synaptic exocytosis of dense-core vesicles in blue crab (callinectes sapidus) stomach muscles
TRANSCRIPT

052415.666 130513.125 130513.250 130513.688 130920.166 131518.937 131520.710 132119.426140518.855 140521.350 140521.416 140521.936 161519.874 161805.830 192015.500 192514.284192514.402 192514.710030112*425
Synaptic exocytosis of dense-core vesicles in blue crab(Callinectes sapidus) stomach musclesVarsha Patel, C.K. Govind
Life Sciences Division, University of Toronto at Scarborough, 1265 Military Trail, Scarborough, Ontario, Canada M1C 1A4
&misc:Received: 14 October 1996 / Accepted: 21 February 1997
&p.1:Abstract. Neuromuscular terminals of a single moto-neuron to four muscles (CPV7a, GM5a, CV2, and CV3)in the stomach of the blue crab Callinectes sapidusshowed structural evidence for the exocytotic release ofdense-core vesicles exclusively at synapses. The primaryevidence was the appearance of dense cores in the syn-aptic cleft, accompanied by indentations of the presyn-aptic or postsynaptic membrane. In their simplest form,these consisted of an omega-shaped figure of the presyn-aptic membrane enclosing one dense core, denoting re-lease of a single dense-core vesicle. A larger indentationof the presynaptic membrane enclosing several densecores denoted multiple release. A more complex form ofmultiple release was where the presynaptic membranewas normal, but the postsynaptic membrane elaboratedinto a sac projecting into the granular sarcoplasm andfilled with dense cores. The postsynaptic sac in some in-stances was compressed into a thin, fingerlike extension,which lacked dense cores and, at its distal end, separatedinto small cisternae, suggesting a mechanism for mem-brane recycling. Profiles depicting single and multiplereleases of dense-core vesicles were found more fre-quently at neuromuscular terminals that release relative-ly large amounts of transmitter with a single stimulus,such as CV2 and CV3, compared to those releasingsmaller amounts, such as CPV7a and GM5a. The dispar-ity in release sites among the four muscles of this singlemotor unit and the fact that many of the multiple-releasefigures were closely adjacent to the active zones fortransmitter release suggest a possible modulatory rolefor dense-core vesicles in synaptic transmission. Suchmodulation may be long lasting, as implied by the post-synaptic sacs, which may permit prolonged release ofthe contents of their dense cores into the synaptic cleft.This is in keeping with the functional role of these stom-ach muscles, which is to be continuously active for longperiods of time.
&kwd:Key words: Neuromuscular junction – Synaptic vesicles– Vesicle fusion – Electron microscopy – Callinectessapidus(Crustacea)
Introduction
The long-standing view of a single-transmitter neuronhas been gradually replaced by a multiple-messengermodel, which incorporates the existence of not only theclassical transmitter but other messengers, such as nucle-otides, neuropeptides, and neurohormones, substancesthat can be synthesized and stored along with the classi-cal transmitter in a variety of neuron types (Thurson-Klein et al. 1988). While the classical transmitter is forrapid signalling, these other messengers serve to modu-late the action of the classical transmitter via presynapticand postsynaptic mechanisms (Hökfelt et al. 1982; Burn-stock 1986). Within nerve terminals the storage of theclassical transmitter is in small (40–50 nm in diameter)clear vesicles, while that of the other messengers is inlarge (100–200 nm in diameter) dense-core vesicles(Thurson-Klein et al. 1988). These two distinct vesiclepopulations are usually found in disproportionate num-bers in nerve terminals, with the clear vesicles formingthe overwhelming majority. Release of classical trans-mitter from these clear vesicles occurs at the active zoneof synaptic contacts where vesicles are highly clustered(reviewed by Heuser and Reese 1977; Burns and Augus-tine 1995). The high concentration of ion channels at theactive zone admits calcium in response to an action po-tential, resulting in the fusion of docked vesicles withthe presynaptic membrane. Thus exocytosis of clear ves-icles results in the quantal release of transmitter. Struc-tural evidence for vesicle exocytosis in nerve terminalsis seen in the form of omega-shaped figures at the pre-synaptic membrane, usually at the active zone (Heuserand Reese 1977; Burns and Augustine 1995).
Structural evidence for the release of the contents ofdense-core vesicles also appears in the form of omega-
Supported by the Natural Sciences and Engineering ResearchCouncil of Canada
Correspondence to:C.K. Govind&/fn-block:
Cell Tissue Res (1997) 289:517–526
© Springer-Verlag 1997

518
shaped figures (Thurson-Klein et al. 1988). However,such fusion sites are usually at nonsynaptic areas of thenerve terminal, although exocytosis at synapses has beennoted in rare instances (Dickinson-Nelson and Reese1983). The regional segregation of release sites for clearand dense-core vesicles implies that the content of theformer acts in rapid signalling while that of the latter ismore slow-acting and in keeping with the role of dense-core vesicles in modulating the action of classical trans-mitters. It was therefore unusual to find exocytosis ofdense-core vesicles at synaptic contacts of crustaceanneuromuscular terminals as described in the present re-port.
