the function of genomes in bioenergetic organelles
TRANSCRIPT

Published online 19 December 2002
The function of genomes in bioenergetic organelles
John F AllenPlant Biochemistry Center for Chemistry and Chemical Engineering Lund University Box 124 SE-221 00 Lund Sweden
( johnallenplantbioluse)
Mitochondria and chloroplasts are energy-transducing organelles of the cytoplasm of eukaryotic cellsThey originated as bacterial symbionts whose host cells acquired respiration from the precursor of themitochondrion and oxygenic photosynthesis from the precursor of the chloroplast The host cells alsoacquired genetic information from their symbionts eventually incorporating much of it into their owngenomes Genes of the eukaryotic cell nucleus now encode most mitochondrial and chloroplast proteinsGenes are copied and moved between cellular compartments with relative ease and there is no obviousobstacle to successful import of any protein precursor from the cytosol So why are any genes at allretained in cytoplasmic organelles One proposal is that these small but functional genomes provide alocation for genes that is close to and in the same compartment as their gene products This co-locationfacilitates rapid and direct regulatory coupling Redox control of synthesis de novo is put forward as thecommon property of those proteins that must be encoded and synthesized within mitochondria andchloroplasts This testable hypothesis is termed CORR for co-location for redox regulation Principlespredictions and consequences of CORR are examined in the context of competing hypotheses and cur-rent evidence
Keywords chloroplasts mitochondria photosynthesis respiration gene expression redox regulation
1 INTRODUCTION
Mitochondria and chloroplasts are cytoplasmic organellesof eukaryotic cells Their primary function is energy trans-duction Mitochondria carry out respiration where energyis provided by differences in chemical redox potentialwhereas chloroplasts carry out photosynthesis where thesource of energy is light Both processes couple vectorialelectron transfer (redox chemistry) to ATP synthesis bymeans of a transmembrane proton motive force Theprimary steps in energy transduction are intrinsic tointernal membranesmdashmitochondrial inner membranesand chloroplast thylakoidsmdashthat are homologous withenergy-transducing cytoplasmic membranes of bacteriaMitochondria and chloroplasts also contain respectivelyassociated metabolic cycles of carbon dioxide production(the oxidative tricarboxylic acid pathway) and assimilation(the reductive pentose phosphate pathway) These cyclesare coupled tightly to energy transduction but involve sol-uble intermediates and are catalysed by soluble enzymesThey take place in internal aqueous phasesmdashthe mito-chondrial matrix and chloroplast stromamdashthat are hom-ologous with the bacterial cytoplasm
A prominent feature of mitochondria and chloroplastsis at first sight apparently unrelated to their primary func-tion their internal aqueous phases also contain discretequasi-autonomous genetic systems Each organellersquos gen-etic system contains DNA RNA and all the componentsnecessary both for DNA replication and for synthesis ofthe proteins encoded therein Nuclearly encoded compo-
One contribution of 21 to a Discussion Meeting Issue lsquoChloroplasts andmitochondria functional genomics and evolutionrsquo
Phil Trans R Soc Lond B (2003) 358 19ndash38 19 Oacute 2002 The Royal SocietyDOI 101098rstb20021191
nents are always required for the operation of an organel-lar genetic system but mitochondria and chloroplasts maynevertheless be described as lsquoquasi-autonomousrsquo theyarise only from pre-existing mitochondria and chloro-plasts and they are capable of performing synthesis ofsome of their own components even transiently after iso-lation in vitro The proteins synthesized that are not partof the genetic system are a subset of proteins with relatedfunctions in electron transport and closely related eventsin photosynthesis and respiration A central core of thissubset of proteins contains proteins that are universallyorganelle-encoded These include membrane-spanningsubunits that are central to protein complexes of electrontransfer and proton translocation
Figure 1 is a schematic diagram of generalized animalmitochondrial inner membrane Figure 2 is the corre-sponding diagram of part of a generalized chloroplast froma green plant In both diagrams protein subunits arepresented as cartoon shapes that are coloured in such asway as to indicate the location of the genes encoding themInventories of mitochondrial (Lang et al 1999) and chlor-oplast (Stoebe et al 1998 Martin et al 2002) genes areavailable and the list of organisms from which they comeis growing rapidly (Korab-Laskowska et al 1998) Table1 presents a summary of the functions of chloroplast genesretained from cyanobacteria
The retention of protein-coding genes in organelleDNA must be the result of an evolutionary process Theevolutionary mechanisms at work and the selection press-ures for loss and retention of genes are open and contro-versial questions A broad spectrum of contrasting viewscan be found in the literature and some of these are con-sidered in sect 4
20 J F Allen Genomes in mitochondria and chloroplasts
I II III IV ATPase
H+ H+ H+
NADH
NAD+ succinate fumarate ADP
ATP
O2 H2O
H+
inter-membrane spaceP-phase
N-phasemitochondrial matrix
Figure 1 Elements of energy transduction in respiration and oxidative phosphorylation in mitochondria The mitochondrialinner membrane is shown in yellow The principal complexes involved in energy transduction are complex I (NADHdehydrogenase) complex II (succinate dehydrogenase) complex III (the cytochrome bndashc1 complex) complex IV (cytochrome coxidase) and the coupling ATPase Vectorial electron transfer is depicted as thin dark-blue arrows Proton (hydrogen ionH1 ) translocation is depicted as thin red arrows Other chemical conversions are given as black arrows The major variableenvironmental input is oxygen (O2 ) shown in blue Subunits of protein complexes are coloured according to the location ofthe genes encoding them Mitochondria are usually pink or reddish-brown the colour of cytochromes and ironndashsulphurproteins so reddish-brown subunits have genes in the mitochondrion and are synthesized in the mitochondrial matrix lightbrown subunits have genes in the nucleus and are imported from the cytosol as precursors The depiction of sites of synthesisis schematic only and corresponds roughly to the arrangement in vertebrates Adapted from Ozawa (1995)
ATP
ADP
H+
NADP+
NADPH
CH2OATP
ADPNADP+
NADPHH2O O2
H+H+ H+
H+O2
H2OH+ H+
CO2
RUBISCO
H+
photosystem II Cyt b6_f photosystem I ATPase
thylakoid lumenP-phase
N-phasechloroplast stroma
light light
light
Figure 2 Elements of energy transduction in photosynthesis in chloroplasts The chloroplast thylakoid membrane is shown inyellow The principal complexes involved in energy transduction are photosystem II the cytochrome b6 ndashf complex (Cyt b6 ndashf )photosystem I the coupling ATPase and RUBISCO Vectorial electron transfer is depicted as thin dark-blue arrows Proton(hydrogen ion H1 ) translocation is depicted as thin red arrows Other chemical conversions are given as black arrows Themajor variable environmental inputs are light (orange arrows) and carbon dioxide (blue) Subunits of protein complexes arecoloured according to the location of the genes encoding them Chloroplasts are usually green so green subunits have genes inthe chloroplast and are synthesized in the chloroplast stroma light brown subunits have genes in the nucleus and areimported from the cytosol as precursors Biochemically green suggests chlorophyll and the colouring is then counterintuitiveonly the reaction centre subunits of photosystems I and II are both chlorophyll-binding and synthesized in the chloroplastThe depiction of sites of synthesis is schematic only and corresponds roughly to the arrangement in land plants Adapted fromMartin amp Herrmann (1998) and Race et al (1999)
Phil Trans R Soc Lond B (2003)
Genomes in mitochondria and chloroplasts J F Allen 21
Table 1 Retention of cyanobacterial genes in chloroplast DNA(Protein-coding genes are sorted into the functional categories assigned by Kaneko et al (1996ab) The genes are named andtheir distribution recorded in 16 completely sequenced chloroplast genomes to date in Martin et al (2002) from which the dataare taken Electronic Appendix A (available on The Royal Societyrsquos Publications Web site) lists the genes sorted into the functionalcategories of their gene products GPC genes per functional category The number of genes present in each category in thecyanobacterium Synechocystis PCC 6803 (Syn) and in any chloroplast genome completely sequenced (cp) Cat Ret categoryretention The number of times that a Synechocystis functional category is represented by a gene in cpDNA divided by the numberof genes in that category in Synechocystis itself Cat Ret = cpSyn For example Synechocystis has 84 genes for amino-acid biosynth-esis 10 of which are still present in at least one of 16 sequenced plastid genomes Hence Cat Ret = 1084 = 119 for amino-acid biosynthesis Deg Ret degree of retention The number of genes present in 16 chloroplast genomes divided by 16 andmultiplied by the number of times that the functional category is represented in chloroplast DNA For example in the 16 genomesanalysed there are 10 different amino-acid biosynthetic genes that occur in at least one cpDNA But of the 160 elements of theresulting matrix that could be positive only 27 are Hence Deg Ret = 27(16 acute 10) = 169 for amino-acid biosynthesis F(ret)fraction of retention The number of genes present in 16 chloroplast genomes divided by 16 and multiplied by the number oftimes that the functional category is represented in Synechocystis For example in the 16 genomes sequenced 27 different amino-acid biosynthetic genes occur in cpDNA Synechocystis has 84 genes for amino-acid biosynthesis that can be assumed to havebeen present in the common ancestor of these 16 plastid genomes Hence for amino-acid biosynthesisF(ret) = 27(16 acute 84) = 20 The genes analysed here correspond to those tabulated by Martin et al (2002) Only genes withknown homologues in cyanobacterial genomes were included in the analysis Values were calculated from the supplementarymaterial of Martin et al (2002) which was downloaded from httpwwwpnasorgcgicontentfull182432999DCI at the PNASWeb site)
no of GPC
Syn cp Cat Ret () Deg Ret () F(ret) ()
amino-acid biosynthesis 84 10 119 169 20cofactorprosthetic group biosynthesis 108 21 194 202 39cell envelope 64 7 109 250 27cellular processes 68 19 279 210 59central metabolism 31 1 32 63 02energy metabolism 86 3 35 125 04fatty-acid biosynthesis 35 8 229 172 39photosynthesis and respiration 139 74 532 603 321photosystem I 12 11 917 625 573photosystem II 25 20 800 806 645purine and pyrimidine biosynthesis 39 1 26 63 02regulatory functions 136 7 51 178 09DNA replication 49 2 41 156 06transcription 23 4 174 1000 130translation 144 56 389 531 207transport 158 2 13 63 01other plus hypothetical 587 43 73 202 15
(a) The bacterial nature of mitochondria andchloroplasts
It is generally accepted that organellar genetic systemsare evidence for the evolutionary origin of mitochondriaand chloroplasts as free-living eubacteria or a-proteobac-teria which entered into close inter-dependency with ahost cell (Douglas 1998 Delwiche 1999 Lang et al 1999McFadden 1999 2001 Roger 1999 Hackstein et al2001 Stoebe amp Maier 2002) The genetic systemsretained by organelles have many distinctively bacterialfeatures and heterologous systems for gene expression canbe reconstituted in vitro from organellar and bacterialcomponents By sharp contrast the genetic systems of theeukaryotic cell nucleus and cytosol are only distantlyrelated to those of bacteria and to those of their own cyto-plasmic organelles Instead nucleo-cytoplasmic geneticsystems more closely resemble the genetic systems ofarchaea (also known as archaebacteria) It is has been sug-gested that eukaryotic cells are an archaeal system of infor-mation storage replication transcription and translationthat is powered by a eubacterial system of energy trans-
Phil Trans R Soc Lond B (2003)
duction (Martin amp Russell 2003) The genetic systems ofmitochondria and chloroplasts can then be viewed aseukaryotic cellsrsquo residual bacterial systems of informationstorage and transmission Eukaryotic cellsrsquo residualarchaeal systems of energy transduction might theninclude direct vectorial ion pumps such as P- and V-ATP-ases and seven-helix transporters now associated prim-arily with information and signalling However the centralprocesses of energy transduction respiration and photo-synthesis are clearly bacterial in origin
(b) Currencies of symbiosisOne hypothesis for the origin of mitochondria states
that the nucleo-cytoplasmic system of eukaryotes isderived from that of an archaeal methanogenic chemo-autotroph that came to rely on the precursor of mitochon-dria for its supply of molecular hydrogen (Martin ampMuller 1998) This lsquohydrogen hypothesis for the firsteukaryotersquo resolves a long-held but paradoxical feature ofthe endosymbiont hypothesis how could a heterotrophichost cell have been able to supply abundant organic sub-
22 J F Allen Genomes in mitochondria and chloroplasts
strates that the bacterial precursor of mitochondria couldnot already obtain for itself Furthermore a second para-dox is resolved or at a least avoided export of ATP fromthe bacterial partner was not an immediate requirementfor the symbiosis to become established and could haveevolved subsequently Thus it is not necessary to postu-late a suicidal bacterium that excretes ATP nor a deferredselective advantage to the partnership implying foresightor an assurance of eventual contract fulfilment
The hydrogen hypothesis (Martin amp Muller 1998) seeksto provide an explanation of the origin of mitochondriathat accounts for the immediate benefit of the primordialsymbiosis to each partner but the implications of thehydrogen hypothesis for the origin of chloroplasts seemnot to have been considered in detail lsquoWhat was in it forthe proto-chloroplastrsquo may be an even more acute ques-tion than lsquowhat was in it for the proto-mitochondrionrsquoas speaking metabolically and energetically cyanobacteriacan do almost anything
Eukaryotes whether autotrophic or heterotrophic havenow mostly adopted the aerobic part of the energy metab-olism of their original bacterial constituents though inter-esting exceptions are known Respiration andphotosynthesis are antiparallel hydrogen transfer (redox)reactions
respiration
light photosynthesis
CH2O + A2 + H2O 2H2A + CO2
As in the Van Niel equation for photosynthesis A maybe an organic moiety for example giving H2A as succi-nate an inorganic element such as sulphur or evenlsquonothingrsquo so that H2A becomes H2 When A is O(oxygen) it becomes obvious that O2 in photosynthesiscomes from H2O not CO2 and the equation simplifies to
respiration
light photosynthesis
CH2O + O2 H2O + CO2
Phosphate group transfer and electron-transfer-linkedATP synthesis are intermediates in all cases In respir-ation CO2 is produced from stepwise oxidative decar-boxylations of pyruvate dehydrogenase and reactions ofthe tricarboxylic acid cycle In most forms of photosyn-thesis CO2 is assimilated to form initially sugar phos-phates
The mitochondrion exports ATP and CO2 and importsADP inorganic phosphate an electron acceptor andpyruvate Energetically the preferred electron acceptor ismolecular oxygen Oxygen comes from the environmentcells and cell membranes are freely permeable to oxygenThe primary metabolic division of labour between thechloroplast and the plant cytosol is that the chloroplastexports oxygen and reduced sugar phosphates (triosephosphate) and imports CO2 less reduced sugars (3-phosphoglycerate) and inorganic phosphate The light
Phil Trans R Soc Lond B (2003)
comes directly from the external environment and in thisrespect the chloroplast thylakoid is the external boundaryof the plant cell where light both as energy and infor-mation is first received The CO2 is also an environmentalinput though its supply may be regulated locally by CO2-or HCO2
3 -concentrating mechanismsAt the cellular level the mitochondrial inner membrane
is the first place where altered O2 supply exerts its pro-found metabolic effects Even granted specialized photore-ceptors the chloroplast thylakoid membrane is first placewhere changes in light quality and intensity affect a plantrsquosprimary input of energy Temperature changes selectivelyaffect diffusion-limited metabolism involving solubleintermediates In both locations abiotic environmentalchanges therefore have immediate and very far-reachingeffects on electron transfer and redox chemistry Theinternal membranes of mitochondria and chloroplastsstand at the energetic interface of the cell with the physicalenvironment and are likely to be first to know about andrespond to its changes
2 THE PROBLEM OF CYTOPLASMIC GENETICSYSTEMS
Given that the genomes of mitochondria and chloro-plasts are derived from those of once free-living bacteriait is clear that most of the bacterial genes that have sur-vived in any form have now been removed to the cellnucleus If most genes can be moved to the nucleus thenwhy not any gene And if any gene can be moved whynot all Why have mitochondria and chloroplasts retainedfunctional genomes at all What do the genes of mitochon-dria and chloroplasts have in common that has resultedin their retention
Alberts et al (1994 p 400) succinctly describe theproblem posed by the persistence of cytoplasmic geneticsystems as follows
Why do mitochondria and chloroplasts require their ownseparate genetic systems when other organelles thatshare the same cytoplasm such as peroxisomes and lyso-somes do nothellip The reason for such a costly arrange-ment is not clear and the hope that the nucleotidesequences of mitochondrial and chloroplast genomeswould provide the answer has proved unfounded Wecannot think of compelling reasons why the proteinsmade in mitochondria and chloroplasts should be madethere rather than in the cytosol
3 CORR CO-LOCATION OF GENES AND GENEPRODUCTS FOR REDOX REGULATION OF GENE
EXPRESSION
This hypothesis states that mitochondria and chloro-plasts contain genes whose expression is required to beunder the direct regulatory control of the redox state oftheir gene products or of electron carriers with whichtheir gene products interact These genes comprise a coreor primary subset of organellar genes The requirementfor redox control of each gene in the primary subset thenconfers a selective advantage upon location of that genewithin the organelle instead of in the cell nucleus Chloro-plast and mitochondrial genomes also contain genes for
Genomes in mitochondria and chloroplasts J F Allen 23
components of the chloroplast and mitochondrial geneticsystems themselves These genes comprise a secondarysubset of organellar genes genetic system genes There isgenerally no requirement for redox control of expressionof genes in this secondary subset though retention ofredox control of genetic system genes may in some casesallow amplification of redox signals acting upon genes inthe primary subset Retention of genes of the secondarysubset (genetic system genes) is necessary for the oper-ation of redox control of expression of genes in the primarysubset (bioenergetic genes) Without genes in the primarysubset the function of genes in the secondary subset iseventually lost and organelles lose their genomes
This hypothesis of co-location for redox regulation ofgene expression CORR was first outlined in a review(Allen 1992) The hypothesis was then systematically putforward in two articles (Allen 1993ab) developed(Allen amp Raven 1996) and has been independentlyreviewed (Race et al 1999) Some of its predictions havebeen tested experimentally as described later
Figure 3 illustrates the general idea of redox regulatorycontrol operating on mitochondrial genes to give the dis-tribution of sites of protein synthesis seen in figure 1 Fig-ure 4 shows the corresponding predicted pattern ofchloroplast redox regulatory control giving rise to the dis-tribution of sites of protein synthesis seen in figure 2Table 1 shows a functional classification of cyanobacterialgenes retained in 16 completely sequenced chloroplastgenomes This analysis (table 1) reveals a strong selectiveretention of genes for subunits of complexes involved inrespiration and photosynthesis and especially in chloro-plast photosystems I and II The pattern (table 1) seemsto be broadly consistent with CORR (figure 4) and incon-sistent with alternative hypotheses (see sect 4) It is reason-able to view photosynthesis as a process where preciserapid and coordinated redox regulatory control of geneexpression repays the cost of maintaining a chloroplastgenetic system
The CORR hypothesis is based on several propositionsassumptions or principles 10 of which are stated explicitlyand listed below Some of the principles are already generallyaccepted some are shared with competing hypothesesand some make direct experimental predictions Howeverother underlying principles of CORR are themselves generalprinciples or subsidiary hypotheses for which no directproof is available but for which predictions are made andpotentially falsifying observations or results can be imagined
The 10 principles that form the basis of the CORRhypothesis follow and each will be critically discussed inthe light of available evidence
(a) The principle of endosymbiotic originAs now generally agreed bioenergetic organelles have
evolved from free-living bacteriaAlthough once controversial (Mahler amp Raff 1975) this
principle will be taken as an axiom and not consideredfurther here The reader is referred to Gray amp Doolittle(1982) Gray (1992) Martin (1999) Martin et al (2001)to sect 1 and to accompanying articles in this journal issue
(b) The principle of unselective gene transferGene transfer between the symbiont or organelle and the cell
nucleus may occur in either direction and is not selective forparticular genes
Phil Trans R Soc Lond B (2003)
This principle takes the form of a testable assertion thatseems to be consistent with current knowledge Nothingcertain seems to be known about the mechanism of intra-cellular gene relocation (Palmer 1997 Martin amp Herrm-ann 1998 Martin et al 1998 Adams et al 2000 Henze ampMartin 2001 Millen et al 2001 Rujan amp Martin 2001Daley et al 2002) Plausible scenarios can be imagined onthe basis of the classical genetical phenomena of trans-formation transduction and transfection However asthere is no agreed mechanism there is no basis forasserting a mechanistic argument for selectivity Genetransfer between the organelle and nucleus seems to be aspecial case of lateral or horizontal gene transfer(Doolittle 1999) and no mechanistic basis seems to becurrently available for selectivity in what may be trans-ferred It should also be pointed out that lateral genetransfer between the organelle and nucleus does not haveto occur within a single cell nor within a multicellularindividual (although here any vertically transmitted genemust be incorporated into the germ line) for the outcomeof copying genes to novel compartments to be securedThus any plausible mechanism for heterologous recombi-nation may apply equally to gene transfer between com-partments Mitochondrial departures from the lsquouniversalrsquogenetic code and biased codon usage in chloroplasts (seelater) may be of selective value in that a barrier is therebyput in place to expression of genes in the lsquowrongrsquo compart-ment This barrier is unlikely to be insuperable especiallyif the lsquowrongrsquo compartment turns out actually to be a bet-ter location Impeded relocation might confer stabilitywithout being a primary reason for the existence of cyto-plasmic genomes
(c) The principle of unselective protein importThere is no barrier to the successful import of any precursor
protein nor to its processing and assembly into a functionalmature form
As with sect 3b this principle takes the form of a testableassertion that seems to be consistent with current knowl-edge The principle is discussed in more detail in sect 4because its negation (some proteins cannot be imported)has previously been put forward in itself as a reason forthe retention of cytoplasmic genomes Good hypothesesare easily falsified and they therefore forbid certain obser-vations The principle of unselective protein import for-bids the existence of a protein even a synthetic one thatcannot be imported by some means as a precursor
(d) The principle of continuity of redox controlDirect redox control of expression of certain genes was present
in the bacterial progenitors of chloroplasts and mitochondriaand was vital for selectively advantageous cell function beforeduring and after the transition from bacterium to organelle Themechanisms of this control have been conserved
This principle has been tested experimentally in the pastfew years and seems to be corroborated by an emergingpicture of redox signalling in bacteria and organellesExperiments carried out in vitro on the products of proteinsynthesis (Allen et al 1995 Galvis et al 1998) or RNAsynthesis (Pearson et al 1993 Konstantinov et al 1995Wilson et al 1996b) are consistent with a general phenom-enon of redox dependency of gene expression in bioener-getic organelles Redox control of chloroplast transcription
24 J F Allen Genomes in mitochondria and chloroplasts
nucleus cytosol
O2
H2O
redox regulation
N-phasemitochondrial matrix
Figure 3 Gene expression and principal pathways of biosynthesis of subunits of protein complexes involved in respiration andoxidative phosphorylation in animal mitochondria As in figure 1 reddish-brown DNA RNA and protein subunits are locatedand synthesized in the mitochondrial matrix light brown protein subunits have genes (also light brown) in the nucleus andare imported from the cytosol as precursors White genes and ribosomal and protein subunits are nuclearndashcytoplasmic and ofarchaebacterial origin Reddish-brown and light-brown genes and ribosomal and protein subunits are of bacterial origin Themajor variable environmental input is oxygen (blue) The essence of the CORR hypothesis is that it is beyond the ability ofthe nuclearndashcytoplasmic system to respond rapidly and directly to changes in oxygen concentration or partial pressure and soredox regulation of gene expression (red arrows) has been retained from the ancestral endosymbiont This redox regulationrequires co-location of certain genes with their gene products within the mitochondrion
nucleus cytosol
CH2O
CO2
redox regulation
N-phasechloroplast stroma
light light
Figure 4 Gene expression and principal pathways of biosynthesis of subunits of protein complexes involved inphotosynthesis in chloroplasts As in figure 2 green DNA RNA and protein subunits are located and synthesized in thechloroplast light brown protein subunits have genes (also light brown) in the nucleus and are imported from the cytosolas precursors White genes and ribosomal and protein subunits are nuclearndashcytoplasmic and of archaebacterial originGreen and light-brown genes and ribosomal and protein subunits are of bacterial origin The major environmental inputsare light (orange arrows) and carbon dioxide (blue) The essence of the CORR hypothesis is that it is beyond the abilityof the nuclearndashcytoplasmic system to respond rapidly and directly to changes in light and carbon dioxide and so redoxregulation of gene expression (red arrows) has been retained from the ancestral endosymbiont This redox regulationrequires co-location of certain genes with their gene products within the chloroplast
Phil Trans R Soc Lond B (2003)
Genomes in mitochondria and chloroplasts J F Allen 25
has been demonstrated (Pfannschmidt et al 1999abAllen amp Pfannschmidt 2000 Tullberg et al 2000) and wasprompted by this principle
The level of redox control of chloroplast transcriptionestablished unequivocally so far is that of plastoquinonean electron and hydrogen atom carrier located strategicallybetween photosystems I and II of photosynthesis Thedirections in which the effects are exerted are functionallyintelligible reduction of plastoquinone increases photosy-stem I transcription and decreases photosystem IItranscription oxidation of plastoquinone decreases photo-system I transcription and increases photosystem II tran-scription Plastoquinone in normal photosynthesis isreduced by the action of photosystem II and oxidized bythat of photosystem I Therefore redox control of chloro-plast gene expression provides a compensatory response(Pfannschmidt et al 1999a) and adds a previously mis-sing part of an explanation for the ability of plants andalgae to adjust the ratio of the two photosystems inresponse to changes in light quality (Chow et al 1990)By the same argument a functionally equivalent post-translational mechanism of redox control of light-harvesting function by photosystems I and II may havebeen conserved during the transition from prokaryotes toeukaryotes (Allen 1992 1995 Allen amp Nilsson 1997Allen amp Race 2002)
The precedent of chloroplast redox control of transcrip-tion (Pfannschmidt et al 1999ab) taken together with theprinciple in sect 3d predicts plastoquinone redox control ofcyanobacterial reaction centre gene transcription and thisis borne out by experiment (El Bissati amp Kirilovsky2001) This complementary self-adjusting transcriptionalresponse (Fujita 1997 Murakami et al 1997 El Bissati ampKirilovsky 2001) indicates strongly that redox control ofphotosynthetic reaction centre gene transcription waspresent in the proto-chloroplast Thus the mechanism ofregulation is likely to have been conserved throughout theevolutionary transition from bacterial symbiont to cyto-plasmic organelle A conserved mechanism of redox regu-lation of transcription in photosynthetic prokaryotes(Unden amp Bongaerts 1997 Bauer et al 1999) and chloro-plasts (Allen 1993c Pfannschmidt et al 1999b) is pre-dicted if evolutionary continuity of the process has beenmaintained
Thus apart from being consistent with experimentaldata the principle of continuity of redox control has beenof heuristic value in pointing to the underlying bacterialcharacter previously unsuspected in the control of geneexpression especially of transcription in chloroplasts ofhigher plants Other possible precedents for redox signal-ling components of cytoplasmic organelles include the prr(Eraso amp Kaplan 2000 2002 Oh amp Kaplan 2000) andreg (Bauer et al 1999) redox regulatory systems of purplenon-sulphur bacteria The TspO lsquooxygen sensorrsquo of Rho-dobacter sphaeroides is present in mammalian mitochondriaas a protein known there as a benzodiazepine receptorand each protein will complement loss of the other(Yeliseev amp Kaplan 2000) As outlined in sect 3j genes forsuch components are to be expected primarily in nucleargenomes This independent evolutionary hypothesis (sect 3d)would be strengthened still further by a demonstration ofone or more decisive redox regulatory links shared by pho-tosynthetic prokaryotes and all eukaryotes with mitochon-
Phil Trans R Soc Lond B (2003)
dria and where present with chloroplasts (Bauer et al2003)
(e) The principle of the selective value of redoxcontrol
For each gene under redox control (sect 3d) it is selectivelyadvantageous for that gene to be retained and expressed onlywithin the organelle
The twin selective advantages of redox control are likelyto be energetic efficiency and the suppression of the harm-ful side effects of electron transport operating on thewrong substrates No direct experimental support for thisprinciple is available and equally there is no evidenceagainst it Allotopic mutants in which genes have beenexperimentally relocated from organelles to the nucleushave been produced in yeast higher plants (Kanevski ampMaliga 1994) and human cells (Manfredi et al 2002) andare predicted by CORR to be poor adapters to environ-mental change Some authors believe there is potentialtherapeutic value in allotopic transformation movingmitochondrial genomes to the nucleus (Kanevski ampMaliga 1994 de Grey 2000) and predict the possibilityof r2 human cell lines in the foreseeable future (de Grey2000) This principle also has implications for ageing andthe evolution of separate sexes (Allen 1996) and possibleconsequences for development (Blackstone 1999 20002001 Harvey et al 2002) and somatic cloning in animals(Allen amp Allen 1999)
(f ) The principle of the selective value of nuclearlocation for genes not under redox control
For each bacterial gene that survives and is not under redoxcontrol it is selectively advantageous for that gene to be locatedin the nucleus and expressed only in the nucleus and cytosolIf the mature gene product functions in chloroplasts or mito-chondria the gene is first expressed in the form of a precursorfor import
There are several possible reasons for the selectiveadvantage of nuclear location of genes for organellar pro-teins One is the decreased probability of mutation arisingfrom free radical by-products of lsquoincorrectrsquo electron trans-fers (Raven et al 1994ab Allen amp Raven 1996) Therewould seem to be obvious economies in resource use andadvantages in coordination of gene expression if genes arekept in one compartment One might say that the bipartiteor tripartite genetic system of eukaryotic cells is untidyand inefficient
Another suggestion is that organellar genes do notundergo recombination and are present in relatively smallcopy numbers so that disadvantageous mutations willspread relatively quickly through a clonal population oforganelles Incorporation of functional gene copiesinstead into the nuclear genome may then be an irrevers-ible step analogous to the click of a ratchet (Saccone et al2000) There are examples of genes shared between allthree compartments (Stern amp Lonsdale 1982 Lonsdale etal 1983)
The cost of retaining an organellar genetic system is notincreased appreciably when additional genes are retainedin organelles if just one gene is retained then the cost ofa cytoplasmic genetic system is incurred and there may berelatively little extra cost for each additional gene retained(Allen amp Raven 1996) For a functional explanation of the
26 J F Allen Genomes in mitochondria and chloroplasts
existence of chloroplast and mitochondrial genomes thefocus of attention should therefore be on the relativelyconstant primary subset of bioenergetic genes (sect 3e)
(g) The principle of contemporary operation ofselection on gene location
For any species the distribution of genes between organelle(by the principle in sect 3e) and nucleus (by the principle in sect 3f)is the result of selective forces that continue to operate
This principle is essentially a statement contradictingboth the lsquofrozen accidentrsquo hypothesis and the hypothesisthat gene relocation is progressive predictable and stillincomplete (sect 4) In support of this principle it is becom-ing increasingly clear that mitochondria that lose theirfunction in aerobic respiration also lose their genomes asseen in the relict mitochondria of microsporidia(Embley amp Martin 1998 Van der Giezen et al 2002) andthe lsquomitosomersquo of Entamoeba histolytica (Tovar et al1999) In chloroplasts loss of photosynthesis results inloss of photosynthetic genes The examples of Epifagus (aparasitic higher plant) (Ems et al 1995 Wolfe et al 1992)and the residual apicomplexan plastids of Plasmodium(Wilson et al 1996a) and Toxoplasma where some plas-tidic genetic system is retained mean that some of theirgene products must be subject to redox regulation of geneexpression if this principle is correct Experimental test ofthis principle is possible using allotopic mutants as forthe predictions of the principle in sect 3f The evidence thatthe apicoplast (Wilson et al 1996a) is a plastid is largelygenetic the residual genome resembles that of photosyn-thetic plastids (chloroplasts) in gene organization ratherthan in content A plastid that has completely lost its gen-ome by virtue of having lost photosynthesis and all otherprocesses requiring redox regulation might be difficult todistinguish from an organelle derived in a similar way froma mitochondrion The persistence of targeted importedchaperones Hsp70 (Tovar et al 1999 Williams et al2002) and cp60 (Roger et al 1998) in residual organellesmight not distinguish a plastid ancestry from a mitochon-drial one
(h) The principle of primary involvement inenergy transduction
Those genes for which redox control is always vital to cellfunction have gene products involved in or closely connectedwith primary electron transfer These genes are always con-tained within the organelle
This principle would be violated and the CORRhypothesis refuted if any photosynthetic reaction centrecore subunit or respiratory chain subunit currentlyorganelle-encoded is found that functions in primary vec-torial electron transport and is nevertheless encoded in thenucleus and synthesized cytosolically as a precursor forimport all without any selective cost to the individualFrom the chloroplast transcriptional experiments ofPfannschmidt et al (1999ab) the chloroplast geneswhose transcription is regulated by the plastoquinoneredox state include psaAB (for photosystem I reactioncentre core polypeptides) psbA (for the photosystem IIpolypeptide D1) and rbcL (for the large subunit of RUB-ISCO interestingly following the same pattern of redoxcontrolmdashupregulated by oxidized plastoquinonemdashaspsbA) Transcription of the chloroplast genes petA
Phil Trans R Soc Lond B (2003)
(cytochrome f ) trnG (glycine tRNA) and rpoB (RNApolymerase) did not respond to plastoquinone redox statepetA for cytochrome f is predicted by this principle to beregulated at a different level of redox control or if at plas-toquinone at a different stage in gene expression Thisprediction might also apply to trnG and rpoB though thesequalify as members of the secondary subset that of geneticsystem genes to which the principle in sect 3i applies
(i) The principle of secondary involvement inenergy transduction
Genes whose products contribute to the organelle genetic sys-tem itself or whose products are associated with secondaryevents in energy transduction may be contained in theorganelle in one group of organisms but not in anotherdepending on the physiology and biochemistry of photosynthesisand respiration in the species concerned
Even phylogenetically moderately related species(within kingdoms) show differences in the location ofsome genes for example the ATP synthase subunits ofNeurospora and Saccharomyces (Attardi amp Schatz 1988)The CORR hypothesis also does not explicitly account forthe wide disparities in the distribution of genetic systemgenes for example the almost complete import of tRNAsin Chlamydomonas (GoldschmidtClermont 1998) Trans-port of nucleic acids across membranes is uncommon andwhy some RNAs should be organelle-encoded in somespecies but nuclearly encoded in others is not currentlyunderstood The only suggestion that can be made alongthe lines of CORR is that in some genetic systems genesmay need to be under the control of a regulatory systemthat responds to a signal arising within the organelle itselfThis signal could be the primary signal of altered redoxstate or some secondary signal Among the latter might befound signals that themselves report on the current stateof gene expression another piece of information that thenucleus might not have at its immediate disposal Suchsecondary signals could result at least in part fromCORR though other developmental and environmentalinputs may be imagined and would not detract fromCORR unless it could be shown that they provided a suf-ficient reason for the retention of the genetic system asa whole
(j) The principle of the nuclear encoding of redoxsignalling components
Components of the redox-signalling pathways upon whichCORR depends are themselves not involved in primary electrontransfer and so their genes have been relocated to the nucleusin accordance with sect 3f
This principle states that the redox signalling pathwaysthat are required for CORR should not themselves beexpected to utilize components whose biosynthesis mustbe under direct redox control Such components shouldtherefore fall into the major category of organellar pro-teins and be imported as precursors from the cytosol inline with the principle in sect 3f However the principleabove might be need to be modified in cases where ampli-fication of the redox signal is useful to the cell In thebacterial ntr system which was a precedent for CORR(Harrison et al 1990 Allen 1992) the histidine kinasentrB and aspartate response regulator ntrC form part ofthe gln operon whose transcription they control Odontella
Genomes in mitochondria and chloroplasts J F Allen 27
and Porphyra have plastids that encode more proteins thanthose of most chlorophytes (Stoebe et al 1998 Turmel etal 1999) and these proteins include two-component sig-nal transduction proteins prr and ycf27 (Stoebe et al1998) Extreme or extremely rapid environmental fluctu-ations might be catered for by amplification of signals thatare transmitted by signalling pathways whose componentsare under their own control It is not known whether ycf27and prr are in operons whose promoters their gene pro-ducts control It is also uncertain whether ycf27 is a redoxsignalling component and its distribution at least in itsplastidic location seems to be correlated with the presenceof a phycobilin-based light-harvesting system as found inred algae and cyanobacteria (Ashby amp Mullineaux1999ab Ashby et al 2002) The best guess of the originalstatement of the CORR hypothesis (Allen 1992 1993ac)was that bacterial two-component systems (Iuchi amp Lin1992 Allen 1993c Grebe amp Stock 1999) provide a plaus-ible means of exerting redox regulatory control over mito-chondrial and chloroplast gene expression andsubsequent research confirms the presence of two-compo-nent genes in genomes of Arabidopsis and yeast SeveralArabidopsis two-component genes are predicted to be tar-geted to mitochondria and chloroplasts (Forsberg et al2001) These must be considered as prime candidates forcomponents involved in redox regulation of chloroplasttranscription seen in the experiments of Pfannschmidt etal (1999a 2001a) The presumably homologous regulat-ory process serving to control photosystem stoichiometryin cyanobacteria the prr system (Li amp Sherman 2000) isnow characterized as a two-component system involvinga redox sensor and redox response regulator as originallydefined (Allen 1992 1993ac)
4 CONTRASTING HYPOTHESES CONCERNINGTHE RETENTION OF CYTOPLASMIC GENOMES
CORR is a recent addition to explanations that havebeen put forward to account for the persistence of gen-omes in organelles These models and their underlyingmechanisms are now discussed and compared
(a) There is no reason that is just how it isOne reaction to the problem of cytoplasmic genetic sys-
tems (sect 2) is that there is no problem and no particularreason for their retention However there is a very widephylogenetic distribution of a common core or primarysubset of organellar-encoded genes and one which haswithstood considerable genomic reorganization over evol-utionary time This constancy is seen for example in thesame genes being present in the chloroplasts of landplants with multiple copies of a single circular chloroplastchromosome and in dinoflagellate chloroplasts with acollection of different DNA minicircles each with essen-tially one gene (Zhang et al 1999 2001 Barbrook et al2001) Another suggested explanation of the retention ofmitochondrial and chloroplast genomes is simply torestate the endosymbiont hypothesis and to point out thatchloroplast and mitochondrial genes lsquocame inrsquo with thebacterial endosymbiont This suggestion however doesnot address the question of why some genes were retainedwhereas others were not It is also now clear that mostgenes that came in and still survive have nuclear and cyto-
Phil Trans R Soc Lond B (2003)
solic gene products whereas many chloroplast and mito-chondria proteins had a eukaryotic origin (Salzberg et al2001 Martin amp Russell 2003) The odds against a con-served pattern in gene location (eg table 1) arising bychance must be astronomical Thus there is a reason forthe retention of organellar genomes and a reason for theretention of the same primary subset of genes Becauseevolution proceeds by natural selection of chance vari-ation lsquoreasonrsquo in this context requires identification of thefunction of the location of a gene that is the selectivevalue of the phenotype produced by that location Evol-ution sometimes retains apparently functionless relics ofpreviously useful features but not for long especially ifthere is a cost
(b) The lock-in hypothesisThe lsquolock-inrsquo hypothesis of Bogorad (1975) was pro-
posed independently of the endosymbiont hypothesis anddoes not depend upon it According to the lock-in hypoth-esis core components of multi-subunit complexes must besynthesized de novo in the correct cellular compartmentotherwise photosystems or respiratory chain complexesfor example might assemble in the plasma membranevacuolar membrane or endoplasmic reticulum The lock-in hypothesis made no clear prediction about the conti-nuity of cytoplasmic genetic systems even though cyto-plasmic uniparental non-Mendelian inheritance for bothmitochondria (Roodyn amp Wilkie 1968) and plastids(Kirk amp Tilney-Bassett 1978) made it likely that organellarDNA the chondriome and the plastome replicated inde-pendently of nuclear DNA (Whitehouse 1969)
Mechanisms of protein import and targeting (Schatz1998 Koehler et al 1999 Leuenberger et al 1999 Gab-riel et al 2001 Jarvis amp Soll 2001 Pfanner amp Geissler2001 Duby amp Boutry 2002 Pfanner amp Wiedemann 2002)now effectively answer the problem of specifying the cor-rect locations for membrane protein complexes for whichthe lock-in hypothesis was proposed as a solution How-ever regulation of synthesis of an entire functional multi-subunit complex by control of synthesis of individual sub-units is still an important principle This seems to be afeature of the lsquoCESrsquo model (control by epistasy ofsynthesis) of Wollman (Wollman et al 1999 Eberhard etal 2002) which envisages nuclear control as all-embracingwith organellar genetic systems doing as it were whatthey are told In the hypothesis of CORR propoundedhere CES is equally important but the essence of CORRis that the organellar genetic system alone knows what arethe immediate and most important signals of environmen-tal change The organelle may keep the nucleus informedbut this is a lower priority than initiating an appropriateresponse CES and CORR agree that assembly of multi-subunit complexes is a complex business and requirescoordinated control in which no step is rate-limiting butmany may be obligatory
(c) Going going gone the evolutionary process oftransfer of genes from organelles to the
nucleus is still underway but incompleteThis hypothesis envisages that for most eukaryotes we
are now witnessing the last stages in a slow evolutionarytransfer of genes to the nucleus and this transfer may beexpected to proceed to completion in what is in evol-
28 J F Allen Genomes in mitochondria and chloroplasts
utionary terms the near future (Palmer 1997 Gray 19992000 Gray et al 1999 Adams et al 2000 Daley et al2002) A clear and compelling statement of this widelyheld position is that of Herrmann amp Westhoff (2001 p10)
hellipthe transfer of geneshelliptells that the partite plant gen-ome is not in a phylogenetic equilibrium All availabledata suggest that the ultimate aim of genome restructur-ing in the plant cell as in the eukaryotic cell in generalis the elimination of genome compartmentation whileretaining physiological compartmentation
One serious problem with this hypothesis is the dis-tinctly non-random sample of genes that remain to betransferred A relatively constant subset of genes is foundin bioenergetically functional mitochondria from fungi tomammals and angiosperms as in photosynthetic plastids(chloroplasts) from unicellular algae to angiosperms Thishypothesis therefore requires an additional ad hocassumption that some gene transfers are inherently moreprobable than others (this too is widely supposed seelater) and thus that gene transfer events proceed in thesame sequence in quite independent lineages Thereseems to be no evidence for this and the order of geneloss from mitochondria and chloroplasts has been used toinfer phylogeny (Martin et al 1998 Adams et al 2000Millen et al 2001) lsquoGoing going gonersquo (Palmer 1997)is difficult to refute categorically but it does not seem tobe consistent with recent evidence For example chromo-some two of Arabidopsis thaliana (Lin et al 1999) has asingle insertion encompassing with about 99 sequencesimilarity the complete Arabidopsis mitochondrial genome(Unseld et al 1997 Marienfeld et al 1999 Stupar et al2001) Unless this single transfer is a freak occurrencethis finding strongly indicates that gene transfer occurswith relative ease (Adams et al 2000 Henze amp Martin2001 Rujan amp Martin 2001 Salzberg et al 2001) Inaddition there is experimental evidence that randomlygenerated polypeptides are mitochondrially imported athigh frequency (Lemire et al 1989) The hypothesis thatwe now see the same few stragglers in diverse organellargenomes is not impossible but should be regarded as alast resort perhaps applicable only in extreme cases suchas the greatly reduced apicomplexan plastomes and chon-driomes Even there the hypothesis carries with it an obli-gation to explain why organisms with few organellar geneshave lsquoevolvedrsquo faster than those with many
(d ) The frozen accidentWhat may be described as lsquothe frozen accident hypoth-
esisrsquo states that the evolutionary process of gene transferfrom organelle to nucleus was underway when somethinghappened that stopped it One example of the frozen acci-dent is a scenario (Von Heijne 1986) in which import intomitochondria of the precursor proteins arising from newlytranslocated genes proceeded for long enough after theprimordial endosymbiosis for most of the symbiont-derived genes to be moved to the cell nucleus The lsquoacci-dentrsquo that halted the process was the evolutionary originof exocytosis and protein secretion after which it becameimpossible for precursors properly destined for mitochon-drial import to be distinguished from those destined forexport from the cell As with the lsquolock-inrsquo hypothesis the
Phil Trans R Soc Lond B (2003)
lsquofrozen accidentrsquo seems incompatible with the remarkableprecision and specificity of protein targeting (Schatz 1998Koehler et al 1999 Leuenberger et al 1999 Gabriel etal 2001 Jarvis amp Soll 2001 Pfanner amp Geissler 2001Duby amp Boutry 2002 Pfanner amp Wiedemann 2002) Pro-tein sorting mechanisms do not seem to forbid controlledimport into any cellular compartment Another lsquoaccidentrsquothat may have impeded subsequent gene transfer in thecase of mitochondria is their departure from what is other-wise a universal genetic code Genetic code differences arediscussed later in the context of whether some genes can-not be moved
(e) Some proteins (with cofactors) cannot beimported
We may describe as the lsquounimportability hypothesisrsquo theproposal that some proteins are inherently unimportableand must therefore be synthesized de novo in the correctcompartment within the cell The unimportability hypoth-esis is most often proposed as its special case the hydro-phobicity hypothesis with hydrophobicity being thereason for unimportability
(i) HydrophobicityA commonly entertained notion is that proteins that are
encoded and synthesized within organelles comprise a pri-mary subset characterized by shared hydrophobicity allare intrinsic membrane proteins (Von Heijne 1986Popot amp de Vitry 1990 Claros et al 1995) As with theCORR hypothesis favoured here the primary subset thenrequires a secondary subset of genetic system componentsthe latter not themselves being hydrophobic but beingrequired for synthesis of the others Certainly the mito-chondrial inner membrane contains some intrinsic proteincomplexes whose subunits broadly satisfy the criterion ofhydrophobicity However the core subunits that are mito-chondrially encoded are also explained by the CORRhypothesis This would make hydrophobicity a require-ment for their function in vectorial electron and protontransfer and not a reason in itself for retention of theirgenes in the cytoplasm
The hydrophobicity hypothesis has some very clearcounter-examples in chloroplasts and cannot thereforestand if one wishes for a single principle that embracesboth types of bioenergetic organelle One clear counter-example is the large (L) subunit of the enzyme RUBISCOan abundant but entirely membrane-extrinsic protein ofca 55 kDa molecular mass The RUBISCO large subunitis generally chloroplast-encoded with the exception of thenuclearly encoded bacterial-type L subunit of L2 RUBI-SCO in peridinin-containing dinoflagellates (Morse et al1995) In most photosynthetic eukaryotes RUBISCO hasa chloroplast encoded large subunit and a nuclearly enco-ded small (S) subunit (Ellis 1984 Gatenby amp Ellis 1990)giving an L8S8 holoenzyme and this RUBISCO is widelytaken as a paradigm for the need for coordination ofnuclear and chloroplast gene expression (Rodermel 2001)
By complete contrast to the RUBISCO large subunitwhich is a chloroplast-encoded and water-soluble proteina second major violation of the hydrophobicity hypothesisis the existence of the universally nuclearly encoded butalways hydrophobic subunits of the chloroplast LHC IIand LHC I (Chitnis amp Thornber 1988) These apopro-
Genomes in mitochondria and chloroplasts J F Allen 29
teins of ca 27ndash30 kDa are quite water-insoluble and areusually fully extracted only in chloroformndashmethanol or inthe presence of detergents The Lhc gene family is diverse(Green amp Kuhlbrandt 1995) but most gene productsor Lhcp (light-harvesting chlorophyll protein) polypeptideshave three membrane-spanning hydrophobic a-helices(Kuhlbrandt 1994) It is interesting that three-helix Lhcproteins seem to be absent from extant cyanobacteria andchloroxybacteria (Green amp Kuhlbrandt 1995)
(ii) The five-helix rulePerhaps with the three-helix model of LHC II in mind
a variation of the hydrophobicity hypothesis is the empiri-cal lsquofive-helix rulersquo (Popot amp de Vitry 1990 Claros et al1995 Wollman et al 1999) which states that no intrinsicmembrane protein with five or more membrane-spanninghelices can be encoded in the nucleus Five is a good guessfor a threshold number of helices demanding organellar-encoding in the context of photosynthetic reaction centres(Heathcote et al 2002) where the earliest known struc-ture that of type II centres (Deisenhofer et al 1985) asin photosystem II (Zouni et al 2001) shows exactly fivehelices with type I centres containing 11 (Jordan et al2001) There are however many nuclearly encoded sub-units in both mitochondria and chloroplasts that havefewer than five membrane helices so the five-helix rulecan only describe a sufficient condition and not a neces-sary condition for nuclear encoding
It should also be noted that ADPndashATP carriers (AACs)are encoded in the nucleus AACs of the mitochondrialinner membrane have six transmembrane helices (van derGiezen et al 2002) while the those of the chloroplast innerenvelope (Winkler amp Neuhaus 1999) have 12 (Tjaden etal 1998)
With the counter-examples of nuclearly encoded AACswith six or 12 membrane-spanning helices many intrinsicmembrane proteins with fewer than five helices somenuclearly encoded and some not and with the RUBISCOlarge subunit as a major product of chloroplast geneexpression with no membrane domain at all the five-helixrule begins to seem rather like a case of special pleadingEven if there are indeed special mitigating circumstancesfor AACs (this argument is outlined later) the five-helixrule as with the hydrophobicity hypothesis of which it isa special case will disappear if it can be shown that pos-session of five or more membrane helices is required forsome other function such as that of CORR that deter-mines the organellar location of the gene concernedHydrophobicity is in some cases a feature of proteins thatare encoded in organelles but it is not in itself the issue
For chloroplasts the hydrophobicity hypothesis predictsretention of cyanobacterial genes in functional categoriesassociated with the chloroplast envelope and with trans-port just as much as with photosynthesis Table 1 showsthat this is not the case
A mechanism for the unimportability of hydrophobicproteins is rarely stated explicitly and it is rather left tothe imagination whether hydrophobic proteins might asit were get stuck halfway through the chloroplast envelopeor mitochondrial outer membrane whether transport can-not get started on a precursor protein that has alreadyspontaneously inserted into the membrane or whetherhydrophobic proteins will be unable to make the journey
Phil Trans R Soc Lond B (2003)
through the cytosol without precipitating there In thecytosol co-translational transport or membrane insertionwould in principle address the latter point but it seemsto be generally true that chloroplast and mitochondrialproteins are post-translationally inserted into the organel-lar outer membrane So the obvious ancillary questionabout unimportability is lsquowhy then cannot hydrophobicproteins be co-translationally imported into organellesrsquoThere is evidence in yeast for co-translational transportinto mitochondria of proteins thought to be derived fromthe original endosymbiont (Marc et al 2002) If there isany truth in hydrophobicity as a barrier to protein importthe CORR principle of unselective import (sect 3c) shouldbe easily refuted for example by a demonstration that noknown targeting presequence is capable of importing poly-phenylalanine encoded by polyU for in vitro import stud-ies
An additional proposal intended to account for theunimportability of some hydrophobic proteins and forimportability of others is suggested by A de Grey(personal communication) who argues that bacteriallyderived protein precursors imported into mitochondriacross the mitochondrial inner membrane completelybefore insertion into that membrane from the mitochon-drial matrix (the N-phase) This is held to be an obstacleto successful import of membrane-intrinsic proteins thatmight en route discharge the proton motive force By con-trast eukaryotically derived proteins can be inserted intothe mitochondrial inner membrane from the inter-mem-brane space (the P-phase) and then there is for somereason no similar uncoupling effect Thus adenylatetransporters or AACs can be nuclearly encoded whereascytochrome oxidase subunits for example cannot It isnot obvious why bacterially derived subunits such asthose of cytochrome oxidase did not acquire eukaryoticinsertion mechanisms Perhaps this acquisition is forbid-den by the need to conserve pre-existing routes ofassembly into multi-subunit complexes or perhaps theneed for insertion from the inside reflects a need to incor-porate a cofactor for example haem or ironndashsulphur clus-ters along the way
(iii) Unimportability of three dimensional structures andcofactors
Accumulating evidence on protein transport mech-anisms seems to make a specific obstacle to successfultransport of certain sorts of protein less and less plausibleespecially given the observation that precursors with N-terminal targeting sequences not mature functional poly-peptides themselves traverse membranes during biosynth-esis
Protein folding inside the living cell is invariably assistedby molecular chaperones (Gatenby amp Ellis 1990 Azem etal 1997 Komiya et al 1997 1990 Jackson-Constan et al2001) proteins that impede the formation of lsquoimproperrsquostructural interactions that will otherwise produce aggre-gation or other non-functional interactions Proteinimport mechanisms appear mostly to rely on specific mol-ecular chaperones for guided unfolding before membraneinsertion and refolding after translocation of the polypep-tide into its destined compartment (Gatenby amp Ellis 1990Azem et al 1997 Komiya et al 1997 Jackson-Constanet al 2001) Because polypeptides are mostly lsquothreadedrsquo
30 J F Allen Genomes in mitochondria and chloroplasts
through membranes no special constraint on transportshould be expected to arise from 3D structure or surfaceproperties of a protein
An exception to this unfolding rule may be providedby holoproteins containing cofactors that are transportedalong with the apoprotein presumably with a largely intact3D structure The lsquoTATrsquo (twin-arginine translocase) sys-tem (Bogsch et al 1998 Dalbey amp Robinson 1999 Berkset al 2000ab Robinson 2000 Robinson amp Bolhuis 2001Sargent et al 2002) works broadly along these linesthough its mechanism is unresolved TAT is relevant herebecause it adds transport of essentially complete holopro-teins to the cellrsquos repertoire of transport mechanisms andeven proteins that may not be allowed to unfold perhapsbecause of the toxicity of the free cofactor are not therebymade candidates for an unimportability rule One variantof the unimportability hypothesis is that transport canwork only by unfolding and dissociated cofactors may betoxic to the cell The bacterially derived TAT system(Bogsch et al 1998 Dalbey amp Robinson 1999 Berks etal 2000ab Sargent et al 2002) overcomes this argumentas would examples of cofactor-free polypeptides such as(C)Fo subunit c which are nevertheless generally encodedin organelles
The presence of a vesicular transport system in chloro-plasts (Westphal et al 2001) counts against the unim-portability hypothesis Vesicular transport between thechloroplast envelope and the thylakoid membrane shouldin principle allow any protein to be ferried across thechloroplast stroma Vesicles might allow transport inparticular of membrane proteins that would discharge theproton motive force if allowed to connect transiently theN and P aqueous phases on each side of a coupling mem-brane
Another observation that seems to count against unim-portability is the increased number of membrane barriersin plastids derived from secondary tertiary or higher-orderendosymbioses These plastids seem to have retained ifanything fewer genes rather than more as unimportabilitypredicts The nucleomorph the residual nucleus of theprimary host retains many genes for molecular chap-erones (Archibald et al 2001 Cavalier-Smith 2002) inagreement with the conclusion that protein import pro-ceeds by conserved and specialized mechanisms If eachnew import added to gain a higher-order endosymbiosisrequires additional targeting presequences then perhapsthere is an advantage in components of an import machin-ery not being imported themselves
It seems that unimportability is less easy an assumptionto make today than in previous years and the onus maynow be on its proponents to cite specific examples of pro-teins that can never be imported into chloroplasts or mito-chondria
(f ) Some genes cannot be moved(i) Differences in the mitochondrial genetic code
The genetic code is the specific relation between ribon-ucleotide triplets of codons and tRNAs that recognizethem and which thus determine which individual aminoacids if any will be incorporated into the growing poly-peptide chain Mitochondria show small departures fromthe otherwise universal genetic code (Attardi amp Schatz1988 Alberts et al 1994) UGA is a lsquostoprsquo codon in the
Phil Trans R Soc Lond B (2003)
universal code and in plant mitochondria but is the codonfor tryptophan in mitochondria of animals and fungiCUA means leucine everywhere except fungal mitochon-dria where it means threonine and AGA and AGU codefor arginine in the lsquouniversalrsquo code but serine in insectmitochondria and lsquostoprsquo in mammalian ones The signifi-cance of this heterogeneity in coding is most unlikely torest in the early origin of mitochondria as the lsquouniversalrsquocode is employed by bacteria and protists Furthermorethe diversity of departures from the lsquouniversalrsquo codeamong mitochondria of different eukaryotic lineages indi-cates they had independent origins A general rule formitochondrial genetic codes is that reliance on the thirdlsquowobblersquo position is decreased which allows all 20 proteinamino acids to be specified by fewer tRNAs (22 in animalmitochondria instead of 31 in the cytosol) This in turnmeans that mtDNA can specify more proteins for a givenlength of DNA or conversely that the mitochondrialgenome can be condensed or lsquostreamlinedrsquo Although thisarrangement does not explain the retention of mitochon-drial genomes and may be a secondary event in mitochon-drial evolution it almost certainly places an extra barrierin place of gene relocation As discussed previously how-ever there seems to be no single barrier that can explainthe retention of a relatively constant subset of mitochon-drial genes
There has clearly been a contraction of mitochondrialDNA especially in animals Animal mitochondrial DNAhas few if any non-coding sequences and animal mito-chondrial genetic codes require fewer tRNAs than the sin-gle code operating in the cytosol (Gray 1999 Gray et al1999 Selosse et al 2001) These may be devices todecrease the total frequency of mutations per mitochon-drial genome assuming a constant frequency per unitlength of mitochondrial DNA It has been proposed thata genersquos removal to the nucleus decreases its susceptibilityto free radical mutagenesis (Raven et al 1994ab Allen ampRaven 1996) If this is the case then the non-standardcode may be a further indication of an overriding need tomaintain certain genes within the mitochondrion What-ever the reason for the retention of genes in organelles ithad better be a good one bioenergetic organelles areclearly the wrong place to keep genetic information andgo to extraordinary lengths to offset the consequences ofit being there
(ii) Molecular refugeesA phenomenon related to differences in the genetic code
is seen in chloroplasts This is a bias in codon usage Thelsquomolecular refugeesrsquo hypothesis (Howe et al 2000) sug-gests that nuclear copies of what were once chloroplastgenes escape constraints on codon usage and thus thatwhat distinguishes chloroplast-encoded proteins is thattheir amino-acid composition is unusual enough for thebias in codon usage not to apply It is difficult to see whatphysico-chemical properties might be uniquely shared bythe amino acids of chloroplast-encoded proteins and inaddition there seem to be no lsquoforbiddenrsquo amino-acid resi-due in chloroplasts As with the departure from the univer-sal genetic code in mitochondria is seems likely that abias in codon usage reflects some secondary property ofchloroplast genomes and is not the primary reason fortheir retention In any case any restriction on gene move-
Genomes in mitochondria and chloroplasts J F Allen 31
ment will act in the wrong direction to explain why chloro-plasts have retained genomes at all provided there is noresulting obstacle to a chloroplast protein being success-fully encoded as a precursor in the nucleus
(iii) Genes are copied and replicated not relocated in onestep
If complete relocation is envisaged as proceeding by thesimultaneous appearance of a gene in one compartmentand its deletion from another then it is possible to imaginethat whatever the vector is there may be selectivity ingene transfer (Henze amp Martin 2001) For example ifviral incorporation replication and recombination in bothorganelle and nucleus were the mechanism of relocationthen successful gene transfer might depend on the abilityof the virus to complete the process in one step This inturn might depend on some property of the gene or ofthe protein it encodes if expression occurs at any stage inthe process of relocation An assumption of the CORRhypothesis favoured here is that genes are not movedphysically but merely copied after which either the copyin the original location or the copy in the new locationmay be lost as a result of selection A useful analogy maybe the action of copying files between computer storagevolumes In this analogy the copy is identical to the orig-inal and either may subsequently be deleted without lossof information
Because any gene that is not usefully expressed willundergo no selection to maintain a functional allele theprevailing location will be the one in which the functionalcopy of the gene resides The problem of which gene sur-vives is then subsumed into the problem of which is theselectively advantageous location for it It seems reason-able to assume that there wasmdashand ismdashan interim periodin which homologous genes in different compartments areessentially in competition to see which functions best Asoutlined above there is no obvious obstacle for copyingDNA sequences within the cell The physico-chemicalproperties of DNA are fairly uniform and not directlydependent on nucleotide sequence though organelles likeintra-cellular parasites seem to prefer A and T to G andC The answer to the question lsquowhich gene location func-tions bestrsquo may have nothing to do with the gene itselfnor directly with the structure and function of its geneproduct Instead the answer may have everything to dowith the required regulatory interaction between the geneproduct and expression of the gene that produces it
(g) CORRThis hypothesis has been propounded earlier (sect 3) and
will be summarized as follows Vectorial electron and pro-ton transfer exerts regulatory control over expression ofgenes encoding proteins directly involved in or affectingredox poise This regulatory coupling is indispensable foradaptation to environmental changes in chemical activitiesof oxygen light and CO2 and requires co-location of thesegenes with their gene products Organelles lsquomake theirown decisionsrsquo on the basis of environmental changesaffecting redox state The hypothesis is abbreviatedCORR for co-location for redox regulation or more com-pletely lsquoco-location (of gene and gene product) for(evolutionary) continuity of redox regulation of geneexpressionrsquo
Phil Trans R Soc Lond B (2003)
5 SUMMARY OF THE CURRENT STATUS OF THEHYPOTHESIS OF CORR
Q Why do mitochondria and chloroplasts have theirown genetic systems
A CORR co-location of genes and gene products per-mits direct and autonomous redox regulation of geneexpression
(a) Objections to CORR(i) There is no regulation of gene expression in chloroplasts
and mitochondriaThis is still commonly stated but incorrect See the prin-
ciple in sect 3d
(ii) There is no regulation of gene expression in chloroplastsand mitochondria at the level of transcription
For chloroplasts there is now clear evidence for a func-tionally intelligible redox control of transcription(Pfannschmidt et al 1999a) This invalidates the previouswidely held assumption that all control of chloroplast geneexpression is post-transcriptional (Gruissem amp Tonkyn1993 GoldschmidtClermont 1998) The evidence formitochondria is less clear but redox control of transcrip-tion certainly cannot be ruled out and there are indi-cations that it may occur (Pearson et al 1993Konstantinov et al 1995 Wilson et al 1996b) It mustalso be restated that transcriptional control is not neces-sary for CORR but serves as a compelling example of aconserved typically bacterial mode of response that oper-ates still in eukaryotic cytoplasmic organelles thus illus-trating the principle of continuity of redox control (sect 3d)Post-transcriptional redox regulation (Danon amp Mayfield1994 Kim amp Mayfield 1997 Brown et al 2001 Treb-itsh amp Danon 2001) is consistent with CORR and thereis nothing to forbid it being a primary mechanism thatsatisfies most or indeed all of the formal requirements ofthe hypothesis in particular cases
(iii) There are no redox signalling components encoded inchloroplast or mitochondrial DNA
This is what CORR predicts See the principle in sect 3j
(iv) There is redox control of nuclear gene expressionThis is correct and many examples are now known
especially for chloroplasts (Escoubas et al 1995 Kovacs etal 2000 Pfannschmidt et al 2001ab Surpin et al 2002)Redox signals from photosynthetic or respiratory electrontransport chains are not at all prohibited by CORR Theessence of CORR is that there is a subset of organellarproteins for which there is an overriding requirement fora redox regulatory control that is initiated within theorganelle and whose effects are exerted by the simplestand most direct route This redox signalling is envisagedto take place without a requirement for prior consultationand coordination with what may be regarded as a nucleo-cytoplasmic bureaucracy
(v) Nuclear genes can be expressed as rapidly as chloroplastand mitochondrial genes
Zerges (2002) for example cites the equal times takenfor complete expression of nuclear and chloroplast genesin Chlamydomonas as evidence against the CORR hypoth-
32 J F Allen Genomes in mitochondria and chloroplasts
esis However the question is not one of the time takento express a gene but the time taken to alter the rate ofits expression in response to a (redox) signal originatingwithin the organelle An equally important additional fac-tor may be the autonomy of the signalling pathway thatis its independence from other inputs as described in theprevious paragraph One might say that mitochondria andchloroplasts require a free hand
(vi) There is no evidence to support this hypothesisThe hypothesis is rich in predictions (sectsect 3 and 6) and
those that have been tested stringently are in agreementwith experimental results None of the predictions of thehypothesis has been found to be incorrect and none of theunderlying principles (sect 3) has been disproved Furtherpredictions will be the subject of future experiments
(b) Evidence for CORR(i) Chloroplasts have selectively retained cyanobacterial genes
for primary events in photosynthesisSee table 1
(ii) There is regulation of gene expression in chloroplastsThere is transcriptional regulation of gene expression
in chloroplasts (sect 3d) The case for mitochondria is notyet proven
(iii) There is redox regulation of gene expression inchloroplasts
There is transcriptional redox regulation of geneexpression in chloroplasts (sect 3d) The case for mitochon-dria is not yet proven though see (Allen et al 1995 Kon-stantinov et al 1995 Galvis et al 1998)
(iv) There are no redox signalling components encoded inchloroplast or mitochondrial DNA
Correct but with possible exceptions discussed in sect 3j
(v) There are lsquobacterialrsquo redox signalling components ineukaryotes
See sect 3j
(vi) Further predictions DNA-array results and phenotypesof allotopic mutants
A systematic test of CORR might be to make an oxid-ized-minus-reduced gene expression profile of the lsquotran-scriptomersquo of a cyanobacterium to see if the set of geneswhose transcription is under most rapid and direct redoxcontrol corresponds to the genes of the chloroplast gen-ome Laboratories with this experimental capability do notseem to have addressed this possibility (Hihara et al 2002Ikeuchi 2002) Likewise if a putative modern representa-tive of the proto-mitochondrion can be found and agreedeg Rhodospirillum rubrum or Paracoccus denitrificans thena similar experiment should reveal a subset of genes corre-sponding to mitochondrial genomes DNA-microarraysalso present the possibility of testing the relative rate ofredox response and dependency on other signallinginputs of transcription of eukaryotic genes To dateorganellar genes have not been included in such investi-gations (Ferea et al 1999) A further experimental possi-bility is to examine the redox regulatory phenotypes ofexisting allotopic mutants as outlined in sect 3e
Phil Trans R Soc Lond B (2003)
6 CONCLUSION AND FUTURE PROSPECTS
The CORR hypothesis states that the function of mito-chondrial and chloroplast genomes is to provide co-location (of gene and gene product) for (evolutionary)continuity of redox regulation of gene expression Thishypothesis is neither self-contradictory nor devoid of test-able predictions It is also consistent with available evi-dence and in principle makes sense of the otherwisepuzzling distribution of genes between compartments ineukaryotic cells The explanation that CORR offersapplies equally to mitochondria and chloroplasts andidentifies selective forces that still operate to determine thelocation of genes No selective barriers need be assumedto gene transfer protein import nor membrane insertionNor do we need to invoke a lsquofrozen accidentrsquo that some-thing happened to stop gene transfer for some reason atsome time in the remote and inaccessible past CORR alsoexplains the absence of genomes from organelles that havelost their primary bioenergetic functions CORR may actas a stimulus to a search for residual bioenergetic functionin those organelles that have lost aerobic respiration andphotosynthesis but where residual genomes remain
CORR also predicts regulatory properties of knowncomponents of chloroplasts and mitochondria that in turnindicate important but as yet uncharacterized flexibility inenergy transduction These components include the RUB-ISCO large subunit and the CFo and Fo subunits of coup-ling ATPase in both chloroplasts and mitochondriaRedox regulation of the relative stoichiometry of compo-nents as seen in regulation of chloroplast photosystemstoichiometry (Pfannschmidt et al 1999a) may extend to(C)Fo-(C)F1-ATPase indicating that the function ofretention of genes for one or more of these componentsis flexibility in the stoichiometries of H1 to e2 and H1 toATP in chemiosmotic coupling (Schemidt et al 1998Stock et al 2000 Allen 2002)
So far explicit tests of CORR have largely been con-fined to chloroplasts and to some extent mitochondria ofhigher plants but this merely reflects the expertise andinterests of the investigators Further attempted disproofof CORR is welcomed and encouraged Novel experi-mental tests can be devised as well as attempted refu-tation through analysis of existing data It is hoped thatexplicit attention will be given to the CORR hypothesis inlaboratories already suitably equipped and that resourcescan be allocated for this purpose in laboratories wherethere is the interest but not currently the technology orexpertise
The heuristic value of the hypothesis already seen inthe role of redox-regulated transcription of photosyntheticreaction centre genes in chloroplasts may be its most use-ful property The central proposal is that mitochondriaand chloroplasts contain genes for proteins whose functionin electron transfer demands rapid direct and uncon-ditional redox regulatory control of their biosynthesis Thefull implications of this hypothesis are very broad and onlyjust beginning to be explored
I thank Carol Allen Aubrey de Grey William Martin John ARaven and Francis-Andre Wollman for discussion and corre-spondence during the preparation of this paper and WilliamMartin for suggestions including table 1 Research grants from
Genomes in mitochondria and chloroplasts J F Allen 33
The Swedish Natural Sciences Research Council (NFR) andthe Crafoord Foundation are gratefully acknowledged
REFERENCES
Adams K L Daley D O Qiu Y L Whelan J amp PalmerJ D 2000 Repeated recent and diverse transfers of a mito-chondrial gene to the nucleus in flowering plants Nature408 354ndash357
Alberts B Bray D Lewis J Raff M Roberts K amp Wat-son J D 1994 The molecular biology of the cell New Yorkand London Garland Publishing
Allen C A Hakansson G amp Allen J F 1995 Redox con-ditions specify the proteins synthesized by isolated chloro-plasts and mitochondria Redox Rep 1 119ndash123
Allen J F 1992 Protein phosphorylation in regulation ofphotosynthesis Biochim Biophys Acta 1098 275ndash335
Allen J F 1993a Control of gene expression by redox poten-tial and the requirement for chloroplast and mitochondrialgenomes J Theor Biol 165 609ndash631
Allen J F 1993b Redox control of gene expression and thefunction of chloroplast genomes a hypothesis PhotosynthRes 36 95ndash102
