the erythrocyte binding motif of plasmodium vivax duffy binding protein is highly polymorphic and...
TRANSCRIPT

Molecular and Biochemical Parasitology 111 (2000) 253–260
The erythrocyte binding motif of Plasmodium 6i6ax Duffybinding protein is highly polymorphic and functionally
conserved in isolates from Papua New Guinea�
Jia Xainli a, John H. Adams b, Christopher L. King a,c,*a Di6ision of Geographic Medicine, Case Western Reser6e Uni6ersity, 2109 Adelbert Road, Room 137, Cle6eland,
OH 44106-4983, USAb Department of Biology, Uni6ersity of Notre Dame, Notre Dame, IN, USA
c Veterans Affairs Medical Center, Cle6eland, OH, USA
Received 21 June 2000; accepted 24 July 2000
Abstract
The Duffy binding protein (DBP) of Plasmodium 6i6ax is a critical adhesion ligand that participates in merozoiteinvasion of human Duffy positive reticulocytes. Binding domains have been shown to lie within a conservedN-terminal cyteine-rich region, region II, that contains 330-aa and the critical binding residues have been recentlymapped to 170-aa stretch within this region. Previous studies on few isolates indicated a significant degree ofpolymorphism in region II (DBPII). To examine further the degree of variability of DBPII, and whether thesevariants produce functional changes, DBPII was amplified by nested PCR from 24 isolates from Papua New Guinea,and the amplicons were cloned and sequenced. One synonymous and 18 non-synonymous mutations were identified.Altogether, 93% of the cumulative polymorphisms lie within the 170-aa region. Targeted surface expression of regionII of two different alleles on the surface of Cos7 cells did not affect their binding to Duffy positive erythrocytes. Theseresults indicate that polymorphisms in the critical binding motifs do not alter its function. If DBPII variation aroseto avert mechanisms of protective immunity targeting the DBP, vaccine development employing the parasite bindingligand may require strategies to minimize the effect of this polymorphism. © 2000 Elsevier Science B.V. All rightsreserved.
Keywords: Malaria; Plasmodium 6i6ax ; Erythrocyte binding proteins; Duffy binding proteins
www.parasitology-online.com.
Abbre6iations: DARC, Duffy antigen receptor for chemokines; DBP, Duffy antigen binding protein; PCR, polymerase chainreaction.� Note: Nucleotide sequence data reported in this paper are available in GenBank under the accession numbers AF289480-483,
AF289635-635 and AF291096.* Corresponding author. Tel.: +1-216-3684817; fax: +1-216-3684825.E-mail address: [email protected] (C.L. King).
0166-6851/00/$ - see front matter © 2000 Elsevier Science B.V. All rights reserved.
PII: S 0166 -6851 (00 )00315 -7

J. Xainli et al. / Molecular and Biochemical Parasitology 111 (2000) 253–260254
1. Introduction
Plasmodium 6i6ax is prevalent in Asia, Centraland Southern America and in many regions it isthe most common cause of malaria [1–3]. P.6i6ax, like other species of human Plasmodium,initiates erythrocyte invasion through expressionof several surface and apical organelles on themerozoite that bind to erythrocyte surfaceproteins [4,5]. One well-characterized ligand-re-ceptor interaction involves the Duffy bindingprotein (DBP) expressed on P. 6i6ax merozoiteand its corresponding receptor on erythrocytes,the Duffy antigen receptor for chemokines(DARC), [5–7]. This interaction is unique amonghuman malaria infections in that this receptor-lig-and interaction is essential for P. 6i6ax invasion ofthe erythrocytes, and alternative erythrocyte inva-sion pathways have not been identified so far [8].Individuals that lack the Duffy surface antigen ontheir erythrocytes are resistant to P. 6i6ax malaria[9]. Over 90% of sub-Saharan Africans lack theDuffy antigen [10] and as a result P. 6i6ax malariais rare in most of Africa. Thus, P. 6i6ax DBPshould be an attractive vaccine candidate antigen.