The co-existence of classical neurotransmitter withother secretory substances is also seen in invertebrateneurons (Golding and Bayraktaroglu 1984; Buma andRoubos 1986; Schürmann et al. 1991; Beltz 1995), andthis is reflected in the presence of different vesicle types,including both clear and dense-core vesicles. The pres-ence of omega figures containing dense cores in the neu-ropilar projections of neurons was taken as evidence forexoytosis of dense-core vesicles in the central nervoussystem (Golding and Bayraktaroglu 1984; Schürmann etal. 1991) and occurred typically at nonsynaptic sites;synaptic sites appeared to be reserved for fusion of clearvesicles. For neuromuscular terminals that are usuallypopulated by a small number of dense-core vesicles inaddition to large numbers of clear vesicles (Atwood etal. 1971), there is considerable evidence for excoytosisof clear vesicles at the active zones of synapses (Rheu-ben and Reese 1978; Pearce et al. 1986), but evidencefor exocytosis of dense-core vesicles is lacking. It wastherefore noteworthy to find exocytosis of dense-corevesicles at neuromuscular terminals of a crustacean mo-toneuron.
The motoneuron that we chose to examine innervatesfour separate muscles in the stomach of the blue crabCallinectes sapidus(Govind et al. 1975). These fourmuscles, CPV7a, GM5a, CV2, and CV3 (named ac-cording to Maynard and Dando 1974), bring aboutmovements of a ridge-like structure that regulates thepassage of food from the anterior (cardiac) region to theposterior (pyloric) region of the stomach (reviewed byClaiborne and Ayers 1987; Factor 1995). Neuromuscu-lar terminals in these four muscles release differentamounts of transmitter in response to a single pulse;those on GM5a and CPV7a generate relatively-small(2–5 mV) excitatory postsynaptic potentials comparedto the several-fold higher (15–30 mV) postsynaptic po-tentials in CV2 and CV3 (Govind et al. 1975). Differ-ences in synaptic performance among the four musclestranslate into differences in activity, despite the fact thatall four are innervated by a single motoneuron. It is thisaspect of the stomach motoneuron that we initially setout to explore, but along the way uncovered exocytosisof dense-core vesicles at synaptic contacts. We alsofound that dense-core vesicle exocytosis occurred morefrequently in muscles generating large excitatory post-synaptic potentials, suggesting differentiation of thesenerve terminals for both clear vesicles and dense-corevesicles.
Materials and methods
Adult blue crabs (Callinectes sapidus) were purchased from a lo-cal fish store and held for a few days without food in an aquariumcontaining artificial seawater (Instant Ocean) at 22°C. The stom-ach muscles were dissected by chipping away the dorsal exoskele-ton to expose the foregut and then cutting the esophagus and mid-gut to isolate the foregut (Govind et al. 1975). The isolated fore-gut was thoroughly rinsed in saline, pinned out in a dish to exposethe intrinsic muscles on the stomach wall, and prepared for elec-tron microscopy by techniques used previously (Atwood et al.1977). The preparation was superfused with primary fixative for30 min in order to minimize contractile movement and damage tothe innervation, and the individual muscles were then processedfor an additional hour. The primary fixative contained 2.5% glu-taraldehyde, 0.5% formaldehyde, 4% sucrose, and 0.1% CaCl2 in0.1 M sodium cacodylate buffer at pH 7.4. The tissue was brieflyrinsed in sodium cacodylate buffer and postfixed in osmium te-troxide for 1 h. Following a brief rinse in buffer, the tissue was de-hydrated in ethanol, transferred through propylene oxide, and em-bedded in resin.
Ultrastructural sections of the muscles were examined to lo-cate nerve terminal regions, and some of these were serially sec-tioned for lengths of 5–20µm. Electron micrographs at a finalmagnification of 25 000× enabled qualitative and quantitative ana-lyses of nerve terminals (Pearce et al. 1986; King et al. 1996). Thenerve-terminal length was determined by addition of the estimat-ed-thickness value of each section traversed by the terminal. Thelength of the terminal branches was also included in the estima-tion of total terminal length. The lengths of all terminals found inan area of serially sectioned tissue were summed to yield a finalvalue for the length of terminal sectioned for a given tissue sam-ple.
For morphometric analysis, an acetate grid-sheet with dotsplaced 0.5 cm apart was superimposed on every fifth section in or-der to determine the percentage of a terminal occupied by mito-chondria, clear vesicles, dense-core vesicles, and axoplasm (Kinget al. 1996). This was accomplished by keeping a record of the to-tal number of dots landing on each of the constituents of the ter-minal. Terminal volume was determined by multiplying the totalnumber of dots occupied by the terminal by [0.5 cm×magnifica-tion factor (µm2)] to yield the surface area of the terminal in thesection. By multiplying the surface area with the length of the ter-minal that included this section and the next few unanalyzed ones[based on section-thickness estimates (µm)], the volume of thatportion of the terminal was calculated. This procedure was repeat-ed for the entire length of the terminal. The total volume of theterminal was obtained by adding together the values for the vol-ume of the terminal that spanned the distance between one ana-lyzed section selected for volumetric analysis to the next.
