Đs vers Đcolor tanisinda kl Đ k Örnekt.c. Çukurova Ün Đvers Đtes Đ sa Ğlik b ĐlĐmler Đ...
TRANSCRIPT

T.C. ÇUKUROVA ÜNĐVERSĐTESĐ SAĞLIK B ĐLĐMLER Đ ENSTĐTÜSÜ MĐKROBĐYOLOJ Đ ANABĐLĐM DALI
PĐTYRĐASĐS VERSĐCOLOR TANISINDA KL ĐNĐK ÖRNEK
ALMA YÖNTEMLER ĐNĐN ve ÇEŞĐTL Đ BESĐYERĐ
SONUÇLARININ KAR ŞILA ŞTIRILMASI
Pınar ALTINTOP
YÜKSEK L ĐSANS TEZĐ
TEZ YÖNET ĐCĐSĐ
Prof. Dr. Macit ĐLK ĐT
ADANA-2009

T.C. ÇUKUROVA ÜNĐVERSĐTESĐ SAĞLIK B ĐLĐMLER Đ ENSTĐTÜSÜ MĐKROBĐYOLOJ Đ ANABĐLĐM DALI
PĐTYRĐASĐS VERSĐCOLOR TANISINDA KL ĐNĐK ÖRNEK
ALMA YÖNTEMLER ĐNĐN ve ÇEŞĐTL Đ BESĐYERĐ
SONUÇLARININ KAR ŞILA ŞTIRILMASI
Pınar ALTINTOP
YÜKSEK L ĐSANS TEZĐ
TEZ YÖNET ĐCĐSĐ
Prof. Dr. Macit ĐLK ĐT
ADANA-2009

iii
TEŞEKKÜR
Eğitim sürem boyunca bilimsel desteklerini esirgemeyen, her zaman hoşgörülü
davranan, her konuda rahatlıkla ulaşıp danıştığım, büyük desteklerini gördüğüm ve
engin deneyimlerinden yararlandığım hocalarım; Sayın Prof Dr Macit Đlkit, Sayın Prof
Dr Mehmet Karakaş ve Sayın Prof Dr Fatih KÖKSAL’a, ve Çukurova Üniversitesi Tıp
Fakültesi Mikrobiyoloji Anabilim Dalı’ndaki diğer tüm hocalarıma, meslektaşım Taner
BOZKURT’a, Teknisyen Seher ÖKSÜZ’e, çalışma arkadaşlarıma ve bölüm
çalışanlarına sonsuz teşekkürlerimi sunarım.
Ayrıca, eğitim sürem boyunca maddi ve manevi desteğini esirgemeyen annem,
babam ve kardeşlerime sonsuz teşekkür ve saygılarımı sunarım.
Pınar ALTINTOP

iv
ĐÇĐNDEKĐLER
Sayfa No
Kabul ve Onay ıı
TEŞEKKÜR ııı
ĐÇĐNDEKĐLER ıv
ŞEKĐLLER DĐZĐNĐ vı
TABLOLAR DĐZĐNĐ vıı
SĐMGE VE KISALTMALAR D ĐZĐNĐ vııı
ÖZET-ANAHTAR SÖZCÜKLER ıx
ABSTRACT-KEYWORDS xııı
1.GĐRĐŞ 1
2.GENEL BĐLGĐLER 2
2.1. Tarihçe 2
2.2. Filogeni ve Taksonomi 5
2.3. Morfoloji 7
2.4. Fizyoloji 7
2.5 Yapı 8
2.6. Biyokimya 9
2.7.Malassezia’nın Karekteristiği 9
2.7.1. Đzoloasyon ve Đn vivo Đdentifikasyon 9
2.7.2. Đn vitro Đdentifikasyon 11
2.8. Malassezia’nın ekolojisi 16

v
2.9. Epidemiyoloji 18
2.10. Koleksiyonlardaki Malassezia’nın Korunması 20
2.11. Tanı 21
2.12. Kommensalizm 22
2.12.1. Normal Derideki Malassezia’nın türlerinin 22
Yayılımı
2.13. Deri Đmmun Sistemi 24
2.14. Malassezia Đle Đlgili Yüzeysel Hastalıklar 24
2.14.1. Pityriasis versicolor 25
2.14.2. Seboreik Dermatit 30
2.14.3. Atopik Dermatit 33
2.14.4. Malassezia (Pityrosporum)Folliküliti 34
2.14.5. Gougerot-Carteaud’un Retikulat (ağsı) ve 36
Birleşik Papillomları
2.14.6. Psöriasis 36
2.14.7. Malassezia Yenidoğan Püstülozu 37
2.14.8. Otitis 37
2.14.9. Onikomikoz 38
2.15. Moleküler Đdentifikasyon Yöntemi 38
2.16.Tedavi 39
2.16.1.Topikal Tedavi 39
2.16.1.1.Non-spesifik antifungal tedavi 39
2.16.1.2. Spesifik topikal anifungal tedavi 39

vi
2.16.2. Oral Tedavi 40
3. GEREÇ VE YÖNTEM 41
3.1. Klinik örnek alma yöntemi 41
3.2. Wood ışığı muayenesi 41
3.3. Mikolojik inceleme 42
3.3.1. Doğrudan mikroskop incelemesi 42
3.3.2. Kültür 42
3.3.3. Tanı Testleri 44
4. BULGULAR 49
5. TARTIŞMA 70
6. SONUÇLAR VE ÖNERĐLER 83
7. KAYNAKLAR 85
8. ÖZGEÇMĐŞ 112

vii
ŞEKĐLLER D ĐZĐNĐ
Şekil no Sayfa no
Şekil 2.1. Wood ışığı 22
Şekil 2.2. Örneğin laktofenol ile görünen köfte- makarna( blastospor ve hif) 26
Şekil 2.3. Pityriasis versicolor (Hipo-pigmente) 30
Şekil 2.4. Pityriasis versicolor (Hiper-pigmente) 30
Şekil 3.1. Lezyondan alınan örneğin %10’luk 42
KOH ile incelemesi (spor ve hif)
Şekil 3.2. Tween identifikasyon testi 45
Şekil 4.1. Pigmentasyon özelliğinin cinsiyete göre dağılımı 50
Şekil 4.2. Hastalığın süresi ile lezyonunu özelliği arasındaki ilişki 52
Şekil 4.3. Malassezia türlerinin pigmentasyon özelliğine göre dağılımı 55
Şekil 4.4. Modifiye-Dixon besiyeri a (Malassezia) 66
Şekil 4.5. Modifiye-Dixon besiyeri b (Malassezia) 66
Şekil 4.6. Modifiye-Dixon besiyeri- presipitasyon 66
Şekil 4.7. Modifiye-Dixon besiyeri-selofan- bant ile alınmış örnek 66
Şekil 4.8. CCA’da pigment ve presipitasyon 67
Şekil 4.9. Leeming-Notman besiyeri 67
Şekil 4.10. Modifiye-Dixon besiyeri-Büyük koloni 67
Bistüri ile alınmış örnek
Şekil 4.11. Tween 60-eskülin besiyeri-siyah zon (3+) 67

viii
Şekil 4.12. Tween-60 eskülin besiyeri siyah ve gri zon 67
Şekil 4.13. Tween 60-eskülin besiyerinde oluşan farklı renkteki zonlar 68
Şekil 4.14. Chremophor EL besyerinde üreme 68
Şekil 4.15. Gram boyamada tomurcuklanan blastosporlar (maya hücreleri) 68
Şekil 4.16. Tween identifikasyon testi, Tween 20, 40, 60 ve 80 (M. furfur) 68
Şekil 4.17. Üre testi (pozitif) 68
Şekil 4.18: %10’luk KOH ile doğrudan mikroskop incelemesi 79
( köfte-makarna)

ix
TABLOLAR D ĐZĐNĐ
Tablo no Sayfa no
Tablo 2.1. modifiye-Dixon agardaki Malassezia spp. nin koloni morfolojisi 16
Tablo 4.1. Pityriasis versicolor’lu olguların yaş gruplarına göre dağılımı 49
Tablo 4.2 Bistirü ve selofan bantla alınan klinik örneklerin 56
mantar üremesine göre dağılımı
Tablo 4.3. Đzolatların CCA, C-EL ve Tween 60 eskülin besiyerindeki 58
üreme özellikleri ile katalaz aktivitelerinin sonuçları
Tablo 4.4. Örneklerin 37°C, 40°C ve SGA besiyerindeki üreme sonuçları 61
Tablo 4.5. Tween identifikasyon testi (Tween 20, 40, 60 ve 80) 63
Tablo 4.6. Pityriasis versicolor’da klinik ve laboratuvar bulguların 65
Pigmentasyon durumuna göre dağılımı

x
KISALTMALAR
PV :Pityriasis versicolor
P :Pityrosporum
M :Malassezia
rRNA : Ribozomal Ribonükleik asit
ITS :Internal transcript sequensing
KOH :Potasyum hidroksit
DNA :Deoksiribonükleik asit
AIDS :Acquired Immune Deficiency Syndrome
PCR :Polymerase chain reaction
SD :Seboreik Dermatit
IgG :Immünglobulin G
AD :Atopik Dermatit
IgE :Immunglobulin E
MF :Malassezia follüküliti
CPR :Gougerot-Carteaud’un retikulat ve birleşik papillomları
ÇÜTF :Çukurova Üniversitesi Tıp Fakültesi
MD :Modifiye-Dixon Agar
LN :Leeming-Notman Agar
SGA :Sabouraud Glikoz Agar
CCA :Chromogenic Candida Agar
T-60 eskülin :Tween-60 eskülin
ID :Đdentifikasyon

xi
ÖZET
Pityriasis versicolor tanısında klinik örnek alma yöntemlerinin
ve çeşitli besiyeri sonuçlarının karşılaştırılması
Pityriasis versicolor (PV, tinea versicolor) daha çok üst kol, boyun gibi üst gövdede ve yüzde görülen derinin yüzeysel mantar infeksiyonudur. Bu
infeksiyonpembeden kahverengi hatta siyah rengine kadar değişen renklerde lezyonlara neden olabilir. Lezyonlarda pullanma tipiktir. Lezyonlar hipo ve hiper-pigmente olabilir. Genelde lezyonlar yuvarlak veya oval, maküler veya papüler olabilir. Bazı hastalarda hafif kaşıntı görülebilir. PV genelde asemptomatiktir ve çoğu hasta kozmetik bozukluktan şikâyet eder.
Çalışmada, PV tanılı 57 hasta incelendi. Đnfeksiyonun tanısı hastaların lezyonlarından alınan klinik örneklerinin %10’luk K OH ile incelemesinde köfte-makarna görünümü ile konuldu. Lezyonlardan bistüri ve selofan-bantla alınan klinik örnekler modifiye Dixon ve Leeming-Notman agara ekildi ve beş gün süre ile 37C°’de bekletildi. Katalaz testi, Chremophor-EL, Sabouraud glikoz agar (SGA) besiyerinde üreme durumu ve Tween-60 eskülin besiyerindeki siyah zon oluşumu değerlendirildi. Candida Chromogenic Agar (CCA) besiyerindeki pigment oluşumu ile Tween 20, 40, 60 ve 80’deki presipitasyon zonlarına göre tür tanımı yapıldı. Đzolatların 37°C ve 40°C’deki ısı toleransları incelendi. Ayrıca, izolatlara laktofenol pamuk mavisi ve gram boyamayla blastospor morfolojisi değerlendirildi. Çalı şmada, lezyonlardan alınan örneklerin KOH ile incelemesinde köfte-makarna görüntüsü belirlendi. Đzolatların hepsi katalaz pozitif idi. Klinik örnekl erin 6’sı (%10.5) hiçbir besiyerinde üremedi, 7’sinde üreme oldu, ancak çoğaltılamadı için tür tayini yapılamadı. Diğer örnekler pasajlarda üredi ve identifiye edildi. Klinik örneklerin 27’sinde M. furfur, 10’unda M. slooffiae, dördünde M.sympodialis, ikisinde M. globosa saptandı. Diğer iki örnekte atipik Tween paterni görüldü ve identifiye edilemedi. Klinik örnek alma yöntemleri karşılaştırıldı ğında bistüri yönteminin daha iyi olduğu ve daha çok kepek alınımı gerçekleştirdi ği ve daha iyi üreme olduğu görüldü. Bant yönteminin ise örnek almada yeterli olmadığı görüldü. Besiyeri performanslarına bakıldığında modifiye-Dixon besiyerinde daha çok koloni ürediği, ve üreyen kolonilerin çaplarının daha büyük olduğu görüldü. Çalışmada, Chremophor-EL ve Tween 60-eskülin testlerinin tür identifikasyonu ile uyumlu olmadığı görüldü. Anket sonuçları değerlendirildi ğinde PV’un daha çok erkeklerde ve en çok 20-30 yaş grubunda olduğu görüldü. PV’un daha çok rekürren ve her iki cinste boyun sırt, göğüste olduğu görüldü. Wood ışığı ve pigmentasyon çeşidinin tür ile bir ili şkisi olmadığı belirlendi. Anahtar kelimeler: Malassezia, Pityriasis versicolor, bistüri, selofan-bant, modifiye-Dixon agar, Leeming- Notman agar, Tween identifikasyon testi

xii
ABSTRACT
Comparison of clinical sample collection methods and different isolation media results in diagnosis of Pityriasis versicolor
Pityriasis versicolor (PV,tinea versicolor) is a superficial fungal infection of the skin, appearing most commonly on the upper trunk, as well as on the upper arms, neck and face. PV may causes in different colours, ranging from pink or tan to dark brown or even black. The lesions have a characteristic flaking appearance, although in larger lesions this can be evident only at the border of the macule, and can be hypopigmented or hyperpigmented. Generally, the lesions take the form of round or oval macules or papules, although in advanced cases the lesions may become confluent. Whereas some patients experience mild pruritus, PV is generally asymptomatic, and most patients complain primarily about the cosmetic appearance of the disease.
In the study, diagnosed that 57 PV patient included. The diagnosis of PV that sampling from PV lesions was established by the characteristic “spaghetti and meatballs” appearance. Samples from with a sterile scrapel and cellotape inoculated on modifiye-Dixon and Leeming- Notman agar. Culture were incubated at 32 °C and were waited for 5 days. After growth, catalase test was tested. Growth in Chromophor-EL, Sabouraud glicose agar was evaluted. Pigment on Chrom Candida Agar(CCA) was examined. As identification of Malassezia was used Tween test which prespitating Tween 20, 40, 60, 80 was determined. To examine blastospor morphology, lactophenol cotton blue and Gram stain was done. In the study, samples from lesions that was examined with KOH direct microscopy were showed “spaghetti and meatball” appearance. All of isolates were catalase pozitive. Six of samples did not grow on both medium. Seven of samples were not determinated because of the fact that they could’t isolated more. Other samples that were isolated well, were applied Tween asimilation test and diagnose tests. 27 of samples were M. furfur, 10 of samples were M. slooffiae, four of them M. sympodialis, two of them M. globosa and another two of them were not compatible with Tween compound.
When comparing collection of samples, scraping method were more beter then cellotape band and when using scraping method collection of dandruff were much more than other. Eventuality much contain of microorganisms were seen. Cellotape method was seen unusefull. As performing media, coloni on modifiye-Dixon media grew much more and very bigger than Leeming- Notman media. In the study, ın addition Chremophor-EL and Tween 60-eskülin test were not compatible with species identification.
When results of survery were evaluated, PV was more commen men. As an age it was commenly seen between 20-30 age. Another results of the study was that PV was more recurren and the lesions in both sex were observed commonly on neck, back and chest. Correlation wood light and pigmentation between species was not associated with. Key words: Malassezia, Pityriasis versicolor, scrapel, cellotape, modifiye-Dixon medium, Leeming- Notman medium, Tween identification test.

1
1. GĐRĐŞ ve AMAÇ
Pityriasis versicolor (PV) Malassezia maya mantarının sebep olduğu kronik
yüzeysel bir infeksiyondur. Bu hastalık bütün dünyada yaygın olarak görülür. PV,
deride hipo ve hiper–pigmentasyona sebep olur. PV vücüdun yağdan zengin olan
boyun, sırt, göğüs gibi bölgelerde görülür. Lezyonlarda kaşıntı meydana gelebilir. PV
asemptomatiktir ve kozmetik bozukluğa sebep olur. PV’nun tropikal bölgelerdeki
prevalansı % 40-60’dır. Ilık ve nemli hava bu hastalığın gelişmesinde önemli bir
faktördür. Hastalık en sık 20-30’lu yaşlarda görülür. Lezyonlardan alınan örneğin KOH
ile incelemesinde tipik “köfte- makarna”(spor ve hif) görüntüsü görülür. PV’ye sebep
olan Malassezia maya mantarı 13 türü vardır: M. furfur, M. sympodialis, M. slooffiae,
M. pachydermatis, M. globosa, M. obtusa, M. dermatis, M.japonica, M. nana, M.
resticta, M. equina, M. caprae.
Bugüne dek ülkemiz adresli çalışmalarda; Pityriasis versicolor etkenlerini
sorgulayan, bu mikro-organizmaların vücut bölgelerine, infeksiyon süresine ve nüks
durumuna göre dağılımını araştıran bir araştırma bulunmamaktadır. Bu çalışma ile
Malassezia cinsi maya mantarlarının laboratuvar tanısı, üreme ve üretilme özelliklerinin
belirlenmesi ve tür düzeyinde tanımlamada çeşitli yöntemler denenecektir. Bu amaçla
laboratuvarımızda geniş bir kolleksiyon oluşturulmuş ve standart kökenler ile kalite
kontrolünün sağlanması amaçlanmıştır.
Bu çalışmada klinik örnek alma tekniklerinin ve izolasyon amaçlı kullanılan
besiyerlerinin laboratuvar tanıda yeri tartışmaya açılacaktır. Pityriasis versicolor tanılı
lezyon bölgesinden hem steril bistüri yardımı ile epitel kazıntı örnekleri ve hem de sellofan
– bant yöntemi ile örnek alınacaktır. Bu örnekler hem Modifiye- Dixon agar’a hemde
Leeming- Notman agar’a inoküle edilecek ve 32 °C’de 7 gün inkübasyon sonunda üreme
özellikleri belirlenmiştir.

2
2. GENEL BĐLGĐLER
2.1. Tarihçe
Sinonimleri pityriasis versikolor’a eş anlamlı olarak, dermatomikosiz furfuraz,
tineaflava, liver spots, kromofitozis ve Eichstedt hastalığı kullanılmıştır1.Đnfeksiyon
başka uzun süreler değişik diskromilerle karıştırılmış, 1801'de Willan tarafından özgül
bir klinik olarak kabul edilmiş ve pityriasis versicolor ismi verilmiştir1.
Pityriasis versikolor (PV)’un etkeni Malassezia cinsi maya mantarlarıdır. Maya
hücrelerinin varlığı ilk kez 1835’de Eichstedt tarafından deri kazıntı örneklerinde
tanımlanmıştır2. Robin 1853’de PV lezyonlarından bir mantar izole etmiş ve bunun bir
dermatofit olduğunu düşünerek Microsporum furfur olarak adlandırmıştır3. Rivolta,
1873’de bir psöriasis lgusunda çift çeperli, tomurcuklu hücreleri tanımlamıştır.4
Malassez, 1874’de mikro-organizmaya ilişkin yumurtamsı ve yuvarlak hücreleri
tanımlamıştır5. Đzleyen yıllarda Bizzozero, Malassez’in sporlarına benzer şekilde hem
küresel hem de yumurtamsı şekilde tomurcuklana maya hücrelerini tanımlamıştır. Bu
organizmalar sırasıyla Saccharomyces sphaericus ve Saccharomyces ovalis olarak
adlandırılmıştır6. Sabouraud, bu maya hücrelerin değişken morfolojilerini incelemiş
maya ve miçelli şekillerin biribirleri ile ilişkili olabileceğini ileri sürmüş ve
Pityrosporum malassezi ismini önermiştir.7 Castellani ve Chalmers 1913’de ilk kültürü
yapmış ve mantar Pityrosporum ovale olarak yeniden isimlendirilmiştir.8 Weidman,
1925’de lipit bağımlı olmayan hayvanlarda kolonize olan ve insan derisinde nadir
olarak bulunan bir tür bulmuş ve Malassezia pachydermatis ismini vermiştir11.
Malassezia’nın ilk tanımlaması 1846’de yapılmasına ve birkaç araştırıcının in vitro
şartlarda mantarı üretmesine karşın, kültürde ilk başarılı izolasyonu 1927’de Panja
tarafından yapılmıştır12.
Mayanın ikinci şekli 1951’de Gordon tarafından isimlendirilmiştir. Gordon hem
lezyonlu hem de lezyonsuz bölgelerden küresel ve yumurtamsı şeklinde olan maya
hücrelerini bulmuş ve P. orbiculare ismini vermiştir9. P. orbiculare ve P.ovale’ nin aynı
mikro- organizmalar veya aynı mikro-organizmanın farklı şekli olup olmadığı uzun bir
süre tartışma konusu olmuştur. P.ovale ve P.orbiculare’ nin prevalansı farklı coğrafik

3
bölgelere göre değişiklik göstermiştir. Örneğin P.orbiculare Đsveç’ten; P.ovale
Đngiltere’den ve Amerika’dan rapor edilmiştir10. Borrelli, tropikal bölgelerde çalışmalar
yapmış ve aynı hastada Pityrosporum’un iki şeklini bulmuştur13. P. orbiculare’ nin esas
olarak göğüs, boyun, yüz ve üst bacakta; P.ovale’ nin ise belden aşağıda olan gluteal
bölgeden izole etmiştir14,15. Borrelli, P.ovale’ nin aynı zamanda yaşlı popülasyonlarda
daha az görüldüğünü belirtmiştir. Bununla beraber, Borelli, PV‘un iki şekli olduğunu
söylemiştir. Bunlardan biri M. furfur diğeri P.ovale’dir. Mikolojik, mikroskobik,
immünolojik çalışmalar; M. furfur ve M. ovalis aynı mayanın morfolojik varyantı
olduğunu ve belki de kompleks hayat siklusunun farklı evresini gösterdiğini işaret
etmiştir. Benham 1939’da organizmanın üretilmesi için yağ maddesine gereksinimi
olduğunu görmüş ve kültürde üretilmesinin zor olduğunu açıklamıştır17. Ayrıca lipit
gereksinimin belirlenmesinden sonra çeşitli besiyerlerinin formülasyonu için
kolaylaştırıcı bir yol olmuştur ve bu organizma kolayca izole edilmiş ve saklanabilmiştir 18,19.
Đlk resmi taksonomik sınıflandırma olarak Pityrosporum cinsi içinde yapılmıştır
ve iki tür tanımlanmıştır: P. ovale ve hayvanlarda bulunan P. pachydermatis20. Gordon
yuvarlak hücre çeperine dayanarak P. orbiculare’ yi eklemiştir 21. 1970‘lere kadar üç tür
bilinmekteydi; P. ovale, P. orbiculare ve P. pachydermatis. Maya ve miselli şekiller
arasında ilişki bulunduğu kabul edilmekle beraber faz dönüşümü hiç gösterilmedi ve iki ayrı
cins olarak sürdürüldü22. Bu karışık durum 1977 yılında çözümlendi. Birbirinden bağımsız
üç grup araştırıcı in vitro şartlarda maya hücrelerinin misel üretmesini başardılar23-25.
Çeşitli kültür koşullarını kullanarak PV’de görülenlerden ayırt edilemeyen hifler
ürettiler.
Hem yuvarlak hem de yumurtamsı hücrelerin hif üretebildiği gözlemlendiğinden yuvarlak ve
yumurtamsı hücrelerle, hiflerin aynı tek bir organizmanın yaşam çevrimindeki stratejiler
olduğuna işaret ettiği ortaya konuldu25. 1986 yılında maya formundan misel formuna
dönüşümü saptanmasıyla ile M. furfur (Robin Baillon(P.orbiculare, P.ovale ve
Malassezia furfur) ve Malassezia pachydermatis (P.pachydermatis) isimlerini
kapsamak üzere bu iki cins birleştirilmi ştir26.Birçok araştırıcı türlerin koloni
morfolojilerinin ve hücre yapılarının farklı olmasına rağmen P.ovale ve P. orbiculare
ismini kullanmaya devam etmişlerdir27.1990 yılında Simmon ve Guého yeni bir lipofilik
tür olan, Malassezia furfur’a oranla daha düşük G+S içeriğine ve simpodiyal
tomurcuklanmasına dayanarak Malassezia sympodialis’i tarif ettiler29. Cunningham ve

4
ark. farklı yüzey antijenleri ile belirlenen, farklı ekolojik özelliklerine karşılık gelen kültür ve
morfoloji özelliklerine göre A, B, C olarak üç serovar ayırt ettiler30. Fakat 1990 yılının
başlarında Malassezia cinsinin sınıflaması hala karışıktı.1995 yılında Simmon ve Gueho’ nun
yayınladıkları yeni bir çalışma ile bu karışıklık sona erdi31. Bu araştırıcılar farklı araştırma
gruplarının farklı şemalarda sınıflandırdığı Malassezia türlerinden 104 tane toplamışlar ve
bunların büyük alt birim rRNA ve nükleer DNA komplementerlerinin sekanslama çalışmaları
ile sonuçlanan fizyoloji, morfoloji ve ultrastrüktürel özellikleri ile birleşerek bilinen üç türe
ilaveten, dört yeni Malassezia türünü tanımladılar. Bu yedi türler: M. furfur, M. sympodialis,
M. obtusa, M. globosa, M. restricta, M. slooffiae, and M. pachydermatis’dir32. Bu
türler başka taksonomistlerce de doğrulandı34-37. Cinsin taksonmisini incelemek için birçok
moleküler yöntemler geliştirilmi ştir ve birçoğu Guillot tarafından tanımlanan ayrı yedi türün
varlığını doğrulamıştır. Karyotipleme, kitin sentez 2 genini dizi analizi, mitokondrial rRNA’ nın
büyük alt biriminin dizi analizi, ITS veya rDNA’ nın dizi analizi, gradient jel elektroforez gibi
birçok moleküler yöntemler bu ayrı yedi türü doğrulamaktadır. Saadatzadeh et al.
(Malassezia türlerini Malassezia furfur serovar A olarak sınıflamıştır38. Ardından,
Sugita ve ark2002’de atopik dermatitli hastalardan ayırdıkları kökenlerden yeni bir tür
M. dermatis’ i tanımladılar39. Yine yakın tarihlerde Sugita ve ark. tarafından sağlıklı bir
japonun deri mikroflorasından Malassezia japonica’ yı izole ettiler402004 yılında Hirai
ve ark. Japonya ve Brezilya’da hayvanlardan ayırdıkları bir grup kökeni fenotipik ve genotipik
yöntemlerle tanımlayarak M. nana yeni türünü tanımlamışlardır41. Aynı yıl Sugita ve ark.
Japonya’ da seboreik dermatitli bir hastadan M. yamatoensis’ i tarif ettiler42. Halen Malassezia
cinsi 13 tür içermektedir; M. japonica, M. globosa, M. restricta, M. obtusa, M.
slooffiae, M. furfur, M. nana, M. sympodialis, M. dermatis, M. pachydermatis, M.
yamatoensis, M.equina, M. caprae. Bunlardan onikisi zorunlu lipofilik olup M.
pachydermatis zorunlu lipofilik bir tür değildir, insan derisinde seyrek olarak
kolonizasyon yapabilmektedir ve birkaç hayvan deri hastalığıyla ilişkilidir.
Birçok çalışmada tanınan türlerin alt grupları da bulunmuştur44(M. furfur45, M.
sympodialis46, M.pachydermatis47). Malassezia cinsinin yeniden sınıflandırılması
önceki yöntemlerle sınıflandırılmış kökenlere dayanan birçok çalışmanın tekrarlanması
gereğini birlikte getirdi. Bir kısım çalışma kökenlerinin halen stok kültür
koleksiyonlarında bulunduğu bilinmektedir. Saadatzadeh38. M. furfur serovar A
kökenlerinin M. sympodialis’e karşılık gelmesi gerekirken her zaman böyle olmadığını
buldu. Böylece M. furfur serovar A, B ve C’nin M. sympodialis, M. globosa,

5
M.restricta’ya karşı geldiği sonucunun da her zaman doğru olmadığı ortaya kondu.
Önceki çalışmalarda kullanılan spesifik suşlar test edilmiştir ve yeni bir taksonomik
sınıflandırma içine alınmıştır. Eski ve yeni sınıflandırma içinde olan taksonmik suşlar
arasındaki direkt ilişki uyumlu olmayabilir. Bu problem kültürde saklanan türlerde daha
çok belirtilir. Amerika’daki kültür saklama koleksiyonundan 42132 suş P.orbiculare
(sinonimi M. globosa) olarak saklanmıştır. Halen, koleksiyonlardaki kökenler yeniden
taksonomik olarak incelenmekte, ayrıca bu türlerin hastalık etkeni veya hastalığı
tetikleyici olarak rolleri araştırılmaktadır.
2.2 Filogeni ve Taksonomi
Malassezia cinsinin bazı türlerinin telemorfu bulunamamasına karşın
basidiomycetes sınıfının karekterlerini taşır. Malassezia cinsi çok tabakalı hücre
duvarına sahiptir. Tekrarlayan tek kutuplu tomurcuklanma yapar. Üreyi hidroliz etme
yeteneği vardır. Diazonyum B mavisi ile kırmızı renk verir.
Ribozomal RNA dizi analizi gibi yeni moleküler yöntemlere geçişle beraber
araştırıcılar yedi tane tür identifiye edilmiştir48. Buna ek olarak bilinen türler Malassezia
furfur, M. pachydermatis, M. sympodialis, iken M. globosa, M. Obtusa, M. restricta ve
M. slooffiae cinse eklenmiştir. Tüm türler fizyolojik ve morfolojik karakterlere
sahiptir49
Sonra Malassezia’nın yedi türünün de filogenetik olarak Basidiomycota sınıfında
olduğu doğrulanmıştır. Bu türler bir kökenden türemiştir. Malasseziales alt sınıfında,
Ustilaginomycetes sınıfında yer alır. Bununla beraber sürprizdir ki bu köken
Exobasidium ve Tilletia olarak bilinen ve bitkilerde patojen olan cinslerle, insan ve
hayvanlarda patojen olan Trichosposporonales ve Tremellales sınıflarında yer alan
Cryptococcus neoformans ve Trichosporon’dan çok daha yakın akrabadır51.
Alem: Fungi
Bölüm: Basidiomycota
Alt bölüm: Agaricomycotina
Sınıf: Heterobasidiomycetes

6
Alt sınıf: Malasseziales
Ordo: Tremellales
Cins: Malassezia
Bu yedi tür medikal laboratuvarlarda rutinde kolayca morfolojik ve fizyolojik
olarak identifiye edilebilir51-53. Şüphesiz ki sequensing (dizi analizi) türlerdeki
karışıklığı ortadan kaldırmıştır54.Üstelik dizi analizinin güvenilir olmasından dolayı,
Malassezia maya mantarının moleküler identifikasyon için birkaç teknikler
geliştirilebilir.
Đdentifikasyondaki bu farklı yöntemler birkaç çalışmaya yol açmıştır ki sağlıklı ve
hastalıklı derilerin her ikisinden de farklı türler arasındaki yakın ilişkiler de
araştırılmıştır. Bu çalışmalardan bazılarında şu sonuç çıkmıştır; M. globosa PV’de esas
türdür. Bu sadece kutanöz hastalıktır ki Malassezia‘nın ili şkisi tartışılmazdır. Ancak bu
türler normal florada da Malassezia furfur, M. sympodialis, M. restricta türlerinde biri
veya karışık olarak ta bulunabilir55( Sugita et al). Malassezia maya mantarı ile ilgili
dermatolojik bozukluklardaki rolü tartışmalıdır. Seboreik dermatit, atopik dermatit,
follükülit gibi hastalıklardaki çalışmalar immunolojik olarak odaklanmıştır. Bu da
patojenik mekanizmayı açıklayabilir56,57,58,59,60,61,62(Ashbee et al 1994a,b;Devos and van
der valk 2000; Kesavan et al 2000;Koyama et al 2000;Tengvall Linder et al
2000;Faergemann et al.2001;Asbee and Evans2002;Faergermann et al 2001). Confluent
retikulat, papilomatosis, yeni doğan püstülosis, psöriasis, bilefarit, otitis ve
onikomikozis gibi hastalıklarda Malassezia’nın varlığı ve önemi hala tartışmada
sorundur.

