social neuroscience erp time course and brain areas of
TRANSCRIPT

PLEASE SCROLL DOWN FOR ARTICLE
This article was downloaded by: [Van Overwalle, Frank][Overwalle, Frank Van]On: 19 January 2011Access details: Access Details: [subscription number 909176132]Publisher Psychology PressInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Social NeurosciencePublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t741771143
ERP time course and brain areas of spontaneous and intentional goalinferencesLaurens Van der Cruyssena; Marijke Van Duynslaegera; Aisha Cortoosa; Frank Van Overwallea
a Vrije Universiteit, Brussels, Belgium
First published on: 19 December 2008
To cite this Article Van der Cruyssen, Laurens , Van Duynslaeger, Marijke , Cortoos, Aisha and Van Overwalle,Frank(2009) 'ERP time course and brain areas of spontaneous and intentional goal inferences', Social Neuroscience, 4: 2,165 — 184, First published on: 19 December 2008 (iFirst)To link to this Article: DOI: 10.1080/17470910802253836URL: http://dx.doi.org/10.1080/17470910802253836
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article may be used for research, teaching and private study purposes. Any substantial orsystematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply ordistribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae and drug dosesshould be independently verified with primary sources. The publisher shall not be liable for any loss,actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directlyor indirectly in connection with or arising out of the use of this material.

ERP time course and brain areas of spontaneous andintentional goal inferences
Laurens Van der Cruyssen, Marijke Van Duynslaeger, Aisha Cortoos, and FrankVan Overwalle
Vrije Universiteit, Brussels, Belgium
This study measured event-related potentials during spontaneous and intentional goal inferences.Participants read sentences describing the behavior of a target person from which a strong goal orintention could be inferred. The last word of each sentence determined the consistency with the goalinduced during preceding sentences. In comparison with behaviors that were consistent with the impliedgoal, a stronger P200 waveform was obtained when the behaviors were irrelevant with that goal or didnot contain goal-directed behavior at all, and this P200 showed considerable parallels betweenspontaneous and intentional inferences. This indicates that goals were inferred rapidly and automaticallywhile reading the behaviors, irrespective of the participants’ spontaneous or intentional instructions. Inline with this, source localization (LORETA) of the event-related potentials shows predominantlyactivation in the temporoparietal junction (TPJ) during and immediately after goal detection (225�300 ms). Before and after this time interval, however, activation is stronger at the TPJ duringspontaneous processing, and stronger at the medial prefrontal cortex (mPFC) during intentionalprocessing. Memory measures taken after the presentation of the stimulus materials support theoccurrence of goal inferences and show significant correlations with the neural components, indicatingthat these components are valid neural indices of spontaneous and intentional goal inferences. Theresults are highly similar to previous ERP research on trait inferences that revealed a similar division ofbrain activation for spontaneous (TPJ) and intentional (mPFC) processes, but appearing later at about600 ms, pointing to similar brain areas recruited for social inferences, but at different timings for differentinference types.
INTRODUCTION
Detecting the intentions of other people is a
fundamental capacity to maneuver in our social
world. Upon meeting other people, we must
immediately infer whether their intentions aregood or bad, so as to approach or avoid them,
because our immediate well-being and survival
may depend on this decision. Is someone making
a fist to hit me or to conceal a present for me? Isan unfamiliar person carrying a gun (to shoot me)
or unarmed? This research explores several
questions on the identification of goals. What
are the cognitive processes driving the detectionof goals and intentions of other people? Do theyreflect rapid automatic reactions or do theyrequire slower reflective deliberations? How fastcan we make goal interpretations? To answerthese questions, we focus on the processingdifferences between spontaneous goal inferences(SGI) versus intentional goal inferences (IGI),and how electrophysiological measures can un-cover the neural components associated with eachprocessing mode. Research into neural responsescan increase our understanding of the timingand localization of the neural processing of
# 2009 Psychology Press, an imprint of the Taylor & Francis Group, an Informa business
Correspondence should be addressed to: Frank Van Overwalle, Department of Psychology, Vrije Universiteit Brussel, Pleinlaan
2, B-1050 Brussels, Belgium. E-mail: [email protected]
This research was supported by an FWO Grant of the Research Foundation*Flanders to Frank Van Overwalle.
SOCIAL NEUROSCIENCE, 2009, 4 (2), 165�184
www.psypress.com/socialneuroscience DOI:10.1080/17470910802253836
Downloaded By: [Van Overwalle, Frank][Overwalle, Frank Van] At: 18:35 19 January 2011

goal-directed behavioral information. Moreover,if the brain responds to the detection of humanintentionality, a neural response can be used as anovel index for the identification of SGI.
In contrast to intentional social inferences thatare made with the explicit goal to form animpression about a target, spontaneous socialinferences are formed without intention or aware-ness. Many dual-process models in person percep-tion and social neuroscience are consistent withthe idea that information processing involveseither spontaneous associative processes or con-trolled symbolic reasoning (Keysers & Gazzola,2007; Satpute & Lieberman, 2006; Smith &DeCoster, 2000). Past research on spontaneousinferences in social cognition has focused mainlyon traits of other persons. It has been amplydemonstrated that trait inferences are activatedspontaneously, in the sense that if an actor’sbehavior is diagnostic of a trait, inferring thattrait requires almost no intention or awareness,involves only little mental effort, and is difficult tosuppress or modify (for a review, see Uleman,Blader, & Todorov, 2005). Only very recentlyhave researchers begun to explore other types ofspontaneous inferences such as on the goals ofsomeone’s actions (Hassin, Aarts, & Ferguson,2005).
This earlier social cognition research, however,is limited by its use of behavioral tasks, which donot allow exploring the type and timing of theunderlying processes related to the detectionsocial inferences. Are goals identified at earlyautomatic stages or at later reflective comprehen-sion stages, and do these timings differ betweenspontaneous and intentional inferences? Event-related brain potentials (ERPs) are ideally suitedto provide evidence on these matters, as theyafford millisecond accuracy in the timing of brainprocesses, and have been applied recently by VanDuynslaeger, Van Overwalle, and Verstraeten(2007) and Van Duynslaeger, Sterken, Van Over-walle, and Verstraeten (2008) for studying spon-taneous and intentional inferences of otherperson’s traits. Moreover, reliable techniques tolocalize the source of ERPs have emerged (LOR-ETA; Pascual-Marqui, Michel, & Lehmann, 1994)and have been applied to social interpretations ofemotions (Esslen, Pascual-Marqui, Hell, Kochi, &Lehmann, 2004) and traits (Van Duynslaeger etal., 2007). Although the spatial resolution of ERPwaves is poorer than functional magnetic reso-nance imaging (fMRI), a prime advantage is thattheir high time resolution allows localizing and
imaging of the electric activity in a millisecondrange, unlike fMRI, which offers a time resolu-tion in the range of a few seconds.
ERP COMPONENTS
ERPs are waveforms that reflect electric activityof the brain during responses to specific stimuli.They are manifestations of information-proces-sing activities, and different types of ERP com-ponents are associated with different functions inthis process. To elicit ERP responses to particularstimuli, researchers typically use an ‘‘oddball’’paradigm, in which infrequent or inconsistentinformation is provided among otherwise fre-quent and consistent information. Specifically,by providing behavioral descriptions that areinconsistent with a prevailing goal or other socialinferences such as traits (Van Duynslaeger et al.,2007, 2008), researchers can identify ERP wave-form deflections that reflect this inconsistency.The timing of this ERP deflection reveals theprocessing stage at which the goal is identified.Research has revealed two main types of ERPswhich index feature and inconsistency detection.The first type is the P200, which reacts to earlyand automatic feature encoding and classification(Peters, Suchan, Zhang, & Daum, 2005). Thesecond type is the P300, which occurs later andresponds to inconsistencies in comprehension.These characteristics make the P200 and P300ideally suited for exploring the neural correlatesof automatic (early) and intentional (late) goalinferences respectively.
P200 and P300 as indices ofinconsistency
The P200 is an early positive waveform that peaksat about 200 ms after the critical stimulus, andtypically attains the highest amplitudes at frontaland central scalp locations. There is evidence thatthe P200 indexes early sensory stages of featuredetection (Luck & Hillyard, 1994), encoding andclassification (Dunn, Dunn, Languis, & Andrews,1998; Raney, 1993). For instance, modulation ofthe P200 amplitude indicates that words areidentified within the first 200 ms during sentencereading (Dambacher, Kliegl, Hofmann, & Jacobs,2006). More importantly for the present purpose,the P200 might also reflect increased attentiondue to unexpected or improbable stimuli such as
166 VAN DER CRUYSSEN ET AL.
Downloaded By: [Van Overwalle, Frank][Overwalle, Frank Van] At: 18:35 19 January 2011

words (Peters et al., 2005) or after experimentalinstruction (Clark & Hillyard, 1996). It has alsobeen reported that increased P200 amplitudes arecorrelated with enhanced retrieval (Dunn et al.,1998; Smith, 1993). These findings have led to theview that the P200 indexes early comprehensionand updating of a representation in response tonew incoming information (Raney, 1993).
The P300 is a late positive peak that typicallyinitiates around 300 ms after the critical stimulusand continues till 600 or 1000 ms (and is thereforealso termed the late positive potential), and attainsthe highest amplitudes mainly at parietal andcentral scalp locations. Research has documentedthat there is a relation between the P300 and theprocessing of anomalous, inconsistent or infre-quent stimuli presented in a context of otherwisenormal or frequent information, as long as thisinformation is relevant for the task. The ampli-tude of the P300 increases as a function of theamount of discrepancy between the stimulus andthe preceding context, and correlates with laterrecall of the discrepant stimuli, especially whenelaborate rehearsal strategies are minimized (Fa-biani, Karis & Donchin, 1986; Fabiani & Donchin,1995). These findings have led to the view that theP300 is an index of online updating of workingmemory after inconsistency detection
P200 and P300 in social cognition
Social processes occur very rapidly. Severalresearchers found enhanced positive ERPs inresponse to emotional images, such as angry,fearful or happy human faces or armed personsin comparison to neutral faces or unarmedpersons, leading to a P200 (Carretie, Mercado,Tapia, & Hinojosa, 2001; Correll, Urland, & Ito,2006; Eimer & Holmes, 2002; Eimer, Holmes, &McGlone, 2003; Kubota & Ito, 2007) or a P300(Keil et al., 2002). The P200 is also sensitive tooutgroup versus ingroup racial and genderdifferences, consistent with the idea that out-group members are often seen as more negativeor threatening than ingroup members. Ito andcolleagues (Correll et al., 2006; Ito & Urland,2003, 2005; Kubota & Ito, 2007) found thatBlack or male outgroup faces evoke largerP200s than White or female ingroup faces. TheP200 race divergence occurs even when tasksdirect attention away from race (Ito & Urland,2005). These findings of greater attention tomore emotional or distinctive social stimuli may
reflect an automatic vigilance effect in whichattention is quickly and relatively automaticallydirected to stimuli with potentially negativeimplications. None of these studies, however,involves social inferences about the intentions orattributes of individual persons.
More relevant evidence on person inferences isavailable on the P300. Cacioppo and coworkers(Cacioppo, Crites, Berntson, & Coles, 1993; Ca-cioppo, Crites, Gardner, & Berntson, 1994) foundthat an evaluative inconsistency between a traitword and previously presented trait words (e.g., anegative trait after a sequence of positive traits)elicited a large P300 between approximately 500and 1000 ms at central and parietal scalp locations.Of more interest for our research, Van Duynslae-ger and colleagues (2007, 2008; see also Bartholow,Fabiani, Gratton, & Bettencourt, 2001; Bartholow,Pearson, Gratton, & Fabiani, 2003) engaged theirparticipants to form spontaneous or intentionaltrait impressions about actors described in shortbehavioral sentences. The behaviors were eitherconsistent with traits implied during precedingbehaviors (e.g., a friendly act after a sequence ofcourteous behaviors) or opposite in valence withthe implied traits (e.g., an impolite act). VanDuynslaeger et al. (2007, 2008) found greaterP300 activation at about 600 ms after presentingthe critical word for trait-inconsistent sentences asopposed to trait-consistent sentences, regardless ofwhether the inference was made spontaneously orintentionally. This result implies that traits areidentified at a late processing stage about 600 mspost-stimulus.
SCALP LOCATIONS OF SOCIALJUDGMENTS
Recent fMRI evidence suggests that two brainareas are involved in the understanding andattribution of mental states (i.e., goals and traits)of others: the temporoparietal junction (TPJ) andthe medial prefrontal cortex (mPFC; for reviewssee Amodio & Frith, 2006; Van Overwalle, 2008).Research seems to indicate that the TPJ is mainlyinvolved in the spontaneous attribution of tem-porary intentions and desires by others. Whenviewing animations of simple objects that move ina human-like fashion, observers have an immedi-ate sense of intentionality (e.g., a triangle‘‘chases’’ a square) that activates the TPJ (Martin& Weisberg, 2003; Ohnishi et al., 2004; Schultz,Imamizu, Kawato, & Frith, 2004). When seeking
GOAL INFERENCES AND P200 167
Downloaded By: [Van Overwalle, Frank][Overwalle, Frank Van] At: 18:35 19 January 2011

