, é de 55,25 ng/ml (intervalo entre 13,26 e 82,95 ng/ml), de
acordo com referência apontada pelo fabricante do kit. Na situação de hipóxia, 14 crianças
estiveram abaixo do intervalo normal; na ausência de hipóxia, somente 4 estavam com
concentrações inferiores. As outras 4 crianças (idade superior a 2 anos), tiveram os EDP
das concentrações séricas de IGF-I calculados. A criança número 6 variou de -2,25 para -
2,13 EDP; a criança número 8 variou de -2,3 para -1,67 EDP; a número 18 de -0,3 para
0,88 EDP; e a número 26 de -2,54 para -2,19 EDP, nas situações de hipóxia e após a
recuperação, respectivamente.
As observações foram realizadas em 2 situações diferentes num mesmo
indivíduo, portanto esses mesmos resultados foram pareados e o teste de Wilcoxon foi
utilizado. Houve diferença significativa entre as situações de hipóxia e sem hipóxia (p <
0,0001).

74
Tabela 3 – Concentrações séricas de IGF-I (ng/ml) dos pacientes durante a hipóxia aguda
associada a infecções respiratórias e, após, nos mesmos indivíduos, na ausência dela.
Valores expressos em média, DP e mediana.
Pacientes
Concentrações séricas
de IGF-I (ng/ml)
Hipóxia Sem
hipóxia
1 11,5 26,0
2 16,3 70,0
3 19,2 49,5
4 12,7 108,8
5 0,62 73,8
6 12,3 22,5
7 1,8 53,7
8 20,4 83,5
9 1,3 24,0
10 6,0 38,5
11 9,9 5,0
12 8,0 9,3
13 34,6 59,5
14 32,2 45,3
15 4,4 3,5
16 5,7 56,0
17 10,5 61,5
18 192,9 300,5
19 25,3 58,2
20 7,1 7,4
21 33,8 72,3
23 13,5 71,2
24 18,3 95,8
26 12,4 49,8
Média 21,3 60,2
DP 37,8 58,6
Mediana 12,3 54,8

75
Concentrações séricas de IGF-I
0
50
100
150
200
250
300
350
Hipóxia Sem hipóxia
ng
/dl
Figura 4 – Concentrações séricas de IGF-I (ng/ml) dos pacientes durante a hipóxia aguda
associada a infecções respiratórias e, após, na ausência dela.
IV.2 – Concentrações séricas de IGFBP-1
As concentrações séricas de IGFBP-1 foram obtidas nas amostras colhidas
durante o momento de hipóxia aguda associada a infecções respiratórias e fora dessa
condição em 26 crianças, sendo que em 1, dentre as 27 selecionadas, a amostra durante a
hipóxia mostrou-se com volume insuficiente para a dosagen, fato que ocasionou a sua
exclusão nessa análise.
Na tabela 4, encontram-se os resultados obtidos das concentrações séricas de
IGFBP-1 nos indivíduos durante a hipóxia aguda associada a infecções respiratórias e no

76
momento quando a normalidade foi restabelecida. Os resultados observados em conjunto de
acordo com a situação hipóxia ou ausência dela apresentaram médias de 135,3 (±79,3) e
75,3 (±55,5) ng/ml, respectivamente. As medianas, sob esse mesmo aspecto, foram 115,5
(77,4 – 171,6) em hipóxia e 68,6 (28,25 – 106,8) ng/ml na ausência dela. Da mesma forma,
a figura 5 ilustra o comportamento das concentrações séricas de IGFBP-1 de cada criança,
inicialmente, na hipóxia e, após, na ausência dela.
Os resultados foram pareados e o teste de Wilcoxon foi utilizado o que
também mostrou diferença significativa entre as situações de hipóxia e sem hipóxia (p =
0,0014).

77
Tabela 4 – Concentrações séricas de IGFBP-1 (ng/ml) dos pacientes durante a hipóxia
aguda associada a infecções respiratórias e, após, nos mesmos indivíduos, na ausência dela.
Valores expressos em média, DP e mediana.
Pacientes
Concentrações séricas
de IGFBP-1(ng/ml)
Hipóxia Sem
hipóxia
1 105,7 137,6
2 193,4 63,2
3 95,4 86,9
4 175,4 12,2
5 136,4 15,5
6 169,7 179,6
7 78,7 37,2
8 170,4 37,8
9 140,9 56,9
10 224,9 103,8
11 109,3 88
12 155,3 93
13 53,2 18,7
14 31,6 30,5
15 65,9 207
16 211,2 81
17 113,5 20,1
18 111,8 124,1
19 330,2 170,2
20 117,5 104,2
21 40,9 73,9
23 157,7 37,1
24 73,6 21,4
25 331,3 114,6
26 10,5 8,6
27 113,3 33,1
Média 135,3 75,3
DP 79,3 55,5
Mediana 115,5 68,6
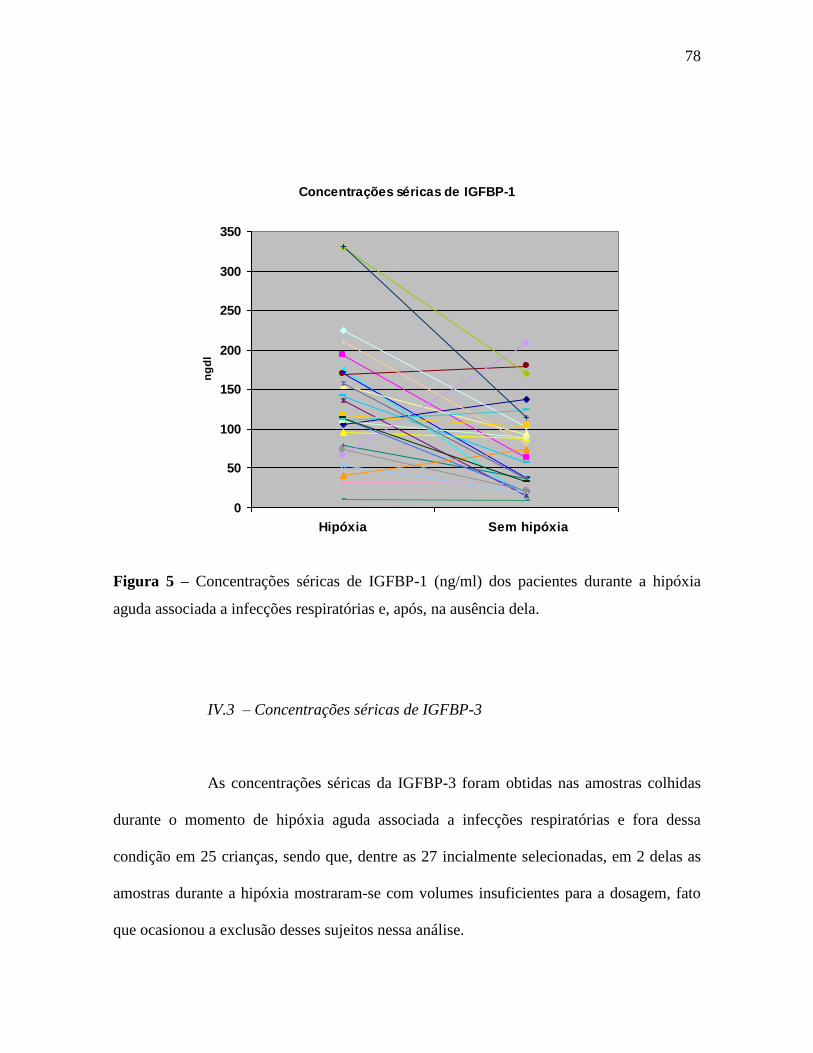
78
Concentrações séricas de IGFBP-1
0
50
100
150
200
250
300
350
Hipóxia Sem hipóxia
ng
dl
Figura 5 – Concentrações séricas de IGFBP-1 (ng/ml) dos pacientes durante a hipóxia
aguda associada a infecções respiratórias e, após, na ausência dela.
IV.3 – Concentrações séricas de IGFBP-3
As concentrações séricas da IGFBP-3 foram obtidas nas amostras colhidas
durante o momento de hipóxia aguda associada a infecções respiratórias e fora dessa
condição em 25 crianças, sendo que, dentre as 27 incialmente selecionadas, em 2 delas as
amostras durante a hipóxia mostraram-se com volumes insuficientes para a dosagem, fato
que ocasionou a exclusão desses sujeitos nessa análise.

