shc, grb2, sosl, protein complexes fms hematopoietic cells
TRANSCRIPT

MOLECULAR AND CELLULAR BIOLOGY, Sept. 1994, p. 5682-56910270-7306/94/$04.00+0Copyright © 1994, American Society for Microbiology
Shc, Grb2, Sosl, and a 150-Kilodalton Tyrosine-PhosphorylatedProtein Form Complexes with Fms in Hematopoietic CellsMARIO N. LIOUBIN,l.2* GARY M. MYLES,' KRISTEN CARLBERG,' DAVID BOWTELL,3
AND LARRY R. ROHRSCHNEIDER' 2
Fred Hutchinson Cancer Research Center, Seattle, Washington 981041; Department of Pathology, University ofWashington, Seattle, Washington 981952; and Howard Florey Institute of Experimental Physiology
and Medicine, University of Melboume, Parkville, Victoria 3052, Australia3
Received 1 March 1994/Returned for modification 4 April 1994/Accepted 31 May 1994
Fms, the macrophage colony-stimulating factor (M-CSF) receptor, is normally expressed in myeloid cellsand initiates signals for both growth and development along the monocyte/macrophage lineage. We haveexamined Fms signal transduction pathways in the murine myeloid progenitor cell line FDC-P1. M-CSFstimulation of FDC-P1 cells expressing exogenous Fms resulted in tyrosine phosphorylation of a variety ofcellular proteins in addition to Fms. M-CSF stimulation also resulted in Fms association with two of thesetyrosine-phosphorylated proteins, one of which was identified as the 55-kDla Shc, which is shown in othersystems to be involved in growth stimulation, and the other was a previously uncharacterized 150-kDa protein(p150). Fms also formed complexes with Grb2 and Sosl, and neither contained phosphotyrosine. Whereas bothGrb2 and Sosl complexed with Fms only after M-CSF stimulation, the amount of Sosl complexed with Grb2was not M-CSF dependent. Shc coimmunoprecipitated Sosl, Grb2, and tyrosine-phosphorylated p150, whileGrb2 immunoprecipitates contained mainly phosphorylated p150, Fms, Shc, and Sosl. Shc interacted withtyrosine-phosphorylated p150 via its SH2 domain, and the Grb2 SH2 domain likewise bound tyrosine-phosphorylated Fms and p150. Analysis of Fms mutated at each of four tyrosine autophosphorylation sitesindicated that none of these sites dramatically affected p150 phosphorylation or its association with Shc andGrb2. M-CSF stimulation of fibroblast cell lines expressing exogenous murine Fms did not phosphorylatep150, and this protein was not detected either in cell lysates or in Grb2 or Shc immunoprecipitates. The p150protein is not related to known signal transduction molecules and may be myeloid cell specific. These resultssuggest that M-CSF stimulation of myeloid cells could activate Ras through the nucleotide exchange factorSosl by Grb2 binding to either Fms, Shc, or p150 and that Fms signal transduction in myeloid cells differs fromthat in fibroblasts.
The fins proto-oncogene encodes the cellular receptor forthe macrophage colony-stimulating factor (M-CSF) (for re-
views, see references 31 and 34). This receptor protein (Fms)contains an extracellular domain composed of five tandemimmunoglobulin-like loops and a cytoplasmic tyrosine kinasedomain split by the insertion of approximately 70 hydrophilicamino acids termed the kinase insert region. These structuralfeatures define a class of growth factor receptors whoseimmediate family members include the platelet-derived growthfactor (PDGF) receptors, c-Kit, and the Flt3/Flk2 proteins (7,19, 20, 28, 35, 54).Fms is expressed in monocytes/macrophages and their pro-
genitor cells and in decidual and trophoblast cells of thedeveloping embryo (34). The primary function of Fms in themyeloid cells is to stimulate the growth and direct the differ-entiation of progenitor cells along the monocyte/macrophagelineage. Mature cell functions of macrophages are also acti-vated upon M-CSF stimulation, and cell survival is augmentedby low levels of M-CSF.The molecular signals that transduce information for Fms
functions are not well understood, and only the initial eventshave been characterized. M-CSF binding to the amino-termi-nal three immunoglobulin-like loops of the extracellular do-main of the receptor starts the process by inducing dimeriza-
* Corresponding author. Mailing address: Fred Hutchinson CancerResearch Center, 1124 Columbia St., Seattle, WA 98104. Phone: (206)667-4437. Fax: (206) 667-6522.
tion of receptor molecules (4, 52). The cytoplasmic tyrosinekinase domain is activated, and receptor molecules are
transphosphorylated on at least four tyrosine residues (30, 37,48) which may then act as recognition sites for binding SH2domain-containing proteins such as the p85 subunit of phos-phatidylinositol 3'-kinase (30) and the adaptor protein Grb2(49). The further steps in the propagation of signals down-stream from Fms are not clear; however, signaling mechanismsfor the related PDGF receptor and for growth factor receptorsbelonging to other classes of tyrosine kinase receptors such as
the epidermal growth factor (EGF) receptor and the insulinreceptor have been described (1, 3, 11, 12, 17, 18, 21, 38, 39, 44,45). Extrapolating from the results with these receptors, we
might expect Src family members to bind to a tyrosine-phosphorylated site near the juxtamembrane region of Fms(23), and additional signaling molecules like Shc (25) and son
of sevenless (Sos) proteins (2) to participate in the growthsignaling pathway leading to Ras activation and nuclear tran-scription via Raf and the mitogen-activated protein kinasepathway (22, 50, 53).The main problem with extrapolating from these other
growth factor receptor systems is that although they are allmembers of a closely related family, Fms behaves differentlyand does not associate with the same set of cellular signaltransduction molecules as the PDGF receptor. Both the GT-Pase-activating protein and phospholipase Cy associate withactivated PDGF receptor but not with the activated M-CSFreceptor (9, 29). Therefore, although one would expect some
5682
Vol. 14, No. 9
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 05
Feb
ruar
y 20
22 b
y 22
2.23
9.16
0.51
.

Fms SIGNAL TRANSDUCTION 5683
common utilization of growth signaling pathways, basic differ-ences must exist in their overall signaling mechanisms. Afterall, Fms functions in myeloid cell development, and anycomparison with signaling mechanisms in fibroblasts maytotally miss cell-type-specific signaling pathways.We therefore examined the signaling mechanisms for Fms
expressed in the myeloid progenitor cell line FDC-P1 (32) andlooked for similarities and differences in these mechanismswith fibroblast signaling mechanisms. Our results indicate theexistence of diverse pathways in myeloid cells and identify anew myeloid cell-specific protein involved in Fms signal trans-duction.
MATERIALS AND METHODS
Materials. Polyclonal antibodies to Shc were provided by T.Pawson. Anti-Grb2 polyclonal antibody 50 was provided by A.Batzer, and monoclonal antibodies to Grb2 were purchasedfrom Transduction Laboratories (Lexington, Ky.). Polyclonalantibodies to Sosl (Earth and DL-10) were prepared topeptides of the murine protein (38), and polyclonal antibodiesto Fms (5674 and 4062) were raised in our laboratory (4, 33).Secondary antibodies conjugated to horseradish peroxidasewere purchased from Boehringer Mannheim. Unless otherwisespecified, all other reagents were purchased from Sigma (St.Louis, Mo.).
Tissue culture. FDC-P1 cells were propagated in Dulbeccomodified Eagle medium supplemented with 10% fetal bovineserum and 2% WEHI 3B cell conditioned medium, whichprovided the interleukin-3 required for growth. Rat2 and T2cells were propagated in Dulbecco modified Eagle mediumsupplemented with 10% calf serum. All cell lines were grown at37°C, 5% C02, and 95% humidity. FDC-P1 and Rat2 cellswere infected with Fms provirus produced by T2 cells aspreviously described (13).
Construction of Fms mutants. Single and multiple pointmutations were introduced in Fms as described by Tapley et al.(48). FDC-P1 cells were infected with retroviral constructsencoding wild-type or mutant Fms. Fms-expressing cells wereselected by fluorescence-activated cell sorting and cultured inmedium containing interleukin-3.