Allen J F 1993c Redox control of transcription sensorsresponse regulators activators and repressors FEBS Lett332 203ndash207
Allen J F 1995 Thylakoid proteinndashphosphorylation state-1ndashstate-2 transitions and photosystem stoichiometry adjust-ment redox control at multiple levels of gene expressionPhysiol Plant 93 196ndash205
Allen J F 1996 Separate sexes and the mitochondrial theoryof ageing J Theor Biol 180 135ndash140
Allen J F 2002 Photosynthesis of ATP-electrons protonpumps rotors and poise Cell 110 273ndash276
Allen J F amp Allen C A 1999 A mitochondrial model forpremature ageing of somatically cloned mammals IUBMBLife 48 369ndash372
Allen J F amp Nilsson A 1997 Redox signalling and the struc-tural basis of regulation of photosynthesis by protein phos-phorylation Physiol Plant 100 863ndash868
Allen J F amp Pfannschmidt T 2000 Balancing the two photo-systems photosynthetic electron transfer governs transcrip-tion of reaction centre genes in chloroplasts Phil TransR Soc Lond B 355 1351ndash1357 (DOI 101098rstb20000697)
Allen J F amp Race H L 2002 Will the real LHC II kinaseplease step forward Sciencersquos STKE httpwwwstkeorgcgicontentfullsigtrans2002155pe43
Allen J F amp Raven J A 1996 Free-radical-induced mutationvs redox regulation costs and benefits of genes in organellesJ Mol Evol 42 482ndash492
Archibald J M Cavalier-Smith T Maier U amp Douglas S2001 Molecular chaperones encoded by a reduced nucleusthe cryptomonad nucleomorph J Mol Evol 52 490ndash501
Ashby M K amp Mullineaux C W 1999a Cyanobacterialycf27 gene products regulate energy transfer from phycobili-somes to photosystems I and II FEMS Microbiol Lett 181253ndash260
Ashby M K amp Mullineaux C W 1999b The role of ApcDand ApcF in energy transfer from phycobilisomes to PSI andPSII in a cyanobacterium Photosynth Res 61 169ndash179
Ashby M K Houmard J amp Mullineaux C W 2002 Theycf27 genes from cyanobacteria and eukaryotic algae distri-bution and implications for chloroplast evolution FEMSMicrobiol Lett 214 25ndash30
Attardi G amp Schatz G 1988 Biogenesis of mitochondria ARev Cell Biol 4 289ndash333
Azem A Oppliger W Lustig A Jeno P Feifel B SchatzG amp Horst M 1997 The mitochondrial hsp70 chaperone
Phil Trans R Soc Lond B (2003)
system effect of adenine nucleotides peptide substrate andmGrpE on the oligomeric state of mhsp70 J Biol Chem272 20 901ndash20 906
Barbrook A C Symington H Nisbet R E R LarkumA amp Howe C J 2001 Organisation and expression of theplastid genome of the dinoflagellate Amphidinium opercula-tum Mol Genet Genomics 266 632ndash638
Bauer C E Elsen S amp Bird T H 1999 Mechanisms forredox control of gene expression A Rev Microbiol 53495ndash523
Bauer C E Elsen S Swem L R Swem D L amp MasudaS 2003 Redox and light regulation of gene expression inphotosynthetic prokaryotes Phil Trans R Soc Lond B 358147ndash154 (DOI 101098rstb20021189)
Berks B C Sargent F De Leeuw E Hinsley A P Stan-ley N R Jack R L Buchanan G amp Palmer T 2000aA novel protein transport system involved in the biogenesisof bacterial electron transfer chains Biochim Biophys ActaBioenerg 1459 325ndash330
Berks B C Sargent F amp Palmer T 2000b The tat proteinexport pathway Mol Microbiol 35 260ndash274
Blackstone N W 1999 Redox control in development andevolution evidence from colonial hydroids J Exp Biol 2023541ndash3553
Blackstone N W 2000 Redox control and the evolution ofmulticellularity Bioessays 22 947ndash953
Blackstone N W 2001 Redox state reactive oxygen speciesand adaptive growth in colonial hydroids J Exp Biol 2041845ndash1853
Bogorad L 1975 Evolution of organelles and eukaryotic gen-omes Science 188 891ndash898
Bogsch E G Sargent F Stanley N R Berks B C Robin-son C amp Palmer T 1998 An essential component of a novelbacterial protein export system with homologues in plastidsand mitochondria J Biol Chem 273 18 003ndash18 006
Brown E C Somanchi A amp Mayfield S P 2001 Interor-ganellar crosstalk new perspectives on signaling from thechloroplast to the nucleus Genome Biol 2 10211ndash10214
Cavalier-Smith T 2002 Chloroplast evolution secondarysymbiogenesis and multiple losses Curr Biol 12 R62ndashR64
Chitnis P R amp Thornber J P 1988 The major light-har-vesting complex of photosystem 2 Aspects of its molecularand cell biology Photosynth Res 16 41ndash63
Chow W S Melis A amp Anderson J M 1990 Adjustmentsof photosystem stoichiometry in chloroplasts improve thequantum efficiency of photosynthesis Proc Natl Acad SciUSA 87 7502ndash7506
Claros M G Perea J Shu Y Samatey F A PopotJ L amp Jacq C 1995 Limitations to in vivo import of hydro-phobic proteins into yeast mitochondria The case of a cyto-plasmically synthesized apocytochrome b Eur J Biochem228 762ndash771
Dalbey R E amp Robinson C 1999 Protein translocation intoand across the bacterial plasma membrane and the plant thy-lakoid membrane Trends Biochem Sci 24 17ndash22
Daley D O Adams K L Clifton R Qualmann S MillarA H Palmer J D Pratje E amp Whelan J 2002 Genetransfer from mitochondrion to nucleus novel mechanismsfor gene activation from Cox2 Plant J 30 11ndash21
Danon A amp Mayfield S P 1994 Light-regulated translationof chloroplast messenger RNAs through redox potentialScience 266 1717ndash1719
de Grey A 2000 Mitochondrial gene therapy an arena for thebiomedical use of inteins Trends Biotechnol 18 394ndash399
Deisenhofer J Epp O Miki K Huber R amp Michel H1985 Structure of the protein subunits in the photosyntheticreaction center of Rhodopseudomonas viridis at 3Ac resolutionNature 318 618ndash624
34 J F Allen Genomes in mitochondria and chloroplasts
Delwiche C F 1999 Tracing the thread of plastid diversitythrough the tapestry of life Am Nat 154 S164ndashS177
Doolittle W F 1999 Phylogenetic classification and the uni-versal tree Science 284 2124ndash2129
Douglas S E 1998 Plastid evolution origins diversitytrends Curr Opin Genet Dev 8 655ndash661
Duby G amp Boutry M 2002 Mitochondrial protein importmachinery and targeting information Plant Sci 162 477ndash490
Eberhard S Drapier D amp Wollman F A 2002 Searchinglimiting steps in the expression of chloroplast encoded pro-teins relations between gene copy number transcriptiontranscript abundance and translation rate in the chloroplastof Chlamydomonas reinhardtii Plant J 31 149ndash160
El Bissati K amp Kirilovsky D 2001 Regulation of psbA andpsaE expression by light quality in Synechocystis species PCC6803 A redox control mechanism Plant Physiol 1251988ndash2000
Ellis R J 1984 The nuclear domination of chloroplast devel-opment Science Progress 69 129ndash142
Embley T M amp Martin W 1998 Molecular evolution ahydrogen-producing mitochondrion Nature 396 517ndash519
Ems S C Morden C W Dixon C K Wolfe K HdePamphilis C W amp Palmer J D 1995 Transcriptionsplicing and editing of plastid RNAs in the non-photosyn-thetic plant Epifagus virginiana Plant Mol Biol 29 721ndash733
Eraso J M amp Kaplan S 2000 From redox flow to gene regu-lation role of the PrrC protein of Rhodobacter sphaeroides241 Biochemistry 39 2052ndash2062
Eraso J M amp Kaplan S 2002 Redox flow as an instrumentof gene regulation Meth Enzymol 348 216ndash229
Escoubas J M Lomas M Laroche J amp Falkowski P G1995 Light-intensity regulation of cab gene-transcription issignaled by the redox state of the plastoquinone pool ProcNatl Acad Sci USA 92 10 237ndash10 241
Ferea T L Botstein D Brown P O amp Rosenzweig R F1999 Systematic changes in gene expression patterns follow-ing adaptive evolution in yeast Proc Natl Acad Sci USA96 9721ndash9726
Forsberg J Rosenquist M Fraysse L amp Allen J F 2001Redox signalling in chloroplasts and mitochondria genomicand biochemical evidence for two-component regulatory sys-tems in bioenergetic organelles Biochem Soc Trans 29403ndash407
Fujita Y 1997 A study on the dynamic features of photosys-tem stoichiometry accomplishments and problems forfuture studies Photosynth Res 53 83ndash93
Gabriel K Buchanan S K amp Lithgow T 2001 The alphaand the beta protein translocation across mitochondrial andplastid outer membranes Trends Biochem Sci 26 36ndash40
Galvis M L E Allen J F amp Hakansson G 1998 Proteinsynthesis by isolated pea mitochondria is dependent on theactivity of respiratory complex II Curr Genet 33 320ndash329
Gatenby A A amp Ellis R J 1990 Chaperone function theassembly of ribulose bisphosphate carboxylase-oxygenase ARev Cell Biol 6 125ndash149
GoldschmidtClermont M 1998 Coordination of nuclear andchloroplast gene expression in plant cells Int Rev CytolSurv Cell Biol 177 115ndash180
Gray M W 1992 The endosymbiont hypothesis revisited IntRev Cytol 141 233ndash357
Gray M W 1999 Evolution of organellar genomes CurrOpin Genet Dev 9 678ndash687
Gray M W 2000 Evolutionary biology mitochondrial geneson the move Nature 408 302ndash305
Gray M W amp Doolittle W F 1982 Has the endosymbionthypothesis been proven Microbiol Rev 46 1ndash42
Gray M W Burger G amp Lang B F 1999 Mitochondrialevolution Science 283 1476ndash1481
Phil Trans R Soc Lond B (2003)
Grebe T W amp Stock J B 1999 The histidine protein kinasesuperfamily Adv Microb Physiol 41 139ndash227
Green B R amp Kuhlbrandt W 1995 Sequence conservationof light-harvesting and stress-response proteins in relation tothe 3-dimensional molecular structure of Lhcii PhotosynthRes 44 139ndash148
Gruissem W amp Tonkyn J C 1993 Control mechanisms ofplastid gene expression Crit Rev Plant Sci 12 19ndash55
Hackstein J H P (and 11 others) 2001 Hydrogenosomesconvergent adaptations of mitochondria to anaerobicenvironments Zool Anal Complex Syst 104 290ndash302
Harrison M A Keen J N Findlay J B C amp Allen J F1990 Modification of a Glnb-like gene-product by photosyn-thetic electron-transport in the cyanobacterium Synechoc-occus 6301 FEBS Lett 264 25ndash28
Harvey A J Kind K L amp Thompson J G 2002 Redoxregulation of early embryo development Reproduction 123479ndash486
Heathcote P Fyfe P K amp Jones M R 2002 Reactioncentres the structure and evolution of biological solarpower Trends Biochem Sci 27 79ndash87
Henze K amp Martin W 2001 How do mitochondrial genesget into the nucleus Trends Genet 17 383ndash387
Herrmann R G amp Westhoff P 2001 Thylakoid biogenesis anddynamics the result of a complex phylogenetic puzzle InRegulation of photosynthesis vol 11 (ed E-M Aro amp BAndersson) pp 1ndash28 Dordrecht The Netherlands Kluwer
Hihara Y Sonoike K Kanehisa M amp Ikeuchi M 2002Search for redox responsive genes in Synechocystis sp PCC6803 by using DNA microarrays Plant Cell Physiol 43S178
Howe C J Barbrook A C amp Lockhart P J 2000 Organellegenes do they jump or are they pushed Trends Genet 1665ndash66
Ikeuchi M 2002 Gene expression and responses to light stressin cyanobacteria Plant Cell Physiol 43 S21
Iuchi S amp Lin E C C 1992 Mutational analysis of signaltransduction by Arcb a membrane sensor protein respon-sible for anaerobic repression of operons involved in the cen-tral aerobic pathways in Escherichia coli J Bacteriol 1743972ndash3980
Jackson-Constan D Akita M amp Keegstra K 2001 Molecu-lar chaperones involved in chloroplast protein importBiochim Biophys Acta Mol Cell Res 1541 102ndash113
Jarvis P amp Soll M 2001 Toc Tic and chloroplast proteinimport Biochim Biophys Acta Mol Cell Res 1541 64ndash79
Jordan P Fromme P Witt H T Klukas O SaengerW amp Krauss N 2001 Three-dimensional structure ofcyanobacterial photosystem I at 25 acngstrom resolutionNature 411 909ndash917
Kaneko T (and 23 others) 1996a Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regionsDNA Res 3 109ndash136
Kaneko T (and 23 others) 1996b Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regions[Supplement] DNA Res 3 185ndash209
Kanevski I amp Maliga P 1994 Relocation of the plastid rbcLgene to the nucleus yields functional ribulose-15-bisphosph-ate carboxylase in tobacco chloroplasts Proc Natl Acad SciUSA 91 1969ndash1973
Kim J amp Mayfield S P 1997 Protein disulfide isomerase as aregulator of chloroplast translational activation Science 2781954ndash1957
Genomes in mitochondria and chloroplasts J F Allen 35
Kirk J T O amp Tilney-Bassett R A E 1978 The plastidsTheir chemistry structure growth and inheritance Amster-dam Elsevier
Koehler C M Merchant S amp Schatz G 1999 How mem-brane proteins travel across the mitochondrial intermem-brane space Trends Biochem Sci 24 428ndash432
Komiya T Rospert S Schatz G amp Mihara K 1997 Bind-ing of mitochondrial precursor proteins to the cytoplasmicdomains of the import receptors Tom70 and Tom20 isdetermined by cytoplasmic chaperones FASEB J 11 1196
Konstantinov Y M Lutsenko G N amp Podsosonny V A1995 Genetic functions of isolated maize mitochondriaunder model changes of redox conditions Biochem MolBiol Int 36 319ndash326
Korab-Laskowska M Rioux P Brossard N LittlejohnT G Gray M W Lang B F amp Burger G 1998 Theorganelle genome database project (GOBASE) Nucleic AcidsRes 26 138ndash144
Kovacs L Wiessner W Kis M Nagy F Mende D ampDemeter S 2000 Short- and long-term redox regulation ofphotosynthetic light energy distribution and photosystemstoichiometry by acetate metabolism in the green alga Chla-mydobotrys stellata Photosynth Res 65 231ndash247
Kuhlbrandt W 1994 Structure and function of the plant light-harvesting complex LHC-II Curr Opin Struct Biol 4519ndash528
Lang B F Gray M W amp Burger G 1999 Mitochondrialgenome evolution and the origin of eukaryotes A RevGenet 33 351ndash397
Lemire B D Fankhauser C Baker A amp Schatz G 1989The mitochondrial targeting function of randomly generatedpeptide sequences correlates with predicted helical amphi-philicity J Biol Chem 264 20 206ndash20 215
Leuenberger D Bally N A Schatz G amp Koehler C M1999 Different import pathways through the mitochondrialintermembrane space for inner membrane proteins EMBOJ 18 4816ndash4822
Li H amp Sherman L A 2000 A redox-responsive regulator ofphotosynthesis gene expression in the cyanobacterium Syne-chocystis sp strain PCC 6803 J Bacteriol 182 4268ndash4277
Lin X Y (and 36 others) 1999 Sequence and analysis of chro-mosome 2 of the plant Arabidopsis thaliana Nature 402761ndash768
Lonsdale D M Hodge T P Howe C J amp Stern D B1983 Maize mitochondrial-DNA contains a sequence hom-ologous to the ribulose-15-bisphosphate carboxylase largesubunit gene of chloroplast DNA Cell 34 1007ndash1014
McFadden G I 1999 Endosymbiosis and evolution of theplant cell Curr Opin Plant Biol 2 513ndash519
McFadden G I 2001 Chloroplast origin and integrationPlant Physiol 125 50ndash53
Mahler H R amp Raff R A 1975 The evolutionary origin ofthe mitochondrion a nonsymbiotic model Int Rev Cytol43 1ndash124
Manfredi G Fu J Ojaimi J Sadlock J E Kwong J QGuy J amp Schon E A 2002 Rescue of a deficiency in ATPsynthesis by transfer of MTATP6 a mitochondrial DNA-encoded gene to the nucleus Nature Genet 30 394ndash399
Marc P Margeot A Devaux F Blugeon C Corral-Debrinski M amp Jacq C 2002 Genome-wide analysis ofmRNAs targeted to yeast mitochondria EMBO Rep 3159ndash164
Marienfeld J Unseld M amp Brennicke A 1999 The mito-chondrial genome of Arabidopsis is composed of both nativeand immigrant information Trends Plant Sci 4 495ndash502
Martin W 1999 A briefly argued case that mitochondria andplastids are descendants of endosymbionts but that thenuclear compartment is not Proc R Soc Lond B 2661387ndash1395 (DOI 101098rspb19990792)
Phil Trans R Soc Lond B (2003)
Martin W amp Herrmann R G 1998 Gene transfer fromorganelles to the nucleus how much what happens andwhy Plant Physiol 118 9ndash17
Martin W amp Muller M 1998 The hydrogen hypothesis forthe first eukaryote Nature 392 37ndash41
Martin W amp Russell M J 2003 On the origins of cells ahypothesis for the evolutionary transitions from abiotic geo-chemistry to chemoautotrophic prokaryotes and from pro-karyotes to nucleated cells Phil Trans R Soc Lond B 35859ndash85 (DOI 101098rstb20021183)
Martin W Stoebe B Goremykin V Hansmann S Hase-gawa M amp Kowallik K V 1998 Gene transfer to thenucleus and the evolution of chloroplasts Nature 393162ndash165
Martin W Hoffmeister M Rotte C amp Henze K 2001 Anoverview of endosymbiotic models for the origins of eukary-otes their ATP-producing organelles (mitochondria andhydrogenosomes) and their heterotrophic lifestyle BiolChem 382 1521ndash1539
Martin W Rujan T Richly E Hansen A Cornelsen SLins T Leister D Stoebe B Hasegawa M amp PennyD 2002 Evolutionary analysis of Arabidopsis cyanobacterialand chloroplast genomes reveals plastid phylogeny and thou-sands of cyanobacterial genes in the nucleus Proc NatlAcad Sci USA 99 12 246ndash12 251
Millen R S (and 12 others) 2001 Many parallel losses of infAfrom chloroplast DNA during angiosperm evolution withmultiple independent transfers to the nucleus Plant Cell 13645ndash658
Morse D Salois P Markovic P amp Hastings J W 1995 Anuclear-encoded form-II RUBISCO in dinoflagellatesScience 268 1622ndash1624
Murakami A Kim S J amp Fujita Y 1997 Changes in photo-system stoichiometry in response to environmental con-ditions for cell growth observed with the cyanophyteSynechocystis PCC 6714 Plant Cell Physiol 38392ndash397
Oh J I amp Kaplan S 2000 Redox signaling globalization ofgene expression EMBO J 19 4237ndash4247
Ozawa T 1995 Mitochondrial-DNA mutations associatedwith aging and degenerative diseases Exp Gerontol 30269ndash290
Palmer J D 1997 Organelle genomes going going goneScience 275 790ndash791
Pearson C K Wilson S B Schaffer R amp Ross A W 1993NAD turnover and utilization of metabolites for RNA syn-thesis in a reaction sensing the redox state of the cyto-chrome-b( 6 )f complex in isolated chloroplasts Eur JBiochem 218 397ndash404
Pfanner N amp Geissler A 2001 Versatility of the mitochon-drial protein import machinery Natl Rev Mol Cell Biol 2339ndash349
Pfanner N amp Wiedemann N 2002 Mitochondrial proteinimport two membranes three translocases Curr Opin CellBiol 14 400ndash411
Pfannschmidt T Nilsson A amp Allen J F 1999a Photosyn-thetic control of chloroplast gene expression Nature 397625ndash628
Pfannschmidt T Nilsson A Tullberg A Link G amp AllenJ F 1999b Direct transcriptional control of the chloroplastgenes psbA and psaAB adjusts photosynthesis to light energydistribution in plants IUBMB Life 48 271ndash276
Pfannschmidt T Allen J F amp Oelmuller R 2001a Prin-ciples of redox control in photosynthesis gene expressionPhysiol Plant 112 1ndash9
Pfannschmidt T Schutze K Brost M amp Oelmuller R2001b A novel mechanism of nuclear photosynthesis generegulation by redox signals from the chloroplast during pho-
36 J F Allen Genomes in mitochondria and chloroplasts
tosystem stoichiometry adjustment J Biol Chem 27636 125ndash36 130
Popot J L amp de Vitry C 1990 On the microassembly of inte-gral membrane proteins A Rev Biophys Biophys Chem 19369ndash403
Race H L Herrmann R G amp Martin W 1999 Why haveorganelles retained genomes Trends Genet 15 364ndash370
Raven J A Johnston A M Parsons R amp Kubler J 1994aThe influence of natural and experimental high O2 concen-trations on O2 evolving phototrophs Biol Rev CambridgePhil Soc 69 61ndash94
Raven J A Johnston A M Parsons R amp Kubler J 1994bThe occurrence and influence on photolithotrophs of highoxygen concentrations Proc R Soc Edinb B 102 193ndash201
Robinson C 2000 The twin-arginine translocation system anovel means of transporting folded proteins in chloroplastsand bacteria Biol Chem 381 89ndash93
Robinson C amp Bolhuis A 2001 Protein targeting by the twin-arginine translocation pathway Natl Rev Mol Cell Biol 2350ndash356
Rodermel S 2001 Pathways of plastid-to-nucleus signalingTrends Plant Sci 6 471ndash478
Roger A J 1999 Reconstructing early events in eukaryoticevolution Am Nat 154 S146ndashS163
Roger A J Svard S G Tovar J Clark C G SmithM W Gillin F D amp Sogin M L 1998 A mitochondrial-like chaperonin 60 gene in Giardia lamblia evidence thatdiplomonads once harbored an endosymbiont related to theprogenitor of mitochondria Proc Natl Acad Sci USA 95229ndash234
Roodyn D B amp Wilkie D 1968 The biogenesis of mitochondriaMethuenrsquos monographs on biological subjects London Methuen
Rujan T amp Martin W 2001 How many genes in Arabidopsiscome from cyanobacteria An estimate from 386 proteinphylogenies Trends Genet 17 113ndash120
Saccone C Gissi C Lanave C Larizza A Pesole G ampReyes A 2000 Evolution of the mitochondrial genetic sys-tem an overview Gene 261 153ndash159
Salzberg S L White O Peterson J amp Eisen J A 2001Microbial genes in the human genome lateral transfer orgene loss Science 292 1903ndash1906
Sargent F Berks B C amp Palmer T 2002 Assembly of mem-brane-bound respiratory complexes by the Tat protein-trans-port system Arch Microbiol 178 77ndash84
Schatz G 1998 Protein transport the doors to organellesNature 395 439ndash440
Schemidt R A Qu J Williams J R amp Brusilow W S1998 Effects of carbon source on expression of F0 genes andon the stoichiometry of the c subunit in the F1 F0 ATPaseof Escherichia coli J Bacteriol 180 3205ndash3208
Selosse M A Albert B R amp Godelle B 2001 Reducing thegenome size of organelles favours gene transfer to thenucleus Trends Ecol Evol 16 135ndash141
Stern D B amp Lonsdale D M 1982 Mitochondrial and chlo-roplast genomes of maize have a 12- kilobase DNA-sequencein common Nature 299 698ndash702
Stock D Gibbons C Arechaga I Leslie A G W ampWalker J E 2000 The rotary mechanism of ATP synthaseCurr Opin Struct Biol 10 672ndash679
Stoebe B amp Maier U G 2002 One two three naturersquos toolbox for building plastids Protoplasma 219 123ndash130
Stoebe B Martin W amp Kowallik K V 1998 Distributionand nomenclature of protein-coding genes in 12 sequencedchloroplast genomes Plant Mol Biol Rep 16 243ndash255
Stupar R M Lilly J W Town C D Cheng Z Kaul SBuell C R amp Jiang J M 2001 Complex mtDNA consti-tutes an approximate 620 kb insertion on Arabidopsis thali-ana chromosome 2 implication of potential sequencing
Phil Trans R Soc Lond B (2003)
errors caused by large-unit repeats Proc Natl Acad SciUSA 98 5099ndash5103
Surpin M Larkin R M amp Chory J 2002 Signal transduc-tion between the chloroplast and the nucleus Plant Cell 14S327ndashS338
Tjaden J Schwoppe C Mohlmann T Quick P W ampNeuhaus H E 1998 Expression of a plastidic ATPADPtransporter gene in Escherichia coli leads to a functional aden-ine nucleotide transport system in the bacterial cytoplasmicmembrane J Biol Chem 273 9630ndash9636
Tovar J Fischer A amp Clark C G 1999 The mitosome a novelorganelle related to mitochondria in the amitochondrial para-site Entamoeba histolytica Mol Microbiol 32 1013ndash1021
Trebitsh T amp Danon A 2001 Translation of chloroplastpsbA mRNA is regulated by signals initiated by both photos-ystems II and I Proc Natl Acad Sci USA 98 12 289ndash12 294
Tullberg A Alexciev K Pfannschmidt T amp Allen J F2000 Photosynthetic electron flow regulates transcription ofthe psaB gene in pea (Pisum sativum L) chloroplasts throughthe redox state of the plastoquinone pool Plant Cell Physiol41 1045ndash1054
Turmel M Lemieux C Burger G Lang B F Otis CPlante I amp Gray M W 1999 The complete mitochondrialDNA sequences of Nephroselmis olivacea and Pedinomonasminor two radically different evolutionary patterns withingreen algae Plant Cell 11 1717ndash1729
Unden G amp Bongaerts J 1997 Alternative respiratory path-ways of Escherichia coli energetics and transcriptional regu-lation in response to electron acceptors Biochim BiophysActa Bioenerg 1320 217ndash234
Unseld M Marienfeld J R Brandt P amp Brennicke A1997 The mitochondrial genome of Arabidopsis thaliana con-tains 57 genes in 366 924 nucleotides Nature Genet 1557ndash61
Van der Giezen M Slotboom D J Horner D S DyalP L Harding M Xue G P Embley T M amp KunjiE R S 2002 Conserved properties of hydrogenosomal andmitochondrial ADPATP carriers a common origin for bothorganelles EMBO J 21 572ndash579
Von Heijne G 1986 Why mitochondria need a genome FEBSLett 198 1ndash4
Westphal S Soll J amp Vothknecht U C 2001 A vesicle trans-port system inside chloroplasts FEBS Lett 506 257ndash261
Whitehouse H L K 1969 Towards an understanding of themechanism of heredity London Edward Arnold Ltd
Williams B A P Hirt R P Lucocq J M amp Embley T M2002 A mitochondrial remnant in the microsporidian Trachi-pleistophora hominis Nature 418 865ndash869
Wilson R J (and 10 others) 1996a Complete gene map of theplastid-like DNA of the malaria parasite Plasmodium falcipa-rum J Mol Biol 261 155ndash172
Wilson S B Davidson G S Thomson L M amp PearsonC K 1996b Redox control of RNA synthesis in potato mito-chondria Eur J Biochem 242 81ndash85
Winkler H H amp Neuhaus H E 1999 Non-mitochondrialATP transport Trends Biochem Sci 24 64ndash68
Wolfe K H Morden C W amp Palmer J D 1992 Small sin-gle-copy region of plastid DNA in the non-photosyntheticangiosperm Epifagus virginiana contains only two genes Dif-ferences among dicots monocots and bryophytes in geneorganization at a non-bioenergetic locus J Mol Biol 22395ndash104
Wollman F A Minai L amp Nechushtai R 1999 The biogen-esis and assembly of photosynthetic proteins in thylakoidmembranes Biochim Biophys Acta Bioenerg 1411 21ndash85
Yeliseev A A amp Kaplan S 2000 TspO of Rhodobactersphaeroides a structural and functional model for the mam-
Genomes in mitochondria and chloroplasts J F Allen 37
malian peripheral benzodiazepine receptor J Biol Chem275 5657ndash5667
Zerges W 2002 Does complexity constrain organelle evol-ution Trends Plant Sci 7 175ndash182
Zhang Z D Green B R amp Cavalier-Smith T 1999 Singlegene circles in dinoflagellate chloroplast genomes Nature400 155ndash159
Zhang Z D Cavalier-Smith T amp Green B R 2001 A fam-ily of selfish minicircular chromosomes with jumbled chloro-plast gene fragments from a dinoflagellate Mol Biol Evol18 1558ndash1565
Zouni A Witt H T Kern J Fromme P Krauss NSaenger W amp Orth P 2001 Crystal structure of photosys-tem II from Synechococcus elongatus at 38 acngstrom resol-ution Nature 409 739ndash743
DiscussionD S Horner (Dipartimento di Fisiologia e Biochimica
Generali University of Milan Milan Italy) Given thatthere are components of these membrane-bound energy-transduction pathways that are consistently nuclear enco-ded not the key ones you pointed out but others do youexpect perhaps a slower perhaps a more crude but yet aredox communication between the organelle andexpression of nuclear-encoded genes
J F Allen There is and there is no question about itI think that Bogoradrsquos lock-in hypothesis has somethingto say here and there are other people who say it is all todo with assembling multi-subunit complexes If you havecontrol over the initial construction of the core aroundwhich the whole complex is eventually built then redoxcontrol or some sort of control over synthesis of that corewill control construction of the whole thing eventuallyand I think that may well be largely correct I think it ispossible in principle for nuclear genes to make precursorsaccording to redox instructions but it is not really soimportant that those precursors are here and now presentin exactly the right quantities because the precursors canto a limited extent hang around and wait Actually thereis proteolytic degradation of unattached precursors and soon but I think the fact that these chloroplast and mito-chondrially DNA-encoded subunits are at the core ofthese energy-transducing complexes is the crucial issue
T Cavalier-Smith (Department of Zoology University ofOxford Oxford UK ) In the case of mitochondria the sub-set of genes that remains is much more variable than youimplied It runs from three proteins in the minimal casesup to I think over 90 That is a big difference and manyof the genes in different lineages of protists that remainhave nothing to do with redox control they are things likeribosomal proteins and so on So I think that even thoughwhat you say about redox control of certain genes mayactually be an important thing it is not the fundamentaloverall reason why the genomes are maintained It may bepart of the explanation for why certain genes are alwaysincluded in that subset but even in mitochondria I thinkthat cytochrome b is the only one that is always presentSo I think that both this great diversity of what has beenretained in different lineages and the nucleomorph caseindicate that evolution was able to do one thing in onelineage 500 million years ago and never got around todoing it for 500 million years in another lineage so thereis a lot more haphazardness and evolutionary accidentsand contingency than a straight functional interpretation
Phil Trans R Soc Lond B (2003)
there is a lot of just sheer lsquohappenedndashorndashdidnrsquotndashhappenrsquoor carryover
J F Allen I was not implying anything about the rangeof mitochondrial genomes I have a limited amount oftime so I just talked about what I could talk about withinthat limit Of course there will be a range and this willdepend upon the physiology and bioenergetics of theparticular organism But what I am trying to do is to seewhat the common properties of the irreducible core areof components that are always encoded in the organellesand cytochrome b is a perfect example because that reallyis the thing that binds these two all-important quinoneswithout which there is no Q cycle without which there isno proton pumping and bioenergetics and in Plasmodiumwhere the mitochondrion is reduced to this bare mini-mum there is still a mitochondrial gene for cytochromeb Now my argument about the ribosomal proteins andall the rest of it is that in order to be able to exert thisall-important redox control over gene expression of certaincore components you need an in situ genetic system andthis can vary in composition for reasons that I cannotdescribe Maybe you need RNA polymerase or maybeyou can import RNA polymerase In plastids you haveboth of these options You are going to need ribosomestransfer RNAs and the rest of the apparatus of geneexpression and that needs to be in situ but we are stillleft with the question why does there have to be any gen-etic system at all The mature tobacco or spinach chloro-plast seems to have an entire genetic system just to makeone protein D1 which is all it ever needs to resynthesizeI think that this is a clue It is worth having this very unec-onomical crazy system because there is an overridingrequirement to connect electron transfer with geneexpression directly and unambiguouslymdashwithout recourseto a bureaucracy if you like which is what I think thenucleus is
K Willison (Institute of Cancer Research Chester BeattyLaboratories London UK ) These photosynthetic reactioncomplexes are a sort of baroque mixture of polypeptidechains and cofactors and my understanding is that someof them in high photon flux can turn over within say 30minutes so I am wondering if there is a correlationbetween gene retention and the turnover rates of some ofthe components and the complexes it seems to me thatthat could really be a limiting factor and the reason thatthe proteins had to be encoded locally in order to get thecomplexes to be assembled rapidly
J F Allen I think that is right You can only alter thesteady-state concentration of a protein complex by affect-ing the synthesis and breakdown and therefore turnoverThere is an extreme example the D1 protein encoded bythe psbA gene of chloroplasts which is always in the chlor-oplast genome without exception High light will tend todegrade this protein and when the high light goes awaythere is an absolute need very quickly to bring it back upagain I think that is a very good example Whether thisargument applies to genes such as cytochrome oxidase Ido not know
C J Leaver (Department of Plant Sciences University ofOxford Oxford UK ) Your experiments tend to relatemainly to the gene expression side What about proteinRNA turnover the whole concept of redox control ofthatmdashdo we know anything
38 J F Allen Genomes in mitochondria and chloroplasts
J F Allen Well quite a lot is known about chloro-plasts There is redox control of messenger RNA stabilityfor this same D1 protein which has been described byStephen Mayfield and his co-workers The evidence is thatthe site of redox control is ferredoxin via thioredoxin andthe implication is that thioredoxin tells whether non-cyclicelectron transport is working at all When it is workingthere is a signal that says lsquoOK now continue with all theprotein synthesisrsquo It is not particularly specific but moreof a global effect I think this is what Mayfield argues butof course this is not inconsistent with my argument I haveconcentrated on quinone pools and medium-potentialredox regulatory control because that is where I can seesignals about the dangerous consequences of getting itwrong but there is control of other levels of geneexpression In mitochondria I am not so sure I wouldbounce the question back to you on that one
W Martin (Institute of Botany III Heinrich-Heine Univ-ersitat Dusseldorf Dusseldorf Germany) The nucleomorphgenome was cited as evidence in conflict with your modelbut is that true The nucleomorph genome is not in abioenergetic organelle in that sense It resides in the cyto-sol so does the size of the nucleomorph genome workagainst your model Is it really evidence against yourmodel
J F Allen I do not know why should it be evidenceI cannot explain why the nucleomorph has a genomeThat is why I clutched at the straw that it is storage fornucleic acid precursors
Phil Trans R Soc Lond B (2003)
T Cavalier-Smith The reason I introduced the nucleo-morph is not to say it is direct evidence against yourhypothesis but to show that there probably was no singlefunctional reason for the retention of every gene in everyorganelle It may be that there are specific physiologicalreasons for the retention of certain genes and thereforeyou might be right about a subset of genes but I verymuch doubt if there are specific physiological reasons forthe retention of every single gene that is retained differen-tially because odd things happen in evolution Somethings are easy and sometimes things are difficult Evol-ution is history
J F Allen Of course I agree with that I do not thinkI can explain everything but I do think I can explainsomething If I have done that then I am content
GLOSSARY
AAC ADPndashATP carrierCES control by epistasy of synthesisCORR co-location for redox regulationLHC light-harvesting complexPCC Pasteur culture collectionORF open reading frameRUBISCO ribulose-15-bisphosphate carboxylasendashoxygenaseTAT twin-arginine translocase
Visit httpwwwpubsroyalsocacuk to see an electronic appendix tothis paper
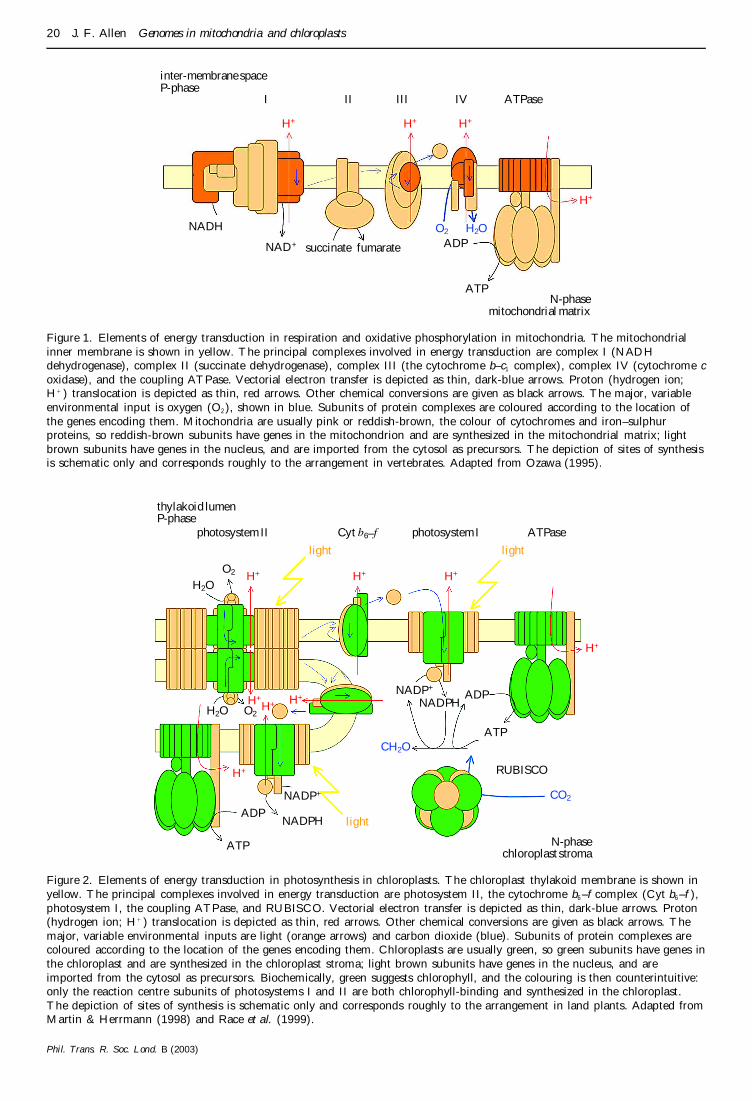
20 J F Allen Genomes in mitochondria and chloroplasts
I II III IV ATPase
H+ H+ H+
NADH
NAD+ succinate fumarate ADP
ATP
O2 H2O
H+
inter-membrane spaceP-phase
N-phasemitochondrial matrix
Figure 1 Elements of energy transduction in respiration and oxidative phosphorylation in mitochondria The mitochondrialinner membrane is shown in yellow The principal complexes involved in energy transduction are complex I (NADHdehydrogenase) complex II (succinate dehydrogenase) complex III (the cytochrome bndashc1 complex) complex IV (cytochrome coxidase) and the coupling ATPase Vectorial electron transfer is depicted as thin dark-blue arrows Proton (hydrogen ionH1 ) translocation is depicted as thin red arrows Other chemical conversions are given as black arrows The major variableenvironmental input is oxygen (O2 ) shown in blue Subunits of protein complexes are coloured according to the location ofthe genes encoding them Mitochondria are usually pink or reddish-brown the colour of cytochromes and ironndashsulphurproteins so reddish-brown subunits have genes in the mitochondrion and are synthesized in the mitochondrial matrix lightbrown subunits have genes in the nucleus and are imported from the cytosol as precursors The depiction of sites of synthesisis schematic only and corresponds roughly to the arrangement in vertebrates Adapted from Ozawa (1995)
ATP
ADP
H+
NADP+
NADPH
CH2OATP
ADPNADP+
NADPHH2O O2
H+H+ H+
H+O2
H2OH+ H+
CO2
RUBISCO
H+
photosystem II Cyt b6_f photosystem I ATPase
thylakoid lumenP-phase
N-phasechloroplast stroma
light light
light
Figure 2 Elements of energy transduction in photosynthesis in chloroplasts The chloroplast thylakoid membrane is shown inyellow The principal complexes involved in energy transduction are photosystem II the cytochrome b6 ndashf complex (Cyt b6 ndashf )photosystem I the coupling ATPase and RUBISCO Vectorial electron transfer is depicted as thin dark-blue arrows Proton(hydrogen ion H1 ) translocation is depicted as thin red arrows Other chemical conversions are given as black arrows Themajor variable environmental inputs are light (orange arrows) and carbon dioxide (blue) Subunits of protein complexes arecoloured according to the location of the genes encoding them Chloroplasts are usually green so green subunits have genes inthe chloroplast and are synthesized in the chloroplast stroma light brown subunits have genes in the nucleus and areimported from the cytosol as precursors Biochemically green suggests chlorophyll and the colouring is then counterintuitiveonly the reaction centre subunits of photosystems I and II are both chlorophyll-binding and synthesized in the chloroplastThe depiction of sites of synthesis is schematic only and corresponds roughly to the arrangement in land plants Adapted fromMartin amp Herrmann (1998) and Race et al (1999)
Phil Trans R Soc Lond B (2003)
Genomes in mitochondria and chloroplasts J F Allen 21
Table 1 Retention of cyanobacterial genes in chloroplast DNA(Protein-coding genes are sorted into the functional categories assigned by Kaneko et al (1996ab) The genes are named andtheir distribution recorded in 16 completely sequenced chloroplast genomes to date in Martin et al (2002) from which the dataare taken Electronic Appendix A (available on The Royal Societyrsquos Publications Web site) lists the genes sorted into the functionalcategories of their gene products GPC genes per functional category The number of genes present in each category in thecyanobacterium Synechocystis PCC 6803 (Syn) and in any chloroplast genome completely sequenced (cp) Cat Ret categoryretention The number of times that a Synechocystis functional category is represented by a gene in cpDNA divided by the numberof genes in that category in Synechocystis itself Cat Ret = cpSyn For example Synechocystis has 84 genes for amino-acid biosynth-esis 10 of which are still present in at least one of 16 sequenced plastid genomes Hence Cat Ret = 1084 = 119 for amino-acid biosynthesis Deg Ret degree of retention The number of genes present in 16 chloroplast genomes divided by 16 andmultiplied by the number of times that the functional category is represented in chloroplast DNA For example in the 16 genomesanalysed there are 10 different amino-acid biosynthetic genes that occur in at least one cpDNA But of the 160 elements of theresulting matrix that could be positive only 27 are Hence Deg Ret = 27(16 acute 10) = 169 for amino-acid biosynthesis F(ret)fraction of retention The number of genes present in 16 chloroplast genomes divided by 16 and multiplied by the number oftimes that the functional category is represented in Synechocystis For example in the 16 genomes sequenced 27 different amino-acid biosynthetic genes occur in cpDNA Synechocystis has 84 genes for amino-acid biosynthesis that can be assumed to havebeen present in the common ancestor of these 16 plastid genomes Hence for amino-acid biosynthesisF(ret) = 27(16 acute 84) = 20 The genes analysed here correspond to those tabulated by Martin et al (2002) Only genes withknown homologues in cyanobacterial genomes were included in the analysis Values were calculated from the supplementarymaterial of Martin et al (2002) which was downloaded from httpwwwpnasorgcgicontentfull182432999DCI at the PNASWeb site)
no of GPC
Syn cp Cat Ret () Deg Ret () F(ret) ()
amino-acid biosynthesis 84 10 119 169 20cofactorprosthetic group biosynthesis 108 21 194 202 39cell envelope 64 7 109 250 27cellular processes 68 19 279 210 59central metabolism 31 1 32 63 02energy metabolism 86 3 35 125 04fatty-acid biosynthesis 35 8 229 172 39photosynthesis and respiration 139 74 532 603 321photosystem I 12 11 917 625 573photosystem II 25 20 800 806 645purine and pyrimidine biosynthesis 39 1 26 63 02regulatory functions 136 7 51 178 09DNA replication 49 2 41 156 06transcription 23 4 174 1000 130translation 144 56 389 531 207transport 158 2 13 63 01other plus hypothetical 587 43 73 202 15
(a) The bacterial nature of mitochondria andchloroplasts
It is generally accepted that organellar genetic systemsare evidence for the evolutionary origin of mitochondriaand chloroplasts as free-living eubacteria or a-proteobac-teria which entered into close inter-dependency with ahost cell (Douglas 1998 Delwiche 1999 Lang et al 1999McFadden 1999 2001 Roger 1999 Hackstein et al2001 Stoebe amp Maier 2002) The genetic systemsretained by organelles have many distinctively bacterialfeatures and heterologous systems for gene expression canbe reconstituted in vitro from organellar and bacterialcomponents By sharp contrast the genetic systems of theeukaryotic cell nucleus and cytosol are only distantlyrelated to those of bacteria and to those of their own cyto-plasmic organelles Instead nucleo-cytoplasmic geneticsystems more closely resemble the genetic systems ofarchaea (also known as archaebacteria) It is has been sug-gested that eukaryotic cells are an archaeal system of infor-mation storage replication transcription and translationthat is powered by a eubacterial system of energy trans-
Phil Trans R Soc Lond B (2003)
duction (Martin amp Russell 2003) The genetic systems ofmitochondria and chloroplasts can then be viewed aseukaryotic cellsrsquo residual bacterial systems of informationstorage and transmission Eukaryotic cellsrsquo residualarchaeal systems of energy transduction might theninclude direct vectorial ion pumps such as P- and V-ATP-ases and seven-helix transporters now associated prim-arily with information and signalling However the centralprocesses of energy transduction respiration and photo-synthesis are clearly bacterial in origin
(b) Currencies of symbiosisOne hypothesis for the origin of mitochondria states
that the nucleo-cytoplasmic system of eukaryotes isderived from that of an archaeal methanogenic chemo-autotroph that came to rely on the precursor of mitochon-dria for its supply of molecular hydrogen (Martin ampMuller 1998) This lsquohydrogen hypothesis for the firsteukaryotersquo resolves a long-held but paradoxical feature ofthe endosymbiont hypothesis how could a heterotrophichost cell have been able to supply abundant organic sub-
22 J F Allen Genomes in mitochondria and chloroplasts
strates that the bacterial precursor of mitochondria couldnot already obtain for itself Furthermore a second para-dox is resolved or at a least avoided export of ATP fromthe bacterial partner was not an immediate requirementfor the symbiosis to become established and could haveevolved subsequently Thus it is not necessary to postu-late a suicidal bacterium that excretes ATP nor a deferredselective advantage to the partnership implying foresightor an assurance of eventual contract fulfilment
The hydrogen hypothesis (Martin amp Muller 1998) seeksto provide an explanation of the origin of mitochondriathat accounts for the immediate benefit of the primordialsymbiosis to each partner but the implications of thehydrogen hypothesis for the origin of chloroplasts seemnot to have been considered in detail lsquoWhat was in it forthe proto-chloroplastrsquo may be an even more acute ques-tion than lsquowhat was in it for the proto-mitochondrionrsquoas speaking metabolically and energetically cyanobacteriacan do almost anything
Eukaryotes whether autotrophic or heterotrophic havenow mostly adopted the aerobic part of the energy metab-olism of their original bacterial constituents though inter-esting exceptions are known Respiration andphotosynthesis are antiparallel hydrogen transfer (redox)reactions
respiration
light photosynthesis
CH2O + A2 + H2O 2H2A + CO2
As in the Van Niel equation for photosynthesis A maybe an organic moiety for example giving H2A as succi-nate an inorganic element such as sulphur or evenlsquonothingrsquo so that H2A becomes H2 When A is O(oxygen) it becomes obvious that O2 in photosynthesiscomes from H2O not CO2 and the equation simplifies to
respiration
light photosynthesis
CH2O + O2 H2O + CO2
Phosphate group transfer and electron-transfer-linkedATP synthesis are intermediates in all cases In respir-ation CO2 is produced from stepwise oxidative decar-boxylations of pyruvate dehydrogenase and reactions ofthe tricarboxylic acid cycle In most forms of photosyn-thesis CO2 is assimilated to form initially sugar phos-phates
The mitochondrion exports ATP and CO2 and importsADP inorganic phosphate an electron acceptor andpyruvate Energetically the preferred electron acceptor ismolecular oxygen Oxygen comes from the environmentcells and cell membranes are freely permeable to oxygenThe primary metabolic division of labour between thechloroplast and the plant cytosol is that the chloroplastexports oxygen and reduced sugar phosphates (triosephosphate) and imports CO2 less reduced sugars (3-phosphoglycerate) and inorganic phosphate The light
Phil Trans R Soc Lond B (2003)
comes directly from the external environment and in thisrespect the chloroplast thylakoid is the external boundaryof the plant cell where light both as energy and infor-mation is first received The CO2 is also an environmentalinput though its supply may be regulated locally by CO2-or HCO2
3 -concentrating mechanismsAt the cellular level the mitochondrial inner membrane
is the first place where altered O2 supply exerts its pro-found metabolic effects Even granted specialized photore-ceptors the chloroplast thylakoid membrane is first placewhere changes in light quality and intensity affect a plantrsquosprimary input of energy Temperature changes selectivelyaffect diffusion-limited metabolism involving solubleintermediates In both locations abiotic environmentalchanges therefore have immediate and very far-reachingeffects on electron transfer and redox chemistry Theinternal membranes of mitochondria and chloroplastsstand at the energetic interface of the cell with the physicalenvironment and are likely to be first to know about andrespond to its changes
2 THE PROBLEM OF CYTOPLASMIC GENETICSYSTEMS
Given that the genomes of mitochondria and chloro-plasts are derived from those of once free-living bacteriait is clear that most of the bacterial genes that have sur-vived in any form have now been removed to the cellnucleus If most genes can be moved to the nucleus thenwhy not any gene And if any gene can be moved whynot all Why have mitochondria and chloroplasts retainedfunctional genomes at all What do the genes of mitochon-dria and chloroplasts have in common that has resultedin their retention
Alberts et al (1994 p 400) succinctly describe theproblem posed by the persistence of cytoplasmic geneticsystems as follows
Why do mitochondria and chloroplasts require their ownseparate genetic systems when other organelles thatshare the same cytoplasm such as peroxisomes and lyso-somes do nothellip The reason for such a costly arrange-ment is not clear and the hope that the nucleotidesequences of mitochondrial and chloroplast genomeswould provide the answer has proved unfounded Wecannot think of compelling reasons why the proteinsmade in mitochondria and chloroplasts should be madethere rather than in the cytosol
3 CORR CO-LOCATION OF GENES AND GENEPRODUCTS FOR REDOX REGULATION OF GENE
EXPRESSION
This hypothesis states that mitochondria and chloro-plasts contain genes whose expression is required to beunder the direct regulatory control of the redox state oftheir gene products or of electron carriers with whichtheir gene products interact These genes comprise a coreor primary subset of organellar genes The requirementfor redox control of each gene in the primary subset thenconfers a selective advantage upon location of that genewithin the organelle instead of in the cell nucleus Chloro-plast and mitochondrial genomes also contain genes for
Genomes in mitochondria and chloroplasts J F Allen 23
components of the chloroplast and mitochondrial geneticsystems themselves These genes comprise a secondarysubset of organellar genes genetic system genes There isgenerally no requirement for redox control of expressionof genes in this secondary subset though retention ofredox control of genetic system genes may in some casesallow amplification of redox signals acting upon genes inthe primary subset Retention of genes of the secondarysubset (genetic system genes) is necessary for the oper-ation of redox control of expression of genes in the primarysubset (bioenergetic genes) Without genes in the primarysubset the function of genes in the secondary subset iseventually lost and organelles lose their genomes
This hypothesis of co-location for redox regulation ofgene expression CORR was first outlined in a review(Allen 1992) The hypothesis was then systematically putforward in two articles (Allen 1993ab) developed(Allen amp Raven 1996) and has been independentlyreviewed (Race et al 1999) Some of its predictions havebeen tested experimentally as described later
Figure 3 illustrates the general idea of redox regulatorycontrol operating on mitochondrial genes to give the dis-tribution of sites of protein synthesis seen in figure 1 Fig-ure 4 shows the corresponding predicted pattern ofchloroplast redox regulatory control giving rise to the dis-tribution of sites of protein synthesis seen in figure 2Table 1 shows a functional classification of cyanobacterialgenes retained in 16 completely sequenced chloroplastgenomes This analysis (table 1) reveals a strong selectiveretention of genes for subunits of complexes involved inrespiration and photosynthesis and especially in chloro-plast photosystems I and II The pattern (table 1) seemsto be broadly consistent with CORR (figure 4) and incon-sistent with alternative hypotheses (see sect 4) It is reason-able to view photosynthesis as a process where preciserapid and coordinated redox regulatory control of geneexpression repays the cost of maintaining a chloroplastgenetic system
The CORR hypothesis is based on several propositionsassumptions or principles 10 of which are stated explicitlyand listed below Some of the principles are already generallyaccepted some are shared with competing hypothesesand some make direct experimental predictions Howeverother underlying principles of CORR are themselves generalprinciples or subsidiary hypotheses for which no directproof is available but for which predictions are made andpotentially falsifying observations or results can be imagined
The 10 principles that form the basis of the CORRhypothesis follow and each will be critically discussed inthe light of available evidence
(a) The principle of endosymbiotic originAs now generally agreed bioenergetic organelles have
evolved from free-living bacteriaAlthough once controversial (Mahler amp Raff 1975) this
principle will be taken as an axiom and not consideredfurther here The reader is referred to Gray amp Doolittle(1982) Gray (1992) Martin (1999) Martin et al (2001)to sect 1 and to accompanying articles in this journal issue
(b) The principle of unselective gene transferGene transfer between the symbiont or organelle and the cell
nucleus may occur in either direction and is not selective forparticular genes
Phil Trans R Soc Lond B (2003)
This principle takes the form of a testable assertion thatseems to be consistent with current knowledge Nothingcertain seems to be known about the mechanism of intra-cellular gene relocation (Palmer 1997 Martin amp Herrm-ann 1998 Martin et al 1998 Adams et al 2000 Henze ampMartin 2001 Millen et al 2001 Rujan amp Martin 2001Daley et al 2002) Plausible scenarios can be imagined onthe basis of the classical genetical phenomena of trans-formation transduction and transfection However asthere is no agreed mechanism there is no basis forasserting a mechanistic argument for selectivity Genetransfer between the organelle and nucleus seems to be aspecial case of lateral or horizontal gene transfer(Doolittle 1999) and no mechanistic basis seems to becurrently available for selectivity in what may be trans-ferred It should also be pointed out that lateral genetransfer between the organelle and nucleus does not haveto occur within a single cell nor within a multicellularindividual (although here any vertically transmitted genemust be incorporated into the germ line) for the outcomeof copying genes to novel compartments to be securedThus any plausible mechanism for heterologous recombi-nation may apply equally to gene transfer between com-partments Mitochondrial departures from the lsquouniversalrsquogenetic code and biased codon usage in chloroplasts (seelater) may be of selective value in that a barrier is therebyput in place to expression of genes in the lsquowrongrsquo compart-ment This barrier is unlikely to be insuperable especiallyif the lsquowrongrsquo compartment turns out actually to be a bet-ter location Impeded relocation might confer stabilitywithout being a primary reason for the existence of cyto-plasmic genomes
(c) The principle of unselective protein importThere is no barrier to the successful import of any precursor
protein nor to its processing and assembly into a functionalmature form
As with sect 3b this principle takes the form of a testableassertion that seems to be consistent with current knowl-edge The principle is discussed in more detail in sect 4because its negation (some proteins cannot be imported)has previously been put forward in itself as a reason forthe retention of cytoplasmic genomes Good hypothesesare easily falsified and they therefore forbid certain obser-vations The principle of unselective protein import for-bids the existence of a protein even a synthetic one thatcannot be imported by some means as a precursor
(d) The principle of continuity of redox controlDirect redox control of expression of certain genes was present
in the bacterial progenitors of chloroplasts and mitochondriaand was vital for selectively advantageous cell function beforeduring and after the transition from bacterium to organelle Themechanisms of this control have been conserved
This principle has been tested experimentally in the pastfew years and seems to be corroborated by an emergingpicture of redox signalling in bacteria and organellesExperiments carried out in vitro on the products of proteinsynthesis (Allen et al 1995 Galvis et al 1998) or RNAsynthesis (Pearson et al 1993 Konstantinov et al 1995Wilson et al 1996b) are consistent with a general phenom-enon of redox dependency of gene expression in bioener-getic organelles Redox control of chloroplast transcription
24 J F Allen Genomes in mitochondria and chloroplasts
nucleus cytosol
O2
H2O
redox regulation
N-phasemitochondrial matrix
Figure 3 Gene expression and principal pathways of biosynthesis of subunits of protein complexes involved in respiration andoxidative phosphorylation in animal mitochondria As in figure 1 reddish-brown DNA RNA and protein subunits are locatedand synthesized in the mitochondrial matrix light brown protein subunits have genes (also light brown) in the nucleus andare imported from the cytosol as precursors White genes and ribosomal and protein subunits are nuclearndashcytoplasmic and ofarchaebacterial origin Reddish-brown and light-brown genes and ribosomal and protein subunits are of bacterial origin Themajor variable environmental input is oxygen (blue) The essence of the CORR hypothesis is that it is beyond the ability ofthe nuclearndashcytoplasmic system to respond rapidly and directly to changes in oxygen concentration or partial pressure and soredox regulation of gene expression (red arrows) has been retained from the ancestral endosymbiont This redox regulationrequires co-location of certain genes with their gene products within the mitochondrion
nucleus cytosol
CH2O
CO2
redox regulation
N-phasechloroplast stroma
light light
Figure 4 Gene expression and principal pathways of biosynthesis of subunits of protein complexes involved inphotosynthesis in chloroplasts As in figure 2 green DNA RNA and protein subunits are located and synthesized in thechloroplast light brown protein subunits have genes (also light brown) in the nucleus and are imported from the cytosolas precursors White genes and ribosomal and protein subunits are nuclearndashcytoplasmic and of archaebacterial originGreen and light-brown genes and ribosomal and protein subunits are of bacterial origin The major environmental inputsare light (orange arrows) and carbon dioxide (blue) The essence of the CORR hypothesis is that it is beyond the abilityof the nuclearndashcytoplasmic system to respond rapidly and directly to changes in light and carbon dioxide and so redoxregulation of gene expression (red arrows) has been retained from the ancestral endosymbiont This redox regulationrequires co-location of certain genes with their gene products within the chloroplast
Phil Trans R Soc Lond B (2003)
Genomes in mitochondria and chloroplasts J F Allen 25
has been demonstrated (Pfannschmidt et al 1999abAllen amp Pfannschmidt 2000 Tullberg et al 2000) and wasprompted by this principle
The level of redox control of chloroplast transcriptionestablished unequivocally so far is that of plastoquinonean electron and hydrogen atom carrier located strategicallybetween photosystems I and II of photosynthesis Thedirections in which the effects are exerted are functionallyintelligible reduction of plastoquinone increases photosy-stem I transcription and decreases photosystem IItranscription oxidation of plastoquinone decreases photo-system I transcription and increases photosystem II tran-scription Plastoquinone in normal photosynthesis isreduced by the action of photosystem II and oxidized bythat of photosystem I Therefore redox control of chloro-plast gene expression provides a compensatory response(Pfannschmidt et al 1999a) and adds a previously mis-sing part of an explanation for the ability of plants andalgae to adjust the ratio of the two photosystems inresponse to changes in light quality (Chow et al 1990)By the same argument a functionally equivalent post-translational mechanism of redox control of light-harvesting function by photosystems I and II may havebeen conserved during the transition from prokaryotes toeukaryotes (Allen 1992 1995 Allen amp Nilsson 1997Allen amp Race 2002)
The precedent of chloroplast redox control of transcrip-tion (Pfannschmidt et al 1999ab) taken together with theprinciple in sect 3d predicts plastoquinone redox control ofcyanobacterial reaction centre gene transcription and thisis borne out by experiment (El Bissati amp Kirilovsky2001) This complementary self-adjusting transcriptionalresponse (Fujita 1997 Murakami et al 1997 El Bissati ampKirilovsky 2001) indicates strongly that redox control ofphotosynthetic reaction centre gene transcription waspresent in the proto-chloroplast Thus the mechanism ofregulation is likely to have been conserved throughout theevolutionary transition from bacterial symbiont to cyto-plasmic organelle A conserved mechanism of redox regu-lation of transcription in photosynthetic prokaryotes(Unden amp Bongaerts 1997 Bauer et al 1999) and chloro-plasts (Allen 1993c Pfannschmidt et al 1999b) is pre-dicted if evolutionary continuity of the process has beenmaintained
Thus apart from being consistent with experimentaldata the principle of continuity of redox control has beenof heuristic value in pointing to the underlying bacterialcharacter previously unsuspected in the control of geneexpression especially of transcription in chloroplasts ofhigher plants Other possible precedents for redox signal-ling components of cytoplasmic organelles include the prr(Eraso amp Kaplan 2000 2002 Oh amp Kaplan 2000) andreg (Bauer et al 1999) redox regulatory systems of purplenon-sulphur bacteria The TspO lsquooxygen sensorrsquo of Rho-dobacter sphaeroides is present in mammalian mitochondriaas a protein known there as a benzodiazepine receptorand each protein will complement loss of the other(Yeliseev amp Kaplan 2000) As outlined in sect 3j genes forsuch components are to be expected primarily in nucleargenomes This independent evolutionary hypothesis (sect 3d)would be strengthened still further by a demonstration ofone or more decisive redox regulatory links shared by pho-tosynthetic prokaryotes and all eukaryotes with mitochon-
Phil Trans R Soc Lond B (2003)
dria and where present with chloroplasts (Bauer et al2003)
(e) The principle of the selective value of redoxcontrol
For each gene under redox control (sect 3d) it is selectivelyadvantageous for that gene to be retained and expressed onlywithin the organelle
The twin selective advantages of redox control are likelyto be energetic efficiency and the suppression of the harm-ful side effects of electron transport operating on thewrong substrates No direct experimental support for thisprinciple is available and equally there is no evidenceagainst it Allotopic mutants in which genes have beenexperimentally relocated from organelles to the nucleushave been produced in yeast higher plants (Kanevski ampMaliga 1994) and human cells (Manfredi et al 2002) andare predicted by CORR to be poor adapters to environ-mental change Some authors believe there is potentialtherapeutic value in allotopic transformation movingmitochondrial genomes to the nucleus (Kanevski ampMaliga 1994 de Grey 2000) and predict the possibilityof r2 human cell lines in the foreseeable future (de Grey2000) This principle also has implications for ageing andthe evolution of separate sexes (Allen 1996) and possibleconsequences for development (Blackstone 1999 20002001 Harvey et al 2002) and somatic cloning in animals(Allen amp Allen 1999)
(f ) The principle of the selective value of nuclearlocation for genes not under redox control
For each bacterial gene that survives and is not under redoxcontrol it is selectively advantageous for that gene to be locatedin the nucleus and expressed only in the nucleus and cytosolIf the mature gene product functions in chloroplasts or mito-chondria the gene is first expressed in the form of a precursorfor import
There are several possible reasons for the selectiveadvantage of nuclear location of genes for organellar pro-teins One is the decreased probability of mutation arisingfrom free radical by-products of lsquoincorrectrsquo electron trans-fers (Raven et al 1994ab Allen amp Raven 1996) Therewould seem to be obvious economies in resource use andadvantages in coordination of gene expression if genes arekept in one compartment One might say that the bipartiteor tripartite genetic system of eukaryotic cells is untidyand inefficient
Another suggestion is that organellar genes do notundergo recombination and are present in relatively smallcopy numbers so that disadvantageous mutations willspread relatively quickly through a clonal population oforganelles Incorporation of functional gene copiesinstead into the nuclear genome may then be an irrevers-ible step analogous to the click of a ratchet (Saccone et al2000) There are examples of genes shared between allthree compartments (Stern amp Lonsdale 1982 Lonsdale etal 1983)
The cost of retaining an organellar genetic system is notincreased appreciably when additional genes are retainedin organelles if just one gene is retained then the cost ofa cytoplasmic genetic system is incurred and there may berelatively little extra cost for each additional gene retained(Allen amp Raven 1996) For a functional explanation of the
26 J F Allen Genomes in mitochondria and chloroplasts
existence of chloroplast and mitochondrial genomes thefocus of attention should therefore be on the relativelyconstant primary subset of bioenergetic genes (sect 3e)
(g) The principle of contemporary operation ofselection on gene location
For any species the distribution of genes between organelle(by the principle in sect 3e) and nucleus (by the principle in sect 3f)is the result of selective forces that continue to operate
This principle is essentially a statement contradictingboth the lsquofrozen accidentrsquo hypothesis and the hypothesisthat gene relocation is progressive predictable and stillincomplete (sect 4) In support of this principle it is becom-ing increasingly clear that mitochondria that lose theirfunction in aerobic respiration also lose their genomes asseen in the relict mitochondria of microsporidia(Embley amp Martin 1998 Van der Giezen et al 2002) andthe lsquomitosomersquo of Entamoeba histolytica (Tovar et al1999) In chloroplasts loss of photosynthesis results inloss of photosynthetic genes The examples of Epifagus (aparasitic higher plant) (Ems et al 1995 Wolfe et al 1992)and the residual apicomplexan plastids of Plasmodium(Wilson et al 1996a) and Toxoplasma where some plas-tidic genetic system is retained mean that some of theirgene products must be subject to redox regulation of geneexpression if this principle is correct Experimental test ofthis principle is possible using allotopic mutants as forthe predictions of the principle in sect 3f The evidence thatthe apicoplast (Wilson et al 1996a) is a plastid is largelygenetic the residual genome resembles that of photosyn-thetic plastids (chloroplasts) in gene organization ratherthan in content A plastid that has completely lost its gen-ome by virtue of having lost photosynthesis and all otherprocesses requiring redox regulation might be difficult todistinguish from an organelle derived in a similar way froma mitochondrion The persistence of targeted importedchaperones Hsp70 (Tovar et al 1999 Williams et al2002) and cp60 (Roger et al 1998) in residual organellesmight not distinguish a plastid ancestry from a mitochon-drial one
(h) The principle of primary involvement inenergy transduction
Those genes for which redox control is always vital to cellfunction have gene products involved in or closely connectedwith primary electron transfer These genes are always con-tained within the organelle
This principle would be violated and the CORRhypothesis refuted if any photosynthetic reaction centrecore subunit or respiratory chain subunit currentlyorganelle-encoded is found that functions in primary vec-torial electron transport and is nevertheless encoded in thenucleus and synthesized cytosolically as a precursor forimport all without any selective cost to the individualFrom the chloroplast transcriptional experiments ofPfannschmidt et al (1999ab) the chloroplast geneswhose transcription is regulated by the plastoquinoneredox state include psaAB (for photosystem I reactioncentre core polypeptides) psbA (for the photosystem IIpolypeptide D1) and rbcL (for the large subunit of RUB-ISCO interestingly following the same pattern of redoxcontrolmdashupregulated by oxidized plastoquinonemdashaspsbA) Transcription of the chloroplast genes petA
Phil Trans R Soc Lond B (2003)
(cytochrome f ) trnG (glycine tRNA) and rpoB (RNApolymerase) did not respond to plastoquinone redox statepetA for cytochrome f is predicted by this principle to beregulated at a different level of redox control or if at plas-toquinone at a different stage in gene expression Thisprediction might also apply to trnG and rpoB though thesequalify as members of the secondary subset that of geneticsystem genes to which the principle in sect 3i applies
(i) The principle of secondary involvement inenergy transduction
Genes whose products contribute to the organelle genetic sys-tem itself or whose products are associated with secondaryevents in energy transduction may be contained in theorganelle in one group of organisms but not in anotherdepending on the physiology and biochemistry of photosynthesisand respiration in the species concerned
Even phylogenetically moderately related species(within kingdoms) show differences in the location ofsome genes for example the ATP synthase subunits ofNeurospora and Saccharomyces (Attardi amp Schatz 1988)The CORR hypothesis also does not explicitly account forthe wide disparities in the distribution of genetic systemgenes for example the almost complete import of tRNAsin Chlamydomonas (GoldschmidtClermont 1998) Trans-port of nucleic acids across membranes is uncommon andwhy some RNAs should be organelle-encoded in somespecies but nuclearly encoded in others is not currentlyunderstood The only suggestion that can be made alongthe lines of CORR is that in some genetic systems genesmay need to be under the control of a regulatory systemthat responds to a signal arising within the organelle itselfThis signal could be the primary signal of altered redoxstate or some secondary signal Among the latter might befound signals that themselves report on the current stateof gene expression another piece of information that thenucleus might not have at its immediate disposal Suchsecondary signals could result at least in part fromCORR though other developmental and environmentalinputs may be imagined and would not detract fromCORR unless it could be shown that they provided a suf-ficient reason for the retention of the genetic system asa whole
(j) The principle of the nuclear encoding of redoxsignalling components
Components of the redox-signalling pathways upon whichCORR depends are themselves not involved in primary electrontransfer and so their genes have been relocated to the nucleusin accordance with sect 3f
This principle states that the redox signalling pathwaysthat are required for CORR should not themselves beexpected to utilize components whose biosynthesis mustbe under direct redox control Such components shouldtherefore fall into the major category of organellar pro-teins and be imported as precursors from the cytosol inline with the principle in sect 3f However the principleabove might be need to be modified in cases where ampli-fication of the redox signal is useful to the cell In thebacterial ntr system which was a precedent for CORR(Harrison et al 1990 Allen 1992) the histidine kinasentrB and aspartate response regulator ntrC form part ofthe gln operon whose transcription they control Odontella
Genomes in mitochondria and chloroplasts J F Allen 27
and Porphyra have plastids that encode more proteins thanthose of most chlorophytes (Stoebe et al 1998 Turmel etal 1999) and these proteins include two-component sig-nal transduction proteins prr and ycf27 (Stoebe et al1998) Extreme or extremely rapid environmental fluctu-ations might be catered for by amplification of signals thatare transmitted by signalling pathways whose componentsare under their own control It is not known whether ycf27and prr are in operons whose promoters their gene pro-ducts control It is also uncertain whether ycf27 is a redoxsignalling component and its distribution at least in itsplastidic location seems to be correlated with the presenceof a phycobilin-based light-harvesting system as found inred algae and cyanobacteria (Ashby amp Mullineaux1999ab Ashby et al 2002) The best guess of the originalstatement of the CORR hypothesis (Allen 1992 1993ac)was that bacterial two-component systems (Iuchi amp Lin1992 Allen 1993c Grebe amp Stock 1999) provide a plaus-ible means of exerting redox regulatory control over mito-chondrial and chloroplast gene expression andsubsequent research confirms the presence of two-compo-nent genes in genomes of Arabidopsis and yeast SeveralArabidopsis two-component genes are predicted to be tar-geted to mitochondria and chloroplasts (Forsberg et al2001) These must be considered as prime candidates forcomponents involved in redox regulation of chloroplasttranscription seen in the experiments of Pfannschmidt etal (1999a 2001a) The presumably homologous regulat-ory process serving to control photosystem stoichiometryin cyanobacteria the prr system (Li amp Sherman 2000) isnow characterized as a two-component system involvinga redox sensor and redox response regulator as originallydefined (Allen 1992 1993ac)
4 CONTRASTING HYPOTHESES CONCERNINGTHE RETENTION OF CYTOPLASMIC GENOMES
CORR is a recent addition to explanations that havebeen put forward to account for the persistence of gen-omes in organelles These models and their underlyingmechanisms are now discussed and compared
(a) There is no reason that is just how it isOne reaction to the problem of cytoplasmic genetic sys-
tems (sect 2) is that there is no problem and no particularreason for their retention However there is a very widephylogenetic distribution of a common core or primarysubset of organellar-encoded genes and one which haswithstood considerable genomic reorganization over evol-utionary time This constancy is seen for example in thesame genes being present in the chloroplasts of landplants with multiple copies of a single circular chloroplastchromosome and in dinoflagellate chloroplasts with acollection of different DNA minicircles each with essen-tially one gene (Zhang et al 1999 2001 Barbrook et al2001) Another suggested explanation of the retention ofmitochondrial and chloroplast genomes is simply torestate the endosymbiont hypothesis and to point out thatchloroplast and mitochondrial genes lsquocame inrsquo with thebacterial endosymbiont This suggestion however doesnot address the question of why some genes were retainedwhereas others were not It is also now clear that mostgenes that came in and still survive have nuclear and cyto-
Phil Trans R Soc Lond B (2003)
solic gene products whereas many chloroplast and mito-chondria proteins had a eukaryotic origin (Salzberg et al2001 Martin amp Russell 2003) The odds against a con-served pattern in gene location (eg table 1) arising bychance must be astronomical Thus there is a reason forthe retention of organellar genomes and a reason for theretention of the same primary subset of genes Becauseevolution proceeds by natural selection of chance vari-ation lsquoreasonrsquo in this context requires identification of thefunction of the location of a gene that is the selectivevalue of the phenotype produced by that location Evol-ution sometimes retains apparently functionless relics ofpreviously useful features but not for long especially ifthere is a cost
(b) The lock-in hypothesisThe lsquolock-inrsquo hypothesis of Bogorad (1975) was pro-
posed independently of the endosymbiont hypothesis anddoes not depend upon it According to the lock-in hypoth-esis core components of multi-subunit complexes must besynthesized de novo in the correct cellular compartmentotherwise photosystems or respiratory chain complexesfor example might assemble in the plasma membranevacuolar membrane or endoplasmic reticulum The lock-in hypothesis made no clear prediction about the conti-nuity of cytoplasmic genetic systems even though cyto-plasmic uniparental non-Mendelian inheritance for bothmitochondria (Roodyn amp Wilkie 1968) and plastids(Kirk amp Tilney-Bassett 1978) made it likely that organellarDNA the chondriome and the plastome replicated inde-pendently of nuclear DNA (Whitehouse 1969)
Mechanisms of protein import and targeting (Schatz1998 Koehler et al 1999 Leuenberger et al 1999 Gab-riel et al 2001 Jarvis amp Soll 2001 Pfanner amp Geissler2001 Duby amp Boutry 2002 Pfanner amp Wiedemann 2002)now effectively answer the problem of specifying the cor-rect locations for membrane protein complexes for whichthe lock-in hypothesis was proposed as a solution How-ever regulation of synthesis of an entire functional multi-subunit complex by control of synthesis of individual sub-units is still an important principle This seems to be afeature of the lsquoCESrsquo model (control by epistasy ofsynthesis) of Wollman (Wollman et al 1999 Eberhard etal 2002) which envisages nuclear control as all-embracingwith organellar genetic systems doing as it were whatthey are told In the hypothesis of CORR propoundedhere CES is equally important but the essence of CORRis that the organellar genetic system alone knows what arethe immediate and most important signals of environmen-tal change The organelle may keep the nucleus informedbut this is a lower priority than initiating an appropriateresponse CES and CORR agree that assembly of multi-subunit complexes is a complex business and requirescoordinated control in which no step is rate-limiting butmany may be obligatory
(c) Going going gone the evolutionary process oftransfer of genes from organelles to the
nucleus is still underway but incompleteThis hypothesis envisages that for most eukaryotes we
are now witnessing the last stages in a slow evolutionarytransfer of genes to the nucleus and this transfer may beexpected to proceed to completion in what is in evol-
28 J F Allen Genomes in mitochondria and chloroplasts
utionary terms the near future (Palmer 1997 Gray 19992000 Gray et al 1999 Adams et al 2000 Daley et al2002) A clear and compelling statement of this widelyheld position is that of Herrmann amp Westhoff (2001 p10)
hellipthe transfer of geneshelliptells that the partite plant gen-ome is not in a phylogenetic equilibrium All availabledata suggest that the ultimate aim of genome restructur-ing in the plant cell as in the eukaryotic cell in generalis the elimination of genome compartmentation whileretaining physiological compartmentation
One serious problem with this hypothesis is the dis-tinctly non-random sample of genes that remain to betransferred A relatively constant subset of genes is foundin bioenergetically functional mitochondria from fungi tomammals and angiosperms as in photosynthetic plastids(chloroplasts) from unicellular algae to angiosperms Thishypothesis therefore requires an additional ad hocassumption that some gene transfers are inherently moreprobable than others (this too is widely supposed seelater) and thus that gene transfer events proceed in thesame sequence in quite independent lineages Thereseems to be no evidence for this and the order of geneloss from mitochondria and chloroplasts has been used toinfer phylogeny (Martin et al 1998 Adams et al 2000Millen et al 2001) lsquoGoing going gonersquo (Palmer 1997)is difficult to refute categorically but it does not seem tobe consistent with recent evidence For example chromo-some two of Arabidopsis thaliana (Lin et al 1999) has asingle insertion encompassing with about 99 sequencesimilarity the complete Arabidopsis mitochondrial genome(Unseld et al 1997 Marienfeld et al 1999 Stupar et al2001) Unless this single transfer is a freak occurrencethis finding strongly indicates that gene transfer occurswith relative ease (Adams et al 2000 Henze amp Martin2001 Rujan amp Martin 2001 Salzberg et al 2001) Inaddition there is experimental evidence that randomlygenerated polypeptides are mitochondrially imported athigh frequency (Lemire et al 1989) The hypothesis thatwe now see the same few stragglers in diverse organellargenomes is not impossible but should be regarded as alast resort perhaps applicable only in extreme cases suchas the greatly reduced apicomplexan plastomes and chon-driomes Even there the hypothesis carries with it an obli-gation to explain why organisms with few organellar geneshave lsquoevolvedrsquo faster than those with many
(d ) The frozen accidentWhat may be described as lsquothe frozen accident hypoth-
esisrsquo states that the evolutionary process of gene transferfrom organelle to nucleus was underway when somethinghappened that stopped it One example of the frozen acci-dent is a scenario (Von Heijne 1986) in which import intomitochondria of the precursor proteins arising from newlytranslocated genes proceeded for long enough after theprimordial endosymbiosis for most of the symbiont-derived genes to be moved to the cell nucleus The lsquoacci-dentrsquo that halted the process was the evolutionary originof exocytosis and protein secretion after which it becameimpossible for precursors properly destined for mitochon-drial import to be distinguished from those destined forexport from the cell As with the lsquolock-inrsquo hypothesis the
Phil Trans R Soc Lond B (2003)
lsquofrozen accidentrsquo seems incompatible with the remarkableprecision and specificity of protein targeting (Schatz 1998Koehler et al 1999 Leuenberger et al 1999 Gabriel etal 2001 Jarvis amp Soll 2001 Pfanner amp Geissler 2001Duby amp Boutry 2002 Pfanner amp Wiedemann 2002) Pro-tein sorting mechanisms do not seem to forbid controlledimport into any cellular compartment Another lsquoaccidentrsquothat may have impeded subsequent gene transfer in thecase of mitochondria is their departure from what is other-wise a universal genetic code Genetic code differences arediscussed later in the context of whether some genes can-not be moved