79
Na tabela 5, estão os resultados obtidos das concentrações séricas da IGFBP-
3 nos pacientes durante a hipóxia aguda associada a infecções respiratórias e após, quando
a normalidade foi restabelecida. Os resultados observados em conjunto de acordo com a
situação hipóxia ou ausência dela apresentaram médias de 1,53 (±0,8) e 2,34 (±0,7) mg/l,
respectivamente. As medianas, sob esse mesmo aspecto, foram 1,32 (0,86 – 2,17) mg/l em
hipóxia e 2,25 (1,9 – 2,78) mg/l em condições normais.
Da mesma forma, a figura 6 ilustra o comportamento das concentrações
séricas da IGFBP-3 de cada indivíduo, inicialmente, na hipóxia aguda associada a infecções
respiratórias e, após, na ausência dela.
Os resultados das concentrações séricas de IGFBP-3 foram pareados e o
teste de Wilcoxon foi utilizado o que mostrou diferença significativa entre as situações de
hipóxia e sem hipóxia (p = 0,0002).
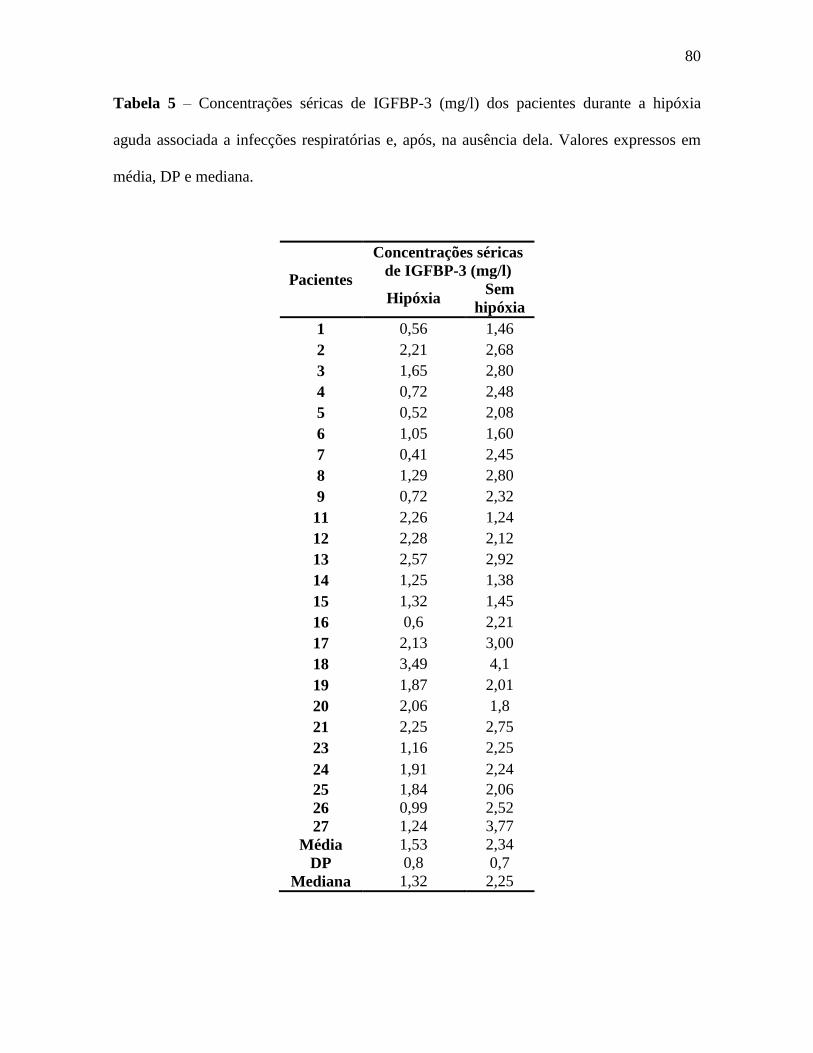
80
Tabela 5 – Concentrações séricas de IGFBP-3 (mg/l) dos pacientes durante a hipóxia
aguda associada a infecções respiratórias e, após, na ausência dela. Valores expressos em
média, DP e mediana.
Pacientes
Concentrações séricas
de IGFBP-3 (mg/l)
Hipóxia Sem
hipóxia
1 0,56 1,46
2 2,21 2,68
3 1,65 2,80
4 0,72 2,48
5 0,52 2,08
6 1,05 1,60
7 0,41 2,45
8 1,29 2,80
9 0,72 2,32
11 2,26 1,24
12 2,28 2,12
13 2,57 2,92
14 1,25 1,38
15 1,32 1,45
16 0,6 2,21
17 2,13 3,00
18 3,49 4,1
19 1,87 2,01
20 2,06 1,8
21 2,25 2,75
23 1,16 2,25
24 1,91 2,24
25 1,84 2,06
26 0,99 2,52
27 1,24 3,77
Média 1,53 2,34
DP 0,8 0,7
Mediana 1,32 2,25
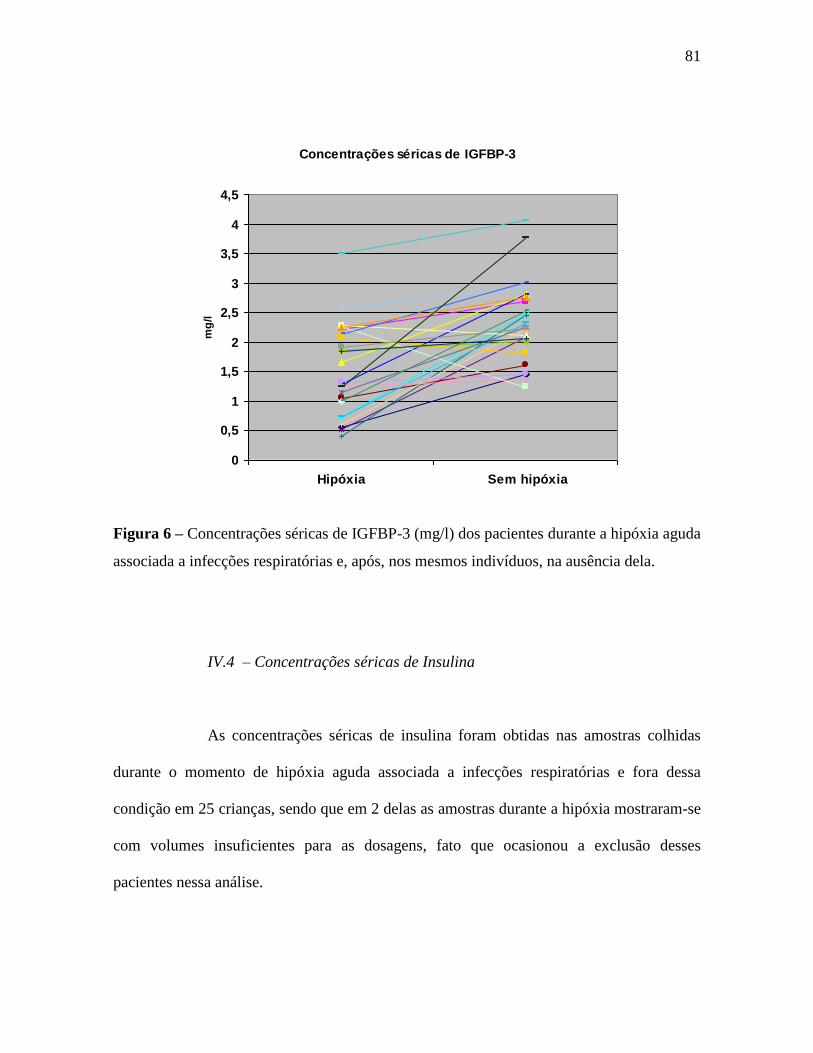
81
Concentrações séricas de IGFBP-3
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
Hipóxia Sem hipóxia
mg
/l
Figura 6 – Concentrações séricas de IGFBP-3 (mg/l) dos pacientes durante a hipóxia aguda
associada a infecções respiratórias e, após, nos mesmos indivíduos, na ausência dela.
IV.4 – Concentrações séricas de Insulina
As concentrações séricas de insulina foram obtidas nas amostras colhidas
durante o momento de hipóxia aguda associada a infecções respiratórias e fora dessa
condição em 25 crianças, sendo que em 2 delas as amostras durante a hipóxia mostraram-se
com volumes insuficientes para as dosagens, fato que ocasionou a exclusão desses
pacientes nessa análise.

82
Na tabela 6, estão os resultados obtidos das concentrações séricas de insulina
nas crianças durante a hipóxia aguda associada a infecções respiratórias e quando a
normalidade foi restabelecida. Os resultados observados em conjunto de acordo com a
situação hipóxia ou ausência dela apresentaram médias de 11,7 (±7) e 14,5 (±9,6) µUI/ml,
respectivamente. As medianas, sob esse mesmo aspecto, foram 9,1 (8,48 – 11,1) em
hipóxia e 9,8 (8,79 – 18,6) µUI/ml em condições normais. Da mesma forma, a figura 7
ilustra o comportamento das concentrações séricas de insulina de cada indivíduo,
inicialmente, na hipóxia e, após, na ausência dela.
Os resultados das concentrações séricas de insulina dos indivíduos foram
pareados e o teste de Wilcoxon foi utilizado. Nessa análise, não houve diferença
significativa entre as situações de hipóxia e sem hipóxia (p = 0,132).

83
Tabela 6 – Concentrações séricas de insulina (µUI/ml) dos pacientes durante a hipóxia
aguda associada a infecções respiratórias e, após, na ausência dela. Valores expressos em
média, DP e mediana.
Pacientes
Concentrações séricas
de insulina (µUI/ml)
Hipóxia Sem
Hipóxia
1 8,0 20,2
2 9,6 8,9
3 11,1 8,4
4 12,8 20,8
5 8,3 11,8
6 8,5 9,1
7 10,2 42,4
9 8,4 11,0
10 8,5 8,6
11 11,9 8,8
12 9,3 8,9
13 20,3 34,2
14 33,7 9,6
15 8,4 8,8
16 9,0 17,0
17 33,2 9,8
18 10,6 8,6
19 9,1 8,8
20 9,0 8,4
21 9,0 10,7
23 7,5 8,1
24 8,7 10,5
25 9,3 21,8
26 7,9 35,2
27 11,0 12,0
Média 11,7 14,5
DP 7,0 9,6
Mediana 9,1 9,8

84
Concentrações séricas de Insulina
0
5
10
15
20
25
30
35
40
45
Hipóxia Sem Hipóxia
mic
roU
I/m
l
Figura 7 – Concentrações séricas de insulina (µUI/ml) dos pacientes durante a hipóxia
aguda associada a infecções respiratórias e, após, na ausência dela.
IV.5 – Expressão do IGF1R
Foram realizadas qPCRs de todas as amostras de 26 indivíduos, sendo que 1
indivíduo foi excluído dessa análise pois, apesar do fato de que a sua amostra apresentou
amplificação adequada quando foi testada utilizando RT-PCR para o gene GUSB, as
amostras, quando submetidas à qPCR, não apresentaram amplificação adequada, além de
apresentarem por diversas vezes duplicatas díspares. Os resultados obtidos de todos os

85
indivíduos nas duas situações (hipóxia e sem hipóxia), quando agrupados em conjunto,
mostraram um Ct, em média, de 32,33 (±2,45) e mediana de 32,34.
A expressão relativa do IGF1R foi quantificada mediante utilização do
cálculo do ∆∆Ct. Assim, os resultados foram analisados em conjunto, os indivíduos quando
na situação de hipóxia apresentaram quantificação relativa da expressão do IGF1R em
média de 1,61 (±1,26) e mediana de 1,28 (0,8 – 2,11). Fora da hipóxia a quantificação
relativa da expressão desse receptor foi em média 1,21 (±1,1) e a mediana foi de 0,93 (0,41
– 1,7).
Na tabela 7, encontram-se os resultados da quantificação relativa do gene
IGF1R de todos os indivíduos nas situações de hipóxia aguda associada a infecções
respiratórias e normalidade. Deve ser ressaltado que esses resultados são relativos à
expressão do gene alvo em comparação a um indivíduo (calibrador). Nessa tabela, a
quantificação da expressão do gene IGF1R é mostrada de acordo com a situação de
oxigenação sanguínea das crianças, bem como os valores da média, desvio-padrão e
mediana.
A figura 8 mostra a evolução da expressão do gene do receptor tipo 1 de IGF
nos indivíduos quando em hipóxia aguda associada a infecções respiratórias e, após
melhora clínica, fora dessa condição.

86
Tabela 7 – Quantificação relativa da expressão do IGF1R em linfócitos de sangue
periférico, de acordo com a situação hipóxia aguda associada a infecções respiratórias ou
sem hipóxia. Valores expressos em média, DP e mediana.
Pacientes
Quantificação
Relativa
Hipóxia Sem
hipóxia
1 2,77 1,98
2 0,33 0,46
3 1,39 0,86
4 1,70 4,60
5 4,54 1,59
6 0,65 0,50
7 2,03 1,46
8 1,81 0,49
9 1,15 0,99
10 0,25 0,10
11 0,27 0,22
12 1,12 0,25
13 1,04 0,05
14 1,62 1,55
15 1,91 0,21
16 2,55 1,10
17 1,31 0,79
18 0,82 1,17
19 5,61 3,14
20 2,36 0,52
21 1,25 2,70
22 0,10 0,27
24 1,17 2,62
25 0,76 0,73
26 0,87 1,01
27 2,45 2,06
Média 1,61 1,21
DP 1,26 1,1
Mediana 1,28 0,93

87
Expressão do gene IGF1R
0
1
2
3
4
5
6
Hipóxia Sem Hipóxia
Figura 8 – Expressão relativa do IGF1R em linfócitos de sangue periférico de pacientes em
hipóxia aguda associada a infecções respiratórias e, após, na ausência dela.
Considerando-se que as mesmas crianças foram observadas em 2 situações
diferentes, os resultados da quantificação relativa do IGF1R demonstraram aumento
significativo na expressão relativa do IGF1R na situação de hipóxia (p = 0,0373).

