studies on the regulation of insulin-like growth factor binding protein 3 secretion in human...
TRANSCRIPT

JOURNAL OF BONE AND MINERAL RESEARCH Volume 9, Number 6,1994 Mary Ann Liebert, he., Publishers
Studies on the Regulation of Insulin-like Growth Factor Binding Protein 3 Secretion in Human Osteosarcoma Cells In Vitro
Y. NAKAO, S. H[LLIKER, D.J. BAYLINK, and S. MOHAN
ABSTRACT
Previous studies demonstrated that insulin-like growth factors (IGFs) are important autocrine and paracrine mitogens for human bone cells in vitro and that IGF binding proteins (IGFBPs) are important regulators of the biologic actions of IGFs. Thus, the actions of IGFs may be determined not only by their concentrations but also by the type and amount of IGFBPs produced by human bone cells at a local site in bone. In this study, we sought to determine the effects of dexamethasone, 1,25-(OH), D3, and parathyroid hormone (PTH) on the secretion of IGFBP-3 in human osteosarcoma cell limes. Serum-free cultures of low- and high-alkaline phosphatase (ALP) SaOS-2, MG-63, and TES9 human osteosarcoma cells were treated for 24 or 48 h with the effectors and the conditioned media used for determination of IGFBP-3 using a radioimmunoassay. We report that (1) the basal rate of IGFBP-3 secretion (ng/mg cellular protein) was dependent upon cell type, with TES9 > low-ALP Saw- 2 > MG-63 > high-ALP SaOS-2 cells, and did not correlate with either basal cell proliferation or basal cellular ALP activity; (2) dexamethasone (10-'2-10-7 M) inhibited IGFBP-3 secretion in a dose-dependent manner in low-ALP SaOS-2, MG-63, and TES9 cells but not in high-ALP SaOS-2 cells; (3) 1,25-(OH),D3 (10-"-10-* M) stimulated IGFBP-3 secretion in a dose-dependent manner in MG-63,low-ALP SaOS-2, and high-ALP SaOS-2 cells, and the coaddition of TGF-f3 and 1 ,25-(OH),D3 increased synergistically IGFBP-3 secretion and cellular ALP activity in MG-63 cells; and (4) human FTH-(1-34) (0.1-100 ng/ml) had no significant effect on IGFBP-3 secretion in MG-63,low-ALP SaOS-2, or high-ALP SaOS-2 cells. We conclude that such agents as dexametha- sone, 1,25-(OH)zD3, and PTH differentially regulate IGFBP-3 secretion in human osteosarcoma cells in vitro.
INTRODUCTION
E AND OTHERS have proposed that insulin-like growth W factors (IGFs) may play important roles in the local regulation of bone metabolism.('-3) Their site of production and activity support such a role. First, IGF-I and I1 have been found in bone extracts of several different species and in conditioned media of human bone cells. Second, IGF-I1 mRNA transcripts have been detected in cultured human bone cells. Third, the exogenous addition of IGFs has been shown to stimulate both proliferative and differentiated functions in human bone cells.('*2' In addition, the activity of IGFs in different tissues, including bone. may be modulated by a family of structurally related proteins that specifically bind to IGFs. Six different IGF
binding proteins (designated IGFBP-1 through IGFBP-6) have been purified from human plasma or tissues, including bone.'4' In previous studies, we showed that human bone cells in culture under serum-free conditions secrete a number of IGFBPs (25, 29,34, and 38.5-41.5 kD forms), of which the 25 kD IGFBP-4 and 38.5-41.5 kD IGFBP-3 are the two major form^.'^*^*^' Studies on the biologic actions of IGFBPs suggest that they play important roles in bone metabolism by regulating IGF activities in both a positive and a negative manner.('*') Thus. it has been proposed that the activity of IGFs at a site in bone is determined not only by the concentration of IGFs but also by the amounts of various IGFBPs produced by bone cells at that site. Moreover, systemic effectors, such as glucocorticoids (e.g., dexametha- sone) and osteotropic hormones [e.g., parathyroid hormone,
Departments of Medicine, Biochemistry, and Physiology, lioma Linda University and Jerry L. Pettis Veterans Administration Hospital. Loma Linda, California.
865

866 NAKAO ET AL.
PTH, and 1,25-dihydroxyvitamin D,, 1 ,25-(OH),D3], may also influence bone cell proliferation and differentiation through modulation of the IGF-induced cellular r e ~ p o n s e . ( ~ - ' ~ )
We and others have begun to test the hypothesis that systemic effectors modulate the IGF-induced cellular response by regu- lating the balance between the major IGFBPs. Schmid and colleagues reported that IGFBP-3 is increased by human growth hormone and estradiol in rat calvarial cells"4' and is increased by growth hormone and IGF-I but decreased by cortisol in a bone-derived rat cell line (PyMS)."" Moriwake and col- leagues"6' reported that 1,25-(OH),D, stimulated the secretion of IGFBP-3 in a human osteosarcoma cell line (MG-63). Recently, we demonstrated that 1 ,25-(OH)*D3 stimulated the production of the inhibitory IGFBP-4 in several normal human bone and osteosarcoma cell types'17) and that PTH stimulated IGFBP-4 protein and mRNA levels in normal human bone cells."*' The current study was undertaken to determine whether IGFBP-3 secretion can be systemically (and possibly differentially) regulated in human osteosarcoma cell types that differ in their relative rates of cell proliferation and alkaline phosphatase activity. We report the effects of a glucocorticoid (dexamethasone) and of the osteotropic hormones PTH and 1,25-(OH),D, (alone and combined with transforming growth factor-@ [TGF-@I) on IGFBP-3 secretion in MG-63, TE89, and low- and high-alkaline phosphatase (ALP) subpopulations of SaOS-2 human osteosarcoma cells.