for or reacting to an appropriate story ending, theTPJ is more strongly activated for stories invol-ving behavioral intentions than mere physicalevents (Blakemore et al., 2007; den Ouden, Frith,Frith, & Blakemore, 2005; Saxe & Wexler, 2005;Vollm et al., 2006; Walter et al., 2004). Whenreading stories, the TPJ is recruited more whenthese stories involve beliefs and reasoning by theactors than their physical appearances (Saxe &Powell, 2006; for an overview, see Van Overwalle,2008). All these tasks do not focus explicitly onthe goal underlying the behaviors, and are thusrelatively spontaneous. In contrast, the mPFC isrecruited for attributing enduring traits (e.g.,Mitchell, Banaji, & Macrae, 2005; Todorov, Gob-bini, Evans, & Haxby, 2007), and is involved incontrolled processing and action monitoring(Miller & Cohen, 2001; Satpute & Lieberman,2006).
This has led to the recent proposal that that theTPJ implements pre-reflective, intuitive and em-pathic representations about other people’s in-tentions and beliefs, as part of an automaticmirror system that recognizes observed intentionsof others by their similarity with own behavioralintentions, whereas the mPFC supports reflectiverepresentations, when one is deliberatively rea-soning about other people (Keysers & Gazzola,2007; Keysers & Perrett, 2004). Consistent withthis, a LORETA source analysis conducted byVan Duynslaeger et al. (2007) revealed thatspontaneous trait inferences recruit mainly theright TPJ, while intentional trait inferences dom-inate activity in the mPFC. Moreover, theseresearchers found a correlation between TPJand mPFC activity and cued recall, but onlyunder spontaneous instructions.
To summarize, previous ERP research suggeststhat an early P200 and a late P300 followinginconsistent information is most likely to befound in frontal and central scalp areas for theP200, and at central and parietal scalp regions forthe P300. Moreover, earlier fMRI and LORETAimaging research suggests that the main brainactivity during social (i.e., trait) inferences islocalized in the TPJ for spontaneous inferences,and in the mPFC for intentional inferences.
RESEARCH
For the present research, we borrowed VanDuynslaeger et al.’s (2007, 2008) paradigm, andprovided short behavioral descriptions that are
consistent, inconsistent or irrelevant with the goalimplied during preceding behaviors. We in-structed our participants either explicitly to inferthe goal of each target person (IGI) or to read thestimulus material carefully, without mentioninganything about the person’s goals or impressions(SGI). If the ERP waveforms show effects ofinconsistency, this indicates that the participantsidentified the implied goal. Because negativeinconsistencies in social inference give rise tomore prominent responses (Cacioppo, Gardner,& Berntson, 1999; Reeder & Brewer, 1979;Ybarra, 2002) and stronger ERP deflections(Bartholow et al., 2003; Cacioppo et al., 1999;Van Duynslaeger et al., 2007, 2008), we providedonly positive goal expectations with negativeinconsistencies (e.g., preparing a party is followedby buying a gun). To study the breadth of the goalinference process, we also included positive eventdescriptions that did not involve any intention orgoal-directedness from the actor. To the extentthat goal inferences are driven by the perceivedintentionality of behaviors, these no-goal eventsshould elicit even stronger ERP inconsistencyeffects.
Like Van Duynslaeger et al. (2007, 2008), wetook several memory measures as behavioralvalidation of the ERPs, taken after the presenta-tion of all stimulus material so that they did notinterfere with the ongoing EEG recording. Thefirst, a cued recall task (Winter & Uleman, 1984),requests participants to recall all informationpresented with the aid of cue words that reflectthe implied goals. When observing or readingabout a goal-implying behavior, the inferred goalsare assumed to be stored in memory togetherwith the behavioral information from which theyare inferred. As a consequence, enhanced recallof consistent behaviors aided by this goal cuesuggests that goal interpretations were madewhile reading the material. The second, a sen-tence completion task (Bartholow et al., 2001,2003), consists of completing the last, criticalgoal-implying word of the original sentences. Inearlier trait research, memory effects on thismeasure heavily depended on the assumptionthat actors have a limited number of enduringtraits, so that inconsistent information receivesmore cognitive processing and is memorizedbetter than consistent behavior (Stangor &McMillan, 1992). However, this measure mightbe less diagnostic for goals because inconsisten-cies are not tied to a unique and stable expecta-tion about a person (for a review, see Fyock &
168 VAN DER CRUYSSEN ET AL.
Downloaded By: [Van Overwalle, Frank][Overwalle, Frank Van] At: 18:35 19 January 2011

Stangor, 1994). Goals are adaptive to newcircumstances, so that inconsistent behavior maysuggest a change of proximal means in the pursuitof the same distal intention (e.g., buying an‘‘inconsistent’’ gun for a party suggests a surpriseparty for Halloween or carnival) rather than realdiscrepancies.
HYPOTHESES
Dual-process models in person perception andsocial neuroscience posit that spontaneous asso-ciative processes occur fast, and subserve andinform later intentional thoughts (for a review,see Smith & DeCoster, 2000). Based on thistheorizing, we expect many parallels betweenspontaneous and intentional processing modesand resulting brain activity involved in goalinferences, although we also anticipate somesmaller differences between SGI and IGI. Inparticular, we make the following predictions.
For the neural measures, we hypothesize that ifgoal inferences are automatic, then goal incon-sistencies generate an early P200 mainly in frontaland central midline scalp areas. In contrast, if goalinferences require deliberate processing, we ex-pect a later P300 mainly at central and parietalmidline scalp regions. In addition, we expect thatgoal inferences activate several distinct brainareas as revealed in LORETA brain maps. Inline with recent evidence in social neuroscience,we propose that spontaneous goal identificationrecruits mainly the TPJ, whereas intentionalprocessing activates predominantly the mPFC(Van Overwalle, 2008). However, if goal infer-ences are automatic and inform later deliberativeprocessing (Keysers & Gazzola, 2007; Smith& DeCoster, 2000), we speculate that thesedifferences in brain activation will be reducedduring the early automatic stage of goal identifi-cation (when the TPJ is active), and will appearonly at very early preparatory or later delibera-tive stages.
For the memory measures that validate theexistence of goal inferences, we assume that SGIare made to the same degree as IGI, and arestored in memory to a similar degree. Further-more, we expect better memory for goal-consis-tent behaviors after recall cued with the inducedgoal than after goal-inconsistent behaviors, but nodifferences on the sentence completion task.Finally, we expect correlations of these memorymeasures with ERP amplitude as well as with
LORETA activation in the TPJ or mPFC, be-cause these neural indices reflect greater brainactivity and thus deeper processing, leading tobetter memory.
METHOD
Participants
Participants were 50 students at the Vrije Uni-versiteit Brussel (VUB), without prior history ofany neurological dysfunction. There were 25women and 25 men, with an age varying between18 and 35, and an average of 21.3493.60. Themajority of participants were recruited via auniversity-wide electronic mailing system for allstudents, while some additional participants wererecruited on a personal basis. In exchange fortheir participation, they were paid 25 euros.Among all participants, 26 received a sponta-neous instruction while another 24 participantsreceived an intentional instruction.
Stimulus material
The general design was adapted from Van Duyn-slaeger et al. (2007, 2008; see also Bartholowet al., 2001, 2003). Participants read severalsentences that described the behavior of a ficti-tious target actor and from which a strong goalcould be inferred or not. The sentences involved25 positive goal-implying behaviors and 7 positiveno-goal events, and each set of sentences wasintroduced by showing the name of the actor onthe computer screen for 2 s. To avoid anyassociation with a familiar and/or existing name,fictitious Star Trek-like names were used (VanDuynslaeger et al., 2007, 2008). For each targetactor, a series of four behavioral sentences waspresented, each consisting of six words shown inthe center of the computer screen. Every wordwas presented during 300 ms followed by a350 ms blank (Osterhout, Bersick, & McLaughlin,1997), except for the last word of a sentencewhich was the critical one, and was thereforepresented for 350 ms to make sure that it wouldnot be missed. The first three sentences implied acommon goal (e.g., preparing a party), whereasthe last word of the last sentence determined thedegree of consistency with the previously impliedgoal: goal-consistent (GC), goal-inconsistent(GI) and goal-irrelevant (GIR). GC-sentences
GOAL INFERENCES AND P200 169
Downloaded By: [Van Overwalle, Frank][Overwalle, Frank Van] At: 18:35 19 January 2011