88
IV.6 – Síntese dos resultados
Na tabela 8 são apresentadas as médias (±DP), as medianas e os níveis de
significância alcançados quando da comparação dos resultados das concentrações séricas de
IGF-I, IGFBP-1, IGFBP-3, insulina e expressão do IGF1R nos linfócitos de sangue
periférico, além da própria oximetria de pulso nas situações de hipóxia associada à infecção
respiratória aguda e sem hipóxia. Nessa tabela, os resultados e suas comparações são
apresentados de forma sintética o que permite maior clareza dos dados encontrados.
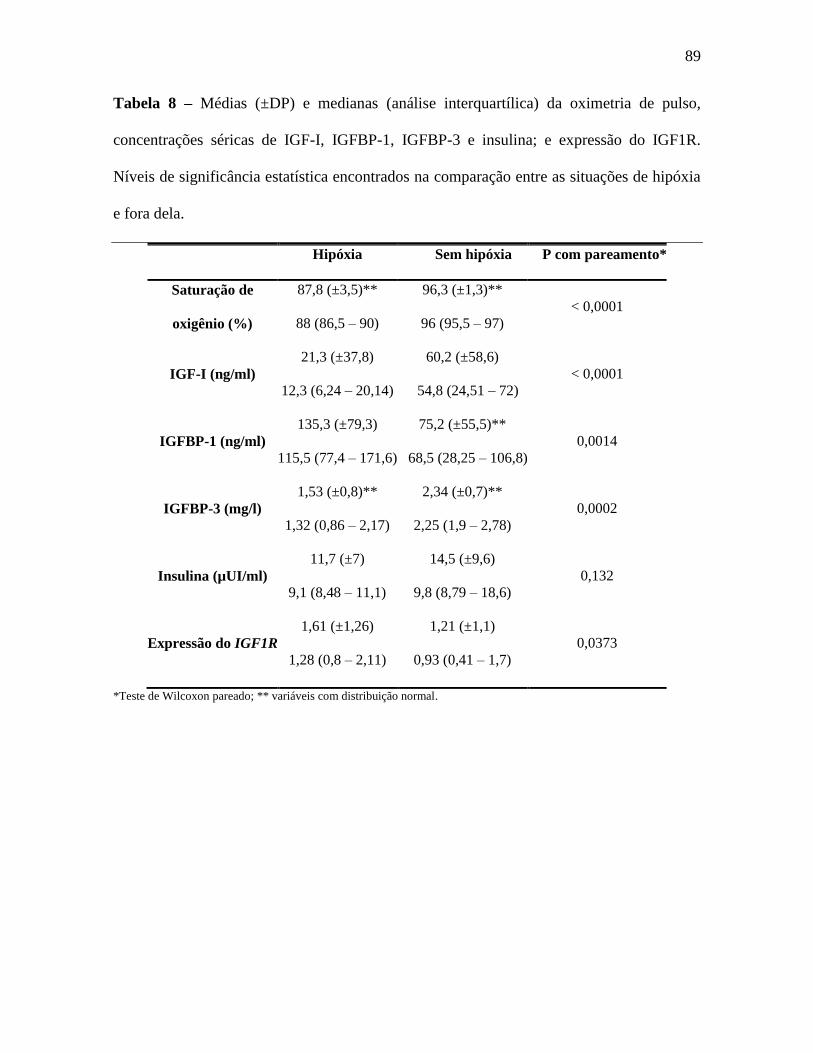
89
Tabela 8 – Médias (±DP) e medianas (análise interquartílica) da oximetria de pulso,
concentrações séricas de IGF-I, IGFBP-1, IGFBP-3 e insulina; e expressão do IGF1R.
Níveis de significância estatística encontrados na comparação entre as situações de hipóxia
e fora dela.
Hipóxia Sem hipóxia P com pareamento*
Saturação de
oxigênio (%)
87,8 (±3,5)**
88 (86,5 – 90)
96,3 (±1,3)**
96 (95,5 – 97)
< 0,0001
IGF-I (ng/ml)
21,3 (±37,8)
12,3 (6,24 – 20,14)
60,2 (±58,6)
54,8 (24,51 – 72)
< 0,0001
IGFBP-1 (ng/ml)
135,3 (±79,3)
115,5 (77,4 – 171,6)
75,2 (±55,5)**
68,5 (28,25 – 106,8)
0,0014
IGFBP-3 (mg/l)
1,53 (±0,8)**
1,32 (0,86 – 2,17)
2,34 (±0,7)**
2,25 (1,9 – 2,78)
0,0002
Insulina (µUI/ml)
11,7 (±7)
9,1 (8,48 – 11,1)
14,5 (±9,6)
9,8 (8,79 – 18,6)
0,132
Expressão do IGF1R
1,61 (±1,26)
1,28 (0,8 – 2,11)
1,21 (±1,1)
0,93 (0,41 – 1,7)
0,0373
*Teste de Wilcoxon pareado; ** variáveis com distribuição normal.

90
V – Discussão

91
O principal objetivo desse estudo foi verificar a resposta de componentes do
sistema IGF à hipóxia aguda em crianças. Os resultados obtidos mostraram uma redução
significativa das concentrações séricas de IGF-I e de IGFBP-3, aumento das concentrações
séricas de IGFBP-1, concentrações séricas de insulina inalteradas e aumento da expressão
do IGF1R em crianças durante a hipóxia aguda.
A RCIU, situação na qual provavelmente há hipóxia crônica, tem sido
instrumento de estudo do comportamento do sistema IGF mediante restrição de oxigênio.
Sob esse aspecto, estudos procuraram relacionar fetos com RCIU e concentrações de IGF-I
e IGFBP-1 no líquido amniótico e no plasma materno, no entanto, não houve correlação
entre RCIU e as concentrações desses fatores (BAHTIA et al., 2002). Em outra análise, as
concentrações amnióticas de IGF-I, IGFBP-1 e leptina foram avaliadas e, novamente, não
houve correlação com a RCIU (ROIG et al., 2005). Alguns modelos animais de hipóxia
intra-uterina também demonstraram que as concentrações séricas de IGF-I e –II nos filhotes
de ovinos e de murinos, apesar da acidose e da RCIU a que foram submetidos, não se
alteravam mediante estímulo hipoxêmico, apesar da redução ocorrida do mRNA hepático
do IGF-I (McLELLAN et al., 1992; TAPANAINEN et al., 1994). As concentrações de
IGF-II tenderam a ser maiores alcançando exatamente o nível de significância (p=0,05) em
filhotes de ratos (TAPANAINEN et al., 1994).
Os efeitos da hipóxia aguda intermitente em fetos ovinos pré e a termo no
sistema IGF foi verificada em estudo no qual a oclusão episódica (90 segundos) do cordão
umbilical era realizada a cada 30 minutos durante 3 a 5 horas por dia durante 4 dias. As
concentrações séricas de IGF-I, IGF-II, IGFBP-1, IGFBP-2, IGFBP-3 e IGFBP-4
permaneceram inalteradas durante todo o período de 4 dias (GREEM et al., 2000). Vale

92
ressaltar que em alguns desses estudos houve também a restrição de nutrientes, uma vez
que houve redução experimental do fluxo sanguíneo uterino ou redução da circulação
umbilical.
A redução das concentrações plasmáticas do IGF-I na hipóxia aguda em
crianças com infecções respiratórias verificada no presente estudo diverge de alguns dados
obtidos em análises realizadas em animais e nas situações de RCIU em humanos. Essas
análises foram obtidas em filhotes de animais submetidos à hipóxia durante a gestação ou
em RNs humanos. As crianças que participaram do presente estudo já não estavam sob a
influência do meio intra-uterino quando foram submetidos à hipóxia, portanto foram
submetidos à restrição aguda de oxigênio diretamente.
O sistema IGF também foi abordado em situações de hipoxemia devido a
cardiopatias congênitas sendo que, dentre essas patologias, aquelas que provocam cianose
estão mais comumente relacionadas a prejuízos importantes do crescimento quando
comparadas às cardiopatias congênitas não cianosantes. Um estudo comparou crianças com
cardiopatias congênitas cianosantes bem nutridas e mal nutridas com controles normais
pareados para idade e sexo abordando as concentrações séricas de IGF-I e sua relação com
a hipoxemia crônica. Observou-se que as concentrações séricas de IGF-I do grupo com
estado nutricional comprometido foram as menores dentre os 3 grupos (p < 0,05).
Entretanto, as crianças cardiopatas consideradas bem nutridas apresentaram concentrações
séricas de IGF-I reduzidas quando comparadas aos controles (p < 0,05) (DÜNDAR;
AKÇORAL; SAYLAM et al., 2000). Salienta-se que nessa análise, não houve influência de
outros fatores, como os estados infecciosos; e, além disso, o efeito hipoxêmico verificado
nessa análise também não foi agudo. Na presente avaliação, além do fato de que processos
infecciosos estiveram envolvidos na etiologia da hipóxia, essa instalou-se de forma aguda e,

93
contudo, esteve também relacionada a reduções significativas das concentrações séricas de
IGF-I em crianças.
Por outro lado, abordando o comportamento do sistema IGF em quadros
infecciosos, as concentrações séricas de GH, IGF-I, IGFBP-1, IGFBP-3 e insulina foram
obtidas em 27 crianças admitidas em unidade de terapia intensiva apresentando choque
séptico por meningococcemia. Em 12 delas, um perfil de secreção de GH de 6 horas
evidenciou que, dentre os não sobreviventes, as medianas foram maiores que dentre aqueles
que sobreviveram. As concentrações séricas de IGF-I total estiveram reduzidas em todas as
crianças participantes do estudo quando comparadas a crianças normais. A média de
escores de desvio-padrão do IGF-I sérico total foi menor entre aqueles que não
sobreviveram, porém os dados obtidos não atingiram significância estatística. Entretanto, as
concentrações séricas de IGF-I livre foram menores naquelas crianças não sobreviventes.
As concentrações séricas de IGFBP-1, no momento da admissão hospitalar, mostraram-se
extremamente elevadas nos indivíduos não sobreviventes, sendo que, nos sobreviventes, as
concentrações séricas dessa proteína reduziram-se significativamente nas 48 horas
seguintes sem correlação com as concentrações séricas de insulina. No momento da
admissão hospitalar, as concentrações séricas de IGFBP-3, tanto em sobreviventes quanto
em não sobreviventes, apresentavam-se reduzidas quanto comparadas com pares saudáveis,
com aumento significativo durante a evolução nos indivíduos sobreviventes (DE GROOF
et al., 2002). A inclusão das crianças nesse estudo baseou-se em achados clínicos
sugestivos de meningococcemia, aliados a culturas positivas para o referido agente
(Neisseria meningitidis). Além disso, os indivíduos deveriam apresentar também
hipotensão ou outros sinais indiretos de hipoperfusão tecidual, tais como, acidose
metabólica inexplicada, hipoxemia arterial (pO2 menor que 75 mmHg ou oximetria de

94
pulso menor que 96%) ou insuficiência renal aguda. Os dados apresentados por De Groof et
al. em 2002 foram semelhantes aos que foram observados no presente estudo quanto à
evolução das concentrações séricas do IGF-I, IGFBP-1, IGFBP-3 e insulina. Apesar do fato
de que um dos principais critérios de inclusão adotados por De Groof et al. em 2002 foi a
infecção grave por Neisseria meningitidis, deve ser ressaltado que para ser considerado
grave o estado clínico do indivíduo incluído, a criança deveria apresentar outras alterações
que per se cursam com graus variados de hipoxemia tecidual.
Em outro estudo, uma coorte de 32 crianças com bronquiolite,
comprovadamente provocada pelo vírus sincicial respiratório, foi avaliada mediante
obtenção das concentrações séricas de cortisol, prolactina, leptina, IGF-I e GH durante o
quadro respiratório agudo e 3 meses após o evento crítico. Dessa coorte, 13 indivíduos
necessitaram de ventilação mecânica e cuidados intensivos; e 19 foram acompanhados sem
esse tipo de assistência, porém, como critério de inclusão no estudo, necessitaram de oferta
suplementar de oxigênio a fim de manter a oximetria de pulso acima de 92%. Nos
indivíduos que utilizaram ventilação mecânica, as concentrações séricas de prolactina e GH
foram maiores quando comparadas com aquelas obtidas em pacientes que foram mantidos
em enfermaria, portanto, sem assistência ventilatória mecânica; o comportamento das
concentrações séricas desses hormônios também foi semelhante quando as suas
concentrações séricas foram comparadas com as concentrações séricas obtidas na situação
de convalescência, ou seja, 3 meses após o evento agudo. Além disso, os pacientes que
necessitaram de ventilação mecânica apresentaram concentrações séricas de IGF-I e leptina
menores que os indivíduos internados que não necessitaram dessa assistência. Dentre esses
hormônios avaliados, somente IGF-I e cortisol apresentaram diferença significativa quando
comparadas as concentrações séricas obtidas na internação sem o auxílio da ventilação

95
mecânica e a convalescência, onde as concentrações séricas do IGF-I foram menores
durante o evento agudo (TASKER et al., 2004). A evolução das concentrações séricas de
IGF-I também foi semelhante aos dados ora observados, porém sem as informações
complementares oriundas de outros componentes do sistema IGF.
Em ambos os estudos supracitados, houve inclusão de indivíduos com algum
grau de hipóxia e, além dos quadros infecciosos bem estabelecidos, diversos participantes
necessitaram de ventilação mecânica. Nessas análises também houve relação entre a
gravidade do quadro clínico e a resposta mais acentuada dos componentes do sistema IGF,
tais como redução das concentrações séricas de IGF-I e aumento das concentrações séricas
de IGFBP-1. Apesar da ausência de crianças que necessitassem de ventilação mecânica ou
que tivessem apresentado choque séptico, esses dados foram semelhantes aos encontrados
no presente estudo, ademais, deve ser ressaltado que há grande dificuldade em controlar os
efeitos no sistema IGF do estado infeccioso, da hipóxia e do tratamento per se, uma vez que
são utilizadas medicações, sedação, oferta suplementar de oxigênio, o que em seu conjunto,
provavelmente, pode representar dificuldade na identificação do real desencadeador da
resposta observada. Além disso, os dados ora apresentados sugerem uma recuperação
precoce das concentrações de IGF-I, visto que a estada hospitalar das crianças foi em média
de apenas 8 dias. Esse intervalo de tempo em que as amostras foram colhidas foi suficiente
para que houvesse aumento significativo em relação às concentrações verificadas no evento
agudo.
Em fetos ovinos submetidos à hipóxia intra-uterina mediante redução do
fluxo sanguíneo do útero, houve queda da saturação e da pressão parcial de oxigênio, além
da redução significativa do pH arterial nas primeiras 8 horas. Sob o efeito dessa hipóxia,
houve aumento de 80% e redução de 50% das concentrações plasmáticas da IGFBP-1 e

