role of insulin-like growth factor binding protein-3 (igfbp-3) in the differentiation of primary...
TRANSCRIPT

JOURNAL OF CELLULAR PHYSIOLOGY 195:70–79 (2003)
Role of Insulin-Like Growth Factor Binding Protein-3(IGFBP-3) in the Differentiation of Primary
Human Adult Skeletal Myoblasts
EMILY J. FOULSTONE,* PAUL B. SAVAGE, ANNA L. CROWN, JEFF M.P. HOLLY, AND
CLAIRE E.H. STEWART
Division of Surgery, University of Bristol, Bristol Royal Infirmary, Bristol, England
Although muscle satellite cells were identified almost 40 years ago, little is knownabout the induction of their proliferation and differentiation in response to physio-logical/pathological stimuli or to growth factors/cytokines. In order to investigatethe role of the insulin-like growth factor (IGF)/IGF binding protein (IGFBP) system inadult humanmyoblast differentiationwe have developed a primary human skeletalmuscle cell model. We show that under low serum media (LSM) differentiatingconditions, the cells secrete IGF binding proteins-2, -3, -4 and -5. Intact IGFBP-5was detected at days 1 and 2 but by day 7 in LSM it was removed by proteolysis.IGFBP-4 levels were also decreased at day 7 in the presence of IGF-I, potentially byproteolysis. In contrast, we observed that IGFBP-3 initially decreased on transfer ofcells into LSMbut then increasedwithmyotube formation. Treatmentwith 20ng/mltumour necrosis factor-alpha (TNFa), which inhibits myoblast differentiation,blocked IGFBP-3 production and secretion whereas 30 ng/ml IGF-I, whichstimulatesmyoblast differentiation, increased IGFBP-3 secretion. The TNFa-induc-ed decrease in IGFBP-3 production and inhibition of differentiation could not berescuedby addition of IGF-I. LongR3IGF-I,which does not bind to the IGFBPs, had asimilar effect on differentiation and IGFBP-3 secretion as IGF-I, both with andwithout TNFa, confirming that increased IGFBP-3 is not purely due to increasedstability conferred by binding to IGF-I. Furthermore reduction of IGFBP-3 secretionusing antisense oligonucleotides led to an inhibition of differentiation. Taken to-gether these data indicate that IGFBP-3 supports myoblast differentiation. J. Cell.Physiol. 195: 70–79, 2003. � 2003 Wiley-Liss, Inc.
Muscle degeneration is recognised as a significantcause of patient disability and death underlying manychronic catabolic illnesses (Calman, 1982), but as yet noeffective therapeutic interventions are available totackle this problem. The normal rate of protein turnoveris high (250–300 g/day) so any small but persistentchange in catabolism/anabolism could culminate insignificant muscle loss. Muscle myofibres are terminallydifferentiated and incapable of replication, thereforeregeneration is dependent on a small population of re-sident cells generally referred to as satellite cells (Olson,1992; Olson and Rosenthal, 1994). In order to elucidatepotential therapeutic targets for muscle wasting, it isessential to understand how these muscle satellite cellsproliferate and differentiate. A thorough understandingof human myoblast growth and differentiation wouldalso have potential relevance to skeletal muscle stemcell therapies, allowing manipulation of the system fortherapeutic effect.
Although many studies have been carried out on thegrowth and differentiation of muscle cells, the majorityof in vitro studies use immortalised rat and mousemuscle cell lines. These studies have highlighted thecritical role of the insulin-like growth factors IGF-Iand IGF-II and their binding proteins in the con-trol of proliferation, differentiation and metabolism
(Florini et al., 1991; Stewart and Rotwein, 1996). Todate, six high affinity IGF binding proteins (IGFBPs)have been identified (Jones and Clemmons, 1995). Theseproteins modulate the activity of the IGFs by extendingtheir half-life and by either potentiating or inhibitingbinding of the IGFs to their receptors (Conover et al.,1990, 1993). Importantly, however, the muscle cell linesused to date differ in which IGF binding proteins theyproduce (James et al., 1993; McCusker and Clemmons,1998; Crown et al., 2000) and are not identical in theirresponses to IGFs (Florini et al., 1991). We havetherefore developed a more physiologically relevant,primary adult human muscle cell model in which toaddress our questions (Crown et al., 2000).
� 2003 WILEY-LISS, INC.
Contract grant sponsor: AICR; Contract grant number: 99-134.
*Correspondence to: Emily J. Foulstone, Division of Surgery,University of Bristol, Bristol Royal Infirmary, Bristol, BS2 8HW,England. E-mail: [email protected]
Received 6 August 2002; Accepted 5 November 2002
DOI: 10.1002/jcp.10227

One major difference between these cultures and thesecondary cell lines that have been studied to date is thatthe adult human muscle cell cultures produce andsecrete large amounts of IGFBP-3 (Crown et al., 2000).Although IGFBP-3 is the major IGFBP found in serum,it is also produced locally in many tissues (Kim et al.,1997). Its activity is modulated not only by its rate ofsynthesis but also by post-translational modificationsand by proteolysis (Collett-Solberg and Cohen, 1996).Recently IGFBP-3 has been shown to have effects oncell growth and apoptosis that are independent of IGFaction (Gill et al., 1997; Fowler et al., 2000; Hollowoodet al., 2000) and high levels of IGFBP-3 decrease theproliferation of chondrocytes (Spagnoli et al., 2001). Inaddition, IGFBP-3 has also been implicated in porcinemuscle cell differentiation (Johnson et al., 1999).Interestingly, IGFBP-5, which we have shown inhibitsdifferentiation and acts as a potent survival factor formurine C2 myoblasts (James et al., 1996; Meadowset al., 2000; Foulstone et al., 2001) is closer to IGFBP-3in sequence than any other IGFBP suggesting poten-tial for overlapping/related actions. Data derived fromother cell models would therefore suggest a potentialand important role for IGFBP-3 in skeletal musclehomeostasis with manipulation of IGFBP-3 possiblyacting as an important tool in influencing skeletalmuscle maintenance and repair in vivo. We thereforesought to investigate what role IGFBP-3 may play inprimary skeletal muscle differentiation, proliferationand survival.
In addition to the IGFs, other growth factors andcytokines can influence muscle growth and mainte-nance (Hawke and Garry, 2001). Production of theinflammatory cytokine tumour necrosis factor a (TNFa)occurs as an important pathophysiological responseduring many critical illnesses and TNFa has beenimplicated in the aetiology of muscle wasting (Matthysand Billiau, 1997). We have shown that TNFa blocks C2myoblast differentiation and decreases IGFBP-5 secre-tion while at the same time increasing proliferation andinducing cell death (Meadows et al., 2000; Foulstoneet al., 2001). We therefore investigated whether TNFahad similar effects in the primary human skeletalmuscle cells with regard to differentiation, death andimpact on the IGFBPs. In light of their importancein differentiation, growth and survival in many cellsystems, including muscle, we also investigated the roleof the IGFs, alone or in combination with TNFa, ondifferentiation and IGFBP-3 secretion in the primarycells. These studies identify the IGFBPs secreted byhuman primary myoblasts during differentiation, theimpact of physiologically important growth factor/cytokine family members on their regulation and im-plicate IGFBP-3 as a potent and positive differentiationfactor.