Synapse size was determined by multiplying the length of thesynaptic profile in a section (using a caliper) by the section thick-ness (Pearce et al. 1986) The surface area of the synapse in thissection was added to those values obtained from all consecutivesections in which the synapse was present to yield the total sur-face area for that synapse. Mean synaptic area was determinedfrom completely serially sectioned synapses (which included theirbeginning and ending) by adding together the areas of these com-plete synapses and dividing by the total number. Total synaptic ar-ea was calculated by adding together the areas of incomplete(which either did not begin or did not end within the sectioned re-gion) and complete synapses.
Four animals were prepared for electron microscopy, and twoof the better-preserved ones were examined in detail. In one ani-mal, a total of 15 different terminal regions was sampled, fivefrom CPV7a, four from GM5a, two from CV2, and four fromCV3. In the other animal, five different terminal regions weresampled from each of the four muscles. In all, 35 separate nerveterminals were sampled via serial sectioning; altogether, 500–800sections were examined to generate the morphometric data.

Results
Neuromuscular terminals on the four muscles makingup the single motor unit of the inferior cardiac moto-neuron (Maynard and Selverston 1975) show the typicalcrustacean pattern (Atwood 1976, 1982) (Fig. 1). Nerveterminals were found around the perimeter of muscle fi-bers in areas consisting primarily of granular sarco-plasm, mitochondria, and glial cell processes. Glial cellprocesses demarcated the boundaries of the synapticcontacts that occurred between the terminal membraneand the granular sarcoplasm. Synapses were character-ized by darkly-stained presynaptic and postsynapticmembranes, uniformly separated by a distance of about20 nm. Dense bars, representing active sites of transmit-ter release, resided on the presynaptic membrane as
electron-opaque hemispherical structures around whichclusters of clear vesicles were seen. These clear vesi-cles, containing the neurotransmitter glutamate, werespherical (40 nm in diameter) and occurred in largenumbers.
In addition to these typical features, nerve terminalsof the four stomach muscles were characterized by asmall but distinct population of dense-core vesicles(Fig. 1). These vesicles were also spherical but larger(100–150 nm in diameter), and they had an outer mem-brane slightly separated from a smaller dense-stainingcore. The dense core was usually located centrally, al-though a small proportion of these vesicles contained ec-centric cores, and a few vesicles were seen in which thecore was completely dense. Dense-core vesicles werenot localized to any particular region of the terminal but
519
Fig. 1. Cross-section of CV2 excitatory terminal (t), characteristi-cally filled with many small clear vesicles, a few large dense-corevesicles, and mitochondria. Synaptic contacts (between short ar-rows) on membrane adjacent to granular sarcoplam (s) are demar-
cated by glial tissue (g). Presynaptic dense bar (long arrows) witha cloud of clear synaptic vesicles indicates the active zone of thesynapse. ×43 700. Bar: 1 µm&/fig.c:

520
were scattered throughout the terminal, intermixed withthe more-abundant clear vesicles.
Exocytosis of dense-core vesicles
In our survey of these four muscles, we observed evi-dence of exocytosis of dense-core vesicles at synaptic
contacts, essentially in the form of dense cores withinthe synaptic cleft (Fig. 2B). At these sites, the presynap-tic membrane showed distinct infoldings, which wereusually seen to incorporate one irregularly-shaped denseobject resembling the dense core of a dense-core vesicle.On occasion, two or more electron-opaque cores werelocated between the invaginated presynaptic membraneand the unaffected postsynaptic membrane.