96
IGFBP-2, respectivamente, sendo que tais alterações já eram presentes após 4 horas da
ligação das artérias uterinas. Ainda nesse estudo, as fontes produtoras de IGFBPs foram
pesquisadas sendo que o fígado (IGFBP-1 e -2) e os rins (IGFBP-2) foram os tecidos onde
o mRNA de cada uma dessas proteínas foi detectado. Em situação de hipóxia, houve
aumento de 10 vezes no mRNA da IGFBP-1 no fígado e, nos fetos que tiveram maior
aumento nesse mRNA, também houve detecção nos rins. Deve ser ressaltado que esse
estudo foi o primeiro a demonstrar in vivo que os genes responsáveis pela IGFBP-1 e pela
IGFBP-2 poderiam ser induzidos ou suprimidos dentro de 2 a 4 horas do início da condição
patológica instalada (McLELLAN et al., 1992). Da mesma forma, ratas submetidas à
hipóxia (atmosfera com 13-14% de oxigênio) por 7 dias durante a gestação apresentaram
filhotes com RCIU. Os filhotes apresentaram redução do peso em 24%, seus fígados e
placentas foram 20% e 10% mais leves, respectivamente, que os controles. As
concentrações séricas da IGFBP-1 e -2 foram 1,75 vezes maior que os controles enquanto
que a concentração sérica de IGFBP-4 foi 2,5 vezes maior. A expressão do mRNA de
IGFBP-1 e IGFBP-2 nos fígados fetais submetidos à hipóxia foram 4 e 6 vezes maiores que
os controles, respectivamente. O mRNA hepático da IGFBP-4 não apresentou alteração
(TAPANAINEN et al., 1994).
A presença de IGFBP-1 em tecidos de peixe Danio rerio em estágios
precoces do desenvolvimento embrionário permitiu o estudo da expressão dessa proteína
em embriões, haja vista que os estudos realizados em mamíferos foram em fetos de estágios
avançados do desenvolvimento intra-uterino, muito provavelmente devido às dificuldades
inerentes ao acesso de fetos em estágios mais precoces. Porém, nesse estudo realizado em
peixes, os embriões vivem livremente em água sendo, portanto, facilmente acessíveis à
pesquisa. Dessa forma, foram detectados transcritos de IGFBP-1 em todo o

97
desenvolvimento embrionário, inicialmente em pequenas quantidades e, subsequentemente,
em maiores quantidades em tecidos esqueléticos da porção anterior do corpo, tornando-se
indetectável ao término do desenvolvimento embrionário. Nos estágios mais tardios, a
produção concentrou-se no fígado. No período embrionário tardio, a hipóxia (ambiente com
10% da concentração de oxigênio) durante 24 horas provocou aumento na detecção
hepática da transcrição da IGFBP-1. O mesmo procedimento em peixes adultos provocou
aumentos de até 280 vezes o mRNA da IGFBP-1 quando comparados com peixes em
condições normais. Por outro lado, a restrição nutricional foi capaz de promover aumentos
significativos nas concentrações séricas de IGFBP-1 em peixes adultos, sendo que após 2
semanas nessa condição, houve aumento de até 10 vezes no mRNA hepático da IGFBP-1
(MAURES; DUAN, 2002). Embora esses estudos tenham sido realizados em peixes, a
análise estrutural da IGFBP-1 demonstra semelhanças estruturais importantes com a
proteína humana, além de constituir o único modelo vertebrado com essa finalidade.
Apesar das controvérsias apontadas nos estudos em relação ao
comportamento das concentrações das IGFBPs e suas expressões nas situações de hipóxia,
em todos esses estudos houve aumento na quantidade de mRNA da IGFBP-1. Portanto,
comparando-se esses estudos prévios, houve semelhança entre os resultados obtidos das
concentrações séricas de IGFBP-1 e de sua expressão na hipóxia, associada ou não, à
restrição de nutrientes em animais adultos ou fetos, havendo aumento tanto da expressão,
principalmente hepática, quanto das concentrações da IGFBP-1 (McLELLAN et al., 1992;
TAPANAINEN et al., 1994; MAURES; DUAN, 2002). Além disso, em nenhuma dessas
análises, foram incluídos animais com infecções. Os resultados do presente estudo no qual
as concentrações séricas de IGFBP-1 estiveram aumentadas em crianças com hipóxia
associada a doenças respiratórias agudas são semelhantes aos encontrados nos modelos

98
animais supracitados de hipóxia intra-uterina. Entretanto, deve ser ressaltado que, durante
restrição nutricional, habitualmente há queda das concentrações plasmáticas de insulina, o
que promove aumento nas concentrações de IGFBP-1, uma vez que a expressão e secreção
da IGFBP-1 têm como um de seus principais reguladores a insulina. No presente estudo, as
concentrações séricas de insulina permaneceram inalteradas quando as situações de hipóxia
e ausência dela foram comparadas. Esse dado sugere que a regulação realizada pela insulina
na produção de IGFBP-1, na presente análise, foi menos importante que outros efeitos
regulatórios, tais como a própria hipóxia ou o insulto infeccioso. Da mesma forma,
pacientes com choque séptico devido à meningococcemia e indivíduos com bronquiolite
por vírus sincicial respiratório internados em unidade de terapia intensiva, apresentaram
aumento significativo das concentrações de IGFBP-1 correlacionadas com a gravidade do
quadro clínico (DE GROOF et al., 2002; TASKER et al., 2004). Portanto, as
concentrações séricas aumentadas de IGFBP-1 ora observadas, apesar da dificuldade
encontrada em controlar a provável influência do componente infeccioso, são semelhantes
àquelas verificadas em outras situações de hipóxia.
A maioria das respostas de transcrição observadas durante a hipóxia é obtida
mediante ativação do sistema que envolve o HIF-1. Na realidade, esse fator é um
heterodímero composto por 2 subunidades (HIF-1α e HIF-1β) (WANG; SEMENZA, 1995)
onde o HIF-1β é expresso constitutiva e independentemente da disponibilidade de oxigênio
no núcleo de todas as células. Entretanto, o HIF-1α é extremamente lábil e, em condições
de oferta adequada de oxigênio, é hidroxilado em resíduos de prolina permitindo a ligação
com o produto do gene de supressão tumoral de von Hippel-Lindau (pVHL), um
componente de reconhecimento do complexo ubiquitina-ligase, o que torna o HIF-1α alvo
de degradação proteassomal (McLELLAN et al., 1992; WANG; SEMENZA, 1995; OHH et

99
al., 2000). Nas situações onde a disponibilidade de oxigênio é reduzida, ocorre bloqueio
dessa hidroxilação dos resíduos de prolina permitindo o acúmulo de HIF-1α e a formação
do complexo α-β no núcleo celular. Com isso, ocorre a ligação às sequências de DNA
chamadas de HRE, que são necessárias à transcrição gênica específica da situação de
hipóxia (KAJIMURA; AIDA; DUAN, 2006).
Em 1998, foi observado aumento significativo das concentrações circulantes
de IGFBP-1 em sangue de artéria umbilical de crianças nascidas com acidose mista
(componentes respiratório e metabólico presentes simultaneamente) (TAZUKE et al.,
1998). Nesse mesmo estudo, células de carcinoma hepatocelular humano (HepG2) foram
encubadas em ambiente contendo pO2 de 2% e tiveram, com isso, aumento significativo na
concentração de IGFBP-1 no meio de cultura e no seu mRNA. Os autores, analisando a
estrutura do gene dessa proteína, consideraram haver 3 prováveis HREs no intron 1 e
concluíram que a resposta da IGFBP-1 à hipóxia poderia ocorrer mediante ativação das
HREs. Além disso, a co-transfecção de plasmídio que expressa HIF-1α constitutivamente
proporcionou aumento de até 4 vezes na atividade do gene repórter. A sequência de
nucleotídeos que compõe o promoter desse gene apresenta diversos elementos regulatórios
responsivos ao AMPc (CRE), aos glicocorticóides (GRE-I e II), à insulina (IRS-A e B),
além do fator nuclear de hepatócito (HNF-1) e do elemento TATA (MURRAY; PERDEW,
2008). Os dados obtidos no presente estudo demonstram que o aumento das concentrações
séricas de IGFBP-1 pode fazer parte do repertório de respostas do sistema IGF mediante
estímulo hipoxêmico nos seres humanos, particularmente em crianças, inclusive com
mecanismos moleculares, ao menos, parcialmente descritos. É importante ressaltar que essa
proteína ligante, dentre as suas diversas ações, é capaz de inibir as ações biológicas do IGF-
I, fato que, aliado à redução das suas concentrações séricas, também colaboraria num

100
possível mecanismo de contenção do crescimento durante o evento hipoxêmico agudo,
dado que não foi avaliado na presente análise. Ademais, ao observar o número de
elementos regulatórios, até o momento conhecidos, aliado à falta de informações precisas
sobre a forma como atuam, fica evidente que a regulação do gene codificador da IGFBP-1 é
complexa e composta por diversos elementos que atuam de forma coordenada ou
independente.
As concentrações séricas de IGF-I e IGFBP-3 foram avaliadas em 94
crianças com cardiopatias congênitas, procurando correlacionar esses parâmetros com a
presença ou ausência de cianose e com o estado nutricional, em um estudo prospectivo e
randomizado. Observou-se que dentre os indivíduos com cardiopatia cianosante havia
número significativamente maior de crianças com estado nutricional comprometido.
Ademais, as concentrações séricas de IGF-I foram menores dentre os cardiopatas quando
comparados aos controles (p < 0,001) e menores dentre aqueles com cardiopatias
cianosantes quando comparados aos cardiopatas sem cianose (p < 0,001). Ambos os
grupos, cianótico e não cianótico, apresentaram concentrações séricas de IGF-I menores
que os controles (p < 0,001). Os autores concluíram que a presença de cianose foi o fator
que mais influenciou a redução nas concentrações séricas de IGF-I (DINLEYICI et al.,
2007).
Sob o mesmo aspecto, as concentrações séricas de IGF-I e IGFBP-3 do
sangue de cordão umbilical foram avaliadas em estudo controlado, no qual 18 recém-
nascidos de termo que sofreram asfixia perinatal aguda, foram comparados a controles
normais. As concentrações séricas, tanto do IGF-I quanto da IGFBP-3, foram menores nos
indivíduos que sofreram asfixia e apresentaram correlação positiva com o escore de Apgar
do primeiro e do quinto minutos de vida e com o pH do sangue arterial umbilical, além de

101
correlação negativa com a pCO2 e com o lactato. Esses dados demonstraram que a
exposição à hipóxia e à acidose ao nascimento relacionaram-se a reduções nas
concentrações séricas do IGF-I e da IGFBP-3 (DINLEYICI et al., 2006). Apesar do quadro
infeccioso presente em todos os indivíduos selecionados para o presente estudo, essas duas
análises (DINLEYICI et al., 2006; DINLEYICI et al., 2007) também registraram reduções
nas concentrações séricas de IGF-I e IGFBP-3 semelhantes àquelas aqui observadas,
demonstrando, na ausência de estados infecciosos, influência da hipóxia no sistema IGF.
Entretanto, ao menos em um desses estudos (DINLEYICI et al., 2006), foram avaliados
indivíduos no momento do nascimento, situação a qual, inclusive sob o aspecto da
oxigenação sanguínea, é marcada por várias alterações inerentes ao nascimento que
culminam com o abandono do padrão de circulação fetal. Por outro lado, as concentrações
séricas de IGFBP-3 e IGFBP-4 não apresentaram variações significativas quando avaliadas
em fetos ovinos submetidos à hipóxia devido à restrição do fluxo sanguíneo após ligadura
das artérias uterinas e, em cultura de hepatócitos fetais, a hipóxia promoveu aumento
significativo das concentrações de IGFBP-3, sendo que as concentrações de IGFBP-2 e
IGFBP-4 também aumentaram, porém não significativamente (McLELLAN et al., 1992;
POPOVICI et al., 2001).
Células normais submetidas à hipoxia apresentam redução significativa da
síntese protéica. O mesmo comportamento é observado em células tumorais. Entretanto,
algumas linhagens celulares oriundas de tumor pancreático (AsPC-1) que são resistentes a
baixas concentrações de oxigênio, apesar da redução de até 20% da síntese protéica durante
a hipóxia, apresentaram aumento na síntese de IGFBP-1 e IGFBP-3 (KOGA et al., 2008).
A influência da hipóxia no metabolismo glicêmico, abordando as
concentrações de insulina nessa situação, foi analisada em 8 adultos jovens expostos a