Immunoprecipitations and Western blot (immunoblot)analysis. Cells were lysed in lysis buffer (20 mM Tris [pH 7.5],10 mM EDTA, 100 mM NaCl, 1 mM ZnCI2, 0.05% NaN3, 2%polyoxyethylene 9 lauryl ether [CI2E9], 2 mM orthovanadate, 1mM phenylmethylsulfonyl fluoride, 10 mM iodoacetamide,and S ,ug of leupeptin per ml). After 15 min on ice, lysates werespun at 25,000 x g for 30 min, and the pellet containing nucleiand other insoluble material was discarded. For Western blotanalysis, samples were electrophoresed on sodium dodecylsulfate (SDS)-7.5 or 8.75% polyacrylamide gels and trans-ferred to nitrocellulose membranes for 2 h at 180 mA with asemidry blotting apparatus (Ellard Instrumentation Ltd.). Af-ter transfer, the membranes were processed as described byCarlberg and Rohrschneider (4). Briefly, the membranes wereblocked overnight at 4°C with block buffer (1% bovine serumalbumin and 1% ovalbumin in rinse buffer [10 mM Tris-HCl{pH 7.5}, 300 mM NaCl, 0.5% Tween 20]). The nitrocellulosemembranes were incubated for 1 h with primary antibodydiluted in block buffer, and the signal was visualized withhorseradish peroxidase-conjugated secondary antibodies andECL (enhanced chemiluminescence) reagents (Amersham). Insome cases, the blots were stripped of antibodies by extensivewashing in strip buffer (100 mM glycine-HCl [pH 2.5]), blockedwith block solution for 1 h at room temperature, and thenreprobed with different antibodies as described above.
For immunoprecipitations, lysates were incubated with theprimary antibody and 10 ,ul of protein G-Sepharose beads(Pharmacia) for 1 h at 4°C. The beads were washed five timeswith wash buffer (50 mM Tris [pH 8.0], 150 mM NaCl, 0.1%CI2E9, 200 ,uM orthovanadate), and proteins were eluted bythe addition of 50 pul of 2x SDS-polyacrylamide gel electro-phoresis (PAGE) sample buffer and heating at 95°C for 5 min.In some experiments, Fms antibodies immobilized to agarosebeads were used (4). Alternatively, cell lysates were incubatedwith the purified glutathione S-transferase (GST) fusion pro-teins at 4°C for 2 h, and GST fusion proteins were immuno-precipitated by addition of 20 pL of glutathione-agarose beads.The beads were washed five times with wash buffer, andproteins were eluted as described above for SDS-PAGEanalysis.GST-Grb2 fusion proteins. The cDNA for the murine Grb2
was PCR cloned from an EL4 library (Stratagene) by usingnondegenerate oligonucleotides based on the nucleotide se-quence of the human cDNA (18). The oligonucleotide se-quences were as follows: Hu-Grb2 5' NDG, 5'-TA GCC ATCGCC AAA TAT GAC TTC AAA GC-3'; and Hu-Grb2 3'NDG, 5'-TTA GAC GTT CCG GTT CAC GGG GGT G-3'.The 651-bp DNA fragment was gel purified and ligated directlyinto the pT7Blue (Novagene) T-overhang vector. Two positiveclones were identified by PCR screening using the T7 universalsequencing primer and Hu-Grb2 3' NDG. The identity of eachof these clones was confirmed by dideoxy sequencing ofsingle-strand phagemid, and one of the clones was sequencedin its entirety. Using this full-length murine Grb2 cDNA cloneas a template, we used the following oligonucleotides in a PCRto clone the Grb2 SH2 domain: Hu-Grb2-SH2 UP (5'-G AGAGAG GAT CCA TGG TTT TTlT GGC AAA ATC CCC AGCGCC-3') and Hu-Grb2-SH2 DN (5'-GAG AGA GAA TTCTCA CGG CTG CTG TGG CAC CTG TTC TAT GTC C-3').The full-length murine Grb2 and/or murine Grb2 SH2 domainDNA was digested with BamHI and EcoRI and cloned into thesame sites of the pGEX-1 vector (Pharmacia).GST-Shc SH2 fusion proteins. The murine Shc SH2 was
PCR cloned from Trisolve (Biotecx, Houston, Tex.)-purifiedBALB/c RNA, using two degenerate oligonucleotides based onthe human Shc sequence (25): Shc SH2 R (5'-GAGAGAGGATCCATGGTTC (T) CAT (C) GGG (C) AAG (A) CT-3')and Shc SH2 LD (5'-GAGAGAGAATTCTCAC (T) TCC (G)ACA (T) (G) GGT (C) TGC (T) TGT (C) AG-3'). The ShcSH2 domain DNA fragment was purified on an agarose gel,digested with BamHI and EcoRI, and cloned into pBSKII+(Stratagene). A positive clone was identified by dideoxy se-quencing, and the DNA fragment was subcloned into theBamHI-EcoRI site of pGEX-1.
Purification ofGST fusion proteins. All GST fusion proteinswere affinity purified from cell lysates of Escherichia coliXLlBlue (Stratagene) by adsorption to glutathione-agaroseresin, followed by elution with 5 mM glutathione as previouslydescribed (46).
RESULTS
We initially examined the temporal sequence of M-CSF-stimulated protein tyrosine phosphorylation in FDC-P1 cellsexpressing exogenous murine Fms (FDfms cells). When FD-fms cells were stimulated with saturating amounts of murineM-CSF, proteins of 165, 150, 120 to 125, 95 to 100, 63, and 55kDa were reactive with a monoclonal antiphosphotyrosineantibody in Western blot analysis (Fig. lA). The mature Fms(165 kDa) was the first protein phosphorylated, reaching amaximum within 1 min and declining to almost basal levels
VOL. 14, 1994
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 05
Feb
ruar
y 20
22 b
y 22
2.23
9.16
0.51
.
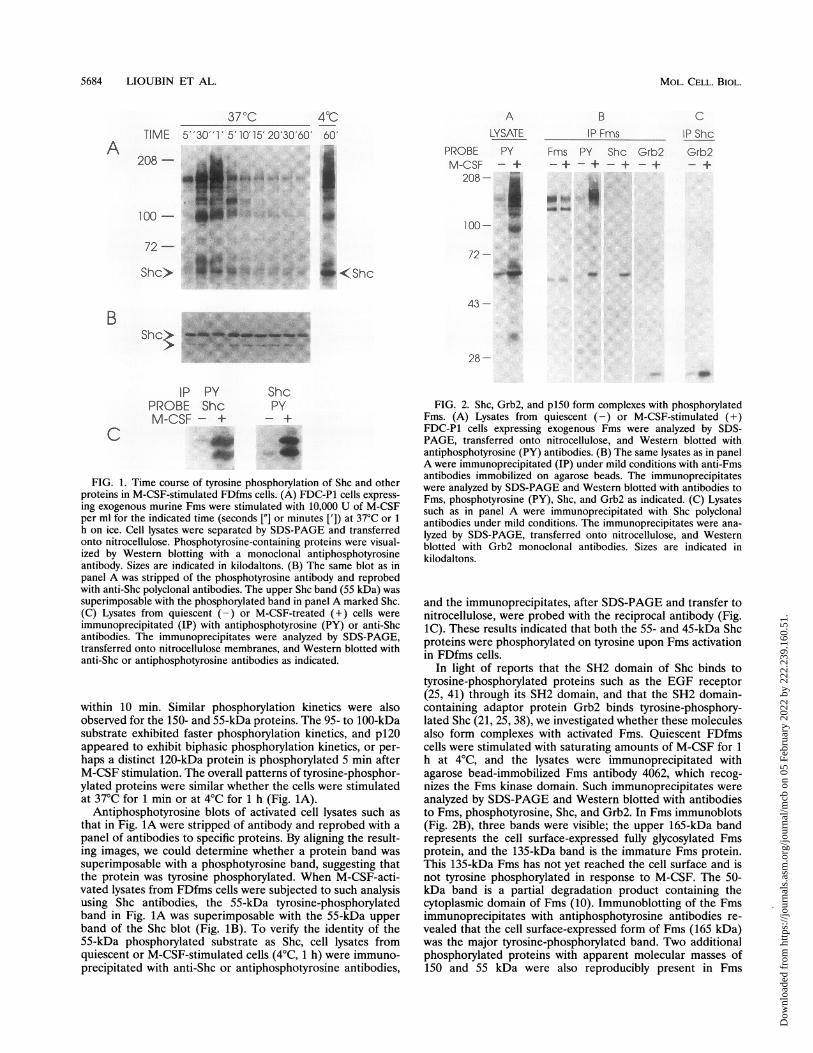
5684 LIOUBIN ET AL.
37 °CTIME 5"30"115'10'
208 -
100 -
72 -
Shc>
Shc;
IPPROBEM-CSF
-1 40C15' 20'30'60' 60'
..
...