MATERIALS AND METHODS
Materials
MG-63 and TE89 human osteosarcoma cell lines were ob- tained from American Type Culture Collection (Rockville, MD); low-ALP SaOS-2 and high-ALP SaOS-2 human osteo- sarcoma cell line subpopulations, which exhibit characteristics of less mature and more mature osteoblast phenotype, respec- tively, were isolated as described."" Dulbecco's modified Eagle's medium (DMEM) with and without phenol red and trypsin were purchased from GIBCO (Santa Clara, CA). Calf serum was purchased from HyClone (Logan, UT), and tissue culture dishes were obtained from Costar (Cambridge, MA) and Coming Glass Works (Coming, NY). Human PTH [hPTH-( 1- 34)] was purchased from Bachem Corp. (Torrance, CA), bovine serum albumin (BSA), radioimmunoassay (RIA) grade, was supplied by Fluka (Ronkonkoma, NJ), and 1 ,25-(OH),D3 was a gift from Dr. M. Uskokovic (Hoffmann-LaRoche, Nutley, NJ). TGF-P was purchased from Austral Biologicals (San Ramon, CA). For the IGFBP-3 RIA, nonglycosylated human recombi- nant IGFBP-3 and rabbit antihuman IGFBP-3 polyclonal anti- bodies were kindly donated by Dr. A. Sommer (Celtrix Pharma- ceuticals, Santa Clara, CA) and Dr. V. Mukku (Genentech, Inc., San Francisco, CA), respectively. Goat antirabbit IgG and normal rabbit serum were provided by Pel-Freez Biologicals (Rogers, AK). IGFBP-3 tracer was prepared using Na['251] purchased from the International Chemical and Nuclear Corp. (Irvine, CA). All other chemicals were obtained from the Sigma Chemical Co. (St. Louis, MO).
Cell culture and collection of conditioned medium
Osteoblast-like human osteosarcoma cell lines (MG-63, TE89, and low- and high-ALP SaOS-2) were maintained in DMEM supplemented with 10% calf serum under the conditions of 5% CO, and air atmosphere and saturated humidity at 37°C. Upon reaching confluence, cells were trypsinized. When suffi- cient cells were available, cells were plated at 5.6 x lo5 cells/60 mm culture dish and incubated in DMEM supplemented with 2% calf serum at 37°C for 24 h, followed by serum-free incubation in DMEM supplemented with 0.1% BSA at 37°C for 24 h. The media were then removed and replaced with 2 ml DMEM containing the effectors. DMEM containing 0.1% BSA was used as vehicle for hPTH-(1-34) (0.1-100 ng/ml) and dexamethasone (10-'2-10-7 M) and also as the control for the effectors. DMEM containing 0.01% ethyl alcohol and 0.1% BSA was used as vehicle for 1 ,25-(OH),D3 ( 10-'L-10-8 M) and also as the control for this effector. For the experiments using dexamethasone, phenol red-free DMEM and water-solu- ble dexamethasone (Sigma Chemical Co., D-2915) were used, since phenol red has been shown to mimic the biologic actions of certain steroid hormones on osteoblast proliferation." At differ- ent times of incubation, the conditioned media were collected in BSA-precoated tubes to decrease the nonspecific binding of IGFBPs to the plastic tube and stored at -20°C for IGFBP-3 RIA. Culture dishes were rinsed with phosphate- buffered saline and stored at -20°C for determination of cellular protein by the Lowry et al. method.'**'
IGFBP-3 radioimmunoassay
For IGFBP-3 RIA, conditioned medium samples were col- lected in BSA-precoated polypropylene tubes and, if necessary, concentrated by SpeedVac (Savant Instruments, Hicksville, NY) centrifugation before assay. IGFBP-3 levels were deter- mined using human recombinant IGFBP-3 as the standards and radioactive tracer and rabbit antihuman IGFBP-3 polyclonal antiserum. Briefly, 0.1 ml sample was incubated with 0.2 mi RIA buffer (0.03 M sodium phosphate buffer, pH 7.5, supple- mented with 0.25% BSA, 0.02% protamine sulfate, 0.02% sodium azide, and 0.01 M EDTA) and 0.1 ml rabbit antihuman IGFBP-3 antiserum (32,000-fold dilution in RIA buffer) for I h at room temperature. About 40,000 cpm [ '251]-labeled IGFBP-3 tracer, iodinated by a modification of the chloramine-T meth- Od,(23) was added to each tube in 0.1 ml RIA buffer and the tubes incubated overnight at 5°C. Then, 0.5 ml secondary antiserum (goat antirabbit serum diluted 20-fold and normal rabbit serum diluted 100-fold in RIA buffer) and 0.2 ml of 8% polyethylene glycol (PEG 8000) were added and the tubes incubated for 4 h at room temperature. Last, the tubes were centrifuged at 1300 X g for 30 minutes, the supernatants decanted and discarded, and the radioactive precipitates quantitated. The detectable range (effec- tive dose) of this IGFBP-3 RIA was usually between 7.5 and 160 nglml (ED,, and ED,, respectively), and there was no signifi- cant cross-reactivity with other known IGFBPs at concentrations of up to 500 ng/ml.
Cell proliferation assay
Cells were plated (10,OOO per well) in serum-free DMEM into 48-well culture plates and incubated for 24 h. Then media were exchanged to fresh serum-free DMEM and incubated for 18 h.