describe intentional behaviors that are consistent
with the implied goal from the introductory
sentences (e.g., ‘‘Vortram searches quickly for
colorful decorations’’), GI-sentences are goal-
inconsistent (e.g., ‘‘Vortram searches quickly for
his rifle’’), while GIR-sentences describe neutral
behaviors (e.g., ‘‘Vortram searches for his brown
pet’’). All these conditions were carefully
matched so that the last sentence always con-
tained the same verb.In addition, no-goal sentences described non-
intentional events during which the actor was
merely observing a phenomenon. The first three
sentences implied a common phenomenon (e.g.,
snowing), whereas the last sentences determined
the consistency with this phenomenon: no-goal-
consistent (NGC), no-goal-inconsistent (NGI)
and no-goal-irrelevant (NGIR). NGC-sentences
are consistent with the three introductory sen-
tences (e.g., ‘‘Boelmok senses cold and wet
flakes’’), NGI-sentences are inconsistent (e.g.,
‘‘Boelmok senses a very oppressive heat’’), while
GIR-sentences describe neutral behaviors (e.g.,
‘‘Boelmok senses itches behind his ears’’). All
these conditions were also carefully matched so
that the last sentence always contained the same
verb.The sentences were newly developed in Dutch
and pilot tested to check if a great majority of
the participants drawn from a similar population
spontaneously inferred the same implied goal.
The participants (n�89) were requested to write
down the goal implied by the introductory and
GC sentences, and if more than 70% indicated
the same goal (or a close synonym) it was
retained for the main study. No-goal observa-
tions were included in the main study if less than
30% of the participants indicated any goal.
Another group of participants (n�120) was
requested to rate to what extent the implied
goal could be applied on the sentences using an
11-point scale (0�less applicable to 10�more
applicable), and GI to GIR sentences showing
less applicability were retained. As a final check,
a small group of participants (n�16) was
additionally requested to rate the valence of
the GI to GIR sentences using an 11-point scale
(0�negative to 10�positive). GI sentences that
were seen as negative (i.e., mean rating below
midpoint 5) were retained, as well as GIR
sentences that were seen as positive (i.e., mean
rating above midpoint 5).
Procedure
After seating the participant, the electrodes forthe electroencephalographs (EEG) and electro-oculograms (EOG) were placed at the correctlocations. The participants were informed thatthey would read stories about several persons andthat each story would start with a name followedby different sentences about the actor. Next, theyreceived the crucial instruction to ‘‘try to famil-iarize yourself with the material of the experi-ment’’ (SGI) or to ‘‘try to infer the goal that theperson wants to reach’’ (IGI; for similar instruc-tions, see also Todorov & Uleman, 2002). Becauseparticipants could not read at their own pace butwere shown each word at a fixed pace, they werealso told to pay as much attention as possible toeach word, because they would get questionsabout them afterwards. It was also emphasizedto move and eye-blink as little as possible to limitartifacts in the EEG (Stern, Ray, & Quigley,2001).
During reading the EEG was recorded. After-wards the electrodes for the EEG and EOG wereremoved. Next, the participants were given thecued recall and sentence completion tasks in thesame order for all participants. In the cued recalltask, participants had to write as many behavioralsentences as possible with the aid of words thatconsisted of all the implied goals (goal-implyingsentences) and all observations (no-goal sen-tences). In the sentence completion task, partici-pants were presented with 19 incomplete GC-,GI- and GIR-sentences (randomly selected fromthe sentence pool) and had to complete the lastword.
Electrophysiological registration andanalysis
The EEG was recorded from 19 scalp sitesaccording to the international 10�20 electrodesystem, using Ag/AgCl electrodes fixed in astretch head cap (Activeshield) from AdvancedNeuro Technology. The montage included threemidline sites (Fz, Cz, Pz) and eight sites overeach hemisphere (Fp1/Fp2, F3/F4, F7/F8, C3/C4,T3/T4, T5/T6, P3/P4, O1/O2), with the averageof all EEG-channels as recording and offlinereference. A ground electrode was located alongthe midline between the Fz and Cz electrodes.Vertical and horizontal eye movements were
170 VAN DER CRUYSSEN ET AL.
Downloaded By: [Van Overwalle, Frank][Overwalle, Frank Van] At: 18:35 19 January 2011

recorded bipolarly via EOGs using electrodesplaced above and below the left eye and 1 cmexternal to the outer canthus of each eye,respectively. Impedance was kept below 10 kV.The EEG was recorded continuously during thewhole experiment, with a digitizing rate of 256Hz. Stimulus presentation, recording and analysiswere done with the hardware (Cognitrace) andsoftware (Eevoke, Eemagine and ASA) fromAdvanced Neuro Technology. The stimulus ma-terial was presented on screen and directly time-locked on the EEG recordings by the Eevokesoftware. The raw EEG data were filtered by a0.03 to 30 Hz band-pass, horizontal and verticalEOG artifacts were corrected using the SOBIalgorithm (Joyce, Gorodnitsky, & Kutas, 2004),and remaining artifacts beyond �75 and 75 mVat each time unit were removed before analysis.For ERP, the relevant EEG sequences wereaveraged per participants, channel and condition.Each relevant sequence began 250 ms prior tothe presentation of the last critical word in eachsentence (prestimulus baseline) and lasted to1125 ms after the presentation of the criticalword, leaving 82% artifact-free sequences. Agrand average was then calculated on theseindividual ERP data across all participants.
RESULTS
ERP data
To statistically analyze the time course of posi-tive and negative peaks in the ERP waveformsand associated cognitive activity, we divided theERP data of each participant in consecutive timeintervals (50�300, 300�450, 450�600, and 600�1000 ms). The largest positive and negative peaks(with maximum and minimum amplitude respec-tively) in each interval were identified for eachparticipant. For the statistical analyses, we com-pared the GC condition with the GI and GIRconditions because these conditions share thesame three introductory sentences, and differonly with respect to consistency with the impliedgoal in the last sentence. We compared the GCconditions also with the NGC sentences (hence-forth also termed no-goal condition) because thiscondition is matched with the GC condition forconsistency with its introductory sentences, anddiffers mainly with respect to the implied goal-directedness or intentionality of the behavior.Specifically, the positive and negative peaks were
statistically analyzed for each channel separatelyby means of a repeated measures analysis ofvariance (ANOVA) with Consistency (GC, GI,GIR, NGC) and Interval (50�300, 300�450, 450�600, 600�1000) as within-participants factors andInstruction (read, goal) as between-participantsfactor. Reported p-levels for this and all otherANOVAs are corrected for violations of thesphericity assumption using the Greenhouse-Geisser correction.
The ANOVA revealed for all midline chan-nels a main effect for Interval, F(3, 126)�5.83�23.74, pB.001, as well as for Consistency, F(3,126)�6.45*29.05, pB.002, and for many chan-nels also a significant interaction between them.In order to test our specific hypothesis concern-ing the P200 and P300 under different instruc-tions, we conducted simple t-tests comparing thepositive peaks in the goal-consistent conditionagainst the inconsistent (GC vs. GI), irrelevant(GC vs. GIR) and no-goal (GC vs. NGC)conditions separately for SGI and IGI. Allsignificant t-tests were also visually inspected toverify that they reflected positive peaks. Forcompleteness, we also tested for negative peaks,but visual and statistical inspection revealed nonegative ERP waveforms unless noted otherwise.In order to control for multiple comparisons, ouranalytical strategy was as follows. To test ourhypothesis, we used a conventional .05 a-level forthe predicted frontal (Fz) and central (Cz) mid-line scalp locations in the 50�300 ms interval forthe P200, and for the predicted central (Cz) andparietal (Pz) midline scalp locations in 300�1000ms interval for the P300. The results are given inTable 1, with the predicted locations indicated inbold type. To explore other possible locations, weadjusted the a-level to .000098 using the Bonfer-roni correction, which takes into account all cellsof the ANOVA (32) and all remaining channels(16). None of the differences at other scalplocations survived this stringent a-level. Forillustrative purposes, however, these differencesexceeding the nonsignificant .10 level are re-ported in Table 1.
Goal inconsistencies
Our hypothesis is that inconsistencies with theimplied goal generate positive deflections in theERP waveforms. We first test this hypothesis forthe GI and GIR conditions which reflect incon-sistencies with the implied goal (but still involvegoal-directed behavior). Table 1 gives the mean
GOAL INFERENCES AND P200 171
Downloaded By: [Van Overwalle, Frank][Overwalle, Frank Van] At: 18:35 19 January 2011
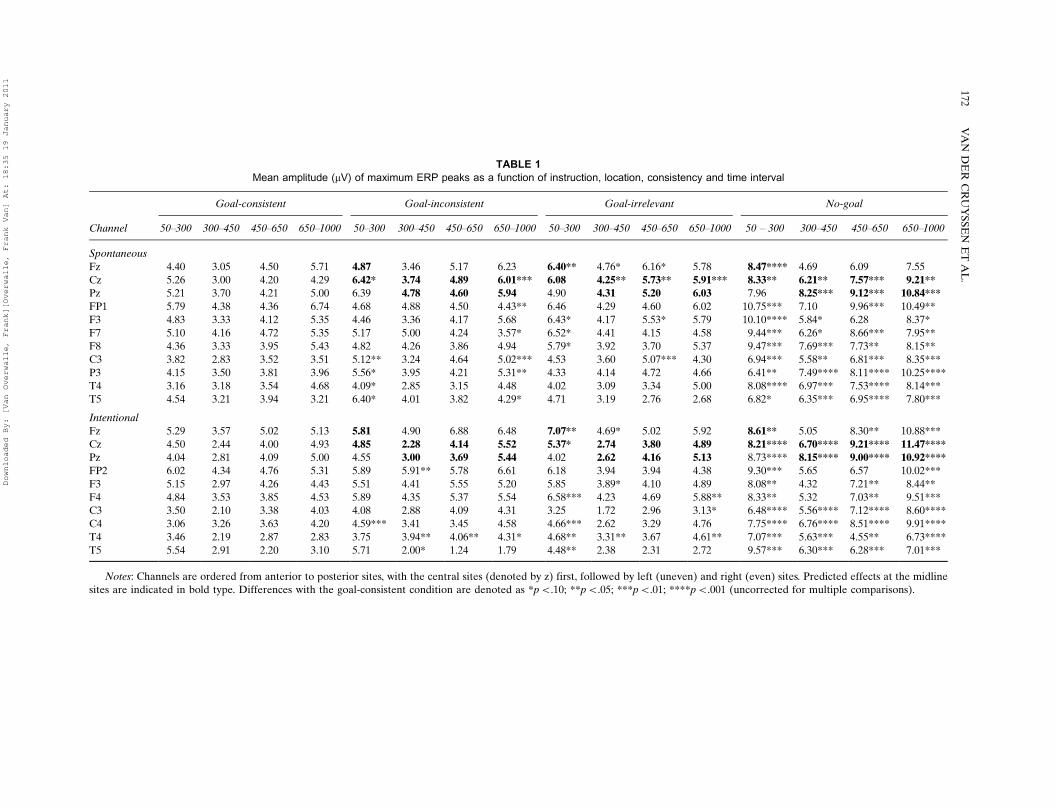
TABLE 1
Mean amplitude (mV) of maximum ERP peaks as a function of instruction, location, consistency and time interval
Goal-consistent Goal-inconsistent Goal-irrelevant No-goal
Channel 50�300 300�450 450�650 650�1000 50�300 300�450 450�650 650�1000 50�300 300�450 450�650 650�1000 50 � 300 300�450 450�650 650�1000
Spontaneous
Fz 4.40 3.05 4.50 5.71 4.87 3.46 5.17 6.23 6.40** 4.76* 6.16* 5.78 8.47**** 4.69 6.09 7.55
Cz 5.26 3.00 4.20 4.29 6.42* 3.74 4.89 6.01*** 6.08 4.25** 5.73** 5.91*** 8.33** 6.21** 7.57*** 9.21**Pz 5.21 3.70 4.21 5.00 6.39 4.78 4.60 5.94 4.90 4.31 5.20 6.03 7.96 8.25*** 9.12*** 10.84***
FP1 5.79 4.38 4.36 6.74 4.68 4.88 4.50 4.43** 6.46 4.29 4.60 6.02 10.75*** 7.10 9.96*** 10.49**
F3 4.83 3.33 4.12 5.35 4.46 3.36 4.17 5.68 6.43* 4.17 5.53* 5.79 10.10**** 5.84* 6.28 8.37*
F7 5.10 4.16 4.72 5.35 5.17 5.00 4.24 3.57* 6.52* 4.41 4.15 4.58 9.44*** 6.26* 8.66*** 7.95**
F8 4.36 3.33 3.95 5.43 4.82 4.26 3.86 4.94 5.79* 3.92 3.70 5.37 9.47*** 7.69*** 7.73** 8.15**
C3 3.82 2.83 3.52 3.51 5.12** 3.24 4.64 5.02*** 4.53 3.60 5.07*** 4.30 6.94*** 5.58** 6.81*** 8.35***
P3 4.15 3.50 3.81 3.96 5.56* 3.95 4.21 5.31** 4.33 4.14 4.72 4.66 6.41** 7.49**** 8.11**** 10.25****
T4 3.16 3.18 3.54 4.68 4.09* 2.85 3.15 4.48 4.02 3.09 3.34 5.00 8.08**** 6.97*** 7.53**** 8.14***
T5 4.54 3.21 3.94 3.21 6.40* 4.01 3.82 4.29* 4.71 3.19 2.76 2.68 6.82* 6.35*** 6.95**** 7.80***
Intentional
Fz 5.29 3.57 5.02 5.13 5.81 4.90 6.88 6.48 7.07** 4.69* 5.02 5.92 8.61** 5.05 8.30** 10.88***
Cz 4.50 2.44 4.00 4.93 4.85 2.28 4.14 5.52 5.37* 2.74 3.80 4.89 8.21**** 6.70**** 9.21**** 11.47****Pz 4.04 2.81 4.09 5.00 4.55 3.00 3.69 5.44 4.02 2.62 4.16 5.13 8.73**** 8.15**** 9.00**** 10.92****
FP2 6.02 4.34 4.76 5.31 5.89 5.91** 5.78 6.61 6.18 3.94 3.94 4.38 9.30*** 5.65 6.57 10.02***
F3 5.15 2.97 4.26 4.43 5.51 4.41 5.55 5.20 5.85 3.89* 4.10 4.89 8.08** 4.32 7.21** 8.44**
F4 4.84 3.53 3.85 4.53 5.89 4.35 5.37 5.54 6.58*** 4.23 4.69 5.88** 8.33** 5.32 7.03** 9.51***
C3 3.50 2.10 3.38 4.03 4.08 2.88 4.09 4.31 3.25 1.72 2.96 3.13* 6.48**** 5.56**** 7.12**** 8.60****
C4 3.06 3.26 3.63 4.20 4.59*** 3.41 3.45 4.58 4.66*** 2.62 3.29 4.76 7.75**** 6.76**** 8.51**** 9.91****
T4 3.46 2.19 2.87 2.83 3.75 3.94** 4.06** 4.31* 4.68** 3.31** 3.67 4.61** 7.07*** 5.63*** 4.55** 6.73****
T5 5.54 2.91 2.20 3.10 5.71 2.00* 1.24 1.79 4.48** 2.38 2.31 2.72 9.57*** 6.30*** 6.28*** 7.01***
Notes: Channels are ordered from anterior to posterior sites, with the central sites (denoted by z) first, followed by left (uneven) and right (even) sites. Predicted effects at the midline
sites are indicated in bold type. Differences with the goal-consistent condition are denoted as *pB.10; **pB.05; ***pB.01; ****pB.001 (uncorrected for multiple comparisons).
17
2V
AN
DE
RC
RU
YS
SE
NE
TA
L.
Downloaded By: [Van Overwalle, Frank][Overwalle, Frank Van] At: 18:35 19 January 2011