MATERIALS AND METHODSMaterials
Unless otherwise stated all chemicals were fromSigma (Poole, UK). Ham’s F-10 media was from BioWhittaker (Wokingham, UK) and heat inactivatedfoetal bovine serum (hiFBS) was from GIBCO (Paisley,UK). TNFa was purchased from Bachem (St. HelensMerseyside, UK). Recombinant human IGF-I and Long-
R3IGF-I were from GroPep (Adelaide, Australia).Recombinant human non-glycosylated IGFBP-3 wasfrom Dr. C. Moot (Brogrowth, CA). Trypsin and gelatinwere from Difco (Oxford, UK). IGFBP-3 anti-sense(CATGACGCCTGCAACCGGGG) and sense (CCCC-GGTTGCAGGCGTCATG) oligonucleotides were pur-chased from Sigma Genosys. Anti-human IGFBP-5antibody was from IBT-Immunological and BiochemicalTestsystems, GmbH (Reutlingen, Germany).
Subjects
The study was approved by the Local Research EthicsCommittee and all patients gave written, informedconsent. Perioperative muscle biopsies were taken fromthe anterior abdominal wall of patients undergoingelective surgery. Biopsies were collected from 41 sub-jects, 24 male and 17 female. Average age was 56 years,(range 36–85 years); BMI 26.71, (range 16.5–40.5).
Cell culture
Cells were isolated following a modified method ofBlau and Webster (1981) (Crown et al., 2000). Briefly,biopsies were dissected into 1 mm3 pieces and digestedin TE (0.05% trypsin, 0.02% EDTA in phosphatebuffered saline (PBS)) for 15 min with gentle mixing.The supernatant was removed and added to hiFBS toneutralize the trypsin before centrifugation at 1,000 rpmfor 5 min. The resultant cell pellet was resuspendedin growth medium (Ham’s F-10 supplemented with20% hiFBS, 50 IU/ml penicillin, 5 mg/ml streptomycinand2.5mg/mlamphotericin B).The processwasrepeateda further two times and the cell suspensions pooled andplated onto T75 flasks coated with 0.2% gelatin. Cellswere grown in a humidified 5% CO2 atmosphere at378C and passaged when 80–90% confluent. Cells wereinduced to differentiate when approximately 80% con-fluent by washing twice in PBS and placing into lowserum media (LSM) (minimum essential medium(MEM) supplemented with 2% hiFBS, 1% L-glutamine,50 IU/ml penicillin, 5 mg/ml streptomycin and 2.5 mg/mlamphotericin B). Experimental cells were dosed attransfer into LSM as described in the text and all ex-periments were terminated after 7 days unless other-wise stated. By day 7, cultures had reached maximumdifferentiation as assessed by measuring increasingcreatine kinase (CK) activity. Experiments were per-formed on cells between passages 2 and 6. The culturesare not homogenous populations of myoblasts butinclude other cell types that were present in the originalbiopsy. The proportion of myoblasts was determined foreach biopsy by desmin staining and cell counting (Crownet al., 2000). Cultures on average contained 48�2.4%myoblasts. Cell death was assessed using Trypan Blueexclusion. Aliquots of cells (50 ml) were loaded onto ahaemocytometer 1:1 with Trypan Blue (50 ml). Total cellnumber was counted and dead cells were identified asthose that had taken up the blue dye. The percentage ofdead cells could then be calculated.
CK assay
Cells were grown on 35-mm dishes. After treatment,media was removed and stored at �808C. The cells werewashed twice with PBS and scraped into 60 ml of TMTbuffer (50 mM Tris-MES, 1% Triton X-100, pH 7.8).
MYOBLAST DIFFERENTIATION: A ROLE FOR IGFBP-3 71

Samples were stored at �808C and were assayed within2 weeks of collection using a commercially available kitfollowing the manufacturer’s instructions (Sigma CK-10). Enzymatic activity was normalised to total proteincontent as determined by the BCA protein assay (Pierce,Chester, UK).
Ligand and Western blotting
Cells were grown on 35-mm dishes. After treatmentmedia was removed and stored at�208C and cell lysateswere prepared. Cells were washed twice with PBS andthen scraped into 100 ml lysis buffer (10 mM Tris-HClpH 7.6, 5 mM EDTA, 50 mM NaCl, 30 mM sodiumpyrophosphate, 50 nM NaF, 100 mM Na2VO4, 0.1 mMPMSF). Samples were stored at �208C until used.Binding proteins in conditioned media or cell lysates,were separated under non-reducing conditions using12.5% SDS–PAGE, transferred to Hybond-C nitro-cellulose membrane (Amersham, Little Chalfont, UK)and analysed either by ligand or Western blotting. Forligand blots, membranes were probed with a mixtureof radiolabelled IGF-I and IGF-II and visualised byautoradiography. For Western blots, membranes wereprobed with an in-house anti-human IGFBP-3 antibody,(1:5,000) or an anti-human IGFBP-5 antibody (1:1,000)in TBST (10 mM Tris base pH 7.4, 150 mM NaCl, 0.2%tween-20) plus 5% milk. Antibodies were incubated atroom temperature over-night. Appropriate secondaryantibody was used (1:10,000) in TBST plus 5% milk andproteins were visualised using enhanced chemilumines-cence (Amersham).
Radioimmunoassay
Secreted IGFBP-3 was quantified by an in-house radioimmunoassay (RIA) as previously described(Yateman et al., 1993), using an in-house antiserum(Cwyfan Hughes et al., 1995) at a final dilution of1:20,000 (interassay % CV 5.1).
Flow cytometry
Cells were grown on 35-mm dishes. Following treat-ment, dead cells in the media were collected and the liveadherent cells were trypsinised in 0.1 ml TE solution.The cells were pooled and fixed following a PBS wash in1 ml of 70% ethanol. Following a minimum of 24 h at 48C,cells were pelleted (6,500 rpm; 5 min) and washed oncewith PBS (6,500 rpm; 5 min). The cells were resus-pended in Krishans reaction buffer (0.05 mg/ml propi-dium iodide, 0.1% sodium citrate, 0.02 mg/ml RNAseA, 0.3% NP-40, pH 8.3) and incubated at 48C for 30 minin the dark. DNA profiles of all cells were measuredon a FacsCalibur flow cytometer (Beckton-Dickinson,Oxford, UK) by an argon laser at 488 nm and analysedusing Cell Quest (Beckton-Dickinson) and Multicycle(Phoenix flow systems, San Diego, CA) softwarepackages.