PY ShcShc PY
FIG. 1. Time course of tyrosine phosphorylation of Shc and otherproteins in M-CSF-stimulated FDfms cells. (A) FDC-P1 cells express-ing exogenous murine Fms were stimulated with 10,000 U of M-CSFper ml for the indicated time (seconds ["] or minutes [']) at 37°C or 1
h on ice. Cell lysates were separated by SDS-PAGE and transferredonto nitrocellulose. Phosphotyrosine-containing proteins were visual-ized by Western blotting with a monoclonal antiphosphotyrosineantibody. Sizes are indicated in kilodaltons. (B) The same blot as inpanel A was stripped of the phosphotyrosine antibody and reprobedwith anti-Shc polyclonal antibodies. The upper Shc band (55 kDa) wassuperimposable with the phosphorylated band in panel A marked Shc.(C) Lysates from quiescent (-) or M-CSF-treated (+) cells wereimmunoprecipitated (IP) with antiphosphotyrosine (PY) or anti-Shcantibodies. The immunoprecipitates were analyzed by SDS-PAGE,transferred onto nitrocellulose membranes, and Western blotted withanti-Shc or antiphosphotyrosine antibodies as indicated.
within 10 min. Similar phosphorylation kinetics were alsoobserved for the 150- and 55-kDa proteins. The 95- to 100-kDasubstrate exhibited faster phosphorylation kinetics, and p120appeared to exhibit biphasic phosphorylation kinetics, or per-haps a distinct 120-kDa protein is phosphorylated 5 min afterM-CSF stimulation. The overall patterns of tyrosine-phosphor-ylated proteins were similar whether the cells were stimulatedat 370C for 1 min or at 40C for 1 h (Fig. 1A).
Antiphosphotyrosine blots of activated cell lysates such asthat in Fig. 1A were stripped of antibody and reprobed with apanel of antibodies to specific proteins. By aligning the result-ing images, we could determine whether a protein band wassuperimposable with a phosphotyrosine band, suggesting thatthe protein was tyrosine phosphorylated. When M-CSF-acti-vated lysates from FDfms cells were subjected to such analysisusing Shc antibodies, the 55-kDa tyrosine-phosphorylatedband in Fig. 1A was superimposable with the 55-kDa upperband of the Shc blot (Fig. 1B). To verify the identity of the55-kDa phosphorylated substrate as Shc, cell lysates fromquiescent or M-CSF-stimulated cells (40C, 1 h) were immuno-precipitated with anti-Shc or antiphosphotyrosine antibodies,
ALYSATE
PROBE PYM-CSF - +208- -
...:..:.
100-
72-
43-
.2
28-
BIP Fms
Fms PY Shc Grb2-+ - + - + - +
... __4,.
CIP ShcGrb2
FIG. 2. Shc, Grb2, and pl50 form complexes with phosphorylatedFms. (A) Lysates from quiescent (-) or M-CSF-stimulated (+)FDC-P1 cells expressing exogenous Fms were analyzed by SDS-PAGE, transferred onto nitrocellulose, and Western blotted withantiphosphotyrosine (PY) antibodies. (B) The same lysates as in panelA were immunoprecipitated (IP) under mild conditions with anti-Fmsantibodies immobilized on agarose beads. The immunoprecipitateswere analyzed by SDS-PAGE and Western blotted with antibodies toFms, phosphotyrosine (PY), Shc, and Grb2 as indicated. (C) Lysatessuch as in panel A were immunoprecipitated with Shc polyclonalantibodies under mild conditions. The immunoprecipitates were ana-lyzed by SDS-PAGE, transferred onto nitrocellulose, and Westernblotted with Grb2 monoclonal antibodies. Sizes are indicated inkilodaltons.
and the immunoprecipitates, after SDS-PAGE and transfer tonitrocellulose, were probed with the reciprocal antibody (Fig.1C). These results indicated that both the 55- and 45-kDa Shcproteins were phosphorylated on tyrosine upon Fms activationin FDfms cells.
In light of reports that the SH2 domain of Shc binds totyrosine-phosphorylated proteins such as the EGF receptor(25, 41) through its SH2 domain, and that the SH2 domain-containing adaptor protein Grb2 binds tyrosine-phosphory-lated Shc (21, 25, 38), we investigated whether these moleculesalso form complexes with activated Fms. Quiescent FDfmscells were stimulated with saturating amounts of M-CSF for 1h at 40C, and the lysates were immunoprecipitated withagarose bead-immobilized Fms antibody 4062, which recog-nizes the Fms kinase domain. Such immunoprecipitates wereanalyzed by SDS-PAGE and Western blotted with antibodiesto Fms, phosphotyrosine, Shc, and Grb2. In Fms immunoblots(Fig. 2B), three bands were visible; the upper 165-kDa bandrepresents the cell surface-expressed fully glycosylated Fmsprotein, and the 135-kDa band is the immature Fms protein.This 135-kDa Fms has not yet reached the cell surface and isnot tyrosine phosphorylated in response to M-CSF. The 50-kDa band is a partial degradation product containing thecytoplasmic domain of Fms (10). Immunoblotting of the Fmsimmunoprecipitates with antiphosphotyrosine antibodies re-vealed that the cell surface-expressed form of Fms (165 kDa)was the major tyrosine-phosphorylated band. Two additionalphosphorylated proteins with apparent molecular masses of150 and 55 kDa were also reproducibly present in Fms
A
B
C
MOL. CELL. BIOL.
UZ < Shc
...:
..:..
..-....%....
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 05
Feb
ruar
y 20
22 b
y 22
2.23
9.16
0.51
.

Fms SIGNAL TRANSDUCTION 5685
LYSATE Grb2M-CSF - + - +
208 -_
s --
IPShc Fms_+- +
IP FmsPHENPHOS - - + +
M-CSF - + - +
Shc_ _-+ +-_
208 -mm w-R < mSosI
.......:: a ad
14
100-100-
.:go <<Shc_ 40 <IgG
FIG. 3. Sosl forms complexes with Grb2, Shc, and Fms. Cell lysatesfrom quiescent (-) or M-CSF-stimulated (+) FDC-P1 cells expressingexogenous murine Fms were immunoprecipitated (IP) with antibodiesto Grb2, Shc, or Fms as indicated. Whole cell lysates as well as theimmunoprecipitates were analyzed by SDS-PAGE and Western blot-ted with polyclonal antibodies to mSosl. Sizes are indicated inkilodaltons.
immunoprecipitates, whereas the phosphorylated band migrat-ing just above the mature Fms was not always observed. The150-kDa phosphorylated protein was less abundant and wasnot very easily visualized because of its migration close to theheavily phosphorylated Fms band. The same blot, whenstripped of antiphosphotyrosine antibodies and reprobed withShc antibodies, revealed that the 55-kDa phosphorylated pro-tein was superimposable with the 55-kDa form of Shc and waspresent in a complex with Fms only after M-CSF stimulation(Fig. 2B). Immunoblots with antibodies to Grb2 also suggestedthat Grb2 was present in a complex with Fms and Shcimmunoprecipitates upon M-CSF stimulation (Fig. 2B and C).Although Fms immunoprecipitates contained Shc proteins,repeated efforts to detect Fms in immune complexes by usingantibodies against Shc produced negative results. One possibleexplanation could be that the Shc antibodies were raisedagainst the Shc SH2 domain and cannot efficiently recognizeShc that is bound through its SH2 domain to other phosphor-ylated proteins. However, we cannot exclude the possibilitythat the amount of Fms coprecipitating with Shc is below ourlevel of detection. Therefore, M-CSF stimulation of FDfmscells resulted in the tyrosine phosphorylation of Fms as well asp150 and Shc, both of which complexed with the activatedreceptor. In addition, Grb2, although not phosphorylated ontyrosine after M-CSF stimulation, also associated with tyrosinephosphorylated Fms and Shc.Grb2 has previously been shown to bind through its SH3
domains to the guanine nucleotide exchange factor Sos (6, 17,24). In the case of the EGF receptor, there are conflictingreports concerning whether Grb2 association with the EGFreceptor results in increased amounts of Sos complexing withGrb2 (3, 6, 17). To test whether Grb2 association with murineSosl (mSosl) was affected by M-CSF stimulation, we examinedthe amount of mSosl (170 kDa) coprecipitating with Grb2from FDfms cells before and after M-CSF stimulation. Theamount of mSosl present in a complex with Shc and Fms wasalso analyzed in similar coprecipitation experiments usinganti-Shc and anti-Fms antibodies. The results indicated thatonly slightly more mSosl was complexed with Grb2 fromM-CSF-treated cells (Fig. 3). However, mSosl was present inShc and or Fms complexes only after M-CSF stimulation.These results suggest that Grb2 was constitutively bound tomSosl and that M-CSF stimulation of cells did not significantly
43 -
FIG. 4. Tyrosine-phosphorylated proteins forming complexes withFms, Shc, and Grb2. (A) Lysates from quiescent (-) or M-CSF-stimulated (+) FDC-P1 cells expressing exogenous Fms were immu-noprecipitated (IP) with antibodies to Fms, Shc, or Grb2 in theabsence (-) or presence (+) of 100 mM phenyl phosphate (PHEN-PHOS). The immunoprecipitates were analyzed by SDS-PAGE andWestern blotted with antiphosphotyrosine monoclonal antibodies. Thepositions of Fms, p150, and Shc are indicated at the right side; sizes inkilodaltons are indicated at the left. Ig, immunoglobulin G.