The P. 6i6ax DBP is a 140 KD protein thatbelongs to a family of erythrocyte bindingproteins characterized by a functionally conservedcysteine-rich region [5,11]. This cysteine-rich re-gion occurs in region II (DBP II), which havebeen shown to contain the binding motifs neces-sary for adherence to DARC on the erythrocyte[12,13]. Critical binding motifs in DBP II has beenmapped to a region between amino acids 291–460[14]. Previous studies have shown that althoughthe positions of the cysteine residues are con-served, other amino acids are highly polymorphic[15,16]. These reports provided a preliminary as-sessment of the DBPII molecular polymorphismsand did not determine whether these changes alterthe capacity to bind DARC-positive erythrocytes.This study identified a selected set of DBPII poly-morphisms from Papua New Guinean P. 6i6axisolates and examines their ability to adhere toDuffy-positive erythrocytes. Two different se-quences have been used to transfect Cos7 cells todetermine if DBPII polymorphisms affect DARC-specific adherence to erythrocytes in vitro.
2. Materials and methods
2.1. Origin of P. 6i6ax samples and DNApreparation
Peripheral blood samples (collected directly intovacutainers containing EDTA) were obtainedfrom individuals that were blood smear positivefor P. 6i6ax residing in the Wosera Area of PapuaNew Guinea near Maprik town, an area that hasbeen described in detail elsewhere [17]. Some ofthe samples were also blood smear positive forPlasodiumm falciparum. Samples were collectedfrom children aged 3–60 months (PNG 5–PNG202, Table 1) whose clinical and parasitologicalcharacteristics have been partially described [18]from an area referred to as the North Wosera.The remaining samples, PNG 284, PNG 393,PNG 778, PNG 1114 (Table 1), were obtainedfrom asymptomatic adults residing in the SouthWosera, an ecologically similar but geographicallydistinct area approximately 15 km away. DNAwas extracted from whole blood (200 ml) usingindividual spin blood kits (Qiagen Inc., Valencia,CA) according to the manufacturer’s protocol.
2.2. PCR amplification of DBPII
The Duffy binding protein genes were amplifiedwith primers complementary to conserved se-quences of the DBP including the entire region II(nest 1 primers; 5% primer-GAT AAA ACT GGGGAG GAA AAA GAT, and 3% primer-CTT ATCGGA TTT GAA TTG GTG GC). The reactionsolution of PCR amplification was 20 mM Tris–HCl, 50 mM KCl, 1.5 mM MgCl2, 200 nM ofeach deoxynucleotide triphosphate, 2.5 U of Taqpolymerase, 160 ng of each primer, and appropri-ate amount of DNA template in a final volume of100 ml with mineral oil on top. PCR was per-formed as following; one cycle of 2 min at 94°Cand 35 cycles of 1 min at 94°C, 2 min at 61°C, 2min at 72°C, and one cycle of 10 min at 72°C. Theprimers for nest 2 PCR upstream, 5% primer-CCTCGA ATG GTG GCA ATC CT-downstream, 3%primer TAC GAT ACC TGC CGT CGT AACwere added to the nest 1 amplicons and amplifiedunder similar conditions except that the annealing

J. Xainli et al. / Molecular and Biochemical Parasitology 111 (2000) 253–260 255
Tab
le1
Sequ
ence
poly
mor
phis
ms
ofP
.6i6a
xD
BP
IIa
381
AA
Con
don
384
191
386
390
394
395
417
424
434
437
446
447
467
503
308
371
377
379
TT
TG
AT
AA
GC
GT
TG
GT
GG
AA
TT
TA
AT
AT
GG
GT
GSa
l-l
TC
AG
CA
AC
TA
TA
AG
GA
AA
TT
GA
GC
(F)
(D)
(K)
(R)
(W)
(W)
(N)
(L)
(I)
(S)
(W)
(G)
(V)
(S)
(T)
(I)
(L)
(K)
(R)
Isol
ate
PN
G16
8.G
..A
.A
....A
AA
.C
....T
(G)
(H)
(I)
(v)
(K)
(P)
(S)
.G.
.A.
A..
..AA
A.
C..
...T
PN
G17
6(G
)(H
)(I
)(V
)(K
)(P
)(P
)(S
).G
..A
.A
..P
NG
5..A
AA
.C
....T
(G)
(H)
(I)
(V)
(K)
(P)
(S)
.G.
.A.
A..
..AA
A.
C..