Statistical analyses
Data were analysed using Microsoft Excel version5.0. Results were expressed as mean�SEM. Signifi-cant effects were determined using Student’s t-test(unpaired, 2-tailed). A statistically significant differencewas considered to be present at P<0.05.
RESULTSDetection of IGFBPs in the conditioned media
The presence of 20% foetal bovine serum in theunconditioned growth media (GM) meant that bovine-derived IGFBP-2, -3, -4 and -5 were detectable by ligandblotting and IGFBP-2 was also detectable in the un-conditioned differentiating media (Fig. 1A). The pri-mary muscle cell cultures were induced to differentiateby transfer into LSM. Media was collected and ligandblotting revealed the presence of IGFBPs at the molec-ular weights for IGFBP-2, -3, and -4 at day 7. At day1 IGFBP-5 was also detected in the media (Fig. 1C).The identity of the IGFBPs was confirmed by West-ern blotting using specific antibodies (Crown et al.,2000 (IGFBP-2 and -4), Fig. 3A (IGFBP-3) and Fig. 4(IGFBP-5)). The antibodies for IGFBP-3 (Fig. 1B),IGFBP-5 (Fig. 4) and IGFBP-4 (Crown et al., 2000)are human specific. Therefore with the possible excep-tion of IGFBP-2, the results indicate that the cul-tures are producing their own human-derived IGFBP-3, -4 and -5. Specifically all the IGFBP-3 detected onWestern immunoblots or by RIA is secreted by themuscle cell cultures and is not derived from the culturemedium.
IGFBP-3 increases with differentiation, intactIGFBP-5 decreases with differentiation
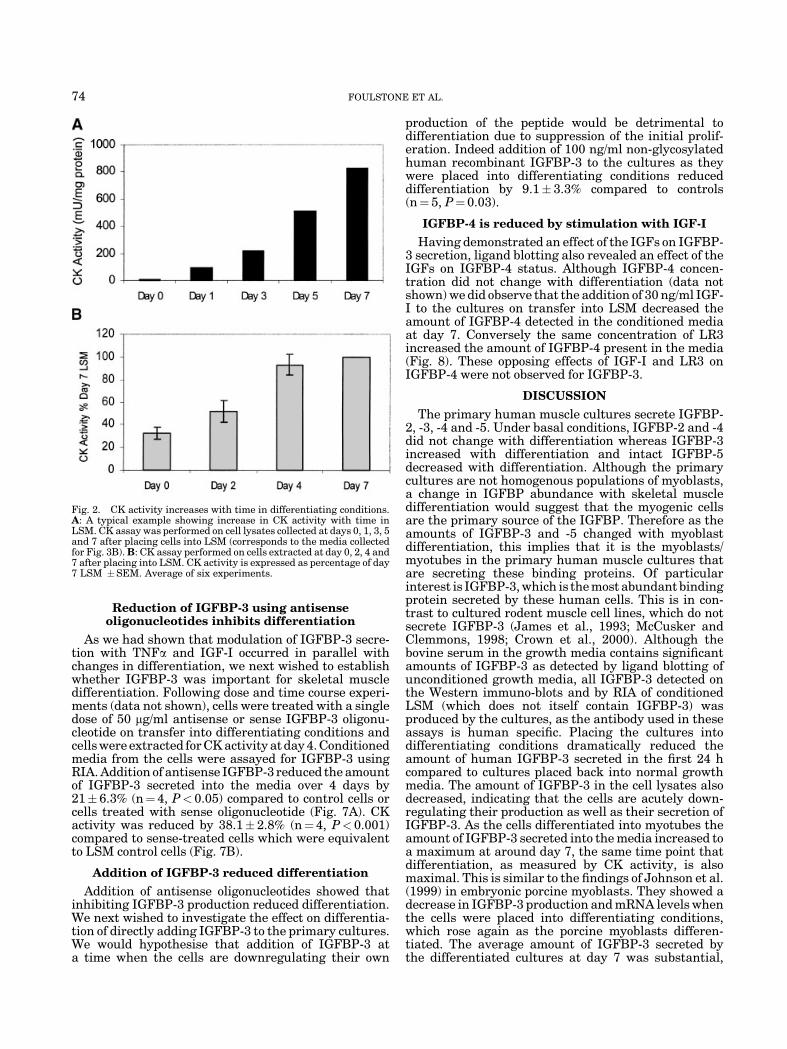
Myoblast differentiation was assessed biochemicallyover the 7 day differentiating period by measuringCK activity, which increases as myoblasts fuse toform multi-nucleated myotubes. CK activity increasedover the 7-day period and begins to plateau by day 7(Fig. 2A,B). Cultures kept in LSM for longer than 7 daysshowed no further increase in CK activity (data notshown). The average percentage myoblasts in the cul-tures was 48�2.4%, this did not significantly changeover the 7 day differentiating period.
We determined that the amount of IGFBP-2 andIGFBP-4 in the media did not alter with differentiation(Fig. 1C) whereas the concentrations of IGFBP-3 andIGFBP-5 did change with the progression to myotubes.The amount of IGFBP-3 secreted into the LSM after24 h was very low (Fig. 1C and 3A). The concentrationof IGFBP-3 in the media then increased over time underdifferentiating conditions. This was established byWestern blotting (Fig. 3A), and confirmed with RIAanalysis (Fig. 3B,C). The average IGFBP-3 concentra-tion in day 7 media was 249�22 ng/mg protein (n¼38).The IGFBP-3 secreted into the conditioned media waspredominantly intact, with very few proteolytic frag-ments being detected on the Western blot (Fig. 3A). Thisapparent lack of proteolytic activity was maintainedover the 7 day differentiating period. The decrease inthe amount of secreted IGFBP-3 which occurred afterplacing cells into LSM was paralleled by a decrease inthe amount of IGFBP-3 detected in the cell lysates after24 h in LSM (Fig. 3D). This reduction in intra-cellularand secreted IGFBP-3 was not observed if cells weretreated the same, but placed back into normal growthmedia instead of LSM (Fig. 3D,E). The amount ofIGFBP-3, which was maximal by day 7, correspondedto the plateau in CK activity also seen at this time point(Fig. 2A,B).