increase this complex formation. Instead, M-CSF stimulationcaused the preformed Grb2-Sosl complexes to associate withtyrosine-phosphorylated proteins such as Fms and Shc.To investigate the nature of the complexes that are formed
in intact cells upon Fms activation, coprecipitation experi-ments with antibodies to Fms, Shc, and Grb2 were performedin the presence or absence of the phosphotyrosine analogphenyl phosphate. The complexes were separated by SDS-PAGE, and phosphotyrosine-containing proteins were de-tected with monoclonal antiphosphotyrosine antibodies (Fig.4). In the Fms immunoprecipitates, both the p150 (faint bandjust below Fms) and Shc proteins were visible in the absence ofphenyl phosphate. The two bands that migrate above Shc werenot consistently seen in other experiments. The band belowShc is the immunoglobulin heavy chain of the antibody usedfor immunoprecipitation. Although Western blot analysis ofFms immunoprecipitates indicated equivalent amounts of Fmsprotein in each lane (data not shown), the phosphotyrosinecontent on Fms in the presence of phenyl phosphate was aboutone-half of that of the control lane (no phenyl phosphate).Longer exposures of the same blot did not show Shc or p150immunoprecipitating with Fms in the presence of phenylphosphate. Shc immunoprecipitates contained only Shc andone additional phosphorylated protein at 150 kDa. The fastest-migrating band present in the M-CSF-stimulated lanes repre-sents the 45-kDa form of Shc. The faint band seen in theunstimulated lanes comigrating with the 45-kDa Shc repre-sents the immunoglobulin heavy chain of the immunoprecipi-tating antibody. The association of Shc with p150 was almostcompletely eliminated with phenyl phosphate competition.Grb2 immunoprecipitates also contained a prominent ty-rosine-phosphorylated band at 150 kDa and additional minorphosphorylated proteins at about 165, 95 to 100, 85, and 55kDa, of which the latter was identified by Western blotting asShc. The broad and faint tyrosine-phosphorylated protein (165kDa) above p150 was identified by Western blot analysis as themature form of Fms (data not shown). These results indicatethat the presence of phenyl phosphate reduced considerablythe ability of Fms, Shc, and Grb2 to form complexes with each
Grb2-+ +
- +- +- +- +
72-
< Fms00 <p150
4.
4.b
VOL. 14, 1994
72-Si.
0. to do
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 05
Feb
ruar
y 20
22 b
y 22
2.23
9.16
0.51
.

5686 LIOUBIN ET AL.
A
PROBE PY FmsM-CSF - + - + - + - +
208--
100- ''.
72-41... K...
43 -
28-
B
M-CSF - - - + + +
208 -
100 -
72-
43 -
FIG. 5. Binding of phosphorylated proteins to Shc- and Grb2-GST fusion proteins. (A) Lysates from quiescent (-) or M-CSF-stimulated (+)FDC-P1 cells expressing exogenous Fms were allowed to bind GST fusion proteins (30 ,ug/ml) containing the murine Shc SH2 domain for 2 h at4°C. The GST fusion proteins were then precipitated by the addition of glutathione-agarose beads. The washed complexes were analyzed bySDS-PAGE and Western blotted with antiphosphotyrosine (PY) or Fms antibodies as indicated. (B) Samples were treated as in panel A exceptthat GST-fusion proteins (3 to 5 p.g/ml) containing murine Grb2 SH2 or full-length Grb2 (Grb2-FL) were used. The phosphorylated proteins werevisualized by Western blot analysis using monoclonal antiphosphotyrosine antibodies. The positions of Fms, p150, and Shc are indicated at theright; sizes in kilodaltons are indicated at the left.
other, suggesting that these interactions occur between SH2domains and phosphotyrosines.To confirm these observations, we prepared GST fusion
proteins containing the SH2 domain of Shc or Grb2 andexamined their binding to potential target proteins. Lysatesprepared from M-CSF-activated FDfms cells were mixed withthe GST fusion proteins, and associated proteins were copre-cipitated with glutathione-agarose beads. The results in Fig.5A indicate that under these conditions, the Shc SH2 domainbound three tyrosine-phosphorylated proteins with apparentmolecular masses of 150, 63, and 55 kDa. Although the 55-kDaprotein appeared to migrate with the same mobility as Shc, wewere not able to verify the identity of this protein because ofgreatly elevated background upon Western blot analysis. Weattributed the background to the large amount of GST-ShcSH2 (30 ,ug/ml) required to detect any phosphoprotein bindingand the reactivity of the antibody with the fusion protein.Western blot analysis using antibodies to Fms did not detectany of the proteins bound to the Shc SH2 domain (Fig. 5A)although Fms proteins were readily detectable in the lysate. Incontrast, the Grb2 SH2- and the full-length Grb2-GST fusionproteins, at concentrations as low as 3 ,ug/ml, readily boundphosphorylated Fms protein, as well as phosphorylated Shcand to a lesser extent p150 (Fig. SB). Although p150 is notreadily visible in the exposure presented here, it was visible inlonger exposures of the same blot. These results suggest thatGrb2 can bind phosphorylated Fms, Shc, and p150 through itsSH2 domain. The interaction of the Grb2 SH2 domain withtyrosine-phosphorylated p150 is probably direct because largeamounts of phosphorylated p150 associated with Grb2 incoprecipitation experiments, as shown in Fig. 4. A summary ofthe molecular interactions observed in M-CSF-stimulated FD-fms cells or interactions that occurred in vitro when GSTfusion proteins of Shc and Grb2 were mixed with cell lysatesfrom M-CSF-activated FDfms cells is presented in Table 1.We were interested in determining whether the known
tyrosine phosphorylation sites on Fms were required for
interactions leading to subsequent phosphorylation of otherproteins, especially p150. Therefore, we expressed mutant Fmsin FDC-P1 cells in which Fms tyrosines 697, 706, and 721 in thekinase insert domain and 807 in the main kinase domain weresubstituted for phenylalanine, singly or in combinations ofthree (FmsY697-706-721F and FmsY697-706-807F). Tyrosine-phosphorylated proteins in whole cell lysates were detectedafter M-CSF stimulation. Equal amounts of total protein wereanalyzed by SDS-PAGE, and phosphotyrosine-containing pro-teins were detected with antiphosphotyrosine antibodies. Thepositions of Fms, p150, and Shc are indicated at the right inFig. 6. The protein phosphorylation pattern in the Fms mutant-infected cells was indistinguishable from that of wild-typeFms-expressing cells. Although the extent of phosphorylationon Fms varied with the individual tyrosine mutations, thedegree of phosphorylation of other target proteins was lessdependent on the individual Fms mutations. However, cellsexpressing the triple mutant FmsY697-706-807F exhibited lesstyrosine phosphorylation of p150. These results indicate that
TABLE 1. Summary of interactions"
Interaction with:Protein
Fms Shc Grb2 p150 Sos
Fms + + + +Shc - + + +Grb2 + + + +bGST-Shc SH2 - - + NDCGST-full-length Grb2 + + - + NDGST-Grb2 SH2 + + - + ND
a Interactions that occurred in vivo were detected by coprecipitation experi-ments of cell lysates, using antibodies to Fms, Shc, and Grb2 as indicated. In vitrointeractions were detected by mixing cell lysates with the indicated GST fusionproteins and precipitating the complexes with glutathione-agarose beads.
b Interaction is not M-CSF dependent.' ND, Not determined.
< Fms< p 150
<9Shc:,
MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 05
Feb
ruar
y 20
22 b
y 22
2.23
9.16
0.51
.

Fms SIGNAL TRANSDUCTION 5687
LL. LA.
N O
0 0
U. Li LA. LL LL Li-Il- (0 - Nl- N- Nl-
LA. (DNN v
0(0(0O< o 04 o 0) cn11> eD ~~~~~r- o w to
205-
0<Fmsii,,' <pl50
100-.. ...'. * '
I-_ ,<
LA. LA. L LA. U-. U.
BLOT
t.............< p150i;,, .:
BLOTpy
eL,:A --A 40-d AM .m < Sh*fl-s.; - . - i
w-me:s~a
BLOTGrb2
72 -
IP: Grb2MMP I4iP <Shc
43 -
FIG. 6. Tyrosine phosphorylation of cellular proteins stimulated byFms mutants. FDC-P1 cells expressing Fms mutants were stimulatedwith M-CSF for 1 h on ice. Aliquots of lysates, normalized for proteincontent, were electrophoresed and immunoblotted with antibodies tophosphotyrosine. FD, uninfected FDC-P1 cells; WT, cells expressingwild-type Fms protein. Sizes are indicated in kilodaltons.
B 2
BLOT::py :.
BLOTpy .......
UL LA.