(e) Some proteins (with cofactors) cannot beimported
We may describe as the lsquounimportability hypothesisrsquo theproposal that some proteins are inherently unimportableand must therefore be synthesized de novo in the correctcompartment within the cell The unimportability hypoth-esis is most often proposed as its special case the hydro-phobicity hypothesis with hydrophobicity being thereason for unimportability
(i) HydrophobicityA commonly entertained notion is that proteins that are
encoded and synthesized within organelles comprise a pri-mary subset characterized by shared hydrophobicity allare intrinsic membrane proteins (Von Heijne 1986Popot amp de Vitry 1990 Claros et al 1995) As with theCORR hypothesis favoured here the primary subset thenrequires a secondary subset of genetic system componentsthe latter not themselves being hydrophobic but beingrequired for synthesis of the others Certainly the mito-chondrial inner membrane contains some intrinsic proteincomplexes whose subunits broadly satisfy the criterion ofhydrophobicity However the core subunits that are mito-chondrially encoded are also explained by the CORRhypothesis This would make hydrophobicity a require-ment for their function in vectorial electron and protontransfer and not a reason in itself for retention of theirgenes in the cytoplasm
The hydrophobicity hypothesis has some very clearcounter-examples in chloroplasts and cannot thereforestand if one wishes for a single principle that embracesboth types of bioenergetic organelle One clear counter-example is the large (L) subunit of the enzyme RUBISCOan abundant but entirely membrane-extrinsic protein ofca 55 kDa molecular mass The RUBISCO large subunitis generally chloroplast-encoded with the exception of thenuclearly encoded bacterial-type L subunit of L2 RUBI-SCO in peridinin-containing dinoflagellates (Morse et al1995) In most photosynthetic eukaryotes RUBISCO hasa chloroplast encoded large subunit and a nuclearly enco-ded small (S) subunit (Ellis 1984 Gatenby amp Ellis 1990)giving an L8S8 holoenzyme and this RUBISCO is widelytaken as a paradigm for the need for coordination ofnuclear and chloroplast gene expression (Rodermel 2001)
By complete contrast to the RUBISCO large subunitwhich is a chloroplast-encoded and water-soluble proteina second major violation of the hydrophobicity hypothesisis the existence of the universally nuclearly encoded butalways hydrophobic subunits of the chloroplast LHC IIand LHC I (Chitnis amp Thornber 1988) These apopro-
Genomes in mitochondria and chloroplasts J F Allen 29
teins of ca 27ndash30 kDa are quite water-insoluble and areusually fully extracted only in chloroformndashmethanol or inthe presence of detergents The Lhc gene family is diverse(Green amp Kuhlbrandt 1995) but most gene productsor Lhcp (light-harvesting chlorophyll protein) polypeptideshave three membrane-spanning hydrophobic a-helices(Kuhlbrandt 1994) It is interesting that three-helix Lhcproteins seem to be absent from extant cyanobacteria andchloroxybacteria (Green amp Kuhlbrandt 1995)
(ii) The five-helix rulePerhaps with the three-helix model of LHC II in mind
a variation of the hydrophobicity hypothesis is the empiri-cal lsquofive-helix rulersquo (Popot amp de Vitry 1990 Claros et al1995 Wollman et al 1999) which states that no intrinsicmembrane protein with five or more membrane-spanninghelices can be encoded in the nucleus Five is a good guessfor a threshold number of helices demanding organellar-encoding in the context of photosynthetic reaction centres(Heathcote et al 2002) where the earliest known struc-ture that of type II centres (Deisenhofer et al 1985) asin photosystem II (Zouni et al 2001) shows exactly fivehelices with type I centres containing 11 (Jordan et al2001) There are however many nuclearly encoded sub-units in both mitochondria and chloroplasts that havefewer than five membrane helices so the five-helix rulecan only describe a sufficient condition and not a neces-sary condition for nuclear encoding
It should also be noted that ADPndashATP carriers (AACs)are encoded in the nucleus AACs of the mitochondrialinner membrane have six transmembrane helices (van derGiezen et al 2002) while the those of the chloroplast innerenvelope (Winkler amp Neuhaus 1999) have 12 (Tjaden etal 1998)
With the counter-examples of nuclearly encoded AACswith six or 12 membrane-spanning helices many intrinsicmembrane proteins with fewer than five helices somenuclearly encoded and some not and with the RUBISCOlarge subunit as a major product of chloroplast geneexpression with no membrane domain at all the five-helixrule begins to seem rather like a case of special pleadingEven if there are indeed special mitigating circumstancesfor AACs (this argument is outlined later) the five-helixrule as with the hydrophobicity hypothesis of which it isa special case will disappear if it can be shown that pos-session of five or more membrane helices is required forsome other function such as that of CORR that deter-mines the organellar location of the gene concernedHydrophobicity is in some cases a feature of proteins thatare encoded in organelles but it is not in itself the issue
For chloroplasts the hydrophobicity hypothesis predictsretention of cyanobacterial genes in functional categoriesassociated with the chloroplast envelope and with trans-port just as much as with photosynthesis Table 1 showsthat this is not the case
A mechanism for the unimportability of hydrophobicproteins is rarely stated explicitly and it is rather left tothe imagination whether hydrophobic proteins might asit were get stuck halfway through the chloroplast envelopeor mitochondrial outer membrane whether transport can-not get started on a precursor protein that has alreadyspontaneously inserted into the membrane or whetherhydrophobic proteins will be unable to make the journey
Phil Trans R Soc Lond B (2003)
through the cytosol without precipitating there In thecytosol co-translational transport or membrane insertionwould in principle address the latter point but it seemsto be generally true that chloroplast and mitochondrialproteins are post-translationally inserted into the organel-lar outer membrane So the obvious ancillary questionabout unimportability is lsquowhy then cannot hydrophobicproteins be co-translationally imported into organellesrsquoThere is evidence in yeast for co-translational transportinto mitochondria of proteins thought to be derived fromthe original endosymbiont (Marc et al 2002) If there isany truth in hydrophobicity as a barrier to protein importthe CORR principle of unselective import (sect 3c) shouldbe easily refuted for example by a demonstration that noknown targeting presequence is capable of importing poly-phenylalanine encoded by polyU for in vitro import stud-ies
An additional proposal intended to account for theunimportability of some hydrophobic proteins and forimportability of others is suggested by A de Grey(personal communication) who argues that bacteriallyderived protein precursors imported into mitochondriacross the mitochondrial inner membrane completelybefore insertion into that membrane from the mitochon-drial matrix (the N-phase) This is held to be an obstacleto successful import of membrane-intrinsic proteins thatmight en route discharge the proton motive force By con-trast eukaryotically derived proteins can be inserted intothe mitochondrial inner membrane from the inter-mem-brane space (the P-phase) and then there is for somereason no similar uncoupling effect Thus adenylatetransporters or AACs can be nuclearly encoded whereascytochrome oxidase subunits for example cannot It isnot obvious why bacterially derived subunits such asthose of cytochrome oxidase did not acquire eukaryoticinsertion mechanisms Perhaps this acquisition is forbid-den by the need to conserve pre-existing routes ofassembly into multi-subunit complexes or perhaps theneed for insertion from the inside reflects a need to incor-porate a cofactor for example haem or ironndashsulphur clus-ters along the way
(iii) Unimportability of three dimensional structures andcofactors
Accumulating evidence on protein transport mech-anisms seems to make a specific obstacle to successfultransport of certain sorts of protein less and less plausibleespecially given the observation that precursors with N-terminal targeting sequences not mature functional poly-peptides themselves traverse membranes during biosynth-esis
Protein folding inside the living cell is invariably assistedby molecular chaperones (Gatenby amp Ellis 1990 Azem etal 1997 Komiya et al 1997 1990 Jackson-Constan et al2001) proteins that impede the formation of lsquoimproperrsquostructural interactions that will otherwise produce aggre-gation or other non-functional interactions Proteinimport mechanisms appear mostly to rely on specific mol-ecular chaperones for guided unfolding before membraneinsertion and refolding after translocation of the polypep-tide into its destined compartment (Gatenby amp Ellis 1990Azem et al 1997 Komiya et al 1997 Jackson-Constanet al 2001) Because polypeptides are mostly lsquothreadedrsquo
30 J F Allen Genomes in mitochondria and chloroplasts
through membranes no special constraint on transportshould be expected to arise from 3D structure or surfaceproperties of a protein
An exception to this unfolding rule may be providedby holoproteins containing cofactors that are transportedalong with the apoprotein presumably with a largely intact3D structure The lsquoTATrsquo (twin-arginine translocase) sys-tem (Bogsch et al 1998 Dalbey amp Robinson 1999 Berkset al 2000ab Robinson 2000 Robinson amp Bolhuis 2001Sargent et al 2002) works broadly along these linesthough its mechanism is unresolved TAT is relevant herebecause it adds transport of essentially complete holopro-teins to the cellrsquos repertoire of transport mechanisms andeven proteins that may not be allowed to unfold perhapsbecause of the toxicity of the free cofactor are not therebymade candidates for an unimportability rule One variantof the unimportability hypothesis is that transport canwork only by unfolding and dissociated cofactors may betoxic to the cell The bacterially derived TAT system(Bogsch et al 1998 Dalbey amp Robinson 1999 Berks etal 2000ab Sargent et al 2002) overcomes this argumentas would examples of cofactor-free polypeptides such as(C)Fo subunit c which are nevertheless generally encodedin organelles
The presence of a vesicular transport system in chloro-plasts (Westphal et al 2001) counts against the unim-portability hypothesis Vesicular transport between thechloroplast envelope and the thylakoid membrane shouldin principle allow any protein to be ferried across thechloroplast stroma Vesicles might allow transport inparticular of membrane proteins that would discharge theproton motive force if allowed to connect transiently theN and P aqueous phases on each side of a coupling mem-brane
Another observation that seems to count against unim-portability is the increased number of membrane barriersin plastids derived from secondary tertiary or higher-orderendosymbioses These plastids seem to have retained ifanything fewer genes rather than more as unimportabilitypredicts The nucleomorph the residual nucleus of theprimary host retains many genes for molecular chap-erones (Archibald et al 2001 Cavalier-Smith 2002) inagreement with the conclusion that protein import pro-ceeds by conserved and specialized mechanisms If eachnew import added to gain a higher-order endosymbiosisrequires additional targeting presequences then perhapsthere is an advantage in components of an import machin-ery not being imported themselves
It seems that unimportability is less easy an assumptionto make today than in previous years and the onus maynow be on its proponents to cite specific examples of pro-teins that can never be imported into chloroplasts or mito-chondria
(f ) Some genes cannot be moved(i) Differences in the mitochondrial genetic code
The genetic code is the specific relation between ribon-ucleotide triplets of codons and tRNAs that recognizethem and which thus determine which individual aminoacids if any will be incorporated into the growing poly-peptide chain Mitochondria show small departures fromthe otherwise universal genetic code (Attardi amp Schatz1988 Alberts et al 1994) UGA is a lsquostoprsquo codon in the
Phil Trans R Soc Lond B (2003)
universal code and in plant mitochondria but is the codonfor tryptophan in mitochondria of animals and fungiCUA means leucine everywhere except fungal mitochon-dria where it means threonine and AGA and AGU codefor arginine in the lsquouniversalrsquo code but serine in insectmitochondria and lsquostoprsquo in mammalian ones The signifi-cance of this heterogeneity in coding is most unlikely torest in the early origin of mitochondria as the lsquouniversalrsquocode is employed by bacteria and protists Furthermorethe diversity of departures from the lsquouniversalrsquo codeamong mitochondria of different eukaryotic lineages indi-cates they had independent origins A general rule formitochondrial genetic codes is that reliance on the thirdlsquowobblersquo position is decreased which allows all 20 proteinamino acids to be specified by fewer tRNAs (22 in animalmitochondria instead of 31 in the cytosol) This in turnmeans that mtDNA can specify more proteins for a givenlength of DNA or conversely that the mitochondrialgenome can be condensed or lsquostreamlinedrsquo Although thisarrangement does not explain the retention of mitochon-drial genomes and may be a secondary event in mitochon-drial evolution it almost certainly places an extra barrierin place of gene relocation As discussed previously how-ever there seems to be no single barrier that can explainthe retention of a relatively constant subset of mitochon-drial genes
There has clearly been a contraction of mitochondrialDNA especially in animals Animal mitochondrial DNAhas few if any non-coding sequences and animal mito-chondrial genetic codes require fewer tRNAs than the sin-gle code operating in the cytosol (Gray 1999 Gray et al1999 Selosse et al 2001) These may be devices todecrease the total frequency of mutations per mitochon-drial genome assuming a constant frequency per unitlength of mitochondrial DNA It has been proposed thata genersquos removal to the nucleus decreases its susceptibilityto free radical mutagenesis (Raven et al 1994ab Allen ampRaven 1996) If this is the case then the non-standardcode may be a further indication of an overriding need tomaintain certain genes within the mitochondrion What-ever the reason for the retention of genes in organelles ithad better be a good one bioenergetic organelles areclearly the wrong place to keep genetic information andgo to extraordinary lengths to offset the consequences ofit being there
(ii) Molecular refugeesA phenomenon related to differences in the genetic code
is seen in chloroplasts This is a bias in codon usage Thelsquomolecular refugeesrsquo hypothesis (Howe et al 2000) sug-gests that nuclear copies of what were once chloroplastgenes escape constraints on codon usage and thus thatwhat distinguishes chloroplast-encoded proteins is thattheir amino-acid composition is unusual enough for thebias in codon usage not to apply It is difficult to see whatphysico-chemical properties might be uniquely shared bythe amino acids of chloroplast-encoded proteins and inaddition there seem to be no lsquoforbiddenrsquo amino-acid resi-due in chloroplasts As with the departure from the univer-sal genetic code in mitochondria is seems likely that abias in codon usage reflects some secondary property ofchloroplast genomes and is not the primary reason fortheir retention In any case any restriction on gene move-
Genomes in mitochondria and chloroplasts J F Allen 31
ment will act in the wrong direction to explain why chloro-plasts have retained genomes at all provided there is noresulting obstacle to a chloroplast protein being success-fully encoded as a precursor in the nucleus
(iii) Genes are copied and replicated not relocated in onestep
If complete relocation is envisaged as proceeding by thesimultaneous appearance of a gene in one compartmentand its deletion from another then it is possible to imaginethat whatever the vector is there may be selectivity ingene transfer (Henze amp Martin 2001) For example ifviral incorporation replication and recombination in bothorganelle and nucleus were the mechanism of relocationthen successful gene transfer might depend on the abilityof the virus to complete the process in one step This inturn might depend on some property of the gene or ofthe protein it encodes if expression occurs at any stage inthe process of relocation An assumption of the CORRhypothesis favoured here is that genes are not movedphysically but merely copied after which either the copyin the original location or the copy in the new locationmay be lost as a result of selection A useful analogy maybe the action of copying files between computer storagevolumes In this analogy the copy is identical to the orig-inal and either may subsequently be deleted without lossof information
Because any gene that is not usefully expressed willundergo no selection to maintain a functional allele theprevailing location will be the one in which the functionalcopy of the gene resides The problem of which gene sur-vives is then subsumed into the problem of which is theselectively advantageous location for it It seems reason-able to assume that there wasmdashand ismdashan interim periodin which homologous genes in different compartments areessentially in competition to see which functions best Asoutlined above there is no obvious obstacle for copyingDNA sequences within the cell The physico-chemicalproperties of DNA are fairly uniform and not directlydependent on nucleotide sequence though organelles likeintra-cellular parasites seem to prefer A and T to G andC The answer to the question lsquowhich gene location func-tions bestrsquo may have nothing to do with the gene itselfnor directly with the structure and function of its geneproduct Instead the answer may have everything to dowith the required regulatory interaction between the geneproduct and expression of the gene that produces it
(g) CORRThis hypothesis has been propounded earlier (sect 3) and
will be summarized as follows Vectorial electron and pro-ton transfer exerts regulatory control over expression ofgenes encoding proteins directly involved in or affectingredox poise This regulatory coupling is indispensable foradaptation to environmental changes in chemical activitiesof oxygen light and CO2 and requires co-location of thesegenes with their gene products Organelles lsquomake theirown decisionsrsquo on the basis of environmental changesaffecting redox state The hypothesis is abbreviatedCORR for co-location for redox regulation or more com-pletely lsquoco-location (of gene and gene product) for(evolutionary) continuity of redox regulation of geneexpressionrsquo
Phil Trans R Soc Lond B (2003)
5 SUMMARY OF THE CURRENT STATUS OF THEHYPOTHESIS OF CORR
Q Why do mitochondria and chloroplasts have theirown genetic systems
A CORR co-location of genes and gene products per-mits direct and autonomous redox regulation of geneexpression
(a) Objections to CORR(i) There is no regulation of gene expression in chloroplasts
and mitochondriaThis is still commonly stated but incorrect See the prin-
ciple in sect 3d
(ii) There is no regulation of gene expression in chloroplastsand mitochondria at the level of transcription
For chloroplasts there is now clear evidence for a func-tionally intelligible redox control of transcription(Pfannschmidt et al 1999a) This invalidates the previouswidely held assumption that all control of chloroplast geneexpression is post-transcriptional (Gruissem amp Tonkyn1993 GoldschmidtClermont 1998) The evidence formitochondria is less clear but redox control of transcrip-tion certainly cannot be ruled out and there are indi-cations that it may occur (Pearson et al 1993Konstantinov et al 1995 Wilson et al 1996b) It mustalso be restated that transcriptional control is not neces-sary for CORR but serves as a compelling example of aconserved typically bacterial mode of response that oper-ates still in eukaryotic cytoplasmic organelles thus illus-trating the principle of continuity of redox control (sect 3d)Post-transcriptional redox regulation (Danon amp Mayfield1994 Kim amp Mayfield 1997 Brown et al 2001 Treb-itsh amp Danon 2001) is consistent with CORR and thereis nothing to forbid it being a primary mechanism thatsatisfies most or indeed all of the formal requirements ofthe hypothesis in particular cases
(iii) There are no redox signalling components encoded inchloroplast or mitochondrial DNA
This is what CORR predicts See the principle in sect 3j
(iv) There is redox control of nuclear gene expressionThis is correct and many examples are now known
especially for chloroplasts (Escoubas et al 1995 Kovacs etal 2000 Pfannschmidt et al 2001ab Surpin et al 2002)Redox signals from photosynthetic or respiratory electrontransport chains are not at all prohibited by CORR Theessence of CORR is that there is a subset of organellarproteins for which there is an overriding requirement fora redox regulatory control that is initiated within theorganelle and whose effects are exerted by the simplestand most direct route This redox signalling is envisagedto take place without a requirement for prior consultationand coordination with what may be regarded as a nucleo-cytoplasmic bureaucracy
(v) Nuclear genes can be expressed as rapidly as chloroplastand mitochondrial genes
Zerges (2002) for example cites the equal times takenfor complete expression of nuclear and chloroplast genesin Chlamydomonas as evidence against the CORR hypoth-
32 J F Allen Genomes in mitochondria and chloroplasts
esis However the question is not one of the time takento express a gene but the time taken to alter the rate ofits expression in response to a (redox) signal originatingwithin the organelle An equally important additional fac-tor may be the autonomy of the signalling pathway thatis its independence from other inputs as described in theprevious paragraph One might say that mitochondria andchloroplasts require a free hand
(vi) There is no evidence to support this hypothesisThe hypothesis is rich in predictions (sectsect 3 and 6) and
those that have been tested stringently are in agreementwith experimental results None of the predictions of thehypothesis has been found to be incorrect and none of theunderlying principles (sect 3) has been disproved Furtherpredictions will be the subject of future experiments
(b) Evidence for CORR(i) Chloroplasts have selectively retained cyanobacterial genes
for primary events in photosynthesisSee table 1
(ii) There is regulation of gene expression in chloroplastsThere is transcriptional regulation of gene expression
in chloroplasts (sect 3d) The case for mitochondria is notyet proven
(iii) There is redox regulation of gene expression inchloroplasts
There is transcriptional redox regulation of geneexpression in chloroplasts (sect 3d) The case for mitochon-dria is not yet proven though see (Allen et al 1995 Kon-stantinov et al 1995 Galvis et al 1998)
(iv) There are no redox signalling components encoded inchloroplast or mitochondrial DNA
Correct but with possible exceptions discussed in sect 3j
(v) There are lsquobacterialrsquo redox signalling components ineukaryotes
See sect 3j
(vi) Further predictions DNA-array results and phenotypesof allotopic mutants
A systematic test of CORR might be to make an oxid-ized-minus-reduced gene expression profile of the lsquotran-scriptomersquo of a cyanobacterium to see if the set of geneswhose transcription is under most rapid and direct redoxcontrol corresponds to the genes of the chloroplast gen-ome Laboratories with this experimental capability do notseem to have addressed this possibility (Hihara et al 2002Ikeuchi 2002) Likewise if a putative modern representa-tive of the proto-mitochondrion can be found and agreedeg Rhodospirillum rubrum or Paracoccus denitrificans thena similar experiment should reveal a subset of genes corre-sponding to mitochondrial genomes DNA-microarraysalso present the possibility of testing the relative rate ofredox response and dependency on other signallinginputs of transcription of eukaryotic genes To dateorganellar genes have not been included in such investi-gations (Ferea et al 1999) A further experimental possi-bility is to examine the redox regulatory phenotypes ofexisting allotopic mutants as outlined in sect 3e
Phil Trans R Soc Lond B (2003)
6 CONCLUSION AND FUTURE PROSPECTS
The CORR hypothesis states that the function of mito-chondrial and chloroplast genomes is to provide co-location (of gene and gene product) for (evolutionary)continuity of redox regulation of gene expression Thishypothesis is neither self-contradictory nor devoid of test-able predictions It is also consistent with available evi-dence and in principle makes sense of the otherwisepuzzling distribution of genes between compartments ineukaryotic cells The explanation that CORR offersapplies equally to mitochondria and chloroplasts andidentifies selective forces that still operate to determine thelocation of genes No selective barriers need be assumedto gene transfer protein import nor membrane insertionNor do we need to invoke a lsquofrozen accidentrsquo that some-thing happened to stop gene transfer for some reason atsome time in the remote and inaccessible past CORR alsoexplains the absence of genomes from organelles that havelost their primary bioenergetic functions CORR may actas a stimulus to a search for residual bioenergetic functionin those organelles that have lost aerobic respiration andphotosynthesis but where residual genomes remain
CORR also predicts regulatory properties of knowncomponents of chloroplasts and mitochondria that in turnindicate important but as yet uncharacterized flexibility inenergy transduction These components include the RUB-ISCO large subunit and the CFo and Fo subunits of coup-ling ATPase in both chloroplasts and mitochondriaRedox regulation of the relative stoichiometry of compo-nents as seen in regulation of chloroplast photosystemstoichiometry (Pfannschmidt et al 1999a) may extend to(C)Fo-(C)F1-ATPase indicating that the function ofretention of genes for one or more of these componentsis flexibility in the stoichiometries of H1 to e2 and H1 toATP in chemiosmotic coupling (Schemidt et al 1998Stock et al 2000 Allen 2002)
So far explicit tests of CORR have largely been con-fined to chloroplasts and to some extent mitochondria ofhigher plants but this merely reflects the expertise andinterests of the investigators Further attempted disproofof CORR is welcomed and encouraged Novel experi-mental tests can be devised as well as attempted refu-tation through analysis of existing data It is hoped thatexplicit attention will be given to the CORR hypothesis inlaboratories already suitably equipped and that resourcescan be allocated for this purpose in laboratories wherethere is the interest but not currently the technology orexpertise
The heuristic value of the hypothesis already seen inthe role of redox-regulated transcription of photosyntheticreaction centre genes in chloroplasts may be its most use-ful property The central proposal is that mitochondriaand chloroplasts contain genes for proteins whose functionin electron transfer demands rapid direct and uncon-ditional redox regulatory control of their biosynthesis Thefull implications of this hypothesis are very broad and onlyjust beginning to be explored
I thank Carol Allen Aubrey de Grey William Martin John ARaven and Francis-Andre Wollman for discussion and corre-spondence during the preparation of this paper and WilliamMartin for suggestions including table 1 Research grants from
Genomes in mitochondria and chloroplasts J F Allen 33
The Swedish Natural Sciences Research Council (NFR) andthe Crafoord Foundation are gratefully acknowledged
REFERENCES
Adams K L Daley D O Qiu Y L Whelan J amp PalmerJ D 2000 Repeated recent and diverse transfers of a mito-chondrial gene to the nucleus in flowering plants Nature408 354ndash357
Alberts B Bray D Lewis J Raff M Roberts K amp Wat-son J D 1994 The molecular biology of the cell New Yorkand London Garland Publishing
Allen C A Hakansson G amp Allen J F 1995 Redox con-ditions specify the proteins synthesized by isolated chloro-plasts and mitochondria Redox Rep 1 119ndash123
Allen J F 1992 Protein phosphorylation in regulation ofphotosynthesis Biochim Biophys Acta 1098 275ndash335
Allen J F 1993a Control of gene expression by redox poten-tial and the requirement for chloroplast and mitochondrialgenomes J Theor Biol 165 609ndash631
Allen J F 1993b Redox control of gene expression and thefunction of chloroplast genomes a hypothesis PhotosynthRes 36 95ndash102
Allen J F 1993c Redox control of transcription sensorsresponse regulators activators and repressors FEBS Lett332 203ndash207
Allen J F 1995 Thylakoid proteinndashphosphorylation state-1ndashstate-2 transitions and photosystem stoichiometry adjust-ment redox control at multiple levels of gene expressionPhysiol Plant 93 196ndash205
Allen J F 1996 Separate sexes and the mitochondrial theoryof ageing J Theor Biol 180 135ndash140
Allen J F 2002 Photosynthesis of ATP-electrons protonpumps rotors and poise Cell 110 273ndash276
Allen J F amp Allen C A 1999 A mitochondrial model forpremature ageing of somatically cloned mammals IUBMBLife 48 369ndash372
Allen J F amp Nilsson A 1997 Redox signalling and the struc-tural basis of regulation of photosynthesis by protein phos-phorylation Physiol Plant 100 863ndash868
Allen J F amp Pfannschmidt T 2000 Balancing the two photo-systems photosynthetic electron transfer governs transcrip-tion of reaction centre genes in chloroplasts Phil TransR Soc Lond B 355 1351ndash1357 (DOI 101098rstb20000697)
Allen J F amp Race H L 2002 Will the real LHC II kinaseplease step forward Sciencersquos STKE httpwwwstkeorgcgicontentfullsigtrans2002155pe43
Allen J F amp Raven J A 1996 Free-radical-induced mutationvs redox regulation costs and benefits of genes in organellesJ Mol Evol 42 482ndash492
Archibald J M Cavalier-Smith T Maier U amp Douglas S2001 Molecular chaperones encoded by a reduced nucleusthe cryptomonad nucleomorph J Mol Evol 52 490ndash501
Ashby M K amp Mullineaux C W 1999a Cyanobacterialycf27 gene products regulate energy transfer from phycobili-somes to photosystems I and II FEMS Microbiol Lett 181253ndash260
Ashby M K amp Mullineaux C W 1999b The role of ApcDand ApcF in energy transfer from phycobilisomes to PSI andPSII in a cyanobacterium Photosynth Res 61 169ndash179
Ashby M K Houmard J amp Mullineaux C W 2002 Theycf27 genes from cyanobacteria and eukaryotic algae distri-bution and implications for chloroplast evolution FEMSMicrobiol Lett 214 25ndash30
Attardi G amp Schatz G 1988 Biogenesis of mitochondria ARev Cell Biol 4 289ndash333
Azem A Oppliger W Lustig A Jeno P Feifel B SchatzG amp Horst M 1997 The mitochondrial hsp70 chaperone
Phil Trans R Soc Lond B (2003)
system effect of adenine nucleotides peptide substrate andmGrpE on the oligomeric state of mhsp70 J Biol Chem272 20 901ndash20 906
Barbrook A C Symington H Nisbet R E R LarkumA amp Howe C J 2001 Organisation and expression of theplastid genome of the dinoflagellate Amphidinium opercula-tum Mol Genet Genomics 266 632ndash638
Bauer C E Elsen S amp Bird T H 1999 Mechanisms forredox control of gene expression A Rev Microbiol 53495ndash523
Bauer C E Elsen S Swem L R Swem D L amp MasudaS 2003 Redox and light regulation of gene expression inphotosynthetic prokaryotes Phil Trans R Soc Lond B 358147ndash154 (DOI 101098rstb20021189)
Berks B C Sargent F De Leeuw E Hinsley A P Stan-ley N R Jack R L Buchanan G amp Palmer T 2000aA novel protein transport system involved in the biogenesisof bacterial electron transfer chains Biochim Biophys ActaBioenerg 1459 325ndash330
Berks B C Sargent F amp Palmer T 2000b The tat proteinexport pathway Mol Microbiol 35 260ndash274
Blackstone N W 1999 Redox control in development andevolution evidence from colonial hydroids J Exp Biol 2023541ndash3553
Blackstone N W 2000 Redox control and the evolution ofmulticellularity Bioessays 22 947ndash953
Blackstone N W 2001 Redox state reactive oxygen speciesand adaptive growth in colonial hydroids J Exp Biol 2041845ndash1853
Bogorad L 1975 Evolution of organelles and eukaryotic gen-omes Science 188 891ndash898
Bogsch E G Sargent F Stanley N R Berks B C Robin-son C amp Palmer T 1998 An essential component of a novelbacterial protein export system with homologues in plastidsand mitochondria J Biol Chem 273 18 003ndash18 006
Brown E C Somanchi A amp Mayfield S P 2001 Interor-ganellar crosstalk new perspectives on signaling from thechloroplast to the nucleus Genome Biol 2 10211ndash10214
Cavalier-Smith T 2002 Chloroplast evolution secondarysymbiogenesis and multiple losses Curr Biol 12 R62ndashR64
Chitnis P R amp Thornber J P 1988 The major light-har-vesting complex of photosystem 2 Aspects of its molecularand cell biology Photosynth Res 16 41ndash63
Chow W S Melis A amp Anderson J M 1990 Adjustmentsof photosystem stoichiometry in chloroplasts improve thequantum efficiency of photosynthesis Proc Natl Acad SciUSA 87 7502ndash7506
Claros M G Perea J Shu Y Samatey F A PopotJ L amp Jacq C 1995 Limitations to in vivo import of hydro-phobic proteins into yeast mitochondria The case of a cyto-plasmically synthesized apocytochrome b Eur J Biochem228 762ndash771
Dalbey R E amp Robinson C 1999 Protein translocation intoand across the bacterial plasma membrane and the plant thy-lakoid membrane Trends Biochem Sci 24 17ndash22
Daley D O Adams K L Clifton R Qualmann S MillarA H Palmer J D Pratje E amp Whelan J 2002 Genetransfer from mitochondrion to nucleus novel mechanismsfor gene activation from Cox2 Plant J 30 11ndash21
Danon A amp Mayfield S P 1994 Light-regulated translationof chloroplast messenger RNAs through redox potentialScience 266 1717ndash1719
de Grey A 2000 Mitochondrial gene therapy an arena for thebiomedical use of inteins Trends Biotechnol 18 394ndash399
Deisenhofer J Epp O Miki K Huber R amp Michel H1985 Structure of the protein subunits in the photosyntheticreaction center of Rhodopseudomonas viridis at 3Ac resolutionNature 318 618ndash624
34 J F Allen Genomes in mitochondria and chloroplasts
Delwiche C F 1999 Tracing the thread of plastid diversitythrough the tapestry of life Am Nat 154 S164ndashS177
Doolittle W F 1999 Phylogenetic classification and the uni-versal tree Science 284 2124ndash2129
Douglas S E 1998 Plastid evolution origins diversitytrends Curr Opin Genet Dev 8 655ndash661
Duby G amp Boutry M 2002 Mitochondrial protein importmachinery and targeting information Plant Sci 162 477ndash490
Eberhard S Drapier D amp Wollman F A 2002 Searchinglimiting steps in the expression of chloroplast encoded pro-teins relations between gene copy number transcriptiontranscript abundance and translation rate in the chloroplastof Chlamydomonas reinhardtii Plant J 31 149ndash160
El Bissati K amp Kirilovsky D 2001 Regulation of psbA andpsaE expression by light quality in Synechocystis species PCC6803 A redox control mechanism Plant Physiol 1251988ndash2000
Ellis R J 1984 The nuclear domination of chloroplast devel-opment Science Progress 69 129ndash142
Embley T M amp Martin W 1998 Molecular evolution ahydrogen-producing mitochondrion Nature 396 517ndash519
Ems S C Morden C W Dixon C K Wolfe K HdePamphilis C W amp Palmer J D 1995 Transcriptionsplicing and editing of plastid RNAs in the non-photosyn-thetic plant Epifagus virginiana Plant Mol Biol 29 721ndash733
Eraso J M amp Kaplan S 2000 From redox flow to gene regu-lation role of the PrrC protein of Rhodobacter sphaeroides241 Biochemistry 39 2052ndash2062
Eraso J M amp Kaplan S 2002 Redox flow as an instrumentof gene regulation Meth Enzymol 348 216ndash229
Escoubas J M Lomas M Laroche J amp Falkowski P G1995 Light-intensity regulation of cab gene-transcription issignaled by the redox state of the plastoquinone pool ProcNatl Acad Sci USA 92 10 237ndash10 241
Ferea T L Botstein D Brown P O amp Rosenzweig R F1999 Systematic changes in gene expression patterns follow-ing adaptive evolution in yeast Proc Natl Acad Sci USA96 9721ndash9726
Forsberg J Rosenquist M Fraysse L amp Allen J F 2001Redox signalling in chloroplasts and mitochondria genomicand biochemical evidence for two-component regulatory sys-tems in bioenergetic organelles Biochem Soc Trans 29403ndash407
Fujita Y 1997 A study on the dynamic features of photosys-tem stoichiometry accomplishments and problems forfuture studies Photosynth Res 53 83ndash93
Gabriel K Buchanan S K amp Lithgow T 2001 The alphaand the beta protein translocation across mitochondrial andplastid outer membranes Trends Biochem Sci 26 36ndash40
Galvis M L E Allen J F amp Hakansson G 1998 Proteinsynthesis by isolated pea mitochondria is dependent on theactivity of respiratory complex II Curr Genet 33 320ndash329
Gatenby A A amp Ellis R J 1990 Chaperone function theassembly of ribulose bisphosphate carboxylase-oxygenase ARev Cell Biol 6 125ndash149
GoldschmidtClermont M 1998 Coordination of nuclear andchloroplast gene expression in plant cells Int Rev CytolSurv Cell Biol 177 115ndash180
Gray M W 1992 The endosymbiont hypothesis revisited IntRev Cytol 141 233ndash357
Gray M W 1999 Evolution of organellar genomes CurrOpin Genet Dev 9 678ndash687
Gray M W 2000 Evolutionary biology mitochondrial geneson the move Nature 408 302ndash305
Gray M W amp Doolittle W F 1982 Has the endosymbionthypothesis been proven Microbiol Rev 46 1ndash42
Gray M W Burger G amp Lang B F 1999 Mitochondrialevolution Science 283 1476ndash1481
Phil Trans R Soc Lond B (2003)
Grebe T W amp Stock J B 1999 The histidine protein kinasesuperfamily Adv Microb Physiol 41 139ndash227
Green B R amp Kuhlbrandt W 1995 Sequence conservationof light-harvesting and stress-response proteins in relation tothe 3-dimensional molecular structure of Lhcii PhotosynthRes 44 139ndash148
Gruissem W amp Tonkyn J C 1993 Control mechanisms ofplastid gene expression Crit Rev Plant Sci 12 19ndash55
Hackstein J H P (and 11 others) 2001 Hydrogenosomesconvergent adaptations of mitochondria to anaerobicenvironments Zool Anal Complex Syst 104 290ndash302
Harrison M A Keen J N Findlay J B C amp Allen J F1990 Modification of a Glnb-like gene-product by photosyn-thetic electron-transport in the cyanobacterium Synechoc-occus 6301 FEBS Lett 264 25ndash28
Harvey A J Kind K L amp Thompson J G 2002 Redoxregulation of early embryo development Reproduction 123479ndash486
Heathcote P Fyfe P K amp Jones M R 2002 Reactioncentres the structure and evolution of biological solarpower Trends Biochem Sci 27 79ndash87
Henze K amp Martin W 2001 How do mitochondrial genesget into the nucleus Trends Genet 17 383ndash387
Herrmann R G amp Westhoff P 2001 Thylakoid biogenesis anddynamics the result of a complex phylogenetic puzzle InRegulation of photosynthesis vol 11 (ed E-M Aro amp BAndersson) pp 1ndash28 Dordrecht The Netherlands Kluwer
Hihara Y Sonoike K Kanehisa M amp Ikeuchi M 2002Search for redox responsive genes in Synechocystis sp PCC6803 by using DNA microarrays Plant Cell Physiol 43S178
Howe C J Barbrook A C amp Lockhart P J 2000 Organellegenes do they jump or are they pushed Trends Genet 1665ndash66
Ikeuchi M 2002 Gene expression and responses to light stressin cyanobacteria Plant Cell Physiol 43 S21
Iuchi S amp Lin E C C 1992 Mutational analysis of signaltransduction by Arcb a membrane sensor protein respon-sible for anaerobic repression of operons involved in the cen-tral aerobic pathways in Escherichia coli J Bacteriol 1743972ndash3980
Jackson-Constan D Akita M amp Keegstra K 2001 Molecu-lar chaperones involved in chloroplast protein importBiochim Biophys Acta Mol Cell Res 1541 102ndash113
Jarvis P amp Soll M 2001 Toc Tic and chloroplast proteinimport Biochim Biophys Acta Mol Cell Res 1541 64ndash79
Jordan P Fromme P Witt H T Klukas O SaengerW amp Krauss N 2001 Three-dimensional structure ofcyanobacterial photosystem I at 25 acngstrom resolutionNature 411 909ndash917
Kaneko T (and 23 others) 1996a Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regionsDNA Res 3 109ndash136
Kaneko T (and 23 others) 1996b Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regions[Supplement] DNA Res 3 185ndash209
Kanevski I amp Maliga P 1994 Relocation of the plastid rbcLgene to the nucleus yields functional ribulose-15-bisphosph-ate carboxylase in tobacco chloroplasts Proc Natl Acad SciUSA 91 1969ndash1973
Kim J amp Mayfield S P 1997 Protein disulfide isomerase as aregulator of chloroplast translational activation Science 2781954ndash1957
Genomes in mitochondria and chloroplasts J F Allen 35
Kirk J T O amp Tilney-Bassett R A E 1978 The plastidsTheir chemistry structure growth and inheritance Amster-dam Elsevier
Koehler C M Merchant S amp Schatz G 1999 How mem-brane proteins travel across the mitochondrial intermem-brane space Trends Biochem Sci 24 428ndash432
Komiya T Rospert S Schatz G amp Mihara K 1997 Bind-ing of mitochondrial precursor proteins to the cytoplasmicdomains of the import receptors Tom70 and Tom20 isdetermined by cytoplasmic chaperones FASEB J 11 1196
Konstantinov Y M Lutsenko G N amp Podsosonny V A1995 Genetic functions of isolated maize mitochondriaunder model changes of redox conditions Biochem MolBiol Int 36 319ndash326
Korab-Laskowska M Rioux P Brossard N LittlejohnT G Gray M W Lang B F amp Burger G 1998 Theorganelle genome database project (GOBASE) Nucleic AcidsRes 26 138ndash144
Kovacs L Wiessner W Kis M Nagy F Mende D ampDemeter S 2000 Short- and long-term redox regulation ofphotosynthetic light energy distribution and photosystemstoichiometry by acetate metabolism in the green alga Chla-mydobotrys stellata Photosynth Res 65 231ndash247
Kuhlbrandt W 1994 Structure and function of the plant light-harvesting complex LHC-II Curr Opin Struct Biol 4519ndash528
Lang B F Gray M W amp Burger G 1999 Mitochondrialgenome evolution and the origin of eukaryotes A RevGenet 33 351ndash397
Lemire B D Fankhauser C Baker A amp Schatz G 1989The mitochondrial targeting function of randomly generatedpeptide sequences correlates with predicted helical amphi-philicity J Biol Chem 264 20 206ndash20 215
Leuenberger D Bally N A Schatz G amp Koehler C M1999 Different import pathways through the mitochondrialintermembrane space for inner membrane proteins EMBOJ 18 4816ndash4822
Li H amp Sherman L A 2000 A redox-responsive regulator ofphotosynthesis gene expression in the cyanobacterium Syne-chocystis sp strain PCC 6803 J Bacteriol 182 4268ndash4277
Lin X Y (and 36 others) 1999 Sequence and analysis of chro-mosome 2 of the plant Arabidopsis thaliana Nature 402761ndash768
Lonsdale D M Hodge T P Howe C J amp Stern D B1983 Maize mitochondrial-DNA contains a sequence hom-ologous to the ribulose-15-bisphosphate carboxylase largesubunit gene of chloroplast DNA Cell 34 1007ndash1014
McFadden G I 1999 Endosymbiosis and evolution of theplant cell Curr Opin Plant Biol 2 513ndash519
McFadden G I 2001 Chloroplast origin and integrationPlant Physiol 125 50ndash53
Mahler H R amp Raff R A 1975 The evolutionary origin ofthe mitochondrion a nonsymbiotic model Int Rev Cytol43 1ndash124
Manfredi G Fu J Ojaimi J Sadlock J E Kwong J QGuy J amp Schon E A 2002 Rescue of a deficiency in ATPsynthesis by transfer of MTATP6 a mitochondrial DNA-encoded gene to the nucleus Nature Genet 30 394ndash399
Marc P Margeot A Devaux F Blugeon C Corral-Debrinski M amp Jacq C 2002 Genome-wide analysis ofmRNAs targeted to yeast mitochondria EMBO Rep 3159ndash164
Marienfeld J Unseld M amp Brennicke A 1999 The mito-chondrial genome of Arabidopsis is composed of both nativeand immigrant information Trends Plant Sci 4 495ndash502
Martin W 1999 A briefly argued case that mitochondria andplastids are descendants of endosymbionts but that thenuclear compartment is not Proc R Soc Lond B 2661387ndash1395 (DOI 101098rspb19990792)
Phil Trans R Soc Lond B (2003)
Martin W amp Herrmann R G 1998 Gene transfer fromorganelles to the nucleus how much what happens andwhy Plant Physiol 118 9ndash17
Martin W amp Muller M 1998 The hydrogen hypothesis forthe first eukaryote Nature 392 37ndash41
Martin W amp Russell M J 2003 On the origins of cells ahypothesis for the evolutionary transitions from abiotic geo-chemistry to chemoautotrophic prokaryotes and from pro-karyotes to nucleated cells Phil Trans R Soc Lond B 35859ndash85 (DOI 101098rstb20021183)
Martin W Stoebe B Goremykin V Hansmann S Hase-gawa M amp Kowallik K V 1998 Gene transfer to thenucleus and the evolution of chloroplasts Nature 393162ndash165
Martin W Hoffmeister M Rotte C amp Henze K 2001 Anoverview of endosymbiotic models for the origins of eukary-otes their ATP-producing organelles (mitochondria andhydrogenosomes) and their heterotrophic lifestyle BiolChem 382 1521ndash1539
Martin W Rujan T Richly E Hansen A Cornelsen SLins T Leister D Stoebe B Hasegawa M amp PennyD 2002 Evolutionary analysis of Arabidopsis cyanobacterialand chloroplast genomes reveals plastid phylogeny and thou-sands of cyanobacterial genes in the nucleus Proc NatlAcad Sci USA 99 12 246ndash12 251
Millen R S (and 12 others) 2001 Many parallel losses of infAfrom chloroplast DNA during angiosperm evolution withmultiple independent transfers to the nucleus Plant Cell 13645ndash658
Morse D Salois P Markovic P amp Hastings J W 1995 Anuclear-encoded form-II RUBISCO in dinoflagellatesScience 268 1622ndash1624
Murakami A Kim S J amp Fujita Y 1997 Changes in photo-system stoichiometry in response to environmental con-ditions for cell growth observed with the cyanophyteSynechocystis PCC 6714 Plant Cell Physiol 38392ndash397
Oh J I amp Kaplan S 2000 Redox signaling globalization ofgene expression EMBO J 19 4237ndash4247
Ozawa T 1995 Mitochondrial-DNA mutations associatedwith aging and degenerative diseases Exp Gerontol 30269ndash290
Palmer J D 1997 Organelle genomes going going goneScience 275 790ndash791
Pearson C K Wilson S B Schaffer R amp Ross A W 1993NAD turnover and utilization of metabolites for RNA syn-thesis in a reaction sensing the redox state of the cyto-chrome-b( 6 )f complex in isolated chloroplasts Eur JBiochem 218 397ndash404
Pfanner N amp Geissler A 2001 Versatility of the mitochon-drial protein import machinery Natl Rev Mol Cell Biol 2339ndash349
Pfanner N amp Wiedemann N 2002 Mitochondrial proteinimport two membranes three translocases Curr Opin CellBiol 14 400ndash411
Pfannschmidt T Nilsson A amp Allen J F 1999a Photosyn-thetic control of chloroplast gene expression Nature 397625ndash628
Pfannschmidt T Nilsson A Tullberg A Link G amp AllenJ F 1999b Direct transcriptional control of the chloroplastgenes psbA and psaAB adjusts photosynthesis to light energydistribution in plants IUBMB Life 48 271ndash276
Pfannschmidt T Allen J F amp Oelmuller R 2001a Prin-ciples of redox control in photosynthesis gene expressionPhysiol Plant 112 1ndash9
Pfannschmidt T Schutze K Brost M amp Oelmuller R2001b A novel mechanism of nuclear photosynthesis generegulation by redox signals from the chloroplast during pho-
36 J F Allen Genomes in mitochondria and chloroplasts
tosystem stoichiometry adjustment J Biol Chem 27636 125ndash36 130
Popot J L amp de Vitry C 1990 On the microassembly of inte-gral membrane proteins A Rev Biophys Biophys Chem 19369ndash403
Race H L Herrmann R G amp Martin W 1999 Why haveorganelles retained genomes Trends Genet 15 364ndash370
Raven J A Johnston A M Parsons R amp Kubler J 1994aThe influence of natural and experimental high O2 concen-trations on O2 evolving phototrophs Biol Rev CambridgePhil Soc 69 61ndash94
Raven J A Johnston A M Parsons R amp Kubler J 1994bThe occurrence and influence on photolithotrophs of highoxygen concentrations Proc R Soc Edinb B 102 193ndash201
Robinson C 2000 The twin-arginine translocation system anovel means of transporting folded proteins in chloroplastsand bacteria Biol Chem 381 89ndash93
Robinson C amp Bolhuis A 2001 Protein targeting by the twin-arginine translocation pathway Natl Rev Mol Cell Biol 2350ndash356
Rodermel S 2001 Pathways of plastid-to-nucleus signalingTrends Plant Sci 6 471ndash478
Roger A J 1999 Reconstructing early events in eukaryoticevolution Am Nat 154 S146ndashS163
Roger A J Svard S G Tovar J Clark C G SmithM W Gillin F D amp Sogin M L 1998 A mitochondrial-like chaperonin 60 gene in Giardia lamblia evidence thatdiplomonads once harbored an endosymbiont related to theprogenitor of mitochondria Proc Natl Acad Sci USA 95229ndash234
Roodyn D B amp Wilkie D 1968 The biogenesis of mitochondriaMethuenrsquos monographs on biological subjects London Methuen
Rujan T amp Martin W 2001 How many genes in Arabidopsiscome from cyanobacteria An estimate from 386 proteinphylogenies Trends Genet 17 113ndash120
Saccone C Gissi C Lanave C Larizza A Pesole G ampReyes A 2000 Evolution of the mitochondrial genetic sys-tem an overview Gene 261 153ndash159
Salzberg S L White O Peterson J amp Eisen J A 2001Microbial genes in the human genome lateral transfer orgene loss Science 292 1903ndash1906
Sargent F Berks B C amp Palmer T 2002 Assembly of mem-brane-bound respiratory complexes by the Tat protein-trans-port system Arch Microbiol 178 77ndash84
Schatz G 1998 Protein transport the doors to organellesNature 395 439ndash440
Schemidt R A Qu J Williams J R amp Brusilow W S1998 Effects of carbon source on expression of F0 genes andon the stoichiometry of the c subunit in the F1 F0 ATPaseof Escherichia coli J Bacteriol 180 3205ndash3208
Selosse M A Albert B R amp Godelle B 2001 Reducing thegenome size of organelles favours gene transfer to thenucleus Trends Ecol Evol 16 135ndash141
Stern D B amp Lonsdale D M 1982 Mitochondrial and chlo-roplast genomes of maize have a 12- kilobase DNA-sequencein common Nature 299 698ndash702
Stock D Gibbons C Arechaga I Leslie A G W ampWalker J E 2000 The rotary mechanism of ATP synthaseCurr Opin Struct Biol 10 672ndash679
Stoebe B amp Maier U G 2002 One two three naturersquos toolbox for building plastids Protoplasma 219 123ndash130
Stoebe B Martin W amp Kowallik K V 1998 Distributionand nomenclature of protein-coding genes in 12 sequencedchloroplast genomes Plant Mol Biol Rep 16 243ndash255
Stupar R M Lilly J W Town C D Cheng Z Kaul SBuell C R amp Jiang J M 2001 Complex mtDNA consti-tutes an approximate 620 kb insertion on Arabidopsis thali-ana chromosome 2 implication of potential sequencing
Phil Trans R Soc Lond B (2003)
errors caused by large-unit repeats Proc Natl Acad SciUSA 98 5099ndash5103
Surpin M Larkin R M amp Chory J 2002 Signal transduc-tion between the chloroplast and the nucleus Plant Cell 14S327ndashS338
Tjaden J Schwoppe C Mohlmann T Quick P W ampNeuhaus H E 1998 Expression of a plastidic ATPADPtransporter gene in Escherichia coli leads to a functional aden-ine nucleotide transport system in the bacterial cytoplasmicmembrane J Biol Chem 273 9630ndash9636
Tovar J Fischer A amp Clark C G 1999 The mitosome a novelorganelle related to mitochondria in the amitochondrial para-site Entamoeba histolytica Mol Microbiol 32 1013ndash1021
Trebitsh T amp Danon A 2001 Translation of chloroplastpsbA mRNA is regulated by signals initiated by both photos-ystems II and I Proc Natl Acad Sci USA 98 12 289ndash12 294
Tullberg A Alexciev K Pfannschmidt T amp Allen J F2000 Photosynthetic electron flow regulates transcription ofthe psaB gene in pea (Pisum sativum L) chloroplasts throughthe redox state of the plastoquinone pool Plant Cell Physiol41 1045ndash1054
Turmel M Lemieux C Burger G Lang B F Otis CPlante I amp Gray M W 1999 The complete mitochondrialDNA sequences of Nephroselmis olivacea and Pedinomonasminor two radically different evolutionary patterns withingreen algae Plant Cell 11 1717ndash1729
Unden G amp Bongaerts J 1997 Alternative respiratory path-ways of Escherichia coli energetics and transcriptional regu-lation in response to electron acceptors Biochim BiophysActa Bioenerg 1320 217ndash234
Unseld M Marienfeld J R Brandt P amp Brennicke A1997 The mitochondrial genome of Arabidopsis thaliana con-tains 57 genes in 366 924 nucleotides Nature Genet 1557ndash61
Van der Giezen M Slotboom D J Horner D S DyalP L Harding M Xue G P Embley T M amp KunjiE R S 2002 Conserved properties of hydrogenosomal andmitochondrial ADPATP carriers a common origin for bothorganelles EMBO J 21 572ndash579
Von Heijne G 1986 Why mitochondria need a genome FEBSLett 198 1ndash4
Westphal S Soll J amp Vothknecht U C 2001 A vesicle trans-port system inside chloroplasts FEBS Lett 506 257ndash261
Whitehouse H L K 1969 Towards an understanding of themechanism of heredity London Edward Arnold Ltd
Williams B A P Hirt R P Lucocq J M amp Embley T M2002 A mitochondrial remnant in the microsporidian Trachi-pleistophora hominis Nature 418 865ndash869
Wilson R J (and 10 others) 1996a Complete gene map of theplastid-like DNA of the malaria parasite Plasmodium falcipa-rum J Mol Biol 261 155ndash172
Wilson S B Davidson G S Thomson L M amp PearsonC K 1996b Redox control of RNA synthesis in potato mito-chondria Eur J Biochem 242 81ndash85
Winkler H H amp Neuhaus H E 1999 Non-mitochondrialATP transport Trends Biochem Sci 24 64ndash68
Wolfe K H Morden C W amp Palmer J D 1992 Small sin-gle-copy region of plastid DNA in the non-photosyntheticangiosperm Epifagus virginiana contains only two genes Dif-ferences among dicots monocots and bryophytes in geneorganization at a non-bioenergetic locus J Mol Biol 22395ndash104
Wollman F A Minai L amp Nechushtai R 1999 The biogen-esis and assembly of photosynthetic proteins in thylakoidmembranes Biochim Biophys Acta Bioenerg 1411 21ndash85
Yeliseev A A amp Kaplan S 2000 TspO of Rhodobactersphaeroides a structural and functional model for the mam-
Genomes in mitochondria and chloroplasts J F Allen 37
malian peripheral benzodiazepine receptor J Biol Chem275 5657ndash5667
Zerges W 2002 Does complexity constrain organelle evol-ution Trends Plant Sci 7 175ndash182
Zhang Z D Green B R amp Cavalier-Smith T 1999 Singlegene circles in dinoflagellate chloroplast genomes Nature400 155ndash159
Zhang Z D Cavalier-Smith T amp Green B R 2001 A fam-ily of selfish minicircular chromosomes with jumbled chloro-plast gene fragments from a dinoflagellate Mol Biol Evol18 1558ndash1565
Zouni A Witt H T Kern J Fromme P Krauss NSaenger W amp Orth P 2001 Crystal structure of photosys-tem II from Synechococcus elongatus at 38 acngstrom resol-ution Nature 409 739ndash743
DiscussionD S Horner (Dipartimento di Fisiologia e Biochimica
Generali University of Milan Milan Italy) Given thatthere are components of these membrane-bound energy-transduction pathways that are consistently nuclear enco-ded not the key ones you pointed out but others do youexpect perhaps a slower perhaps a more crude but yet aredox communication between the organelle andexpression of nuclear-encoded genes
J F Allen There is and there is no question about itI think that Bogoradrsquos lock-in hypothesis has somethingto say here and there are other people who say it is all todo with assembling multi-subunit complexes If you havecontrol over the initial construction of the core aroundwhich the whole complex is eventually built then redoxcontrol or some sort of control over synthesis of that corewill control construction of the whole thing eventuallyand I think that may well be largely correct I think it ispossible in principle for nuclear genes to make precursorsaccording to redox instructions but it is not really soimportant that those precursors are here and now presentin exactly the right quantities because the precursors canto a limited extent hang around and wait Actually thereis proteolytic degradation of unattached precursors and soon but I think the fact that these chloroplast and mito-chondrially DNA-encoded subunits are at the core ofthese energy-transducing complexes is the crucial issue
T Cavalier-Smith (Department of Zoology University ofOxford Oxford UK ) In the case of mitochondria the sub-set of genes that remains is much more variable than youimplied It runs from three proteins in the minimal casesup to I think over 90 That is a big difference and manyof the genes in different lineages of protists that remainhave nothing to do with redox control they are things likeribosomal proteins and so on So I think that even thoughwhat you say about redox control of certain genes mayactually be an important thing it is not the fundamentaloverall reason why the genomes are maintained It may bepart of the explanation for why certain genes are alwaysincluded in that subset but even in mitochondria I thinkthat cytochrome b is the only one that is always presentSo I think that both this great diversity of what has beenretained in different lineages and the nucleomorph caseindicate that evolution was able to do one thing in onelineage 500 million years ago and never got around todoing it for 500 million years in another lineage so thereis a lot more haphazardness and evolutionary accidentsand contingency than a straight functional interpretation
Phil Trans R Soc Lond B (2003)
there is a lot of just sheer lsquohappenedndashorndashdidnrsquotndashhappenrsquoor carryover
J F Allen I was not implying anything about the rangeof mitochondrial genomes I have a limited amount oftime so I just talked about what I could talk about withinthat limit Of course there will be a range and this willdepend upon the physiology and bioenergetics of theparticular organism But what I am trying to do is to seewhat the common properties of the irreducible core areof components that are always encoded in the organellesand cytochrome b is a perfect example because that reallyis the thing that binds these two all-important quinoneswithout which there is no Q cycle without which there isno proton pumping and bioenergetics and in Plasmodiumwhere the mitochondrion is reduced to this bare mini-mum there is still a mitochondrial gene for cytochromeb Now my argument about the ribosomal proteins andall the rest of it is that in order to be able to exert thisall-important redox control over gene expression of certaincore components you need an in situ genetic system andthis can vary in composition for reasons that I cannotdescribe Maybe you need RNA polymerase or maybeyou can import RNA polymerase In plastids you haveboth of these options You are going to need ribosomestransfer RNAs and the rest of the apparatus of geneexpression and that needs to be in situ but we are stillleft with the question why does there have to be any gen-etic system at all The mature tobacco or spinach chloro-plast seems to have an entire genetic system just to makeone protein D1 which is all it ever needs to resynthesizeI think that this is a clue It is worth having this very unec-onomical crazy system because there is an overridingrequirement to connect electron transfer with geneexpression directly and unambiguouslymdashwithout recourseto a bureaucracy if you like which is what I think thenucleus is
K Willison (Institute of Cancer Research Chester BeattyLaboratories London UK ) These photosynthetic reactioncomplexes are a sort of baroque mixture of polypeptidechains and cofactors and my understanding is that someof them in high photon flux can turn over within say 30minutes so I am wondering if there is a correlationbetween gene retention and the turnover rates of some ofthe components and the complexes it seems to me thatthat could really be a limiting factor and the reason thatthe proteins had to be encoded locally in order to get thecomplexes to be assembled rapidly
J F Allen I think that is right You can only alter thesteady-state concentration of a protein complex by affect-ing the synthesis and breakdown and therefore turnoverThere is an extreme example the D1 protein encoded bythe psbA gene of chloroplasts which is always in the chlor-oplast genome without exception High light will tend todegrade this protein and when the high light goes awaythere is an absolute need very quickly to bring it back upagain I think that is a very good example Whether thisargument applies to genes such as cytochrome oxidase Ido not know
C J Leaver (Department of Plant Sciences University ofOxford Oxford UK ) Your experiments tend to relatemainly to the gene expression side What about proteinRNA turnover the whole concept of redox control ofthatmdashdo we know anything
38 J F Allen Genomes in mitochondria and chloroplasts
J F Allen Well quite a lot is known about chloro-plasts There is redox control of messenger RNA stabilityfor this same D1 protein which has been described byStephen Mayfield and his co-workers The evidence is thatthe site of redox control is ferredoxin via thioredoxin andthe implication is that thioredoxin tells whether non-cyclicelectron transport is working at all When it is workingthere is a signal that says lsquoOK now continue with all theprotein synthesisrsquo It is not particularly specific but moreof a global effect I think this is what Mayfield argues butof course this is not inconsistent with my argument I haveconcentrated on quinone pools and medium-potentialredox regulatory control because that is where I can seesignals about the dangerous consequences of getting itwrong but there is control of other levels of geneexpression In mitochondria I am not so sure I wouldbounce the question back to you on that one
W Martin (Institute of Botany III Heinrich-Heine Univ-ersitat Dusseldorf Dusseldorf Germany) The nucleomorphgenome was cited as evidence in conflict with your modelbut is that true The nucleomorph genome is not in abioenergetic organelle in that sense It resides in the cyto-sol so does the size of the nucleomorph genome workagainst your model Is it really evidence against yourmodel
J F Allen I do not know why should it be evidenceI cannot explain why the nucleomorph has a genomeThat is why I clutched at the straw that it is storage fornucleic acid precursors
Phil Trans R Soc Lond B (2003)
T Cavalier-Smith The reason I introduced the nucleo-morph is not to say it is direct evidence against yourhypothesis but to show that there probably was no singlefunctional reason for the retention of every gene in everyorganelle It may be that there are specific physiologicalreasons for the retention of certain genes and thereforeyou might be right about a subset of genes but I verymuch doubt if there are specific physiological reasons forthe retention of every single gene that is retained differen-tially because odd things happen in evolution Somethings are easy and sometimes things are difficult Evol-ution is history
J F Allen Of course I agree with that I do not thinkI can explain everything but I do think I can explainsomething If I have done that then I am content
GLOSSARY
AAC ADPndashATP carrierCES control by epistasy of synthesisCORR co-location for redox regulationLHC light-harvesting complexPCC Pasteur culture collectionORF open reading frameRUBISCO ribulose-15-bisphosphate carboxylasendashoxygenaseTAT twin-arginine translocase
Visit httpwwwpubsroyalsocacuk to see an electronic appendix tothis paper

Genomes in mitochondria and chloroplasts J F Allen 21
Table 1 Retention of cyanobacterial genes in chloroplast DNA(Protein-coding genes are sorted into the functional categories assigned by Kaneko et al (1996ab) The genes are named andtheir distribution recorded in 16 completely sequenced chloroplast genomes to date in Martin et al (2002) from which the dataare taken Electronic Appendix A (available on The Royal Societyrsquos Publications Web site) lists the genes sorted into the functionalcategories of their gene products GPC genes per functional category The number of genes present in each category in thecyanobacterium Synechocystis PCC 6803 (Syn) and in any chloroplast genome completely sequenced (cp) Cat Ret categoryretention The number of times that a Synechocystis functional category is represented by a gene in cpDNA divided by the numberof genes in that category in Synechocystis itself Cat Ret = cpSyn For example Synechocystis has 84 genes for amino-acid biosynth-esis 10 of which are still present in at least one of 16 sequenced plastid genomes Hence Cat Ret = 1084 = 119 for amino-acid biosynthesis Deg Ret degree of retention The number of genes present in 16 chloroplast genomes divided by 16 andmultiplied by the number of times that the functional category is represented in chloroplast DNA For example in the 16 genomesanalysed there are 10 different amino-acid biosynthetic genes that occur in at least one cpDNA But of the 160 elements of theresulting matrix that could be positive only 27 are Hence Deg Ret = 27(16 acute 10) = 169 for amino-acid biosynthesis F(ret)fraction of retention The number of genes present in 16 chloroplast genomes divided by 16 and multiplied by the number oftimes that the functional category is represented in Synechocystis For example in the 16 genomes sequenced 27 different amino-acid biosynthetic genes occur in cpDNA Synechocystis has 84 genes for amino-acid biosynthesis that can be assumed to havebeen present in the common ancestor of these 16 plastid genomes Hence for amino-acid biosynthesisF(ret) = 27(16 acute 84) = 20 The genes analysed here correspond to those tabulated by Martin et al (2002) Only genes withknown homologues in cyanobacterial genomes were included in the analysis Values were calculated from the supplementarymaterial of Martin et al (2002) which was downloaded from httpwwwpnasorgcgicontentfull182432999DCI at the PNASWeb site)
no of GPC
Syn cp Cat Ret () Deg Ret () F(ret) ()
amino-acid biosynthesis 84 10 119 169 20cofactorprosthetic group biosynthesis 108 21 194 202 39cell envelope 64 7 109 250 27cellular processes 68 19 279 210 59central metabolism 31 1 32 63 02energy metabolism 86 3 35 125 04fatty-acid biosynthesis 35 8 229 172 39photosynthesis and respiration 139 74 532 603 321photosystem I 12 11 917 625 573photosystem II 25 20 800 806 645purine and pyrimidine biosynthesis 39 1 26 63 02regulatory functions 136 7 51 178 09DNA replication 49 2 41 156 06transcription 23 4 174 1000 130translation 144 56 389 531 207transport 158 2 13 63 01other plus hypothetical 587 43 73 202 15
(a) The bacterial nature of mitochondria andchloroplasts
It is generally accepted that organellar genetic systemsare evidence for the evolutionary origin of mitochondriaand chloroplasts as free-living eubacteria or a-proteobac-teria which entered into close inter-dependency with ahost cell (Douglas 1998 Delwiche 1999 Lang et al 1999McFadden 1999 2001 Roger 1999 Hackstein et al2001 Stoebe amp Maier 2002) The genetic systemsretained by organelles have many distinctively bacterialfeatures and heterologous systems for gene expression canbe reconstituted in vitro from organellar and bacterialcomponents By sharp contrast the genetic systems of theeukaryotic cell nucleus and cytosol are only distantlyrelated to those of bacteria and to those of their own cyto-plasmic organelles Instead nucleo-cytoplasmic geneticsystems more closely resemble the genetic systems ofarchaea (also known as archaebacteria) It is has been sug-gested that eukaryotic cells are an archaeal system of infor-mation storage replication transcription and translationthat is powered by a eubacterial system of energy trans-
Phil Trans R Soc Lond B (2003)
duction (Martin amp Russell 2003) The genetic systems ofmitochondria and chloroplasts can then be viewed aseukaryotic cellsrsquo residual bacterial systems of informationstorage and transmission Eukaryotic cellsrsquo residualarchaeal systems of energy transduction might theninclude direct vectorial ion pumps such as P- and V-ATP-ases and seven-helix transporters now associated prim-arily with information and signalling However the centralprocesses of energy transduction respiration and photo-synthesis are clearly bacterial in origin
(b) Currencies of symbiosisOne hypothesis for the origin of mitochondria states
that the nucleo-cytoplasmic system of eukaryotes isderived from that of an archaeal methanogenic chemo-autotroph that came to rely on the precursor of mitochon-dria for its supply of molecular hydrogen (Martin ampMuller 1998) This lsquohydrogen hypothesis for the firsteukaryotersquo resolves a long-held but paradoxical feature ofthe endosymbiont hypothesis how could a heterotrophichost cell have been able to supply abundant organic sub-
22 J F Allen Genomes in mitochondria and chloroplasts
strates that the bacterial precursor of mitochondria couldnot already obtain for itself Furthermore a second para-dox is resolved or at a least avoided export of ATP fromthe bacterial partner was not an immediate requirementfor the symbiosis to become established and could haveevolved subsequently Thus it is not necessary to postu-late a suicidal bacterium that excretes ATP nor a deferredselective advantage to the partnership implying foresightor an assurance of eventual contract fulfilment
The hydrogen hypothesis (Martin amp Muller 1998) seeksto provide an explanation of the origin of mitochondriathat accounts for the immediate benefit of the primordialsymbiosis to each partner but the implications of thehydrogen hypothesis for the origin of chloroplasts seemnot to have been considered in detail lsquoWhat was in it forthe proto-chloroplastrsquo may be an even more acute ques-tion than lsquowhat was in it for the proto-mitochondrionrsquoas speaking metabolically and energetically cyanobacteriacan do almost anything
Eukaryotes whether autotrophic or heterotrophic havenow mostly adopted the aerobic part of the energy metab-olism of their original bacterial constituents though inter-esting exceptions are known Respiration andphotosynthesis are antiparallel hydrogen transfer (redox)reactions
respiration
light photosynthesis
CH2O + A2 + H2O 2H2A + CO2
As in the Van Niel equation for photosynthesis A maybe an organic moiety for example giving H2A as succi-nate an inorganic element such as sulphur or evenlsquonothingrsquo so that H2A becomes H2 When A is O(oxygen) it becomes obvious that O2 in photosynthesiscomes from H2O not CO2 and the equation simplifies to
respiration
light photosynthesis
CH2O + O2 H2O + CO2
Phosphate group transfer and electron-transfer-linkedATP synthesis are intermediates in all cases In respir-ation CO2 is produced from stepwise oxidative decar-boxylations of pyruvate dehydrogenase and reactions ofthe tricarboxylic acid cycle In most forms of photosyn-thesis CO2 is assimilated to form initially sugar phos-phates
The mitochondrion exports ATP and CO2 and importsADP inorganic phosphate an electron acceptor andpyruvate Energetically the preferred electron acceptor ismolecular oxygen Oxygen comes from the environmentcells and cell membranes are freely permeable to oxygenThe primary metabolic division of labour between thechloroplast and the plant cytosol is that the chloroplastexports oxygen and reduced sugar phosphates (triosephosphate) and imports CO2 less reduced sugars (3-phosphoglycerate) and inorganic phosphate The light
Phil Trans R Soc Lond B (2003)
comes directly from the external environment and in thisrespect the chloroplast thylakoid is the external boundaryof the plant cell where light both as energy and infor-mation is first received The CO2 is also an environmentalinput though its supply may be regulated locally by CO2-or HCO2
3 -concentrating mechanismsAt the cellular level the mitochondrial inner membrane
is the first place where altered O2 supply exerts its pro-found metabolic effects Even granted specialized photore-ceptors the chloroplast thylakoid membrane is first placewhere changes in light quality and intensity affect a plantrsquosprimary input of energy Temperature changes selectivelyaffect diffusion-limited metabolism involving solubleintermediates In both locations abiotic environmentalchanges therefore have immediate and very far-reachingeffects on electron transfer and redox chemistry Theinternal membranes of mitochondria and chloroplastsstand at the energetic interface of the cell with the physicalenvironment and are likely to be first to know about andrespond to its changes
2 THE PROBLEM OF CYTOPLASMIC GENETICSYSTEMS
Given that the genomes of mitochondria and chloro-plasts are derived from those of once free-living bacteriait is clear that most of the bacterial genes that have sur-vived in any form have now been removed to the cellnucleus If most genes can be moved to the nucleus thenwhy not any gene And if any gene can be moved whynot all Why have mitochondria and chloroplasts retainedfunctional genomes at all What do the genes of mitochon-dria and chloroplasts have in common that has resultedin their retention
Alberts et al (1994 p 400) succinctly describe theproblem posed by the persistence of cytoplasmic geneticsystems as follows
Why do mitochondria and chloroplasts require their ownseparate genetic systems when other organelles thatshare the same cytoplasm such as peroxisomes and lyso-somes do nothellip The reason for such a costly arrange-ment is not clear and the hope that the nucleotidesequences of mitochondrial and chloroplast genomeswould provide the answer has proved unfounded Wecannot think of compelling reasons why the proteinsmade in mitochondria and chloroplasts should be madethere rather than in the cytosol
3 CORR CO-LOCATION OF GENES AND GENEPRODUCTS FOR REDOX REGULATION OF GENE
EXPRESSION
This hypothesis states that mitochondria and chloro-plasts contain genes whose expression is required to beunder the direct regulatory control of the redox state oftheir gene products or of electron carriers with whichtheir gene products interact These genes comprise a coreor primary subset of organellar genes The requirementfor redox control of each gene in the primary subset thenconfers a selective advantage upon location of that genewithin the organelle instead of in the cell nucleus Chloro-plast and mitochondrial genomes also contain genes for
Genomes in mitochondria and chloroplasts J F Allen 23
components of the chloroplast and mitochondrial geneticsystems themselves These genes comprise a secondarysubset of organellar genes genetic system genes There isgenerally no requirement for redox control of expressionof genes in this secondary subset though retention ofredox control of genetic system genes may in some casesallow amplification of redox signals acting upon genes inthe primary subset Retention of genes of the secondarysubset (genetic system genes) is necessary for the oper-ation of redox control of expression of genes in the primarysubset (bioenergetic genes) Without genes in the primarysubset the function of genes in the secondary subset iseventually lost and organelles lose their genomes
This hypothesis of co-location for redox regulation ofgene expression CORR was first outlined in a review(Allen 1992) The hypothesis was then systematically putforward in two articles (Allen 1993ab) developed(Allen amp Raven 1996) and has been independentlyreviewed (Race et al 1999) Some of its predictions havebeen tested experimentally as described later
Figure 3 illustrates the general idea of redox regulatorycontrol operating on mitochondrial genes to give the dis-tribution of sites of protein synthesis seen in figure 1 Fig-ure 4 shows the corresponding predicted pattern ofchloroplast redox regulatory control giving rise to the dis-tribution of sites of protein synthesis seen in figure 2Table 1 shows a functional classification of cyanobacterialgenes retained in 16 completely sequenced chloroplastgenomes This analysis (table 1) reveals a strong selectiveretention of genes for subunits of complexes involved inrespiration and photosynthesis and especially in chloro-plast photosystems I and II The pattern (table 1) seemsto be broadly consistent with CORR (figure 4) and incon-sistent with alternative hypotheses (see sect 4) It is reason-able to view photosynthesis as a process where preciserapid and coordinated redox regulatory control of geneexpression repays the cost of maintaining a chloroplastgenetic system
The CORR hypothesis is based on several propositionsassumptions or principles 10 of which are stated explicitlyand listed below Some of the principles are already generallyaccepted some are shared with competing hypothesesand some make direct experimental predictions Howeverother underlying principles of CORR are themselves generalprinciples or subsidiary hypotheses for which no directproof is available but for which predictions are made andpotentially falsifying observations or results can be imagined
The 10 principles that form the basis of the CORRhypothesis follow and each will be critically discussed inthe light of available evidence
(a) The principle of endosymbiotic originAs now generally agreed bioenergetic organelles have
evolved from free-living bacteriaAlthough once controversial (Mahler amp Raff 1975) this
principle will be taken as an axiom and not consideredfurther here The reader is referred to Gray amp Doolittle(1982) Gray (1992) Martin (1999) Martin et al (2001)to sect 1 and to accompanying articles in this journal issue
(b) The principle of unselective gene transferGene transfer between the symbiont or organelle and the cell
nucleus may occur in either direction and is not selective forparticular genes
Phil Trans R Soc Lond B (2003)
This principle takes the form of a testable assertion thatseems to be consistent with current knowledge Nothingcertain seems to be known about the mechanism of intra-cellular gene relocation (Palmer 1997 Martin amp Herrm-ann 1998 Martin et al 1998 Adams et al 2000 Henze ampMartin 2001 Millen et al 2001 Rujan amp Martin 2001Daley et al 2002) Plausible scenarios can be imagined onthe basis of the classical genetical phenomena of trans-formation transduction and transfection However asthere is no agreed mechanism there is no basis forasserting a mechanistic argument for selectivity Genetransfer between the organelle and nucleus seems to be aspecial case of lateral or horizontal gene transfer(Doolittle 1999) and no mechanistic basis seems to becurrently available for selectivity in what may be trans-ferred It should also be pointed out that lateral genetransfer between the organelle and nucleus does not haveto occur within a single cell nor within a multicellularindividual (although here any vertically transmitted genemust be incorporated into the germ line) for the outcomeof copying genes to novel compartments to be securedThus any plausible mechanism for heterologous recombi-nation may apply equally to gene transfer between com-partments Mitochondrial departures from the lsquouniversalrsquogenetic code and biased codon usage in chloroplasts (seelater) may be of selective value in that a barrier is therebyput in place to expression of genes in the lsquowrongrsquo compart-ment This barrier is unlikely to be insuperable especiallyif the lsquowrongrsquo compartment turns out actually to be a bet-ter location Impeded relocation might confer stabilitywithout being a primary reason for the existence of cyto-plasmic genomes
(c) The principle of unselective protein importThere is no barrier to the successful import of any precursor
protein nor to its processing and assembly into a functionalmature form
As with sect 3b this principle takes the form of a testableassertion that seems to be consistent with current knowl-edge The principle is discussed in more detail in sect 4because its negation (some proteins cannot be imported)has previously been put forward in itself as a reason forthe retention of cytoplasmic genomes Good hypothesesare easily falsified and they therefore forbid certain obser-vations The principle of unselective protein import for-bids the existence of a protein even a synthetic one thatcannot be imported by some means as a precursor
(d) The principle of continuity of redox controlDirect redox control of expression of certain genes was present
in the bacterial progenitors of chloroplasts and mitochondriaand was vital for selectively advantageous cell function beforeduring and after the transition from bacterium to organelle Themechanisms of this control have been conserved