Fig. 2A–F.Putative exocytosis of dense-core vesicles at CV2 neu-romuscular synapses. Nerve terminals (t) populated with clear anddense-core vesicles make synaptic contact (between short arrows)with muscle membranes in areas of granular sarcoplasm (s). AInitial fusion of dense-core vesicle to presynaptic membrane. BSlight infolding of presynaptic membrane enclosing dense-corematerial, indicative of a single dense-core vesicle which has fusedwith presynaptic membrane. C Synapse with presynaptic densebar (long arrow), next to which the synaptic cleft is enlarged andcontains many dense cores, indicative of multiple exocytosis. Thepresynaptic membrane in this area stains typically dark but has a
ruffled appearance. D Synapse, next to which is a large invagina-tion of postsynaptic membrane projecting as a sac into the granu-lar sarcoplasm and containing several dense cores. E Synapsewith two presynaptic dense bars (long arrows), between which thesynaptic cleft is enlarged and contains three dense cores. Thepostsynaptic membrane in this area extends as a slender fingerlikeinvagination into granular sarcoplasm. F The granular sarcoplasmadjacent to a synapse has a slender folded membranous structure(double-headed arrow) resembling the collapsed invagination ofthe postsynaptic sac in E above. ×70 000. Bar: 0.5µm&/fig.c:
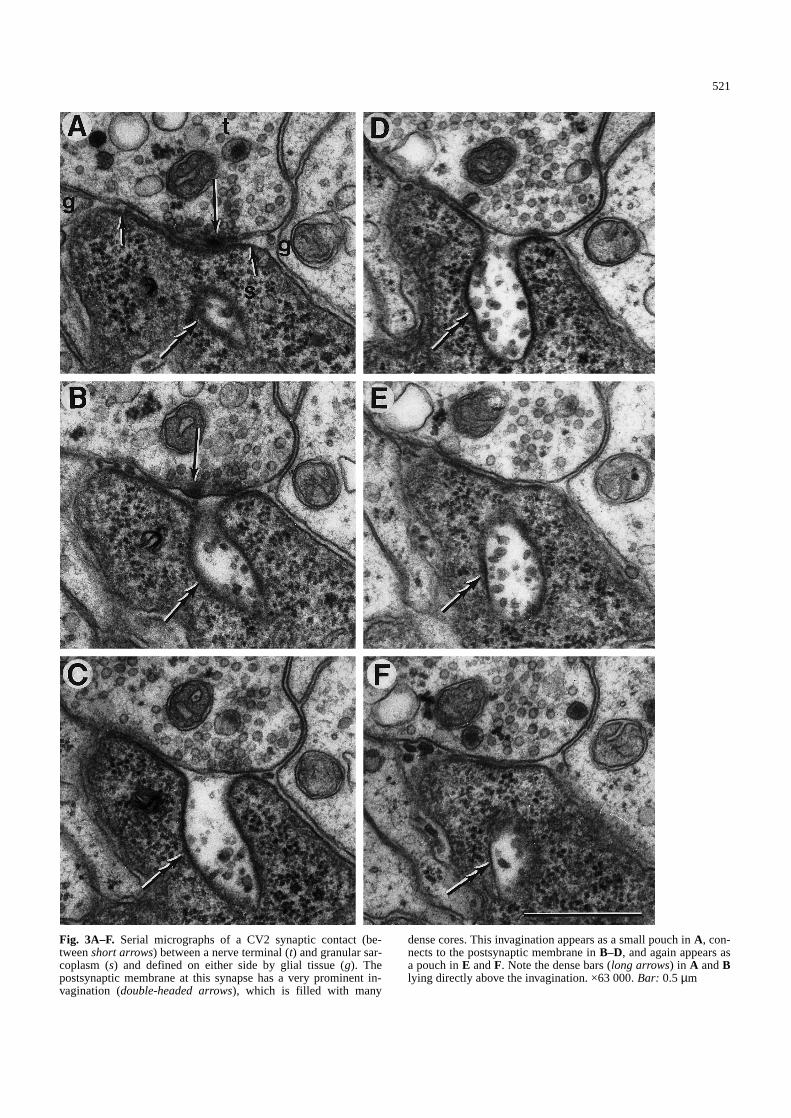
Fig. 3A–F. Serial micrographs of a CV2 synaptic contact (be-tween short arrows) between a nerve terminal (t) and granular sar-coplasm (s) and defined on either side by glial tissue (g). Thepostsynaptic membrane at this synapse has a very prominent in-vagination (double-headed arrows), which is filled with many
dense cores. This invagination appears as a small pouch in A, con-nects to the postsynaptic membrane in B–D, and again appears asa pouch in E and F. Note the dense bars (long arrows) in A and Blying directly above the invagination. ×63 000. Bar: 0.5µm&/fig.c:
521

522
Fig. 4. Selected micrographs from a long series of 16 serial sec-tions beginning in A with a CV2 nerve terminal (t) making synap-tic contact (between short arrows) with granular sarcoplasm (s)and showing two distinct presynaptic dense bars (long arrows).The postsynaptic membrane forms a large sac (double-headed ar-
rows in B, C) filled with dense cores. The sac becomes a thin, fin-gerlike extension (double-headed arrowsin D, E), which at itsdistal end begins to lose its dense staining and separates into un-specialized membrane-bound structures (double-headed arrowsinF–H). ×47 800. Bar: 1 µm

523
Although it was difficult to find a dense-core vesiclebeing attached to the presynaptic membrane at its earlieststage of exocytosis, a few examples were found where adense-core vesicle was in very close association with thepresynaptic membrane (Fig. 2A). These examples arehighly suggestive of an ensuing attachment process,which might have been observed had the tissue been ap-propriately sectioned. It can be inferred that the chance ofcapturing the initial fusion of the dense-core vesicle tothe presynaptic membrane is highly improbable giventhat the attachment point (i.e., neck region connecting thedense-core vesicle to the synapse) is likely to be less thanthe thickness of an ultrastructural section.