DIFFERENTIAL REGULATION OF IGFBP-3
100
90
80
7 0
60
effector. IGFBP-3 values were standardized as ng/dish or ng/mg cellular protein and are given as mean ? standard deviation (SD). Comparison of group means was made by analysis of variance (ANOVA); comparison of control and each treatment group was made using Student's t-test. A dose-dependent change in IGFBP-3 was defined as a significant correlation (p < 0.05) between IGFBP-3 secretion (nglmg protein) and the logarithm of the effector dose as determined by regression
2 50
bp 40
analysis (CSS; Stat Soft, Inc., Tulsa, OK). 30
20
10 RESULTS
TABLE I . BASAL IGFBP-3 SECRETION, CELL PROLIFERATION, AND ALP ACTIVITY IN VARIOUS HUMAN OSTEOSARCOMA CELL LINES"
IGFBP-3 Cell prolgerution ALP (mUlmg protein) Cell lines (nglmg protein) (70 high-ALP SaOS-2) (% high-ALP SuOS-2)
-
-
-
-
-
-
-
-
~~ ~
High-ALP SaOS-2 73 f 31 100 ? 23.7 100 * 16.6 MG-63 146 ? 43 450 * 218 0.0295 2 0.0035 Low-ALP SaOS-2 215 f 70 237 * 80.6 TE89 776 f 99 69.2 2 37.6 0.0545 * 0.0078
0.337 5 0.035
Basal amounts of IGFBP-3 secreted by various human osteosarcoma cell lines were deter- mined by performing several cxperiments (MG-63, five experiments; low-ALP SaOS-2, six experiments; high-ALP SaOS-2, three experiments; and TE89, four experiments). Conditioned media collected from 24 h serum-free cultures of MG-63 and high-ALP SaOS-2 cells were concentrated 2.5- and %-fold, respectively, by SpeedVac centrifugation before IGFBP-3 RIA. For low-ALP SaOS-2 and TE89 cells, 100 pI straight conditioned medium was used for RIA. Cell proliferation and ALP activity were measured as described in Materials and Methods and are expressed as percentage of high-ALP SaOS-2 cells (mean 2 SD; n = 3-5 experiments). The net values of incorporated [ 'Hlthymidine and specific activity of ALP in high-ALP SaOS-2 cells were 763 2 181 cpm and 1820 f 300 mU/mg protein (mean * SD).
Basal selwtion of IGFBP-3, cellular ALP activity, and cell proliferution
Before determination of the effects of the systemic regulators on IGFBP-3 production in human bone cells, we first measured the basal level of IGFBP-3 secretion in four osteoblast-like human osteosarcoma cell lines: MG-63, TE89, and low- and high-ALP subpopulations of SaOS-2 cells. The IGFBP-3 con- tent of the 24 h conditioned medium from these cell lines differed by a.. much as 10-fold (Table I). TE89 cells secreted the most IGFBP-3 (776 * 99 ng/mg protein), followed by the rapidly proliferating low-ALP SaOS-2 cells (215 * 70 ng/mg protein) and MG-63 cells (146 2 43 nglmg protein). The slowly dividing high-ALP SaOS-2 cells secreted the least IGFBP-3
O k '1'0 20 40 60 90100 1 IGFBP-3 (ns/ml)
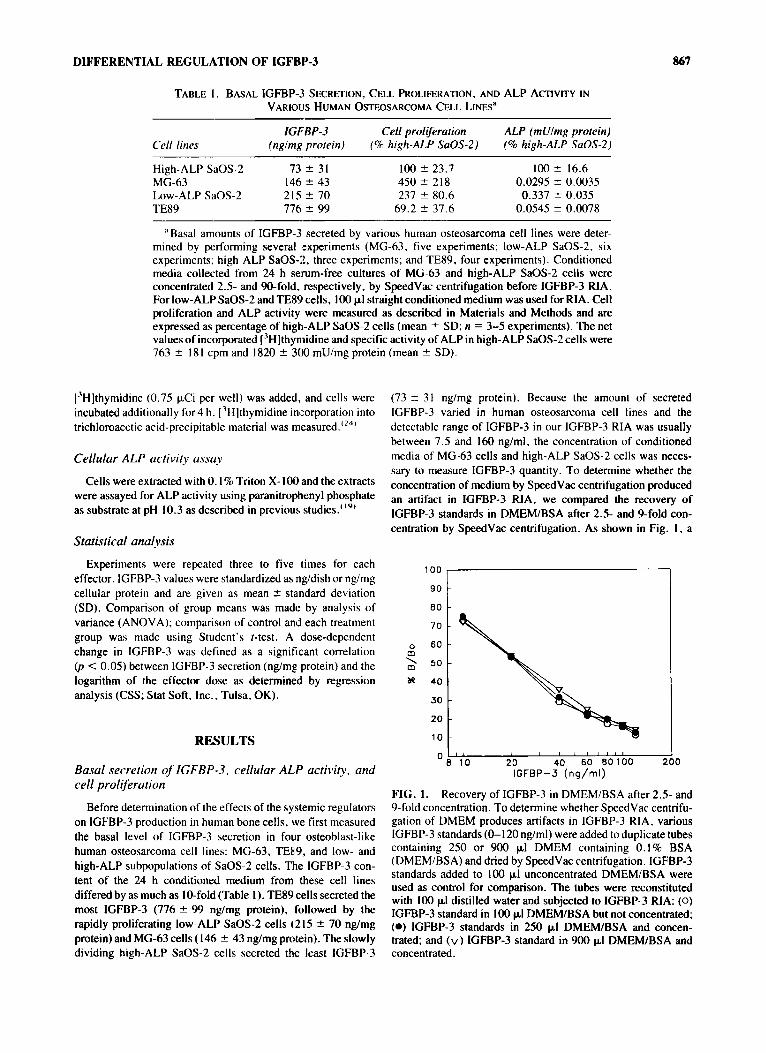
0
FIG. 1. Recovery of IGFBP-3 in DMEM/BSA after 2.5- and 9-fold concentration. To determine whether SpeedVac centrifu- gation of DMEM produces artifacts in IGFBP-3 RIA, various IGFBP-3 standards (0-120 nglml) were added to duplicate tubes containing 250 or 900 p1 DMEM containing 0.1% BSA (DMEM/BSA) and dried by SpeedVac centrifugation. IGFBP-3 standards added to 100 pl unconcentrated DMEM/BSA were used as control for comparison. The tubes were reconstituted with 100 pl distilled water and subjected to IGFBP-3 RIA: (0) IGFBP-3 standard in 100 pI DMEMlBSA but not concentrated; (0) IGFBP-3 standards in 250 pI DMEMlBSA and concen- trated; and (v) IGFBP-3 standard in 900 pI DMEMlBSA and concentrated.