7
2.3. Morfoloji
Malassezia maya mantarı dimorfiktir, saprofit ve parazitik miçel formu vardır.
Malassezia hem maya hem hif şeklinde bulunabilen, normal deride ekseri maya şekli ile
karşılaşılan bir mantardır. Bazı kökenlerde hifle karşılaşılsa da bu şekil kültürde de önde gelen
şekildir49,63.Çeşitli besiyerlerinde124,25,64in vitro miselli şekile dönüşüm gösterilebilir, bazı
kökenler dönüşüm yeteneğinden yoksundur.38 Eşeysiz üreme tek kutuplu enteroblastik
tomurcuklanma iledir; ana hücre yavru hücreden bir bölme ile ayrılır. Tomurcuk ana hücreden
fission ile (transversal yönde bölme duvarı oluşturarak) ayrılır ve ana hücrede belirgin bir
kopma yeri izi kalır65. Tomurcuğun geniş dipli62 olması karakteristiktir, ancak dar da olabilir66.
Kültürler karakteristik olarak meyve kokar25. Sekiz Malassezia türü tipik morfoloji ve fizyoloji
özellikleri göstermektedir49,51. Maya hücreleri küremsi, yumurtamsı, şişe veya silindir biçimli
olabilir. Cinsin en önemli özelliği en iç yüzeyi dalgalı olan çok katlı kalın bir hücre duvarına
sahip olmasıdır. Protoplazma zarı hücre duvarının iç yüzüne sıkıca bağlıdır. Hücre duvarının
başlıca bileşenleri mannoproteinler (%75-80), lipidler (%15-20) ve kitin 67(%1-2)’dir.
2.4 Fizyoloji
Malassezia’nın fizyolojik özellikleri, kültür zorlukları sebebiyle tam olarak
açıklanamamıştır. Benham 1939’da şekerleri fermente etmediğini bildirmiştir68.Mantar, lipidi
tek karbon kaynağı olarak kullanabilir68,vitamin gereksinimi yoktur69. In vitro aerop koşullarda
normal olarak üreyebildiği gibi mikroaerofilik ve anaerop koşullarda da üreyebilir70. Lipidleri
tek karbon kaynağı olarak kullanabildiklerinden M. pachydermatis dışındaki türler lipofiliktir.
Uzun zincirli yağ asitlerini oluşturamaz ve üreme sırasında mutlak gereksinim duyduğu yağ
asitlerini enerji için olmaktan çok metabolize etmeden doğrudan hücre lipidlerinin yerine
koyar62. Normal insan saçlı derisinde bulunan yağların mantarın lipid isteğini yerine koyabildiği
gösterilmiştir62. Yapılan çalışmalarda yağ asitlerindeki C zincirinin uzunluğu 10’dan
fazla olması üremeyi desteklediğini rapor etmişlerdir ve zincir uzunluğunun tek tek
veya artarak olup olmadığını sorun olmadığını vurgulamışlardır71.Büyüme sırasında
kullanılan lipit kaynağı organizmanın bileşimindeki yağ asitlerini etkiler. Yağ asitleri
enerji kaynağı olarak kullanılmaz. Bunlar metabolize edilmeksizin direkt olarak
hücresel lipitlerle birleştirilir 72. Wilde ve Stewart insanların normal kafa derisindeki lipit
varlığının organizmanın lipit gereksinimini doldurbildiğini buldular. Malassezia türleri

8
birçok enzimleri metabolize eder. Đn vitro ve in vivo şartlarda lipolitik aktivitesi vardır
ve lipaz üretimi belirtilmiştir. Lipaz hücre duvarında lokalizedir ve /veya sitoplâzma
membrana yerleşmiştir73,74. Ran ve ark. optimum ph‘nın 5.0 olduğunu bulmuşlar ve
lipaz üretiminin üremenin logaritmik fazı sırasında en fazla olduğunu ve bunun substrat
konsantrasyonu ile ilişkili olduğunu buldular. Bu araştırıcılar Malassezia’nın üremesi
için en az üç lipite ihtiyacı olduğunu buldular. Malassezia in vitro da ayrıca fosfolipaz
üretir75. Bu fosfolipaz aktivitesi Hep-2 hücresindeki arakidonik asitin yeniden
metabolitleri deride inflamasyonu içerir76. Bu durum Malassezia’yı bir mekanizma yolu
harekete geçirir. Malassezia türleri lipoksigenaz aktivitesi ile bir enzim üretir. Bu olay
kolestrol, skualen, doymamuş ve serbest yağ asitlerini okside ederek bu aktivitesi
gösterilmiştir24. Lipoperoksidazların üretimi hücre zarına zarar verir ve sonuç olarak
derideki pigmentasyon değişikli ğine sebep olur77. Malassezia tipik olarak meyve
kokusu verir. Bu ilk olarak Van Abbe tarafından tanımlanmıştır78.Lipit içeren
besiyerinde Malassezia’dan gelen gazın uçucu gamma laktonlar oluşturduğunu
göstermiştir. Bu karekter Malassezia için tektir ve diğerlerinden bu genusu ayırmak için
mümkün olan bir yoldur. Bir başka metabolitte azeleik astittir. Azeleik asit dikarboksilit
asit üretir. Malassezia oleik asit varlığında üretildiği zaman tirozinaz enziminin
inhibitörünü üretir. Bu enzim melanin üretiminde görev alır79. Azeleik asit birkaç tümör
hücresinin metabolizmasını inhibe ederek nötrofillerdeki reaktif oksijen üretimini
azaltır80. Mayser ve ark81., M. furfur, M. sympodialis, M. globosa, M. obtusa, M. restricta,
M. slooffiae, ve M. pachydermatis kökenlerini, başlıca nitrojen kaynağı olarak triptofan
içeren besiyerinde pigment ve florokrom oluşturmaları bakımından incelemişler;
yalnızca M. furfur kökenlerinin bu özelliğe sahip olduklarını; bunun dışındaki tüm
lipofilik türlerin bu besiyerinde üremediğini, oysa triptofan yerine pepton içeren Dixon
agar (DA)’da hepsinin geliştiğini ve hızla kahverengi bir hale oluşturduklarını
gözlemlemişlerdir.
2.5. Yapı
Malassezia türleri monopolar üreme yoluyla seksüel olmayan bir bölünme geçirir ve
geniş karekteristik bir tomurcuklanma yapar. Anna ve kız evlad hücre bir septum (bölme)
yoluyla ayrılır. Malassezia cinsinin hücre duvarı zayıf bir şekilde belirtilmiştir, diğer birçok

9
mayalar gibi bileşimi çok kalındır (0,12 µm) ve hücre hacminin % 26 -37’ye hadar
oluşmaktadır82. Hücre duvarının major bileşeni şeker (%70), protein (%10), lipid (%15-20) ve
az miktarda nitrojen ve sülfürdür83,84. Bir çok araştırıcı hücre duvarının iki tabakadan
oluştuğunu söylerken, diğer araştırıcılar kompleks bir duvarın olduğunu bulmuşlardır85,86.
Ayrıca önceki çalışmalarda hücre duvarınınetrafında dış lameller tabaknın olduğunu
söylerler87,88. Lameller tabakası membran benzeri yapıdadır. Lameller yapı insan derisine invaze
olmasına hemde katatere kolonize olmasına yardımcı olur89. Sitoplazmik membran hücre
yüzeyüne yakından bağlanır ve körfezi takib eder90,89,86,91. Her bir hücrenin mitokondrisin şekli
ve sayısı değişkenlik gösterebilir, yuvarlak ve oval şekilde olabilir82. Çekirdekte iyi tanımlanmış
bir membran vardır ve granüler homojen bir nükleoplazma ile çevrilidir. Vakuoler hücre içinde
lipitle çevrilmiştir ve hücrenin yapısına göre büyüklüğü değişkenlik gösterir90.
2.6. Biyokimya
Malassezia türleri zengin enzimlere ve metabolitlere sahiptir. Mantar hem in vitro62 hem in
vivo 73,92 lipaz üretimine işaret eden lipolitik aktivite gösterir. Lipaz hücre duvarında ve/veya
sitoplazma zarında yerleşmiştir73,93. Malassezia türleri in vitro fosfolipaz da üretirler96. Ayrıca
antibakteriyel94 ve antifungal95 özellik gösteren metabolitlere de sahiptir.
2.7. Malassezia’nın Karekteristi ği
2.7.1.Đzoloasyon ve Đn vivo da Đdentifikasyon
Malassezia maya mantarının sebep olduğu kutanöz patoloji yeterince
karekteristiktir ki dermatologlar tarafından kolayca tanınır. Genelde kültürde üretmek
için ekipmanlara ihtiyaç olmadığından dolayı dermatologlar lezyonlardan mantarı izole
etmezler fakat sık sık lezyonlardan alınan materyalden direk mikroskobik incelemeyle
mantarın varlığını doğrularlar. Aslında mayalar dokular arasında çok küçük görünür
fakat tekrarlayan tek kutuplu tomurcuklanma ana hücrede çok tipik kalıcı bir iz (skar)
bırakır. Bu olguda görünen en tatmin edici metod mürekkeple (Parker Kalıcı mavi ve
siyah boyası) boyandığı zaman görünür. Eşit şartlarda KOH ile % 20-30 oranında

10
birleştirilir 97. Malassezia’nın hücreleri (maya ve hif) hemen boyayı alır ve beyaz
zeminde mavi görünür. Daha selektif boyama olarak kalkaflor beyazı kullanılır. Bu
boya sadece floresan mikoskobunda görünür. Bu teknik seboreik dermatitte güzel bir
görüntü sağlar. Dokulardaki mantarın görünümü maya görünümündedir. Hiflerin
varlığı, yokluğu ve sıklığı hastalık hakkında bilgi verir. Sadece M. furfur insanlarda98
(Yarrow ve Ahearn 1984) ve hayvanlarda, M. pachydermatis tanımlandığı zaman
kültürden daha çok izolasyon gerekli değildi. Malassezia’ nın patolojisindeki ilginçliğin
yeniden tanımlanmasıyla, birkaç lipit bağımlı Malassezia türleri insanlarda ve
hayvanlarda görülmeye başlandı. Ve bunlar sağlıklı ve hastalıklı deriden izole edildi.
Yıllarca Malassezia lipofilik maya mantarı derideki kepeklerden, tırnaktan, diğer
infekte materyallerden Sabouraud besi yerinde ( %2glukoz, % 1pepton, % 2agar ) veya
zeytinyağı eklenmiş Sabouraud besiyerinde üretildi. Her iki besiyerinde bakteri ve maya
kontaminasyonunu önlemek için %0.5 kloramfenikol ve %0.5 siklohekzimit ilave
edildi. Sabouraud besiyeri lipofilikleri izole etmek için yeterliydi. Fakat lipit bağımlı
olmayan türler yani Malassezia pachydermatis yeterli değildi. Oysaki aynı besiyerine %
1 zeytinyağı ilave edilirse, lipit bağımlı M. furfur’un izolasyonu için çok kullanışlıydı99
(Gueho ve Meyer 1989). Taksonomi incelemede zeytinyağı birkaç tür için toksik
olduğunu vurgulamıştır. Bundan dolayı bu besiyeri çok seçicidir ama çok uzun sürede
kullanılamaz. Bu iki besiyeri şu anda Malassezia türlerini üretmek için geniş ölçüde
kullanılmaktadır. Dikson besiyeri 1964’te Van Abbe tarafından tanımlanmış ve formüle
edilmiştir. Bu formül; % 3.6 malt ekstrat, % 0.6 pepton, % 2.0 oksbile,% 1 tween 40, %
2 gliserol, % 0.2 oleik asit ve %1.2 agar. 1987’de Leeming ve Notman besiyeri
tanımlamışlardır. Malassezia sıcakkanlı hayvanların normal flora üyesinin bir parçası
olduğu için bu besiyerine kloramfenikol ve siklohekzimit ilave edilmesi tavsiye edilir.
Modifiye diskon ve Leeming- Notman besiyerlerinin avantajı koyu renkleriyle koloni
sayımına kolaylık sağlar. Özelliklede birkaç tür karışık olduğu zaman morfolojilerini
daha iyi tanımlanmasının sağlar. Önce kültürler sistematik olarak 37°C de inkübe
ediliyordu. Yapılan çalışmalarda zeytinyağının 37°C de birkaç türü inhibe ettiğini
göstermiştir. Böylece bütün kültürler simdi laboratuar özelliklerine göre 30 ile 35°C
lerde inkübe edilmektedir. Üremenin maksimum sıcaklığını göstermenin dışında 37°C
ve yukarısı asla kullanılmamaktadır.

11
2.7.2. Đn vitro da Đdentifikasyon
Malassezia’nın yedi türü genetik olarak açıkça ayrılmıştır100( Guillot ve Guého
1995). Bununla beraber karekterlerine, vücuttaki yerlerine, koloni morfolojilerine,
üreme sıcaklıklarına, Tween asimilasyonlarına, cremophor-EL kullanma ve β-
glukosidaz (eskülini ayırma) aktivitelerine dayanarak bazı mikrobiyoloji
laboratuarlarında rutin teknikler kullanarak kolaylıkla ayrılabilir.
Taksonomide101(Guého 1996) çeşitli konsantrasyonlarda olan Tween’deki (polietilen
sorbitan yağları) üremeleri değerlendirilmiştir. Çalışmalarda maksimum üreme ve
katalaz aktivitesine bakılmıştır. Kolaylaştırılan identifikasyon metodu Tween ilave
edilmiş Sabouraud agar plaklarındaki sonuçları ile anlaşılmıştır51(guillot).
Đdentifikasyon şeması Mayser ve ark. tarafından geliştirilmi ştir52. Sabouraud difüzyon
plaklarına ilave edilen cremophor EL ve eskülin kullanılarak bakılan β-glukosidaz
aktivitesinin değerlendirilmesini desteklemiştir. Tamamlanan identifikasyon sistemi
Guého ve ark. tarafından gösterilmiştir. Malassezia türlerini tanımlamak için birçok
yaklaşımlar olmuştur. Kültür odaklı meteodlar, guanin sitozin yüzdesini, DNA‘yı,
hücre morfolojisini, farklı non-iyonik Tween deterjanları, katalaz varlığı, üreme
sıcaklığı, β-glukozidaz aktivitesini, eskülin ayırımını ve kromofor EL’de üreme gibi
paremetreleri içerir50,101,102,103,52.
Malassezia furfur kalın, konveks, yumru şeklinde ve 4-5 mm çapında olan
kolonileri vardır. Yapısı yumuşaktır ve hücreleri kolaylıkla emülsiyon yaparlar. Bu
durum bütün türler için böyle değildir. Tür morfolojik olarak heterojendir ki
yumurtamsı, yuvarlak (globose) ve silindirik olabilir. 4.0-5.0 ve 6.0 µm uzunluğundadır.
Bazı suşlar kendiliğinden veya kültür koşulları altında hif oluşturabilir. Bu hifler
mantarın doliporlarının olmadığından beri pseudohif olarak bilinmektedir ve bunlar
Basidiomycetes içindeki gerçek hifleri kapatır. Bu tür genellikle genetik olarak yüksek
oranda ribozomal RNA ‘ya benzerdir.
Fakat iki tanede karyotipi vardır103(Boekhout ve ark.1998). Buna karşılık M. furfur
fizyolojik olarak homojendir. Bütün karekterleri ile beraber her zaman identifiye
edilebilir. Bu karekterler şunlardır: 37°C de ürer, güçlü katalaz aktivitesi vardır, β-
glukosidaz aktivitesi zayıftır veya hiç yoktur, Tween 20, 40, 60, 80 ‘i eşit olarak asimile
eder ve chremophor-EL gibi lipit bileşiğini kullanır. Gerçekte M. furfur’un üremesi için

12
birkaç lipitin yeterli olduğu için lipit bağımlılığı zayıftır. Mayser ve ark52. kastor yağı ve
risinoleik asit gibi yağların M. furfur’un üremesi için yeterli olduğunu vurgulamıştır.
2002 de Murai ve ark.104 M. furfur’un glisinin üreme üzerinde en az etkisi olan tek tür
olduğunu göstermişlerdir. Spesifik karekterler bütün türler için sabittir. Ancak atipik
varyantlar meydana gelebilir. Örneğin M. furfur köpek izolatı gibi. Bu varyant rDNA
dizi analizi yoluyla doğrulanmıştır fakat chremophor-EL de üremez54. Türün normal
izolatları düzenlidir fakat fazla miktarda hayvanlardan izole edilmez105,106. M.furfur
doymuş yağ asitlerini 20 karbonlu yağ asitlerinden daha çok ihtiyaç duyar71.
Malassezia pachydermatis tarihsel olarak tanımlanan ikinci türdür. Rutin
kullanılan zengin besiyerinde107(Sabouraud ve mikosel) üreyebilir. M. pacyhdermatis
mDixon besiyerinde de ürer, kolonileri kalın, krem renginde, mat bir yüzeyde konveks
ve genellikle nazik bir yapısı vardır bu yüzden emülsiyonu zordur. Hücreleri küçüktür
ve 2.5-4.0 µm çapında, yumurtamsı veya silindiriktir. Ayrıca tomurcukları geniştir.
Midgley108 1998’de hayvanlardan pembe izolatlar rapor etmiştir. Köpeklerden izole
edilen bu izolatların küçük kolonileri olduğunu ve kültürde lipit bağımlı olduğunu
göstermiştir. Duarte ve ark54.(2002) M. pachydermatis’in atipik lipit bağımlı olan
suşunu izole etmiştir ki bu atopik suş transferden sonra lipit bağımlı olarak kalmıştır. Bu
yüzden Malassezia’nın insan ve hayvan kaynaklı bütün epidemiyolojik kaynakları lipit
içerikli besiyerini kullanmalıdır. Taksonomik inceleme tek bir genetik türü
korumaktadır100. Fakat karekterlerin birçoğu değişebilmektedir konakçı içindeki
spesifiteye adaptasyon olabilir109. Bütün izolatlar 37° C de ürer ama farklılıkları katalaz
aktivitesiyle, β-glukosidaz aktivitesi, chremophor-EL deki üreme özelliği ve Tween
20,40, 60, 80 asimilasyonu ile meydana gelir110. Bu bileşiklerden özelliklede Tween 20
ve kromophor-EL bu türün üremesini inhibe eder. M.pachydermatis epidemik sepsise
sebep olsada insanlarda nadir rastlanır ve genellikle prematürlerde komplikasyona sebep
olur111(Mickelsen ve ark.) Đnsanların derilerinden izole edilmesi tesadüf olarak
düşünülmektedir. Bunun aksine bu tür sıcakkanlı hayvanlarda özellikle köpeklerde
kulak iltihabı ve seboreik dermatitten sorumludur112.

13
M. sympodialis identifiye edilmesi kolay olan bir türdür. Şekli geniştir ki çapı 5-
6 µm çapındadır. Kolonileri düzdür ve sık sık bir merkezden yükselir. Bunlar krem
renklidir, yumuşak ve parlak bir yüzeyi vardır. Yapısı homojendir ve emülsiyon
yapması çok kolaydır. Hücreleri küçüktür ve 2.5 – 5.0 µm, şekli ovaldir. Aynı zamanda
simpodiyal tomurcuklanmanın bir sonunu olarak maya hücresinin bazıları “yonca
yaprağı” şeklinde kalabilir. Bu tür Malassezia furfur serovar A ile ilişkilidir 113.37°C’de
ürer ve rutinde β-glukosidaz aktivitesi ile kolaylıkla karekteriz edilir. Eskülin
besiyerinde 37°C de 24 saatte siyah zon yapar ve Tween 40, 60 ve 80 de asimile eder.
Ancak Tween 20’yi asimile edemez. Kromofor aktivitesi zayıftır. Đlk izolatlar kromofor
kaynağından uzak olan bölgelerde küçük koloniler meydana getirebilirler.
Sugita ve ark114. 2002 de atopik dermatitli olan bir hastadan yeni bir tür olan
Malassezia dermais’i izole ettiler. 26 S lik ribozamal DNA analizi ve ITS( iç trankripte
bölgeleri) dizi analizi bu türün M. sympodialis ile yakından ilişkili olduğunu
görülmüştür. Bu iki türün rDNA’ları sadece %1.2 oranında farklıdır. Bunların koloni
özellikleri arasında tek bir farklılık vardır115.Fakat onlar basit laboratuar testleri ile
ayrılamayabilirler5. M. dermatis gibi M. sympodialis güçlü katalaz aktivitesi vardır.
Dört çeşit Tween’i de asimile eder. Bununla beraber eskülin ve kromofor EL
besiyerlerinde üremesi zayıftır. Aynı şekilde atlardaki Malassezia izolatları da rDNA
dizi analizi ile bulunmuştur ki bunlar yeni iki yeni tür gösterilirler116. Malassezia
equi’nin yeni bir türlerden biri olarak adlandırılmasına karşılık, kâğıt üzerinde henüz
tanımlanmamıştır. M. dermatis gibi bu hayvansal kökenli (zoofilik) türler filogenetik
olarak M. sympodialis ile yakından ilişkilidir. Birçok çalışma ile bunların daha iyi
tanımlanmasına olanak verecektir. M. nana hayvanlardan izole edilen başka yeni bir
türdür, M. sympodiyalis’e benzemektedir. Fakat farklı olarak eskülini
ayıramazlar117(hirai ve ark2004). Sensu stricto türü kedilerde dış kulak yolu iltihabı
(otitis eksterna) ile ilişkili olduğu bulunmuştur 118.
M. globosa morfolojik olarak Gordon tarafından tanımlanan P.orbiculare’nin
orijinal tanımlaması ile benzerdir119. Bu spesifik isim korunmamıştır. Çünkü geçmişte
P.orbiculare’nin M. globosa ile veya diğer lipit bağımlı türlerle ilişkili olduğunu
belirten yayınlar gösterilmiştir. Đmmünolojik olarak M. globosa, serovar B ile
ili şkilidir 1. Bu tür tanınabilen koloniler üretir ki 4 mm çapındadır. Yüzeyi pürtüklü ve

14
buruşuk, krem rengindedir. Yapısı kırılgan, özelliklede emülsiyonu zordur. Hücreleri
küre biçiminde; bazılarının 6 ile 8 µm çapında tomurcukları vardır. PV lezyonlarında,
bu maya düzenli olarak yalancı hifler (pseudohif) oluştururlar. Bunlar çoğunlukla ilk
kültürlerde jerm tüp gibidir. Fakat birkaç pasajdan sonra bu hifler görünmezler. M.
globosa 37°C de üremez veya çok zayıf ürer, eskülini ayırmaz ve herhangi bir lipit
eklenmiş besiyerlerinde üremez. Taze izolatlar Tween varlığında özelliklede Tween 20
çok küçük koloniler üretebilirler. M. globosa çok sık olarak M. sympodialis’le ili şkilidir
fakat M. furfur, M. slooffiae ve M. restricta ile de ilişkilidir 105. Bazen geviş getiren
hayvanlardan ve atlardan izole edilmiştir106.
M. obtusa morfolojik olarak M. furfur ile karıştırılabilir fakat fizyolojik olarak
değil. Bu tür yavaş ürer. Böylece koloniler küçük çaplı ve nemli olur. Bunun aksine
silindirik hücreler cinsin içersinde en büyük kolonilerdi ki ana hücreye bağlandığı
zaman 10 µm çapındadır. M.obtusa 37°C de üremez. M. globosa ve M. restricta gibi
beş lipitlerin bazılarını identifikasyon kriteri olarak kullanır. Fakat güçlü β-glukosidaz
aktivitesi vardır. Şu ana kadar bilinen en az rastlanan türdür ki sadece sağlıklı insan
derisinden görülür. Hayvanlarda M. furfur ile köpeklerde kulak iltihabı olgusundan120ve
sağlıklı atlar ve keçilerden106 izole edilmiştir.
M. restricta, P.ovale’nin bir formu olabilir fakat eski literatürle ile bağlantısı
azdır. Restricta ismi onun zayıf perfomansından dolayı verilmiştir. Çok yavaş ürer,
kolonisi küçüktür (2mm çapında), krem rengindedir ve zor bir yapısı vardır.
Emülsiyonu zordur. Küreselden yumurtamsı kadar değişen hücreleri vardır ve 2.0-4.0
µm çapındadır. Mikromorfolojisi M. sympodialis’e benzer fakat makromorfolojileri
farklıdır. Bu yeni türler serovar C ile ilişkili olabilir. Üçüncü serotip Cunningham ve
ark. 113(1990) tarafından lipit bağımlı Malassezia maya mantarı tarafından
tanımlanmıştır. Bu noktada serotipi belirtmek için önemlidir. Aspiroz ve ark. 121(1999)
biyotiplemesini önererek sadece üç tür tanımlamıştır. Bunlar: M. sympodialis, M.
restricta, M.globosa. Diğerlerini (M. furfur ve M. slooffiae) önemsememiştir.
Đdentifikasyonda ilginç bir durum vardır ki lipit bağımlı olduğu halde katalaz aktivitesi
negatiftir. Bu test ilk identifikasyonda özellikle de kafadan çok sayıda sınırlı Malassezia
kolonisi elde edildiğinde ilk adım olarak tavsiye edilir. Ek olarak bu tür 37°C de
üremez, eskülini ayırmaz, Tween ve cromophor varlığında üremez. Bu tür M. obtusa

15
gibi ilk defa evcil hayvanlardan izole edilmiştir122.Bu türün sayımındaki büyük
zorluktan dolayı, insanlardaki ve belki hayvanlardaki varlığı muhtemelen azdır. Ayrıca
izolasyondaki birçok zorluklar, yeterli kültür elde etmede ve korumada seboreik
dermatit, atopik dermatit gibi birçok dermatolojik hastalıkları anlamada bir engel
olmaktadır.
M. slooffiae, dört yeni türden birisidir ve daha az sıklıkla insanlardan izole edilir
fakat düzenli olarak domuzların derisiyle ilgilidir. Morfolojik olarak M. furfur gibi
yanlış tanımlanabilir. Kolonilerin kenarları katlanmış, 3 mm çapında krem renklidir ve
hassas bir yapısı vardır. Hücreleri küçük, silindirik, 1.5-3.5 µm uzunluğundadır. Asla
küresel, uzun veya Malassezia furfur gibi değişken değildir. Bununla beraber fizyolojik
olarak identifikasyonda belirsizliği yoktur. Türün katalaz aktivitesi vardır ve M. furfur
gibi 40°C ye kadar ürer. Fakat eskülini ayırmaz, Tween 80 de üremesi zayıftır ve
chremophor-EL de üremez. M. slooffiae sağlıklı insan derisinde bulunabilir. Ama her
zaman deride az sıklıkla bulunur. Đnsanlarda ana patojen olması muhtemel değildi.

16
Tablo 2.1.: Dixon agardaki Malassezia spp. nin koloni morfolojisi
2.8. Malassezia’ nın Ekolojisi
Malassezia’nın üremesi ve gelişmesi için kesinlikle lipite ihtiyacı vardır ve
Malassezia mezofiliktir. Optimal üreme ısıları 30-35°C’dir. Bu iki özellik Malassezia
’nın neden sadece sıcakkanlı hayvanlardan izole edildiğini gösterir. Laboratuvarda
spesifik maddeler ilave edilirse ve uygun sıcaklı sağlanırsa, üretilebilirler ve
saklanabilirler. Veterinerler, köpeklerde kulak infeksiyonuna yol açan M.
pachydermatis’i uzun bir süre araştırmışlardır123,112. Son zamanlarda M. furfur ve M.
obtusa’nın da köpeklerde dış kulak infeksiyonundan izole edildiği rapolar vardır ve
kedilerden de M. sympodalis elde edilmiştir105,118.Önceki birçok çalışmalarda lipit

17
bağımlı Malassezia’nın birçok hayvanın (maymun, domuz, ayı, kuş, gergedan)
derisinden izole edildiği gösterilmiştir 108,124,125.
Sağlıklı derideki Malassezia spp. varlığı 19. yüzyılın başlarında fark edilmiştir.
Kolonizasyonun sıklığı ve yoğunluğu kişinin yaşıyla ve yağ bezleriyle ilgilidir126
(Marcon ve Powell 1992). Yeni tanımlamalardan beri bazı çalışmalarda insan
derisindeki Malassezia’nın tam yayılımına odaklanmıştır. Crespo Erchiga ve
ark105,127,106 yaklaşık olarak 300 kişide üç başarılı çalışma yapmışlardır ve sağlıklı
deride özelliklede M. sympodialis‘in baskın tür olduğunu göstermişlerdir. Bundan başka
M. globosa ile beraber düşük oranda bulunur. Diğer türler vücutta % 10 kadar bulunur.
Devam eden yeni çalışmalarda M. restricta kafa derisinde baskın olan tür olarak
bulunmuştur. Yetişkinlerin kafa derisinde M. restricta % 20 oranında bulunur ve bu
durum genelde kepeklerin şiddetiyle ilgilidir. Bundan başka, Aspiroz ve ark121. 1990’da
M. restricta türünün özellikle kafa derisinde, M. sympodialis’ in ise sırtta yaygın olarak
bulmuştur. Ancak M.globosa’ yı kafa derisinde, alın ve gövde de eşit oranda bulmuştur.
Bu veriler Midgley ‘in128 (UK) 2000’de M. globosa türünü kafada % 45-50 gibi yüksek
oranında bulmasına rağmen yayınladığı verilerle benzerdir. Bu farklı durumda, M.
furfur sensu stricto izolatının sayısı çok düşüktür128,106 ve hatta Japonya’da 35 normal
objede yapılan çalışmada gövdeden sadece M. globosa ( % 51) ve M. sympodialis (%
26) izole edilmiştir. Kafa ve yüzden alınan örneklerin % 60’ı kültürde negatif iken, M.
globosa, M. furfur ve M. sympodialis çok az olgudan izole edilmiştir129.
Rusya’da 32 tane sağlıklı bireyin derisinden alınan örnekler arasında M. sympodialis en
yaygın tür iken M. globosa çok az miktarda bulunmuştur130.
Kanada’da 20 tane sağlıklı bireyden yapılan bir çalışmada M. sympodialis en yaygın tür
olarak bulunmuştur131. Bu çalışmada M. globosa kafa, alın ve gövdede eşit olarak
bulunmuş, omuzlarda ve bacaklarda çok az izole edilmiştir. Oysaki M. slooffiae ve M.
restricta kafa ve alında diğer vücut bölgelerine göre daha fazla izole edilmiştir. Bu
çeşitli çalışmalar arasındaki karşılaşılan farklılık; farklı örnek alma yöntemleri,
kullanılan besiyerleri 131, 128,129,122(Gupta ve ark. 2001a tarafından Leeming-Notman
besiyeri, Midgley 2000, Nakabayashi 2002 ve Crespo Erchiga ve ark.2002’de mDikson
besiyeri) ayrıca etnik ve coğrafik faktörlerle açıklanmıştır.