102
atmosferas pobres em oxigênio. Durante a oferta normal e baixa de oxigênio, esses
indivíduos ingeriram 75 g de glicose, sendo avaliados diversos parâmetros a cada 30
minutos até 2 horas após a ingestão. Comparando-se as diferentes situações de oferta de
oxigênio, não houve diferença significativa entre as concentrações séricas de insulina e
peptídeo C, assim como não houve alteração na sensibilidade insulínica, abordada mediante
obtenção do HOMA-IR (KELLY et al. 2009). As concentrações séricas de insulina também
permaneceram inalteradas durante a evolução clínica das crianças que participaram do
presente estudo, dado que foi semelhante ao observado em adultos submetidos à hipóxia,
que simulou a exposição desses indivíduos a altitudes elevadas, o que apontou, em seu
conjunto, para a ausência de efeitos da hipóxia sobre as concentrações séricas de insulina.
Existem diversas relações observadas entre o IGF1R e a RCIU associada à
baixa estatura, porém há poucas análises do comportamento da expressão do IGF1R em
situações de hipóxia, tanto em fetos quanto em crianças ou indivíduos adultos. Porém,
ovelhas submetidas à hipóxia aguda durante a gestação apresentaram aumento na presença
e na expressão do IGF1R nos cones de crescimento neuronais do cérebro e cerebelo nos
seus fetos. É importante ressaltar que a mistura de gases oferecida às ovelhas gestantes
apresentavam concentrações de oxigênio entre 20% e 0%. As reduções da quantidade de
oxigênio foram de 5% a cada 30 minutos, até o momento no qual, os fetos apresentaram
hipóxia grave observada em gasometrias colhidas seriadamente. A concentração de IGF1R
nos cones de crescimento neuronais em condições normais é alta, dessa forma, os autores
propuseram que esse comportamento observado do IGF1R provavelmente seria uma
resposta adaptativa à situação de hipóxia na tentativa de manter o crescimento neuronal em
padrões normais, através da facilitação da ligação entre receptor e IGF-1 (MORGAN;
CHAO, 2004). Em tumores, incluindo melanomas, câncer de cólon, pâncreas, próstata e

103
rins, observou-se expressão elevada do IGF1R (BOHULA; PLAYFORD; MACAULAY,
2003). Além disso, a ativação desse receptor foi capaz de proteger as células tumorais de
diversos agentes favorecedores da apoptose nos tumores sólidos presentes no meio
extracelular, tais como, estresse osmolar, deprivação de nutrientes e hipóxia no micro-
ambiente tumoral (PERETZ et al., 2002). Parte dessas observações foi realizada em tecidos
tumorais e, apesar disso, elas apontam para a existência de uma relação entre os IGFs, o
IGF1R e as adaptações ocorridas nas situações de hipóxia. Nesse mesmo sentido, IGFs e
insulina foram capazes de aumentar a resposta à hipóxia mediante a ativação das vias AKT
e MAPK de sinalização, o que promoveu a estabilização dos fatores HIF-α e β e a
consequente ligação às sequências HRE, necessária à transcrição de diversos genes nas
situações de hipóxia (TREINS et al., 2005). Os dados do presente estudo também
evidenciaram o aumento da expressão do gene IGF1R em linfócitos de sangue periférico de
crianças em hipóxia aguda associada a quadros infecciosos respiratórios.
A expressão do IGF1R já foi demonstrada em todas as subpopulações de
células mononucleares de sangue periférico, em estudos que utilizaram a citometria de
fluxo e anticorpos monoclonais contra o IGF1R (KOOIJMAN et al., 1992). Não houve
diferença nessa expressão quando diferentes indivíduos foram comparados entre si, no
entanto, houve grandes diferenças entre os vários tipos celulares de um mesmo indivíduo.
Em geral, a expressão do IGF1R é baixa nas células sanguíneas, quando são comparadas
com outros tecidos do organismo. Os monócitos e os linfócitos CD4+ apresentaram
expressão do IGF1R significativamente maior que linfócitos CD8+ e linfócitos B, sendo
que esses últimos compuseram a subpopulação que menos expressa o IGF1R (KOOIJMAN
et al., 1992). Outras análises também verificaram que o IGF-I foi capaz de modular a ação
de células T in vitro (TAPSON et al., 1988; JOHNSON, JONES, KOZAK, 1992).

104
As células NK (natural killer) são linfócitos envolvidos na vigilância
direcionada a infecções virais e doenças neoplásicas. A atividade citotóxica desse tipo
celular foi estimulada, in vitro, quando no meio de cultura foi adicionado IGF-I o que
demonstrou que os IGF1R são funcionais em células NK (KOOIJMAN et al., 1992).
Posteriormente, a cinética da expressão do IGF1R foi avaliada, mediante técnicas de
citometria de fluxo e RT-PCR, em linfócitos T ativados por um desafio imunológico no
período entre 0 e 48 horas após essa ativação. Nos linfócitos T estimulados, observou-se
um movimento de internalização dos IGF1Rs que se encontravam na membrana celular
alcançando o mínimo de receptores entre 1 e 6 horas após o início da ativação imunológica
associada à concomitante redução do mRNA do IGF1R nesse período. Seguiu-se a esse
fenômeno, um aumento do número de IGF1R na superfície celular acompanhado pelo
aumento no mRNA no citoplasma que ao final da avaliação (48 horas) atingiu patamares
superiores aos encontrados no momento da ativação, fato que levou os autores a concluir
que houve uma síntese de novo do IGF1R após a ativação dos linfócitos T (SEGRETIN et
al., 2003). O uso da fitohemaglutinina no estímulo de linfócitos T humanos in vitro,
demonstrou que a síntese de novo do IGF1R foi observada em linfócitos T CD4+ e CD8+
ativados, e que as células não ativadas não apresentaram esse processamento do IGF1R
(STENTZ; KITABCHI, 2004). Portanto, os linfócitos na condição de células pertencentes
ao sistema imunológico, têm suas funções habitualmente estimuladas quando são expostos
a desafios infecciosos. As crianças que compuseram a amostra ora avaliada apresentaram
diagnósticos clínicos que, na sua totalidade, apresentavam agressões ao organismo de
etiologia infecciosa. Apesar do fato de não ter sido avaliada a resposta imunológica das
crianças participantes, seus linfócitos provavelmente foram estimulados, haja vista os
quadros infecciosos que cursaram com a hipóxia. O aumento na expressão do gene

105
codificador do IGF1R verificado nesses pacientes pode fazer parte da ativação dessas
células, conforme estudos supracitados. Portanto, os resultados do presente estudo apontam
no mesmo sentido, porém a técnica utilizada para avaliar a expressão do IGF1R foi
diferente, além de que a observação foi realizada em crianças in vivo.
A resposta tecidual à hipóxia aguda, no presente estudo, foi analisada
somente em células sanguíneas. Nessas condições, não há dado prévio na literatura que
permita comparação, porém esses resultados, por outro lado, também podem sugerir um
provável mecanismo de compensação mediante a redução das concentrações séricas de
IGF-I e IGFBP-3, e o aumento concomitante das concentrações séricas de IGFBP-1,
sugerindo um fenômeno de up regulation, como descrito em estudo recente (MILANI,
2009).
O conjunto de alterações que foram observadas na presente análise apontou
para uma provável redução das ações biológicas do IGF-I durante o evento hipoxêmico
agudo associado à infecção respiratória, haja vista a redução da concentração sérica desse
fator associada à redução da concentração sérica de IGFBP-3 e ao aumento na concentração
sérica de IGFBP-1. Considerando-se também os resultados obtidos em outros estudos
encontrados na literatura, surge a necessidade de se refletir sobre a real natureza desse
comportamento do sistema IGF. Sob esse aspecto, tais alterações poderiam fazer parte de
um repertório maior de respostas adaptativas, que visam à proteção do indivíduo durante
um episódio de estresse orgânico, e, portanto, provavelmente sem necessidade de
intervenção. Por outro lado, esse ambiente proporcionado por essas oscilações dos
componentes do sistema IGF, durante um evento agudo, poderia ser uma resposta ao estado
hipoxêmico associada a resultados realmente danosos ao crescimento e desenvolvimento.
Nessa eventualidade, pode-se especular a validade da utilização do IGF-I como terapêutica

106
para essas situações na tentativa de minimizar os efeitos dessa resposta do sistema IGF
sobre o desenvolvimento infantil. Entretanto, é importante salientar que estudos anteriores,
nos quais foi utilizado GH em pacientes criticamente enfermos, mostraram resultados que
contra-indicam o seu uso, podendo ser considerado somente em algumas subclasses desses
indivíduos (TAKALA et al., 1999; CARROLL et al., 2004; SCHUETZ et al., 2009).
Concluindo, o presente estudo demonstrou a redução das concentrações
séricas de IGF-I e IGFBP-3, o aumento das concentrações séricas de IGFBP-1 associadas a
concentrações séricas de insulina inalteradas e o aumento da expressão do IGF1R, em
crianças em situação de hipóxia aguda. Os dados apresentados, analisados em conjunto,
sugerem que a hipóxia aguda, provavelmente, associada a fatores infecciosos, foi capaz de
estimular um padrão de alterações que estaria relacionado à redução do estímulo anabólico
provocado pelo IGF-I. Ademais, o aumento na expressão do IGF1R sugere um mecanismo
de proteção tecidual, compensando a redução das concentrações séricas do IGF-I e o
aumento das concentrações séricas da IGFBP-1, não associado a alterações nas
concentrações séricas de insulina. Porém, para que os diversos mecanismos envolvidos
sejam conhecidos, inclusive com melhor controle dos diferentes fatores envolvidos na
resposta do sistema IGF, são necessárias análises mais profundas do comportamento desse
sistema perante estímulos hipoxêmicos isolados em seres humanos.

107
VI - Conclusões

108
VI – Conclusões
1) Crianças com hipóxia aguda associada a infecções respiratórias
apresentaram durante o evento agudo redução das concentrações séricas de IGF-I e IGFBP-
3, aumento das concentrações séricas de IGFBP-1 e da expressão do gene do IGF1R em
linfócitos periféricos;
2) Não houve diferença nas concentrações séricas de insulina entre as
situações de hipóxia aguda associada a infecções respiratórias em crianças, e após sua
recuperação;
3) Não foi possível, nesse modelo experimental, controlar o provável efeito
dos estados infecciosos presentes nos indivíduos participantes;
4) Os dados apresentados, analisados em conjunto, sugerem que a hipóxia
aguda, provavelmente, associada a fatores infecciosos, foi capaz de estimular um padrão de
alterações do sistema IGF, que estaria relacionado à redução do estímulo anabólico
provocado pelo IGF-I e essa redução associada ao aumento na expressão do gene
codificador do IGF1R sugere um mecanismo compensatório de ativação tecidual em
resposta a necessidade do organismo.

