stabilization of affinity-tagged recombinant protein during/after its production in a cell-free...
TRANSCRIPT

JOURNAL OF BIOSCIENCE AND BIOENGINEERING Vol. 95, No. 3,209-214. 2003
Stabilization of Affinity-Tagged Recombinant Protein during/after Its Production in a Cell-Free System
Using Wheat-Germ Extract YASUAKI KAWARASAKI , 1~* YASUHIRO YAMADA, a MAKI ICHIMORI, I
T O M O Y A SHINBATA, 2 KATSUNORI KOHDA, 3 HIDEO NAKANO, 1 AND TSUNEO Y A MA N E 1
Laboratory of Molecular Biotechnology, Division of Molecular and Cellular Mechanisms, Graduate School of Bio and Agricultural Sciences, Nagoya University, Furo-cho, Chikusa-ku, Nagoya 464-8601, Japan, 1
Nippon Flour Mills, Co. Ltd., 2114-2 Nurumizu, AtugL Kanagawa 243-0033, Japan, 2 and Toyota Central R&D Laboratories Inc., Nagakute, Aichi 480-1192, Japan 3
Received 22 July 2002/Accepted 21 October 2002
We found that the affinity tag fused to the carboxyl (C-) terminal of a single-chain Fv (seFv) antibody was proteolytically degraded in a wheat germ cell-free protein synthesis system. The ad- dition of two extra residues of glycine to the tail of the cMyc tag significantly increased the stabil- ity of the tag, suggesting that wheat endogenous carboxypeptidase(s) play a primary role in the C- terminal tag-specific degradation. In addition to the modification of the tag sequence, addition of diisopropyl fluorophosphate, which is known as an inhibitor of carboxypeptidases, prevented the cMyc tag sequence degradation. The effects of other protease inhibitors on the translation reac- tion and stability of the synthesized protein are also reported.
[Key words: cell-free protein synthesis system, proteolytic degradation, carboxypeptidase, affinity tag, protease inhibitors]
A combination of the cell-free expression of an affinity- tagged recombinant gene with an assay system based on the tag-specific antibody facilitates rapid analysis of gene prod- uct function, or rapid purification of synthesized protein. Recent advances in the productivity of cell-free protein syn- thesis systems (1-3), including the system using wheat- germ extract (4), have facilitated the rapid preparation of gene products in substantial amounts, sufficient for a variety of biochemical analyses such as protein-protein interac- tions. These highly productive cell-free systems have been generally accepted, and are currently commercially avail- able.
The productivity of the cell-free systems, however, has been evaluated using canonical reporter genes, such as those encoding dihydrofolate reductase (DHFR), chlorampheni- col acetyltransferase, and green fluorescent protein. The products of these genes are considered to be robust in the cell-free extracts as well as in the cytoplasm of living cells. Although a list of proteins which can be synthesized in vitro is available on manufacturers' websites (e.g., http://www. roche-applied-science.corn/), the stability of synthesized proteins has rarely been discussed. In particular, the stability of the affinity tag sequence fused to the heterologous pro-
* Corresponding author, e-mail: [email protected] phone: +81-(0)52-789-4144 fax: +81-(0)52-789-4145
Present address: Georgiou Laboratory, the Institute for Cellular and Molecular Biology, University of Texas at Austin, Austin, Texas 78712-1095, USA.
209
tein remains unknown, despite its critical irn~portance in de- tection (5), separation (6), or affinity immobilizalion (7) of the products.
We discovered that in vitro synthesized polypeptides which have a His 6 affinity tag at their carboxyl (C-) terminal are hardly detected in immunochemical assays using tag- specific antibodies. This, however, was not obse~eed when the tag is located at the N-terminal of the same gene prod- uct.
In this study, we performed a series of analyses concern- ing the stability of tag peptide sequences in the cell-free system using wheat-germ extract (8). We found that the poor signal intensity of the C-terminal-tagged polypeptides was due to C-terminal-specific degradation caused by endo- genous carboxypeptidase(s). In addition, we demonstrated how to prevent the degradation of the tag sequence. The manner of degradation of the synthesized polypeptides will also be discussed in this report.
MATERIALS AND METHODS
Template constructions The structures of the tagged scFv gene constructs are summarized in Fig. 1. Plasmid pMID2-ahSA, which carried the T7 promoter, a cap-independent translation en- hancing sequence TE(37-65) (9), the anti-human serum albumin single chain Fv-(anti-HSA-scFv, patent no. P2000-139460A) cod- ing region, a C-terminal His 6 tag sequence and the T7 terminator, was constructed using standard procedures including PCR am- plification and conventional genetic manipulations (10). Plasmid pYEx2-ahSA was prepared by inserting the Ncol and NotI frag-

210 KAWARASAKI ET AL. J. BIOSCI. B:OENG.,
TE(37-65)
+1
TE(37-65)
~ PCR
Itis 6
, ' ) k , cMyc GIyGIy
cMyc-G0 cMyc-G 1
cMyc-G2 EcoRI
EcoRI
Eco RI
FIG. 1. Schematic of gene structures used in this study. Each scFv mRNA for cell-free protein synthesis was prepared by in vitro tran- scription using pMID2-ahSA (for scFv-His6) or PCR products from pYEx2-ahSA (for scFv-cMyc-G0, -G1, or -G2) as a template. For each template, an open arrowhead (PTT), a white bar, a closed box and an open box represent the T7 promoter sequence, 5' cap-independent trans- lation enhancing sequence (TE(37-65) [9]), single-chain Fv-coding re- gion (derived from gene fragments of the anti-human serum albumin (HSA) monoclonal antibody, PAT: P2000-139460A), and the affinity tag sequence, respectively. The hatched boxes in scFv-cMyc-G1 and -G2 represent the additional glycine residue(s).
merits of pMID2-ahSA into the expression vector pYEx2 (pRSETB-derived plasmid vector, unpublished work) to obtain the cMyc-tagged scFv template. The resulting pYEx2-ahSA plasmid has the T7 promotel, TE(37-65), the scFv-coding region fused to the cMyc tag-coding sequence at its 3'-end with the additional two residues of glycine, and the T7 termination sequence. The DNA templates for cMyc-tagged scFv with various additional glycine residues (designated as anti-HSA scFv-cMyc-G0, -G1, and -G2, according to the number of added Gly residues) were prepared by PCR. The PCR amplifications were carried out using a set of specific primers: rset-uT7 (ATTCATTAATGCAGATCTCGATCC, which anneals upstream of the T7 promoter sequence) and cMyc- GO (gggaattcctatccttaTAAATCTTCTTCACTTAT, for scFv-cMyc- G0), cMyc-G1 (gggaattcctattaTCC TAAATCTTCTTCACTTAT, for scFv-cMyc-G1), or cMyc-G2 (gggaattcctaTCCTCC TAAATCTTC TTCACTTAT, for scFv-cMyc-G2). The uppercase and underlined letters in the above three primers represent the sequence comple- mentary to the cMyc sequence and termination codon, respec- tively. The sequences for the additional Gly residues are repre- sented in italicized letters.
In vitro expression of affinity-tagged genes In vitro tran- scription of the tagged scFv genes was carried out using the MEGA script in vitro transcription kit (Ambion, Austin, TX, USA) according to the manufacturer's protocol. In the case of PCR- amplified templates such as anti-HSA-scFv-cMycG0, -cMycG1, or -cMycG2, the PCR product was digested with EcoRI at a unique site located downstream of the termination codon (depicted in Fig. 1), then used as a template for transcription. Cell-free protein syn- thesis using wheat-germ extract was carried out as reported pre- viously (8). The wheat germ for the extract with high translation efficiency was prepared from domestically harvested wheat seeds in Nippon Flour Mills (Kanagawa).
Degradation of translation product The translation prod- ucts were specifically labeled with [35S]-methionine (Amersham Biosciences, Piscataway, N J, USA), then analyzed by SDS-PAGE
and autoradiography. The effect of a protease inhibitor on the translation reaction was tested using dhfr mRNA as a template, and evaluated by quantifying the enzymatic activity of the synthe- sized DHFR (8).
Affinity tag degradation The cell-free translation reaction mixture (5 ~d/lane) was subjected to SDS-PAGE, then electro- transferred onto a nitrocellulose membrane. The tagged scFv pro- tein was detected by the anti-His antibody (Amersham Bioscience, for the C-His tag) or anti cMyc IgG (Roche Diagnostic, Indiana- polis, IN, USA, for the cMyc tag): The bands representing reaction of proteins with the tag-specific antibody were then visualized us- ing horseradish peroxidase-conjugated anti-mouse IgG rabbit IgG with the ECL Western blotting detection kit (Amersham Bio- sciences) or VectorStain Western blotting detection kit (Vector Laboratory, Burlingame, CA, USA). The intensity of the band was analyzed by a bioluminescence analysis system (ATTO, Tokyo).
RESULTS AND DISCUSSION
C-terminal tag-specific degradation in a cell-free pro- tein synthesis system The affinity tag fused at the C- terminal of the recombinant protein was hardly detected by immunoblot analysis using the tag-specific monoclonal anti- body, despite a sufficient amount o f polypeptide o f the cor- rect size being synthesized. As an example, the results o f the in vitro expression and analysis o f affinity-tagged single chain Fv against human serum albumin (anti-HSA scFv) is shown in Fig. 2. The scFv protein fused to six residues of histidine (-HHHHHH-OH), or to the cMyc affinity tag se- quence (-EQKLISEEDL-OH) at the C-terminal was synthe- sized in the presence o f [3:S]-methionine. The translation product was detected as a single band upon SDS-PAGE (Fig. 2A). The mobility o f the product was consistent with that deduced from its amino acid sequence (30 kDa). Fur- ther incubation (for 6 h) o f the product in the reaction mix- ture after stopping translation by adding puromycin affected neither the mobility nor intensity o f the band. This result in- dicates that the synthesized polypeptide is apparently robust in the reaction mixture. The Western blot analysis of the same proteins using tag-specific antibodies, however, re- sulted in significantly reduced or poor signal intensities (Fig. 2B).
The weak intensity o f the signals was not due to the in- trinsic instabilities o f the affinity tags, since the same His 6 tag located at the N-terminal o f target proteins were clearly detected, as reported previously (9). The results suggest that the affinity tag is labile when it is located at the C-terminal of the target proteins.
The signal intensity o f the C-terminal cMyc tag was, however, markedly improved when two glycine residues were added onto the tag sequence (-EQKLISEEDLOG-OH, where the underlined sequence represents the additional glycine residues), whereas a single glycine residue was not sufficient (Fig. 2B). As shown in Fig. 2A, changes in the mRNA sequence at the C-terminal coding region did not affect the yield o f the translation products. Therefore, the increase in the signal intensity in the Western blotting analy- sis was not due to an increase in the amount o f the synthe- sized protein, but due to the stabilization of the affinity tag.
It should be noted here that the results shown in Fig. 2B were obtained by exposing the blotted membrane to the sub-

VOL. 95, 2003 TAG STABILIZATION IN A CELL-FREE EXPRESSION SYSTEM 211
A His 6 cMycG0 cMycGlcMycG2
M 0 6 0 6 0 6 0 6 c
......
t 4 . 3 ~
B
81.8 68'4 55.0
41,6
His 6 cMycG0 cMycG1 cMycG2 M 0 6 0 6 0 6 0 6
FIG 2. Carboxyl-terminal tag-specific degradation during/after cell- free protein synthesis. Stabilities of the synthesized seFv proteins and their C-terminal alTmity tags were analyzed by SDS-PAGE/autoradio- graphy (A) and SDS-PAGE/Western blotting using affinity tag-spe- cific monoclonal antibodies (B). The affinity tags fused to the scFv proteins are indicated on the top of the images. Arrows on the right of the images indicate the migration point of the synthesized scFv pro- tein. The number on each lane indicates the incubation time (h) after stopping of translation by the addition of puromycin. Lanes M and C contain molecular size markers and a sample of the cell-free protein synthesis reaction without the addition of exogenous mRNA, respec- tively. The numbers on the left of the size markers indicate the molec- ular masses of the size markers (kDa). The detection of the tagged polypeptides on the blotted membrane (B) was carried out using a hy- persensitive chemical luminescence detection kit (Vector Laboratory) and a longer (30 min) reaction time.
strate solution for a longer period (30 min) to detect the very weak signals of anti-HSA-scFv-cMycG0, or -cMycG1. As a result, many faint bands due to the nonspecific binding of antibodies were also detected, while the two signals in the -cMycG2 lanes were fully saturated so that a reduction in the amount of the tagged polypeptides during the post-trans- lational incubation would be difficult to distinguish from this figure. The degradation of the -cMycG2 tag during the post-translational incubation will be shown in the following sections.
These results indicate that at least two additional glycine residues are necessary to protect the preceding affinity tag sequence from degradation. They also suggest that the deg- radation of affinity tag is not due to the intrinsic instability of the tag sequence, but to sequential degradation from its C-terminal. The primary candidate responsible for this deg- radation is carboxypeptidase(s).
Carboxypeptidase C, carboxypeptidase Y (EC 3.4.16.5), and carboxypeptidase D (EC 3.4.16.6) are serine proteases that specifically release carboxyl terminal amino acid resi-
dues from substrate polypeptides with broad specificities (11). The carboxypeptidases C and D are widely distributed among fungi, animals, and higher plants including wheat and barley. They are divided into the two subgroups according to their preference regarding the last amino acid residue of the substrate (the X1 position of [-X2-X1-OH]): Carboxy- peptidases C and D prefer hydrophobic and basic amino acid residues in the X1 position, respectively (11). In wheat germ, the source of the cell-free extract of this gene expres- sion system, little information on the storage levels of these carboxypeptidases in germ cells and their enzymatic proper- ties is available. However, it was reported that one wheat mRNA for a carboxypeptidase, which showed potypeptide sequence similarity to the active site and substrate-binding regions of yeast carboxypeptidase C, was expressed in the premature grains as welt as in the germinating grain (12). In barley grain, two distinct carboxypeptidase C (CPD-MI and -MIII) and a carboxypeptidase D (CPD-MII) were expressed in the aleurone layer during germination (13). This suggests that wheat grain also likely possesses both types of carboxy- peptidases, although it is not known whether these enzymes are present in the germ fraction.
The amino acid residue preferences for the X2 and X1 positions of several carboxypeptidases, including the three barley enzymes, were evaluated by Degan et al. (13). In their report, the amino acid residue preferences for positions of X2 and X1 of the barley enzymes showed that a mixture of these enzymes covered a broad range of peptide bonds. However, it was also shown that none of the barley enzymes hydrolyzed peptide bonds efficiently when Asp, Pro, or Gly was located at position X2 and Gly at position X1. This preference is consistent with our result that two additional glycine residues in the C-terminal of the tagged protein effi- ciently blocked the degradation (Fig. 2B). Thus, this evi- dence strongly suggests that it is wheat endogenous carboxy- peptidase(s) that degrade the C-terminal tag during/after translation reaction.
Effect of protease inhibitors on cell-free protein syn- thesis Further stabilization of the tag sequence should be achieved because the two glycines did not block the degra- dation completely. In fact, a gradual decrease in the signal intensity during post-translational incubation was still ob- served upon Western blotting analysis using the cMyc-tag- specific antibody. Figure 3 indicates that there was nearly a 50% reduction in the signal intensity of the cMyc-tagged fu- sion protein during the 6 h of post-translational incubation.
In addition to the carboxypeptidases, the presence of other proteases such as phytepsin ([EC 3.4.23.40], aspartic pro- teinase), cathepsin H (EC 3.4.22.16), and peptidase K (EC 3.4.21.64) in the cell-free extract is likely, since they were reported to be involved in the germination of cereal seed. Therefore, possible inhibitors of these potential proteases should also be tested.
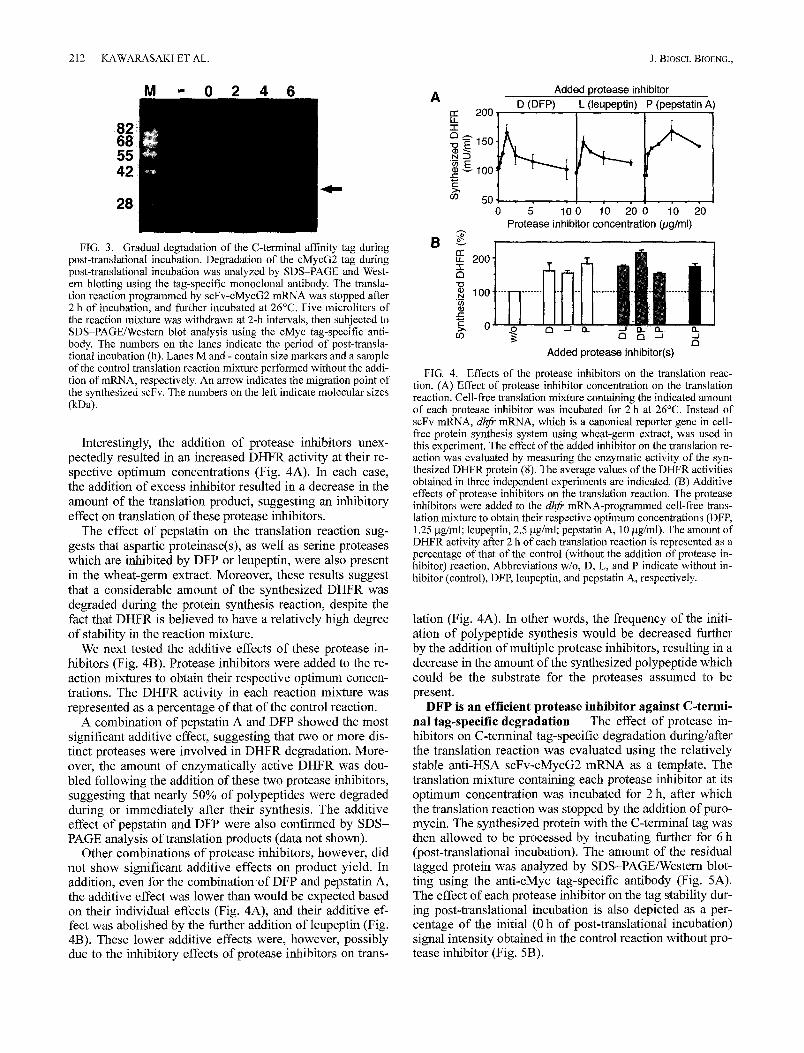
The effects of the concentration of protease inhibitors, such as diisopropylfluorophosphate (DFP, inhibits various serine proteases including carboxypeptidases), leupeptin (in- hibits cysteine proteases and various serine proteases) and pepstatin (aspartic proteinase inhibitor), on the translation reaction were evaluated using in vitro transcripts of the ca- nonical reporter gene DHFR as mRNA (Fig. 4).
212 KAWARASAKI ET AL. J. Biosci. BIOENG.,
M - 0 2 4 6
FIG. 3. Gradual degradation of the C-terminal affinity tag during post-translational incubation. Degradation of the cMycG2 tag during post-translational incubation was analyzed by SDS-PAGE and West- em blotting using the tag-specific monoclonal antibody. The transla- tion reaction programmed by scFv-cMycG2 mRNA was stopped after 2 h of incubation, and further incubated at 26°C. Five microliters of the reaction mixture was withdrawn at 2-h intervals, then subjected to SDS-PAGE/Western blot analysis using the cMyc tag-specific anti- body. The numbers on the lanes indicate the period of post-transla- tional incubation (h). Lanes M and - contain size markers and a sample of the control translation reaction mixture performed without the addi- tion of mRNA, respectively. An arrow indicates the migration point of the synthesized scFv. The numbers on the left indicate molecular sizes (~a).
Interestingly, the addition of protease inhibitors unex- pectedly resulted in an increased DHFR activity at their re- spective optimum concentrations (Fig. 4A). In each case, the addition of excess inhibitor resulted in a decrease in the amount of the translation product, suggesting an inhibitory effect on translation of these protease inhibitors.
The effect of pepstatin on the translation reaction sug- gests that aspartic proteinase(s), as well as serine proteases which are inhibited by DFP or leupeptin, were also present in the wheat-germ extract. Moreover, these results suggest that a considerable amount of the synthesized DHFR was degraded during the protein synthesis reaction, despite the fact that DHFR is believed to have a relatively high degree of stability in the reaction mixture.
We next tested the additive effects of these protease in- hibitors (Fig. 4B). Protease inhibitors were added to the re- action mixtures to obtain their respective optimum concen- trations. The DHFR activity in each reaction mixture was represented as a percentage of that of the control reaction.
A combination of pepstatin A and DFP showed the most significant additive effect, suggesting that two or more dis- tinct proteases were involved in DHFR degradation. More- over, the amount of enzymatically active DHFR was dou- bled following the addition of these two protease inhibitors, suggesting that nearly 50% of polypeptides were degraded during or immediately after their synthesis. The additive effect of pepstatin and DFP were also confirmed by SDS- PAGE analysis of translation products (data not shown).
Other combinations of protease inhibitors, however, did not show significant additive effects on product yield• In addition, even for the combination of DFP and pepstatin A, the additive effect was lower than would be expected based on their individual effects (Fig. 4A), and their additive ef- fect was abolished by the further addition of leupeptin (Fig. 4B). These lower additive effects were, however, possibly due to the inhibitory effects of protease inhibitors on trans-
A re 200
O ~150
~ g ~oo ¢ -
o~ 50
Added protease inhibitor D (DFP) L (leupeptin) P (pepstatin A)
5" 10 0 10 20 0 10 Protease inhibitor concentration (pg/ml)
2O
B o~
G!! !,1 200
• . . . . . . . . . . . . . . . _ . . . . . . . . .
03 a Added protease inhibitor(s)
FIG. 4. Effects of the protease inhibitors on the translation reac- tion. (A) Effect of protease inhibitor concentration on the translation reaction. Cell-free translation mixture containing the indicated amount of each protease inhibitor was incubated for 2 h at 26°C. Instead of scFv mRNA, dhfr mRNA, which is a canonical reporter gene in cell- free protein synthesis system using wheat-germ extract, was used in this experiment. The effect of the added inhibitor on the translation re- action was evaluated by measuring the enzymatic activity of the syn- thesized DHFR protein (8). The average values of the DHFR activities obtained in three independent experiments are indicated. (B) Additive effects of protease inhibitors on the translation reaction. The protease inhibitors were added to the dhfr mRNA-programmed cell-free trans- lation mixture to obtain their respective optimum concentrations (DFP, 1.25 gg/ml; leupeptin, 2.5 gg/ml; pepstatin A, 10 gg/ml). The amount of DHFR activity after 2 h of each translation reaction is represented as a percentage of that of the control (without the addition of protease in- hibitor) reaction. Abbreviations w/o, D, L, and P indicate without in- hibitor (control), DFP, leupeptin, and pepstatin A, respectively.
lation (Fig. 4A). In other words, the frequency of the initi- ation of polypeptide synthesis would be decreased further by the addition of multiple protease inhibitors, resulting in a decrease in the amount of the synthesized polypeptide which could be the substrate for the proteases assumed to be present.
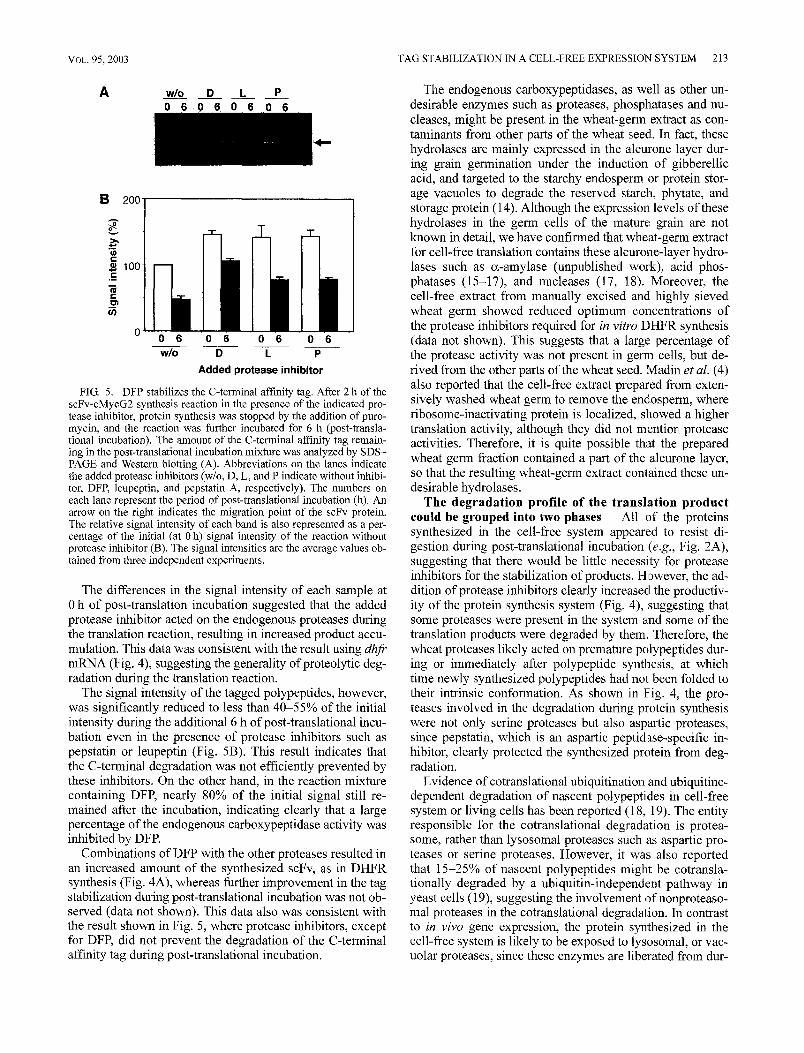
DFP is an efficient protease inhibitor against C-termi- nal tag-specific degradation The effect of protease in- hibitors on C-terminal tag-specific degradation during/after the translation reaction was evaluated using the relatively stable anti-HSA scFv-cMycG2 mRNA as a template. The translation mixture containing each protease inhibitor at its optimum concentration was incubated for 2 h, after which the translation reaction was stopped by the addition ofpuro- mycin. The synthesized protein with the C-terminal tag was then allowed to be processed by incubating further for 6 h (post-translational incubation). The amount of the residual tagged protein was analyzed by SDS-PAGE/Western blot- ting using the anti-cMyc tag-specific antibody (Fig. 5A). The effect of each protease inhibitor on the tag stability dur- ing post-translational incubation is also depicted as a per- centage of the initial (0 h of post-translational incubation) signal intensity obtained in the control reaction without pro- tease inhibitor (Fig. 5B).
VOL. 95, 2003 T A G S T A B I L I Z A T I O N 1N A C E L L - F R E E EXPRESSION SYSTEM 213
A w/o n o l , - l i o i , T l o n , ' l l o i ; Li l l - . . ' l ffi:-'l
B~ 200 I
e- l 0 0 ¢-
¢- ._~
FIG. 5.
0 0 6 0 6 0 6 0 6 w/o D L P
Added protease inhibitor
DFP stabilizes the C-tenninal affinity tag. After 2 h of the scFv-cMycG2 synthesis reaction in the presence of the indicated pro- tease inhibitor, protein synthesis was stopped by the addition of puro- mycin, and the reaction was further incubated for 6 h (post-transla- tional incubation). The amount of the C-terminal affinity tag remain- ing in the post-translational incubation mixture was analyzed by SDS- PAGE and Western blotting (A). Abbreviations on the lanes indicate the added protease inhibitors (w/o, D, L, mad P indicate without inhibi- tor, DFR leupeptin, and pepstatin A, respectively). The numbers on each lane represent the period of post-translational incubation (11). An arrow on the right indicates the migration point of the scFv protein. The relative signal intensity of each band is also represented as a per- centage of the initial (at 0 h) signal intensity of the reaction without protease inhibitor (B). The signal intensities are the average values ob- tained from three independent experiments.
The differences in the signal intensity of each sample at 0 h of post-translation incubation suggested that the added protease inhibitor acted on the endogenous proteases during the translation reaction, resulting in increased product accu- mulation. This data was consistent with the result using dhfr mRNA (Fig. 4), suggesting the generality ofproteolytic deg- radation during the translation reaction.
The signal intensity of the tagged polypeptides, however, was significantly reduced to less than 40-55% of the initial intensity during the additional 6 h of post-translational incu- bation even in the presence of protease inhibitors such as pepstatin or leupeptin (Fig. 5B). This result indicates that the C-terminal degradation was not efficiently prevented by these inhibitors. On the other hand, in the reaction mixture containing DFP, nearly 80% of the initial signal still re- mained after the incubation, indicating clearly that a large percentage of the endogenous carboxypeptidase activity was inhibited by DFR
Combinations of DFP with the other proteases resulted in an increased amount of the synthesized scFv, as in DHFR synthesis (Fig. 4A), whereas further improvement in the tag stabilization during post-translational incubation was not ob- served (data not shown). This data also was consistent with the result shown in Fig. 5, where protease inhibitors, except for DFP, did not prevent the degradation of the C-terminal affinity tag during post-translational incubation.
The endogenous carboxypeptidases, as well as other un- desirable enzymes such as proteases, phosphatases and nu- cleases, might be present in the wheat-germ extract as con- taminants from other parts of the wheat seed. In fact, these hydrolases are mainly expressed in the alem'one layer dur- ing grain germination under the induction of gibberellic acid, and targeted to the starchy endosperm or protein stor- age vacuoles to degrade the reserved starch, phytate, and storage protein (14). Although the expression levels of these hydrolases in the germ cells of the mature grain are not known in detail, we have confirmed that wheat-germ extract for cell-free translation contains these aleurone-layer hydro- lases such as a-amylase (unpublished work), acid phos- phatases (15-17), and nucleases (17, 18). Moreover, the cell-free extract from manually excised and highly sieved wheat germ showed reduced optimum concentrations of the protease inhibitors required for in vitro DHFR synthesis (data not shown). This suggests that a large: percentage of the protease activity was not present in germ cells, but de- rived from the other parts of the wheat seed. Madin et al. (4) also reported that the cell-free extract prepared from exten- sively washed wheat germ to remove the endosperm, where ribosome-inactivating protein is localized, showed a higher translation activity, although they did not mention protease activities. Therefore, it is quite possible that the prepared wheat germ fraction contained a part of the aleurone layer, so that the resulting wheat-germ extract contained these un- desirable hydrolases.
The degradation profile of the translation product could be grouped into two phases All of the proteins synthesized in the cell-free system appeared to resist di- gestion during post-translational incubation (e.g., Fig. 2A), suggesting that there would be little necessity for protease inhibitors for the stabilization of products. However, the ad- dition of protease inhibitors clearly increased the productiv- ity of the protein synthesis system (Fig. 4), suggesting that some proteases were present in the system and some of the translation products were degraded by them. Therefore, the wheat proteases likely acted on premature polypeptides dur- ing or immediately after polypeptide synthesis, at which time newly synthesized polypeptides had not been folded to their intrinsic conformation. As shown in Fig. 4, the pro- teases involved in the degradation during protein synthesis were not only serine proteases but also aspartic proteases, since pepstatin, which is an aspartic peptidase-specific in- hibitor, clearly protected the synthesized protein from deg- radation.
Evidence of cotranslational ubiquitination and ubiquitine- dependent degradation of nascent polypeptides in cell-free system or living cells has been reported (18, 19). The entity responsible for the cotranslational degradation is protea- some, rather than lysosomal proteases such as aspartic pro- teases or serine proteases. However, it was also reported that 15-25% of nascent polypeptides migh,t be cotransla- tionally degraded by a ubiquitin-independent pathway in yeast cells (19), suggesting the involvement of nonproteaso- mal proteases in the cotranslational degradation. In contrast to in vivo gene expression, the protein synthesiTed in the cell-free system is likely to be exposed to lysosomal, or vac- uolar proteases, since these enzymes are liberated from dur-

214 KAWARASAKI ET AL. J. BIOSCL BIOENG.,
ing the homogenization of the resource cells. Cotransla- tional degradation in the cell-free protein synthesis system, therefore, might be intensified by these undesirable pro- teases. Nevertheless, it should be emphasized that the ad- dition of protease inhibitors to the translation mixture effi- ciently prevented cotranslational degradation without ad- versely affecting the translation activity (Fig. 5). This is of benefit particularly when the translation system is applied to the preparation of gene products the stability of which is un- known.
On the other hand, the carboxypeptidase acted on the target polypeptide post-translationally, since carboxypepti- dase could not access the C-terminal tag until it was re- leased from the ribosome. However, once liberated from the ribosome, the C-terminal tag would be exposed to the car- boxypeptidases. In the advanced cell-free translation system using wheat-germ extract, the translation reaction lasts nearly 10 h (15) and can be extended to more than 60 h us- ing a substrate supplementation system (1-3, 5, 20). The exposure to carboxypeptidases continues in the subsequent functional analysis unless the product is separated from the translation mixture. The presence of the carboxypeptidases in the system, therefore, can be the cause of low recovery in the purification of the product, since the C-terminal affinity tag-specific degradation would proceed during such a long reaction period. It should be noted here again that the C-ter- minal tag-specific degradation can be prevented efficiently by the addition of two glycine residues at the C-terminal of the target protein and the addition of appropriate amount of DFP to the cell-free translation system.
A C K N O W L E D G M E N T S
This work was financially supported by a Grant-in-Aid for Scientific Research from the Japanese Society for the Promotion of Science (no. 13750734). We especially thank Dr. Toshiki Nakamura, Tohoku National Agricultural Experiment Station, and Ms. Setsuko Nanba, Nippon Flour Mills, Co., Ltd., for the produc- tion of wheat seeds with high translation activity, and the prepara- tion of precisely sieved wheat germ, respectively.
REFERENCES
1. Jermutus, L., Ryabova, L.A., and Pliiekthun, A.: Recent advances in producing and selecting functional proteins by using cell-free translation. Curr. Opin. Biotechnol., 9, 534- 548 (1998).
2. Spirin, A.S. (ed.).: Cell-free translation systems. Springer, Berlin, Germany (2002).
3. Ryabova, L.A., Morozov, I.Y., Spirin, A. S.: Continuous- flow cell-free translation, transcription-translation, and repli- cation-translation systems. Method. Mol. Biol., 77, 179-193 (1998).
4. Madin, K., Sawasaki, T., Ogasawara, T., and Endo, Y.: A
highly efficient and robust cell-free protein synthesis system prepared from wheat embryos: plants apparently contain a suicide system directed at ribosomes. Proc. Natl. Acad. Sci. USA, 97, 559-564 (2000).
5. Rungpragayphan, S., Kawarasaki, Y., Imaeda, T., Kohda, K., Nakano, H., and Yamane, T.: High-throughput, cloning- independent protein library construction by combining single- molecule DNA amplification with in vitro expression, J. Mol. Biol., 318, 395405 (2002).
6. Alimov, A.P., Khmelnitsky, A.Yu., Simonenko, E N., Spirin, A. S., and Chetverin, A.B.: Cell-free synthesis and affinity isolation of proteins on a nanomole scale. Biotech- niques, 28, 338-344 (2000).
7. He, M. and Taussig, M. J.: Single step generation of protein arrays from DNA by cell-free expression and in situ immobili- sation (PISA method). Nucleic Acids Res., 29, e73 (2001).
8. Kawarasaki, Y., Kawai, T., Nakano, H., and Yamane, T.: A long-lived batch reaction system of cell-free protein syn- thesis. Anal. Biochem., 226, 320-324 (1995).
9. Kawarasaki, Y., Kasahara, S, Kodera, N., Shinbata, T., Sekiguchi, S., Nakano, H,, and Yamane, T.: A trimmed viral cap-independent translation enhancing sequence for rapid in vitro gene expression. Biotechnol. Prog., 16, 517-521 (2OOO).
10. Sambrook, J. and Russell, D. W.: Molecular cloning: a labo- ratory manual, 3rd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (2001).
11. Remington, S.J. and Breddam, K.: Carboxypeptidases C and D. Method. Enzymol., 244, 231-248 (1994).
12. Banlcombe, D.C., Barker, R.F., and Jarvis, M.G.: A gibberellin responsive wheat gene has homology to yeast car- boxypeptidase Y. J. Biol. Chem., 262, 13726-13735 (1987).
13. Degan, F. D., Ribadeau-Dumas, B., and Breddam, K.: Pu- rification and Characterization of two serine carboxypepti- dases from Aspergillus niger and their use in C-terminal se- quencing of proteins and peptide synthesis. Appl. Environ. Microbiol., 58, 2144-2152 (1992).
14. Fath, A., Bathke, P., Lonsdale, J., Meza-Romero, R., and Jones, R.: Programmed cell death in cereal aleurone. Plant Mol. Biol., 44, 255-266 (2000).
15. Kawarasaki, Y., Nakano, H., and Yamane, T.: Purification and some properties of wheat germ acid phosphatases. Plant Sci., 119, 67-77 (1996).
16. Nakano, H., Tanaka, T., Kawarasaki, Y., and Yamane, T.: Highly productive cell-free protein synthesis system using condensed wheat-germ extract. J. Biotechnol., 46, 275-282 (1996).
17. Kawarasaki, Y., Nakano, H., and Yamane, Y.: Phosphatase- immunodepleted cell-free protein synthesis system. J. Bio- technol., 61, 199-208 (1998).
18. Sato, S., Ward, C.L., and Kopito, R.R.: Cotranslational ubiquitination of cystic fibrosis transmembrane conductance regulator in vitro. J. Biol. Chem., 273, 7189-7192 (1998).
19. Turner, G. C. and Varshavsky, A.: Detecting and measuring cotranslational protein degradation in vivo. Science, 289, 2117-2120 (2000).
20. Sawasaki, T., Hasegawa, Y., Tsuchimochi, M., Kamura, N., Ogasawara, T., and Endo, Y.: A bilayer cell-free protein synthesis system for high-throughput screening of gene prod- ucts. FEBS Lett., 514, 102-105 (2002).