18
Son olarak Sugita ve ark.132 2001’de nested polimeraz zincir reaksiyonu(PCR)
metodunu kullanarak, sağlıklı derinin baş ve ense kısmından alınan 18 tane örnekteki
Malassezia DNA’sını % 78‘ini ortaya çıkarmıştır. Bu izolatlar arasında M. restricta, M.
sympodialis, M. globosa ve M. furfur % 61’den 11’e kadar dizilen sıklıkta bulunmuştur.
Bu durum en az dört tane Malasezia türünün; M. sympodialis, M. restricta, M. globosa
ve M. furfur insan derisinde flora elemanı olduğu sonucunu doğrulamaktadır.
Đsveç’te Hellgren ve Vincent’in133 yaptığı bir çalışmada bu hastalığın insidansında
iklimin önemini belirtmişlerdir ve onlar PV’yi Đsveç’te yaygın bir hastalık olmadığını
vurgulamıştır.
Bu araştırıcıların açıkladığı verilere göre, tropikal iklimler göz önüne alındığında, en
yüksek prevalansı Yeni Gineden Mc Lennan ve O’Keeffe (1996) (% 42) rapor
etmişlerdir.
2.9. Epidemiyoloji
Pityriasis versicolor genellikle 13-19 yaş arası gençlerde ve genç erişkinlerde
görülür. Malassezia’lar lipit gereksinimi olduğundan PV küçük çocuklar ve yaşlılarda
nadiren görülür. Yağ miktarı arttığı dönemde PV görülmeye başlar. Normal floradaki
yayılımı seböz bezlerinin yoğunlu ile ilgilidir ve yüz, göğüs ve sırtta yüksek oranda
mantar bulunur. Saçta, tırnakta ve mukozalarda bulunmaz. Đnsan derisinin normal
florasında bu maya mantarı vardır. Mantarın maya formundan hif formuna dönüşümü
ile birlikte PV oluşur. Bu dönüşüm, endojen ve ekzojen faktörlerle ilgilidir134,135.
Pityriasis versicolor’a yol açan endojen faktörler; beslenme bozukluğu136, doğum
kontrol hapı alımı, kortikosteroitlerin kullanımı, bağışıklık sistemi bozuk olanlar ve aşırı
terleme137,138 dir. PV, ılıman iklimlere oranla tropik iklimlerde sık görülür. Ayrıca, yaz
aylarında daha yaygındır. Sıcaklığın ve nemin artması patogenezde rol oynar. Özellikle
sporcular bu hastalığa yatkındır. Bu durumun aşırı terlemeden olup olmadığı açık
değildir139,140. (Hijyen bu hastalığın gelişmesinde önemli bir etmen değildir. Ancak,
deriye krem veya losyon uygulaması bu hastalığın gelişmesinde kolaylaştırıcı bir
etkendir141,142,143. PV ile beslenme bozukluğu ve sosyo-ekonomik düzey arasındaki
ili şki yoktur. Ancak PV’un gelişimi organizmanın bağışık yanıtının değişmesiyle de

19
ilgili olabilir 144,147. AIDS146,147, visseral Layşmanyoz gibi bağışık sistemi baskılanmış
insanlarda PV daha sık görülür145,146,147,148,149. Bununla beraber, AIDS’lilerde diğer
mantar hastalıklarının çok sık görülmesine karşılık PV’un daha sık görülmediğini
gösteren çalışmalarda vardır. PV’un, ışın tedavisi ve lityum tedavisi alan kişilerde
görüldüğü rapor edilmiştir150,151.
Endojen faktörler PV gelişimini kolaylaştırır. Birçok çalışma PV’un erkeklerde
daha sık görüldüğünü belirtmesine karşılık137,152,153 farklı görüşlerde bildirilmiştir21,154.
Ancak, birçok çalışma PV’un gelişiminde cinsiyetin çok önemli olmadığını
göstermiştir. Bir kere PV görülürse, multifaktoryal kalıtımdan dolayı ikinci ve üçüncü
akrabalarda da görülmeye başlar156.
PV, tropikal iklimlerde yaygın görülmesine karşılık, çocuklarda nadirdir157.
Çocuklarda PV Kuzey Amerikaya160 oranla Avrupa’dan158,159daha sık rapor edilmiştir.
Çocukların yüzünde erişkinlerden daha fazla görülür158,159,160,161. PV iki yaşın altındaki
çocuklarda ve prematürlerde nadiren görülür162. Bir olgu sunumunda iki haftalık
yenidoğanın göğüs bölgesinde saptandığı rapor edilmiştir163. Bu olguda ortamın
sıcaklığı ve neminin sebep olduğu düşünülmüştür. Prematüre doğanların derilerinde bu
maya mantarı vardır, ancak lezyonların gelişip gelişmediği gözardı edilir. Bu durumun
bebeklerin doğumundan sonra sağlık personeli tarafından elle dokunulmasından ileri
geldiği düşünülmüştür164.Birçok sağlıklı yenidoğanda yaşamın ilk altı ayında deri
florasında Malassezia türleri gelişmeye başlar. Đklim ve genetik faktörlerinde etkisi ile
Malassezia’ ların ilk kolonizasyonu meydana gelir165.
PV, yaşlılarda ender görülür166. Bunun sebebi yaşlılıkla beraber yağ üretiminin
azalmasıdır. Bununla beraber, hastanede yatan yaşlı insanlar PV’un gelişimi için yüksek
risk oluştururlar167. Bu durum şu şekilde açıklanır; bu insanların bağışıklık sistemi
baskılanmıştır ve vücutları daha çok ısınır ve terler. Özellikle de giysilerini
değiştirmezlerse daha çok risk altına girerler. Bu durum, sık sık banyo yapılması ile
azalır.
Pityriasis versicolor, Malassezia’ların sebep olduğu kozmetik bir
infeksiyondur168. Hastalığın oluşumu mikro-organizmanın kolonizasyonun
yoğunluğunun artışı ile ilgilidir. Pityrosporum folliküliti saç folliküllerinin

20
inflamasyonu ile tarif edilir. Akne benzeri kızarıklıkla sonuçlanır169,170. Seboreik
dermatit Malassezia cinsi maya mantarlarının artışıyla ilgili 162 iken, seboreik dermatitli
hastaların lezyonlu ve lezyonsuz bölgelerindeki Malassezia’ ların miktarında bir
farklılık bulunmamıştır. Bu araştırıcılar, Malassezia’ ların deri üzerinde aşırı üremesi
sonucu olduğunu söylemişlerdir171.
Pityriasis versicolor’un prevalansı Đskandinavya ülkelerinde popülasyonun %1’i
kadardır 154,133, 172. Tropikal ülkelerde PV oranı daha sık görülür ve bazı bölgelerde %
50’ye ulaşır173,174. Hatta bu ülkeler arasında hassasiyeti değişkenlik gösterebilir 175 ve bu
ülkeler infeksiyona eğilimlidir ve devamlı nüks eder. Afrika’da yapılan bir çalışmada
kronik PV lezyonlarının kendiliğinden kaybolduğu gösterilmiştir. Ancak diğer
çalışmalar bunun mümkün olmadığını ileri sürmüşlerdir.
Pityriasis versicolor lezyonlarının görüldüğü bölgeler sırt, boyun, ense, yüz ve
üst kollardır. Ancak diğer bölgelerde de görüldüğü rapor edilmiştir. Pityriasis versicolor
saçlı deride de meydana gelebilir, lezyonları kepekli veya kepeksiz olabilir176,177.
Yetişkinlerin yüzünde olan lezyonlar genelde gövde ve üst kollarda görülen PV’a ikincil
gelişir 178,179.
Lezyonlar peniste te görülebilir149,180,181. Bundan başka lezyonlar gövdeye dahil
olmayan bölgelerde de rapor edilmiştir. Bu bölgeler boyun, kasık, meme145, periareolar
bölge (erkek hastalarda)183, koltuk altı, genital bölge182, ellerde, antekubital bölge,
önkol184. Bu bölgelerde PV lezyonları ender görülmesine karşılık, PV tanısı kabul
edilmemektedir. Çünkü klinik deri erüpsiyonu anatomik yayılımı yansıtmaz.
2.10. Koleksiyonlardaki Malassezia’nın Korunması
Malassezia cinsinin uzun yıllar boyunca neden tartışma konusu olduğunu
açıklayabilen birkaç sebep vardır. Bunlardan birincisi; lipofilik aktivitesini
anlaşılamaması ve yıllarca mantar koleksiyonu olmadan yalnızca literatürdeki
özelliklerinin bilinmesidir. Mantar kültüründe izolasyonunun yapılabilmesi ile izolatlar
saklanabilmiştir. Bununla beraber, zamanla sadece daha az kalan türler özelliklede M.
furfur ve M. pachydermatis sağ kalabilmiştir. Guého ve Meyer 1989’da99 Malassezia

21
cinsini ilk kez genom karşılaştırmasına dayanarak yeniden değerlendirmiş ve
bulgularını yayınlamıştır( liyofilize suşlar ve Utrecht Hollanda’daki Centralbureau voor
Schimmelcultures mantar koleksiyonunda). Ayrıca lipit bağımlı suşlar zeytinyağı
eklenmiş besiyerinde üremiştir. Gerçekte, yalnızca M. furfur, M. pachydermatis ve M.
slooffiae liyofilizasyonla yaşar. Yarrow ve Ahearn, 1984’te bu cinsi yalnızca iki türle
sınırlamıştır. Guého ve ark. 1996’da101 koleksiyonlarda saklanan suşlarla, taze izolatları
karşılaştırmış ve başarılı bir taksonomik çalışma yapmışlardır. Bütün türler, modifiye-
dixon besiyerinde değerlendirilmiş ve bu durum mikro-organizmaların daha iyi
saklanmasına olanak vermiştir. Bu araştırıcı asla mikroorganizmayı oda sıcaklığında
inkübe etmemiş ve saklamamıştır. Ancak, her ay 34°C’de yeniden kültürünü yapmıştır.
Koleksiyonu daha sonra dondurup kurutarak -80°C’de saklanmıştır. Crespo ve ark.185
Malassezia için kullanılan saklama koşullarının önemini doğrulamıştır.
2.11.Tanı
Genelde pityriasis versikolor’un tanısı kolaydır. Bununla beraber lezyonların
görünümündeki farklılık deneyimli olmayan bir klinisyen için karışık olabilir. Wood
ışığı muayenesi pityriasis versicolor’un tanısına yardımcı olabilir. Wood ışığı altında
(filtre edilmiş UV ışını, 365nm) pityriasis versicolor lezyonları sarı-yeşil renkte floresan
verir. Bu floresan klinik olarak görülebilir lezyonları içine alır56,186.Wood ışığı ayırıcı
tanı için faydalı olabilir. Çünkü floresanın rengi Malassezia’nın hifleri için
karekteristiktir. Bununla beraber Wood ışığı olguların sadece 1/3’inde pozitif bir cevap
sağlar ve daha çok bu Malassezia furfur’ dur186,187.
Mikolojik incelemede pityriasis versicolor’un tanısını doğrulanabilir. Lezyonlardan
alınan klinik materyal ışık mikroskobunda incelenmelidir. Lezyonlu bölgeler fazla
miktarda mikroorganizma içerir. Lezyonlardan bistirü ile kazıma yönteminden başka
selofan bant yöntemi de kullanılabilir. Đlk önce, lezyonlardan alınan klinik materyal
%10-15 ilave edilmiş KOH ile lam lamel arası preparat yapılarak ışık mikroskobunda
incelenir. KOH keratini eriterek mantar elemanlarının görülmesini kolaylaştırır. KOH
ile muameleden sonra 15-20 dakika beklenmelidir. Lam hafif ısıtılırsa keratinin erimesi
hızlanır, sonra ışık mikroskobunda incelenir. Işık mikroskobu muayanesinde hif ve

22
maya hücreleri görünür. Bu görüntü karekteristiktir ki buna “köfte- makarna” görüntüsü
denir. Potasyum hidroksitin yerine Albert solüsyonu189 ve toludin mavisi( 0.15gr,
malaşit yeşili 0.2 gr, 1ml asetik asit, 2 ml % 95 lik etanol, 110 ml distile su 190)
kullanılabilir.
PV’un tanısında deri biyopsisine gerek yoktur. PV etkeni mikroorganizmanın üretilmesi
için lipitten zengin bir besiyerine gereksinim vardır. Bu yüzden Malassezia’lar standart
besiyerlerinde üremez. Ayrıca, Malassezia maya mantarı normal popülasyonun hemen
tamamında (%90-100) bulunur191. Bu nedenle mikroorganizmanın besiyerinde üremesi
PV tanısını doğrulamaz. PV, etken mantarın mantarın maya şeklinden mantarın
patolojik olan hif şekline dönüşümü ile ilgilidir. Bu durum, daha çok kişiye özel
endojen faktörlerle ilişkilidir 192 .
Şekil 2.1: Wood ışığı
2.12. Kommensalizm
2.12.1. Normal Derideki Malassezia türlerinin Yayılımı
Malassezia normal insan derisinin flora üyesi olan bir mikroorganizmadır ve
özelliklede göğüs, sırt ve kafa bölgesi gibi yağ bakımından zengin bölgelerden izole
edilebilir193,194. Birçok çalışmalarda farklı populasyonlardaki ve farklı yaş gruplarındaki
taşıyıcılık oranı incelenmiştir. Bununla beraber ilkönce yapılan çalışmalarda örnek alma
yöntemlerinin ve besiyerlerinin sınırlı olmasından dolayı, taşıyıcılık oranı sık sık düşük
bulunmuştur. Leeming ve ark. tarafından, çeşitli vücut bölgelerindeki Malassezia
türlerinin dağolımıyla ilgili geniş bir çalışma yapmıştır. Bu çalışmada optimize edilmiş
besiyeri kullanmıştır194. Bütün vücut yüzeyinden 20 tane klinik örnek almışlardır.
Malassezia türleri gögüs, sıttan, kafa derisinden, kulaktan, kalçadan izole edilmiştir. Bu
bölgeler arasında gögüs, kulak, sırtın üst kısmında, alında ve yanaktaki izolasyon en
yüksektir. Erkekler ve bayanlar arasındaki farklılılar not edildi. Populasyonun yüksek
yoğunluğuyla beraber erkeklerin belinde ve gövdesinde daha düşüktü. Aynı besiyerini

23
kullanan daha sonraki çalışmalar büyük ölçüde bu bulguları doğruladı195. Bergbrant ve
Faergemann196 derideki Malassezia yoğunluğunun yaş arttıkça derideki yağ miktarından
dolayı azaldığını belirtmiştir. Bu yüzden 30’ lu yaşlarda Malassezia türlerinin sayısı, 40
-80 yaş arası kişilerden önemli ölçüde daha fazladır. M. pachydermatis genellikle insan
derisinden izole edilir ama bu türün varlığı geçicidir ve insanların derilerinde
kommensal olarak bulunmaz195.
Son zamanlardaki çalışmalar sağlıklı insan derisindeki yeni tanımlanan Malassezia
türlerinin dağılımını incelemişlerdir. Bu durum çeşitli çalışmalar arasında farklılık
göstermektedir ve bunun için muhtemel olan iki tane açıklama vardır. Birincisi farklı
ülkelerdeki kişilerin derilerindeki Malassezia yayılımının farklı olmasıdır. Çünkü farklı
coğrafik bölgelerdeki insanların kökenleri farklıdır. Bu durum önceki çalışmalarda
açıklamıştır. Bununla beraber Đspanya’da iki çalışma yapılmış ve farklı sonuçlar
çıkmıştır. Đkinci açıklama ise kullanılan örnek alma yöntemleri, sayılamayan ve göreceli
olarak yapılan yöntemlerdir. Bu yöntemler, farklı yerlerde yapılan çalışmalarla hangi
türün baskın olduğunu tanımlamak için ve sayılabilir veriler elde etmek için yetersizdir.
Bu yüzden insan derisindeki yeni türlerin yayılımındaki sayılabilir veriler hala
beklemededir.
Çocuklardaki kolonizasyon oranı, tartışma konusudur. Erişkinlerle beraber rapor
edilen kolonizasyon oranı, kullanılan besiyerini ve örnek alma yönteminin kısmen
yansıtır. Özellikle genç çocuklarda ve yeni doğanlarda, uygun olmayan tekniklerden
beri uygun örnek alma yöntemleri pratik olabilir. 60 sağlıklı çocuktan biri, sırasıyla 2
aylıktan 14 yaşına kadar olan çocuklarda Malassezia türleri pozitif bulunmamıştır. Bu
diğer çalışmalarla çelişmektedir ki sağlıklı çocuklardaki taşıyıcılık oranı kafada %74,
sırtta %93 ve alında % 87 olarak bulunmuştur. Genelde Malassezia’ nın taşıyıcılık oranı
puberteyle arttığı görülmüştür. Bu durum aynı zamanda seböz bezlerinin artmasıyla
ilgilidir 197.
Prematüre doğanlarda kataterle ilişkili fungemiyaların olarak Malassezia’nın
tanınmasının artması, prematürlerde ve 40 haftayı doldurmamış bebeklerdeki
kolonizasyon oranını çalışmak için hızlı desteklemiştir. Hastanedeki yenidoğanlarda
kolonizasyon oranı % 37’den % 100’e kadar olduğu rapor edilmiştir. Küçük yaşlardaki
gebelik, düşük doğum ağırlıklı bebeklerin olması ve uzun süre hastanede kalınması gibi

24
faktörler bu grubun kolonizasyonu için kolaylaştırıcı olabilir. Bununla beraber sağlıklı
yenidoğanlardaki kolonizasyon orandaki hiçbir sistematik inceleme olmamıştır. Bu
yüzden bu durum hala kapalıdır.
2.13. Deri Đmmun Sistemi
Malassezia kutanöz kommensal bir mikroorganizmadır. Bu yüzden onun immun
sistemle ilişkisinin ilk noktası muhtemelen deri yoluyla olmaktadır. Deri vücudun en
geniş olmaktadır. Đnsan ve çevre arasındaki ara yüzeydir198. Deri mikroorganizmaların
geçiçi ve kalıcı florasından dolayı geniş antijenlerle karşı karşıya kalır. Yıllarca derinin
yıllarca duran bir bariyer olarak bilinmesine rağmen, şimdi deri kompleks ve immun
sistemin parçası olan bir organ olarak bilinmektedir ki hem spesifik hem de nonspesifik
immun cevapta rol oynar.
Deride Malassezia gibi birçok mikroorganizma kommensal olarak bulunur. Bu
mikroorganizmalar besin ve yer ihtiyacı için yarışırlar. Bu mikroorganizmalar ayrıca
patojenlerle de yarışırlar. Sonunda epidermisteki hücrelerin şekli sık sık değişir ve
inflamasyon sırasında hücrelerin değişme oranı artar. Bu değişiklik mikroorganizmalara
zarar verir ki infekte olmuş veya kolonize olmuş bu hücreler, deri tabakasının daha
derinine invaze olmayı engeller. Ayrıca derinin bariyer fonksiyonu ve onun kommensal
florası, fagositik hücreler nonspesifik kutanöz immun cevapta önemlidir.
2.14. Malassezia Đle Đlgili Yüzeysel Hastalıklar
Malassezia cinsi maya mantarları, insanlarda çeşitli hastalıklara sebep olur.
Bunlar arasında başta PV olmak üzere, seboreik dermatit, atopik dermatit, follikülit,
Gougerot-Carteaud’un retikülat ve birleşik papillomları, psöriasis, yenidoğan püstülozu,
otitis eksterna ve onikomikoz yer alır. Malassezia geçmiş yıllarda prematürlerde ve
kataterden lipitle beslenenlerde sistemik infeksiyonlara111, sebep olmasına rağmen esas
olarak deriyi etkiler. M. pachydermatis her ne kadar kalalikülitise, yara ve yeni doğanda
sistemik infeksiyonlara sebep olsa da, insanlardaki hastalıkla ilişkili değildir. Ayrıca

25
hayvanlarda önemli bir patajendir ki dermatis, otitis eksternaya sebep olur. Bu yüzden
insanlarda bu organizmaya karşı oluşan immun cevap çalışılmamıştır.
2.14.1. Pityriasis Versicolor
Malassezia, Pityriasis versicolor PV (sinomimi Tinea versicolor)’un etiyolojik
ajanı olarak bilinen kronik, iyi huylu kutanöz hastalıktır ki çoğunlukla asemptomatik
olan hafif ve kronik durumlu olan bir hastalıktır. Gövdenin üzerinde yer alan
değişebilen renklerde (pembe, beyaz, kahverengi) görülen kepeklerle karekterize edilir.
Lezyonlar genellikle sırt, göğüs, karın ve üst ekstremitelere yerleşir. 199Mantar, melanositleri
bozar, hücreler ya pigmentlerini kaybederler ve akromik bir tablo ortaya çıkar ya da pigment
artışı olur ve koyu sütlü kahverenginde lezyonlar meydana gelir. Gövdede ve omuzlarda Wood
ışığı (365 nm dalga boylu UV ışığı) altında ekseri altın rengi floresans verir. Kronik ve
tekrarlayıcı bir mikozdur199. Mantarın maya fazından miçel fazına dönüşmesiyle meydana
gelmektedir. Daha sonra mantar stratum corneum tabakasına geçer. Korneositlerin
arasına penetre olur hafif kaşıntıyla beraber hipo-veya hiperpigmentasyolu lezyonlara
sebep olur. Bu durum sebase (yağ) bezlerinin çok aktif olduğu zaman meydana gelir200.
Çocuklarda ve yaşlılarda rapor edilmesine rağmen çoğunlukla adolesanlarda ve orta
yaşlılarda meydana gelir. Kolaylaştırıcı etmenler; genetik hassaslık137, hastalık veya
yetersiz beslenme, malnütrisyon62 plazmadaki artmış kortizol düzeyi137 yüksek çevre
sıcaklığı ve nem. Ilıman iklimlerdeki insidansı % 1 dir138. Fakat tropikal bölgelerdeki
insidansı % 40-60 gibi yüksek bir oran rapor edilmiştir. Tropikal bölgelerdeki lezyonlar
daha çok yoğundur201 ve lezyonlardan alınan örneklerin mikroskobik görüntüsü ılıman
iklimlerde alınan lezyonlardan farklı görülebilir. PV gövde üzerindeki klasik görüntüsü,
muhtemelen giyinmeden dolayı oluşan kapanmayla ilgilidir. Bu durum genellikle
sağlıklı ki şilerde de görülür ve insidansı AIDS’liler kadar yüksek değildir. Tanı
genellikle direkt olarak tipik 3-8 µm çapında olan maya hücrelerinin gözlemlenmesiyle
konulur ve lezyonlardan kepeklerin potasyum hidroksitle incelemesinde 1.5-4 µm
çapındaki kısa güdük pseudo-hiler görülür. Bu tipik görüntüye “köfte-makarna” denir
(Şekil 2). Ilıman bölgelerde mikroskobik görüntüde hifle beraber kümelenmiş maya
hücresi görülür. Tropikal bölgelerde ise hifle beraber yumurtamsı ve silindirik hücreler
görülür201. Mayalar esas ajan olan M. globosa gibi küremsidir bazen M. furfur gibi

26
yumurtamsı olabilir. Taksonomik sınıflandırmada sadece M. globosa ve M. furfur
kendiliğinden az veya çok hif oluşturabilir49.
Şekil 2.2: Örneğin laktofenol ile görünen köfte- makarna( blastospor ve hif)
PV’nin mikolojisi yoğun olarak çalışılmıştır. PV lezyonlarındaki Malassezia’nın
hif formu, Neuman 1871 yılında lezyonlardan hifi gördüğü zaman tanımlamıştır.
Malassezia ile daha başarılı çalışma62 1938 yılında PV lezyonlarından izole edilmiştir.
Gordon hem PV lezyonlarından hem de normal deriden yuvarlak maya hücresi
tanımlanmıştır119 Robert(1969) PV’li 25 hastayı incelemiştir ve lezyonlarını kazımıştır
ve 25 tane hastada Malassezia maya mantarını bulmuştur. Bununla beraber maya ve
hiflerin sayısı değişkenlik göstermiştir. 25 tane örnek zeytinyağı eklenmiş malt agarda
üretmiştir. 25 tane izolatın hepsinde P. orbiculare izole etmiştir. Ayrıca bunların 6 tane
sinde P. ovale de üretildi. Robert hastaların normal derilerindeki klinik izolattan hifleri
bulmuştur.62
Takip edilen yıllarda McGinley ve ark. 31 tane PV’li hasta çalışmıştır. Hem
normal deride ve hem de lezyonlarda, maya ve hiflerin miktarını hesaplamıştır.
Lezyonlardaki hiflerin miktarı 295,300 cm-2 iken mayalar 155,900 cm-2, oran 2:1:1dir.
Normal derideki örnekte mayalar 18,900 cm-2 ve hif ise 5,800, oran 1:2:1 dir. Ayrıca
McGinley ve ark. lezyonlardaki korneositleri saymışlar ve korneosit miktarının normal
deriden üç kez daha yüksek olduğunu bulmuştur. Ve bu durum organizmanın kolonize
olmasını destekler. Sonuç olarak Malassezia’nın sebep olan bir mikroorganizmadır202.
Faergemann ve Bernander (1979) 30 tane PV’li hasta örnekleri toplamışlar. Hem
normal deriden hem de lezyonlu bölgeden bistirü ile kazıyarak örnek almışlardır. Bütün
hastaların lezyonlarından mantarı izole etmişlerdir 24 hastanın normal derisinden P.
orbiculare’yi izole etmişlerdir. Fakat P. ovale’yi hiçbir örnekten izole etmemişlerdir.
Bu araştırıcılar yaptıkları çalışmada Đsveç populasyonunda P. ovale’nin yaygın
olmadığını söylemişlerdir.

27
Ashbee ve ark. PV’nin mikrobiyolojik olarak çalışmışlardır. 10 tane PV’li
hastanın gövdesinden ve kafasından örnekler almışlardır ve her bir vücut
Malassezia’nın toplam sayımını ve özellikle sevovarların hastalıkla ilgisi olup
olmadığını anlamak için de her bir serovar (A, B ve C) miktarını ölçmüşlerdir. Bu
çalışmanın sonucunda; serovar A’nın gövdede baskın olduğunu ve hem hasta grubunda
hem de kontrol grubunda aynı olduğunu ve bu sonucun hastalıkla ilgisi olmadığını
bulmuşlardır. Hiçbir serovarın kafada baskın olmadığını bulmuşlardır. Sonuçta hem
kontrol grubunda hem de lezyonlu deride aynı bölgedeki serovarların dağılımı veya
Malassezia’nın yoğunluğu açısından farkı olmadığını rapor etmişlerdir. Bu bulgular
McGinley’in çalışmasını doğrulamaktadır fakat McGinley ve ark. mikroskop yoluyla
toplam sayımı rapor etmişlerdir. Ashbee ise kültürdeki yaşayabilen sayımı çalışmıştır bu
yüzden sonuçlar direkt olarak karşılaştırılabilir değildir.
Yeni taksonomideki izleyen tanımlamalarda, birkaç epidemiyolojik çalışmalar
farklı ismlendirmelere yol açmıştır. Japonya’da Nakabayashi ve ark. (2000) 22 tane PV’
li hastanın lezyonlarının %55‘inde M. globosa’yı bulmuştur. Ayrıca diğer türleri %
10’un altında bulmuştur. Bu tür patojenitede rol oynar. Bundan başka Crespo Erchiga
(1999) bu türü patojen olarak tanımlamıştır.
Kanada’da Gupta ve ark.(20001) yaptığı çalışmada 111 tane PV’li hastadan M.
sympodialis %59, M. globosa %25 ve M. furfur %11 bulmuştur. Bu çalışmada
mikroskobisi pozitif olan olgular kültürde üretmeyi başarmıştır.
Güney Đspanya’da Crespo Erchiga ve ark. (2000) üç tane başarılı epidemiyolojik
çalışma yapmışlardır. Bu çalışmada PV, seboreik dermatitli aynı hastaların lezyonlu
bölgelerinden ve sağlıklı deriden klinik örnek alarak Malassezia maya mantarını
izolasyonu karşılaştırmışlardır. Lezyonlardan alınan örneklerin mikroskobik
incelemesinde tipik yuvarlak maya hücreleri ve hifleri görmüşlerdir. Araştırıcılar bu
çalışmanın sonucunda, lezyonun merkezinden alınan klinik örneğin, kenardan alınan
örneğe göre daha az uygun olduğunu bulmamışlardır. Bunun aksine tinea gibi
dermatomikolarda, maya mantarı ve pseudohif etkilenmiş vücudun her tarafına aynı
sayıda yayılır. Bu çalışmadaki son grup ise 96 hastadan erişkin hastaların %97‘sinde M.
globosa izole edildi. Olguların sadece 60 M. globosa, % 29’unda M. sympodialis,
%7’sinde M. slooffiae izole edildi. Bu iki tür (M. sympodialis ve M. slooffiae) klinik

28
olarak etkilenmemiş gövdede benzer oranda bulunmuştur. Fakat M. globosa lezyonsuz
bölgelerden izole edilmemiştir. Bununla beraber alında olguların %12’sinde M. globosa
az miktarda izole edilmiştir. Bu türün seboreik dermatitte ve sağlıklı derideki maya
hücrelerinin varlığı lokal faktörleri (nem, terleme, ısı, yağ) işaret eder. Bununla beraber
kişinin yatkınlığı ve özel durumları hif formuna dönüşümünden ve klinik lezyonların
gelişmesinden sorumludur. Crespo Erchiga ve ark. (2002) 210 PV’li olgudan yapılan bir
çalışmada yeni sonuçlar elde etmişlerdir. Bu yeni sonuçlar önceki verilerle uyumludur
ki PV etiyolojisinde ılıman iklimlerde M. globosa’nın rolü olduğunu doğrulamaktadır.
Üç başka çalışma bu sonucu doğrulamaktadır. Aspiroz ve ark.203 (2002, Đspanya,
Zaragoza), 79 PV’ li hastanın % 90’ında M. globosa ve % 41’inde M. sympodialis
bulmuştur. Japonya’da Nakabayashi(2002) Malassezia türlerini PCR ile identifiye etmiş
ve M. globosa % 97 oranında bularak en yaygın tür olarak tanımlamıştır. Sonra M.
restricta (%79) ve M. sympodialis (%68) takip etmiştir. Son olarak Yunanistan’dan
Gaitanis ve ark. (2002) DNA-PCR metodunu kullanarak PV lezyonlarından sadece M.
globosa‘ yı identifiye etmiş ve tanımlamıştır. Ayrıca bu sonuç kültür ve biyokimyasal
testleriyle de doğrulanmıştır.
Önceki yapılan çalışmalarda ilginç bir nokta tartışmaya sebep olmuştur. PV’a sebep
olan türlerin klinik ve coğrafik yayılımına göre değişiklik gösterebileceği
vurgulanmıştır. Ingham ve Cunningham 1993’de bu tartışmayı doğrulamıştır. Castelleni
1925’te tropikal bölgelerde görülen Pityriasis’i bağımsız olarak ayırdı ve Panja(1927)
hastalığı iki gruba ayırdı; Pityriasis versicolor (Malassezia furfur) ve Pityriasis flava (M
.tropica). Ilıman bölgelerde lezyonlar üst gövdeyi etkiler ve miksoskop altında farklılık
göstermesine rağmen genelde küresel hücreler görülür. Ancak tropikal iklimlerde bunun
aksine lezyonlar daha çok yüzü ve enseyi etkiler. Mikroskopta yumurtamsı veya
silindirik hücreler görülür. Midgley PV’den sorumlu olan türlerin vücut bölgelerine
göre değişiklik gösterdiğini belirtmiştir. Crespo Erchiga’ya göre PV olguların sadece %
1’i görünür, direkt mikroskop altında oval ve silindirik hücreler görülür. Tropikal
bölgelerdeki hastalarda lezyonlar yüz, bacak gibi atipik bölgeleri etkilediği görülmüştür.
M. furfur’ un sebep olduğu PV olgularının sayısı, lezyonlardan alınan örnekler 37° C de
üretildiği zaman önemli ölçüde gösterilebilir99(Guého ve Meyer1989). M. furfur deride
düzenli olarak bulunur ve bu tür lipit bağımlılığından dolayı derin infeksiyonlardan

29
sorumludur43 (Boekhout ve ark.1998; Theelen ve ark. 2001). Bununla beraber bu tür son
zamanlardaki araştırmalarda nadirdir. M. furfur ayrıca düzenli bir tür olarak
düşünülmesine rağmen, insan derisinde seyrek olarak bulunmaktadır.
PV hastalarında Malassezia’nın immünolojik cevabı belirsiz şartlarda
incelenmiştir. Wu ve Chen(1985) artmış humoral Malassezia antikorları bulmuş,
kontrol grubu ile karşılaştırmıştır. Ancak diğer çalışmalar bu iki grup arasında bir fark
bulamamıştır56 (Faergemann 1983; Midgley ve Hay 1988; Asbee ve ark.1994). Çelişkili
sonuçlar hücresel immünite araştırmalarda sonuçlandırılmıştır.
PV’nun başka bir bakış açısı ise lezyonlardaki deri pigmentasyondaki
değişikliktir. Birkaç teoride derideki hipopigmentasyonu açıklama amaçlanmıştır. Buna
göre lipoksigenaz veya azeleik asitin melanin üretiminin ihibisyonu ve keratinositin
melanozomlara transferinin engellenmesine sebep olur. Diğer araştırıcılar
hiperpigmente lezyonlarının inflamasyondan204, deri kalınlığının artması veya derideki
mikroorganizmanın sayısının artmasından dolayı olduğunu savunmuşlardır. Bu
çalışmaya rağmen pigmentasyon mekanizmasının değişmesi çoğunlukla çözülmemiştir.
PV tedavisi basittir. Farklı antifungal ilaçlarla tedaviden (oral veya topikal)
sonra lezyonlar hızlıca kaybolur. Akromik alanlar haftalarca bir kaç ay devam edebilir.
Topikal olarak en sık kullanılan ilaçlar selenyum sülfit, propilen gliserol ve antifungal
azoller. Yoğun olgularda ketakonazol ve itrokonazol gibi kısa zamanlı sistemik ilaçlar
kullanılır. Bununla beraber tedavinin çok yüksek olmasına rağmen, bir yıldan sonra %
60, iki yıldan sonra % 80 oranında rekürrens meydana gelir. Çünkü altta yatan koşullar
ve lokal faktörler değiştirilmemiş olarak kalır. Yağ bezlerinin içinde bulunan mayalar
çoğunlukla KOH ile ortaya çıkar. Bu durum rekürrensin sıklık oranını açıklar.
Profilaktik itrakonazol tedavisi 6 ay boyunca günde 200 mg verilir. Olguların % 88’
inde mikolojik iyileşme görülür.