72 FOULSTONE ET AL.

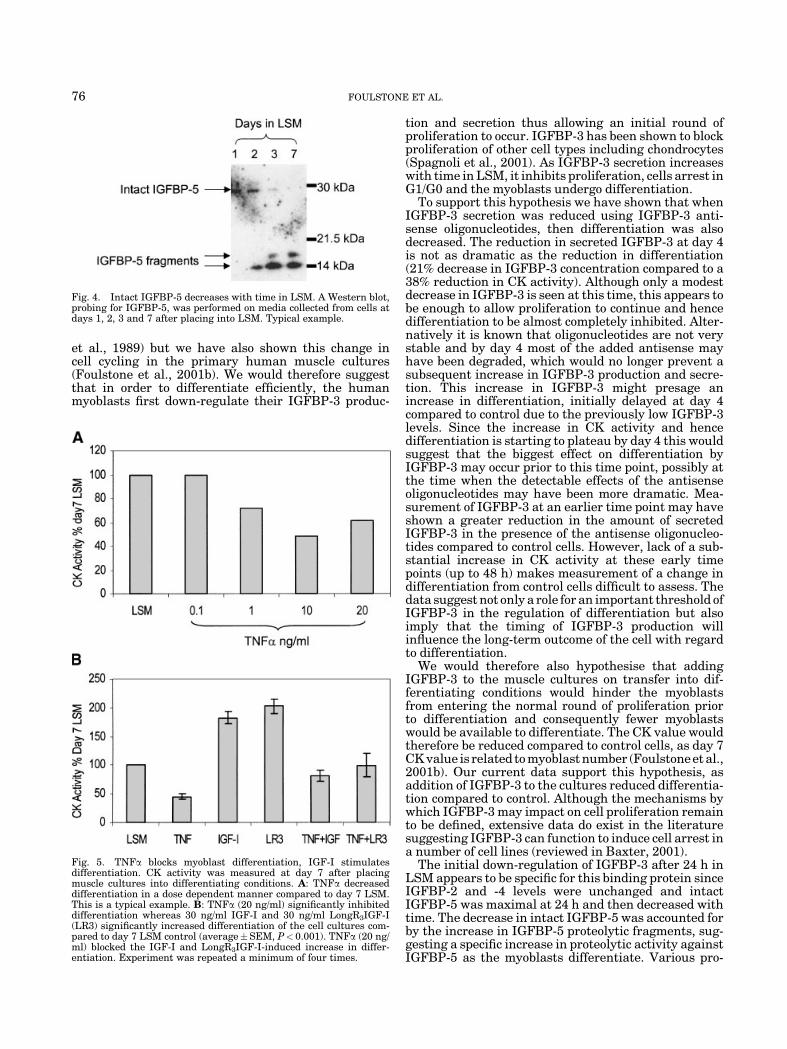
The initial decrease followed by an increase inIGFBP-3 production with differentiation was specificto this binding protein. In contrast to IGFBP-3, intactIGFBP-5, which is seen in the differentiating mediaat day 1, decreased with time over the 7 days in LSM(Fig. 4). IGFBP-5 proteolytic fragments increased overthe same time period such that by day 7 no intactIGFBP-5 was present in the media (Fig. 4).
In contrast to the IGFBPs, both IGF-I and IGF-II wereundetectable by RIA in the conditioned media from theprimary cultures (data not shown).
TNFa blocks myoblast differentiation,IGF-I stimulates differentiation
Having characterised the IGFBPs secreted by theprimary skeletal muscle cultures, we next wished toinvestigate the impact of modulators of skeletal muscledifferentiation in this model. The inhibition of differ-entiation by TNFa was dose dependent, with maximuminhibition occurring at 10–20 ng/ml (Fig 5A). Addition of20 ng/ml TNFa significantly inhibited differentiation by54.6�4.6% (P<0.001) (Fig. 5B). As TNFa can induceapoptosis of murine C2 skeletal myoblasts (Foulstoneet al., 2001), we assessed the effect of TNFa on apoptosisin the primary human muscle cultures. Using TrypanBlue exclusion and Flow cytometry we found thatstimulation with TNFa did not induce any apoptosis(data not shown). Stimulation with 30 ng/ml IGF-I or
30 ng/ml LongR3IGF-I (LR3) (maximal doses, data notshown) on the other hand significantly increased dif-ferentiation of the myoblasts by 82.8�10.9% and102.6� 13.1%, respectively (P< 0.001) (Fig. 5B). Therewas no significant difference between IGF-I and LR3stimulation of differentiation. Co-incubation of thesegrowth factors with TNFa blocked the IGF-I/LR3-induced increase in differentiation (Fig. 5B).
TNFa inhibits IGFBP-3 secretion whereasIGF-I increases secretion
Since TNFa blocked differentiation we wished todetermine the impact of TNFa on the increase inIGFBP-3 secretion which occurs with differentiation.As with differentiation, TNFa decreased IGFBP-3secretion in a dose dependant manner (Fig. 6A). As wellas reducing the amount of secreted IGFBP-3, 20 ng/mlTNFa also reduced the amount of IGFBP-3 in thecell lysate (Fig. 6B) suggesting that synthesis as wellas secretion of IGFBP-3 was inhibited by treatmentwith this cytokine. Stimulation of the cultures with30 ng/ml IGF-I or 30 ng/ml LR3, which increaseddifferentiation, also significantly increased IGFBP-3secretion at day 7 compared to control (P<0.001)(Fig 6A,C). Addition of the growth factors could notreverse the TNFa-induced inhibition of IGFBP-3 secre-tion (Fig. 6A). This was confirmed with RIA analysis(Fig. 6C).
Fig. 1. Primary muscle cultures secrete IGFBP-2, -3, -4 and -5 under differentiating conditions.A: Ligand blot of unconditioned growth media (GM) and unconditioned LSM. Normal human serum(NHS) was used as a positive control for IGFBP-3. B: Ligand blot was re-probed with anti-humanIGFBP-3 antibody. C: Ligand blot of media collected from differentiating cells after 1 day and 7 days indifferentiating conditions.
MYOBLAST DIFFERENTIATION: A ROLE FOR IGFBP-3 73
Reduction of IGFBP-3 using antisenseoligonucleotides inhibits differentiation
As we had shown that modulation of IGFBP-3 secre-tion with TNFa and IGF-I occurred in parallel withchanges in differentiation, we next wished to establishwhether IGFBP-3 was important for skeletal muscledifferentiation. Following dose and time course experi-ments (data not shown), cells were treated with a singledose of 50 mg/ml antisense or sense IGFBP-3 oligonu-cleotide on transfer into differentiating conditions andcells were extracted for CK activity at day 4. Conditionedmedia from the cells were assayed for IGFBP-3 usingRIA. Addition of antisense IGFBP-3 reduced the amountof IGFBP-3 secreted into the media over 4 days by21�6.3% (n¼ 4, P< 0.05) compared to control cells orcells treated with sense oligonucleotide (Fig. 7A). CKactivity was reduced by 38.1�2.8% (n¼4, P<0.001)compared to sense-treated cells which were equivalentto LSM control cells (Fig. 7B).
