suillus quiescens, a new species commonly found in the ... · of california, berkeley, california...
TRANSCRIPT

Suillus quiescens, a new species commonly foundin the spore bank in California and Oregon
Thomas D. Bruns1
Department of Plant and Microbial Biology, Universityof California, Berkeley, California 94720-3102
Lisa C. GrubishaUSDA-ARS, Department of Plant Sciences, University ofArizona, P.O. Box 210036, Tucson, Arizona 85721
James M. TrappeDepartment of Forest Science, Oregon State University,Corvallis, Oregon 97331-7501
Jennifer F. KerekesElse C. Vellinga
Department of Plant and Microbial Biology, Universityof California, Berkeley, California 94720-3120
Abstract: Suillus quiescens sp. nov. is common underPinus muricata on Santa Cruz and Santa Rosa Islandsin the northern Channel Islands of California, and wesubsequently found it fruiting at Point Reyes NationalSeashore on the central coast of California. Sequenc-es from the internal transcribed spacer region showthat it is distinct from all 44 species of Suillus tested,and features of its morphology separate it from allother unsequenced species. Suillus quiescens has abroader distribution than coastal California because italso was encountered as ectomycorrhizae on roots ofpine seedlings from the eastern Sierra Nevada, coastalOregon and the southern Cascade Mountains. Thereason it had not been identified from these areasmight be due to its resemblance to S. brevipes atmaturity or it might be a rare fruiter that persists inthe spore bank.
Key words: bioassay, suilloid-specific primers,Suillus brevipes, S. occidentalis, S. volcanalis
INTRODUCTION
We describe a new species, Suillus quiescens, that weencountered in 2002 while collecting on Santa CruzIsland where it and S. pungens Thiers & Smith werethe only two Suillus species that were abundantlyfruiting in association with native Pinus muricata(Grubisha et al. 2005).
We determined the nucleotide sequence from theinternal transcribed spacer region (hereafter ITS) of
several specimens of Suillus quiescens to compare it toother similar Suillus species. We turned to the ITSregion because it had been sequenced from a broadsample of genus Suillus (Kretzer and Bruns 1997,Kretzer et al. 1996, Manian et al. 2001, Wu et al.2000). In addition we acquired ITS sequence fromcollections of S. volcanalis Thiers and S. occidentalisThiers, two species that are morphologically similar toS. quiescens but previously were unsequenced.
MATERIALS AND METHODS
Standard methods for describing the basidiocarps wereapplied with the terminology of Smith and Thiers (1971).Color annotations in the macroscopic descriptions arefrom Kelly and Judd (1976). The notation [72, 7, 7]indicates that measurements were made on 72 spores inseven samples in seven collections. These abbreviationsare used: avl for average length, avw for average width, Qfor quotient of length and width and avQ for averagequotient.
Amplification and sequencing.—DNA was extracted with amodified protocol of the REDExtract-N-AmpTM Plant PCRKit (Sigma-Aldrich Co., St Louis, Missouri) as follows: 10 mLextraction solution, incubated at 65 C for 10 min followedby 95 C for 30 min, 30 mL dilution solution was added andleft at room temperature 3 h. The ITS region was generallyamplified with the ITS-1f/ITS-4b primer set with anEppendorf Mastercycler Gradient thermocycler underconditions described by Gardes and Bruns (1993). In thecase of older specimens or slightly moldy specimens, such asholotypes of S. volcanalis and S. occidentalis, we anticipatedproblems with degraded target DNA or DNA from molds.To circumvent these problems we designed Suilloid specificprimers ITS-2S 59-AAGATTCGATGATTCACTGTAG-39 andITS-3S 59GTAAATTCTCAACCCCTCTCGA-3 that are re-spectively 31 and 108 bp upstream from the ITS 2/ITS3primer site. We designed these new primers by eye from analignment with nine Suillus species and seven moredistantly related basidiomycetes (Tapinella, Sebacina, Thele-phora, Cortinarius, Tricholoma, Coltricia and Chalciporus).These primers were designed to be perfect matches with allSuillus, while mismatching nonsuilloid taxa at the 39 end.BLAST analyses revealed that the sequences selected wereperfect matches only to Suillus and the related genera,Rhizopogon, Truncocolumella, Chroogomphus and Gomphi-dius. These two primers were used to amplify the ITS1 andITS2 regions (White et al. 1990) with primer sets ITS-1f/ITS-2S and ITS-3S/ITS-4b under conditions described byGardes and Bruns (1993). PCR products were cleaned with0.5 mL ExoSAP IT (USB Corp., Cleveland, Ohio) and cycledat 37 C for 45 min, followed by 80 C for 15 min. Sequenc-
Submitted 23 Jun 2009; accepted for publication 9 Sep2009.1 Corresponding author. E-mail: [email protected]
Mycologia, 102(2), 2010, pp. 438–446. DOI: 10.3852/09-149# 2010 by The Mycological Society of America, Lawrence, KS 66044-8897
438

ing was performed with an ABI PRISM 3100 GeneticAnalyzer (Applied Biosystems, Foster City, California).Sequences were edited and contigs assembled with Se-quencher 4.2.2 (Gene Codes Corp., Ann Arbor, Michigan).