30
Şekil 2.3 : Pityriasis versicolor (Hipo-pigmente)
Şekil 2.4: Pityriasis versicolor ( Hiper- pigmente)
2.14.2. Seboreik Dermatit
Seboreik dermatit (SD) kronik, nüksedici bir deri hastalığıdır. Kafa, kirpik,
burun gibi yağ bakımından zengin bölgelerde meydana gelen ve bunun sonucu olarak
yağlı görünüm veren, pullanmaya sebep olan ve kırmızı lekelerin oluşmasıyla
karekterize edilen bir hastalıktır. Seboreik dermatit belki Malassezia ile ilgili en yaygın
hastalıktır ki toplumun % 1-3‘ünde görülür62. Kepek son yıllarda daha çok dikkat
çekmeye başladı. Kepeğin varlığı toplumda saygınlığın azalmasına ve negatif imaj
olmasına yol açabilir. Bunun yanı sıra SD bir hastalıktır ki genelde kafadaki
kepeklerden dolayı tartışma konusudur. SD ve kepek arasındaki ilişki tartışma
konusudur. Bazı araştırmacılar, kepeğin şiddetiyle kafadaki SD‘in tanısına bakmışlardır.
Buna karşılık diğer araştırıcılar kaynağı ne olursa olsun, kepek adının kafadaki dökülen
pul için kullanılması gerektiğine inanmışlardır205. Kepekler erişkin populasyonunun %

31
5-10’unu etkiler. Bu durum hastalığın başlangıç veya orta aşaması olduğunu
düşündürür. SD’ nin gelişiminde Malassezia’nın yeniden aktiflik kazanmasının rolü,
kanıtlarla desteklenmiştir ki birçok olguda kepek SD’ in ılımlı bir formudur. Bazı
araştırıcılar kepek SD’in non-inflamatuvar formu olduğunu düşünmüşlerdir56. Tüm
mevcut olan verilere bakıldığında, SD ve kepek benzer orijinlerin farklı görünümleri
olarak düşünülmüştür.
Son zamanlardaki verilerin çoğunda Malassezia ile kepek arasında direk bir
bağlantı olduğunu destekler. Birincisi, etkili tedavi şartlarında geniş bir madde türlerini
çinko tuzları, selenyum tuzları gibi yüksek spesifik azoller meydana getirebilir. Bu
materyaller mantarın antifungal aktivitesi olarak bilinmektedir. Đkinci faktör ise, SD’
deki gelişmeler kafadaki Malassezia üretimi miktarı beraber olmaktadır.
SD’nin insidansı AIDS gibi immun supresif kişilerde daha yüksektir. Bu oran
AIDS’ lilerde % 30- 33 oranında görülür206. Bu durum immün sistem, hastalığın
patogenezinde önemli bir rol oynar. Đnsidansı Parkinson hastalığı, mutiple skleroz ve ay
depresyonu gibi hastalarda daha yüksektir. Nadirde olsa Malassezia blefarit olgusu
hastalıkla ilgisi olabilir.
SD’nin gelişiminde Malassezia’nın yeniden aktiflik kazanmasının rolü,
kanıtlarla desteklenmiştir ki birçok olguda kepek SD’in ılımlı bir formudur. Bazı
araştırıcılar kepek SD’ in non-inflamatuvar formu olduğunu düşünmüşlerdir56.Tüm
mevcut olan verilere bakıldığında, SD ve kepek benzer orijinlerin farklı görünümleri
olarak düşünülmüştür.
Son zamanlardaki verilerin çoğunda Malassezia ile kepek arasında direk bir bağlantı
olduğunu destekler. Birincisi, etkili tedavi şartlarında geniş bir madde türlerini çinko
tuzları, selenyum tuzları gibi yüksek spesifik azoller meydana getirebilir. Bu
materyaller mantarın antifungal aktivitesi olarak bilinmektedir. Đkinci faktör ise, SD’
deki gelişmeler kafadaki Malassezia üretimi miktarı beraber olmaktadır.
SD’in diğer hastalıklarla da ilgisi vardır. Bu hastalıklar Parkinson Hastalığı,
depresyon, spinal yaralanmalar ve pityriasis versicolor. SD genelde erkeklerde daha
yaygın görülmektedir ve çoğunlukla gençlerde ve genç erişkinlerde daha sık görülür ve
50 yaşın üzerindeki insanlarda da yeniden görülür. Adolesan çağındaki insanlarda ve

32
genç erişkinlerdeki SD’in ilgisi çoğunlukla puberte dönemindeki artmış yağ
miktarından dolayıdır207. Hastalık mevsimlerde de etkilenmektedir. En kötü lezyonlar
kış ayında görülür ama güneş ışığı hastalığın klinik görünümünü iyileştirir. Malassezia
güçlü bir UV koruyucusu (Pityriacitrin) sekrete eder208.
SD’nin lezyonları görünüşte değişiklik gösterir. Karekteristik özelliği kırmızı
leke, pul pul dökülen ve yağlı cilttir. Özelliklede kafada, burun ve dudak kısmında,
kulaklarda, kirpiklerde ve göğüste görülür. SD hastaları yağlı cilde sahip olmasına
rağmen, bu önemli bir sebep değildir.
Đlk yapılan çalışmalarda, Malassezia maya mantarını SD’ye yol açan bir etmen
olarak görülmüştür. Bununla beraber daha sonra araştırmacılar hiperproliferasyonun bir
sonucu olarak bu koşulları incelemeye başlamışlardır. Bu hipotez keratolitiğin
etkisinden ve SD’nin tedavisindeki anti-inflamasyon ajanıdır (salisilik asit ve
kortikosteroid). Keratolitik terapi inflamayon ve hiperproliferasyonun daha az
şiddetinden dolayı SD’ye göre kepekte daha az etkilidir. Selenyum sülfit ve çinko
pitrion gibi geniş spekturumlu antifungallar topikal tedavide etkilidir.
Malassezia maya mantarı ile SD arasındaki ilişki hala bir tartışma konusudur.
Sabouraund (1932) geçen yüzyılın ilk yarısında, Pityrosporum malassezii’nin bu
hastalığa yol açan etmen olarak düşünmüştür. Literatürde yeniden yapılan
incelemelerde Shuster(1984) P. ovale’yi etiyolojik ajan olarak düşünmüştür. Sonra bir
çok çalışmalar SD‘nin şiddeti ve kepeklerde görülen Malassezia hücreleri arasında bir
yakın bir ilişki olduğunu göstermiştir. Taksonomik incelemeler birkaç çalışmayı
meydana getirmiştir ve SD’deki lezyonlarında bulunan farklı türlerin dağılımını
bulmayı amaçlamıştır. Japonya’da Nakabayashi ve ark.(2000) 42 SD’li hastanın
yüzünden %35 M. furfur’u, %22 M. globosa izole etmiştir ve normal kişilerden daha az
izole etmiştir.
Kanada’da Gupta ve ark102.(2001) kontak plaklar kullanarak 28 SD’li hastadan
yaptığı çalışmada M. globosa, M. sympodialis, M. slooffiae azalan oranlarda bulmuştur.
Đspanya’da Crespo Erchiga ve ark.(1999) 75 SD’li hastanın kafa ve yüzünde M
.restricta (% 43), M. globosa (% 34) ‘yı baskın bulmuştur. Gerçekte M. sympodialis(%
19), M. slooffiae (% 2.5) ve M. furfur ( % 0.8) daha az baskındır. Buna karşılık sağlıklı

33
derinin bazı bölgelerinde( alın) örneklerin % 90’ı steril kaldı, bunun aksine aynı
hastanın sağlıklı gövde örneklerinin % 62’sinde baskın tür M. sympodialis’ti. Burda
farklı olarak dikkat çeken şey, antifungal içeren şampuanların kulanımından dolayıdır.
Crespo Erchiga ve Delgado Florencio (2002) SD’li 100 hastadan yeni bir
çalışma yapmışlardır. Bunun sonucunda SD lezyonlarında % 65 M. restricta, % 64 M.
globosa‘ yı baskın bulmuşlardır. Sonra esas olarak gövdede M. sympodialis (%27)
bulmuştur. Gövdede M. furfur’u % 4 oranında bulmuştur. Sağlıklı derinin alın
bölgesinde, hastaların % 20’ sinde M. sympodialis, % 10 M. globosa iken omuzlarda
kişilerin % 58’ inde M. sympodialis ve % 7 M. globosa bulmuştur. Aynı yıl Amerika’
da( Cincinnati) Gemmer ve ark. SD’li 70 hastanın kafa örneklerinden, direk floresan,
nested DNA PCR yöntemini kullanarak alınan kepeklerde % 50-72 M. restricta % 33-
45 M. globosa’yı baksın bulmuş iken % 7-8 gibi az miktarda M. sympodialis’i
bulmuştur ama M. furfur’u hiçbir örnekte bulmamıştır.
Tedavi seçenekleri olarak selenyum sülfit, priton çinko, prakton olamin veya
ketokonazol içeren şampuanlar, topikal terbinafin solusyonu veya ketakonazol krem
aynı zamanda topikal kortikosteroidler uygulanabilir. Kortikosteroidler hatta yumuşak
olan hidrokorsiton asetat kafa ve deri lezyonlarındaki iyileşmesi hızlı sağlar ve nüksü
kortikosteroid içeren ketakonazol tarafından geciktirilebilir. Bu iyi tedavi, Malassezia
maya mantarının özellikle M.restricta ve M globosa‘ nın SD etiyolojisinde ve kepekte
önemli olduğunu gösterir.
2.14.3. Atopik Dermatit
Atopik dermatit veya atopik egzema etiyolojisi bilinmeyen kronik, kaşıntılı
yaygın bir inflamasyon deri hastalığıdır. Karektersitik özelliği eritremayla beraber
egzamatöz lezyonlar ve epidermiste kalınlaşmadır. Bu hastalığa ilk defa Clemmensen
ve Hjorth 1983’de Malasezia maya mantarının yol açtığını belirtmişlerdir. Dahası bu
araştırıcılar AD’li hastaların tedavisinde oral ketakonazollerin önemli etkisini de
bulmuşlardır. Bundan sonra birçok raporda AD’deki mayaların önemliliği belirtilmiştir.
Bu hastalıkta genetik faktörler önemli bir rol oynamaktadır ve birçok hastanın ailesinde
AD, alerjik rinit, astım veya bunların kombinasyonları vardır. Eğer evli çiftlerin her

34
ikisi bu hastalığı varsa, onların çocukları % 70 gibi büyük bir oranda risk taşır. Birçok
hastada AD çocukluk çağında görülür ve büyüme bozukluğu olan çocuklarda % 60-70
oranında görülür. Erişkinlerde AD’nin insidansı % 2 olarak bilinmektedir.
Erişkinlerdeki AD başlangıcı göreceli olarak yaygın değildir. Batı ülkelerinde AD’in
insidansı yüksektir209,210
Erişkinlerdeki AD’in meydana gelmesinde Malassezia maya mantarı önemli bir rol
oynamaktadır. Özellikle bu hastalık başta ve boyunda meydana gelir. AD’li erişkin
hastaların % 83’ünde Malassezia izole edilebilir ve bu hastalık sistemik ketakonazole
cevap verir212. Mayalar kontrol grubunda da sıklıkla izole edilebilir. Đnfeksiyoz ajanlara
kıyasla mayalar hassas kişilerde alerjen olarak rastlanırlar. Bu hipotez gösterilerek
desteklenmiştir ki AD’li hastalar mayalara karşı pozitif reaksiyon gösterirler. Son
yıllarda moleküler çalışmalar Malassezia’dan çıkrılan bazı alerjenlerin yapısı
açıklanmıştır211.
Sugita ve ark. 2002’ de AD’li üç hastadan M. sympodialis ile yakın ilişkili olan M.
dermatis’i tanımladılar59. AD’li hastalardan Malassezia’nın kutanöz kolonizsyonu
incelemesi sırasında, hastaların derileinde yeni bir tür buldular. Toplam 19 AD’li hasta
bu çalışmaya dahil edilmiştir. Toplam beş tane Malassezia dermatis suşundan iki
hastadan izole etmişlerdir. Üç suş tek bir hastadan izole edilmiştir. Ancak diğer ikisi her
bir hastadan izole edilmiştir. M. dermatis’ in fizyolojik olarak karekteristik özelliği M.
furfur’ la özdeştir fakat taksonomik olarak M. sympodialis’ e yakındır. 26 S’lik ve ITS
bölgelerinden rekombinant DNA dizi analizi ile bunu tanımlamışlardır. Beş suş M.
sympodialis’ in bir varyantından farklı türü göstermiştir39. Sonraki tarihlerde yine Sugita
ve ark. AD’lilerde yeni bir tür olan M. japonica‘yı tanımladılar. Bu tür Tween 40,
Tween 60’ı asimile edebilirken; Tween 20 ve Tween 80‘i asimile edemez37 .
2.14.4. Malassezia (Pityrosporum )Folliküliti
Pityriasis versicolor gibi Malassezia follükülitide Malassezia’nın kolonizasyou
ile ilgilir. Malassezia follüküliti (MF), iyi huylu bir hastalıktır ki özellikle boyun ve
gögüste püstüller ve papüller şeklinde karaekterize edilir. Şartlar genellikle yaygındır ve
sık sık akne gibi iyi tanınmaz. MF’yi ilk olarak Weary 1969’da tanımlamıştır ve Potter

35
ve ark. 1973’de histolojik ve klinik olarak ayırarak identifiye etmiştir. Kaşıntı,
komedonların varlığı ile ilgilidir ve yüzdeki lezyonlar iki hastalık arasındaki farkı
kolaylaştırır. MF çoğunlukla tropikal ülkelerde veya ılıman bölgelerde yazın meydana
gelir. Derideki tıkanma bu hastalıkta önemli bir rol oynar ve antibiyotik tedavisi
(tetrasiklin), kortikosteroid ve immunsupresiflikle ilgilidir. Diabetes mellitus,
immunsistemi bozuk olan kişiler, Cushing ve Hodgkin hastaları, sistemik kandidozis gibi
altta yatan bazı durumlar bu hastalığa zemin hazırlar ve bu kişilerde mortaliteye sebep
olabilir. Hastalık gençlerde ve orta yaşlılrda görülmekle beraber en sık 13-45 yaş arası
kişilerde görülür213.
Her ne kadar maya hürelerinin hiflere transformasyonu sadece PV görülsede, MF’li
hastaların histolojik muayenesinde da benzer şey görülür. Wood ışığı muayenesinde
beyaz mavi veya beyaz renkte floresan verir. Malassezia maya mantarı’ nın büyük bir
miktarı saç folüküllerine invaze olur. Bu invazyon, eritramatöz papüllerin gelişimi ile
sonuçlanır ve bazen papüller hem asemptomatik veya kaşıntılı olabilir. Folüküllerdeki
steroid aknenin bileşiminde ve akne vulgariste M. furfur (orbiculare ve ovale)
tanımlanmıştır. Genelllikle folüküllerde Staphylococcus ve Propionibacteri ile birlikte
Malassezia maya mantarı da görülür. Bazı araştırıcılara göre Malassezia follükülütleri
polimorf bir bozukluktur. Onlar en yaygın lezyonları, molluskoid ve kubbe
komedopapül (2-3 mm çapında) olarak tanımlamışlardır. Bununla beraber, birkaç oluda
hastalar nodül, püstül ve kist içerir. Bu durum tropikal iklimlerde görüldüğü
için(Filipinler), Malassezia follükülleri ılıman iklimlerde daha yaygın görülür214,215.
Eğer follükülitli olgularda, follükülüten biyopsi alınırsa, folüküllerde bol miktarda
tomurcuklanmış maya hücreleri ve birkaç hifler görülür. Aynı zamanda direk
mikroskobik incelemeyle mikroorganizma de görülmektedir. Bu tartışmalar bu
hastalıkta Malassezia’nın patojenik rolü olduğunu göstermektedir. Pityriasis versicolor
gibi Malassezia follükülütleri daha çok sırt, gögüs, omuzların üst kısmında görülür214,216
.Bazı coğrafik bölgelerde özellikle nemli bölgelerde ve tropikal bölgelerde yüzde de
görülür.
Saç folükülleri de ayrıca kendine ait karakteristik bir görünümü vardır. Saç folükülleri
genişlemiş ve şişme eğilimindedir ve sık sık keratin materyali ile doludur. Ayrıca
mayalar aşırı ürer ve folükülleri tıkamasına izin verir.

36
Tedavide topikal veya oral antifungallar MF‘nin tedavisinde en etkili ajanlardır.
Hastalar oral itrakonazolle veya haftalık flukonazolle başarıyla tedavi edilebilir.
2.14.5. Gougerot-Carteaud’un Retikulat (ağsı) ve Birleşik Papillomları
Malassezia’nın deride yapmış olduğu nadir kutanöz hastalıklardan biridir.
Birbirine karışan hiperkeratik papüller, gri-kahverenkli pigmentleri ile karekterize
edilir. Gövde üzerinde lokalize olur ki bu durum tartışmaya yol açar. Gougerot-
Carteaud’ un retikulat ve birleşik papillomları (CRP)’nın tanısı MF gibidir. Her ikisi de
tanısı stratum corneumda lipofilik maya mantarının direk mikroskop ve histopatoloji
incelemesine dayanır ve sistemik topikal ve antifungal cevaba karşı olumlu cevap verir.
Robert ve Lachapelle 1969’ da, CRP’ de P. orbiculare’ nin kolonizasyonuna karşı
konakçıda özel bir cevap oluşturabileceğini göstermişlerdir. Ancak, Nordby ve Mitchell
986’ da sadece 19 hastanın 5’ inde selenyum sülfitli topikal tedaviye karşı cevap
verdiklerini söylemişlerdir. Ayrıca iki olguda vitamin D’ nin analogları topikal ile
birlikte kalsipotriol ve takalsitol başarıyla uygulanmıştır217.
2.14.6. Psöriasis
Psöriasis hastalığındaki Malassezia’nın rolü hala tanımlanmamıştır. Fakat
birkaç raporda psöriasin gelişimi ile Malassezia maya mantarı arasında bir ilişki vardır.
Psöriasis epidermisteki hiperkeratinizayon ve hiperproliferasyon olarak tanımlanır.
Genelde en yayagın olgu mayayla ilgilidir ki maya hücreleri kafa bölgesini kapsama
eğilimindedir. Bu hipotez, kafa psöriaslilerde ketakonazole karşı oluşan cevabı destekler
ve kafada olan M. ovalis ( M. restricta) olduğunda kafa psöriasis ile aralarında bir ilişki
olduğu bulunmuştur. Ama psöriasislilerde sünnet derisinde Malassezia maya mantarı
rol oynayabilir218 .
Klinik olarak psöriasis lezyonları SD’ye benzer ancak lezyonların histolojik
görünüşü farklıdır. SD’lilerden alınan biyopsi örnekleri, eski lezyonlar bu karakterlerini
kaybetse ve psöriasise benzemeye başlamasına rağmen, SD süngerimsi şekilde görünür.

37
Bu lezyonlar sık sık ortokeratotik ve parakeratotik hücrelerin tıkanmasıyla ve sinir
hücrelerinin düzensiz olmasıyla tanımlanır220.
Psöriasis ayrıca genetik faktörler etkilidir. Bu yüzden araştırmacılar psöriasisli
hastaların immun reaksiyonlarını incelemişledir. Bu kişiler hem Malassezia maya
mantarına hem de bu mantarın ayrılmış proteinlerine karşı immunolojik cevap
vermişlerdir.
Gupta ve ark., SD ve psöriasisli hastaların hepsinde altı tane Malassezia türünü
izole etmişlerdir ama en çok ta M. globosa türünü izole etmişlerdir. Bu tür eşit sıklıkla
gövde, kafa ve alından izole etmiştir. Ancak son zamanlarda148 yapılan çalışmada hem
sağlıklı ki şilerde hem de psöriasisli kişilerde yapılan çalışmalarda Malassezia türlerinin
yayılımı önemli ölçüde farklı yayılım göstermiştir. M. globosa ( %55) ve onun maya
formu psörisisli hastalarda en baskın türdür. Sonra M. slooffiae ( % 18) ve M. restricta (
%10) en yaygın tür olarak izole edilmiştir219.
2.14.7. Malassezia Yenidoğan Püstülozu
1999 yılından beri çok az olguda yenidoğanın yüzünde ve boynunda non-
folüküler papüllopüstüller tanımlanmıştır. Tanı direk mikroskobi ve histopatoloji
yoluyla yapıldığında tipik Malassezia maya mantarı gösterir. Dahası, lezyonlar
ketakonazolle topikal terapiden sonra ortadan kalkar. Rapalanoro ve ark. bu durumu
neonatal akne olarak farztmişlerdir. Đki çalışmada M. sympodialis bu lekelerin
başlamasına neden olduğu gösterilmiştir221.
2.14.8. Otitis
2000’de Avusturalya’ da M. sympodialis’in otitis eksternanda maling bir tümör
olsusu rapor edilmiştir. Bu hasta tip II diabetes mellitusu vardı. Bu kişide birkaç
infeksiyon gelişti. Bu infeksiyon dış kulak kanalı etrafında olmaya başladı ve temporal
kemiğik ve yanındaki yumuşak dokunun yıkımıyla ilerledi. Đnfeksiyon intravenöz
amfoterisin B ve flukonazol terapisinden sonra total iyileşme gösterdi.

38
Hayvanlarda özellikle köpeklerde otitiste M. pachydermatis türlerinin rolü veteriner
takımları tarafından iyice tanımlanmıştır118,.
2.14.9. Onikomikoz
Malassezia’nın da dahil olduğu birkaç onikomikozis rapor edilmiştir. Ayrıca,
tırnak materyalinde Malassezia’nın varlığı seyrek olarak bulunmaktadır. Kutanöz
floranın bir üyesi olarak subungual kolonizasyon tarafından sebep olur ki birçok belirsiz
olgunun ilişkisi belirtilmiştir. Malassezia maya mantarı, sıklıkla Candida spp. ile ilgili
tırnağın distalinde olan onikolizislilerden elde edimiştir. AIDS’ lilerin infekte
tırnağından M. furfur izole edilmiştir. Ve Brezilya’da onikomikozisli hastalarda ciddi
bir şekilde izole edilmiştir222.
2.15. Moleküler Đdentifikasyon Yöntemi
Lipofilik olan Malassezia cinsi maya mantarı yıllarca kompleks ve
tanımlanmayan bir grup olarak kalmıştır ve mikolojik özellikten çok epidemiyolojik ve
klinik özelliklere dayanmıştır. Önceden dört tür gösterilmiştir: P. ovale, P. orbiculare,
P. pachydermatis ve P. canis. Seksüel döngü yokluğunda maya morfolojisinin uygun
sınıflandırması ve üreme için kompleks besi yeri gereksinimi bu durum için bir sebeptir.
Moleküler metodolojiye geçince, özellikle geniş ribozomal RNA altbirim dizi
analizi(LSU) bu pronlemlerin çözümüne yol açmıştır. Bir kez veriler genetik olarak
tanımlandı. Rutin mikrobiyoloji laboratuarında kullanılan karekteristik anahtar olan
uygulamalar tanımlamak için moleküler yöntem mümkündür. Bununla beraber birçok
metod zaman alıcıdır( kültürse üremeden 4 gün sonra) ve laboratuar çalışmasındaki
yüksek bir standart ister. Git gide laboratuvarlar PCR ile donanımlıdır. Bununla beraber
moleküler yöntemler çalışmanın çalışmanın amacına(tür identifikasyona, epidemiyoloji
için tür tayini) odaklanmalıdır 51,99,100.

39
2.16.Tedavi
Pityriasis versicolor topikal ve oral antifungal ajanların geniş bir kısmına cevap
verir. Malassezia’nın maya formundan hif formuna dönüşümü endojen faktörlerden
dolayı düşünülür. Rekürrensi yaygındır ve klinisyenler bunun için profilaktik tedaviyi
düşünürler.
2.16.1.Topikal Tedavi
2.16.1.1. Non- spesifik antifungal tedavi
Bu ajanlar göreceli olarak konuşulur, pityriasis versicolor için eski tedaviler
mümkündür ve büyük bir kısmı esas olarak fungal ajanlara karşı direk olarak aktiviteye
sahip değildir. Genel olarak fiziki olarak davranırlar ve/veya kimyasal olarak stratum
korneumdaki dokuları uzaklaştırırlar.
Selenyum sülfid, %2.5 losyon içerir, krem veya şampuandır. Pityriasis versicolor‘
un tedavisinde etkilidir. Bazı hastalar bu ilaçların tatsız kokusundan şikayeçidir.
Propilen glikol, a keratolitik ajan diğer topikal medikasyonlar içinde baz alarak
sık sık kullanılır. Kendi kendine pityriasis versicolor’ da etkili olduğu gösterilmiştir.
Salisilik asitli merhemler, benzoik asit ve salisilik asit içerir. Benzoik asit fungisitik
aktivitesi için salisilik asit kertolitik asit içindir. Bu tedavi pityrisis versicolor’ a karşı
etkili olduğu gösterilmiştir.
Sülfür ve salisilik asit, birkaç çalışmada PV’a karşı bu kombinasyonun etkili olduğu
gösterilmiştir.223-225
2.16.1.2 Spesifik topikal antifungal tedavi
Azoller, azoller fungisitik bir etkisi vardır. Ergesterol biyosentezini inhibe eder ve mantar hücre membranını formasyonunu parçalar.
Ketakonazol: % 2 krem formülasyonu rapor edilmiştir. % 2’lik ketakonazol şampuan ve jeli tedavinin ilk gününde etkili olabilir.

40
Flukonazol: Diğer topikal azoller gibi pityriasis versicolor’ un tedavisinde kullanılan bir triazoldur. % 2’ lik şampuanı tedavide etkilidir.
Terbinafin: Terbinafin alkil amin sınıfının bir antifungal üyesidir. Bu ilaç
tiyokarbonat gibi sterol sentezi sırasında squalen oksitlenmesini inhibe eder. Bu yüzden
mantarlardaki membran sentezini parçalar226.
2.16.2. Oral tedavi
Oral tedavinin avantajı artmış hasta uyumunu içerebilir. Oral tedavi daha uygun ve
hasta için daha az zaman alıcıdır. Bununla beraber bazı klinisyenler pityriasis versicolor
derinin geniş bir kısmını etkilediği zaman oral tedaviyi sistemik tedaviden ayırmak için
tercih edebilirler.
Ketakonazol: Pityriasis versicolor’a karşı geliştirilen ilk etkili oral tedavidir.
Đtrakonzol: Đtrakonazolun ilaç absorbsiyonu, ağızdan alındığında artar. Đtrakonazol
5 veya 7 gün 200mg/dozda alındığında etkilidir.
Flukonazol: Pityriasis versicolor’un tedavisinde potansiyel kullanımı araştırılmıştır.
Flukonazol etkili bir tedavidir.

41
3.GEREÇ VE YÖNTEM
Çalışmada, Çukurova Üniversitesi Tıp Fakültesi Dermatoloji Anabilim Dalı
Polikliniğine pityriasis versicolor nedeni ile Eylül 2008 - Mayıs 2009 tarihleri arasında
başvuran 24’ü kadın, 33’ü erkek toplam 57 olgu incelendi. Bu olgularda, farklı klinik
alma yöntemleri (bistüri ve selofan bant) ve farklı besiyerleri (modifiye-Dixon agar ve
Leeming-Notman agar) karşılaştırıldı. Tür düzeyinde tanı, Çukurova Üniversitesi Tıp
Fakültesi Mikrobiyoloji Anabilim Dalı, Mikoloji Bilim Dalı laboratuvarında yapıldı.
Çalışmada katılımcıların oluru alındı ve önceden hazırlanan anket formunda yer alan
soruları yanıtlamaları istendi. Çalışma öncesinde Çukurova Üniversitesi Tıp Fakültesi
Etik Kurulunun onayı alındı.
3.1. Klinik örneklerin alınması
Pityriasis versicolor’lu olguların lezyonlu bölgesi önce %70’lik etil alkolle
silindi ve kuruması için 3-5 dakika beklendi. Sonra steril bir bistüri yardımı ile klinik
örnekler alındı. Epitel kazıntı örnekleri modifiye-Dixon ve Leeming-Notman agar’a
ekildi ve 32°C’de etüvde 5 gün bekletildi. Sonra, aynı olgunun aynı lezyonlu bölgesine
steril bir selofan-bant yapıştırıldı. 10-15 saniye kadar bekletilip çekildi ve banta gelen
kepek örnekleri yine modifiye-Dixon ve Leeming Notman agarların yüzeyine bastırıldı
ve 3-5 sn. sonra çekidi.
3.2. Wood ışığı muayenesi
Pityriasis versicolor’lu olguların lezyonlu bölgesi karanlık bir odada Wood
ışığında incelendi. Sarı-yeşil renkte flöresans varlığı araştırıldı. Bu renkte flöresans
oluşturanlar pozitif olarak kabul edilirken, renk oluşturmayanlar ise negatif olarak
değerlendirildi.