PN
G18
..T(G
)(H
)(I
)(V
)(K
)(P
)(S
).G
..A
.A
..P
NG
180
AA
.C
....T
(G)
(H)
(I)
(K)
(P)
(S)
.C.
.G.
.A.
A..
AA
...T
PN
G7
(S)
(G)
(H)
(I)
(K)
(S)
.G.
.A.
A..
AA
.P
NG
23..T
(G)
(H)
(I)
(K)
(S)
.G.
.A.
A..
AA
...T
PN
G24
(G)
(H)
(I)
(K)
(S)
PN
G15
2.G
..A
.A
..A
A.
..T(G
)(H
)(I
)(K
)(S
).G
.C
..C
..A
..A
A.
..SP
NG
202
(G)
(H)
(R)
(I)
(K)
(S)
.G.
.A.
A..
..AA
A.
PN
G19
6.C
...T
(G)
(H)
(I)
(V)
(K)
(S)
(S)
.G.
.A.
..CA
..A
A.
..TP
NG
15(G
)(H
)(C
)(I
)(K
)(S
).G
..A
...C
A..
AA
...T
PN
G16
(G)
(H)
(C)
(I)
(K)
(S)
.A.
A..
PN
G18
4A
A.
..T(H
)(I
)(K
)(S
).A
.A
...C
.C
..A
A.
.A.
PN
G19
4.G
...T
(H)
(I)
(T)
(R)
(K)
(K)
(S)
(S)
.G.
A..
.C.
AA
...T
PN
G19
0(G
)(I
)(T
)(K
)(S
).G
.A
..P
NG
183
..TA
b
(G)
(I)
(N)
(S)
.G.
.A.
A..
..TP
NG
20(G
)(H
)(I
)(S
).G
..A
.A
...C
.P
NG
185
..T

J. Xainli et al. / Molecular and Biochemical Parasitology 111 (2000) 253–260256
Tab
le1
(Con
tinu
ed)
381
384
386
390
394
395
417
424
434
437
446
447
467
AA
Con
don
503
191
308
371
377
379
(G)
(H)
(I)
(T)
(S)
.G.
.A.
..AC
..P
NG
8..T
G..
(G)
(H)
(K)
(R)
(S)
(E)
.G.
.A.
A..
PN
G28
4.A
.(G
)(H
)(I
)(E
).G
..A
.P
NG
393
.A.
A..
AA
.(G
)(H
)(I
)(K
)(E
).C
..G
..A
.A
..P
NG
778
AA
.(S
)(G
)(H
)(I
)(K
)P
NG
1114
.G.
C..
..AA
..C
..A
A.
.A.
G..
(G)
(T)
(K)
(G)
(R)
(K)
(E)
222
21
211
22
233
35
195
220
22
1
aC
odon
s17
5th
roug
h57
5w
ere
sequ
ence
dan
don
lyth
ose
resi
dues
for
whi
cha
mut
atio
nw
asid
enti
fied
are
show
n.A
llot
her
sequ
ence
sex
amin
edw
ere
iden
tica
lto
the
prev
ious
lyse
quen
ced
Sal-
Iis
olat
e.b
Var
iant
sin
dica
ted
inbo
ldar
epr
esen
tin
only
asi
ngle
isol
ate,
but
the
sam
em
utat
ion
has
been
iden
tifie
dfr
omot
her
repo
rts.

J. Xainli et al. / Molecular and Biochemical Parasitology 111 (2000) 253–260 257
temperature was 59°C. Agarose gel electrophore-sis was performed to visualize the amplicons of990 bp. The PCR products were purified from thelow melting agarose gel, and cloned directly intopCR2.1-TOPO cloning vector using a TOPO-TAcloning kit (In Vitrogen, La Jolla, CA). Clonedspecies-specific inserts were sequenced using vec-tor-based extended M13 forward and reverseprimers. In preparation for DNA sequencing ofthe species-specific amplicons, excess dNTPs andunincorporated primers were eliminated using theQIAquick PCR purification kit (Qiagen Inc.).DNA sequencing was performed by fluorescence-based methodologies using Applied Biosystems377 automated DNA sequencer.