109
VII – Referências bibliográficas

110
Abbott AM, Bueno R, Pedrini MT, Murray JM, Smith RJ. Insulin-like
growth factor I receptor gene structure. J Biol Chem (Baltimore) 1992;267(15):10759-63.
Abuzzahab MJ, Schneider A, Goddard A, Grigorescu F, Lautier C, Keller E
et al. IGF-I receptor mutations resulting in intrauterine and postnatal growth retardation. N
Engl J Med (Boston) 2003;349(23):2211-22.
Aguiar-Oliveira MH, Gill MS, de A Barreto ES, Alcântara MR, Miraki-
Moud F, Menezes CA et al. Effect of severe growth hormone (GH) deficiency due to a
mutation in the GH-releasing hormone receptor on insulin-like growth factors (IGFs), IGF-
binding proteins, and ternary complex formation throughout life. J Clin Endocrinol Metab
(Chevy Chase) 1999;84(11):4118-26.
Airaksinen MS, Saarma M. The GDNF family: signaling, biological
functions and therapeutic value. Nat Rev Neurosci (London) 2002;3(5):383-94.
Akaogi K, Sato J, Okabe Y, Sakamoto Y, Yasumitsu H, Miyazaki K.
Synergistic growth stimulation of mouse fibroblasts by tumor-derived adhesion factor with
insulin-like growth factors and insulin. Cell Growth Differ (Philadelphia) 1996;7(12):1671-
7.
Andress DL, Loop SM, Zapf J, Keifer MC. Carboxytruncated insulin-like
growth factor binding protein-5 stimulates mitogenesis in osteoblast cells. Biochem
Biophys Res Commun (San Diego) 1993;195(1):25-30.
Band CJ, Posner BI. Phosphatidylinositol 3‟-kinase and p70s6k are required
for insulin but not bisperoxovanadium 1,10-phenanthroline (bpV-(phen)) inhibition of
insulin-like growth factor binding protein gene expression. Evidence for MEK-independent
activation of mitogen-activated protein kinase by bpV(phen). J Biol Chem (Baltimore)
1997;272(1):138-45.
Baxter RC. Insulin-like growth factor (IGF)-binding proteins: interactions
with IGFs and intrinsic bioactivities. Am J Physiol Endocrinol Metab (Bethesda)
2000;278(6):E967-E976.

111
Baxter RC, Binoux M, Clemmons DR, Conover C, Drop SLS, Holly JMP et
al. Recommendations for nomenclature for the insulin-like growth factor binding protein
(IGFBP) superfamily. J Clin Endocrinol Metab (Chevy Chase) 1998;8(3):273-4.
Baxter RC, Dai J. Purification and characterization of the acid-labile subunit
of rat serum insulin-like growth factor binding protein complex. Endocrinology (Chevy
Chase) 1994;134(2):848-52.
Baxter RC, Martin JL. Structure of the Mr 140,000growth hormone
dependent insulin-like growth factor binding protein complex: determination by
reconstitution and affinity-labeling. Proc Natl Acad Sci USA (Washington)
1989;86(18):6898-902.
Baxter RC, Meka S, Firth SM. Molecular distribution of IGF binding
protein-5 in human serum. J Clin Endocrinol Metab (Chevy Chase) 2002;87(1):271-6.
Beillard E, Pallisgaard N, Van der Velden VHJ, Bi W, Dee R, Van der
Schoot et al. Evaluation of candidate control genes for diagnosis and residual disease
detection in leukemic patients using „real-time‟ quantitative reverse-transcriptase
polymerase chain reaction (RQ-PCR) – a Europe against cancer program. Leukemia
(London) 2003;17(12):2474-86.
Bernabei P, Bosticardo M, Losana G, Regis G, Paola FD, Angelis SD et al.
IGF-I down-regulates IFN-gamma R2 chain surface expression and desensitizes IFN-
gamma/STAT-1 signaling in human T lymphocytes. Blood (Washington)
2003;102(8):2933-9.
Bhatia S, Faessen GH, Carland G, Balise RL, Gargosky SE, Druzin M et al.
A longitudinal analysis of maternal serum insulin-like growth factor I (IGF-I) and total and
nonphosphorylated IGF-binding protein-1 in human pregnancies complicated by
intrauterine growth restriction. J Clin Endocrinol Metab (Chevy Chase) 2002;87(4):1864-
70.
Blum WF, Albertsson-Wikland K, Rosberg S, Rank MB. Serum levels of
insulin-like growth factor I (IGF-I) and IGF binding protein 3 reflect spontaneous growth
hormone secretion. J Clin Endocrinol Metab (Chevy Chase) 1993;76(6):1610-6.

112
Bohula EA, Playford MP, Macaulay VM. Targeting the type 1 insulin-like
growth factor receptor as anti-cancer treatment. Anticancer Drugs (London)
2003;14(9):669-82.
Boisclair Y, Rhoads RP, Ueki I, Wang J, Ooi GT. The acid-labile subunit
(ALS) of the 150 kDa IGF-binding protein complex: an important but forgotten component
of the circulating IGF system. J Endocrinol (Bristol) 2001;170(1):63-70.
Bouhaddioui L, Brun JF, Jacquemin JL, Bailly A, Bouhaddioui N, Kabbaji
K et al. Immunoreactive somatomedin C in children from Morocco: a biological marker of
nutritional growth retardation? Biomed Pharmacother (Paris) 1989;43(1):59-63.
Brismar K, Fernqvist-Forbes E, Wahren J, Hall K. Effect of insulin on the
hepatic production of insulin-like growth factor binding protein-1 (IGFBP-1), IGFBP-3 and
IGF-I in insulin-dependent diabetes. J Clin Endocrinol Metab (Chevy Chase)
1994;79(3):872-8.
Burren CP, Wilson EM, Hwa V, Oh Y, Rosenfeld RG. Binding properties
and distribution of insulin-like growth factor binding protein-related protein 3 (IGFBP-
rP3)/NovH), an additional member of the IGFBP superfamily. J Clin Endocrinol Metab
(Chevy Chase) 1999;84(3):1096-103.
Busby WH Jr, Nam TJ, Moralez A, Smith C, Jennings M, Clemmons DR.
The complement component C1s is the protease that accounts for cleavage of insulin-like
growth factor-binding protein-5 in fibroblast medium. J Biol Chem (Baltimore)
2000;275(48):37638-44.
Bustin SA. Absolute quantification of mRNA using real-time reverse
transcription polymerase chain reaction assays. J Mol Endocrinol (Bristol) 2000;25(2):169-
93.
Byun D, Mohan S, Yoo M, Sexton C, Baylink DJ, Qin X. Pregnancy-
associated plasma protein-A accounts for the insulin-like growth factor (IGF)-binding
protein-4 (IGFBP-4) proteolytic activity in human pregnancy serum and enhances the
mitogenic activity of IGF by degrading IGFBP-4 in vitro. J Clin Endocrinol Metab (Chevy
Chase) 2001;86(2):847-54.

113
Camacho-Hübner C, McCusker RH, Clemmons DR. Secretion and
biological action of insulin-like growth factor binding proteins in two human tumor-derived
cell lines in vitro. J Cell Physiol (New York) 1991;148(2):281-9.
Camacho-Hübner C, Busby WH Jr, McCusker RH, Wright G, Clemmons
DR. Identification of the forms of insulin-like growth factor-binding proteins produced by
human fibroblast and the mechanisms that regulate their secretion. J Biol Chem (Baltimore)
1992;267(17):11949-56.
Carroll PV, Jackson NC, Russell-Jones DL, Treacher DF, Sönksen PH,
Umpleby AM. Combined growth hormone/insulin-like growth factor I in addition to
glutamine-supplemented TNP results in net protein anabolism in critical illness. Am J
Physiol Endocrinol Metab (Bethesda) 2004;286(1):E151-7.
Centers for Disease Control and Prevention. Growth charts for the United
States: methods and development. Atlanta: Series report 11(246);2000.201p.
Clayton PE, Gill MS. Normal growth and its endocrine control. In: Brook
CGD Hindmarsh PC. Clinical Paediatric Endocrinology. 4a. ed. Oxford: Blackwell Science
Ltd; 2001. p. 95-114.
Clemmons DR. Use of mutagenesis to probe IGF-binding proteins
structure/function relationships. Endocr Rev (Chevy Chase) 2001;22(6):800-17.
Clemmons DR, Snyder DK, Busby WH Jr. Variables controlling the
secretion of insulin-like growth factor binding protein-2 in normal humans subjects. J Clin
Endocrinol Metab (Chevy Chase) 1991;73(4):727-33.
Cohick WS, Clemmons DR. The insulin-like growth factors. Annu Rev
Physiol (Palo Alto) 1993;55:131-53.
Conover CA, Divertie GD, Lee PDK. Cortisol increases plasma insulin-like
growth factor binding protein-1 in humans. Acta Endocrinol (Copenhagen)
1993;128(2):140-3.
Conover CA, Faessen GF, Ilg KE, Chandrasekher YA, Christiansen M,
Overgaard MT et al. Pregnancy-associated plasma protein-a is the insulin-like growth
factor binding protein-4 protease secreted by human ovarian granulosa cells and is a marker

114
of dominant follicle selection and the corpus luteum. Endocrinology (Chevy Chase)
2001;142(5):2155-8.
Conover CA, Kiefer MC. Regulation and biological effect of endogenous
insulin-like growth factor binding proteins-5 in human osteoblastic cells. J Clin Endocrinol
Metab (Chevy Chase) 1993;76(5):1153-9.
Conover CA, Powell DR. Insulin-like growth factor (IGF)-binding protein-3
blocks IGF-induced receptor down-regulation and cell desensitization in cultured bovine
fibroblasts. Endocrinology (Chevy Chase) 1991;129(2):710-6.
Coverley JA, Baxter RC. Phosphorylation of insulin-like growth factor
binding proteins. Mol Cell Endocrinol (Limerick) 1997;128(1-2):1-5.
Coterril AM, Cowell CT, Baxter RC, McNeil D, Silinik M. Regulation of the
growth hormone-independent growth factor-binding protein in children. J Clin Endocrinol
Metab (Chevy Chase) 1988;67(5):882-7.
Cotterill AM, Holly JMP, Taylor AM, Davies SC, Coulson VJ, Preece MA
et al. The insulin-like growth factor (IGF)-binding proteins and IGF bioactivity in Laron-
type dwarfism. J Clin Endocrinol Metab (Chevy Chase) 1992;74(1):56-63.
Daughaday WH. Growth hormone, insulin-like growth factors and
acromegaly. In: De Groot LJ. Endocrinology. 3a. ed. Philadelphia: Saunders; 1995. p. 303-
29.
De Groof F, Joosten KFM, Janssen, JAMJL, De Kleijn ED, Hazelset JA,
Hop WCJ et al. Acute stress response in children with meningococcal sepsis: important
differences in the growth hormone/insulin-like growth factor I axis between nonsurvivors
and survivors. J Clin Endocrinol Metab (Chevy Chase) 2002;87(7):3118-24.
Dinleyici EC, Kilic Z, Buyukkaragoz B, Ucar B, Alatas O, Aydogdu SD et
al. Serum IGF-I, IGFBP-3 and growth hormone levels in children with congenital heart
disease: relationship with nutritional status, cyanosis and left ventricular functions. Neuro
Endocrinol Lett (Stockholm) 2007;28(3):279-83.

115
Dinleyici EC, Tekin N, Colak O, Aksit MA. Cord blood IGF-I and IGFBP-3
levels in asphyxiated term newborns. Neuro Endocrinol Lett (Stockholm) 2006;27(6):745-
7.
Dündar B, Akçoral A, Saylam G, Unal N, Mese T, Hüdaoğlu S et al.
Chronic hypoxemia leads to reduced serum IGF-I levels in cyanotic congenital heart
disease. J Pediatr Endocrinol Metab (London) 2000;13(4):431-6.
Dupont J, Dunn SE, Barrett C, LeRoith D. Microarray analysis and
identification of novel molecules involved in insulin-like growth factor-1 receptor signaling
and gene expression. Recent Prog Horm Res (New York) 2003;58:325-42.
Forbes BE, Turner D, Hodge SJ, McNeil KA, Forsberg G, Wallace JC.
Localization of an insulin-like growth factor (IGF) binding site of bovine IGF binding
protein-2 using disulfide mapping and deletion mutation analysis of the C-terminal domain.
J Biol Chem (Baltimore) 1998;273(8):4647-52.
Froesch ER, Bürgi H, Ramseier EB, Bally P, Labhart A. Antibody-
suppressible and nonsuppressible insulin-like activities in human serum and their
physiologic significance. An insulin assay with adipose tissue of increased precision and
specificity. J Clin Invest (Ann Arbor) 1963;42(11):1816-34.
Garrett TPJ, McKern NM, Lou M, Frenkel MJ, Bentley JD, Lovrecz GO et
al. Crystal structure of the first three domains of the type-1 insulin-like growth factor
receptor. Nature (London) 1998;394(6691):395-9.
Giudice LC, Conover CA, Bale L, Faessen GH, Ilg K, Sun I et al.
Identification and regulation of the IGFBP-4 protease and its physiological inhibitor in
human trophoblasts and endometrial stroma: evidence for paracrine regulation of IGF-II
bioavailability in the placental bed during human implantation. J Clin Endocrinol Metab
(Chevy Chase) 2002;87(5):2359-66.
Giudice LC, Zegher F, Gargosky SE, Dsupin BA, de las Fuentes L, Cristal
RA, et al. Insulin-like growth factors and their binding proteins in the term and preterm
human fetus and neonate with normal and extremes of intrauterine growth. J Clin
Endocrinol Metab (Chevy Chase) 1995;80(5):1548-55.