Putative fusion sites with several dense cores repre-sented exocytosis of multiple dense-core vesicles, andthese sites were usually associated closely with a presyn-aptic dense bar at which clear vesicles were clustered(Fig. 2C). At these multiple-fusion sites, the presynapticmembrane assumed a ruffled appearance. Alternatively,the presynaptic membrane appeared normal, but therewas an invagination of the postsynaptic membranewhich contained several irregular dense cores (Fig. 2D).With serial sectioning, the postsynaptic invagination wasuncovered as a sac projecting into the granular sarco-plasm and populated with dense cores (Fig. 3). The neckof a postsynaptic sac varied in width, but the sac itselfprojected perpendicularly for some distance, from as fewas four sections to as many as 18.
All such sacs were closely associated with presynap-tic dense bars, one to three of which occurred around theorigin of the sac (Figs. 2D,E; 3A,B). The presynapticdense bars showed the usual clustering of clear vesicles,suggesting release of conventional transmitter.
An equally striking finding was that of postysynapticsacs in which the proximal region was intact and con-tained dense cores (Figs. 2E, 4A–C) but the distal regionappeared collapsed, forming a slender fingerlike invagi-nation with the opposing membranes very close to eachother (Fig. 4D–H). These invaginations lacked densecores, but the apposing membranes showed the charac-teristic staining of regular postsynaptic membrane. Theyusually went through several consecutive sections, andwhen visualized in three dimensions, they appeared as a
rectangular sheet connected at some point to the syn-apse. In a few cases, the most-distal end fragmented intosmall sacs, which did not have the characteristic darkstaining of postsynaptic membranes (Fig. 4F–H). Pro-files similar to the fingerlike invagination of the postsyn-aptic sacs described above were on occasion seen in thegranular sarcoplasm in the vicinity of synaptic contacts(Fig. 2F). They, however, were not connected to a post-synaptic sac or the postsynaptic membrane and appearedto be the distal remnants of postsynaptic sacs.
Differences in frequency of dense-core vesicle exocytosis
Profiles of dense-core vesicles were observed in all seri-ally sectioned terminals of both animals, although theywere more prevalent in one animal where they appearedto differ in frequency among the four muscles. Hence, acount of their number was made in this animal, as an in-dicator of release of neuromodulatory substance, andcompared to the number and area of synapses, as an in-dicator of release of conventional transmitter substance(Table 1). Dense-core vesicles occupy a small but rela-tively constant volume of the terminal (3%) in each ofthe four muscles. Clear vesicles, on the other hand, con-stitute a much larger volume (20–30%) of the terminal.Mitochondria, as the only other prevalent component ofterminals, make up 15–20% of the volume. The totalnumber of fusion profiles of dense-core vesicles variedconsiderably among the four muscles, showing an in-creasing trend beginning with CPV7a and followed byGM5a, CV2, and CV3. A similar trend was seen whenthe number of fusions per synapse was calculated, withthe lowest number for CPV7a, followed by GM5a, CV2,and CV3. The number of fusion profiles was also nor-malized to synaptic area and here, too, the lowest fre-quency was found in CPV7a, a slightly higher frequencyin GM5a, and a considerably higher frequency in bothCV2 and CV3. Specifically, there were 0.6 fusion/µm2
of synaptic area in CPV7a, 0.8 fusion/µm2 in GM5a, and1.4 fusions/µm2 in CV2 and CV3.
Because release of conventional transmitter differsamong the four muscles, with CPV7a and GM5a as low-
Table 1.Quantitative comparison of dense-core vesicle fusions in synaptic terminals of a motoneuron to four stomach muscles in the bluecrab Callinectes sapidus&/tbl.c:&tbl.b:
Stomach muscle
CPV7a GM5a CV2 CV3
Terminal length sectioned (µm) 44.56 28.75 33.43 73.3Volume% of dense-core vesicles 2.5 3.6 2.9 2.0Volume% of clear vesicles 22.0 31.2 23.7 27.1Volume% of mitochondria 19.2 20.5 17.1 16.4Total number of fusions 35 49 76 114Number of synapses 93 34 46 56Number of fusions/synapse 0.376 1.441 1.652 2.036Area (µm2) of synapse (mean±SD) 0.686±0.387 1.860±1.783 1.163±0.730 1.453±0.652Total synaptic area (µm2) 63.820 63.251 53.488 81.379Number of fusions/µm2 of synapse 0.548 0.775 1.421 1.401
&/tbl.b:

524
output types and CV2 and CV3 as high-output types(Govind et al. 1975), the number of fusions was com-pared to these two synaptic types (Table 2). When com-bined in this manner, the number of fusions per synapsewas three-fold greater for the high-output terminalscompared to their low-output counterparts. When nor-malized for synaptic area, the number of fusions wastwo-fold higher for the high-output terminals than forthe low-output ones. Clearly, the number of fusions inthis one animal occurred at a higher frequency in termi-nals releasing relatively large amounts of transmittercompared to their counterparts releasing smalleramounts.