Phylogenetic analyses.—Suillus ITS sequences were retrievedfrom GenBank with the Emerencia interface Website(http://andromeda.botany.gu.se/genussearch.html). Se-quences that did not contain both ITS1 and ITS 2 spacerswere removed, and when large numbers of nearly identicalspecies were encountered a subset of these were selected.Preference was given to published sequences and those inthe UNITE database (Koljalg et al. 2005).
These identified Suillus sequences from GenBank wereused in the analysis: S. amaranthii: U74615.1; S. americanus:L54103.1; S. asiaticus: AF166504, L54090.1; S. bellinii:AJ419216.1; S. bovinus: AJ272401.1, AJ272402.1, AJ272403.1,AJ419215.1, AJ419934.1, AJ419935.1, AJ493679.1, L54077.1;S. bresadolae: L54084.1; S. brevipes: AY880941.1, L54111.1;U74620.1; S. cavipes: L54085.1, L54105.1, L54119.1; S.collinitus: AJ410857.1, L54089.1; S. cothurnatus: AJ419217.1,AJ419218.1, L54092.1; S. decipiens: AF166508, AF166510,L54079.1; S. flavidus: AY641461.1, UDB001214, UDB001648,UDB001649; S. fluryi: AJ272407.1; S. glandulosipes: L54087.1;S. granulatus: AB284447.1, AJ272408.1, AJ272409.1,AJ272410.1, L54076.1, L54113.1, L54121.1, UDB000650,UDB000666; S. grevillei: M91612.1, M91614.1, M91616.1;S. intermedius: L54074.1; S. lakei L54086.1; S. laricinus:AJ272400.1, L54099.1, L54102.1; S. c. f. laricinus L54120.1; S.luteus AF166511, AJ272411.1, AJ272413.1, AJ272414.1,AJ272415.1, AJ272416.1, AJ419219.1, L54083.1, L54100.1,L54110.1, UDB001650; S. mediterraneensis: AJ410860.1; S.neoalbidipes: AY880940.1, L54112.1; S. ochraceoroseus:L54093.1; S. paluster: L54080.1; S. placidus: AB284441.1,AB284443.1, DQ407265.1, L54118.1, L54108.1; S. cf. placi-dus: L54118.1; S. plorans: AJ272417.1; S. pseudobrevipes:AY880938.1, L54107.1; S. punctipes: L54098.1; S. pungensL54095.1; S. serotinus: L54116.1; S. spraguei: AF166519,AF166520, AF166522, AF166525, AY854069.1, M91617.1; S.spectabilis: L54104.1; S. sibiricus: AF166512, AF166513,L54117.1; S. sinuspaulianus: L54078.1; S. subaureus:L54109.1; S. subluteus: L54075.1, L54088.1; S. suilloides:U74616.1, U74617.1, U74618.1; S. tomentosus: AY880937.1,L54106.1, U74614.1; S. tridentinus: AJ419220.1, L54082.1; S.umbonatus: AF166526, AY880939.1, L54115.1; S. variegatusAF231914, AJ272419.1, AJ272420.1, AJ272421.1, L54081.1; S.weaverae L54091.1.
These environmental Suillus sequences were retrievedfrom GenBank: AF476994.1, AJ272405.1, AJ272406.1,AJ410857.1, AY097052.1, AY587754.1, AY880932.1,EF458012.1, EF619768.1, DQ351501.1, DQ351502,DQ351503.1, DQ351504.
These newly derived sequences were included: S brevipes(UC18060328) GQ249388, (UC18060327) GQ249389; S.quiescens (Holotype UC1860306) GQ249402, (UC180310)GQ249390, (UC1860312) GQ249391, (UC1860308)GQ249393, (UC1859739) GQ249401, (bioassay-no speci-men) GQ249392; S. occidentalis (HDT28175) GQ249394,(STB86) GQ249395, (STB82) GQ249396, (STB1418)GQ249397; S. volcanalis (HDT39873) GQ249398,
(HDT12800 holotype) GQ249399; S. megaporinus(UC18060326) GQ249400.