amplitudes for each condition, as well as t-tests ofthe GI and GIR conditions against the GCcondition. The results were generally in accor-dance with our predictions. As predicted, t-testsof the GI and GIR sentences revealed differenceswith the GC sentences that were significant at thefrontal (Fz) and central (Cz) midline sites. Nosignificant differences were found at the parietalmidline (Pz) site. Of most importance is thatsignificant differences were found already at thefirst time interval at the frontal midline, for bothSGI and IGI. The mean latency for the earliestpositive peaks at Fz in the three conditions rangesbetween 166�178 ms for SGI and 201�205 ms forIGI. This suggests a P200 and indicates that goalinconsistencies are detected at a very earlyprocessing stage. At the left and right hemisphere,similar differences were found mainly at frontal,central, and temporal sites and less so at parietalsites (see Table 1), none of which survived theBonferroni correction. As can be seen from Table1, for the midline sites, the difference with theGIR condition is slightly stronger than with theGI condition, which might be due to our earliersuggestion that goal inconsistencies suggest achange of means or type of the same distal goal(i.e., searching for a rifle may imply a Halloweenparty rather than an ordinary party) whereasirrelevant behaviors imply less goal-directedness(i.e., searching for a spoon is not indicative of aparty at all).
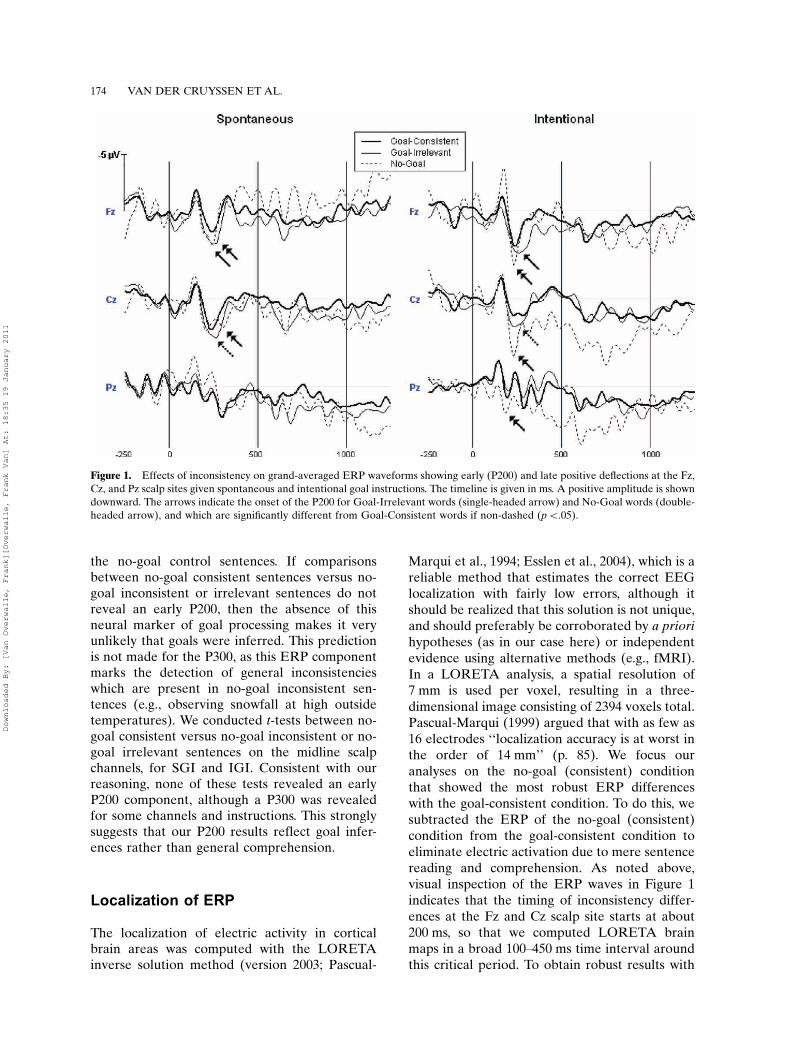
Because stronger results were found for GIRsentences, the ERP waveform of this condition isdepicted together with the GC condition inFigure 1. As can be seen, goal-irrelevant beha-viors (light line) elicited a larger positive ERPthan goal-consistent behaviors (heavy line) start-ing at about 200 ms. These P200 deflections(marked by a single-headed arrow) are significantfor Fz, but not for Cz, for both SGI and IGI. TheCz midline site reveals further positive ERPdeflections at later time intervals that are sig-nificant for SGI (at pB.05; see Table 1), but notfor IGI. There are no other substantial differ-ences in the waveforms between SGI and IGI.
Goals vs. no-goals
The no-goal consistent (NGC) condition re-flects a major difference with the GC condition inthat it involves no intentional behavior, althoughboth conditions reflect consistency with the pre-ceding sentences describing the actor. As onewould expect, these comparisons reveal much
stronger effects. Most differences are significantat all midline sites. Of most importance is thatearly significant differences are found again, nowat all frontal (Fz) and central (Cz) midline sitesfor both SGI and IGI, as predicted. The meanlatency for the earliest positive peaks in the no-goal condition at Fz and Cz ranges between 142�177 ms for SGI and 177�178 ms for IGI. Thisagain suggests a P200 indicating that goals aredetected at a very early processing stage. Also aspredicted, significant differences at later intervalsare found at all central (Cz) and parietal (Pz)midline sites. At the left and right hemisphere(see Table 1), similar differences were foundmainly at frontal, central and temporal sites andless so at parietal sites, none of which survived theBonferroni correction.
The ERP waveform of the no-goal conditionis also depicted in Figure 1. As can be seen, no-goal behaviors (broken line) elicited a largerpositive ERP than goal-consistent behaviors(heavy line) starting at about 200 ms. TheseP200 deflections (marked with a double-headedarrow) are significant for SGI and IGI, andoverlap substantially with the P200 of the goal-irrelevant condition (single-headed arrow). Theno-goal sentences reveal further deflections atlater time intervals. An interesting point is thatthese deflections at the Fz site are more nega-tive-going for SGI (pB.01 uncorrected), whilethey are positive-going for all other midline sitesand for IGI (pB.05, uncorrected). There is alsoan early negative peak at around 150 ms in thePz channel for SGI and in the Fz channel forIGI. Given that these negative peaks were notanticipated and not corroborated by other chan-nels or by the goal-irrelevant waveform, they arefurther ignored.
Taken together, all sentences generate a P200,which is substantially stronger for sentences thatare inconsistent or irrelevant with the expectedgoal or involve no goal at all. These findingssuggest that prior goal expectations are devel-oped automatically at an early processing stage,so that inconsistent information is rapidly de-tected, irrespective of the SGI or IGI instruction.
No-goal inconsistencies
One conceptual question that remains is thatthe preceding findings may reflect processing ofgeneral comprehension rather than processing ofgoals. In order to rule out this alternative inter-pretation, we conducted additional analyses on
GOAL INFERENCES AND P200 173
Downloaded By: [Van Overwalle, Frank][Overwalle, Frank Van] At: 18:35 19 January 2011
the no-goal control sentences. If comparisonsbetween no-goal consistent sentences versus no-goal inconsistent or irrelevant sentences do notreveal an early P200, then the absence of thisneural marker of goal processing makes it veryunlikely that goals were inferred. This predictionis not made for the P300, as this ERP componentmarks the detection of general inconsistencieswhich are present in no-goal inconsistent sen-tences (e.g., observing snowfall at high outsidetemperatures). We conducted t-tests between no-goal consistent versus no-goal inconsistent or no-goal irrelevant sentences on the midline scalpchannels, for SGI and IGI. Consistent with ourreasoning, none of these tests revealed an earlyP200 component, although a P300 was revealedfor some channels and instructions. This stronglysuggests that our P200 results reflect goal infer-ences rather than general comprehension.
Localization of ERP
The localization of electric activity in corticalbrain areas was computed with the LORETAinverse solution method (version 2003; Pascual-
Marqui et al., 1994; Esslen et al., 2004), which is areliable method that estimates the correct EEGlocalization with fairly low errors, although itshould be realized that this solution is not unique,and should preferably be corroborated by a priorihypotheses (as in our case here) or independentevidence using alternative methods (e.g., fMRI).In a LORETA analysis, a spatial resolution of7 mm is used per voxel, resulting in a three-dimensional image consisting of 2394 voxels total.Pascual-Marqui (1999) argued that with as few as16 electrodes ‘‘localization accuracy is at worst inthe order of 14 mm’’ (p. 85). We focus ouranalyses on the no-goal (consistent) conditionthat showed the most robust ERP differenceswith the goal-consistent condition. To do this, wesubtracted the ERP of the no-goal (consistent)condition from the goal-consistent condition toeliminate electric activation due to mere sentencereading and comprehension. As noted above,visual inspection of the ERP waves in Figure 1indicates that the timing of inconsistency differ-ences at the Fz and Cz scalp site starts at about200 ms, so that we computed LORETA brainmaps in a broad 100�450 ms time interval aroundthis critical period. To obtain robust results with
Figure 1. Effects of inconsistency on grand-averaged ERP waveforms showing early (P200) and late positive deflections at the Fz,
Cz, and Pz scalp sites given spontaneous and intentional goal instructions. The timeline is given in ms. A positive amplitude is shown
downward. The arrows indicate the onset of the P200 for Goal-Irrelevant words (single-headed arrow) and No-Goal words (double-
headed arrow), and which are significantly different from Goal-Consistent words if non-dashed (pB.05).
174 VAN DER CRUYSSEN ET AL.
Downloaded By: [Van Overwalle, Frank][Overwalle, Frank Van] At: 18:35 19 January 2011

