the chemistry val. 24, 25, 14806-14810,1982 in u. s ... · val. 257. no. 24, issue of december 25,...
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY
Prmted in U. S. A. Val. 257. NO. 24, Issue of December 25, pp. 14806-14810,1982
Inhibition of HeLa Cell Protein Synthesis Following Poliovirus Infection Correlates with the Proteolysis of a 220,000-dalton Polypeptide Associated with Eucaryotic Initiation Factor 3 and a Cap Binding Protein Complex*
(Received for publication, August 16, 1982)
Diane Etchison$, Susan C. Milburn, Isaac Ederys, Nahum Sonenberggl, and John W. B. Hershey11 From the Department of Biological Chemistry, School of Medicine, University of California, Davis, California 95616 and the §Department of Biochemistry, McGill University, Montreal, PQ, Canada H3G 1 Y6
Following poliovirus infection of HeLa cells, the syn- thesis of cellular proteins is inhibited but translation of poliovirus mRNA proceeds. The defect in the recogni- tion of host cell mRNA may be due to a change in a cap recognition complex which, when added to an infected cell lysate, restores the ability to translate capped mRNAs. We employed immunoblotting techniques to examine initiation factors in crude lysates from unin- fected and poliovirus-infected HeLa cells. Using an an- tiserum against eucaryotic initiation factor 3, we de- tected an antigen of approximate molecular weight 220,000 in uninfected cell lysates but not in infected cell lysates. Antigenically related polypeptides of 100,000 to 130,000 daltons, presumably degradation products, were detected in the infected cell lysate, The time course for degradation of the 220,000-dalton poly- peptide correlates with that for inhibition of cellular protein synthesis in vivo. A portion of the population of 220,000-dalton polypeptides apparently associates with initiation factor eIF3 but is readily dissociated in buffers containing high salt. Affinity-purified antibod- ies against the polypeptide recognize a protein of the same size in a purified preparation of a cap binding protein complex obtained by cap-affinity chromatog- raphy. We postulate that the 220,000-dalton poly- peptide is an essential component of the cap recognition complex and that its degradation in poliovirus-infected cells results in the inhibition of host cell translation. These results are the first demonstration of a specific structural defect in an initiation factor resulting from poliovirus infection.
Poliovirus infection of HeLa cells results in inhibition of the translation of capped mRNAs (for a review, see Ehrenfeld (1)). Early after infection, the rate of protein synthesis de- creases, but after about 2 h postinfection, the translation rate increases again. The latter phase of protein synthesis involves exclusively poliovirus mRNA, which is not capped (2, 3); capped cellular mRNA is not degraded (4) but is not trans- lated. In order to determine where the translational machinery
* The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. * Supported by Postdoctoral Fellowship PF-1919 from the Ameri- can Cancer Society.
1 Supported by the Canadian National Cancer Institute and the Medical Research Council.
(1 Supported by United States Public Health Service Grant GM- 22135.
might be defective, Kaufman et al. ( 5 ) compared the high salt ribosomal wash fractions (which contain crude initiation fac- tors) from uninfected and infected cells and found that the fraction derived from infected cells did not stimulate protein synthesis in host polysomes. Further fractionation of the crude initiation factors implicated a defect in eIF3’ (6). Using a different approach, Rose et al. (7) prepared an infected HeLa cell-fi-ee protein synthesizing system dependent on exogenous mRNA, added capped vesicular stomatitis virus mRNA, and asked whether the addition of purified initiation factors might restore translational activity. Of the factors tested, only the eIF4B preparation possessed restoration activity; however, such preparations were later shown to contain a 24K-CBP (8). From the work of Tahara et al. (9), it is now thought that restoration activity is associated with a larger CBP complex which contains 24K-CBP and higher molecular weight pro- teins. The CBP complex is presumed to be inactive in polio- virus-infected cells.
A defect in the cap recognition step of initiation of protein synthesis in poliovirus-infected HeLa cells is an appealing hypothesis since poliovirus mRNA is not capped and, there- fore, is translated by a cap independent mechanism. Sonen- berg and Shatkin (10) devised an assay to detect cap binding proteins which involves cross-linking of the proteins to radio- labeled, oxidized cap structures in mRNA. The 24K-CBP is the major polypeptide to cross-link specifically in the absence of ATP. Specificity is defined by inhibition of cross-linking in the presence of cap analogues. In the presence of ATP, how- ever, several additional proteins (M, = 80,000, 50,000, and 28,000) are also cross-linked (11). Recent evidence from Mer- rick’s and Shatkin’s laboratories (12) indicates that 2 of these larger proteins may be eIF4A (M, = 50,000) and eIF4B (Mr = 80,000) since purified preparations cross-link to caps when combined with each other and with ATP. The relationship of eIF4A and eIF4B to the CBP complex is not clear, however.