868 NAKAO ET AL.
2.5- or %fold concentration did not affect the recovery of IGFBP-3, thus suggesting that substances in concentrated DMEM did not interfere with the IGFBP-3 RIA.
Cell proliferation and ALP activity were also measured in these cell lines (Table 1) in an attempt to understand better the significance of different basal levels of IGFBP-3 secretion in human osteosarcoma cell lines. The basal level of IGFBP-3 secretion appeared to be directly related to cell proliferation and inversely related to ALP activity in subpopulations of the same cell line (SaOS-2). However, these relationships did not hold when the comparison was expanded to include TE89 and MG-63 cell lines (Table 1).
Effect of dexamethasone on secretion of IGFBP-3
Treatment of cells with dexamethasone for 24 h inhibited IGFBP-3 secretion in three of the four human osteosarcoma cell lines (Fig. 2). Dexamethasone ( 10-12-10-7 M) inhibited the secretion of IGFBP-3 in MG-63. low-ALP SaOS-2, and TE89 cell lines in a dosedependent m n e r (correlation and regres- sion: r = -0.60, p < 0.001; r = -0.65, p < 0.005; and r = -0.89, p < 0.001, respectively) but had no effect on IGFBP-3 secretion by high-ALP SaOS-2 cells. At M dexamethasone, IGFBP-3 secretion decreased maximally by 37% in MG-63 cells, 4 1 % in low-ALP SaOS-2 cells, and 46% in
140 I
\ F 20
0 0 ;1 -12 -11
A) MG63 I I 1 I I
-10 -9 -8 -7 Dexamethasone Conc. (log[M])
e 120 T I 1 I C) High ALP SoOS-2
- 2 100
*: 80 g .;
c
h g 6o p 40 \ p 20 v
0 0 -12 - 1 1 -10 -9 -8 -7
Dexamethosone Conc. (log[M])
TE89 cell lines. No changes in cellular protein content were observed following 24 h dexamethasone treatment, but a slight decrease was observed after 48 h treatment (data not shown).
To determine whether this was truly a regulatory effect, not an artifact (e.g., toxicity), a time course of the effect of dexametha- sone treatment on IGFBP-3 secretion was canied out in one of the responsive cell lines, low-ALP SaOS-2 (Fig. 3). In this experiment, low-ALP SaOS-2 cells were incubated without or with M dexamethasone and the conditioned medium collected after0.5,4,8,24, and48 h. IGFBP-3 levels measured by RIA increased with time in the conditioned medium of both control and dexamethasone-treated cultures. In comparison with the control, however, there was a significant reduction in the level of IGFBP-3 in M dexamethasone-treated cultures after 24 h.
Effect of 1,25-(OH),D, on IGFBP-3 secretion
Treatment of cells with 1,25-(OH),D, for 48 h stimulated IGFBP-3 secretion in all three human osteosarcoma cell lines tested (Fig. 4). 1,25-(OH)2D, (lO-'l-lO-s M) increased IGFBP-3 secretion in a dose-dependent manner in MG-63, low-ALP SaOS-2, and high-ALP SaOS-2 cells (correlation and regression: r = 0.56, p < 0.05; r = 0.75. p < 0.005; and r = 0.85, p < 0.001, respectively). 1,25-(OH),D, increased
160
140
120
100
80
60
40
20
0
B) LOW ALP SOOS-2
140
2 120 5
C - 8 100
7 : 80 2 3 " g
c
a c 60
p 4 0
\ p 20 v
0
0 -12 -11 -10 -9 -8 -7 Oexamethosone Conc. (log[M])
I 0) TE89
Dexomethosone Conc. (log[M])
FIG. 2. Effect of dexamethasone on IGFBP-3 secretion by various human osteosarcoma cell lines. Conditioned medium was collected fromcultures treated with 10-12-10-7 M dexamethasone for 24 h. Conditioned media from MG-63 and high-ALP SaOS-2 cells were concentrated 2.5- and 9-fold using SpeedVac centrifugation before the IGFBP-3 RIA. The amount of IGFBP-3 secreted was standardized on the basis of cellular protein content and expressed as percentage of vehicle-treated control. Values are mean 2 SD (n = 4). Results shown here are from arepresentative experiment (n = 3-5). The difference between group means was significant (by ANOVA) for low-ALP SaOS-2 @ < 0.05) and TE89 cells @ < 0.001) but did not quite attain significance for MG-63 cells. Furthermore, a dosedependent inhibition of IGFBP-3 production by dexamethasone was demonstrated by correlation and regression (MG-63, r = -0.60, p < 0.01; low-ALP SaOS-2, r = -0.65, p < 0.005; TE89, r = -0.89, p <: 0.001). Statistical difference from the control level (Student's t-test): *p < 0.05; **p < 0.01; and ***p < 0.001.