less noise, LORETA solutions were averagedover time intervals of 50 ms each, except for thecritical 200�250 ms interval, where averages weretaken for each 25 ms interval to allow visualinspection of shorter changes.
Figure 2 displays the LORETA solutions of themaximum activation for two predicted brain areas(TPJ and mPFC), and for another area that wasalso frequently activated and identified as themedial paracentral cortex (mPC) extending to theposterior mPFC, which reacts to conflicting sti-muli (e.g., Stroop task; Van Overwalle, 2008). Thefigure displays the LORETA brain maps for SGI(left panels) and IGI (middle panels). The activa-tion of the TPJ was mainly at the right hemi-sphere and is therefore shown on the right lateralbrain surface, whereas the activation of the mPFCand mPC was medial and therefore shown on themedial surface. As can be seen, the strongestactivation in the TPJ is confined mainly to theinferior part (e.g., Talairach x�y�z coordinates 53,�53, 15 in SGI at 200�225 ms), which is typicalfor goal inferences (mean coordinates 51, �57,25 in the meta-analysis by Van Overwalle, 2008).The strongest activation of the mPFC is confinedmainly to its ventral part (e.g., coordinates 3, 52,�6 in IGI at 300�350 ms), which is typical forinferences on familiar others (�1, 49, 4; VanOverwalle, 2008). This latter result may be due tothe fact that four sentences were presented aboutthe same actor, so that while reading the lastsentence the participants were already somewhatfamiliar with the actor.
We tested differences between SGI and IGI bya voxel-by-voxel t-test of the LORETA images(see Esslen et al., 2004). To identify the areas ofmost interest, we consider differences in thepredicted TPJ and mPFC areas at a somewhatmore stringent .025 significance level (jtj�2.25).The t-values are displayed via LORETA imagesin the right panels of Figure 2, in blue for t-valuesthat are stronger in IGI. We also calculateddifferences at other areas with a nonparametricrandomization test (Nichols & Holmes, 2002)which corrects for multiple comparisons.
We predicted that the TPJ would be relativelymore involved in SGI while the mPFC would bemore involved in IGI. In general, these predic-tions were confirmed. Figure 2 shows that beforegoal identification (100�200 ms), activation wasmaximal in the TPJ during SGI and maximal inthe mPFC (and other medial areas) during IGI.During goal identification (�200 ms) and 100 msafterwards, the TPJ was most active under both
SGI and IGI processing modes. After 300�350 ms,the mPFC activation was strongest for bothprocessing modes, perhaps reflecting deliberativethoughts related to the initial goal identificationor other thoughts.
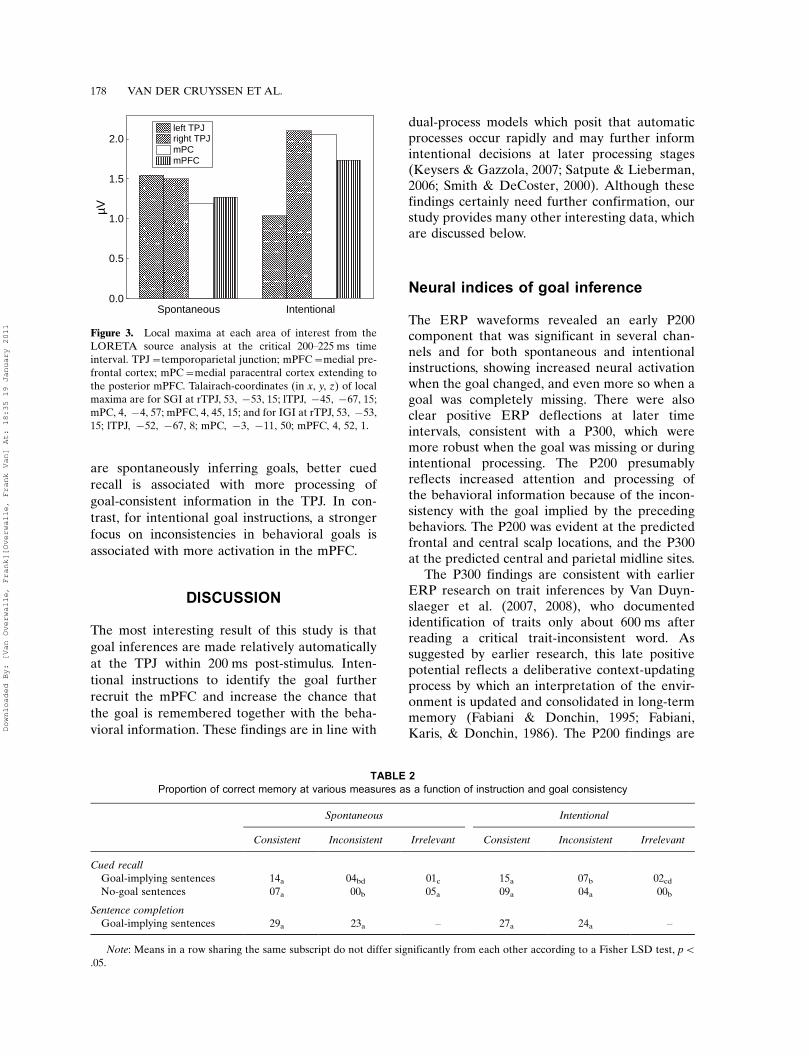
These results are generally confirmed by thet-values in Figure 2. The activation of the mPFC issignificant stronger for IGI than SGI at the early100�200 interval, while no reliable differencesappear in subsequent intervals. At around 300 ms,the t-values show more activation at the TPJbilaterally during IGI. At first sight, this mayseem surprising as the left (SGI) vs. middle (IGI)panels do not show this difference. However, thisis due to the fact that the t-tests calculate absolutedifferences, while the activation in the SGI andIGI brain maps is shown in terms of localmaxima. To illustrate this, Figure 3 comparesthe local maxima of activation at the critical200�225 ms time interval in the three brain areasof interest. As can be seen, although some areasdominate, substantial activation is observed forall areas. There were no reliable differencesbetween SGI and IGI for other areas usingcorrections for multiple comparisons (Nichols &Holmes, 2002), except for significantly moreactivation in the mPC in the intervals 225�250and 300�350, pB.05.
Memory measures
The responses on the cued recall and sentencecompletion tasks were scored on the basis ofverbatim accuracy of the sentence (without theactor’s name), although synonyms were allowed.The proportion of correct responses is given inTable 2.
Cued recall
This memory measure was taken to verify thehypothesis that goal inferences were made on thebasis of goal-consistent behavioral descriptions,and requires higher goal-cued recall for GC ascompared to GI, GIR and NGC behaviors. Therecall data were analyzed with an ANOVA withInstruction (read, goal) as between-participantsfactor and Consistency (GC, GI, GIR, NGC) aswithin-participants factors. The ANOVA revealedthe predicted main effect of Consistency, F(3,144)�26.38, pB.001, h2�.355. There was nomain or interaction effect of Instruction, aspredicted. Planned comparisons indicated that
GOAL INFERENCES AND P200 175
Downloaded By: [Van Overwalle, Frank][Overwalle, Frank Van] At: 18:35 19 January 2011
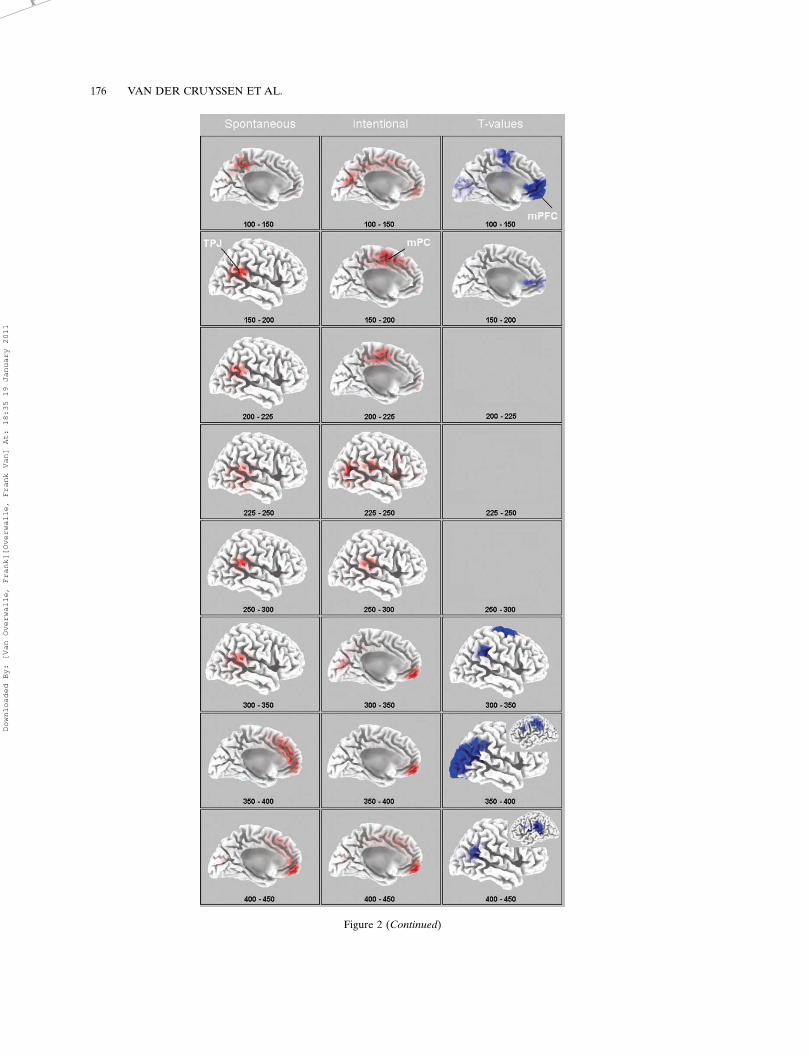
Figure 2 (Continued)
p
176 VAN DER CRUYSSEN ET AL.
Downloaded By: [Van Overwalle, Frank][Overwalle, Frank Van] At: 18:35 19 January 2011