The initial analysis used 134 ITS sequences representingall Suillus species and Suillus environmental sequencesavailable (see METHODS AND MATERIALS for list). These werealigned with Clustal X 2.09 (Larkin et al. 2007) andtrimmed to 800 characters (including gaps). The sequenceswere analyzed by neighbor joining and by parsimony withPAUP 4.0d81 (Swofford 2002) running in classic mode on aMac (1.9 GHz PowerPC). Neighbor joining trees usedlikelihood distances with a transition/transversion rate setto 2 and gaps treated as missing data. Parsimony analysisemployed a heuristic searches from a random starting tree,TBR branch swapping and MAXTREES set to 10 000. Theconsensus parsimony tree and the neighbor joining tree(not shown) grouped all Suillus quiescens sequences with asmall set of unidentified environmental sequences in aunique clade nested among other species in the granulatusgroup (S. granulatus [L.] Roussel, S. granulatus sensuThiers & A.H. Sm. 1971, S. brevipes, S. glandulosipes Thiers& A.H. Sm., S. neoalbidipes M.E. Palm & E.L. Stewart, S.luteus [L.] Roussel, S. pseudobrevipes A.H. Sm. & Thiers, S.flavidus [Fr.] J. Presl, S. mediterraneensis [Jacquet. & J.Blum] Redeuilh, S. bellinii [Inzenga] Watling, S. collinitus[Fr.] Kuntze, S. volcanalis).
All latter granulatus-like sequences inclusive of S.quiescens sequences, plus additional sequences of S.collinitus, S. fluryi Huijsman, S. occidentalis and S. volcanalis(63 in total), where realigned with Clustal X and visuallyadjusted. This alignment, which had many fewer gaps, wastrimmed to 695 positions. The dataset was analyzed withneighbor joining and parsimony as indicated above. Inaddition branch strengths were assessed by 10 000 bootstrapreplicates with the fast step-wise addition option and with10 000 jackknife with 33% deletion and the fast step-wiseaddition. Both data matrices and the tree shown weresubmitted to TreeBASE (study S2452; matrices M4663,M4664)
RESULTS AND DISCUSSION
The four ITS sequences of S. quiescens fruiting bodies,combined with five environmental sequences derivedfrom mycorrhizal root tips, form a distinct cladeamong the granulatus-like species (FIG. 1). Branchlengths, bootstrap and jackknife values that supportthe S. quiescens clade are similar to those associatedwith other well recognized species in the group.These results show that S. quiescens is clearly distinctfrom S. brevipes, S. volcanalis and S. occidentalis, thethree species that it macroscopically resembles.
We initially thought that S. quiescens could be avariant of S. brevipes because S. quiescens differsmorphologically from S. brevipes primarily by a muchpaler colored immature pileus and by minute,colored glands on the stipe that develop with age.Although initial BLAST and phylogenetic analysesshowed the two to be different all the available S.
BRUNS ET AL.: SUILLUS QUIESCENS 439

FIG. 1. Parsimony analysis of the ITS sequences for the granulatus group of Suillus. Arrow indicates Suillus quiescens clade.A random tree from the 10 000 trees of 305 length is shown. Bold branches indicated those present in the strict consensus ofall 10 000 trees. Numbers indicate bootstrap support; jackknife values (not shown) were almost identical. GenBank accessionnumbers for the ITS sequences analyzed are derived from basidiocarps (BC), root tips (RT) of forest trees, soil mycelium (MY)and seedling bioassays of spores (BA).
440 MYCOLOGIA

brevipes sequences were from eastern North Americaand it seemed possible that the western materialmight be different and perhaps synonymous with ourcollections. For that reason we added two sequencesof S. brevipes from western North America. We foundthat the sequences derived from western S. brevipeswere slightly different from those derived from theeastern material, but none were closely related to S.quiescens (FIG. 1).
Suillus volcanalis presented a problem for compar-ison because the key characters of the species seemedlike our material (Thiers 1979), but the name S.volcanalis is not often applied and is represented byonly a few collections. The first ITS sequence that weacquired for it was from a collection by Harry Thiersin 1979 at a site near the holotype location. The nearperfect match of this sequence to S. pseudobrevipesbrought up the possibility that the species identifica-tion for the S. volcanalis collection was wrong.However we were able to acquire partial ITS sequencefrom the holotype (from 1965) and this too was foundto be nearly identical to both S. pseudobrevipes and tothe later collection of S. volcanalis, but all thesesequences were different from S. quiescens.
Although S. occidentalis is not known from Califor-nia, it does appear to be reasonably common inArizona, the area from which it originally wasdescribed (Thiers 1976b). Recent photos posted onthe Web by Dr Scott Bates of Arizona State Universitylooked very similar to what we were calling S.quiescens. However the sequences we obtained froma Thiers collection of S. occidentalis and three othersderived from material sent to us from Dr Bates provedto be quite similar to one another but distinct from S.quiescens (FIG. 1).