Discussion
Exocytotic release of dense-core vesicles, evidenced byomega-shaped profiles, is common in endocrine tissue ofboth vertebrates (Normann 1976) and invertebrates (Ber-lind 1977). This secretory mechanism has also been re-ported for the nervous systems of vertebrates (Thurson-Klein et al. 1988) and invertebrates (Golding and Bay-raktaroglu 1984; Buma and Roubos 1986; Schurmann etal. 1991), and its discovery at the crustacean neuromus-cular junction (present report) reveals several new facetsin the release of this vesicle type.
Most reports for exocytosis of dense-core vesicles,both in vertebrates and invertebrates, have concerned thecentral nervous system; similar reports are lacking forthe neuromuscular junction. Among invertebrates, andparticularly crustaceans, exocytosis is commonly ob-served for clear vesicles but not for dense-core vesicles(Atwood 1976, 1982). Our findings, however, show thatthe small population of dense-core vesicles at neuromus-cular terminals of a blue crab use exocytosis for releas-ing their contents, much as dense-core vesicles do in thecentral nervous system (Cuadras 1989; Schurmann et al.1991). Since fusion profiles were observed in all fourmuscles of this motor unit in both animals sampled, exo-cytosis probably represents a normal event in these ter-minals. Bearing in mind that movements of the stomachwall are frequent and long lasting (Claiborne and Ayers1987; Factor 1995), it is expected that exocytosis ofdense-core vesicles would occur more often in thesemuscles than in some others, such as limb muscles,which have been the object of more intensive studies(Atwood 1976, 1982).
In nerve terminals of both vertebrates and inverte-brates, exocytosis of dense-core vesicles has been rou-tinely observed at nonsynaptic sites; synaptic sites, al-though present in these nerve terminals, appear to be re-served for clear vesicles. In rare cases, exocytosis ofdense-core vesicles has been observed at synaptic con-tacts of vertebrates (Dickinson-Nelson and Reese 1983;Castel et al. 1996), but not for invertebrates. Segregatedsites in the nerve terminal for the release of clear anddense-core vesicles not only imply different specializa-tions of the axolemma for the docking and fusion ofthese two vesicle classes, but also a mechanism for sepa-rating their functions. However, in blue crab stomachmuscles, dense-core vesicle exocytosis was found exclu-sively at synaptic contacts, denoting the presence of nec-essary docking and fusion proteins at these sites. Sincesynaptic contacts are specialized for release of clear ves-icles, it would be efficient to parlay release of other mes-sengers at this same site. Whether proteins for dockingand fusion of clear and dense-core vesicles are similar isunknown, but unlikely, since exocytosis of clear vesiclesoccurs at active zones, whereas that of dense-core vesi-cles occurs elsewhere on the synapse.
Dense-core vesicle exocytosis in blue crab stomachmuscles was not observed at active zones, although fu-sion sites of these vesicles were often immediately adja-cent to active zones. This juxtaposition of fusion sitesfor the two vesicle classes may have functional implica-tions, at least in speeding up any modulatory effects ofdense-core vesicles on the conventional transmitter.More profound actions of the chemical messenger ofdense-core vesicles would involve modulation of trans-mitter release and its action at the presynaptic or post-synaptic site or modulation of the contractile mechanismof the muscle itself. The latter has been shown to occurfor tonic motoneurons in the crayfish abdomen (Bishopet al. 1984, 1987), where release of proctolin enhancesconductance-generated contraction of the slow fibers ofthe superficial flexor muscle, an action which eminentlysuits the functional nature of these motor units in main-taining posture.
Previous reports usually describe exocytosis of singledense-core vesicles and only occasionally of multiplevesicles, either fused into a packet or aligned in rows(Golding and Bayraktaroglu 1984; Schürmann et al.1991). Such packets are also observed in blue crab stom-ach muscles with multiple dense cores in the synapticcleft resembling the packets described in the crayfish
Table 2.Quantitative comparison ofdense-core vesicle fusions between low-output and high-output synaptic termi-nals of a stomach motoneuron in the bluecrab Callinectes sapidus&/tbl.c:&tbl.b:
Synaptic terminals
Low-output High-output(CPV7a, GM5a) (CV2, CV3)
Terminal length sectioned (µm) 73.31 106.73Total number of fusions 84 190Number of synapses 127 102Number of fusions/synapse 0.661 1.863Total synaptic area (µm2) 127.071 134.867Number of fusions/µm2 of synapse 0.661 1.409
&/tbl.b:

central nervous system (Schurmann et al. 1991). Howev-er, in addition to these packets, we observed distinct in-vaginations of the postsynaptic membrane as sacs hous-ing several dense cores and projecting into the granularsarcoplasm. These sacs, in effect, represent an enlarge-ment of the postsynaptic site to accomodate multiple re-lease of vesicles, while the postsynaptic membrane re-tains its receptor function, as deduced by the fact that themembrane of a sac has the same appearance as the regu-lar postsynaptic membrane. Not only does a postsynapticsac house multiple dense-core vesicles, but it may alsoserve to increase the surface area of the receptors and inthis way augment the action of the conventional trans-mitter, as well as that of modulatory messengers if that istheir site of action. In keeping with this view is the factthat postsynaptic sacs were found in close proximity todense bars representing active zones for release of con-ventional transmitter.