conforming to predictions, the goal cues elicitedstronger recall of GC sentences than any of theother sentences, F(1, 48)�71.68, pB.001, whilethere was a further significant difference betweenGI and GIR sentences, F(1, 48)�8.10, pB.01. Noother differences were significant. This suggeststhat the implied goal was more strongly associatedin memory with consistent behaviors regardless ofthe spontaneous or intentional instruction.
Sentence completion
We used a similar ANOVA with Instruction(read, goal) as between-participants factor andConsistency (GC, GI) as within-participants fac-tors (without the GIR and NGC cells; in analogywith Bartholow et al., 2001, 2003, and VanDuynslaeger et al., 2007, 2008, the latter werenot included in the sentence completion task). Inline with our predictions, we found no significanteffects, F(1, 48)B1.5, ns. It is instructive to note,however, that all proportions were significantlyhigher than zero, p�.001, indicating that asubstantial proportion of the critical words asso-ciated or inconsistent with the implied goals werememorized.
Correlations between neuralcomponents and memory measures
The results thus far reveal that the processing ofinconsistent or irrelevant information is asso-ciated with a P200 waveform and other positivedeflections, and that the processing of goal-directed information is associated with activationin two major brain areas, the TPJ and the mPFC.In order to validate these neural results asindicators of goal inferences, we calculated sev-eral correlations between the memory tasks andthese neural measures.
P200 and other positive ERP deflections
Our prediction was that the P200 is indicative ofgoal detection. Because we found no substantial
differences between the correlations of GC and GIinformation, for each participant, we computed anaverage score for the GC and GI conditions on allcued recall and sentence completion tasks. Wethen computed Pearson correlations betweenthese memory indices and the ERP peaks at eachinterval. The results are given in Table 3. Forspontaneous instructions, there are no significantpositive correlations (pB.05) between the mem-ory tasks and ERP deflections. In contrast, forintentional instructions, there were reliable posi-tive correlations between the memory measuresand ERP deflections, including the early P200,mainly at the Cz scalp location. Taken together,this indicates that the ERP deflections are validindicators of goal processing as measured bytraditional behavioral memory tasks.
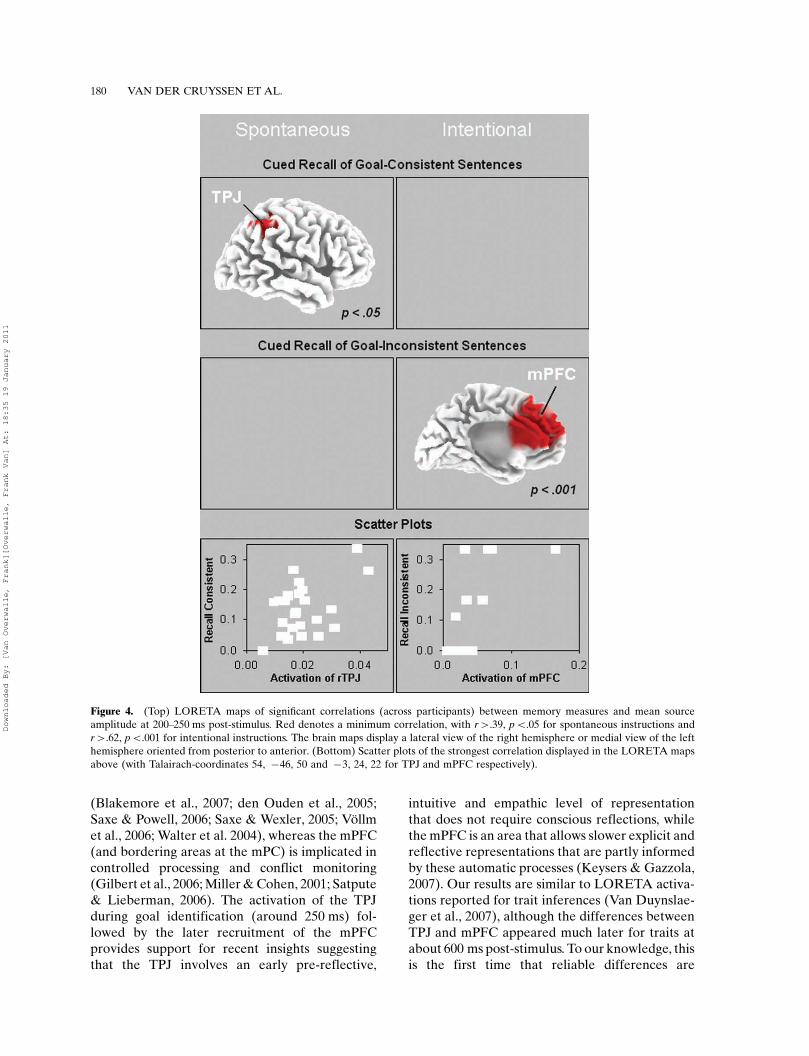
LORETA
Our prediction was that the LORETA activa-tion in the TPJ is mainly indicative of sponta-neous goal inference, while the mPFC reflectsfurther reflective processing of goal inferences. Toconfirm this statistically, we computed a Pearsoncorrelation across all participants between thememory tasks and the activation at all LORETAvoxels from the critical 200�250 ms post-stimulusinterval as presented above (i.e., GC minusNGC). To protect against multiple comparisons,we explored only the hypothesized brain areas(TPJ and mPFC) and used minimally a conven-tional .05 level. The results are depicted inFigure 4. Under SGI, cued recall of consistentinformation shows significant correlations withactivation in the TPJ, pB.05. Under IGI, cuedrecall of inconsistent information shows strongcorrelations in a large mPFC area, pB.001. Theseareas of significant correlation are somewhatmore dorsal than in Figure 2, and this might bedue to the fact that LORETAs extracted fromcorrelations on individual ERPs are probablysomewhat less reliable than LORETAs based onERPs averaged across all participants (as inFigure 2). With this caveat in mind, the resultsnevertheless seem to suggest that when people
Figure 2 (opposite). LORETA source analysis. The first two columns depict the amplitudes of Goal-Consistent conditions minus
No-Goal conditions under spontaneous and intentional instructions, averaged during consecutive post-stimulus time segments. The
maps are scaled with respect to their minimum and maximum amplitude. The third column depicts maps of t-values, pB.05
uncorrected, for the statistical comparison between spontaneous and intentional LORETA maps, with blue indicating greater
activation under intentional instructions. The brain maps are oriented from the posterior to the anterior cortex and display either a
medial view of the left hemisphere or a lateral view of the right or left (inserts) hemisphere. Talairach-coordinates (in x, y, z) of the
local maxima of significant differences between SGI and IGI: at 100�150 ms, �3, 38, 1; at 150�200 ms, �3, 45, 8; at 300�350 ms, 60,
�39, 36; at 350�400 ms, 53, �53, 22 and �59, �39, 36; at 400�450 ms, 46, �46, 22 and �59, �46, 22.
GOAL INFERENCES AND P200 177
Downloaded By: [Van Overwalle, Frank][Overwalle, Frank Van] At: 18:35 19 January 2011
are spontaneously inferring goals, better cued
recall is associated with more processing of
goal-consistent information in the TPJ. In con-
trast, for intentional goal instructions, a stronger
focus on inconsistencies in behavioral goals is
associated with more activation in the mPFC.
DISCUSSION
The most interesting result of this study is that
goal inferences are made relatively automatically
at the TPJ within 200 ms post-stimulus. Inten-
tional instructions to identify the goal further
recruit the mPFC and increase the chance that
the goal is remembered together with the beha-
vioral information. These findings are in line with
dual-process models which posit that automaticprocesses occur rapidly and may further informintentional decisions at later processing stages(Keysers & Gazzola, 2007; Satpute & Lieberman,2006; Smith & DeCoster, 2000). Although thesefindings certainly need further confirmation, ourstudy provides many other interesting data, whichare discussed below.
Neural indices of goal inference
The ERP waveforms revealed an early P200component that was significant in several chan-nels and for both spontaneous and intentionalinstructions, showing increased neural activationwhen the goal changed, and even more so when agoal was completely missing. There were alsoclear positive ERP deflections at later timeintervals, consistent with a P300, which weremore robust when the goal was missing or duringintentional processing. The P200 presumablyreflects increased attention and processing ofthe behavioral information because of the incon-sistency with the goal implied by the precedingbehaviors. The P200 was evident at the predictedfrontal and central scalp locations, and the P300at the predicted central and parietal midline sites.
The P300 findings are consistent with earlierERP research on trait inferences by Van Duyn-slaeger et al. (2007, 2008), who documentedidentification of traits only about 600 ms afterreading a critical trait-inconsistent word. Assuggested by earlier research, this late positivepotential reflects a deliberative context-updatingprocess by which an interpretation of the envir-onment is updated and consolidated in long-termmemory (Fabiani & Donchin, 1995; Fabiani,Karis, & Donchin, 1986). The P200 findings are
TABLE 2
Proportion of correct memory at various measures as a function of instruction and goal consistency
Spontaneous Intentional
Consistent Inconsistent Irrelevant Consistent Inconsistent Irrelevant
Cued recall
Goal-implying sentences 14a 04bd 01c 15a 07b 02cd
No-goal sentences 07a 00b 05a 09a 04a 00b
Sentence completion
Goal-implying sentences 29a 23a � 27a 24a �
Note: Means in a row sharing the same subscript do not differ significantly from each other according to a Fisher LSD test, pB
.05.
µV
Spontaneous Intentional0.0
0.5
1.0
1.5
2.0 left TPJ right TPJ mPC mPFC
Figure 3. Local maxima at each area of interest from the
LORETA source analysis at the critical 200�225 ms time
interval. TPJ�temporoparietal junction; mPFC�medial pre-
frontal cortex; mPC�medial paracentral cortex extending to
the posterior mPFC. Talairach-coordinates (in x, y, z) of local
maxima are for SGI at rTPJ, 53, �53, 15; lTPJ, �45, �67, 15;
mPC, 4, �4, 57; mPFC, 4, 45, 15; and for IGI at rTPJ, 53, �53,
15; lTPJ, �52, �67, 8; mPC, �3, �11, 50; mPFC, 4, 52, 1.
178 VAN DER CRUYSSEN ET AL.
Downloaded By: [Van Overwalle, Frank][Overwalle, Frank Van] At: 18:35 19 January 2011

