the differentiation of cardiomyocytes from mouse embryonic stem cells is altered by dioxin
TRANSCRIPT

Ta
TRa
b
c
d
e
f
g
a
ARRAA
KCEDMM
1
(iocee
d
s
G
0d
Toxicology Letters 202 (2011) 226–236
Contents lists available at ScienceDirect
Toxicology Letters
journa l homepage: www.e lsev ier .com/ locate / tox le t
he differentiation of cardiomyocytes from mouse embryonic stem cells isltered by dioxin
ui Neria,1,2, Valeria Mericoa,b,2, Fabio Fiordalisoc, Monica Salioc, Paola Rebuzzinia, Lucia Sacchid,iccardo Bellazzid,e, Carlo Alberto Redia,b,f, Maurizio Zuccotti g,∗, Silvia Garagnaa,e,f,∗∗
Laboratorio di Biologia dello Sviluppo, Dipartimento di Biologia Animale, Universita’ degli Studi di Pavia, Via Ferrata 9, 27100 Pavia, ItalyFondazione I.R.C.C.S. Policlinico San Matteo, Piazzale Golgi, 19, 27100 Pavia, ItalyUnit of Bio-imaging, Department of Cardiovascular Research, Istituto di Ricerche Farmacologiche “Mario Negri”, Via La Masa 19, 20156 Milan, ItalyDipartimento di Informatica e Sistemistica, Universita’ degli Studi di Pavia, ItalyCentro di Ricerca Interdipartimentale di Ingegneria Tissutale, Universita’ degli Studi di Pavia, Via Ferrata 1, 27100 Pavia, ItalyCentro di Eccellenza in Biologia Applicata, Universita’ degli Studi di Pavia, Via Ferrata 1, 27100 Pavia, ItalySezione di Istologia ed Embriologia, Dipartimento di Medicina Sperimentale, Universita’ degli Studi di Parma, Via Volturno 39, 43100 Parma, Italy
r t i c l e i n f o
rticle history:eceived 16 July 2010eceived in revised form 10 February 2011ccepted 11 February 2011vailable online 24 February 2011
eywords:ardiomyocytes differentiationmbryonic stem cells
a b s t r a c t
2,3,7,8-Tetrachlorodibenzo-para-dioxin (TCDD) causes abnormalities during heart development. Car-diomyocytes derived from embryonic stem (ES) cells are a robust model for the study of earlycardiomyogenesis. Here, we evaluated the effects of TCDD at key stages during the differentiation ofmouse ES cells into cardiomyocytes analysing: (i) the transcription of lineage differentiation (Brachyury,Nkx-2.5, Actc-1), cardiac-specific (Alpk3, cTnT, cTnI, cTnC) and detoxification phase I (Cyp1A1, Cyp1A2 andCyp1B1) and phase II (Nqo1, Gsta1 and Ugt1a6) genes; (ii) the global gene expression; (iii) the ultra-structure of ES-derived cardiomyocytes; (iv) level of ATP production and (v) the immunolocalisation ofsarcomeric �-actinin, �-myosin heavy chain and cTnT proteins. We show that TCDD affects the differen-
ioxinyofibrilsouse
tiation of ES cells into cardiomyocytes at several levels: (1) induces the expression of phase I genes; (2)down-regulates a group of heart-specific genes, some involved in the oxidative phosphorylation path-way; (3) reduces the efficiency of differentiation; (4) alters the arrangement of mitochondria, that showtwisted and disrupted cristae, and of some sarcomeres, with misalignement or disarrangement of themyofibrillar organisation and (5) reduces ATP production. This study provides novel evidences that TCDD
fferenting a
impairs cardiomyocyte ditheir disruption represen
. Introduction
The heart is considered one of the target organs of dioxin2,3,7,8-tetrachlorodibenzo-para-dioxin or TCDD) toxicity. Dioxins one of the most dangerous and diffused man-made xenobi-
tics. In the environment, dioxins tend to accumulate in the foodhain and humans, being at the top of it, accumulate the high-st quantities. Dioxins are of concern to the public, as short-termxposure to high levels may result in skin lesions, such as chlo-∗ Corresponding author.∗∗ Corresponding author at: Laboratorio di Biologia dello Sviluppo, Dipartimentoi Biologia Animale, Universita’ degli Studi di Pavia, Via Ferrata 9, 27100 Pavia, Italy.
E-mail addresses: [email protected] (M. Zuccotti),[email protected] (S. Garagna).
1 Present address: INSERM Avenir Program, Stem cells and Cardiogenesis, Campusenopole 1, 4 rue Pierre Fontaine, 91058 Evry, France.2 These authors contributed equally to this work.
378-4274/$ – see front matter © 2011 Elsevier Ireland Ltd. All rights reserved.oi:10.1016/j.toxlet.2011.02.008
tiation. Sarcomeres and mitochondria could be a target for dioxin toxicity,possible mechanism developing cardiac injury.
© 2011 Elsevier Ireland Ltd. All rights reserved.
racne, patchy darkening of the skin and altered liver function.Long-term exposure induces impairment of the immune system,liver damage, cancer, disruption of hormone signalling pathways,reproductive and developmental defects (Mandal, 2005), includ-ing altered heart formation (Yeager et al., 2006). Like the liverand thymus, the heart expresses the AhR-Arnt-DRE system. TheAhR (aryl hydrocarbon receptor) is a ligand-activated transcriptionfactor belonging to the basic helix-loop-helix/Per-AhR-Arnt-Sim(bHLH/PAS) superfamily of proteins. The mechanism of induction ofgene transcription by TCDD involves ligand recognition and bindingby the AhR, nuclear translocation, and dimerization with the AhRnuclear translocator (Arnt). The TCDD-AhR-Arnt heterodimer bindsto Dioxin-Responsive Elements present in dioxin-inducible genesand modulates their expression (Nebert et al., 2000; Kelling et al.,
1987; Schmidt et al., 1996; Xie et al., 2006). Chronic exposure of ratsto TCDD determines a dose-dependent increase in the incidenceof degenerative cardiovascular lesions and ultimately cardiomy-opathy (Jokinen et al., 2003). Adult mice exposed subchronicallyto TCDD developed high blood pressure and cardiac enlargement
Lette
(dSses
v2sozASytod
fsffae(
fedemstod
dftepeo(iiwa
ttsypdp(oeCUdEet
T. Neri et al. / Toxicology
Kopf et al., 2008), whilst exposure to an acute high dose of TCDDetermined an increase in triglyceride levels (Dalton et al., 2001).ome of these changes may be ascribed to altered gene expres-ion, inflammation and oxidative stress (Lund et al., 2005; Arzuagat al., 2007), whereas others are related to direct effects on calciumignalling in cardiomyocytes (Xie et al., 2006).
During development, TCDD has been shown to cause heart andascular abnormalities in a variety of animal models (Jokinen et al.,003; Kopf et al., 2008; Dalton et al., 2001; Lind et al., 2004). Thesetudies have demonstrated dioxin-related decrease in cardiomy-cyte proliferation very early during development in chick andebrafish embryos and in the murine fetus (Ivnitski et al., 2001;ntkiewicz et al., 2005; Thackaberry et al., 2005a; Goldstone andtegeman, 2006). Thackaberry et al. (2005b), by microarrays anal-sis of gene expression, have shown global changes in cardiac generanscription following TCDD exposure during in vivo developmentf the fetal murine heart, indicating possible alterations during car-iac cell differentiation.
The effects that TCDD exerts on cardiomyocytes whilst they dif-erentiate is still poorly known. To this end, the use of embryonictem (ES) cells, an established model of in vitro cardiomyocyte dif-erentiation (Kehat et al., 2001; Xu et al., 2002) has been exploitedor the first time in a recent study, in which TCDD has been found tolter the expression of developmentally regulated genes in a vari-ty of differentiation pathways and in a number of homeotic genesWang et al., 2010).
Mouse ES cells differentiate into cardiomyocytes through theormation of embryoid bodies (EBs) that recapitulate, among oth-rs, the developmental stages characteristic of mouse fetal heartevelopment (Boheler et al., 2002). ES cell-derived cardiomyocytesxpress cardiac gene transcripts in a developmentally controlledanner at time-points corresponding to three main developmental
tages: (1) early, primary myocardial-like cells, at 4 days of differen-iation; (2) intermediate, myofibrillogenic commitment, at 7 daysf differentiation and (3) terminal, cardiomyocytes at 12 days ofifferentiation.
Compared to in vivo studies, the use of an in vitro model of car-iomyocyte differentiation (cardiomyocytes may be obtained alsorom human ES cells, Kehat et al., 2001; Schuldiner et al., 2000) hashe advantage of allowing a more precise definition of the effectsxerted by TCDD on a specific cell type (i.e., cardiomyocytes) com-ared to the entire heart and permits the identification of primaryffects that in vivo would be more difficult to distinguish from sec-ndary effects resulting from damages to other tissues and organse.g., liver and kidney). Furthermore, this in vitro model allows thenvestigation of the effects of TCDD at different time points dur-ng cardiomyocyte differentiation and to study their functionality
ithout quantitative limitations, a difficult task to carry out usingnimal models.
