targeting virulence to prevent infection: to kill or not to kill?
TRANSCRIPT

THERAPEUTICSTRATEGIES
DRUG DISCOVERY
TODAY
Drug Discovery Today: Therapeutic Strategies Vol. 1, No. 4 2004
Editors-in-Chief
Raymond Baker – formerly University of Southampton, UK and Merck Sharp & Dohme, UK
Eliot Ohlstein – GlaxoSmithKline, USA
Infectious diseases
Targeting virulence to preventinfection: to kill or not to kill?Michael N. Alekshun1,*, Stuart B. Levy1,2
1Paratek Pharmaceuticals, 75 Kneeland Street, Boston, MA 02111, USA2The Center for Adaptation Genetics and Drug Resistance, and the Departments of Molecular Biology and Microbiology and of Medicine,
Tufts University School of Medicine, 136 Harrison Avenue, Boston, MA 02111, USA
Targeting components of the infectious process as a
means to prevent infection has long been considered as
an alternative to classic antimicrobial therapies.
Although no clinically used drugs have yet emerged
from these efforts, the dwindling supply of anti-infec-
tive treatment options within the physician’s arma-
mentarium has stoked a renewed interest in the
identification and development of novel strategies to
prevent infection.
*Corresponding author: (M.N. Alekshun) [email protected]
1740-6773/$ � 2004 Elsevier Ltd. All rights reserved. DOI: 10.1016/j.ddstr.2004.10.006
Section Editor:Gary Woodnutt – Diversa Corp., San Diego, CA, USA
There are multiple virulence factors that are required to initiate andmaintain infection in the host. As many of these are external to the
infecting cell, they provide clear target opportunities that wouldcircumvent the need for cellular penetration. However, by definition,
compounds that inhibit these targets would not kill the organism andthus many of the screening processes currently used will not be
applicable. The difficulties associated with progressing targets of thistype are discussed with some key examples of areas that might become
clinical candidates in the near future. The suggestion that virulenceinhibiting compounds might be less likely to generate resistance is
intriguing but unproven. The lack of a clear path through regulatorybodies and the inability to test activity by traditional, and well accepted,
clinical microbiology add to the complexity of achieving success withthese novel interventions. However, as the clinical options for
treatment of infectious disease are eroded by resistance the time mightbe ripe for exploiting these strategies.
an immeasurable benefit to human health, but the wide-
Introduction
The use of antibiotics to treat infectious diseases has provided
spread emergence of bacteria that are resistant to these ther-
apeutics has raised grave concern about the future of an
antimicrobial approach. The pharmaceutical industry has,
for decades, responded by synthesizing derivatives of existing
chemical classes to obtain drugs with ‘expanded’ spectrums
of antibacterial activity. Besides derivatives of other antibio-
tics, only two novel agents (a cyclic lipopeptide and an
oxazolidinone) and a new streptogramin combination have
reached clinical availability in the past two decades [1].
Current development efforts have made little headway in
resolving the resistance problem, and large pharmaceutical
companies are exiting the field.
As an alternative to antibiotics, targeting of VIRULENCE FAC-
TORS (see Glossary) has been viewed cautiously, but repeat-
edly. For this approach to be successful, the novel agents need
to have a reduced propensity to select for resistance, as a
consequence of lacking intrinsic antibacterial activity, and
must act on bacterial-specific targets. Although no one agent
boasts all of these qualifications, recent data suggest that the
time might be ripe for fully exploiting this new therapeutic
paradigm.
Here, we discuss small molecules or proteins that target
gene products involved in infection or VIRULENCE (see Glos-
sary), and those that use other strategies not involving
growth inhibition. Vaccines against specific microbial anti-
gens or toxins are not addressed.
www.drugdiscoverytoday.com 483

Drug Discovery Today: Therapeutic Strategies | Infectious diseases Vol. 1, No. 4 2004
Glossary
Pathogens: organisms that are capable of causing disease, including
classical pathogens, opportunists and commensals.
Small-molecule inhibitors: low molecular weight organic chemicals
that target and inhibit the function of a virulence factor.
Virulence: the capacity of a bacterium to cause disease.
Virulence factors: traits expressed by bacteria that aid the infectious
process. Might encompass an antibiotic-resistance determinant, an
adhesin, an invasion, a toxin, biofilm formation and so on.
‘Broad-spectrum’ approaches
Strategies that target transcription factors
Given that infection is regulated primarily at the level of
transcription, SMALL-MOLECULE INHIBITORS (see Glossary) of bac-
terial transcription factors can be expected to exhibit pleio-
tropic effects on the virulence phenotype. Proteins within the
AraC (MarA) and MarR protein families are attractive targets
for these efforts because they regulate virulence in many
medically important Gram-negative [2] and Gram-positive
[3] PATHOGENS (see Glossary). With regard to the former, inac-
tivation of the gene specifying an AraC (MarA) family
member in Pseudomonas aeruginosa [4], Vibrio cholerae [5]
and Yersinia pestis [6] renders these organisms avirulent
(Table 1).