This principle has been tested experimentally in the pastfew years and seems to be corroborated by an emergingpicture of redox signalling in bacteria and organellesExperiments carried out in vitro on the products of proteinsynthesis (Allen et al 1995 Galvis et al 1998) or RNAsynthesis (Pearson et al 1993 Konstantinov et al 1995Wilson et al 1996b) are consistent with a general phenom-enon of redox dependency of gene expression in bioener-getic organelles Redox control of chloroplast transcription
24 J F Allen Genomes in mitochondria and chloroplasts
nucleus cytosol
O2
H2O
redox regulation
N-phasemitochondrial matrix
Figure 3 Gene expression and principal pathways of biosynthesis of subunits of protein complexes involved in respiration andoxidative phosphorylation in animal mitochondria As in figure 1 reddish-brown DNA RNA and protein subunits are locatedand synthesized in the mitochondrial matrix light brown protein subunits have genes (also light brown) in the nucleus andare imported from the cytosol as precursors White genes and ribosomal and protein subunits are nuclearndashcytoplasmic and ofarchaebacterial origin Reddish-brown and light-brown genes and ribosomal and protein subunits are of bacterial origin Themajor variable environmental input is oxygen (blue) The essence of the CORR hypothesis is that it is beyond the ability ofthe nuclearndashcytoplasmic system to respond rapidly and directly to changes in oxygen concentration or partial pressure and soredox regulation of gene expression (red arrows) has been retained from the ancestral endosymbiont This redox regulationrequires co-location of certain genes with their gene products within the mitochondrion
nucleus cytosol
CH2O
CO2
redox regulation
N-phasechloroplast stroma
light light
Figure 4 Gene expression and principal pathways of biosynthesis of subunits of protein complexes involved inphotosynthesis in chloroplasts As in figure 2 green DNA RNA and protein subunits are located and synthesized in thechloroplast light brown protein subunits have genes (also light brown) in the nucleus and are imported from the cytosolas precursors White genes and ribosomal and protein subunits are nuclearndashcytoplasmic and of archaebacterial originGreen and light-brown genes and ribosomal and protein subunits are of bacterial origin The major environmental inputsare light (orange arrows) and carbon dioxide (blue) The essence of the CORR hypothesis is that it is beyond the abilityof the nuclearndashcytoplasmic system to respond rapidly and directly to changes in light and carbon dioxide and so redoxregulation of gene expression (red arrows) has been retained from the ancestral endosymbiont This redox regulationrequires co-location of certain genes with their gene products within the chloroplast
Phil Trans R Soc Lond B (2003)
Genomes in mitochondria and chloroplasts J F Allen 25
has been demonstrated (Pfannschmidt et al 1999abAllen amp Pfannschmidt 2000 Tullberg et al 2000) and wasprompted by this principle
The level of redox control of chloroplast transcriptionestablished unequivocally so far is that of plastoquinonean electron and hydrogen atom carrier located strategicallybetween photosystems I and II of photosynthesis Thedirections in which the effects are exerted are functionallyintelligible reduction of plastoquinone increases photosy-stem I transcription and decreases photosystem IItranscription oxidation of plastoquinone decreases photo-system I transcription and increases photosystem II tran-scription Plastoquinone in normal photosynthesis isreduced by the action of photosystem II and oxidized bythat of photosystem I Therefore redox control of chloro-plast gene expression provides a compensatory response(Pfannschmidt et al 1999a) and adds a previously mis-sing part of an explanation for the ability of plants andalgae to adjust the ratio of the two photosystems inresponse to changes in light quality (Chow et al 1990)By the same argument a functionally equivalent post-translational mechanism of redox control of light-harvesting function by photosystems I and II may havebeen conserved during the transition from prokaryotes toeukaryotes (Allen 1992 1995 Allen amp Nilsson 1997Allen amp Race 2002)
The precedent of chloroplast redox control of transcrip-tion (Pfannschmidt et al 1999ab) taken together with theprinciple in sect 3d predicts plastoquinone redox control ofcyanobacterial reaction centre gene transcription and thisis borne out by experiment (El Bissati amp Kirilovsky2001) This complementary self-adjusting transcriptionalresponse (Fujita 1997 Murakami et al 1997 El Bissati ampKirilovsky 2001) indicates strongly that redox control ofphotosynthetic reaction centre gene transcription waspresent in the proto-chloroplast Thus the mechanism ofregulation is likely to have been conserved throughout theevolutionary transition from bacterial symbiont to cyto-plasmic organelle A conserved mechanism of redox regu-lation of transcription in photosynthetic prokaryotes(Unden amp Bongaerts 1997 Bauer et al 1999) and chloro-plasts (Allen 1993c Pfannschmidt et al 1999b) is pre-dicted if evolutionary continuity of the process has beenmaintained
Thus apart from being consistent with experimentaldata the principle of continuity of redox control has beenof heuristic value in pointing to the underlying bacterialcharacter previously unsuspected in the control of geneexpression especially of transcription in chloroplasts ofhigher plants Other possible precedents for redox signal-ling components of cytoplasmic organelles include the prr(Eraso amp Kaplan 2000 2002 Oh amp Kaplan 2000) andreg (Bauer et al 1999) redox regulatory systems of purplenon-sulphur bacteria The TspO lsquooxygen sensorrsquo of Rho-dobacter sphaeroides is present in mammalian mitochondriaas a protein known there as a benzodiazepine receptorand each protein will complement loss of the other(Yeliseev amp Kaplan 2000) As outlined in sect 3j genes forsuch components are to be expected primarily in nucleargenomes This independent evolutionary hypothesis (sect 3d)would be strengthened still further by a demonstration ofone or more decisive redox regulatory links shared by pho-tosynthetic prokaryotes and all eukaryotes with mitochon-
Phil Trans R Soc Lond B (2003)
dria and where present with chloroplasts (Bauer et al2003)
(e) The principle of the selective value of redoxcontrol
For each gene under redox control (sect 3d) it is selectivelyadvantageous for that gene to be retained and expressed onlywithin the organelle
The twin selective advantages of redox control are likelyto be energetic efficiency and the suppression of the harm-ful side effects of electron transport operating on thewrong substrates No direct experimental support for thisprinciple is available and equally there is no evidenceagainst it Allotopic mutants in which genes have beenexperimentally relocated from organelles to the nucleushave been produced in yeast higher plants (Kanevski ampMaliga 1994) and human cells (Manfredi et al 2002) andare predicted by CORR to be poor adapters to environ-mental change Some authors believe there is potentialtherapeutic value in allotopic transformation movingmitochondrial genomes to the nucleus (Kanevski ampMaliga 1994 de Grey 2000) and predict the possibilityof r2 human cell lines in the foreseeable future (de Grey2000) This principle also has implications for ageing andthe evolution of separate sexes (Allen 1996) and possibleconsequences for development (Blackstone 1999 20002001 Harvey et al 2002) and somatic cloning in animals(Allen amp Allen 1999)
(f ) The principle of the selective value of nuclearlocation for genes not under redox control
For each bacterial gene that survives and is not under redoxcontrol it is selectively advantageous for that gene to be locatedin the nucleus and expressed only in the nucleus and cytosolIf the mature gene product functions in chloroplasts or mito-chondria the gene is first expressed in the form of a precursorfor import
There are several possible reasons for the selectiveadvantage of nuclear location of genes for organellar pro-teins One is the decreased probability of mutation arisingfrom free radical by-products of lsquoincorrectrsquo electron trans-fers (Raven et al 1994ab Allen amp Raven 1996) Therewould seem to be obvious economies in resource use andadvantages in coordination of gene expression if genes arekept in one compartment One might say that the bipartiteor tripartite genetic system of eukaryotic cells is untidyand inefficient
Another suggestion is that organellar genes do notundergo recombination and are present in relatively smallcopy numbers so that disadvantageous mutations willspread relatively quickly through a clonal population oforganelles Incorporation of functional gene copiesinstead into the nuclear genome may then be an irrevers-ible step analogous to the click of a ratchet (Saccone et al2000) There are examples of genes shared between allthree compartments (Stern amp Lonsdale 1982 Lonsdale etal 1983)
The cost of retaining an organellar genetic system is notincreased appreciably when additional genes are retainedin organelles if just one gene is retained then the cost ofa cytoplasmic genetic system is incurred and there may berelatively little extra cost for each additional gene retained(Allen amp Raven 1996) For a functional explanation of the
26 J F Allen Genomes in mitochondria and chloroplasts
existence of chloroplast and mitochondrial genomes thefocus of attention should therefore be on the relativelyconstant primary subset of bioenergetic genes (sect 3e)
(g) The principle of contemporary operation ofselection on gene location
For any species the distribution of genes between organelle(by the principle in sect 3e) and nucleus (by the principle in sect 3f)is the result of selective forces that continue to operate
This principle is essentially a statement contradictingboth the lsquofrozen accidentrsquo hypothesis and the hypothesisthat gene relocation is progressive predictable and stillincomplete (sect 4) In support of this principle it is becom-ing increasingly clear that mitochondria that lose theirfunction in aerobic respiration also lose their genomes asseen in the relict mitochondria of microsporidia(Embley amp Martin 1998 Van der Giezen et al 2002) andthe lsquomitosomersquo of Entamoeba histolytica (Tovar et al1999) In chloroplasts loss of photosynthesis results inloss of photosynthetic genes The examples of Epifagus (aparasitic higher plant) (Ems et al 1995 Wolfe et al 1992)and the residual apicomplexan plastids of Plasmodium(Wilson et al 1996a) and Toxoplasma where some plas-tidic genetic system is retained mean that some of theirgene products must be subject to redox regulation of geneexpression if this principle is correct Experimental test ofthis principle is possible using allotopic mutants as forthe predictions of the principle in sect 3f The evidence thatthe apicoplast (Wilson et al 1996a) is a plastid is largelygenetic the residual genome resembles that of photosyn-thetic plastids (chloroplasts) in gene organization ratherthan in content A plastid that has completely lost its gen-ome by virtue of having lost photosynthesis and all otherprocesses requiring redox regulation might be difficult todistinguish from an organelle derived in a similar way froma mitochondrion The persistence of targeted importedchaperones Hsp70 (Tovar et al 1999 Williams et al2002) and cp60 (Roger et al 1998) in residual organellesmight not distinguish a plastid ancestry from a mitochon-drial one
(h) The principle of primary involvement inenergy transduction
Those genes for which redox control is always vital to cellfunction have gene products involved in or closely connectedwith primary electron transfer These genes are always con-tained within the organelle
This principle would be violated and the CORRhypothesis refuted if any photosynthetic reaction centrecore subunit or respiratory chain subunit currentlyorganelle-encoded is found that functions in primary vec-torial electron transport and is nevertheless encoded in thenucleus and synthesized cytosolically as a precursor forimport all without any selective cost to the individualFrom the chloroplast transcriptional experiments ofPfannschmidt et al (1999ab) the chloroplast geneswhose transcription is regulated by the plastoquinoneredox state include psaAB (for photosystem I reactioncentre core polypeptides) psbA (for the photosystem IIpolypeptide D1) and rbcL (for the large subunit of RUB-ISCO interestingly following the same pattern of redoxcontrolmdashupregulated by oxidized plastoquinonemdashaspsbA) Transcription of the chloroplast genes petA
Phil Trans R Soc Lond B (2003)
(cytochrome f ) trnG (glycine tRNA) and rpoB (RNApolymerase) did not respond to plastoquinone redox statepetA for cytochrome f is predicted by this principle to beregulated at a different level of redox control or if at plas-toquinone at a different stage in gene expression Thisprediction might also apply to trnG and rpoB though thesequalify as members of the secondary subset that of geneticsystem genes to which the principle in sect 3i applies
(i) The principle of secondary involvement inenergy transduction
Genes whose products contribute to the organelle genetic sys-tem itself or whose products are associated with secondaryevents in energy transduction may be contained in theorganelle in one group of organisms but not in anotherdepending on the physiology and biochemistry of photosynthesisand respiration in the species concerned
Even phylogenetically moderately related species(within kingdoms) show differences in the location ofsome genes for example the ATP synthase subunits ofNeurospora and Saccharomyces (Attardi amp Schatz 1988)The CORR hypothesis also does not explicitly account forthe wide disparities in the distribution of genetic systemgenes for example the almost complete import of tRNAsin Chlamydomonas (GoldschmidtClermont 1998) Trans-port of nucleic acids across membranes is uncommon andwhy some RNAs should be organelle-encoded in somespecies but nuclearly encoded in others is not currentlyunderstood The only suggestion that can be made alongthe lines of CORR is that in some genetic systems genesmay need to be under the control of a regulatory systemthat responds to a signal arising within the organelle itselfThis signal could be the primary signal of altered redoxstate or some secondary signal Among the latter might befound signals that themselves report on the current stateof gene expression another piece of information that thenucleus might not have at its immediate disposal Suchsecondary signals could result at least in part fromCORR though other developmental and environmentalinputs may be imagined and would not detract fromCORR unless it could be shown that they provided a suf-ficient reason for the retention of the genetic system asa whole
(j) The principle of the nuclear encoding of redoxsignalling components
Components of the redox-signalling pathways upon whichCORR depends are themselves not involved in primary electrontransfer and so their genes have been relocated to the nucleusin accordance with sect 3f
This principle states that the redox signalling pathwaysthat are required for CORR should not themselves beexpected to utilize components whose biosynthesis mustbe under direct redox control Such components shouldtherefore fall into the major category of organellar pro-teins and be imported as precursors from the cytosol inline with the principle in sect 3f However the principleabove might be need to be modified in cases where ampli-fication of the redox signal is useful to the cell In thebacterial ntr system which was a precedent for CORR(Harrison et al 1990 Allen 1992) the histidine kinasentrB and aspartate response regulator ntrC form part ofthe gln operon whose transcription they control Odontella
Genomes in mitochondria and chloroplasts J F Allen 27
and Porphyra have plastids that encode more proteins thanthose of most chlorophytes (Stoebe et al 1998 Turmel etal 1999) and these proteins include two-component sig-nal transduction proteins prr and ycf27 (Stoebe et al1998) Extreme or extremely rapid environmental fluctu-ations might be catered for by amplification of signals thatare transmitted by signalling pathways whose componentsare under their own control It is not known whether ycf27and prr are in operons whose promoters their gene pro-ducts control It is also uncertain whether ycf27 is a redoxsignalling component and its distribution at least in itsplastidic location seems to be correlated with the presenceof a phycobilin-based light-harvesting system as found inred algae and cyanobacteria (Ashby amp Mullineaux1999ab Ashby et al 2002) The best guess of the originalstatement of the CORR hypothesis (Allen 1992 1993ac)was that bacterial two-component systems (Iuchi amp Lin1992 Allen 1993c Grebe amp Stock 1999) provide a plaus-ible means of exerting redox regulatory control over mito-chondrial and chloroplast gene expression andsubsequent research confirms the presence of two-compo-nent genes in genomes of Arabidopsis and yeast SeveralArabidopsis two-component genes are predicted to be tar-geted to mitochondria and chloroplasts (Forsberg et al2001) These must be considered as prime candidates forcomponents involved in redox regulation of chloroplasttranscription seen in the experiments of Pfannschmidt etal (1999a 2001a) The presumably homologous regulat-ory process serving to control photosystem stoichiometryin cyanobacteria the prr system (Li amp Sherman 2000) isnow characterized as a two-component system involvinga redox sensor and redox response regulator as originallydefined (Allen 1992 1993ac)
4 CONTRASTING HYPOTHESES CONCERNINGTHE RETENTION OF CYTOPLASMIC GENOMES
CORR is a recent addition to explanations that havebeen put forward to account for the persistence of gen-omes in organelles These models and their underlyingmechanisms are now discussed and compared
(a) There is no reason that is just how it isOne reaction to the problem of cytoplasmic genetic sys-
tems (sect 2) is that there is no problem and no particularreason for their retention However there is a very widephylogenetic distribution of a common core or primarysubset of organellar-encoded genes and one which haswithstood considerable genomic reorganization over evol-utionary time This constancy is seen for example in thesame genes being present in the chloroplasts of landplants with multiple copies of a single circular chloroplastchromosome and in dinoflagellate chloroplasts with acollection of different DNA minicircles each with essen-tially one gene (Zhang et al 1999 2001 Barbrook et al2001) Another suggested explanation of the retention ofmitochondrial and chloroplast genomes is simply torestate the endosymbiont hypothesis and to point out thatchloroplast and mitochondrial genes lsquocame inrsquo with thebacterial endosymbiont This suggestion however doesnot address the question of why some genes were retainedwhereas others were not It is also now clear that mostgenes that came in and still survive have nuclear and cyto-
Phil Trans R Soc Lond B (2003)
solic gene products whereas many chloroplast and mito-chondria proteins had a eukaryotic origin (Salzberg et al2001 Martin amp Russell 2003) The odds against a con-served pattern in gene location (eg table 1) arising bychance must be astronomical Thus there is a reason forthe retention of organellar genomes and a reason for theretention of the same primary subset of genes Becauseevolution proceeds by natural selection of chance vari-ation lsquoreasonrsquo in this context requires identification of thefunction of the location of a gene that is the selectivevalue of the phenotype produced by that location Evol-ution sometimes retains apparently functionless relics ofpreviously useful features but not for long especially ifthere is a cost
(b) The lock-in hypothesisThe lsquolock-inrsquo hypothesis of Bogorad (1975) was pro-
posed independently of the endosymbiont hypothesis anddoes not depend upon it According to the lock-in hypoth-esis core components of multi-subunit complexes must besynthesized de novo in the correct cellular compartmentotherwise photosystems or respiratory chain complexesfor example might assemble in the plasma membranevacuolar membrane or endoplasmic reticulum The lock-in hypothesis made no clear prediction about the conti-nuity of cytoplasmic genetic systems even though cyto-plasmic uniparental non-Mendelian inheritance for bothmitochondria (Roodyn amp Wilkie 1968) and plastids(Kirk amp Tilney-Bassett 1978) made it likely that organellarDNA the chondriome and the plastome replicated inde-pendently of nuclear DNA (Whitehouse 1969)
Mechanisms of protein import and targeting (Schatz1998 Koehler et al 1999 Leuenberger et al 1999 Gab-riel et al 2001 Jarvis amp Soll 2001 Pfanner amp Geissler2001 Duby amp Boutry 2002 Pfanner amp Wiedemann 2002)now effectively answer the problem of specifying the cor-rect locations for membrane protein complexes for whichthe lock-in hypothesis was proposed as a solution How-ever regulation of synthesis of an entire functional multi-subunit complex by control of synthesis of individual sub-units is still an important principle This seems to be afeature of the lsquoCESrsquo model (control by epistasy ofsynthesis) of Wollman (Wollman et al 1999 Eberhard etal 2002) which envisages nuclear control as all-embracingwith organellar genetic systems doing as it were whatthey are told In the hypothesis of CORR propoundedhere CES is equally important but the essence of CORRis that the organellar genetic system alone knows what arethe immediate and most important signals of environmen-tal change The organelle may keep the nucleus informedbut this is a lower priority than initiating an appropriateresponse CES and CORR agree that assembly of multi-subunit complexes is a complex business and requirescoordinated control in which no step is rate-limiting butmany may be obligatory
(c) Going going gone the evolutionary process oftransfer of genes from organelles to the
nucleus is still underway but incompleteThis hypothesis envisages that for most eukaryotes we
are now witnessing the last stages in a slow evolutionarytransfer of genes to the nucleus and this transfer may beexpected to proceed to completion in what is in evol-
28 J F Allen Genomes in mitochondria and chloroplasts
utionary terms the near future (Palmer 1997 Gray 19992000 Gray et al 1999 Adams et al 2000 Daley et al2002) A clear and compelling statement of this widelyheld position is that of Herrmann amp Westhoff (2001 p10)
hellipthe transfer of geneshelliptells that the partite plant gen-ome is not in a phylogenetic equilibrium All availabledata suggest that the ultimate aim of genome restructur-ing in the plant cell as in the eukaryotic cell in generalis the elimination of genome compartmentation whileretaining physiological compartmentation
One serious problem with this hypothesis is the dis-tinctly non-random sample of genes that remain to betransferred A relatively constant subset of genes is foundin bioenergetically functional mitochondria from fungi tomammals and angiosperms as in photosynthetic plastids(chloroplasts) from unicellular algae to angiosperms Thishypothesis therefore requires an additional ad hocassumption that some gene transfers are inherently moreprobable than others (this too is widely supposed seelater) and thus that gene transfer events proceed in thesame sequence in quite independent lineages Thereseems to be no evidence for this and the order of geneloss from mitochondria and chloroplasts has been used toinfer phylogeny (Martin et al 1998 Adams et al 2000Millen et al 2001) lsquoGoing going gonersquo (Palmer 1997)is difficult to refute categorically but it does not seem tobe consistent with recent evidence For example chromo-some two of Arabidopsis thaliana (Lin et al 1999) has asingle insertion encompassing with about 99 sequencesimilarity the complete Arabidopsis mitochondrial genome(Unseld et al 1997 Marienfeld et al 1999 Stupar et al2001) Unless this single transfer is a freak occurrencethis finding strongly indicates that gene transfer occurswith relative ease (Adams et al 2000 Henze amp Martin2001 Rujan amp Martin 2001 Salzberg et al 2001) Inaddition there is experimental evidence that randomlygenerated polypeptides are mitochondrially imported athigh frequency (Lemire et al 1989) The hypothesis thatwe now see the same few stragglers in diverse organellargenomes is not impossible but should be regarded as alast resort perhaps applicable only in extreme cases suchas the greatly reduced apicomplexan plastomes and chon-driomes Even there the hypothesis carries with it an obli-gation to explain why organisms with few organellar geneshave lsquoevolvedrsquo faster than those with many
(d ) The frozen accidentWhat may be described as lsquothe frozen accident hypoth-
esisrsquo states that the evolutionary process of gene transferfrom organelle to nucleus was underway when somethinghappened that stopped it One example of the frozen acci-dent is a scenario (Von Heijne 1986) in which import intomitochondria of the precursor proteins arising from newlytranslocated genes proceeded for long enough after theprimordial endosymbiosis for most of the symbiont-derived genes to be moved to the cell nucleus The lsquoacci-dentrsquo that halted the process was the evolutionary originof exocytosis and protein secretion after which it becameimpossible for precursors properly destined for mitochon-drial import to be distinguished from those destined forexport from the cell As with the lsquolock-inrsquo hypothesis the
Phil Trans R Soc Lond B (2003)
lsquofrozen accidentrsquo seems incompatible with the remarkableprecision and specificity of protein targeting (Schatz 1998Koehler et al 1999 Leuenberger et al 1999 Gabriel etal 2001 Jarvis amp Soll 2001 Pfanner amp Geissler 2001Duby amp Boutry 2002 Pfanner amp Wiedemann 2002) Pro-tein sorting mechanisms do not seem to forbid controlledimport into any cellular compartment Another lsquoaccidentrsquothat may have impeded subsequent gene transfer in thecase of mitochondria is their departure from what is other-wise a universal genetic code Genetic code differences arediscussed later in the context of whether some genes can-not be moved
(e) Some proteins (with cofactors) cannot beimported
We may describe as the lsquounimportability hypothesisrsquo theproposal that some proteins are inherently unimportableand must therefore be synthesized de novo in the correctcompartment within the cell The unimportability hypoth-esis is most often proposed as its special case the hydro-phobicity hypothesis with hydrophobicity being thereason for unimportability
(i) HydrophobicityA commonly entertained notion is that proteins that are
encoded and synthesized within organelles comprise a pri-mary subset characterized by shared hydrophobicity allare intrinsic membrane proteins (Von Heijne 1986Popot amp de Vitry 1990 Claros et al 1995) As with theCORR hypothesis favoured here the primary subset thenrequires a secondary subset of genetic system componentsthe latter not themselves being hydrophobic but beingrequired for synthesis of the others Certainly the mito-chondrial inner membrane contains some intrinsic proteincomplexes whose subunits broadly satisfy the criterion ofhydrophobicity However the core subunits that are mito-chondrially encoded are also explained by the CORRhypothesis This would make hydrophobicity a require-ment for their function in vectorial electron and protontransfer and not a reason in itself for retention of theirgenes in the cytoplasm
The hydrophobicity hypothesis has some very clearcounter-examples in chloroplasts and cannot thereforestand if one wishes for a single principle that embracesboth types of bioenergetic organelle One clear counter-example is the large (L) subunit of the enzyme RUBISCOan abundant but entirely membrane-extrinsic protein ofca 55 kDa molecular mass The RUBISCO large subunitis generally chloroplast-encoded with the exception of thenuclearly encoded bacterial-type L subunit of L2 RUBI-SCO in peridinin-containing dinoflagellates (Morse et al1995) In most photosynthetic eukaryotes RUBISCO hasa chloroplast encoded large subunit and a nuclearly enco-ded small (S) subunit (Ellis 1984 Gatenby amp Ellis 1990)giving an L8S8 holoenzyme and this RUBISCO is widelytaken as a paradigm for the need for coordination ofnuclear and chloroplast gene expression (Rodermel 2001)
By complete contrast to the RUBISCO large subunitwhich is a chloroplast-encoded and water-soluble proteina second major violation of the hydrophobicity hypothesisis the existence of the universally nuclearly encoded butalways hydrophobic subunits of the chloroplast LHC IIand LHC I (Chitnis amp Thornber 1988) These apopro-
Genomes in mitochondria and chloroplasts J F Allen 29
teins of ca 27ndash30 kDa are quite water-insoluble and areusually fully extracted only in chloroformndashmethanol or inthe presence of detergents The Lhc gene family is diverse(Green amp Kuhlbrandt 1995) but most gene productsor Lhcp (light-harvesting chlorophyll protein) polypeptideshave three membrane-spanning hydrophobic a-helices(Kuhlbrandt 1994) It is interesting that three-helix Lhcproteins seem to be absent from extant cyanobacteria andchloroxybacteria (Green amp Kuhlbrandt 1995)
(ii) The five-helix rulePerhaps with the three-helix model of LHC II in mind
a variation of the hydrophobicity hypothesis is the empiri-cal lsquofive-helix rulersquo (Popot amp de Vitry 1990 Claros et al1995 Wollman et al 1999) which states that no intrinsicmembrane protein with five or more membrane-spanninghelices can be encoded in the nucleus Five is a good guessfor a threshold number of helices demanding organellar-encoding in the context of photosynthetic reaction centres(Heathcote et al 2002) where the earliest known struc-ture that of type II centres (Deisenhofer et al 1985) asin photosystem II (Zouni et al 2001) shows exactly fivehelices with type I centres containing 11 (Jordan et al2001) There are however many nuclearly encoded sub-units in both mitochondria and chloroplasts that havefewer than five membrane helices so the five-helix rulecan only describe a sufficient condition and not a neces-sary condition for nuclear encoding
It should also be noted that ADPndashATP carriers (AACs)are encoded in the nucleus AACs of the mitochondrialinner membrane have six transmembrane helices (van derGiezen et al 2002) while the those of the chloroplast innerenvelope (Winkler amp Neuhaus 1999) have 12 (Tjaden etal 1998)
With the counter-examples of nuclearly encoded AACswith six or 12 membrane-spanning helices many intrinsicmembrane proteins with fewer than five helices somenuclearly encoded and some not and with the RUBISCOlarge subunit as a major product of chloroplast geneexpression with no membrane domain at all the five-helixrule begins to seem rather like a case of special pleadingEven if there are indeed special mitigating circumstancesfor AACs (this argument is outlined later) the five-helixrule as with the hydrophobicity hypothesis of which it isa special case will disappear if it can be shown that pos-session of five or more membrane helices is required forsome other function such as that of CORR that deter-mines the organellar location of the gene concernedHydrophobicity is in some cases a feature of proteins thatare encoded in organelles but it is not in itself the issue
For chloroplasts the hydrophobicity hypothesis predictsretention of cyanobacterial genes in functional categoriesassociated with the chloroplast envelope and with trans-port just as much as with photosynthesis Table 1 showsthat this is not the case
A mechanism for the unimportability of hydrophobicproteins is rarely stated explicitly and it is rather left tothe imagination whether hydrophobic proteins might asit were get stuck halfway through the chloroplast envelopeor mitochondrial outer membrane whether transport can-not get started on a precursor protein that has alreadyspontaneously inserted into the membrane or whetherhydrophobic proteins will be unable to make the journey
Phil Trans R Soc Lond B (2003)
through the cytosol without precipitating there In thecytosol co-translational transport or membrane insertionwould in principle address the latter point but it seemsto be generally true that chloroplast and mitochondrialproteins are post-translationally inserted into the organel-lar outer membrane So the obvious ancillary questionabout unimportability is lsquowhy then cannot hydrophobicproteins be co-translationally imported into organellesrsquoThere is evidence in yeast for co-translational transportinto mitochondria of proteins thought to be derived fromthe original endosymbiont (Marc et al 2002) If there isany truth in hydrophobicity as a barrier to protein importthe CORR principle of unselective import (sect 3c) shouldbe easily refuted for example by a demonstration that noknown targeting presequence is capable of importing poly-phenylalanine encoded by polyU for in vitro import stud-ies
An additional proposal intended to account for theunimportability of some hydrophobic proteins and forimportability of others is suggested by A de Grey(personal communication) who argues that bacteriallyderived protein precursors imported into mitochondriacross the mitochondrial inner membrane completelybefore insertion into that membrane from the mitochon-drial matrix (the N-phase) This is held to be an obstacleto successful import of membrane-intrinsic proteins thatmight en route discharge the proton motive force By con-trast eukaryotically derived proteins can be inserted intothe mitochondrial inner membrane from the inter-mem-brane space (the P-phase) and then there is for somereason no similar uncoupling effect Thus adenylatetransporters or AACs can be nuclearly encoded whereascytochrome oxidase subunits for example cannot It isnot obvious why bacterially derived subunits such asthose of cytochrome oxidase did not acquire eukaryoticinsertion mechanisms Perhaps this acquisition is forbid-den by the need to conserve pre-existing routes ofassembly into multi-subunit complexes or perhaps theneed for insertion from the inside reflects a need to incor-porate a cofactor for example haem or ironndashsulphur clus-ters along the way
(iii) Unimportability of three dimensional structures andcofactors
Accumulating evidence on protein transport mech-anisms seems to make a specific obstacle to successfultransport of certain sorts of protein less and less plausibleespecially given the observation that precursors with N-terminal targeting sequences not mature functional poly-peptides themselves traverse membranes during biosynth-esis
Protein folding inside the living cell is invariably assistedby molecular chaperones (Gatenby amp Ellis 1990 Azem etal 1997 Komiya et al 1997 1990 Jackson-Constan et al2001) proteins that impede the formation of lsquoimproperrsquostructural interactions that will otherwise produce aggre-gation or other non-functional interactions Proteinimport mechanisms appear mostly to rely on specific mol-ecular chaperones for guided unfolding before membraneinsertion and refolding after translocation of the polypep-tide into its destined compartment (Gatenby amp Ellis 1990Azem et al 1997 Komiya et al 1997 Jackson-Constanet al 2001) Because polypeptides are mostly lsquothreadedrsquo
30 J F Allen Genomes in mitochondria and chloroplasts
through membranes no special constraint on transportshould be expected to arise from 3D structure or surfaceproperties of a protein
An exception to this unfolding rule may be providedby holoproteins containing cofactors that are transportedalong with the apoprotein presumably with a largely intact3D structure The lsquoTATrsquo (twin-arginine translocase) sys-tem (Bogsch et al 1998 Dalbey amp Robinson 1999 Berkset al 2000ab Robinson 2000 Robinson amp Bolhuis 2001Sargent et al 2002) works broadly along these linesthough its mechanism is unresolved TAT is relevant herebecause it adds transport of essentially complete holopro-teins to the cellrsquos repertoire of transport mechanisms andeven proteins that may not be allowed to unfold perhapsbecause of the toxicity of the free cofactor are not therebymade candidates for an unimportability rule One variantof the unimportability hypothesis is that transport canwork only by unfolding and dissociated cofactors may betoxic to the cell The bacterially derived TAT system(Bogsch et al 1998 Dalbey amp Robinson 1999 Berks etal 2000ab Sargent et al 2002) overcomes this argumentas would examples of cofactor-free polypeptides such as(C)Fo subunit c which are nevertheless generally encodedin organelles
The presence of a vesicular transport system in chloro-plasts (Westphal et al 2001) counts against the unim-portability hypothesis Vesicular transport between thechloroplast envelope and the thylakoid membrane shouldin principle allow any protein to be ferried across thechloroplast stroma Vesicles might allow transport inparticular of membrane proteins that would discharge theproton motive force if allowed to connect transiently theN and P aqueous phases on each side of a coupling mem-brane
Another observation that seems to count against unim-portability is the increased number of membrane barriersin plastids derived from secondary tertiary or higher-orderendosymbioses These plastids seem to have retained ifanything fewer genes rather than more as unimportabilitypredicts The nucleomorph the residual nucleus of theprimary host retains many genes for molecular chap-erones (Archibald et al 2001 Cavalier-Smith 2002) inagreement with the conclusion that protein import pro-ceeds by conserved and specialized mechanisms If eachnew import added to gain a higher-order endosymbiosisrequires additional targeting presequences then perhapsthere is an advantage in components of an import machin-ery not being imported themselves
It seems that unimportability is less easy an assumptionto make today than in previous years and the onus maynow be on its proponents to cite specific examples of pro-teins that can never be imported into chloroplasts or mito-chondria
(f ) Some genes cannot be moved(i) Differences in the mitochondrial genetic code
The genetic code is the specific relation between ribon-ucleotide triplets of codons and tRNAs that recognizethem and which thus determine which individual aminoacids if any will be incorporated into the growing poly-peptide chain Mitochondria show small departures fromthe otherwise universal genetic code (Attardi amp Schatz1988 Alberts et al 1994) UGA is a lsquostoprsquo codon in the
Phil Trans R Soc Lond B (2003)
universal code and in plant mitochondria but is the codonfor tryptophan in mitochondria of animals and fungiCUA means leucine everywhere except fungal mitochon-dria where it means threonine and AGA and AGU codefor arginine in the lsquouniversalrsquo code but serine in insectmitochondria and lsquostoprsquo in mammalian ones The signifi-cance of this heterogeneity in coding is most unlikely torest in the early origin of mitochondria as the lsquouniversalrsquocode is employed by bacteria and protists Furthermorethe diversity of departures from the lsquouniversalrsquo codeamong mitochondria of different eukaryotic lineages indi-cates they had independent origins A general rule formitochondrial genetic codes is that reliance on the thirdlsquowobblersquo position is decreased which allows all 20 proteinamino acids to be specified by fewer tRNAs (22 in animalmitochondria instead of 31 in the cytosol) This in turnmeans that mtDNA can specify more proteins for a givenlength of DNA or conversely that the mitochondrialgenome can be condensed or lsquostreamlinedrsquo Although thisarrangement does not explain the retention of mitochon-drial genomes and may be a secondary event in mitochon-drial evolution it almost certainly places an extra barrierin place of gene relocation As discussed previously how-ever there seems to be no single barrier that can explainthe retention of a relatively constant subset of mitochon-drial genes
There has clearly been a contraction of mitochondrialDNA especially in animals Animal mitochondrial DNAhas few if any non-coding sequences and animal mito-chondrial genetic codes require fewer tRNAs than the sin-gle code operating in the cytosol (Gray 1999 Gray et al1999 Selosse et al 2001) These may be devices todecrease the total frequency of mutations per mitochon-drial genome assuming a constant frequency per unitlength of mitochondrial DNA It has been proposed thata genersquos removal to the nucleus decreases its susceptibilityto free radical mutagenesis (Raven et al 1994ab Allen ampRaven 1996) If this is the case then the non-standardcode may be a further indication of an overriding need tomaintain certain genes within the mitochondrion What-ever the reason for the retention of genes in organelles ithad better be a good one bioenergetic organelles areclearly the wrong place to keep genetic information andgo to extraordinary lengths to offset the consequences ofit being there
(ii) Molecular refugeesA phenomenon related to differences in the genetic code
is seen in chloroplasts This is a bias in codon usage Thelsquomolecular refugeesrsquo hypothesis (Howe et al 2000) sug-gests that nuclear copies of what were once chloroplastgenes escape constraints on codon usage and thus thatwhat distinguishes chloroplast-encoded proteins is thattheir amino-acid composition is unusual enough for thebias in codon usage not to apply It is difficult to see whatphysico-chemical properties might be uniquely shared bythe amino acids of chloroplast-encoded proteins and inaddition there seem to be no lsquoforbiddenrsquo amino-acid resi-due in chloroplasts As with the departure from the univer-sal genetic code in mitochondria is seems likely that abias in codon usage reflects some secondary property ofchloroplast genomes and is not the primary reason fortheir retention In any case any restriction on gene move-
Genomes in mitochondria and chloroplasts J F Allen 31
ment will act in the wrong direction to explain why chloro-plasts have retained genomes at all provided there is noresulting obstacle to a chloroplast protein being success-fully encoded as a precursor in the nucleus
(iii) Genes are copied and replicated not relocated in onestep
If complete relocation is envisaged as proceeding by thesimultaneous appearance of a gene in one compartmentand its deletion from another then it is possible to imaginethat whatever the vector is there may be selectivity ingene transfer (Henze amp Martin 2001) For example ifviral incorporation replication and recombination in bothorganelle and nucleus were the mechanism of relocationthen successful gene transfer might depend on the abilityof the virus to complete the process in one step This inturn might depend on some property of the gene or ofthe protein it encodes if expression occurs at any stage inthe process of relocation An assumption of the CORRhypothesis favoured here is that genes are not movedphysically but merely copied after which either the copyin the original location or the copy in the new locationmay be lost as a result of selection A useful analogy maybe the action of copying files between computer storagevolumes In this analogy the copy is identical to the orig-inal and either may subsequently be deleted without lossof information
Because any gene that is not usefully expressed willundergo no selection to maintain a functional allele theprevailing location will be the one in which the functionalcopy of the gene resides The problem of which gene sur-vives is then subsumed into the problem of which is theselectively advantageous location for it It seems reason-able to assume that there wasmdashand ismdashan interim periodin which homologous genes in different compartments areessentially in competition to see which functions best Asoutlined above there is no obvious obstacle for copyingDNA sequences within the cell The physico-chemicalproperties of DNA are fairly uniform and not directlydependent on nucleotide sequence though organelles likeintra-cellular parasites seem to prefer A and T to G andC The answer to the question lsquowhich gene location func-tions bestrsquo may have nothing to do with the gene itselfnor directly with the structure and function of its geneproduct Instead the answer may have everything to dowith the required regulatory interaction between the geneproduct and expression of the gene that produces it
(g) CORRThis hypothesis has been propounded earlier (sect 3) and
will be summarized as follows Vectorial electron and pro-ton transfer exerts regulatory control over expression ofgenes encoding proteins directly involved in or affectingredox poise This regulatory coupling is indispensable foradaptation to environmental changes in chemical activitiesof oxygen light and CO2 and requires co-location of thesegenes with their gene products Organelles lsquomake theirown decisionsrsquo on the basis of environmental changesaffecting redox state The hypothesis is abbreviatedCORR for co-location for redox regulation or more com-pletely lsquoco-location (of gene and gene product) for(evolutionary) continuity of redox regulation of geneexpressionrsquo
Phil Trans R Soc Lond B (2003)
5 SUMMARY OF THE CURRENT STATUS OF THEHYPOTHESIS OF CORR
Q Why do mitochondria and chloroplasts have theirown genetic systems
A CORR co-location of genes and gene products per-mits direct and autonomous redox regulation of geneexpression
(a) Objections to CORR(i) There is no regulation of gene expression in chloroplasts
and mitochondriaThis is still commonly stated but incorrect See the prin-
ciple in sect 3d
(ii) There is no regulation of gene expression in chloroplastsand mitochondria at the level of transcription
For chloroplasts there is now clear evidence for a func-tionally intelligible redox control of transcription(Pfannschmidt et al 1999a) This invalidates the previouswidely held assumption that all control of chloroplast geneexpression is post-transcriptional (Gruissem amp Tonkyn1993 GoldschmidtClermont 1998) The evidence formitochondria is less clear but redox control of transcrip-tion certainly cannot be ruled out and there are indi-cations that it may occur (Pearson et al 1993Konstantinov et al 1995 Wilson et al 1996b) It mustalso be restated that transcriptional control is not neces-sary for CORR but serves as a compelling example of aconserved typically bacterial mode of response that oper-ates still in eukaryotic cytoplasmic organelles thus illus-trating the principle of continuity of redox control (sect 3d)Post-transcriptional redox regulation (Danon amp Mayfield1994 Kim amp Mayfield 1997 Brown et al 2001 Treb-itsh amp Danon 2001) is consistent with CORR and thereis nothing to forbid it being a primary mechanism thatsatisfies most or indeed all of the formal requirements ofthe hypothesis in particular cases
(iii) There are no redox signalling components encoded inchloroplast or mitochondrial DNA
This is what CORR predicts See the principle in sect 3j
(iv) There is redox control of nuclear gene expressionThis is correct and many examples are now known
especially for chloroplasts (Escoubas et al 1995 Kovacs etal 2000 Pfannschmidt et al 2001ab Surpin et al 2002)Redox signals from photosynthetic or respiratory electrontransport chains are not at all prohibited by CORR Theessence of CORR is that there is a subset of organellarproteins for which there is an overriding requirement fora redox regulatory control that is initiated within theorganelle and whose effects are exerted by the simplestand most direct route This redox signalling is envisagedto take place without a requirement for prior consultationand coordination with what may be regarded as a nucleo-cytoplasmic bureaucracy
(v) Nuclear genes can be expressed as rapidly as chloroplastand mitochondrial genes
Zerges (2002) for example cites the equal times takenfor complete expression of nuclear and chloroplast genesin Chlamydomonas as evidence against the CORR hypoth-
32 J F Allen Genomes in mitochondria and chloroplasts
esis However the question is not one of the time takento express a gene but the time taken to alter the rate ofits expression in response to a (redox) signal originatingwithin the organelle An equally important additional fac-tor may be the autonomy of the signalling pathway thatis its independence from other inputs as described in theprevious paragraph One might say that mitochondria andchloroplasts require a free hand
(vi) There is no evidence to support this hypothesisThe hypothesis is rich in predictions (sectsect 3 and 6) and
those that have been tested stringently are in agreementwith experimental results None of the predictions of thehypothesis has been found to be incorrect and none of theunderlying principles (sect 3) has been disproved Furtherpredictions will be the subject of future experiments
(b) Evidence for CORR(i) Chloroplasts have selectively retained cyanobacterial genes
for primary events in photosynthesisSee table 1
(ii) There is regulation of gene expression in chloroplastsThere is transcriptional regulation of gene expression
in chloroplasts (sect 3d) The case for mitochondria is notyet proven
(iii) There is redox regulation of gene expression inchloroplasts
There is transcriptional redox regulation of geneexpression in chloroplasts (sect 3d) The case for mitochon-dria is not yet proven though see (Allen et al 1995 Kon-stantinov et al 1995 Galvis et al 1998)
(iv) There are no redox signalling components encoded inchloroplast or mitochondrial DNA
Correct but with possible exceptions discussed in sect 3j
(v) There are lsquobacterialrsquo redox signalling components ineukaryotes
See sect 3j
(vi) Further predictions DNA-array results and phenotypesof allotopic mutants
A systematic test of CORR might be to make an oxid-ized-minus-reduced gene expression profile of the lsquotran-scriptomersquo of a cyanobacterium to see if the set of geneswhose transcription is under most rapid and direct redoxcontrol corresponds to the genes of the chloroplast gen-ome Laboratories with this experimental capability do notseem to have addressed this possibility (Hihara et al 2002Ikeuchi 2002) Likewise if a putative modern representa-tive of the proto-mitochondrion can be found and agreedeg Rhodospirillum rubrum or Paracoccus denitrificans thena similar experiment should reveal a subset of genes corre-sponding to mitochondrial genomes DNA-microarraysalso present the possibility of testing the relative rate ofredox response and dependency on other signallinginputs of transcription of eukaryotic genes To dateorganellar genes have not been included in such investi-gations (Ferea et al 1999) A further experimental possi-bility is to examine the redox regulatory phenotypes ofexisting allotopic mutants as outlined in sect 3e
Phil Trans R Soc Lond B (2003)
6 CONCLUSION AND FUTURE PROSPECTS
The CORR hypothesis states that the function of mito-chondrial and chloroplast genomes is to provide co-location (of gene and gene product) for (evolutionary)continuity of redox regulation of gene expression Thishypothesis is neither self-contradictory nor devoid of test-able predictions It is also consistent with available evi-dence and in principle makes sense of the otherwisepuzzling distribution of genes between compartments ineukaryotic cells The explanation that CORR offersapplies equally to mitochondria and chloroplasts andidentifies selective forces that still operate to determine thelocation of genes No selective barriers need be assumedto gene transfer protein import nor membrane insertionNor do we need to invoke a lsquofrozen accidentrsquo that some-thing happened to stop gene transfer for some reason atsome time in the remote and inaccessible past CORR alsoexplains the absence of genomes from organelles that havelost their primary bioenergetic functions CORR may actas a stimulus to a search for residual bioenergetic functionin those organelles that have lost aerobic respiration andphotosynthesis but where residual genomes remain
CORR also predicts regulatory properties of knowncomponents of chloroplasts and mitochondria that in turnindicate important but as yet uncharacterized flexibility inenergy transduction These components include the RUB-ISCO large subunit and the CFo and Fo subunits of coup-ling ATPase in both chloroplasts and mitochondriaRedox regulation of the relative stoichiometry of compo-nents as seen in regulation of chloroplast photosystemstoichiometry (Pfannschmidt et al 1999a) may extend to(C)Fo-(C)F1-ATPase indicating that the function ofretention of genes for one or more of these componentsis flexibility in the stoichiometries of H1 to e2 and H1 toATP in chemiosmotic coupling (Schemidt et al 1998Stock et al 2000 Allen 2002)
So far explicit tests of CORR have largely been con-fined to chloroplasts and to some extent mitochondria ofhigher plants but this merely reflects the expertise andinterests of the investigators Further attempted disproofof CORR is welcomed and encouraged Novel experi-mental tests can be devised as well as attempted refu-tation through analysis of existing data It is hoped thatexplicit attention will be given to the CORR hypothesis inlaboratories already suitably equipped and that resourcescan be allocated for this purpose in laboratories wherethere is the interest but not currently the technology orexpertise
The heuristic value of the hypothesis already seen inthe role of redox-regulated transcription of photosyntheticreaction centre genes in chloroplasts may be its most use-ful property The central proposal is that mitochondriaand chloroplasts contain genes for proteins whose functionin electron transfer demands rapid direct and uncon-ditional redox regulatory control of their biosynthesis Thefull implications of this hypothesis are very broad and onlyjust beginning to be explored
I thank Carol Allen Aubrey de Grey William Martin John ARaven and Francis-Andre Wollman for discussion and corre-spondence during the preparation of this paper and WilliamMartin for suggestions including table 1 Research grants from
Genomes in mitochondria and chloroplasts J F Allen 33
The Swedish Natural Sciences Research Council (NFR) andthe Crafoord Foundation are gratefully acknowledged
REFERENCES
Adams K L Daley D O Qiu Y L Whelan J amp PalmerJ D 2000 Repeated recent and diverse transfers of a mito-chondrial gene to the nucleus in flowering plants Nature408 354ndash357
Alberts B Bray D Lewis J Raff M Roberts K amp Wat-son J D 1994 The molecular biology of the cell New Yorkand London Garland Publishing
Allen C A Hakansson G amp Allen J F 1995 Redox con-ditions specify the proteins synthesized by isolated chloro-plasts and mitochondria Redox Rep 1 119ndash123
Allen J F 1992 Protein phosphorylation in regulation ofphotosynthesis Biochim Biophys Acta 1098 275ndash335
Allen J F 1993a Control of gene expression by redox poten-tial and the requirement for chloroplast and mitochondrialgenomes J Theor Biol 165 609ndash631
Allen J F 1993b Redox control of gene expression and thefunction of chloroplast genomes a hypothesis PhotosynthRes 36 95ndash102
Allen J F 1993c Redox control of transcription sensorsresponse regulators activators and repressors FEBS Lett332 203ndash207
Allen J F 1995 Thylakoid proteinndashphosphorylation state-1ndashstate-2 transitions and photosystem stoichiometry adjust-ment redox control at multiple levels of gene expressionPhysiol Plant 93 196ndash205
Allen J F 1996 Separate sexes and the mitochondrial theoryof ageing J Theor Biol 180 135ndash140
Allen J F 2002 Photosynthesis of ATP-electrons protonpumps rotors and poise Cell 110 273ndash276
Allen J F amp Allen C A 1999 A mitochondrial model forpremature ageing of somatically cloned mammals IUBMBLife 48 369ndash372
Allen J F amp Nilsson A 1997 Redox signalling and the struc-tural basis of regulation of photosynthesis by protein phos-phorylation Physiol Plant 100 863ndash868
Allen J F amp Pfannschmidt T 2000 Balancing the two photo-systems photosynthetic electron transfer governs transcrip-tion of reaction centre genes in chloroplasts Phil TransR Soc Lond B 355 1351ndash1357 (DOI 101098rstb20000697)
Allen J F amp Race H L 2002 Will the real LHC II kinaseplease step forward Sciencersquos STKE httpwwwstkeorgcgicontentfullsigtrans2002155pe43
Allen J F amp Raven J A 1996 Free-radical-induced mutationvs redox regulation costs and benefits of genes in organellesJ Mol Evol 42 482ndash492
Archibald J M Cavalier-Smith T Maier U amp Douglas S2001 Molecular chaperones encoded by a reduced nucleusthe cryptomonad nucleomorph J Mol Evol 52 490ndash501
Ashby M K amp Mullineaux C W 1999a Cyanobacterialycf27 gene products regulate energy transfer from phycobili-somes to photosystems I and II FEMS Microbiol Lett 181253ndash260
Ashby M K amp Mullineaux C W 1999b The role of ApcDand ApcF in energy transfer from phycobilisomes to PSI andPSII in a cyanobacterium Photosynth Res 61 169ndash179
Ashby M K Houmard J amp Mullineaux C W 2002 Theycf27 genes from cyanobacteria and eukaryotic algae distri-bution and implications for chloroplast evolution FEMSMicrobiol Lett 214 25ndash30
Attardi G amp Schatz G 1988 Biogenesis of mitochondria ARev Cell Biol 4 289ndash333
Azem A Oppliger W Lustig A Jeno P Feifel B SchatzG amp Horst M 1997 The mitochondrial hsp70 chaperone
Phil Trans R Soc Lond B (2003)
system effect of adenine nucleotides peptide substrate andmGrpE on the oligomeric state of mhsp70 J Biol Chem272 20 901ndash20 906
Barbrook A C Symington H Nisbet R E R LarkumA amp Howe C J 2001 Organisation and expression of theplastid genome of the dinoflagellate Amphidinium opercula-tum Mol Genet Genomics 266 632ndash638
Bauer C E Elsen S amp Bird T H 1999 Mechanisms forredox control of gene expression A Rev Microbiol 53495ndash523
Bauer C E Elsen S Swem L R Swem D L amp MasudaS 2003 Redox and light regulation of gene expression inphotosynthetic prokaryotes Phil Trans R Soc Lond B 358147ndash154 (DOI 101098rstb20021189)
Berks B C Sargent F De Leeuw E Hinsley A P Stan-ley N R Jack R L Buchanan G amp Palmer T 2000aA novel protein transport system involved in the biogenesisof bacterial electron transfer chains Biochim Biophys ActaBioenerg 1459 325ndash330
Berks B C Sargent F amp Palmer T 2000b The tat proteinexport pathway Mol Microbiol 35 260ndash274
Blackstone N W 1999 Redox control in development andevolution evidence from colonial hydroids J Exp Biol 2023541ndash3553
Blackstone N W 2000 Redox control and the evolution ofmulticellularity Bioessays 22 947ndash953
Blackstone N W 2001 Redox state reactive oxygen speciesand adaptive growth in colonial hydroids J Exp Biol 2041845ndash1853
Bogorad L 1975 Evolution of organelles and eukaryotic gen-omes Science 188 891ndash898
Bogsch E G Sargent F Stanley N R Berks B C Robin-son C amp Palmer T 1998 An essential component of a novelbacterial protein export system with homologues in plastidsand mitochondria J Biol Chem 273 18 003ndash18 006
Brown E C Somanchi A amp Mayfield S P 2001 Interor-ganellar crosstalk new perspectives on signaling from thechloroplast to the nucleus Genome Biol 2 10211ndash10214
Cavalier-Smith T 2002 Chloroplast evolution secondarysymbiogenesis and multiple losses Curr Biol 12 R62ndashR64
Chitnis P R amp Thornber J P 1988 The major light-har-vesting complex of photosystem 2 Aspects of its molecularand cell biology Photosynth Res 16 41ndash63
Chow W S Melis A amp Anderson J M 1990 Adjustmentsof photosystem stoichiometry in chloroplasts improve thequantum efficiency of photosynthesis Proc Natl Acad SciUSA 87 7502ndash7506
Claros M G Perea J Shu Y Samatey F A PopotJ L amp Jacq C 1995 Limitations to in vivo import of hydro-phobic proteins into yeast mitochondria The case of a cyto-plasmically synthesized apocytochrome b Eur J Biochem228 762ndash771
Dalbey R E amp Robinson C 1999 Protein translocation intoand across the bacterial plasma membrane and the plant thy-lakoid membrane Trends Biochem Sci 24 17ndash22
Daley D O Adams K L Clifton R Qualmann S MillarA H Palmer J D Pratje E amp Whelan J 2002 Genetransfer from mitochondrion to nucleus novel mechanismsfor gene activation from Cox2 Plant J 30 11ndash21
Danon A amp Mayfield S P 1994 Light-regulated translationof chloroplast messenger RNAs through redox potentialScience 266 1717ndash1719
de Grey A 2000 Mitochondrial gene therapy an arena for thebiomedical use of inteins Trends Biotechnol 18 394ndash399
Deisenhofer J Epp O Miki K Huber R amp Michel H1985 Structure of the protein subunits in the photosyntheticreaction center of Rhodopseudomonas viridis at 3Ac resolutionNature 318 618ndash624
34 J F Allen Genomes in mitochondria and chloroplasts
Delwiche C F 1999 Tracing the thread of plastid diversitythrough the tapestry of life Am Nat 154 S164ndashS177
Doolittle W F 1999 Phylogenetic classification and the uni-versal tree Science 284 2124ndash2129
Douglas S E 1998 Plastid evolution origins diversitytrends Curr Opin Genet Dev 8 655ndash661
Duby G amp Boutry M 2002 Mitochondrial protein importmachinery and targeting information Plant Sci 162 477ndash490
Eberhard S Drapier D amp Wollman F A 2002 Searchinglimiting steps in the expression of chloroplast encoded pro-teins relations between gene copy number transcriptiontranscript abundance and translation rate in the chloroplastof Chlamydomonas reinhardtii Plant J 31 149ndash160
El Bissati K amp Kirilovsky D 2001 Regulation of psbA andpsaE expression by light quality in Synechocystis species PCC6803 A redox control mechanism Plant Physiol 1251988ndash2000
Ellis R J 1984 The nuclear domination of chloroplast devel-opment Science Progress 69 129ndash142
Embley T M amp Martin W 1998 Molecular evolution ahydrogen-producing mitochondrion Nature 396 517ndash519
Ems S C Morden C W Dixon C K Wolfe K HdePamphilis C W amp Palmer J D 1995 Transcriptionsplicing and editing of plastid RNAs in the non-photosyn-thetic plant Epifagus virginiana Plant Mol Biol 29 721ndash733
Eraso J M amp Kaplan S 2000 From redox flow to gene regu-lation role of the PrrC protein of Rhodobacter sphaeroides241 Biochemistry 39 2052ndash2062
Eraso J M amp Kaplan S 2002 Redox flow as an instrumentof gene regulation Meth Enzymol 348 216ndash229
Escoubas J M Lomas M Laroche J amp Falkowski P G1995 Light-intensity regulation of cab gene-transcription issignaled by the redox state of the plastoquinone pool ProcNatl Acad Sci USA 92 10 237ndash10 241
Ferea T L Botstein D Brown P O amp Rosenzweig R F1999 Systematic changes in gene expression patterns follow-ing adaptive evolution in yeast Proc Natl Acad Sci USA96 9721ndash9726
Forsberg J Rosenquist M Fraysse L amp Allen J F 2001Redox signalling in chloroplasts and mitochondria genomicand biochemical evidence for two-component regulatory sys-tems in bioenergetic organelles Biochem Soc Trans 29403ndash407
Fujita Y 1997 A study on the dynamic features of photosys-tem stoichiometry accomplishments and problems forfuture studies Photosynth Res 53 83ndash93
Gabriel K Buchanan S K amp Lithgow T 2001 The alphaand the beta protein translocation across mitochondrial andplastid outer membranes Trends Biochem Sci 26 36ndash40
Galvis M L E Allen J F amp Hakansson G 1998 Proteinsynthesis by isolated pea mitochondria is dependent on theactivity of respiratory complex II Curr Genet 33 320ndash329
Gatenby A A amp Ellis R J 1990 Chaperone function theassembly of ribulose bisphosphate carboxylase-oxygenase ARev Cell Biol 6 125ndash149
GoldschmidtClermont M 1998 Coordination of nuclear andchloroplast gene expression in plant cells Int Rev CytolSurv Cell Biol 177 115ndash180
Gray M W 1992 The endosymbiont hypothesis revisited IntRev Cytol 141 233ndash357
Gray M W 1999 Evolution of organellar genomes CurrOpin Genet Dev 9 678ndash687
Gray M W 2000 Evolutionary biology mitochondrial geneson the move Nature 408 302ndash305
Gray M W amp Doolittle W F 1982 Has the endosymbionthypothesis been proven Microbiol Rev 46 1ndash42
Gray M W Burger G amp Lang B F 1999 Mitochondrialevolution Science 283 1476ndash1481
Phil Trans R Soc Lond B (2003)
Grebe T W amp Stock J B 1999 The histidine protein kinasesuperfamily Adv Microb Physiol 41 139ndash227
Green B R amp Kuhlbrandt W 1995 Sequence conservationof light-harvesting and stress-response proteins in relation tothe 3-dimensional molecular structure of Lhcii PhotosynthRes 44 139ndash148
Gruissem W amp Tonkyn J C 1993 Control mechanisms ofplastid gene expression Crit Rev Plant Sci 12 19ndash55
Hackstein J H P (and 11 others) 2001 Hydrogenosomesconvergent adaptations of mitochondria to anaerobicenvironments Zool Anal Complex Syst 104 290ndash302
Harrison M A Keen J N Findlay J B C amp Allen J F1990 Modification of a Glnb-like gene-product by photosyn-thetic electron-transport in the cyanobacterium Synechoc-occus 6301 FEBS Lett 264 25ndash28
Harvey A J Kind K L amp Thompson J G 2002 Redoxregulation of early embryo development Reproduction 123479ndash486
Heathcote P Fyfe P K amp Jones M R 2002 Reactioncentres the structure and evolution of biological solarpower Trends Biochem Sci 27 79ndash87
Henze K amp Martin W 2001 How do mitochondrial genesget into the nucleus Trends Genet 17 383ndash387
Herrmann R G amp Westhoff P 2001 Thylakoid biogenesis anddynamics the result of a complex phylogenetic puzzle InRegulation of photosynthesis vol 11 (ed E-M Aro amp BAndersson) pp 1ndash28 Dordrecht The Netherlands Kluwer
Hihara Y Sonoike K Kanehisa M amp Ikeuchi M 2002Search for redox responsive genes in Synechocystis sp PCC6803 by using DNA microarrays Plant Cell Physiol 43S178
Howe C J Barbrook A C amp Lockhart P J 2000 Organellegenes do they jump or are they pushed Trends Genet 1665ndash66
Ikeuchi M 2002 Gene expression and responses to light stressin cyanobacteria Plant Cell Physiol 43 S21
Iuchi S amp Lin E C C 1992 Mutational analysis of signaltransduction by Arcb a membrane sensor protein respon-sible for anaerobic repression of operons involved in the cen-tral aerobic pathways in Escherichia coli J Bacteriol 1743972ndash3980
Jackson-Constan D Akita M amp Keegstra K 2001 Molecu-lar chaperones involved in chloroplast protein importBiochim Biophys Acta Mol Cell Res 1541 102ndash113
Jarvis P amp Soll M 2001 Toc Tic and chloroplast proteinimport Biochim Biophys Acta Mol Cell Res 1541 64ndash79
Jordan P Fromme P Witt H T Klukas O SaengerW amp Krauss N 2001 Three-dimensional structure ofcyanobacterial photosystem I at 25 acngstrom resolutionNature 411 909ndash917
Kaneko T (and 23 others) 1996a Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regionsDNA Res 3 109ndash136
Kaneko T (and 23 others) 1996b Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regions[Supplement] DNA Res 3 185ndash209
Kanevski I amp Maliga P 1994 Relocation of the plastid rbcLgene to the nucleus yields functional ribulose-15-bisphosph-ate carboxylase in tobacco chloroplasts Proc Natl Acad SciUSA 91 1969ndash1973
Kim J amp Mayfield S P 1997 Protein disulfide isomerase as aregulator of chloroplast translational activation Science 2781954ndash1957
Genomes in mitochondria and chloroplasts J F Allen 35
Kirk J T O amp Tilney-Bassett R A E 1978 The plastidsTheir chemistry structure growth and inheritance Amster-dam Elsevier
Koehler C M Merchant S amp Schatz G 1999 How mem-brane proteins travel across the mitochondrial intermem-brane space Trends Biochem Sci 24 428ndash432
Komiya T Rospert S Schatz G amp Mihara K 1997 Bind-ing of mitochondrial precursor proteins to the cytoplasmicdomains of the import receptors Tom70 and Tom20 isdetermined by cytoplasmic chaperones FASEB J 11 1196
Konstantinov Y M Lutsenko G N amp Podsosonny V A1995 Genetic functions of isolated maize mitochondriaunder model changes of redox conditions Biochem MolBiol Int 36 319ndash326
Korab-Laskowska M Rioux P Brossard N LittlejohnT G Gray M W Lang B F amp Burger G 1998 Theorganelle genome database project (GOBASE) Nucleic AcidsRes 26 138ndash144
Kovacs L Wiessner W Kis M Nagy F Mende D ampDemeter S 2000 Short- and long-term redox regulation ofphotosynthetic light energy distribution and photosystemstoichiometry by acetate metabolism in the green alga Chla-mydobotrys stellata Photosynth Res 65 231ndash247
Kuhlbrandt W 1994 Structure and function of the plant light-harvesting complex LHC-II Curr Opin Struct Biol 4519ndash528
Lang B F Gray M W amp Burger G 1999 Mitochondrialgenome evolution and the origin of eukaryotes A RevGenet 33 351ndash397
Lemire B D Fankhauser C Baker A amp Schatz G 1989The mitochondrial targeting function of randomly generatedpeptide sequences correlates with predicted helical amphi-philicity J Biol Chem 264 20 206ndash20 215
Leuenberger D Bally N A Schatz G amp Koehler C M1999 Different import pathways through the mitochondrialintermembrane space for inner membrane proteins EMBOJ 18 4816ndash4822
Li H amp Sherman L A 2000 A redox-responsive regulator ofphotosynthesis gene expression in the cyanobacterium Syne-chocystis sp strain PCC 6803 J Bacteriol 182 4268ndash4277
Lin X Y (and 36 others) 1999 Sequence and analysis of chro-mosome 2 of the plant Arabidopsis thaliana Nature 402761ndash768
Lonsdale D M Hodge T P Howe C J amp Stern D B1983 Maize mitochondrial-DNA contains a sequence hom-ologous to the ribulose-15-bisphosphate carboxylase largesubunit gene of chloroplast DNA Cell 34 1007ndash1014
McFadden G I 1999 Endosymbiosis and evolution of theplant cell Curr Opin Plant Biol 2 513ndash519
McFadden G I 2001 Chloroplast origin and integrationPlant Physiol 125 50ndash53
Mahler H R amp Raff R A 1975 The evolutionary origin ofthe mitochondrion a nonsymbiotic model Int Rev Cytol43 1ndash124
Manfredi G Fu J Ojaimi J Sadlock J E Kwong J QGuy J amp Schon E A 2002 Rescue of a deficiency in ATPsynthesis by transfer of MTATP6 a mitochondrial DNA-encoded gene to the nucleus Nature Genet 30 394ndash399
Marc P Margeot A Devaux F Blugeon C Corral-Debrinski M amp Jacq C 2002 Genome-wide analysis ofmRNAs targeted to yeast mitochondria EMBO Rep 3159ndash164
Marienfeld J Unseld M amp Brennicke A 1999 The mito-chondrial genome of Arabidopsis is composed of both nativeand immigrant information Trends Plant Sci 4 495ndash502
Martin W 1999 A briefly argued case that mitochondria andplastids are descendants of endosymbionts but that thenuclear compartment is not Proc R Soc Lond B 2661387ndash1395 (DOI 101098rspb19990792)
Phil Trans R Soc Lond B (2003)
Martin W amp Herrmann R G 1998 Gene transfer fromorganelles to the nucleus how much what happens andwhy Plant Physiol 118 9ndash17
Martin W amp Muller M 1998 The hydrogen hypothesis forthe first eukaryote Nature 392 37ndash41
Martin W amp Russell M J 2003 On the origins of cells ahypothesis for the evolutionary transitions from abiotic geo-chemistry to chemoautotrophic prokaryotes and from pro-karyotes to nucleated cells Phil Trans R Soc Lond B 35859ndash85 (DOI 101098rstb20021183)
Martin W Stoebe B Goremykin V Hansmann S Hase-gawa M amp Kowallik K V 1998 Gene transfer to thenucleus and the evolution of chloroplasts Nature 393162ndash165
Martin W Hoffmeister M Rotte C amp Henze K 2001 Anoverview of endosymbiotic models for the origins of eukary-otes their ATP-producing organelles (mitochondria andhydrogenosomes) and their heterotrophic lifestyle BiolChem 382 1521ndash1539
Martin W Rujan T Richly E Hansen A Cornelsen SLins T Leister D Stoebe B Hasegawa M amp PennyD 2002 Evolutionary analysis of Arabidopsis cyanobacterialand chloroplast genomes reveals plastid phylogeny and thou-sands of cyanobacterial genes in the nucleus Proc NatlAcad Sci USA 99 12 246ndash12 251
Millen R S (and 12 others) 2001 Many parallel losses of infAfrom chloroplast DNA during angiosperm evolution withmultiple independent transfers to the nucleus Plant Cell 13645ndash658
Morse D Salois P Markovic P amp Hastings J W 1995 Anuclear-encoded form-II RUBISCO in dinoflagellatesScience 268 1622ndash1624
Murakami A Kim S J amp Fujita Y 1997 Changes in photo-system stoichiometry in response to environmental con-ditions for cell growth observed with the cyanophyteSynechocystis PCC 6714 Plant Cell Physiol 38392ndash397
Oh J I amp Kaplan S 2000 Redox signaling globalization ofgene expression EMBO J 19 4237ndash4247
Ozawa T 1995 Mitochondrial-DNA mutations associatedwith aging and degenerative diseases Exp Gerontol 30269ndash290
Palmer J D 1997 Organelle genomes going going goneScience 275 790ndash791
Pearson C K Wilson S B Schaffer R amp Ross A W 1993NAD turnover and utilization of metabolites for RNA syn-thesis in a reaction sensing the redox state of the cyto-chrome-b( 6 )f complex in isolated chloroplasts Eur JBiochem 218 397ndash404
Pfanner N amp Geissler A 2001 Versatility of the mitochon-drial protein import machinery Natl Rev Mol Cell Biol 2339ndash349
Pfanner N amp Wiedemann N 2002 Mitochondrial proteinimport two membranes three translocases Curr Opin CellBiol 14 400ndash411
Pfannschmidt T Nilsson A amp Allen J F 1999a Photosyn-thetic control of chloroplast gene expression Nature 397625ndash628
Pfannschmidt T Nilsson A Tullberg A Link G amp AllenJ F 1999b Direct transcriptional control of the chloroplastgenes psbA and psaAB adjusts photosynthesis to light energydistribution in plants IUBMB Life 48 271ndash276
Pfannschmidt T Allen J F amp Oelmuller R 2001a Prin-ciples of redox control in photosynthesis gene expressionPhysiol Plant 112 1ndash9
Pfannschmidt T Schutze K Brost M amp Oelmuller R2001b A novel mechanism of nuclear photosynthesis generegulation by redox signals from the chloroplast during pho-
36 J F Allen Genomes in mitochondria and chloroplasts
tosystem stoichiometry adjustment J Biol Chem 27636 125ndash36 130
Popot J L amp de Vitry C 1990 On the microassembly of inte-gral membrane proteins A Rev Biophys Biophys Chem 19369ndash403
Race H L Herrmann R G amp Martin W 1999 Why haveorganelles retained genomes Trends Genet 15 364ndash370
Raven J A Johnston A M Parsons R amp Kubler J 1994aThe influence of natural and experimental high O2 concen-trations on O2 evolving phototrophs Biol Rev CambridgePhil Soc 69 61ndash94
Raven J A Johnston A M Parsons R amp Kubler J 1994bThe occurrence and influence on photolithotrophs of highoxygen concentrations Proc R Soc Edinb B 102 193ndash201
Robinson C 2000 The twin-arginine translocation system anovel means of transporting folded proteins in chloroplastsand bacteria Biol Chem 381 89ndash93
Robinson C amp Bolhuis A 2001 Protein targeting by the twin-arginine translocation pathway Natl Rev Mol Cell Biol 2350ndash356
Rodermel S 2001 Pathways of plastid-to-nucleus signalingTrends Plant Sci 6 471ndash478
Roger A J 1999 Reconstructing early events in eukaryoticevolution Am Nat 154 S146ndashS163
Roger A J Svard S G Tovar J Clark C G SmithM W Gillin F D amp Sogin M L 1998 A mitochondrial-like chaperonin 60 gene in Giardia lamblia evidence thatdiplomonads once harbored an endosymbiont related to theprogenitor of mitochondria Proc Natl Acad Sci USA 95229ndash234
Roodyn D B amp Wilkie D 1968 The biogenesis of mitochondriaMethuenrsquos monographs on biological subjects London Methuen
Rujan T amp Martin W 2001 How many genes in Arabidopsiscome from cyanobacteria An estimate from 386 proteinphylogenies Trends Genet 17 113ndash120
Saccone C Gissi C Lanave C Larizza A Pesole G ampReyes A 2000 Evolution of the mitochondrial genetic sys-tem an overview Gene 261 153ndash159
Salzberg S L White O Peterson J amp Eisen J A 2001Microbial genes in the human genome lateral transfer orgene loss Science 292 1903ndash1906
Sargent F Berks B C amp Palmer T 2002 Assembly of mem-brane-bound respiratory complexes by the Tat protein-trans-port system Arch Microbiol 178 77ndash84
Schatz G 1998 Protein transport the doors to organellesNature 395 439ndash440
Schemidt R A Qu J Williams J R amp Brusilow W S1998 Effects of carbon source on expression of F0 genes andon the stoichiometry of the c subunit in the F1 F0 ATPaseof Escherichia coli J Bacteriol 180 3205ndash3208
Selosse M A Albert B R amp Godelle B 2001 Reducing thegenome size of organelles favours gene transfer to thenucleus Trends Ecol Evol 16 135ndash141
Stern D B amp Lonsdale D M 1982 Mitochondrial and chlo-roplast genomes of maize have a 12- kilobase DNA-sequencein common Nature 299 698ndash702
Stock D Gibbons C Arechaga I Leslie A G W ampWalker J E 2000 The rotary mechanism of ATP synthaseCurr Opin Struct Biol 10 672ndash679
Stoebe B amp Maier U G 2002 One two three naturersquos toolbox for building plastids Protoplasma 219 123ndash130
Stoebe B Martin W amp Kowallik K V 1998 Distributionand nomenclature of protein-coding genes in 12 sequencedchloroplast genomes Plant Mol Biol Rep 16 243ndash255
Stupar R M Lilly J W Town C D Cheng Z Kaul SBuell C R amp Jiang J M 2001 Complex mtDNA consti-tutes an approximate 620 kb insertion on Arabidopsis thali-ana chromosome 2 implication of potential sequencing
Phil Trans R Soc Lond B (2003)
errors caused by large-unit repeats Proc Natl Acad SciUSA 98 5099ndash5103
Surpin M Larkin R M amp Chory J 2002 Signal transduc-tion between the chloroplast and the nucleus Plant Cell 14S327ndashS338
Tjaden J Schwoppe C Mohlmann T Quick P W ampNeuhaus H E 1998 Expression of a plastidic ATPADPtransporter gene in Escherichia coli leads to a functional aden-ine nucleotide transport system in the bacterial cytoplasmicmembrane J Biol Chem 273 9630ndash9636
Tovar J Fischer A amp Clark C G 1999 The mitosome a novelorganelle related to mitochondria in the amitochondrial para-site Entamoeba histolytica Mol Microbiol 32 1013ndash1021
Trebitsh T amp Danon A 2001 Translation of chloroplastpsbA mRNA is regulated by signals initiated by both photos-ystems II and I Proc Natl Acad Sci USA 98 12 289ndash12 294
Tullberg A Alexciev K Pfannschmidt T amp Allen J F2000 Photosynthetic electron flow regulates transcription ofthe psaB gene in pea (Pisum sativum L) chloroplasts throughthe redox state of the plastoquinone pool Plant Cell Physiol41 1045ndash1054
Turmel M Lemieux C Burger G Lang B F Otis CPlante I amp Gray M W 1999 The complete mitochondrialDNA sequences of Nephroselmis olivacea and Pedinomonasminor two radically different evolutionary patterns withingreen algae Plant Cell 11 1717ndash1729
Unden G amp Bongaerts J 1997 Alternative respiratory path-ways of Escherichia coli energetics and transcriptional regu-lation in response to electron acceptors Biochim BiophysActa Bioenerg 1320 217ndash234
Unseld M Marienfeld J R Brandt P amp Brennicke A1997 The mitochondrial genome of Arabidopsis thaliana con-tains 57 genes in 366 924 nucleotides Nature Genet 1557ndash61
Van der Giezen M Slotboom D J Horner D S DyalP L Harding M Xue G P Embley T M amp KunjiE R S 2002 Conserved properties of hydrogenosomal andmitochondrial ADPATP carriers a common origin for bothorganelles EMBO J 21 572ndash579
Von Heijne G 1986 Why mitochondria need a genome FEBSLett 198 1ndash4
Westphal S Soll J amp Vothknecht U C 2001 A vesicle trans-port system inside chloroplasts FEBS Lett 506 257ndash261
Whitehouse H L K 1969 Towards an understanding of themechanism of heredity London Edward Arnold Ltd
Williams B A P Hirt R P Lucocq J M amp Embley T M2002 A mitochondrial remnant in the microsporidian Trachi-pleistophora hominis Nature 418 865ndash869
Wilson R J (and 10 others) 1996a Complete gene map of theplastid-like DNA of the malaria parasite Plasmodium falcipa-rum J Mol Biol 261 155ndash172
Wilson S B Davidson G S Thomson L M amp PearsonC K 1996b Redox control of RNA synthesis in potato mito-chondria Eur J Biochem 242 81ndash85
Winkler H H amp Neuhaus H E 1999 Non-mitochondrialATP transport Trends Biochem Sci 24 64ndash68
Wolfe K H Morden C W amp Palmer J D 1992 Small sin-gle-copy region of plastid DNA in the non-photosyntheticangiosperm Epifagus virginiana contains only two genes Dif-ferences among dicots monocots and bryophytes in geneorganization at a non-bioenergetic locus J Mol Biol 22395ndash104
Wollman F A Minai L amp Nechushtai R 1999 The biogen-esis and assembly of photosynthetic proteins in thylakoidmembranes Biochim Biophys Acta Bioenerg 1411 21ndash85
Yeliseev A A amp Kaplan S 2000 TspO of Rhodobactersphaeroides a structural and functional model for the mam-
Genomes in mitochondria and chloroplasts J F Allen 37
malian peripheral benzodiazepine receptor J Biol Chem275 5657ndash5667
Zerges W 2002 Does complexity constrain organelle evol-ution Trends Plant Sci 7 175ndash182