11 L10LLL LL
t~t:LA. LA U.- L L LL070 0 0S 0)N.D r Nl CD 'I0
the phosphorylation of p150 on tyrosine was relatively inde-pendent of most tyrosine autophosphorylation sites on Fmswith the exception of the triple mutant FmsY697-706-807F.The effect of tyrosine-to-phenylalanine mutations of Fms on
potentially altering interactions between p150 and Shc or Grb2was next analyzed in M-CSF-stimulated FDC-P1 cells express-ing the Fms mutants. Lysates containing equal amounts ofprotein were immunoprecipitated with antibodies to Grb2(Fig. 7A) or Shc (Fig. 7B). Coprecipitating tyrosine-phosphor-ylated proteins were detected by immunoblotting with an-
tiphosphotyrosine antibodies, and Grb2 was detected by prob-ing with a monoclonal antibody to Grb2. The results indicatethat interactions between Grb2, Shc, and p150 occurred re-gardless of these mutations in Fms (Fig. 7A). The elevatedlevels of p150 coprecipitating with Grb2 in mutant FmsY721Fand FmsY697-706-721F (Fig. 7A) paralleled the amount ofFms expression (Fig. 7C). The levels of p150 precipitating withGrb2 from the M-CSF-activated triple mutant FmsY697-706-807F was the most severely decreased but was not completelyabolished (Fig. 7A). Similar results were obtained from the Shcimmunoprecipitation experiments (Fig. 7B). Our results showthat tyrosine phosphorylation of p150 and its association withGrb2 and Shc was not abolished by any of the four tyrosineautophosphorylation site mutations; however, these interac-tions were somewhat decreased in cells expressing the triplemutant FmsY697-706-807F.The tyrosine phosphorylation of a 150-kDa protein and its
association with Shc and Grb2 have not been described in thesignal transduction mechanisms of the growth factor receptorsfor EGF and PDGF. We also have detected this p150 inmyeloid 32D cells expressing exogenous murine Fms afterM-CSF stimulation (data not shown), and therefore we inves-tigated the possibility that the involvement of p150 was specificfor signal transduction in myeloid cells. Because other growthsignaling mechanisms have been investigated primarily infibroblasts, we looked for the association of a tyrosine-phos-phorylated protein of 150 kDa with Grb2 in lysates preparedfrom M-CSF-stimulated Rat2 cells expressing murine Fms
IP: ShcLL LLs
LA LL LL LL LA ULCNI N NI
C cr CN1 0rC CD( N1 N co MD
BLOTFms l< Fms
FIG. 7. Complex formation between p150 and Grb2 or Shc in cellsexpressing Fms mutants. FDC-P1 cells expressing Fms mutants werestimulated with M-CSF for 1 h on ice. Lysates were equalized forprotein content and immunoprecipitated (IP) with antibodies to Grb2(A) or Shc (B), and the precipitates were analyzed by SDS-PAGE. Theblots were immunoblotted as indicated at the left with antiphosphoty-rosine (PY) or Grb2 antibodies. The positions of p150, Shc, and Grb2are indicated at the right. (C) Whole cell lysates, equalized for proteincontent, were immunoblotted with antibodies to Fms. The upper bandrepresents the mature cell surface-expressed 165-kDa form of Fms.FD, uninfected FDC-P1 cells; WT, cells expressing wild-type Fmsprotein.
from a retroviral vector (Rat2fms cells). Figure 8 demonstratesthat immunoprecipitates of Grb2 from M-CSF-activatedRat2fms cells contained a tyrosine-phosphorylated protein atabout 160 kDa, a less prominent band at 100 kDa, and a56-kDa protein which is probably Shc. Although the 56-kDaband is not visible in the exposure presented here because ofthe intensity of the antibody heavy-chain band, it was visible inlighter exposures. These proteins were not detected in immu-noprecipitates from quiescent Rat2fms cells, which suggeststhat their phosphorylation and association with Grb2 wereM-CSF specific. The 150-kDa tyrosine-phosphorylated proteinthat was easily seen coprecipitating with Grb2 from FDfms
< Grb2
<pl50
< Shc
VOL. 14, 1994
N...... ... nt o,
BLOT A iiiliw'i < Grb2Grb2
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 05
Feb
ruar
y 20
22 b
y 22
2.23
9.16
0.51
.

5688 LIOUBIN ET AL.
Rat2 FD
IP C,'M-CSF - + - + + +
205-
100-
72-
43
FIG. 8. Analysis of p150 complexed with Shc or Grb2 in Fms-expressing fibroblasts. Cell lysates from quiescent (-) or M-CSF-stimulated (+) Rat2 cells expressing exogenous murine Fms wereimmunoprecipitated (IP) with polyclonal antibodies to Shc, Grb2, oragarose-immobilized Fms antibodies as indicated. The samples were
analyzed by SDS-PAGE and Western blotted with monoclonal an-
tiphosphotyrosine antibodies. Results for tyrosine-phosphorylated pro-teins coprecipitating with Grb2 from M-CSF-stimulated FDfms (FD)cells are also presented as a reference. The locations of Fms and p150is indicated at the right. Sizes are indicated in kilodaltons.
lysates (Fig. 8, rightmost lane) was not visible in Grb2 immu-noprecipitates from Rat2fms cells, even on very long expo-sures. The 160-kDa tyrosine-phosphorylated protein that asso-
ciated with Grb2 displayed the same mobility on SDS-PAGEas the mature Fms protein immunoprecipitated from Rat2fmscells after M-CSF stimulation (Fig. 8). Furthermore, this160-kDa protein was immunoreactive with Fms antibodies inWestern blot analysis (data not shown). In contrast to theresults obtained from FDfms cells (Fig. 4), Shc immunopre-cipitates from Rat2fms cells failed to coprecipitate any ty-rosine-phosphorylated proteins other than Shc from eitherquiescent or M-CSF-stimulated cells. The phosphorylated Shcprotein can be seen in the M-CSF-stimulated lane just abovethe intensely stained immunoglobulin heavy-chain band. Theseresults suggest that the 150-kDa tyrosine-phosphorylated pro-tein participated in Fms-induced signal transduction of my-eloid cells but not in fibroblasts. Consistent with this possibility,we have been unable to detect a similar Grb2-binding proteinin BALB/c-fms cells and have not observed p150 in phospho-tyrosine blots of total cell lysates prepared from either BALB/c-fms or Rat2fms cells (data not shown), whereas p150 was
readily detectable in total cell lysates of stimulated FDfms cells(Fig. 1).
DISCUSSION
Upon M-CSF stimulation of FDC-P1 cells expressing Fms, a
variety of hyperphosphorylated proteins are observed withina very short time. At 37°C, maximum phosphorylation isachieved within 1 min after stimulation, and the proteins aredephosphorylated (or degraded) to almost basal levels within10 min. A 63-kDa protein appeared to exhibit maximumphosphorylation within 30 s, and phosphotyrosine was rapidlyreduced to almost basal levels within 1 min. Other proteins of100 and 150 kDa exhibited kinetics similar to those of p63, butwith slightly slower dephosphorylation. It is not known whetherany of these proteins are kinases or phosphatases or are simply
more susceptible to dephosphorylation or degradation by otherphosphatases or proteases. Clearly, the detectability of Fms islost because of rapid degradation following endocytosis; how-ever, the half-life for degradation in FDfms cells is about 5 min(5), and therefore dephosphorylation occurs more rapidly thandegradation. It seems likely that the other proteins alsoundergo cycles of phosphorylation and dephosphorylation.Also, most of the same proteins appear to be phosphorylatedat 4°C as at 37°C. The lower temperature is not permissive forinternalization, suggesting that internalization of Fms is notrequired for the initiation of signal transduction.
Activation of tyrosine kinase receptors, such as the EGFreceptor (25, 40), insulin receptor (15, 27), and PDGF receptor(27), results in Shc phosphorylation and Ras activation. Rasactivation to its GTP-bound form is facilitated by the nucle-otide exchange factor Sos (3, 11, 17, 18, 24, 38, 39). Grb2, a23-kDa protein composed entirely of one SH2 domain flankedby two SH3 domains, binds through both of its SH3 domains toproline-rich sequences of Sos (6, 17, 24) and through its SH2domain to other tyrosine-phosphorylated proteins such asactivated EGF receptors (3, 18) or tyrosine kinase substratessuch as Shc (21, 25, 38, 39) and insulin receptor substrate 1(IRS1) (1, 44, 45). Interactions of Grb2 with any of thesetyrosine-phosphorylated proteins result in the translocation ofSos to the plasma membrane, where it stimulates Ras toexchange GDP for GTP (3, 18). In this report, we showed thatupon M-CSF stimulation of FDfms cells, Shc proteins arephosphorylated on tyrosine and participate in a complexformation with Fms, Grb2, and p150 by directly interactingwith Grb2 and p150.