Naturally occurring small molecule modulators are known
to affect many AraC (MarA) family members, for example,
Escherichia coli AraC (arabinose) and Rob (bile salts, fatty acids
and dipyridyl) and V. cholerae ToxT (TcpN) (bile salts). Thus, it
is reasonable to envisage additional medicinal chemistry
efforts that would exploit these natural scaffolds. Using a
murine model of ascending pyelonephritis (urinary tract
infection [UTI]), we have found that removal of MarA and
its paralogs from multi-drug resistant uropathogenic E. coli
Table 1. Effect(s) of deleting a gene(s) specifying an AraC (Mar
Organism Protein (alternative designation) Phenotype
Escherichia coli MarA, SoxS, Rob Reduced kid
Proteus mirabilis UreR Reduced co
and urine in
Yersinia pestis LcrF Reduced let
Pseudomonas aeruginosa ExsA Reduced let
Vibrio cholerae ToxT Reduced let
Staphylococcus aureus Uncharacterized Reduced ab
Streptococcus pneumoniae rr09 (SPr0578 or SP0661) Reduced let
SP1433 Reduced CI
Mycobacterium tuberculosis Rv1395 Reduced lun
Rv1931c Reduced lun
Abbreviations: CI, competitive index; UTI, urinary tract infection.a In the cochallenge experiment, mice are infected with an �1:1 ratio of wild-type and mutanb The competitive index represents the ratio of wild-type to mutant bacteria recovered from a h
relative to the wild-type organism the mutant is less virulent.
484 www.drugdiscoverytoday.com
dramatically prevented these organisms from colonizing the
mouse kidney (M.N. Alekshun et al., unpublished). These
results prompted efforts to identify small molecule AraC
(MarA) inhibitors. Many of these newly identified agents
have activity in vitro against proteins from E. coli, Salmonella
enterica serovar Typhimurium, Proteus vulgaris and P. aerugi-
nosa and many exhibited infection prevention in the murine
UTI model (M.N. Alekshun et al., unpublished).
Members of the SarA protein family in Staphylococcus aureus
are MarR orthologs. Recent data have indicated a beneficial
effect of acetylsalicylate in an experimental rabbit model of S.
aureus endocarditis [7]. More current studies have shown that
salicylate attenuated virulence in both laboratory and clini-
cally derived S. aureus isolates by negatively affecting the
interaction of the pathogen with fibronectin and fibrinogen
and exotoxin production in vitro [8]. This effect was depen-
dent on the presence of SarA. Thus, the modulating effects of
salicylate on the virulence of S. aureus in vitro correlated with a
positive outcome in vivo.
Strategies that target quorum-sensing systems
Both Gram-negative and Gram-positive bacteria, including P.
aeruginosa, E. coli, S. aureus and the streptococci, use quorum
sensing (QS) to regulate the expression of many virulence
factors [9]. In Gram-negative bacteria, small molecules called
acylated homoserine lactones (AHLs) act as the QS signals,
whereas small peptides fill this role in Gram-positive organ-
isms. Some virulence factors regulated by QS include biofilm
formation, the development of competence, secretion of
exotoxins and enzymes and sporulation. There are several
recent developments that have documented the efficacy of
AHL antagonists in animal models of infection [10–12].
Hentzer et al. [11] and Wu et al. [10] used a murine model of
P. aeruginosa pulmonary infection to show that treatment
A) protein in an animal model(s) of infection
of mutant organisms in infection models in vivo Refs
ney colonization in UTI model M.N. Alekshun
et al., unpublished
lonization of kidneys, bladder
a cochallenge UTI modela[42]
hality and CI in a model of bacteremiab [6]
hality in a pneumonia model [4]
hality following orogastric infection [43]
scess formation and systemic dissemination [44]
hality and bacteremia [45,46]b [47]
g burden in a model of bacteremia [48]
g and spleen burden [49]
t bacteria. Total CFU/g of tissue is then determined for each organism.
ost infected with a mixture of both organisms. A reduced competitive index indicates that

Vol. 1, No. 4 2004 Drug Discovery Today: Therapeutic Strategies | Infectious diseases
with synthetic furanones (initially derived from the marine
alga Delisea pulchra) resulted in a three-log decrease in the
colony forming units (CFU) per gram of lung tissue. The
pathologic response of the lungs to the infection was less
severe and fewer abscesses were found in infected mice trea-
ted with these agents [10].
Initial studies with naturally occurring thiolactone-con-
taining peptides demonstrated infection prevention activity
in a subcutaneous S. aureus abscess mouse model of infection
[13]. More recently, Dell’Acqua et al. [12] used a vascular-graft
rat model of S. aureus and Staphylococcus epidermidis (includ-
ing both susceptible and multi-drug resistant strains) infec-
tion to show that the delivery of a QS-peptide antagonist,
either by local or parenteral administration, reduced signifi-
cantly the bacterial load on an implanted Dacron graft. The
combination of local and parenteral treatments was success-
ful in effecting complete protection in this model [12].