ITS sequences are not yet available for several otherspecies of granulatus-like Suillus in western NorthAmerica, but the morphological characters for thesespecies are distinctly different from S. quiescens.Among those species with sparsely glandular, non-annulate stipes and glutinous nonfibrillose pilei thefollowing species were considered and rejected aspossible identifications for our material. Suillusbrunnescens A.H. Sm. & Thiers is different by itsinitially white pileus that stains brown, by slightlybroader spores (3.8–4.2 vs. 2.4–3.7 mm) and by itsassociation with five-needle pines (Smith and Thiers1964). Similarly S. pallidiceps A.H. Sm. & Thiers hasan initially white pileus and a stipe that lacks darkglands even at maturity (Smith and Thiers 1964).Suillus wasatchicus Thiers, although possibly similarin color, has more conspicuous glands on the stipe. S.kaibabensis Thiers has pinkish pileus and tubes and astrongly glandular stipe (Thiers 1976a).
Sequences of S. quiescens were found to be close
matches to some unidentified Suillus sequences thatwere retrieved from mycorrhizae of pine seedlingsfrom multiple locations and studies in Oregon andCalifornia (FIG. 1, TABLE I). All these reports werefrom pine seedlings and most were from bioassays inwhich pines were planted in test soils and grownunder laboratory or greenhouse conditions to assayfor spores or other resistant propagules. AlthoughSuillus species are not uncommon in such assays(TABLE I) their frequency of occurrence generallyreaches double digits only when the soil is assayedduring the fruiting season, without drying or dilution,as was done in the Oregon study by Ashkannejhadand Horton (2006). In California, which tends to bedrier than the Oregon site sampled by Ashkannejhadand Horton (2006), such assays usually are dominatedby Rhizopogon and retrieve only a low percentage ofSuillus (Kjøller and Bruns 2003, Rusca et al. 2006).Furthermore spatial frequency of Suillus sporeswithin a site appears to be much lower than that ofRhizopogon (Izzo et al. 2006) and there is someevidence that Suillus spores do not retain theirviability as long as Rhizopogon (Ashkannejhad andHorton 2006). However spores of some Suillusspecies, including S. quiescens, do appear to be fairlyresistant to soil heating (Peay et al. 2009), and so it isinteresting that sequences of S. quiescens also wereretrieved from pine seedlings planted in steam-pasteurized soil that then were planted in the fieldnear Bend, Oregon (Warren et al. 2008).
The above shows that S. quiescens is distributedfrom at least the northern Channel Islands (nearSanta Barbara) to coastal and central Oregon, and theeastern Sierra Nevada (near Mammoth, California),and in most of these areas it is known only from thespore bank. The lack of observed fruiting could bebecause the species has been confused with S. brevipesand simply overlooked. However in Point ReyesNational Seashore, where the senior author hascollected for two decades, it does not appear to be acommon fruiter, even though it has been retrievedfrom the spore bank there in two independentstudies. Furthermore on Santa Cruz Island, where itis a common fruiter, 18% of the seedling bioassaysyielded S. quiescens instead of the intended Rhizopo-gon species. In contrast S. pungens, which was also acommon fruiter on the island, colonized only 2% ofthe seedlings (Grubisha et al. 2007). These resultssuggest that S. quiescens is over-represented in thespore bank studies relative to other Suillus species,and we hypothesize that S. quiescens is an earlysuccessional species that fruits primarily in youngforests and resides in the spore bank for extendedperiods. Some species in the sister genus Rhizopogonare known to have long lived spores and thought to
BRUNS ET AL.: SUILLUS QUIESCENS 441

exhibit such strategies (Bruns et al. 2009), but so farthe limited data on Suillus suggest that they haveshort lived spores (Ashkannejhad and Horton 2006).
Suillus is among the best known genera in NorthAmerica and Europe, but as shown by S. quiescensspecies remain undescribed even in regions that havebeen fairly well collected. Some apparently unde-scribed Suillus species are known and common butcurrently are lumped in morphological concepts thatare too broad. S. granulatus and S. placidus are stellarexamples of such species complexes that are clearlyillustrated by ITS data shown here (FIG. 1) andreported elsewhere (Kretzer et al. 1996, Manian etal. 2001). Suillus also contains species that have beendescribed but are not often collected or perhaps arenot recognized when they are collected; ITS can bevery useful in these cases as well. In the current studyITS data retrieved from historical collections of S.vulcanalis and S. occidentalis were essential toestablish that S. quiescens had not already beendescribed. Although some of the specimens involvedwere more than 40 y old amplifying the ITS region astwo pieces and using specific primers let us workaround the problems of degraded DNA and contri-butions from secondary molds. The limitations of ITSfor species-level determinations also are illustratedhere because some pairs of species are not distin-guished by this locus (S. glandulosipes and S.neoalbidipes, S. pseudobrevipes and S. vulcanalis, S.brevipes and S. weaverae). These pairs might be due toover-description (i.e. synonymy) or to the lack of ITSdivergence among sibling species, but until other
useful loci are developed for the genus this willremain an unresolved issue.