42
3.3. Mikolojik inceleme
3.3.1 Doğrudan mikroskop incelemesi
Bu amaçla, lezyonlu bölgeden alınan klinik örnekten %15’lik KOH ile lam-
lamel arası preparat hazırlandı. Kuru zeminde 20 dakika süre ile bekletildi. Daha sonra
ışık mikroskobunda x10’luk ve x40’lık büyütme ile köfte-makarna (kalın duvarlı spor
ve kısa-güdük hif) varlığı yönünden incelendi.
Şekil 3.1: Lezyondan alınan örneğin %10’nluk KOH ile incelemesi (spor ve hif)
3.3.2 Kültür
Bistüri ve selofan-bant ile alınan örnekler steril petride hazırlanan modifiye
Dixon ve Leeming- Notman agar’a ekildi ve en çok sonra iki saat içerisinde ÇÜTF
Mikrobiyoloji Anabilim Dalı Mikoloji Bilim Dalı laboratuvarına ulaştırıldı. Besiyerleri
aerop koşullarda, 32°C’de 5 gün süre ile inkübe edildi. Üreme varlığı yönünden günlük
olarak incelendi. Bu süre sonunda üreme sonuçları değerlendirildi ve tanı testleri
yapıldı.
modifiye-Dixon besiyeri (MD)
% 3.6 Malt extract
% 0.6 Pepton
% 2 Öküz safrası
% 1 Tween 40
Hif
Spor

43
% 0.2 Gliserol
% 0.2 Oleik asit
% 1.2 Agar
% 0.5 Kloramfenikol (Sigma, Steinheim, Germany)
1g/ L Siklohekzimit (Sigma, Steinheim, Germany)
pH= 6
Hazırlanmış karışım otoklavda 121°C’de 15 dk. steril edildi. Daha sonra besiyeri
42°C‘ye soğutulduktan sonra oleik asit, gliserol, Tween 40, kloramfenikol ve
siklohekzimit ilave edildi. Steril petrilere 4 mm kalınlığında döküldü. Aerop koşullarda,
32 °C’de 5 gün inkübasyona bırakıldı.
Leeming-Notman besiyeri (LN)
Pepton 10 gr
Glikoz 5 gr
Yeast extract 0,1 gr
Öküz safrası 4 gr
Gliserol 1 ml
Gliserol monosterat 0,5 gr
Agar 12 gr
Tween 60 0,5 ml
Đnek sütü 10 ml
Kloramfenikol 50 gr/ ml (Sigma, Steinheim, Germany )
Siklohekzimit 20 mg (Sigma, Steinheim, Germany)
pH= 6,2
Hazırlanan karışım otoklavda 121°C’ de 15 dk. steril edildi. Daha sonra besiyeri
42°C‘ye soğutulduktan sonra gliserol, Tween 60, inek sütü, kloramfenikol ve
siklohekzimit ilave edildi steril petrilere 4 mm kalınlığında döküldü. Besiyeri

44
soğuduktan sonra tüm klinik izolatların pasajları yapıldı. Aerop koşullarda 32°C’de bir
hafta inkübe edildi. Leeming-Notman besiyeri hem klinik izolatların saklanması hem de
besiyerlerinin karşılaştırılması amacı ile kullanıldı.
3.3.3.Tanı testleri
Sabouraud glikoz agar (SGA, Merck, Darmstadt, Germany)
Glikoz 40 gr
Pepton 10 gr
Agar 15 gr
Damıtık su 1000 ml
Ticari olarak elde edilen besiyeri otoklavda 121°C’de 15 dk. steril edildi. Klinik
örneklerden izole edilen maya mantarlarının SGA’a pasajları yapıldı. Aerop koşullarda,
32°C’de 5 gün süre ile inkübe edildi. Üreme olmaması lipofilik maya mantarının varlığı
açısından kanıt oluşturdu.
Katalaz aktivitesi
Katalaz reaksiyonu mikro-organizmada hidrojen peroksidaz enziminin varlığını
belirlemek amacıyla yapıldı. Bunun için steril bir lama bir damla hidrojen peroksit
damlatıldı. Üzerine bir öze maya kolonisi ilave edildi. Gaz çıkışına göre oksijenin
varlığına bakıldı.

45
Tween identifikasyon testi
Klinik izolatların modifiye-Dixon veya Leeming-Notman besiyerinde pasajı
yapıldı. Önce SGA besiyeri hazırlanıp otoklavda 121°C’de steril edildi. Yeteri kadar
soğuduktan sonra % 0.05 kloramfenikol ve % 0.05 siklohekzimit ilave edildi. Sonra
steril tüplere 5 ml steril distile su kondu ve üzerine bir öze dolusu maya kolonisi alınıp
10-15 saniye kadar vortekslendi. Hazırlanan maya karışımında steril enjektör
yardımıyla 2 ml alınıp steril petrilere dağıtıldı. Petrilere 16 ml SGA dağıtılıp hafifçe
karıştırılıp soğumaya bırakıldı. Soğuma işleminden sonra steril pipet ucu ile petride
karşılıklı dört tane ve tam ortasına da bir kuyu açıldı. Bu kuyuları sırası ile Tween 20,
Tween 40, Tween 60 ve Tween 80 ve ayrıca tam ortadaki ortasındaki kuyuya
chremophor-EL’den bir damla ilave edildi. Aerop koşullarda, 32°C’de bir hafta
inkübasyona bırakıldı ve presipitasyon zonu varlığı yönünden incelendi. Sonuçlar
günlük olarak kaydedildi.
O Şekil 3.2: Tween identifiksyon testi
Chremophor-EL slant (polyether of oil and ethylene )
65 gr SGA (SGA, Merck, Darmstadt, Germany)
10 ml chremophor EL (Fluka Biochemika Germany)
Ticari olarak hazırlanmış olan SGA besiyeri otoklavda 121°C’de 15 dk. steril
edildi. 45°C’ye soğuduktan sonra steril bir pipet yardımıyla Chremophor-EL (polyether
T 20
2222
T40
C-EL
T 60 T80

46
of oil and ethylene) 10 ml damlatıldı. Karıştırıldıktan sonra steril tüplere dağıtıldı ve
yatık olarak soğumaya bırakıldı. Tüm izolatlar steril bistüri ile yatık olarak ekildi.
Aerop koşullarda, 32°C’de 15 gün inkübasyona bırakıldı. Üreme sonuçları günlük
olarak takip edildi ve Chremophor-EL’yi kullanma özelliği araştırıldı.
Tween 60 eskülin besiyeri
10 gr pepton (Acumedia, Baltimore, Maryland)
10 gr glikoz
2 gr yeast extract
0.5 gr ferric amonyum sitrat
5 ml Tween 60
15 gr agar
Karışımlar hazırlandı. Ticari olarak hazırlanmış olan Tween-60 eskülin besiyeri
otoklavda 121°C’de 15 dk. steril edildi. 45°C’ye soğuduktan sonra steril bir pipet
yardımıyla Tween 60 ilave edildi. Steril tüplere dağıtılıp yatık olarak donmaya bırakıldı.
Soğuma işleminden sonra tüm izolatlar steril öze yardımıyla dibe batık olarak ekildi.
32°C ve 37°C’lerde 15 gün etüvde inkübasyona bırakılarak β- glikozidaz aktivitesine
bakıldı ve siyah zon oluşumu günlük olarak izlendi.
Chromogenic Candida Agar (Oxoid, Basingstoke, Hampshire, England)
15.6 gr Chromogenic Candida Agar (Oxoid, Basingstoke, Hampshire, England)
500 ml Distile su
Hazırlanan karışım 95°C’de benmaride 1.5-2 saat bekletilip steril edildi. Sonra
45°C’ye kadar soğutuldu ve üzerine etil alkolde çözülmüş Chromogenic Candida
selektif suplementi konuldu. Steril petrilere 4 mm kalınlığında döküldü. Besiyeri

47
donduktan sonra klinik izolatlar ekildi ve aerop koşullarda, 32°C’de 5 gün
inkübasyona bırakıldı. Koloninin rengi ve presipitasyon yönünden incelendi.
modifiye-Dixon besiyerinde presipitasyon testi
% 3.6 Malt extract (Oxoid, Basingstoke, Hampshire, UK)
% 0.6 pepton (Oxoid)
% 2 Ox bile (Oxoid)
% 1 Tween 40 (Sigma, St Louis, Missouri, USA)
% 0.2 gliserol (Tokyo, Tokyo, Japan)
% 0.2 Oleik asit
% 1.2 Agar (Oxoid)
% 0.5 Kloramfenikol (Sigma, Steinheim, Germany)
1g/ L Siklohekzimit (Sigma, Steinheim, Germany)
pH: 6
Hazırlanan karışıma belirli oranda distile su ilave edildi ve otoklavda besiyeri
otoklavda 121° C’de 15 dk. steril edildi. 45°C’ye soğuduktan sonra steril bir pipet
yardımıyla Tween 40, gliserol, oleik asit ilave edildi. Steril petrilere 4 mm kalınlığında
döküldü ve soğumaya bırakıldı. Tüm klinik izolatlar aerop koşullarda, 32°C’ de 5 gün
inkübasyona bırakıldı. Süre sonunda koloninin etrafındaki presipitasyon zonu varlığı
araştırıldı.
Isı testi
Klinik izolatlar her iki çalışma besiyerlerine pasajlandı ve 32°C, 37°C ve 40°
C’lerde üreme yetenekleri yönünden 7 gün süre ile değerlendirildi.

48
Laktofenol pamuk mavisi (Merck, Darmstadt, Germany)
Temiz bir lama bir damla laktofenol pamuk mavisi damlatıldı. Üzerine steril bir
öze ile alınan bir miktar maya kolonisi konup karıştırıldı ve temiz bir lamel ile kapatıldı,
x10’luk ve x 40’lık objektifte incelenerek küremsi, silindirik ve/veya oval şekilde olan
maya hücrelerinin varlığı incelendi.
Gram boyama
Maya hücrelerinin morfolojisi ayrıca Gram boyası ile incelendi.
Virkon
Mikoloji laboratuvarlarında sık görülen kontaminasyon sorununun önlenmesi
amacı ile Virkon’un 10gr’nın 1 lt distile su içerisinde çözülmesi sonucu elde edilen
karışım haftada iki kez, güvenlik kabini her çalışmadan sonra düzenli olarak kullanıldı.
Đstatistiksel inceleme
Đstatistiksel incelemede SPSS 10.0 paket programı ve Epi Đnfo version 3.2 kullanıldı.
Gruplar arasında farkın değerlendirilmesinde Chi-Square testi, Kruskal-Wallis ve
Mann-Whitney testi kullanıldı. P < 0.05 anlamlı kabul edildi.

49
4. BULGULAR
Çalışmada, pityriasis versicolor tanılı 57 olgu incelendi. Olguların yaş ortalaması
± 26,2 idi. Hastaların 34’ü (%56.1) erkek ve 23’ü (%43.9) bayan idi. Hastalar yaş
gruplarına göre beş gruba ayrıldı: grup I,1-15 yaş; grup II, 16–30 yaş; grup III, 31–45
yaş; grup IV, 46–60 yaş ve grup V>60 yaş. Hastların yaş gruplarına göre dağılımı Tablo
1’de gösterilmiştir. PV’lu hastaların klinik özellikleri Tablo 2’de, bistirü ve selofan
bantla alınan klinik örneklerin modifiye-Dixon ve Leeming-Notman agar’da üreme
durumu Tablo 3’de; katalaz tepkimesi, Candida Chromogenic agarda pigment oluşumu,
Chremophor EL ve Tween 60 eskülin besiyerindeki üreme sonuçları ve presipitasyon
oluşumu Tablo 4’de; Sabouraud glikoz agarda, 37°C ve 40 C°’deki üreme özellikleri
tablo 5’de gösterilmiştir. Tween asimilasyon test sonuçları Tablo 6’da gösterilmiştir.
Tablo 4.1. Pityriasis versicolor’lu olguların yaş gruplarına göre dağılımı
Grup Yaş Sayı
I 1-15 10
II 16-30 29
III 31-45 11
IV 46-60 6
V >60 1
Toplam - 57

50
Şekil 4.1: Pigmentasyon özelliğinin cinsiyete göre dağılımı
Olguların lezyonlara göre dağılımı incelendiğinde hastaların; 18’i (%31,6) hipo-
pigmente iken, 39’u (%68,4) ise hiper-pigmente idi. Hastaların hiçbirinde hem hipo-
hem de hiper-pigmentasyon görülmedi.
Wood ışığı sonuçları incelendiğinde; hastaların 32’sinde (%56.1) olumlu (+) iken,
25’inde (% 43.9) olumsuz (-) idi. Wood ışığı olumsuz olanların % 68.0’ı hiperpigmente
iken % 32.0’ı hipopigmente olarak bulundu. Wood ışığı pozitif olanların % 68.8’i
hiperpigmente, % 31.2’si hipopigmente idi. Wood ışığı ile lezyon özelliği arasındaki
ili şki incelendiğinde anlamlı bir ilişki olmadığı belirlendi (p=0.952).
Doğrudan mikroskop incelemesi
Pityriasis versicolor tanılı 57 olgunun boyun, sırt, göğüs, yüz ve diğer
bölgelerinden alınan örneklerin, KOH ile lam-lamel arası incelemesinde; 48’inde

51
(%84.2) köfte-makarna (spor ve hif) görüntüsüne rastlandı, ancak 9’unda (%15.8)
yalnızca spor görüldü. Doğrudan mikroskopi ile lezyon özelliği arasındaki ilişki
incelendiğinde direkt mikroskopisi negatif olanların %55.6’sı hiperpigmente iken,
%44.4’ü hipopigmente idi. Doğrudan mikroskopisi pozitif olanların %70.8’i
hiperpigmente iken %29.2’si hipopigmente idi. Doğrudan mikroskopi ile lezyon özelliği
açısından anlamlı olmadığı görüldü (p=0.366).
Klinik örnekler
Pityriasis versicolor tanılı 57 olgudan alınan klinik örneklerin üreme sonuçlarına
göre dağılımının incelemesinde; yalnızca bistüri ile alınan epitel kazıntısı örneklerinin
48’inde (%84.2), yalnızca selofan bantla alınan örneklerin 22’sinde (%38.6), hem
bistüri hem de selofan bantla alınan örneklerin 26’sında (%45.6) modifiye Dixon veya
Leeming-Notman agarda üreme oldu. Bistüri ve selofan bantla alınan klinik örneklerin
6’sı (%10.5) ne modifiye Dixon ne de Leeming-Notman agarda üremedi. Ayrıca,
yalnızca bistüri ile alınan örneklerin 7’si (%12.3) modifiye-Dixon besiyerinde üredi
ancak pasajlarında üreme olmadığı için tür tanısı yapılamadı ve Malassezia sp. olarak
adlandırıldı.
Modifiye-Dixon ve Leeming-Notman besiyerlerinde oluşan presipitasyonun
araştırılmasında; 37 izolatta (%64.9) 5 günde presipitasyon saptandı, ancak 17 izolatta
(% 29.8) presipitasyon görülmedi. Üreme olmayan izolatlarda presipitasyon oluşumu
değerlendirilemedi.
Hastalarda kepeklenme varlığı araştırıldığında; 55’inde (% 96.5) kepek vardı, ikisinde
(%3.5) ise kepek yoktu. Kepeklenme olmayan 2 olgununda lezyonları hipogigmente idi.
Kepeği olanların % 67.3’ü hiperpigmente, 32.7’si hipopigmente idi. Bu sonuca göre
kepek ile lezyon arasında istatistiksel açıdan bir ili şki olmadığı görüldü (p=0.328).
Hastalığın süresi incelendiğinde; hastaların %52.6’sı rekürren, %47.4’ü primer
infeksiyondu. Hastalığın süresi ile lezyon arasındaki ilişki incelendiğinde primer
infeksiyonların %51.9’u hiperpigmente, %35.9’u hipopigmente idi. Rekürren
infeksiyonların %83.3’ü hiperpigmente iken %16.7’si hipopigmente idi (Şekil 5).

52
Hastalığın süresi ile lezyonun özelliği arasında istatiksel olarak anlamlı ilişki olduğu
belirlendi (p=0.01).
Primer/ Rekürren
RekürrenPrimer
Per
cent
80
60
40
20
0
Lezyon Özelliğ
Hiperpigmente
Hipopigmente
28
72
64
36
Şekil 4.2: Hastalığın süresi ile lezyonunu özelliği arasındaki ilişki
Biyokimyasal Testler
Katalaz aktivitesi
Üreme saptanan 52 klinik örneğin katalaz aktivitesi olumlu (%100) bulundu
(Tablo 4).

53
Tanı testlerinin sonuçları
Chremophor EL besiyerinde üreme
Chremophor EL besiyerinde izolatların 45‘i 32°C’de 6-7 günde presipitasyon
oluşturdu, chremophor EL’yi kullandı (Tablo4).
Tween 60-eskülin agar’da üreme
Tween 60-eskülin besiyerindeki β-glukozidaz aktivitesi 32°C’de beş günlük
inkübasyon ile değerlendirildi. Đzolatların 21’i eskülini kullandı ve tüm besiyerinin
rengini siyaha çevirdi. Örneklerin 14’ü besiyerini rengini griye çevirdi ve tam siyah
renk oluşumu gözlenmedi ve hafif siyah zon oluşturdu. Bu besiyerinde izolatlar
besiyerinin hemen üst kısmında hafif siyah zon oluştururken, dip kısmında herhangi bir
zon oluşturmadı. Đzolatların 10 tanesi eskülini kullanmadı ve siyah renk vermedi ve
besiyerinin renginde bir değişiklik olmadı (Tablo 4).
Chromogenic Candida Agar’da Üreme
Candida Chromogenic Agar’da (CCA) izolatların pigment oluşumu incelendiğinde;
izolatların 45’i 32°C’de dört günlük inkübasyondan sonra pembe mor rengini verdi ve
başka bir renk oluşturmadı (Tablo 4).
Isı testi ve SGA’da Üreme
Klinik izolatların 37°C ve 40°C’deki üreme performanslarına incelendiğinde; 45
örneğin hepsi 37°C’de 5. günde iyi ürediği; 40°C’de örneklerin 20‘si iyi, 5’inin ise
hafif ürediği görüldü (Tablo 5). Diğer 27 izolatın 40°C’de ürediği saptandı. Isı testinin
tür dağılımına göre incelendiğinde; 37°C deki üremenin anlamlı olduğu, 40°C ile
uyumlu olmadığı görülmüştür.

54
Tween 20, 40, 60 ve 80 ile üreme
Malassezia’ların identifikasyon testi olan Tween testinin sonuçları
incelendiğinde; izolatların 26’sı (%45,6) Tween 20, 40, 60 ve 80’nin hepsinde
presipitasyon görüldü. Bu sonuca göre 26 izolat M. furfur olarak tanımlandı. Đzolatların
9 tanesi (%15.8) Tween 20, 40, 60 da presipitasyon zonları oluşturdu ve M.slooffiae
olarak identifiye edildi. 4’ü (%7.0) Tween 40, 60, 80 de presipitaston zonu oluşturdu ve
M. sympodialis olarak tanımlandı. Đki klinik (%3,5) izolat Tween paternlerinin
hiçbirinde presipitasyon zonu oluşturmadı ve M. globosa olarak değerlendirildi. Üç
izolat (%19.7) ise Tween 40 ve 60’da presipitasyon zonu oluşturdu, ancak tür
identifikasyonu yapılamadı. Đzolatların 10’un (%19.6) tür identifikasyonu yapılamadı.
Bu sonuçlar Tablo 6’da gösterilmiştir. Tür ile lezyonun özelliği arasındaki ilişki
incelendiğinde tür tanımı yapılamayan hastaların %75.0’ı hiperpigmente, %25.0’ı
hipopigmente idi. Malassezia slooffiae olanların %77,8’i hiperpigmente, %25,0’ı
hipopigmente idi. Malassezia furfur olanların %57,7’si hiperpigmente, %42,3’ü
hipopigmente idi. Malassezia sympodialis olanların %100’ü hiperpigmente idi.
Malassezia globosa olanların %50’si hiperpigmente iken %50’si hipopigmente idi.
(Şekil 4.6). Đstatistiksel açıdan incelendiğinde tür ile lezyon özelliği arasında anlamlı bir
ili şki olmadığı görüldü (p=0.4).

55
Lezyon Özelliði
HipopigmenteHiperpigmente
Cou
nt
16
14
12
10
8
6
4
2
0
TUR
Tür yok
M.slooffiae
M. furfur
M.globosa
M.sympodialis
Şekil 4.3: Malassezia türlerinin pigmentasyon özelliğine göre dağılımı
Örnek alma yöntemleri karşılaştırıldığında Pityriasis versicolor’lu hastalardan
bistirü ile alınan klinik örneklerin daha iyi ve anlamlı olduğu; selofan bant ile alınan
örnekteki üremelerin daha zayıf olduğu belirlendi.
Örnek alma yöntemleri ile lezyon özelliği arasındaki ilişki incelendiğinde
hiperpigmente olan lezyonların %66.7’si (n=6) bistüri ile alınan örnekte üremedi;
%68.8’i (n=33) üredi. Hipopigmente olanların %33.3’ü (n=3) bistüri ile alınan örnekte
üremedi; %31.2’si (n=15) üredi. Bantta üreyen örneklerin %72.7’si hiperpigmente,
%27,.’ü hipopigmente idi. Bant ile alınan örneklerin fakat üremeyen örneklerin %
65.7’si hiperpigmente, % 34.3’ü hipopigmente idi.
Besiyeri performansları karşılaştırıldığında; klinik örneklerin modifiye-Dixon
besiyerinde daha iyi ürediği (% 82.5), Leeming-Notman besiyerinde ise daha az ürediği
(%73.7) belirlendi.

56
Besiyerlerine göre koloni morfolojisi ve sayısı incelendiğinde; modifiye Dixon
besiyerinde üreyen mayaların kolonilerinin daha büyük olduğu ve bu besiyerinde daha
çok ürediği; Leeming-Notman besiyerinde kolonilerin daha küçük ve az olduğu
gözlendi.
Modifiye-Dixon besiyerinde üreyen kolonilerin zon çaplarının tür dağılımı ile
ili şkisi incelendiğinde; literatürde belirtilen tür dağılımı ile uyumlu olduğu görüldü.
Çalışmada, vücudun güneş gören ve görmeyen bölgelerinin lezyon ile ilişkisi
araştırılmış ve istatiksel olarak güneş gören bölgeler ile lezyonun özelliği arasında
anlamlı bir ilişki bulunmuştur (p=0.096). Güneş gören bölgelerin %59.4’ü
hiperpigmente, %40.6’sı hipopigmente idi. Güneş görmeyenlerin %80’i hiperpigmente,
%20’si hipopigmente olarak bulundu.
Ayrıca, çalışmada saptanan bulgular Tablo 4.2-6’da verilmiştir.
Tablo 4.2. Bistirü ve selofan bantla alınan klinik örneklerin üreme sonuçlarına göre dağılımı
Klinik
izolat
Bistüri
(MD,LN) Selofan-bant(MD,LN)
Đzolat 1 Ø Ø
Đzolat 2 + +
Đzolat 3 + +
Đzolat 4 + +
Đzolat 5 + -
Đzolat 6 + -
Đzolat 7 + +
Đzolat 8 + +
Đzolat 9 + -
Đzolat 10 + -

57
Đzolat 11 + +
Đzolat 12 Ø Ø
Đzolat 13 + +
Đzolat 14 + +
Đzolat 15 + +
Đzolat 16 + -
Đzolat 17 + +
Đzolat 18 + -
Đzolat 19 + +
Đzolat 20 + +
Đzolat 21 + +
Đzolat 22 + -
Đzolat 23 + -
Đzolat 24 + +
Đzolat 25 + +
Đzolat 26 + -
Đzolat 27 + -
Đzolat 28 Ø Ø
Đzolat 29 + -
Đzolat 30 + -
Đzolat 31 + -
Đzolat 32 + +
Đzolat 33 + -
Đzolat 34 + +
Đzolat 35 + +
Đzolat 36 Ø Ø
Đzolat 37 + -
Đzolat 38 + +
Đzolat 39 + -
Đzolat 40 + -
Đzolat 41 + -

58
Đzolat 42 + -
Đzolat 43 + -
Đzolat 44 + +
Đzolat 45 + -
Đzolat 46 + -
Đzolat 47 + +
Đzolat 48 + -
Đzolat 49 + -
Đzolat 50 + -
Đzolat 51 + +
Đzolat 52 + -
Đzolat 53 + -
Đzolat 54 + -
Đzolat 55 + +
Đzolat 56 Ø Ø
Đzolat 57 Ø Ø
MD: Modifiye-Dixon agar;
LN: Leeming-Notman agar
Tablo 4.3. Đzolatların CCA, C-EL ve Tween 60 eskülin besiyerindeki üreme özellikleri, katalaz
aktivitelerinin sonuçları
Klinik
izolat CCA C-EL T-60 Eskülin Prespitasyon(MD,LN) Katalaz
Đzolat 1 Ø Ø Ø Ø Ø
Đzolat 2 Pembe-mor + 2+ ++ +
Đzolat 3 Pembe-mor + 3+ ++ +
Đzolat 4 Pembe-mor + 3+ ++ +
Đzolat 5 Pembe-mor + - ++ +
Đzolat 6 Pembe-mor + 2+ ++ +

59
Đzolat 7 ØØ ØØ ØØ -- ØØ
Đzolat 8 ØØ ØØ ØØ ++ ØØ
Đzolat 9 Pembe-mor + - ++ +
Đzolat 10 Pembe-mor + 3+ ++ +
Đzolat 11 Pembe-mor + - ++ +
Đzolat 12 Ø Ø Ø Ø +
Đzolat 13 Pembe-mor + 2+ ++ +
Đzolat 14 Pembe-mor + 3+ ++ +
Đzolat 15 Pembe-mor + - ++ +
Đzolat 16 Pembe-mor + - ++ +
Đzolat 17 Pembe-mor + 2+ ++ +
Đzolat 18 Pembe-mor + 2+ -- +
Đzolat 19 Pembe-mor + 3+ -- +
Đzolat 20 Pembe-mor + 3+ ++ +
Đzolat 21 Pembe-mor + 3+ ++ +
Đzolat 22 Pembe-mor + 2+ ++ +
Đzolat 23 ØØ ØØ ØØ ++ ØØ
Đzolat 24 Pembe-mor + 1+ -- +
Đzolat 25 Pembe-mor + - ++ +
Đzolat 26 Pembe-mor + 2+ ++ +
Đzolat 27 Pembe-mor + 2+ ++ +
Đzolat 28 Ø Ø Ø Ø Ø
Đzolat 29 Pembe-mor + 2+ ++ +
Đzolat 30 Pembe-mor + 3+ ++ +
Đzolat 31 Pembe-mor + 3+ ++ +
Đzolat 32 Pembe-mor + 3+ ++ +
Đzolat 33 Pembe-mor + 3+ ++ +
Đzolat 34 Pembe-mor + 3+ ++ +
Đzolat 35 Pembe-mor + 3+ -- +
Đzolat 36 Ø Ø Ø Ø Ø
Đzolat 37 Pembe-mor + 3+ -- +

60
Đzolat 38 Pembe-mor + 3+ ++ +
Đzolat 39 Pembe-mor + - -- +
Đzolat 40 Pembe-mor + 3+ ++ +
Đzolat 41 Pembe-mor + - -- +
Đzolat 42 Pembe-mor ± 1+ -- +
Đzolat 43 Pembe-mor + 2+ -- +
Đzolat 44 Pembe-mor + 3+ --
Đzolat 45 ØØ ØØ ØØ -- +
Đzolat 46 ØØ ØØ ØØ ++ +
Đzolat 47 Pembe-mor + 3+ -- +
Đzolat 48 ØØ ØØ ØØ ++ +
Đzolat 49 ØØ ØØ ØØ -- +
Đzolat 50 Pembe-mor + 3+ -- +
Đzolat 51 Pembe-mor + 2+ ++ +
Đzolat 52 Pembe-mor + 2+ -- +
Đzolat 53 Pembe-mor + 3+ -- +
Đzolat 54 Pembe-mor + 3+ ++ +
Đzolat 55 Pembe-mor + 3+ ++ +
Đzolat 56 Ø Ø Ø Ø Ø
Đzolat 57 Ø Ø Ø Ø Ø
+: pozitif üreme, -: negatif zon, 3+:simsiyah zon,
2+:hafif siyah zon,++: presipitasyon var,--
:presipitasyon yok, 1+:çok hafif siyah zon Ø:
üreme olmadı, ØØ: üredi ancak çoğaltılamadı
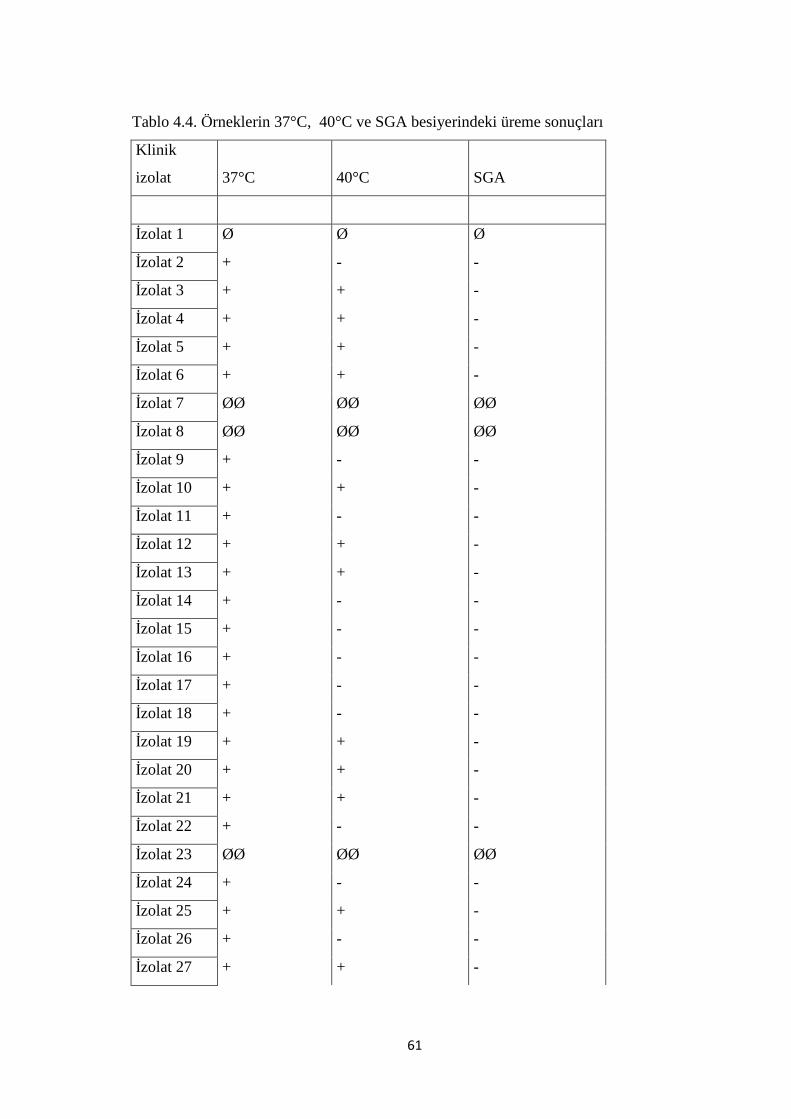
61
Tablo 4.4. Örneklerin 37°C, 40°C ve SGA besiyerindeki üreme sonuçları
Klinik
izolat 37°C 40°C SGA
Đzolat 1 Ø Ø Ø
Đzolat 2 + - -
Đzolat 3 + + -
Đzolat 4 + + -
Đzolat 5 + + -
Đzolat 6 + + -
Đzolat 7 ØØ ØØ ØØ
Đzolat 8 ØØ ØØ ØØ
Đzolat 9 + - -
Đzolat 10 + + -
Đzolat 11 + - -
Đzolat 12 + + -
Đzolat 13 + + -
Đzolat 14 + - -
Đzolat 15 + - -
Đzolat 16 + - -
Đzolat 17 + - -
Đzolat 18 + - -
Đzolat 19 + + -
Đzolat 20 + + -
Đzolat 21 + + -
Đzolat 22 + - -
Đzolat 23 ØØ ØØ ØØ
Đzolat 24 + - -
Đzolat 25 + + -
Đzolat 26 + - -
Đzolat 27 + + -