‘Narrow-spectrum’ approaches
Strategies that target toxins
The selective targeting of bacterial toxins has been viewed as a
precise alternative to classic antibiotic therapy and has
received renewed interest in light of the anthrax attacks in
the US in 2001. Two groups have demonstrated efficacy of
peptide-based inhibitors of anthrax toxin in a rat model of
toxicity (intoxication) [14,15]. In this model, rats are given a
mixture of the anthrax protective antigen and lethal factor
[PA and LF; the combination of which is referred to as lethal
toxin (LeTx)] and death of the host occurs within hours after
inoculation. More recently, three other groups have identi-
fied small-molecule inhibitors of the anthrax LF. Two of these
groups have solved the 3D structures of toxin-inhibitor co-
crystal complexes thereby paving the way for approaches in
structure based drug design (SBDD; see Ref. [16] and refer-
ences therein). Although some of these compounds offer
protection against LF-mediated cytotoxicity in vitro, only
one has shown small-molecule efficacy in the LeTx rat model
[17]. In this particular experiment, administration of the
small-molecule inhibitor in conjunction with, or separate
from, the LeTx demonstrated efficacy in vivo [17].
Like many Gram-negative pathogens, Y. pestis uses a type-
III secretion system (TTSS) to deliver host effector proteins
(Yops) into mammalian cells during infection. YopH is a
potent tyrosine phosphatase that interferes with phagocyto-
sis; Y. pestis mutants lacking yopH are avirulent. Liang et al.
[18] screened a small library of commercially available car-
boxylic acids and identified a potent YopH inhibitor that,
relative to other mammalian tyrosine phosphatases, exhib-
ited specificity for the bacterial protein. A recent YopH-inhi-
bitor co-crystal structure should facilitate SDBB in this area
[19] but the activity in vivo of these inhibitors remains to be
determined.
Strategies that target cell-surface modification
Using bacteriophage as an alternative anti-infective thera-
peutic strategy has received renewed interest but the use of
live bacteriophage, or phage-derived lysins, would ultimately
result in death (lysis) of the infecting organism and thereby
not fall within the anti-infection paradigm. Recent experi-
ments, however, have used a phage-derived endosialidase
(endoE), which specifically and selectively hydrolyzes the
E. coli K1 capsular polysaccharide and so alters pathogen
virulence without killing the organism [20]. Intraperitoneal
administration of endoE in a neonatal rat model of E. coli
bacteremia results in almost complete protection from bac-
teremia and blood samples were pathogen-free within 24 h
following the initiation of therapy [20].
Strategies that target surface proteins: pili, adhesins,
chaperons and sortase
E. coli type-I and type-P pili are virulence factors that have
important roles in cystitis and pyelonephritis, respectively.
Both possess domains (adhesins) that are responsible for the
binding to polysaccharide receptors that are located on the
surfaces of host cells. The adhesins are assembled onto the pili
by chaperones and efforts to identify small-molecule-chaper-
one inhibitors (pilicides) have been described using a cha-
peron–adhesin co-crystal structure [21]. These compounds
(bicyclic b-lactams, 2-pyridones and N-substituted amino
acids [22]) appear to effectively dissociate the chaperone–
adhesin complex in vitro by targeting the active site of the
chaperone [21]. Efficacy in vivo, however, has not been deter-
mined. Carbohydrate-based compounds that target the active
site of the adhesin have also been described [23,24]. These
agents exhibit modest activity in vitro, as measured using both
biophysical and hemagglutination assays [23,24]. The effi-
cacy of these particular compounds in an animal model of
UTI has not been shown, but an effect in vivo has been shown
with glycolipid analogs [25].
The Streptococcus mutans SpaA is an adhesin that functions
in the colonization of the oral cavity and the development of
dental caries. Recent studies have identified a synthetic pep-
tide (p1025) that inhibited binding of SpaA to salivary agglu-
tinin in vitro [26]. The efficacy of p1025 in human volunteers
was then tested in a small double-blind placebo controlled
trial [26]. The oral cavity of all volunteers was first deconta-
minated using chlorhexidine gluconate and, subsequently,
four patients were treated with p1025, four with a buffer
placebo and three with an inactive peptide control [26].
Bacterial recolonization was monitored for a period of 120
days and, although the presence of S. mutans was evident in
patients receiving either the placebo (at 21 days) or the
inactive peptide control (at 58 days), 75% of the individuals
in the p1025 treatment group remained free of S. mutans [26].
Importantly, recolonization of Actinomyces naeslundii, a
www.drugdiscoverytoday.com 485

Drug Discovery Today: Therapeutic Strategies | Infectious diseases Vol. 1, No. 4 2004
Gram-positive oral commensal, was unaffected by the control
peptide and the therapeutic peptide [26].