2.3. Cell surface expression of DBPII in Cos7cells and erythrocyte-binding assays
The plasmid construct used to express P. 6i6axDBPII protein, pHVDR22, was kindly providedby Dr Chetan Chitnis [12]. This construct con-tains DNA sequences encoding region II of theparasite ligand fused with signal sequence andtransmembrane segment of herpes simplex virusglycoprotein D (HSV gD). The fusion proteins aretargeted to the Cos7 cell surface by the transmem-brane segment. The pHVDR22 provided con-tained DBPII sequences (amino acids 198–522)from the Sal-I isolate [11] whose partial sequenceis indicated in Table 1. To replace the Sal-I se-quences with isolates from PNG, the DBPII re-gion was removed from the pHVDR22 bydigestion with the restriction enzymes P6uII andApaI, at 5% and 3% of the insert respectively. Inser-tion of the variant sequences were accomplishedby amplification of the cloned isolates with up-stream, 5% primer-TCT CGT CAG CTG ACGATC TCT AGT GCT ATT-downstream, 3%primer-ACG AGT GGG CCC TGT CAC AACTTC CTG AGT to generate the necessary P6uIIand ApaI restriction sites, respectively. PCR con-ditions were one cycle at 94°C for 2 min, and 30cycles at 94°C 1 min., at 55°C 1 min., at 72°C 2min., and one cycle of 10 min. at 72°C. After gelpurification, and restriction digest with P6uII andApaI, the PCR products were ligated intopHVDR22.
pHVDR22 constructs containing variant DBPIIwere transfected into Cos7 cells by lipo-fectAMINE (Life Technologies Inc., www.lifetech.com) according to manufacturer’s proto-cols. Transfection efficiencies were measured byimmunofluoresence 48–62 h after transfection asdescribed [13,14] using the mAbs ID3 and DL6 asthe primary antibodies (kindly provided by DrGary Cohen, University Pennsylvania, Philadel-phia, PA) that reacts with HSV gD sequencesadjacent to the DBPII region of the chimericmolecule.
Erythrocyte binding assays were performed 48h after transfection of Cos7 cells as describedpreviously [13]. Briefly, 100 ml of a 10% erythro-cyte suspension was added to 0.9 ml of media inwells containing 1×105 viable transfected Cos7cells. Plates were swirled to mix the erythrocyteswell, and the erythrocytes were allowed to settlefor 2 h at room temperature. Non-adherent ery-throcytes were removed by washing the Cos7 cellsthree times with PBS, and the number of trans-fected Cos7 cells with rosettes of erythrocytes wasscored in at least 30 fields at 40× magnificationusing an inverted microscope.
2.4. Gene analysis
The sequence alignments were done withALIGNMENT (Geneworks 2.1, Intelligenetics)and were manually adjusted to give the best fit.The nucleotide sequence of isolates was comparedwith that of P. 6i6ax Sal-I [11].
3. Results and discussion
3.1. Genetic polymorphisms
Eighteen non-synonymous and one synony-mous mutations were observed in 24 isolates fromPNG (Table 1). Fifteen non-synonymous muta-tions were present in at least two isolates, threemutations were present in only one isolate (e.g.codons 379, 386, and 394). These latter mutationshave been identified in prior studies [15,16] Fif-teen additional non-synonymous mutations wereidentified in one isolate only (not shown) and the

J. Xainli et al. / Molecular and Biochemical Parasitology 111 (2000) 253–260258
possibility that these represent Taq polymerase-in-duced error cannot be excluded. No additionalsynonymous mutations were observed. The pre-dominance of non-synonymous polymorphismssuggest that these mutations do not occur atrandom, and they may be maintained in the para-site population by immune selection.
Thirteen of 24 isolates (54%) had the samemutations at codons 308, 384, 390, 424, and 447compared with the Sal-I isolate suggesting someevolutionary divergence of parasites comparedwith those in Central America (represented bySal-I). Most isolates examined had 5–7 residuesthat differed from Sal-I, and had at least oneunique mutation (Table 1). The DBP II genesequence for PNG 168, 176, 005 isolates wereidentical to each other as were PNG 23, PNG 24and PNG 15, PNG 16 (Table 1). It is notable thatthe pairs PNG 23, PNG 24, and PNG 15, PNG16 each represent sibling pairs residing in thesame household that may be infected with sameparasite clone. Serine at codon 308 was present inall isolates in children residing in the North Wos-era compared with arginine at the same residueamong adults living in South Wosera. This sug-gests DBPII alleles may vary between locations.