TAXONOMY
Suillus quiescens T.D. Bruns, and E.C. Vellinga sp.nov. FIGS. 2–3
MycoBank MB 515081.A Suillo brevipede juventute pileo pallide luteibrunneola
sed maturitate fuscicinnamomescens, punctis glandulosistenuissimis in stipite producentibus, apice luteolo stipitis, etordinatione DNA differt. Typus hic designatus: L. Grubisha747.
Holotype: LG747 (UC1860306) University of Cali-fornia Herbarium (UC).
Etymology. Quiescens refers to the species’ ability tolay quiescent in the spore bank until it encounterspine roots.
Pileus broad, hemispheric to broadly convex,moderate to large, 6–12 cm diam; color approachingstrong brown (s. Br 55) or deep brown (deep Br 56)on mature pilei, sometimes with olivaceous patches ortones, much paler when young but still some shade oflight Brown (l Br 57) or between light brown (57) andlight orange (l.o. 52), or grayish yellow; covered inglutinous layer, but glutin often drying in strands togive a slightly fibrillose look to older specimens;margin in-rolled when young with a limited sterilezone of a millimeter or less. Pileus context white,generally unchanging, sometimes with brownishstains just under cuticle, and pale yellow tones justabove tubes. Tubes when young pale yellow (near 89,p.Y.) and with light brown or yellowish brown
TABLE I. Reports of Suillus species identified from bioassay studies in western USA
Species % Region Reference
S. brevipes 13a Oregon central coast Ashkannejhad and Horton 20062 California central coast (Point Reyes) Peay et al. 2009
S. pungens ,1 Southeastern Sierra Nevada Rusca et al. 20062 California central coast (Point Reyes) Peay et al. 20098b California central coast (Point Reyes) Kjøller and Bruns 20032 Santa Cruz Island, California Unpubl from Grubisha et al. 2007
S. pseudobrevipes ,1 Southwestern Sierra Nevada, California Izzo et al. 2006California central coast Peay et al. 2009
S. quiescens 8a Oregon central coast Ashkannejhad and Horton 2006,1 Southeastern Sierra Nevada Rusca et al. 2006
3 California central coast (Point Reyes) unpubl from Bruns et al. 20091 California central coast (Point Reyes) Peay et al. 2009
18 Santa Cruz Island, California unpubl from Grubisha et al. 2007S. tomentosus 20a Oregon central coast Ashkannejhad and Horton 2006
3 California central coast (Point Reyes) Peay et al. 200917b Northern California coast (Salt Point) Kjøller and Bruns 2003
S. umbonatus 15a Oregon central coast Ashkannejhad and Horton 2006
a Averaged from their three site types.b Based on culture; numbers not fully comparable to others.
442 MYCOLOGIA

glandular secretions; older tubes slightly lighter thanbrilliant orange yellow (67 Brill OY), becomingolivaceous yellow (near 84, s.Y.) Mature tube mouthsradially elongate but generally less than 1 mm wide.Stipe usually short, typically 2–4 cm long, but some-times longer (6–8 cm), even, or very slightly bulbous,or tapered to the base, pale yellow (p. Y 89) to lightyellow (l. Y. 86) on apical fifth; lower part of stipesame color at apex or white or overlaid with a lightbrown (l.Br. 57) layer or streaks of glutin as on pileus,occasionally this overlay gives the impression of aannular zone (FIG. 2C); stipe surface finely glandular;
glands initially only slightly darker than stipe surface,but sometimes light brown, becoming nearly blackafter drying. Stipe context white and unchanging, butsometimes with orangish stains in older or insect-damaged parts. Spore deposit not determined, butprobably between strong yellow brown (s.y Br 74) andstrong brown (s Br 55) as seen on pileus surface ofseveral specimens.
Spores [72, 7, 7] in side view 6.1–14.7 3 2.4–3.7 mm,avl 3 avw 5 7.7–8.7 3 2.8–3.0 mm, Q 5 2.4–3.3, avQ 5
2.7–2.9, elongate, with a slight suprahilar depressionin side view, and oblong in face view, most with a
FIG. 2. Suillus quiescens basidiomes. A. Holotype UC1860306 collection from Santa Cruz Island. B. Collection UC1860307from Santa Cruz Island showing in-rolled margin on young specimen. C. Collection UC1859739 from Point Reyes NationalSeashore showing brown overlay on lower stipe.