It is important to note that postsynaptic sacs weresubstantial structures extending 0.5–1µm in length, giv-ing the impression that they lasted for longer than thetime needed for the release of a single dense-core vesi-cle. In this case, the sacs would provide a mechanism forthe slow, timed release of the chemical messenger in thedense cores rather than an all-or-none release of a singledense-core vesicle.
Once the dense cores within a sac are depleted, thesac would collapse into the fingerlike extensions that weobserved, and these would separate from the postsynap-tic membrane, lose their specialization, and fragment in-to small cisternae as a mechanism for membrane retriev-al. Since invagination of the postsynaptic membrane inthese sacs extends deep into the granular sarcoplasm,membrane retrieval would best be carried out by the tar-get muscle fiber. Membrane recapture by the musclemight be a more conservative and efficient (i.e., requir-ing less energy) method of removing large amounts ofmembrane from the terminal than would its internaliza-tion by the terminal. This resembles receptor-mediatedendocytosis in which target cells internalize hormone-re-ceptor complexes prior to recycling of the latter (Gold-ing and Bayraktaroglu 1984).
Our observation of differences in the numbers of exo-cytotic profiles of dense-core vesicles demonstrates, forthe first time, putative differentiation in the release ofthese vesicles from a single motoneuron. This is not un-expected as there is considerable differentiation in therelease of conventional transmitter from single crusta-cean motoneurons at their neuromuscular junctions (re-viewed by Atwood 1976, 1982). If the chemical messen-gers from dense-core vesicles are to modulate the actionof the conventional transmitter, we may anticipate paral-lel methods for differentiation for both classes of chemi-cal messengers. Certainly, release of the neuromodulatorproctolin in crayfish tonic motoneurons is stimulus de-pendent and may be depleted with sufficient stimulation(Bishop et al. 1987), much as conventional transmitter incrustacean motoneurons. There is also the observationthat long-term in vivo stimulation of a crayfish-limb ton-ic motoneuron increases the number of both clear vesi-cles and dense-core vesicles in its nerve terminals (Mea-
row and Govind 1989), suggesting parallel mechanismsfor neuronal plasticity.
The generation and control of motor patterns to thestomach muscles in crustaceans are areas of intensivestudy (Selverston and Moulins 1987), and several neuro-modulatory substances have been identified in the sto-matogastric ganglion housing the motoneurons (Marder1987). Several varieties of dense-core vesicles character-ize central neuropilar projections of the motoneurons(King 1976). Modulation of motor patterns by thesechemical messengers occurs with their release fromdense-core vesicles in the neuropile of the stomatogasticganglion. The execution of these motor patterns at theperiphery provides another level of control, because dif-ferences in the amount of transmitter released by theneuromuscular synapses of motoneurons governs thelevel of depolarization of the muscle fiber and hence itsdegree of contraction (Govind and Lingle 1987). Neuro-modulatory substances in dense-core vesicles at thesemotor nerve terminals provide an additional level of con-trol (e.g., proctolin’s effect on crayfish tonic abdominalmuscles), and our finding of synaptic exocytosis ofdense-core vesicles, as simple and complex fusion sites,in these neuromuscular terminals provides some provoc-ative insights as to how this may be done.
&p.2:Acknowledgements.We thank Ms. Joanne Pearce for technical as-sistance and criticism of the manuscript and Dr. Ted Brown forcritically reading an earlier draft.