Zhang Z D Green B R amp Cavalier-Smith T 1999 Singlegene circles in dinoflagellate chloroplast genomes Nature400 155ndash159
Zhang Z D Cavalier-Smith T amp Green B R 2001 A fam-ily of selfish minicircular chromosomes with jumbled chloro-plast gene fragments from a dinoflagellate Mol Biol Evol18 1558ndash1565
Zouni A Witt H T Kern J Fromme P Krauss NSaenger W amp Orth P 2001 Crystal structure of photosys-tem II from Synechococcus elongatus at 38 acngstrom resol-ution Nature 409 739ndash743
DiscussionD S Horner (Dipartimento di Fisiologia e Biochimica
Generali University of Milan Milan Italy) Given thatthere are components of these membrane-bound energy-transduction pathways that are consistently nuclear enco-ded not the key ones you pointed out but others do youexpect perhaps a slower perhaps a more crude but yet aredox communication between the organelle andexpression of nuclear-encoded genes
J F Allen There is and there is no question about itI think that Bogoradrsquos lock-in hypothesis has somethingto say here and there are other people who say it is all todo with assembling multi-subunit complexes If you havecontrol over the initial construction of the core aroundwhich the whole complex is eventually built then redoxcontrol or some sort of control over synthesis of that corewill control construction of the whole thing eventuallyand I think that may well be largely correct I think it ispossible in principle for nuclear genes to make precursorsaccording to redox instructions but it is not really soimportant that those precursors are here and now presentin exactly the right quantities because the precursors canto a limited extent hang around and wait Actually thereis proteolytic degradation of unattached precursors and soon but I think the fact that these chloroplast and mito-chondrially DNA-encoded subunits are at the core ofthese energy-transducing complexes is the crucial issue
T Cavalier-Smith (Department of Zoology University ofOxford Oxford UK ) In the case of mitochondria the sub-set of genes that remains is much more variable than youimplied It runs from three proteins in the minimal casesup to I think over 90 That is a big difference and manyof the genes in different lineages of protists that remainhave nothing to do with redox control they are things likeribosomal proteins and so on So I think that even thoughwhat you say about redox control of certain genes mayactually be an important thing it is not the fundamentaloverall reason why the genomes are maintained It may bepart of the explanation for why certain genes are alwaysincluded in that subset but even in mitochondria I thinkthat cytochrome b is the only one that is always presentSo I think that both this great diversity of what has beenretained in different lineages and the nucleomorph caseindicate that evolution was able to do one thing in onelineage 500 million years ago and never got around todoing it for 500 million years in another lineage so thereis a lot more haphazardness and evolutionary accidentsand contingency than a straight functional interpretation
Phil Trans R Soc Lond B (2003)
there is a lot of just sheer lsquohappenedndashorndashdidnrsquotndashhappenrsquoor carryover
J F Allen I was not implying anything about the rangeof mitochondrial genomes I have a limited amount oftime so I just talked about what I could talk about withinthat limit Of course there will be a range and this willdepend upon the physiology and bioenergetics of theparticular organism But what I am trying to do is to seewhat the common properties of the irreducible core areof components that are always encoded in the organellesand cytochrome b is a perfect example because that reallyis the thing that binds these two all-important quinoneswithout which there is no Q cycle without which there isno proton pumping and bioenergetics and in Plasmodiumwhere the mitochondrion is reduced to this bare mini-mum there is still a mitochondrial gene for cytochromeb Now my argument about the ribosomal proteins andall the rest of it is that in order to be able to exert thisall-important redox control over gene expression of certaincore components you need an in situ genetic system andthis can vary in composition for reasons that I cannotdescribe Maybe you need RNA polymerase or maybeyou can import RNA polymerase In plastids you haveboth of these options You are going to need ribosomestransfer RNAs and the rest of the apparatus of geneexpression and that needs to be in situ but we are stillleft with the question why does there have to be any gen-etic system at all The mature tobacco or spinach chloro-plast seems to have an entire genetic system just to makeone protein D1 which is all it ever needs to resynthesizeI think that this is a clue It is worth having this very unec-onomical crazy system because there is an overridingrequirement to connect electron transfer with geneexpression directly and unambiguouslymdashwithout recourseto a bureaucracy if you like which is what I think thenucleus is
K Willison (Institute of Cancer Research Chester BeattyLaboratories London UK ) These photosynthetic reactioncomplexes are a sort of baroque mixture of polypeptidechains and cofactors and my understanding is that someof them in high photon flux can turn over within say 30minutes so I am wondering if there is a correlationbetween gene retention and the turnover rates of some ofthe components and the complexes it seems to me thatthat could really be a limiting factor and the reason thatthe proteins had to be encoded locally in order to get thecomplexes to be assembled rapidly
J F Allen I think that is right You can only alter thesteady-state concentration of a protein complex by affect-ing the synthesis and breakdown and therefore turnoverThere is an extreme example the D1 protein encoded bythe psbA gene of chloroplasts which is always in the chlor-oplast genome without exception High light will tend todegrade this protein and when the high light goes awaythere is an absolute need very quickly to bring it back upagain I think that is a very good example Whether thisargument applies to genes such as cytochrome oxidase Ido not know
C J Leaver (Department of Plant Sciences University ofOxford Oxford UK ) Your experiments tend to relatemainly to the gene expression side What about proteinRNA turnover the whole concept of redox control ofthatmdashdo we know anything
38 J F Allen Genomes in mitochondria and chloroplasts
J F Allen Well quite a lot is known about chloro-plasts There is redox control of messenger RNA stabilityfor this same D1 protein which has been described byStephen Mayfield and his co-workers The evidence is thatthe site of redox control is ferredoxin via thioredoxin andthe implication is that thioredoxin tells whether non-cyclicelectron transport is working at all When it is workingthere is a signal that says lsquoOK now continue with all theprotein synthesisrsquo It is not particularly specific but moreof a global effect I think this is what Mayfield argues butof course this is not inconsistent with my argument I haveconcentrated on quinone pools and medium-potentialredox regulatory control because that is where I can seesignals about the dangerous consequences of getting itwrong but there is control of other levels of geneexpression In mitochondria I am not so sure I wouldbounce the question back to you on that one
W Martin (Institute of Botany III Heinrich-Heine Univ-ersitat Dusseldorf Dusseldorf Germany) The nucleomorphgenome was cited as evidence in conflict with your modelbut is that true The nucleomorph genome is not in abioenergetic organelle in that sense It resides in the cyto-sol so does the size of the nucleomorph genome workagainst your model Is it really evidence against yourmodel
J F Allen I do not know why should it be evidenceI cannot explain why the nucleomorph has a genomeThat is why I clutched at the straw that it is storage fornucleic acid precursors
Phil Trans R Soc Lond B (2003)
T Cavalier-Smith The reason I introduced the nucleo-morph is not to say it is direct evidence against yourhypothesis but to show that there probably was no singlefunctional reason for the retention of every gene in everyorganelle It may be that there are specific physiologicalreasons for the retention of certain genes and thereforeyou might be right about a subset of genes but I verymuch doubt if there are specific physiological reasons forthe retention of every single gene that is retained differen-tially because odd things happen in evolution Somethings are easy and sometimes things are difficult Evol-ution is history
J F Allen Of course I agree with that I do not thinkI can explain everything but I do think I can explainsomething If I have done that then I am content
GLOSSARY
AAC ADPndashATP carrierCES control by epistasy of synthesisCORR co-location for redox regulationLHC light-harvesting complexPCC Pasteur culture collectionORF open reading frameRUBISCO ribulose-15-bisphosphate carboxylasendashoxygenaseTAT twin-arginine translocase
Visit httpwwwpubsroyalsocacuk to see an electronic appendix tothis paper

22 J F Allen Genomes in mitochondria and chloroplasts
strates that the bacterial precursor of mitochondria couldnot already obtain for itself Furthermore a second para-dox is resolved or at a least avoided export of ATP fromthe bacterial partner was not an immediate requirementfor the symbiosis to become established and could haveevolved subsequently Thus it is not necessary to postu-late a suicidal bacterium that excretes ATP nor a deferredselective advantage to the partnership implying foresightor an assurance of eventual contract fulfilment
The hydrogen hypothesis (Martin amp Muller 1998) seeksto provide an explanation of the origin of mitochondriathat accounts for the immediate benefit of the primordialsymbiosis to each partner but the implications of thehydrogen hypothesis for the origin of chloroplasts seemnot to have been considered in detail lsquoWhat was in it forthe proto-chloroplastrsquo may be an even more acute ques-tion than lsquowhat was in it for the proto-mitochondrionrsquoas speaking metabolically and energetically cyanobacteriacan do almost anything
Eukaryotes whether autotrophic or heterotrophic havenow mostly adopted the aerobic part of the energy metab-olism of their original bacterial constituents though inter-esting exceptions are known Respiration andphotosynthesis are antiparallel hydrogen transfer (redox)reactions
respiration
light photosynthesis
CH2O + A2 + H2O 2H2A + CO2
As in the Van Niel equation for photosynthesis A maybe an organic moiety for example giving H2A as succi-nate an inorganic element such as sulphur or evenlsquonothingrsquo so that H2A becomes H2 When A is O(oxygen) it becomes obvious that O2 in photosynthesiscomes from H2O not CO2 and the equation simplifies to
respiration
light photosynthesis
CH2O + O2 H2O + CO2
Phosphate group transfer and electron-transfer-linkedATP synthesis are intermediates in all cases In respir-ation CO2 is produced from stepwise oxidative decar-boxylations of pyruvate dehydrogenase and reactions ofthe tricarboxylic acid cycle In most forms of photosyn-thesis CO2 is assimilated to form initially sugar phos-phates
The mitochondrion exports ATP and CO2 and importsADP inorganic phosphate an electron acceptor andpyruvate Energetically the preferred electron acceptor ismolecular oxygen Oxygen comes from the environmentcells and cell membranes are freely permeable to oxygenThe primary metabolic division of labour between thechloroplast and the plant cytosol is that the chloroplastexports oxygen and reduced sugar phosphates (triosephosphate) and imports CO2 less reduced sugars (3-phosphoglycerate) and inorganic phosphate The light
Phil Trans R Soc Lond B (2003)
comes directly from the external environment and in thisrespect the chloroplast thylakoid is the external boundaryof the plant cell where light both as energy and infor-mation is first received The CO2 is also an environmentalinput though its supply may be regulated locally by CO2-or HCO2
3 -concentrating mechanismsAt the cellular level the mitochondrial inner membrane
is the first place where altered O2 supply exerts its pro-found metabolic effects Even granted specialized photore-ceptors the chloroplast thylakoid membrane is first placewhere changes in light quality and intensity affect a plantrsquosprimary input of energy Temperature changes selectivelyaffect diffusion-limited metabolism involving solubleintermediates In both locations abiotic environmentalchanges therefore have immediate and very far-reachingeffects on electron transfer and redox chemistry Theinternal membranes of mitochondria and chloroplastsstand at the energetic interface of the cell with the physicalenvironment and are likely to be first to know about andrespond to its changes
2 THE PROBLEM OF CYTOPLASMIC GENETICSYSTEMS
Given that the genomes of mitochondria and chloro-plasts are derived from those of once free-living bacteriait is clear that most of the bacterial genes that have sur-vived in any form have now been removed to the cellnucleus If most genes can be moved to the nucleus thenwhy not any gene And if any gene can be moved whynot all Why have mitochondria and chloroplasts retainedfunctional genomes at all What do the genes of mitochon-dria and chloroplasts have in common that has resultedin their retention
Alberts et al (1994 p 400) succinctly describe theproblem posed by the persistence of cytoplasmic geneticsystems as follows
Why do mitochondria and chloroplasts require their ownseparate genetic systems when other organelles thatshare the same cytoplasm such as peroxisomes and lyso-somes do nothellip The reason for such a costly arrange-ment is not clear and the hope that the nucleotidesequences of mitochondrial and chloroplast genomeswould provide the answer has proved unfounded Wecannot think of compelling reasons why the proteinsmade in mitochondria and chloroplasts should be madethere rather than in the cytosol
3 CORR CO-LOCATION OF GENES AND GENEPRODUCTS FOR REDOX REGULATION OF GENE
EXPRESSION
This hypothesis states that mitochondria and chloro-plasts contain genes whose expression is required to beunder the direct regulatory control of the redox state oftheir gene products or of electron carriers with whichtheir gene products interact These genes comprise a coreor primary subset of organellar genes The requirementfor redox control of each gene in the primary subset thenconfers a selective advantage upon location of that genewithin the organelle instead of in the cell nucleus Chloro-plast and mitochondrial genomes also contain genes for
Genomes in mitochondria and chloroplasts J F Allen 23
components of the chloroplast and mitochondrial geneticsystems themselves These genes comprise a secondarysubset of organellar genes genetic system genes There isgenerally no requirement for redox control of expressionof genes in this secondary subset though retention ofredox control of genetic system genes may in some casesallow amplification of redox signals acting upon genes inthe primary subset Retention of genes of the secondarysubset (genetic system genes) is necessary for the oper-ation of redox control of expression of genes in the primarysubset (bioenergetic genes) Without genes in the primarysubset the function of genes in the secondary subset iseventually lost and organelles lose their genomes
This hypothesis of co-location for redox regulation ofgene expression CORR was first outlined in a review(Allen 1992) The hypothesis was then systematically putforward in two articles (Allen 1993ab) developed(Allen amp Raven 1996) and has been independentlyreviewed (Race et al 1999) Some of its predictions havebeen tested experimentally as described later
Figure 3 illustrates the general idea of redox regulatorycontrol operating on mitochondrial genes to give the dis-tribution of sites of protein synthesis seen in figure 1 Fig-ure 4 shows the corresponding predicted pattern ofchloroplast redox regulatory control giving rise to the dis-tribution of sites of protein synthesis seen in figure 2Table 1 shows a functional classification of cyanobacterialgenes retained in 16 completely sequenced chloroplastgenomes This analysis (table 1) reveals a strong selectiveretention of genes for subunits of complexes involved inrespiration and photosynthesis and especially in chloro-plast photosystems I and II The pattern (table 1) seemsto be broadly consistent with CORR (figure 4) and incon-sistent with alternative hypotheses (see sect 4) It is reason-able to view photosynthesis as a process where preciserapid and coordinated redox regulatory control of geneexpression repays the cost of maintaining a chloroplastgenetic system
The CORR hypothesis is based on several propositionsassumptions or principles 10 of which are stated explicitlyand listed below Some of the principles are already generallyaccepted some are shared with competing hypothesesand some make direct experimental predictions Howeverother underlying principles of CORR are themselves generalprinciples or subsidiary hypotheses for which no directproof is available but for which predictions are made andpotentially falsifying observations or results can be imagined
The 10 principles that form the basis of the CORRhypothesis follow and each will be critically discussed inthe light of available evidence
(a) The principle of endosymbiotic originAs now generally agreed bioenergetic organelles have
evolved from free-living bacteriaAlthough once controversial (Mahler amp Raff 1975) this
principle will be taken as an axiom and not consideredfurther here The reader is referred to Gray amp Doolittle(1982) Gray (1992) Martin (1999) Martin et al (2001)to sect 1 and to accompanying articles in this journal issue
(b) The principle of unselective gene transferGene transfer between the symbiont or organelle and the cell
nucleus may occur in either direction and is not selective forparticular genes
Phil Trans R Soc Lond B (2003)
This principle takes the form of a testable assertion thatseems to be consistent with current knowledge Nothingcertain seems to be known about the mechanism of intra-cellular gene relocation (Palmer 1997 Martin amp Herrm-ann 1998 Martin et al 1998 Adams et al 2000 Henze ampMartin 2001 Millen et al 2001 Rujan amp Martin 2001Daley et al 2002) Plausible scenarios can be imagined onthe basis of the classical genetical phenomena of trans-formation transduction and transfection However asthere is no agreed mechanism there is no basis forasserting a mechanistic argument for selectivity Genetransfer between the organelle and nucleus seems to be aspecial case of lateral or horizontal gene transfer(Doolittle 1999) and no mechanistic basis seems to becurrently available for selectivity in what may be trans-ferred It should also be pointed out that lateral genetransfer between the organelle and nucleus does not haveto occur within a single cell nor within a multicellularindividual (although here any vertically transmitted genemust be incorporated into the germ line) for the outcomeof copying genes to novel compartments to be securedThus any plausible mechanism for heterologous recombi-nation may apply equally to gene transfer between com-partments Mitochondrial departures from the lsquouniversalrsquogenetic code and biased codon usage in chloroplasts (seelater) may be of selective value in that a barrier is therebyput in place to expression of genes in the lsquowrongrsquo compart-ment This barrier is unlikely to be insuperable especiallyif the lsquowrongrsquo compartment turns out actually to be a bet-ter location Impeded relocation might confer stabilitywithout being a primary reason for the existence of cyto-plasmic genomes
(c) The principle of unselective protein importThere is no barrier to the successful import of any precursor
protein nor to its processing and assembly into a functionalmature form
As with sect 3b this principle takes the form of a testableassertion that seems to be consistent with current knowl-edge The principle is discussed in more detail in sect 4because its negation (some proteins cannot be imported)has previously been put forward in itself as a reason forthe retention of cytoplasmic genomes Good hypothesesare easily falsified and they therefore forbid certain obser-vations The principle of unselective protein import for-bids the existence of a protein even a synthetic one thatcannot be imported by some means as a precursor
(d) The principle of continuity of redox controlDirect redox control of expression of certain genes was present
in the bacterial progenitors of chloroplasts and mitochondriaand was vital for selectively advantageous cell function beforeduring and after the transition from bacterium to organelle Themechanisms of this control have been conserved
This principle has been tested experimentally in the pastfew years and seems to be corroborated by an emergingpicture of redox signalling in bacteria and organellesExperiments carried out in vitro on the products of proteinsynthesis (Allen et al 1995 Galvis et al 1998) or RNAsynthesis (Pearson et al 1993 Konstantinov et al 1995Wilson et al 1996b) are consistent with a general phenom-enon of redox dependency of gene expression in bioener-getic organelles Redox control of chloroplast transcription
24 J F Allen Genomes in mitochondria and chloroplasts
nucleus cytosol
O2
H2O
redox regulation
N-phasemitochondrial matrix
Figure 3 Gene expression and principal pathways of biosynthesis of subunits of protein complexes involved in respiration andoxidative phosphorylation in animal mitochondria As in figure 1 reddish-brown DNA RNA and protein subunits are locatedand synthesized in the mitochondrial matrix light brown protein subunits have genes (also light brown) in the nucleus andare imported from the cytosol as precursors White genes and ribosomal and protein subunits are nuclearndashcytoplasmic and ofarchaebacterial origin Reddish-brown and light-brown genes and ribosomal and protein subunits are of bacterial origin Themajor variable environmental input is oxygen (blue) The essence of the CORR hypothesis is that it is beyond the ability ofthe nuclearndashcytoplasmic system to respond rapidly and directly to changes in oxygen concentration or partial pressure and soredox regulation of gene expression (red arrows) has been retained from the ancestral endosymbiont This redox regulationrequires co-location of certain genes with their gene products within the mitochondrion
nucleus cytosol
CH2O
CO2
redox regulation
N-phasechloroplast stroma
light light
Figure 4 Gene expression and principal pathways of biosynthesis of subunits of protein complexes involved inphotosynthesis in chloroplasts As in figure 2 green DNA RNA and protein subunits are located and synthesized in thechloroplast light brown protein subunits have genes (also light brown) in the nucleus and are imported from the cytosolas precursors White genes and ribosomal and protein subunits are nuclearndashcytoplasmic and of archaebacterial originGreen and light-brown genes and ribosomal and protein subunits are of bacterial origin The major environmental inputsare light (orange arrows) and carbon dioxide (blue) The essence of the CORR hypothesis is that it is beyond the abilityof the nuclearndashcytoplasmic system to respond rapidly and directly to changes in light and carbon dioxide and so redoxregulation of gene expression (red arrows) has been retained from the ancestral endosymbiont This redox regulationrequires co-location of certain genes with their gene products within the chloroplast
Phil Trans R Soc Lond B (2003)
Genomes in mitochondria and chloroplasts J F Allen 25
has been demonstrated (Pfannschmidt et al 1999abAllen amp Pfannschmidt 2000 Tullberg et al 2000) and wasprompted by this principle
The level of redox control of chloroplast transcriptionestablished unequivocally so far is that of plastoquinonean electron and hydrogen atom carrier located strategicallybetween photosystems I and II of photosynthesis Thedirections in which the effects are exerted are functionallyintelligible reduction of plastoquinone increases photosy-stem I transcription and decreases photosystem IItranscription oxidation of plastoquinone decreases photo-system I transcription and increases photosystem II tran-scription Plastoquinone in normal photosynthesis isreduced by the action of photosystem II and oxidized bythat of photosystem I Therefore redox control of chloro-plast gene expression provides a compensatory response(Pfannschmidt et al 1999a) and adds a previously mis-sing part of an explanation for the ability of plants andalgae to adjust the ratio of the two photosystems inresponse to changes in light quality (Chow et al 1990)By the same argument a functionally equivalent post-translational mechanism of redox control of light-harvesting function by photosystems I and II may havebeen conserved during the transition from prokaryotes toeukaryotes (Allen 1992 1995 Allen amp Nilsson 1997Allen amp Race 2002)
The precedent of chloroplast redox control of transcrip-tion (Pfannschmidt et al 1999ab) taken together with theprinciple in sect 3d predicts plastoquinone redox control ofcyanobacterial reaction centre gene transcription and thisis borne out by experiment (El Bissati amp Kirilovsky2001) This complementary self-adjusting transcriptionalresponse (Fujita 1997 Murakami et al 1997 El Bissati ampKirilovsky 2001) indicates strongly that redox control ofphotosynthetic reaction centre gene transcription waspresent in the proto-chloroplast Thus the mechanism ofregulation is likely to have been conserved throughout theevolutionary transition from bacterial symbiont to cyto-plasmic organelle A conserved mechanism of redox regu-lation of transcription in photosynthetic prokaryotes(Unden amp Bongaerts 1997 Bauer et al 1999) and chloro-plasts (Allen 1993c Pfannschmidt et al 1999b) is pre-dicted if evolutionary continuity of the process has beenmaintained
Thus apart from being consistent with experimentaldata the principle of continuity of redox control has beenof heuristic value in pointing to the underlying bacterialcharacter previously unsuspected in the control of geneexpression especially of transcription in chloroplasts ofhigher plants Other possible precedents for redox signal-ling components of cytoplasmic organelles include the prr(Eraso amp Kaplan 2000 2002 Oh amp Kaplan 2000) andreg (Bauer et al 1999) redox regulatory systems of purplenon-sulphur bacteria The TspO lsquooxygen sensorrsquo of Rho-dobacter sphaeroides is present in mammalian mitochondriaas a protein known there as a benzodiazepine receptorand each protein will complement loss of the other(Yeliseev amp Kaplan 2000) As outlined in sect 3j genes forsuch components are to be expected primarily in nucleargenomes This independent evolutionary hypothesis (sect 3d)would be strengthened still further by a demonstration ofone or more decisive redox regulatory links shared by pho-tosynthetic prokaryotes and all eukaryotes with mitochon-
Phil Trans R Soc Lond B (2003)
dria and where present with chloroplasts (Bauer et al2003)
(e) The principle of the selective value of redoxcontrol
For each gene under redox control (sect 3d) it is selectivelyadvantageous for that gene to be retained and expressed onlywithin the organelle
The twin selective advantages of redox control are likelyto be energetic efficiency and the suppression of the harm-ful side effects of electron transport operating on thewrong substrates No direct experimental support for thisprinciple is available and equally there is no evidenceagainst it Allotopic mutants in which genes have beenexperimentally relocated from organelles to the nucleushave been produced in yeast higher plants (Kanevski ampMaliga 1994) and human cells (Manfredi et al 2002) andare predicted by CORR to be poor adapters to environ-mental change Some authors believe there is potentialtherapeutic value in allotopic transformation movingmitochondrial genomes to the nucleus (Kanevski ampMaliga 1994 de Grey 2000) and predict the possibilityof r2 human cell lines in the foreseeable future (de Grey2000) This principle also has implications for ageing andthe evolution of separate sexes (Allen 1996) and possibleconsequences for development (Blackstone 1999 20002001 Harvey et al 2002) and somatic cloning in animals(Allen amp Allen 1999)
(f ) The principle of the selective value of nuclearlocation for genes not under redox control
For each bacterial gene that survives and is not under redoxcontrol it is selectively advantageous for that gene to be locatedin the nucleus and expressed only in the nucleus and cytosolIf the mature gene product functions in chloroplasts or mito-chondria the gene is first expressed in the form of a precursorfor import
There are several possible reasons for the selectiveadvantage of nuclear location of genes for organellar pro-teins One is the decreased probability of mutation arisingfrom free radical by-products of lsquoincorrectrsquo electron trans-fers (Raven et al 1994ab Allen amp Raven 1996) Therewould seem to be obvious economies in resource use andadvantages in coordination of gene expression if genes arekept in one compartment One might say that the bipartiteor tripartite genetic system of eukaryotic cells is untidyand inefficient
Another suggestion is that organellar genes do notundergo recombination and are present in relatively smallcopy numbers so that disadvantageous mutations willspread relatively quickly through a clonal population oforganelles Incorporation of functional gene copiesinstead into the nuclear genome may then be an irrevers-ible step analogous to the click of a ratchet (Saccone et al2000) There are examples of genes shared between allthree compartments (Stern amp Lonsdale 1982 Lonsdale etal 1983)
The cost of retaining an organellar genetic system is notincreased appreciably when additional genes are retainedin organelles if just one gene is retained then the cost ofa cytoplasmic genetic system is incurred and there may berelatively little extra cost for each additional gene retained(Allen amp Raven 1996) For a functional explanation of the
26 J F Allen Genomes in mitochondria and chloroplasts
existence of chloroplast and mitochondrial genomes thefocus of attention should therefore be on the relativelyconstant primary subset of bioenergetic genes (sect 3e)
(g) The principle of contemporary operation ofselection on gene location
For any species the distribution of genes between organelle(by the principle in sect 3e) and nucleus (by the principle in sect 3f)is the result of selective forces that continue to operate
This principle is essentially a statement contradictingboth the lsquofrozen accidentrsquo hypothesis and the hypothesisthat gene relocation is progressive predictable and stillincomplete (sect 4) In support of this principle it is becom-ing increasingly clear that mitochondria that lose theirfunction in aerobic respiration also lose their genomes asseen in the relict mitochondria of microsporidia(Embley amp Martin 1998 Van der Giezen et al 2002) andthe lsquomitosomersquo of Entamoeba histolytica (Tovar et al1999) In chloroplasts loss of photosynthesis results inloss of photosynthetic genes The examples of Epifagus (aparasitic higher plant) (Ems et al 1995 Wolfe et al 1992)and the residual apicomplexan plastids of Plasmodium(Wilson et al 1996a) and Toxoplasma where some plas-tidic genetic system is retained mean that some of theirgene products must be subject to redox regulation of geneexpression if this principle is correct Experimental test ofthis principle is possible using allotopic mutants as forthe predictions of the principle in sect 3f The evidence thatthe apicoplast (Wilson et al 1996a) is a plastid is largelygenetic the residual genome resembles that of photosyn-thetic plastids (chloroplasts) in gene organization ratherthan in content A plastid that has completely lost its gen-ome by virtue of having lost photosynthesis and all otherprocesses requiring redox regulation might be difficult todistinguish from an organelle derived in a similar way froma mitochondrion The persistence of targeted importedchaperones Hsp70 (Tovar et al 1999 Williams et al2002) and cp60 (Roger et al 1998) in residual organellesmight not distinguish a plastid ancestry from a mitochon-drial one
(h) The principle of primary involvement inenergy transduction
Those genes for which redox control is always vital to cellfunction have gene products involved in or closely connectedwith primary electron transfer These genes are always con-tained within the organelle
This principle would be violated and the CORRhypothesis refuted if any photosynthetic reaction centrecore subunit or respiratory chain subunit currentlyorganelle-encoded is found that functions in primary vec-torial electron transport and is nevertheless encoded in thenucleus and synthesized cytosolically as a precursor forimport all without any selective cost to the individualFrom the chloroplast transcriptional experiments ofPfannschmidt et al (1999ab) the chloroplast geneswhose transcription is regulated by the plastoquinoneredox state include psaAB (for photosystem I reactioncentre core polypeptides) psbA (for the photosystem IIpolypeptide D1) and rbcL (for the large subunit of RUB-ISCO interestingly following the same pattern of redoxcontrolmdashupregulated by oxidized plastoquinonemdashaspsbA) Transcription of the chloroplast genes petA
Phil Trans R Soc Lond B (2003)
(cytochrome f ) trnG (glycine tRNA) and rpoB (RNApolymerase) did not respond to plastoquinone redox statepetA for cytochrome f is predicted by this principle to beregulated at a different level of redox control or if at plas-toquinone at a different stage in gene expression Thisprediction might also apply to trnG and rpoB though thesequalify as members of the secondary subset that of geneticsystem genes to which the principle in sect 3i applies
(i) The principle of secondary involvement inenergy transduction
Genes whose products contribute to the organelle genetic sys-tem itself or whose products are associated with secondaryevents in energy transduction may be contained in theorganelle in one group of organisms but not in anotherdepending on the physiology and biochemistry of photosynthesisand respiration in the species concerned
Even phylogenetically moderately related species(within kingdoms) show differences in the location ofsome genes for example the ATP synthase subunits ofNeurospora and Saccharomyces (Attardi amp Schatz 1988)The CORR hypothesis also does not explicitly account forthe wide disparities in the distribution of genetic systemgenes for example the almost complete import of tRNAsin Chlamydomonas (GoldschmidtClermont 1998) Trans-port of nucleic acids across membranes is uncommon andwhy some RNAs should be organelle-encoded in somespecies but nuclearly encoded in others is not currentlyunderstood The only suggestion that can be made alongthe lines of CORR is that in some genetic systems genesmay need to be under the control of a regulatory systemthat responds to a signal arising within the organelle itselfThis signal could be the primary signal of altered redoxstate or some secondary signal Among the latter might befound signals that themselves report on the current stateof gene expression another piece of information that thenucleus might not have at its immediate disposal Suchsecondary signals could result at least in part fromCORR though other developmental and environmentalinputs may be imagined and would not detract fromCORR unless it could be shown that they provided a suf-ficient reason for the retention of the genetic system asa whole
(j) The principle of the nuclear encoding of redoxsignalling components
Components of the redox-signalling pathways upon whichCORR depends are themselves not involved in primary electrontransfer and so their genes have been relocated to the nucleusin accordance with sect 3f
This principle states that the redox signalling pathwaysthat are required for CORR should not themselves beexpected to utilize components whose biosynthesis mustbe under direct redox control Such components shouldtherefore fall into the major category of organellar pro-teins and be imported as precursors from the cytosol inline with the principle in sect 3f However the principleabove might be need to be modified in cases where ampli-fication of the redox signal is useful to the cell In thebacterial ntr system which was a precedent for CORR(Harrison et al 1990 Allen 1992) the histidine kinasentrB and aspartate response regulator ntrC form part ofthe gln operon whose transcription they control Odontella
Genomes in mitochondria and chloroplasts J F Allen 27
and Porphyra have plastids that encode more proteins thanthose of most chlorophytes (Stoebe et al 1998 Turmel etal 1999) and these proteins include two-component sig-nal transduction proteins prr and ycf27 (Stoebe et al1998) Extreme or extremely rapid environmental fluctu-ations might be catered for by amplification of signals thatare transmitted by signalling pathways whose componentsare under their own control It is not known whether ycf27and prr are in operons whose promoters their gene pro-ducts control It is also uncertain whether ycf27 is a redoxsignalling component and its distribution at least in itsplastidic location seems to be correlated with the presenceof a phycobilin-based light-harvesting system as found inred algae and cyanobacteria (Ashby amp Mullineaux1999ab Ashby et al 2002) The best guess of the originalstatement of the CORR hypothesis (Allen 1992 1993ac)was that bacterial two-component systems (Iuchi amp Lin1992 Allen 1993c Grebe amp Stock 1999) provide a plaus-ible means of exerting redox regulatory control over mito-chondrial and chloroplast gene expression andsubsequent research confirms the presence of two-compo-nent genes in genomes of Arabidopsis and yeast SeveralArabidopsis two-component genes are predicted to be tar-geted to mitochondria and chloroplasts (Forsberg et al2001) These must be considered as prime candidates forcomponents involved in redox regulation of chloroplasttranscription seen in the experiments of Pfannschmidt etal (1999a 2001a) The presumably homologous regulat-ory process serving to control photosystem stoichiometryin cyanobacteria the prr system (Li amp Sherman 2000) isnow characterized as a two-component system involvinga redox sensor and redox response regulator as originallydefined (Allen 1992 1993ac)
4 CONTRASTING HYPOTHESES CONCERNINGTHE RETENTION OF CYTOPLASMIC GENOMES
CORR is a recent addition to explanations that havebeen put forward to account for the persistence of gen-omes in organelles These models and their underlyingmechanisms are now discussed and compared
(a) There is no reason that is just how it isOne reaction to the problem of cytoplasmic genetic sys-
tems (sect 2) is that there is no problem and no particularreason for their retention However there is a very widephylogenetic distribution of a common core or primarysubset of organellar-encoded genes and one which haswithstood considerable genomic reorganization over evol-utionary time This constancy is seen for example in thesame genes being present in the chloroplasts of landplants with multiple copies of a single circular chloroplastchromosome and in dinoflagellate chloroplasts with acollection of different DNA minicircles each with essen-tially one gene (Zhang et al 1999 2001 Barbrook et al2001) Another suggested explanation of the retention ofmitochondrial and chloroplast genomes is simply torestate the endosymbiont hypothesis and to point out thatchloroplast and mitochondrial genes lsquocame inrsquo with thebacterial endosymbiont This suggestion however doesnot address the question of why some genes were retainedwhereas others were not It is also now clear that mostgenes that came in and still survive have nuclear and cyto-
Phil Trans R Soc Lond B (2003)
solic gene products whereas many chloroplast and mito-chondria proteins had a eukaryotic origin (Salzberg et al2001 Martin amp Russell 2003) The odds against a con-served pattern in gene location (eg table 1) arising bychance must be astronomical Thus there is a reason forthe retention of organellar genomes and a reason for theretention of the same primary subset of genes Becauseevolution proceeds by natural selection of chance vari-ation lsquoreasonrsquo in this context requires identification of thefunction of the location of a gene that is the selectivevalue of the phenotype produced by that location Evol-ution sometimes retains apparently functionless relics ofpreviously useful features but not for long especially ifthere is a cost
(b) The lock-in hypothesisThe lsquolock-inrsquo hypothesis of Bogorad (1975) was pro-
posed independently of the endosymbiont hypothesis anddoes not depend upon it According to the lock-in hypoth-esis core components of multi-subunit complexes must besynthesized de novo in the correct cellular compartmentotherwise photosystems or respiratory chain complexesfor example might assemble in the plasma membranevacuolar membrane or endoplasmic reticulum The lock-in hypothesis made no clear prediction about the conti-nuity of cytoplasmic genetic systems even though cyto-plasmic uniparental non-Mendelian inheritance for bothmitochondria (Roodyn amp Wilkie 1968) and plastids(Kirk amp Tilney-Bassett 1978) made it likely that organellarDNA the chondriome and the plastome replicated inde-pendently of nuclear DNA (Whitehouse 1969)
Mechanisms of protein import and targeting (Schatz1998 Koehler et al 1999 Leuenberger et al 1999 Gab-riel et al 2001 Jarvis amp Soll 2001 Pfanner amp Geissler2001 Duby amp Boutry 2002 Pfanner amp Wiedemann 2002)now effectively answer the problem of specifying the cor-rect locations for membrane protein complexes for whichthe lock-in hypothesis was proposed as a solution How-ever regulation of synthesis of an entire functional multi-subunit complex by control of synthesis of individual sub-units is still an important principle This seems to be afeature of the lsquoCESrsquo model (control by epistasy ofsynthesis) of Wollman (Wollman et al 1999 Eberhard etal 2002) which envisages nuclear control as all-embracingwith organellar genetic systems doing as it were whatthey are told In the hypothesis of CORR propoundedhere CES is equally important but the essence of CORRis that the organellar genetic system alone knows what arethe immediate and most important signals of environmen-tal change The organelle may keep the nucleus informedbut this is a lower priority than initiating an appropriateresponse CES and CORR agree that assembly of multi-subunit complexes is a complex business and requirescoordinated control in which no step is rate-limiting butmany may be obligatory
(c) Going going gone the evolutionary process oftransfer of genes from organelles to the
nucleus is still underway but incompleteThis hypothesis envisages that for most eukaryotes we
are now witnessing the last stages in a slow evolutionarytransfer of genes to the nucleus and this transfer may beexpected to proceed to completion in what is in evol-
28 J F Allen Genomes in mitochondria and chloroplasts
utionary terms the near future (Palmer 1997 Gray 19992000 Gray et al 1999 Adams et al 2000 Daley et al2002) A clear and compelling statement of this widelyheld position is that of Herrmann amp Westhoff (2001 p10)
hellipthe transfer of geneshelliptells that the partite plant gen-ome is not in a phylogenetic equilibrium All availabledata suggest that the ultimate aim of genome restructur-ing in the plant cell as in the eukaryotic cell in generalis the elimination of genome compartmentation whileretaining physiological compartmentation
One serious problem with this hypothesis is the dis-tinctly non-random sample of genes that remain to betransferred A relatively constant subset of genes is foundin bioenergetically functional mitochondria from fungi tomammals and angiosperms as in photosynthetic plastids(chloroplasts) from unicellular algae to angiosperms Thishypothesis therefore requires an additional ad hocassumption that some gene transfers are inherently moreprobable than others (this too is widely supposed seelater) and thus that gene transfer events proceed in thesame sequence in quite independent lineages Thereseems to be no evidence for this and the order of geneloss from mitochondria and chloroplasts has been used toinfer phylogeny (Martin et al 1998 Adams et al 2000Millen et al 2001) lsquoGoing going gonersquo (Palmer 1997)is difficult to refute categorically but it does not seem tobe consistent with recent evidence For example chromo-some two of Arabidopsis thaliana (Lin et al 1999) has asingle insertion encompassing with about 99 sequencesimilarity the complete Arabidopsis mitochondrial genome(Unseld et al 1997 Marienfeld et al 1999 Stupar et al2001) Unless this single transfer is a freak occurrencethis finding strongly indicates that gene transfer occurswith relative ease (Adams et al 2000 Henze amp Martin2001 Rujan amp Martin 2001 Salzberg et al 2001) Inaddition there is experimental evidence that randomlygenerated polypeptides are mitochondrially imported athigh frequency (Lemire et al 1989) The hypothesis thatwe now see the same few stragglers in diverse organellargenomes is not impossible but should be regarded as alast resort perhaps applicable only in extreme cases suchas the greatly reduced apicomplexan plastomes and chon-driomes Even there the hypothesis carries with it an obli-gation to explain why organisms with few organellar geneshave lsquoevolvedrsquo faster than those with many
(d ) The frozen accidentWhat may be described as lsquothe frozen accident hypoth-
esisrsquo states that the evolutionary process of gene transferfrom organelle to nucleus was underway when somethinghappened that stopped it One example of the frozen acci-dent is a scenario (Von Heijne 1986) in which import intomitochondria of the precursor proteins arising from newlytranslocated genes proceeded for long enough after theprimordial endosymbiosis for most of the symbiont-derived genes to be moved to the cell nucleus The lsquoacci-dentrsquo that halted the process was the evolutionary originof exocytosis and protein secretion after which it becameimpossible for precursors properly destined for mitochon-drial import to be distinguished from those destined forexport from the cell As with the lsquolock-inrsquo hypothesis the
Phil Trans R Soc Lond B (2003)
lsquofrozen accidentrsquo seems incompatible with the remarkableprecision and specificity of protein targeting (Schatz 1998Koehler et al 1999 Leuenberger et al 1999 Gabriel etal 2001 Jarvis amp Soll 2001 Pfanner amp Geissler 2001Duby amp Boutry 2002 Pfanner amp Wiedemann 2002) Pro-tein sorting mechanisms do not seem to forbid controlledimport into any cellular compartment Another lsquoaccidentrsquothat may have impeded subsequent gene transfer in thecase of mitochondria is their departure from what is other-wise a universal genetic code Genetic code differences arediscussed later in the context of whether some genes can-not be moved
(e) Some proteins (with cofactors) cannot beimported
We may describe as the lsquounimportability hypothesisrsquo theproposal that some proteins are inherently unimportableand must therefore be synthesized de novo in the correctcompartment within the cell The unimportability hypoth-esis is most often proposed as its special case the hydro-phobicity hypothesis with hydrophobicity being thereason for unimportability
(i) HydrophobicityA commonly entertained notion is that proteins that are
encoded and synthesized within organelles comprise a pri-mary subset characterized by shared hydrophobicity allare intrinsic membrane proteins (Von Heijne 1986Popot amp de Vitry 1990 Claros et al 1995) As with theCORR hypothesis favoured here the primary subset thenrequires a secondary subset of genetic system componentsthe latter not themselves being hydrophobic but beingrequired for synthesis of the others Certainly the mito-chondrial inner membrane contains some intrinsic proteincomplexes whose subunits broadly satisfy the criterion ofhydrophobicity However the core subunits that are mito-chondrially encoded are also explained by the CORRhypothesis This would make hydrophobicity a require-ment for their function in vectorial electron and protontransfer and not a reason in itself for retention of theirgenes in the cytoplasm
The hydrophobicity hypothesis has some very clearcounter-examples in chloroplasts and cannot thereforestand if one wishes for a single principle that embracesboth types of bioenergetic organelle One clear counter-example is the large (L) subunit of the enzyme RUBISCOan abundant but entirely membrane-extrinsic protein ofca 55 kDa molecular mass The RUBISCO large subunitis generally chloroplast-encoded with the exception of thenuclearly encoded bacterial-type L subunit of L2 RUBI-SCO in peridinin-containing dinoflagellates (Morse et al1995) In most photosynthetic eukaryotes RUBISCO hasa chloroplast encoded large subunit and a nuclearly enco-ded small (S) subunit (Ellis 1984 Gatenby amp Ellis 1990)giving an L8S8 holoenzyme and this RUBISCO is widelytaken as a paradigm for the need for coordination ofnuclear and chloroplast gene expression (Rodermel 2001)
By complete contrast to the RUBISCO large subunitwhich is a chloroplast-encoded and water-soluble proteina second major violation of the hydrophobicity hypothesisis the existence of the universally nuclearly encoded butalways hydrophobic subunits of the chloroplast LHC IIand LHC I (Chitnis amp Thornber 1988) These apopro-
Genomes in mitochondria and chloroplasts J F Allen 29
teins of ca 27ndash30 kDa are quite water-insoluble and areusually fully extracted only in chloroformndashmethanol or inthe presence of detergents The Lhc gene family is diverse(Green amp Kuhlbrandt 1995) but most gene productsor Lhcp (light-harvesting chlorophyll protein) polypeptideshave three membrane-spanning hydrophobic a-helices(Kuhlbrandt 1994) It is interesting that three-helix Lhcproteins seem to be absent from extant cyanobacteria andchloroxybacteria (Green amp Kuhlbrandt 1995)
(ii) The five-helix rulePerhaps with the three-helix model of LHC II in mind
a variation of the hydrophobicity hypothesis is the empiri-cal lsquofive-helix rulersquo (Popot amp de Vitry 1990 Claros et al1995 Wollman et al 1999) which states that no intrinsicmembrane protein with five or more membrane-spanninghelices can be encoded in the nucleus Five is a good guessfor a threshold number of helices demanding organellar-encoding in the context of photosynthetic reaction centres(Heathcote et al 2002) where the earliest known struc-ture that of type II centres (Deisenhofer et al 1985) asin photosystem II (Zouni et al 2001) shows exactly fivehelices with type I centres containing 11 (Jordan et al2001) There are however many nuclearly encoded sub-units in both mitochondria and chloroplasts that havefewer than five membrane helices so the five-helix rulecan only describe a sufficient condition and not a neces-sary condition for nuclear encoding
It should also be noted that ADPndashATP carriers (AACs)are encoded in the nucleus AACs of the mitochondrialinner membrane have six transmembrane helices (van derGiezen et al 2002) while the those of the chloroplast innerenvelope (Winkler amp Neuhaus 1999) have 12 (Tjaden etal 1998)
With the counter-examples of nuclearly encoded AACswith six or 12 membrane-spanning helices many intrinsicmembrane proteins with fewer than five helices somenuclearly encoded and some not and with the RUBISCOlarge subunit as a major product of chloroplast geneexpression with no membrane domain at all the five-helixrule begins to seem rather like a case of special pleadingEven if there are indeed special mitigating circumstancesfor AACs (this argument is outlined later) the five-helixrule as with the hydrophobicity hypothesis of which it isa special case will disappear if it can be shown that pos-session of five or more membrane helices is required forsome other function such as that of CORR that deter-mines the organellar location of the gene concernedHydrophobicity is in some cases a feature of proteins thatare encoded in organelles but it is not in itself the issue
For chloroplasts the hydrophobicity hypothesis predictsretention of cyanobacterial genes in functional categoriesassociated with the chloroplast envelope and with trans-port just as much as with photosynthesis Table 1 showsthat this is not the case
A mechanism for the unimportability of hydrophobicproteins is rarely stated explicitly and it is rather left tothe imagination whether hydrophobic proteins might asit were get stuck halfway through the chloroplast envelopeor mitochondrial outer membrane whether transport can-not get started on a precursor protein that has alreadyspontaneously inserted into the membrane or whetherhydrophobic proteins will be unable to make the journey
Phil Trans R Soc Lond B (2003)
through the cytosol without precipitating there In thecytosol co-translational transport or membrane insertionwould in principle address the latter point but it seemsto be generally true that chloroplast and mitochondrialproteins are post-translationally inserted into the organel-lar outer membrane So the obvious ancillary questionabout unimportability is lsquowhy then cannot hydrophobicproteins be co-translationally imported into organellesrsquoThere is evidence in yeast for co-translational transportinto mitochondria of proteins thought to be derived fromthe original endosymbiont (Marc et al 2002) If there isany truth in hydrophobicity as a barrier to protein importthe CORR principle of unselective import (sect 3c) shouldbe easily refuted for example by a demonstration that noknown targeting presequence is capable of importing poly-phenylalanine encoded by polyU for in vitro import stud-ies
An additional proposal intended to account for theunimportability of some hydrophobic proteins and forimportability of others is suggested by A de Grey(personal communication) who argues that bacteriallyderived protein precursors imported into mitochondriacross the mitochondrial inner membrane completelybefore insertion into that membrane from the mitochon-drial matrix (the N-phase) This is held to be an obstacleto successful import of membrane-intrinsic proteins thatmight en route discharge the proton motive force By con-trast eukaryotically derived proteins can be inserted intothe mitochondrial inner membrane from the inter-mem-brane space (the P-phase) and then there is for somereason no similar uncoupling effect Thus adenylatetransporters or AACs can be nuclearly encoded whereascytochrome oxidase subunits for example cannot It isnot obvious why bacterially derived subunits such asthose of cytochrome oxidase did not acquire eukaryoticinsertion mechanisms Perhaps this acquisition is forbid-den by the need to conserve pre-existing routes ofassembly into multi-subunit complexes or perhaps theneed for insertion from the inside reflects a need to incor-porate a cofactor for example haem or ironndashsulphur clus-ters along the way
(iii) Unimportability of three dimensional structures andcofactors
Accumulating evidence on protein transport mech-anisms seems to make a specific obstacle to successfultransport of certain sorts of protein less and less plausibleespecially given the observation that precursors with N-terminal targeting sequences not mature functional poly-peptides themselves traverse membranes during biosynth-esis
Protein folding inside the living cell is invariably assistedby molecular chaperones (Gatenby amp Ellis 1990 Azem etal 1997 Komiya et al 1997 1990 Jackson-Constan et al2001) proteins that impede the formation of lsquoimproperrsquostructural interactions that will otherwise produce aggre-gation or other non-functional interactions Proteinimport mechanisms appear mostly to rely on specific mol-ecular chaperones for guided unfolding before membraneinsertion and refolding after translocation of the polypep-tide into its destined compartment (Gatenby amp Ellis 1990Azem et al 1997 Komiya et al 1997 Jackson-Constanet al 2001) Because polypeptides are mostly lsquothreadedrsquo
30 J F Allen Genomes in mitochondria and chloroplasts
through membranes no special constraint on transportshould be expected to arise from 3D structure or surfaceproperties of a protein
An exception to this unfolding rule may be providedby holoproteins containing cofactors that are transportedalong with the apoprotein presumably with a largely intact3D structure The lsquoTATrsquo (twin-arginine translocase) sys-tem (Bogsch et al 1998 Dalbey amp Robinson 1999 Berkset al 2000ab Robinson 2000 Robinson amp Bolhuis 2001Sargent et al 2002) works broadly along these linesthough its mechanism is unresolved TAT is relevant herebecause it adds transport of essentially complete holopro-teins to the cellrsquos repertoire of transport mechanisms andeven proteins that may not be allowed to unfold perhapsbecause of the toxicity of the free cofactor are not therebymade candidates for an unimportability rule One variantof the unimportability hypothesis is that transport canwork only by unfolding and dissociated cofactors may betoxic to the cell The bacterially derived TAT system(Bogsch et al 1998 Dalbey amp Robinson 1999 Berks etal 2000ab Sargent et al 2002) overcomes this argumentas would examples of cofactor-free polypeptides such as(C)Fo subunit c which are nevertheless generally encodedin organelles
The presence of a vesicular transport system in chloro-plasts (Westphal et al 2001) counts against the unim-portability hypothesis Vesicular transport between thechloroplast envelope and the thylakoid membrane shouldin principle allow any protein to be ferried across thechloroplast stroma Vesicles might allow transport inparticular of membrane proteins that would discharge theproton motive force if allowed to connect transiently theN and P aqueous phases on each side of a coupling mem-brane
Another observation that seems to count against unim-portability is the increased number of membrane barriersin plastids derived from secondary tertiary or higher-orderendosymbioses These plastids seem to have retained ifanything fewer genes rather than more as unimportabilitypredicts The nucleomorph the residual nucleus of theprimary host retains many genes for molecular chap-erones (Archibald et al 2001 Cavalier-Smith 2002) inagreement with the conclusion that protein import pro-ceeds by conserved and specialized mechanisms If eachnew import added to gain a higher-order endosymbiosisrequires additional targeting presequences then perhapsthere is an advantage in components of an import machin-ery not being imported themselves
It seems that unimportability is less easy an assumptionto make today than in previous years and the onus maynow be on its proponents to cite specific examples of pro-teins that can never be imported into chloroplasts or mito-chondria
(f ) Some genes cannot be moved(i) Differences in the mitochondrial genetic code
The genetic code is the specific relation between ribon-ucleotide triplets of codons and tRNAs that recognizethem and which thus determine which individual aminoacids if any will be incorporated into the growing poly-peptide chain Mitochondria show small departures fromthe otherwise universal genetic code (Attardi amp Schatz1988 Alberts et al 1994) UGA is a lsquostoprsquo codon in the
Phil Trans R Soc Lond B (2003)
universal code and in plant mitochondria but is the codonfor tryptophan in mitochondria of animals and fungiCUA means leucine everywhere except fungal mitochon-dria where it means threonine and AGA and AGU codefor arginine in the lsquouniversalrsquo code but serine in insectmitochondria and lsquostoprsquo in mammalian ones The signifi-cance of this heterogeneity in coding is most unlikely torest in the early origin of mitochondria as the lsquouniversalrsquocode is employed by bacteria and protists Furthermorethe diversity of departures from the lsquouniversalrsquo codeamong mitochondria of different eukaryotic lineages indi-cates they had independent origins A general rule formitochondrial genetic codes is that reliance on the thirdlsquowobblersquo position is decreased which allows all 20 proteinamino acids to be specified by fewer tRNAs (22 in animalmitochondria instead of 31 in the cytosol) This in turnmeans that mtDNA can specify more proteins for a givenlength of DNA or conversely that the mitochondrialgenome can be condensed or lsquostreamlinedrsquo Although thisarrangement does not explain the retention of mitochon-drial genomes and may be a secondary event in mitochon-drial evolution it almost certainly places an extra barrierin place of gene relocation As discussed previously how-ever there seems to be no single barrier that can explainthe retention of a relatively constant subset of mitochon-drial genes
There has clearly been a contraction of mitochondrialDNA especially in animals Animal mitochondrial DNAhas few if any non-coding sequences and animal mito-chondrial genetic codes require fewer tRNAs than the sin-gle code operating in the cytosol (Gray 1999 Gray et al1999 Selosse et al 2001) These may be devices todecrease the total frequency of mutations per mitochon-drial genome assuming a constant frequency per unitlength of mitochondrial DNA It has been proposed thata genersquos removal to the nucleus decreases its susceptibilityto free radical mutagenesis (Raven et al 1994ab Allen ampRaven 1996) If this is the case then the non-standardcode may be a further indication of an overriding need tomaintain certain genes within the mitochondrion What-ever the reason for the retention of genes in organelles ithad better be a good one bioenergetic organelles areclearly the wrong place to keep genetic information andgo to extraordinary lengths to offset the consequences ofit being there
(ii) Molecular refugeesA phenomenon related to differences in the genetic code
is seen in chloroplasts This is a bias in codon usage Thelsquomolecular refugeesrsquo hypothesis (Howe et al 2000) sug-gests that nuclear copies of what were once chloroplastgenes escape constraints on codon usage and thus thatwhat distinguishes chloroplast-encoded proteins is thattheir amino-acid composition is unusual enough for thebias in codon usage not to apply It is difficult to see whatphysico-chemical properties might be uniquely shared bythe amino acids of chloroplast-encoded proteins and inaddition there seem to be no lsquoforbiddenrsquo amino-acid resi-due in chloroplasts As with the departure from the univer-sal genetic code in mitochondria is seems likely that abias in codon usage reflects some secondary property ofchloroplast genomes and is not the primary reason fortheir retention In any case any restriction on gene move-
Genomes in mitochondria and chloroplasts J F Allen 31
ment will act in the wrong direction to explain why chloro-plasts have retained genomes at all provided there is noresulting obstacle to a chloroplast protein being success-fully encoded as a precursor in the nucleus
(iii) Genes are copied and replicated not relocated in onestep
If complete relocation is envisaged as proceeding by thesimultaneous appearance of a gene in one compartmentand its deletion from another then it is possible to imaginethat whatever the vector is there may be selectivity ingene transfer (Henze amp Martin 2001) For example ifviral incorporation replication and recombination in bothorganelle and nucleus were the mechanism of relocationthen successful gene transfer might depend on the abilityof the virus to complete the process in one step This inturn might depend on some property of the gene or ofthe protein it encodes if expression occurs at any stage inthe process of relocation An assumption of the CORRhypothesis favoured here is that genes are not movedphysically but merely copied after which either the copyin the original location or the copy in the new locationmay be lost as a result of selection A useful analogy maybe the action of copying files between computer storagevolumes In this analogy the copy is identical to the orig-inal and either may subsequently be deleted without lossof information
Because any gene that is not usefully expressed willundergo no selection to maintain a functional allele theprevailing location will be the one in which the functionalcopy of the gene resides The problem of which gene sur-vives is then subsumed into the problem of which is theselectively advantageous location for it It seems reason-able to assume that there wasmdashand ismdashan interim periodin which homologous genes in different compartments areessentially in competition to see which functions best Asoutlined above there is no obvious obstacle for copyingDNA sequences within the cell The physico-chemicalproperties of DNA are fairly uniform and not directlydependent on nucleotide sequence though organelles likeintra-cellular parasites seem to prefer A and T to G andC The answer to the question lsquowhich gene location func-tions bestrsquo may have nothing to do with the gene itselfnor directly with the structure and function of its geneproduct Instead the answer may have everything to dowith the required regulatory interaction between the geneproduct and expression of the gene that produces it
(g) CORRThis hypothesis has been propounded earlier (sect 3) and
will be summarized as follows Vectorial electron and pro-ton transfer exerts regulatory control over expression ofgenes encoding proteins directly involved in or affectingredox poise This regulatory coupling is indispensable foradaptation to environmental changes in chemical activitiesof oxygen light and CO2 and requires co-location of thesegenes with their gene products Organelles lsquomake theirown decisionsrsquo on the basis of environmental changesaffecting redox state The hypothesis is abbreviatedCORR for co-location for redox regulation or more com-pletely lsquoco-location (of gene and gene product) for(evolutionary) continuity of redox regulation of geneexpressionrsquo
Phil Trans R Soc Lond B (2003)
5 SUMMARY OF THE CURRENT STATUS OF THEHYPOTHESIS OF CORR
Q Why do mitochondria and chloroplasts have theirown genetic systems
A CORR co-location of genes and gene products per-mits direct and autonomous redox regulation of geneexpression
(a) Objections to CORR(i) There is no regulation of gene expression in chloroplasts
and mitochondriaThis is still commonly stated but incorrect See the prin-
ciple in sect 3d
(ii) There is no regulation of gene expression in chloroplastsand mitochondria at the level of transcription
For chloroplasts there is now clear evidence for a func-tionally intelligible redox control of transcription(Pfannschmidt et al 1999a) This invalidates the previouswidely held assumption that all control of chloroplast geneexpression is post-transcriptional (Gruissem amp Tonkyn1993 GoldschmidtClermont 1998) The evidence formitochondria is less clear but redox control of transcrip-tion certainly cannot be ruled out and there are indi-cations that it may occur (Pearson et al 1993Konstantinov et al 1995 Wilson et al 1996b) It mustalso be restated that transcriptional control is not neces-sary for CORR but serves as a compelling example of aconserved typically bacterial mode of response that oper-ates still in eukaryotic cytoplasmic organelles thus illus-trating the principle of continuity of redox control (sect 3d)Post-transcriptional redox regulation (Danon amp Mayfield1994 Kim amp Mayfield 1997 Brown et al 2001 Treb-itsh amp Danon 2001) is consistent with CORR and thereis nothing to forbid it being a primary mechanism thatsatisfies most or indeed all of the formal requirements ofthe hypothesis in particular cases
(iii) There are no redox signalling components encoded inchloroplast or mitochondrial DNA
This is what CORR predicts See the principle in sect 3j
(iv) There is redox control of nuclear gene expressionThis is correct and many examples are now known
especially for chloroplasts (Escoubas et al 1995 Kovacs etal 2000 Pfannschmidt et al 2001ab Surpin et al 2002)Redox signals from photosynthetic or respiratory electrontransport chains are not at all prohibited by CORR Theessence of CORR is that there is a subset of organellarproteins for which there is an overriding requirement fora redox regulatory control that is initiated within theorganelle and whose effects are exerted by the simplestand most direct route This redox signalling is envisagedto take place without a requirement for prior consultationand coordination with what may be regarded as a nucleo-cytoplasmic bureaucracy
(v) Nuclear genes can be expressed as rapidly as chloroplastand mitochondrial genes
Zerges (2002) for example cites the equal times takenfor complete expression of nuclear and chloroplast genesin Chlamydomonas as evidence against the CORR hypoth-
32 J F Allen Genomes in mitochondria and chloroplasts
esis However the question is not one of the time takento express a gene but the time taken to alter the rate ofits expression in response to a (redox) signal originatingwithin the organelle An equally important additional fac-tor may be the autonomy of the signalling pathway thatis its independence from other inputs as described in theprevious paragraph One might say that mitochondria andchloroplasts require a free hand
(vi) There is no evidence to support this hypothesisThe hypothesis is rich in predictions (sectsect 3 and 6) and
those that have been tested stringently are in agreementwith experimental results None of the predictions of thehypothesis has been found to be incorrect and none of theunderlying principles (sect 3) has been disproved Furtherpredictions will be the subject of future experiments
(b) Evidence for CORR(i) Chloroplasts have selectively retained cyanobacterial genes
for primary events in photosynthesisSee table 1
(ii) There is regulation of gene expression in chloroplastsThere is transcriptional regulation of gene expression
in chloroplasts (sect 3d) The case for mitochondria is notyet proven
(iii) There is redox regulation of gene expression inchloroplasts
There is transcriptional redox regulation of geneexpression in chloroplasts (sect 3d) The case for mitochon-dria is not yet proven though see (Allen et al 1995 Kon-stantinov et al 1995 Galvis et al 1998)
(iv) There are no redox signalling components encoded inchloroplast or mitochondrial DNA
Correct but with possible exceptions discussed in sect 3j
(v) There are lsquobacterialrsquo redox signalling components ineukaryotes
See sect 3j
(vi) Further predictions DNA-array results and phenotypesof allotopic mutants
A systematic test of CORR might be to make an oxid-ized-minus-reduced gene expression profile of the lsquotran-scriptomersquo of a cyanobacterium to see if the set of geneswhose transcription is under most rapid and direct redoxcontrol corresponds to the genes of the chloroplast gen-ome Laboratories with this experimental capability do notseem to have addressed this possibility (Hihara et al 2002Ikeuchi 2002) Likewise if a putative modern representa-tive of the proto-mitochondrion can be found and agreedeg Rhodospirillum rubrum or Paracoccus denitrificans thena similar experiment should reveal a subset of genes corre-sponding to mitochondrial genomes DNA-microarraysalso present the possibility of testing the relative rate ofredox response and dependency on other signallinginputs of transcription of eukaryotic genes To dateorganellar genes have not been included in such investi-gations (Ferea et al 1999) A further experimental possi-bility is to examine the redox regulatory phenotypes ofexisting allotopic mutants as outlined in sect 3e
Phil Trans R Soc Lond B (2003)
6 CONCLUSION AND FUTURE PROSPECTS
The CORR hypothesis states that the function of mito-chondrial and chloroplast genomes is to provide co-location (of gene and gene product) for (evolutionary)continuity of redox regulation of gene expression Thishypothesis is neither self-contradictory nor devoid of test-able predictions It is also consistent with available evi-dence and in principle makes sense of the otherwisepuzzling distribution of genes between compartments ineukaryotic cells The explanation that CORR offersapplies equally to mitochondria and chloroplasts andidentifies selective forces that still operate to determine thelocation of genes No selective barriers need be assumedto gene transfer protein import nor membrane insertionNor do we need to invoke a lsquofrozen accidentrsquo that some-thing happened to stop gene transfer for some reason atsome time in the remote and inaccessible past CORR alsoexplains the absence of genomes from organelles that havelost their primary bioenergetic functions CORR may actas a stimulus to a search for residual bioenergetic functionin those organelles that have lost aerobic respiration andphotosynthesis but where residual genomes remain
CORR also predicts regulatory properties of knowncomponents of chloroplasts and mitochondria that in turnindicate important but as yet uncharacterized flexibility inenergy transduction These components include the RUB-ISCO large subunit and the CFo and Fo subunits of coup-ling ATPase in both chloroplasts and mitochondriaRedox regulation of the relative stoichiometry of compo-nents as seen in regulation of chloroplast photosystemstoichiometry (Pfannschmidt et al 1999a) may extend to(C)Fo-(C)F1-ATPase indicating that the function ofretention of genes for one or more of these componentsis flexibility in the stoichiometries of H1 to e2 and H1 toATP in chemiosmotic coupling (Schemidt et al 1998Stock et al 2000 Allen 2002)
So far explicit tests of CORR have largely been con-fined to chloroplasts and to some extent mitochondria ofhigher plants but this merely reflects the expertise andinterests of the investigators Further attempted disproofof CORR is welcomed and encouraged Novel experi-mental tests can be devised as well as attempted refu-tation through analysis of existing data It is hoped thatexplicit attention will be given to the CORR hypothesis inlaboratories already suitably equipped and that resourcescan be allocated for this purpose in laboratories wherethere is the interest but not currently the technology orexpertise
The heuristic value of the hypothesis already seen inthe role of redox-regulated transcription of photosyntheticreaction centre genes in chloroplasts may be its most use-ful property The central proposal is that mitochondriaand chloroplasts contain genes for proteins whose functionin electron transfer demands rapid direct and uncon-ditional redox regulatory control of their biosynthesis Thefull implications of this hypothesis are very broad and onlyjust beginning to be explored
I thank Carol Allen Aubrey de Grey William Martin John ARaven and Francis-Andre Wollman for discussion and corre-spondence during the preparation of this paper and WilliamMartin for suggestions including table 1 Research grants from
Genomes in mitochondria and chloroplasts J F Allen 33
The Swedish Natural Sciences Research Council (NFR) andthe Crafoord Foundation are gratefully acknowledged
REFERENCES
Adams K L Daley D O Qiu Y L Whelan J amp PalmerJ D 2000 Repeated recent and diverse transfers of a mito-chondrial gene to the nucleus in flowering plants Nature408 354ndash357
Alberts B Bray D Lewis J Raff M Roberts K amp Wat-son J D 1994 The molecular biology of the cell New Yorkand London Garland Publishing
Allen C A Hakansson G amp Allen J F 1995 Redox con-ditions specify the proteins synthesized by isolated chloro-plasts and mitochondria Redox Rep 1 119ndash123
Allen J F 1992 Protein phosphorylation in regulation ofphotosynthesis Biochim Biophys Acta 1098 275ndash335
Allen J F 1993a Control of gene expression by redox poten-tial and the requirement for chloroplast and mitochondrialgenomes J Theor Biol 165 609ndash631
Allen J F 1993b Redox control of gene expression and thefunction of chloroplast genomes a hypothesis PhotosynthRes 36 95ndash102
Allen J F 1993c Redox control of transcription sensorsresponse regulators activators and repressors FEBS Lett332 203ndash207
Allen J F 1995 Thylakoid proteinndashphosphorylation state-1ndashstate-2 transitions and photosystem stoichiometry adjust-ment redox control at multiple levels of gene expressionPhysiol Plant 93 196ndash205
Allen J F 1996 Separate sexes and the mitochondrial theoryof ageing J Theor Biol 180 135ndash140
Allen J F 2002 Photosynthesis of ATP-electrons protonpumps rotors and poise Cell 110 273ndash276
Allen J F amp Allen C A 1999 A mitochondrial model forpremature ageing of somatically cloned mammals IUBMBLife 48 369ndash372
Allen J F amp Nilsson A 1997 Redox signalling and the struc-tural basis of regulation of photosynthesis by protein phos-phorylation Physiol Plant 100 863ndash868
Allen J F amp Pfannschmidt T 2000 Balancing the two photo-systems photosynthetic electron transfer governs transcrip-tion of reaction centre genes in chloroplasts Phil TransR Soc Lond B 355 1351ndash1357 (DOI 101098rstb20000697)
Allen J F amp Race H L 2002 Will the real LHC II kinaseplease step forward Sciencersquos STKE httpwwwstkeorgcgicontentfullsigtrans2002155pe43
Allen J F amp Raven J A 1996 Free-radical-induced mutationvs redox regulation costs and benefits of genes in organellesJ Mol Evol 42 482ndash492
Archibald J M Cavalier-Smith T Maier U amp Douglas S2001 Molecular chaperones encoded by a reduced nucleusthe cryptomonad nucleomorph J Mol Evol 52 490ndash501
Ashby M K amp Mullineaux C W 1999a Cyanobacterialycf27 gene products regulate energy transfer from phycobili-somes to photosystems I and II FEMS Microbiol Lett 181253ndash260
Ashby M K amp Mullineaux C W 1999b The role of ApcDand ApcF in energy transfer from phycobilisomes to PSI andPSII in a cyanobacterium Photosynth Res 61 169ndash179
Ashby M K Houmard J amp Mullineaux C W 2002 Theycf27 genes from cyanobacteria and eukaryotic algae distri-bution and implications for chloroplast evolution FEMSMicrobiol Lett 214 25ndash30
Attardi G amp Schatz G 1988 Biogenesis of mitochondria ARev Cell Biol 4 289ndash333
Azem A Oppliger W Lustig A Jeno P Feifel B SchatzG amp Horst M 1997 The mitochondrial hsp70 chaperone
Phil Trans R Soc Lond B (2003)
system effect of adenine nucleotides peptide substrate andmGrpE on the oligomeric state of mhsp70 J Biol Chem272 20 901ndash20 906
Barbrook A C Symington H Nisbet R E R LarkumA amp Howe C J 2001 Organisation and expression of theplastid genome of the dinoflagellate Amphidinium opercula-tum Mol Genet Genomics 266 632ndash638
Bauer C E Elsen S amp Bird T H 1999 Mechanisms forredox control of gene expression A Rev Microbiol 53495ndash523
Bauer C E Elsen S Swem L R Swem D L amp MasudaS 2003 Redox and light regulation of gene expression inphotosynthetic prokaryotes Phil Trans R Soc Lond B 358147ndash154 (DOI 101098rstb20021189)
Berks B C Sargent F De Leeuw E Hinsley A P Stan-ley N R Jack R L Buchanan G amp Palmer T 2000aA novel protein transport system involved in the biogenesisof bacterial electron transfer chains Biochim Biophys ActaBioenerg 1459 325ndash330
Berks B C Sargent F amp Palmer T 2000b The tat proteinexport pathway Mol Microbiol 35 260ndash274
Blackstone N W 1999 Redox control in development andevolution evidence from colonial hydroids J Exp Biol 2023541ndash3553
Blackstone N W 2000 Redox control and the evolution ofmulticellularity Bioessays 22 947ndash953
Blackstone N W 2001 Redox state reactive oxygen speciesand adaptive growth in colonial hydroids J Exp Biol 2041845ndash1853
Bogorad L 1975 Evolution of organelles and eukaryotic gen-omes Science 188 891ndash898
Bogsch E G Sargent F Stanley N R Berks B C Robin-son C amp Palmer T 1998 An essential component of a novelbacterial protein export system with homologues in plastidsand mitochondria J Biol Chem 273 18 003ndash18 006
Brown E C Somanchi A amp Mayfield S P 2001 Interor-ganellar crosstalk new perspectives on signaling from thechloroplast to the nucleus Genome Biol 2 10211ndash10214
Cavalier-Smith T 2002 Chloroplast evolution secondarysymbiogenesis and multiple losses Curr Biol 12 R62ndashR64
Chitnis P R amp Thornber J P 1988 The major light-har-vesting complex of photosystem 2 Aspects of its molecularand cell biology Photosynth Res 16 41ndash63