BRUNS ET AL.: SUILLUS QUIESCENS 443
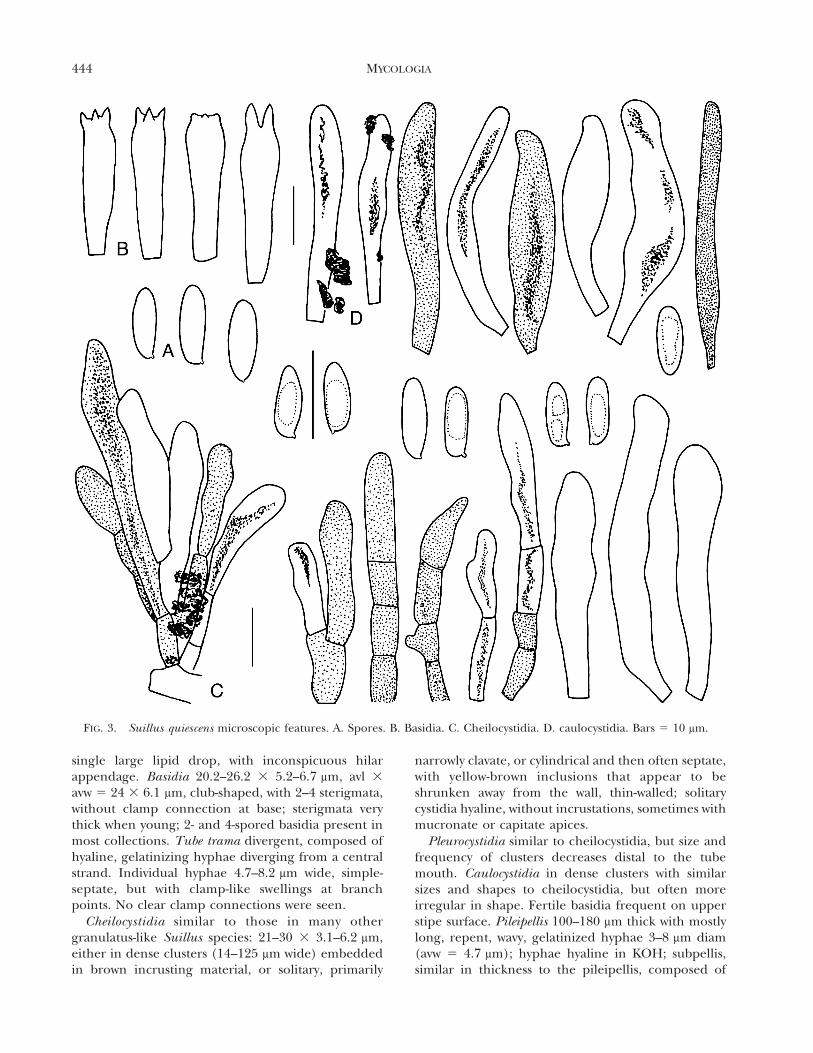
single large lipid drop, with inconspicuous hilarappendage. Basidia 20.2–26.2 3 5.2–6.7 mm, avl 3
avw 5 24 3 6.1 mm, club-shaped, with 2–4 sterigmata,without clamp connection at base; sterigmata verythick when young; 2- and 4-spored basidia present inmost collections. Tube trama divergent, composed ofhyaline, gelatinizing hyphae diverging from a centralstrand. Individual hyphae 4.7–8.2 mm wide, simple-septate, but with clamp-like swellings at branchpoints. No clear clamp connections were seen.
Cheilocystidia similar to those in many othergranulatus-like Suillus species: 21–30 3 3.1–6.2 mm,either in dense clusters (14–125 mm wide) embeddedin brown incrusting material, or solitary, primarily
narrowly clavate, or cylindrical and then often septate,with yellow-brown inclusions that appear to beshrunken away from the wall, thin-walled; solitarycystidia hyaline, without incrustations, sometimes withmucronate or capitate apices.
Pleurocystidia similar to cheilocystidia, but size andfrequency of clusters decreases distal to the tubemouth. Caulocystidia in dense clusters with similarsizes and shapes to cheilocystidia, but often moreirregular in shape. Fertile basidia frequent on upperstipe surface. Pileipellis 100–180 mm thick with mostlylong, repent, wavy, gelatinized hyphae 3–8 mm diam(avw 5 4.7 mm); hyphae hyaline in KOH; subpellis,similar in thickness to the pileipellis, composed of
FIG. 3. Suillus quiescens microscopic features. A. Spores. B. Basidia. C. Cheilocystidia. D. caulocystidia. Bars 5 10 mm.
444 MYCOLOGIA

hyphae with dark yellow-brown contents and oftenwith granular brownish incrustations in KOH. Pileuscontext composed of inflated (10–24 mm wide),branched hyphae; some branches inflated near thesepta and in some views giving the impression ofclamps, but no clear clamp connections were seen.
Habitat and distribution. Fruiting in small groupsespecially with young Bishop pine on Santa CruzIsland, where it is the most commonly encounteredSuillus species. It is also encountered when one usespine seedlings to bioassay soil from pine forests of theislands, coastal and montane areas of California andOregon. In the Channel Islands it was collectedfruiting Jan–Mar, and on the central coast at PointReyes National Seashore it has been collected fruitingwith young Bishop pine mid-November to earlyDecember. Given its distribution in Oregon andeastern California other hosts such as lodgepole,jeffrey and ponderosa pines are expected.