62
Đzolat 28 Ø Ø Ø
Đzolat 29 + - -
Đzolat 30 + - -
Đzolat 31 + - -
Đzolat 32 + + -
Đzolat 33 + + -
Đzolat 34 + - -
Đzolat 35 + + -
Đzolat 36 Ø Ø Ø
Đzolat 37 + ± -
Đzolat 38 + + -
Đzolat 39 + - -
Đzolat 40 + ± -
Đzolat 41 + - -
Đzolat 42 + ± -
Đzolat 43 + ± -
Đzolat 44 + + -
Đzolat 45 ØØ ØØ ØØ
Đzolat 46 ØØ ØØ ØØ
Đzolat 47 + + -
Đzolat 48 ØØ ØØ ØØ
Đzolat 49 ØØ ØØ ØØ
Đzolat 50 + + -
Đzolat 51 + - -
Đzolat 52 + - -
Đzolat 53 + + -
Đzolat 54 + - -
Đzolat 55 + + -
Đzolat 56 Ø Ø Ø Đzolat 57 Ø Ø Ø
+: pozitif üreme, -: negatif üreme, Ø: Üreme olmadı, ØØ: Üredi ancak çoğaltılamadı, ±: zayıf üreme

63
Tablo 4.5. Tween identifikasyon testi (Tween 20, 40, 60 ve 80)
Klinik
izolat Tween 20 Tween 40 Tween 60 Tween 80 Tür
Đzolat 1 Ø Ø Ø Ø Ø
Đzolat 2 + + + - M. slooffiae
Đzolat 3 + + + + M. furfur
Đzolat 4 + + + + M. furfur
Đzolat 5 + + + + M. furfur
Đzolat 6 + + + + M. furfur
Đzolat 7 ØØ ØØ ØØ ØØ ØØ
Đzolat 8 ØØ ØØ ØØ ØØ ØØ
Đzolat 9 + + + + M. furfur
Đzolat 10 + + + + M. furfur
Đzolat 11 + + + - M. slooffiae
Đzolat 12 Ø Ø Ø Ø Ø
Đzolat 13 + + + + M. furfur
Đzolat 14 - + + + M. sympodialis
Đzolat 15 + + + - M. slooffiae
Đzolat 16 + + + - M. slooffiae
Đzolat 17 + + + + M. furfur
Đzolat 18 + + + + M. furfur
Đzolat 19 + + + + M. furfur
Đzolat 20 + + + + M. furfur
Đzolat 21 + + + + M. furfur
Đzolat 22 + + + - M. slooffiae
Đzolat 23 ØØ ØØ ØØ ØØ Malassezia sp.
Đzolat 24 + + + + M. furfur
Đzolat 25 + + + + M. furfur
Đzolat 26 + + + + M. furfur
Đzolat 27 + + + + M. furfur

64
Đzolat 28 Ø Ø Ø Ø Ø
Đzolat 29 + + + + M. furfur
Đzolat 30 + + + + M. furfur
Đzolat 31 + + + + M. furfur
Đzolat 32 + + + + M. furfur
Đzolat 33 + + + + M. furfur
Đzolat 34 + + + - M. slooffiae
Đzolat 35 + + + - M. slooffiae
Đzolat 36 Ø Ø Ø Ø Ø
Đzolat 37 + + + + M. furfur
Đzolat 38 - + + + M. sypodialis
Đzolat 39 - - - - M. globosa
Đzolat 40 - + + - Malassezia sp.
Đzolat 41 + + + + M. furfur
Đzolat 42 + + + - M. slooffiae
Đzolat 43 - - - - M. globosa
Đzolat 44 + + + - M. slooffiae
Đzolat 45 ØØ ØØ ØØ ØØ Malassezia sp.
Đzolat 46 ØØ ØØ ØØ ØØ Malassezia sp.
Đzolat 47 - + + + M. sympodialis
Đzolat 48 ØØ ØØ ØØ ØØ Malassezia sp.
Đzolat 49 ØØ ØØ ØØ ØØ Malassezia sp.
Đzolat 50 + + + + M. furfur
Đzolat 51 + + + + M. furfur Đzolat 52 + + + + M. furfur Đzolat 53 + + + - M. slooffiae Đzolat 54 + + + + M. furfur Đzolat 55 - + + - Malassezia sp. Đzolat 56 Ø Ø Ø Ø Ø Đzolat 57 Ø Ø Ø Ø Ø
+: presipitasyon olanlar, -: presipitasyonu olmayanlar, Ø: üreme olmadı, ØØ:
üredi ancak çoğaltılamadı.

65
Tablo 4.6. Pityriasis versicolor’da klinik ve laboratuar bulguların pigmentasyon durumuna göre
dağılımı
Lezyon özelliği
Hiperpigmente
n(%)
Hipopigmente
n(%)
p
Cinsiyet 0,277
Kadın 19(76,0) 6(24,0)
Erkek 20(62,5) 12(37,5)
Güneş 0.096
güneş gören 19(59,4) 13(40,6)
güneş görmeyen 20(80,0) 5(20,0)
Primer/rekürren 0.011
Primer 14(51,9) 13(48,1)
Rekürren 25(83,3) 5(16,7)
Kepek var/yok 0.328
Kepek var 37(67,3) 18(32,7)
Kepek yok 2(5,1) 0(0,0)
Wood ışığı 0.952
Wood + 22(68,8) 10(31,3)
Wood - 17(68,0) 8(32,0)
Direk mikroskopi 0.366
+ 34(70,8) 14(29,2)
- 5(55,6) 4(44,4)
Bistüri örneği 0.902
üredi 33(68,8) 15(31,3) üremedi 6(66,7) 3(33,3) Bant örneği 0.579
üredi 16(72,7) 6(27,3) üremedi 23(65,7) 12(34,3) ModifiyeDixon
besiyeri
0.906
üredi 32(68,1) 15(31,9) üremedi 7(70) 3(30) Leemig-Notman
besiyeri 0.633

66
üredi 28(66,7) 14(33,3) üremedi 11(73,3) 4(26,7) Tür 0.4
ID yapılamadı 12(75,0) 14(25,0) M. slooffiae 7(77,8) 2(22,2) M. furfur 15(37,7) 11(42,3) M. globosa 1(50,0) 1(50,0) M. sympodialis 4(100) 0(0,0)
ID: Đdentifikasyon
Şekil 4.4. Şekil 4.5.
Şekil 4.6. Şekil 4.7.

67
Şekil 4. 8. Şekil 4.9.
Şekil 4. 10.
Şekil 4.11. Şekil 4.12.

68
Şekil 4.13. Tween 60- eskülin besiyeri Şekil 4.14.C-EL besiyerinde üreme
farklı renkteki zonlar
Şekil 4.15. Gram boyamadaki tomurcuklanan Şekil 4.16. Tween identifikasyon testi
Blastosporlar (maya hücreleri) Tween 20, 40, 60 ve 80 M. furfur
Şekil 4.17. Üre testi

69
Şekil 4.18. %10’luk KOH ile doğrudan mikroskop incelemesi (köfte-makarna görüntüsü)

70
5. TARTI ŞMA
Pityriasis versicolor (PV) derinin normal florasında bulunan ve dimorfik bir
maya mantarı olan Malassezia spp.’nin stratum corneum tabakasına kolonize olarak
sebep olduğu kronik ve yüzeysel bir deri hastalığıdır1. Robert Willian 19. yüzyılın
başlarında bu infeksiyona PV ismini vermiştir 229. David Gruby’nin 1846’da tinea
capitis ve favus’un etiyolojisi üzerindeki çalışmasından sonra, Eichstedt deri
lezyonlarında mantar elemanlarının varlığını bildirmiştir. Bu maya mantarı deri
pigmentinin değişmesine sebep olarak hipo ve/veya hiperpigmentasyonlu lezyonların
ortaya çıkmasına yol açar. PV genelde kozmetik bir infeksiyondur. Hastalığın en
belirgin semptomlarından biriside derinin kepek kepek dökülmesidir229. Lezyonlar
yumurtamsı veya yuvarlak şekilde olabilir. Bazı olgularda kaşıntı oluşabilir230.
Bu hastalık dünyanın hemen her bölgesinde görülmesine karşılık, tropikal iklimlerdeki
insidansı %40-60 gibi yüksek oranlardadır. Sıcak ve nemli havanın bu infeksiyonun
ortaya çıkmasında etkili olduğu anlaşılmaktadır228. Hastalık her yaşta görülmekle
birlikte özellikle genç erişkinleri etkiler. Çünkü yağ bezleri en çok gençlerde aktiftir.
Çocuklarda ve yaşlılarda nadiren görülür. Bu hastalık normal florada kolonize olan
Malassezia maya mantarının maya formundan patojen hif şekline dönüşmesi ve stratum
corneum tabakasına invazyonu sonucunda oluşur. PV daha çok yaz aylarında ve
tropikal bölgelerde meydana gelir. Bu hastalığın gelişmesinde endojen faktörlerin
önemi büyüktür. Bu faktörler; malnütrüasyon, doğum kontrol haplarının ve sistemik
kortikosteroidlerin alınması, immünsüpresyon ve aşırı terlemedir230.
Pityriasis versicolor’un tanısı kolaydır ve genellikle klinik bulgular yeterlidir.
Đnfeksiyon yağdan zengin bölgelerinde daha çok gövde ve omuzlarda görülür. Ayrıca,
yüz, ense, boyun, sırt veya vücüdun diğer bölgelerinde de görülebilir. PV’nun tanısı
doğrudan mikroskopi veya Wood ışığı (filtre edilmiş 365 nm dalga boyundaki UV ışını)
ile kolayca doğrulanır. Lezyonlu bölgenin epitelyum kazıntısı etken mikro-
organizmanın tanısı yararlanılır. Mikroorganizmayı elde etmek için örnekler lezyonlu
bölgeden kazıma yoluyla alınabilir. Klinik örnekteki keratin ve deri kalıntıları %10-
15’lik KOH ile kolaylıkla çözülür. Mantar elemanları, spor ve hif, açığa çıkar.

71
Lezyonlardaki spor miktarı normal floradan daha fazladır. Bu tipik bulguya “köfte-
makarna” görünümü denir. Buna ek olarak PV’ nin Wood ışığı muayenesinde
lezyonların olduğu bölge sarı-yeşil renkte flöresans verir. Bununla beraber, Wood ışığı
pozitif olan olguların sadece üçte birinde görülür. Son yıllardaki çalışmalarda sadece
Malassezia furfur’ un indol bileşenleri ürettiği belirtmiş ve yalnızca bu türün Wood
ışığında altında flöresans oluşturduğunu göstermiştir230.
Genelde PV lezyonlarından izole edilen en yaygın Malassezia türü M. globosa
ve M. sympodialis’dir. M. slooffiae ve M. furfur gibi türlere daha az rastlanır. Bu
çalışmalardaki farklı sonuçların farklı coğrafik bölgelerin sonuçlarının
karşılaştırılmasından da kaynaklanabileceği vurgulanmıştır230.
Son yıllarda, bu mantarın Seboreik dermatit, atopik dermatit, follikülit ve
parenteral beslenenlen yeni doğanlarda görülen sistemik hastalıklarla ilişkili olabileceği
bildirilmi ştir Malassezia pachydermatis dışındaki türler lipofiliktir. Bu maya mantarı
lipit bağımlı olduğu için in vitro uzun zincirli yağ asitlerinin olması gerekmektedir.
modifiye-Dixon ve Leeming-Notman agarın (LNA) kullanımı gelişimi Malassezia
spp.’nin izolasyonu büyük ölçüde gelişmiştir. Malassezia cinsinden yumurtamsı,
elipsoidal ve silindirik hücrelere sahiptir. Malassezia genusunudan M. furfur’un
pleomorfik hücreleri vardır, büyüklüğü değişkendir, silindirik, yumurtamsı maya
hücreleri geliştirir. Bu tür sağlıklı insan ve hayvan derisinde görülebilir. M. globosa
tipik küremsi hücre morfolojisi ile tanınır. Maya hücreleri dar bir noktadan
tomurcuklanır. M. sympodialis insan derisinde en yaygın olan maya mantarıdır. M.
sympodialis kültür ve biyokimyasal özelliği ile tanınır ve esas olarak güçlü β-
glukozidaz aktivitesi vardır ve eskülin hidrolizi hızlıdır. M. slooffiae Midgley tarafından
Pityrosporum ovale olarak tanımlanmıştır. M. restricta fizyolojik ve biyokimyasal
aktivitesi sınırlı bir türdür. Kültürde zor ürer ve zor saklanır. Katalaz aktivitesi olmayan
tek türdür. Saçlı deri ve yüz bölgesinden izole edilir. Oval ve silindirik hücreden oluşur.
M. obtusa çok nadir görülür. Morfolojisi M. furfur’la benzerdir, ancak fiziksel olarak M.
restricta’ya yakındır231.
Pityrisis versicolor’un tanısı kolaydır, klinik ve mikroskobik muayene ile hemen
tanınır.

72
Erchiga ve ark. PV’lu 96 hastanın %10’luk KOH incelemesinde, tüm örneklerin
doğrudan mikroskobisinde köfte-makarna (blastospor ve hif) görüntüsünü bulmuştur229.
Gupta ve ark., Kanada’da PV’li 523 hastanın 386’sının (%68.1)
mikroskopisinin pozitif olduğunu bulmuşlardır232.Aspiroz ve ark. Đspanya’da yaptıkları
bir çalışmada PV’lu 79 hastanın örneklerinin KOH incelemesinde hepsinde tipik köfte-
makarna görünümüne rastlamışlardır.231.Jena ve ark.’nın çocuklarda yaptığı çalışmada
olguların %72.3’ünde köfte-makarna görüntüsünün olduğunu belirtmiştir235. Payle ve
ark. Pityriasis versicolor’lu 20 hastada KOH solüsyonu ile Albert solüsyonunu
karşılaştırmışlardır. KOH ile görüntülenen 20 klinik izolatın hepsinde tipik maya
hücresi ve hifleri görmüşlerdir. Tarazooie ve ark. PV’ li 94 hastanın % 89.4’ünde spor
ve hife rastlamışlardır237. S. Maheswari Amma PV’lu tanılı 100 hasta ve 100 kontrol
olgusunda, lezyonlu bölgeden aldıkları %100’ünün KOH ile doğrudan mikroskop
incelemesinde blastospor ve hifleri görmüştür238.
Prohic ve ark., 90 PV’lu olguyu incelemiş; lezyonlu bölgelerden alınan örnekler
%30’luk KOH incelemişler ve örneklerin 88’inde (%98) tipik spor ve hifleri
görmüşlerdir. Đki olguda hem spor hem de hifleri görmemişlerdir39.
McGincley ve ark,. 30 PV’lu olguyu çalışmış lezyonlardan alınan örneğin
doğrudan mikroskop incelemesinde 23 olguda sadece maya hücresini, 18 olguda hif
elemenlarını görmüşlerdir202.
Çalışmada, PV tanılı 57 hasta incelendi ve 52’sinde üreme gözlendi. Örneklerin
48’ ünde (%84.2) % 10’luk KOH ile doğrudan mikroskop incelemesinde tipik köfte-
makarna (spor ve hif) görüldü. 9’unda (%15.8) ise belirlenemedi. Doğrudan
mikroskopisi negatif olan 8 örneğin 3’ünde (%37.5) hiç üreme olmadı. Doğrudan
mikroskop sonuçları ile lezyon özelliği arasındaki ilişki incelendiğinde istatistiksel
olarak anlamlı olmadığı görüldü (p=0.366). Çalışmada PV’ nun tanısı direkt bakı ile
kolayca konuldu. Ancak, klinik bulguların olmasına karşın, doğrudan mikroskop
incelemesinin olumsuz olmasının birkaç sebepten dolayı olabileceği düşünüldü. Bunlar
arasında, duş alma, terleme, lezyonlu bölgeye krem benzeri ürünlerin sürülmesi veya
herhangi bir antifungal ilacın kullanılmasının kepeklenmeyi azaltabileceğine ve
kepeklerdeki mantar elemanların görünmemesine neden olabileceği düşünüldü.

73
Mantar kültürü, Malassezia türlerinin morfolojik, fizyolojik ve biyokimyasal
özelliklerine göre ayrılması için gereklidir237. Bu nedenle, Malassezia identifikasyon
testlerinin yapılabilmesi için ilk önce kültürde üretilmesi gereklidir.
Erchiga ve ark, PV tanılı 96 hasta çalışmış 93’ünün (%97) kültür pozitif olarak
bulmuştur229. Aspiroz ve ark. PV’li 79 hastanın %99’unda kültür pozitifliğini
bildirmişlerdir231. Gupta ve ark., PV kuşkulu 523 hastanın 129‘unda kültürde mantar
izole etmiştir. Bu örneklerin 111’ini saf kültür olarak üretmiş, kültür sonucuna göre
bunların 84’ünü PV olarak tanımlamıştır, 27’sinde kuşkulu PV olarak tanımlanmıştır.
Çünkü, bu olguların mikolojik incelemesinde hif saptanmış ve kültürlerinde üreme
olmamıştır.232.
Amma SM’nin 100 PV’lu hastada yapmış olduğu bir çalışmada lezyonlu
bölgeden alınan 100 örneğin %70’inde kültür pozitifliğini bulmuştur. Klinik örneklerin
% 30’unda ise üreme olmamıştır. Buna gerekçe olarakta besiyerinde kontaminan olan
mikroorganizmaların Malassezia maya mantarlarının üremesini engellemesi ve
besiyerindeki yetersiz yağ miktarının olduğunu vurgulamıştır. Daha iyi besiyeri ve
üretme yöntemi kullanarak mantarın izolasyon oranının artacağını belirtmiştir238.
Krisanty ve ark., PV’lu 98 hastanın 91’ini (%92.8) kültürde izole etmiş ve
identifiye etmişlerdir. Diğer 7 (%7.1) örnek biyokimyasal ve morfolojik olarak
uyuşmazlık göstermiş ve identifiye edilememiştir. Gupta ve ark., Kanada’da PV’ lu 245
hastanın %70’inin kültürünü pozitif bulmuşlardır. Neves ve ark., Brezilya‘da PV’lu 15
klinik örnek toplamışlardır. Ve 5’inin (%33.33) kültürünü pozitif bularak identifiye
testlerini yapmışlardır. Prohic ve ark., PV’lu 90 olguyu çalışmış ve hepsini kültürde
üretmiştir239.
Çalışmada PV’lu 57 olgu incelendi. Bu olguların 6’sında (%10.5) her iki
besiyerinde (MD ve LN) hiç üreme olmadı, 7’sinde (%12.2) bu iki besiyerinin birinde
üreme oldu, ancak koloninin pasajı yapıldığında çoğalmadı. Diğer 44 (%77.1) örnek ise
kültürde üredi. Diğer çalışmalarda lezyonlardan alınan örneklerin çoğu kültürde üreme
gösterdi. Ancak, çalışmamızda hiç üremeyen örnekler olduğu gibi üreyip
çoğaltılamayan klinik örnekler de vardır.

74
Çalışmada lezyonlardan alınan örneklerin hepsinde üreme olmamasının sebebi
mevsimsel sebeplerden dolayı olabilir. Klinik örneklerin yapıldığı mevsim sonbahar ve
kış mevsimi yani infeksiyonun prevalansının en az olduğu mevsimde üreme oranının az
olabileceği düşünülmüştür.
PV ile yapılan çalışmalarda değişik örnek alma yöntemleri karşılaştırılmıştır.
1ml TritonX-100203 ile emdirilmiş pamuk çubukları; Nakabayashi ve arkbez, bant ve
saç fırçasını; Gupta ve ark., Rodac plaklarını kullamışlardır240,241.
Gupta ve ark.,Kanada’da PV etkeni olarak tropikal bölgelerde M. globosa’nın (%62)
baskın olmasına rağmen, ılıman iklimlerde M. sympodialis’i (%59) baskın tür olarak
rapor etmişlerdir232.
Buna karşılık, Prohic ve ark., PV’lu 79 hastanın % 100’ünde etkeni üretmiş ve en çok
M. globosa’yı (%63), sonra sırası ile, M. sympodialis’i (%14), M. obtusa (%8) ve M.
slooffiae (%4) izole etmişlerdir. M. restricta ve M. pachydermatis ise izole
edilmemiştir. M. sympodialis şu ana kadar en yaygın tür olarak bildirilmesine karşılık
bu araştırıcılar 33 (%37) olguda bulmuşlardır. Yine M. furfur’u beş (%5) olguda izole
etmişlerdir. Araştırıcılar M. sympodialis’i infekte deride olduğu kadar sağlıklı deride
bulmuş ve ikinci baskın tür olarak bulmuşlardır. Bu tür diğer PV çalışmalarında da en
yaygın ikinci tür olarak bulunmuştur. Bu türün patojen olarak rolü hala
açıklanamamıştır239. Yazarların bulduğu bu sonuçlar, Salah ve ark., Nakabayashi ve
ark., nın PV lezyonlarında, sırasıyla ile, %65 ve %55 oranında M. globosa’yı izole
etmeleri ile uyumludur 241. M. globosa’nın yüksek patojenitisi, PV lezyonlarındaki
yüksek oranda lipolitik aktivitesiyle ilgili. Bununla beraber M. globosa sağlıklı deride
baskın türdür (% 49)239.
M. furfur’un özellikle tropikal iklimlerde deri hastalıklarından sorumlu olarak
doğrulanmıştır. Bu tür, Gupta ve ark., ile Prohic ve ark. yaptıkları çalışmalar %10
oranında izole edilmiştir..Ancak Tunus’ta yapılan bir çalışmada M. furfur %25 oranında
bulunmuş ve ikinci baskın tür olduğu saptanmıştır242. Son yıllarda yapılan çalışmalarda
M. obtusa ve M. slooffiae insan derisinde yaygın olarak bulunmamış ancak PV ve
sağlıklı bireylerde ki olgulardan izole edilmiştir. Buna karşın Gupta ve ark.232, Crespo
ve ark229. bu türlere rastlamamıştır. M. restricta, seboreik dermatitli hastaların ve

75
normal bireylerin yüz ve saçlı deri kafa bölgelerinden izole edilmiştir. Ancak, gövde de
nadir görülür. M. pachydermatis prematürlerde septik epidemilere sebep olabilir, ancak
bu tür hayvanlarda daha çok görülür. Đnsanlarda bu türün varlığı nadir ve geçicidir.
Crespo ve ark., PV’lu hastalarda olguların %97’sinde M. globosa’yı izole
etmişlerdir. Yazarlar M. sympodialis %32 ve M. globosa %28 oranında bulmuştur. 7
olguda M. slooffiae izole etmişlerdir. Bu sonuçlara göre M. globosa baskın bir rol
oynar229.
Aspiroz ve ark. PV lezyonlarında M. globosa’yı (%90) izole etmişlerdir.
Lezyonların KOH incelemesinde maya hücreleri ve kısa hifleri görmüşlerdir. Đkinci
baskın tür olarak M. sympodialis (%40.5), bir olguda ise M. slooffiae’yi bulmuşlardır.
M. restricta’yı ayrıca M. obtusa’yı hiç izole etmemişlerdir. Đlginç olarak patojen tür
olarak bilinen M. furfur’u, PV lezyonlarından hiç izole etmemişlerdir. Bununla beraber
M. furfur lipit bağımlı mikroorganizmadır. Hızlı ürer, büyük kolonileri vardır, saklama
koşullarına dayanıklıdır. 37°C’de iyi ürer; 32°C ve 41°C’lerde üreyebilir. Yazarların
bulduğu bu sonuçlar Crespo ve ark.’ nın yaptığı çalışma ile uyumludur231.
Tarazooie ve ark. PV lezyonlarında en sık M. globosa’yı (% 53.3) sonra sırası ile
M. furfur ( % 25.3), M. sympodialis (% 9.3), M. obtusa (%8.1) ve M. slooffiae’yi (%4.0)
bulmuşlardır. Araştırıcılar PV lezyonlarında M. globosa’ yı baskın tür olarak
bulmuşlardır. Bu sonuçlar dünyanın farklı bölgelerinde yapılan başka çalışmaların
bulguları ile uyumludur tartış.19, 31, 32, 38. Makimura ve ark. PV lezyonlarında M. furfur ve
M.sympodialis’i baskın tür olarak bulmuştur240. Gupta’nın203 ılıman iklimlerde M.
sympodialis’i baskın tür bulmasına karşılık, tropikal iklimlerde M. globosa ilk sıradadır.
Dahası, M. furfur yeni çalışmalarda PV lezyonlarından izole edilen ikinci türdür ki bu
sonuç Dutta’ nın çalışmasıyla benzerdir.Salah ve ark., PV lezyonlarında M. globosa’nın
prevalansını yüksek oranda (%55) bulmuşlardır ve bu sonuç birçok çalışmayla
benzedir242.
Lezyonlu deride M. globosa’nın yüksek prevalansı yüksek lipolitik aktivitesiyle
ve bu türün aşırı üremesiyle açıklanabilir. M. furfur suşları Kanada ve Japonya’da
yapılan çalışmalarda daha çok izole edilmiştir. Kanada ve Japonya’da M. furfur %5-
10.8 oranında rapor edilmiştir. Bununla beraber, M. sympodialis diğer çalışmalardan

76
daha az sıklıkla izole edilmiştir. M. restricta zayıf üremesinden dolayı PV
lezyonlarından seyrek olarak izole edilmiştir.
Gaitanis ve Velegraki’nin yaptığı bir çalışmada, PV lezyonlarından M.
globosa’yı (%90) en yaygın olan tür olarak bulmuşlardır. Olguların %77’sine M.
globosa tek olarak izole edilmiştir. M. sympodialis, M. furfur ve M. slooffiae türlerini de
diğerlerine (%13) eşlik etmişlerdir243.
Guillot ve ark., PV’ lu 22 hastanın 12’sinde (%55) M. globosa, 2’sinde M.
sympodialis ve bir olguda M. furfur, M. slooffiae, M. pachydermatis’i izole etmişlerdir.
M. obtusa ve M. restricta’ yı hiç izole etmemişledir129. Gueho ve ark.. PV lezyonlarında
M globosa’yı sık rastladıklarını bildirmişlerdir101. Hindistan’da yapılan çalışmada PV
lezyonlarından M. globosa’ yı baskın tür olarak bulunmuştur 234.
Çalışmada 57 tane PV olgulu hastadan örnek alındı. Üremesi gerçekleşen ve
çalışılan 45 tane örneğin 26’sında (%45.6) M. furfur, 9’unda (%15.8) M. slooffiae,
4’ünde (%7.0) M. sympodialis, 2’sinde (%2) M. globosa izole edildi. Üç izolat ise
Tween testleri ve diğer testlerin sonununa göre identifikasyonuna göre tanımlanamadı.
Çalışmamızda elde edilen sonuçlar, diğer çalışmalarla kıyaslandığında tür
identifikasyonu yönünden farklı bulundu. Birçok çalışmada, M. globosa ve M.
sympodialis baskın tür olarak bulunmuşken, çalışmamızda M. furfur baskın tür olarak
saptandı. Yine birçok çalışmada M. globosa baskın tür olarak görülürken, çalışmamızda
sadece iki örnekte izole edilmiştir. M. slooffiae son yıllarda yapılan çalışmalarda seyrek
olarak görülürken, çalışmada 4 olguda etken olduğu görülmüştür. Çalışmamızda ayrıca,
M. obtusa, M. restricta, M. japonica, M. dermatis ve M. yamatoensis izole edilmemiştir.
Çalışmada elde edilen sonuçların literatür verilerinden farklı olmasına neden olarak,
klinik örneklerin değişik mevsimlerde alınması, çalışılan bölgenin subtropikal iklime
sahip olması, farklı örnek alma yöntemleri, farklı coğrafik özelliklere ve farklı etnik
kökene sahip olmasından dolayı olabileceği düşünülmüştür.
Çalışmada tür dağılımı ile lezyonun özelliği arasındaki ilişki de incelenmiştir.
Çalışmada en çok izole edilen tür olan M. furfur’un %57.7’si hiperpigmente, %42.3’ü
hipopigmente lezyonlarından izole edildiği görüldü. Đstatiksel olarak tür dağılımı ile

77
lezyonunu özelliği arasındaki ilişki incelendiğinde anlamlı bir fark bulunmamıştır
(p=4).
Çalışmada, klinik örneklerin alınmasında bistüri yöntemi (%84.2) selofan-bant
yöntemine göre daha başarılı bulunmuştur. Pityriasis versicolor deride kepek üreten bir
hastalıktır ve bu özelliği ile klinik tanısı konulur. Mikroorganizmayı lezyonlardan alınan
kepeklerle izole edilir. Bistüri ile lezyonlardan daha fazla kepek alınabilmiştir. Bu
nedenle, kepek fazla olduğundan daha iyi üreme elde edilmiştir (%84.2). Selofan-bant
yönteminde ise lezyonlardan alınan kepekler fazla olmadığından (%38.6.) ve alınan
kepekler selofan-bantta kaldığından iyi üreme olmadı (%61.4). Sonuç olarak, PV’de
klinik örnek alma yöntemi olarak bistüri yönteminin daha iyi olduğu bulunmuştur.
Litertüre bakıldığında örnek alma yöntemlerinin karşılaştırılmasına ilişkin çalışmaya
rastlanılmamaktadır.
Çalışmada, klinik örneklerin alınmasında selofan bantla örnek alma yönteminin
sonuçları Crespo ve ark. ve Sidharta ve ark yapmış olduğu çalışma ile uyumlu değildir.
Yazarlar, selofan bantla alınan örneklerin çoğunda maya kolonisi izole etmişlerdir229
Çalışmada incelemeye alınan diğer bir konuda besiyerlerinin performansların
karşılaştırılmasıdır. Bu amaçla, modifiye-Dixon ve Leeming-Notman besiyerlerinin
mikroorganizmayı üretme derecesine bakıldı ve modifiye- Dixon besiyerinin daha iyi
olduğu gözlenmiştir (%82.5). Bu sonuç Mayser ve ark. yaptığı sonuçla pek
uyuşmamaktadır 244.Yazarlar bu iki besiyerinin Malassezia’yı üretme bakımından
benzer özellikte olduğunu vurgulamışlardır. Literatürde besiyeri performanslarının
karşılaştırılması ile ilgili bir çalışmaya pek rastlanılmamaktadır.
Lezyonlardan izole edilen maya kolonisinin miktarı bazı faktörlere bağlıdır.
Hastadan örnek almadan önce; terleme, duş alma, krem kullanma veya hastanın son 15
gün içerisinde herhangi bir antifungal kullanması hem kepeklenmeyi azaltır hem de
maya izolasyonunu etkilemiş olabileceği düşünüldü.
Çalışmada diğer önemli birkaç sonuç ortaya çıkmıştır. Bazı örneklerde
lezyonlardan alınan ilk örneklerin pasajlandıktan sonra koloniler çoğalmamıştır. Fakat