more surprising. Such early differences based on
inconsistent information (Clark & Hillyard, 1996;
Peters et al., 2005) were not expected on the basis
of earlier trait inference research (Bartholow
et al., 2001, 2003; Van Duynslaeger et al., 2007,
2008), and strongly suggest that goals are inferred
automatically. Indeed, assuming that the semantic
interpretation of common (i.e., high-frequency)
words takes about 170 ms (Dambacher et al.,
2006), it is fascinating to see how extremely fast
goals are identified, requiring a mere extra 50 ms
processing time. Given the lack of P200 deflec-
tions in the no-goal control conditions, we can
rule out that goal inferences are simply part of
semantic comprehension, and state that they
mark a distinct social inference process. The fact
that the P200 was strongest for goal-irrelevant or
no-goal sentences, which are relatively neutral to
positive in affective content, also rules out
negative affect as an alternative explanation for
our P200 or P300 deflections (e.g., Correll et al.,
2006; Eimer & Holmes, 2002; Eimer et al., 2003;
Keil et al., 2002; Kubota & Ito, 2007).The fast identification of goals is consistent
with the growing evidence that goal identification
on the basis of behavioral information is part of
an automatic mirror system involving the TPJ, so
that people immediately recognize intentions of
others by their similarity with own behavioral
intentions (cf. Keysers & Perrett, 2004; Van
Overwalle, 2008). Behavioral studies have also
demonstrated that people can infer goals auto-
matically without awareness or intent (Hassin
et al., 2005) and even tend to adopt automatically
the perceived goals of others in their own goalpursuit (Aarts, Gollwitzer & Hassin, 2004).
A more dramatic difference between proces-sing modes is seen when the LORETA activationin different brain areas is considered. The resultsshow that at the very early 100�200 ms interval,spontaneous goal identification activates the righttemporoparietal junction (TPJ), whereas inten-tional goal instructions generate stronger activa-tion at the medial prefrontal cortex (mPFC) and,nonsignificantly, areas at the medial paracentralcortex (mPC). Perhaps this differential activationindexes different attentional mechanisms or at-tentional boosts in different brain areas during apreparatory phase when ‘‘searching’’ for mean-ingful information (SGI) or goals (IGI). After thispreparatory stage, and especially in the 225�300 ms interval, the most active brain areairrespective of instruction is the TPJ. This isconsistent with our prediction that the TPJdominates during early and automatic goal pro-cessing, and therefore reduces differences be-tween spontaneous and intentional mind sets.Afterwards, further processing mainly activatesthe mPFC, first beginning under intentionalinstructions (at 300 ms) and somewhat later underspontaneous instructions (at 350 ms), probablyreflecting conscious reasoning about the goal orother task-related information.
Overall, the results are consistent with earlierfMRI research documenting that the TPJ isrecruited during implicit goal identification whenobserving human-like goal-directed movement(Martin & Weisberg, 2003; Ohnishi et al., 2004;Schultz et al., 2004) or goal-directed behavior
TABLE 3
Pearson correlations (across participants) between memory measures and positive ERP-deflections at different scalp sites and
time segments (including the P200 in the 0�300 ms interval)
Spontaneous Intentional
Interval Fz Cz Pz Fz Cz Pz
Goal-cued recall of (in)consistent sentences
50�300 0.19 �0.14 0.06 0.32 0.55*** 0.23
300�450 0.21 0.13 0.20 0.53*** 0.41** 0.21
450�650 0.10 0.13 0.24 0.30 0.38** 0.34
650�1000 �0.04 0.14 0.19 0.49** 0.51*** 0.52***
Sentence completion of (in)consistent sentences
50�300 �0.28 �0.19 0.16 0.05 0.48** 0.00
300�450 �0.02 �0.08 0.35* 0.44** 0.48** 0.18
450�650 �0.11 0.11 0.34* 0.02 0.35* 0.23
650�1000 �0.21 0.16 0.32 0.42** 0.49** 0.41**
Notes: For cued recall and sentence completion, the percentage correct memory for goal-consistent and goal-inconsistent
sentences was averaged. *pB.10; **pB.05; ***pB.01.
GOAL INFERENCES AND P200 179
Downloaded By: [Van Overwalle, Frank][Overwalle, Frank Van] At: 18:35 19 January 2011
(Blakemore et al., 2007; den Ouden et al., 2005;Saxe & Powell, 2006; Saxe & Wexler, 2005; Vollmet al., 2006; Walter et al. 2004), whereas the mPFC(and bordering areas at the mPC) is implicated incontrolled processing and conflict monitoring(Gilbert et al., 2006; Miller & Cohen, 2001; Satpute& Lieberman, 2006). The activation of the TPJduring goal identification (around 250 ms) fol-lowed by the later recruitment of the mPFCprovides support for recent insights suggestingthat the TPJ involves an early pre-reflective,
intuitive and empathic level of representationthat does not require conscious reflections, whilethe mPFC is an area that allows slower explicit andreflective representations that are partly informedby these automatic processes (Keysers & Gazzola,2007). Our results are similar to LORETA activa-tions reported for trait inferences (Van Duynslae-ger et al., 2007), although the differences betweenTPJ and mPFC appeared much later for traits atabout 600 ms post-stimulus. To our knowledge, thisis the first time that reliable differences are
Figure 4. (Top) LORETA maps of significant correlations (across participants) between memory measures and mean source
amplitude at 200�250 ms post-stimulus. Red denotes a minimum correlation, with r�.39, pB.05 for spontaneous instructions and
r�.62, pB.001 for intentional instructions. The brain maps display a lateral view of the right hemisphere or medial view of the left
hemisphere oriented from posterior to anterior. (Bottom) Scatter plots of the strongest correlation displayed in the LORETA maps
above (with Talairach-coordinates 54, �46, 50 and �3, 24, 22 for TPJ and mPFC respectively).
180 VAN DER CRUYSSEN ET AL.
Downloaded By: [Van Overwalle, Frank][Overwalle, Frank Van] At: 18:35 19 January 2011

reported between brain areas after spontaneousand intentional goal inferences.
Several authors have recently questioned therole of the TPJ and argued that it is not selectivefor social cognition, but has evolved from per-ceptual mechanisms that subserve non-socialattentional processes. In a meta-analysis of brainimaging studies (Decety & Lamm, 2007) andwithin single participants using fMRI (Mitchell,2008), a considerable overlap was found in TPJactivation across social inference tasks and non-social cue-orienting tasks, suggesting that the twotasks share important computational functionsthat generalize across different content domains.The social tasks require mentalizing about po-tentially false beliefs and agency of goal-directedbehaviors, while the orienting tasks involvereorienting one’s attention after incorrect cuesor unexpected changes about upcoming events. Ithas been proposed that a common mechanismunderlying TPJ activation is the detection ofinconsistencies in external sensory events (Dec-ety & Lamm, 2007) or, alternatively, the detec-tion of a possible trajectory and end-state ofbehaviors (Van Overwalle, 2008). In support ofthis latter view, from a visual perceptive, not onlyorienting but also social judgments such asidentifying the goal of a movement, taking theviewpoint of another person or deciding aboutthe agent of a behavior often require that weestimate the most likely trajectory of a targetmovement. Questions have also arisen concern-ing the role of mirror neurons in the TPJ, sincemost research identifying mirror neurons hasrevealed evidence for the involvement of theinferior parietal lobe, an area somewhat dorsalbut not identical to the TPJ (Van Overwalle,2008). Interpretation difficulties also exist withrespect to the role of the mPFC, because thisarea may not only serve the integration of socialinformation over time to allow for trait infer-ences but, more drastically, it may altogetherstore these enduring trait or script inferences(Huey, Krueger & Grafman, 2006; Van Over-walle, 2008). Although these questions clearlyopen up the interpretation of the functionsunderlying the TPJ and mPFC, this does notinvalidate the present finding that these areas areinvolved in social judgment.
Although localization of ERP activity usingLORETA is less precise and limited to the cortexin comparison with fMRI, its millisecond timeprecision offers more insight in the time courseof brain activation. Currently, such processing
differences in the time domain cannot be detectedwith the fMRI technique because blood deoxy-genation, on which it rests, requires severalseconds. Other techniques such as magneto-elec-troencephalogram (MEG) might provide a moretime-sensitive resolution combined with a morereliable brain location.
Memory validation
To validate that our neural measures reflect themental content of goals, we borrowed severalmemory measures from earlier behavioral socialcognition research. We presented a large amountof behavioral sentences to minimize strategicretrieval strategies given spontaneous instruc-tions, in line with recent spontaneous inferenceresearch using memory tasks (e.g., Todorov &Uleman, 2002).
Consistent with our predictions, the meanperformance on the memory measures was highlyparallel between spontaneous and intentionalgoal inference, and suggests the involvement ofbroadly similar encoding and storage processes.Cued recall was stronger for goal-consistentbehavior than for inconsistent or irrelevant beha-vior irrespective of instruction, further confirmingthat goals were encoded and stored in memory.Similar findings were observed in recent ERPresearch involving trait inferences by VanDuynslaeger et al. (2007, 2008). It is unlikelythat the absence of memory differences is due to alack of statistical power, as the number ofparticipants in each group (�25) was consider-ably larger than earlier ERP research (i.e.,Bartholow et al., 2001, 2003, 2006). One differ-ence with trait research*which we anticipated*is that memory in the sentence completion task isnot higher for inconsistent behaviors, becausegoals are more flexible than traits, so that actorscan easily switch from one goal to another with-out being really incongruent in their overallbehavioral intentions or characteristics.
Correlations with the memory measures es-tablish more firmly the reliability of our ERPs asindices of goal inference. Although correlationsbetween the P200 and memory measures failedto be significant under spontaneous instructions,we observed reliable correlations between LOR-ETA brain activation (at the TPJ and mPFC)and goal-cued recall under both instructions,indicating that better memory for the goalsimplied by behavioral information is associated
GOAL INFERENCES AND P200 181
Downloaded By: [Van Overwalle, Frank][Overwalle, Frank Van] At: 18:35 19 January 2011

with increased brain activation in these two areasknown to be critical in mentalizing. It is inter-esting to note that LORETA activations in theTPJ during spontaneous instructions were corre-lated with increased recall of consistent beha-viors, whereas activations in the mPFC duringintentional processing were correlated with en-hanced cued recall of inconsistent behaviors,indicating that spontaneous versus intentionalinstructions direct attention to different aspectsof the behavior that are processed in differentareas of the brain. That no memory correlationswere found with the P200 under spontaneousinstructions contradicts the finding by Van Duyn-slaeger et al. (2007, 2008) that these correlationswere pronounced under spontaneous trait in-structions. This difference is probably due tothe early and automatic nature of goal inferencesas opposed to more deliberative trait inferences.Assuming that goals are inferred automaticallywithout any conscious thought, it seems plausiblethat little correlations are found with memorymeasures. Perhaps goals are so routinely de-tected without extra mental effort that they arenot remembered very well. In the light of this,the significant correlations with the LORETAactivation are therefore all the more remarkable.Many fMRI studies in this field do not corrobo-rate the psychological validity of their neurallocalizations with memory measures, and so theycannot speak to this issue.
CONCLUSION
In contrast to trait inferences, which are identifiedat a late 600 ms post-stimulus stage, the P200deflections in this study provide evidence on therapid and automatic nature of the neural compu-tations involved in the identification of goals. TheP200 shows many parallels between spontaneousand intentional modes of goal identification,which are corroborated by parallels in TPJactivation during goal extraction. In contrast,before and after goal identification, there isgreater activation of the TPJ during spontaneousprocessing, and more activation in the mPFCduring intentional processing, in line with fMRIresearch on the proposed role of the TPJ inspontaneous attributions of intentions and of themPFC in reflective reasoning (Keysers & Gaz-zola, 2007). Our results are highly similar toearlier ERP research on trait inferences thatrevealed a similar division of brain activation
for spontaneous (TPJ) and intentional (mPFC)processes, but appearing later at about 600 ms(e.g., Van Duynslaeger et al., 2007). The similarityof brain areas recruited for social inferences, butat a different timing for goals and traits, is aunique contribution of recent neurophysiologicalresearch. Pathologies of person inference, such asin autism and paranoia, might benefit greatlyfrom the insights gained from this and similarstudies on the neural correlates of spontaneousand intentional goal detection.
Manuscript received 13 February 2008
Manuscript accepted 29 May 2008
First published online 19 December 2008
REFERENCES
Aarts, H., Gollwitzer, P. M., & Hassin, R. R. (2004).Goal contagion: Perceiving is for pursuing. Journalof Personality and Social Psychology, 87, 23�37.
Amodio, D. M., & Frith, C. D. (2006). Meeting ofminds: The medial frontal cortex and social cogni-tion. Nature Reviews Neuroscience, 7, 268�277.
Bartholow, B. D., Dickter, C. L., & Sestir, M. A. (2006).Stereotype activation and control of race bias:Cognitive control of inhibition and its impairmentby alcohol. Journal of Personality and Social Psy-chology, 90, 272�286.
Bartholow, B. D., Fabiani, M., Gratton, G., & Betten-court, B. A. (2001). A psychophysiological examina-tion of cognitive processing of and affectiveresponses to social expectancy violations. Psycholo-gical Science, 12, 197�204.
Bartholow, B. D., Pearson, M. A., Gratton, G., &Fabiani, M. (2003). Effects of alcohol on personperception: A social cognitive neuroscience ap-proach. Journal of Personality and Social Psychol-ogy, 85, 627�638.
Blakemore, J. S., den Ouden, H., Choudhury, S., &Frith, C. (2007). Adolescent development of theneural circuitry for thinking about intentions. SocialCognitive Affective Neuroscience, 2, 130�139.
Cacioppo, J. T., Crites, S. L., Berntson, G. G., & Coles,M. G. H. (1993). If attitudes affect how stimuli areprocessed, should they not affect the event-relatedbrain potential? Psychological Science, 4, 108�112.
Cacioppo, J. T., Crites, S. L., Jr., Gardner, W. L., &Berntson, G. G. (1994). Bioelectrical echoes formevaluative categorizations: I. A late positive brainpotential that varies as a function of trait negativityand extremity. Journal of Personality and SocialPsychology, 67, 115�125.
Cacioppo, J. T., Gardner, W. L., & Berntson, G. G.(1999). The affect system has parallel and integra-tive processing components: Form follows function.Journal of Personality and Social Psychology, 76,839�855.
Carretie, L., Mercado, F., Tapia, M., & Hinojosa, J. A.(2001). Emotion, attention, and the ‘negativity bias’,
182 VAN DER CRUYSSEN ET AL.
Downloaded By: [Van Overwalle, Frank][Overwalle, Frank Van] At: 18:35 19 January 2011