Collections studied. USA, CALIFORNIA, Santa Bar-bara County, Santa Cruz Island, junction of RidgeRoad and Sauces Canyon Road, 119u48.6929W,34u00.499N, 388 m, under young to moderate agedPinus muricata, 13 Jan 2002, Lisa Grubisha LG747HOLOTYPE UC (1860306); USA, CALIFORNIA,Santa Barbara County, Santa Cruz Island, junction ofRidge Road and Sauces Canyon Road, 119u48.9209W,34u00.089N, 399 m, under young Pinus muricata, 1Mar 2001, Lisa Grubisha LG359, UC (1860310); USA,CALIFORNIA, Santa Barbara County, Santa CruzIsland, junction of Ridge Road and Sauces CanyonRoad. Eastern most pines along Ridge Road,119u47.6509W, 34u00.779N, 388 m, under young tomoderate aged Pinus muricata, 1 Mar 2001, LisaGrubisha LG360, UC (1860312); USA, CALIFORNIA,Santa Barbara County, Santa Cruz Island, along SaucesCanyon Road. Eastern most pines along Ridge Road,119u48.9909W, 34u00.119N, 399 m, under young Pinusmuricata, 1 Mar 2001, Lisa Grubisha LG370, UC(1860311); USA, CALIFORNIA, Santa Barbara Coun-ty, Santa Cruz Island, along Sauces Canyon Road.Eastern most pines along Ridge Road, 119u48.9909W,34u00.119N, 388 m, under young to moderate agedPinus muricata with a well developed organic layer, 1Mar 2001, Lisa Grubisha LG373, UC (1860309); USA,CALIFORNIA, Santa Barbara County, Santa CruzIsland, along Sauces Canyon Road. Eastern most pinesalong Sauces Canyon Road, 119u48.9989W,34u00.119N, 388 m, under young to moderate agedPinus muricata with a well developed organic layer, 1Mar 2001, Lisa Grubisha LG372, UC (1860308); USA,CALIFORNIA, Santa Barbara County, Santa CruzIsland, intersection of Ridge Road and LagunitasSecus Road. 119u 47.7509W, 34u00.7979N, 423 m,Open forest shrub community of young bishop pine
and oak, 13 Jan 2002, Lisa Grubisha LG830, UC(1860307).
Summary comparison. Suillus quiescens is most dis-tinctive when young and fresh and then only by itsmacroscopic characteristics. It gives one the impres-sion of a pale version of the eastern North American S.brevipes because of the short stipe and the glutinouspileus. However the light brown pileus when young,the fine glandular dots at the top of the stipe atmaturity and the yellowish stipe apex separate S.quiescens from S. brevipes, which has an almostchocolate (59 d Br.) pileus and a pure white, glandlessstipe when young. Suillus pungens, which occurs in thesame habitats is white when immature and has strongolivaceous tones before becoming similar in color toolder S. quiescens. The stipe of S. pungens is muchmore prominently glandular. The in-rolled margin ofyoung S. quiescens is similar to S. glandulosipes or S.neoalbidipes, but in contrast to the latter two species ithas only a small sterile zone that one might need ahand lens to see. Suillus occidentalis appears to besimilar in appearance, but at maturity the pileus islighter colored than that of S. quiescens.
ACKNOWLEDGMENTS
We thank the Harry Thiers Herbarium (SFSU) for loans ofS. occidentalis and S. volcanalis specimens, Peter Kennedyfor determining the ITS sequences of S. quiescens,TDB3205, the University of California Santa Cruz IslandNatural Reserve and Channel Islands National Park forpermission to collect fungal specimens, Lyndal Laughrinand Sarah Chaney for arranging logistical support on SantaCruz Island and Santa Rosa Island respectively, and RoyHalling, Manfred Binder and an anonomous reviewer formany useful suggestions on the manuscript Financialsupport was supplied in part by NSF DEB-0618293 to TDBand ECV. We dedicate this work to Harry Thiers, whosecommand of the western North American Suillus speciesremains unsurpassed.
LITERATURE CITED
Ashkannejhad S, Horton TR. 2006. Ectomycorrhizal ecologyunder primary succession on coastal sand dunes:interactions involving Pinus contorta, suilloid fungiand deer. New Phytol 169:345–354.
Bruns TD, Peay KG, Boynton PJ, Grubisha LC, Hynson NA,Nguyen NH, Rosenstock NP. 2009. Inoculum potentialof Rhizopogon spores increases with time over the first4 y of a 99 y spore burial experiment. New Phytol 181:463–470.