Out of the 170-aa representing the critical bind-ing motif only a relatively small number ofresidues are targeted for change. The amino acidsubstitutions, however, usually produced signifi-cant changes in charge and/or polarity likely toaffect protein structure. For example, tworesidues changed from arginine to serine (codon308) or to histidine (codon 390), and three fromtryptophan to arginine (codons 394 and 437) or tocyteine (395). Overall, 14 of the 18 mutationsproduced a change in the family of amino acidsencoded compared with the Sal-I isolate. Elevenof these amino acid substitutions resulted in achange in charge. Changes in the protein structureat certain residues may alter antibody or T cellbinding of a particular epitope, thereby escapingthe host’s protective immune response. Cysteineresidues were highly conserved and most residueswere invariant suggesting that they are probablycritical for functional integrity of the ligand.
3.2. Relationship of polymorphisms to DBPII6ariation from other locations
Many of the DBPII polymorphisms identifiedin the present study have been observed previ-ously [15,16]. In Madang, PNG, 12 isolates weresequenced and 30 unique non-synonymous muta-tions identified [15]. In Colombia, 10 isolates weresequenced and 19 unique non-synonymous muta-tions described [16]. Twelve of 18 (66%) non-syn-onymous mutations shown in Table 1 areidentical to those observed from Madang, PNG(codons 308, 379, 381, 384, 386, 390, 394, 417,424, 437, 447, 503) and eight (44%) are sharedwith those from Colombia, South America(codons 191, 371, 384, 386, 417, 424, 437, 503).Base substitutions at codons 384, 386, 417, 424,427 and 503 are found in all three locations. Thissuggests certain alleles for the DBPII have a widegeographic distribution. The greater number ofshared mutations between isolates collected at twodifferent locations in PNG, compared with thosefrom Colombia, is consistent with closer geo-graphic proximity of Maprik and Madang (about350 km apart) compared with Colombia. It is alsopossible that the difference in Colombian andPNG polymorphisms may be attributed to differ-ences in the Duffy receptor on erythrocytes inColombian and PNG populations.
3.3. Polymorphisms are concentrated in thecritical binding region of DBPII
The cumulative polymorphisms identified fromisolates in the present study and in the two previ-ous reports show 124 of 133 (93%) individualmutations occur in the critical binding regionbetween cysteines 4 and 7 (aa 291–460, Fig. 1).All but one of these mutations is non-synony-mous. Compared with the Sal-I isolate, mostpolymorphisms (105, 80%) are located at fiveresidues: codons 308, 384, 390, 424 and 447. Sincethis region of the parasite molecule is criticalpoint of contact between the parasite and thehost, and contains a number of hydrophilicresidues to which antibody responses are likely todevelop, it is possible that this region of themolecule is targeted by the host immune response.
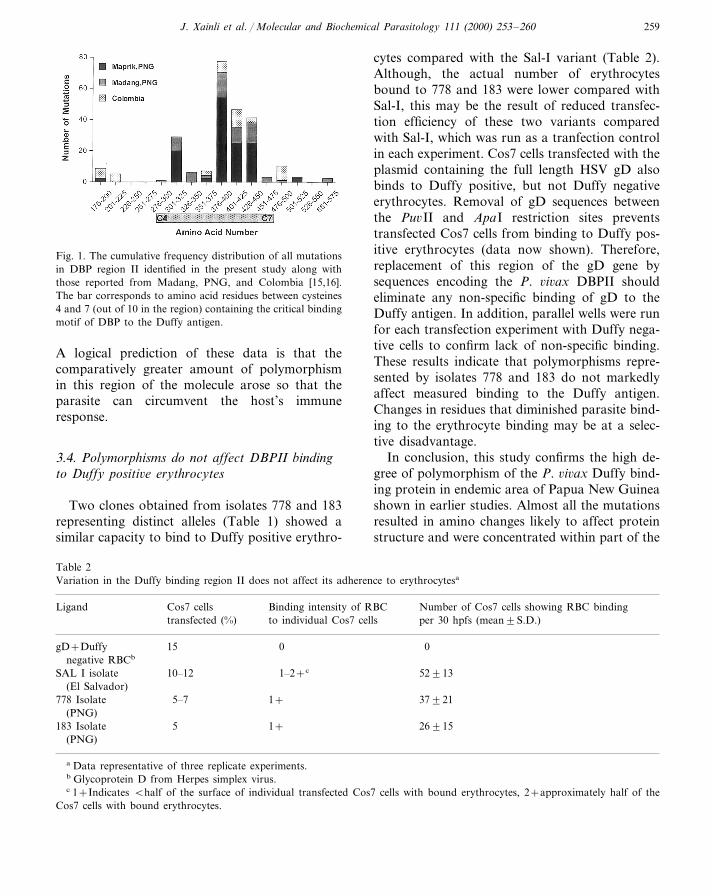
J. Xainli et al. / Molecular and Biochemical Parasitology 111 (2000) 253–260 259
Fig. 1. The cumulative frequency distribution of all mutationsin DBP region II identified in the present study along withthose reported from Madang, PNG, and Colombia [15,16].The bar corresponds to amino acid residues between cysteines4 and 7 (out of 10 in the region) containing the critical bindingmotif of DBP to the Duffy antigen.