Whilst some of the molecular changes induced by TCDD duringhe differentiation of ES cells into cardiomyocytes are beginningo emerge (Wang et al., 2010), its effects on the cellular organi-ation and on global gene expression have not been investigatedet. Thus, in the presence or absence of TCDD, at specific timeoints along the process of differentiation of ES cells into car-iomyocytes we have analysed: (1) on day 4, 7 and 12, theattern of transcription of Brachyury (mesoderm marker), Nkx-2.5cardiac mesoderm marker) and alpha c actin (Actc-1, cardiomy-cyte maker) genes; (2) on day 4, 7 and 12, the constitutivexpression and TCDD-inducibility of phase I (cytochromes P450yp1A1, Cyp1A2 and Cyp1B1) and phase II (Nqo1, Gsta1 and
gt1a6) genes, involved in the detoxification response; (3) onay 12 from EBs formation, the ultrastructural characteristics ofS-derived cardiomyocytes; the immunolocalisation of sarcom-ric �-actinin, �-myosin heavy chain (�-myosin) and cardiacroponin T (cTnT) proteins, markers of their differentiated sta-rs 202 (2011) 226–236 227
tus (Boheler et al., 2002) and by microarrays the transcriptomeprofile.
2. Materials and methods
2.1. ES cells culture
The mouse low-affinity R1 ES cell line [established from (129X1/SvJ x 129S1)F1 3.5 days post coitum (dpc) blastocyst; kindly provided by Dr. Nagy, SamuelLunenfeld Research Institute, Mount Sinai Hospital, Toronto, Ontario, Canada], wascultured as previously described (Neri et al., 2008). Briefly, ES cells were main-tained on a mitomycin C-treated STO feeder-layer (ATCC, CRL-2225) in a completeES medium supplemented with 15% ES Cell Qualified Fetal Bovine Serum (Gibco)and 500 U/ml ESGRO-LIF (Chemicon International). ES cells were routinely passagedevery 48 h and maintained in an incubator at 37 ◦C with 7.5% CO2 in air. Cultureof ES cells in medium containing TCDD (Cambridge Isotope Laboratories, Inc.) wasbegun after three passages on gelatin, allowing the elimination of the feeder layer offibroblasts, as confirmed by microscopic observations. TCDD, dissolved in dimethylsulphoxide (DMSO, used as vehicle, Sigma–Aldrich), was added to the completeES medium to a final concentration of 100 nmol/l. This concentration was chosenbased on the results of our earlier study that showed the highest TCDD-induction ofCyp1A1 expression in undifferentiated mouse ES cells obtained from a TCDD low-affinity strain (Neri et al., 2008). Control samples, cells cultured in the absence (Ctrl)or presence (DMSO) of 0.06% DMSO, were carried out and analysed in parallel.
2.2. Embryoid bodies formation and cardiomyocytes differentiation
To ensure sample uniformity EBs were formed from 1 × 103 ES cells using thehanging drop method (Wobus et al., 2002) in an ES cell medium without LIF, con-taining 100 nmol/l TCDD; the medium was changed regularly and used throughoutdifferentiation. After 3 days of culture in drops, EBs were passaged to 100 mm Petridishes (Corning) for culture in suspension and on day 5, 3 EBs were plated in each1.9 cm2 well of a 24 wells plate (Corning). On day 4, 7 and 12 of EBs differentiation,cells were harvested for RT-PCR and immunocytochemistry (on day 12) analyses.Controls (see above) were carried out and analysed in parallel.
2.3. Contraction assay
On day 7 and 12 of differentiation the number of beating EBs was counted (zurNieden et al., 2004). To evaluate the frequency of contraction, the number of beatsper min was counted for every ES-derived cardiomyocyte sample in at least 10 beat-ing areas. The experiment was repeated three times. The results are presented asmean ± standard deviation.
2.4. RT-PCR analysis
RT-PCR analyses were performed as previously described (Neri et al., 2008).Briefly, total RNA was isolated using the GenElute Mammalian Total RNAKit (Sigma–Aldrich), following the manufacturer’s instructions. All traces ofDNA contamination were eliminated using an on-column DNaseI digestion kit(Sigma–Aldrich). 1 �g of RNA was retrotranscribed and the cycles used for the PCRof all the genes under study were optimised. The optimal number of PCR cycles foreach gene was selected within the linear range of the regression curve. The follow-ing cycle programme was used for PCR amplification: 94 ◦C for 5 min, followed byn cycles (Cyp1A1, Gsta1 and Nkx-2.5 n = 35; Cyp1A2, n = 38; Cyp1B1 and Brachyury,n = 33; Nqo1, n = 28; Actc-1, n = 29; Ugt1a6, n = 40; Gapdh, n = 25) at 94 ◦C for 30 sec,and Tm (Cyp1A at 61 ◦C; Cyp1A2, Cyp1B1, Gsta1, Nqo1 and Ugt1a6 at 60 ◦C; Gapdh at55 ◦C) for 30 s, 72 ◦C for 30 s, followed by a single step at 72 ◦C for 7 min. The primersequences for phase I and phase II genes are those described earlier (Neri et al.,2008). PCR products were sequenced and showed 99% homology with the sequencefound in ENSEMBL. 10 �l of PCR product were mixed with 2 �l of loading bufferand were electrophoresed on a 2.5% agarose gel in 0.5× TBE containing 0.1 �g/mlethidium bromide at 6 V/cm for 45 min. The PCR products were visualised undershort-wave length UV and quantified using a Bio-Rad Gel Doc system by comparing,after normalisation on the internal control Gapdh product, the known quantity ofthe low mass ladder marker bands with those obtained from the amplification ofthe gene sequences.
For quantitative real-time PCR, one twentieth of the cDNA resulting from 1 �gof the retrotranscribed RNA was amplified in triplicate with a Rotorgene 6000(Corbett Life Science) in 20 �l reaction mixture containing 200 nM of each spe-cific primer and the MESA GREEN qPCR MasterMix Plus for SYBR assay no ROXsample (Eurogentec) at 1× final concentration. The amplification reaction was asfollows: 95 ◦C for 5 min, followed by 30 cycles at 95 ◦C for 10 s, 60 ◦C for 15 s, 72 ◦Cfor 20 s. The Rotorgene 6000 Series Software 1.7 was used for comparative concen-
tration analysis. �-2-Microglobulin expression was used for sample normalisation.PCR products specificity from each sequence analysed was confirmed using meltingcurve analysis (from 56 ◦C to 95 ◦C) and subsequent agarose gel electrophore-sis. Primer sequences were as follows: Alpk3, F 5′ AACGGCAGCTTTCTTGTCAC3′ , R 5′ CTCTCCTTGAGGCCTTGGTA 3′; cTnC, F 5′ CAGCAAAGGGAAGTCTGAGG 3′ ,R 5′ TGCAGCATCATCTTCAGCTC 3′; cTnT, F 5′ GAAGTTCGACCTGCAGGAAA 3′ , R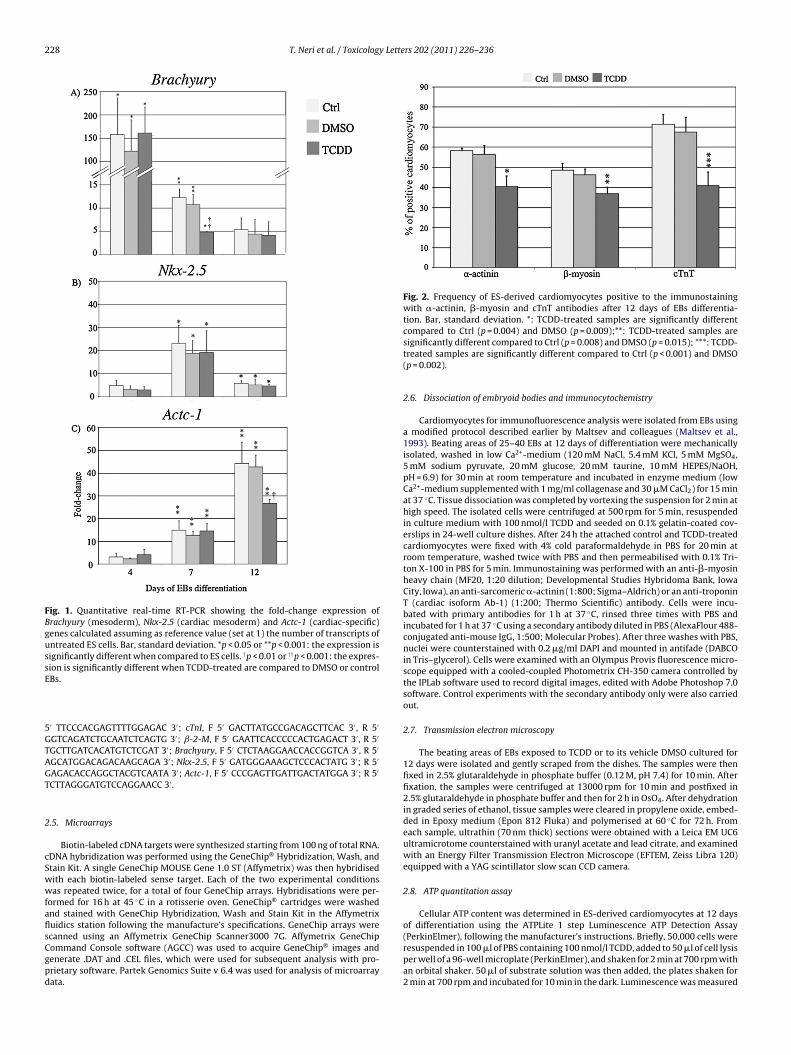
228 T. Neri et al. / Toxicology Letters 202 (2011) 226–236
Fig. 1. Quantitative real-time RT-PCR showing the fold-change expression ofBrachyury (mesoderm), Nkx-2.5 (cardiac mesoderm) and Actc-1 (cardiac-specific)genes calculated assuming as reference value (set at 1) the number of transcripts ofuntreated ES cells. Bar, standard deviation. *p < 0.05 or **p < 0.001: the expression issignificantly different when compared to ES cells. †p < 0.01 or ††p < 0.001: the expres-sE
5GTAGT
2
cSwwfaflsCgpd
Fig. 2. Frequency of ES-derived cardiomyocytes positive to the immunostainingwith �-actinin, �-myosin and cTnT antibodies after 12 days of EBs differentia-tion. Bar, standard deviation. *: TCDD-treated samples are significantly different
of differentiation using the ATPLite 1 step Luminescence ATP Detection Assay(PerkinElmer), following the manufacturer’s instructions. Briefly, 50,000 cells were
ion is significantly different when TCDD-treated are compared to DMSO or controlBs.
′ TTCCCACGAGTTTTGGAGAC 3′; cTnI, F 5′ GACTTATGCCGACAGCTTCAC 3′ , R 5′
GTCAGATCTGCAATCTCAGTG 3′; ˇ-2-M, F 5′ GAATTCACCCCCACTGAGACT 3′ , R 5′
GCTTGATCACATGTCTCGAT 3′; Brachyury, F 5′ CTCTAAGGAACCACCGGTCA 3′ , R 5′
GCATGGACAGACAAGCAGA 3′; Nkx-2.5, F 5′ GATGGGAAAGCTCCCACTATG 3′; R 5′
AGACACCAGGCTACGTCAATA 3′; Actc-1, F 5′ CCCGAGTTGATTGACTATGGA 3′; R 5′
CTTAGGGATGTCCAGGAACC 3′ .
.5. Microarrays
Biotin-labeled cDNA targets were synthesized starting from 100 ng of total RNA.DNA hybridization was performed using the GeneChip® Hybridization, Wash, andtain Kit. A single GeneChip MOUSE Gene 1.0 ST (Affymetrix) was then hybridisedith each biotin-labeled sense target. Each of the two experimental conditionsas repeated twice, for a total of four GeneChip arrays. Hybridisations were per-
ormed for 16 h at 45 ◦C in a rotisserie oven. GeneChip® cartridges were washednd stained with GeneChip Hybridization, Wash and Stain Kit in the Affymetrixuidics station following the manufacture’s specifications. GeneChip arrays werecanned using an Affymetrix GeneChip Scanner3000 7G. Affymetrix GeneChip
ommand Console software (AGCC) was used to acquire GeneChip® images andenerate .DAT and .CEL files, which were used for subsequent analysis with pro-rietary software. Partek Genomics Suite v 6.4 was used for analysis of microarrayata.compared to Ctrl (p = 0.004) and DMSO (p = 0.009);**: TCDD-treated samples aresignificantly different compared to Ctrl (p = 0.008) and DMSO (p = 0.015); ***: TCDD-treated samples are significantly different compared to Ctrl (p < 0.001) and DMSO(p = 0.002).
2.6. Dissociation of embryoid bodies and immunocytochemistry
Cardiomyocytes for immunofluorescence analysis were isolated from EBs usinga modified protocol described earlier by Maltsev and colleagues (Maltsev et al.,1993). Beating areas of 25–40 EBs at 12 days of differentiation were mechanicallyisolated, washed in low Ca2+-medium (120 mM NaCl, 5.4 mM KCl, 5 mM MgSO4,5 mM sodium pyruvate, 20 mM glucose, 20 mM taurine, 10 mM HEPES/NaOH,pH = 6.9) for 30 min at room temperature and incubated in enzyme medium (lowCa2+-medium supplemented with 1 mg/ml collagenase and 30 �M CaCl2) for 15 minat 37 ◦C. Tissue dissociation was completed by vortexing the suspension for 2 min athigh speed. The isolated cells were centrifuged at 500 rpm for 5 min, resuspendedin culture medium with 100 nmol/l TCDD and seeded on 0.1% gelatin-coated cov-erslips in 24-well culture dishes. After 24 h the attached control and TCDD-treatedcardiomyocytes were fixed with 4% cold paraformaldehyde in PBS for 20 min atroom temperature, washed twice with PBS and then permeabilised with 0.1% Tri-ton X-100 in PBS for 5 min. Immunostaining was performed with an anti-�-myosinheavy chain (MF20, 1:20 dilution; Developmental Studies Hybridoma Bank, IowaCity, Iowa), an anti-sarcomeric �-actinin (1:800; Sigma–Aldrich) or an anti-troponinT (cardiac isoform Ab-1) (1:200; Thermo Scientific) antibody. Cells were incu-bated with primary antibodies for 1 h at 37 ◦C, rinsed three times with PBS andincubated for 1 h at 37 ◦C using a secondary antibody diluted in PBS (AlexaFlour 488-conjugated anti-mouse IgG, 1:500; Molecular Probes). After three washes with PBS,nuclei were counterstained with 0.2 �g/ml DAPI and mounted in antifade (DABCOin Tris–glycerol). Cells were examined with an Olympus Provis fluorescence micro-scope equipped with a cooled-coupled Photometrix CH-350 camera controlled bythe IPLab software used to record digital images, edited with Adobe Photoshop 7.0software. Control experiments with the secondary antibody only were also carriedout.
2.7. Transmission electron microscopy
The beating areas of EBs exposed to TCDD or to its vehicle DMSO cultured for12 days were isolated and gently scraped from the dishes. The samples were thenfixed in 2.5% glutaraldehyde in phosphate buffer (0.12 M, pH 7.4) for 10 min. Afterfixation, the samples were centrifuged at 13000 rpm for 10 min and postfixed in2.5% glutaraldehyde in phosphate buffer and then for 2 h in OsO4. After dehydrationin graded series of ethanol, tissue samples were cleared in propylene oxide, embed-ded in Epoxy medium (Epon 812 Fluka) and polymerised at 60 ◦C for 72 h. Fromeach sample, ultrathin (70 nm thick) sections were obtained with a Leica EM UC6ultramicrotome counterstained with uranyl acetate and lead citrate, and examinedwith an Energy Filter Transmission Electron Microscope (EFTEM, Zeiss Libra 120)equipped with a YAG scintillator slow scan CCD camera.
2.8. ATP quantitation assay
Cellular ATP content was determined in ES-derived cardiomyocytes at 12 days
resuspended in 100 �l of PBS containing 100 nmol/l TCDD, added to 50 �l of cell lysisper well of a 96-well microplate (PerkinElmer), and shaken for 2 min at 700 rpm withan orbital shaker. 50 �l of substrate solution was then added, the plates shaken for2 min at 700 rpm and incubated for 10 min in the dark. Luminescence was measured

T. Neri et al. / Toxicology Letters 202 (2011) 226–236 229
F l (Ctrl)c ruptec tively
wa
2
aawp
3
3c
o
ig. 3. Immunocytochemical localisation of �-actinin, �-myosin and cTnT in controharacteristic striated organisation (arrows and insets) found in Ctrl and DMSO is disonfocal sections of �-actinin localisation in Ctrl and TCDD-treated samples, respec
ith a luminometer (GENios Plus, Tecan). Each sample was measured in triplicatend the experiment was repeated three times.
.9. Statistical analysis
Experiments were independently replicated at least three times. For RT-PCRnalysis, at least two repeats of the same sample were analysed. Data are presenteds means ± standard deviation (SD). Statistical significance was determined by one-ay ANOVA for multiple groups with post hoc testing using Fisher’s LSD method. A< 0.05 value was considered statistically significant.
. Results
.1. Pattern of expression of mesoderm-, cardiac mesoderm- andardiac-specific genes
The process of cardiomyocyte differentiation was investigatedn day 4, 7 and 12 of EBs culture in the presence of TCDD and in con-
, DMSO or TCDD-treated ES-derived cardiomyocytes counterstained with DAPI. Thed in TCDD-treated cardiomyocytes (arrowheads and insets). b’ and c’, representative. Bar, 10 �m.
trol samples, analysing by real-time RT-PCR the expression of genemarkers of mesoderm (Brachyury), cardiac mesoderm (Nkx-2.5) andcardiac cells (Actc-1) differentiation (Fig. 1). The expression of thethree genes in ES cells cultured without TCDD for 2 days was usedas reference value (set at 1) for calculating the n-fold change duringEBs differentiation in the presence or absence of TCDD. After 4 daysof EBs differentiation, the expression of Brachyury dramaticallyincreased, up to more than 125-fold, in all the three experimen-tal conditions and then it decreases to basal levels after 12 days ofdifferentiation. At 7 days of differentiation the TCDD-treated sam-ples showed a more evident decrease compared to both controls
(p < 0.001). The expression of both Nkx-2.5 and Actc-1 genes wasup-regulated starting from 7 days of differentiation; then whilstthe former was down-regulated (although maintaining a significantdifferent expression compared to ES cells), the latter continued toincrease. At 12 days of differentiation, the expression of Actc-1 in
230 T. Neri et al. / Toxicology Letters 202 (2011) 226–236
Fig. 4. ES-derived cardiomyocytes obtained after 12 days of culture without (a) and (b) or with (c) and (d) TCDD. (a) Control-DMSO cardiomyocytes show organised sarcomeresc H bana fascia( eric pa ith tw
Tb
3
3ˇ
ed
Fdtlts
haracterised by parallel myofilaments anchored to Z bands (white arrowheads) andre perfectly aligned and packed (Bar, 1 �m). (b) Desmosome junction (arrows) andc) Few interrupted myofilaments are slightly orientated without any sign of sarcomnd show disarray of myofilaments (arrowheads); mitochondria are disorganised w
CDD-treated samples was significantly lower when compared tooth control and DMSO samples (p < 0.01).
.2. Morphological analysis of ES-derived cardiomyocytes
.2.1. Immunocytochemical assessment of sarcomeric ˛-actinin,-myosin heavy chain and cardiac troponin T proteins
The immunocytochemical analysis, carried out to evaluate theffects of TCDD on the efficiency of cardiomyocyte differentiation,emonstrated a lower frequency of TCDD-treated cardiomyocytes
ig. 5. Relative amount of ATP content in control, DMSO and TCDD-treated ES-erived cardiomyocytes after 12 days of EBs differentiation. The histogram showshe fold-change calculated assuming as reference value (set at 1) the cellular ATPevel of Ctrl. Bar, standard deviation. *: The amount of cellular ATP level in TCDD-reated is significantly lower compared to Ctrl (p = 0.02) and DMSO (p = 0.004)amples.
ds (black arrows); mitochondria (white arrows) in close association with myofibrilsadherens (arrowheads) were observed in ES-derived cardiomyocytes (Bar, 500 nm).attern, only one Z band (arrows) is visible (Bar, 1 �m). (d) Myofibrils are misalignedisted and disrupted cristae (arrows) (Bar, 1 �m).
positive to �-actinin, �-myosin and cTnT proteins compared toCtrl and DMSO samples (Fig. 2 and Table 1S). Furthermore, TCDD-treated cardiomyocytes positive for �-actinin, �-myosin or cTnTproteins were characterised by a disruption of these myofibrillarcomponents. In fact, whilst ES-derived cardiomyocytes, obtainedafter enzymatic dispersion of EBs cultured for 12 days, showed thestriated cardiomyocyte-specific pattern of �-actinin, �-myosin andcTnT distribution in Ctrl (Fig. 3a, d and s) and DMSO (Fig. 3g, j andv) samples, following TCDD treatment, the antibody still revealedthe presence of the protein, but the characteristic striated con-figuration was lost (�-actinin, Fig. 3m; �-myosin, 3p and cTnT,3y).
3.2.2. Ultrastructural characteristics of ES-derivedcardiomyocytes
A detailed ultrastructural characterisation was carried out incontrol-DMSO and TCDD-treated ES-derived cardiomyocytes after12 days of culture. Control cardiomyocytes (Fig. 4a) showed organ-ised sarcomeres characterised by myofilaments ordered in parallelto the long axis of the cell and anchored to Z bands; H bandsare also clearly visible. Mitochondria are in close association withmyofibrils and perfectly aligned and packed. Intercalated diskscomposed of desmosome junctions and fascia adherens are also vis-
ible between cardiomyocytes (Fig. 4b). TCDD-treated ES-derivedcardiomyocytes contained both normally organised or disruptedmyofibrils that lacked alignment and were distributed through-out the cytoplasm in a disoriented manner (Fig. 4c) or, in somecases, they revealed the presence of Z and H bands, but with dis-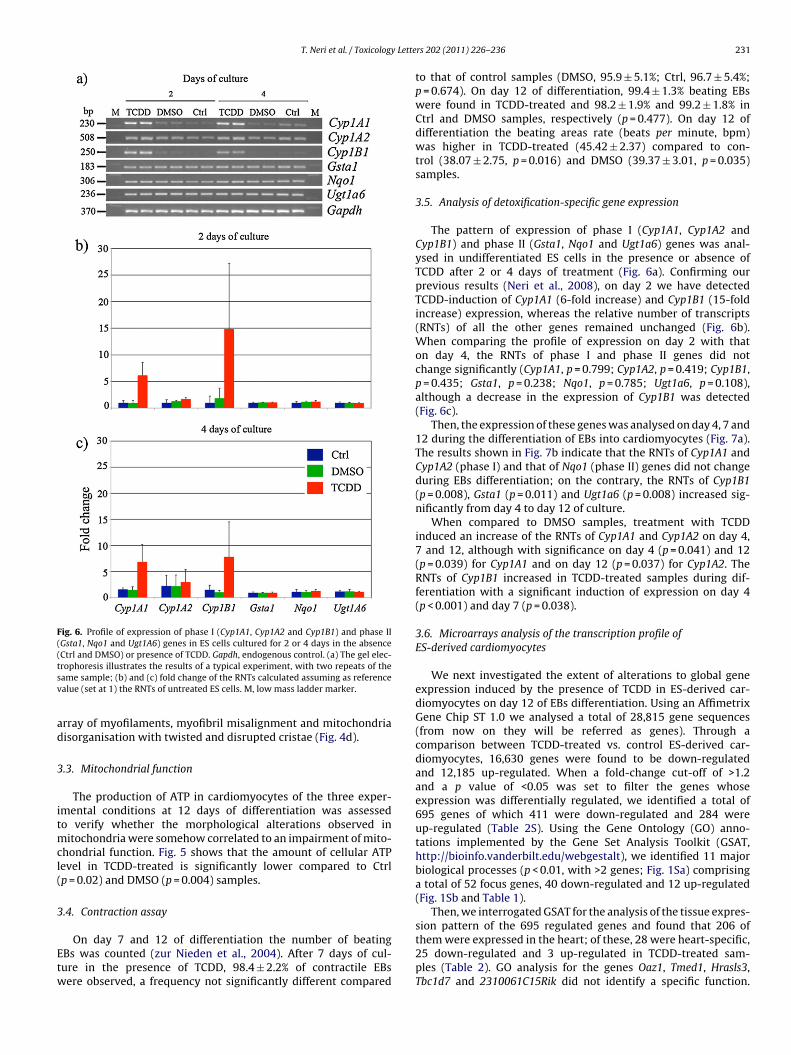
T. Neri et al. / Toxicology Lette
Fig. 6. Profile of expression of phase I (Cyp1A1, Cyp1A2 and Cyp1B1) and phase II(Gsta1, Nqo1 and Ugt1A6) genes in ES cells cultured for 2 or 4 days in the absence(tsv
ad
3
itmcl(
3
Etw
Ctrl and DMSO) or presence of TCDD. Gapdh, endogenous control. (a) The gel elec-rophoresis illustrates the results of a typical experiment, with two repeats of theame sample; (b) and (c) fold change of the RNTs calculated assuming as referencealue (set at 1) the RNTs of untreated ES cells. M, low mass ladder marker.
rray of myofilaments, myofibril misalignment and mitochondriaisorganisation with twisted and disrupted cristae (Fig. 4d).
.3. Mitochondrial function
The production of ATP in cardiomyocytes of the three exper-mental conditions at 12 days of differentiation was assessedo verify whether the morphological alterations observed in
itochondria were somehow correlated to an impairment of mito-hondrial function. Fig. 5 shows that the amount of cellular ATPevel in TCDD-treated is significantly lower compared to Ctrlp = 0.02) and DMSO (p = 0.004) samples.
.4. Contraction assay
On day 7 and 12 of differentiation the number of beatingBs was counted (zur Nieden et al., 2004). After 7 days of cul-ure in the presence of TCDD, 98.4 ± 2.2% of contractile EBsere observed, a frequency not significantly different compared
rs 202 (2011) 226–236 231
to that of control samples (DMSO, 95.9 ± 5.1%; Ctrl, 96.7 ± 5.4%;p = 0.674). On day 12 of differentiation, 99.4 ± 1.3% beating EBswere found in TCDD-treated and 98.2 ± 1.9% and 99.2 ± 1.8% inCtrl and DMSO samples, respectively (p = 0.477). On day 12 ofdifferentiation the beating areas rate (beats per minute, bpm)was higher in TCDD-treated (45.42 ± 2.37) compared to con-trol (38.07 ± 2.75, p = 0.016) and DMSO (39.37 ± 3.01, p = 0.035)samples.
3.5. Analysis of detoxification-specific gene expression
The pattern of expression of phase I (Cyp1A1, Cyp1A2 andCyp1B1) and phase II (Gsta1, Nqo1 and Ugt1a6) genes was anal-ysed in undifferentiated ES cells in the presence or absence ofTCDD after 2 or 4 days of treatment (Fig. 6a). Confirming ourprevious results (Neri et al., 2008), on day 2 we have detectedTCDD-induction of Cyp1A1 (6-fold increase) and Cyp1B1 (15-foldincrease) expression, whereas the relative number of transcripts(RNTs) of all the other genes remained unchanged (Fig. 6b).When comparing the profile of expression on day 2 with thaton day 4, the RNTs of phase I and phase II genes did notchange significantly (Cyp1A1, p = 0.799; Cyp1A2, p = 0.419; Cyp1B1,p = 0.435; Gsta1, p = 0.238; Nqo1, p = 0.785; Ugt1a6, p = 0.108),although a decrease in the expression of Cyp1B1 was detected(Fig. 6c).
Then, the expression of these genes was analysed on day 4, 7 and12 during the differentiation of EBs into cardiomyocytes (Fig. 7a).The results shown in Fig. 7b indicate that the RNTs of Cyp1A1 andCyp1A2 (phase I) and that of Nqo1 (phase II) genes did not changeduring EBs differentiation; on the contrary, the RNTs of Cyp1B1(p = 0.008), Gsta1 (p = 0.011) and Ugt1a6 (p = 0.008) increased sig-nificantly from day 4 to day 12 of culture.
When compared to DMSO samples, treatment with TCDDinduced an increase of the RNTs of Cyp1A1 and Cyp1A2 on day 4,7 and 12, although with significance on day 4 (p = 0.041) and 12(p = 0.039) for Cyp1A1 and on day 12 (p = 0.037) for Cyp1A2. TheRNTs of Cyp1B1 increased in TCDD-treated samples during dif-ferentiation with a significant induction of expression on day 4(p < 0.001) and day 7 (p = 0.038).
3.6. Microarrays analysis of the transcription profile ofES-derived cardiomyocytes
We next investigated the extent of alterations to global geneexpression induced by the presence of TCDD in ES-derived car-diomyocytes on day 12 of EBs differentiation. Using an AffimetrixGene Chip ST 1.0 we analysed a total of 28,815 gene sequences(from now on they will be referred as genes). Through acomparison between TCDD-treated vs. control ES-derived car-diomyocytes, 16,630 genes were found to be down-regulatedand 12,185 up-regulated. When a fold-change cut-off of >1.2and a p value of <0.05 was set to filter the genes whoseexpression was differentially regulated, we identified a total of695 genes of which 411 were down-regulated and 284 wereup-regulated (Table 2S). Using the Gene Ontology (GO) anno-tations implemented by the Gene Set Analysis Toolkit (GSAT,http://bioinfo.vanderbilt.edu/webgestalt), we identified 11 majorbiological processes (p < 0.01, with >2 genes; Fig. 1Sa) comprisinga total of 52 focus genes, 40 down-regulated and 12 up-regulated(Fig. 1Sb and Table 1).
Then, we interrogated GSAT for the analysis of the tissue expres-
sion pattern of the 695 regulated genes and found that 206 ofthem were expressed in the heart; of these, 28 were heart-specific,25 down-regulated and 3 up-regulated in TCDD-treated sam-ples (Table 2). GO analysis for the genes Oaz1, Tmed1, Hrasls3,Tbc1d7 and 2310061C15Rik did not identify a specific function.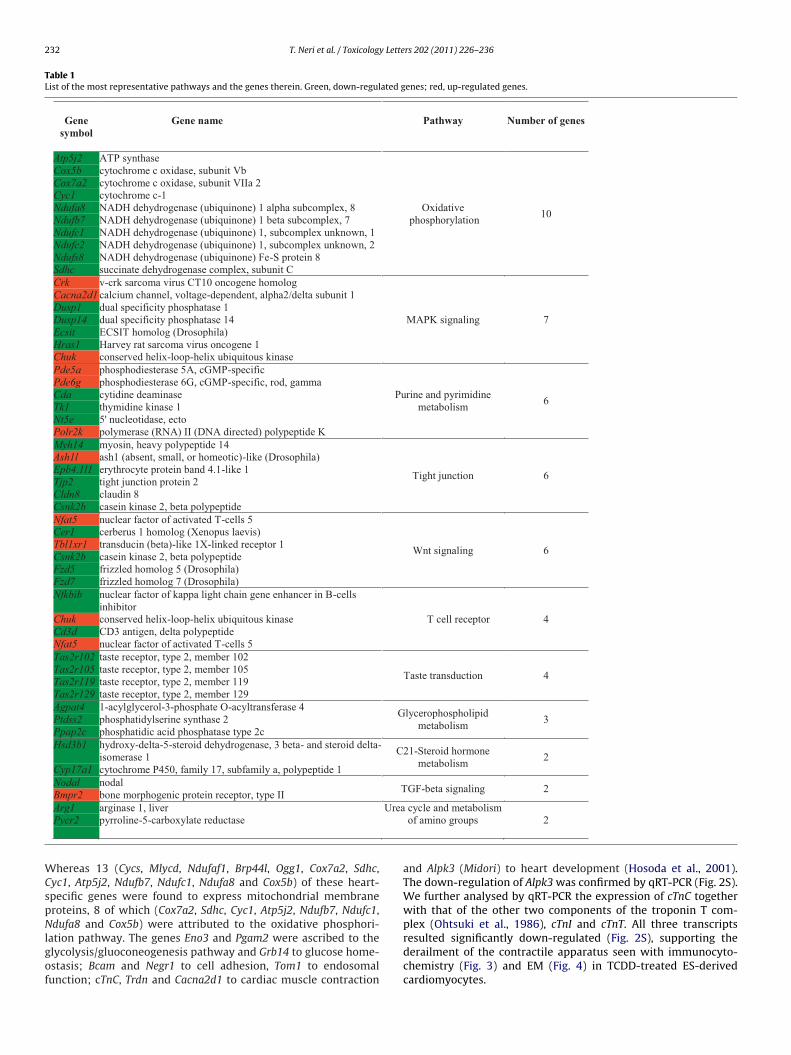
232 T. Neri et al. / Toxicology Letters 202 (2011) 226–236
Table 1List of the most representative pathways and the genes therein. Green, down-regulated genes; red, up-regulated genes.
Gene symbol
Gene name Pathway Number of genes
Atp5j2 ATP synthase
Oxidativephosphorylation 10
Cox5b cytochrome c oxidase, subunit VbCox7a2 cytochrome c oxidase, subunit VIIa 2Cyc1 cytochrome c-1Ndufa8 NADH dehydrogenase (ubiquinone) 1 alpha subcomplex, 8Ndufb7 NADH dehydrogenase (ubiquinone) 1 beta subcomplex, 7Ndufc1 NADH dehydrogenase (ubiquinone) 1, subcomplex unknown, 1Ndufc2 NADH dehydrogenase (ubiquinone) 1, subcomplex unknown, 2Ndufs8 NADH dehydrogenase (ubiquinone) Fe-S protein 8Sdhc succinate dehydrogenase complex, subunit CCrk v-crk sarcoma virus CT10 oncogene homolog
MAPK signaling 7
Cacna2d1 calcium channel, voltage-dependent, alpha2/delta subunit 1Dusp1 dual specificity phosphatase 1Dusp14 dual specificity phosphatase 14Ecsit ECSIT homolog (Drosophila)Hras1 Harvey rat sarcoma virus oncogene 1Chuk conserved helix-loop-helix ubiquitous kinasePde5a phosphodiesterase 5A, cGMP-specific
Purine and pyrimidine metabolism 6
Pde6g phosphodiesterase 6G, cGMP-specific, rod, gammaCda cytidine deaminaseTk1 thymidine kinase 1Nt5e 5' nucleotidase, ectoPolr2k polymerase (RNA) II (DNA directed) polypeptide KMyh14 myosin, heavy polypeptide 14
Tight junction 6
Ash1l ash1 (absent, small, or homeotic)-like (Drosophila)Epb4.1l1 erythrocyte protein band 4.1-like 1Tjp2 tight junction protein 2Cldn8 claudin 8Csnk2b casein kinase 2, beta polypeptideNfat5 nuclear factor of activated T-cells 5
Wnt signaling 6
Cer1 cerberus 1 homolog (Xenopus laevis)Tbl1xr1 transducin (beta)-like 1X-linked receptor 1Csnk2b casein kinase 2, beta polypeptideFzd5 frizzled homolog 5 (Drosophila)Fzd7 frizzled homolog 7 (Drosophila)Nfkbib nuclear factor of kappa light chain gene enhancer in B-cells
inhibitorT cell receptor 4Chuk conserved helix-loop-helix ubiquitous kinase
Cd3d CD3 antigen, delta polypeptideNfat5 nuclear factor of activated T-cells 5Tas2r102 taste receptor, type 2, member 102
Taste transduction 4Tas2r105 taste receptor, type 2, member 105Tas2r119 taste receptor, type 2, member 119Tas2r129 taste receptor, type 2, member 129Agpat4 1-acylglycerol-3-phosphate O-acyltransferase 4 Glycerophospholipid
metabolism 3Ptdss2 phosphatidylserine synthase 2Ppap2c phosphatidic acid phosphatase type 2cHsd3b1 hydroxy-delta-5-steroid dehydrogenase, 3 beta- and steroid delta-
isomerase 1 C21-Steroid hormone metabolism 2
Cyp17a1 cytochrome P450, family 17, subfamily a, polypeptide 1Nodal nodal TGF-beta signaling 2Bmpr2 bone morphogenic protein receptor, type II
Urea
WCspNlgof
Arg1Pycr2
arginase 1, liverpyrroline-5-carboxylate reductase
hereas 13 (Cycs, Mlycd, Ndufaf1, Brp44l, Ogg1, Cox7a2, Sdhc,yc1, Atp5j2, Ndufb7, Ndufc1, Ndufa8 and Cox5b) of these heart-pecific genes were found to express mitochondrial membraneroteins, 8 of which (Cox7a2, Sdhc, Cyc1, Atp5j2, Ndufb7, Ndufc1,