We have prepared antisera in goats against eIF2, eIF3, eIF4A, and eIF4B and have used the antisera or affnity- purified antibodies to characterize the initiation factors in crude HeLa cell lysates (13). Using immunoblot analyses and the two-dimensional isoelectric focusing/SDS-polyacrylamide gel method of O’Farrell (14), we have shown that there are no detectable changes in the abundance, molecular weight, or charge forms of eIF4A and eIF4B following poliovirus infec-
’ The abbreviations used are: eIF, eucaryotic initiation factor; SDS, sodium dodecyl sulfate; CBP, cap binding protein; A cut, the 0-40s saturated ammonium sulfate fraction from the high salt ribosomal wash; RSW, the high salt ribosomal wash; HEPES, 442-hydroxy- ethyl)-1-piperazineethanesulfonic acid; 24 K represents 24 kilodaltons, for example.
14806

Polio-induced Proteolysis of a CBP-associated Polypeptide 14807
tion.* We have also examined infected HeLa cell lysates by immunoblot analysis with ant i -eIF3 antiserum, in an effort to detect possible changes in the factor . We descr ibe here the detection of a 220,000-dalton protein (p220) in uninfected cell lysates which is degraded following infection. The p220 ap- pears to associate in part wi th eIF3, and i s p resent in the CBP complex prepared by m'G-affinity chromatography (9). W e propose that proteolysis of this protein may be responsible for the defect in the cap recognition process in poliovirus-infected cells.
EXPERIMENTAL PROCEDURES Cells and Virus-HeLa S:, cells and poliovirus type 1 (Mahoney
strain) were grown and virus was purified as previously described (15). For infection, HeLa cells were suspended a t a density of 6 X 10" cells/ml in Joklik's minimal essential medium supplemented with 5 mM HEPES, pH 7.4. After about 30 min, poliovirus was added a t a multiplicity of infection of 30-50 or as indicated in the figures, and calf serum was added to 6%. Infected cells were harvested by centrif- ugation a t times indicated, lysed, and analyzed as described below or in figure legends.
Cell Subfractions and Initiation Factors-HeLa cells were grown a t densities of 2-8 X 10" cells/ml and harvested a t 5-6 X 10" cells/ml during exponential growth (6). They were washed 3 times with Earle's salts, resuspended in 2 cell pellet volumes of lysis buffer (20 mM HEPES, pH 7.5,lO mM KCI, 1.5 mM Mg(OAc)n, 2.5 mM dithiothreitol), and stored frozen a t -70 "C until use. Upon thawing, phenylmethyl- sulfonyl fluoride was added to a final concentration of 5 mM and the cells were broken by homogenization with a Tekmar tissu- mizer. Postmitochondrial supernatants (SlO), RSW, and ammonium sulfate subfraction, A cut (0-40% saturation), were prepared as de- scribed (6). eIF3 was purified from infected or uninfected HeLa cells through the phosphocellulose step (step 4) as described for rabbit reticulocyte eIF3 (16).
The cap binding protein complex was purified from the A cut of rabbit reticulocytes. A cut (10-20 A ~ X O units) was sedimented in a 10- 35% sucrose gradient in a Beckman SW 40 rotor a t 38,000 rpm for 24 h, in buffer I (20 mM HEPES, pH 7.5, 0.2 mM EDTA, 7 mM 2- mercaptoethanol, and 0.5 mM phenylmethylsulfonyl fluoride) contain- ing 0.5 M KCI. Fractions from the top third of the gradient that did not include eIF3 were pooled and dialyzed against buffer containing 0.1 M KCI. For affinity chromatogr_aphy. 6-10 AM, units were loaded onto a column (1.5 X 0.7 cm) of m'GTP-agarose, synthesized as will be described elsewhere, and equilibrated with buffer I containing 0.1 M KC1 and 10% glycerol. The column was washed with 35 ml of the same buffer, followed by 4 ml of 0.1 M GTP in buffer I and 10% glycerol. Elution of cap binding protein complex was accomplished by passing 4 ml of 70 p~ m'GDP in buffer A and 10°C glycerol. The yield of cap binding protein complex was 50-150 pg and consisted of several polypeptides (approximately 24,50, 100. and 210 kilodaltons). This fraction was active in restoring translation of capped mRNA in poliovirus-infected cell-free extracts. Bovine serum albumin was added to the cap binding protein complex (1 mg/ml) to stabilize biological activity.