The sortase enzymes (SrtA and SrtB) are cysteine protease-
transpeptidase Gram-positive virulence factors that are
responsible for the covalent attachment of surface-exposed
proteins, including protein A, fibronectin and collagen-bind-
ing proteins. Compared with wild-type strains, S. aureus that
lack srtA are less lethal, produce a reduced inflammatory
response, and are attenuated in mouse kidney and rat endo-
carditis infection models [27,28]. Peptide- [29], carbohydrate-
[30] and small-molecule- [31–33] based SrtA inhibitors have
been described. Although compounds that have evolved
from these efforts possess activity in vitro, their efficacy in
vivo awaits experimental verification.
Probiotics
Bacterial interference by competitive exclusion
Probiotics [beneficial (commensal) bacteria, for example,
Lactobacillus spp., Bifidobacterium spp. and E. coli Nissle
1917] have long been touted as an alterative to pharmaco-
logic intervention for the treatment of many diseases includ-
ing inflammatory bowel disease, UTI, wound infections,
atopic eczema, vaginitis, arthritis and travellers’ diarrhea.
Probiotics are most commonly administered by oral con-
sumption and data from many recent randomized, double-
blind and comparative or placebo-controlled clinical trials are
very encouraging [34].
From the clinical perspective, Lactobacillus rhamnosus strain
GG (LGG) is probably the most extensively studied probiotic
[35]. LGG is ‘non-pathogenic’ and the beneficial effects of this
and other probiotic organisms are multifactorial. LGG adheres
strongly to tissue culture cells in vitro, and colonizes the human
gastrointestinal tract. Experiments in vivo suggest that it com-
petes with the normal flora and pathogenic bacteria for host
colonization and has positive effects on intestinal and sys-
temic immunity [35]. Notably, LGG produce antimicrobial
compounds that exhibit a broad spectrum of activity in vitro
[36], but their possible role in the multifaceted competitive
exclusion activity of the organism is unclear.
In a fascinating set of recent experiments, subcutaneous
delivery of Lactobacillus salivarius 118 to interleukin-10 (IL-
10) deficient mice had a statistically significant anti-inflam-
matory effect on the development of colitis and arthritis in
this species [37]. Lactobacillus fermentum RC-14 has been
shown to prevent abscess formation in a rat model of surgical
implant S. aureus infection [38]. The protective effect was
subsequently attributed to a secreted biosurfactant lacking
intrinsic antimicrobial activity and containing many col-
lagen-binding proteins [38].
Agents in clinical development
The vast majority of anti-virulence therapies are yet to be
studied in the clinical setting, but there is hope that one will
486 www.drugdiscoverytoday.com
make it. Clostridium difficile is an important cause of
nosocomial diarrhea and the treatment of infected patients
usually requires therapy with either vancomycin or metro-
nidazole, two drugs that have profound effects on the intest-
inal flora. GT160-246 (Tolevamer1) is a high molecular
weight anionic polymer that lacks antimicrobial activity
and targets the A and B toxins of C. difficile [39]. This drug
has recently completed a randomized, double-blind phase-II
clinical trial in which the efficacy of two separate doses of
Tolevamer1 was compared with oral vancomycin. Interest-
ingly, the time to resolution of diarrhea and rate of recurrence
was similar for both therapies, that is, one antibacterial and
the other not. A larger phase-III clinical trial is currently being
planned.
Conclusions
The concept of targeting virulence as a new therapeutic
strategy to prevent (or treat) infection has relied on the
premise that affecting the ability of an organism to cause
disease, for example, preventing the function of a virulence
factor(s), would render it non-infectious. The approach, in
theory, seems easily achievable but in reality there are several
outstanding issues.
Because virulence factors are generally required for growth
in vivo, the in vitro whole-cell screens that would normally be
employed in classical antibiotic discovery programs are of
limited use. The adoption of this new preventative (or ther-
apeutic) concept, therefore, requires a paradigm shift in the
research and development (R&D) infrastructure of an orga-
nization. Standard minimal inhibitory concentrations (MIC)
determinations will need to be replaced with other assays that
are predictive of virulence factor expression (e.g. toxin excre-
tion and cytotoxicity towards tissue culture cells). Also, a
change in the mindset of the regulatory authorities (e.g. the
Food and Drug Administration and the European Medicines
Agency) will be needed. These factors ultimately affect the
economic considerations that drive modern-day R&D efforts
and, collectively, are major reasons why the discovery and
development of these new agents has been so difficult to
achieve.
Many of the approaches described herein have relied on
peptides to demonstrate feasibility. Although these agents are
suitable from the basic science perspective, the production
costs, stability in vivo, pharmacokinetic considerations, for
example, of small-peptide-derived therapeutics it is unlikely
to garner enthusiastic support for real-life treatment scenar-
ios. Fortunately, many researchers are moving in the
direction of small-molecule therapeutics for anthrax toxin
(Table 2).