cytes compared with the Sal-I variant (Table 2).Although, the actual number of erythrocytesbound to 778 and 183 were lower compared withSal-I, this may be the result of reduced transfec-tion efficiency of these two variants comparedwith Sal-I, which was run as a tranfection controlin each experiment. Cos7 cells transfected with theplasmid containing the full length HSV gD alsobinds to Duffy positive, but not Duffy negativeerythrocytes. Removal of gD sequences betweenthe Pu6II and ApaI restriction sites preventstransfected Cos7 cells from binding to Duffy pos-itive erythrocytes (data now shown). Therefore,replacement of this region of the gD gene bysequences encoding the P. 6i6ax DBPII shouldeliminate any non-specific binding of gD to theDuffy antigen. In addition, parallel wells were runfor each transfection experiment with Duffy nega-tive cells to confirm lack of non-specific binding.These results indicate that polymorphisms repre-sented by isolates 778 and 183 do not markedlyaffect measured binding to the Duffy antigen.Changes in residues that diminished parasite bind-ing to the erythrocyte binding may be at a selec-tive disadvantage.
In conclusion, this study confirms the high de-gree of polymorphism of the P. 6i6ax Duffy bind-ing protein in endemic area of Papua New Guineashown in earlier studies. Almost all the mutationsresulted in amino changes likely to affect proteinstructure and were concentrated within part of the
A logical prediction of these data is that thecomparatively greater amount of polymorphismin this region of the molecule arose so that theparasite can circumvent the host’s immuneresponse.
3.4. Polymorphisms do not affect DBPII bindingto Duffy positi6e erythrocytes
Two clones obtained from isolates 778 and 183representing distinct alleles (Table 1) showed asimilar capacity to bind to Duffy positive erythro-
Table 2Variation in the Duffy binding region II does not affect its adherence to erythrocytesa
Ligand Cos7 cells Number of Cos7 cells showing RBC bindingBinding intensity of RBCto individual Cos7 cells per 30 hpfs (mean9S.D.)transfected (%)
0gD+Duffy 15 0negative RBCb
529131–2+cSAL I isolate 10–12(El Salvador)
1+5–7 37921778 Isolate(PNG)
26915183 Isolate 5 1+(PNG)
a Data representative of three replicate experiments.b Glycoprotein D from Herpes simplex virus.c 1+Indicates Bhalf of the surface of individual transfected Cos7 cells with bound erythrocytes, 2+approximately half of the
Cos7 cells with bound erythrocytes.

J. Xainli et al. / Molecular and Biochemical Parasitology 111 (2000) 253–260260
gene encoding critical sequences necessary forbinding to the Duffy antigen. This natural varia-tion may have arisen from immune selection tocircumvent host immunity to block P. 6i6axmerozoite adherence to erythrocytes. Whetherthese polymorphic residues represent critical Band/or T cell epitopes associated with a protectiveimmune response remains to be established. Thereappeared to be limitations on the degree of vari-ability. Little or no polymorphism was observedat most residues, notably those encoding cysteine,suggesting that they are necessary for functionalcapacity of the parasite ligand. A vaccine thatincorporates DBPII sequences should be designedto include the most common variants or by in-creasing the immunogenicity of conserved regionsof the molecule provided they are accessible to thehost’s immune response.