Addition of IGFBP-3 reduced differentiation
Addition of antisense oligonucleotides showed thatinhibiting IGFBP-3 production reduced differentiation.We next wished to investigate the effect on differentia-tion of directly adding IGFBP-3 to the primary cultures.We would hypothesise that addition of IGFBP-3 ata time when the cells are downregulating their own
production of the peptide would be detrimental todifferentiation due to suppression of the initial prolif-eration. Indeed addition of 100 ng/ml non-glycosylatedhuman recombinant IGFBP-3 to the cultures as theywere placed into differentiating conditions reduceddifferentiation by 9.1� 3.3% compared to controls(n¼ 5, P¼0.03).
IGFBP-4 is reduced by stimulation with IGF-I
Having demonstrated an effect of the IGFs on IGFBP-3 secretion, ligand blotting also revealed an effect of theIGFs on IGFBP-4 status. Although IGFBP-4 concen-tration did not change with differentiation (data notshown) we did observe that the addition of 30 ng/ml IGF-I to the cultures on transfer into LSM decreased theamount of IGFBP-4 detected in the conditioned mediaat day 7. Conversely the same concentration of LR3increased the amount of IGFBP-4 present in the media(Fig. 8). These opposing effects of IGF-I and LR3 onIGFBP-4 were not observed for IGFBP-3.
DISCUSSION
The primary human muscle cultures secrete IGFBP-2, -3, -4 and -5. Under basal conditions, IGFBP-2 and -4did not change with differentiation whereas IGFBP-3increased with differentiation and intact IGFBP-5decreased with differentiation. Although the primarycultures are not homogenous populations of myoblasts,a change in IGFBP abundance with skeletal muscledifferentiation would suggest that the myogenic cellsare the primary source of the IGFBP. Therefore as theamounts of IGFBP-3 and -5 changed with myoblastdifferentiation, this implies that it is the myoblasts/myotubes in the primary human muscle cultures thatare secreting these binding proteins. Of particularinterest is IGFBP-3, which is the most abundant bindingprotein secreted by these human cells. This is in con-trast to cultured rodent muscle cell lines, which do notsecrete IGFBP-3 (James et al., 1993; McCusker andClemmons, 1998; Crown et al., 2000). Although thebovine serum in the growth media contains significantamounts of IGFBP-3 as detected by ligand blotting ofunconditioned growth media, all IGFBP-3 detected onthe Western immuno-blots and by RIA of conditionedLSM (which does not itself contain IGFBP-3) wasproduced by the cultures, as the antibody used in theseassays is human specific. Placing the cultures intodifferentiating conditions dramatically reduced theamount of human IGFBP-3 secreted in the first 24 hcompared to cultures placed back into normal growthmedia. The amount of IGFBP-3 in the cell lysates alsodecreased, indicating that the cells are acutely down-regulating their production as well as their secretion ofIGFBP-3. As the cells differentiated into myotubes theamount of IGFBP-3 secreted into the media increased toa maximum at around day 7, the same time point thatdifferentiation, as measured by CK activity, is alsomaximal. This is similar to the findings of Johnson et al.(1999) in embryonic porcine myoblasts. They showed adecrease in IGFBP-3 production and mRNA levels whenthe cells were placed into differentiating conditions,which rose again as the porcine myoblasts differen-tiated. The average amount of IGFBP-3 secreted bythe differentiated cultures at day 7 was substantial,
Fig. 2. CK activity increases with time in differentiating conditions.A: A typical example showing increase in CK activity with time inLSM. CK assay was performed on cell lysates collected at days 0, 1, 3, 5and 7 after placing cells into LSM (corresponds to the media collectedfor Fig. 3B). B: CK assay performed on cells extracted at day 0, 2, 4 and7 after placing into LSM. CK activity is expressed as percentage of day7 LSM �SEM. Average of six experiments.
74 FOULSTONE ET AL.

249�22 ng IGFBP-3/mg protein as measured by RIA.These specific alterations in IGFBP-3 production sug-gest it may play an integral role in the control ofmyoblast growth and differentiation.
When myoblasts are placed into differentiating con-ditions they undergo a round of proliferation before thecells arrest in G1/G0, prior to differentiation. This hasbeen shown not only in the myoblast cell lines (Tollefsen
Fig. 3. The amount of IGFBP-3 increases with differentiation.Placing muscle cultures into LSM but not into GM decreasesIGFBP-3 in the cell lysate. A: A Western blot, probing for IGFBP-3,was performed on media collected at days 1, 3, 5 and 7 after placingcells into LSM. Conditioned growth media (GM) collected from cellsprior to placing into differentiating conditions was also run. Typicalexample. B: A typical example of increasing IGFBP-3 concentrationwith time in LSM. Media was collected at days 2, 3, 5 and 7 afterplacing cells into LSM and RIA was performed. RIA was also
performed on unconditioned differentiating media (LSM). IGFBP-3concentration has been corrected for protein. C: RIA was performed onmedia collected at days 1, 3, 5 and 7. IGFBP-3 was corrected forprotein and is expressed as percentage of day 7 IGFBP-3�SEM.Average of five experiments. D: A Western blot was performed on celllysates extracted at time 0 (T0) and 24 h after placing cells into eitherLSM or GM. E: RIA was performed on media from cells after 24 h ineither LSM or GM. Typical examples, experiment was repeated atleast three times.
MYOBLAST DIFFERENTIATION: A ROLE FOR IGFBP-3 75
et al., 1989) but we have also shown this change incell cycling in the primary human muscle cultures(Foulstone et al., 2001b). We would therefore suggestthat in order to differentiate efficiently, the humanmyoblasts first down-regulate their IGFBP-3 produc-
tion and secretion thus allowing an initial round ofproliferation to occur. IGFBP-3 has been shown to blockproliferation of other cell types including chondrocytes(Spagnoli et al., 2001). As IGFBP-3 secretion increaseswith time in LSM, it inhibits proliferation, cells arrest inG1/G0 and the myoblasts undergo differentiation.