The development of resistance to these new approaches
has also been the subject of discussion. History has taught
that the overuse (and misuse) of any (small-molecule) anti-
infective will ultimately lead to resistance [40]. Because the
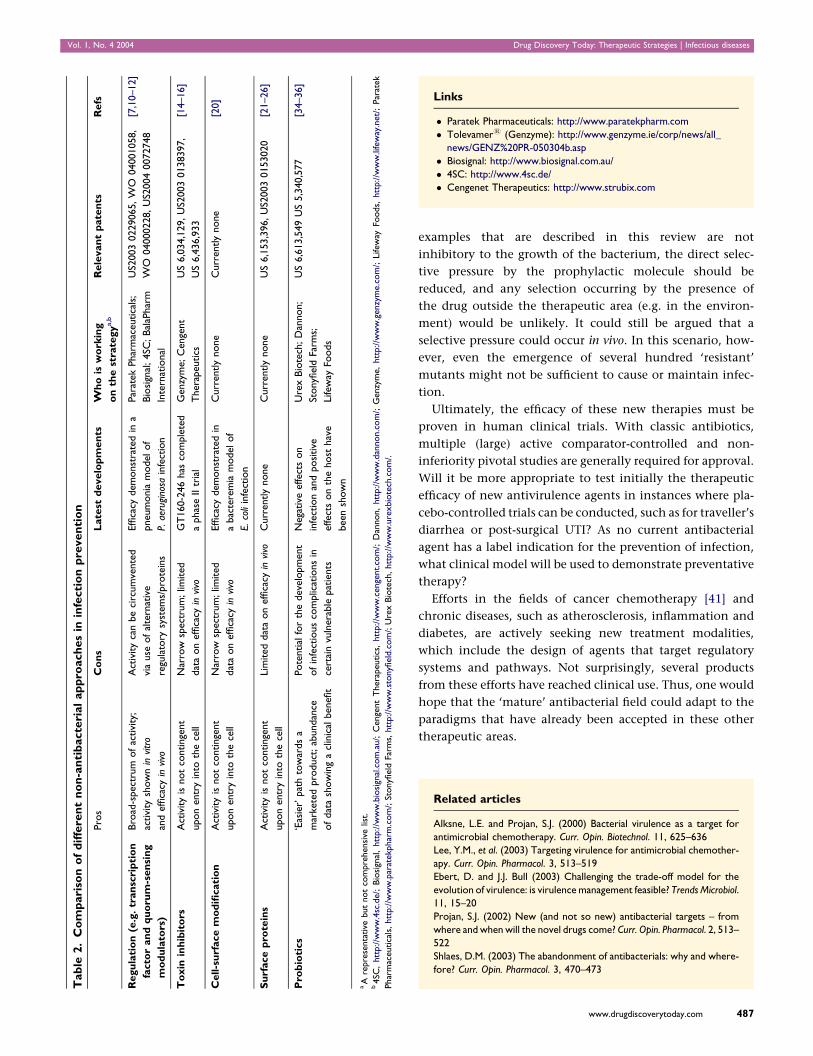
Vol. 1, No. 4 2004 Drug Discovery Today: Therapeutic Strategies | Infectious diseases
Tab
le2.
Co
mp
ari
son
of
dif
fere
nt
no
n-a
nti
bacte
rial
ap
pro
ach
es
inin
fecti
on
pre
ven
tio
n
Pro
sC
on
sL
ate
std
evelo
pm
en
tsW
ho
isw
ork
ing
on
the
stra
tegy
a,b
Rele
van
tp
ate
nts
Refs
Regu
lati
on
(e.g
.tr
an
scri
pti
on
facto
ran
dq
uo
rum
-sen
sin
g
mo
du
lato
rs)
Bro
ad-s
pec
trum
of
activi
ty;
activi
tysh
ow
nin
vitr
o
and
effica
cyin
vivo
Act
ivity
can
be
circ
um
vente
d
via
use
of
alte
rnat
ive
regu
lato
rysy
stem
s/pro
tein
s
Effi
cacy
dem
onst
rate
din
a
pneu
monia
model
of
P.ae
rugi
nosa
infe
ctio
n
Par
atek
Pha
rmac
eutica
ls;
Bio
sign
al;4SC
;B
alaP
har
m
Inte
rnat
ional
US2
003
0229065,
WO
04001
058,
WO
04000
228,
US2
004
00727
48
[7,1
0–12
]
To
xin
inh
ibit
ors
Act
ivity
isnot
continge
nt
upon
entr
yin
toth
ece
ll
Nar
row
spec
trum
;lim
ited
dat
aon
effica
cyin
vivo
GT
160-2
46
has
com
ple
ted
aphas
eII
tria
l
Gen
zym
e;C
enge
nt
Ther
apeu
tics
US
6,0
34,1
29,U
S2003
01383
97,
US
6,4
36,9
33
[14–16
]
Cell-s
urf
ace
mo
difi
cati
on
Act
ivity
isnot
continge
nt
upon
entr
yin
toth
ece
ll
Nar
row
spec
trum
;lim
ited
dat
aon
effica
cyin
vivo
Effi
cacy
dem
onst
rate
din
abac
tere
mia
model
of
E.co
liin
fect
ion
Curr
ently
none
Curr
ently
none
[20]
Su
rface
pro
tein
sA
ctiv
ity
isnot
continge
nt
upon
entr
yin
toth
ece
ll
Lim
ited
dat
aon
effica
cyin
vivo
Curr
ently
none
Curr
ently
none
US
6,1
53,3
96,U
S2003
01530
20
[21–26
]
Pro
bio
tics
‘Eas
ier’
pat
hto
war
ds
a
mar
kete
dpro
duct
;ab
undan
ce
of
dat
ash
ow
ing
acl
inic
alben
efit
Pote
ntial
for
the
dev
elopm
ent
of
infe
ctio
us
com
plic
atio
ns
in
cert
ain
vuln
erab
lepat
ients
Neg
ativ
eef
fect
son
infe
ctio
nan
dposi
tive
effe
cts
on
the
host
hav
e
bee
nsh
ow
n
Ure
xB
iote
ch;D
anno
n;
Stony
fiel
dFa
rms;
Life
way
Foods
US
6,6
13,5
49
US
5,3
40,5
77[3
4–36
]
aA
repre
senta
tive
but
not
com
pre
hen
sive
list.