DIFFERENTIAL REGULATION OF IGFBP-3 869
A) MG63 . 1 1
- -- 2 150 .- 5 - 100 r) I a m
& 50
- a
" 0.5 4 8 24 48
Incubation Time ( hours )
.. C) High ALP SaOS-2
FIG. 3. Time course of dexamethasone effect on IGFBP-3 secretion in low-ALP SaOS-2 cells. Low-ALP SaOS-2 cells were incubated without or with lo-' M dexamethasone. After 0.5,4, 8, 24. and 48 h, conditioned medium wits collected and subjected to IGFBP-3 RIA without prior SpeedVac concentra- tion. Values are expressed as mean 5 SD ( n == 4). Statistical difference from the control level (Student's r-test): **p < 0.01; ***p < 0.001 *
IGFBP-3 secretion maximally by 144% in MG-63 cells (lo-' M), 164% in low-ALP SaOS-2 cells (lo-' M). and 210% in high-ALP SaOS-2 cells ( lo-* M).
Synergistic efect of 1,25-(OH),D, and TGF-#I on IGFBP-3 secretion und ALP activity
Recently, two separate groups reported that I .25-(OH),D, and TGF-P increase synergistically ALP expression in normal human bone cells or MG-63 celIs.'2s.2h' We therefore sought to determine whether TGF-P modulates the I .25-(OH),D,-in- duced IGFBP-3 secretion in MG-63 cells (Tattle 2). Although I ,25-(OH),D, ( M) and TGF-f3 ( I nghnl) individually increased IGFBP-3 secretion up to 41 and 247O/c, respectively, they did not increase cellular ALP activity significantly. In contrast, the combination of 1,25-(OH),D, and TGF-P increased synergistically both IGFBP-3 secretion and ALP activity.
Effect of hPTH-(1-34) on IGFBP-3 seertition
Treatment of cells with hPTH-( 1-34) (0.1-100 ng/ml) for 48 h had no significant effect on the secretion of IGFBP-3 in MG-63,low-ALP SaOS-2, or high-ALP SaOS-2 cells (Table 3). TE89 cells were not evaluated in this experiment because previous ligand blot studies failed to indicate the FTH effect on IGFBP-3 secretion in this cell line (unpublished data).
DISCUSSION
This study demonstrates that the basal late of IGFBP-3 secretion varies as much as 10-fold depending on the cell type studied. Hassager et al.'27' also reported that the basal secretion of various IGFBPs is cell type dependent. The significance of this finding is complicated by reports that in a variety of cell types IGFBP-3 can either enhance or inhibit the metabolic or mitogenic effects of the IGFs, depending on the study condi-
180
160
140
- c
'ij 120
b 5- L'u 80 P g
60
4 0 a 5 20
n
7 iR 100
0 - 1 1 -10 -9 -8 1,25(OH)2 D3 Conc. (log[M])
200 c e c s 150 c 0 n
a - I X m & 100
a
E 50
$: OI
> v
0 0 - 1 1 -10 -9 -8
1 , 2 ~ 5 ( 0 H ) ~ D ~ Conc. (log[M])
T I
0 - 1 1 -10 -9 -8 1,25(0ti)2D3 Conc. (log[M])
FIG. 4. Effect of 1,25-(OH),D, on IGFBP-3 secretion by various human osteosarcoma cell lines. Cells were incubated with various concentrations of I ,25-(OH),D, ( 10~'2-10-" M) for 48 h. Conditioned medium was collected and subjected to IGFBP-3 RIA. The IGFBP-3 concentrations were standardized against protein content, and the values are expressed as percent- age of alcohol-treated control, mean 2 SD (n = 4). The differ- ence between group means was significant (by ANOVA) for all cell lines tested (MG-63, p < 0.05; low-ALP SaOS-2, p < 0.001; high-ALP SaOS-2, p < 0.001). Furthermore, a dose-dependent inhibition demonstrated by correlation and re- gression of IGFBP-3 production by 1,25-(OH),D, was observed (MG-63, r = 0.56; p < 0.05; low-ALP SaOS2; r = 0.75, p < 0.005; high-ALP SaOS-2, r = 0.85, p < 0.001). Statisti- cal difference from control level (by Student's r-test): *p < 0.05; **p < 0.01; ***p < 0.001.
tions .(6,28' Potential explanations for the differential endogenous IGFBP-3 secretion in different osteosarcoma cell lines can only be speculated upon at this time. It is possible that the differences in the IGFBP-3 secretion can be attributed to the different stages of osteoblast differentiation these osteosarcoma cell lines may

870 NAKAO ET AL.
TABLE 2. EFFEcr OF 1,25-(OH)@, AND TGF-p ON IGFBP-3 SECRETION AND ALP ACTIVITY IN MG-63 CELLS'
IGFBP-3 secretion ALP activity Treatment (nglmg protein) (mUlmg protein)
Control 119 2 26 1.07 2 0.37 1,25-(oH)@, 168 * 29b 1.15 2 0.11 TGF-@ 413 2 40' 1.45 2 0.37 1,25-(OH)& + TGF-P 1010 2 30d 4.88 2 0.57'
'MG-63 cells (5.6 X l@ cellsl60 mm dish) were cultured in DMEM supplemented with 2% calf serum for 24 h, followed by incubation in DMEM containing 0.1% BSA (DMEWBSA) for 24 h. Cells were then incubated with DMEMlBSA containing lo-' M 1 ,25-(OH),D3 and/or 1 nglml of TGF-p or appropriate vehicle for 48 h. The values are expressed as mean 2 SD (n = 4).