To support this hypothesis we have shown that whenIGFBP-3 secretion was reduced using IGFBP-3 anti-sense oligonucleotides, then differentiation was alsodecreased. The reduction in secreted IGFBP-3 at day 4is not as dramatic as the reduction in differentiation(21% decrease in IGFBP-3 concentration compared to a38% reduction in CK activity). Although only a modestdecrease in IGFBP-3 is seen at this time, this appears tobe enough to allow proliferation to continue and hencedifferentiation to be almost completely inhibited. Alter-natively it is known that oligonucleotides are not verystable and by day 4 most of the added antisense mayhave been degraded, which would no longer prevent asubsequent increase in IGFBP-3 production and secre-tion. This increase in IGFBP-3 might presage anincrease in differentiation, initially delayed at day 4compared to control due to the previously low IGFBP-3levels. Since the increase in CK activity and hencedifferentiation is starting to plateau by day 4 this wouldsuggest that the biggest effect on differentiation byIGFBP-3 may occur prior to this time point, possibly atthe time when the detectable effects of the antisenseoligonucleotides may have been more dramatic. Mea-surement of IGFBP-3 at an earlier time point may haveshown a greater reduction in the amount of secretedIGFBP-3 in the presence of the antisense oligonucleo-tides compared to control cells. However, lack of a sub-stantial increase in CK activity at these early timepoints (up to 48 h) makes measurement of a change indifferentiation from control cells difficult to assess. Thedata suggest not only a role for an important threshold ofIGFBP-3 in the regulation of differentiation but alsoimply that the timing of IGFBP-3 production willinfluence the long-term outcome of the cell with regardto differentiation.
We would therefore also hypothesise that addingIGFBP-3 to the muscle cultures on transfer into dif-ferentiating conditions would hinder the myoblastsfrom entering the normal round of proliferation priorto differentiation and consequently fewer myoblastswould be available to differentiate. The CK value wouldtherefore be reduced compared to control cells, as day 7CK value is related to myoblast number (Foulstone et al.,2001b). Our current data support this hypothesis, asaddition of IGFBP-3 to the cultures reduced differentia-tion compared to control. Although the mechanisms bywhich IGFBP-3 may impact on cell proliferation remainto be defined, extensive data do exist in the literaturesuggesting IGFBP-3 can function to induce cell arrest ina number of cell lines (reviewed in Baxter, 2001).
The initial down-regulation of IGFBP-3 after 24 h inLSM appears to be specific for this binding protein sinceIGFBP-2 and -4 levels were unchanged and intactIGFBP-5 was maximal at 24 h and then decreased withtime. The decrease in intact IGFBP-5 was accounted forby the increase in IGFBP-5 proteolytic fragments, sug-gesting a specific increase in proteolytic activity againstIGFBP-5 as the myoblasts differentiate. Various pro-
Fig. 4. Intact IGFBP-5 decreases with time in LSM. A Western blot,probing for IGFBP-5, was performed on media collected from cells atdays 1, 2, 3 and 7 after placing into LSM. Typical example.
Fig. 5. TNFa blocks myoblast differentiation, IGF-I stimulatesdifferentiation. CK activity was measured at day 7 after placingmuscle cultures into differentiating conditions. A: TNFa decreaseddifferentiation in a dose dependent manner compared to day 7 LSM.This is a typical example. B: TNFa (20 ng/ml) significantly inhibiteddifferentiation whereas 30 ng/ml IGF-I and 30 ng/ml LongR3IGF-I(LR3) significantly increased differentiation of the cell cultures com-pared to day 7 LSM control (average�SEM, P< 0.001). TNFa (20 ng/ml) blocked the IGF-I and LongR3IGF-I-induced increase in differ-entiation. Experiment was repeated a minimum of four times.
76 FOULSTONE ET AL.

teases specific to IGFBP-5 have been described in theliterature and include serine and metallo-proteinases(Nam et al., 1996; Loechel et al., 2000; Overgaard et al.,2001). It is feasible that the protease in our primaryhuman cultures is one of these, however, future studieswould need to be initiated to begin to investigate whichones are present. IGFBP-5 has been reported to stim-ulate proliferation in chondrocytes (Kiepe et al., 2001)and in osteoblasts (Andress, 2001). In addition Kiepeet al. (2001) report that an amino-terminal IGFBP-5fragment blocked chondrocyte proliferation. Thesefindings complement the data from our model whereintact IGFBP-5 is present during the initial round ofproliferation with the proteolytic fragments appearingas the cells arrest and start to differentiate. This raisesthe possibility that IGFBP-5 as well as IGFBP-3 mayinfluence myoblast proliferation and differentiation.In contrast to IGFBP-5, IGFBP-3 showed very littleproteolysis over the 7 day differentiating period as seenby the lack of significant fragments on the Western blots.This lack of proteolysis was unexpected, as a prominent30 kDa proteolytic fragment of IGFBP-3 is usuallyevident on Western immuno-blots of normal humanserum.
Both TNFa and IGF-I, which we have shown mod-ulate differentiation of the muscle cultures, are seen toalter IGFBP-3 secretion. TNFa, which inhibits differ-entiation and promotes proliferation, also reducedthe production and the secretion of IGFBP-3 into thedifferentiating media. TNFa has been shown to in-crease proliferation of myoblasts both in the C2 cell line(Foulstone et al., 2001) and also in the primary musclecultures (Foulstone et al., 2001b). One way that TNFamay be able to continue to stimulate proliferation is bysuppressing IGFBP-3 levels. In contrast IGF-I, whichincreases differentiation measured as an increase in CKactivity at day 7, also increased the amount of IGFBP-3secretion seen at this time point. The initial decrease inIGFBP-3 at 24 h, however, is still seen with addition ofIGF-I (data not shown). The fact that LR3, which doesnot bind to the IGFBPs, also increased IGFBP-3secretion would suggest that the increase in IGFBP-3in the media is not due to an increase in stabilityconferred by binding to IGF-I.
The amount of IGFBP-2 and IGFBP-4 detected usingligand blotting did not change over the 7 day differ-entiating period. However, addition of IGF-I to the cellsas they were placed into differentiating media reduced
Fig. 6. TNFa inhibits IGFBP-3 secretion whereas IGF-Iincreases secretion A: A typical Western blot of mediacollected after 7 days in LSM, probed for IGFBP-3. Cellswere either untreated (LSM), stimulated with TNFa at thedoses specified or treated with 30 ng/ml IGF-I (IGF) or30 ng/ml LongR3IGF-I (LR3) with or without 20 ng/ml TNFa.B: Western blot of cell lysates at day 7. Cells wereuntreated (LSM) or treated with 20 ng/ml TNFa (a) ontransfer into LSM. C: RIAs for IGFBP-3 were performed onmedia collected from cells after 7 days in LSM. Cells wereuntreated (LSM) or stimulated with 30 ng/ml IGF-I, 30 ng/ml LongR3IGF-I (LR3), 20 ng/ml TNFa, or a combination ofTNFa plus growth factor. IGFBP-3 concentration is ex-pressed as a percentage of day 7 LSM�SEM. All conditionswere significantly different from LSM, P< 0.001, experi-ment was repeated at least four times.