In contrast to fibroblasts, which express all three forms ofShc, myeloid cells express only the 45- and 55-kDa forms.Although both of these proteins are tyrosine phosphorylatedafter M-CSF stimulation of FDfms cells, only the 55-kDaphosphorylated Shc is readily detected in cell lysates becauseof its much higher abundance over the 45-kDa form. The66-kDa Shc protein was not detectable in these cells either inwhole cells lysates or in immunoprecipitates with Shc antibod-ies.Although the presence of the 55-kDa Shc in Fms immuno-
precipitates was dependent on M-CSF stimulation, its associ-ation with the activated receptor is probably indirect (or atleast not via the Shc SH2 domain) since the Shc SH2 domainfusion protein failed to bind phosphorylated Fms. Instead,GST-Shc SH2 bound a 150-kDa tyrosine-phosphorylated pro-tein which was also present in Fms immunoprecipitations. It ispossible that Shc is present in a Fms complex because of itsassociation with p150. Although we showed in this study thatthe Shc SH2 domain directly binds a tyrosine phosphorylationsite of p150, we do not yet know the nature of the associationbetween p150 and Fms. It is unlikely that Grb2 serves as anadaptor for Shc-Fms association since Grb2 has only one SH2domain that can bind only one phosphorylation site on eitherFms or Shc but not both at the same time, and there is noevidence that Grb2 SH3 domains bind Shc or Fms.Grb2 and Sosl were associated in unstimulated FDfms cells,
and M-CSF stimulation did not greatly alter the amount ofSosl bound to Grb2. In contrast, Sosl was found in a complexwith Fms and/or Shc only after M-CSF stimulation, and theamount of Sosl present in such complexes paralleled theamount of Grb2. GST-Grb2 SH2 or full-length Grb2 fusionproteins readily bound phosphorylated Fms and Shc proteins,indicating that the Grb2 SH2 domain interacts directly with atyrosine phosphorylation site on Fms and Shc. Shc has only onetyrosine phosphorylation site in the collagen homology domain(25), and therefore one phosphorylated Shc molecule can bind
MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 05
Feb
ruar
y 20
22 b
y 22
2.23
9.16
0.51
.

Fms SIGNAL TRANSDUCTION 5689
only one Grb2 molecule through SH2 domain interactions.Fms, on the other hand, possesses at least four tyrosineautophosphorylation sites (positions 697, 706, 721, and 807 formurine Fms) (30, 37, 48). A tyrosine-to-phenylalanine muta-tion of murine Fms at position 697 abolishes the binding ofGrb2 to Fms (49) as well as the presence of Sosl in Fmscomplexes (unpublished data). These results suggest that Grb2exists in a constitutive complex with Sosl and that upon Fmsactivation, preformed Grb2-Sosl complexes may bind throughthe Grb2SH2 domain to phosphorylated Fms as well as Shcand p150. In hematopoietic cells, overexpression of activatedRas results in both proliferation and differentiation (14). Inaddition, expression of dominant negative mutants of Ras infibroblasts blocks Fms transformation (47). Consistent withthese observations, and by analogy with other growth factorreceptor systems (1, 3, 24, 38, 42-44), our work suggests thatSosl is probably the Ras activator in M-CSF-stimulated Fms-expressing hematopoietic cells.Grb2 immunoprecipitates from M-CSF-activated FDfms
cells contained a prominent tyrosine-phosphorylated proteinof 150 kDa which exhibited the same SDS-PAGE mobility asthe-150 kDa protein coprecipitating with Shc (Fig. 4) via itsSH2 domain (Fig. 5). Immunoprecipitation experiments (Fig.4) indicated that considerable amounts of p150 precipitatedwith both Grb2 and Shc. In addition, Grb2 immunoprecipitatesalso contained small amounts of phosphorylated Fms and Shc.The p150 present in the Grb2 immunoprecipitates most likelyis not due to its association with Fms since Fms is present inlow amounts, and only minor amounts of p150 coimmunopre-cipitated with Fms. Comparison of the ratios of phosphory-lated Shc to p150 in the Shc and Grb2 immunoprecipitatesdemonstrates that the presence of Shc-p150 complexes couldnot account for all the p150 coprecipitating with Grb2. Fur-thermore, GST-Grb2 SH2 domain fusion proteins also copre-cipitated p150, although it appeared to bind Fms preferen-tially. Although we cannot entirely exclude the possibility thatp150 coprecipitates with GST-Grb2 SH2 because of its associ-ation with Shc, the possibility that p150 is present in a complexwith Grb2-SH2 because of its association with Fms can beexcluded since GST-Grb2 SH2 precipitates p150 from lysatesof mutant Fms that itself cannot bind Grb2 (unpublished data).When cell lysates were separated by SDS-PAGE, transferredonto nitrocellulose membranes, and then probed with the GSTfusion proteins of Shc SH2 or Grb2 SH2 domains, onlyphosphorylated Shc bound to the GST-Grb2 SH2 fusion withhigh affinity (data not shown). These results, taken togetherwith the fact that Shc also binds through its SH2 domain top150, suggest that p150 possesses at least two putative tyrosinephosphorylation sites, one of which binds Shc and another ofwhich binds Grb2. p150 may form two distinct complexes, onewith Shc and another with Grb2, or alternatively, all threeproteins may be in the same complex. Our results do notclearly distinguish between these two possibilities.
Analysis of p150 phosphorylation in cells expressing Fmstyrosine autophosphorylation site mutants indicated that thetyrosine phosphorylation of p150 was not entirely dependenton any of these sites. The triple mutant FmsY697-706-807Fstimulated less tyrosine phosphorylation of p150 than wild-typeFms, but this may be due to the slightly reduced kinase activitythat we have observed with Fms triple site mutants (3a). Thelack of effect of the FmsY697F mutation also suggests thatphosphorylation of p150 is not facilitated by Grb2 acting as anadaptor protein linking p150 to Fms at the Y697 site (49).Also, in contrast to the FmsY697F mutant expressed infibroblasts (49), the same mutant expressed in FDC-P1 cellsdid not impair M-CSF-dependent growth stimulation (la).
Fms
I,/ /
IIi
p\ 15
'\\\""5OIshv4Wko)FIG. 9. Schematic representation of possible interactions of signal
transduction proteins in M-CSF-stimulated myeloid cells. The diagramillustrates some of the interactions occurring among the signal trans-duction proteins identified in this study of M-CSF-stimulated FDfmscells. M-CSF is shown bound to the extracellular domain of thehomodimerized Fms at the plasma membrane. Phosphorylated ty-rosine residues are represented by a Y with an attached small graycircle. The single SH2 domain of both Shc and Grb2 is shown boundto phosphotyrosine residues. The two SH3 domains of Grb2 contactSos at proline-rich sequences. Not all possible complexes of theseproteins are shown, and others are discussed in the text.
This finding suggests that the pl50-Grb2 complex in FDC-P1cells could be substituting for the Fms-Grb2 signaling observedin fibroblasts. p150 may be phosphorylated by Fms because itassociates with Fms through an as yet unknown tyrosinephosphorylation site or a site not dependent on phosphoryla-tion. Perhaps more likely, however, is the possibility that p150is phosphorylated by a kinase other than Fms.p150 has not been described previously for any other
tyrosine kinase receptor signaling pathway such as EGF recep-tor, insulin receptor, PDGF receptor, or Fms, mainly studiedin fibroblasts. In this study, we showed that p150 is phosphor-ylated in response to M-CSF and participates in Fms signaltransduction in the myeloid cell line FDC-P1. We have alsoobserved this 150-kDa tyrosine-phosphorylated protein afterM-CSF stimulation of another myeloid cell line, 32D, and twoerythroid cell lines all expressing exogenous Fms (17a). Incontrast, neither the 150-kDa protein nor a similar proteinbinding to Shc and/or Grb2 was detectable in M-CSF-stimu-lated fibroblast cell lines (Rat2 and BALB/c) expressing exog-enous Fms. These observations are consistent with the hypoth-esis that tyrosine phosphorylation of p150 is hematopoieticspecific and in these cells might participate in a Fms signalingpathway that is absent in fibroblasts.The phosphorylation of the 150-kDa protein also appears to
be receptor specific. Tyrosine phosphorylation of p150 was notobserved after insulin stimulation of 32D cells that expressexogenous insulin receptors, and these cells did not respond toinsulin unless IRS1 was also introduced together with theinsulin receptor (51). In contrast, M-CSF stimulation of32Dfms cells readily resulted in the tyrosine phosphorylationof the 150-kDa protein, and the cells respond with prolifera-tion in the absence of IL-3 and maturation toward the macro-phage differentiation pathway (la).We have already demonstrated that the 150-kDa protein is
not identical to the mSosl protein because of its size andtyrosine phosphorylation. It is probably also not equivalent tothe mSos2 protein (2) because, despite their similar sizes, the
VOL. 14, 1994
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 05
Feb
ruar
y 20
22 b
y 22
2.23
9.16
0.51
.