78
daha sonra koloniler sıvı modifiye-Dixon ve Leeming-Notman besiyerinde 10 günlük
inkübasyondan sonra üretildiği görüldü. Aynı şekilde kuruyan kolonilerde sıvı
besiyerlerinde aynı süre zarfında inkübe edildiğinde yeniden canlandığı belirlendi.
Gupta ve ark. yaptığı çalışmada PV’un yaşa göre dağılımını incelemişlerdir.
Yazarlar hastaların yaşa göre dağılımına bakıldığında 60’dan büyük (%93.3) yaştaki
bireylerde bu hastalığın fazla olduğu görülmüştür. En düşük oran 0-14 yaş arası (%36.6)
bireylerde görmüştür102.
Karakaş ve ark., PV’un daha çok 16-30 yaş arasında arası görüldüğünü
vurgulamışlardır243. Salahi-Moghaddam ve ark. hapishanedeki bireylerde yaptığı PV
çalışmasında, hastalığın en fazla 28-32 yaş arasında olduğunu belirtmiştir251. He ve ark.,
yaptığı epidemiyolojik çalışmada PV’ nin hem erkek hem de bayanlarda 20-29 yaş
arasında daha fazla olduğunu belirtmiştir252. Silvia ve ark. 8-10 aylık bebeklerde
Malassezia maya mantarının oranını % 23.3, 11-15 yaş arası çocuklarda ise % 26.7
olarak bulmuştur. Bu sonuç yukarıda belirtilen Gupta ve ark. yaptığı sonuç ile
uyumludur102..Prohic ve ark. PV’yi yaş grubuna göre incelemişler ve hastaların
çoğunluğunun 16-30 yaş arasında olduğunu belirtmişlerdir239. Faergemann ve
Fredrickson beş yaştan daha küçük çocuklarda Malassezia maya mantarlarının
kolonisini bulmamışlardır 246. Hindistan’da yapılan bir çalışmada 11-30 yaş arası
bireylerde Malassezia’nın kolonizasyonun en yüksek oranda bulunmuştur. Ben Salah
ve ark. PV’nun daha çok 15-30 yaş arasını (%65) etkilediğini vurgulamışlardır. PV
sıklığını 0-5 yaş grubunda çocuklarda % 0,5-10 yaş arası çocuklarda ise % 2, 45 yaş
üstü bireylerde ise %11 olarak bulmuştur242. Kyriakis ve ark., 16-25 yaş arası
bireylerde PV’ nu yaygın olduğunu, 70 yaş üstü bireylerde ve 6-10 yaş arası çocuklarda
daha az olduğunu, 0-5 yaş arası çocuklarda ise hiç PV’un görülmediğini rapor
etmişlerdir. Jena ve ark. PV’u çocukluk yaş gruplarında (2-14 yaş) incelemiş; hastaların
çoğunluğunun 8-12 yaş arasında olduğunu, ayrıca 10 bebekte infeksiyon görüldüğünü
belirtmişlerdir. Yazarlar sonuçta PV’un çocuklarda yaygın olmadığını rapor etmişlerdir.
Silvia ve ark. çocuk yaş grubunda infeksiyonun en çok 11-15 yaş arasında olduğunu
belirtmiştir. Di silverio ve ark. PV’un infantlarda da görüldüğünü belirtmiştir.
Yaşa göre farklı gruplandırmalardan dolayı çeşitli çalışmalardaki
karşılaştırmalarda farklıdır. Bergbrant ve Broberg, Malassezia miktarını çocuklarda

79
oldukça yüksek bulmuştur. Bu durum, çocukların derilerindeki sebum miktarının çok
az olduğuna işaraet eder. Derideki lipit miktarı ve bileşimi puberte döneminde
değişiklik gösterir.16 Puberte döneminde artan lipit miktarı, derideki Malassezia maya
mantarının kolonizasyonunu artırabilir.4 Çocuklarda 14 yaşına kadar Malassezia
kolonizasyonu erişkinlere göre daha azdır. Önceki çalışmalar yaş artışı ile deride
Malassezia kolonizasyonunu azaldığını berlirtmiştir. Koloni miktarının 30’lu yaşlarda
40, 50, 60 ve 70’li yaşlardan daha yüksek olduğu belirtilmiştir. Bunun da sebebi olarak
artan yaşla beraber derideki sebum miktarı azalmaktadır.95 Bununla beraeber hastanede
yatan yaşlılarda PV’ nun gelişmesi de yüksektir.95 Bu durum bağışıklık sisteminin
baskılanması ile açıklanmaktadır. Bu hastalar kıyafetlerini değiştirmezlerse daha çok
terleyebilirler ve vücutları daha çok nemli kalır. Bu durum, PV’ye neden olabilir.
Çalışmamızda, 57 PV’un en çok 16-30 yaş grubunda sonra 31-45 yaş ve 1-15
yaş grubunda saptandı. Çalışmanın yaş dağılımına göre sonucu diğer çalışmalarla
uyumludur. Çalışmamızda ilginç olarak üç aylık erkek bebekten selofan-bantla alınan
örnekte ve 72 yaşındaki erkek hastadan alınan bistüri ile alınan örnekte Malassezia cinsi
maya mantarları iyi bir şekilde üretilmiştir. Bu sonuç diğer çalışmalardan farklıdır. Bu
durum, iki hastanın derideki sebum miktarının fazla olmasından veya genetik
faktörlerden dolayı olabilir.
Çalışmada incelemeye alınan diğer bir konu ise PV ile cinsiyet arasındaki
ili şkidir. Bu konu ile ilgili çok çalışma vardır. PV’un gelişiminde cinsiyetin rolü hala
anlaşılmamaktadır. Madras’ta PV’nin erkek bayan oranı 3.3:1 olarak bulunmuştur.20
Bazı çalışmalarda bu oranın bayanlarda yüksek olduğu vurgulanmıştır6,22-24. Gupta ve
ark. PV’ nin bayan ve erkek arasındaki ilişkide farklılık olmadığını, her iki cinsiyette de
aynı oranda görüldüğünü bildirmiştir.
Aspiroz ve ark., yaptığı PV çalışmasında bayan oranını daha yüksek olarak
bulmuştur251.
Ingordo ve ark., Khosravi ve ark., Endonezya’da, Çin’de, Kyriakis ve ark.
PV’un prevalansının erkeklerde daha fazla görüldüğünü rapor etmiştir.
Tarazooi ve ark.‘nın yaptığı çalışmada PV’nin gelişiminde cinsiyetin önemli
olmadığını vurgulamıştır237.

80
Çalışmada PV prevalansının erkeklerde (%56.1) bayanlara (%43.9) göre olduğu
gözlendi. Bu durum literatürle uyuşmaktadır. Erkekelerde PV prevalansının yüksekliği,
yağ aktivitesinin bayanlardan daha yüksek olması, hormonların aktvitesi, yağ miktarı
daha fazla olduğu için daha çok terleme ve androjenler bu infeksiyonun oluşumundan
sorumlu tutulmuştur252. Ayrıca, bayanlar daha çok hijyene ve güzelliğe dikkat ettikleri
için bayanlarda daha az görülebilir. Cinsiyetin insidansını açıklamak zordur. Çünkü
hastaların çoğu deri hijyenini ve güzelliğini sağlamak için çeşitli kozmetikler
kullanmaktadır.
Çalışmada cinsiyetle lezyon özelliği arasındaki ilişki de araştırılmıştır. Buna
göre erkeklerin % 62.2’si hiperpigmente, %24.0’ı hipopigmente bulunmuştur.
Bayanların % 76’sı hiperpigmente, %24’ü hipopigmente bulunmuştur. Đstatistiksel
açıdan cinsiyetle lezyonun özelliği arasında bir ilişki bulunmamıştır (p=0.277).
Bir çalışmada PV’un diabetle ilişkisi araştırılmıştır. Keddie, PV’lu hastalarda
epidermis tabakasında ekstrasellüler glikojen depoları bulmuştur. Bu infeksiyona
duyarlılığın derinin glikojen metabolizması ile ilgili olabileceğini ileri sürülmüştür.
Keddie PV’un, diabetle ilişkisinin düşük oranda olduğunu bulmuştur. Sonuç olarak,
diabetin bu hastalığın patogenezinde önemli bir faktör olmadığını belirtmiştir246.
Çalışmamız yalnızca bir hastada diabet öyküsü vardı. Bu sonuç, Keddie ve ark.’nın
yaptığı çalışma ile uyumludur.
Pityriasis versicolor’un hamile kadınlarla ilişkisine bakıldığında literatürde fazla
çalışma yoktur. Zampino ve ark. Đtalya’da hamile bayanlarda PV’un prevalansını
araştırmış ve hamile bayanları üç trimester olarak incelemiştir. Sıcak iklime bağlı olarak
hamileliğin birinci ve üçüncü trimesterinde Malassezia’nın kolonizasyonunu bulmuştur.
Sonuçta hamileliğin PV’nin prevalansını artırmadığını rapor etmişlerdir. Bununla
beraber, maya mantarının kolonizasyonunun hamileliğin sonunda artacağını
belirtmişlerdir247. Hamilelik boyunca derideki değişmeler, artan yağ bezlerinin aktivitesi
Malassezia infeksiyonunun oluşmasına sebep olabilir. Hamile bayanlardaki yağ
bezlerini aktivitesi sınırlıdır. Henderson ve ark. doğumdan birkaç hafta önce ve sonra
yağ bezlerinin oranını ölçmüşlerdir. Hamileliğin sonuçlandıktan sonra 10 anneden
üçünde yüksek oranda olduğunu belirtmişlerdir.8 Buna ek olarak yağ bezinin
aktivitesindeki artış, tiroid fonksiyonunu artırabilir ki bu durum PV’ye sebep olabilir.4

81
Çalışmamızda bir tane hamile PV’lu hasta vardı ve hamileliğin ilk trimesterinde
idi. Bu durum yapılan çalışmayla uyumludur. PV’un gebelikle ilişkisi yoktur 247.
Çalışmaya dahil edilen hastaların 55’inin (%96.5) kepek vardı; 2’si ise (%3.5)
kepek yoktu. Bu durumun, Malassezia maya mantarının izolasyonu ile uyumlu olduğu
görülmüştür. Kepek ne kadar çok olursa besiyerindeki koloni sayısının o kadar fazla
olduğu görülmüştür. Çalışmada ayrıca, kepek ile lezyonun özelliği arasındaki ilişki de
incelendi ve istatistiksel olarak kepek ile lezyon arasında anlamlı bir ilişki olmadığı
belirlendi (p=0.328).
Çalışmaya dahil edilen hastaların 39’u (%68.4) hiperpigmente, 18’i (%31.6)
hipopigmente idi. PV’deki hiperpigmentasyonun mekanizmasına dair iki teori vardır.
Birincisi hiperpigmente bireylerde keratin tabakasının kalınlığının artması; ikincisi bu
bireylerde hücre inflamasyon olması ve bu durumun melanositleri stimüle etmesi ve
sonuçta daha çok pigment üretiminin olmasıdır.
Prohic ve ark., PV olgularının %53’ünde lezyonun hiper-pigmente, %40’ında
hipo-pigmente, %7’sinde hem hipo hem de hiper-pigmente olduğunu vurgulamışlardır.
Çalışmamızda, mantar türü lezyonunun özelliğinin istatistiksel olarak anlamlı
olmadığı görüldü (p=4) ve spesifik bir bulgu bulunmadı.
Çalışmada, PV’un anatomik bölgelere göre dağılımı incelendiğinde; en çok
boyun, sırt, göğüs gibi üst bölgede görüldüğü gözlenmiştir. PV güneş gören bölgelerde
%56.1, güneş görmeyen bölgelerde ise % 43.9’oranında bulundu. Bu bölgelerdeki yağ
bezlerinin aktivitesinin fazla olması yanında bu bölgelerde aşırı terlemeye bağlı olarak
lezyonların görülebildiği düşünüldü.
Çalışmada ayrıca, güneş gören bölgelerle lezyonun özelliği de araştırıldı ve
istatistiksel olarak güneş gören bölge ile lezyonun özelliği arasında anlamlı bir ilişki
bulundu (p=0.096).
Literatürde, yalnızca M. furfur’un flöresans oluşturduğu bildirilmektedir.
Çalışmamızda, Wood ışığı ile tür dağılımının ile ilişkisi incelendi ve Wood ışığının tür
düzeyinde tanıya yardımcı olmadığı görüldü. Ayrıca Wood ışığı ile lezyonun özelliği
arasında ilişkiye bakıldığında wood ışığı negatif olanların %68’i hiperpigmente iken

82
%32’si hipopigmentedir. Wood ışığı pozitif olan %68.8’i hiperpigmente, %31.3’ü
hipopigmentedir. Đstatistiksel olarak wood ışığı ile lezyonun özelliği arasında anlamlı
bir ili şki bulunmadı (p=0.952).
PV’nun gelişmesinde ailesel faktörler de önemlidir. He ve ark., PV’lu
hastalarda yaptığı çalışmada ailenin önemli bir faktör olmadığını bulmuştur248.
Çalışmadan elde edilen sonuçlardan birisi de hiperpigmente bölgelerin; Wood ışığında
flöresans verdiği (%68.8), doğrudan mikroskop incelemesinde pozitiflik oranının daha
yüksek olduğu ve daha çok rekürren infeksiyona yol açtığı belirlenmiştir.
Sonuç olarak PV olgularında en sık izole edilen lipofilik maya mantarının M.
furfur olduğu, klinik örneklerin alınmasında bistüri yönteminin daha uygun olduğu;
izolasyonda modifiye-Dixon besiyerinin daha iyi üreme sağlandığı, infeksiyonun en çok
16-30 yaş grubunda görüldüğü, rekürren infeksiyonların daha çok hiperpigmentasyon
ile özellendiği saptanmıştır. Malassezia’ların tür tanısında klasik yöntemlerin
laboratuvar tanıda her zaman yeterli olmadığı, bu nedenle bazı türlerin tanınamadığı
görüldü. Bu nedenle, moleküler biyolojik temelli yöntemlerin kullanımına gerek olduğu
düşünüldü.

83
6. SONUÇ ve ÖNERĐLER
1. Bu çalışmada, Pityriasis versicolor (PV) tanılı 33’ü erkek, 24’ü kadın toplam 57
hasta çalışmaya dahil edildi.
2. Klinik örnek alma yöntemleri karşılaştırıldığında; bistüri yönteminin selofan-bant
yöntemine göre daha iyi olduğu görüldü. Selofan-bant yönteminin ise kullanışlı
olmadığı, kepek azlığında üremeyi etkilediği ve kepeklerin bantta kalarak besiyerine
inoküle edilemediği veya az sayıda koloni ürediği görüldü. Bistüri ile alınan klinik
örneklerde çok sayıda kepek alındığından mantar izolasyonunun daha iyi olduğu
görüldü. Çalışmada kepek ile mantarın izolasyonu arasında bir ilişki olduğu kepeklenme
ne kadar çoksa mantar üremesinin iyi olduğu görüldü
3. Çalışmada kullanılan besiyerlerinin performansları karşılaştırılıp mantarı izole etme,
çoğaltma ve saklama koşulları değerlendirildiğinde; modifiye-Dixon besiyerinin daha
üstün olduğu görüldü. Modifiye- Dixon besiyerinde maya kolonisinin Leeming-
Notmana göre daha çok, sayıda ve daha büyük olduğu görüldü. Leeming-Notman
besiyerinde üreyen kolonilerin az sayıda ve küçük olduğu belirlendi.
4. Çalışmada 57 klinik örneğin 7’sinde hiç üreme olmadı. 7’sinde modifiye-Dixon veya
Leeming-Notman besiyerinden yalnızca birinde üredi. Ancak, çoğaltılamadığı için tür
identifikasyonu yapılamadı.
5. Đdentifikasyonu yapılan örneklerin 27’si M. furfur, 10’u M. slooffiae, 4’ü M.
sympodialis ve 2’si M. globosa idi.
6. Wood ışığı ile Malassezia türleri arasında istatistiksel olarak anlamlı bir ili şkinin
bulunmadığı belirlendi.
7. Deride görülen pigmentasyon çeşidi (hipo-hiper-pigmente) ile mantar türü bir ilişki
arasında bir olmadığı görüldü.
8. Lezyonlarda çoklu mikro-organizma görülmedi. Lezyonlardan tek bir tür izole edildi,
mikst mikro-organimaya rastlanmadı.

84
9. Đnfeksiyon ile anatomik bölge arasındaki ilişki incelendiğinde; lezyonların sıklığa
göre boyun, sırt ve göğüs bölgesinde yer aldığı görüldü.
13. Hastalığın cinsiyete ve yaşa göre dağılımı incelendiğinde; infeksiyonun en çok
erkeklerde ve 16-30 yaş grubunda görüldüğü belirlendi.
14. Đnfeksiyonun daha çok rekürren özellkikte olduğu görüldü ve bazı hastalarda PV’un
yıllardır var olduğu ortaya kondu.
15. Çalışmanın sonucunda hastalığın kaşıntıya yol açmadığı görüldü.
16. Moleküler biyolojik temelli çalışmalar, tür tanısı yapılamayan maya mantarlarınında
belirlenmesini sağlayabilecektir. Bu nedenle, bu yöntemlerin kullanılması doğru ve
akılcı olacaktır.

85
7. KAYNAKLAR
1. Rippon JW. Medical Mycology 3rd ed. Philadelphia: WB Saunders Company, 1988: 154-9.
2. Baillon H. Traite de Botanique Medicale Cryptogamique. Paris: Octave Doin, 1846;1889: 234.
3. Robin C. Historie Naturelle des Vegeaux Parasites. London: Balliere, 1853: 438.
4. Rivolta S. Dei Parassiti Vegetali come Introduzione allo Studio delle Malattie Parassitarei. E
delle Alterazioni dell’alimento degli Aimali Domestici. Torino, Italy: Speir, 1873: 469–479.
5. Gordon MA. The lipophilic mycoflora of the skin. Mycologica 1951; 43: 524–534.
6. Sloof WC. Pityrosporum sabouraud. In: Lodder J, editor. The Yeasts: a Taxonomic Study. North-
Holland Publishing Co., Amsterdam, 1970: 1167–1186.
7. Sabouraud R. Maladies du cuir chevelu. II. Les maladies desquamatives. Masson, Paris, 1904.
8. Castellani C, Chalmers AJ. Manual of Tropical Medicine, 2nd edn. Wm Wood, New York,
1913.
9. Gordon MA . Lipophilic yeastlike organisms associated with tinea versicolor. J Invest Dermatol,
1951; 17: 267–272.
10. Faergemann J, Fredriksson T. Experimental infections in rabbits and humans with Pityrosporum
orbuiculare and P. ovale. J Invest Dermatol 1981; 77: 314–318.
11. Weidmann FD. Exfoliative dermatitis in the Indian rhinoceros (Rhinoceros unicornis) with
description of a new yeast species Pityrosporum pachydermatis. In: Fox H, editor. Report of the
Laboratory and Museum of Comparative Pathology of the Zoological Society of Philadelphia.
Zoological Society of Philadelphia, Philadelphia, 1925; 36–44.

86
12. Panja, G. The Malassezia of the skin, their cultivation, morphology and species. Trans. 7th
Congr. Far East. Assoc. Trop. Med. 1927;2: 442 456.
13. Borelli D. Pityriasis versicolor due to Malassezia ovalis. Mycopathologia, 1985; 89: 147–153.
14. Faergemann J. Tinea versicolor and Pityrosporum orbiculare: mycological investigations,
experimental infections and epidemiological surveys. Acta Derm Venereol (Suppl)(Stockh) 1979;
86: 1– 23.
15. Faergemann J, Fredriksson T. Age incidence of Pityrosporum orbiculare on human skin. Acta
Derm Venereol, 1980; 60:531–533.
16. Borelli D . Pityriasis versicolor: unicity or duality? Mycopathologia, 1990; 111: 3–(Letter).
17. Benham, RW.The cultural characteristics of Pityrosporum ovale—a lipophilic fungus. J. Investig.
Dermatol. 1939; 2:187–203.
18. Faergemann, J., and T. Fredriksson. 1979. Tinea versicolor with regard to seborrhoeic
dermatitis. Arch. Dermatol. 115:966–968.
19. Leung, DYM. Atopic dermatitis: the skin as a window into the pathogenesis of chronic allergic
diseases. J. Allergy Clin. Immunol. 1995; 96:302–318.
20. Lodder, J., and NJW. Kreger-Van Rij. C. P. Kurtzman and Jack W. Fell (ed.). The yeasts: a
taxonomic study. North-Holland Publishing Co., Amsterdam, The Netherlands. Pityrosporum,
1952;. 440–445.
21. Gordon, M. A.. Lipophilic yeast organism associated with tinea versicolor. J. Investig. Dermatol.
1951;17:267–272.

87
22. Lodder J.. A Taxonomic Study. Amsterdam: North Holland.. The Yeasts, 1979; 1167- 1186.
23. Dorn M, Roehnert K. Dimorphism of Pityrosporum orbiculare in a defined culture medium. J
InvestiDermatol, 1977; 69: 244-248.
24. Nazzaro-Porro M, Passi S, Caprilli F. Induction of hyphae in cultures in Pityrosporum by
cholesterol and cholesterol esters. J Investig Dermatol, 1977; 69:531-534.
25. Salkin F, Gordon MA. Polymorphism of Malassezia furfur. Can J Microbiol, 1977; 23: 471-475.
26. Cannon, P. F.. International Commission on the taxonomy of fungi changes in fungi of
microbiological, industrial and medical importance. Microbiol. Sci. 1986; 3:285–287.
27. Faergemann, J.. Pityrosporum ovale and skin diseases. Keio J. Med, 1993 ;42:91–94.
28. Midgley, G.The diversity of Pityrosporum (Malassezia) yeasts in vivo and in vitro.
Mycopathologia, 1989; 106:143–155.
29. Simmons, R. B., and E. Gueho.. A new species of Malassezia. Mycol. Res. 1990; 94: 1146–1149.
30. Cunningham AC, Leeming JP, Ingham E, Gowland G. Differentiation of three serovars of
Malassezia furfur. J Appl Bacteriol, 1990; 68: 439-446.
31. Guillot, J., and E. Gueho. The diversity of Malassezia yeasts confirmed by rRNA sequence and
nuclear DNA comparisons. Antonie Leeuwenhoek, 1995; 67:297–314.
32. Gueho E, Midgley G, and Guillot J. 1996. The genus Malassezia with description of four new
species. Antonie Leeuwenhoek, 1996; 69:337–355.

88
33. Gupta AK, Kohli Y, Summerbell RC. Molecular differentiation of seven Malassezia species. J
Clin Microbiol, 2000; 38: 1869-1875.
34. Ahern DG, Simmons RB. Malassezia Baillon. CP Kutzman, Fell JW (eds). The Yeasts-a
taxonomic study, 4th ed. Elsevier, Amsterdam, the Netherlands, 1998:782-784.
35. Kano R, Aizawa T, Nakamura Y, Watanabe S, Hasegawa A. Chitin synthase 2 gene sequence
of Malassezia species. Microbiol Immunol, 1999; 43: 813-815.
36. Makimura K, Tamura y, Kudo M, Uchida K, Saito H, Ya maguchi H. Species identification
and strain typing of Malassezia species stock strains and clinical isolates based on the DNA
sequences of nuclear ribosomal internal transcribed spacer 1 regions. J Med Microbiol 2000; 49:
29-35.
37. Sugita T, Kodama M, Saito M, Ito T, Kato Y, Tsuboi R, Nishikawa A. Sequence diversity of
the intergenic spacer region of the rRNA gene of Malassezia globosa colonizing the skin of
patients with atopic dermatitis and healthy individuals. J Clin Microbiol, 2003; 41: 3022- 3027.
38. Saadatzadeh, MR, Ashbee HR, KT Holland, and Ingham E. Production of the mycelial phase
of Malassezia species in vitro. Med. Mycol, 2001;39:487-493.
39. Sugita T, Takashima M, Shinoda T, Suto H, Unno T, Tsuboi R, Ogawa H, Nishikawa A.
New yeast species, Malassezia dermatis, isolated from patients with atopic dermatitis. J Clin
Microbiol, 2002; 40: 1363-1367.
40. Sugita T, Takashima M, Kodama M, Tsuboi R, Nishikawa A. Description of a new yeast
species, Malassezia japonica, and its detection in patients with atopic dermatitis and healthy
subjects. J Clin Microbiol, 2003; 41: 4695-4699.

89
41. Hirai A, Kano R, Makimura K, Duarte ER, Hamdan JS, Lachance MA, Yamaguchi H,
Hasegawa A. Malassezia nana sp. nov., a novel lipid-dependent yeast species isolated from
animals. Int J Syst Evol Microbiol, 2004; 54: 623-627.
42. Takashi S., Tajima M., Takashima M., Amaya M., Saito M., Ryoyi T., and Nisihikawa A. A
New Yeast, Malassezia yamatoensis, Isolated from a Patient with Seborrheic Dermatitis, and Its
Distribution in Patients and Healthy Subjects. Microbiology and immunology, 48,2004;.579-583.
43. Boekhout T, Kamp M, Gueho E. Molecular typing of Malassezia species with PFGE and
RAPD.Med Mycol, 1998; 36:365-372.
44. Theelen B, Silvestri M, Gueho E, Van Belkum A, Boekhout T.Identification and typing of
Malassezia yeasts using amplified fragment length polymorphism (AFLP), random amplified
polymorphic DNA (RAPD) and denaturing gradient gel electrophoresis. FEMS Yeast Res, 2001; 1:
79-86.
45. Gupta AK, Boekhout T, Theelen B, Summerbell R, Batra R. Identification and typing of
Malassezia species by amplified fragment length polymorphism and sequence analyses of the
internal transcribed spacer and large-subunit regions of ribosomal DNA. J Clin Microbiol, 2004;
42: 4253-4260.
46. Cabanes FJ, Hernandez JJ, Castella G. Molecular analysis of Malassezia sympodialis -related
strains from domestic animals. J Clin Microbiol 2005; 43: 277-283.
47. Aizawa T, Kano R, Nakamura Y, Watanabe S, Hasegawa A. The genetic diversity of clinical
isolates of Malassezia pachydermatis from dogs and cats. Med Mycol, 2001; 39: 329-334.
48. Guillot J, Gueho E. The diversity of Malassezia yeasts confirmed by rRNA sequence and nuclear
DNA comparisons. Antonie vanLeeuwenhoek, 1995; 67: 297-314.

90
49. Gueho E, Midgley G, Guillot J. The genus Malassezia with description of four new species.
Antonie van Leeuwenhoek, 1996; 69: 337-355.
50. Boekhout Tand Guého, E. Basidiomycetous yeasts. In: Howard, D.H.(ed.), Pathogenic fungi in
humans and animals, 2nd edition. New York: Marcel Dekker, Inc., 2003;535-64.
51. Guillot, J., E., et al.. Identification of Malassezia species. A practical approach. J Mycal Méd,
1996; 6, 103-10.
52. Mayser, P., Haze,P., et al Differantion of Malassezia species:selectivity of cremophor EL,
castor oil and ricinoleic acid for M. furfur. Br J Dermatol, 1997; 137,208-13.
53. Guého E, Boekhout Tet al. The role of Malassezia species in the ecology of humans skin and as
pathogen. Med Mycol, 1998; 3(1), 220-9.
54. Duarte ER, Lanhance, MA and Hamdan, JS. Identification of atypical strains of Malassezia
spp. from cattle and dog. Can J Microbiol, 2002; 48,749-52.
55. Sugita T, Kodama M, et al. Sequence diversity of intergenic spacer region of the rRNA of
Malassezia globosa colonizing the skin of patients with atopic dermatitis and healthy individuals.
J Clin Microbiol, 2003a; 43, 3022-7.
56. Ashbee HR, Ingham E, et al.. Cell- mediated immune responce to Malassezia furfur serovars A,
B and C in patientes with pityriasis versicolor, seborrheic dermatis and controls. Exp. Dermatol,
1994a; 3, 227-33.
57. Devos, SA and van der valk PG.. The relevance of skin prick tests for Pityrosporum ovale in
patients with head and neck dermatitis. Allergy, 2000;55, 1056-8.

91
58. Kesavan S, Holland KT and Ingham E. The effects of lipid extraction on the
immunomodulatory activity of Malassezia species in vitro. Med Mycol, 2000; 38, 239-47.
59. Koyama T, Kanbe T, et al Isolation and characterization of a major antigenic component of
Malassezia globosa to IgE antibodies in sera of patients with aopic dermatitis. Microbiol Immunol,
2000;44, 373-9.
60. Tengvall Linder M, Johansson C, et al. Positive atopy patch test reactions to Pityrosporum
orbiculare in aopic dermatitis patients. Clin Exp. Allergy, 2000;30, 122-31.
61. Faergemann J, Bergbrant IM, et al.. Seborrhoeic dermatitis and Pityrosporum (Malassezia)
folliculitis: characterzation of inflammatory cells and mediators in the skin by
immunohistochemistry. Br J Dermatol, 2001; 144. 549-56.
62. Ashbee HR and Evans EG. Immunology of diseases associated with Malassezia species. Clin
Microbiol Rev, 2002; 15,21-57.
63. Midgley G. The diversity of Pityrosporum (Malassezia) yeasts in vivo and in vitro.
Mycopathologia, 1989; 106: 143-155.
64. Dorn M, Roehnert K. Dimorphism of Pityrosporum orbiculare in a defined culture medium. J
Investig Dermatol, 1977; 69: 244-248.
65. Hoog GS, Guarro J, Gené J, Figueras MJ (ED)., 2nd ed.Centraalbureau voor
Schimmelcultures/Universitat Rovira i Virgili, Reus, Spain. Atlas of Clinical Fungi, 2000;144-
155.
66. Faergemann J. Atopic dermatitis and fungi. Clin Microbiol Rev, 2002; 15: 543-563.

92
67. Mittag H. Fine structural investigations of Malassezia furfur. II. The envelope of the yeast cells.
Mycoses, 1995; 38: 13-21.
68. Lodder J. A Taxonomic Study. Amsterdam: North Holland Publishing Co . The Yeasts 1979:
1167- 1186.
69. Mayser P, Imkampe A, Winkeler M, Papvassilis C. Growth requirements and nitrogen
metabolism of Malassezia furfur. Arch Dermatol Res 1998; 290: 277- 282. 44.
70. Faergeman J, Bernander S. Microaerophylic and anaerobic growth of Pityrosporum species.
Sabouradia, 1981; 19: 117-121.
71. Wilde PF, and PS Stewart. A study of the fatty acid metabolism of the yeast Pityrosporum
ovale. Biochem. J, 1968; 108:225–23.
72. Caprilli F, Mercantini R, Nazzaro-Porro M, Passi S, and Tonolo A.. Studies of the genus
Pityrosporum in submerged culture. Mycopathol. Mycol. Appl. 1973;51:171–189.
73. Catterall MD, ME. Ward, and Jacobs P. A reappraisal of the role of Pityrosporum orbiculare in
pityriasis versicolor and the significance of extracellular lipase. J. Investig.
Dermatol,1978;71:398–401.
74. Ran Y, Yoshike T, and H. Ogawa.. Lipase of M. furfur: some properties and their relationship to
cell growth. J. Med. Vet. Mycol. 1993;31:77–85.
75. Riciputo RM, Oliveri S, Micali G, and Sapuppo A. Phospholipase activity in Malassezia furfur
pathogenic strains. Mycoses,1996; 39:233–235.