MYOBLAST DIFFERENTIATION: A ROLE FOR IGFBP-3 77

the amount of IGFBP-4 seen in the media at day 7. Thisdecrease is not seen when the same concentration of LR3is added, in fact the levels of IGFBP-4 increased. Wehave previously shown that in a fibroblast cell culturesystem, IGFBP-4 is degraded by a protease that isinhibited by IGFBP-3, when IGFBP-3 is in a non-IGFbound state (Donnelly and Holly, 1996). We suggest thatthis may be what is occurring in the primary muscle cellcultures. IGFBP-3 levels in the media increase asthe myoblasts differentiate to form myotubes. In thepresence of IGF-I this IGFBP-3 would be sequestered bythe growth factor and would therefore be unavailable toinhibit the proteolysis of IGFBP-4. LR3 is capable ofstimulating IGFBP-3 secretion, but as it does not bind tothe IGFBPs it would not sequester the IGFBP-3 thus
allowing it to bind to the protease and protect IGFBP-4from being proteolysed. As LR3 increased the concen-tration of IGFBP-3 over control levels so by defaultthe activity of the protease would be blocked and theconcentration of IGFBP-4 would be increased comparedto day 7 control media. We suggest that the addition ofLR3 increased the protection of IGFBP-4, alternativelyhowever, LR3 may have a direct effect on production ofIGFBP-4. To clarify this further, future studies inves-tigating the impact of LR3 versus IGF-I on IGFBP-4message would provide informative data.
Although the liver is thought to be the major source ofcirculating IGFBP-3 the evidence relies on the correla-tion between IGFBP-3 levels and liver function (Schalchet al., 1998; Shaaravy et al., 1998). In the circulation theIGFBP-3 is bound in a ternary complex to IGF-I and theacid labile subunit (ALS). Liver specific IGF-I knockoutmodels show a reduction in circulating IGF-I but noeffect on skeletal muscle growth and development(Butler and LeRoith, 2001). ALS knockout mice alsoshow a reduction in circulating IGF-I and at the sametime a 90% reduction in circulating IGFBP-3 butanimals have a very similar phenotype to the liverspecific IGF-I knockouts (Butler and LeRoith, 2001).These knockouts which reduce the circulating IGF-I,suggest that local autocrine production of IGF-I byskeletal muscle is important for its growth and main-tenance. The data from the ALS knockout suggest thatthis may also be the case for IGFBP-3. It may be that, asfor IGF-I, the local production of IGFBP-3 in the skeletalmuscle rather than the systemic levels of IGFBP-3are important to skeletal muscle physiology/pathophy-siology. Indeed skeletal muscle, as has been acknowl-edged for adipose tissue, may itself act as an autocrine/endocrine organ for the normal production of IGFBP-3.
Our results show that the amount and timing of IGBP-3 production is critical. Too much IGFBP-3 present tooearly during differentiation will prevent the initialround of proliferation. The eventual number of myo-blasts will be lower and the myotubes themselves willtherefore be smaller. Reduced IGFBP-3 on the otherhand is equally detrimental, as the myoblasts willcontinue to proliferate, preventing arrest in G0/G1 andhence preventing differentiation. Understanding therole that IGFBP-3 plays in the switch between myoblastproliferation and differentiation is therefore vital ifskeletal muscle stem cell systems are going to be im-plemented as viable therapeutic approaches. Alteringlocal IGFBP-3 may also allow manipulation of musclesatellite cells in vivo. This could have fundamentalclinical implications for conditions where muscle regen-eration is impaired. Furthermore, the presence of largeamounts of IGFBP-3 being produced by the musclecultures raises the interesting possibility that skeletalmuscle itself may be a source for circulating IGFBP-3.
ACKNOWLEDGMENTS
We thank the upper GI surgical team lead by Prof. DAlderson at the Bristol Royal Infirmary and Mr. P Nivenat St Michaels Hospital, Bristol for muscle biopsysamples. Many thanks to Moira Hunt, research nurseat the Bristol Royal Infirmary, for patient consentingand data collection. This work was funded by theAssociation of International Cancer Research (AICR).
Fig. 7. IGFBP-3 antisense oligonucleotides block IGFBP-3 secretionand myoblast differentiation. A: A total of 50 mg/ml antisense IGFBP-3or 50 mg/ml sense IGFBP-3 oligonucleotides were added to the musclecultures when they were placed into differentiating conditions. Mediawas collected at day 4 and assayed for IGFBP-3 using RIA. Valueswere corrected for protein and expressed as percentage of day 4 LSM.Average of four experiments. B: Cells were extracted at day 4 andassayed for CK activity. Values were expressed as percentage of day 4LSM. Average of four experiments.
Fig. 8. IGFBP-4 proteolysis is inhibited by IGFBP-3. A ligand blotwas performed using media collected at day 7 after placing cells intoLSM. Cells were untreated (LSM) or stimulated with 30 ng/ml IGF-I(IGF) or 30 ng/ml LongR3IGF-I (LR3).
78 FOULSTONE ET AL.

Extra funding was obtained from The Charitable trustfor the United Bristol Hospitals and from the PeelMedical Research Trust.
LITERATURE CITED
Andress DL. 2001. IGF-binding protein-5 stimulates osteoblastactivity and bone accretion in ovariectomized mice. Am J PhysiolEndocrinol Metab 281:E283–288.
Baxter RC. 2001. Signalling pathways involved in antiproliferativeeffects of IGFBP-3: A review. Mol Pathol 54:145–148.
Blau HM, Webster C. 1981. Isolation and characterisation of humanmuscle cells. Proc Natl Acad Sci USA 78:5623–5627.
Butler AA, LeRoith D. 2001. Tissue-specific versus generalized genetargeting of the igfI and igfIr genes and their roles in insulin-likegrowth physiology. Endocrinology 142:1685–1688.
Calman KC. 1982. Malignancy. Cancer cachexia. Br J Hosp Med 27:28–29, 33–34.
Collett-Solberg PF, Cohen P. 1996. The role of the insulin-like growthfactor binding proteins and the IGFBP proteases in modulating IGFaction. Endocrinol Metab Clin North Am 25:591–614.
Conover CA, Ronk M, Lombana F, Powell DR. 1990. Structural andbiological characterisation of bovine insulin-like growth factorbinding protein-3. Endocrinology 127:2795–2803.
Conover CA, Bale LK, Clarkson JT, Durham SK. 1993. Potentiationof insulin-like growth factor action by insulin-like growth factorbinding protein-3: Studies of underlying mechanism. Growth Regul3:87–88.
Crown AL, He XL, Holly JMP, Lightman SL, Stewart CEH. 2000.Characterisation of the IGF system in a primary adult humanskeletal muscle cell model, and comparison of the effects of insulinand IGF-I on protein metabolism. J Endocrinol 167:403–415.
Cwyfan Hughes SC, Johnson MR, Heinrich G, Holly JMP. 1995.Could the abnormalities in insulin-like growth factors and theirbinding proteins during pregnancy result in gestational diabetes?J Endocrinol 147:517–524.
Donnelly MJ, Holly JMP. 1996. The role of IGFBP-3 in the regulationof IGFBP-4 proteolysis. J Endocrinol 149:R1–7.