116
Greem LR, Kawagoe Y, Hill DJ, Richardson BS, Han VKM. The effect of
intermittent umbilical cord occlusion on insulin-like growth factors and their binding
proteins in preterm and near-term ovine fetuses. J Endocrinol (Bristol) 2000;166(3):565-77.
Hall K. Human somatomedin. Determination, occurrence, biological activity
and purification. Acta Endocrinol Suppl (Copenhagen) 1972;163:1052.
Hilding A, Brismar K, Thorén M, Hall K. Glucagon stimulates insulin-like
growth factor binding protein-1 secretion in healthy subjects, patients with pituitary
insufficiency, and patients with insulin-dependent diabetes mellitus. J Clin Endocrinol
Metab (Chevy Chase) 1993;77(5):1142-7.
Holly JMP, Dunger DB, Edge JA, Smith CP, Chard T, Wass JAH. Insulin-
like growth factor binding protein-1 levels in diabetic adolescents and their relationship to
metabolic control. Diabet Med (Oxford) 1990;7(7):618-23.
Hossemlopp P, Seurin D, Segovia-Quinson B, Hardouin S, Binoux M.
Analysis of serum insulin-like growth factor binding proteins using western blotting: use of
method for titration of the binding proteins and competitive binding studies. Anal Biochem
(Orlando) 1986;154(1):138–43.
Hwa V, Oh Y, Rosenfeld RG. The insulin-like growth factor binding protein
(IGFBP) superfamily. Endocr Rev (Chevy Chase) 1999;20(6):761-87.
Hwa V, Haeusler G, Pratt KL, Little BM, Frisch H, Koller D et al. Total
absence of functional acid labile subunit resulting in severe insulin-like growth factor
deficiency and moderate growth failure. J Clin Endocrinol Metab (Chevy Chase)
2006;91(5):1826-31.
Johnson EW, Jones LA, Kozak RW. Expression and function of insulin-like
growth factor receptors on anti-CD3-activated human T lymphocytes. J Immunol
(Baltimore) 1992;148(1):63-71.
Jones JI, Busby WH, Wrigh G, Smith CE, Kimack NM, Clemmons DR.
Identification of the sites of phosphorylation in insulin-like growth factor binding protein-1:
regulation of its affinity by phosphorylation of serine 101. J Biol Chem (Baltimore)
1993c;268(2):1125-31.

117
Jones JI, Clemmons DR. Insulin-like growth factors and their binding
proteins: biological actions. Endocr Rev (Chevy Chase) 1995;16(1):3-34.
Jones JI, D‟Ercole AJ, Camacho-Hubner C, Clemmons DR. Phosphorylation
of insulin-like growth factor (IGF)-binding protein 1 in cell culture and in vivo: effects on
affinity for IGF-I. Proc Natl Acad Sci USA (Washington) 1991;88(17):7481-5.
Jones JI, Gockerman A, Busby WH Jr, Camacho-Hübner C, Clemmons DR.
Extracellular matrix contains insulin-like growth factor binding protein-5: potentiation of
the effects of IGF-I. J Cell Biol (New York) 1993a;121(3):679-87.
Jones JI, Gockerman A, Busby WH Jr, Wright G, Clemmons DR. Insulin-
like growth factor binding protein 1 stimulates cell migration and binds to the alpha 5 beta
1 integrin by means of its Arg-Gly-Asp sequence. Proc Natl Acad Sci USA (Washington)
1993b;90(22):10553-7.
Juul A. Serum levels of insulin-like growth factor I and its binding proteins
in health and disease. Growth Horm IGF Res (London) 2003;13(4):113-70.
Kajimura S, Aida K, Duan C. Understanding hypoxia-induced gene
expression in early development: in vitro and in vivo analysis of hypoxia-inducible factor 1-
regulated Zebra fish insulin-like growth factor binding protein 1 gene expression. Mol Cell
Biol (Washington) 2006;26(3):1142-55.
Kawashima Y, Kanzaki S, Yang F, Kinoshita T, Hanaki K, Nagaishi J et al.
Mutation at cleavage site of insulin-like growth factor receptor in a short-stature child born
with intrauterine growth retardation. J Clin Endocrinol Metab (Chevy Chase)
2005;90(8):4679-87.
Keegan AD, Nelms K, Wang LM, Pierce JH, Paul WE. Interleukin 4
receptor: signaling mechanisms. Immunol Today (Amsterda) 1994;15(9):423-32.
Kelley KM, Oh Y, Gargosky SE, Gucev Z, Matsumoto T, Hwa V et al.
Insulin-like growth factor-binding proteins (IGFBPs) and their regulatory dynamics. Int J
Biochem Cell Biol (Tarrytown) 1996; 28(6):619-37.

118
Kelly KR, Williamson DL, Fealy CE, Kriz DA, Krishnan RK, Huang H et
al. Acute altitude-induced hypoxia suppresses plasma glucose and leptin in healthy humans.
Metabolism (Philadelphia) 2009;(ahead of print).
Kooijman R, Willems M, De Haas CJC, Rijkers GT, Schuurmans ALG, Van
Buul-Offers S et al. Expression of type I insulin-like growth factor receptors on human
peripheral blood mononuclear cells. Endocrinology (Chevy Chase) 1992,131(5):2244-50.
Koga T, Endo H, Miyamoto Y, Mukai M, Akira S, Inoue M. IGFBPs
contribute to survival of pancreatic cancer cells under severely hypoxic conditions. Cancer
Lett (Limerick) 2008;268(1):82-8..
Langford K, Nicolaides K, Miell JP. Maternal and fetal insulin-like growth
factors and their binding proteins in the second and third trimesters of human pregnancy.
Hum Reprod (Oxford) 1998;13(5):1389-93.
Latour D, Mohan S, Linkhart TA, Baylink DJ, Strong DD. Inhibitory
insulin-like growth factor-binding protein: cloning complete sequence and physiological
regulation. Mol Endocrinol (Baltimore) 1990;4(12):1806-14.
Lawrence JB, Oxvig C, Overgaard MT, Sottrup-Jensen L, Gleich GJ Hays
LG et al. The insulin-like growth factor (IGF)-dependent IGF binding protein-4 protease
secreted by human fibroblasts is pregnancy-associated plasma protein-A. Proc Natl Acad
Sci U S A (Washington) 1999;96(6):3149-53.
Lee PDK, Giudice LC, Conover CA, Powell DR. Insulin-like growth factor
binding protein-1: recent findings and new directions. Proc Soc Exp Biol Med (Malden)
1997;216(3):319-57.
LeRoith D, Werner H, Beitner-Johnson D, Roberts CT Jr. Molecular and
cellular aspects of the insulin-like growth factor I receptor. Endocr Rev (Chevy Chase)
1995;16(2):143-63.
Liu JL, Grinberg A, Westphal H, Sauer B, Accili D, Karas M, et al. Insulin-
like growth factor-I affects perinatal lethality and postnatal development in a gene dosage-
dependent manner: manipulation using the Cre/loxP system in transgenic mice. Mol
Endocrinol (Baltimore) 1998;12(9):1452-62.

119
Malthiéry Y, Lissitzky S. Primary structure of human thuroglobulin deduced
from the sequence of its 8448-base complementary DNA. Eur J Biochem (Oxford)
1987;165(3):491-8.
Mauras N, Beaufrere B. Recombinant human insulin-like growth factor-I
enhances whole body protein anabolism and significantly diminishes the protein catabolic
effects of prednisone in humans without a diabetogenic effect. J Clin Endocrinol Metab
(Chevy Chase) 1995;80(3):869-74.
Maures TJ, Duan C. Structure, developmental expression, and physiological
regulation of zebrafish IGF binding protein-1. Endocrinology (Chevy Chase)
2002;143(7):2722-31.
McLellan KC, Hooper SB, Bocking AD, Delhanty PJD, Phillips ID, Hill DJ,
et al. Prolonged hypoxia induced by the reduction of maternal uterine blood flow alters
insulin-like growth factor-binding protein-1 (IGFBP-1) and IGFBP-2 gene expression in
the ovine fetus. Endocrinology (Chevy Chase) 1992;131(4):1619-28.
Miele C, Rochford JJ, Fillippa N, Giorgetti-Peraldi S, Van Obberghen E.
Insulin and insuli-like growth factor-I induce vascular endothelial growth factor mRNA
expression via different signaling pathways. J Biol Chem (Baltimore) 2000;275(28)21695-
702.
Milani, SLS. Caracterização clínica e laboratorial de crianças e adolescents
com insensibilidade parcial ao hormônio de crescimento ou ao IGF-I [dissertação]. Ribeirão
Preto: Universidade de São Paulo, Faculdade de Medicina de Ribeirão Preto; 2009.
Mohan S, Baylink DJ. IGF system components and their role in bone
metabolism. In: Rosenfeld RG, Roberts CT. IGFs in Health and Disease. New Jersey:
Human Press; 1999. p. 457-96.
Mohan S, Baylink DJ. IGF-binding proteins are multifunctional and act via
IGF-dependent and independent mechanisms. J Endocrinol (Bristol) 2002;175(1):19-31.
Mohan S, Strong DD, Lempert UG, Tremollieres F, Wergedal JE, Baylink
DJ. Studies on regulation of insulin-like growth factor binding protein (IGFBP)-3 and
IGFBP-4 production in human bone cells. Acta Endocrinol (Copenhagen)
1992;127(6):555-64.

120
Morgan BLG, Chao CR. The effects of hypoxia on growth cones in the
ovine fetal brain. J Matern Fetal Neonatal Med (Boca Raton) 2004;16(1):55-9.
Mounier C, Dumas V, Posner BI. Regulation of hepatic insulin-like growth
factor-binding protein-1 gene expression by insulin: central role for mammalian target of
rapamycin independent of forkhead box O proteins. Endocrinology (Chevy Chase)
2006;147(5):2383-91.
Murray IA, Perdew GH. Omeprazole stimulates the induction of human
insulin-like growth factor binding protein-1 through aryl hydrocarbon receptor activation. J
Pharmacol Exp Ther (Bethesda) 2008;324(3):1102-10.
Neely EK, Beukers MW, Oh Y, Cohen P, Rosenfield RG. Insulin-like
growth factors receptors. Acta Paediatr Scand (Stockholm) 1991;(supp372):116-23.
Neumann GM, Marinaro JA, Bach LA. Identification of O-glycosylation
sites and partial characterization of carbohydrate structure and disulfide linkages of human
insulin-like growth factor binding protein 6. Biochem J (London) 1998;37(18):6572-85.
Nielsen FC. The molecular and cellular biology of insulin-like growth factor
II. Prog Growth Factor Res (Oxford) 1992;4(3):257-90.
O‟Connor R, Kauffmann-Zeh A, Liu YM, Lehar S, Evan GI, Baserga R et
al. Identification of domains fo the insulin-like growth factor I receptor that are required for
protection from apoptosis. Mol Cell Biol (Washington) 1997;17(1):427-35.
Oh Y, Muller HL, Lamson G, Rosenfeld RG. Insulin-like growth factor
(IGF)-independent action of IGF-binding protein-3 in Hs578T human breast cancer cells.
Cell surface binding and growth inhibition. J Biol Chem (Baltimore) 1993;268(20):14964-
71.
Ohh M, Park CW, Ivan M, Hoffman MA, Kim TY, Huang LE et al.
Ubiquitination of hypoxia inducible factor requires direct binding to the beta-domain of the
von Hippel-Lindau protein. Nat Cell Biol (London) 2000;2(7):423-7.
Olson EN, Klein WH. bHLH factors in muscle development: dead lines and
commitments, what to leave in and what to leave out. Genes Dev (Cold Spring Harbor)
1994;8(1):1-8.