dufa8 and Cox5b) were attributed to the oxidative phosphori-ation pathway. The genes Eno3 and Pgam2 were ascribed to thelycolysis/gluoconeogenesis pathway and Grb14 to glucose home-stasis; Bcam and Negr1 to cell adhesion, Tom1 to endosomalunction; cTnC, Trdn and Cacna2d1 to cardiac muscle contraction
cycle and metabolism of amino groups 2
and Alpk3 (Midori) to heart development (Hosoda et al., 2001).The down-regulation of Alpk3 was confirmed by qRT-PCR (Fig. 2S).We further analysed by qRT-PCR the expression of cTnC togetherwith that of the other two components of the troponin T com-
plex (Ohtsuki et al., 1986), cTnI and cTnT. All three transcriptsresulted significantly down-regulated (Fig. 2S), supporting thederailment of the contractile apparatus seen with immunocyto-chemistry (Fig. 3) and EM (Fig. 4) in TCDD-treated ES-derivedcardiomyocytes.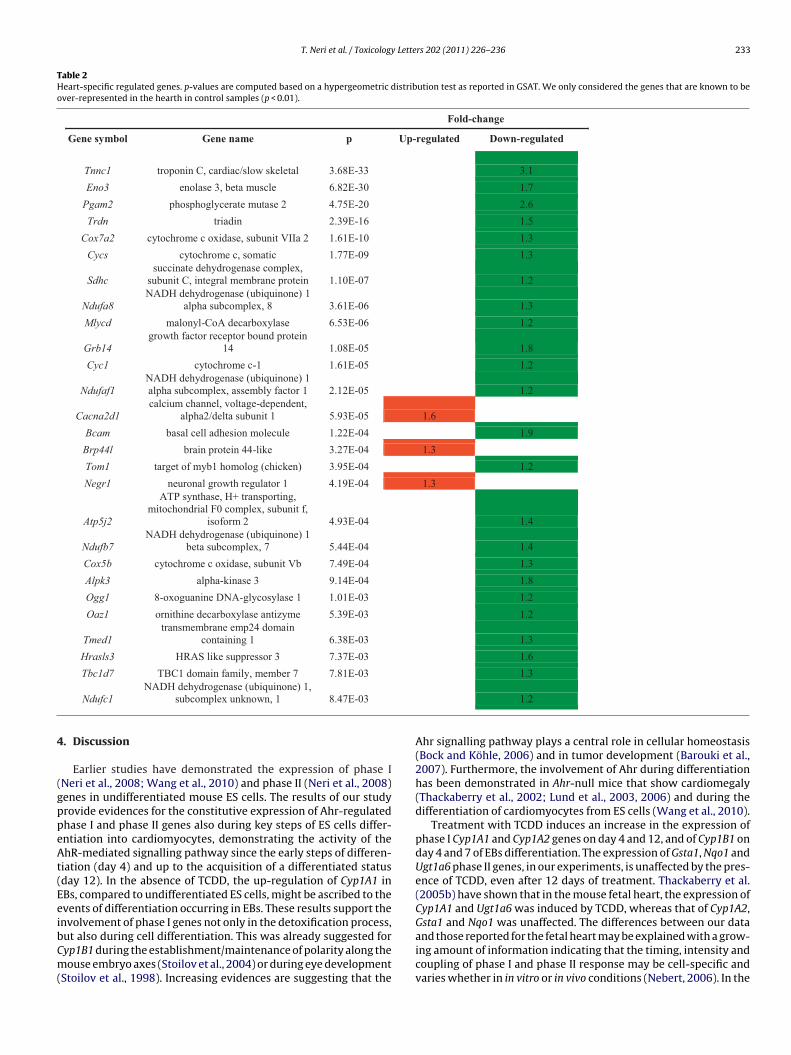
T. Neri et al. / Toxicology Letters 202 (2011) 226–236 233
Table 2Heart-specific regulated genes. p-values are computed based on a hypergeometric distribution test as reported in GSAT. We only considered the genes that are known to beover-represented in the hearth in control samples (p < 0.01).
Fold-change
Gene symbol Gene name p Up-regulated Down-regulated
Tnnc1 troponin C, cardiac/slow skeletal 3.68E-33 3.1Eno3 enolase 3, beta muscle 6.82E-30 1.7Pgam2 phosphoglycerate mutase 2 4.75E-20 2.6Trdn triadin 2.39E-16 1.5Cox7a2 cytochrome c oxidase, subunit VIIa 2 1.61E-10 1.3Cycs cytochrome c, somatic 1.77E-09 1.3
Sdhcsuccinate dehydrogenase complex,
subunit C, integral membrane protein 1.10E-07 1.2
Ndufa8NADH dehydrogenase (ubiquinone) 1
alpha subcomplex, 8 3.61E-06 1.3Mlycd malonyl-CoA decarboxylase 6.53E-06 1.2
Grb14growth factor receptor bound protein
14 1.08E-05 1.8Cyc1 cytochrome c-1 1.61E-05 1.2
Ndufaf1NADH dehydrogenase (ubiquinone) 1 alpha subcomplex, assembly factor 1 2.12E-05 1.2
Cacna2d1calcium channel, voltage-dependent,
alpha2/delta subunit 1 5.93E-05 1.6Bcam basal cell adhesion molecule 1.22E-04 1.9Brp44l brain protein 44-like 3.27E-04 1.3Tom1 target of myb1 homolog (chicken) 3.95E-04 1.2Negr1 neuronal growth regulator 1 4.19E-04 1.3
Atp5j2
ATP synthase, H+ transporting, mitochondrial F0 complex, subunit f,
isoform 2 4.93E-04 1.4
Ndufb7NADH dehydrogenase (ubiquinone) 1
beta subcomplex, 7 5.44E-04 1.4Cox5b cytochrome c oxidase, subunit Vb 7.49E-04 1.3Alpk3 alpha-kinase 3 9.14E-04 1.8Ogg1 8-oxoguanine DNA-glycosylase 1 1.01E-03 1.2Oaz1 ornithine decarboxylase antizyme 5.39E-03 1.2
Tmed1transmembrane emp24 domain
containing 1 6.38E-03 1.3Hrasls3 HRAS like suppressor 3 7.37E-03 1.6
4
(gppeAt(EeibCm(
Tbc1d7 TBC1 domain family, member 7 7.81E-03
Ndufc1NADH dehydrogenase (ubiquinone) 1,
subcomplex unknown, 1 8.47E-03
. Discussion
Earlier studies have demonstrated the expression of phase INeri et al., 2008; Wang et al., 2010) and phase II (Neri et al., 2008)enes in undifferentiated mouse ES cells. The results of our studyrovide evidences for the constitutive expression of Ahr-regulatedhase I and phase II genes also during key steps of ES cells differ-ntiation into cardiomyocytes, demonstrating the activity of thehR-mediated signalling pathway since the early steps of differen-
iation (day 4) and up to the acquisition of a differentiated statusday 12). In the absence of TCDD, the up-regulation of Cyp1A1 inBs, compared to undifferentiated ES cells, might be ascribed to thevents of differentiation occurring in EBs. These results support the
nvolvement of phase I genes not only in the detoxification process,ut also during cell differentiation. This was already suggested foryp1B1 during the establishment/maintenance of polarity along theouse embryo axes (Stoilov et al., 2004) or during eye developmentStoilov et al., 1998). Increasing evidences are suggesting that the
1.3
1.2
Ahr signalling pathway plays a central role in cellular homeostasis(Bock and Köhle, 2006) and in tumor development (Barouki et al.,2007). Furthermore, the involvement of Ahr during differentiationhas been demonstrated in Ahr-null mice that show cardiomegaly(Thackaberry et al., 2002; Lund et al., 2003, 2006) and during thedifferentiation of cardiomyocytes from ES cells (Wang et al., 2010).
Treatment with TCDD induces an increase in the expression ofphase I Cyp1A1 and Cyp1A2 genes on day 4 and 12, and of Cyp1B1 onday 4 and 7 of EBs differentiation. The expression of Gsta1, Nqo1 andUgt1a6 phase II genes, in our experiments, is unaffected by the pres-ence of TCDD, even after 12 days of treatment. Thackaberry et al.(2005b) have shown that in the mouse fetal heart, the expression ofCyp1A1 and Ugt1a6 was induced by TCDD, whereas that of Cyp1A2,
Gsta1 and Nqo1 was unaffected. The differences between our dataand those reported for the fetal heart may be explained with a grow-ing amount of information indicating that the timing, intensity andcoupling of phase I and phase II response may be cell-specific andvaries whether in in vitro or in vivo conditions (Nebert, 2006). In the
234 T. Neri et al. / Toxicology Letters 202 (2011) 226–236
F ta1, N( eats ov ass ladt
aC
cadcrtPfdetdd
eo
ig. 7. Profile of expression of phase I (Cyp1A1, Cyp1A2 and Cyp1B1) and phase II (Gsa) The gel electrophoresis shows the results of a typical experiment, with two repalue (set at 1) the RNTs of untreated ES cells. M, low mass ladder marker. M, low mranscriptase but no RNA; PCR blank.
bsence of phase II genes activation, as in our TCDD-treated cells,yp1A1, Cyp1A2 and Cyp1B1 may be competent to detoxification.