5690 LIOUBIN ET AL.
mSos2 protein is expressed in fibroblasts (as is Sosl) yet the150-kDa protein does not participate in Fms signaling in thesecells. Likewise, the 150-kDa protein differs from other putativesignal transducers such as IRS1, Arg, Abl, and 4PS (16, 26, 36,51) with respect to size, cell type distribution, and/or immuno-logical reactivity. A 145-kDa tyrosine-phosphorylated proteinhas been reported to participate in erythropoietin receptor
signaling in erythroid cells (8), and we are currently investigat-ing potential relationships to p150. Thus, the 150-kDa proteintyrosine phosphorylated after M-CSF stimulation appears torepresent a unique component of the Fms signal transductionmechanism present in the hematopoietic lineage.A schematic representation of the molecular interactions
observed in the myeloid progenitor cell line FDC-P1 express-
ing exogenous murine Fms after M-CSF stimulation is pre-
sented in Fig. 9. Upon M-CSF stimulation of FDfms cells, Fmsas well as Shc and a 150-kDa protein are tyrosine phosphory-lated. This modification initiates the complex formation ofpreformed Grb2-Sosl heterodimers with Fms, Shc, and p150.Shc also associates through its SH2 domain with a phosphor-ylated tyrosine of p150. Although Fms was also shown to existin a complex with Shc and p150, their mechanism of interac-tion is not yet clear. For simplicity, the schematic representa-tion shows direct interactions of Shc and Grb2 on differentp150 molecules. The possibility that Shc and Grb2 bothassociate with different phosphorylated tyrosines on the same
p150 molecule also exists and should not be excluded. Growthstimulation in fibroblasts may utilize the Fms-Grb2-Sosl com-
plex for activation of Ras. Tyrosine phosphorylation of Shcmay also participate in fibroblast growth signaling. In myeloidcells, however, additional signaling mechanisms may occur
through tyrosine phosphorylation of p150.
ACKNOWLEDGMENTS
We thank Tony Pawson for generously providing polyclonal anti-bodies to Shc, Andreas Batzer for providing polyclonal antibodies to
Grb2, Tina Loucks for preparation of the manuscript, and Allen Chenand Paul Algate for critical evaluation of the manuscript.
This work was supported by PHS grants CA20551 and CA40987from the National Cancer Institute. M.N.L. was supported by PHSgrant NRSA T32 GM07270 from NIGMS.
REFERENCES1. Baltensperger, K., L. M. Kozma, A. D. Cherniack, J. K. Klarlund,
A. Chawla, U. Banerjee, and M. P. Czech. 1993. Binding of the Rasactivator son of sevenless to insulin receptor substrate-1 signalingcomplexes. Science 260:1950-1952.
la.Bourette, R. P., et al. Unpublished data.2. Bowtell, D., P. Fu, M. Simon, and P. Senior. 1992. Identification of
murine homologues of the Drosophila Son of Sevenless gene:
potential activators of ras. Proc. Natl. Acad. Sci. USA 89:6511-6515.
3. Buday, L. 1993. Epidermal growth factor regulates p21ras throughthe formation of a complex of receptor, Grb2 adapter protein, andSOS nucleotide exchange factor. Cell 73:611-620.
3a.Carlberg, K., et al. Unpublished data.4. Carlberg, K., and L. R. Rohrschneider. 1994. The effect of
activating mutations on dimerization, tyrosine phosphorylation,and internalization of the macrophage colony stimulating factorreceptor. Mol. Biol. Cell 5:81-95.
5. Carlberg, K., P. Tapley, C. Haystead, and L. R. Rohrschneider.1991. Role of kinase activity and the kinase insert region inligand-induced internalization and degradation of the c-fins pro-
tein. EMBO J. 10:877-883.6. Chardin, P., J. H. Camonis, N. W. Gale, L. Van Aelst, J. Schles-
singer, M. H. Wigler, and D. Bar-Sagi. 1993. Human Sos 1: a
guanine nucleotide exchange factor for Ras that binds to GRB2.Science 260:1338-1343.
7. Claesson-Welch, L., A. Eriksson, B. Westermark, and C. H.
Heldin. 1989. cDNA cloning and expression of the human A-typeplatelet-derived growth factor (PDGF) receptor establishes struc-tural similarity to the B-type PDGF receptor. Proc. Natl. Acad.Sci. USA 86:4917-4921.
8. Damen, J. E., L. Liu, R. L. Cutler, and G. Krystal. 1993.Erythropoietin stimulates the tyrosine phosphorylation of Shc andits association with Grb2 and a 145-Kd tyrosine phosphorylatedprotein. Blood 82:2296-2303.
9. Downing, J. R., B. L. Margolis, A. Zilberstein, R. A. Ashmun, A.Ullrich, C. J. Sherr, and J. Schlessinger. 1989. PhospholipaseC-gamma, a substrate for PDGF receptor kinase, is not phosphor-ylated on tyrosine during the mitogenic response to CSF-1. EMBOJ. 8:3345-3350.
10. Downing, J. R., M. F. Roussel, and C. J. Sherr. 1989. Ligand andprotein kinase C downmodulate the colony-stimulating factor 1receptor by independent mechanisms. Mol. Cell. Biol. 9:2890-2896.
11. Egan, S. E., B. W. Giddings, M. W. Brooks, L. Buday, A. M.Sizeland, and R. A. Weinberg. 1993. Association of SOS Rasexchange protein with Grb2 is implicated in tyrosine kinase signaltransduction and transformation. Nature (London) 363:45-51.
12. Gale, N. W., S. Kaplan, E. J. Lowenstein, J. Schlessinger, and D.Bar-Sagi. 1993. Grb2 mediates the EGF-dependent activation ofguanine nucleotide exchange on Ras. Nature (London) 363:88-92.
13. Heard, J. M., M. F. Roussel, C. W. Rettenmier, and C. J. Sherr.1987. Multilineage hematopoietic disorders induced by transplan-tation of bone marrow cells expressing the v-fms oncogene. Cell51:663-673.
14. Hibi, S., J. Lohler, J. Friel, C. Stocking, and W. Ostertag. 1993.Induction of monocytic differentiation and tumorigenicity byv-Ha-ras in differentiation-arrested hematopoietic cells. Blood81:1841-1848.
15. Kovacina, K. S., and R. A. Roth. 1993. Identification of SHC as asubstrate of the insulin receptor kinase distinct from the GAP-associated 62 kDa tyrosine phosphoprotein. Biochem. Biophys.Res. Commun. 192:1303-1311.
16. Kruh, G. D., R. Perego, T. Miki, and S. A. Aaronson. 1990. Thecomplete coding sequence of arg defines the Abelson subfamily ofcytoplasmic tyrosine kinases. Proc. Natl. Acad. Sci. USA 87:5802-5806.
17. Li, N., A. Batzer, R. Daly, V. Yajnik, E. Skolnik, P. Chardin, D.Bar-Sagi, B. Margolis, and J. Schlessinger. 1993. Guanine-nucle-otide-releasing factor hSOS1 binds to Grb2 and links receptortyrosine kinases to Ras signalling. Nature (London) 363:85-88.
17/a.Lioubin, M. N., and R. P. Bourette. Unpublished data.18. Lowenstein, E. J., R. J. Daly, A. G. Batzer, W. Li, B. Margolis, R.
Lammers, A. Ullrich, E. Y. Skolnik, D. Bar-Sagi, and J. Schles-singer. 1992. The SH2 and SH3 domain-containing protein GRB2links receptor tyrosine kinases to ras signaling. Cell 70:431-442.
19. Mathews, W., C. T. Jordan, G. W. Wiegand, D. Pardoll, and I. R.Lemischka. 1991. A receptor tyrosine kinase specific to hemato-poietic stem and progenitor cell-enriched populations. Cell 65:1143-1152.
20. Matsui, T., M. Heidaran, T. Miki, N. Popescu, W. LaRochelle, M.Kraus, S. Pierce, and S. Aaronson. 1989. Isolation of a novelreceptor cDNA establishes the existence of two PDGF receptorgenes. Science 243:800-804.
21. McGlade, J., A. Cheng, G. Pelicci, P. G. Pelicci, and T. Pawson.1992. Shc proteins are phosphorylated and regulated by the v-srcand v-fps protein-tyrosine-kinases. Proc. Natl. Acad. Sci. USA89:8869-8873.
22. Moodie, S. A., B. M. Willumsen, M. J. Weber, and A. Wolfman.1993. Complexes of Ras-GTP and Raf-1 and mitogen-activatedprotein kinase kinase. Science 260:1658-1661.
23. Mori, S., L. Ronnstrand, K. Yokote, A. Engstr8m, S. A. Court-neidge, L. Claesson-Welsh, and C.-H. Heldin. 1993. Identificationof two juxtamembrane autophosphorylation sites in the PDGF3-receptor; involvement in the interaction with Src family tyrosine
kinases. EMBO J. 12:2257-2264.24. Olivier, J. P., T. Raabe, M. Henkemeyer, B. Dickson, G. Mbamalu,
B. Margolis, J. Schlessinger, E. Hafen, and T. Pawson. 1993. ADrosophila SH2-SH3 adaptor protein implicated in coupling thesevenless tyrosine kinase to an activator of Ras guanine nucleotide
MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 05
Feb
ruar
y 20
22 b
y 22
2.23
9.16
0.51
.