bVersus control, p < 0.05 (by Student's t-test). 'Versus contro1.p < 0.001; versus 1,25-(OH),D,.p < 0.001 (by Student's t-test). dVersus control, p < 0.001; versus 1,25-(OH),D,, p < 0.001; versus TGF-P, p < 0.001 (by
eVersus control, p < 0.001; versus 1,25-(OH),D,, p < 0.001; versus TGF-P, p < 0.001 (by Student's r-test).
Student's r-test).
represent. As proposed by Farley et al.,"'' more mature osteo- blasts are characterized by higher ALP activity and lower cell proliferation, whereas less mature osteoblasts are characterized by lower ALP activity and higher cell proliferation. Indeed, high ALP activity appeared to correlate with low cell proliferation in three of the four cell types evaluated. Only TE89 cells were low in both ALP activity and cell proliferation. Furthermore, no correlation exists between the level of IGFBP-3 secretion and either ALP activity or cell proliferation across all four cell types. This finding suggests that IGFBP-3 secretion in different human OsteOsarcoma cell lines may be influenced by factors other than stage of differentiation. Moreover, recent studies showed that the cellular response to a given hormonal signal depends not only upon the stage of cell differentiation but also upon the stage of the cell cycle.'29' Another potential explanation for the differential IGFBP-3 secretion among these various human osteosarcoma cell lines is that the autocrine and paracrine repertoire produced by various osteosarcoma cell lines may be responsible for the differential IGFBP-3 secretion. Recent studies supporting this explanation showed that IGFBP-3 secretion is regulated by IGF-I and IGF-11, the major growth factors produced by bone cells.'3o' Last, it is also possible that IGFBP-3 gene may be expressed to varying levels in different
human osteosarcoma cell lines as a result of the transforma- tion process itself. Additional studies are needed to sort out which of these mechanism(s) may be responsible for the differ- ence in basal levels of IGFBP-3 secretion and the differential regulation of IGFBP-3 secretion in various human osteosarcoma cell lines.
This study further supports the conclusion of other published reports that IGFBP-3 secretion is regulated in a complex man- ner. For example, dexamethasone inhibited IGFBP-3 secretion in low-ALP SaOS-2, MG-63, and TE89 cells but not in high- ALP SaOS-2 cells. Dexamethasone promotes a more highly differentiated phenotype in untransformed normal human osteo- blast-like cells.'3') Thus, the differential response of IGFBP-3 secretion to dexamethasone treatment in low-ALP SaOS-2 and high-ALP SaOS-2 cell subpopulations perhaps may be ex- plained by the fact that high-ALP SaOS-2 cells already possess a more highly differentiated osteoblast phenotype."') It does not explain, however, the relative sensitivities of the responsive cell types to the glucocorticoid. The apparent trend is that the responsiveness to dexamethasone increases as the basal level of IGFBP-3 secretion increases across all cell lines. Determining whether this is truly a significant relationship requires further study. 1 ,25-(OH),D, increases differentiation and decreases
TABLE 3. EFFECT OF hPTH-( 1-34) ON IGFBP-3 SECRETION IN HUMAN OSTEOSARCOMA CELL LINE^
IGFBP-3 (nglmg protein, % of control) hPTH-( 1-34) ( n g W MG-63 Low-ALP SaOS-2 High-ALP SaOS-2
0 100.0 2 7.2 100.0 2 11.4 100.0 ? 8.1 0.1 97.6 2 4.8 96.6 f 3.0 89.7 f 6.6 1 .o 98.1 2 4.8 101.9 f 9.5 96.6 * 2.5 10.0 101.0 2 8.7 112.1 2 5.7 107.0 2 3.7 100 104.3 2 12.6 100.0 2 12.5 96.3 % 22.0
'Cells were incubated with various concentrations of hPTH-( 1-34) for 48 h. Conditioned media were collected and subjected to IGFBP-3 RIA. The IGFBP-3 contents were standardized by protein content, and the values are expressed as mean 2 SD (n = 4), percentage of control.

DIFFERENTIAL REGULATION OF IGFBP-3 87 1
cell proliferation." i.25.26' I ,25-(OH),D, stimulated IGFBP-3 secretion in all cell types studied. Furthermore. the degree of stimulation appeared to be directly related to level of basal ALP activity And inversely related to the relative cell proliferation. Thus, the more mature osteoblast cell types appear to be more responsive to I ,25-(OH),D,. A 1,25-(OH),D, stimulation of IGFBP-3 secretion in MG-63 cells was also previously reported by Moriwake and colleagues.""' Furthermore, this study dem- onstrated that 1,25-(OH),D, combined with TGF-P increased IGFBP-3 secretion in a synergistic manner. This finding is consistent with previous findings that I ,25-(OH)2D, combined with TGF-P produced synergistic effects on ALP activity.'2s.2"' At this time, it is not known whether this synergism occurs through a similar mechanism for both ALP activity and IGFBP-3 secretion.