In this study we give evidence of a direct effect of TCDD on theellular organisation of cardiomyocytes derived from ES cells. Theyre a robust model of in vitro cardiomyocyte differentiation; whilstifferentiating, they express cardiac genes in a developmentallyontrolled manner. In our experiments, the transcription profileeported for Brachyury, Nkx-2.5 and Actc-1 well describe the majorime-points of this pathway of differentiation (Boheler et al., 2002;ucéat, 2008). Brachyury (a marker of early mesoderm, vital for theormation and differentiation of posterior mesoderm and for axialevelopment in all vertebrates), as expected, shows a decreasingxpression, whereas that of Nkx-2.5 (a recognised transcription fac-or during vertebrate embryonic heart development) peaks after 7
ays and that of Actc-1 (marker of cardiac cells) increases duringifferentiation from 4 to 12 days.Addition of TCDD to the culture medium did not alter thexpression profile of these three genes, although a lower profilef expression was observed at 7 days for Brachyury and at 12 days
qo1 and Ugt1A6) genes at 4, 7 and 12 days of embryoid bodies (EBs) differentiation.f the same sample; (b) fold change of the RNTs calculated assuming as referenceder marker;∅1, RT blank without reverse transcriptase;∅2, RT blank with reverse
of differentiation for the Actc-1 gene. In contrast to our data, in car-diomyocytes differentiated from mouse ES cells, Wang et al. (2010)have found down-regulation of Nkx-2.5, a homeobox transcriptionfactor regulator of genes in the cardiomyocyte lineage and a markerof cardiac progenitor cells (Yano et al., 2008). The lack of a down-regulative effect of TCDD on the expression of Nkx-2.5 (our results)may explain the higher number of differentiated cardiomyocyteswith a beating phenotype compared to that obtained by Wanget al. (2010). These contrasting results perhaps highlight a differentresponse of the low ligand-affinity Ah receptor ES cell line used inour study compared to the high ligand-affinity line used by Wanget al. (2010).
Although the process of ES cells differentiation into cardiomy-ocytes with a beating phenotype is not inhibited, a reduced number
of differentiated cells was observed in TCDD-treated comparedto control samples. This reduction may be explained with thedown-regulation of Alpk3 (Midori) (as determined by both ourmicroarrays analysis and qRT-PCR at 12 days of differentiation)which over-expression has been shown to promote more effi-
Lette
cideclocfmomTomlo
sacadlrbtcdtwaOicim
oaTsAae
edtewaaoa
liaiE
C
T. Neri et al. / Toxicology
iently the differentiation of P19CL6 embryonal carcinoma cellsnto cardiomyocytes (Hosoda et al., 2001) and perhaps also by theown-regulation of Brachyury at 7 days of differentiation. The pres-nce of TCDD clearly has disruptive consequences on the cellularomponents that characterise cardiomyocytes. At the molecularevel, we described a down-regulated transcription of Actc-1 andf the three components (cTnC, cTnI and cTnT) of the troponinomplex, localised along the thin filaments and regulating theorce-generating interaction of myosin heads with the thin fila-
ents. At the cytological level, the immunocytochemical analysisf the antibodies against cTnT and also against �-actinin and �-yosin showed a disrupted pattern of localisation of these proteins.
he ultrastructural analysis confirmed an altered organisationf some of the sarcomeres, with significant myofibrils misalign-ent or complete disarrangement of the myofibrillar organisation
eading to an alteration of the characteristic striated phenotypebserved in control samples.
The EM observation of ES-derived cardiomyocytes has alsohown mitochondria which organisation and structure wereffected by TCDD. The observation of twisted and disruptedristae in TCDD-treated ES-derived cardiomyocytes, suggests anlteration of mitochondria functionality, supported by both theown-regulation of genes involved in the oxidative phosphori-
ation pathway (our microarrays analysis) and by the observededuced ATP levels. The metabolic deficiency might be responsi-le for the impaired contractile apparatus, as it has been shownhat an alteration of the respiratory chain function prevents mito-hondrial organisation, causes energetic deficiency and thereforeetermines defective sarcomerogenesis and alterations to contrac-ility (Chung et al., 2007). In our ES-derived cardiomyocytes treatedith TCDD, the presence of a fair amount of intact sarcomeres,
longside damaged sarcomeres, ensures the beating of these cells.n the other hand, the structural disorganization of sarcomeres
nduced by TCDD might account for an asynchronous and less effi-ient contractility responsible of the higher beating rate observedn our study, also reported by Nishii et al. (2008) in cTnT−/−
ice.Although molecular and morphological alterations were
bserved on cardiomyocytes at an advanced phase of differenti-tion, they are likely the result of deregulative effects induced byCDD at earlier stages. The TCDD-induced alteration of the expres-ion of a group of homeobox genes, whose promoters containhr-binding sites, might be crucial in determining cell differenti-tion abnormalities, as early as 4 days from EBs formation (Wangt al., 2010).
In summary, in this study we have shown: (1) a constitutivexpression of Ahr-regulated phase I and phase II genes during car-iomyocyte differentiation and the induction of phase I genes inhe presence of TCDD; (2) a TCDD-induced down-regulation of thexpression of a group of genes expressed in the heart, some ofhich are involved in the oxidative phosphorylation pathway; (3)TCDD-induced alteration of the efficiency of ES cells differenti-
tion and (4) of the characteristic myofibrillar and mitochondriarganisation; (5) an altered energetic mitochondrial metabolismnd cardiomyocytes contractility.
The results of our study and those reported in a recent pub-ication (Wang et al., 2010) open the opportunity to study, on ann vitro model of cardiomyocytes differentiation, the cellular mech-nisms and signalling pathways known to play a major role in thenduction of environmental cardiovascular diseases (Korashy andl-Kadi, 2006).
onflict of interest statement
The authors declare that there are no conflicts of interest.
rs 202 (2011) 226–236 235
Acknowledgments
This work was supported by: Fondazione Banca del Monte diLombardia contributo per la ricerca 2009, Fondazione I.R.C.C.S.Policlinico San Matteo di Pavia, Regione Lombardia, Alma MaterTicinensis and ‘Bando Giovani Ricercatori 2007’ to C.A.R. “The(hybridoma or monoclonal antibody) developed by Fischman D.A.was obtained from the Developmental Studies Hybridoma Bankdeveloped under the auspices of the NICHD and maintained by TheUniversity of Iowa, Department of Biology, Iowa City, IA 52242”.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.toxlet.2011.02.008.
References
Antkiewicz, D., Burns, C., Carney, S., Peterson, R., Heideman, W., 2005. Heart mal-formation is an early response to TCDD in embryonic zebrafish. Toxicol. Sci. 84,368–377.
Arzuaga, X., Reiterer, G., Majkova, Z., Kilgore, M., Toborek, M., Hennig, B., 2007. PPA-Ralpha ligands reduce PCB-induced endothelial activation: possible interactionsin inflammation and atherosclerosis. Cardiovasc. Toxicol. 7, 264–272.
Barouki, R., Coumoul, X., Fernandez-Salguero, P.M., 2007. The aryl hydrocarbonreceptor, more than a xenobiotic-interacting protein. FEBS Lett. 581, 3608–3615.
Bock, K.W., Köhle, C., 2006. Ah receptor: dioxin-mediated toxic responses as hintsto deregulated physiologic functions. Biochem. Pharmacol. 72, 393–404.
Boheler, K., Czyz, J., Tweedie, D., Yang, H., Anisimov, S., Wobus, A., 2002. Differen-tiation of pluripotent embryonic stem cells into cardiomyocytes. Circ. Res. 91,189–201.
Chung, S., Dzeja, P.P., Faustino, R.S., Perez-Terzic, C., Behfar, A., Terzic, A., 2007. Mito-chondrial oxidative metabolism is required for the cardiac differentiation ofstem cells. Nat. Clin. Pract. Cardiovasc. Med. 4 (Suppl. 1), S60–67.
Dalton, T., Kerzee, J., Wang, B., Miller, M., Dieter, M., Lorenz, J., Shertzer, H., Nerbert,D., Puga, A., 2001. Dioxin exposure is an environmental risk factor for ischemicheart disease. Cardiovasc. Toxicol. 1, 285–298.
Goldstone, H., Stegeman, J., 2006. Molecular mechanisms of 2,3,7,8-tetrachlorodibenzo-p-dioxin cardiovascular embryotoxicity. Drug Metab.Rev. 38, 261–289.
Hosoda, T., Monzen, K., Hiroi, Y., Oka, T., Takimoto, E., Yazaki, Y., Nagai, R., Komuro,I., 2001. A novel myocyte-specific gene Midori promotes the differentiation ofP19CL6 cells into cardiomyocytes. J. Biol. Chem. 276, 35978–35989.