Chow W S Melis A amp Anderson J M 1990 Adjustmentsof photosystem stoichiometry in chloroplasts improve thequantum efficiency of photosynthesis Proc Natl Acad SciUSA 87 7502ndash7506
Claros M G Perea J Shu Y Samatey F A PopotJ L amp Jacq C 1995 Limitations to in vivo import of hydro-phobic proteins into yeast mitochondria The case of a cyto-plasmically synthesized apocytochrome b Eur J Biochem228 762ndash771
Dalbey R E amp Robinson C 1999 Protein translocation intoand across the bacterial plasma membrane and the plant thy-lakoid membrane Trends Biochem Sci 24 17ndash22
Daley D O Adams K L Clifton R Qualmann S MillarA H Palmer J D Pratje E amp Whelan J 2002 Genetransfer from mitochondrion to nucleus novel mechanismsfor gene activation from Cox2 Plant J 30 11ndash21
Danon A amp Mayfield S P 1994 Light-regulated translationof chloroplast messenger RNAs through redox potentialScience 266 1717ndash1719
de Grey A 2000 Mitochondrial gene therapy an arena for thebiomedical use of inteins Trends Biotechnol 18 394ndash399
Deisenhofer J Epp O Miki K Huber R amp Michel H1985 Structure of the protein subunits in the photosyntheticreaction center of Rhodopseudomonas viridis at 3Ac resolutionNature 318 618ndash624
34 J F Allen Genomes in mitochondria and chloroplasts
Delwiche C F 1999 Tracing the thread of plastid diversitythrough the tapestry of life Am Nat 154 S164ndashS177
Doolittle W F 1999 Phylogenetic classification and the uni-versal tree Science 284 2124ndash2129
Douglas S E 1998 Plastid evolution origins diversitytrends Curr Opin Genet Dev 8 655ndash661
Duby G amp Boutry M 2002 Mitochondrial protein importmachinery and targeting information Plant Sci 162 477ndash490
Eberhard S Drapier D amp Wollman F A 2002 Searchinglimiting steps in the expression of chloroplast encoded pro-teins relations between gene copy number transcriptiontranscript abundance and translation rate in the chloroplastof Chlamydomonas reinhardtii Plant J 31 149ndash160
El Bissati K amp Kirilovsky D 2001 Regulation of psbA andpsaE expression by light quality in Synechocystis species PCC6803 A redox control mechanism Plant Physiol 1251988ndash2000
Ellis R J 1984 The nuclear domination of chloroplast devel-opment Science Progress 69 129ndash142
Embley T M amp Martin W 1998 Molecular evolution ahydrogen-producing mitochondrion Nature 396 517ndash519
Ems S C Morden C W Dixon C K Wolfe K HdePamphilis C W amp Palmer J D 1995 Transcriptionsplicing and editing of plastid RNAs in the non-photosyn-thetic plant Epifagus virginiana Plant Mol Biol 29 721ndash733
Eraso J M amp Kaplan S 2000 From redox flow to gene regu-lation role of the PrrC protein of Rhodobacter sphaeroides241 Biochemistry 39 2052ndash2062
Eraso J M amp Kaplan S 2002 Redox flow as an instrumentof gene regulation Meth Enzymol 348 216ndash229
Escoubas J M Lomas M Laroche J amp Falkowski P G1995 Light-intensity regulation of cab gene-transcription issignaled by the redox state of the plastoquinone pool ProcNatl Acad Sci USA 92 10 237ndash10 241
Ferea T L Botstein D Brown P O amp Rosenzweig R F1999 Systematic changes in gene expression patterns follow-ing adaptive evolution in yeast Proc Natl Acad Sci USA96 9721ndash9726
Forsberg J Rosenquist M Fraysse L amp Allen J F 2001Redox signalling in chloroplasts and mitochondria genomicand biochemical evidence for two-component regulatory sys-tems in bioenergetic organelles Biochem Soc Trans 29403ndash407
Fujita Y 1997 A study on the dynamic features of photosys-tem stoichiometry accomplishments and problems forfuture studies Photosynth Res 53 83ndash93
Gabriel K Buchanan S K amp Lithgow T 2001 The alphaand the beta protein translocation across mitochondrial andplastid outer membranes Trends Biochem Sci 26 36ndash40
Galvis M L E Allen J F amp Hakansson G 1998 Proteinsynthesis by isolated pea mitochondria is dependent on theactivity of respiratory complex II Curr Genet 33 320ndash329
Gatenby A A amp Ellis R J 1990 Chaperone function theassembly of ribulose bisphosphate carboxylase-oxygenase ARev Cell Biol 6 125ndash149
GoldschmidtClermont M 1998 Coordination of nuclear andchloroplast gene expression in plant cells Int Rev CytolSurv Cell Biol 177 115ndash180
Gray M W 1992 The endosymbiont hypothesis revisited IntRev Cytol 141 233ndash357
Gray M W 1999 Evolution of organellar genomes CurrOpin Genet Dev 9 678ndash687
Gray M W 2000 Evolutionary biology mitochondrial geneson the move Nature 408 302ndash305
Gray M W amp Doolittle W F 1982 Has the endosymbionthypothesis been proven Microbiol Rev 46 1ndash42
Gray M W Burger G amp Lang B F 1999 Mitochondrialevolution Science 283 1476ndash1481
Phil Trans R Soc Lond B (2003)
Grebe T W amp Stock J B 1999 The histidine protein kinasesuperfamily Adv Microb Physiol 41 139ndash227
Green B R amp Kuhlbrandt W 1995 Sequence conservationof light-harvesting and stress-response proteins in relation tothe 3-dimensional molecular structure of Lhcii PhotosynthRes 44 139ndash148
Gruissem W amp Tonkyn J C 1993 Control mechanisms ofplastid gene expression Crit Rev Plant Sci 12 19ndash55
Hackstein J H P (and 11 others) 2001 Hydrogenosomesconvergent adaptations of mitochondria to anaerobicenvironments Zool Anal Complex Syst 104 290ndash302
Harrison M A Keen J N Findlay J B C amp Allen J F1990 Modification of a Glnb-like gene-product by photosyn-thetic electron-transport in the cyanobacterium Synechoc-occus 6301 FEBS Lett 264 25ndash28
Harvey A J Kind K L amp Thompson J G 2002 Redoxregulation of early embryo development Reproduction 123479ndash486
Heathcote P Fyfe P K amp Jones M R 2002 Reactioncentres the structure and evolution of biological solarpower Trends Biochem Sci 27 79ndash87
Henze K amp Martin W 2001 How do mitochondrial genesget into the nucleus Trends Genet 17 383ndash387
Herrmann R G amp Westhoff P 2001 Thylakoid biogenesis anddynamics the result of a complex phylogenetic puzzle InRegulation of photosynthesis vol 11 (ed E-M Aro amp BAndersson) pp 1ndash28 Dordrecht The Netherlands Kluwer
Hihara Y Sonoike K Kanehisa M amp Ikeuchi M 2002Search for redox responsive genes in Synechocystis sp PCC6803 by using DNA microarrays Plant Cell Physiol 43S178
Howe C J Barbrook A C amp Lockhart P J 2000 Organellegenes do they jump or are they pushed Trends Genet 1665ndash66
Ikeuchi M 2002 Gene expression and responses to light stressin cyanobacteria Plant Cell Physiol 43 S21
Iuchi S amp Lin E C C 1992 Mutational analysis of signaltransduction by Arcb a membrane sensor protein respon-sible for anaerobic repression of operons involved in the cen-tral aerobic pathways in Escherichia coli J Bacteriol 1743972ndash3980
Jackson-Constan D Akita M amp Keegstra K 2001 Molecu-lar chaperones involved in chloroplast protein importBiochim Biophys Acta Mol Cell Res 1541 102ndash113
Jarvis P amp Soll M 2001 Toc Tic and chloroplast proteinimport Biochim Biophys Acta Mol Cell Res 1541 64ndash79
Jordan P Fromme P Witt H T Klukas O SaengerW amp Krauss N 2001 Three-dimensional structure ofcyanobacterial photosystem I at 25 acngstrom resolutionNature 411 909ndash917
Kaneko T (and 23 others) 1996a Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regionsDNA Res 3 109ndash136
Kaneko T (and 23 others) 1996b Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regions[Supplement] DNA Res 3 185ndash209
Kanevski I amp Maliga P 1994 Relocation of the plastid rbcLgene to the nucleus yields functional ribulose-15-bisphosph-ate carboxylase in tobacco chloroplasts Proc Natl Acad SciUSA 91 1969ndash1973
Kim J amp Mayfield S P 1997 Protein disulfide isomerase as aregulator of chloroplast translational activation Science 2781954ndash1957
Genomes in mitochondria and chloroplasts J F Allen 35
Kirk J T O amp Tilney-Bassett R A E 1978 The plastidsTheir chemistry structure growth and inheritance Amster-dam Elsevier
Koehler C M Merchant S amp Schatz G 1999 How mem-brane proteins travel across the mitochondrial intermem-brane space Trends Biochem Sci 24 428ndash432
Komiya T Rospert S Schatz G amp Mihara K 1997 Bind-ing of mitochondrial precursor proteins to the cytoplasmicdomains of the import receptors Tom70 and Tom20 isdetermined by cytoplasmic chaperones FASEB J 11 1196
Konstantinov Y M Lutsenko G N amp Podsosonny V A1995 Genetic functions of isolated maize mitochondriaunder model changes of redox conditions Biochem MolBiol Int 36 319ndash326
Korab-Laskowska M Rioux P Brossard N LittlejohnT G Gray M W Lang B F amp Burger G 1998 Theorganelle genome database project (GOBASE) Nucleic AcidsRes 26 138ndash144
Kovacs L Wiessner W Kis M Nagy F Mende D ampDemeter S 2000 Short- and long-term redox regulation ofphotosynthetic light energy distribution and photosystemstoichiometry by acetate metabolism in the green alga Chla-mydobotrys stellata Photosynth Res 65 231ndash247
Kuhlbrandt W 1994 Structure and function of the plant light-harvesting complex LHC-II Curr Opin Struct Biol 4519ndash528
Lang B F Gray M W amp Burger G 1999 Mitochondrialgenome evolution and the origin of eukaryotes A RevGenet 33 351ndash397
Lemire B D Fankhauser C Baker A amp Schatz G 1989The mitochondrial targeting function of randomly generatedpeptide sequences correlates with predicted helical amphi-philicity J Biol Chem 264 20 206ndash20 215
Leuenberger D Bally N A Schatz G amp Koehler C M1999 Different import pathways through the mitochondrialintermembrane space for inner membrane proteins EMBOJ 18 4816ndash4822
Li H amp Sherman L A 2000 A redox-responsive regulator ofphotosynthesis gene expression in the cyanobacterium Syne-chocystis sp strain PCC 6803 J Bacteriol 182 4268ndash4277
Lin X Y (and 36 others) 1999 Sequence and analysis of chro-mosome 2 of the plant Arabidopsis thaliana Nature 402761ndash768
Lonsdale D M Hodge T P Howe C J amp Stern D B1983 Maize mitochondrial-DNA contains a sequence hom-ologous to the ribulose-15-bisphosphate carboxylase largesubunit gene of chloroplast DNA Cell 34 1007ndash1014
McFadden G I 1999 Endosymbiosis and evolution of theplant cell Curr Opin Plant Biol 2 513ndash519
McFadden G I 2001 Chloroplast origin and integrationPlant Physiol 125 50ndash53
Mahler H R amp Raff R A 1975 The evolutionary origin ofthe mitochondrion a nonsymbiotic model Int Rev Cytol43 1ndash124
Manfredi G Fu J Ojaimi J Sadlock J E Kwong J QGuy J amp Schon E A 2002 Rescue of a deficiency in ATPsynthesis by transfer of MTATP6 a mitochondrial DNA-encoded gene to the nucleus Nature Genet 30 394ndash399
Marc P Margeot A Devaux F Blugeon C Corral-Debrinski M amp Jacq C 2002 Genome-wide analysis ofmRNAs targeted to yeast mitochondria EMBO Rep 3159ndash164
Marienfeld J Unseld M amp Brennicke A 1999 The mito-chondrial genome of Arabidopsis is composed of both nativeand immigrant information Trends Plant Sci 4 495ndash502
Martin W 1999 A briefly argued case that mitochondria andplastids are descendants of endosymbionts but that thenuclear compartment is not Proc R Soc Lond B 2661387ndash1395 (DOI 101098rspb19990792)
Phil Trans R Soc Lond B (2003)
Martin W amp Herrmann R G 1998 Gene transfer fromorganelles to the nucleus how much what happens andwhy Plant Physiol 118 9ndash17
Martin W amp Muller M 1998 The hydrogen hypothesis forthe first eukaryote Nature 392 37ndash41
Martin W amp Russell M J 2003 On the origins of cells ahypothesis for the evolutionary transitions from abiotic geo-chemistry to chemoautotrophic prokaryotes and from pro-karyotes to nucleated cells Phil Trans R Soc Lond B 35859ndash85 (DOI 101098rstb20021183)
Martin W Stoebe B Goremykin V Hansmann S Hase-gawa M amp Kowallik K V 1998 Gene transfer to thenucleus and the evolution of chloroplasts Nature 393162ndash165
Martin W Hoffmeister M Rotte C amp Henze K 2001 Anoverview of endosymbiotic models for the origins of eukary-otes their ATP-producing organelles (mitochondria andhydrogenosomes) and their heterotrophic lifestyle BiolChem 382 1521ndash1539
Martin W Rujan T Richly E Hansen A Cornelsen SLins T Leister D Stoebe B Hasegawa M amp PennyD 2002 Evolutionary analysis of Arabidopsis cyanobacterialand chloroplast genomes reveals plastid phylogeny and thou-sands of cyanobacterial genes in the nucleus Proc NatlAcad Sci USA 99 12 246ndash12 251
Millen R S (and 12 others) 2001 Many parallel losses of infAfrom chloroplast DNA during angiosperm evolution withmultiple independent transfers to the nucleus Plant Cell 13645ndash658
Morse D Salois P Markovic P amp Hastings J W 1995 Anuclear-encoded form-II RUBISCO in dinoflagellatesScience 268 1622ndash1624
Murakami A Kim S J amp Fujita Y 1997 Changes in photo-system stoichiometry in response to environmental con-ditions for cell growth observed with the cyanophyteSynechocystis PCC 6714 Plant Cell Physiol 38392ndash397
Oh J I amp Kaplan S 2000 Redox signaling globalization ofgene expression EMBO J 19 4237ndash4247
Ozawa T 1995 Mitochondrial-DNA mutations associatedwith aging and degenerative diseases Exp Gerontol 30269ndash290
Palmer J D 1997 Organelle genomes going going goneScience 275 790ndash791
Pearson C K Wilson S B Schaffer R amp Ross A W 1993NAD turnover and utilization of metabolites for RNA syn-thesis in a reaction sensing the redox state of the cyto-chrome-b( 6 )f complex in isolated chloroplasts Eur JBiochem 218 397ndash404
Pfanner N amp Geissler A 2001 Versatility of the mitochon-drial protein import machinery Natl Rev Mol Cell Biol 2339ndash349
Pfanner N amp Wiedemann N 2002 Mitochondrial proteinimport two membranes three translocases Curr Opin CellBiol 14 400ndash411
Pfannschmidt T Nilsson A amp Allen J F 1999a Photosyn-thetic control of chloroplast gene expression Nature 397625ndash628
Pfannschmidt T Nilsson A Tullberg A Link G amp AllenJ F 1999b Direct transcriptional control of the chloroplastgenes psbA and psaAB adjusts photosynthesis to light energydistribution in plants IUBMB Life 48 271ndash276
Pfannschmidt T Allen J F amp Oelmuller R 2001a Prin-ciples of redox control in photosynthesis gene expressionPhysiol Plant 112 1ndash9
Pfannschmidt T Schutze K Brost M amp Oelmuller R2001b A novel mechanism of nuclear photosynthesis generegulation by redox signals from the chloroplast during pho-
36 J F Allen Genomes in mitochondria and chloroplasts
tosystem stoichiometry adjustment J Biol Chem 27636 125ndash36 130
Popot J L amp de Vitry C 1990 On the microassembly of inte-gral membrane proteins A Rev Biophys Biophys Chem 19369ndash403
Race H L Herrmann R G amp Martin W 1999 Why haveorganelles retained genomes Trends Genet 15 364ndash370
Raven J A Johnston A M Parsons R amp Kubler J 1994aThe influence of natural and experimental high O2 concen-trations on O2 evolving phototrophs Biol Rev CambridgePhil Soc 69 61ndash94
Raven J A Johnston A M Parsons R amp Kubler J 1994bThe occurrence and influence on photolithotrophs of highoxygen concentrations Proc R Soc Edinb B 102 193ndash201
Robinson C 2000 The twin-arginine translocation system anovel means of transporting folded proteins in chloroplastsand bacteria Biol Chem 381 89ndash93
Robinson C amp Bolhuis A 2001 Protein targeting by the twin-arginine translocation pathway Natl Rev Mol Cell Biol 2350ndash356
Rodermel S 2001 Pathways of plastid-to-nucleus signalingTrends Plant Sci 6 471ndash478
Roger A J 1999 Reconstructing early events in eukaryoticevolution Am Nat 154 S146ndashS163
Roger A J Svard S G Tovar J Clark C G SmithM W Gillin F D amp Sogin M L 1998 A mitochondrial-like chaperonin 60 gene in Giardia lamblia evidence thatdiplomonads once harbored an endosymbiont related to theprogenitor of mitochondria Proc Natl Acad Sci USA 95229ndash234
Roodyn D B amp Wilkie D 1968 The biogenesis of mitochondriaMethuenrsquos monographs on biological subjects London Methuen
Rujan T amp Martin W 2001 How many genes in Arabidopsiscome from cyanobacteria An estimate from 386 proteinphylogenies Trends Genet 17 113ndash120
Saccone C Gissi C Lanave C Larizza A Pesole G ampReyes A 2000 Evolution of the mitochondrial genetic sys-tem an overview Gene 261 153ndash159
Salzberg S L White O Peterson J amp Eisen J A 2001Microbial genes in the human genome lateral transfer orgene loss Science 292 1903ndash1906
Sargent F Berks B C amp Palmer T 2002 Assembly of mem-brane-bound respiratory complexes by the Tat protein-trans-port system Arch Microbiol 178 77ndash84
Schatz G 1998 Protein transport the doors to organellesNature 395 439ndash440
Schemidt R A Qu J Williams J R amp Brusilow W S1998 Effects of carbon source on expression of F0 genes andon the stoichiometry of the c subunit in the F1 F0 ATPaseof Escherichia coli J Bacteriol 180 3205ndash3208
Selosse M A Albert B R amp Godelle B 2001 Reducing thegenome size of organelles favours gene transfer to thenucleus Trends Ecol Evol 16 135ndash141
Stern D B amp Lonsdale D M 1982 Mitochondrial and chlo-roplast genomes of maize have a 12- kilobase DNA-sequencein common Nature 299 698ndash702
Stock D Gibbons C Arechaga I Leslie A G W ampWalker J E 2000 The rotary mechanism of ATP synthaseCurr Opin Struct Biol 10 672ndash679
Stoebe B amp Maier U G 2002 One two three naturersquos toolbox for building plastids Protoplasma 219 123ndash130
Stoebe B Martin W amp Kowallik K V 1998 Distributionand nomenclature of protein-coding genes in 12 sequencedchloroplast genomes Plant Mol Biol Rep 16 243ndash255
Stupar R M Lilly J W Town C D Cheng Z Kaul SBuell C R amp Jiang J M 2001 Complex mtDNA consti-tutes an approximate 620 kb insertion on Arabidopsis thali-ana chromosome 2 implication of potential sequencing
Phil Trans R Soc Lond B (2003)
errors caused by large-unit repeats Proc Natl Acad SciUSA 98 5099ndash5103
Surpin M Larkin R M amp Chory J 2002 Signal transduc-tion between the chloroplast and the nucleus Plant Cell 14S327ndashS338
Tjaden J Schwoppe C Mohlmann T Quick P W ampNeuhaus H E 1998 Expression of a plastidic ATPADPtransporter gene in Escherichia coli leads to a functional aden-ine nucleotide transport system in the bacterial cytoplasmicmembrane J Biol Chem 273 9630ndash9636
Tovar J Fischer A amp Clark C G 1999 The mitosome a novelorganelle related to mitochondria in the amitochondrial para-site Entamoeba histolytica Mol Microbiol 32 1013ndash1021
Trebitsh T amp Danon A 2001 Translation of chloroplastpsbA mRNA is regulated by signals initiated by both photos-ystems II and I Proc Natl Acad Sci USA 98 12 289ndash12 294
Tullberg A Alexciev K Pfannschmidt T amp Allen J F2000 Photosynthetic electron flow regulates transcription ofthe psaB gene in pea (Pisum sativum L) chloroplasts throughthe redox state of the plastoquinone pool Plant Cell Physiol41 1045ndash1054
Turmel M Lemieux C Burger G Lang B F Otis CPlante I amp Gray M W 1999 The complete mitochondrialDNA sequences of Nephroselmis olivacea and Pedinomonasminor two radically different evolutionary patterns withingreen algae Plant Cell 11 1717ndash1729
Unden G amp Bongaerts J 1997 Alternative respiratory path-ways of Escherichia coli energetics and transcriptional regu-lation in response to electron acceptors Biochim BiophysActa Bioenerg 1320 217ndash234
Unseld M Marienfeld J R Brandt P amp Brennicke A1997 The mitochondrial genome of Arabidopsis thaliana con-tains 57 genes in 366 924 nucleotides Nature Genet 1557ndash61
Van der Giezen M Slotboom D J Horner D S DyalP L Harding M Xue G P Embley T M amp KunjiE R S 2002 Conserved properties of hydrogenosomal andmitochondrial ADPATP carriers a common origin for bothorganelles EMBO J 21 572ndash579
Von Heijne G 1986 Why mitochondria need a genome FEBSLett 198 1ndash4
Westphal S Soll J amp Vothknecht U C 2001 A vesicle trans-port system inside chloroplasts FEBS Lett 506 257ndash261
Whitehouse H L K 1969 Towards an understanding of themechanism of heredity London Edward Arnold Ltd
Williams B A P Hirt R P Lucocq J M amp Embley T M2002 A mitochondrial remnant in the microsporidian Trachi-pleistophora hominis Nature 418 865ndash869
Wilson R J (and 10 others) 1996a Complete gene map of theplastid-like DNA of the malaria parasite Plasmodium falcipa-rum J Mol Biol 261 155ndash172
Wilson S B Davidson G S Thomson L M amp PearsonC K 1996b Redox control of RNA synthesis in potato mito-chondria Eur J Biochem 242 81ndash85
Winkler H H amp Neuhaus H E 1999 Non-mitochondrialATP transport Trends Biochem Sci 24 64ndash68
Wolfe K H Morden C W amp Palmer J D 1992 Small sin-gle-copy region of plastid DNA in the non-photosyntheticangiosperm Epifagus virginiana contains only two genes Dif-ferences among dicots monocots and bryophytes in geneorganization at a non-bioenergetic locus J Mol Biol 22395ndash104
Wollman F A Minai L amp Nechushtai R 1999 The biogen-esis and assembly of photosynthetic proteins in thylakoidmembranes Biochim Biophys Acta Bioenerg 1411 21ndash85
Yeliseev A A amp Kaplan S 2000 TspO of Rhodobactersphaeroides a structural and functional model for the mam-
Genomes in mitochondria and chloroplasts J F Allen 37
malian peripheral benzodiazepine receptor J Biol Chem275 5657ndash5667
Zerges W 2002 Does complexity constrain organelle evol-ution Trends Plant Sci 7 175ndash182
Zhang Z D Green B R amp Cavalier-Smith T 1999 Singlegene circles in dinoflagellate chloroplast genomes Nature400 155ndash159
Zhang Z D Cavalier-Smith T amp Green B R 2001 A fam-ily of selfish minicircular chromosomes with jumbled chloro-plast gene fragments from a dinoflagellate Mol Biol Evol18 1558ndash1565
Zouni A Witt H T Kern J Fromme P Krauss NSaenger W amp Orth P 2001 Crystal structure of photosys-tem II from Synechococcus elongatus at 38 acngstrom resol-ution Nature 409 739ndash743
DiscussionD S Horner (Dipartimento di Fisiologia e Biochimica
Generali University of Milan Milan Italy) Given thatthere are components of these membrane-bound energy-transduction pathways that are consistently nuclear enco-ded not the key ones you pointed out but others do youexpect perhaps a slower perhaps a more crude but yet aredox communication between the organelle andexpression of nuclear-encoded genes
J F Allen There is and there is no question about itI think that Bogoradrsquos lock-in hypothesis has somethingto say here and there are other people who say it is all todo with assembling multi-subunit complexes If you havecontrol over the initial construction of the core aroundwhich the whole complex is eventually built then redoxcontrol or some sort of control over synthesis of that corewill control construction of the whole thing eventuallyand I think that may well be largely correct I think it ispossible in principle for nuclear genes to make precursorsaccording to redox instructions but it is not really soimportant that those precursors are here and now presentin exactly the right quantities because the precursors canto a limited extent hang around and wait Actually thereis proteolytic degradation of unattached precursors and soon but I think the fact that these chloroplast and mito-chondrially DNA-encoded subunits are at the core ofthese energy-transducing complexes is the crucial issue
T Cavalier-Smith (Department of Zoology University ofOxford Oxford UK ) In the case of mitochondria the sub-set of genes that remains is much more variable than youimplied It runs from three proteins in the minimal casesup to I think over 90 That is a big difference and manyof the genes in different lineages of protists that remainhave nothing to do with redox control they are things likeribosomal proteins and so on So I think that even thoughwhat you say about redox control of certain genes mayactually be an important thing it is not the fundamentaloverall reason why the genomes are maintained It may bepart of the explanation for why certain genes are alwaysincluded in that subset but even in mitochondria I thinkthat cytochrome b is the only one that is always presentSo I think that both this great diversity of what has beenretained in different lineages and the nucleomorph caseindicate that evolution was able to do one thing in onelineage 500 million years ago and never got around todoing it for 500 million years in another lineage so thereis a lot more haphazardness and evolutionary accidentsand contingency than a straight functional interpretation
Phil Trans R Soc Lond B (2003)
there is a lot of just sheer lsquohappenedndashorndashdidnrsquotndashhappenrsquoor carryover
J F Allen I was not implying anything about the rangeof mitochondrial genomes I have a limited amount oftime so I just talked about what I could talk about withinthat limit Of course there will be a range and this willdepend upon the physiology and bioenergetics of theparticular organism But what I am trying to do is to seewhat the common properties of the irreducible core areof components that are always encoded in the organellesand cytochrome b is a perfect example because that reallyis the thing that binds these two all-important quinoneswithout which there is no Q cycle without which there isno proton pumping and bioenergetics and in Plasmodiumwhere the mitochondrion is reduced to this bare mini-mum there is still a mitochondrial gene for cytochromeb Now my argument about the ribosomal proteins andall the rest of it is that in order to be able to exert thisall-important redox control over gene expression of certaincore components you need an in situ genetic system andthis can vary in composition for reasons that I cannotdescribe Maybe you need RNA polymerase or maybeyou can import RNA polymerase In plastids you haveboth of these options You are going to need ribosomestransfer RNAs and the rest of the apparatus of geneexpression and that needs to be in situ but we are stillleft with the question why does there have to be any gen-etic system at all The mature tobacco or spinach chloro-plast seems to have an entire genetic system just to makeone protein D1 which is all it ever needs to resynthesizeI think that this is a clue It is worth having this very unec-onomical crazy system because there is an overridingrequirement to connect electron transfer with geneexpression directly and unambiguouslymdashwithout recourseto a bureaucracy if you like which is what I think thenucleus is
K Willison (Institute of Cancer Research Chester BeattyLaboratories London UK ) These photosynthetic reactioncomplexes are a sort of baroque mixture of polypeptidechains and cofactors and my understanding is that someof them in high photon flux can turn over within say 30minutes so I am wondering if there is a correlationbetween gene retention and the turnover rates of some ofthe components and the complexes it seems to me thatthat could really be a limiting factor and the reason thatthe proteins had to be encoded locally in order to get thecomplexes to be assembled rapidly
J F Allen I think that is right You can only alter thesteady-state concentration of a protein complex by affect-ing the synthesis and breakdown and therefore turnoverThere is an extreme example the D1 protein encoded bythe psbA gene of chloroplasts which is always in the chlor-oplast genome without exception High light will tend todegrade this protein and when the high light goes awaythere is an absolute need very quickly to bring it back upagain I think that is a very good example Whether thisargument applies to genes such as cytochrome oxidase Ido not know
C J Leaver (Department of Plant Sciences University ofOxford Oxford UK ) Your experiments tend to relatemainly to the gene expression side What about proteinRNA turnover the whole concept of redox control ofthatmdashdo we know anything
38 J F Allen Genomes in mitochondria and chloroplasts
J F Allen Well quite a lot is known about chloro-plasts There is redox control of messenger RNA stabilityfor this same D1 protein which has been described byStephen Mayfield and his co-workers The evidence is thatthe site of redox control is ferredoxin via thioredoxin andthe implication is that thioredoxin tells whether non-cyclicelectron transport is working at all When it is workingthere is a signal that says lsquoOK now continue with all theprotein synthesisrsquo It is not particularly specific but moreof a global effect I think this is what Mayfield argues butof course this is not inconsistent with my argument I haveconcentrated on quinone pools and medium-potentialredox regulatory control because that is where I can seesignals about the dangerous consequences of getting itwrong but there is control of other levels of geneexpression In mitochondria I am not so sure I wouldbounce the question back to you on that one
W Martin (Institute of Botany III Heinrich-Heine Univ-ersitat Dusseldorf Dusseldorf Germany) The nucleomorphgenome was cited as evidence in conflict with your modelbut is that true The nucleomorph genome is not in abioenergetic organelle in that sense It resides in the cyto-sol so does the size of the nucleomorph genome workagainst your model Is it really evidence against yourmodel
J F Allen I do not know why should it be evidenceI cannot explain why the nucleomorph has a genomeThat is why I clutched at the straw that it is storage fornucleic acid precursors
Phil Trans R Soc Lond B (2003)
T Cavalier-Smith The reason I introduced the nucleo-morph is not to say it is direct evidence against yourhypothesis but to show that there probably was no singlefunctional reason for the retention of every gene in everyorganelle It may be that there are specific physiologicalreasons for the retention of certain genes and thereforeyou might be right about a subset of genes but I verymuch doubt if there are specific physiological reasons forthe retention of every single gene that is retained differen-tially because odd things happen in evolution Somethings are easy and sometimes things are difficult Evol-ution is history
J F Allen Of course I agree with that I do not thinkI can explain everything but I do think I can explainsomething If I have done that then I am content
GLOSSARY
AAC ADPndashATP carrierCES control by epistasy of synthesisCORR co-location for redox regulationLHC light-harvesting complexPCC Pasteur culture collectionORF open reading frameRUBISCO ribulose-15-bisphosphate carboxylasendashoxygenaseTAT twin-arginine translocase
Visit httpwwwpubsroyalsocacuk to see an electronic appendix tothis paper

Genomes in mitochondria and chloroplasts J F Allen 23
components of the chloroplast and mitochondrial geneticsystems themselves These genes comprise a secondarysubset of organellar genes genetic system genes There isgenerally no requirement for redox control of expressionof genes in this secondary subset though retention ofredox control of genetic system genes may in some casesallow amplification of redox signals acting upon genes inthe primary subset Retention of genes of the secondarysubset (genetic system genes) is necessary for the oper-ation of redox control of expression of genes in the primarysubset (bioenergetic genes) Without genes in the primarysubset the function of genes in the secondary subset iseventually lost and organelles lose their genomes
This hypothesis of co-location for redox regulation ofgene expression CORR was first outlined in a review(Allen 1992) The hypothesis was then systematically putforward in two articles (Allen 1993ab) developed(Allen amp Raven 1996) and has been independentlyreviewed (Race et al 1999) Some of its predictions havebeen tested experimentally as described later
Figure 3 illustrates the general idea of redox regulatorycontrol operating on mitochondrial genes to give the dis-tribution of sites of protein synthesis seen in figure 1 Fig-ure 4 shows the corresponding predicted pattern ofchloroplast redox regulatory control giving rise to the dis-tribution of sites of protein synthesis seen in figure 2Table 1 shows a functional classification of cyanobacterialgenes retained in 16 completely sequenced chloroplastgenomes This analysis (table 1) reveals a strong selectiveretention of genes for subunits of complexes involved inrespiration and photosynthesis and especially in chloro-plast photosystems I and II The pattern (table 1) seemsto be broadly consistent with CORR (figure 4) and incon-sistent with alternative hypotheses (see sect 4) It is reason-able to view photosynthesis as a process where preciserapid and coordinated redox regulatory control of geneexpression repays the cost of maintaining a chloroplastgenetic system
The CORR hypothesis is based on several propositionsassumptions or principles 10 of which are stated explicitlyand listed below Some of the principles are already generallyaccepted some are shared with competing hypothesesand some make direct experimental predictions Howeverother underlying principles of CORR are themselves generalprinciples or subsidiary hypotheses for which no directproof is available but for which predictions are made andpotentially falsifying observations or results can be imagined
The 10 principles that form the basis of the CORRhypothesis follow and each will be critically discussed inthe light of available evidence
(a) The principle of endosymbiotic originAs now generally agreed bioenergetic organelles have
evolved from free-living bacteriaAlthough once controversial (Mahler amp Raff 1975) this
principle will be taken as an axiom and not consideredfurther here The reader is referred to Gray amp Doolittle(1982) Gray (1992) Martin (1999) Martin et al (2001)to sect 1 and to accompanying articles in this journal issue
(b) The principle of unselective gene transferGene transfer between the symbiont or organelle and the cell
nucleus may occur in either direction and is not selective forparticular genes
Phil Trans R Soc Lond B (2003)
This principle takes the form of a testable assertion thatseems to be consistent with current knowledge Nothingcertain seems to be known about the mechanism of intra-cellular gene relocation (Palmer 1997 Martin amp Herrm-ann 1998 Martin et al 1998 Adams et al 2000 Henze ampMartin 2001 Millen et al 2001 Rujan amp Martin 2001Daley et al 2002) Plausible scenarios can be imagined onthe basis of the classical genetical phenomena of trans-formation transduction and transfection However asthere is no agreed mechanism there is no basis forasserting a mechanistic argument for selectivity Genetransfer between the organelle and nucleus seems to be aspecial case of lateral or horizontal gene transfer(Doolittle 1999) and no mechanistic basis seems to becurrently available for selectivity in what may be trans-ferred It should also be pointed out that lateral genetransfer between the organelle and nucleus does not haveto occur within a single cell nor within a multicellularindividual (although here any vertically transmitted genemust be incorporated into the germ line) for the outcomeof copying genes to novel compartments to be securedThus any plausible mechanism for heterologous recombi-nation may apply equally to gene transfer between com-partments Mitochondrial departures from the lsquouniversalrsquogenetic code and biased codon usage in chloroplasts (seelater) may be of selective value in that a barrier is therebyput in place to expression of genes in the lsquowrongrsquo compart-ment This barrier is unlikely to be insuperable especiallyif the lsquowrongrsquo compartment turns out actually to be a bet-ter location Impeded relocation might confer stabilitywithout being a primary reason for the existence of cyto-plasmic genomes
(c) The principle of unselective protein importThere is no barrier to the successful import of any precursor
protein nor to its processing and assembly into a functionalmature form
As with sect 3b this principle takes the form of a testableassertion that seems to be consistent with current knowl-edge The principle is discussed in more detail in sect 4because its negation (some proteins cannot be imported)has previously been put forward in itself as a reason forthe retention of cytoplasmic genomes Good hypothesesare easily falsified and they therefore forbid certain obser-vations The principle of unselective protein import for-bids the existence of a protein even a synthetic one thatcannot be imported by some means as a precursor
(d) The principle of continuity of redox controlDirect redox control of expression of certain genes was present
in the bacterial progenitors of chloroplasts and mitochondriaand was vital for selectively advantageous cell function beforeduring and after the transition from bacterium to organelle Themechanisms of this control have been conserved
This principle has been tested experimentally in the pastfew years and seems to be corroborated by an emergingpicture of redox signalling in bacteria and organellesExperiments carried out in vitro on the products of proteinsynthesis (Allen et al 1995 Galvis et al 1998) or RNAsynthesis (Pearson et al 1993 Konstantinov et al 1995Wilson et al 1996b) are consistent with a general phenom-enon of redox dependency of gene expression in bioener-getic organelles Redox control of chloroplast transcription
24 J F Allen Genomes in mitochondria and chloroplasts
nucleus cytosol
O2
H2O
redox regulation
N-phasemitochondrial matrix
Figure 3 Gene expression and principal pathways of biosynthesis of subunits of protein complexes involved in respiration andoxidative phosphorylation in animal mitochondria As in figure 1 reddish-brown DNA RNA and protein subunits are locatedand synthesized in the mitochondrial matrix light brown protein subunits have genes (also light brown) in the nucleus andare imported from the cytosol as precursors White genes and ribosomal and protein subunits are nuclearndashcytoplasmic and ofarchaebacterial origin Reddish-brown and light-brown genes and ribosomal and protein subunits are of bacterial origin Themajor variable environmental input is oxygen (blue) The essence of the CORR hypothesis is that it is beyond the ability ofthe nuclearndashcytoplasmic system to respond rapidly and directly to changes in oxygen concentration or partial pressure and soredox regulation of gene expression (red arrows) has been retained from the ancestral endosymbiont This redox regulationrequires co-location of certain genes with their gene products within the mitochondrion
nucleus cytosol
CH2O
CO2
redox regulation
N-phasechloroplast stroma
light light
Figure 4 Gene expression and principal pathways of biosynthesis of subunits of protein complexes involved inphotosynthesis in chloroplasts As in figure 2 green DNA RNA and protein subunits are located and synthesized in thechloroplast light brown protein subunits have genes (also light brown) in the nucleus and are imported from the cytosolas precursors White genes and ribosomal and protein subunits are nuclearndashcytoplasmic and of archaebacterial originGreen and light-brown genes and ribosomal and protein subunits are of bacterial origin The major environmental inputsare light (orange arrows) and carbon dioxide (blue) The essence of the CORR hypothesis is that it is beyond the abilityof the nuclearndashcytoplasmic system to respond rapidly and directly to changes in light and carbon dioxide and so redoxregulation of gene expression (red arrows) has been retained from the ancestral endosymbiont This redox regulationrequires co-location of certain genes with their gene products within the chloroplast
Phil Trans R Soc Lond B (2003)
Genomes in mitochondria and chloroplasts J F Allen 25
has been demonstrated (Pfannschmidt et al 1999abAllen amp Pfannschmidt 2000 Tullberg et al 2000) and wasprompted by this principle
The level of redox control of chloroplast transcriptionestablished unequivocally so far is that of plastoquinonean electron and hydrogen atom carrier located strategicallybetween photosystems I and II of photosynthesis Thedirections in which the effects are exerted are functionallyintelligible reduction of plastoquinone increases photosy-stem I transcription and decreases photosystem IItranscription oxidation of plastoquinone decreases photo-system I transcription and increases photosystem II tran-scription Plastoquinone in normal photosynthesis isreduced by the action of photosystem II and oxidized bythat of photosystem I Therefore redox control of chloro-plast gene expression provides a compensatory response(Pfannschmidt et al 1999a) and adds a previously mis-sing part of an explanation for the ability of plants andalgae to adjust the ratio of the two photosystems inresponse to changes in light quality (Chow et al 1990)By the same argument a functionally equivalent post-translational mechanism of redox control of light-harvesting function by photosystems I and II may havebeen conserved during the transition from prokaryotes toeukaryotes (Allen 1992 1995 Allen amp Nilsson 1997Allen amp Race 2002)
The precedent of chloroplast redox control of transcrip-tion (Pfannschmidt et al 1999ab) taken together with theprinciple in sect 3d predicts plastoquinone redox control ofcyanobacterial reaction centre gene transcription and thisis borne out by experiment (El Bissati amp Kirilovsky2001) This complementary self-adjusting transcriptionalresponse (Fujita 1997 Murakami et al 1997 El Bissati ampKirilovsky 2001) indicates strongly that redox control ofphotosynthetic reaction centre gene transcription waspresent in the proto-chloroplast Thus the mechanism ofregulation is likely to have been conserved throughout theevolutionary transition from bacterial symbiont to cyto-plasmic organelle A conserved mechanism of redox regu-lation of transcription in photosynthetic prokaryotes(Unden amp Bongaerts 1997 Bauer et al 1999) and chloro-plasts (Allen 1993c Pfannschmidt et al 1999b) is pre-dicted if evolutionary continuity of the process has beenmaintained
Thus apart from being consistent with experimentaldata the principle of continuity of redox control has beenof heuristic value in pointing to the underlying bacterialcharacter previously unsuspected in the control of geneexpression especially of transcription in chloroplasts ofhigher plants Other possible precedents for redox signal-ling components of cytoplasmic organelles include the prr(Eraso amp Kaplan 2000 2002 Oh amp Kaplan 2000) andreg (Bauer et al 1999) redox regulatory systems of purplenon-sulphur bacteria The TspO lsquooxygen sensorrsquo of Rho-dobacter sphaeroides is present in mammalian mitochondriaas a protein known there as a benzodiazepine receptorand each protein will complement loss of the other(Yeliseev amp Kaplan 2000) As outlined in sect 3j genes forsuch components are to be expected primarily in nucleargenomes This independent evolutionary hypothesis (sect 3d)would be strengthened still further by a demonstration ofone or more decisive redox regulatory links shared by pho-tosynthetic prokaryotes and all eukaryotes with mitochon-
Phil Trans R Soc Lond B (2003)
dria and where present with chloroplasts (Bauer et al2003)
(e) The principle of the selective value of redoxcontrol
For each gene under redox control (sect 3d) it is selectivelyadvantageous for that gene to be retained and expressed onlywithin the organelle
The twin selective advantages of redox control are likelyto be energetic efficiency and the suppression of the harm-ful side effects of electron transport operating on thewrong substrates No direct experimental support for thisprinciple is available and equally there is no evidenceagainst it Allotopic mutants in which genes have beenexperimentally relocated from organelles to the nucleushave been produced in yeast higher plants (Kanevski ampMaliga 1994) and human cells (Manfredi et al 2002) andare predicted by CORR to be poor adapters to environ-mental change Some authors believe there is potentialtherapeutic value in allotopic transformation movingmitochondrial genomes to the nucleus (Kanevski ampMaliga 1994 de Grey 2000) and predict the possibilityof r2 human cell lines in the foreseeable future (de Grey2000) This principle also has implications for ageing andthe evolution of separate sexes (Allen 1996) and possibleconsequences for development (Blackstone 1999 20002001 Harvey et al 2002) and somatic cloning in animals(Allen amp Allen 1999)
(f ) The principle of the selective value of nuclearlocation for genes not under redox control
For each bacterial gene that survives and is not under redoxcontrol it is selectively advantageous for that gene to be locatedin the nucleus and expressed only in the nucleus and cytosolIf the mature gene product functions in chloroplasts or mito-chondria the gene is first expressed in the form of a precursorfor import
There are several possible reasons for the selectiveadvantage of nuclear location of genes for organellar pro-teins One is the decreased probability of mutation arisingfrom free radical by-products of lsquoincorrectrsquo electron trans-fers (Raven et al 1994ab Allen amp Raven 1996) Therewould seem to be obvious economies in resource use andadvantages in coordination of gene expression if genes arekept in one compartment One might say that the bipartiteor tripartite genetic system of eukaryotic cells is untidyand inefficient
Another suggestion is that organellar genes do notundergo recombination and are present in relatively smallcopy numbers so that disadvantageous mutations willspread relatively quickly through a clonal population oforganelles Incorporation of functional gene copiesinstead into the nuclear genome may then be an irrevers-ible step analogous to the click of a ratchet (Saccone et al2000) There are examples of genes shared between allthree compartments (Stern amp Lonsdale 1982 Lonsdale etal 1983)
The cost of retaining an organellar genetic system is notincreased appreciably when additional genes are retainedin organelles if just one gene is retained then the cost ofa cytoplasmic genetic system is incurred and there may berelatively little extra cost for each additional gene retained(Allen amp Raven 1996) For a functional explanation of the
26 J F Allen Genomes in mitochondria and chloroplasts
existence of chloroplast and mitochondrial genomes thefocus of attention should therefore be on the relativelyconstant primary subset of bioenergetic genes (sect 3e)
(g) The principle of contemporary operation ofselection on gene location
For any species the distribution of genes between organelle(by the principle in sect 3e) and nucleus (by the principle in sect 3f)is the result of selective forces that continue to operate
This principle is essentially a statement contradictingboth the lsquofrozen accidentrsquo hypothesis and the hypothesisthat gene relocation is progressive predictable and stillincomplete (sect 4) In support of this principle it is becom-ing increasingly clear that mitochondria that lose theirfunction in aerobic respiration also lose their genomes asseen in the relict mitochondria of microsporidia(Embley amp Martin 1998 Van der Giezen et al 2002) andthe lsquomitosomersquo of Entamoeba histolytica (Tovar et al1999) In chloroplasts loss of photosynthesis results inloss of photosynthetic genes The examples of Epifagus (aparasitic higher plant) (Ems et al 1995 Wolfe et al 1992)and the residual apicomplexan plastids of Plasmodium(Wilson et al 1996a) and Toxoplasma where some plas-tidic genetic system is retained mean that some of theirgene products must be subject to redox regulation of geneexpression if this principle is correct Experimental test ofthis principle is possible using allotopic mutants as forthe predictions of the principle in sect 3f The evidence thatthe apicoplast (Wilson et al 1996a) is a plastid is largelygenetic the residual genome resembles that of photosyn-thetic plastids (chloroplasts) in gene organization ratherthan in content A plastid that has completely lost its gen-ome by virtue of having lost photosynthesis and all otherprocesses requiring redox regulation might be difficult todistinguish from an organelle derived in a similar way froma mitochondrion The persistence of targeted importedchaperones Hsp70 (Tovar et al 1999 Williams et al2002) and cp60 (Roger et al 1998) in residual organellesmight not distinguish a plastid ancestry from a mitochon-drial one
(h) The principle of primary involvement inenergy transduction
Those genes for which redox control is always vital to cellfunction have gene products involved in or closely connectedwith primary electron transfer These genes are always con-tained within the organelle
This principle would be violated and the CORRhypothesis refuted if any photosynthetic reaction centrecore subunit or respiratory chain subunit currentlyorganelle-encoded is found that functions in primary vec-torial electron transport and is nevertheless encoded in thenucleus and synthesized cytosolically as a precursor forimport all without any selective cost to the individualFrom the chloroplast transcriptional experiments ofPfannschmidt et al (1999ab) the chloroplast geneswhose transcription is regulated by the plastoquinoneredox state include psaAB (for photosystem I reactioncentre core polypeptides) psbA (for the photosystem IIpolypeptide D1) and rbcL (for the large subunit of RUB-ISCO interestingly following the same pattern of redoxcontrolmdashupregulated by oxidized plastoquinonemdashaspsbA) Transcription of the chloroplast genes petA
Phil Trans R Soc Lond B (2003)
(cytochrome f ) trnG (glycine tRNA) and rpoB (RNApolymerase) did not respond to plastoquinone redox statepetA for cytochrome f is predicted by this principle to beregulated at a different level of redox control or if at plas-toquinone at a different stage in gene expression Thisprediction might also apply to trnG and rpoB though thesequalify as members of the secondary subset that of geneticsystem genes to which the principle in sect 3i applies
(i) The principle of secondary involvement inenergy transduction
Genes whose products contribute to the organelle genetic sys-tem itself or whose products are associated with secondaryevents in energy transduction may be contained in theorganelle in one group of organisms but not in anotherdepending on the physiology and biochemistry of photosynthesisand respiration in the species concerned
Even phylogenetically moderately related species(within kingdoms) show differences in the location ofsome genes for example the ATP synthase subunits ofNeurospora and Saccharomyces (Attardi amp Schatz 1988)The CORR hypothesis also does not explicitly account forthe wide disparities in the distribution of genetic systemgenes for example the almost complete import of tRNAsin Chlamydomonas (GoldschmidtClermont 1998) Trans-port of nucleic acids across membranes is uncommon andwhy some RNAs should be organelle-encoded in somespecies but nuclearly encoded in others is not currentlyunderstood The only suggestion that can be made alongthe lines of CORR is that in some genetic systems genesmay need to be under the control of a regulatory systemthat responds to a signal arising within the organelle itselfThis signal could be the primary signal of altered redoxstate or some secondary signal Among the latter might befound signals that themselves report on the current stateof gene expression another piece of information that thenucleus might not have at its immediate disposal Suchsecondary signals could result at least in part fromCORR though other developmental and environmentalinputs may be imagined and would not detract fromCORR unless it could be shown that they provided a suf-ficient reason for the retention of the genetic system asa whole
(j) The principle of the nuclear encoding of redoxsignalling components
Components of the redox-signalling pathways upon whichCORR depends are themselves not involved in primary electrontransfer and so their genes have been relocated to the nucleusin accordance with sect 3f
This principle states that the redox signalling pathwaysthat are required for CORR should not themselves beexpected to utilize components whose biosynthesis mustbe under direct redox control Such components shouldtherefore fall into the major category of organellar pro-teins and be imported as precursors from the cytosol inline with the principle in sect 3f However the principleabove might be need to be modified in cases where ampli-fication of the redox signal is useful to the cell In thebacterial ntr system which was a precedent for CORR(Harrison et al 1990 Allen 1992) the histidine kinasentrB and aspartate response regulator ntrC form part ofthe gln operon whose transcription they control Odontella
Genomes in mitochondria and chloroplasts J F Allen 27
and Porphyra have plastids that encode more proteins thanthose of most chlorophytes (Stoebe et al 1998 Turmel etal 1999) and these proteins include two-component sig-nal transduction proteins prr and ycf27 (Stoebe et al1998) Extreme or extremely rapid environmental fluctu-ations might be catered for by amplification of signals thatare transmitted by signalling pathways whose componentsare under their own control It is not known whether ycf27and prr are in operons whose promoters their gene pro-ducts control It is also uncertain whether ycf27 is a redoxsignalling component and its distribution at least in itsplastidic location seems to be correlated with the presenceof a phycobilin-based light-harvesting system as found inred algae and cyanobacteria (Ashby amp Mullineaux1999ab Ashby et al 2002) The best guess of the originalstatement of the CORR hypothesis (Allen 1992 1993ac)was that bacterial two-component systems (Iuchi amp Lin1992 Allen 1993c Grebe amp Stock 1999) provide a plaus-ible means of exerting redox regulatory control over mito-chondrial and chloroplast gene expression andsubsequent research confirms the presence of two-compo-nent genes in genomes of Arabidopsis and yeast SeveralArabidopsis two-component genes are predicted to be tar-geted to mitochondria and chloroplasts (Forsberg et al2001) These must be considered as prime candidates forcomponents involved in redox regulation of chloroplasttranscription seen in the experiments of Pfannschmidt etal (1999a 2001a) The presumably homologous regulat-ory process serving to control photosystem stoichiometryin cyanobacteria the prr system (Li amp Sherman 2000) isnow characterized as a two-component system involvinga redox sensor and redox response regulator as originallydefined (Allen 1992 1993ac)
4 CONTRASTING HYPOTHESES CONCERNINGTHE RETENTION OF CYTOPLASMIC GENOMES
CORR is a recent addition to explanations that havebeen put forward to account for the persistence of gen-omes in organelles These models and their underlyingmechanisms are now discussed and compared
(a) There is no reason that is just how it isOne reaction to the problem of cytoplasmic genetic sys-
tems (sect 2) is that there is no problem and no particularreason for their retention However there is a very widephylogenetic distribution of a common core or primarysubset of organellar-encoded genes and one which haswithstood considerable genomic reorganization over evol-utionary time This constancy is seen for example in thesame genes being present in the chloroplasts of landplants with multiple copies of a single circular chloroplastchromosome and in dinoflagellate chloroplasts with acollection of different DNA minicircles each with essen-tially one gene (Zhang et al 1999 2001 Barbrook et al2001) Another suggested explanation of the retention ofmitochondrial and chloroplast genomes is simply torestate the endosymbiont hypothesis and to point out thatchloroplast and mitochondrial genes lsquocame inrsquo with thebacterial endosymbiont This suggestion however doesnot address the question of why some genes were retainedwhereas others were not It is also now clear that mostgenes that came in and still survive have nuclear and cyto-
Phil Trans R Soc Lond B (2003)
solic gene products whereas many chloroplast and mito-chondria proteins had a eukaryotic origin (Salzberg et al2001 Martin amp Russell 2003) The odds against a con-served pattern in gene location (eg table 1) arising bychance must be astronomical Thus there is a reason forthe retention of organellar genomes and a reason for theretention of the same primary subset of genes Becauseevolution proceeds by natural selection of chance vari-ation lsquoreasonrsquo in this context requires identification of thefunction of the location of a gene that is the selectivevalue of the phenotype produced by that location Evol-ution sometimes retains apparently functionless relics ofpreviously useful features but not for long especially ifthere is a cost
(b) The lock-in hypothesisThe lsquolock-inrsquo hypothesis of Bogorad (1975) was pro-
posed independently of the endosymbiont hypothesis anddoes not depend upon it According to the lock-in hypoth-esis core components of multi-subunit complexes must besynthesized de novo in the correct cellular compartmentotherwise photosystems or respiratory chain complexesfor example might assemble in the plasma membranevacuolar membrane or endoplasmic reticulum The lock-in hypothesis made no clear prediction about the conti-nuity of cytoplasmic genetic systems even though cyto-plasmic uniparental non-Mendelian inheritance for bothmitochondria (Roodyn amp Wilkie 1968) and plastids(Kirk amp Tilney-Bassett 1978) made it likely that organellarDNA the chondriome and the plastome replicated inde-pendently of nuclear DNA (Whitehouse 1969)
Mechanisms of protein import and targeting (Schatz1998 Koehler et al 1999 Leuenberger et al 1999 Gab-riel et al 2001 Jarvis amp Soll 2001 Pfanner amp Geissler2001 Duby amp Boutry 2002 Pfanner amp Wiedemann 2002)now effectively answer the problem of specifying the cor-rect locations for membrane protein complexes for whichthe lock-in hypothesis was proposed as a solution How-ever regulation of synthesis of an entire functional multi-subunit complex by control of synthesis of individual sub-units is still an important principle This seems to be afeature of the lsquoCESrsquo model (control by epistasy ofsynthesis) of Wollman (Wollman et al 1999 Eberhard etal 2002) which envisages nuclear control as all-embracingwith organellar genetic systems doing as it were whatthey are told In the hypothesis of CORR propoundedhere CES is equally important but the essence of CORRis that the organellar genetic system alone knows what arethe immediate and most important signals of environmen-tal change The organelle may keep the nucleus informedbut this is a lower priority than initiating an appropriateresponse CES and CORR agree that assembly of multi-subunit complexes is a complex business and requirescoordinated control in which no step is rate-limiting butmany may be obligatory
(c) Going going gone the evolutionary process oftransfer of genes from organelles to the
nucleus is still underway but incompleteThis hypothesis envisages that for most eukaryotes we
are now witnessing the last stages in a slow evolutionarytransfer of genes to the nucleus and this transfer may beexpected to proceed to completion in what is in evol-
28 J F Allen Genomes in mitochondria and chloroplasts
utionary terms the near future (Palmer 1997 Gray 19992000 Gray et al 1999 Adams et al 2000 Daley et al2002) A clear and compelling statement of this widelyheld position is that of Herrmann amp Westhoff (2001 p10)
hellipthe transfer of geneshelliptells that the partite plant gen-ome is not in a phylogenetic equilibrium All availabledata suggest that the ultimate aim of genome restructur-ing in the plant cell as in the eukaryotic cell in generalis the elimination of genome compartmentation whileretaining physiological compartmentation
One serious problem with this hypothesis is the dis-tinctly non-random sample of genes that remain to betransferred A relatively constant subset of genes is foundin bioenergetically functional mitochondria from fungi tomammals and angiosperms as in photosynthetic plastids(chloroplasts) from unicellular algae to angiosperms Thishypothesis therefore requires an additional ad hocassumption that some gene transfers are inherently moreprobable than others (this too is widely supposed seelater) and thus that gene transfer events proceed in thesame sequence in quite independent lineages Thereseems to be no evidence for this and the order of geneloss from mitochondria and chloroplasts has been used toinfer phylogeny (Martin et al 1998 Adams et al 2000Millen et al 2001) lsquoGoing going gonersquo (Palmer 1997)is difficult to refute categorically but it does not seem tobe consistent with recent evidence For example chromo-some two of Arabidopsis thaliana (Lin et al 1999) has asingle insertion encompassing with about 99 sequencesimilarity the complete Arabidopsis mitochondrial genome(Unseld et al 1997 Marienfeld et al 1999 Stupar et al2001) Unless this single transfer is a freak occurrencethis finding strongly indicates that gene transfer occurswith relative ease (Adams et al 2000 Henze amp Martin2001 Rujan amp Martin 2001 Salzberg et al 2001) Inaddition there is experimental evidence that randomlygenerated polypeptides are mitochondrially imported athigh frequency (Lemire et al 1989) The hypothesis thatwe now see the same few stragglers in diverse organellargenomes is not impossible but should be regarded as alast resort perhaps applicable only in extreme cases suchas the greatly reduced apicomplexan plastomes and chon-driomes Even there the hypothesis carries with it an obli-gation to explain why organisms with few organellar geneshave lsquoevolvedrsquo faster than those with many
(d ) The frozen accidentWhat may be described as lsquothe frozen accident hypoth-
esisrsquo states that the evolutionary process of gene transferfrom organelle to nucleus was underway when somethinghappened that stopped it One example of the frozen acci-dent is a scenario (Von Heijne 1986) in which import intomitochondria of the precursor proteins arising from newlytranslocated genes proceeded for long enough after theprimordial endosymbiosis for most of the symbiont-derived genes to be moved to the cell nucleus The lsquoacci-dentrsquo that halted the process was the evolutionary originof exocytosis and protein secretion after which it becameimpossible for precursors properly destined for mitochon-drial import to be distinguished from those destined forexport from the cell As with the lsquolock-inrsquo hypothesis the
Phil Trans R Soc Lond B (2003)
lsquofrozen accidentrsquo seems incompatible with the remarkableprecision and specificity of protein targeting (Schatz 1998Koehler et al 1999 Leuenberger et al 1999 Gabriel etal 2001 Jarvis amp Soll 2001 Pfanner amp Geissler 2001Duby amp Boutry 2002 Pfanner amp Wiedemann 2002) Pro-tein sorting mechanisms do not seem to forbid controlledimport into any cellular compartment Another lsquoaccidentrsquothat may have impeded subsequent gene transfer in thecase of mitochondria is their departure from what is other-wise a universal genetic code Genetic code differences arediscussed later in the context of whether some genes can-not be moved
(e) Some proteins (with cofactors) cannot beimported
We may describe as the lsquounimportability hypothesisrsquo theproposal that some proteins are inherently unimportableand must therefore be synthesized de novo in the correctcompartment within the cell The unimportability hypoth-esis is most often proposed as its special case the hydro-phobicity hypothesis with hydrophobicity being thereason for unimportability
(i) HydrophobicityA commonly entertained notion is that proteins that are
encoded and synthesized within organelles comprise a pri-mary subset characterized by shared hydrophobicity allare intrinsic membrane proteins (Von Heijne 1986Popot amp de Vitry 1990 Claros et al 1995) As with theCORR hypothesis favoured here the primary subset thenrequires a secondary subset of genetic system componentsthe latter not themselves being hydrophobic but beingrequired for synthesis of the others Certainly the mito-chondrial inner membrane contains some intrinsic proteincomplexes whose subunits broadly satisfy the criterion ofhydrophobicity However the core subunits that are mito-chondrially encoded are also explained by the CORRhypothesis This would make hydrophobicity a require-ment for their function in vectorial electron and protontransfer and not a reason in itself for retention of theirgenes in the cytoplasm
The hydrophobicity hypothesis has some very clearcounter-examples in chloroplasts and cannot thereforestand if one wishes for a single principle that embracesboth types of bioenergetic organelle One clear counter-example is the large (L) subunit of the enzyme RUBISCOan abundant but entirely membrane-extrinsic protein ofca 55 kDa molecular mass The RUBISCO large subunitis generally chloroplast-encoded with the exception of thenuclearly encoded bacterial-type L subunit of L2 RUBI-SCO in peridinin-containing dinoflagellates (Morse et al1995) In most photosynthetic eukaryotes RUBISCO hasa chloroplast encoded large subunit and a nuclearly enco-ded small (S) subunit (Ellis 1984 Gatenby amp Ellis 1990)giving an L8S8 holoenzyme and this RUBISCO is widelytaken as a paradigm for the need for coordination ofnuclear and chloroplast gene expression (Rodermel 2001)
By complete contrast to the RUBISCO large subunitwhich is a chloroplast-encoded and water-soluble proteina second major violation of the hydrophobicity hypothesisis the existence of the universally nuclearly encoded butalways hydrophobic subunits of the chloroplast LHC IIand LHC I (Chitnis amp Thornber 1988) These apopro-
Genomes in mitochondria and chloroplasts J F Allen 29
teins of ca 27ndash30 kDa are quite water-insoluble and areusually fully extracted only in chloroformndashmethanol or inthe presence of detergents The Lhc gene family is diverse(Green amp Kuhlbrandt 1995) but most gene productsor Lhcp (light-harvesting chlorophyll protein) polypeptideshave three membrane-spanning hydrophobic a-helices(Kuhlbrandt 1994) It is interesting that three-helix Lhcproteins seem to be absent from extant cyanobacteria andchloroxybacteria (Green amp Kuhlbrandt 1995)
(ii) The five-helix rulePerhaps with the three-helix model of LHC II in mind
a variation of the hydrophobicity hypothesis is the empiri-cal lsquofive-helix rulersquo (Popot amp de Vitry 1990 Claros et al1995 Wollman et al 1999) which states that no intrinsicmembrane protein with five or more membrane-spanninghelices can be encoded in the nucleus Five is a good guessfor a threshold number of helices demanding organellar-encoding in the context of photosynthetic reaction centres(Heathcote et al 2002) where the earliest known struc-ture that of type II centres (Deisenhofer et al 1985) asin photosystem II (Zouni et al 2001) shows exactly fivehelices with type I centres containing 11 (Jordan et al2001) There are however many nuclearly encoded sub-units in both mitochondria and chloroplasts that havefewer than five membrane helices so the five-helix rulecan only describe a sufficient condition and not a neces-sary condition for nuclear encoding
It should also be noted that ADPndashATP carriers (AACs)are encoded in the nucleus AACs of the mitochondrialinner membrane have six transmembrane helices (van derGiezen et al 2002) while the those of the chloroplast innerenvelope (Winkler amp Neuhaus 1999) have 12 (Tjaden etal 1998)
With the counter-examples of nuclearly encoded AACswith six or 12 membrane-spanning helices many intrinsicmembrane proteins with fewer than five helices somenuclearly encoded and some not and with the RUBISCOlarge subunit as a major product of chloroplast geneexpression with no membrane domain at all the five-helixrule begins to seem rather like a case of special pleadingEven if there are indeed special mitigating circumstancesfor AACs (this argument is outlined later) the five-helixrule as with the hydrophobicity hypothesis of which it isa special case will disappear if it can be shown that pos-session of five or more membrane helices is required forsome other function such as that of CORR that deter-mines the organellar location of the gene concernedHydrophobicity is in some cases a feature of proteins thatare encoded in organelles but it is not in itself the issue
For chloroplasts the hydrophobicity hypothesis predictsretention of cyanobacterial genes in functional categoriesassociated with the chloroplast envelope and with trans-port just as much as with photosynthesis Table 1 showsthat this is not the case
A mechanism for the unimportability of hydrophobicproteins is rarely stated explicitly and it is rather left tothe imagination whether hydrophobic proteins might asit were get stuck halfway through the chloroplast envelopeor mitochondrial outer membrane whether transport can-not get started on a precursor protein that has alreadyspontaneously inserted into the membrane or whetherhydrophobic proteins will be unable to make the journey
Phil Trans R Soc Lond B (2003)
through the cytosol without precipitating there In thecytosol co-translational transport or membrane insertionwould in principle address the latter point but it seemsto be generally true that chloroplast and mitochondrialproteins are post-translationally inserted into the organel-lar outer membrane So the obvious ancillary questionabout unimportability is lsquowhy then cannot hydrophobicproteins be co-translationally imported into organellesrsquoThere is evidence in yeast for co-translational transportinto mitochondria of proteins thought to be derived fromthe original endosymbiont (Marc et al 2002) If there isany truth in hydrophobicity as a barrier to protein importthe CORR principle of unselective import (sect 3c) shouldbe easily refuted for example by a demonstration that noknown targeting presequence is capable of importing poly-phenylalanine encoded by polyU for in vitro import stud-ies
An additional proposal intended to account for theunimportability of some hydrophobic proteins and forimportability of others is suggested by A de Grey(personal communication) who argues that bacteriallyderived protein precursors imported into mitochondriacross the mitochondrial inner membrane completelybefore insertion into that membrane from the mitochon-drial matrix (the N-phase) This is held to be an obstacleto successful import of membrane-intrinsic proteins thatmight en route discharge the proton motive force By con-trast eukaryotically derived proteins can be inserted intothe mitochondrial inner membrane from the inter-mem-brane space (the P-phase) and then there is for somereason no similar uncoupling effect Thus adenylatetransporters or AACs can be nuclearly encoded whereascytochrome oxidase subunits for example cannot It isnot obvious why bacterially derived subunits such asthose of cytochrome oxidase did not acquire eukaryoticinsertion mechanisms Perhaps this acquisition is forbid-den by the need to conserve pre-existing routes ofassembly into multi-subunit complexes or perhaps theneed for insertion from the inside reflects a need to incor-porate a cofactor for example haem or ironndashsulphur clus-ters along the way
(iii) Unimportability of three dimensional structures andcofactors
Accumulating evidence on protein transport mech-anisms seems to make a specific obstacle to successfultransport of certain sorts of protein less and less plausibleespecially given the observation that precursors with N-terminal targeting sequences not mature functional poly-peptides themselves traverse membranes during biosynth-esis
Protein folding inside the living cell is invariably assistedby molecular chaperones (Gatenby amp Ellis 1990 Azem etal 1997 Komiya et al 1997 1990 Jackson-Constan et al2001) proteins that impede the formation of lsquoimproperrsquostructural interactions that will otherwise produce aggre-gation or other non-functional interactions Proteinimport mechanisms appear mostly to rely on specific mol-ecular chaperones for guided unfolding before membraneinsertion and refolding after translocation of the polypep-tide into its destined compartment (Gatenby amp Ellis 1990Azem et al 1997 Komiya et al 1997 Jackson-Constanet al 2001) Because polypeptides are mostly lsquothreadedrsquo
30 J F Allen Genomes in mitochondria and chloroplasts
through membranes no special constraint on transportshould be expected to arise from 3D structure or surfaceproperties of a protein
An exception to this unfolding rule may be providedby holoproteins containing cofactors that are transportedalong with the apoprotein presumably with a largely intact3D structure The lsquoTATrsquo (twin-arginine translocase) sys-tem (Bogsch et al 1998 Dalbey amp Robinson 1999 Berkset al 2000ab Robinson 2000 Robinson amp Bolhuis 2001Sargent et al 2002) works broadly along these linesthough its mechanism is unresolved TAT is relevant herebecause it adds transport of essentially complete holopro-teins to the cellrsquos repertoire of transport mechanisms andeven proteins that may not be allowed to unfold perhapsbecause of the toxicity of the free cofactor are not therebymade candidates for an unimportability rule One variantof the unimportability hypothesis is that transport canwork only by unfolding and dissociated cofactors may betoxic to the cell The bacterially derived TAT system(Bogsch et al 1998 Dalbey amp Robinson 1999 Berks etal 2000ab Sargent et al 2002) overcomes this argumentas would examples of cofactor-free polypeptides such as(C)Fo subunit c which are nevertheless generally encodedin organelles
The presence of a vesicular transport system in chloro-plasts (Westphal et al 2001) counts against the unim-portability hypothesis Vesicular transport between thechloroplast envelope and the thylakoid membrane shouldin principle allow any protein to be ferried across thechloroplast stroma Vesicles might allow transport inparticular of membrane proteins that would discharge theproton motive force if allowed to connect transiently theN and P aqueous phases on each side of a coupling mem-brane
Another observation that seems to count against unim-portability is the increased number of membrane barriersin plastids derived from secondary tertiary or higher-orderendosymbioses These plastids seem to have retained ifanything fewer genes rather than more as unimportabilitypredicts The nucleomorph the residual nucleus of theprimary host retains many genes for molecular chap-erones (Archibald et al 2001 Cavalier-Smith 2002) inagreement with the conclusion that protein import pro-ceeds by conserved and specialized mechanisms If eachnew import added to gain a higher-order endosymbiosisrequires additional targeting presequences then perhapsthere is an advantage in components of an import machin-ery not being imported themselves
It seems that unimportability is less easy an assumptionto make today than in previous years and the onus maynow be on its proponents to cite specific examples of pro-teins that can never be imported into chloroplasts or mito-chondria
(f ) Some genes cannot be moved(i) Differences in the mitochondrial genetic code
The genetic code is the specific relation between ribon-ucleotide triplets of codons and tRNAs that recognizethem and which thus determine which individual aminoacids if any will be incorporated into the growing poly-peptide chain Mitochondria show small departures fromthe otherwise universal genetic code (Attardi amp Schatz1988 Alberts et al 1994) UGA is a lsquostoprsquo codon in the
Phil Trans R Soc Lond B (2003)
universal code and in plant mitochondria but is the codonfor tryptophan in mitochondria of animals and fungiCUA means leucine everywhere except fungal mitochon-dria where it means threonine and AGA and AGU codefor arginine in the lsquouniversalrsquo code but serine in insectmitochondria and lsquostoprsquo in mammalian ones The signifi-cance of this heterogeneity in coding is most unlikely torest in the early origin of mitochondria as the lsquouniversalrsquocode is employed by bacteria and protists Furthermorethe diversity of departures from the lsquouniversalrsquo codeamong mitochondria of different eukaryotic lineages indi-cates they had independent origins A general rule formitochondrial genetic codes is that reliance on the thirdlsquowobblersquo position is decreased which allows all 20 proteinamino acids to be specified by fewer tRNAs (22 in animalmitochondria instead of 31 in the cytosol) This in turnmeans that mtDNA can specify more proteins for a givenlength of DNA or conversely that the mitochondrialgenome can be condensed or lsquostreamlinedrsquo Although thisarrangement does not explain the retention of mitochon-drial genomes and may be a secondary event in mitochon-drial evolution it almost certainly places an extra barrierin place of gene relocation As discussed previously how-ever there seems to be no single barrier that can explainthe retention of a relatively constant subset of mitochon-drial genes
There has clearly been a contraction of mitochondrialDNA especially in animals Animal mitochondrial DNAhas few if any non-coding sequences and animal mito-chondrial genetic codes require fewer tRNAs than the sin-gle code operating in the cytosol (Gray 1999 Gray et al1999 Selosse et al 2001) These may be devices todecrease the total frequency of mutations per mitochon-drial genome assuming a constant frequency per unitlength of mitochondrial DNA It has been proposed thata genersquos removal to the nucleus decreases its susceptibilityto free radical mutagenesis (Raven et al 1994ab Allen ampRaven 1996) If this is the case then the non-standardcode may be a further indication of an overriding need tomaintain certain genes within the mitochondrion What-ever the reason for the retention of genes in organelles ithad better be a good one bioenergetic organelles areclearly the wrong place to keep genetic information andgo to extraordinary lengths to offset the consequences ofit being there
(ii) Molecular refugeesA phenomenon related to differences in the genetic code
is seen in chloroplasts This is a bias in codon usage Thelsquomolecular refugeesrsquo hypothesis (Howe et al 2000) sug-gests that nuclear copies of what were once chloroplastgenes escape constraints on codon usage and thus thatwhat distinguishes chloroplast-encoded proteins is thattheir amino-acid composition is unusual enough for thebias in codon usage not to apply It is difficult to see whatphysico-chemical properties might be uniquely shared bythe amino acids of chloroplast-encoded proteins and inaddition there seem to be no lsquoforbiddenrsquo amino-acid resi-due in chloroplasts As with the departure from the univer-sal genetic code in mitochondria is seems likely that abias in codon usage reflects some secondary property ofchloroplast genomes and is not the primary reason fortheir retention In any case any restriction on gene move-
Genomes in mitochondria and chloroplasts J F Allen 31
ment will act in the wrong direction to explain why chloro-plasts have retained genomes at all provided there is noresulting obstacle to a chloroplast protein being success-fully encoded as a precursor in the nucleus
(iii) Genes are copied and replicated not relocated in onestep
If complete relocation is envisaged as proceeding by thesimultaneous appearance of a gene in one compartmentand its deletion from another then it is possible to imaginethat whatever the vector is there may be selectivity ingene transfer (Henze amp Martin 2001) For example ifviral incorporation replication and recombination in bothorganelle and nucleus were the mechanism of relocationthen successful gene transfer might depend on the abilityof the virus to complete the process in one step This inturn might depend on some property of the gene or ofthe protein it encodes if expression occurs at any stage inthe process of relocation An assumption of the CORRhypothesis favoured here is that genes are not movedphysically but merely copied after which either the copyin the original location or the copy in the new locationmay be lost as a result of selection A useful analogy maybe the action of copying files between computer storagevolumes In this analogy the copy is identical to the orig-inal and either may subsequently be deleted without lossof information
Because any gene that is not usefully expressed willundergo no selection to maintain a functional allele theprevailing location will be the one in which the functionalcopy of the gene resides The problem of which gene sur-vives is then subsumed into the problem of which is theselectively advantageous location for it It seems reason-able to assume that there wasmdashand ismdashan interim periodin which homologous genes in different compartments areessentially in competition to see which functions best Asoutlined above there is no obvious obstacle for copyingDNA sequences within the cell The physico-chemicalproperties of DNA are fairly uniform and not directlydependent on nucleotide sequence though organelles likeintra-cellular parasites seem to prefer A and T to G andC The answer to the question lsquowhich gene location func-tions bestrsquo may have nothing to do with the gene itselfnor directly with the structure and function of its geneproduct Instead the answer may have everything to dowith the required regulatory interaction between the geneproduct and expression of the gene that produces it