Gardes M, Bruns TD. 1993. ITS primers with enhancedspecificity for basidiomycetes—application to the iden-tification of mycorrhizae and rusts. Mol Ecol 2:113–118.
Grubisha LC, Bergemann SE, Bruns TD. 2007. Host islandswithin the California northern Channel Islands create
BRUNS ET AL.: SUILLUS QUIESCENS 445

fine-scale genetic structure in two sympatric species ofthe symbiotic ectomycorrhizal fungus Rhizopogon. MolEcol 16:1811–1822.
———, Trappe JM, Bruns TD. 2005. Preliminary record ofectomycorrhizal fungi on two California ChannelIslands. In: Garcelon D, Schwemm C, eds. Proceedingsof the 6th California Islands Symposium. NPS Tech.Pub. CHIS-05-01, Institute for Wildlife Studies, Arcataand Ventura, California. p 171–183.
Izzo A, Nguyen DT, Bruns TD. 2006. Spatial structure andrichness of the ectomycorrhizal resistant propagulecommunity colonizing hosts with differing seedlingestablishment patterns. Mycologia 98:374–383.
Kenn L, Kelly KL, Judd DB. 1976. ISCC-NBS Color-namecharts illustrated with centroid colors. In: Supplementto US Natl. Bureau Standards. Circular 553: The ISCC-NBS method of designing colors and a dictionary ofcolor names. 18 p.
Kjøller R, Bruns TD. 2003. Rhizopogon spore bank commu-nities: within and among Californian pine forests.Mycologia 95:603–613.
Koljalg U, Larsson KH, Abarenkov K, Nilsson RH, Alexan-der IJ, Eberhardt U, Erland S, Høiland K, Kjøller R,Larsson E, Pennanen T, Sen R, Taylor AFS, Tedersoo L,Vralstad T, Ursing BM. 2005. UNITE: a databaseproviding Web-based methods for the molecularidentification of ectomycorrhizal fungi. New Phytol166:1063–1068.
Kretzer A, Bruns TD. 1997. Molecular revisitation of thegenus Gastrosuillus. Mycologia 89:586–589.
———, Li Y, Szaro T, Bruns TD. 1996. Internal transcribedspacer sequences from 38 recognized species of Suillussensu lato: phylogenetic and taxonomic implications.Mycologia 88:776–785.
Larkin MA, Blackshields G, Brown NP, Chenna R, McGetti-gan PA, McWilliam H, Valentin F, Wallace IM, Wilm A,Lopez R, Thompson JD, Gibson TJ, Higgins DG. 2007.Clustal W and Clustal X version 2.0. Bioinformatics 23:2947–2948.
Manian S, Sreenivasaprasad S, Bending GD, Mills PR. 2001.
Genetic diversity and interrelationships among com-mon European Suillus species based on ribosomalDNA sequences. FEMS Microbiol Lett 204:117–121.
Peay KG, Garbelotto M, Bruns TD. 2009. Spore heatresistance plays an important role in disturbance-mediated assemblage shift of ectomycorrhizal fungicolonizing Pinus muricata seedlings. J Ecol 97:537–547.
Rusca TA, Kennedy PG, Bruns TD. 2006. The effect ofdifferent pine hosts on the sampling of Rhizopogonspore banks in five eastern Sierra Nevada forests. NewPhytol 170:551–560.
Smith AH, Thiers HD. 1964. A contribution toward amonograph of the genus Suillus. Ann Arbor, Michigan:privately printed.
———, ———. 1971. The Boletes of Michigan. Ann Arbor:Univ. Michigan Press. 428 p.
Swofford DL. 2002. PAUP*: phylogenetic analysis usingparsimony (*and other methods). Version 4. Sunder-land, Massachusetts: Sinauer Associates.
Thiers HD. 1976a. Boletes of the southwestern UnitedStates. Mycotaxon 3:261–273.
———. 1979. The genus Suillus in western United States.Mycotaxon 9:285–296.
Warren JM, Brooks JR, Meinzer FC, Eberhart JL. 2008.Hydraulic redistribution of water from Pinus ponderosatrees to seedlings: evidence for an ectomycorrhizalpathway. New Phytol 178:382–394.
White TJ, Bruns TD, Lee SB, Taylor JW. 1990. Amplificationand direct sequencing of fungal ribosomal RNA genesfor phylogenetics. In: Innis MA, Gelfand DH, SninskyJJ, White TJ, eds. PCR protocols: a guide to methodsand applications. New York: Academic Press. p 315–322.
Wu Q-X, Mueller GM, Lutzoni FM, Huang Y-Q, Guo S-Y.2000. Phylogenetic and biogeographic relationships ofeastern Asian and eastern North American disjunctSuillus species (Fungi) as inferred from nuclearribosomal RNA ITS sequences. Mol Phylogen Evol 17:37–47.
446 MYCOLOGIA