Ivnitski, I., Elmaoued, R., Walker, M., 2001. 2,3,7,8-Tetrachlorodibenzo-p-dioxin(TCDD) inhibition of coronary development is preceded by a decrease in myocyteproliferation and an increase in cardiac apoptosis. Teratology 64, 201–212.
Jokinen, M., Walker, N., Brix, A., Sells, D., Haseman, J., Nyska, A., 2003.Increase in cardiovascular pathology in female Sprague–Dawley rats followingchronic treatment with 2,3,7,8-tetrachlorodibenzo-p-dioxin and 3,3′ ,4,4′ ,5-pentachlorobiphenyl. Cardiovasc. Toxicol. 3, 299–310.
Kehat, I., Kenyagin-Karsenti, D., Snir, M., Segev, H., Amit, M., Gepstein, A., Livne, E.,Binah, O., Itskovitz-Eldor, J., Gepstein, L., 2001. Human embryonic stem cellscan differentiate into myocytes with structural and functional properties ofcardiomyocytes. J. Clin. Invest. 108, 407–414.
Kelling, C., Menahan, L., Peterson, R., 1987. Effects of 2,3,7,8-tetrachlorodibenzo-p-dioxin treatment on mechanical function of the rat heart. Toxicol. Appl.Pharmacol. 91, 497–501.
Kopf, P., Huwe, J., Walker, M., 2008. Hypertension, cardiac hypertrophy, andimpaired vascular relaxation induced by 2,3,7,8-tetrachlorodibenzo-p-dioxinare associated with increased superoxide. Cardiovasc. Toxicol. 8, 181–193.
Korashy, H., El-Kadi, A., 2006. The role of aryl hydrocarbon receptor in the patho-genesis of cardiovascular diseases. Drug Metab. Rev. 38, 411–450.
Lind, P., Orberg, J., Edlund, U., Sjöblom, L., Lind, L., 2004. The dioxin-like pollutantPCB 126 (3,3′ ,4,4′ ,5-pentachlorobiphenyl) affects risk factors for cardiovasculardisease in female rats. Toxicol. Lett. 150, 293–299.
Lund, A., Goens, M., Kanagy, N., Walker, M., 2003. Cardiac hypertrophy in arylhydrocarbon receptor null mice is correlated with elevated angiotensin II,endothelin-1, and mean arterial blood pressure. Toxicol. Appl. Pharmacol. 193,177–187.
Lund, A., Goens, M., Nunez, B., Walker, M., 2006. Characterizing the role ofendothelin-1 in the progression of cardiac hypertrophy in aryl hydrocarbonreceptor (AhR) null mice. Toxicol. Appl. Pharmacol. 212, 127–135.
Lund, A., Peterson, S., Timmins, G., Walker, M., 2005. Endothelin-1-mediatedincrease in reactive oxygen species and NADPH oxidase activity in hearts of
aryl hydrocarbon receptor (AhR) null mice. Toxicol. Sci. 88, 265–273.Maltsev, V., Rohwedel, J., Hescheler, J., Wobus, A., 1993. Embryonic stem cellsdifferentiate in vitro into cardiomyocytes representing sinusnodal, atrial andventricular cell types. Mech. Dev. 44, 41–50.
Mandal, P., 2005. Dioxin: a review of its environmental effects and its aryl hydro-carbon receptor biology. J. Comp. Physiol. B 175, 221–230.

2 y Lette
N
N
N
N
O
P
S
S
S
S
36 T. Neri et al. / Toxicolog
ebert, D., 2006. Comparison of gene expression in cell culture to that in the intactanimal: relevance to drugs and environmental toxicants. Focus on “developmentof a transactivator in hepatoma cells that allows expression of phase I, phase II,and chemical defense genes”. Am. J. Physiol. Cell Physiol. 290, C37–41.
ebert, D., Roe, A., Dieter, M., Solis, W., Yang, Y., Dalton, T., 2000. Role of the aromatichydrocarbon receptor and [Ah] gene battery in the oxidative stress response, cellcycle control, and apoptosis. Biochem. Pharmacol. 59, 65–85.
eri, T., Merico, V., Garagna, S., Redi, C., Zuccotti, M., 2008. Expression of phase Iand phase II genes in mouse embryonic stem cells cultured in the presence of2,3,7,8-tetrachlorodibenzo-para-dioxin. Biochim. Biophys. Acta 1780, 826–836.
ishii, K., Morimoto, S., Minakami, R., Miyano, Y., Hashizume, K., Ohta, M., Zhan, D.Y.,Lu, Q.W., Shibata, Y., 2008. Targeted disruption of the cardiac troponin T genecauses sarcomere disassembly and defects in heartbeat within the early mouseembryo. Dev. Biol. 322, 65–73.
htsuki, I., Maruyama, K., Ebashi, S., 1986. Regulatory and cytoskeletal proteins ofvertebrate skeletal muscle. Adv. Protein Chem. 38, 1–67.
ucéat, M., 2008. Protocols for cardiac differentiation of embryonic stem cells. Meth-ods 45, 168–171.
chmidt, J., Su, G., Reddy, J., Simon, M., Bradfield, C., 1996. Characterization of amurine Ahr null allele: involvement of the Ah receptor in hepatic growth anddevelopment. Proc. Natl. Acad. Sci. U.S.A. 93, 6731–6736.
chuldiner, M., Yanuka, O., Itskovitz-Eldor, J., Melton, D., Benvenisty, N., 2000. Effectsof eight growth factors on the differentiation of cells derived from humanembryonic stem cells. Proc. Natl. Acad. Sci. U.S.A. 97, 11307–11312.
toilov, I., Akarsu, A., Alozie, I., Child, A., Barsoum-Homsy, M., Turacli, M., Or, M.,Lewis, R., Ozdemir, N., Brice, G., Aktan, S., Chevrette, L., Coca-Prados, M., Sarfarazi,M., 1998. Sequence analysis and homology modeling suggest that primary con-
genital glaucoma on 2p21 results from mutations disrupting either the hingeregion or the conserved core structures of cytochrome P4501B1. Am. J. Hum.Genet. 62, 573–584.toilov, I., Rezaie, T., Jansson, I., Schenkman, J., Sarfarazi, M., 2004. Expression ofcytochrome P4501b1 (Cyp1b1) during early murine development. Mol. Vis. 10,629–636.
rs 202 (2011) 226–236
Thackaberry, E., Gabaldon, D., Walker, M., Smith, S., 2002. Aryl hydrocarbon receptornull mice develop cardiac hypertrophy and increased hypoxia-inducible factor-1alpha in the absence of cardiac hypoxia. Cardiovasc. Toxicol. 2, 263–274.
Thackaberry, E., Jiang, Z., Johnson, C., Ramos, K., Walker, M., 2005a. Toxicogenomicprofile of 2,3,7,8-tetrachlorodibenzo-p-dioxin in the murine fetal heart: modu-lation of cell cycle and extracellular matrix genes. Toxicol. Sci. 88, 231–241.
Thackaberry, E., Nunez, B., Ivnitski-Steele, I., Friggins, M., Walker, M., 2005b. Effect of2,3,7,8-tetrachlorodibenzo-p-dioxin on murine heart development: alterationin fetal and postnatal cardiac growth, and postnatal cardiac chronotropy. Toxi-col. Sci. 88, 242–249.
Wang, Y., Fan, Y., Puga, A., 2010. Dioxin exposure disrupts the differentiation ofmouse embryonic stem cells into cardiomyocytes. Toxicol. Sci. 115, 225–237.
Wobus, A., Guan, K., Yang, H., Boheler, K., 2002. Embryonic stem cells as a model tostudy cardiac, skeletal muscle, and vascular smooth muscle cell differentiation.Methods Mol. Biol. 185, 127–156.
Xie, A., Walker, N., Wang, D., 2006. Dioxin (2,3,7,8-tetrachlorodibenzo-p-dioxin)enhances triggered after depolarizations in rat ventricular myocytes. Cardiovasc.Toxicol. 6, 99–110.
Xu, R., Chen, X., Li, D., Li, R., Addicks, G., Glennon, C., Zwaka, T., Thomson, J., 2002.BMP4 initiates human embryonic stem cell differentiation to trophoblast. Nat.Biotechnol. 20, 1261–1264.
Yano, S., Miake, J., Mizuta, E., Manabe, K., Bahrudin, U., Morikawa, K., Arakawa,K., Sasaki, N., Igawa, O., Shigemasa, C., Yamamoto, Y., Morisaki, T., Hidaka, K.,Kurata, Y., Yoshida, A., Shiota, G., Higaki, K., Ninomiya, H., Lee, J.K., Shirayoshi,Y., Hisatome, I., 2008. Changes of HCN gene expression and I(f) currents inNkx2.5-positive cardiomyocytes derived from murine embryonic stem cellsduring differentiation. Biomed. Res. 29, 195–203.
Yeager, R., Oleske, D., Millsap, D., Henshel, D., 2006. Severe craniofacial malfor-mations resulting from developmental exposure to dioxin. Reprod. Toxicol. 22,811–812.
zur Nieden, N., Kempka, G., Ahr, H., 2004. Molecular multiple endpoint embry-onic stem cell test-a possible approach to test for the teratogenic potential ofcompounds. Toxicol. Appl. Pharmacol. 194, 257–269.