121
Patel S, Lochhead PA, Rena G, Fumagalli S, Pende M, Kozma SC et al.
Insulin regulation of insulin-like growth factor-binding protein-1 gene expression is
dependent on the mammalian target of rapamycin, but independent of ribosomal S6 kinase
activity. J Biol Chem (Baltimore) 2002;277(12):9889-95.
Peretz S, Kim C, Rockwell S, Baserga R, Glazer PM. IGF1 receptor
expression protects against microenvironmental stress found in the solid tumor. Radiat Res
(Charlottesville) 2002;158(2):174-80.
Popovici RM, Lu M, Bhatia S, Faessen GH, Giaccia AJ, Giudice LC.
Hypoxia regulates insulin-like growth factor-binding protein 1 in human fetal hepatocytes
in primary culture: suggestive molecular mechanisms for in utero fetal growth restriction
caused by uteroplacental insufficiency. J Clin Endocrinol Metab (Chevy Chase)
2001;86(6):2653-9.
Povoa G, Enberg G, Jornvall H, Hall K. Isolation and characterization of
somatomedin-binding protein from midterm human amniotic fluid. Eur J Biochem (Oxford)
1984;144(2):199-204.
Powell-Braxton L, Hollingshead P, Giltinan D, Pitts-Meek S, Stewart T.
Inactivation of the IGF-I gene in mice results in perinatal lethality. Ann N Y Acad Sci
(New York) 1993;692:300-1.
Qin X, Byun D, Lau KW, Baylink DJ, Mohan S. Evidence that the
interaction between insulin-like growth factor (IGF)-II and IGF binding protein (IGFBP)-4
is essential for the action of the IGF-II dependent IGFBP-4 protease. Arch Biochem
Biophys (New York) 2000;379(2):209-16.
Raile K, Klammt A, Schneider A, Keller S, Laue R, Smith R et al. Clinical
and functional characteristics of the human Arg59Ter insulin-like growth factor I receptor
(IGF1R) mutation: implications for a gene dosage effect of the human IGF1R. J Clin
Endocrinol Metab (Chevy Chase) 2006;91(6):2264-71.
Rechler MM. Insulin-like growth factor binding proteins. Vitam Horm (New
York) 1993;47:1-114.

122
Rechler MM, Nissley SP. The nature and regulation of the receptor for
insulin-like growth factors. Annu Rev Physiol (Palo Alto) 1985;47:425-42.
Resnicoff M, Burgaud JL, Rotman HL, Abraham D, Baserga R. Correlation
between apoptosis, tumorigenesis, and levels of insulin-like growth factor I receptors.
Cancer (Hoboken) 1995;55(17):3739-41.
Roghani M, Lassarre C, Zapf J, Póvoa G, Binoux M. Two insulin-like
growth factor (IGF)-binding proteins are responsible for the selective affinity for IGF-II of
cerebrospinal fluid binding proteins. J Clin Endocrinol Metab (Chevy Chase)
1991;73(3):658-66.
Roig MD, Sabrià J, Valls C, Borràs M, Miró E, Ponce J et al. The use of
biochemical markers in prenatal diagnosis of intrauterine growth retardation: insulin-like
growth factor I, Leptin, and alpha-fetoprotein. Eur J Obstet Gynecol Reprod Biol
(Limerick) 2005;120(1):27-32.
Rosenfeld RG, Pham H, Cohen P, Fielder P, Gargosky SE, Muller H et al.
Insulin-like growth factor binding proteins and their regulation. Acta Paediatrica
(Stockholm) 1994;(suppl 399):154-8.
Rudman D, Kutner MH, Rogers CM, Lubin MF, Fleming GA, Bain RP.
Impaired growth hormone secretion in the adult population: relation to age and adiposity. J
Clin Invest (Ann Arbor) 1981;67(5):1361-7.
Sara VR, Hall K. Insulin-like growth factors and their binding proteins.
Physiol Rev (Bethesda) 1990;70(3):591-614.
Salmon WD Jr., Daughday WH. A hormonally controlled serum factor which
stimulates sulfate incorporation by cartilage in vitro. J Lab Clin Med (Saint Louis)
1957;49(6), 825-36.
Schofield PN. Molecular biology of the insulin-like growth factor gene
structure and expression. Acta Paediatr Scand (Stockholm) 1991;(suppl 372):83-90.
Schuetz P, Muller B, Nusbaumer C, Wieland M, Crist-Crain M. Circulating
levels of GH predict mortality and complement prognostic scores in critically ill medical
patients. Eur J Endocrinol (Oslo) 2009;160:157-63.

123
Scrideli CA, Tone LG. Técnicas de biologia molecular em doenças onco-
hematológicas da infância. In: Braga, JAP; Loggetto, SR; Tone, LG. Hematologia para o
Pediatra. São Paulo:Ateneu; 2007. p. 457-71.
Segretin ME, Galeano A, Roldán A, Schillaci R. Insulin-like growth factor-1
receptor regulation in activated human T lymphocytes. Horm Res (New York)
2003;59(6):276-80.
Smith WJ, Underwood LE, Clemmons DR. Effects of caloric or protein
restriction on insulin-like growth factor-I (IGF-I) and IGF-binding proteins in children and
adults. J Clin Endocrinol Metab (Chevy Chase) 1995;80(2):443-9.
Soos MA, Siddle K. Immunological relationships between receptors for
insulin and insulin-like growth factor I: evidence for structural heterogeneity of insulin-like
growth factor-I receptors involving hybrids with insulin receptors. Biochem J (London)
1989;263:553-63.
Soos MA, Field CE, Siddle K. Purified hybrid insulin/insulin-like growth
factor-I receptors bind insulin-like growth factor-I, but not insulin with high affinity.
Biochem J (London) 1993;290(Pt2):419-26.
Stentz FB, Kitabchi AE. De novo emergence of growth factor receptors in
activated human CD4+ and CD8+ lymphocytes. Metabolism (Philadelphia)
2004;53(1):117-22.
Sussenbach JS, Steenbergh PH, Jansen E, Meinsma D, Van Dijk MA,
Holthuizen P et al. Structure and post-transcriptional regulation of expression of the human
IGF-I and –II genes. In: Spencer EM. Modern concepts of Insulin-like Growth Factors.
New York: Elsevier; 1991. p. 639-54.
Takala J, Ruolonen E, Webster NR, Nielsen MS, Zandstra DF, Vundelinckx
G et al. Increased mortality associated with growth hormone treatment in critically ill
adults. N Engl J Med (Massachusetts) 1999;341(11):785-92.
Tapanainen PJ, Bang P, Wilson K, Unterman TG, Vreman HJ, Rosenfeld
RG. Maternal hypoxia as a model for intrauterine growth retardation: effects on insulin-like
growth factors and their binding proteins. Pediatr Res (Hagerstown) 1994;36(2):152-8.

124
Tapson VF, Boni-Schnetzler M, Pilch PF, Center DM, Berman JS. Structural
and functional characterization of the human T lymphocyte receptor for insulin-like growth
factor I in vitro. J Clin Invest (Ann Arbor) 1988;82(3):950-7.
Tasker RC, Roe MFE, Bloxham DM, White DK, Ross-Russell RI,
O‟Donnell. The neuroendocrine stress response and severity of acute respiratory syncytial
virus bronchiolitis in infancy. Intensive Care Med (New York) 2004;30(12):2257-62.
Tazuke SI, Mazure NM, Sugawara J, Carland G, Faessen GH, Suen LF et al.
Hypoxia stimulates inulin-like growth factor binding protein 1 (IGFBP-1) gene expression
in HepG2 cells: A possible model for IGFBP-1 expression in fetal hypoxia. Proc Natl Acad
Sci USA (Washington) 1998;95(17):10188-93.
Thissen JP, Ketelslegers JM, Underwood LE. Nutritional regulation of the
insulin-like growth factors. Endocr Rev (Chevy Chase) 1994;15(1):80-101.
Treins C, Giorgette-Peraldi S, Murdaca J, Monthouel-Kartmann MN, Van
Obberghen E. Regulation of hypoxia-inducible factor (HIF)-1 activity and expression of
HIF hydroxylases in response to insulin-like growth factor I. Mol Endocrinol (Baltimore)
2005;19(5):1304-17.
Twigg SM, Kiefer MC, Zapf J, Baxter RC. Insulin-like growth factor-
binding protein 5 complexes with the acid-labile subunit. Role of the carboxyl-terminal
domain. J Biol Chem (Baltimore) 1998;273(44):28791-8.
Ullrich A, Gray A, Tam AW, Yang-Feng T, Tsubokawa M, Collins C et al.
Insulin-like growth factor I receptor primary structure: comparison with insulin receptor
suggests structural determinants that define functional specificity. EMBO J (Oxford)
1986;5(10):2503-12.
Villaudy J, Delbe J, Blat C, Desauty G, Golde A, Harel L. An IGF binding
protein is an inhibitor of FGF stimulation. J Cell Physiol (New York) 1991;149(3):492-6.
Vincent AM, Feldman EL. Control of cell survival by IGF signaling
pathways. Growth Horm IGF Res (Edinburgh) 2002;12(4):193-7.

125
Wang GL, Semenza GL. Purification and characterization of hypoxia-
inducible factor 1. J Biol Chem (Baltimore) 1995;270(3):1230-7.
Woods KA, Camacho-Hubner C, Savage MO, Clark AJ. Intrauterine growth
retardation and postnatal growth failure associated with deletion of the insulin-like growth
factor I gene. N Engl J Med (Boston) 1996;335(18):1363-7.
World Health Organization. Physical status: the use and interpretation of
anthropometry. Geneva: WHO (Technical Report Series, WHO/854); 1995.452p.
Zapf J. Physiological role of the insulin-like growth factor binding proteins.
Eur J Endocrinol (Bristol) 1995;132(6):645-54.
Zapf J, Born W, Chang JY, James P, Froesch ER, Fischer JA. Isolation and
NH2-terminal amino acid sequences of rat serum carrier proteins for insulin-like growth
factors. Biochem Biophys Res Commun (San Diego) 1988;156(3):1187-94.
Zapf J, Rinderknecht E, Humbel RE, Froesch ER. Nonsuppressible insulin-
like activity (NSILA) from human serum: recent accomplishments and their physiologic
implications. Metabolism (Philadelphia) 1978;27(12):1803-28.

126
VIII – Apêndices e anexos

127
Apêndice A - Protocolos utilizados no Laboratório de Oncologia Pediátrica do
HCFMRPUSP
1 – Preparação do tampão PBS (Phosphate-buffered saline)
Dissolver 8 g de NaCl, 0,2 g de KCl, 1,44 g de Na2HPO4 em 800 ml de água
destilada.
Ajustar o pH para 7,4 com HCl.
Acrescentar água quantidade suficiente para 1 litro.
Aliquotar a solução e esterilizar em autoclave por 20 minutos.
Armazenar em geladeira.
2 – Tratamento da água com DEPC
Acrescentar DEPC (diethyl pyrocarbonate) ao volume de água desejado em
concentração final de 0,1%.
Deixar a solução overnight e, no dia seguinte, esterilizar em autoclave por 20
minutos.
3 – Preparação da solução tampão TBE (10 X)

128
Dissolver 108g de Tris;
55 g de ácido bórico;
40 ml de EDTA 0,5 M pH 8,0.
Acrescentar água suficiente para 1 litro.
4 – Preparação do gel de agarose a 1% para RNA
Lavar a cuba e pentes a serem utilizados com solução contendo removedores
de RNase.
Enxaguar com água destilada.
Acrescentar 1 g de agarose a 100 ml de solução tampão (TBE) preparada
com água tratada com DEPC e aquecer até a dissolução completa da agarose.
Acrescentar 3 µl de brometo de etídio em concentração de 10 mg/ml.
Verter gentilmente o gel para polimerização.
5 – Preparação do gel de agarose a 2%
Acrescentar 2 g de agarose a 100 ml de solução tampão (TBE) e aquecer até
a dissolução completa da agarose.
Acrescentar 3 µl de brometo de etídio em concentração de 10 mg/ml.
Verter gentilmente o gel para polimerização.

129
Anexo A - Parecer do Comitê de Ética em Pesquisa do HCFMRP-USP

130
Artigo para publicação

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo