stepwise chilling: tender pork without compromising … · stepwise chilling: tender pork without...
TRANSCRIPT

K. Rosenvold, U. Borup and M. TherkildsenStepwise chilling: Tender pork without compromising water-holding capacity
doi: 10.2527/jas.2009-2468 originally published online January 29, 20102010, 88:1830-1841.J ANIM SCI
http://jas.fass.org/content/88/5/1830the World Wide Web at:
The online version of this article, along with updated information and services, is located on
www.asas.org
at Serials/Acq. Dept., Library on November 24, 2011jas.fass.orgDownloaded from

ABSTRACT: The current pork slaughter process is primarily optimized to reduce cooler shrink and the in-cidence of PSE pork. Elimination of the halothane gene and improved preslaughter handling have decreased the incidence of PSE pork and improved the water-holding capacity of the muscle; however, the chilling process has not been optimized to accommodate these changes. The hypothesis that stepwise chilling could improve tender-ness without compromising water-holding capacity was tested in this study. The stepwise chilling treatments were composed of a rapid chilling to 10 or 15°C (in a chilling tunnel) and a 6-h holding period at 10 or 15°C, followed by rapid chilling to 4°C. Both treatments were compared directly with a chilling treatment that simu-lated conventional tunnel chilling; one carcass half from each pig was allocated to a stepwise chilling treatment, whereas the other carcass half was allocated to the con-trol treatment. A total of 42 pigs were slaughtered on 6 slaughter days. Biopsies were collected for analysis of glycogen degradation and glycogen debranching enzyme activity from slaughter until 72 h postmortem, and samples for color, sarcomere length, drip loss, Warner-Bratzler shear force, and sensory analysis were removed from the carcass 24 h postmortem. Substantial tem-
perature differences were obtained during the holding period between the stepwise and conventionally chilled carcass halves. These had almost, but not completely, disappeared by 22 h postmortem, and although the dif-ferences were small, pH was significantly (P < 0.01) less in the stepwise-chilled carcasses compared with the control carcasses. The stepwise chilling treatments led to significantly improved (P < 0.01) tenderness in LM without compromising quality indicators or attributes such as pH, drip loss, or ham processing yield, although color of the stepwise-chilled pork was affected. Neither the tenderness of processed semimembranosus muscle nor the shear force of biceps femoris muscle was affected (P > 0.05) because of the smaller temperature differ-ences in these muscles. The improvements in tenderness could be solely attributed to the increased proteolysis postmortem in the stepwise-chilled carcasses, with the greater temperatures favoring proteolytic enzymes in-volved in muscle protein degradation. Furthermore, the results for glycogen metabolism successfully revealed that both pro- and macroglycogen contributed to the energy generation in postmortem muscles, with degra-dation of both forms early postmortem.
Key words: chilling, drip loss, macroglycogen, pork, proglycogen, tenderness
©2010 American Society of Animal Science. All rights reserved. J. Anim. Sci. 2010. 88:1830–1841 doi:10.2527/jas.2009-2468
INTRODUCTION
Although the beef and lamb slaughter processes are optimized to produce tender meat by using tech-nologies such as electrical stimulation (Devine et al., 2004), hip suspension (Sørheim and Hildrum, 2002), and conditioning (Smulders et al., 1992), the slaughter process for pork is primarily optimized to reduce cooler shrink and the incidence of PSE pork. This is primar-ily done by decreasing the temperature quickly after slaughter to prevent the detrimental effect of increased temperatures in combination with low pH (Wismer-Pedersen and Briskey, 1961). Today, the halothane gene has largely been eliminated and, in combination
Stepwise chilling: Tender pork without compromising water-holding capacity1
K. Rosenvold,*2,3 U. Borup,* and M. Therkildsen†
*Danish Meat Research Institute, Department of Meat Quality, DK-4000 Roskilde, Denmark; and †Faculty of Agricultural Science, Department of Food Science,
University of Aarhus, DK-8830 Tjele, Denmark
1 The authors thank Jonna Anderson, Maiken Baltzer, Camilla Bejerholm, Mianne Darré, and Peter Vorup from the Danish Meat Research Institute, Roskilde, Denmark, and Jens Askov Jensen and Ivan Nielsen from the University of Aarhus, Tjele, Denmark for ex-cellent technical assistance, and acknowledgement for financing this work is given to The Danish Bacon and Meat Council (Copenhagen, Denmark) and the Danish Food Industry Agency under the Ministry of Food, Agriculture and Fisheries (Copenhagen, Denmark).
2 Present address: AgResearch MIRINZ, Ruakura Research Cen-tre, Hamilton, 3240 New Zealand.
3 Corresponding author: [email protected] September 10, 2009.Accepted January 25, 2010.
1830
at Serials/Acq. Dept., Library on November 24, 2011jas.fass.orgDownloaded from

with a more gentle treatment of the pigs preslaughter, the incidence of PSE has decreased and water-holding capacity (WHC) has improved. However, the chilling process has not always been optimized to accommodate these changes.
The prerigor temperature is crucial for tenderness, with temperatures between 10 to 15°C resulting in maximal tenderness, which can be explained by both minimal muscle contraction and maximal proteolysis (Locker and Hagyard, 1963). When suboptimal, both factors limit the extent of postmortem proteolysis and affect the final tenderness. A cooling profile that aims at optimizing tenderness should therefore both mini-mize contraction and optimize the conditions for pro-teolysis.
Water-holding capacity, another crucial meat qual-ity trait, is described as multifactorial in its origin; however, final pH and rate of pH change are critical (Wismer-Pedersen, 1987). The rate of pH change, re-sulting from the breakdown of glycogen to lactate, is a function of temperature (Bendall, 1973). Glycogen is broken down by glycogen phosphorylase and glycogen debranching enzyme (GDE; Brown and Brown, 1966). Kylä-Puhju et al. (2005) has shown that the activity of GDE decreases with decreasing temperatures and that the activity of GDE is identical at 15 and 4°C. It can be speculated that the pH changes will be identical at these 2 temperatures.
Finally, muscle temperature at the time of slaugh-ter affects WHC (Schäfer et al., 2002), and group-wise handling of pigs preslaughter has reduced the carcass temperatures at the time of slaughter (K. Rosenvold, unpublished data), with improved WHC (Støier et al., 2001).
Combining the observations that temperature at 1 min postmortem (T1min) is reduced because of group-wise preslaughter handling, that the temperature in-terval of 10 to 15°C prerigor results in maximal ten-derness, and that glycogen breakdown at 15 and 4°C is identical led to the following hypothesis: a stepwise chilling method, composed of a rapid temperature re-duction to 10 to 15°C and a 6-h holding period at 10 or 15°C, followed by chilling to 4°C, will improve tender-ness without compromising WHC.
MATERIALS AND METHODS
The experiment was approved by and the animals treated in accordance with the guidelines outlined by The Danish Inspectorate of Animal Experimentation.
Experimental Design
Two stepwise chilling treatments were investigated in the study. The stepwise chilling treatments were achieved using a commercial prechilling process time (40 min postmortem) and chilling conditions of com-mercial chilling tunnels, modeled for the mean car-
cass temperature to reach 10°C (Step10) and 15°C (Step15), respectively. The carcasses were then held at each of these temperatures for 6 h to allow the muscles to go into rigor mortis (knowing that rigor is reached within 3 to 6 h postmortem) by using either batch chill-ing (Taylor and Dant, 1971) or tunnel chilling (Maribo et al., 1998). With greater carcass temperatures in the stepwise chilling treatments, rigor mortis was expected to be reached within the 6 h. Subsequently, the mean carcass temperature was decreased to 4°C, using chill-ing tunnel conditions for this temperature to be reached as quickly as possible.
Both stepwise chilling treatments were compared di-rectly with a chilling treatment that simulated conven-tional tunnel chilling (control), with one carcass half from each pig allocated to 1 of the 2 stepwise chilling treatments and the other carcass half allocated to the control treatment (Con10 and Con15, respectively).
Three chillers were used in the study. Chiller 1 oper-ated at −22°C and a fan with a frequency transform-er ensured an air flow of 3 m·s−1, thereby simulating the temperature and air speed of commercial chilling tunnels used in Danish commercial slaughter plants. Chiller 2 operated at either 10°C or 15°C, depending on which of the 2 stepwise chilling treatments was being investigated on that day. Chiller 3 operated at 4°C, and temperature equilibration took place there.
Treatment Step10. Forty minutes postmortem, the carcass half was placed in chiller 1 (−22°C, 3 m·s−1). After 69 min in chiller 1, the carcass half was moved to chiller 2 (10°C, 0.1 m·s−1), where it stayed for 6 h. The carcass half was then returned to chiller 1 for an addi-tional 12 min, and finally it was moved to chiller 3 (4°C, 0.1 m·s−1), where it was left for the carcass temperature to equilibrate until the next day.
Treatment Step15. Forty minutes postmortem, the carcass half was placed in chiller 1 (−22°C, 3 m·s−1). After 47 min in chiller 1, the carcass half was moved to chiller 2 (15°C, 0.1 m·s−1), where it stayed for 6 h. The carcass half was then returned to chiller 1 for an addi-tional 24 min, and finally it was moved to chiller 3 (4°C, 0.1 m·s−1), where it was left for the carcass temperature to equilibrate until the next day.
Control Treatment (Con10 and Con15). For-ty minutes postmortem, the carcass half was placed in chiller 1 (−22°C, 3 m·s−1). After 75 min in chiller 1, the carcass half was moved to chiller 3 (4°C, 0.1 m·s−1), where it was left for the carcass temperature to equili-brate until the next day.
Animals
The pigs used in the study (21 castrated males and 21 females) were reared at the experimental farm of Foulum Research Centre, The Faculty of Agricultural Sciences, University of Aarhus, Denmark. They were crossbreeds between Danish Landrace × Danish York-shire sows and Duroc boars, and all were noncarriers
Stepwise chilling of pork 1831
at Serials/Acq. Dept., Library on November 24, 2011jas.fass.orgDownloaded from

of the halothane gene. The sex distribution for the 2 stepwise chilling treatments was as shown in Table 1.
Slaughter Procedure and Sampling
Each of the 2 stepwise chilling treatments was re-peated 3 times on separate slaughter days, giving a to-tal of 6 slaughter days. Seven pigs were slaughtered on each slaughter day. One carcass half was chilled using either Step10 or Step15 (alternating between the right and the left side), and the other half was chilled using the control treatment. Control carcass halves, corre-sponding to either the Step10 or the Step15 carcass halves, are referred to as Con10 and Con15, respec-tively. On the day of slaughter, the animals were trans-ported from the porcine facilities to the experimental slaughter plant (200 m) at Research Centre Foulum, University of Aarhus, Denmark, with as little stress as possible. Pigs were stunned by 80% CO2 for 3 min, exsanguinated, scalded at 62°C for 3 min, cleaned, and eviscerated within 30 min. At 40 min postmortem, the chilling process was initiated.
For analysis of glycogen degradation, GDE activity, and desmin degradation, muscle biopsies were taken from the LM at 1 min, 40 min, 2 h, 8 h, 24 h, and 72 h postmortem and were immediately snap-frozen in liq-uid nitrogen and stored at −80°C. At 24 h postmortem, 2-cm-thick samples were cut from the LM and the M. biceps femoris (BF) for measurement of instrumental color. After color measurements, the LM sample was vacuum-packed and stored at 4°C until 72 h postmor-tem, snap-frozen in liquid nitrogen, and then stored at −80°C until measurement of the myofibrillar frag-mentation index (MFI). In addition, samples for sar-comere length determination in LM were removed and fixed in borate solutions (fix 1: 0.039 M boric acid, 0.1 M KCl, 5 mM EDTA, and 2.5% glutaraldehyde; fix 2: 0.039 M boric acid, 0.025 M KCl, 5 mM EDTA, and 2.5% glutaraldehyde). Other 2-cm samples from LM and BF were used for determination of drip loss, and 7-cm samples from LM and BF were used for determi-nation of Warner-Bratzler shear force. All samples were vacuum-packed and aged at 4°C until 72 h postmortem, and then stored at −20°C until measurement. A 25-cm
sample from LM was removed from the carcass and used for sensory analysis, and the M. semimembranosus (SM) was used to produce ham.
Temperature and pH Measurements
Temperature was measured continuously in LM, in the ham adjacent to the aitch bone, and in the shoulder at the blade bone from 30 min to 22 h postmortem, using temperature loggers that logged the temperature every 2 min (StowAway TidbiT, Bourne, MA). Individ-ual temperature and pH measurements were captured in the LM adjacent to the last rib at 1 min postmortem and in the LM, BF, and SM at 35 min postmortem. These data were also collected when carcasses were moved from one chiller to the other and at 4, 6, and 22 h postmortem. Temperature was measured using a Testo 901 thermometer (Testo, Lenzkirch, Germany) and pH was measured using a Knick pH-meter (Model 913, Knick, Berlin, Germany) equipped with an inser-tion glass electrode (Ingold LOT glass electrode Ø 6 mm type 3120, Mettler Toledo, Columbus, OH). The pH meter was calibrated in buffers with pH 4.01 and 7.00 (Radiometer, Copenhagen, Denmark) at 35°C for the measurements made up to 35 min postmortem in LM and up to 4 h postmortem in BF and SM. The pH electrode was calibrated at 15°C for the remaining pH measurements made on the day of slaughter. For measurements carried out at 24 h postmortem, the cali-bration was conducted at 4°C. The mean of 2 measure-ments was used for both pH and temperature.
Percentage of Lean and Cooler Shrink
Carcass percentage of lean was determined using a Fat-O-Meat’er (SFK-Technologies, Herlev, Denmark). Cooler shrink was determined by weighing the carcass-es immediately before chilling and then when the car-casses were boned at 24 h postmortem.
Drip Loss
Drip loss was determined using the EZ-DripLoss method (Christensen, 2003). From the 2-cm-thick LM
Table 1. Slaughter weight and percentage of lean of the 42 pigs that were subjected to either stepwise chilling (Step10 or Step15) or control (Con10 or Con15) chilling1
Item
Step10/Con10 Step15/Con15
SEMCastrates Females Castrates Females
n 11 10 10 11 Slaughter wt, kg 104.0 102.7 106.6 100.4 5.3Lean, % 60.8 60.7 61.7 60.8 0.6
1Values presented as least squares means and SEM. The stepwise chilling treatments were applied 40 min postmortem by subjecting one-half of the carcass to chilling tunnel conditions (−22°C, 2 m·s−1) until the mean carcass temperature reached 10°C (69 min, Step10) and 15°C (47 min, Step15), respectively. The carcasses were then held at 10°C or 15°C, respectively, for 6 h before the temperature was reduced to 4°C. The other one-half of the carcass was subjected to chilling tunnel conditions (−22°C, 2 m·s−1) for 75 min and then placed at 4°C (Con10 and Con15, respectively).
Rosenvold et al.1832
at Serials/Acq. Dept., Library on November 24, 2011jas.fass.orgDownloaded from

and BF samples, 2 circular samples (25 mm in diam-eter) were punched out using a plug center bit. The circular samples were then placed in a special cup and closed hermetically with a lid to prevent evaporation, which allowed unimpeded drip loss from the meat. Af-ter 24 h of storage at 4 to 6°C, the drip loss, inversely related to WHC, was determined.
Color Measurements
Color was measured on the LM and BF samples us-ing a Minolta CR-300 Chroma Meter (Minolta, Osaka, Japan) calibrated against a white tile (L* = 92.30, a* = 0.32, and b* = 0.33). The aperture was 8 mm, and illuminant D65 and a 10° standard observer were used. Samples were allowed to bloom for 1 h at 4°C before the measurements. The 3 variables L*, a*, and b*, rep-resenting lightness, redness, and yellowness, were mea-sured on 5 sites of each sample, and the mean of the 5 measurements was reported.
Glycogen Degradation Postmortem
Degradation of glycogen was followed in Step15 and Con15 LM samples by measuring the contents of free glucose, proglycogen, and macroglycogen and the activ-ity of GDE. The contents of free glucose, proglycogen, and macroglycogen were measured in biopsies sampled at 0, 2, 8, 24, and 72 h postmortem, using the method described by Ylä-Ajos et al. (2007). Additionally, the content of free glucose was measured in the supernatant after precipitation of proglycogen and before hydroly-sis with HCl to measure the content of macroglycogen. Thus, in the present study, data for free glucose, progly-cogen, and macroglycogen are presented.
The activity of GDE was measured in LM biopsies sampled at 0, 2, 8, and 24 h postmortem by the method described by Nelson et al. (1970), with the modifications described by Kylä-Puhju et al. (2005). The method was scaled to fit measurement in 96-well ELISA plates so that in the present study, 20 μL of muscle extract was added to the wells together with 5 μL of 0.5 M sodium maleate in triplicate, and the reaction was begun by adding 25 μL of 1% limit dextrin. After incubation for 5, 10, and 15 min at 39°C, the reaction was stopped by boiling in a microwave oven, followed by storage on ice. Thereafter, 65 μL of the iodine reagent (3.8%) was added to each well and the absorbance at 525 nm was measured after 20 min. The activity is expressed per gram of muscle tissue per minute from the slope of the absorbance curve.
Texture Characteristics
Warner-Bratzler shear force was determined in LM and BF at 72 h postmortem, as described by Hansen et al. (2006) according to the procedure described by Honikel (1998). However, in this study, the samples
(trimmed to be 5 cm long in a longitudinal direction, 8 cm wide, and 4 cm high) were heated in a water bath maintained at 70°C for 90 min, to reach an internal temperature of 70°C.
Sarcomere length, degradation of desmin, and the MFI were measured in the LM of both carcass halves subjected to either the Step15 or the Con15 chilling treatments and in the control carcass halves. Sarcomere length was measured as described by Kristensen et al. (2004). Degradation of desmin was measured by West-ern blotting according to the sample preparation pro-cedure described by Wheeler and Koohmaraie (1999) and the method described by Kristensen et al. (2004). Ten-microgram protein samples (5 μL, at 0, 2, 8, 24, and 72 h) from LM of both carcass halves from 1 pig were loaded on the same 10.0% 18-well Criterion gel (Bio-Rad Laboratories, Sundbyberg, Sweden) and run at 200 V for 1.5 h at 4°C. The transfer of the proteins to polyvinylidene fluoride membranes was done accord-ing to the method of Towbin et al. (1979) at 1.5 mA for 1.5 h at 4°C. After blotting, the membranes were rinsed and blocked as described by Kristensen et al. (2004), and then incubated with mouse monoclonal an-ti-desmin at a dilution of 1:10,000 (Sigma D1033, Sig-ma-Aldrich Denmark A/S, Brøndby, Denmark) for 1.5 h at room temperature. After washing, the membranes were incubated with Alexa Flour 488 rabbit anti-mouse (Invitrogen A/S, Taastrup, Denmark) at a dilution of 1:4,000 for 1 h. The percentage of intact desmin was quantified with an FX Molecular Imager using Quan-tity One software (Bio-Rad Laboratories) at an excita-tion wavelength of 488 nm and an emission wavelength of 530 nm. The amount of intact desmin is expressed as a percentage of that observed at 0 h postmortem in the corresponding LM sample. The MFI was measured according to the method of Culler et al. (1978), with minor modifications as described in Therkildsen et al. (2002).
Processing of Ham
The SM were vacuum-packed and stored at 4°C until 48 h postmortem, when they were frozen and stored at −20°C until processing. Before processing, the SM were defrosted at 7°C for 48 h. They were then brine-injected (see brine composition in Table 2) to a weight gain of 14%, using a multineedle injector. After brine injec-tion, the SM were tumbled as one batch in a 3-chamber tumbler (model TRI, machine No. 9775-2004, Fomaco, Køge, Denmark). They were tumbled under full vacuum at 7°C for 12 h, with 30 min of tumbling (10 rotations/min) and 30-min breaks (total of 3,600 rotations). After tumbling, the SM were vacuum-packed and cooked at 85°C for 30 min and then at 75°C until an internal tem-perature of 71°C was reached, sprinkled with water at 10°C for 2 h, and chilled at 2°C for 24 h. After storage at 4°C for 5 to 11 d, the sensory analysis was carried out. The SM were weighed after thawing, brine injec-
Stepwise chilling of pork 1833
at Serials/Acq. Dept., Library on November 24, 2011jas.fass.orgDownloaded from

tion, and tumbling, and again immediately before the sensory analysis.
Sensory Analysis
The sensory panel consisted of 9 assessors (2 males and 7 females), all citizens from the Roskilde area of Denmark. The panel had received basic sensory training based on ISO 4121, ASTM-MNL 13, DIN 10964, and the assessors were all familiar with pork and descriptive sensory analyses. Before the analysis, each panelist was trained in 2 sessions on the samples presented in the experiment.
Pork Chops. The 25-cm LM sample for sensory analysis was vacuum-packed and stored at 4°C un-til sensory analysis was carried out 3 d postmortem. Two-centimeter-thick chops were cut and tempered to a core temperature of 10 to 15°C before cooking in a preheated frying pan (155°C); chops were turned every 2 min until an internal temperature of 65 to 68°C was reached. From each chop, two 2.5-cm-wide and 4-cm-long slices were cut. Each assessor was served 1 slice on a preheated plate. The sensory attributes of tender-ness, juiciness, and hardness at first bite were assessed on a continuous scale from 0 (no intensity) to 15 (high intensity).
Processed Ham. The sensory analysis of the pro-cessed SM was carried out 5 to 11 d after processing. The SM were randomized across the 7 sessions, with the exception that the 2 SM from the same pig (Step10 and Con10, or Step15 and Con15) were analyzed in the same session. The hams were weighed when they were removed from the vacuum bags to determine cooking loss, and thereafter were cut into 5-mm-thick slices and served cold. The sensory attributes of surface holes, wet surface, slice cohesiveness, tenderness, and juiciness were assessed on a continuous scale from 0 (no inten-sity) to 15 (high intensity).
Data Analysis
The MIXED procedure (SAS Inst. Inc., Cary, NC) was applied when calculating the least squares means and SE for all the variables. The effect of each chill-ing treatment (Step10 and Step15) was analyzed sepa-rately. The model included the fixed effects of chill-ing treatment, sex, and their interactions, with BW at slaughter as a covariate and the random effects of day of slaughter and pig for all the attributes, with the ex-
ception of glycogen, GDE, and the sensory attributes. In the analysis of glycogen and GDE, the model also included time postmortem and the interactions of time postmortem × chilling method and time postmortem × sex. In the analysis of the sensory attributes, the model including the fixed effects of chilling treatment, sex, and their interactions, with BW at slaughter as a covariate and the random effects of day of slaughter, pig, and sensory assessor. In none of the statistical tests was sex found to have any direct effect (P > 0.05) or to interact with the chilling treatments, so these were subsequently removed from further analysis.
RESULTS
Slaughter Weight, Percentage of Lean, and Cooler Shrink
Slaughter weight and percentage of lean of the 42 pigs included in the study are shown in Table 1. There were no significant differences (P > 0.05) in slaughter weight or percentage of lean between chilling treatments or between sexes. The were no differences in cooler shrink between the Step10 and Con10 carcasses, whereas the cooling shrink was 1 percentage point greater in the Step15 carcasses than in the Con15 carcasses (P < 0.01; Table 3).
Temperature and pH
Temperatures measured continuously in LM, in the ham at the aitch bone, and in the shoulder at the blade bone from 30 min to 22 h postmortem, are shown in Figure 1. The mean LM at T1min in the 42 pigs was 39.5 ± 0.4°C. The maximal T1min was measured to 40.6°C. Significant temperature differences were obtained be-tween Step10 and Con10 (change in temperature at 8 h was 5.4°C in LM, 3.5°C in BF, and 3.7°C in SM) and between Step15 and Con15 (change in temperature at 7.5 h was 7.3°C in LM, 5.6°C in BF, and 5.6°C in SM). The temperature differences had almost, but not com-pletely, disappeared at 22 h postmortem.
The pH in LM at 1 min postmortem (time of slaugh-ter) and in LM, BF, and SM at 35 min, 4 h, 6 h, and 22 h postmortem as well as when the carcass halves were moved among the 3 chillers are shown in Table 4. The pH measured in Step10 and Con10 were not different (P > 0.05) in LM in the first 4 h postmortem, whereas pH at 6, 8, and 22 h were significantly less (P < 0.05) in LM with Step10 than with Con10. However, the dif-ference in pH at 22 h was only 0.05 pH units. In BF, pH at 4 and 8 h was significantly less (P < 0.05) with Step10 than with Con10. In SM, only pH at 8 h was significantly different (P < 0.01) between the 2 treat-ments. The pH measured in Step15 and Con15 was not different (P > 0.05) in LM in the first 4 h postmortem, whereas pH at 6, 8, and 22 h in LM, pH at 6 and 8 h in BF, and pH at 2 and 4 h in SM was significantly less (P < 0.01) with Step15 compared with Con15.
Table 2. Brine composition
Ingredient Amount, %
Water 81.29NaCl containing 0.58% nitrite 17.65Dextrose 0.82Ascorbate 0.24Total 100
Rosenvold et al.1834
at Serials/Acq. Dept., Library on November 24, 2011jas.fass.orgDownloaded from
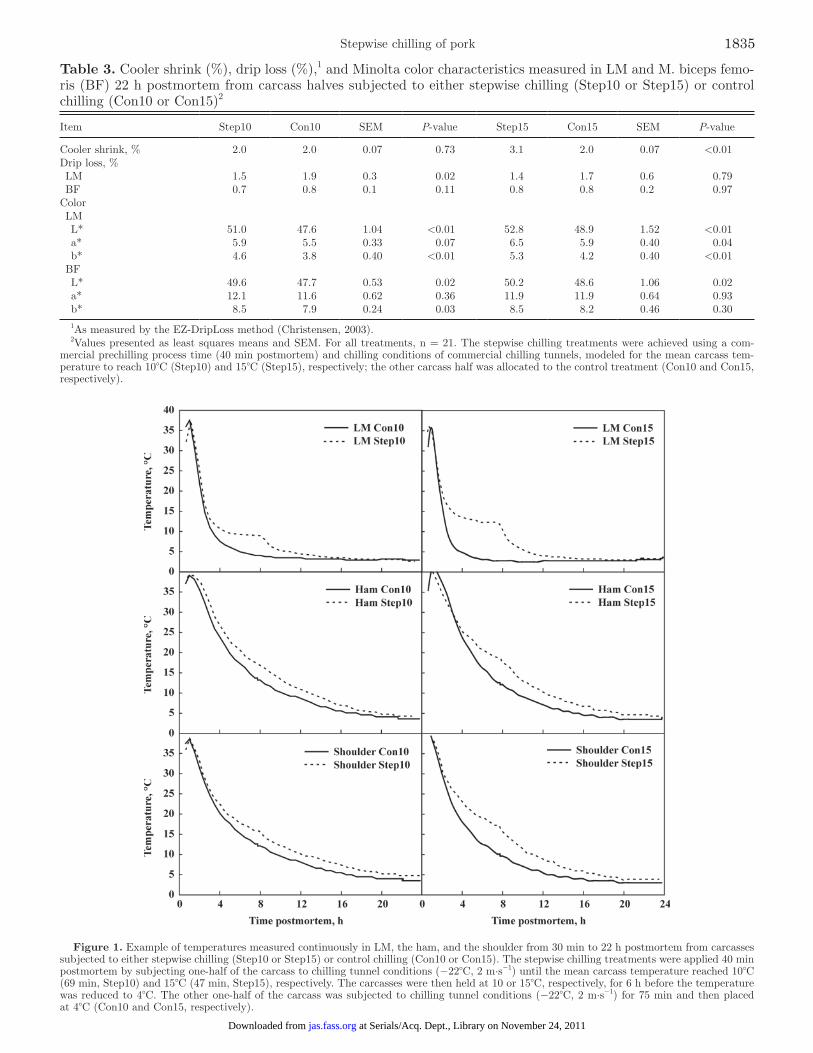
Table 3. Cooler shrink (%), drip loss (%),1 and Minolta color characteristics measured in LM and M. biceps femo-ris (BF) 22 h postmortem from carcass halves subjected to either stepwise chilling (Step10 or Step15) or control chilling (Con10 or Con15)2
Item Step10 Con10 SEM P-value Step15 Con15 SEM P-value
Cooler shrink, % 2.0 2.0 0.07 0.73 3.1 2.0 0.07 <0.01Drip loss, % LM 1.5 1.9 0.3 0.02 1.4 1.7 0.6 0.79 BF 0.7 0.8 0.1 0.11 0.8 0.8 0.2 0.97Color LM L* 51.0 47.6 1.04 <0.01 52.8 48.9 1.52 <0.01 a* 5.9 5.5 0.33 0.07 6.5 5.9 0.40 0.04 b* 4.6 3.8 0.40 <0.01 5.3 4.2 0.40 <0.01 BF L* 49.6 47.7 0.53 0.02 50.2 48.6 1.06 0.02 a* 12.1 11.6 0.62 0.36 11.9 11.9 0.64 0.93 b* 8.5 7.9 0.24 0.03 8.5 8.2 0.46 0.30
1As measured by the EZ-DripLoss method (Christensen, 2003).2Values presented as least squares means and SEM. For all treatments, n = 21. The stepwise chilling treatments were achieved using a com-
mercial prechilling process time (40 min postmortem) and chilling conditions of commercial chilling tunnels, modeled for the mean carcass tem-perature to reach 10°C (Step10) and 15°C (Step15), respectively; the other carcass half was allocated to the control treatment (Con10 and Con15, respectively).
Figure 1. Example of temperatures measured continuously in LM, the ham, and the shoulder from 30 min to 22 h postmortem from carcasses subjected to either stepwise chilling (Step10 or Step15) or control chilling (Con10 or Con15). The stepwise chilling treatments were applied 40 min postmortem by subjecting one-half of the carcass to chilling tunnel conditions (−22°C, 2 m·s−1) until the mean carcass temperature reached 10°C (69 min, Step10) and 15°C (47 min, Step15), respectively. The carcasses were then held at 10 or 15°C, respectively, for 6 h before the temperature was reduced to 4°C. The other one-half of the carcass was subjected to chilling tunnel conditions (−22°C, 2 m·s−1) for 75 min and then placed at 4°C (Con10 and Con15, respectively).
Stepwise chilling of pork 1835
at Serials/Acq. Dept., Library on November 24, 2011jas.fass.orgDownloaded from
Glycogen Degradation Postmortem
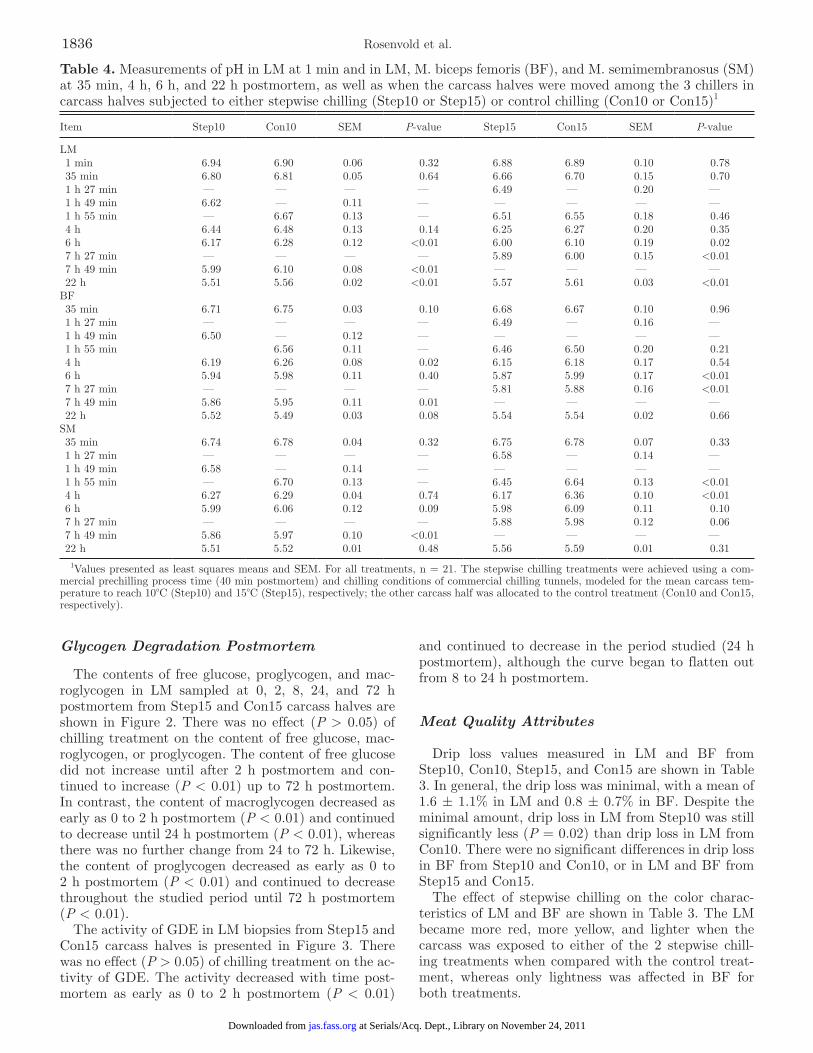
The contents of free glucose, proglycogen, and mac-roglycogen in LM sampled at 0, 2, 8, 24, and 72 h postmortem from Step15 and Con15 carcass halves are shown in Figure 2. There was no effect (P > 0.05) of chilling treatment on the content of free glucose, mac-roglycogen, or proglycogen. The content of free glucose did not increase until after 2 h postmortem and con-tinued to increase (P < 0.01) up to 72 h postmortem. In contrast, the content of macroglycogen decreased as early as 0 to 2 h postmortem (P < 0.01) and continued to decrease until 24 h postmortem (P < 0.01), whereas there was no further change from 24 to 72 h. Likewise, the content of proglycogen decreased as early as 0 to 2 h postmortem (P < 0.01) and continued to decrease throughout the studied period until 72 h postmortem (P < 0.01).
The activity of GDE in LM biopsies from Step15 and Con15 carcass halves is presented in Figure 3. There was no effect (P > 0.05) of chilling treatment on the ac-tivity of GDE. The activity decreased with time post-mortem as early as 0 to 2 h postmortem (P < 0.01)
and continued to decrease in the period studied (24 h postmortem), although the curve began to flatten out from 8 to 24 h postmortem.
Meat Quality Attributes
Drip loss values measured in LM and BF from Step10, Con10, Step15, and Con15 are shown in Table 3. In general, the drip loss was minimal, with a mean of 1.6 ± 1.1% in LM and 0.8 ± 0.7% in BF. Despite the minimal amount, drip loss in LM from Step10 was still significantly less (P = 0.02) than drip loss in LM from Con10. There were no significant differences in drip loss in BF from Step10 and Con10, or in LM and BF from Step15 and Con15.
The effect of stepwise chilling on the color charac-teristics of LM and BF are shown in Table 3. The LM became more red, more yellow, and lighter when the carcass was exposed to either of the 2 stepwise chill-ing treatments when compared with the control treat-ment, whereas only lightness was affected in BF for both treatments.
Table 4. Measurements of pH in LM at 1 min and in LM, M. biceps femoris (BF), and M. semimembranosus (SM) at 35 min, 4 h, 6 h, and 22 h postmortem, as well as when the carcass halves were moved among the 3 chillers in carcass halves subjected to either stepwise chilling (Step10 or Step15) or control chilling (Con10 or Con15)1
Item Step10 Con10 SEM P-value Step15 Con15 SEM P-value
LM 1 min 6.94 6.90 0.06 0.32 6.88 6.89 0.10 0.78 35 min 6.80 6.81 0.05 0.64 6.66 6.70 0.15 0.70 1 h 27 min — — — — 6.49 — 0.20 — 1 h 49 min 6.62 — 0.11 — — — — — 1 h 55 min — 6.67 0.13 — 6.51 6.55 0.18 0.46 4 h 6.44 6.48 0.13 0.14 6.25 6.27 0.20 0.35 6 h 6.17 6.28 0.12 <0.01 6.00 6.10 0.19 0.02 7 h 27 min — — — — 5.89 6.00 0.15 <0.01 7 h 49 min 5.99 6.10 0.08 <0.01 — — — — 22 h 5.51 5.56 0.02 <0.01 5.57 5.61 0.03 <0.01BF 35 min 6.71 6.75 0.03 0.10 6.68 6.67 0.10 0.96 1 h 27 min — — — — 6.49 — 0.16 — 1 h 49 min 6.50 — 0.12 — — — — — 1 h 55 min 6.56 0.11 — 6.46 6.50 0.20 0.21 4 h 6.19 6.26 0.08 0.02 6.15 6.18 0.17 0.54 6 h 5.94 5.98 0.11 0.40 5.87 5.99 0.17 <0.01 7 h 27 min — — — — 5.81 5.88 0.16 <0.01 7 h 49 min 5.86 5.95 0.11 0.01 — — — — 22 h 5.52 5.49 0.03 0.08 5.54 5.54 0.02 0.66SM 35 min 6.74 6.78 0.04 0.32 6.75 6.78 0.07 0.33 1 h 27 min — — — — 6.58 — 0.14 — 1 h 49 min 6.58 — 0.14 — — — — — 1 h 55 min — 6.70 0.13 — 6.45 6.64 0.13 <0.01 4 h 6.27 6.29 0.04 0.74 6.17 6.36 0.10 <0.01 6 h 5.99 6.06 0.12 0.09 5.98 6.09 0.11 0.10 7 h 27 min — — — — 5.88 5.98 0.12 0.06 7 h 49 min 5.86 5.97 0.10 <0.01 — — — — 22 h 5.51 5.52 0.01 0.48 5.56 5.59 0.01 0.31
1Values presented as least squares means and SEM. For all treatments, n = 21. The stepwise chilling treatments were achieved using a com-mercial prechilling process time (40 min postmortem) and chilling conditions of commercial chilling tunnels, modeled for the mean carcass tem-perature to reach 10°C (Step10) and 15°C (Step15), respectively; the other carcass half was allocated to the control treatment (Con10 and Con15, respectively).
Rosenvold et al.1836
at Serials/Acq. Dept., Library on November 24, 2011jas.fass.orgDownloaded from
Texture Characteristics
To understand the background of the sensory evalu-ation of tenderness, the sarcomere length, MFI, and degradation of desmin in LM and the Warner-Bratzler shear force of LM and BF were measured from Step15
and Con15 carcass halves (Table 5). The sarcomere length was 1.80 μm for both Step15 and Con15, sug-gesting that neither the stepwise chilling treatment nor the control treatment induced cold shortening. In con-trast, the MFI, measured on LM samples after 3 d of aging, showed a larger MFI value in samples exposed to stepwise chilling (P = 0.04), suggesting a greater proteolysis in these samples compared with the control chilling. This was supported by a tendency toward a larger degradation of desmin at 72 h postmortem (P = 0.08) in Step15 compared with Con15, whereas there was no significant effect (P > 0.05) on desmin degra-dation after 8 and 24 h postmortem. Warner-Bratzler shear force values for the LM samples from Step15 were less compared with those from Con15 (P < 0.01). There was also a tendency for reduced thawing loss in Step15 compared with Con15 (P = 0.06), but there was no ef-fect on cooking loss. In contrast, for the BF, there was no difference (P > 0.05) for any attributes. For Step10 and Con10 samples, Warner-Bratzler shear force values for LM and BF showed the same pattern as for Step15 and Con15 (data not shown).
Ham Process Yield
Process yields after multineedle injection of brine and tumbling, as well as the total process yield after cooking of hams produced from SM are shown in Table 6. There was no significant (P > 0.05) effect of stepwise chill-ing on yield at any time. A yield loss of approximately 3% is normal for this product type (unpublished data, Danish Meat Research Institute, Roskilde, Denmark).
Figure 3. Glycogen debranching enzyme (GDE) activity in LM samples from carcasses subjected to either stepwise chilling (Step15, n = 21) or control chilling (Con 15, n = 21). The stepwise chilling treatments were applied 40 min postmortem by subjecting one-half of the carcass to chilling tunnel conditions (−22°C, 2 m·s−1) until the mean carcass temperature reached 10°C (69 min, Step10) and 15°C (47 min, Step15), respectively. The carcasses were then held at 10 or 15°C, respectively, for 6 h before the temperature was reduced to 4°C. The other one-half of the carcass was subjected to chilling tunnel conditions (−22°C, 2 m·s−1) for 75 min and then placed at 4°C (Con10 and Con15, respectively). Values presented are least squares means and SEM. Differences between all times postmortem: P < 0.05. abs = absorbance.
Figure 2. Content of free glucose, proglycogen, and macroglycogen in LM samples from carcasses subjected to either stepwise chilling (Step15, n = 21) or control chilling (Con15, n = 21). The stepwise chilling treatments were applied 40 min postmortem by subjecting one-half of the carcass to chilling tunnel conditions (−22°C, 2 m·s−1) until the mean carcass temperature reached 10°C (69 min, Step10) and 15°C (47 min, Step15), respectively. The carcasses were then held at 10 or 15°C, respectively, for 6 h before the temperature was reduced to 4°C. The other one-half of the carcass was subjected to chilling tunnel conditions (−22°C, 2 m·s−1) for 75 min and then placed at 4°C (Con10 and Con15, respectively). Values presented are least squares means and SEM. Free glucose: differences between all times postmortem (P < 0.01), except at 0 and 2 h postmortem. Proglycogen: differences between all times postmortem (P < 0.01). Macroglycogen: differences between all times postmortem (P < 0.01), except 24 and 72 h post-mortem.
Stepwise chilling of pork 1837
at Serials/Acq. Dept., Library on November 24, 2011jas.fass.orgDownloaded from
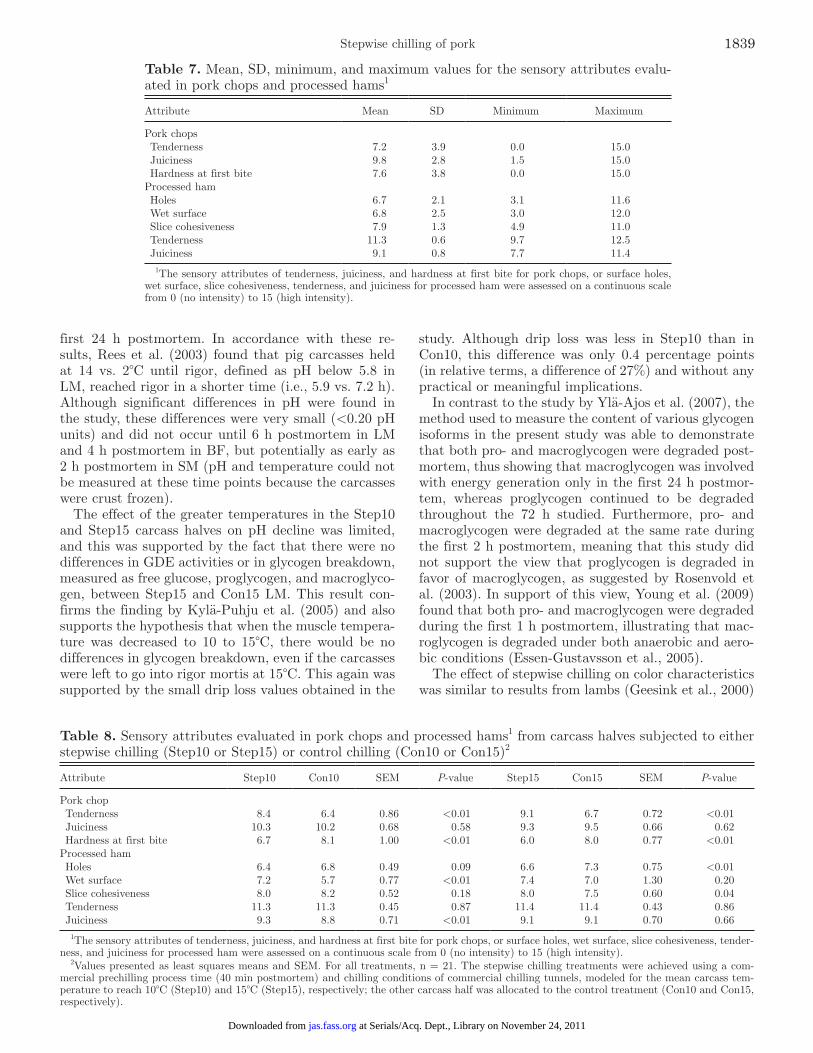
Sensory Analysis
Pork Chops. The mean, SD, minimum, and maxi-mum values for the sensory attributes evaluated for pork chops are presented in Table 7. The sensory analy-sis showed that LM from Step10 was significantly more tender than LM from Con10, with a difference of al-most 2 sensory units (P < 0.01). The sensory values for the LM from Step15 were also significantly more tender than those from Con15, with a difference in tenderness of more than 2 sensory units (P < 0.01). Hardness at first bite supported the tenderness results. Juiciness of the meat was not affected by stepwise chilling (Table 8).
Processed Hams. The mean, SD, minimum, and maximum values for the sensory attributes evaluated for the processed hams are shown in Table 7. There was a certain variation in the attributes of holes and wet surface, whereas the variation was minimal in the at-tributes of slice cohesiveness, tenderness, and juiciness. The processed ham from Step10 carcass halves had a wetter surface (P < 0.01) and were juicier (P < 0.01) than the ham from Con10 carcass halves. Ham slices from Step15 carcass halves had fewer holes (P < 0.01)
and the slices were more cohesive (P = 0.04) compared with those from Con15 carcass halves.
DISCUSSION
The 2 stepwise chilling treatments (Step10 and Step15) investigated in this study were modeled for the mean time to reach carcass temperatures of 10 and 15°C as soon as possible after slaughter (1 h 49 min, and 1 h 27 min, respectively). The carcasses were then held at their respective 10 or 15°C temperatures for 6 h to allow the muscle to enter rigor mortis at these temperatures before the mean carcass temperature was decreased to 4°C. The temperature in the center of the LM was quite close to the target mean carcass tempera-ture. The temperatures in the ham and shoulder were greater than the target mean temperature because of the thickness of the muscle, but a clear difference was observed between the Step10 or Step15 and the Con10 or Con15 muscles. We evaluated that the targeted chill-ing treatments were successfully obtained.
Both stepwise chilling treatments resulted in smaller pH values than their control counterparts during the
Table 5. Sarcomere length, myofibrillar fragmentation index (MFI), and desmin deg-radation at 8, 24, and 72 h postmortem in LM, and Warner-Bratzler shear force, thaw, and cooking loss in LM and M. biceps femoris (BF) from carcass halves subjected to either stepwise chilling (Step15) or control chilling (Con15)1
Item Step15 Con15 SEM P-value
LM Sarcomere length, μm 1.80 1.80 0.09 0.87 MFI 102 95 6.0 0.04 Desmin degradation Intact desmin 8 h postmortem, % 93.1 101.4 5.5 0.30 Intact desmin 24 h postmortem, % 76.5 90.5 7.4 0.18 Intact desmin 72 h postmortem, % 38.8 53.3 6.2 0.08 Warner-Bratzler shear force, kg 3.1 4.0 0.5 <0.01 Thaw loss, % 5.4 6.1 0.3 0.06 Cooking loss, % 19.7 20.2 0.6 0.12BF Warner-Bratzler shear force, kg 3.6 3.7 0.1 0.73 Thaw loss, % 5.5 5.7 0.4 0.43 Cooking loss, % 23.0 23.1 1.2 0.91
1Values presented as least squares means and SEM. For both treatments, n = 21. The stepwise chilling treat-ment was achieved using a commercial prechilling process time (40 min postmortem) and chilling conditions of commercial chilling tunnels, modeled for the mean carcass temperature to reach 15°C (Step15), respectively; the other carcass half was allocated to the control treatment (Con15).
Table 6. Yield after brine injection and tumbling, and total processes yield of ham produced from M. semimem-branosus (SM) from carcass halves subjected to either stepwise chilling (Step10 or Step15) or control chilling (Con10 or Con15)1
Item Step10 Con10 SEM P-value Step15 Con15 SEM P-value
Yield after brine injection, % 11.5 11.0 0.46 0.21 11.4 11.1 0.29 0.58Yield after tumbling, % 13.9 13.5 0.55 0.33 14.3 13.7 0.45 0.15Total yield, % −0.6 −3.3 1.8 0.16 −2.6 −2.7 0.46 0.72
1Values presented as least squares means and SEM. For all treatments, n = 21. The stepwise chilling treatments were achieved using a com-mercial prechilling process time (40 min postmortem) and chilling conditions of commercial chilling tunnels, modeled for the mean carcass tem-perature to reach 10°C (Step10) and 15°C (Step15), respectively; the other carcass half was allocated to the control treatment (Con10 and Con15, respectively).
Rosenvold et al.1838
at Serials/Acq. Dept., Library on November 24, 2011jas.fass.orgDownloaded from
first 24 h postmortem. In accordance with these re-sults, Rees et al. (2003) found that pig carcasses held at 14 vs. 2°C until rigor, defined as pH below 5.8 in LM, reached rigor in a shorter time (i.e., 5.9 vs. 7.2 h). Although significant differences in pH were found in the study, these differences were very small (<0.20 pH units) and did not occur until 6 h postmortem in LM and 4 h postmortem in BF, but potentially as early as 2 h postmortem in SM (pH and temperature could not be measured at these time points because the carcasses were crust frozen).
The effect of the greater temperatures in the Step10 and Step15 carcass halves on pH decline was limited, and this was supported by the fact that there were no differences in GDE activities or in glycogen breakdown, measured as free glucose, proglycogen, and macroglyco-gen, between Step15 and Con15 LM. This result con-firms the finding by Kylä-Puhju et al. (2005) and also supports the hypothesis that when the muscle tempera-ture was decreased to 10 to 15°C, there would be no differences in glycogen breakdown, even if the carcasses were left to go into rigor mortis at 15°C. This again was supported by the small drip loss values obtained in the
study. Although drip loss was less in Step10 than in Con10, this difference was only 0.4 percentage points (in relative terms, a difference of 27%) and without any practical or meaningful implications.
In contrast to the study by Ylä-Ajos et al. (2007), the method used to measure the content of various glycogen isoforms in the present study was able to demonstrate that both pro- and macroglycogen were degraded post-mortem, thus showing that macroglycogen was involved with energy generation only in the first 24 h postmor-tem, whereas proglycogen continued to be degraded throughout the 72 h studied. Furthermore, pro- and macroglycogen were degraded at the same rate during the first 2 h postmortem, meaning that this study did not support the view that proglycogen is degraded in favor of macroglycogen, as suggested by Rosenvold et al. (2003). In support of this view, Young et al. (2009) found that both pro- and macroglycogen were degraded during the first 1 h postmortem, illustrating that mac-roglycogen is degraded under both anaerobic and aero-bic conditions (Essen-Gustavsson et al., 2005).
The effect of stepwise chilling on color characteristics was similar to results from lambs (Geesink et al., 2000)
Table 7. Mean, SD, minimum, and maximum values for the sensory attributes evalu-ated in pork chops and processed hams1
Attribute Mean SD Minimum Maximum
Pork chops Tenderness 7.2 3.9 0.0 15.0 Juiciness 9.8 2.8 1.5 15.0 Hardness at first bite 7.6 3.8 0.0 15.0Processed ham Holes 6.7 2.1 3.1 11.6 Wet surface 6.8 2.5 3.0 12.0 Slice cohesiveness 7.9 1.3 4.9 11.0 Tenderness 11.3 0.6 9.7 12.5 Juiciness 9.1 0.8 7.7 11.4
1The sensory attributes of tenderness, juiciness, and hardness at first bite for pork chops, or surface holes, wet surface, slice cohesiveness, tenderness, and juiciness for processed ham were assessed on a continuous scale from 0 (no intensity) to 15 (high intensity).
Table 8. Sensory attributes evaluated in pork chops and processed hams1 from carcass halves subjected to either stepwise chilling (Step10 or Step15) or control chilling (Con10 or Con15)2
Attribute Step10 Con10 SEM P-value Step15 Con15 SEM P-value
Pork chop Tenderness 8.4 6.4 0.86 <0.01 9.1 6.7 0.72 <0.01 Juiciness 10.3 10.2 0.68 0.58 9.3 9.5 0.66 0.62 Hardness at first bite 6.7 8.1 1.00 <0.01 6.0 8.0 0.77 <0.01Processed ham Holes 6.4 6.8 0.49 0.09 6.6 7.3 0.75 <0.01 Wet surface 7.2 5.7 0.77 <0.01 7.4 7.0 1.30 0.20 Slice cohesiveness 8.0 8.2 0.52 0.18 8.0 7.5 0.60 0.04 Tenderness 11.3 11.3 0.45 0.87 11.4 11.4 0.43 0.86 Juiciness 9.3 8.8 0.71 <0.01 9.1 9.1 0.70 0.66
1The sensory attributes of tenderness, juiciness, and hardness at first bite for pork chops, or surface holes, wet surface, slice cohesiveness, tender-ness, and juiciness for processed ham were assessed on a continuous scale from 0 (no intensity) to 15 (high intensity).
2Values presented as least squares means and SEM. For all treatments, n = 21. The stepwise chilling treatments were achieved using a com-mercial prechilling process time (40 min postmortem) and chilling conditions of commercial chilling tunnels, modeled for the mean carcass tem-perature to reach 10°C (Step10) and 15°C (Step15), respectively; the other carcass half was allocated to the control treatment (Con10 and Con15, respectively).
Stepwise chilling of pork 1839
at Serials/Acq. Dept., Library on November 24, 2011jas.fass.orgDownloaded from

exposed to increased prerigor temperatures from 5 to 35°C when the color was measured 1 d postmortem, whereas the color differences had disappeared at 14 d postmortem after aging at 2°C. The variation in color characteristics in the present study was most probably related to the changes in myoglobin forms, which were affected by temperature and pH, and to the internal reflectance (Lindahl et al., 2001) because the content of pigment within the carcass would not change with vari-ous chilling procedures. Thus, it seems that a greater fraction of oxymyoglobin is responsible for the lighter and more yellow meat from stepwise-chilled meat. How-ever, as seen in the study by Geesink et al. (2000), this effect may not persist after extended storage.
The SM was processed into ham by using a commer-cial recipe containing NaCl as the only binder. This was chosen specifically to explore whether the stepwise chill-ing had affected the water-binding capacity of the meat without being masked by other binding agents (e.g., phosphates). Because neither the processing yield nor the sensory quality was negatively affected by stepwise chilling, this outcome supports the view that stepwise chilling did not compromise the WHC of the meat.
The sarcomere lengths were relatively long, 1.8 μm for both Step15 and Con15, indicating that neither the control chilling nor the stepwise chilling induced cold shortening. Thus, the differences in shear force and sensory tenderness between the control and stepwise-chilled carcasses were not caused by cold shortening, confirming that the current industry practice of blast chilling does not induce cold shortening in LM.
Tenderness of LM (smaller shear force values and greater MFI values) was improved by more than 2 sen-sory units for both stepwise chilling treatments and was accompanied by a tendency toward increased desmin degradation, indicating increased proteolysis in LM for stepwise chilling. This agrees with the results of Rees et al. (2003), who found increased MFI values in LM stored at 14 vs. 2°C until rigor, and supports increased proteolysis as the reason for improved tenderness in the stepwise-chilled carcasses. This is consistent with the results of Josell et al. (2004), who reported im-proved tenderness upon slow chilling, compared with fast-chilled pork carcasses, again with no effect on sarcomere length. It would appear that the stepwise chilling methods used here result in an increased activ-ity of proteolytic enzymes (e.g., the calpain system), stimulating the activity of the enzymes and increasing the degradation of the inhibiting protein calpastatin because of the greater temperature, as suggested by Zamora et al. (1998). Further studies to elucidate the full protein degradation pattern postmortem and the enzymes involved would be very valuable.
Food safety and microbial shelf life are important aspects of the chilling process and should be investi-gated before the proposed stepwise chilling treatments can be implemented commercially. Before conducting this study, we determined that the suggested treat-ments would not cause a food safety or microbial shelf
life risk. This conclusion was based on the fact that the surface of the carcass decreased rapidly to −20°C once the carcass was placed in chiller 1. The surface temperature then increased to 10 to 15°C while the car-casses were held at 10 or 15°C, but it decreased again to −20°C once the carcasses were placed in chiller 1 for the second time for the mean carcass temperature to decrease to 4°C before finally being placed in chiller 3 (4°C). A subsequent study was conducted in a commer-cial slaughter plant and therefore had to omit the step in which the carcasses were placed in the tunnel chiller after being held at 10°C for 6 h. That study investi-gated the effects of stepwise chilling on food safety and microbial shelf life, and confirmed that stepwise chilling had no negative effects on carcass microbiology (un-published data, L. Kristensen, Danish Meat Research Institute, Roskilde, Denmark).
The hypothesis that a stepwise chilling method com-posed of a rapid temperature reduction to 10 or 15°C and a 6-h holding period at 10 or 15°C, followed by rapid chilling to 4°C, would improve the tenderness of pork without compromising the WHC was tested in this study. Not only was the tenderness of LM significantly improved, but there was also no compromise regarding any quality indicators or attributes, such as pH, drip loss, or ham processing yield, although it should be noted that the color of the stepwise-chilled pork was affected. Neither the tenderness of processed SM nor the shear force of BF was affected, possibly because of the smaller temperature differences in these muscles. Furthermore, the results for glycogen metabolism suc-cessfully revealed that both pro- and macroglycogen contribute to the energy generation in postmortem muscles, with degradation of both forms early post-mortem.
It is traditionally believed that pig carcasses must be chilled down to 4°C as fast as possible postmortem to prevent PSE and reduce drip loss and cooler shrink. In contrast, this study shows it is possible to hold car-casses at temperatures optimal for tenderization dur-ing the chilling process without significantly affecting WHC and cooler shrink, and this will lead to improved tenderness. The authors believe that there is further scope to optimize the slaughter and chilling process for quality through the application of well-established meat science principles combined with new discoveries.
LITERATURE CITED
Bendall, J. R. 1973. Postmortem changes in muscle. Pages 243–309 in Structure and Function of Muscle. Vol. 2. G. H. Bourne, ed. Academic Press, New York, NY.
Brown, D. H., and B. I. Brown. 1966. Enzymes of glycogen deb-ranching: Amylo-1,6-glucosidase (I) and oligo-1,4→1,4-glucantransferase (II). Pages 515–524 in Methods in Enzymol-ogy. Vol. 8. S. P. Colowick and N. O. Kaplan, ed. Academic Press, New York, NY.
Christensen, L. B. 2003. Drip loss sampling in porcine M. longis-simus dorsi. Meat Sci. 63:469–477.
Culler, R. D., F. C. Parrish, G. C. Smith, and H. R. Cross. 1978. Relationship of myofibril fragmentation index to certain chemi-
Rosenvold et al.1840
at Serials/Acq. Dept., Library on November 24, 2011jas.fass.orgDownloaded from

cal, physical and sensory characteristics of bovine longissimus muscle. J. Food Sci. 43:1177–1180.
Devine, C. E., D. L. Hopkins, I. H. Hwang, D. M. Ferguson, and I. Richards. 2004. Electrical stimulation. Pages 413–423 in Ency-clopedia of Meat Sciences. M. Dikeman, W. Jensen, C. Devine, and M. Dikeman, ed. Elsevier, Oxford, UK.
Essén-Gustavsson, B., M. Jensen-Waern, R. Jonasson, and L. An-dersson. 2005. Effect of exercise on proglycogen and macrogly-cogen content in skeletal muscles of pigs with the Rendement Napole mutation. Am. J. Vet. Res. 66:1197–1201.
Geesink, G. H., A. D. Bekhit, and R. Bickerstaffe. 2000. Rigor tem-perature and meat quality characteristics of lamb longissimus muscle. J. Anim. Sci. 78:2842–2848.
Hansen, S., M. Therkildsen, and D. V. Byrne. 2006. Effects of a com-pensatory growth strategy on sensory and physical properties of meat from young bulls. Meat Sci. 74:628–643.
Honikel, K. O. 1998. Reference methods for the assessment of physi-cal characteristics of meat. Meat Sci. 49:447–457.
Josell, Å., G. von Seth, and E. Tornberg. 2004. Sensory and meat quality traits of pork in relation to post-slaughter treatment and RN genotype. Meat Sci. 66:113–124.
Kristensen, L., M. Therkildsen, M. D. Aaslyng, N. Oksbjerg, and P. Ertbjerg. 2004. Compensatory growth improves meat tender-ness in gilts but not in barrows. J. Anim. Sci. 82:3617–3624.
Kylä-Puhju, M., M. Ruusunen, and E. Puolanne. 2005. Activity of porcine muscle glycogen debranching enzyme in relation to pH and temperature. Meat Sci. 69:143–149.
Lindahl, G., K. Lundström, and E. Tornberg. 2001. Contribution of pigment content, myoglobin forms and internal reflectance to the colour of pork loin and ham from pure breed pigs. Meat Sci. 59:141–151.
Locker, R. H., and C. J. Hagyard. 1963. A cold shortening effect in beef muscles. J. Sci. Food Agric. 14:787–793.
Maribo, H., E. V. Olsen, P. Barton-Gade, A. J. Møller, and A. Karls-son. 1998. Effect of early post-mortem cooling on temperature, pH fall and meat quality in pigs. Meat Sci. 50:115–129.
Nelson, T. E., D. H. Palmer, and J. Larner. 1970. An investigation of the properties of rabbit muscle oligo-1,4→1,4-glucantransferase. Biochim. Biophys. Acta 212:269–280.
Rees, M. P., G. R. Trout, and R. D. Warner. 2003. The influence of the rate of pH decline on the rate of ageing for pork. II: Interac-tion with chilling temperature. Meat Sci. 65:805–818.
Rosenvold, K., B. Essen-Gustavsson, and H. J. Andersen. 2003. Di-etary manipulation of pro- and macroglycogen in porcine skel-etal muscle. J. Anim. Sci. 81:130–134.
Schäfer, A., K. Rosenvold, P. P. Purslow, H. J. Andersen, and P. Henckel. 2002. Physiological and structural events post mortem of importance for drip loss in pork. Meat Sci. 61:355–366.
Smulders, F. J. M., F. Toldra, J. Flores, and M. Prieto. 1992. New technologies for meat and meat products. Audet Tijdschr. 182:186–188.
Sørheim, O., and K. I. Hildrum. 2002. Muscle stretching techniques for improving meat tenderness. Trends Food Sci. Technol. 13:127–135.
Støier, S., M. D. Aaslyng, E. V. Olsen, and P. Henckel. 2001. The ef-fect of stress during lairage and stunning on muscle metabolism and drip loss in Danish pork. Meat Sci. 59:127–131.
Taylor, A. A., and S. J. Dant. 1971. Influence of carcass cooling rate on drip loss in pigment. J. Food Technol. 6:131–139.
Therkildsen, M., L. M. Larsen, H. G. Bang, and M. Vestergaard. 2002. Effect of growth rate on tenderness development and final tenderness of meat from Friesian calves. Anim. Sci. 74:253–264.
Towbin, H., T. Staehelin, and J. Gordon. 1979. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets—Procedure and some applications. Proc. Natl. Acad. Sci. USA 76:4350–4354.
Wheeler, T. L., and M. Koohmaraie. 1999. The extent of proteolysis is independent of sarcomere length in lamb longissimus and psoas major. J. Anim. Sci. 77:2444–2451.
Wismer-Pedersen, J. 1987. Water. Pages 141–154 in The Science of Meat and Meat Products. J. F. Price and B. S. Schweigert, ed. W. H. Freeman and Co., San Francisco, CA.
Wismer-Pedersen, J., and E. J. Briskey. 1961. Rate of anaerobic gly-colysis versus structure in pork muscle. Nature 189:318–320.
Ylä-Ajos, M. S. K., G. Lindahl, J. F. Young, P. K. Theil, E. Puo-lanne, A. C. Enfält, H. J. Andersen, and N. Oksbjerg. 2007. Post-mortem activity of the glycogen debranching enzyme and change in the glycogen pools in porcine M. longissimus dorsi from carriers and non-carriers of the RN− gene. Meat Sci. 75:112–119.
Young, J. F., H. C. Bertram, and N. Oksbjerg. 2009. Rest before slaughter ameliorates pre-slaughter stress-induced increased drip loss but not stress-induced increase in texture of pork. Meat Sci. 83:634–641.
Zamora, F., F. Chaïb, and E. Dransfield. 1998. Calpains and cal-pastatin from cold-shortened bovine M. longissimus lumborum. Meat Sci. 49:127–133.
Stepwise chilling of pork 1841
at Serials/Acq. Dept., Library on November 24, 2011jas.fass.orgDownloaded from

Referenceshttp://jas.fass.org/content/88/5/1830#BIBLThis article cites 27 articles, 5 of which you can access for free at:
at Serials/Acq. Dept., Library on November 24, 2011jas.fass.orgDownloaded from