References
Atwood HL (1976) Organization and synaptic physiology of crus-tacean neuromuscular systems. Prog Neurobiol 7:291–391
Atwood HL (1982) Synapses and neurotransmitters. In: AtwoodHL, Sandeman DC (eds) The biology of Crustacea, vol 3(Neurobiology: structure and function). Academic Press, NewYork, pp 105-150
Atwood HL, Luff AR, Morin WA, Sherman RE (1971) Dense-cored vesicles at neuromuscular synapses of arthropods andvertebrates. Experientia 27:816
Atwood HL, Govind CK, Jahromi SS (1977) Excitatory synapsesof blue crab gastric mill muscles. Cell Tissue Res 177:145–158
Beltz B (1995) Neurobiology and neuroendocrinology. In: FactorJR (ed) Biology of the lobster Homarus americanus. Academ-ic Press, San Diego, pp 267–289
Berlind A (1977) Cellular dynamics in invertebrate neurosecretorysystems. Int Rev Cytol 49:171–251
Bishop CA, Wine JJ, O’Shea M (1984) Neuropeptide proctolin inpostural motoneurons of the crayfish. J Neurosci 4:2001–2004
Bishop CA, Wine JJ, Nagy F, O’Shea M (1987) Physiologicalconsequences of a peptide cotransmitter in a crayfish nerve-muscle preparation. J Neurosci 7:1769–1779
Buma P, Roubos EW (1986) Ultrastuctural demonstration of non-synaptic release sites in the central nervous system of the snailLymnaea stagnalis, the insect Periplaneta americana, and therat. Neuroscience 17:867–879
Burns ME, Augustine GJ (1995) Synaptic structure and function:dynamic organization yields architectural precision. Cell 83:187–194
Burnstock C (1986) The changing face of autonomic neurotrans-mission. Acta Physiol Scand 126:67–91
Castel M, Morris J, Belenky M (1996) Non-synaptic and dendriticexocytosis from dense-cored vesicles in the suprachiasmaticnucleus. Neuroreport 7:543–547
525

526
Claiborne BJ, Ayers J (1987) Functional anatomy and behavior.In: Selverston AI, Moulins M (eds) The crustacean stomato-gastric system. Springer, Berlin Heidelberg New York, pp9–29
Cuadras J (1989) Non-synaptic release from dense-cored vesiclesoccurs at all terminal types in crayfish neuropile. Brain Res477:332–335
Dickinson-Nelson A, Reese TS (1983) Structural changes duringtransmitter release at synapses in the frog sympathetic ganglia.J Neurosci 3:42–52
Factor JR (1995) The digestive system. In: Factor JR (ed) Biologyof the lobster Homarus americanus. Academic Press, SanDiego, pp 395–440
Golding DW, Bayraktaroglu E (1984) Exocytosis of secretorygranules – a probable mechanism for the release of neuromod-ulators in invertebrate neuropiles. Experientia 40:1277–1280
Govind CK, Lingle C (1987) Neuromuscular organization andpharmacology. In: Selverston AI, Moulins M (eds) The crusta-cean stomatogastric system. Springer, Berlin Heidelberg NewYork, pp 31–48
Govind CK, Atwood HL, Maynard DM (1975) Innervation andneuromuscular physiology of intrinsic foregut muscles in theblue crab and spiny lobster. J Comp Physiol 96:185–204
Heuser JE, Reese TS (1977) Structure of the synapse. In: KandelER (ed) Handbook of physiology. I. The nervous system.Amercian Physiological Society, Baltimore, pp 261–294
Hökfelt T, Lundberg JM, Skirboll L, Johansson O, Schultzberg M,Vincent SR (1982) Co-existence of classical transmitters andpeptides in neurones. In: Cuello AC (ed) Co-transmission.Macmillan Press, London, pp 77–119
King DG (1976) Organization of the crustacean neuropil. I. Pat-terns of synaptic connections in lobster stomatogastric gangli-on. J Neurocytol 5:207–237
King MJR, Atwood HL, Govind CK (1996) Structural features ofcrayfish phasic and tonic neuromuscular terminals. J CompNeurol 372:618–626
Marder E (1987) Neurotransmitters and neuromodulators. In:Selverston AI, Moulins M (eds) The crustacean stomatogastricsystem. Springer, Berlin Heidelberg New York, pp 263–300
Maynard DM, Dando MR (1974) The structure of the stomatogas-tric neuromuscular system in Callinectes sapidus, Homarusamericanus, and Panulirus argus(Decapoda Crustacea). Phi-los Trans R Soc Lond 268:161–220
Maynard DM, Selverston AI (1975) Organization of the stomato-gastric system of the spiny lobster. IV. The pyloric system. JComp Physiol 100:161–182
Mearow KM, Govind CK (1989) Stimulation-induced changes atcrayfish (Procambarus clarkii) neuromuscular terminals. CellTissue Res 256:119–123
Normann, TC (1976) Neurosecretion by exocytosis. Int Rev Cytol46:1–77
Pearce J, Govind CK, Shivers RR (1986) Intramembranous orga-nization of lobster excitatory neuromuscular synapses. J Neu-rocytol 15:241–252
Rheuben MB, Reese TS (1978) Three-dimensional structure andmembrane specialization of moth excitatory neuromuscularsynapse. J Ultrastruct Res 65:95–118
Schürmann F-W, Sandeman R, Sandeman D (1991) Dense-corevesicles and non-synaptic exocytosis in the central body of thecrayfish brain. Cell Tissue Res 265:493–501
Selverston AI, Moulins M (1987) The crustacean stomatogastricsystem. Springer, Berlin Heidelberg New York
Thurson-Klein A, Klein RL, Zhu P-C, Kong J-Y (1988) Differen-tial release of transmitters and neuropeptides co-stored in cen-tral and peripheral neurons. In: Zimmerman H (ed) Cellularand molecular basis of synaptic transmission. Springer, BerlinHeidelberg New York, pp 137–151