Antisera-Goat antiserum against purified rabbit reticulocyte eIF3 is that characterized by Meyer et al. (13). Antibodies against the 220,000-dalton protein (p220) were affinity-purified by modifications (13) of the method of Olmsted (19). The antigen preparation was prepared as follows. Uninfected HeLa cell A cut (approximately 30 Aw, units) was loaded onto a sucrose gradient containing 100 mM KC1 and centrifuged according to conditions described in Fig. 3. Material in a gradient fraction containing p220, but little detectable p170 (as in fraction 16, Fig. 3A) was fractionated on a 10% SDS-polyacrylamide slab gel, then electrophoretically transferred to ABM paper. A hori- zontal strip corresponding to proteins of apparent molecular weight 220,000 (located by immunoblotting with anti-eIF3) was excised and used as the antigen for the affinity purification. The excised ~ 2 2 0 - containing strips were incubated with anti-eIF3 antiserum, then washed with 4 M guanidinium chloride, 10 mM K phosphate, pH 7.2, and 150 mM NaCl to remove the p220 antibodies. The eluted anti- p220 antibody solution was adjusted to pH 7.5 and dialyzed overnight against 10 mM K phosphate, pH 7.5, and 150 mM NaCI.
Polyacrylamide Gel Electrophoresis and Immunoblot Analysis- Material to be analyzed by immunoblot was prepared as described in
'R. Duncan, D. Etchison, and J. W. B. Hershey, manuscript in ~. -
preparation.
the figure legends. Samples were boiled 2 min with sample buffer, loaded onto 10 or 12% SIX-polyacrylamide slab gels (1.4 X 140 X 160 or 1.4 X 140 X 100 mm) and subjected to electrophoresis (17). Proteins in the gel were transferred electrophoretically to nitrocellulose sheets as described previously (13, 18). Briefly, the nitrocellulose paper was stained to locate protein bands, incubated with blocking buffer con- taining 3% bovine serum albumin, and incubated in a solution con- taining goat anti-eIF3 antiserum or affinity-purified antibodies. After washing in a detergent-containing buffer to remove excess antibodies, the paper was incubated with a second antibody, rabbit anti-goat IgG, which was radiolabeled with "'1 (specific activity 2-5 X 10' cpm/pg). It was then washed to remove excess radiolabel, dried, and exposed to Kodak X-Omat SB5 film for 1 to 3 days.
RESULTS
Immunob lo t Analyses of eIF3"The identification of a de- fect in eIF3 activity after poliovirus infection (6) led us to analyze for possible differences in the structure of purified eIF3 from infected and uninfected cells. Stained SDS-poly- acrylamide gel patterns of eIF3 from infected (I-eIF3) or uninfected (U-eIF3) cells looked similar. Furthermore, stained gel patterns from different purified preparations of U-eIF3 varied to such an extent that it was difficult to conclude that any differences in patterns of U-eIF3 as compared to I-eIF3 were due specifically to infection (data not shown).
The development of polyclonal antibodies against rabbit reticulocyte eIF3 (13) provided us with a tool to analyze whole cytoplasmic extracts for alterations in eIF3 proteins after infection. This antiserum cross-reacts with HeLa eIF3 a n d
A. A n t i - e I F 3 E. A ~ ~ - a n t i - p 2 2 0 C. A F F - a n t i - p 2 2 0 I 2 3 4 1 2 3 4 1 2 3 4 5
p220- . 200.
p,15- ==I = p170- . - - - 200p22~- t. i 4
117. 117 93 68
== 93-
68. p 6 6 - 43
p47- - - " 29
p36- "
29 - u-L CBP
U I
u + + + + - I - + + + + 30' 0' 15' 30' 30'
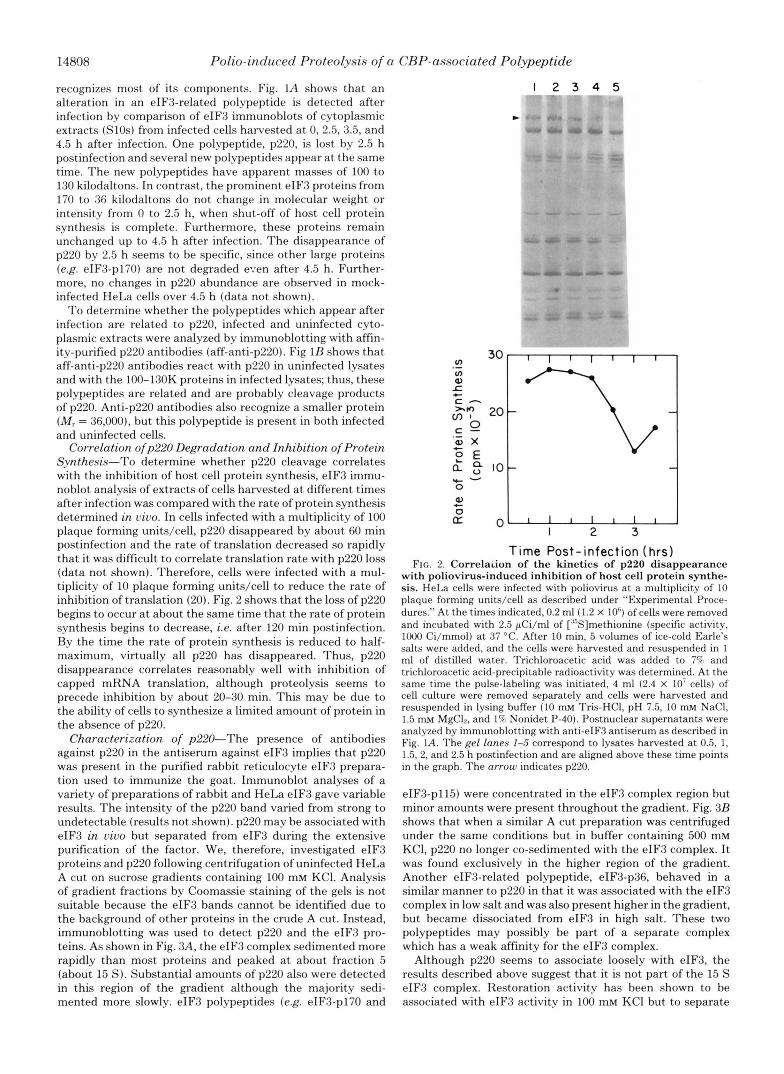
FIG. 1. Immunoblot analyses with anti-eIF3 and affhi ty-pu- rified anti-p220. Uninfected and poliovirus-infected HeLa cell ly- sates, RSWs, and CBP complex were prepared, fractionated by SDS- polyacrylamide gel electrophoresis, and analyzed by immunoblotting as described under "Experimental Procedures." The figure shows photographs of autoradiograms exposed for 1 to 3 days. A , lysates (SlOs) from HeLa cells infected a t 30 plaque forming units/cell were prepared a t 0,2.5,3.5, and 4.5 h postinfection (lanes 2-4 , respectively) and aliquots each containing 0.3 A M units were analyzed. Immuno- blotting was done with anti-eIF3 antiserum. Major eIF3 components and p220 are identified on the left by their apparent molecular weights in this gel system. The arroux on the right point to new cross-reactive proteins, presumably degradation products of p220. B , immuno- blotting was done with affinity-purified anti-p220. Lane I , about 0.3 A m units of lysate from uninfected HeLa cells ( U ) ; lane 2, about 0.3 AZRO units of lysate from poliovirus-infected cells harvested a t 4.5 h postinfection ( I ) ; lane 3, about 0.2 A M , units of uninfected lysate; lane 4, 1.2 pg of CBP complex. Molecular weight markers (in kilodaltons) were myosin heavy chain (200). /&galactosidase (21 7), phosphorylase a (93), bovine serum albumin (68). ovalbumin (43), and carbonic anhvdrase (29). C, about 0.2 AM units each of U-RSW (lane I ) and I-RSW (lane 5) were incubated a t 37 "C separately for 30 min or together for 0. 15, or 30 min (Innes 2-4, respectively) and then analyzed by immunoblotting with aff-anti-p220.
14808 Polio-induced Proteolysis of a CBP-associated Polypeptide
recognizes most of i t s components. Fig. 1A shows that an alteration in an eIF3-related polypeptide is detected after infection by comparison of eIF3 immunoblots of cytoplasmic extracts (SlOs) from infected cells harvested at 0, 2.5, 3.5, and 4.5 h after infection. One polypeptide, p220, is lost by 2.5 h postinfection and several new polypeptides appear at the same time. The new polypeptides have apparent masses of 100 to 130 kilodaltons. In contrast, the prominent eIF3 proteins from 170 to 36 kilodaltons do not change in molecular weight or intensity from 0 to 2.5 h, when shut-off of host cell protein synthesis is complete. Furthermore, these proteins remain unchanged up to 4.5 h after infection. The disappearance of p220 by 2.5 h seems to be specific, since other large proteins (e.g. eIF3-pl70) are not degraded even after 4.5 h. Further- more, no changes in p220 abundance are observed in mock- infected HeLa cells over 4.5 h (data not shown).
To determine whether the pol-ypeptides which appear after infection are related to p220, infected and uninfected cyto- plasmic extracts were analyzed by immunoblotting with affin- ity-purified p220 antibodies (aff-anti-p220). Fig 1B shows that aff-anti-p220 antibodies react with p220 in uninfected lysates and with the 100-130K proteins in infected lysates; thus, these polypeptides are related and are probably cleavage products of p220. Anti-p220 antibodies also recognize a smaller protein ( M , = 36,000), but this polypeptide is present in both infected and uninfected cells.
Correlation ofp220 Degradation and Inhibition of Protein Synthesis-To determine whether p220 cleavage correlates with the inhibition of host cell protein synthesis, eIF3 immu- noblot analysis of extracts of cells harvested at different times after infection was compared with the rate of protein synthesis determined in civo. In cells infected with a multiplicity of 100 plaque forming units/cell, p220 disappeared by about 60 min postinfection and the rate of translation decreased so rapidly that it was difficult to correlate translation rate with p220 loss (data not shown). Therefore, cells were infected with a mul- tiplicity of 10 plaque forming units/cell to reduce the rate of inhibition of translation (20). Fig. 2 shows that the loss of p220 begins to occur.at about the same time that the rate of protein synthesis begins to decrease, i.e. after 120 min postinfection. By the time the rate of protein synthesis is reduced to half- maximum, virtually all p220 has disappeared. Thus, p220 disappearance correlates reasonably well with inhibition of capped mRNA translation, although proteolysis seems to precede inhibition by about 20-30 min. This may be due to the ability of cells to synthesize a limited amount of protein in the absence of p220.
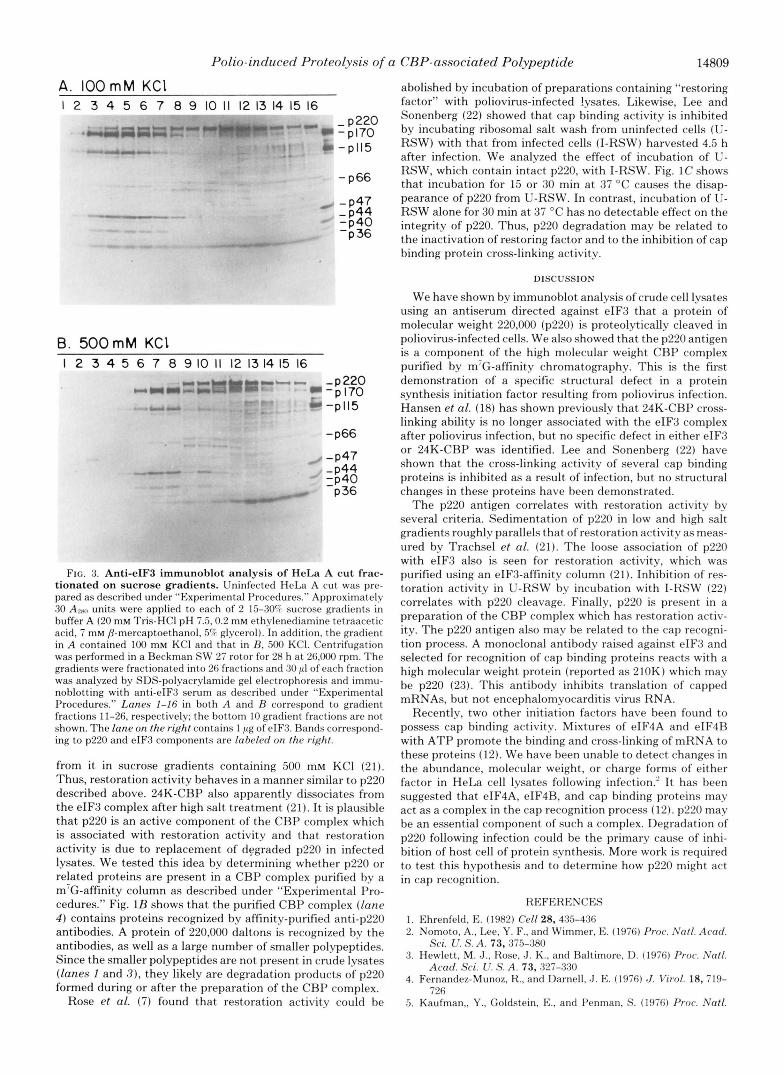
Characterization of p220"The presence of antibodies against p220 in the antiserum against eIF3 implies that p220 was present in the purified rabbit reticulocyte eIF3 prepara- tion used to immunize the goat. Immunoblot analyses of a variety of preparations of rabbit and HeLa eIF3 gave variable results. The intensity of the p220 band varied from strong to undetectable (results not shown). p220 may be associated with eIF3 in vivo but separated from eIF3 during the extensive purification of the factor. We, therefore, investigated eIF3 proteins and p220 following centrifugation of uninfected HeLa A cut on sucrose gradients containing 100 mM KCl. Analysis of gradient fractions by Coomassie staining of the gels is not suitable because the eIF3 bands cannot be identified due to the background of other proteins in the crude A cut. Instead, immunoblotting was used to detect p220 and the eIF3 pro- teins. As shown in Fig. 3A, the eIF3 complex sedimented more rapidly than most proteins and peaked at about fraction 5 (about 15 S). Substantial amounts of p220 also were detected in this region of the gradient although the majority sedi- mented more slowly. eIF3 polypeptides ( e g . eIF3-pl70 and
I 2 3 4 5
c
c a3 0
' L o I 2 3
Time Post-infection (hrs) FIG. 2. Correlaiion of the kinetics of p220 disappearance
with poliovirus-induced inhibition of host cell protein synthe- sis. HeLa cells were infected with poliovirus at a multiplicity of 10 plaque forming units/cell as described under "Experimental Proce- dures." At the times indicated, 0.2 ml (1.2 X 10') of cells were removed and incubated with 2.5 pCi/ml of ["'Slmethionine (specific activity, lo00 Ci/mmol) at 37 "C. After 10 min, 5 volumes of ice-cold Earle's salts were added, and the cells were harvested and resuspended in 1 ml of distilled water. Trichloroacetic acid was added to 7% and trichloroacetic acid-precipitable radioactivity was determined. At the same time the pulse-labeling was initiated, 4 ml (2.4 X 10' cells) of cell culture were removed separately and cells were harvested and resuspended in lysing buffer (10 mM Tris-HCI, pH 7.5, 10 m~ NaCI, 1.5 mM MgC12, and 1% Nonidet P-40). Postnuclear supernatants were analyzed by immunoblotting with anti-eIF3 antiserum as described in Fig. L4. The gel lanes 1-5 correspond to lysates harvested at 0.5, 1, 1.5.2, and 2.5 h postinfection and are aligned above these time points in the graph. The arrow indicates p220.
eIF3-pl15) were concentrated in the eIF3 complex region but minor amounts were present throughout the gradient. Fig. 3B shows that when a similar A cut preparation was centrifuged under the same conditions but in buffer containing 500 mM KC1, p220 no longer co-sedimented with the eIF3 complex. It was found exclusively in the higher region of the gradient. Another eIF3-related polypeptide, eIF3-p36, behaved in a similar manner to p220 in that it was associated with the eIF3 complex in low salt and was also present higher in the gradient, but became dissociated from eIF3 in high salt. These two polypeptides may possibly be part of a separate complex which has a weak affinity for the eIF3 complex.
Although p220 seems to associate loosely with eIF3, the results described above suggest that it is not part of the 15 S eIF3 complex. Restoration activity has been shown to be associated with eIF3 activity in 100 mM KC1 but to separate
Polio-induced Proteolysis of a CBP-associated Polypeptide 14809
A. 100 mM KC\ I 2 3 4 5 6 7 8 9 1011 1213141516
-. - . .. - P220 - pl70 -PI15
- p66
- p47 - p44 -p40 - p 36
B. 500mM KC1 I 2 3 4 5 6 7 8 9 IO I I 12 1314 15 16
-p 220 -p I70 -PI15
- p66
- D47
FIG. 3. Anti-eIF3 immunohlot analysis of HeLa A cut frac- tionated on sucrose gradients. Uninfected HeLa A cut was pre- pared as described under “Experimental I’rocedures.” Approximately 30 A ~ P , units were applied to each of 2 15-30? sucrose gradients in buffer A (20 rnM Tris-HCI pH 7.5.0.2 rnM ethylenediamine tetraacetic acid, 7 mM 8-mercaptoethanol, 5% glycerol). In addition, the gradient in A contained 100 mM KC1 and that in B, 500 KCI. Centrifugation was performed in a Beckrnan SW 27 rotor for 28 h at 26,000 rpm. The gradients were fractionated into 26 fractions and 30 pI of each fraction was analyzed by SDS-polyacrylamide gel electrophoresis and imrnu- noblotting with anti-eIF3 serum as described under “Experimental Procedures.” Lanes I-Z6 in both A and B correspond to gradient fractions 11-26, respectively; the bottom 10 gradient fractions are not shown. The lane on the right contains 1 pg of eIF3. Bands correspond- ing to p220 and eIF3 components are laheled on the right.
from it in sucrose gradients containing 500 mM KC1 (21). Thus, restoration activity behaves in a manner similar to p220 described above. 24K-CBP also apparently dissociates from the eIF3 complex after high salt treatment (21). It is plausible that p220 is an active component of the CBP complex which is associated with restoration activity and that restoration activity is due to replacement of degraded p220 in infected lysates. We tested this idea by determining whether p220 or related proteins are present in a CBP complex purified by a m’G-affinity column as described under “Experimental Pro- cedures.” Fig. 1B shows that the purified CBP complex (lane 4 ) contains proteins recognized by affinity-purified anti-p220 antibodies. A protein of 220,000 daltons is recognized by the antibodies, as well as a large number of smaller polypeptides. Since the smaller polypeptides are not present in crude lysates (lanes 1 and 3 ) , they likely are degradation products of p220 formed during or after the preparation of the CBP complex.
Rose et al. (7) found that restoration activity could be
abolished by incubation of preparations containing “restoring factor” with poliovirus-infected !ysates. Likewise, Lee and Sonenberg (22) showed that cap binding activity is inhibited by incubating ribosomal salt wash from uninfected cells (U- RSW) with that from infected cells (I-RSW) harvested 4.5 h after infection. We analyzed the effect of incubation of U- RSW, which contain intact p220, with I-RSW. Fig. 1C shows that incubation for 15 or 30 min at 37 “C causes the disap- pearance of p220 from U-RSW. In contrast, incubation of U- RSW alone for 30 min at 37 “C has no detectable effect on the integrity of p220. Thus, p220 degradation may be related to the inactivation of restoring factor and to the inhibition of cap binding protein cross-linking activity.
DISCUSSION
We have shown by immunoblot analysis of crude cell lysates using an antiserum directed against eIF3 that a protein of molecular weight 220,000 (p220) is proteolytically cleaved in poliovirus-infected cells. We also showed that the p220 antigen is a component of the high molecular weight CRP complex purified by m’G-affinity chromatography. This is the first demonstration of a specific structural defect in a protein synthesis initiation factor resulting from poliovirus infection. Hansen et al. (18) has shown previously that 24K-CBP cross- linking ability is no longer associated with the eIF3 complex after poliovirus infection, but no specific defect in either eIF3 or 24K-CBP was identified. Lee and Sonenberg (22) have shown that the cross-linking activity of several cap binding proteins is inhibited as a result of infection, but no structural changes in these proteins have been demonstrated.