(g) CORRThis hypothesis has been propounded earlier (sect 3) and
will be summarized as follows Vectorial electron and pro-ton transfer exerts regulatory control over expression ofgenes encoding proteins directly involved in or affectingredox poise This regulatory coupling is indispensable foradaptation to environmental changes in chemical activitiesof oxygen light and CO2 and requires co-location of thesegenes with their gene products Organelles lsquomake theirown decisionsrsquo on the basis of environmental changesaffecting redox state The hypothesis is abbreviatedCORR for co-location for redox regulation or more com-pletely lsquoco-location (of gene and gene product) for(evolutionary) continuity of redox regulation of geneexpressionrsquo
Phil Trans R Soc Lond B (2003)
5 SUMMARY OF THE CURRENT STATUS OF THEHYPOTHESIS OF CORR
Q Why do mitochondria and chloroplasts have theirown genetic systems
A CORR co-location of genes and gene products per-mits direct and autonomous redox regulation of geneexpression
(a) Objections to CORR(i) There is no regulation of gene expression in chloroplasts
and mitochondriaThis is still commonly stated but incorrect See the prin-
ciple in sect 3d
(ii) There is no regulation of gene expression in chloroplastsand mitochondria at the level of transcription
For chloroplasts there is now clear evidence for a func-tionally intelligible redox control of transcription(Pfannschmidt et al 1999a) This invalidates the previouswidely held assumption that all control of chloroplast geneexpression is post-transcriptional (Gruissem amp Tonkyn1993 GoldschmidtClermont 1998) The evidence formitochondria is less clear but redox control of transcrip-tion certainly cannot be ruled out and there are indi-cations that it may occur (Pearson et al 1993Konstantinov et al 1995 Wilson et al 1996b) It mustalso be restated that transcriptional control is not neces-sary for CORR but serves as a compelling example of aconserved typically bacterial mode of response that oper-ates still in eukaryotic cytoplasmic organelles thus illus-trating the principle of continuity of redox control (sect 3d)Post-transcriptional redox regulation (Danon amp Mayfield1994 Kim amp Mayfield 1997 Brown et al 2001 Treb-itsh amp Danon 2001) is consistent with CORR and thereis nothing to forbid it being a primary mechanism thatsatisfies most or indeed all of the formal requirements ofthe hypothesis in particular cases
(iii) There are no redox signalling components encoded inchloroplast or mitochondrial DNA
This is what CORR predicts See the principle in sect 3j
(iv) There is redox control of nuclear gene expressionThis is correct and many examples are now known
especially for chloroplasts (Escoubas et al 1995 Kovacs etal 2000 Pfannschmidt et al 2001ab Surpin et al 2002)Redox signals from photosynthetic or respiratory electrontransport chains are not at all prohibited by CORR Theessence of CORR is that there is a subset of organellarproteins for which there is an overriding requirement fora redox regulatory control that is initiated within theorganelle and whose effects are exerted by the simplestand most direct route This redox signalling is envisagedto take place without a requirement for prior consultationand coordination with what may be regarded as a nucleo-cytoplasmic bureaucracy
(v) Nuclear genes can be expressed as rapidly as chloroplastand mitochondrial genes
Zerges (2002) for example cites the equal times takenfor complete expression of nuclear and chloroplast genesin Chlamydomonas as evidence against the CORR hypoth-
32 J F Allen Genomes in mitochondria and chloroplasts
esis However the question is not one of the time takento express a gene but the time taken to alter the rate ofits expression in response to a (redox) signal originatingwithin the organelle An equally important additional fac-tor may be the autonomy of the signalling pathway thatis its independence from other inputs as described in theprevious paragraph One might say that mitochondria andchloroplasts require a free hand
(vi) There is no evidence to support this hypothesisThe hypothesis is rich in predictions (sectsect 3 and 6) and
those that have been tested stringently are in agreementwith experimental results None of the predictions of thehypothesis has been found to be incorrect and none of theunderlying principles (sect 3) has been disproved Furtherpredictions will be the subject of future experiments
(b) Evidence for CORR(i) Chloroplasts have selectively retained cyanobacterial genes
for primary events in photosynthesisSee table 1
(ii) There is regulation of gene expression in chloroplastsThere is transcriptional regulation of gene expression
in chloroplasts (sect 3d) The case for mitochondria is notyet proven
(iii) There is redox regulation of gene expression inchloroplasts
There is transcriptional redox regulation of geneexpression in chloroplasts (sect 3d) The case for mitochon-dria is not yet proven though see (Allen et al 1995 Kon-stantinov et al 1995 Galvis et al 1998)
(iv) There are no redox signalling components encoded inchloroplast or mitochondrial DNA
Correct but with possible exceptions discussed in sect 3j
(v) There are lsquobacterialrsquo redox signalling components ineukaryotes
See sect 3j
(vi) Further predictions DNA-array results and phenotypesof allotopic mutants
A systematic test of CORR might be to make an oxid-ized-minus-reduced gene expression profile of the lsquotran-scriptomersquo of a cyanobacterium to see if the set of geneswhose transcription is under most rapid and direct redoxcontrol corresponds to the genes of the chloroplast gen-ome Laboratories with this experimental capability do notseem to have addressed this possibility (Hihara et al 2002Ikeuchi 2002) Likewise if a putative modern representa-tive of the proto-mitochondrion can be found and agreedeg Rhodospirillum rubrum or Paracoccus denitrificans thena similar experiment should reveal a subset of genes corre-sponding to mitochondrial genomes DNA-microarraysalso present the possibility of testing the relative rate ofredox response and dependency on other signallinginputs of transcription of eukaryotic genes To dateorganellar genes have not been included in such investi-gations (Ferea et al 1999) A further experimental possi-bility is to examine the redox regulatory phenotypes ofexisting allotopic mutants as outlined in sect 3e
Phil Trans R Soc Lond B (2003)
6 CONCLUSION AND FUTURE PROSPECTS
The CORR hypothesis states that the function of mito-chondrial and chloroplast genomes is to provide co-location (of gene and gene product) for (evolutionary)continuity of redox regulation of gene expression Thishypothesis is neither self-contradictory nor devoid of test-able predictions It is also consistent with available evi-dence and in principle makes sense of the otherwisepuzzling distribution of genes between compartments ineukaryotic cells The explanation that CORR offersapplies equally to mitochondria and chloroplasts andidentifies selective forces that still operate to determine thelocation of genes No selective barriers need be assumedto gene transfer protein import nor membrane insertionNor do we need to invoke a lsquofrozen accidentrsquo that some-thing happened to stop gene transfer for some reason atsome time in the remote and inaccessible past CORR alsoexplains the absence of genomes from organelles that havelost their primary bioenergetic functions CORR may actas a stimulus to a search for residual bioenergetic functionin those organelles that have lost aerobic respiration andphotosynthesis but where residual genomes remain
CORR also predicts regulatory properties of knowncomponents of chloroplasts and mitochondria that in turnindicate important but as yet uncharacterized flexibility inenergy transduction These components include the RUB-ISCO large subunit and the CFo and Fo subunits of coup-ling ATPase in both chloroplasts and mitochondriaRedox regulation of the relative stoichiometry of compo-nents as seen in regulation of chloroplast photosystemstoichiometry (Pfannschmidt et al 1999a) may extend to(C)Fo-(C)F1-ATPase indicating that the function ofretention of genes for one or more of these componentsis flexibility in the stoichiometries of H1 to e2 and H1 toATP in chemiosmotic coupling (Schemidt et al 1998Stock et al 2000 Allen 2002)
So far explicit tests of CORR have largely been con-fined to chloroplasts and to some extent mitochondria ofhigher plants but this merely reflects the expertise andinterests of the investigators Further attempted disproofof CORR is welcomed and encouraged Novel experi-mental tests can be devised as well as attempted refu-tation through analysis of existing data It is hoped thatexplicit attention will be given to the CORR hypothesis inlaboratories already suitably equipped and that resourcescan be allocated for this purpose in laboratories wherethere is the interest but not currently the technology orexpertise
The heuristic value of the hypothesis already seen inthe role of redox-regulated transcription of photosyntheticreaction centre genes in chloroplasts may be its most use-ful property The central proposal is that mitochondriaand chloroplasts contain genes for proteins whose functionin electron transfer demands rapid direct and uncon-ditional redox regulatory control of their biosynthesis Thefull implications of this hypothesis are very broad and onlyjust beginning to be explored
I thank Carol Allen Aubrey de Grey William Martin John ARaven and Francis-Andre Wollman for discussion and corre-spondence during the preparation of this paper and WilliamMartin for suggestions including table 1 Research grants from
Genomes in mitochondria and chloroplasts J F Allen 33
The Swedish Natural Sciences Research Council (NFR) andthe Crafoord Foundation are gratefully acknowledged
REFERENCES
Adams K L Daley D O Qiu Y L Whelan J amp PalmerJ D 2000 Repeated recent and diverse transfers of a mito-chondrial gene to the nucleus in flowering plants Nature408 354ndash357
Alberts B Bray D Lewis J Raff M Roberts K amp Wat-son J D 1994 The molecular biology of the cell New Yorkand London Garland Publishing
Allen C A Hakansson G amp Allen J F 1995 Redox con-ditions specify the proteins synthesized by isolated chloro-plasts and mitochondria Redox Rep 1 119ndash123
Allen J F 1992 Protein phosphorylation in regulation ofphotosynthesis Biochim Biophys Acta 1098 275ndash335
Allen J F 1993a Control of gene expression by redox poten-tial and the requirement for chloroplast and mitochondrialgenomes J Theor Biol 165 609ndash631
Allen J F 1993b Redox control of gene expression and thefunction of chloroplast genomes a hypothesis PhotosynthRes 36 95ndash102
Allen J F 1993c Redox control of transcription sensorsresponse regulators activators and repressors FEBS Lett332 203ndash207
Allen J F 1995 Thylakoid proteinndashphosphorylation state-1ndashstate-2 transitions and photosystem stoichiometry adjust-ment redox control at multiple levels of gene expressionPhysiol Plant 93 196ndash205
Allen J F 1996 Separate sexes and the mitochondrial theoryof ageing J Theor Biol 180 135ndash140
Allen J F 2002 Photosynthesis of ATP-electrons protonpumps rotors and poise Cell 110 273ndash276
Allen J F amp Allen C A 1999 A mitochondrial model forpremature ageing of somatically cloned mammals IUBMBLife 48 369ndash372
Allen J F amp Nilsson A 1997 Redox signalling and the struc-tural basis of regulation of photosynthesis by protein phos-phorylation Physiol Plant 100 863ndash868
Allen J F amp Pfannschmidt T 2000 Balancing the two photo-systems photosynthetic electron transfer governs transcrip-tion of reaction centre genes in chloroplasts Phil TransR Soc Lond B 355 1351ndash1357 (DOI 101098rstb20000697)
Allen J F amp Race H L 2002 Will the real LHC II kinaseplease step forward Sciencersquos STKE httpwwwstkeorgcgicontentfullsigtrans2002155pe43
Allen J F amp Raven J A 1996 Free-radical-induced mutationvs redox regulation costs and benefits of genes in organellesJ Mol Evol 42 482ndash492
Archibald J M Cavalier-Smith T Maier U amp Douglas S2001 Molecular chaperones encoded by a reduced nucleusthe cryptomonad nucleomorph J Mol Evol 52 490ndash501
Ashby M K amp Mullineaux C W 1999a Cyanobacterialycf27 gene products regulate energy transfer from phycobili-somes to photosystems I and II FEMS Microbiol Lett 181253ndash260
Ashby M K amp Mullineaux C W 1999b The role of ApcDand ApcF in energy transfer from phycobilisomes to PSI andPSII in a cyanobacterium Photosynth Res 61 169ndash179
Ashby M K Houmard J amp Mullineaux C W 2002 Theycf27 genes from cyanobacteria and eukaryotic algae distri-bution and implications for chloroplast evolution FEMSMicrobiol Lett 214 25ndash30
Attardi G amp Schatz G 1988 Biogenesis of mitochondria ARev Cell Biol 4 289ndash333
Azem A Oppliger W Lustig A Jeno P Feifel B SchatzG amp Horst M 1997 The mitochondrial hsp70 chaperone
Phil Trans R Soc Lond B (2003)
system effect of adenine nucleotides peptide substrate andmGrpE on the oligomeric state of mhsp70 J Biol Chem272 20 901ndash20 906
Barbrook A C Symington H Nisbet R E R LarkumA amp Howe C J 2001 Organisation and expression of theplastid genome of the dinoflagellate Amphidinium opercula-tum Mol Genet Genomics 266 632ndash638
Bauer C E Elsen S amp Bird T H 1999 Mechanisms forredox control of gene expression A Rev Microbiol 53495ndash523
Bauer C E Elsen S Swem L R Swem D L amp MasudaS 2003 Redox and light regulation of gene expression inphotosynthetic prokaryotes Phil Trans R Soc Lond B 358147ndash154 (DOI 101098rstb20021189)
Berks B C Sargent F De Leeuw E Hinsley A P Stan-ley N R Jack R L Buchanan G amp Palmer T 2000aA novel protein transport system involved in the biogenesisof bacterial electron transfer chains Biochim Biophys ActaBioenerg 1459 325ndash330
Berks B C Sargent F amp Palmer T 2000b The tat proteinexport pathway Mol Microbiol 35 260ndash274
Blackstone N W 1999 Redox control in development andevolution evidence from colonial hydroids J Exp Biol 2023541ndash3553
Blackstone N W 2000 Redox control and the evolution ofmulticellularity Bioessays 22 947ndash953
Blackstone N W 2001 Redox state reactive oxygen speciesand adaptive growth in colonial hydroids J Exp Biol 2041845ndash1853
Bogorad L 1975 Evolution of organelles and eukaryotic gen-omes Science 188 891ndash898
Bogsch E G Sargent F Stanley N R Berks B C Robin-son C amp Palmer T 1998 An essential component of a novelbacterial protein export system with homologues in plastidsand mitochondria J Biol Chem 273 18 003ndash18 006
Brown E C Somanchi A amp Mayfield S P 2001 Interor-ganellar crosstalk new perspectives on signaling from thechloroplast to the nucleus Genome Biol 2 10211ndash10214
Cavalier-Smith T 2002 Chloroplast evolution secondarysymbiogenesis and multiple losses Curr Biol 12 R62ndashR64
Chitnis P R amp Thornber J P 1988 The major light-har-vesting complex of photosystem 2 Aspects of its molecularand cell biology Photosynth Res 16 41ndash63
Chow W S Melis A amp Anderson J M 1990 Adjustmentsof photosystem stoichiometry in chloroplasts improve thequantum efficiency of photosynthesis Proc Natl Acad SciUSA 87 7502ndash7506
Claros M G Perea J Shu Y Samatey F A PopotJ L amp Jacq C 1995 Limitations to in vivo import of hydro-phobic proteins into yeast mitochondria The case of a cyto-plasmically synthesized apocytochrome b Eur J Biochem228 762ndash771
Dalbey R E amp Robinson C 1999 Protein translocation intoand across the bacterial plasma membrane and the plant thy-lakoid membrane Trends Biochem Sci 24 17ndash22
Daley D O Adams K L Clifton R Qualmann S MillarA H Palmer J D Pratje E amp Whelan J 2002 Genetransfer from mitochondrion to nucleus novel mechanismsfor gene activation from Cox2 Plant J 30 11ndash21
Danon A amp Mayfield S P 1994 Light-regulated translationof chloroplast messenger RNAs through redox potentialScience 266 1717ndash1719
de Grey A 2000 Mitochondrial gene therapy an arena for thebiomedical use of inteins Trends Biotechnol 18 394ndash399
Deisenhofer J Epp O Miki K Huber R amp Michel H1985 Structure of the protein subunits in the photosyntheticreaction center of Rhodopseudomonas viridis at 3Ac resolutionNature 318 618ndash624
34 J F Allen Genomes in mitochondria and chloroplasts
Delwiche C F 1999 Tracing the thread of plastid diversitythrough the tapestry of life Am Nat 154 S164ndashS177
Doolittle W F 1999 Phylogenetic classification and the uni-versal tree Science 284 2124ndash2129
Douglas S E 1998 Plastid evolution origins diversitytrends Curr Opin Genet Dev 8 655ndash661
Duby G amp Boutry M 2002 Mitochondrial protein importmachinery and targeting information Plant Sci 162 477ndash490
Eberhard S Drapier D amp Wollman F A 2002 Searchinglimiting steps in the expression of chloroplast encoded pro-teins relations between gene copy number transcriptiontranscript abundance and translation rate in the chloroplastof Chlamydomonas reinhardtii Plant J 31 149ndash160
El Bissati K amp Kirilovsky D 2001 Regulation of psbA andpsaE expression by light quality in Synechocystis species PCC6803 A redox control mechanism Plant Physiol 1251988ndash2000
Ellis R J 1984 The nuclear domination of chloroplast devel-opment Science Progress 69 129ndash142
Embley T M amp Martin W 1998 Molecular evolution ahydrogen-producing mitochondrion Nature 396 517ndash519
Ems S C Morden C W Dixon C K Wolfe K HdePamphilis C W amp Palmer J D 1995 Transcriptionsplicing and editing of plastid RNAs in the non-photosyn-thetic plant Epifagus virginiana Plant Mol Biol 29 721ndash733
Eraso J M amp Kaplan S 2000 From redox flow to gene regu-lation role of the PrrC protein of Rhodobacter sphaeroides241 Biochemistry 39 2052ndash2062
Eraso J M amp Kaplan S 2002 Redox flow as an instrumentof gene regulation Meth Enzymol 348 216ndash229
Escoubas J M Lomas M Laroche J amp Falkowski P G1995 Light-intensity regulation of cab gene-transcription issignaled by the redox state of the plastoquinone pool ProcNatl Acad Sci USA 92 10 237ndash10 241
Ferea T L Botstein D Brown P O amp Rosenzweig R F1999 Systematic changes in gene expression patterns follow-ing adaptive evolution in yeast Proc Natl Acad Sci USA96 9721ndash9726
Forsberg J Rosenquist M Fraysse L amp Allen J F 2001Redox signalling in chloroplasts and mitochondria genomicand biochemical evidence for two-component regulatory sys-tems in bioenergetic organelles Biochem Soc Trans 29403ndash407
Fujita Y 1997 A study on the dynamic features of photosys-tem stoichiometry accomplishments and problems forfuture studies Photosynth Res 53 83ndash93
Gabriel K Buchanan S K amp Lithgow T 2001 The alphaand the beta protein translocation across mitochondrial andplastid outer membranes Trends Biochem Sci 26 36ndash40
Galvis M L E Allen J F amp Hakansson G 1998 Proteinsynthesis by isolated pea mitochondria is dependent on theactivity of respiratory complex II Curr Genet 33 320ndash329
Gatenby A A amp Ellis R J 1990 Chaperone function theassembly of ribulose bisphosphate carboxylase-oxygenase ARev Cell Biol 6 125ndash149
GoldschmidtClermont M 1998 Coordination of nuclear andchloroplast gene expression in plant cells Int Rev CytolSurv Cell Biol 177 115ndash180
Gray M W 1992 The endosymbiont hypothesis revisited IntRev Cytol 141 233ndash357
Gray M W 1999 Evolution of organellar genomes CurrOpin Genet Dev 9 678ndash687
Gray M W 2000 Evolutionary biology mitochondrial geneson the move Nature 408 302ndash305
Gray M W amp Doolittle W F 1982 Has the endosymbionthypothesis been proven Microbiol Rev 46 1ndash42
Gray M W Burger G amp Lang B F 1999 Mitochondrialevolution Science 283 1476ndash1481
Phil Trans R Soc Lond B (2003)
Grebe T W amp Stock J B 1999 The histidine protein kinasesuperfamily Adv Microb Physiol 41 139ndash227
Green B R amp Kuhlbrandt W 1995 Sequence conservationof light-harvesting and stress-response proteins in relation tothe 3-dimensional molecular structure of Lhcii PhotosynthRes 44 139ndash148
Gruissem W amp Tonkyn J C 1993 Control mechanisms ofplastid gene expression Crit Rev Plant Sci 12 19ndash55
Hackstein J H P (and 11 others) 2001 Hydrogenosomesconvergent adaptations of mitochondria to anaerobicenvironments Zool Anal Complex Syst 104 290ndash302
Harrison M A Keen J N Findlay J B C amp Allen J F1990 Modification of a Glnb-like gene-product by photosyn-thetic electron-transport in the cyanobacterium Synechoc-occus 6301 FEBS Lett 264 25ndash28
Harvey A J Kind K L amp Thompson J G 2002 Redoxregulation of early embryo development Reproduction 123479ndash486
Heathcote P Fyfe P K amp Jones M R 2002 Reactioncentres the structure and evolution of biological solarpower Trends Biochem Sci 27 79ndash87
Henze K amp Martin W 2001 How do mitochondrial genesget into the nucleus Trends Genet 17 383ndash387
Herrmann R G amp Westhoff P 2001 Thylakoid biogenesis anddynamics the result of a complex phylogenetic puzzle InRegulation of photosynthesis vol 11 (ed E-M Aro amp BAndersson) pp 1ndash28 Dordrecht The Netherlands Kluwer
Hihara Y Sonoike K Kanehisa M amp Ikeuchi M 2002Search for redox responsive genes in Synechocystis sp PCC6803 by using DNA microarrays Plant Cell Physiol 43S178
Howe C J Barbrook A C amp Lockhart P J 2000 Organellegenes do they jump or are they pushed Trends Genet 1665ndash66
Ikeuchi M 2002 Gene expression and responses to light stressin cyanobacteria Plant Cell Physiol 43 S21
Iuchi S amp Lin E C C 1992 Mutational analysis of signaltransduction by Arcb a membrane sensor protein respon-sible for anaerobic repression of operons involved in the cen-tral aerobic pathways in Escherichia coli J Bacteriol 1743972ndash3980
Jackson-Constan D Akita M amp Keegstra K 2001 Molecu-lar chaperones involved in chloroplast protein importBiochim Biophys Acta Mol Cell Res 1541 102ndash113
Jarvis P amp Soll M 2001 Toc Tic and chloroplast proteinimport Biochim Biophys Acta Mol Cell Res 1541 64ndash79
Jordan P Fromme P Witt H T Klukas O SaengerW amp Krauss N 2001 Three-dimensional structure ofcyanobacterial photosystem I at 25 acngstrom resolutionNature 411 909ndash917
Kaneko T (and 23 others) 1996a Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regionsDNA Res 3 109ndash136
Kaneko T (and 23 others) 1996b Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regions[Supplement] DNA Res 3 185ndash209
Kanevski I amp Maliga P 1994 Relocation of the plastid rbcLgene to the nucleus yields functional ribulose-15-bisphosph-ate carboxylase in tobacco chloroplasts Proc Natl Acad SciUSA 91 1969ndash1973
Kim J amp Mayfield S P 1997 Protein disulfide isomerase as aregulator of chloroplast translational activation Science 2781954ndash1957
Genomes in mitochondria and chloroplasts J F Allen 35
Kirk J T O amp Tilney-Bassett R A E 1978 The plastidsTheir chemistry structure growth and inheritance Amster-dam Elsevier
Koehler C M Merchant S amp Schatz G 1999 How mem-brane proteins travel across the mitochondrial intermem-brane space Trends Biochem Sci 24 428ndash432
Komiya T Rospert S Schatz G amp Mihara K 1997 Bind-ing of mitochondrial precursor proteins to the cytoplasmicdomains of the import receptors Tom70 and Tom20 isdetermined by cytoplasmic chaperones FASEB J 11 1196
Konstantinov Y M Lutsenko G N amp Podsosonny V A1995 Genetic functions of isolated maize mitochondriaunder model changes of redox conditions Biochem MolBiol Int 36 319ndash326
Korab-Laskowska M Rioux P Brossard N LittlejohnT G Gray M W Lang B F amp Burger G 1998 Theorganelle genome database project (GOBASE) Nucleic AcidsRes 26 138ndash144
Kovacs L Wiessner W Kis M Nagy F Mende D ampDemeter S 2000 Short- and long-term redox regulation ofphotosynthetic light energy distribution and photosystemstoichiometry by acetate metabolism in the green alga Chla-mydobotrys stellata Photosynth Res 65 231ndash247
Kuhlbrandt W 1994 Structure and function of the plant light-harvesting complex LHC-II Curr Opin Struct Biol 4519ndash528
Lang B F Gray M W amp Burger G 1999 Mitochondrialgenome evolution and the origin of eukaryotes A RevGenet 33 351ndash397
Lemire B D Fankhauser C Baker A amp Schatz G 1989The mitochondrial targeting function of randomly generatedpeptide sequences correlates with predicted helical amphi-philicity J Biol Chem 264 20 206ndash20 215
Leuenberger D Bally N A Schatz G amp Koehler C M1999 Different import pathways through the mitochondrialintermembrane space for inner membrane proteins EMBOJ 18 4816ndash4822
Li H amp Sherman L A 2000 A redox-responsive regulator ofphotosynthesis gene expression in the cyanobacterium Syne-chocystis sp strain PCC 6803 J Bacteriol 182 4268ndash4277
Lin X Y (and 36 others) 1999 Sequence and analysis of chro-mosome 2 of the plant Arabidopsis thaliana Nature 402761ndash768
Lonsdale D M Hodge T P Howe C J amp Stern D B1983 Maize mitochondrial-DNA contains a sequence hom-ologous to the ribulose-15-bisphosphate carboxylase largesubunit gene of chloroplast DNA Cell 34 1007ndash1014
McFadden G I 1999 Endosymbiosis and evolution of theplant cell Curr Opin Plant Biol 2 513ndash519
McFadden G I 2001 Chloroplast origin and integrationPlant Physiol 125 50ndash53
Mahler H R amp Raff R A 1975 The evolutionary origin ofthe mitochondrion a nonsymbiotic model Int Rev Cytol43 1ndash124
Manfredi G Fu J Ojaimi J Sadlock J E Kwong J QGuy J amp Schon E A 2002 Rescue of a deficiency in ATPsynthesis by transfer of MTATP6 a mitochondrial DNA-encoded gene to the nucleus Nature Genet 30 394ndash399
Marc P Margeot A Devaux F Blugeon C Corral-Debrinski M amp Jacq C 2002 Genome-wide analysis ofmRNAs targeted to yeast mitochondria EMBO Rep 3159ndash164
Marienfeld J Unseld M amp Brennicke A 1999 The mito-chondrial genome of Arabidopsis is composed of both nativeand immigrant information Trends Plant Sci 4 495ndash502
Martin W 1999 A briefly argued case that mitochondria andplastids are descendants of endosymbionts but that thenuclear compartment is not Proc R Soc Lond B 2661387ndash1395 (DOI 101098rspb19990792)
Phil Trans R Soc Lond B (2003)
Martin W amp Herrmann R G 1998 Gene transfer fromorganelles to the nucleus how much what happens andwhy Plant Physiol 118 9ndash17
Martin W amp Muller M 1998 The hydrogen hypothesis forthe first eukaryote Nature 392 37ndash41
Martin W amp Russell M J 2003 On the origins of cells ahypothesis for the evolutionary transitions from abiotic geo-chemistry to chemoautotrophic prokaryotes and from pro-karyotes to nucleated cells Phil Trans R Soc Lond B 35859ndash85 (DOI 101098rstb20021183)
Martin W Stoebe B Goremykin V Hansmann S Hase-gawa M amp Kowallik K V 1998 Gene transfer to thenucleus and the evolution of chloroplasts Nature 393162ndash165
Martin W Hoffmeister M Rotte C amp Henze K 2001 Anoverview of endosymbiotic models for the origins of eukary-otes their ATP-producing organelles (mitochondria andhydrogenosomes) and their heterotrophic lifestyle BiolChem 382 1521ndash1539
Martin W Rujan T Richly E Hansen A Cornelsen SLins T Leister D Stoebe B Hasegawa M amp PennyD 2002 Evolutionary analysis of Arabidopsis cyanobacterialand chloroplast genomes reveals plastid phylogeny and thou-sands of cyanobacterial genes in the nucleus Proc NatlAcad Sci USA 99 12 246ndash12 251
Millen R S (and 12 others) 2001 Many parallel losses of infAfrom chloroplast DNA during angiosperm evolution withmultiple independent transfers to the nucleus Plant Cell 13645ndash658
Morse D Salois P Markovic P amp Hastings J W 1995 Anuclear-encoded form-II RUBISCO in dinoflagellatesScience 268 1622ndash1624
Murakami A Kim S J amp Fujita Y 1997 Changes in photo-system stoichiometry in response to environmental con-ditions for cell growth observed with the cyanophyteSynechocystis PCC 6714 Plant Cell Physiol 38392ndash397
Oh J I amp Kaplan S 2000 Redox signaling globalization ofgene expression EMBO J 19 4237ndash4247
Ozawa T 1995 Mitochondrial-DNA mutations associatedwith aging and degenerative diseases Exp Gerontol 30269ndash290
Palmer J D 1997 Organelle genomes going going goneScience 275 790ndash791
Pearson C K Wilson S B Schaffer R amp Ross A W 1993NAD turnover and utilization of metabolites for RNA syn-thesis in a reaction sensing the redox state of the cyto-chrome-b( 6 )f complex in isolated chloroplasts Eur JBiochem 218 397ndash404
Pfanner N amp Geissler A 2001 Versatility of the mitochon-drial protein import machinery Natl Rev Mol Cell Biol 2339ndash349
Pfanner N amp Wiedemann N 2002 Mitochondrial proteinimport two membranes three translocases Curr Opin CellBiol 14 400ndash411
Pfannschmidt T Nilsson A amp Allen J F 1999a Photosyn-thetic control of chloroplast gene expression Nature 397625ndash628
Pfannschmidt T Nilsson A Tullberg A Link G amp AllenJ F 1999b Direct transcriptional control of the chloroplastgenes psbA and psaAB adjusts photosynthesis to light energydistribution in plants IUBMB Life 48 271ndash276
Pfannschmidt T Allen J F amp Oelmuller R 2001a Prin-ciples of redox control in photosynthesis gene expressionPhysiol Plant 112 1ndash9
Pfannschmidt T Schutze K Brost M amp Oelmuller R2001b A novel mechanism of nuclear photosynthesis generegulation by redox signals from the chloroplast during pho-
36 J F Allen Genomes in mitochondria and chloroplasts
tosystem stoichiometry adjustment J Biol Chem 27636 125ndash36 130
Popot J L amp de Vitry C 1990 On the microassembly of inte-gral membrane proteins A Rev Biophys Biophys Chem 19369ndash403
Race H L Herrmann R G amp Martin W 1999 Why haveorganelles retained genomes Trends Genet 15 364ndash370
Raven J A Johnston A M Parsons R amp Kubler J 1994aThe influence of natural and experimental high O2 concen-trations on O2 evolving phototrophs Biol Rev CambridgePhil Soc 69 61ndash94
Raven J A Johnston A M Parsons R amp Kubler J 1994bThe occurrence and influence on photolithotrophs of highoxygen concentrations Proc R Soc Edinb B 102 193ndash201
Robinson C 2000 The twin-arginine translocation system anovel means of transporting folded proteins in chloroplastsand bacteria Biol Chem 381 89ndash93
Robinson C amp Bolhuis A 2001 Protein targeting by the twin-arginine translocation pathway Natl Rev Mol Cell Biol 2350ndash356
Rodermel S 2001 Pathways of plastid-to-nucleus signalingTrends Plant Sci 6 471ndash478
Roger A J 1999 Reconstructing early events in eukaryoticevolution Am Nat 154 S146ndashS163
Roger A J Svard S G Tovar J Clark C G SmithM W Gillin F D amp Sogin M L 1998 A mitochondrial-like chaperonin 60 gene in Giardia lamblia evidence thatdiplomonads once harbored an endosymbiont related to theprogenitor of mitochondria Proc Natl Acad Sci USA 95229ndash234
Roodyn D B amp Wilkie D 1968 The biogenesis of mitochondriaMethuenrsquos monographs on biological subjects London Methuen
Rujan T amp Martin W 2001 How many genes in Arabidopsiscome from cyanobacteria An estimate from 386 proteinphylogenies Trends Genet 17 113ndash120
Saccone C Gissi C Lanave C Larizza A Pesole G ampReyes A 2000 Evolution of the mitochondrial genetic sys-tem an overview Gene 261 153ndash159
Salzberg S L White O Peterson J amp Eisen J A 2001Microbial genes in the human genome lateral transfer orgene loss Science 292 1903ndash1906
Sargent F Berks B C amp Palmer T 2002 Assembly of mem-brane-bound respiratory complexes by the Tat protein-trans-port system Arch Microbiol 178 77ndash84
Schatz G 1998 Protein transport the doors to organellesNature 395 439ndash440
Schemidt R A Qu J Williams J R amp Brusilow W S1998 Effects of carbon source on expression of F0 genes andon the stoichiometry of the c subunit in the F1 F0 ATPaseof Escherichia coli J Bacteriol 180 3205ndash3208
Selosse M A Albert B R amp Godelle B 2001 Reducing thegenome size of organelles favours gene transfer to thenucleus Trends Ecol Evol 16 135ndash141
Stern D B amp Lonsdale D M 1982 Mitochondrial and chlo-roplast genomes of maize have a 12- kilobase DNA-sequencein common Nature 299 698ndash702
Stock D Gibbons C Arechaga I Leslie A G W ampWalker J E 2000 The rotary mechanism of ATP synthaseCurr Opin Struct Biol 10 672ndash679
Stoebe B amp Maier U G 2002 One two three naturersquos toolbox for building plastids Protoplasma 219 123ndash130
Stoebe B Martin W amp Kowallik K V 1998 Distributionand nomenclature of protein-coding genes in 12 sequencedchloroplast genomes Plant Mol Biol Rep 16 243ndash255
Stupar R M Lilly J W Town C D Cheng Z Kaul SBuell C R amp Jiang J M 2001 Complex mtDNA consti-tutes an approximate 620 kb insertion on Arabidopsis thali-ana chromosome 2 implication of potential sequencing
Phil Trans R Soc Lond B (2003)
errors caused by large-unit repeats Proc Natl Acad SciUSA 98 5099ndash5103
Surpin M Larkin R M amp Chory J 2002 Signal transduc-tion between the chloroplast and the nucleus Plant Cell 14S327ndashS338
Tjaden J Schwoppe C Mohlmann T Quick P W ampNeuhaus H E 1998 Expression of a plastidic ATPADPtransporter gene in Escherichia coli leads to a functional aden-ine nucleotide transport system in the bacterial cytoplasmicmembrane J Biol Chem 273 9630ndash9636
Tovar J Fischer A amp Clark C G 1999 The mitosome a novelorganelle related to mitochondria in the amitochondrial para-site Entamoeba histolytica Mol Microbiol 32 1013ndash1021
Trebitsh T amp Danon A 2001 Translation of chloroplastpsbA mRNA is regulated by signals initiated by both photos-ystems II and I Proc Natl Acad Sci USA 98 12 289ndash12 294
Tullberg A Alexciev K Pfannschmidt T amp Allen J F2000 Photosynthetic electron flow regulates transcription ofthe psaB gene in pea (Pisum sativum L) chloroplasts throughthe redox state of the plastoquinone pool Plant Cell Physiol41 1045ndash1054
Turmel M Lemieux C Burger G Lang B F Otis CPlante I amp Gray M W 1999 The complete mitochondrialDNA sequences of Nephroselmis olivacea and Pedinomonasminor two radically different evolutionary patterns withingreen algae Plant Cell 11 1717ndash1729
Unden G amp Bongaerts J 1997 Alternative respiratory path-ways of Escherichia coli energetics and transcriptional regu-lation in response to electron acceptors Biochim BiophysActa Bioenerg 1320 217ndash234
Unseld M Marienfeld J R Brandt P amp Brennicke A1997 The mitochondrial genome of Arabidopsis thaliana con-tains 57 genes in 366 924 nucleotides Nature Genet 1557ndash61
Van der Giezen M Slotboom D J Horner D S DyalP L Harding M Xue G P Embley T M amp KunjiE R S 2002 Conserved properties of hydrogenosomal andmitochondrial ADPATP carriers a common origin for bothorganelles EMBO J 21 572ndash579
Von Heijne G 1986 Why mitochondria need a genome FEBSLett 198 1ndash4
Westphal S Soll J amp Vothknecht U C 2001 A vesicle trans-port system inside chloroplasts FEBS Lett 506 257ndash261
Whitehouse H L K 1969 Towards an understanding of themechanism of heredity London Edward Arnold Ltd
Williams B A P Hirt R P Lucocq J M amp Embley T M2002 A mitochondrial remnant in the microsporidian Trachi-pleistophora hominis Nature 418 865ndash869
Wilson R J (and 10 others) 1996a Complete gene map of theplastid-like DNA of the malaria parasite Plasmodium falcipa-rum J Mol Biol 261 155ndash172
Wilson S B Davidson G S Thomson L M amp PearsonC K 1996b Redox control of RNA synthesis in potato mito-chondria Eur J Biochem 242 81ndash85
Winkler H H amp Neuhaus H E 1999 Non-mitochondrialATP transport Trends Biochem Sci 24 64ndash68
Wolfe K H Morden C W amp Palmer J D 1992 Small sin-gle-copy region of plastid DNA in the non-photosyntheticangiosperm Epifagus virginiana contains only two genes Dif-ferences among dicots monocots and bryophytes in geneorganization at a non-bioenergetic locus J Mol Biol 22395ndash104
Wollman F A Minai L amp Nechushtai R 1999 The biogen-esis and assembly of photosynthetic proteins in thylakoidmembranes Biochim Biophys Acta Bioenerg 1411 21ndash85
Yeliseev A A amp Kaplan S 2000 TspO of Rhodobactersphaeroides a structural and functional model for the mam-
Genomes in mitochondria and chloroplasts J F Allen 37
malian peripheral benzodiazepine receptor J Biol Chem275 5657ndash5667
Zerges W 2002 Does complexity constrain organelle evol-ution Trends Plant Sci 7 175ndash182
Zhang Z D Green B R amp Cavalier-Smith T 1999 Singlegene circles in dinoflagellate chloroplast genomes Nature400 155ndash159
Zhang Z D Cavalier-Smith T amp Green B R 2001 A fam-ily of selfish minicircular chromosomes with jumbled chloro-plast gene fragments from a dinoflagellate Mol Biol Evol18 1558ndash1565
Zouni A Witt H T Kern J Fromme P Krauss NSaenger W amp Orth P 2001 Crystal structure of photosys-tem II from Synechococcus elongatus at 38 acngstrom resol-ution Nature 409 739ndash743
DiscussionD S Horner (Dipartimento di Fisiologia e Biochimica
Generali University of Milan Milan Italy) Given thatthere are components of these membrane-bound energy-transduction pathways that are consistently nuclear enco-ded not the key ones you pointed out but others do youexpect perhaps a slower perhaps a more crude but yet aredox communication between the organelle andexpression of nuclear-encoded genes
J F Allen There is and there is no question about itI think that Bogoradrsquos lock-in hypothesis has somethingto say here and there are other people who say it is all todo with assembling multi-subunit complexes If you havecontrol over the initial construction of the core aroundwhich the whole complex is eventually built then redoxcontrol or some sort of control over synthesis of that corewill control construction of the whole thing eventuallyand I think that may well be largely correct I think it ispossible in principle for nuclear genes to make precursorsaccording to redox instructions but it is not really soimportant that those precursors are here and now presentin exactly the right quantities because the precursors canto a limited extent hang around and wait Actually thereis proteolytic degradation of unattached precursors and soon but I think the fact that these chloroplast and mito-chondrially DNA-encoded subunits are at the core ofthese energy-transducing complexes is the crucial issue
T Cavalier-Smith (Department of Zoology University ofOxford Oxford UK ) In the case of mitochondria the sub-set of genes that remains is much more variable than youimplied It runs from three proteins in the minimal casesup to I think over 90 That is a big difference and manyof the genes in different lineages of protists that remainhave nothing to do with redox control they are things likeribosomal proteins and so on So I think that even thoughwhat you say about redox control of certain genes mayactually be an important thing it is not the fundamentaloverall reason why the genomes are maintained It may bepart of the explanation for why certain genes are alwaysincluded in that subset but even in mitochondria I thinkthat cytochrome b is the only one that is always presentSo I think that both this great diversity of what has beenretained in different lineages and the nucleomorph caseindicate that evolution was able to do one thing in onelineage 500 million years ago and never got around todoing it for 500 million years in another lineage so thereis a lot more haphazardness and evolutionary accidentsand contingency than a straight functional interpretation
Phil Trans R Soc Lond B (2003)
there is a lot of just sheer lsquohappenedndashorndashdidnrsquotndashhappenrsquoor carryover
J F Allen I was not implying anything about the rangeof mitochondrial genomes I have a limited amount oftime so I just talked about what I could talk about withinthat limit Of course there will be a range and this willdepend upon the physiology and bioenergetics of theparticular organism But what I am trying to do is to seewhat the common properties of the irreducible core areof components that are always encoded in the organellesand cytochrome b is a perfect example because that reallyis the thing that binds these two all-important quinoneswithout which there is no Q cycle without which there isno proton pumping and bioenergetics and in Plasmodiumwhere the mitochondrion is reduced to this bare mini-mum there is still a mitochondrial gene for cytochromeb Now my argument about the ribosomal proteins andall the rest of it is that in order to be able to exert thisall-important redox control over gene expression of certaincore components you need an in situ genetic system andthis can vary in composition for reasons that I cannotdescribe Maybe you need RNA polymerase or maybeyou can import RNA polymerase In plastids you haveboth of these options You are going to need ribosomestransfer RNAs and the rest of the apparatus of geneexpression and that needs to be in situ but we are stillleft with the question why does there have to be any gen-etic system at all The mature tobacco or spinach chloro-plast seems to have an entire genetic system just to makeone protein D1 which is all it ever needs to resynthesizeI think that this is a clue It is worth having this very unec-onomical crazy system because there is an overridingrequirement to connect electron transfer with geneexpression directly and unambiguouslymdashwithout recourseto a bureaucracy if you like which is what I think thenucleus is
K Willison (Institute of Cancer Research Chester BeattyLaboratories London UK ) These photosynthetic reactioncomplexes are a sort of baroque mixture of polypeptidechains and cofactors and my understanding is that someof them in high photon flux can turn over within say 30minutes so I am wondering if there is a correlationbetween gene retention and the turnover rates of some ofthe components and the complexes it seems to me thatthat could really be a limiting factor and the reason thatthe proteins had to be encoded locally in order to get thecomplexes to be assembled rapidly
J F Allen I think that is right You can only alter thesteady-state concentration of a protein complex by affect-ing the synthesis and breakdown and therefore turnoverThere is an extreme example the D1 protein encoded bythe psbA gene of chloroplasts which is always in the chlor-oplast genome without exception High light will tend todegrade this protein and when the high light goes awaythere is an absolute need very quickly to bring it back upagain I think that is a very good example Whether thisargument applies to genes such as cytochrome oxidase Ido not know
C J Leaver (Department of Plant Sciences University ofOxford Oxford UK ) Your experiments tend to relatemainly to the gene expression side What about proteinRNA turnover the whole concept of redox control ofthatmdashdo we know anything
38 J F Allen Genomes in mitochondria and chloroplasts
J F Allen Well quite a lot is known about chloro-plasts There is redox control of messenger RNA stabilityfor this same D1 protein which has been described byStephen Mayfield and his co-workers The evidence is thatthe site of redox control is ferredoxin via thioredoxin andthe implication is that thioredoxin tells whether non-cyclicelectron transport is working at all When it is workingthere is a signal that says lsquoOK now continue with all theprotein synthesisrsquo It is not particularly specific but moreof a global effect I think this is what Mayfield argues butof course this is not inconsistent with my argument I haveconcentrated on quinone pools and medium-potentialredox regulatory control because that is where I can seesignals about the dangerous consequences of getting itwrong but there is control of other levels of geneexpression In mitochondria I am not so sure I wouldbounce the question back to you on that one
W Martin (Institute of Botany III Heinrich-Heine Univ-ersitat Dusseldorf Dusseldorf Germany) The nucleomorphgenome was cited as evidence in conflict with your modelbut is that true The nucleomorph genome is not in abioenergetic organelle in that sense It resides in the cyto-sol so does the size of the nucleomorph genome workagainst your model Is it really evidence against yourmodel
J F Allen I do not know why should it be evidenceI cannot explain why the nucleomorph has a genomeThat is why I clutched at the straw that it is storage fornucleic acid precursors
Phil Trans R Soc Lond B (2003)
T Cavalier-Smith The reason I introduced the nucleo-morph is not to say it is direct evidence against yourhypothesis but to show that there probably was no singlefunctional reason for the retention of every gene in everyorganelle It may be that there are specific physiologicalreasons for the retention of certain genes and thereforeyou might be right about a subset of genes but I verymuch doubt if there are specific physiological reasons forthe retention of every single gene that is retained differen-tially because odd things happen in evolution Somethings are easy and sometimes things are difficult Evol-ution is history
J F Allen Of course I agree with that I do not thinkI can explain everything but I do think I can explainsomething If I have done that then I am content
GLOSSARY
AAC ADPndashATP carrierCES control by epistasy of synthesisCORR co-location for redox regulationLHC light-harvesting complexPCC Pasteur culture collectionORF open reading frameRUBISCO ribulose-15-bisphosphate carboxylasendashoxygenaseTAT twin-arginine translocase
Visit httpwwwpubsroyalsocacuk to see an electronic appendix tothis paper

24 J F Allen Genomes in mitochondria and chloroplasts
nucleus cytosol
O2
H2O
redox regulation
N-phasemitochondrial matrix
Figure 3 Gene expression and principal pathways of biosynthesis of subunits of protein complexes involved in respiration andoxidative phosphorylation in animal mitochondria As in figure 1 reddish-brown DNA RNA and protein subunits are locatedand synthesized in the mitochondrial matrix light brown protein subunits have genes (also light brown) in the nucleus andare imported from the cytosol as precursors White genes and ribosomal and protein subunits are nuclearndashcytoplasmic and ofarchaebacterial origin Reddish-brown and light-brown genes and ribosomal and protein subunits are of bacterial origin Themajor variable environmental input is oxygen (blue) The essence of the CORR hypothesis is that it is beyond the ability ofthe nuclearndashcytoplasmic system to respond rapidly and directly to changes in oxygen concentration or partial pressure and soredox regulation of gene expression (red arrows) has been retained from the ancestral endosymbiont This redox regulationrequires co-location of certain genes with their gene products within the mitochondrion
nucleus cytosol
CH2O
CO2
redox regulation
N-phasechloroplast stroma
light light
Figure 4 Gene expression and principal pathways of biosynthesis of subunits of protein complexes involved inphotosynthesis in chloroplasts As in figure 2 green DNA RNA and protein subunits are located and synthesized in thechloroplast light brown protein subunits have genes (also light brown) in the nucleus and are imported from the cytosolas precursors White genes and ribosomal and protein subunits are nuclearndashcytoplasmic and of archaebacterial originGreen and light-brown genes and ribosomal and protein subunits are of bacterial origin The major environmental inputsare light (orange arrows) and carbon dioxide (blue) The essence of the CORR hypothesis is that it is beyond the abilityof the nuclearndashcytoplasmic system to respond rapidly and directly to changes in light and carbon dioxide and so redoxregulation of gene expression (red arrows) has been retained from the ancestral endosymbiont This redox regulationrequires co-location of certain genes with their gene products within the chloroplast
Phil Trans R Soc Lond B (2003)
Genomes in mitochondria and chloroplasts J F Allen 25
has been demonstrated (Pfannschmidt et al 1999abAllen amp Pfannschmidt 2000 Tullberg et al 2000) and wasprompted by this principle
The level of redox control of chloroplast transcriptionestablished unequivocally so far is that of plastoquinonean electron and hydrogen atom carrier located strategicallybetween photosystems I and II of photosynthesis Thedirections in which the effects are exerted are functionallyintelligible reduction of plastoquinone increases photosy-stem I transcription and decreases photosystem IItranscription oxidation of plastoquinone decreases photo-system I transcription and increases photosystem II tran-scription Plastoquinone in normal photosynthesis isreduced by the action of photosystem II and oxidized bythat of photosystem I Therefore redox control of chloro-plast gene expression provides a compensatory response(Pfannschmidt et al 1999a) and adds a previously mis-sing part of an explanation for the ability of plants andalgae to adjust the ratio of the two photosystems inresponse to changes in light quality (Chow et al 1990)By the same argument a functionally equivalent post-translational mechanism of redox control of light-harvesting function by photosystems I and II may havebeen conserved during the transition from prokaryotes toeukaryotes (Allen 1992 1995 Allen amp Nilsson 1997Allen amp Race 2002)
The precedent of chloroplast redox control of transcrip-tion (Pfannschmidt et al 1999ab) taken together with theprinciple in sect 3d predicts plastoquinone redox control ofcyanobacterial reaction centre gene transcription and thisis borne out by experiment (El Bissati amp Kirilovsky2001) This complementary self-adjusting transcriptionalresponse (Fujita 1997 Murakami et al 1997 El Bissati ampKirilovsky 2001) indicates strongly that redox control ofphotosynthetic reaction centre gene transcription waspresent in the proto-chloroplast Thus the mechanism ofregulation is likely to have been conserved throughout theevolutionary transition from bacterial symbiont to cyto-plasmic organelle A conserved mechanism of redox regu-lation of transcription in photosynthetic prokaryotes(Unden amp Bongaerts 1997 Bauer et al 1999) and chloro-plasts (Allen 1993c Pfannschmidt et al 1999b) is pre-dicted if evolutionary continuity of the process has beenmaintained
Thus apart from being consistent with experimentaldata the principle of continuity of redox control has beenof heuristic value in pointing to the underlying bacterialcharacter previously unsuspected in the control of geneexpression especially of transcription in chloroplasts ofhigher plants Other possible precedents for redox signal-ling components of cytoplasmic organelles include the prr(Eraso amp Kaplan 2000 2002 Oh amp Kaplan 2000) andreg (Bauer et al 1999) redox regulatory systems of purplenon-sulphur bacteria The TspO lsquooxygen sensorrsquo of Rho-dobacter sphaeroides is present in mammalian mitochondriaas a protein known there as a benzodiazepine receptorand each protein will complement loss of the other(Yeliseev amp Kaplan 2000) As outlined in sect 3j genes forsuch components are to be expected primarily in nucleargenomes This independent evolutionary hypothesis (sect 3d)would be strengthened still further by a demonstration ofone or more decisive redox regulatory links shared by pho-tosynthetic prokaryotes and all eukaryotes with mitochon-
Phil Trans R Soc Lond B (2003)
dria and where present with chloroplasts (Bauer et al2003)
(e) The principle of the selective value of redoxcontrol
For each gene under redox control (sect 3d) it is selectivelyadvantageous for that gene to be retained and expressed onlywithin the organelle
The twin selective advantages of redox control are likelyto be energetic efficiency and the suppression of the harm-ful side effects of electron transport operating on thewrong substrates No direct experimental support for thisprinciple is available and equally there is no evidenceagainst it Allotopic mutants in which genes have beenexperimentally relocated from organelles to the nucleushave been produced in yeast higher plants (Kanevski ampMaliga 1994) and human cells (Manfredi et al 2002) andare predicted by CORR to be poor adapters to environ-mental change Some authors believe there is potentialtherapeutic value in allotopic transformation movingmitochondrial genomes to the nucleus (Kanevski ampMaliga 1994 de Grey 2000) and predict the possibilityof r2 human cell lines in the foreseeable future (de Grey2000) This principle also has implications for ageing andthe evolution of separate sexes (Allen 1996) and possibleconsequences for development (Blackstone 1999 20002001 Harvey et al 2002) and somatic cloning in animals(Allen amp Allen 1999)
(f ) The principle of the selective value of nuclearlocation for genes not under redox control
For each bacterial gene that survives and is not under redoxcontrol it is selectively advantageous for that gene to be locatedin the nucleus and expressed only in the nucleus and cytosolIf the mature gene product functions in chloroplasts or mito-chondria the gene is first expressed in the form of a precursorfor import
There are several possible reasons for the selectiveadvantage of nuclear location of genes for organellar pro-teins One is the decreased probability of mutation arisingfrom free radical by-products of lsquoincorrectrsquo electron trans-fers (Raven et al 1994ab Allen amp Raven 1996) Therewould seem to be obvious economies in resource use andadvantages in coordination of gene expression if genes arekept in one compartment One might say that the bipartiteor tripartite genetic system of eukaryotic cells is untidyand inefficient
Another suggestion is that organellar genes do notundergo recombination and are present in relatively smallcopy numbers so that disadvantageous mutations willspread relatively quickly through a clonal population oforganelles Incorporation of functional gene copiesinstead into the nuclear genome may then be an irrevers-ible step analogous to the click of a ratchet (Saccone et al2000) There are examples of genes shared between allthree compartments (Stern amp Lonsdale 1982 Lonsdale etal 1983)
The cost of retaining an organellar genetic system is notincreased appreciably when additional genes are retainedin organelles if just one gene is retained then the cost ofa cytoplasmic genetic system is incurred and there may berelatively little extra cost for each additional gene retained(Allen amp Raven 1996) For a functional explanation of the
26 J F Allen Genomes in mitochondria and chloroplasts
existence of chloroplast and mitochondrial genomes thefocus of attention should therefore be on the relativelyconstant primary subset of bioenergetic genes (sect 3e)
(g) The principle of contemporary operation ofselection on gene location
For any species the distribution of genes between organelle(by the principle in sect 3e) and nucleus (by the principle in sect 3f)is the result of selective forces that continue to operate
This principle is essentially a statement contradictingboth the lsquofrozen accidentrsquo hypothesis and the hypothesisthat gene relocation is progressive predictable and stillincomplete (sect 4) In support of this principle it is becom-ing increasingly clear that mitochondria that lose theirfunction in aerobic respiration also lose their genomes asseen in the relict mitochondria of microsporidia(Embley amp Martin 1998 Van der Giezen et al 2002) andthe lsquomitosomersquo of Entamoeba histolytica (Tovar et al1999) In chloroplasts loss of photosynthesis results inloss of photosynthetic genes The examples of Epifagus (aparasitic higher plant) (Ems et al 1995 Wolfe et al 1992)and the residual apicomplexan plastids of Plasmodium(Wilson et al 1996a) and Toxoplasma where some plas-tidic genetic system is retained mean that some of theirgene products must be subject to redox regulation of geneexpression if this principle is correct Experimental test ofthis principle is possible using allotopic mutants as forthe predictions of the principle in sect 3f The evidence thatthe apicoplast (Wilson et al 1996a) is a plastid is largelygenetic the residual genome resembles that of photosyn-thetic plastids (chloroplasts) in gene organization ratherthan in content A plastid that has completely lost its gen-ome by virtue of having lost photosynthesis and all otherprocesses requiring redox regulation might be difficult todistinguish from an organelle derived in a similar way froma mitochondrion The persistence of targeted importedchaperones Hsp70 (Tovar et al 1999 Williams et al2002) and cp60 (Roger et al 1998) in residual organellesmight not distinguish a plastid ancestry from a mitochon-drial one
(h) The principle of primary involvement inenergy transduction
Those genes for which redox control is always vital to cellfunction have gene products involved in or closely connectedwith primary electron transfer These genes are always con-tained within the organelle
This principle would be violated and the CORRhypothesis refuted if any photosynthetic reaction centrecore subunit or respiratory chain subunit currentlyorganelle-encoded is found that functions in primary vec-torial electron transport and is nevertheless encoded in thenucleus and synthesized cytosolically as a precursor forimport all without any selective cost to the individualFrom the chloroplast transcriptional experiments ofPfannschmidt et al (1999ab) the chloroplast geneswhose transcription is regulated by the plastoquinoneredox state include psaAB (for photosystem I reactioncentre core polypeptides) psbA (for the photosystem IIpolypeptide D1) and rbcL (for the large subunit of RUB-ISCO interestingly following the same pattern of redoxcontrolmdashupregulated by oxidized plastoquinonemdashaspsbA) Transcription of the chloroplast genes petA
Phil Trans R Soc Lond B (2003)
(cytochrome f ) trnG (glycine tRNA) and rpoB (RNApolymerase) did not respond to plastoquinone redox statepetA for cytochrome f is predicted by this principle to beregulated at a different level of redox control or if at plas-toquinone at a different stage in gene expression Thisprediction might also apply to trnG and rpoB though thesequalify as members of the secondary subset that of geneticsystem genes to which the principle in sect 3i applies
(i) The principle of secondary involvement inenergy transduction
Genes whose products contribute to the organelle genetic sys-tem itself or whose products are associated with secondaryevents in energy transduction may be contained in theorganelle in one group of organisms but not in anotherdepending on the physiology and biochemistry of photosynthesisand respiration in the species concerned
Even phylogenetically moderately related species(within kingdoms) show differences in the location ofsome genes for example the ATP synthase subunits ofNeurospora and Saccharomyces (Attardi amp Schatz 1988)The CORR hypothesis also does not explicitly account forthe wide disparities in the distribution of genetic systemgenes for example the almost complete import of tRNAsin Chlamydomonas (GoldschmidtClermont 1998) Trans-port of nucleic acids across membranes is uncommon andwhy some RNAs should be organelle-encoded in somespecies but nuclearly encoded in others is not currentlyunderstood The only suggestion that can be made alongthe lines of CORR is that in some genetic systems genesmay need to be under the control of a regulatory systemthat responds to a signal arising within the organelle itselfThis signal could be the primary signal of altered redoxstate or some secondary signal Among the latter might befound signals that themselves report on the current stateof gene expression another piece of information that thenucleus might not have at its immediate disposal Suchsecondary signals could result at least in part fromCORR though other developmental and environmentalinputs may be imagined and would not detract fromCORR unless it could be shown that they provided a suf-ficient reason for the retention of the genetic system asa whole
(j) The principle of the nuclear encoding of redoxsignalling components
Components of the redox-signalling pathways upon whichCORR depends are themselves not involved in primary electrontransfer and so their genes have been relocated to the nucleusin accordance with sect 3f
This principle states that the redox signalling pathwaysthat are required for CORR should not themselves beexpected to utilize components whose biosynthesis mustbe under direct redox control Such components shouldtherefore fall into the major category of organellar pro-teins and be imported as precursors from the cytosol inline with the principle in sect 3f However the principleabove might be need to be modified in cases where ampli-fication of the redox signal is useful to the cell In thebacterial ntr system which was a precedent for CORR(Harrison et al 1990 Allen 1992) the histidine kinasentrB and aspartate response regulator ntrC form part ofthe gln operon whose transcription they control Odontella
Genomes in mitochondria and chloroplasts J F Allen 27
and Porphyra have plastids that encode more proteins thanthose of most chlorophytes (Stoebe et al 1998 Turmel etal 1999) and these proteins include two-component sig-nal transduction proteins prr and ycf27 (Stoebe et al1998) Extreme or extremely rapid environmental fluctu-ations might be catered for by amplification of signals thatare transmitted by signalling pathways whose componentsare under their own control It is not known whether ycf27and prr are in operons whose promoters their gene pro-ducts control It is also uncertain whether ycf27 is a redoxsignalling component and its distribution at least in itsplastidic location seems to be correlated with the presenceof a phycobilin-based light-harvesting system as found inred algae and cyanobacteria (Ashby amp Mullineaux1999ab Ashby et al 2002) The best guess of the originalstatement of the CORR hypothesis (Allen 1992 1993ac)was that bacterial two-component systems (Iuchi amp Lin1992 Allen 1993c Grebe amp Stock 1999) provide a plaus-ible means of exerting redox regulatory control over mito-chondrial and chloroplast gene expression andsubsequent research confirms the presence of two-compo-nent genes in genomes of Arabidopsis and yeast SeveralArabidopsis two-component genes are predicted to be tar-geted to mitochondria and chloroplasts (Forsberg et al2001) These must be considered as prime candidates forcomponents involved in redox regulation of chloroplasttranscription seen in the experiments of Pfannschmidt etal (1999a 2001a) The presumably homologous regulat-ory process serving to control photosystem stoichiometryin cyanobacteria the prr system (Li amp Sherman 2000) isnow characterized as a two-component system involvinga redox sensor and redox response regulator as originallydefined (Allen 1992 1993ac)
4 CONTRASTING HYPOTHESES CONCERNINGTHE RETENTION OF CYTOPLASMIC GENOMES
CORR is a recent addition to explanations that havebeen put forward to account for the persistence of gen-omes in organelles These models and their underlyingmechanisms are now discussed and compared
(a) There is no reason that is just how it isOne reaction to the problem of cytoplasmic genetic sys-
tems (sect 2) is that there is no problem and no particularreason for their retention However there is a very widephylogenetic distribution of a common core or primarysubset of organellar-encoded genes and one which haswithstood considerable genomic reorganization over evol-utionary time This constancy is seen for example in thesame genes being present in the chloroplasts of landplants with multiple copies of a single circular chloroplastchromosome and in dinoflagellate chloroplasts with acollection of different DNA minicircles each with essen-tially one gene (Zhang et al 1999 2001 Barbrook et al2001) Another suggested explanation of the retention ofmitochondrial and chloroplast genomes is simply torestate the endosymbiont hypothesis and to point out thatchloroplast and mitochondrial genes lsquocame inrsquo with thebacterial endosymbiont This suggestion however doesnot address the question of why some genes were retainedwhereas others were not It is also now clear that mostgenes that came in and still survive have nuclear and cyto-
Phil Trans R Soc Lond B (2003)
solic gene products whereas many chloroplast and mito-chondria proteins had a eukaryotic origin (Salzberg et al2001 Martin amp Russell 2003) The odds against a con-served pattern in gene location (eg table 1) arising bychance must be astronomical Thus there is a reason forthe retention of organellar genomes and a reason for theretention of the same primary subset of genes Becauseevolution proceeds by natural selection of chance vari-ation lsquoreasonrsquo in this context requires identification of thefunction of the location of a gene that is the selectivevalue of the phenotype produced by that location Evol-ution sometimes retains apparently functionless relics ofpreviously useful features but not for long especially ifthere is a cost
(b) The lock-in hypothesisThe lsquolock-inrsquo hypothesis of Bogorad (1975) was pro-
posed independently of the endosymbiont hypothesis anddoes not depend upon it According to the lock-in hypoth-esis core components of multi-subunit complexes must besynthesized de novo in the correct cellular compartmentotherwise photosystems or respiratory chain complexesfor example might assemble in the plasma membranevacuolar membrane or endoplasmic reticulum The lock-in hypothesis made no clear prediction about the conti-nuity of cytoplasmic genetic systems even though cyto-plasmic uniparental non-Mendelian inheritance for bothmitochondria (Roodyn amp Wilkie 1968) and plastids(Kirk amp Tilney-Bassett 1978) made it likely that organellarDNA the chondriome and the plastome replicated inde-pendently of nuclear DNA (Whitehouse 1969)
Mechanisms of protein import and targeting (Schatz1998 Koehler et al 1999 Leuenberger et al 1999 Gab-riel et al 2001 Jarvis amp Soll 2001 Pfanner amp Geissler2001 Duby amp Boutry 2002 Pfanner amp Wiedemann 2002)now effectively answer the problem of specifying the cor-rect locations for membrane protein complexes for whichthe lock-in hypothesis was proposed as a solution How-ever regulation of synthesis of an entire functional multi-subunit complex by control of synthesis of individual sub-units is still an important principle This seems to be afeature of the lsquoCESrsquo model (control by epistasy ofsynthesis) of Wollman (Wollman et al 1999 Eberhard etal 2002) which envisages nuclear control as all-embracingwith organellar genetic systems doing as it were whatthey are told In the hypothesis of CORR propoundedhere CES is equally important but the essence of CORRis that the organellar genetic system alone knows what arethe immediate and most important signals of environmen-tal change The organelle may keep the nucleus informedbut this is a lower priority than initiating an appropriateresponse CES and CORR agree that assembly of multi-subunit complexes is a complex business and requirescoordinated control in which no step is rate-limiting butmany may be obligatory
(c) Going going gone the evolutionary process oftransfer of genes from organelles to the
nucleus is still underway but incompleteThis hypothesis envisages that for most eukaryotes we
are now witnessing the last stages in a slow evolutionarytransfer of genes to the nucleus and this transfer may beexpected to proceed to completion in what is in evol-
28 J F Allen Genomes in mitochondria and chloroplasts
utionary terms the near future (Palmer 1997 Gray 19992000 Gray et al 1999 Adams et al 2000 Daley et al2002) A clear and compelling statement of this widelyheld position is that of Herrmann amp Westhoff (2001 p10)
hellipthe transfer of geneshelliptells that the partite plant gen-ome is not in a phylogenetic equilibrium All availabledata suggest that the ultimate aim of genome restructur-ing in the plant cell as in the eukaryotic cell in generalis the elimination of genome compartmentation whileretaining physiological compartmentation
One serious problem with this hypothesis is the dis-tinctly non-random sample of genes that remain to betransferred A relatively constant subset of genes is foundin bioenergetically functional mitochondria from fungi tomammals and angiosperms as in photosynthetic plastids(chloroplasts) from unicellular algae to angiosperms Thishypothesis therefore requires an additional ad hocassumption that some gene transfers are inherently moreprobable than others (this too is widely supposed seelater) and thus that gene transfer events proceed in thesame sequence in quite independent lineages Thereseems to be no evidence for this and the order of geneloss from mitochondria and chloroplasts has been used toinfer phylogeny (Martin et al 1998 Adams et al 2000Millen et al 2001) lsquoGoing going gonersquo (Palmer 1997)is difficult to refute categorically but it does not seem tobe consistent with recent evidence For example chromo-some two of Arabidopsis thaliana (Lin et al 1999) has asingle insertion encompassing with about 99 sequencesimilarity the complete Arabidopsis mitochondrial genome(Unseld et al 1997 Marienfeld et al 1999 Stupar et al2001) Unless this single transfer is a freak occurrencethis finding strongly indicates that gene transfer occurswith relative ease (Adams et al 2000 Henze amp Martin2001 Rujan amp Martin 2001 Salzberg et al 2001) Inaddition there is experimental evidence that randomlygenerated polypeptides are mitochondrially imported athigh frequency (Lemire et al 1989) The hypothesis thatwe now see the same few stragglers in diverse organellargenomes is not impossible but should be regarded as alast resort perhaps applicable only in extreme cases suchas the greatly reduced apicomplexan plastomes and chon-driomes Even there the hypothesis carries with it an obli-gation to explain why organisms with few organellar geneshave lsquoevolvedrsquo faster than those with many
(d ) The frozen accidentWhat may be described as lsquothe frozen accident hypoth-
esisrsquo states that the evolutionary process of gene transferfrom organelle to nucleus was underway when somethinghappened that stopped it One example of the frozen acci-dent is a scenario (Von Heijne 1986) in which import intomitochondria of the precursor proteins arising from newlytranslocated genes proceeded for long enough after theprimordial endosymbiosis for most of the symbiont-derived genes to be moved to the cell nucleus The lsquoacci-dentrsquo that halted the process was the evolutionary originof exocytosis and protein secretion after which it becameimpossible for precursors properly destined for mitochon-drial import to be distinguished from those destined forexport from the cell As with the lsquolock-inrsquo hypothesis the
Phil Trans R Soc Lond B (2003)
lsquofrozen accidentrsquo seems incompatible with the remarkableprecision and specificity of protein targeting (Schatz 1998Koehler et al 1999 Leuenberger et al 1999 Gabriel etal 2001 Jarvis amp Soll 2001 Pfanner amp Geissler 2001Duby amp Boutry 2002 Pfanner amp Wiedemann 2002) Pro-tein sorting mechanisms do not seem to forbid controlledimport into any cellular compartment Another lsquoaccidentrsquothat may have impeded subsequent gene transfer in thecase of mitochondria is their departure from what is other-wise a universal genetic code Genetic code differences arediscussed later in the context of whether some genes can-not be moved
(e) Some proteins (with cofactors) cannot beimported
We may describe as the lsquounimportability hypothesisrsquo theproposal that some proteins are inherently unimportableand must therefore be synthesized de novo in the correctcompartment within the cell The unimportability hypoth-esis is most often proposed as its special case the hydro-phobicity hypothesis with hydrophobicity being thereason for unimportability
(i) HydrophobicityA commonly entertained notion is that proteins that are
encoded and synthesized within organelles comprise a pri-mary subset characterized by shared hydrophobicity allare intrinsic membrane proteins (Von Heijne 1986Popot amp de Vitry 1990 Claros et al 1995) As with theCORR hypothesis favoured here the primary subset thenrequires a secondary subset of genetic system componentsthe latter not themselves being hydrophobic but beingrequired for synthesis of the others Certainly the mito-chondrial inner membrane contains some intrinsic proteincomplexes whose subunits broadly satisfy the criterion ofhydrophobicity However the core subunits that are mito-chondrially encoded are also explained by the CORRhypothesis This would make hydrophobicity a require-ment for their function in vectorial electron and protontransfer and not a reason in itself for retention of theirgenes in the cytoplasm
The hydrophobicity hypothesis has some very clearcounter-examples in chloroplasts and cannot thereforestand if one wishes for a single principle that embracesboth types of bioenergetic organelle One clear counter-example is the large (L) subunit of the enzyme RUBISCOan abundant but entirely membrane-extrinsic protein ofca 55 kDa molecular mass The RUBISCO large subunitis generally chloroplast-encoded with the exception of thenuclearly encoded bacterial-type L subunit of L2 RUBI-SCO in peridinin-containing dinoflagellates (Morse et al1995) In most photosynthetic eukaryotes RUBISCO hasa chloroplast encoded large subunit and a nuclearly enco-ded small (S) subunit (Ellis 1984 Gatenby amp Ellis 1990)giving an L8S8 holoenzyme and this RUBISCO is widelytaken as a paradigm for the need for coordination ofnuclear and chloroplast gene expression (Rodermel 2001)
By complete contrast to the RUBISCO large subunitwhich is a chloroplast-encoded and water-soluble proteina second major violation of the hydrophobicity hypothesisis the existence of the universally nuclearly encoded butalways hydrophobic subunits of the chloroplast LHC IIand LHC I (Chitnis amp Thornber 1988) These apopro-
Genomes in mitochondria and chloroplasts J F Allen 29
teins of ca 27ndash30 kDa are quite water-insoluble and areusually fully extracted only in chloroformndashmethanol or inthe presence of detergents The Lhc gene family is diverse(Green amp Kuhlbrandt 1995) but most gene productsor Lhcp (light-harvesting chlorophyll protein) polypeptideshave three membrane-spanning hydrophobic a-helices(Kuhlbrandt 1994) It is interesting that three-helix Lhcproteins seem to be absent from extant cyanobacteria andchloroxybacteria (Green amp Kuhlbrandt 1995)
(ii) The five-helix rulePerhaps with the three-helix model of LHC II in mind
a variation of the hydrophobicity hypothesis is the empiri-cal lsquofive-helix rulersquo (Popot amp de Vitry 1990 Claros et al1995 Wollman et al 1999) which states that no intrinsicmembrane protein with five or more membrane-spanninghelices can be encoded in the nucleus Five is a good guessfor a threshold number of helices demanding organellar-encoding in the context of photosynthetic reaction centres(Heathcote et al 2002) where the earliest known struc-ture that of type II centres (Deisenhofer et al 1985) asin photosystem II (Zouni et al 2001) shows exactly fivehelices with type I centres containing 11 (Jordan et al2001) There are however many nuclearly encoded sub-units in both mitochondria and chloroplasts that havefewer than five membrane helices so the five-helix rulecan only describe a sufficient condition and not a neces-sary condition for nuclear encoding
It should also be noted that ADPndashATP carriers (AACs)are encoded in the nucleus AACs of the mitochondrialinner membrane have six transmembrane helices (van derGiezen et al 2002) while the those of the chloroplast innerenvelope (Winkler amp Neuhaus 1999) have 12 (Tjaden etal 1998)
With the counter-examples of nuclearly encoded AACswith six or 12 membrane-spanning helices many intrinsicmembrane proteins with fewer than five helices somenuclearly encoded and some not and with the RUBISCOlarge subunit as a major product of chloroplast geneexpression with no membrane domain at all the five-helixrule begins to seem rather like a case of special pleadingEven if there are indeed special mitigating circumstancesfor AACs (this argument is outlined later) the five-helixrule as with the hydrophobicity hypothesis of which it isa special case will disappear if it can be shown that pos-session of five or more membrane helices is required forsome other function such as that of CORR that deter-mines the organellar location of the gene concernedHydrophobicity is in some cases a feature of proteins thatare encoded in organelles but it is not in itself the issue
For chloroplasts the hydrophobicity hypothesis predictsretention of cyanobacterial genes in functional categoriesassociated with the chloroplast envelope and with trans-port just as much as with photosynthesis Table 1 showsthat this is not the case
A mechanism for the unimportability of hydrophobicproteins is rarely stated explicitly and it is rather left tothe imagination whether hydrophobic proteins might asit were get stuck halfway through the chloroplast envelopeor mitochondrial outer membrane whether transport can-not get started on a precursor protein that has alreadyspontaneously inserted into the membrane or whetherhydrophobic proteins will be unable to make the journey
Phil Trans R Soc Lond B (2003)
through the cytosol without precipitating there In thecytosol co-translational transport or membrane insertionwould in principle address the latter point but it seemsto be generally true that chloroplast and mitochondrialproteins are post-translationally inserted into the organel-lar outer membrane So the obvious ancillary questionabout unimportability is lsquowhy then cannot hydrophobicproteins be co-translationally imported into organellesrsquoThere is evidence in yeast for co-translational transportinto mitochondria of proteins thought to be derived fromthe original endosymbiont (Marc et al 2002) If there isany truth in hydrophobicity as a barrier to protein importthe CORR principle of unselective import (sect 3c) shouldbe easily refuted for example by a demonstration that noknown targeting presequence is capable of importing poly-phenylalanine encoded by polyU for in vitro import stud-ies
An additional proposal intended to account for theunimportability of some hydrophobic proteins and forimportability of others is suggested by A de Grey(personal communication) who argues that bacteriallyderived protein precursors imported into mitochondriacross the mitochondrial inner membrane completelybefore insertion into that membrane from the mitochon-drial matrix (the N-phase) This is held to be an obstacleto successful import of membrane-intrinsic proteins thatmight en route discharge the proton motive force By con-trast eukaryotically derived proteins can be inserted intothe mitochondrial inner membrane from the inter-mem-brane space (the P-phase) and then there is for somereason no similar uncoupling effect Thus adenylatetransporters or AACs can be nuclearly encoded whereascytochrome oxidase subunits for example cannot It isnot obvious why bacterially derived subunits such asthose of cytochrome oxidase did not acquire eukaryoticinsertion mechanisms Perhaps this acquisition is forbid-den by the need to conserve pre-existing routes ofassembly into multi-subunit complexes or perhaps theneed for insertion from the inside reflects a need to incor-porate a cofactor for example haem or ironndashsulphur clus-ters along the way
(iii) Unimportability of three dimensional structures andcofactors
Accumulating evidence on protein transport mech-anisms seems to make a specific obstacle to successfultransport of certain sorts of protein less and less plausibleespecially given the observation that precursors with N-terminal targeting sequences not mature functional poly-peptides themselves traverse membranes during biosynth-esis
Protein folding inside the living cell is invariably assistedby molecular chaperones (Gatenby amp Ellis 1990 Azem etal 1997 Komiya et al 1997 1990 Jackson-Constan et al2001) proteins that impede the formation of lsquoimproperrsquostructural interactions that will otherwise produce aggre-gation or other non-functional interactions Proteinimport mechanisms appear mostly to rely on specific mol-ecular chaperones for guided unfolding before membraneinsertion and refolding after translocation of the polypep-tide into its destined compartment (Gatenby amp Ellis 1990Azem et al 1997 Komiya et al 1997 Jackson-Constanet al 2001) Because polypeptides are mostly lsquothreadedrsquo
30 J F Allen Genomes in mitochondria and chloroplasts
through membranes no special constraint on transportshould be expected to arise from 3D structure or surfaceproperties of a protein