studied through event-related brain potentials. Inter-national Journal of Psychophysiology, 41, 75�85.
Clark, V. P., & Hillyard, S. A. (1996). Spatial selectiveattention affects early extrastriate but not striatecomponents of the visual evoked potential. Journalof Cognitive Neuroscience, 8, 387�402.
Correll, J., Urland, G. R., & Ito, T. A. (2006). Event-related potentials and the decision to shoot: The roleof threat perception and cognitive control. Journalof Experimental Social Psychology, 42, 120�128.
Dambacher, M., Kliegl, R., Hofmann, M, & Jacobs, A.M. (2006). Frequency and predictability effects onevent-related potentials during reading. Brain Re-search, 1084, 89�103.
Decety, J., & Lamm, C. (2007). The role of the righttemporoparietal junction in social interaction: Howlow-level computational processes contribute tometa-cognition. Neuroscientist, 13, 580�593.
den Ouden, H. E. M, Frith, U., Frith, C., & Blakemore,S.-J. (2005). Thinking about intentions. NeuroImage,28, 787�796.
Dunn, B. R., Dunn, D. A., Languis, M., & Andrews, D.(1998). The relation of ERP components to complexmemory processing. Brain and Cognition, 36, 355�376.
Eimer, M., & Holmes, A. (2002). An ERP study on thetime course of emotional face processing. NeuroRe-port, 13, 427�431.
Eimer, M., Holmes, A., & McGlone, F. P. (2003). Therole of spatial attention in the processing of facialexpression: An ERP study of rapid brain responsesto six basic emotions. Cognitive, Affective. Beha-vioral Neuroscience, 3, 97�110.
Esslen, M., Pascual-Marqui, R. D., Hell, D., Kochi, K.,& Lehmann, D. (2004). Brain areas and time courseof emotional processing. NeuroImage, 21, 1189�1203.
Fabiani, M., & Donchin, E. (1995). Encoding processesand memory organization: A model of the vonRestorff effect. Journal of Experimental Psychology:Learning, Memory and Cognition, 21, 224�240.
Fabiani, M., Karis, D., & Donchin, E. (1986). P300 andrecall in an incidental memory paradigm. Psycho-physiology, 23, 298�308.
Fyock, J., & Stangor, C. (1994). The role of memorybiases in stereotype maintenance. British Journal ofSocial Psychology, 33, 331�343.
Gilbert, S. J., Spengler, S., Simons, J. S., Steele, D.,Stephen, M., Lawrie, S.M., et al. (2006). Functionalspecialization within rostral prefrontal cortex (Area10): A meta-analysis. Journal of Cognitive Neu-roscience, 18, 932�948.
Hassin, R. R., Aarts, H., & Ferguson, M. J. (2005).Automatic goal inferences. Journal of ExperimentalSocial Psychology, 41, 129�140.
Huey, E. D., Krueger, F., & Grafman, J. (2006).Representations in the human prefrontal cortex.Association for Psychological Science, 15, 167�171.
Ito, T. A., & Urland, G. R. (2003). Race and gender onthe brain: Electrocortical measures of attention tothe race and gender of multiply categorizableindividuals. Journal of Personality and Social Psy-chology, 85, 616�626.
Ito, T. A., & Urland, G. R. (2005). The influence ofprocessing objectives on the perception of faces: AnERP study of race and gender perception. Cogni-tive, Affective. and Behavioral Neuroscience, 5, 21�36.
Joyce, C. A., Gorodnitsky, I. F., & Kutas, M. (2004).Automatic removal of eye movement and blinkartifacts from EEG data using blind componentseparation. Psychophysiology, 41, 313�325.
Keil, A., Bradley, M. M., Hauk, O., Rockstroh, B.,Elbert, T., & Lang, P. J. (2002). Large-scale neuralcorrelates of affective picture processing. Psycho-physiology, 39, 641�649.
Keysers, C., & Gazzola, V. (2007). Integrating simula-tion and theory of mind: From self to socialcognition. Trends in Cognitive Sciences, 11, 194�196.
Keysers, C., & Perrett, D. I. (2004). Demystifying socialcognition: A Hebbian perspective. Trends in Cogni-tive Sciences, 8, 501�507.
Kubota, J. T., & Ito, T. A. (2007). Multiple cues in socialperception: The time course of processing race andfacial expression. Journal of Experimental SocialPsychology, 43, 738�752.
Luck, S. J., & Hillyard, S. A. (1994). Electrophysiolo-gical correlates of feature analysis during visualsearch. Psychophysiology, 31, 291�308.
Martin, A., & Weisberg, J. (2003). Neural foundationsfor understanding social and mechanical concepts.Cognitive Neuropsychology, 20, 575�587.
Miller, E. K., & Cohen, J. D. (2001). An integrativetheory of prefrontal cortex function. Annual Reviewof Neuroscience, 24, 167�202.
Mitchell, J. P. (2008). Activity in right temporo-parietaljunction is not selective for theory of mind. CerebralCortex, 18, 262�271.
Mitchell, J. P., Banaji, M. R., & Macrae, C. N. (2005).The link between social cognition and self-referen-tial thought in the medial prefrontal cortex. Journalof Cognitive Neuroscience, 17, 1306�1315.
Nichols, T. E., & Holmes, A. P. (2002). Nonparametricpermutations tests for functional neuroimaging: Aprimer with examples. Human Brain Mapping, 15,1�25.
Ohnishi, T., Moriguchi, Y., Matsuda, H., Mori, T.,Hirakata, M., Imabayashi, E., et al. (2004). Theneural network for the mirror system and mentaliz-ing in normally developed children: An fMRI study.NeuroReport, 15, 1483�1487.
Osterhout, L., Bersick, M., & McLaughlin, J. (1997).Brain potentials reflect violations of gender stereo-types. Memory & Cognition, 25, 273�285.
Pascual-Marqui, R. D. (1999). Review of methods forsolving the EEG inverse problem. InternationalJournal of Bioelectromagnetism, 1, 75�86.
Pascual-Marqui, R. D., Michel, C. M., & Lehmann, D.(1994). Low resolution electromagnetic tomogra-phy: A new method for localizing electrical activityin the brain. International Journal of Psychophysiol-ogy, 18, 49�65.
Peters, J., Suchan, B., Zhang, Y., & Daum, I. (2005).Visuo-verbal interactions in working memory: Evi-dence from event-related potentials. Cognitive BrainResearch, 25, 406�415.
GOAL INFERENCES AND P200 183
Downloaded By: [Van Overwalle, Frank][Overwalle, Frank Van] At: 18:35 19 January 2011

Raney, G. E. (1993). Monitoring changes in cognitiveload during reading: An event-related brain poten-tial and reaction time analysis. Journal of Experi-mental Psychology: Learning, Memory, andCognition, 19, 51�69.
Reeder, G. D., & Brewer, M. B. (1979). A schematicmodel of dispositional attribution in interpersonalperception. Psychological Review, 86, 61�79.
Satpute, A. B., & Lieberman, M. D. (2006). Integratingautomatic and controlled processes into neurocog-nitive models of social cognition. Brain Research,1079, 86�97.
Saxe, R., & Powell, L. J. (2006). It’s the thought thatcounts: Specific brain regions for one component oftheory of mind. Psychological Science, 17, 692�699.
Saxe, R., & Wexler, A. (2005). Making sense of anothermind: The role of the right temporo-parietal junc-tion. Neuropsychologia, 43, 1391�1399.
Schultz, J., Imamizu, H., Kawato, M., & Frith, C.D.(2004). Activation of the human superior temporalgyrus during observation of goal attribution byintentional objects. Journal of Cognitive Neu-roscience, 16, 1695�1705.
Smith, M. E. (1993). Neurophysiological manifestationof recollective experience during recognition mem-ory judgments. Journal of Cognitive Neuroscience, 5,1�13.
Smith, E. R., & DeCoster, J. (2000). Dual-processmodels in social and cognitive psychology: Concep-tual integration and links to underlying memorysystems. Personality and Social Psychology Review,4, 108�131.
Stangor, C., & McMillan, D. (1992). Memory forexpectancy-congruent and expectancy-incongruentinformation: A review of the social and socialdevelopmental literatures. Psychological Bulletin,111, 42�61.
Stern, R. M., Ray, W. R., & Quigley, K. S. (2001).Psychophysiological recording (2nd ed.). New York:Oxford University Press.
Todorov, A., Gobbini, M. I., Evans, K. K., & Haxby, J.V. (2007). Spontaneous retrieval of affective person
knowledge in face perception. Neuropsychologia,45, 163�173.
Todorov, A., & Uleman, J. S. (2002). Spontaneous traitinferences are bound to actors’ faces: Evidence froma false recognition paradigm. Journal of Personalityand Social Psychology, 83, 1051�1064.
Uleman, J. S., Blader, S. L., & Todorov, A. (2005).Implicit impressions. In R. R. Hassin, J. S. Uleman,& J. A. Bargh (Eds.), The new unconscious (pp. 362�392). New York: Oxford University Press.
Van Duynslaeger, M., Sterken, C., Van Overwalle, F., &Verstraeten, E. (2008). EEG components of sponta-neous trait inferences. Social NeuroScience, 3, 164�177.
Van Duynslaeger, M., Van Overwalle, F., & Verstrae-ten, E. (2007). Electrophysiological time course andbrain areas of spontaneous and intentional traitinferences. Social Cognitive and Affective Neu-roScience, 2, 174�188.
Van Overwalle, F. (2008). Social cognition and thebrain: A meta-analysis. Human Brain Mapping, inpress.
Vollm, B. A., Taylor, A. N. W., Richardson, P.,Corcoran, R., Stirling, J., McKie, S., et al. (2006).Neuronal correlates of theory of mind and empathy:A functional magnetic resonance imaging study in anonverbal task. NeuroImage, 29, 90�98.
Walter, H., Adenzato, M., Ciaramidaro, A., Enrici, I.,Pia, L., & Bara, B. G. (2004). Understandingintentions in social interaction: The role of theanterior paracingulate cortex. Journal of CognitiveNeuroscience, 16, 1854�1863.
Winter, L., & Uleman, J. S. (1984). When are socialjudgments made? Evidence for the spontaneousnessof trait inferences. Journal of Personality and SocialPsychology, 47, 237�252.
Ybarra, O. (2002). Naıve causal understanding ofvalenced behaviors and its implications for socialinformation processing. Psychological Review, 128,421�441.
184 VAN DER CRUYSSEN ET AL.
Downloaded By: [Van Overwalle, Frank][Overwalle, Frank Van] At: 18:35 19 January 2011