The p220 antigen correlates with restoration activity by several criteria. Sedimentation of p220 in low and high salt gradients roughlv parallels that of restoration activity as meas- ured by Trachsel et al. (21). The loose association of p220 with eIF3 also is seen for restoration activity, which was purified using an eIF3-affinity column (21). Inhibition of res- toration activity in U-RSW by incubation with I-RSW (22) correlates with p220 cleavage. Finally, p220 is present in a preparation of the CRP complex which has restoration activ- ity. The p220 antigen also may be related to the cap recogni- tion process. A monoclonal antibody raised against eIF3 and selected for recognition of cap binding proteins reacts with a high molecular weight protein (reported as 210K) which may be p220 (23). This antibody inhibits translation of capped mRNAs, but not encephalomyocarditis virus RNA.
Recently, two other initiation factors have been found to possess cap binding activity. Mixtures of eIF4A and eIF4R with ATP promote the binding and cross-linking of mRNA to these proteins (12). We have been unable to detect changes in the abundance, molecular weight, or charge forms of either factor in HeLa cell lysates following infection.2 It has been suggested that eIF4A, eIF4B, and cap binding proteins may act as a complex in the cap recognition process (12). p220 may be an essential component of such a complex. Degradation of p220 following infection could be the primary cause of inhi- bition of host cell of protein synthesis. More work is required to test this hypothesis and to determine how p220 might act in cap recognition.
REFERENCES 1. Ehrenfeld, E. (1982) Cell 28, 435-4313 2. Nomoto, A., Lee, Y. F.. and Wimmer, E. (1976) Proc. Natl. Acad.
3. Hewlett, M. .I.. Rose, d . K., and Baltimore, 11. (1976) Proc. Natl.
4. Fernandez-Munoz, R., and Darnell, J . E. (1976) J. Virol. 18, 719-
5. Kaufman,, Y., Goldstein, E., and Penman, S. (1976) Proc. Natl.
Sei. U. S. A. 73, 375-380
Acad. Sei. U. S. A . 73, 327-330
726

14810 Polio-induced Proteolysis of a CBP-associated Polypeptide
Acad. Sci. U. S. A . 73, 1834-1838
J. W. B. (1979) J. Biol. Chem. 254, 10973-10978
Proc. Natl. Acad. Sci. U. S. A. 75,2732-2736
(1978) Proc. Natl. Acad. Sci. U . S. A . 75,4843-4847
Chem. 256,7691-7694
6. Helentjaris, T., Ehrenfeld, E., Brown-Luedi, M. L., and Hershey,
7. Rose, J. K., Trachsel, H., Leong, K., and Baltimore, D. (1978)
8. Sonenberg, N., Morgan, M. A., Merrick, W. C., and Shatkin, A. J.
9. Tahara, S. M., Morgan, M. A,, and Shatkin, A. J. (1981) J. Biol.
10. Sonenberg, N., and Shatkin, A. J. (1977) Proc. Natl. Acud. Sci.
11. Sonenberg, N. (1981) Nucleic Acids Res. 9, 1643-1656 12. Grifo, J. A,, Tahara, S. M., Leis, J. P., Morgan, M. A., Shatkin, A.
13. Meyer, L. J., Milburn, S. C., and Hershey, J. W. B. (1982)
U. S. A. 74,4288-4292
J., and Merrick, W. C . (1982) J. Biol. Chem. 257, 5246-5252
Biochemistry 21,4206-4211
14. OFarrell, P. H. (1975) J. Biol. Chem. 250, 4007-4021 15. Etchison, D., and Ehrenfeld, E. (1980) Virology 107, 135-143 16. Benne, R., Brown-Luedi, M. L., and Hershey, J. W. B. (1979)
17. Laemmli, U. K. (1970) Nature (Lond.) 227,680-685 18. Hansen, J., Etchison, D., Hershey, J. W. B., and Ehrenfeld, E.
19. Olmsted, J. B. (1981) J. Biol. Chem. 256, 11955-11957 20. Helentjaris, T., and Ehrenfeld, E. (1978) J. Virol. 26, 510-521 21. Trachsel, H., Sonenberg, N., Shatkin, A. J., Rose, J. K., Leong,
K., Bergmann, J. E., Gordon, J., and Baltimore, D. (1980) Proc. Natl. Acad. Sci. U. S. A. 77, 770-774
22. Lee, K. A. W., and Sonenberg, N. (1982) Proc. Natl. Acad. Sci. U. S. A . 79, 3447-3451
23. Sonenberg, N., and Trachsel, H. (1982) Curr. Top. Cell. Regul.
Methods Enzymol. 60, 15-35
(1982) J. Virol. 42,200-207
21,65-88