Moreover, this study now provides evidence that the secretion of IGFBP-3 and IGFBP-4 is regulated independently and not necessarily in the same manner across specie!;. This study demonstrated that PTH had no significant effecl. on IGFBP-3 secretion in any cell type tested. Similarly, Hassager et al.'27' recently reported that PTH did not influence IGF13P-3 secretion in normal human bone cells or in various human osteosarcoma cell lines. However, the reason for the null effcct of PTH on IGFBP-3 secretion may not be a lack of PTH reczptors in these cells because we found that PTH stimulates cell proliferation in both low- and high-ALP SaOS-2 cell lines (unpublished obser- vations). In contrast, PTH and dibutyryl cyclic AMP have been shown to stimulate inhibitory IGFBP-4 production significantly in normal human bone cells as well as in various human osteosarcoma cell lines."H.2'.32' In addition, IGFs have been shown to increase and decrease IGFBP-3 and IGFBP-4 secre- tion, respectively. in various human bone cell types.',"' These findings suggest that the secretion of various IGFBPs is regu- lated independently of one another in human bone cells. More- over, the regulation of IGFBP-3 secretion in human bone cells by PTH appears to be different from the previous findings reported for rat osteoblast-like cells. In rat osteoblasts, PTH has been shown to increase the production of IGF-I a.nd IGFBP-3; in human bone cells, PTH had no significant effect on the produc- tion of IGF-I, IGF-II, or IGFBP-3.'"~".34' Becwse the relative distribution of IGF-I and IGF-I1 has been shown to be different between rats and humans,"' it is possible that the responsiveness of human and rat bone cells to certain systemic effectors may vary.
In conclusion, the findings of this study demonstrate that ( I ) basal IGFBP-3 secretion is cell type dependent, and (2) IGFBP-3 secretion is regulated by systemic effectors in both positive and negative manner. Based on these and other find- ings, we speculate that the response of osteoblasts to a given effector may be regulated in a versatile manner. through IGFBP modulation of IGF-induced cellular response (luring osteoblast maturation.
ACKNOWLEDGMENTS
The authors acknowledge Jerry L. Pettis Veterans' Medical Media for photography. This work was supported by funds from the National Institutes of Health (AR 3 1062) and the Department of Medicine, Loma Linda University. This study was presented
in abstract form at the American Society for Bone and Mineral Research meeting in Minneapolis, 1992.
REFERENCES
I . Mohan S. Baylink DJ 1991 Bone growth factors. Clin Orthop 263:30-48.
2. Mohan S. Baylink DJ 1991 The role of insulin-like growth factor-11 in the coupling of bone formation to resorption. In: Spencer EM (ed.) Modem Concepts of Insulin-like Growth Factors. Elsevier, New York. pp. 169-184.
3. Raisz LG 1988 Local and systemic factors in the pathogenesis of osteoporosis. N Engl J Med 318:818-828.
4. Shimasaki S, Ling N 1991 Identification and molecular character- ization of insulin-like growth factor binding proteins (IGFBP- I ,-2. -3,-4,-5 and -6). Prog Growth Factor Res 3:243-266.
5. Mohan S. Baylink DJ 1991 Isolation and characterization of insulin-like growth factor binding proteins produced by human bone cells in vitro. J Bone Miner Res 5:S141.
6. Mohan S 1993 Insulin-like growth factors binding proteins in bone cell regulation. Growth Regul3:67-70.
7. Mohan S, BautistaCM, Wergedal JE, Baylink DJ 1989 Isolation of an inhibitory insulin-like growth factor (IGF) binding protein from bone cell-conditioned medium: A potential local regulator of IGF action. Proc Natl Acad Sci USA 86:833X-8341.
8. Bautista CM. Baylink DJ, Mohan S 1991 Isolation of novel insulin-like growth factor (IGF) binding protein from bone: A potential candidatea for fixing IGF-11 in human bone. Biochem Biophys Res Commun 176:756-763.
9. Canalis E, Centrella M, Burch W, McCarthy TL 1989 Insulin-like growth factor I mediates selective anabolic effects of parathyroid hormone in bone cultures. J Clin Invest 1)4:6045.
10. Linkhart TA. Mohan S 1989 Parathyroid hormone stimulates release of insulin-like growth factor-] (IGF-I) and IGF-II from neonatal mouse calvaria in organ culture. Endocrinology 125: 14x4- I49 I .
I I . Scharla SH, Strong DD, Mohan S, Baylink DJ. Linkhart TA 199 I I .25-Dihydroxy D, differentially regulates the production of insu- lin-like growth factor I (IGF-I) and IGF-binding protein-4 in mouse osteoblasts. Endocrinology 129:3 139-3 146.
2 . Kream BE, Peterhen DN, Raisz LG 1990 Cortisol enhances the anabolic effects of insulin-like growth factor 1 on collagen synthesis and procollagen messenger ribonucleic acid levels in cultured 21-day fetal rat calvariae. Endocrinology 126:1576-1583.
3. McCarthy TL, Centrella M, Canalis E 1990 Cortisol inhibits the synthesis of insulin-like growth factor-I in skeletal cells. Endtrri- nology 126: 1569-1575.
14. Schmid C, Ernst M, Zapf J . Froesch ER 1989 Release of insulin- like growth factor carrier proteins by osteoblasts: Stimulation by estradiol and growth hormone. Biochem Biophys Res Commun 160: 788-794.
15. Schmid C. Zapf I. Froesch ER 1989 Production of carrier proteins for insulin-like growth factors (IGFs) by rat osteoblastic cells: Regulation by IGF I and cortisol. FEBS Lett 244:328-332.