Florini JR, Magri KA, Ewton DZ, James PL, Grindstaff K,Rotwein PS. 1991. ‘‘Spontaneous’’ differentiation of skeletal myo-blasts is dependent upon autocrine secretion of insulin-like growthfactor-II. J Biol Chem 266:15917–15923.
Foulstone EJ, Meadows KA, Holly JMP, Stewart CEH. 2001. Insulin-like growth factors (IGF-I and IGF-II) inhibit C2 skeletal myoblastdifferentiation and enhance TNFa-induced apoptosis. J Cell Physiol189:207–215.
Foulstone EJ, Crown AL, Holly JMP, Stewart CEH. 2001b. Com-parison of primary skeletal muscle cultures from patients with andwithout cancer. In: Proceedings of the 83rd annual meeting of theAmerican Endocrine Society. Denver. p 1–198.
Fowler CA, Perks CM, Newcomb PV, Savage PB, Farndon JR,Holly JMP. 2000. Insulin-like growth factor binding protein-3(IGFBP-3) potentiates paclitaxel induced apoptosis in human breastcancer cells. Int J Cancer 88:448–453.
Gill ZP, Perks CM, Newcomb PV, Holly JMP. 1997. Insulin-likegrowth factor binding protein-3 (IGFBP-3) predisposes breast cancercells to programmed cell death in a non-IGF dependent manner.J Biol Chem 272:25602–25607.
Hawke TJ, Garry DJ. 2001. Myogenic satellite cells: Physiology tomolecular biology. J Appl Physiol 91:534–551.
Hollowood AD, Lai T, Perks CM, Newcomb PV, Alderson D,Holly JMP. 2000. IGFBP-3 prolongs the P53 response and enhancesapoptosis following UV irradiation. Int J Cancer 88:336–341.
James PL, Jones SB, Busby Jr., WH, Clemmons DR, Rotwein P. 1993.A highly conserved insulin-like growth factor binding protein(IGFBP-5) is expressed during myoblast differentiation. J BiolChem 268:22305–22312.
James PL, Stewart CEH, Rotwein P. 1996. Insulin-like growth factorbinding protein-5 modulates muscle differentiation through an
insulin-like growth factor-dependent mechanism. J Cell Biol 133:683–693.
Johnson BJ, White ME, Hathaway MR, Dayton WR. 1999. Decreasedsteady-state insulin-like binding protein-3 (IGFBP-3) mRNA levelis associated with differentiation of cultured porcine myogenic cells.J Cell Physiol 179:237–243.
Jones JI, Clemmons DR. 1995. Insulin-like growth factors and theirbinding proteins: Biological actions. Endocr Rev 16:3–34.
Kiepe D, Andress DL, Mohan S, Standker L, Ulinski T, Himmele R,Mehls O, Tonshoff B. 2001. Intact IGF-binding protein-4 and -5 andtheir respective fragments isolated from chronic renal failure serumdifferentially modulate IGF-I actions in cultured growth platechondrocytes. J Am Soc Nephrol 12:2400–2410.
Kim H-S, Rosenfeld RG, Oh Y. 1997. Biological roles of insulin-like growth factor binding proteins (IGFBPs). Exp Mol Med 29:85–96.
Loechel F, Fox JW, Murphy G, Albrechtsen R, Wewer UM. 2000.ADAM 12-S cleaves IGFBP-3 and IGFBP-5 and is inhibited byTIMP-3. Biochem Biophys Res Commun 278:511–515.
Matthys P, Billiau A. 1997. Cytokines and cachexia. Nutrition 13:763–770.
McCusker RH, Clemmons DR. 1998. Role for cyclic adenosinemonophosphate in modulating insulin-like growth factor bindingprotein secretion by muscle cells. J Cell Physiol 174:293–300.
Meadows KA, Holly JMP, Stewart CEH. 2000. Tumour nec-rosis factor a-induced apoptosis is associated with suppression ofinsulin-like growth factor binding protein-5 secretion in dif-ferentiating murine skeletal myoblasts. J Cell Physiol 183:330–337.
Nam TJ, Busby WH, Jr. Clemmons DR. 1996. Characterization anddetermination of the relative abundance of two types of insulin-likegrowth factor binding protein-5 proteases that are secreted byhuman fibroblasts. Endocrinology 137:5530–5536.
Olson EN. 1992. Interplay between proliferation and differentiationwithin the myogenic lineage. Dev Biol 154:261–272.
Olson EN, Rosenthal N. 1994. Homeobox genes and muscle pattern-ing. Cell 79:9–12.
Overgaard Mt, Boldt HB, Laursen LS, Sottrup-Jensen L, Conover CA,Oxvig C. 2001. Pregnancy-associated plasma protein-A2 (PAPP-A2),a novel insulin-like growth factor-binding protein-5 proteinase.J Biol Chem 276:21849–21853.
Schalch DS, Kalayoglu M, Pirsch JD, Yang H, Raslich M. 1998. Seruminsulin-like growth factors and their binding proteins in patientswith hepatic failure and after liver transplantation. Metabolism47:200–206.
Shaaravy M, Fikry MA, Massoud BA, Lofty S. 1998. Insulin-likegrowth factor binding protein-3: A novel biomarker for the assess-ment of the synthetic capacity of hepatocytes in liver cirrhosis. J ClinEndocrinol Metab 83:3316–3319.
Spagnoli A, Hwa V, Horton WA, Lunstrum GP, Roberts CT Jr,Chiarelli F, Torello M, Rosenfeld RG. 2001. Antiproliferative effectsof insulin-like growth factor-binding protein-3 in mesenchymalchondrogenic cell line RCJ3.1C5.18. Relationship to differentiationstage. J Biol Chem 276:5533–5540.
Stewart CEH, Rotwein P. 1996. Growth, differentiation, and survival:Multiple physiological functions for insulin-like growth factors.Physiol Rev 76:1005–1026.
Tollefsen SE, Sadow JL, Rotwein P. 1989. Co-ordinate expression ofinsulin-like growth factor-II and its receptor during muscle celldifferentiation. Proc Natl Acad Sci 86:1543–1547.
Yateman M, Claffey D, Cwyfan Huges S, Frost V, Wass J, Holly J.1993. Cytokines modulate the sensitivity of human fibroblasts tostimulation with insulin-like growth factor-I (IGF-I) by alteringendogenous IGF-binding protein production. J Endocrinol 137:151–159.
Yi Z, Hathaway MR, Dayton WR, White ME. 2001. Effects of growthfactors on insulin-like growth factor binding protein (IGFBP)secretion by primary porcine satellite cell cultures. J Anim Sci79:2820–2826.
MYOBLAST DIFFERENTIATION: A ROLE FOR IGFBP-3 79