Acknowledgements
This work was supported by the office of Re-search and Development, Medical Research, De-partment of Veteran’s Affairs. We thank Dr AnuShankar and the Staff at the Papua New GuineaInstitute of Medical Research in Madang forproviding the samples. We appreciate thethoughtful comments and critical reading of themanuscript by Dr Peter Zimmerman and Dr Ra-jeev Mehlotra.
References
[1] Sharma VP. Current scenario of malaria in India. Parassi-tologia 1999;41:349–53.
[2] San Sebastian M, Jativa R, Goicolea I. Epidemiology ofmalaria in the Amazon basin of Ecuador. Rev PanamSalud Publica 2000;7:24–8.
[3] Palmer CJ, Makler M, Klaskala WI, Lindo JF, BaumMK, Ager AL. Increased prevalence of Plasmodium falci-parum malaria in Honduras, Central America. RevPanam Salud Publica 1998;4:40–2.
[4] Barnwell JW, Nichols ME, Rubinstein P. In vitro evalua-tion of the role of the Duffy blood group in erythrocyteinvasion by Plasmodium 6i6ax. J Exp Med1989;169:1795–802.
[5] Adams JH, Sim BKL, Doland SA, Fang X, Kaslow DC,Miller LH. A family of erythrocyte binding proteins ofmalaria parasites. Proc Natl Acad Sci USA1992;89:7085–9.
[6] Wertheimer SP, Barnwell JW. Plasmodium 6i6ax interac-tion with the human Duffy blood group glycoprotein:identification of a parasite receptor-like protein. Exp Par-asitol 1989;69:340–50.
[7] Horuk R, Chitnis C, Darbonne W, et al. A receptor forthe malarial parasite Plasmodium 6i6ax : the erythrocytechemokine receptor. Science 1993;261:1182–4.
[8] Galinski M, Barnwell JW. Plasmodium 6i6ax : merozoites,invasion of reticulocytes and consideration for malariavaccine development. Parasitol Today 1996;12:20–9.
[9] Miller LH, Mason SJ, Clyde DF, McGinniss MH. Theresistance factor to Plasmodium 6i6ax in blacks. TheDuffy-blood-group. New Engl J Med 1976;295:302–4.
[10] Miller LH, Carter R. A review. Innate resistance inmalaria. Exp Parasitol 1976;40:132–46.
[11] Fang X, Kaslow DC, Adams JH, Miller LH. Cloning ofthe Plasmodium 6i6ax Duffy receptor. Mol Bochem Para-sitol 1991;44:125–32.
[12] Chitnis CE, Miller LH. Identification of the erythrocytesbinding domains of Plasmodium 6i6ax and Plasmodiumknowlesi proteins involved in erythrocyte invasion. J ExpMed 1994;180:497–506.
[13] Chitnis CE, Chaudhuri A, Horuk R, Pogo AO, MillerLH. The domain on the Duffy blood group antigen forbinding. J Exp Med 1996;184:1531–6.
[14] Ranjan A, Chitnis CE. Mapping regions containing bind-ing residues within functional domains of Plasmodium6i6ax and Plasmodium knowlesi erythrocyte-bindingproteins. Proc Natl Acad Sci USA 1999;96:14067–72.
[15] Tsuboi T, Kappe SH, Al-Yaman F, Prickett MD, AlpersM, Adams JH. Natural variation within the principaladhesion domain of the Plasmodium 6i6ax Duffy bindingprotein. Infect Immun 1994;62:5581–6.
[16] Ampudia E, Patarroyo M, Murillo L. Genetic polymor-phism of the Duffy receptor binding domain of Plasmod-ium 6i6ax in Colombian wild isolates. Mol BiochemParasitol 1996;78:269–72.
[17] Genton B, Al-Yaman F, Beck H-P, et al. The epidemiol-ogy of malaria in the Wosera area, East Sepik Province,in preparation for vaccine trials. Ann Trop Med Parasitol1995;89:369–90.
[18] Shankar AH, Genton B, Semba RD, et al. Effect ofvitamin A supplementation on morbidity due to Plasmod-ium falciparum in young children in Papua New Guinea:a randomised trial. Lancet 1999;354:203–9.
.