Fms SIGNAL TRANSDUCTION 5691
exchange, Sos. Cell 73:179-191.25. Pelicci, G., L. Lanfrancone, F. Grignani, J. McGlade, F. Cavallo,
G. Forni, I. Nicoletti, F. Grignani, T. Pawson, and P. G. Pelicci.1992. A novel transforming protein (SHC) with an SH2 domain isimplicated in mitogenic signal transduction. Cell 70:93-104.
26. Perego, R., D. Ron, and G. D. Kruh. 1991. Arg encodes a widelyexpressed 145 kDa protein-tyrosine kinase. Oncogene 6:1899-1902.
27. Pronk, G. J., J. McGlade, G. Pelicci, T. Pawson, and J. L. Bos.1993. Insulin-induced phosphorylation of the 46- and 52-kDa Shcproteins. J. Biol. Chem. 268:5748-5753.
28. Qiu, F., P. Ray, K. Brown, P. E. Barker, S. Jhawer, R. II. Ruddle,and P. Besmer. 1988. Primary structure of c-kit: relationship withthe CSF-1/PDGF receptor kinase family: oncogenic activation ofv-kit involves deletion of extracellular domain and C-terminus.EMBO J. 7:1003-1011.
29. Reedijk, M., X. Liu, and T. Pawson. 1990. Interactions of phos-phatidylinositol kinase, GTPase-activating protein(GAP), andGAP-associated proteins with the colony-stimulating factor 1receptor. Mol. Cell. Biol. 10:5601-5608.
30. ReediJk, M., X. Liu, P. van der Geer, K. Letwin, M. D. Waterfield,T. Hunter, and T. Pawson. 1992. Tyr721 regulates specific bindingof the CSF-1 receptor kinase insert to PI 3'-kinase SH2 domains:a model for SH2-mediated receptor-target interactions. EMBO J.11:1365-1372.
31. Rohrschneider, L. R. The macrophage-colony stimulating factor(M-CSF) receptor. In J. Sambrook and J. Tooze (ed.), Guidebookto cytokines and their receptors, in press. Oxford University Press,New York.
32. Rohrschneider, L. R., and D. Metcalf. 1989. Induction of macro-phage colony stimulating factor-dependent growth and differenti-ation after introduction of the murine c-fins gene into FDC-P1cells. Mol. Cell. Biol. 9:5081-5092.
33. Rohrschneider, L. R., V. M. Rothwell, and N. A. Nicola. 1989.Transformation of murine fibroblasts by a retrovirus encoding themurine c-fms proto-oncogene. Oncogene 4:1015-1022.
34. Rohrschneider, L. R., and J. F. Woolford. 1992. Structural andfunctional comparison of viral and cellular fins. Semin. Virol.2:385-395.
35. Rosnet, O., S. Marchetto, 0. deLapeyriere, and D. Birnbaum.1991. Murine Elt3, a gene encoding a novel tyrosine kinasereceptor of the PDGFR/CSF1R family. Oncogene 6:1641-1650.
36. Rothenberg, P. L., W. S. Lane, A. Karasik, J. Backer, M. White,and R. C. Kahn. 1991. Purification and partial sequence analysis ofppl85, the major cellular substrate of the insulin receptor tyrosinekinase. J. Biol. Chem. 266:8302-8311.
37. Roussel, M. F., S. A. Shurtleff, J. R. Downing, and C. J. Sherr.1990. A point mutation at tyrosine 809 in the human colony-stimulating factor 1 receptor impairs mitogenesis without abrogat-ing tyrosine kinase activity, association with phosphatidylinositol3-kinase, or induction of fos and junB genes. Proc. Natl. Acad. Sci.USA 87:6738-6742.
38. Rozakis-Adcock, M., R. Fernley, J. Wade, T. Pawson, and D.Bowtell. 1993. The SH2 and SH3 domains of mammalian Grb2couple the EGF receptor to the Ras activator mSOSl. Nature(London) 363:83-85.
39. Rozakis-Adcock, M., J. McGlade, G. Mbamalu, G. Pelicci, R. Daly,W. Li, A. Batzer, S. Thomas, J. Brugge, G. Pelicci, J. Schlessinger,and T. Pawson. 1992. Association of the Shc and Grb2/Sem5SH2-containing proteins is implicated in activation of the Ras
pathway by tyrosine kinases. Nature (London) 360:689-692.40. Ruff-Jamison, S., J. McGlade, T. Pawson, K. Chen, and S. Cohen.
1993. Epidermal growth factor stimulates the tyrosine phosphor-ylation of SHC in the mouse. J. Biol. Chem. 268:7610-7612.
41. Segatto, O., G. Pelicci, S. Giuli, G. Digiesi, P. P. Di Fiore, J.McGlade, T. Pawson, and P. G. Pelicci. 1993. Shc products aresubstrates of erbB-2 kinase. Oncogene 8:2105-2112.
42. Simon, M. A., D. D. L. Bowtell, G. S. Dodson, T. R. Laverty, andG. M. Rubin. 1991. Rasl and a putative guanine nucleotideexchange factor perform crucial steps in signaling by the sevenlessprotein tyrosine kinase. Cell 67:701-716.
43. Simon, M. A., G. S. Dodson, and G. M. Rubin. 1993. AnSH3-SH2-SH3 protein is required for p2lRasl activation andbinds to sevenless and SOS proteins in vitro. Cell 73:169-177.
44. Skolnik, E. Y., A. Batzer, N. Li, C.-H. Lee, L. E. Lowenstein, M.Mohammadi, B. Margolis, and J. Schlessinger. 1993. The functionof GRB2 in linking the insulin receptor to Ras signaling pathways.Science 260:1953-1955.
45. Skolnik, E. Y., C.-H. Lee, A. Batzer, L. M. Vincentini, M. Zhou, R.Daly, M. J. Myers, Jr., J. M. Backer, A. Ullrich, M. F. White, andJ. Schlessinger. 1993. The SH2/SH3 domain-containing proteinGRB2 interacts with tyrosine-phosphorylated IRS-1 and Shc:implications for insulin control of ras signaling. EMBO J. 12:1929-1936.
46. Smith, D. B., and K. S. Johnson. 1988. Single-step purification ofpolypeptides expressed in Escherichia coli as fusions with glutathi-one S-transferase. Gene 67:31-40.
47. Stacey, D. W., M. Roudebush, R. Day, S. D. Mosser, J. B. Gibbs,and L. A. Feig. 1991. Dominant inhibitory Ras mutants demon-strate the requirement for Ras activity in the action of tyrosinekinase oncogenes. Oncogene 6:2297-2304.
48. Tapley, P., A. Kazlauskas, J. A. Cooper, and L. R. Rohrschneider.1990. Macrophage colony-stimulating factor-induced tyrosinephosphorylation of c-Fms proteins expressed in FDC-PI andBALB/c-3T3 cells. Mol. Cell. Biol. 10:2527-2537.
49. van der Geer, P., and T. Hunter. 1993. Mutation of Tyr697, aGRB2-binding site, and Tyr721, a PI 3-kinase binding site, abro-gates signal transduction by the murine CSF-1 receptor expressedin Rat-2 fibroblasts. EMBO J. 12:5161-5172.
50. Vojtek, A. B., S. M. Hollenberg, and J. A. Cooper. 1993. Mamma-lian Ras interacts directly with the serine/threonine kinase Raf.Cell 74:205-214.
51. Wang, L.-M., M. G. Myers, Jr., X.-J. Sun, S. A. Aaronson, M.White, and J. Pierce. 1993. IRS-1: essential for insulin- andIL-4-stimulated mitogenesis in hematopoietic cells. Science 261:1591-1594.
52. Wang, Z., G. Myles, C. S. Brandt, M. N. Lioubin, and L. R.Rohrschneider. 1993. Identification of the ligand-binding regionsin the macrophage colony-stimulating factor receptor extracellulardomain. Mol. Cell. Biol. 13:5348-5359.
53. Warne, P. H., and P. R. Viciana. 1993. Direct interaction of Rasand the amino-terminal region of Raf-1 in vitro. Nature (London)364:352-355.
54. Yarden, Y., J. A. Escobedo, W. J. Kuang, T. L. Yang-Feng, T. 0.Daniel, P. M. Tremble, E. Y. Chen, M. E. Ando, R. N. Harkins, U.Francke, V. A. Fried, A. Ullrich, and L. T. Williams. 1986.Structure of the receptor for platelet-derived growth factor helpsdefine a family of closely related growth factor receptors. Nature(London) 323:226-232.
VOL. 14, 1994
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 05
Feb
ruar
y 20
22 b
y 22
2.23
9.16
0.51
.