93
76. Plotkin LI, Mathov I, Squiquera L, and Leoni J.. Arachidonic acid released from epithelial
cells by Malassezia furfur phospholipase A(2): a potential pathophysiologic mechanism.
Mycologia,. 1998;90:163–169.
77. De Luca C, M Picardo, A Breathnach, and S Passi. Lipoperoxidase activity of Pityrosporum:
charactersiation of by-products and possible role in pityriasis versicolor. Exp. Dermatol. 1996;
5:49–56.
78. Van Abbe NJ. The investigation of dandruff. J. Soc. Cosmet. Chem,1964;15:609-630.
79. Labows JN, J. McGinley JJ Leyden, and GF Webster. Characteristic gamma lactone odour
production of the genus Pityrosporum. Appl. Environ. Microbiol, 1979;38:412–415.
80. Akamatsu H, J Komura, Y Asada, Y Miyachi, and Y Niwa.. Inhibitory effect of azelaic acid on
neutrophil functions: a possible cause for its efficacy in treating pathogenetically unrelated
diseases. Arch. Dermatol. Res, 1991;283:162–166.
81. Benham RW. The cultural characteristics of Pityrosporum ovale a lipophilic fungus. J. Investig.
Dermatol,1939; 2:187–203.
82. Keddie FM, and L. Barajas. Quantitative ultrastructural variations between P. ovale and P.
orbiculare based on serial section electron microscopy. Int. J. Dermatol,1972; 11:40–48.
83. Hechemy KE, and RW Vanderwyk.. Isolation, purification and chemical analysis of P. ovale
cell wall. Bacteriol. Proc, 1968;68:42.
84. Thompson E, and JR Colvin. Composition of the cell wall of Pityrosporum ovale (Bizzozero)
Castellani and Chalmers. Can. J. Microbiol, 1970;16:263–265.

94
85. Simmons RB, and Ahearn DG.. Cell wall ultrastructure and diazonium blue B reactions of
Speropachydermia quercuum, Bulleo tsugae and Malassezia spp. Mycologia,1987;79:38–43.
86. Swift JA, and Dunbar SF.. Ultrastructure of Pityrosporum ovale and P. canis. Nature
,1965;206:1174–1175.
87. Kreger-van Rij, NJW, and Veenhuis M.. An electron microscope study of the yeast
Pityrosporum ovale. Arch. Microbiol, 1970;71:123–131.
88. Winiarczyk S. The ultrastructure of Pityrosporum pachydermatis. Arch. Vet. Pol, 1992 ;32:4–13.
89. Mittag H. . Fine structural investigations of Malassezia furfur. II. The envelope of the yeast cells.
Mycoses, 1995;38:13–21.
90. Barfatani M, Munn RJ, and Schjeide DA. An ultrastructural study of Pityrosporum orbiculare.
J. Investig. Dermatol, 1964; 43:231–233.
91. Wikler JR, Janssen N, Bruynzeel DP, and Nieboer C. The effect of UV light on Pityrosporum
yeasts: ultrastructural changes and inhibition of growth. Acta Dermato-Venereol. . 1990;70:69–72.
92. Marples RR, Downing DT, Kligman AM. Influence of Pityrosporum species in the generation of
free fatty acids in human surface lipid. J Invest Dermatol, 1972; 58: 155- 159.
93. Ran Y, Yoshike T, Ogawa H. Lipase of M.furfur, some properties and their relationship to cell
growth. J Med Vet Mycol, 1993; 31: 77-85.

95
94. Leeming JP, Holland KT, Bojar RA. The in vitro antimicrobial effect of azalaic acid. Br J
Dermatol, 1996; 115: 351-356.
95. Brasch J, Christophers E. Azalaic acid has antimycotic properties in vitro. Dermatology 1993;
186: 55-58.
96. Rider ED, Christensen RD, Hall DC, Rothstein G. Myeloperoxidase deficiency in neutrophils
of neonates. J Pediatr, 1988; 112: 648-651.
97. Cohen, MM. A simple of procedure for staining tinea versicolor ( M. furfur) with fountain pen
ink. J. Invest Dermatol, 1954;22, 26-9.
98. Yarow D, and Ahearn DG.. Genus 7 Malassezia Baillon. In: Kreger-van Rij, N.J.W. (ed), The
yeasts: a taxanomic study, 3rd edition. Amsterdam: elsevier Science Publ BV, 1984;882-5.
99. Guého, E.and Meyer, SA.. A reevaluation of the genus Malassezia by means of genome
comparison. Antonie van Leeuwenhoek, 1989;55, 245-51.
100. Guillot J and Guého E.. The diversity of Malassezia yeasts confirmed by rRNA sequence and
nuclear DNA comparisons. Antonie van leeuwenhoek, 1995; 67, 297-314.
101. Guého E, Midgley G and Guillot J.. The genus Malassezia with description of four new species.
Anyonie van Leeuwenhoek, 1996;69, 337-55.
102. Gupta AK, Kohli Y. Prevalence of Malassezia species on various body sites in clinically healthy
subjects representing different age groups. Medical Mycology, 2004; 42, 35-42.

96
103. Boekhout T, Kamp M and Guého E.. Molecular typing of Malasseia species with PFGE and
RAPD. Med Mycol, 1998;36,365-72.
104. Murai T, Nakamura Y, et al . Differentiation of Malassezia furfur and Malassezia sympodialis by
glicine utilization. Mycoses,2002; 45,180-3.
105. Crespo Erchiga V, Ojeda Martos A, et al. Mycology of pityriasis versicolor. J Mycol Méd,
1999b; 9, 143-8.
106. Crespo ErchigaV, De Toro I, et al. Pityriasis versicolor. Etude épidémiologique sur 210 cas dans
le Sud de I’Espagne. Proc Cong Soc Fr Mycol Méd (Paris) 2002;
107. Sabouraud R. Diagnostic et traitement des affections du cuir chevelu. Paris: Masson et Cie. 1932.
108. Migley G and Clayton YM. The yest flora of birds and mammals in cativity. Antonie van
Leewenhoek, 1969;35, Suppl E, 23-4.
109. Midreuil F, Guillot J, et al. Genetic diversity in the yeast species Malassezia pachydermatis
analysed by multilocus enzyme electrophpresis. Int J Syst Bacteriol, 1999. 49,1287-94.
110. Guého E And Guillot J. Comments on Malassezia species from dogs and cats. Mycoses, 1999;42,
673-4.
111. Mickelsen PA, Viano-Paulson, MC, et al. Clinical and microbiological feature of infection with
Malassezia pachydermatis in high-risk infants. J Infect Dis, 1998; 157,1163-8.
112. Guillot J And Bond R. Malassezia pachydermatis: a review. Med Mycol,1999; 37,295-306.

97
113. Cunningham AC, Leeming JP, et al.. Differentiation of three serovars of Malassezia furfur. J
Apple Bacteriol, 1990; 68,439-46.
114. Sugita T, Takashima M, et al.New yeasts species, Malassezia dermatis, isolated from patients
with atopic dermatitis. J Clin Microbiol, 2002; 40,1363-7.
115. Kurtzman CP and Blanz PA. Ribosomal RNA/DNA sequence comparisons for assessing
phylogenetic relationships. In: Kurtzman, C.P. and Fell, J.W. (eds), The yeasts-a taxonomic study,
4th revised edition. Amsterdam: Elsevier, 1998,69-74.
116. Nell A, James SA, et al. Identification and distribution of a novel Malassezia species yeasts on
normal equine skin. Vet Rec,2002; 150, 395-8.
117. Hirai A, Kano R, et al. Malassezia nana sp.nov., a novel lipid-dependant yeast species isolated
from animals. Int J Syst Evol Microbiol, 2004;54, 623-7.
118. Crespo MJ, Abarca M. and Cabańes FJ. Otitis externa associated with Malassezia sympodialis
in two cats. J Clin Microbiol, 2000a; 38, 1263-6.
119. Gordon M.A . Lipofilic yeast organism associated with tinea versicolor. J Invest Dermatol, 1951a;
17, 267-72.
120. Crespo MJ, Abarca ML and Cabańes FJ. A typical lipid-dependent Malassezia species isolated
from dogs with otitis externa. J Clin Microbiol, 2000b; 38, 23.
121. Aspiroz C, Moreno LA, et al. Differantion of three biotypes of Malassezia species on normal
human skin. Correspondence with M. globosa, M. sympodialis, and M. restricta. Mycopathologia,
1999; 145,69-74.

98
122. Crespo M., Abarca, ML and Cabańes, F:J. Occurence of Malassezia spp. in horses and
domestic ruminanats. Mycoses, 2002; 45,333-7.
123. Guillot J, Guého E, et al. Importance des levures du genre Malassezia en dermatologie
vétérinaire. Point Vét,1998; 29,21-31.
124. Dufait R. Presence de Malassezia pachydermatis(syn Pityrosporum canis) sur les poils et les
plumes des animaux domestiques. Bull Soc Fr Mycol Méd, 1985; 14,19-22.
125. Guillot J, Chermette R.And Gueho E. Prevalence du genre Malassezia chez les mammiferes. J
Mycol Med,1994; 4, 72-4.
126. Marcon MJ and Powell D.A. Humans infections due to Malassezia spp.. Clin Microbiol Rev,
1992; 5, 101-19.
127. Crespo Erchiga V, Odeja Martos A, et al. Aislomiento e identificatiŏn de Malassezia spp.. en
pityriasis versicolor, dermatitis seborreica y piel sana. Rev Ioberoam Micol, 1999a; S16-21.
128. Midgley G. The lipofilic yeasts: state of the art and prospects. Med Mycol, 2000; 38, Suppl 1,9-16.
129. Nakabayashi A, Sei Ya and Guillot J. Identification of Malassezia species isolated from patients
with seborrhoeic dermatitis, atopic dermatitis, pityriasis versicolor and normal subjects. Med
Mycol, 2000; 38,337-41.
130. Arzumanyan V. The yeast Malassezia on the skin of healty individuals and patients with atopic
dermatitis. Vestn Ross Akad Med Nauk, 2001; 2, 29-31.

99
131. Gupta, AK, Kohli Y, et al . Quantitative culture of Malassezia species from different body sites of
individulas with or without dermatoses. Med Mycol, 2001a; 39,243-51.
132. Sugita, T, Suto, H., et al .Molecular analysis of Malassezia microflora on thr skin of atopic
dermatitis patients and healty subjects. J Clin Microbiol, 2001; 39,3486-90.
133. Hellgren L, and J Vincent. The incidence of tinea versicolor in central Sweden. J Med. Microbiol
,1983; 16:501–502.
134. Midgley G, Gueho E, Guilot J. Disease caused by Malassezia species. In: Ajello L, hay RJ,
editors. Tobley and Wilson’s Micbobiology and microbial infections. Medical mycology. 9th ed.
London: Arnold; 1998; p. 201-14.
135. Robets SOB. Pityrosporum orbiculare incidance and distribution on clinically normal skin. Br J
dermatol, 1969; 81,26-9.
136. Stein DH. Superficial fungal infections. Pediatr Clin North Am 1983; 30: 545–561.
137. Burke RC. Tinea versicolor: susceptibility factors and experimental infection in human beings. J
Invest Dermatol 1961; 36: 389 402.
138. Faergemann J, Bernander S. Tina versicolor and Pityrosporum orbiculare: a mycological
investigation. Sabouraudia 1979; 17: 171–179.
139. McDaniel WE. Tinea versicolor. Arch Dermatol, 1977; 113: 519–520 (Letter).
140. Conklin RJ . Common cutaneous disorders in athletes. Sports Med, 1990; 9: 100–119.

100
141. Pontasch MJ, Kyanko ME, Brodell RT. Tinea versicolor of the face in black children in a
temperate region. Cutis, 1989; 43: 81–84.
142. Roed-Petersen J. Tinea versicolor and body lotions. Acta Derm Venereol, 1980; 60: 439–440
(Letter).
143. Faergemann J, Fredriksson T. Tinea versicolor: some new aspects on etiology, pathogenesis and
treatment. Int J Dermatol 1982; 21: 8–11.
144. Shuttleworth D, Philpot CM, Salaman JR. Cutaneous fungal infection following renal
transplantation: a case control study. Br J Dermatol, 1987; 117: 585–590.
145. Burkhart CG, Dvorak N, Stockard H . An unusual case of tinea versicolor in an
immunosuppressed patient. Cutis, 1981; 27: 56–58.
146. Elmets CA. Management of common superficial fungal infections in patients with AIDS. J Am
Acad Dermatol, 1994; 31: 60–63.
147. Schechtman RC, Midgley G, Hay RJ. HIV disease and Malassezia yeasts: a quantitative study
of patients presenting with seborrhoeic dermatitis. Br J Dermatol, 1995; 133: 694–698.
148. Hashim FA, Elhassan AM. Tinea versicolor and visceral leishmaniasis. Int J Dermatol ,1994; 33:
258–259.
149. Daneshvar SA, Hashimoto K. An unusual presentation of tinea versicolor in an
immunosuppressed patient. J Am Acad Dermatol, 1987; 17 (2 Part 1): 304–305 (Letter).

101
150. Conill C, Azon-Masoliver A, Verger E et al. Pityriasis versicolor confined to the radiation
therapy field. Acta Oncol, 1990; 71: 9–10 (Letter).
151. Fearfield LA, Bunker CB . Pityriasis versicolor associated with oral lithium therapy. Clin Exp
Dermatol, 1997; 22: 57–59 (Letter).
152. Roberts SOB. Pityriasis versicolor: a clinical and mycological investigation. Br J Dermatol, 1969;
81: 315–311.
153. Abdul Razack EM, Thambiah AS. A clinical study of pityriasis versicolor in Madras.
Sabouraudia, 1977; 14: 129–148.
154. Faergemann J, Fredriksson T. Tinea versicolor with regard to seborrheic dermatitis. An
epidemiological investigation. Arch Dermatol, 1979; 115: 966–968.
155. Hafez M, el Shamy S. Genetic susceptibility in pityriasis versicolor. Dermatologica, 1985; 171:
86–88.
156. Roberts SOB. Pityriasis versicolor. In: Verbov JL, editor. Superficial Fungal Infections: New
Clinical Applications in Dermatology. MTP Press, Lancaster, 1986: 47–72.
157. Terragni L, Lasagni A, Oriani A et al. Pityriasis versicolor in the pediatric age. Pediatr
Dermatol, 1991; 8: 9–12.
158. Michalowski R, Rodziewicz H. Pityriasis versicolor in children. Br J Dermatol, 1963; 75: 397–
400.
159. Cullen SI. Age of patients with pityriasis versicolor. J Am Acad Dermatol 1983; 9: 464–465.

102
160. Silverman RA.. In: Elewski BE, editor. Cutaneous Fungal Infections. Igakv Shoin, New York,
Pediatric mycoses, 1992: 212–218.
161. Congly H. Pityriasis versicolor in a 3-month old boy. Can Med Assoc J, 1984; 130: 844- 845
(Letter).
162. Wyre HW . Neonatal pityriasis versicolor. Arch Dermatol, 1981; 117: 752–753.
163. Welbel SF, McNeil MM, Pramanik A et al. Nosocomial Malassezia pachydermatis bloodstream
infections in a neonatal intensive care unit. Pediatr Infect Dis, 1994; 13: 104–108.
164. Ashbee HR, Leck AK, Puntis JW, Parson EJ, Evans EG. Skin colonization by Malassezia in
neonates and infants. Infect control Hosp. Epidemiol, 2002; 23:21-6.
165. Michalowski R, Rodziewicz HP. Versicolor in the aged. Br J Dermatol, 1965; 77: 388–390.
166. Di Silverio A, Mosca M, Brandozzi G et al. Pityriasis versicolor in the aged: a clinical
investigation and epidemiological survey in 190 elderly hospitalized patients. Mycopathologia
1989; 105: 187–190.
167. Schmidt A. Malassezia furfur: a fungus belonging to the physiological skin flora and its relevance
in skin diseases. Cutis, 1997; 59: 197–215.
168. Borton LK, Schwartz RA. Pityrosporum foliculitis: a common acneiform condition of middle
age. Ariz Med, 1981; 38: 598–601.

103
169. Bäck O, Faergemann J, Hörnqvist R. Pityrosporum folliculitis: a common disease of the young
and middle-aged. J Am Acad Dermatol, 1985; 12: 56–61.
170. Bergbrant I-M, Faergemann J. Seborrhoeic dermatitis and Pityrosporum ovale: a cultural and
immunological study. Acta Derm Venereol (Stockh), 1989; 69: 332–335.
171. Svejgaard E. Epidemiology and clinical features of dermatomycoses and dermatophytoses. Acta
Derm Venereol (Suppl) (Stockh), 1986; 121: 19–26.
172. Gonzales Ochoa A. Pityriasis versicolor. Rev Med (Mex,) 1956; 2: 81–88.
173. Marples MJ . The incidence of certain skin diseases in Western Samoa: a preliminary survey.
Trans R Soc Trop Med Hyg, 1950; 44: 319–322.
174. Shrum JP, Millikan LE, Bataineh O. Superficial fungal infections in the tropics. Dermatol Clin
,1994; 12: 687–693.
175. Goncalves AP. Unusual locations of pityriasis versicolor with particular reference to scalp lesions.
Dermatol Trop, 1963; 2: 211–216.
176. el Gothamy Z, Ghozzi M. Tinea versicolor of the scalp. Int J Dermatol, 1995; 34: 533–534.
177. Hughes BR. Tinea versicolor in immunosuppressed patients. J Am Acad Dermatol, 1988; 19 (2
Part 1): 357–358 (Letter).
178. Glazer SD, Zugerman C. Tinea versicolor of the face. Cutis 1980; 26: 87 (Letter).

104
179. Blumenthal HL . Tinea versicolor of penis. Arch Dermatol, 1971; 103: 461–462 (Letter).
180. Nia AK, Smith EL . Pityriasis versicolor of the glans penis. Br J Vener Dis, 1979; 55: 320
(Letter).
181. Rudolph RI, Holzwanger JM. Inverse tinea versicolor. Arch Dermatol, 1975; 111: 1213 (Letter).
182. Anthony JL, Schosser RH, Gross DJ. Unilateral areolar and periareolar tinea versicolor. Int J
Dermatol, 1991; 30: 600 (Letter).
183. Kaur I, Handa S, Kumar B . Tinea versicolor: involvement of unusual sites. Int J Dermatol,
1996; 35: 604–605 (Letter).
184. Crespo MJ, Abarca ML and Cabańes FJ. Evaluation of different preservation and storage
methods for Malassezia spp. J Clin Microbiol, 2000c; 38, 3872-5.
185. Payne DE. How I treat tinea versicolor. Postgrad Med, 1968; 44: 213.
186. Mayser P, Wille G, Imkampe A et al. Synthesis of fluorochromes and pigments in Malassezia
furfur by use of tryptophan as the single nitrogen source. Mycoses 1998; 41: 265–271.
187. Weiss R, Raabe P, Mayser P. Yeasts of the genus Malassezia: taxonomic classification and
significance in veterinary and clinical medicine. Mycoses 2000; 43 (Suppl. 1): 69–72.
188. Albert H A new stain for diphtheria bacilli. JAMA 1920; 74: 28.

105
189. Payle B, Serrano L, Bieley HC et al. Albert’s solution versus potassium hydroxide solution in the
diagnosis of tinea versicolor. Int J Dermatol 1994; 33: 182–183.
190. Leeming JP, Notman FH, Holland KT. The distribution and ecology of Malassezia furfur and
cutaneous bacteria on human skin. J Appl Bacteriol 1989; 67: 47–52.
191. Faergemann J. Tinea versicolor and Pityrosporum orbiculare: mycological investigations,
experimental infections and epidemiological surveys. Acta Derm Venereol (Suppl)(Stockh) 1979;
86: 1– 23.
192. Roberts SOB. Pityrosporum orbiculare: incidence and distribution on clinically normal skin. Br.
J. Dermatol.1969; 81:264–269.
193. Leeming JP, Notman FH, and KT. Holland. The distribution and ecology of Malassezia furfur
and cutaneous bacteria on human skin. J. Appl. Bacteriol,1989;67:47–52.
194. Korting HC, Loferer S, and Hamm N.The detergent scrub method for quantitative determination
of Malassezia furfur on chest and back: comparative evaluation of three different media.
Mycoses,1991; 34:267–271.
195. Bergbrant IM, and Faergemann J. Variations of Pityrosporum orbiculare in middle-aged and
elderly individuals. Acta Dermato-Venereol,1988; 68:537–540.
196. Cunningham ACE, Ingham, and Gowland G. Humoral responses to Malassezia furfur serovars
A, B and C in normal individuals of various ages. Br. J. Dermatol. 1992;127:476–481.
197. Bos JD, and ML Kapsenberg. The skin immune system: its cellular constituents and their inter
actions. Immunol,1989;7:235–240.

106
198. Midgley G, Gueho E, Guillot J. Disease caused by Malassezia species. Ajello L, Hay RJ (eds).
Topley and Wilson’s Microbiology and Microbial Infections, vol 4. Arnold, London, United
Kingdom’da 1998; 201-21.
199. Marcon MJ, and Powell DA. Human infections due to Malassezia spp. Clin. Microbiol
Rev,1992; 5:101–119.
200. Midgley G. The diversity of Pityrosporum (Malassezia) yeasts in vivo and in vitro.
Mycopathologia ,1989;106:143–155.
201. McGinley, KJ, Lantis LR, and Marples RR. Microbiology of tinea versicolor. Arch.
Dermatol,1970; 102:168–171.
202. Aspiroz C, Moreno LA, et al. Differentiation of three biotypes of Malassezia species on normal
human skin. Correspondance with M. globosa, M. sympodialis and M. restricta. Mycopathologia,
1999;145, 69-74.
203. Dotz, WI., D M. Henrikson, and GSM Yu. 1985. Tinea versicolor: a light and electron
microscopic study of hyperpigmented skin. J. Am. Acad. Dermatol,1985;12:37–44.
204. Ashbee HR, Gunning J,Holland KT, Cunliffe WJ, and E. Ingham. Titres of IgE specific to
Malassezia furfur serovars A, B and C in patients with pityriasis versicolor and controls. J.
Investig. Dermatol,1985;105: 492.
205. Farthing CF, Staughton RCD, Rowland Payne CME. Skin disease in homosexual patients with
Acquired Immune Deficiency Syndrome (AIDS) and lesser forms of human T cell Leukaemia
Virus (HTLV III) disease. Clin Exp Dermatol 1985;10:3-12.
206. Ramasastry P, Downing DT, Pochi PE, Strauss JS. Chemical composition of human skin
surface lipids from birth to puberty. J Invest Dermatol, 1970;54:139-44.

107
207. Mayser P, Schafer U, Kramer HJ, Irlinger B, Steglich W. Pityriacitrinean ultraviolet-absorbing
indole alkaloid from the yeast Malassezia furfur. Arch Dermatol Res, 2002;294: 131-4.
208. Faergemann J. Atopic dermatitis and fungi. Clin Microbiol Rev, 2002;15:545-63.
209. Bäck O, Faergemann J, Hörnqvist R. Pityrosporum folliculitis: a common disease of the young
and middle-aged. J Am Acad Dermatol,1985; 12: 56–61.
210. el Gothamy Z, Ghozzi M. Tinea versicolor of the scalp. Int J Dermatol, 1995; 34: 533–534.
211. Goncalves AP. Unusual locations of pityriasis versicolor with particular reference to scalp lesions.
Dermatol Trop, 1963; 2: 211–216.
212. Alves EV, Martins JE, et. Al. Pityrosporum folliculitis: renal transplantation case report. J.
Dermatol,2000; 27,49-51.
213. Back O, Faergemann J, Hornqvist R. Pityrosporum folliculitis: a common disease of the young
and middle-aged. J Am Acad Dermatol, 1985;12:56-61.
214. Katoh T, Irimajiri J . Pityriasis versicolor and Malassezia folliculitis. Nippon Ishinkin Gakkai
Zasshi 1999;40:69-71.
215. Faergemann J, Johansson S, Back O, Scheynius A. An immunologic and cultural study of
Pityrosporum folliculitis.J Am Acad Dermatol, 1986;14:429-33.

108
216. Nordby CA and Mitchell AJ. Confleuent and reticulated papillomatis responsive to selenium
sulfide. Int J Dermatol,1986; 25, 194-9.
217. Mayser P, Schutz M, Schuppe HC, Jung A, Schill WB. Frequency and spectrum of Malassezia
yeasts in the area of the prepuce and glans penis. BJU Int, 2001;88:554-8.
218. Prohic A. Identification of Malassezia species isolated from scalp skin of patients with psoriasis
and healthy subjects. Acta Dermatovenerol Croat, 2003;11:10-6.
219. Ackerman AB. Histopathologic differentiation of eczematous dermatitides from psoriasis and
seborrheic dermatitis. Cutis 1977;20:619-23.
220. Rapelanoro R, Mortureux P, et al. Neonatal Malassezia furfur pustulosis. Arch Dermatol,1996;
132, 190-3.
221. Silvia V, Moreno GA, et.al. Isolation of Malassezia furfur from patients with onychomycosis. J
Med Vet Mycol, 1979; 35,73-74.
222. Hersle K. Selenium sulphide treatment of tinea versicolor. Acta Derm Venereol, 1971; 51: 476–
478.
223. Sanchez JL, Torres VM. Double-blind efficacy study of selenium sulfide in tinea versicolor. J
Am Acad Dermatol, 1984; 11 (2 Part 1): 235–238.
224. Chu AC. Comparative clinical trial of bifonazole solution versus selenium sulfide shampoo in the
treatment of pityriasis versicolor. Dermatologica 1984; 169 (Suppl. 1): 81–86.

109
225. Petranyi G, Ryder NS, Stütz A. Allylamine derivatives: new class of synthetic antifungal agents
inhibiting fungal squalene epoxidase. Science 1984; 224: 1239–1241.
226. Sadeque JB, Shahidullah M, Shah OR et al. Systemic ketoconazole in the treatment of tinea
versicolor. Int J Dermatol 1995; 34: 504–505.
227. Hagler AN and Ahearn DG. Rapid diazonium blue B test to detect basidiomycetous yeasts. Int J
Syst Bacteriol 1981;31, 204–208.
228. Crespo VE, Martos AO, Casańo AV, Fajardo FS. Malassezia globosa as the causative agent of
pityriasis versicolor. British journal of Dermatology, 2000; 143: 799-803.
229. Gupta AK, Bluhm R, Summerbel R. Pityriasis versicolor. JEADV, 2002;16 , 19–33.
230. Aspiroz C, Ara M, Varea M, Rezusta A, Rubio C. Isolation of Malassezia globosa and M.
sympodialis from patients with pityriasis versicolor in Spa. Mycopathologia, 2001;154: 111–117.
231. Gupta AK, Kohli Y, Faergemann J et al. Epidemiology of Malassezia yeasts associated with
pityriasis versicolor in Ontario, Canada. Med Mycol 2001; 39: 199–206.
232. Dikshit A, Dutta S, Bajaj AK . Pityriasis versicolor socio-economic and clinico-economic study
in India.
233. Jena DK, Sengupta S, Dwari BC, Ram MK. Pityriasis versicolor in the pediatric age group.
Indian J Dermatol Venereol Leprol, 2005; 71 (4): 259-61.
234. Payle B, Serrano L, Bieley HC et al. Albert’s solution versus potassium hydroxide solution in the
diagnosis of tinea versicolor. Int J Dermatol, 1994; 33: 182–183.

110
235. Tarazooie B, Kordbacheh P, Zaini F, Zomorodian K, Saadat F, Zeraati H, Hallaji Z, Rezaie
S. Study of the distribution of Malassezia species in patients with pityriasis versicolor and healthy
individuals in Tehran, Iran. BMC Dermatol, 2004; 4: 1-6.
236. Scorzetti, G, Fell, JW, Fonseca, A.& Statzell-Tallman, A. Systematics of basidiomycetous
yeasts: a comparison of large subunit D1/D2 and internal transcribed spacer rDNA regions. FEMS
Yeast Res,2002 2, 495–517.
237. Prohic A, Ozegovic L. Malassezia species from lesional and non- lesional skin in patient with
pityriasis versicolor. Mycoses, 2006;50:58–63.
238. Makimura K, Tamura Y, Kudo M, Uchida K, Saito H, Ya maguchi H. Species identification
and strain typing of Malassezia species stock strains and clinical isolates based on the DNA
sequences of nuclear ribosomal internal transcribed spacer 1 regions. J Med Microbiol, 2000; 49:
29-35.
239. Nakabayashi A, Y Sei, and J Guillot. Identification of Malassezia species isolated from patients
with seborrhoeic dermatitis, atopic dermatitis, pityriasis versicolor and normal subjects. Med.
Mycol,2000; 38:337–341.
240. Ben Salah S, Makni F, Marrakchi S, Sellami H, Cheikhrouhou F, Bouassida S, Zahaf A,
and Ayadi A. Identification of Malassezia species from Tunisian patients with pityriasis
versicolor and normal subjects. Mycoses, 2005; 48, 242–245.
241. Gaitanis G, Velegraki A, et.al. Identification of Malassezia species from skin scales by PCR-
RFLP. Clin Microbial Infect, 2002; 162-73.
242. Mayser P, Schütz H, Schuppe C, Jung A and Schill B. Frequency and spectrum of Malassezia
yeasts in the area of the prepuce and glans penis. BJU international, 2001; 88,554-558.

111
243. Karakaş M, Đlkit M, Turaç-Biçer A. Epidemiology of pityriasis versicolor in Adana. Journal of
Dermatology, 2009; 36: 377–382.
244. Salahi-Moghaddam A, Davoodian P, Jafari A, Nikoo MA. Evaluation of pityriasis versicolor in
prisoners. Indian J Dermatol Leprol , 2009; 75(4):379-81.
245. He SM, Du WD, Yang S, Zhou SM, Li We. The genetic epidemiology of tinea versicolor in
China. Mycoses,2007; 51, 55–62.
246. Faergeman J, Fredrikson T. Age of Pityrosporum orbiculare on humn skin. Acta Derm Venerol,
1980; 60:53-533.
247. Mandel EH, Ores RO. The Incidence of Diabetes Mellitus inPatients with Tinea Versicolor.
Journal of the national medical association.1974;66(3),198-200.
248. Zampino MR, Osti F, Corazza M, Virgil A. Prevalence of pityriasis versicolor in a group of
Italian pregnant women. European Academy of Dermatology and Venereology. 2007,21:1249–
1252.

112
8. ÖZGEÇMĐŞ
Kırşehir’de 1982’de doğdu. Đlk eğitimini Van ve Şanlıurfa’da, orta ve lise
eğitimini Şanlıurfa’da tamamladı. Çukurova Üniversitesi Fen-Edebiyat Fakültesi
Biyoloji Bölümünde 2001’de Lisans eğitimine başladı. 2005’de mezun oldu. Çukurova
Üniversitesi Sağlık Bilimleri Enstitüsü Mikrobiyoloji Anabilim Dalında 2005 yılında
Yüksek Lisans eğitimine başlayıp, halen aynı anabilim dalında eğitimine devam
etmektedir.
Pınar ALTINTOP