b4SC
,htt
p://w
ww
.4sc
.de/
;B
iosi
gnal
,htt
p://w
ww
.bio
sign
al.c
om
.au/;
Cen
gent
Ther
apeu
tics
,htt
p://w
ww
.cen
gent.co
m/;
Dan
non,
htt
p://w
ww
.dan
non.c
om
/;G
enzy
me,
htt
p://w
ww
.gen
zym
e.co
m/;
Life
way
Foods,
htt
p://w
ww
.life
way
.net
/;Par
atek
Phar
mac
eutica
ls,
htt
p://w
ww
.par
atek
phar
m.c
om
/;St
onyfi
eld
Farm
s,htt
p://w
ww
.sto
nyfi
eld.c
om
/;U
rex
Bio
tech
,htt
p://w
ww
.ure
xbio
tech
.com
/.
Links
� Paratek Pharmaceuticals: http://www.paratekpharm.com
� Tolevamer1 (Genzyme): http://www.genzyme.ie/corp/news/all_
news/GENZ%20PR-050304b.asp
� Biosignal: http://www.biosignal.com.au/
� 4SC: http://www.4sc.de/
� Cengenet Therapeutics: http://www.strubix.com
examples that are described in this review are not
inhibitory to the growth of the bacterium, the direct selec-
tive pressure by the prophylactic molecule should be
reduced, and any selection occurring by the presence of
the drug outside the therapeutic area (e.g. in the environ-
ment) would be unlikely. It could still be argued that a
selective pressure could occur in vivo. In this scenario, how-
ever, even the emergence of several hundred ‘resistant’
mutants might not be sufficient to cause or maintain infec-
tion.
Ultimately, the efficacy of these new therapies must be
proven in human clinical trials. With classic antibiotics,
multiple (large) active comparator-controlled and non-
inferiority pivotal studies are generally required for approval.
Will it be more appropriate to test initially the therapeutic
efficacy of new antivirulence agents in instances where pla-
cebo-controlled trials can be conducted, such as for traveller’s
diarrhea or post-surgical UTI? As no current antibacterial
agent has a label indication for the prevention of infection,
what clinical model will be used to demonstrate preventative
therapy?
Efforts in the fields of cancer chemotherapy [41] and
chronic diseases, such as atherosclerosis, inflammation and
diabetes, are actively seeking new treatment modalities,
which include the design of agents that target regulatory
systems and pathways. Not surprisingly, several products
from these efforts have reached clinical use. Thus, one would
hope that the ‘mature’ antibacterial field could adapt to the
paradigms that have already been accepted in these other
therapeutic areas.
Related articles
Alksne, L.E. and Projan, S.J. (2000) Bacterial virulence as a target for
antimicrobial chemotherapy. Curr. Opin. Biotechnol. 11, 625–636
Lee, Y.M., et al. (2003) Targeting virulence for antimicrobial chemother-
apy. Curr. Opin. Pharmacol. 3, 513–519
Ebert, D. and J.J. Bull (2003) Challenging the trade-off model for the
evolution of virulence: is virulence management feasible? Trends Microbiol.
11, 15–20
Projan, S.J. (2002) New (and not so new) antibacterial targets – from
where and when will the novel drugs come? Curr. Opin. Pharmacol. 2, 513–
522
Shlaes, D.M. (2003) The abandonment of antibacterials: why and where-
fore? Curr. Opin. Pharmacol. 3, 470–473
www.drugdiscoverytoday.com 487

Drug Discovery Today: Therapeutic Strategies | Infectious diseases Vol. 1, No. 4 2004
Outstanding questions
� Will the stand-alone efficacy of these novel agents be sufficient, or will
they need to be used in conjunction with antibiotics?
� If approved, could these agents be used in areas in which infection is
thought to be linked to chronic disease, for example, atherosclerosis,
Alzheimer’s disease and ulcer disease?
� What effect(s) would this new therapeutic approach have on the
colonization of protective commensal organisms?
� Will small-molecule inhibitors of virulence factors work in immuno-
compromised patients?
References1 Shlaes, D.M. (2003) The abandonment of antibacterials: why and where-
fore? Curr. Opin. Pharmacol. 3, 470–473
2 Finlay, B.B. and Falkow, S. (1997) Common themes in microbial patho-
genicity revisited. Microbiol. Mol. Biol. Rev. 61, 136–169
3 Cheung, A.L. and Zhang, G. (2002) Global regulation of virulence
determinants in Staphylococcus aureus by the SarA protein family. Front
Biosci. 7, d1825–d1842
4 Hauser, A.R. et al. (1998) PepA, a secreted protein of Pseudomonas
aeruginosa, is necessary for cytotoxicity and virulence. Mol. Microbiol.
27, 807–818
5 Skorupski, K. and Taylor, R.K. (1997) Control of the ToxR virulence
regulon in Vibrio cholerae by environmental stimuli. Mol. Microbiol. 25,
1003–1009
6 Flashner, Y. et al. (2004) Generation of Yersinia pestis attenuated strains by
signature-tagged mutagenesis in search of novel vaccine candidates. Infect.
Immun. 72, 908–915
7 Kupferwasser, L.I. et al. (1999) Acetylsalicylic acid reduces
vegetation bacterial density, hematogenous bacterial dissemination, and
frequency of embolic events in experimental Staphylococcus aureus
endocarditis through antiplatelet and antibacterial effects. Circulation
99, 2791–2797
8 Kupferwasser, L.I. et al. (2003) Salicylic acid attenuates virulence in
endovascular infections by targeting global regulatory pathways in Sta-
phylococcus aureus. J. Clin. Invest. 112, 222–233
9 Miller, M.B. and Bassler, B.L. (2001) Quorum sensing in bacteria. Annu.
Rev. Microbiol. 55, 165–199
10 Wu, H. et al. (2004) Synthetic furanones inhibit quorum-sensing and
enhance bacterial clearance in Pseudomonas aeruginosa lung infection
in mice. J. Antimicrob. Chemother. 53, 1054–1061
11 Hentzer, M. et al. (2003) Attenuation of Pseudomonas aeruginosa viru-
lence by quorum sensing inhibitors. EMBO J. 22, 3803–3815
12 Dell’Acqua, G. et al. (2004) Suppression of drug-resistant Staphylococcal
infections by the quorum-sensing inhibitor RNAIII-inhibiting peptide. J.
Infect. Dis. 190, 318–320
13 Mayville, P. et al. (1999) Structure-activity analysis of synthetic autoindu-
cing thiolactone peptides from Staphylococcus aureus responsible for
virulence. Proc. Natl. Acad. Sci. USA 96, 1218–1223
14 Mourez, M. et al. (2001) Designing a polyvalent inhibitor of anthrax toxin.
Nat. Biotechnol. 19, 958–961
15 Sarac, M.S. et al. (2004) Protection against anthrax toxemia by hexa-D-
arginine in vitro and in vivo. Infect. Immun. 72, 602–605
16 Montecucco, C. et al. (2004) Stop the killer: how to inhibit the
anthrax lethal factor metalloprotease. Trends Biochem. Sci. 29, 282–285
17 Dell’Aica, I. et al. (2004) Potent inhibitors of anthrax lethal factor from
green tea. EMBO Rep. 5, 418–422
18 Liang, F. et al. (2003) Aurintricarboxylic acid blocks in vitro and in vivo
activity of YopH, an essential virulent factor of Yersinia pestis, the agent of
plague. J. Biol. Chem. 278, 41734–41741
19 Sun, J.P. et al. (2003) Crystal structure of the Yersinia protein-tyrosine
phosphatase YopH complexed with a specific small molecule inhibitor. J.
Biol. Chem. 278, 33392–33399
488 www.drugdiscoverytoday.com
20 Mushtaq, N. et al. (2004) Prevention and cure of systemic Escherichia coli
K1 infection by modification of the bacterial phenotype. Antimicrob.
Agents Chemother. 48, 1503–1508
21 Svensson, A. et al. (2001) Design and evaluation of pilicides: potential
novel antibacterial agents directed against uropathogenic Escherichia coli.
Chembiochem 2, 915–918
22 Lee, Y.M. et al. (2003) Targeting virulence for antimicrobial chemother-
apy. Curr. Opin. Pharmacol. 3, 513–519
23 Larsson, A. et al. (2003) Quantitative studies of the binding of the class II
PapG adhesin from uropathogenic Escherichia coli to oligosaccharides.