16. Moriwake T. Tanaka H, Kansaki S, Higuchi J . Seino Y I992 1.25-Dihydroxyvitamin D, stimulates the secretion of insulin-like growth factor binding protein 3 (IGFBP-3) by cultured human osteosarcoma cells. Endocrinology 130: 107 1-1073.
17. Linkhart TA. Scharla SH, Kyeyune-Nyombi E. Mohan S, Baylink DJ, Strong DD 1992 I .25-(OH)?D3 increases insulin-like growth factor binding protein4 (IGFBP-4) secretion, mRNA levels, tran- scription and mRNA stability in human osteoblast-like cells. J Bone Miner Res 7(Suppl. I):S158.
18. LaTour D. Mohan S, Linkhart TA. Baylink DJ, Strong DD 1990

872 NAKAO ET AL.
Inhibitory insulin-like growth factor binding protein: Cloning, complete sequence, and physiological regulation. Mol Endocrinol
19. Farley JR, Hall SL, Herring S, Tarbaux NM, Matsuyama T, Wergedal JE 1991 Skeletal alkaline phosphatase specific activity is index of osteoblastic phenotype in subpopulations of the human osteosarcoma cell line SaOS-2. Metabolism 40(7):664471.
20. Ernst M, Schmid C, Froesch ER 1989 Phenol red mimics biological actions of estradiol: Enhancement of osteoblast proliferation in vitro and of type I collagen gene expression in bone and uterus of rats in vitro. J Steroid Biochem 33907-914.
21. Mohan S, Baylink DJ 1991 Evidence that the inhibition of TE85 human bone cell proliferation by agents which stimulate cAMP production may in part be mediated by changes in the IGF-II regulatory system. Growth Reg l:IlO-l18.
22. Lowry OH, Rosenbrough NJ, Farr AL, Randall RJ 1951 Protein measurement with the Fohn phenol reagent. J Biol Chem 193:265 269.
23. Mohan S, Jennings JC, Linkhart TA, Baylink DJ 1988 Primary
4 1806-1 8 14.
24
25
26
27
structure of human insulin-like growth factor: Homology with human insulin-like growth factor-11. Biochim Biophys Acta 966538-547. Wergedal JE, Mohan S, Lundy M, Baylink DJ 1990 Skeletal growth factor and other growth factors known to be present in bone matrix stimulate proliferation and protein synthesis in human bone cells. J Bone Miner Res 5179-186. Wergedal JE, Matsuyama T, Strong DD 1992 Differentiation of normal human bone cells by transforming growth factor-beta and I ,25-(OH), vitamin D,. Metabolism 41:42-48a. Bonewald LF, Kester MB, Schwartz 2, Swain LD, Khare A, Johnson T, Leach RJ, Boyan BD 1991 Effect of combining transforming growth factor beta and 1,25-dihydroxyvitamin D, on differentiation of a human osteosarcoma (MG-63). J Biol Chem 267:8943-8949. Hassager C, Fitzpatrick LA, Spencer EM, Riggs BL, Conover CA 1992 Basal and regulated secretion of insulin-like growth factor binding proteins in osteoblast-like cells is cell line specific. J Clin Endocrinol Metab 7 3 1 k228-233.
potentiates IGF-I stimulated DNA synthesis in human skin fibro- blasts. Biochem Biophys Res Commun 156:199-204.
29. Chakrdborty M, Chatterjee D, Kelloukumpu S, Rasmussen H, Baron R 1991 Cell cycle-dependent coupling of the calcitonin receptor to different G proteins. Science 251: 1078-1082.
30. Mohan S, Strong DD, Lempert UG, Tremollieres F, Wergedal JE, Baylink DJ 1992 Studies on regulation of insulin-like growth factor binding protein (IGFBP)-3 and IGFBP-4 production in human bone cells. Acta Endocrinol (Copenh) 122555464.
31. Wong M-M, Rao LG, Ly H, Hamilton L, Tong J, Sturtridge W, McBroom R, Aubin JE, Murray TM 1990 Long-term effects of physiological concentrations of dexamethasone on human bone- derived cells. J Bone Miner Res 5(8):803-813.
32. Mohan S. Strong DD, Hilliker S, Malpe R, Lee K, Farley J, Baylink DJ 1993 Dibutyryl cyclic adenosine monophosphate differ- entially regulates cell proliferation in low and high alkaline phos- phatase SaOS-2 human osteosarcoma cells: Evidence for mediation by the insulin-like growth factor4 system. J Cell Physiol 156:462- 468.
33. Schmid C, Schalapfer I, Waldvogel M, Peter M, Zapf J, Froesch ER 1993 Production of insulin-like growth factor binding protein-3 by connective tissue and endothelial cells. Growth Regul 3( 1):65- 67.
34. Finkelman RD, Mohan S, Linkhart TA, Abraham SM, Boussy JP, Baylink DJ 1992 PTH stimulates the proliferation of TE-85 human osteosarcoma cells by a mechanism not involving either increased cAMP or increased secretion of IGF-I, IGF-I1 or TGF beta. Bone Miner 1689-100.
Address reprint requests to: Subburaman Mohan
Research Service (151) Pettis V . A . Hospital I1201 Benton Street
Loma Linda. CA 92357 . .
28. De Mellow JSM, Baxter RC 1988 Growth hormone-dependent insulin-like growth factor (IGF) binding protein both inhibits and
Received in original form April 16, 1993; in revised form September 21, 1993; accepted October 14, 1993.