Bioorg. Med. Chem. 11, 2255–2261
24 Ohlsson, J. et al. (2002) Discovery of potent inhibitors of PapG adhesins
from uropathogenic Escherichia coli through synthesis and evaluation of
galabiose derivatives. Chembiochem 3, 772–779
25 Eden, C.S. et al. (1982) Inhibition of experimental ascending urinary tract
infection by an epithelial cell-surface receptor analogue. Nature 298, 560–
562
26 Kelly, C.G. et al. (1999) A synthetic peptide adhesion epitope as a novel
antimicrobial agent. Nat. Biotechnol. 17, 42–47
27 Weiss, W.J. et al. (2004) Effect of srtA and srtB gene expression on the
virulence of Staphylococcus aureus in animal models of infection. J.
Antimicrob. Chemother. 53, 480–486
28 Mazmanian, S.K. et al. (2000) Staphylococcus aureus sortase mutants
defective in the display of surface proteins and in the pathogenesis of
animal infections. Proc. Natl. Acad. Sci. USA 97, 5510–5515
29 Scott, C.J. et al. (2002) Irreversible inhibition of the bacterial cysteine
protease-transpeptidase sortase (SrtA) by substrate-derived affinity labels.
Biochem. J. 366, 953–958
30 Kim, S.H. et al. (2003) Inhibition of sortase, a bacterial surface protein
anchoring transpeptidase, by beta-sitosterol-3-O-glucopyranoside from
Fritillaria verticillata. Biosci. Biotechnol. Biochem. 67, 2477–2479
31 Oh, K.B. et al. (2004) Discovery of diarylacrylonitriles as a novel series of
small molecule sortase A inhibitors. J. Med. Chem. 47, 2418–2421
32 Kim, S.H. et al. (2004) Inhibition of the bacterial surface protein anchoring
transpeptidase sortase by isoquinoline alkaloids. Biosci. Biotechnol. Bio-
chem. 68, 421–424
33 Kim, S.W. et al. (2002) Inhibition of the bacterial surface protein anchoring
transpeptidase sortase by medicinal plants. Biosci. Biotechnol. Biochem.
66, 2751–2754
34 Sartor, R.B. (2004) Therapeutic manipulation of the enteric microflora in
inflammatory bowel diseases: antibiotics, probiotics, and prebiotics. Gas-
troenterology 126, 1620–1633
35 Alvarez-Olmos, M.I. and Oberhelman, R.A. (2001) Probiotic agents and
infectious diseases: a modern perspective on a traditional therapy. Clin.
Infect. Dis. 32, 1567–1576
36 Silva, M. et al. (1987) Antimicrobial substance from a human Lactoba-
cillus strain. Antimicrob. Agents Chemother. 31, 1231–1233
37 Sheil, B. et al. (2004) Is the mucosal route of administration essential for
probiotic function? Subcutaneous administration is associated with
attenuation of murine colitis and arthritis Gut 53, 694–700
38 Gan, B.S. et al. (2002) Lactobacillus fermentum RC-14 inhibits Staphy-
lococcus aureus infection of surgical implants in rats. J. Infect. Dis. 185,
1369–1372
39 Kurtz, C.B. et al. (2001) GT160-246, a toxin binding polymer for treatment
of Clostridium difficile colitis. Antimicrob. Agents Chemother. 45, 2340–
2347
40 Levy, S.B. (1998) The challenge of antibiotic resistance. Sci. Am. 278, 46–
53
41 Darnell, J.E. Jr. (2002) Transcription factors as targets for cancer therapy.
Nat. Rev. Cancer 2, 740–749
42 Dattelbaum, J.D. et al. (2003) UreR, the transcriptional activator of the
Proteus mirabilis urease gene cluster, is required for urease activity and
virulence in experimental urinary tract infections. Infect. Immun. 71, 1026–
1030
43 Champion, G.A. et al. (1997) A branch in the ToxR regulatory cascade of
Vibrio cholerae revealed by characterization of toxT mutant strains. Mol.
Microbiol. 23, 323–331

Vol. 1, No. 4 2004 Drug Discovery Today: Therapeutic Strategies | Infectious diseases
44 Coulter, S.N. et al. (1998) Staphylococcus aureus genetic loci impacting
growth and survival in multiple infection environments. Mol. Microbiol.
30, 393–404
45 Blue, C.E. and Mitchell, T.J. (2003) Contribution of a response regulator to
the virulence of Streptococcus pneumoniae is strain dependent. Infect.
Immun. 71, 4405–4413
46 Throup, J.P. et al. (2000) A genomic analysis of two-component signal
transduction in Streptococcus pneumoniae. Mol. Microbiol. 35, 566–576
47 Hava, D. and Camilli, A. (2002) Large-scale identificationof serotype 4
Streptococcus pneumoniae virulence factors. Mol. Microbiol. 45, 1389–1406
48 Camacho, L.R. et al. (1999) Identification of a virulence gene cluster of
Mycobacterium tuberculosis by signature-tagged transposon mutagenesis.
Mol. Microbiol. 34, 257–267
49 Frota, C.C. et al. (2004) The AraC family transcriptional regulator
Rv1931c plays a role in the virulence of Mycobacterium tuberculosis.
Infect. Immun. 72, 5483–5486
www.drugdiscoverytoday.com 489