An exception to this unfolding rule may be providedby holoproteins containing cofactors that are transportedalong with the apoprotein presumably with a largely intact3D structure The lsquoTATrsquo (twin-arginine translocase) sys-tem (Bogsch et al 1998 Dalbey amp Robinson 1999 Berkset al 2000ab Robinson 2000 Robinson amp Bolhuis 2001Sargent et al 2002) works broadly along these linesthough its mechanism is unresolved TAT is relevant herebecause it adds transport of essentially complete holopro-teins to the cellrsquos repertoire of transport mechanisms andeven proteins that may not be allowed to unfold perhapsbecause of the toxicity of the free cofactor are not therebymade candidates for an unimportability rule One variantof the unimportability hypothesis is that transport canwork only by unfolding and dissociated cofactors may betoxic to the cell The bacterially derived TAT system(Bogsch et al 1998 Dalbey amp Robinson 1999 Berks etal 2000ab Sargent et al 2002) overcomes this argumentas would examples of cofactor-free polypeptides such as(C)Fo subunit c which are nevertheless generally encodedin organelles
The presence of a vesicular transport system in chloro-plasts (Westphal et al 2001) counts against the unim-portability hypothesis Vesicular transport between thechloroplast envelope and the thylakoid membrane shouldin principle allow any protein to be ferried across thechloroplast stroma Vesicles might allow transport inparticular of membrane proteins that would discharge theproton motive force if allowed to connect transiently theN and P aqueous phases on each side of a coupling mem-brane
Another observation that seems to count against unim-portability is the increased number of membrane barriersin plastids derived from secondary tertiary or higher-orderendosymbioses These plastids seem to have retained ifanything fewer genes rather than more as unimportabilitypredicts The nucleomorph the residual nucleus of theprimary host retains many genes for molecular chap-erones (Archibald et al 2001 Cavalier-Smith 2002) inagreement with the conclusion that protein import pro-ceeds by conserved and specialized mechanisms If eachnew import added to gain a higher-order endosymbiosisrequires additional targeting presequences then perhapsthere is an advantage in components of an import machin-ery not being imported themselves
It seems that unimportability is less easy an assumptionto make today than in previous years and the onus maynow be on its proponents to cite specific examples of pro-teins that can never be imported into chloroplasts or mito-chondria
(f ) Some genes cannot be moved(i) Differences in the mitochondrial genetic code
The genetic code is the specific relation between ribon-ucleotide triplets of codons and tRNAs that recognizethem and which thus determine which individual aminoacids if any will be incorporated into the growing poly-peptide chain Mitochondria show small departures fromthe otherwise universal genetic code (Attardi amp Schatz1988 Alberts et al 1994) UGA is a lsquostoprsquo codon in the
Phil Trans R Soc Lond B (2003)
universal code and in plant mitochondria but is the codonfor tryptophan in mitochondria of animals and fungiCUA means leucine everywhere except fungal mitochon-dria where it means threonine and AGA and AGU codefor arginine in the lsquouniversalrsquo code but serine in insectmitochondria and lsquostoprsquo in mammalian ones The signifi-cance of this heterogeneity in coding is most unlikely torest in the early origin of mitochondria as the lsquouniversalrsquocode is employed by bacteria and protists Furthermorethe diversity of departures from the lsquouniversalrsquo codeamong mitochondria of different eukaryotic lineages indi-cates they had independent origins A general rule formitochondrial genetic codes is that reliance on the thirdlsquowobblersquo position is decreased which allows all 20 proteinamino acids to be specified by fewer tRNAs (22 in animalmitochondria instead of 31 in the cytosol) This in turnmeans that mtDNA can specify more proteins for a givenlength of DNA or conversely that the mitochondrialgenome can be condensed or lsquostreamlinedrsquo Although thisarrangement does not explain the retention of mitochon-drial genomes and may be a secondary event in mitochon-drial evolution it almost certainly places an extra barrierin place of gene relocation As discussed previously how-ever there seems to be no single barrier that can explainthe retention of a relatively constant subset of mitochon-drial genes
There has clearly been a contraction of mitochondrialDNA especially in animals Animal mitochondrial DNAhas few if any non-coding sequences and animal mito-chondrial genetic codes require fewer tRNAs than the sin-gle code operating in the cytosol (Gray 1999 Gray et al1999 Selosse et al 2001) These may be devices todecrease the total frequency of mutations per mitochon-drial genome assuming a constant frequency per unitlength of mitochondrial DNA It has been proposed thata genersquos removal to the nucleus decreases its susceptibilityto free radical mutagenesis (Raven et al 1994ab Allen ampRaven 1996) If this is the case then the non-standardcode may be a further indication of an overriding need tomaintain certain genes within the mitochondrion What-ever the reason for the retention of genes in organelles ithad better be a good one bioenergetic organelles areclearly the wrong place to keep genetic information andgo to extraordinary lengths to offset the consequences ofit being there
(ii) Molecular refugeesA phenomenon related to differences in the genetic code
is seen in chloroplasts This is a bias in codon usage Thelsquomolecular refugeesrsquo hypothesis (Howe et al 2000) sug-gests that nuclear copies of what were once chloroplastgenes escape constraints on codon usage and thus thatwhat distinguishes chloroplast-encoded proteins is thattheir amino-acid composition is unusual enough for thebias in codon usage not to apply It is difficult to see whatphysico-chemical properties might be uniquely shared bythe amino acids of chloroplast-encoded proteins and inaddition there seem to be no lsquoforbiddenrsquo amino-acid resi-due in chloroplasts As with the departure from the univer-sal genetic code in mitochondria is seems likely that abias in codon usage reflects some secondary property ofchloroplast genomes and is not the primary reason fortheir retention In any case any restriction on gene move-
Genomes in mitochondria and chloroplasts J F Allen 31
ment will act in the wrong direction to explain why chloro-plasts have retained genomes at all provided there is noresulting obstacle to a chloroplast protein being success-fully encoded as a precursor in the nucleus
(iii) Genes are copied and replicated not relocated in onestep
If complete relocation is envisaged as proceeding by thesimultaneous appearance of a gene in one compartmentand its deletion from another then it is possible to imaginethat whatever the vector is there may be selectivity ingene transfer (Henze amp Martin 2001) For example ifviral incorporation replication and recombination in bothorganelle and nucleus were the mechanism of relocationthen successful gene transfer might depend on the abilityof the virus to complete the process in one step This inturn might depend on some property of the gene or ofthe protein it encodes if expression occurs at any stage inthe process of relocation An assumption of the CORRhypothesis favoured here is that genes are not movedphysically but merely copied after which either the copyin the original location or the copy in the new locationmay be lost as a result of selection A useful analogy maybe the action of copying files between computer storagevolumes In this analogy the copy is identical to the orig-inal and either may subsequently be deleted without lossof information
Because any gene that is not usefully expressed willundergo no selection to maintain a functional allele theprevailing location will be the one in which the functionalcopy of the gene resides The problem of which gene sur-vives is then subsumed into the problem of which is theselectively advantageous location for it It seems reason-able to assume that there wasmdashand ismdashan interim periodin which homologous genes in different compartments areessentially in competition to see which functions best Asoutlined above there is no obvious obstacle for copyingDNA sequences within the cell The physico-chemicalproperties of DNA are fairly uniform and not directlydependent on nucleotide sequence though organelles likeintra-cellular parasites seem to prefer A and T to G andC The answer to the question lsquowhich gene location func-tions bestrsquo may have nothing to do with the gene itselfnor directly with the structure and function of its geneproduct Instead the answer may have everything to dowith the required regulatory interaction between the geneproduct and expression of the gene that produces it
(g) CORRThis hypothesis has been propounded earlier (sect 3) and
will be summarized as follows Vectorial electron and pro-ton transfer exerts regulatory control over expression ofgenes encoding proteins directly involved in or affectingredox poise This regulatory coupling is indispensable foradaptation to environmental changes in chemical activitiesof oxygen light and CO2 and requires co-location of thesegenes with their gene products Organelles lsquomake theirown decisionsrsquo on the basis of environmental changesaffecting redox state The hypothesis is abbreviatedCORR for co-location for redox regulation or more com-pletely lsquoco-location (of gene and gene product) for(evolutionary) continuity of redox regulation of geneexpressionrsquo
Phil Trans R Soc Lond B (2003)
5 SUMMARY OF THE CURRENT STATUS OF THEHYPOTHESIS OF CORR
Q Why do mitochondria and chloroplasts have theirown genetic systems
A CORR co-location of genes and gene products per-mits direct and autonomous redox regulation of geneexpression
(a) Objections to CORR(i) There is no regulation of gene expression in chloroplasts
and mitochondriaThis is still commonly stated but incorrect See the prin-
ciple in sect 3d
(ii) There is no regulation of gene expression in chloroplastsand mitochondria at the level of transcription
For chloroplasts there is now clear evidence for a func-tionally intelligible redox control of transcription(Pfannschmidt et al 1999a) This invalidates the previouswidely held assumption that all control of chloroplast geneexpression is post-transcriptional (Gruissem amp Tonkyn1993 GoldschmidtClermont 1998) The evidence formitochondria is less clear but redox control of transcrip-tion certainly cannot be ruled out and there are indi-cations that it may occur (Pearson et al 1993Konstantinov et al 1995 Wilson et al 1996b) It mustalso be restated that transcriptional control is not neces-sary for CORR but serves as a compelling example of aconserved typically bacterial mode of response that oper-ates still in eukaryotic cytoplasmic organelles thus illus-trating the principle of continuity of redox control (sect 3d)Post-transcriptional redox regulation (Danon amp Mayfield1994 Kim amp Mayfield 1997 Brown et al 2001 Treb-itsh amp Danon 2001) is consistent with CORR and thereis nothing to forbid it being a primary mechanism thatsatisfies most or indeed all of the formal requirements ofthe hypothesis in particular cases
(iii) There are no redox signalling components encoded inchloroplast or mitochondrial DNA
This is what CORR predicts See the principle in sect 3j
(iv) There is redox control of nuclear gene expressionThis is correct and many examples are now known
especially for chloroplasts (Escoubas et al 1995 Kovacs etal 2000 Pfannschmidt et al 2001ab Surpin et al 2002)Redox signals from photosynthetic or respiratory electrontransport chains are not at all prohibited by CORR Theessence of CORR is that there is a subset of organellarproteins for which there is an overriding requirement fora redox regulatory control that is initiated within theorganelle and whose effects are exerted by the simplestand most direct route This redox signalling is envisagedto take place without a requirement for prior consultationand coordination with what may be regarded as a nucleo-cytoplasmic bureaucracy
(v) Nuclear genes can be expressed as rapidly as chloroplastand mitochondrial genes
Zerges (2002) for example cites the equal times takenfor complete expression of nuclear and chloroplast genesin Chlamydomonas as evidence against the CORR hypoth-
32 J F Allen Genomes in mitochondria and chloroplasts
esis However the question is not one of the time takento express a gene but the time taken to alter the rate ofits expression in response to a (redox) signal originatingwithin the organelle An equally important additional fac-tor may be the autonomy of the signalling pathway thatis its independence from other inputs as described in theprevious paragraph One might say that mitochondria andchloroplasts require a free hand
(vi) There is no evidence to support this hypothesisThe hypothesis is rich in predictions (sectsect 3 and 6) and
those that have been tested stringently are in agreementwith experimental results None of the predictions of thehypothesis has been found to be incorrect and none of theunderlying principles (sect 3) has been disproved Furtherpredictions will be the subject of future experiments
(b) Evidence for CORR(i) Chloroplasts have selectively retained cyanobacterial genes
for primary events in photosynthesisSee table 1
(ii) There is regulation of gene expression in chloroplastsThere is transcriptional regulation of gene expression
in chloroplasts (sect 3d) The case for mitochondria is notyet proven
(iii) There is redox regulation of gene expression inchloroplasts
There is transcriptional redox regulation of geneexpression in chloroplasts (sect 3d) The case for mitochon-dria is not yet proven though see (Allen et al 1995 Kon-stantinov et al 1995 Galvis et al 1998)
(iv) There are no redox signalling components encoded inchloroplast or mitochondrial DNA
Correct but with possible exceptions discussed in sect 3j
(v) There are lsquobacterialrsquo redox signalling components ineukaryotes
See sect 3j
(vi) Further predictions DNA-array results and phenotypesof allotopic mutants
A systematic test of CORR might be to make an oxid-ized-minus-reduced gene expression profile of the lsquotran-scriptomersquo of a cyanobacterium to see if the set of geneswhose transcription is under most rapid and direct redoxcontrol corresponds to the genes of the chloroplast gen-ome Laboratories with this experimental capability do notseem to have addressed this possibility (Hihara et al 2002Ikeuchi 2002) Likewise if a putative modern representa-tive of the proto-mitochondrion can be found and agreedeg Rhodospirillum rubrum or Paracoccus denitrificans thena similar experiment should reveal a subset of genes corre-sponding to mitochondrial genomes DNA-microarraysalso present the possibility of testing the relative rate ofredox response and dependency on other signallinginputs of transcription of eukaryotic genes To dateorganellar genes have not been included in such investi-gations (Ferea et al 1999) A further experimental possi-bility is to examine the redox regulatory phenotypes ofexisting allotopic mutants as outlined in sect 3e
Phil Trans R Soc Lond B (2003)
6 CONCLUSION AND FUTURE PROSPECTS
The CORR hypothesis states that the function of mito-chondrial and chloroplast genomes is to provide co-location (of gene and gene product) for (evolutionary)continuity of redox regulation of gene expression Thishypothesis is neither self-contradictory nor devoid of test-able predictions It is also consistent with available evi-dence and in principle makes sense of the otherwisepuzzling distribution of genes between compartments ineukaryotic cells The explanation that CORR offersapplies equally to mitochondria and chloroplasts andidentifies selective forces that still operate to determine thelocation of genes No selective barriers need be assumedto gene transfer protein import nor membrane insertionNor do we need to invoke a lsquofrozen accidentrsquo that some-thing happened to stop gene transfer for some reason atsome time in the remote and inaccessible past CORR alsoexplains the absence of genomes from organelles that havelost their primary bioenergetic functions CORR may actas a stimulus to a search for residual bioenergetic functionin those organelles that have lost aerobic respiration andphotosynthesis but where residual genomes remain
CORR also predicts regulatory properties of knowncomponents of chloroplasts and mitochondria that in turnindicate important but as yet uncharacterized flexibility inenergy transduction These components include the RUB-ISCO large subunit and the CFo and Fo subunits of coup-ling ATPase in both chloroplasts and mitochondriaRedox regulation of the relative stoichiometry of compo-nents as seen in regulation of chloroplast photosystemstoichiometry (Pfannschmidt et al 1999a) may extend to(C)Fo-(C)F1-ATPase indicating that the function ofretention of genes for one or more of these componentsis flexibility in the stoichiometries of H1 to e2 and H1 toATP in chemiosmotic coupling (Schemidt et al 1998Stock et al 2000 Allen 2002)
So far explicit tests of CORR have largely been con-fined to chloroplasts and to some extent mitochondria ofhigher plants but this merely reflects the expertise andinterests of the investigators Further attempted disproofof CORR is welcomed and encouraged Novel experi-mental tests can be devised as well as attempted refu-tation through analysis of existing data It is hoped thatexplicit attention will be given to the CORR hypothesis inlaboratories already suitably equipped and that resourcescan be allocated for this purpose in laboratories wherethere is the interest but not currently the technology orexpertise
The heuristic value of the hypothesis already seen inthe role of redox-regulated transcription of photosyntheticreaction centre genes in chloroplasts may be its most use-ful property The central proposal is that mitochondriaand chloroplasts contain genes for proteins whose functionin electron transfer demands rapid direct and uncon-ditional redox regulatory control of their biosynthesis Thefull implications of this hypothesis are very broad and onlyjust beginning to be explored
I thank Carol Allen Aubrey de Grey William Martin John ARaven and Francis-Andre Wollman for discussion and corre-spondence during the preparation of this paper and WilliamMartin for suggestions including table 1 Research grants from
Genomes in mitochondria and chloroplasts J F Allen 33
The Swedish Natural Sciences Research Council (NFR) andthe Crafoord Foundation are gratefully acknowledged
REFERENCES
Adams K L Daley D O Qiu Y L Whelan J amp PalmerJ D 2000 Repeated recent and diverse transfers of a mito-chondrial gene to the nucleus in flowering plants Nature408 354ndash357
Alberts B Bray D Lewis J Raff M Roberts K amp Wat-son J D 1994 The molecular biology of the cell New Yorkand London Garland Publishing
Allen C A Hakansson G amp Allen J F 1995 Redox con-ditions specify the proteins synthesized by isolated chloro-plasts and mitochondria Redox Rep 1 119ndash123
Allen J F 1992 Protein phosphorylation in regulation ofphotosynthesis Biochim Biophys Acta 1098 275ndash335
Allen J F 1993a Control of gene expression by redox poten-tial and the requirement for chloroplast and mitochondrialgenomes J Theor Biol 165 609ndash631
Allen J F 1993b Redox control of gene expression and thefunction of chloroplast genomes a hypothesis PhotosynthRes 36 95ndash102
Allen J F 1993c Redox control of transcription sensorsresponse regulators activators and repressors FEBS Lett332 203ndash207
Allen J F 1995 Thylakoid proteinndashphosphorylation state-1ndashstate-2 transitions and photosystem stoichiometry adjust-ment redox control at multiple levels of gene expressionPhysiol Plant 93 196ndash205
Allen J F 1996 Separate sexes and the mitochondrial theoryof ageing J Theor Biol 180 135ndash140
Allen J F 2002 Photosynthesis of ATP-electrons protonpumps rotors and poise Cell 110 273ndash276
Allen J F amp Allen C A 1999 A mitochondrial model forpremature ageing of somatically cloned mammals IUBMBLife 48 369ndash372
Allen J F amp Nilsson A 1997 Redox signalling and the struc-tural basis of regulation of photosynthesis by protein phos-phorylation Physiol Plant 100 863ndash868
Allen J F amp Pfannschmidt T 2000 Balancing the two photo-systems photosynthetic electron transfer governs transcrip-tion of reaction centre genes in chloroplasts Phil TransR Soc Lond B 355 1351ndash1357 (DOI 101098rstb20000697)
Allen J F amp Race H L 2002 Will the real LHC II kinaseplease step forward Sciencersquos STKE httpwwwstkeorgcgicontentfullsigtrans2002155pe43
Allen J F amp Raven J A 1996 Free-radical-induced mutationvs redox regulation costs and benefits of genes in organellesJ Mol Evol 42 482ndash492
Archibald J M Cavalier-Smith T Maier U amp Douglas S2001 Molecular chaperones encoded by a reduced nucleusthe cryptomonad nucleomorph J Mol Evol 52 490ndash501
Ashby M K amp Mullineaux C W 1999a Cyanobacterialycf27 gene products regulate energy transfer from phycobili-somes to photosystems I and II FEMS Microbiol Lett 181253ndash260
Ashby M K amp Mullineaux C W 1999b The role of ApcDand ApcF in energy transfer from phycobilisomes to PSI andPSII in a cyanobacterium Photosynth Res 61 169ndash179
Ashby M K Houmard J amp Mullineaux C W 2002 Theycf27 genes from cyanobacteria and eukaryotic algae distri-bution and implications for chloroplast evolution FEMSMicrobiol Lett 214 25ndash30
Attardi G amp Schatz G 1988 Biogenesis of mitochondria ARev Cell Biol 4 289ndash333
Azem A Oppliger W Lustig A Jeno P Feifel B SchatzG amp Horst M 1997 The mitochondrial hsp70 chaperone
Phil Trans R Soc Lond B (2003)
system effect of adenine nucleotides peptide substrate andmGrpE on the oligomeric state of mhsp70 J Biol Chem272 20 901ndash20 906
Barbrook A C Symington H Nisbet R E R LarkumA amp Howe C J 2001 Organisation and expression of theplastid genome of the dinoflagellate Amphidinium opercula-tum Mol Genet Genomics 266 632ndash638
Bauer C E Elsen S amp Bird T H 1999 Mechanisms forredox control of gene expression A Rev Microbiol 53495ndash523
Bauer C E Elsen S Swem L R Swem D L amp MasudaS 2003 Redox and light regulation of gene expression inphotosynthetic prokaryotes Phil Trans R Soc Lond B 358147ndash154 (DOI 101098rstb20021189)
Berks B C Sargent F De Leeuw E Hinsley A P Stan-ley N R Jack R L Buchanan G amp Palmer T 2000aA novel protein transport system involved in the biogenesisof bacterial electron transfer chains Biochim Biophys ActaBioenerg 1459 325ndash330
Berks B C Sargent F amp Palmer T 2000b The tat proteinexport pathway Mol Microbiol 35 260ndash274
Blackstone N W 1999 Redox control in development andevolution evidence from colonial hydroids J Exp Biol 2023541ndash3553
Blackstone N W 2000 Redox control and the evolution ofmulticellularity Bioessays 22 947ndash953
Blackstone N W 2001 Redox state reactive oxygen speciesand adaptive growth in colonial hydroids J Exp Biol 2041845ndash1853
Bogorad L 1975 Evolution of organelles and eukaryotic gen-omes Science 188 891ndash898
Bogsch E G Sargent F Stanley N R Berks B C Robin-son C amp Palmer T 1998 An essential component of a novelbacterial protein export system with homologues in plastidsand mitochondria J Biol Chem 273 18 003ndash18 006
Brown E C Somanchi A amp Mayfield S P 2001 Interor-ganellar crosstalk new perspectives on signaling from thechloroplast to the nucleus Genome Biol 2 10211ndash10214
Cavalier-Smith T 2002 Chloroplast evolution secondarysymbiogenesis and multiple losses Curr Biol 12 R62ndashR64
Chitnis P R amp Thornber J P 1988 The major light-har-vesting complex of photosystem 2 Aspects of its molecularand cell biology Photosynth Res 16 41ndash63
Chow W S Melis A amp Anderson J M 1990 Adjustmentsof photosystem stoichiometry in chloroplasts improve thequantum efficiency of photosynthesis Proc Natl Acad SciUSA 87 7502ndash7506
Claros M G Perea J Shu Y Samatey F A PopotJ L amp Jacq C 1995 Limitations to in vivo import of hydro-phobic proteins into yeast mitochondria The case of a cyto-plasmically synthesized apocytochrome b Eur J Biochem228 762ndash771
Dalbey R E amp Robinson C 1999 Protein translocation intoand across the bacterial plasma membrane and the plant thy-lakoid membrane Trends Biochem Sci 24 17ndash22
Daley D O Adams K L Clifton R Qualmann S MillarA H Palmer J D Pratje E amp Whelan J 2002 Genetransfer from mitochondrion to nucleus novel mechanismsfor gene activation from Cox2 Plant J 30 11ndash21
Danon A amp Mayfield S P 1994 Light-regulated translationof chloroplast messenger RNAs through redox potentialScience 266 1717ndash1719
de Grey A 2000 Mitochondrial gene therapy an arena for thebiomedical use of inteins Trends Biotechnol 18 394ndash399
Deisenhofer J Epp O Miki K Huber R amp Michel H1985 Structure of the protein subunits in the photosyntheticreaction center of Rhodopseudomonas viridis at 3Ac resolutionNature 318 618ndash624
34 J F Allen Genomes in mitochondria and chloroplasts
Delwiche C F 1999 Tracing the thread of plastid diversitythrough the tapestry of life Am Nat 154 S164ndashS177
Doolittle W F 1999 Phylogenetic classification and the uni-versal tree Science 284 2124ndash2129
Douglas S E 1998 Plastid evolution origins diversitytrends Curr Opin Genet Dev 8 655ndash661
Duby G amp Boutry M 2002 Mitochondrial protein importmachinery and targeting information Plant Sci 162 477ndash490
Eberhard S Drapier D amp Wollman F A 2002 Searchinglimiting steps in the expression of chloroplast encoded pro-teins relations between gene copy number transcriptiontranscript abundance and translation rate in the chloroplastof Chlamydomonas reinhardtii Plant J 31 149ndash160
El Bissati K amp Kirilovsky D 2001 Regulation of psbA andpsaE expression by light quality in Synechocystis species PCC6803 A redox control mechanism Plant Physiol 1251988ndash2000
Ellis R J 1984 The nuclear domination of chloroplast devel-opment Science Progress 69 129ndash142
Embley T M amp Martin W 1998 Molecular evolution ahydrogen-producing mitochondrion Nature 396 517ndash519
Ems S C Morden C W Dixon C K Wolfe K HdePamphilis C W amp Palmer J D 1995 Transcriptionsplicing and editing of plastid RNAs in the non-photosyn-thetic plant Epifagus virginiana Plant Mol Biol 29 721ndash733
Eraso J M amp Kaplan S 2000 From redox flow to gene regu-lation role of the PrrC protein of Rhodobacter sphaeroides241 Biochemistry 39 2052ndash2062
Eraso J M amp Kaplan S 2002 Redox flow as an instrumentof gene regulation Meth Enzymol 348 216ndash229
Escoubas J M Lomas M Laroche J amp Falkowski P G1995 Light-intensity regulation of cab gene-transcription issignaled by the redox state of the plastoquinone pool ProcNatl Acad Sci USA 92 10 237ndash10 241
Ferea T L Botstein D Brown P O amp Rosenzweig R F1999 Systematic changes in gene expression patterns follow-ing adaptive evolution in yeast Proc Natl Acad Sci USA96 9721ndash9726
Forsberg J Rosenquist M Fraysse L amp Allen J F 2001Redox signalling in chloroplasts and mitochondria genomicand biochemical evidence for two-component regulatory sys-tems in bioenergetic organelles Biochem Soc Trans 29403ndash407
Fujita Y 1997 A study on the dynamic features of photosys-tem stoichiometry accomplishments and problems forfuture studies Photosynth Res 53 83ndash93
Gabriel K Buchanan S K amp Lithgow T 2001 The alphaand the beta protein translocation across mitochondrial andplastid outer membranes Trends Biochem Sci 26 36ndash40
Galvis M L E Allen J F amp Hakansson G 1998 Proteinsynthesis by isolated pea mitochondria is dependent on theactivity of respiratory complex II Curr Genet 33 320ndash329
Gatenby A A amp Ellis R J 1990 Chaperone function theassembly of ribulose bisphosphate carboxylase-oxygenase ARev Cell Biol 6 125ndash149
GoldschmidtClermont M 1998 Coordination of nuclear andchloroplast gene expression in plant cells Int Rev CytolSurv Cell Biol 177 115ndash180
Gray M W 1992 The endosymbiont hypothesis revisited IntRev Cytol 141 233ndash357
Gray M W 1999 Evolution of organellar genomes CurrOpin Genet Dev 9 678ndash687
Gray M W 2000 Evolutionary biology mitochondrial geneson the move Nature 408 302ndash305
Gray M W amp Doolittle W F 1982 Has the endosymbionthypothesis been proven Microbiol Rev 46 1ndash42
Gray M W Burger G amp Lang B F 1999 Mitochondrialevolution Science 283 1476ndash1481
Phil Trans R Soc Lond B (2003)
Grebe T W amp Stock J B 1999 The histidine protein kinasesuperfamily Adv Microb Physiol 41 139ndash227
Green B R amp Kuhlbrandt W 1995 Sequence conservationof light-harvesting and stress-response proteins in relation tothe 3-dimensional molecular structure of Lhcii PhotosynthRes 44 139ndash148
Gruissem W amp Tonkyn J C 1993 Control mechanisms ofplastid gene expression Crit Rev Plant Sci 12 19ndash55
Hackstein J H P (and 11 others) 2001 Hydrogenosomesconvergent adaptations of mitochondria to anaerobicenvironments Zool Anal Complex Syst 104 290ndash302
Harrison M A Keen J N Findlay J B C amp Allen J F1990 Modification of a Glnb-like gene-product by photosyn-thetic electron-transport in the cyanobacterium Synechoc-occus 6301 FEBS Lett 264 25ndash28
Harvey A J Kind K L amp Thompson J G 2002 Redoxregulation of early embryo development Reproduction 123479ndash486
Heathcote P Fyfe P K amp Jones M R 2002 Reactioncentres the structure and evolution of biological solarpower Trends Biochem Sci 27 79ndash87
Henze K amp Martin W 2001 How do mitochondrial genesget into the nucleus Trends Genet 17 383ndash387
Herrmann R G amp Westhoff P 2001 Thylakoid biogenesis anddynamics the result of a complex phylogenetic puzzle InRegulation of photosynthesis vol 11 (ed E-M Aro amp BAndersson) pp 1ndash28 Dordrecht The Netherlands Kluwer
Hihara Y Sonoike K Kanehisa M amp Ikeuchi M 2002Search for redox responsive genes in Synechocystis sp PCC6803 by using DNA microarrays Plant Cell Physiol 43S178
Howe C J Barbrook A C amp Lockhart P J 2000 Organellegenes do they jump or are they pushed Trends Genet 1665ndash66
Ikeuchi M 2002 Gene expression and responses to light stressin cyanobacteria Plant Cell Physiol 43 S21
Iuchi S amp Lin E C C 1992 Mutational analysis of signaltransduction by Arcb a membrane sensor protein respon-sible for anaerobic repression of operons involved in the cen-tral aerobic pathways in Escherichia coli J Bacteriol 1743972ndash3980
Jackson-Constan D Akita M amp Keegstra K 2001 Molecu-lar chaperones involved in chloroplast protein importBiochim Biophys Acta Mol Cell Res 1541 102ndash113
Jarvis P amp Soll M 2001 Toc Tic and chloroplast proteinimport Biochim Biophys Acta Mol Cell Res 1541 64ndash79
Jordan P Fromme P Witt H T Klukas O SaengerW amp Krauss N 2001 Three-dimensional structure ofcyanobacterial photosystem I at 25 acngstrom resolutionNature 411 909ndash917
Kaneko T (and 23 others) 1996a Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regionsDNA Res 3 109ndash136
Kaneko T (and 23 others) 1996b Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regions[Supplement] DNA Res 3 185ndash209
Kanevski I amp Maliga P 1994 Relocation of the plastid rbcLgene to the nucleus yields functional ribulose-15-bisphosph-ate carboxylase in tobacco chloroplasts Proc Natl Acad SciUSA 91 1969ndash1973
Kim J amp Mayfield S P 1997 Protein disulfide isomerase as aregulator of chloroplast translational activation Science 2781954ndash1957
Genomes in mitochondria and chloroplasts J F Allen 35
Kirk J T O amp Tilney-Bassett R A E 1978 The plastidsTheir chemistry structure growth and inheritance Amster-dam Elsevier
Koehler C M Merchant S amp Schatz G 1999 How mem-brane proteins travel across the mitochondrial intermem-brane space Trends Biochem Sci 24 428ndash432
Komiya T Rospert S Schatz G amp Mihara K 1997 Bind-ing of mitochondrial precursor proteins to the cytoplasmicdomains of the import receptors Tom70 and Tom20 isdetermined by cytoplasmic chaperones FASEB J 11 1196
Konstantinov Y M Lutsenko G N amp Podsosonny V A1995 Genetic functions of isolated maize mitochondriaunder model changes of redox conditions Biochem MolBiol Int 36 319ndash326
Korab-Laskowska M Rioux P Brossard N LittlejohnT G Gray M W Lang B F amp Burger G 1998 Theorganelle genome database project (GOBASE) Nucleic AcidsRes 26 138ndash144
Kovacs L Wiessner W Kis M Nagy F Mende D ampDemeter S 2000 Short- and long-term redox regulation ofphotosynthetic light energy distribution and photosystemstoichiometry by acetate metabolism in the green alga Chla-mydobotrys stellata Photosynth Res 65 231ndash247
Kuhlbrandt W 1994 Structure and function of the plant light-harvesting complex LHC-II Curr Opin Struct Biol 4519ndash528
Lang B F Gray M W amp Burger G 1999 Mitochondrialgenome evolution and the origin of eukaryotes A RevGenet 33 351ndash397
Lemire B D Fankhauser C Baker A amp Schatz G 1989The mitochondrial targeting function of randomly generatedpeptide sequences correlates with predicted helical amphi-philicity J Biol Chem 264 20 206ndash20 215
Leuenberger D Bally N A Schatz G amp Koehler C M1999 Different import pathways through the mitochondrialintermembrane space for inner membrane proteins EMBOJ 18 4816ndash4822
Li H amp Sherman L A 2000 A redox-responsive regulator ofphotosynthesis gene expression in the cyanobacterium Syne-chocystis sp strain PCC 6803 J Bacteriol 182 4268ndash4277
Lin X Y (and 36 others) 1999 Sequence and analysis of chro-mosome 2 of the plant Arabidopsis thaliana Nature 402761ndash768
Lonsdale D M Hodge T P Howe C J amp Stern D B1983 Maize mitochondrial-DNA contains a sequence hom-ologous to the ribulose-15-bisphosphate carboxylase largesubunit gene of chloroplast DNA Cell 34 1007ndash1014
McFadden G I 1999 Endosymbiosis and evolution of theplant cell Curr Opin Plant Biol 2 513ndash519
McFadden G I 2001 Chloroplast origin and integrationPlant Physiol 125 50ndash53
Mahler H R amp Raff R A 1975 The evolutionary origin ofthe mitochondrion a nonsymbiotic model Int Rev Cytol43 1ndash124
Manfredi G Fu J Ojaimi J Sadlock J E Kwong J QGuy J amp Schon E A 2002 Rescue of a deficiency in ATPsynthesis by transfer of MTATP6 a mitochondrial DNA-encoded gene to the nucleus Nature Genet 30 394ndash399
Marc P Margeot A Devaux F Blugeon C Corral-Debrinski M amp Jacq C 2002 Genome-wide analysis ofmRNAs targeted to yeast mitochondria EMBO Rep 3159ndash164
Marienfeld J Unseld M amp Brennicke A 1999 The mito-chondrial genome of Arabidopsis is composed of both nativeand immigrant information Trends Plant Sci 4 495ndash502
Martin W 1999 A briefly argued case that mitochondria andplastids are descendants of endosymbionts but that thenuclear compartment is not Proc R Soc Lond B 2661387ndash1395 (DOI 101098rspb19990792)
Phil Trans R Soc Lond B (2003)
Martin W amp Herrmann R G 1998 Gene transfer fromorganelles to the nucleus how much what happens andwhy Plant Physiol 118 9ndash17
Martin W amp Muller M 1998 The hydrogen hypothesis forthe first eukaryote Nature 392 37ndash41
Martin W amp Russell M J 2003 On the origins of cells ahypothesis for the evolutionary transitions from abiotic geo-chemistry to chemoautotrophic prokaryotes and from pro-karyotes to nucleated cells Phil Trans R Soc Lond B 35859ndash85 (DOI 101098rstb20021183)
Martin W Stoebe B Goremykin V Hansmann S Hase-gawa M amp Kowallik K V 1998 Gene transfer to thenucleus and the evolution of chloroplasts Nature 393162ndash165
Martin W Hoffmeister M Rotte C amp Henze K 2001 Anoverview of endosymbiotic models for the origins of eukary-otes their ATP-producing organelles (mitochondria andhydrogenosomes) and their heterotrophic lifestyle BiolChem 382 1521ndash1539
Martin W Rujan T Richly E Hansen A Cornelsen SLins T Leister D Stoebe B Hasegawa M amp PennyD 2002 Evolutionary analysis of Arabidopsis cyanobacterialand chloroplast genomes reveals plastid phylogeny and thou-sands of cyanobacterial genes in the nucleus Proc NatlAcad Sci USA 99 12 246ndash12 251
Millen R S (and 12 others) 2001 Many parallel losses of infAfrom chloroplast DNA during angiosperm evolution withmultiple independent transfers to the nucleus Plant Cell 13645ndash658
Morse D Salois P Markovic P amp Hastings J W 1995 Anuclear-encoded form-II RUBISCO in dinoflagellatesScience 268 1622ndash1624
Murakami A Kim S J amp Fujita Y 1997 Changes in photo-system stoichiometry in response to environmental con-ditions for cell growth observed with the cyanophyteSynechocystis PCC 6714 Plant Cell Physiol 38392ndash397
Oh J I amp Kaplan S 2000 Redox signaling globalization ofgene expression EMBO J 19 4237ndash4247
Ozawa T 1995 Mitochondrial-DNA mutations associatedwith aging and degenerative diseases Exp Gerontol 30269ndash290
Palmer J D 1997 Organelle genomes going going goneScience 275 790ndash791
Pearson C K Wilson S B Schaffer R amp Ross A W 1993NAD turnover and utilization of metabolites for RNA syn-thesis in a reaction sensing the redox state of the cyto-chrome-b( 6 )f complex in isolated chloroplasts Eur JBiochem 218 397ndash404
Pfanner N amp Geissler A 2001 Versatility of the mitochon-drial protein import machinery Natl Rev Mol Cell Biol 2339ndash349
Pfanner N amp Wiedemann N 2002 Mitochondrial proteinimport two membranes three translocases Curr Opin CellBiol 14 400ndash411
Pfannschmidt T Nilsson A amp Allen J F 1999a Photosyn-thetic control of chloroplast gene expression Nature 397625ndash628
Pfannschmidt T Nilsson A Tullberg A Link G amp AllenJ F 1999b Direct transcriptional control of the chloroplastgenes psbA and psaAB adjusts photosynthesis to light energydistribution in plants IUBMB Life 48 271ndash276
Pfannschmidt T Allen J F amp Oelmuller R 2001a Prin-ciples of redox control in photosynthesis gene expressionPhysiol Plant 112 1ndash9
Pfannschmidt T Schutze K Brost M amp Oelmuller R2001b A novel mechanism of nuclear photosynthesis generegulation by redox signals from the chloroplast during pho-
36 J F Allen Genomes in mitochondria and chloroplasts
tosystem stoichiometry adjustment J Biol Chem 27636 125ndash36 130
Popot J L amp de Vitry C 1990 On the microassembly of inte-gral membrane proteins A Rev Biophys Biophys Chem 19369ndash403
Race H L Herrmann R G amp Martin W 1999 Why haveorganelles retained genomes Trends Genet 15 364ndash370
Raven J A Johnston A M Parsons R amp Kubler J 1994aThe influence of natural and experimental high O2 concen-trations on O2 evolving phototrophs Biol Rev CambridgePhil Soc 69 61ndash94
Raven J A Johnston A M Parsons R amp Kubler J 1994bThe occurrence and influence on photolithotrophs of highoxygen concentrations Proc R Soc Edinb B 102 193ndash201
Robinson C 2000 The twin-arginine translocation system anovel means of transporting folded proteins in chloroplastsand bacteria Biol Chem 381 89ndash93
Robinson C amp Bolhuis A 2001 Protein targeting by the twin-arginine translocation pathway Natl Rev Mol Cell Biol 2350ndash356
Rodermel S 2001 Pathways of plastid-to-nucleus signalingTrends Plant Sci 6 471ndash478
Roger A J 1999 Reconstructing early events in eukaryoticevolution Am Nat 154 S146ndashS163
Roger A J Svard S G Tovar J Clark C G SmithM W Gillin F D amp Sogin M L 1998 A mitochondrial-like chaperonin 60 gene in Giardia lamblia evidence thatdiplomonads once harbored an endosymbiont related to theprogenitor of mitochondria Proc Natl Acad Sci USA 95229ndash234
Roodyn D B amp Wilkie D 1968 The biogenesis of mitochondriaMethuenrsquos monographs on biological subjects London Methuen
Rujan T amp Martin W 2001 How many genes in Arabidopsiscome from cyanobacteria An estimate from 386 proteinphylogenies Trends Genet 17 113ndash120
Saccone C Gissi C Lanave C Larizza A Pesole G ampReyes A 2000 Evolution of the mitochondrial genetic sys-tem an overview Gene 261 153ndash159
Salzberg S L White O Peterson J amp Eisen J A 2001Microbial genes in the human genome lateral transfer orgene loss Science 292 1903ndash1906
Sargent F Berks B C amp Palmer T 2002 Assembly of mem-brane-bound respiratory complexes by the Tat protein-trans-port system Arch Microbiol 178 77ndash84
Schatz G 1998 Protein transport the doors to organellesNature 395 439ndash440
Schemidt R A Qu J Williams J R amp Brusilow W S1998 Effects of carbon source on expression of F0 genes andon the stoichiometry of the c subunit in the F1 F0 ATPaseof Escherichia coli J Bacteriol 180 3205ndash3208
Selosse M A Albert B R amp Godelle B 2001 Reducing thegenome size of organelles favours gene transfer to thenucleus Trends Ecol Evol 16 135ndash141
Stern D B amp Lonsdale D M 1982 Mitochondrial and chlo-roplast genomes of maize have a 12- kilobase DNA-sequencein common Nature 299 698ndash702
Stock D Gibbons C Arechaga I Leslie A G W ampWalker J E 2000 The rotary mechanism of ATP synthaseCurr Opin Struct Biol 10 672ndash679
Stoebe B amp Maier U G 2002 One two three naturersquos toolbox for building plastids Protoplasma 219 123ndash130
Stoebe B Martin W amp Kowallik K V 1998 Distributionand nomenclature of protein-coding genes in 12 sequencedchloroplast genomes Plant Mol Biol Rep 16 243ndash255
Stupar R M Lilly J W Town C D Cheng Z Kaul SBuell C R amp Jiang J M 2001 Complex mtDNA consti-tutes an approximate 620 kb insertion on Arabidopsis thali-ana chromosome 2 implication of potential sequencing
Phil Trans R Soc Lond B (2003)
errors caused by large-unit repeats Proc Natl Acad SciUSA 98 5099ndash5103
Surpin M Larkin R M amp Chory J 2002 Signal transduc-tion between the chloroplast and the nucleus Plant Cell 14S327ndashS338
Tjaden J Schwoppe C Mohlmann T Quick P W ampNeuhaus H E 1998 Expression of a plastidic ATPADPtransporter gene in Escherichia coli leads to a functional aden-ine nucleotide transport system in the bacterial cytoplasmicmembrane J Biol Chem 273 9630ndash9636
Tovar J Fischer A amp Clark C G 1999 The mitosome a novelorganelle related to mitochondria in the amitochondrial para-site Entamoeba histolytica Mol Microbiol 32 1013ndash1021
Trebitsh T amp Danon A 2001 Translation of chloroplastpsbA mRNA is regulated by signals initiated by both photos-ystems II and I Proc Natl Acad Sci USA 98 12 289ndash12 294
Tullberg A Alexciev K Pfannschmidt T amp Allen J F2000 Photosynthetic electron flow regulates transcription ofthe psaB gene in pea (Pisum sativum L) chloroplasts throughthe redox state of the plastoquinone pool Plant Cell Physiol41 1045ndash1054
Turmel M Lemieux C Burger G Lang B F Otis CPlante I amp Gray M W 1999 The complete mitochondrialDNA sequences of Nephroselmis olivacea and Pedinomonasminor two radically different evolutionary patterns withingreen algae Plant Cell 11 1717ndash1729
Unden G amp Bongaerts J 1997 Alternative respiratory path-ways of Escherichia coli energetics and transcriptional regu-lation in response to electron acceptors Biochim BiophysActa Bioenerg 1320 217ndash234
Unseld M Marienfeld J R Brandt P amp Brennicke A1997 The mitochondrial genome of Arabidopsis thaliana con-tains 57 genes in 366 924 nucleotides Nature Genet 1557ndash61
Van der Giezen M Slotboom D J Horner D S DyalP L Harding M Xue G P Embley T M amp KunjiE R S 2002 Conserved properties of hydrogenosomal andmitochondrial ADPATP carriers a common origin for bothorganelles EMBO J 21 572ndash579
Von Heijne G 1986 Why mitochondria need a genome FEBSLett 198 1ndash4
Westphal S Soll J amp Vothknecht U C 2001 A vesicle trans-port system inside chloroplasts FEBS Lett 506 257ndash261
Whitehouse H L K 1969 Towards an understanding of themechanism of heredity London Edward Arnold Ltd
Williams B A P Hirt R P Lucocq J M amp Embley T M2002 A mitochondrial remnant in the microsporidian Trachi-pleistophora hominis Nature 418 865ndash869
Wilson R J (and 10 others) 1996a Complete gene map of theplastid-like DNA of the malaria parasite Plasmodium falcipa-rum J Mol Biol 261 155ndash172
Wilson S B Davidson G S Thomson L M amp PearsonC K 1996b Redox control of RNA synthesis in potato mito-chondria Eur J Biochem 242 81ndash85
Winkler H H amp Neuhaus H E 1999 Non-mitochondrialATP transport Trends Biochem Sci 24 64ndash68
Wolfe K H Morden C W amp Palmer J D 1992 Small sin-gle-copy region of plastid DNA in the non-photosyntheticangiosperm Epifagus virginiana contains only two genes Dif-ferences among dicots monocots and bryophytes in geneorganization at a non-bioenergetic locus J Mol Biol 22395ndash104
Wollman F A Minai L amp Nechushtai R 1999 The biogen-esis and assembly of photosynthetic proteins in thylakoidmembranes Biochim Biophys Acta Bioenerg 1411 21ndash85
Yeliseev A A amp Kaplan S 2000 TspO of Rhodobactersphaeroides a structural and functional model for the mam-
Genomes in mitochondria and chloroplasts J F Allen 37
malian peripheral benzodiazepine receptor J Biol Chem275 5657ndash5667
Zerges W 2002 Does complexity constrain organelle evol-ution Trends Plant Sci 7 175ndash182
Zhang Z D Green B R amp Cavalier-Smith T 1999 Singlegene circles in dinoflagellate chloroplast genomes Nature400 155ndash159
Zhang Z D Cavalier-Smith T amp Green B R 2001 A fam-ily of selfish minicircular chromosomes with jumbled chloro-plast gene fragments from a dinoflagellate Mol Biol Evol18 1558ndash1565
Zouni A Witt H T Kern J Fromme P Krauss NSaenger W amp Orth P 2001 Crystal structure of photosys-tem II from Synechococcus elongatus at 38 acngstrom resol-ution Nature 409 739ndash743
DiscussionD S Horner (Dipartimento di Fisiologia e Biochimica
Generali University of Milan Milan Italy) Given thatthere are components of these membrane-bound energy-transduction pathways that are consistently nuclear enco-ded not the key ones you pointed out but others do youexpect perhaps a slower perhaps a more crude but yet aredox communication between the organelle andexpression of nuclear-encoded genes
J F Allen There is and there is no question about itI think that Bogoradrsquos lock-in hypothesis has somethingto say here and there are other people who say it is all todo with assembling multi-subunit complexes If you havecontrol over the initial construction of the core aroundwhich the whole complex is eventually built then redoxcontrol or some sort of control over synthesis of that corewill control construction of the whole thing eventuallyand I think that may well be largely correct I think it ispossible in principle for nuclear genes to make precursorsaccording to redox instructions but it is not really soimportant that those precursors are here and now presentin exactly the right quantities because the precursors canto a limited extent hang around and wait Actually thereis proteolytic degradation of unattached precursors and soon but I think the fact that these chloroplast and mito-chondrially DNA-encoded subunits are at the core ofthese energy-transducing complexes is the crucial issue
T Cavalier-Smith (Department of Zoology University ofOxford Oxford UK ) In the case of mitochondria the sub-set of genes that remains is much more variable than youimplied It runs from three proteins in the minimal casesup to I think over 90 That is a big difference and manyof the genes in different lineages of protists that remainhave nothing to do with redox control they are things likeribosomal proteins and so on So I think that even thoughwhat you say about redox control of certain genes mayactually be an important thing it is not the fundamentaloverall reason why the genomes are maintained It may bepart of the explanation for why certain genes are alwaysincluded in that subset but even in mitochondria I thinkthat cytochrome b is the only one that is always presentSo I think that both this great diversity of what has beenretained in different lineages and the nucleomorph caseindicate that evolution was able to do one thing in onelineage 500 million years ago and never got around todoing it for 500 million years in another lineage so thereis a lot more haphazardness and evolutionary accidentsand contingency than a straight functional interpretation
Phil Trans R Soc Lond B (2003)
there is a lot of just sheer lsquohappenedndashorndashdidnrsquotndashhappenrsquoor carryover
J F Allen I was not implying anything about the rangeof mitochondrial genomes I have a limited amount oftime so I just talked about what I could talk about withinthat limit Of course there will be a range and this willdepend upon the physiology and bioenergetics of theparticular organism But what I am trying to do is to seewhat the common properties of the irreducible core areof components that are always encoded in the organellesand cytochrome b is a perfect example because that reallyis the thing that binds these two all-important quinoneswithout which there is no Q cycle without which there isno proton pumping and bioenergetics and in Plasmodiumwhere the mitochondrion is reduced to this bare mini-mum there is still a mitochondrial gene for cytochromeb Now my argument about the ribosomal proteins andall the rest of it is that in order to be able to exert thisall-important redox control over gene expression of certaincore components you need an in situ genetic system andthis can vary in composition for reasons that I cannotdescribe Maybe you need RNA polymerase or maybeyou can import RNA polymerase In plastids you haveboth of these options You are going to need ribosomestransfer RNAs and the rest of the apparatus of geneexpression and that needs to be in situ but we are stillleft with the question why does there have to be any gen-etic system at all The mature tobacco or spinach chloro-plast seems to have an entire genetic system just to makeone protein D1 which is all it ever needs to resynthesizeI think that this is a clue It is worth having this very unec-onomical crazy system because there is an overridingrequirement to connect electron transfer with geneexpression directly and unambiguouslymdashwithout recourseto a bureaucracy if you like which is what I think thenucleus is
K Willison (Institute of Cancer Research Chester BeattyLaboratories London UK ) These photosynthetic reactioncomplexes are a sort of baroque mixture of polypeptidechains and cofactors and my understanding is that someof them in high photon flux can turn over within say 30minutes so I am wondering if there is a correlationbetween gene retention and the turnover rates of some ofthe components and the complexes it seems to me thatthat could really be a limiting factor and the reason thatthe proteins had to be encoded locally in order to get thecomplexes to be assembled rapidly
J F Allen I think that is right You can only alter thesteady-state concentration of a protein complex by affect-ing the synthesis and breakdown and therefore turnoverThere is an extreme example the D1 protein encoded bythe psbA gene of chloroplasts which is always in the chlor-oplast genome without exception High light will tend todegrade this protein and when the high light goes awaythere is an absolute need very quickly to bring it back upagain I think that is a very good example Whether thisargument applies to genes such as cytochrome oxidase Ido not know
C J Leaver (Department of Plant Sciences University ofOxford Oxford UK ) Your experiments tend to relatemainly to the gene expression side What about proteinRNA turnover the whole concept of redox control ofthatmdashdo we know anything
38 J F Allen Genomes in mitochondria and chloroplasts
J F Allen Well quite a lot is known about chloro-plasts There is redox control of messenger RNA stabilityfor this same D1 protein which has been described byStephen Mayfield and his co-workers The evidence is thatthe site of redox control is ferredoxin via thioredoxin andthe implication is that thioredoxin tells whether non-cyclicelectron transport is working at all When it is workingthere is a signal that says lsquoOK now continue with all theprotein synthesisrsquo It is not particularly specific but moreof a global effect I think this is what Mayfield argues butof course this is not inconsistent with my argument I haveconcentrated on quinone pools and medium-potentialredox regulatory control because that is where I can seesignals about the dangerous consequences of getting itwrong but there is control of other levels of geneexpression In mitochondria I am not so sure I wouldbounce the question back to you on that one
W Martin (Institute of Botany III Heinrich-Heine Univ-ersitat Dusseldorf Dusseldorf Germany) The nucleomorphgenome was cited as evidence in conflict with your modelbut is that true The nucleomorph genome is not in abioenergetic organelle in that sense It resides in the cyto-sol so does the size of the nucleomorph genome workagainst your model Is it really evidence against yourmodel
J F Allen I do not know why should it be evidenceI cannot explain why the nucleomorph has a genomeThat is why I clutched at the straw that it is storage fornucleic acid precursors
Phil Trans R Soc Lond B (2003)
T Cavalier-Smith The reason I introduced the nucleo-morph is not to say it is direct evidence against yourhypothesis but to show that there probably was no singlefunctional reason for the retention of every gene in everyorganelle It may be that there are specific physiologicalreasons for the retention of certain genes and thereforeyou might be right about a subset of genes but I verymuch doubt if there are specific physiological reasons forthe retention of every single gene that is retained differen-tially because odd things happen in evolution Somethings are easy and sometimes things are difficult Evol-ution is history
J F Allen Of course I agree with that I do not thinkI can explain everything but I do think I can explainsomething If I have done that then I am content
GLOSSARY
AAC ADPndashATP carrierCES control by epistasy of synthesisCORR co-location for redox regulationLHC light-harvesting complexPCC Pasteur culture collectionORF open reading frameRUBISCO ribulose-15-bisphosphate carboxylasendashoxygenaseTAT twin-arginine translocase
Visit httpwwwpubsroyalsocacuk to see an electronic appendix tothis paper

Genomes in mitochondria and chloroplasts J F Allen 25
has been demonstrated (Pfannschmidt et al 1999abAllen amp Pfannschmidt 2000 Tullberg et al 2000) and wasprompted by this principle
The level of redox control of chloroplast transcriptionestablished unequivocally so far is that of plastoquinonean electron and hydrogen atom carrier located strategicallybetween photosystems I and II of photosynthesis Thedirections in which the effects are exerted are functionallyintelligible reduction of plastoquinone increases photosy-stem I transcription and decreases photosystem IItranscription oxidation of plastoquinone decreases photo-system I transcription and increases photosystem II tran-scription Plastoquinone in normal photosynthesis isreduced by the action of photosystem II and oxidized bythat of photosystem I Therefore redox control of chloro-plast gene expression provides a compensatory response(Pfannschmidt et al 1999a) and adds a previously mis-sing part of an explanation for the ability of plants andalgae to adjust the ratio of the two photosystems inresponse to changes in light quality (Chow et al 1990)By the same argument a functionally equivalent post-translational mechanism of redox control of light-harvesting function by photosystems I and II may havebeen conserved during the transition from prokaryotes toeukaryotes (Allen 1992 1995 Allen amp Nilsson 1997Allen amp Race 2002)
The precedent of chloroplast redox control of transcrip-tion (Pfannschmidt et al 1999ab) taken together with theprinciple in sect 3d predicts plastoquinone redox control ofcyanobacterial reaction centre gene transcription and thisis borne out by experiment (El Bissati amp Kirilovsky2001) This complementary self-adjusting transcriptionalresponse (Fujita 1997 Murakami et al 1997 El Bissati ampKirilovsky 2001) indicates strongly that redox control ofphotosynthetic reaction centre gene transcription waspresent in the proto-chloroplast Thus the mechanism ofregulation is likely to have been conserved throughout theevolutionary transition from bacterial symbiont to cyto-plasmic organelle A conserved mechanism of redox regu-lation of transcription in photosynthetic prokaryotes(Unden amp Bongaerts 1997 Bauer et al 1999) and chloro-plasts (Allen 1993c Pfannschmidt et al 1999b) is pre-dicted if evolutionary continuity of the process has beenmaintained
Thus apart from being consistent with experimentaldata the principle of continuity of redox control has beenof heuristic value in pointing to the underlying bacterialcharacter previously unsuspected in the control of geneexpression especially of transcription in chloroplasts ofhigher plants Other possible precedents for redox signal-ling components of cytoplasmic organelles include the prr(Eraso amp Kaplan 2000 2002 Oh amp Kaplan 2000) andreg (Bauer et al 1999) redox regulatory systems of purplenon-sulphur bacteria The TspO lsquooxygen sensorrsquo of Rho-dobacter sphaeroides is present in mammalian mitochondriaas a protein known there as a benzodiazepine receptorand each protein will complement loss of the other(Yeliseev amp Kaplan 2000) As outlined in sect 3j genes forsuch components are to be expected primarily in nucleargenomes This independent evolutionary hypothesis (sect 3d)would be strengthened still further by a demonstration ofone or more decisive redox regulatory links shared by pho-tosynthetic prokaryotes and all eukaryotes with mitochon-
Phil Trans R Soc Lond B (2003)
dria and where present with chloroplasts (Bauer et al2003)
(e) The principle of the selective value of redoxcontrol
For each gene under redox control (sect 3d) it is selectivelyadvantageous for that gene to be retained and expressed onlywithin the organelle
The twin selective advantages of redox control are likelyto be energetic efficiency and the suppression of the harm-ful side effects of electron transport operating on thewrong substrates No direct experimental support for thisprinciple is available and equally there is no evidenceagainst it Allotopic mutants in which genes have beenexperimentally relocated from organelles to the nucleushave been produced in yeast higher plants (Kanevski ampMaliga 1994) and human cells (Manfredi et al 2002) andare predicted by CORR to be poor adapters to environ-mental change Some authors believe there is potentialtherapeutic value in allotopic transformation movingmitochondrial genomes to the nucleus (Kanevski ampMaliga 1994 de Grey 2000) and predict the possibilityof r2 human cell lines in the foreseeable future (de Grey2000) This principle also has implications for ageing andthe evolution of separate sexes (Allen 1996) and possibleconsequences for development (Blackstone 1999 20002001 Harvey et al 2002) and somatic cloning in animals(Allen amp Allen 1999)
(f ) The principle of the selective value of nuclearlocation for genes not under redox control
For each bacterial gene that survives and is not under redoxcontrol it is selectively advantageous for that gene to be locatedin the nucleus and expressed only in the nucleus and cytosolIf the mature gene product functions in chloroplasts or mito-chondria the gene is first expressed in the form of a precursorfor import
There are several possible reasons for the selectiveadvantage of nuclear location of genes for organellar pro-teins One is the decreased probability of mutation arisingfrom free radical by-products of lsquoincorrectrsquo electron trans-fers (Raven et al 1994ab Allen amp Raven 1996) Therewould seem to be obvious economies in resource use andadvantages in coordination of gene expression if genes arekept in one compartment One might say that the bipartiteor tripartite genetic system of eukaryotic cells is untidyand inefficient
Another suggestion is that organellar genes do notundergo recombination and are present in relatively smallcopy numbers so that disadvantageous mutations willspread relatively quickly through a clonal population oforganelles Incorporation of functional gene copiesinstead into the nuclear genome may then be an irrevers-ible step analogous to the click of a ratchet (Saccone et al2000) There are examples of genes shared between allthree compartments (Stern amp Lonsdale 1982 Lonsdale etal 1983)
The cost of retaining an organellar genetic system is notincreased appreciably when additional genes are retainedin organelles if just one gene is retained then the cost ofa cytoplasmic genetic system is incurred and there may berelatively little extra cost for each additional gene retained(Allen amp Raven 1996) For a functional explanation of the
26 J F Allen Genomes in mitochondria and chloroplasts
existence of chloroplast and mitochondrial genomes thefocus of attention should therefore be on the relativelyconstant primary subset of bioenergetic genes (sect 3e)
(g) The principle of contemporary operation ofselection on gene location
For any species the distribution of genes between organelle(by the principle in sect 3e) and nucleus (by the principle in sect 3f)is the result of selective forces that continue to operate
This principle is essentially a statement contradictingboth the lsquofrozen accidentrsquo hypothesis and the hypothesisthat gene relocation is progressive predictable and stillincomplete (sect 4) In support of this principle it is becom-ing increasingly clear that mitochondria that lose theirfunction in aerobic respiration also lose their genomes asseen in the relict mitochondria of microsporidia(Embley amp Martin 1998 Van der Giezen et al 2002) andthe lsquomitosomersquo of Entamoeba histolytica (Tovar et al1999) In chloroplasts loss of photosynthesis results inloss of photosynthetic genes The examples of Epifagus (aparasitic higher plant) (Ems et al 1995 Wolfe et al 1992)and the residual apicomplexan plastids of Plasmodium(Wilson et al 1996a) and Toxoplasma where some plas-tidic genetic system is retained mean that some of theirgene products must be subject to redox regulation of geneexpression if this principle is correct Experimental test ofthis principle is possible using allotopic mutants as forthe predictions of the principle in sect 3f The evidence thatthe apicoplast (Wilson et al 1996a) is a plastid is largelygenetic the residual genome resembles that of photosyn-thetic plastids (chloroplasts) in gene organization ratherthan in content A plastid that has completely lost its gen-ome by virtue of having lost photosynthesis and all otherprocesses requiring redox regulation might be difficult todistinguish from an organelle derived in a similar way froma mitochondrion The persistence of targeted importedchaperones Hsp70 (Tovar et al 1999 Williams et al2002) and cp60 (Roger et al 1998) in residual organellesmight not distinguish a plastid ancestry from a mitochon-drial one
(h) The principle of primary involvement inenergy transduction
Those genes for which redox control is always vital to cellfunction have gene products involved in or closely connectedwith primary electron transfer These genes are always con-tained within the organelle
This principle would be violated and the CORRhypothesis refuted if any photosynthetic reaction centrecore subunit or respiratory chain subunit currentlyorganelle-encoded is found that functions in primary vec-torial electron transport and is nevertheless encoded in thenucleus and synthesized cytosolically as a precursor forimport all without any selective cost to the individualFrom the chloroplast transcriptional experiments ofPfannschmidt et al (1999ab) the chloroplast geneswhose transcription is regulated by the plastoquinoneredox state include psaAB (for photosystem I reactioncentre core polypeptides) psbA (for the photosystem IIpolypeptide D1) and rbcL (for the large subunit of RUB-ISCO interestingly following the same pattern of redoxcontrolmdashupregulated by oxidized plastoquinonemdashaspsbA) Transcription of the chloroplast genes petA
Phil Trans R Soc Lond B (2003)
(cytochrome f ) trnG (glycine tRNA) and rpoB (RNApolymerase) did not respond to plastoquinone redox statepetA for cytochrome f is predicted by this principle to beregulated at a different level of redox control or if at plas-toquinone at a different stage in gene expression Thisprediction might also apply to trnG and rpoB though thesequalify as members of the secondary subset that of geneticsystem genes to which the principle in sect 3i applies
(i) The principle of secondary involvement inenergy transduction
Genes whose products contribute to the organelle genetic sys-tem itself or whose products are associated with secondaryevents in energy transduction may be contained in theorganelle in one group of organisms but not in anotherdepending on the physiology and biochemistry of photosynthesisand respiration in the species concerned
Even phylogenetically moderately related species(within kingdoms) show differences in the location ofsome genes for example the ATP synthase subunits ofNeurospora and Saccharomyces (Attardi amp Schatz 1988)The CORR hypothesis also does not explicitly account forthe wide disparities in the distribution of genetic systemgenes for example the almost complete import of tRNAsin Chlamydomonas (GoldschmidtClermont 1998) Trans-port of nucleic acids across membranes is uncommon andwhy some RNAs should be organelle-encoded in somespecies but nuclearly encoded in others is not currentlyunderstood The only suggestion that can be made alongthe lines of CORR is that in some genetic systems genesmay need to be under the control of a regulatory systemthat responds to a signal arising within the organelle itselfThis signal could be the primary signal of altered redoxstate or some secondary signal Among the latter might befound signals that themselves report on the current stateof gene expression another piece of information that thenucleus might not have at its immediate disposal Suchsecondary signals could result at least in part fromCORR though other developmental and environmentalinputs may be imagined and would not detract fromCORR unless it could be shown that they provided a suf-ficient reason for the retention of the genetic system asa whole
(j) The principle of the nuclear encoding of redoxsignalling components
Components of the redox-signalling pathways upon whichCORR depends are themselves not involved in primary electrontransfer and so their genes have been relocated to the nucleusin accordance with sect 3f
This principle states that the redox signalling pathwaysthat are required for CORR should not themselves beexpected to utilize components whose biosynthesis mustbe under direct redox control Such components shouldtherefore fall into the major category of organellar pro-teins and be imported as precursors from the cytosol inline with the principle in sect 3f However the principleabove might be need to be modified in cases where ampli-fication of the redox signal is useful to the cell In thebacterial ntr system which was a precedent for CORR(Harrison et al 1990 Allen 1992) the histidine kinasentrB and aspartate response regulator ntrC form part ofthe gln operon whose transcription they control Odontella
Genomes in mitochondria and chloroplasts J F Allen 27
and Porphyra have plastids that encode more proteins thanthose of most chlorophytes (Stoebe et al 1998 Turmel etal 1999) and these proteins include two-component sig-nal transduction proteins prr and ycf27 (Stoebe et al1998) Extreme or extremely rapid environmental fluctu-ations might be catered for by amplification of signals thatare transmitted by signalling pathways whose componentsare under their own control It is not known whether ycf27and prr are in operons whose promoters their gene pro-ducts control It is also uncertain whether ycf27 is a redoxsignalling component and its distribution at least in itsplastidic location seems to be correlated with the presenceof a phycobilin-based light-harvesting system as found inred algae and cyanobacteria (Ashby amp Mullineaux1999ab Ashby et al 2002) The best guess of the originalstatement of the CORR hypothesis (Allen 1992 1993ac)was that bacterial two-component systems (Iuchi amp Lin1992 Allen 1993c Grebe amp Stock 1999) provide a plaus-ible means of exerting redox regulatory control over mito-chondrial and chloroplast gene expression andsubsequent research confirms the presence of two-compo-nent genes in genomes of Arabidopsis and yeast SeveralArabidopsis two-component genes are predicted to be tar-geted to mitochondria and chloroplasts (Forsberg et al2001) These must be considered as prime candidates forcomponents involved in redox regulation of chloroplasttranscription seen in the experiments of Pfannschmidt etal (1999a 2001a) The presumably homologous regulat-ory process serving to control photosystem stoichiometryin cyanobacteria the prr system (Li amp Sherman 2000) isnow characterized as a two-component system involvinga redox sensor and redox response regulator as originallydefined (Allen 1992 1993ac)
4 CONTRASTING HYPOTHESES CONCERNINGTHE RETENTION OF CYTOPLASMIC GENOMES
CORR is a recent addition to explanations that havebeen put forward to account for the persistence of gen-omes in organelles These models and their underlyingmechanisms are now discussed and compared
(a) There is no reason that is just how it isOne reaction to the problem of cytoplasmic genetic sys-
tems (sect 2) is that there is no problem and no particularreason for their retention However there is a very widephylogenetic distribution of a common core or primarysubset of organellar-encoded genes and one which haswithstood considerable genomic reorganization over evol-utionary time This constancy is seen for example in thesame genes being present in the chloroplasts of landplants with multiple copies of a single circular chloroplastchromosome and in dinoflagellate chloroplasts with acollection of different DNA minicircles each with essen-tially one gene (Zhang et al 1999 2001 Barbrook et al2001) Another suggested explanation of the retention ofmitochondrial and chloroplast genomes is simply torestate the endosymbiont hypothesis and to point out thatchloroplast and mitochondrial genes lsquocame inrsquo with thebacterial endosymbiont This suggestion however doesnot address the question of why some genes were retainedwhereas others were not It is also now clear that mostgenes that came in and still survive have nuclear and cyto-
Phil Trans R Soc Lond B (2003)
solic gene products whereas many chloroplast and mito-chondria proteins had a eukaryotic origin (Salzberg et al2001 Martin amp Russell 2003) The odds against a con-served pattern in gene location (eg table 1) arising bychance must be astronomical Thus there is a reason forthe retention of organellar genomes and a reason for theretention of the same primary subset of genes Becauseevolution proceeds by natural selection of chance vari-ation lsquoreasonrsquo in this context requires identification of thefunction of the location of a gene that is the selectivevalue of the phenotype produced by that location Evol-ution sometimes retains apparently functionless relics ofpreviously useful features but not for long especially ifthere is a cost
(b) The lock-in hypothesisThe lsquolock-inrsquo hypothesis of Bogorad (1975) was pro-
posed independently of the endosymbiont hypothesis anddoes not depend upon it According to the lock-in hypoth-esis core components of multi-subunit complexes must besynthesized de novo in the correct cellular compartmentotherwise photosystems or respiratory chain complexesfor example might assemble in the plasma membranevacuolar membrane or endoplasmic reticulum The lock-in hypothesis made no clear prediction about the conti-nuity of cytoplasmic genetic systems even though cyto-plasmic uniparental non-Mendelian inheritance for bothmitochondria (Roodyn amp Wilkie 1968) and plastids(Kirk amp Tilney-Bassett 1978) made it likely that organellarDNA the chondriome and the plastome replicated inde-pendently of nuclear DNA (Whitehouse 1969)
Mechanisms of protein import and targeting (Schatz1998 Koehler et al 1999 Leuenberger et al 1999 Gab-riel et al 2001 Jarvis amp Soll 2001 Pfanner amp Geissler2001 Duby amp Boutry 2002 Pfanner amp Wiedemann 2002)now effectively answer the problem of specifying the cor-rect locations for membrane protein complexes for whichthe lock-in hypothesis was proposed as a solution How-ever regulation of synthesis of an entire functional multi-subunit complex by control of synthesis of individual sub-units is still an important principle This seems to be afeature of the lsquoCESrsquo model (control by epistasy ofsynthesis) of Wollman (Wollman et al 1999 Eberhard etal 2002) which envisages nuclear control as all-embracingwith organellar genetic systems doing as it were whatthey are told In the hypothesis of CORR propoundedhere CES is equally important but the essence of CORRis that the organellar genetic system alone knows what arethe immediate and most important signals of environmen-tal change The organelle may keep the nucleus informedbut this is a lower priority than initiating an appropriateresponse CES and CORR agree that assembly of multi-subunit complexes is a complex business and requirescoordinated control in which no step is rate-limiting butmany may be obligatory
(c) Going going gone the evolutionary process oftransfer of genes from organelles to the
nucleus is still underway but incompleteThis hypothesis envisages that for most eukaryotes we
are now witnessing the last stages in a slow evolutionarytransfer of genes to the nucleus and this transfer may beexpected to proceed to completion in what is in evol-
28 J F Allen Genomes in mitochondria and chloroplasts
utionary terms the near future (Palmer 1997 Gray 19992000 Gray et al 1999 Adams et al 2000 Daley et al2002) A clear and compelling statement of this widelyheld position is that of Herrmann amp Westhoff (2001 p10)
hellipthe transfer of geneshelliptells that the partite plant gen-ome is not in a phylogenetic equilibrium All availabledata suggest that the ultimate aim of genome restructur-ing in the plant cell as in the eukaryotic cell in generalis the elimination of genome compartmentation whileretaining physiological compartmentation
One serious problem with this hypothesis is the dis-tinctly non-random sample of genes that remain to betransferred A relatively constant subset of genes is foundin bioenergetically functional mitochondria from fungi tomammals and angiosperms as in photosynthetic plastids(chloroplasts) from unicellular algae to angiosperms Thishypothesis therefore requires an additional ad hocassumption that some gene transfers are inherently moreprobable than others (this too is widely supposed seelater) and thus that gene transfer events proceed in thesame sequence in quite independent lineages Thereseems to be no evidence for this and the order of geneloss from mitochondria and chloroplasts has been used toinfer phylogeny (Martin et al 1998 Adams et al 2000Millen et al 2001) lsquoGoing going gonersquo (Palmer 1997)is difficult to refute categorically but it does not seem tobe consistent with recent evidence For example chromo-some two of Arabidopsis thaliana (Lin et al 1999) has asingle insertion encompassing with about 99 sequencesimilarity the complete Arabidopsis mitochondrial genome(Unseld et al 1997 Marienfeld et al 1999 Stupar et al2001) Unless this single transfer is a freak occurrencethis finding strongly indicates that gene transfer occurswith relative ease (Adams et al 2000 Henze amp Martin2001 Rujan amp Martin 2001 Salzberg et al 2001) Inaddition there is experimental evidence that randomlygenerated polypeptides are mitochondrially imported athigh frequency (Lemire et al 1989) The hypothesis thatwe now see the same few stragglers in diverse organellargenomes is not impossible but should be regarded as alast resort perhaps applicable only in extreme cases suchas the greatly reduced apicomplexan plastomes and chon-driomes Even there the hypothesis carries with it an obli-gation to explain why organisms with few organellar geneshave lsquoevolvedrsquo faster than those with many
(d ) The frozen accidentWhat may be described as lsquothe frozen accident hypoth-
esisrsquo states that the evolutionary process of gene transferfrom organelle to nucleus was underway when somethinghappened that stopped it One example of the frozen acci-dent is a scenario (Von Heijne 1986) in which import intomitochondria of the precursor proteins arising from newlytranslocated genes proceeded for long enough after theprimordial endosymbiosis for most of the symbiont-derived genes to be moved to the cell nucleus The lsquoacci-dentrsquo that halted the process was the evolutionary originof exocytosis and protein secretion after which it becameimpossible for precursors properly destined for mitochon-drial import to be distinguished from those destined forexport from the cell As with the lsquolock-inrsquo hypothesis the
Phil Trans R Soc Lond B (2003)
lsquofrozen accidentrsquo seems incompatible with the remarkableprecision and specificity of protein targeting (Schatz 1998Koehler et al 1999 Leuenberger et al 1999 Gabriel etal 2001 Jarvis amp Soll 2001 Pfanner amp Geissler 2001Duby amp Boutry 2002 Pfanner amp Wiedemann 2002) Pro-tein sorting mechanisms do not seem to forbid controlledimport into any cellular compartment Another lsquoaccidentrsquothat may have impeded subsequent gene transfer in thecase of mitochondria is their departure from what is other-wise a universal genetic code Genetic code differences arediscussed later in the context of whether some genes can-not be moved
(e) Some proteins (with cofactors) cannot beimported
We may describe as the lsquounimportability hypothesisrsquo theproposal that some proteins are inherently unimportableand must therefore be synthesized de novo in the correctcompartment within the cell The unimportability hypoth-esis is most often proposed as its special case the hydro-phobicity hypothesis with hydrophobicity being thereason for unimportability
(i) HydrophobicityA commonly entertained notion is that proteins that are
encoded and synthesized within organelles comprise a pri-mary subset characterized by shared hydrophobicity allare intrinsic membrane proteins (Von Heijne 1986Popot amp de Vitry 1990 Claros et al 1995) As with theCORR hypothesis favoured here the primary subset thenrequires a secondary subset of genetic system componentsthe latter not themselves being hydrophobic but beingrequired for synthesis of the others Certainly the mito-chondrial inner membrane contains some intrinsic proteincomplexes whose subunits broadly satisfy the criterion ofhydrophobicity However the core subunits that are mito-chondrially encoded are also explained by the CORRhypothesis This would make hydrophobicity a require-ment for their function in vectorial electron and protontransfer and not a reason in itself for retention of theirgenes in the cytoplasm
The hydrophobicity hypothesis has some very clearcounter-examples in chloroplasts and cannot thereforestand if one wishes for a single principle that embracesboth types of bioenergetic organelle One clear counter-example is the large (L) subunit of the enzyme RUBISCOan abundant but entirely membrane-extrinsic protein ofca 55 kDa molecular mass The RUBISCO large subunitis generally chloroplast-encoded with the exception of thenuclearly encoded bacterial-type L subunit of L2 RUBI-SCO in peridinin-containing dinoflagellates (Morse et al1995) In most photosynthetic eukaryotes RUBISCO hasa chloroplast encoded large subunit and a nuclearly enco-ded small (S) subunit (Ellis 1984 Gatenby amp Ellis 1990)giving an L8S8 holoenzyme and this RUBISCO is widelytaken as a paradigm for the need for coordination ofnuclear and chloroplast gene expression (Rodermel 2001)
By complete contrast to the RUBISCO large subunitwhich is a chloroplast-encoded and water-soluble proteina second major violation of the hydrophobicity hypothesisis the existence of the universally nuclearly encoded butalways hydrophobic subunits of the chloroplast LHC IIand LHC I (Chitnis amp Thornber 1988) These apopro-
Genomes in mitochondria and chloroplasts J F Allen 29
teins of ca 27ndash30 kDa are quite water-insoluble and areusually fully extracted only in chloroformndashmethanol or inthe presence of detergents The Lhc gene family is diverse(Green amp Kuhlbrandt 1995) but most gene productsor Lhcp (light-harvesting chlorophyll protein) polypeptideshave three membrane-spanning hydrophobic a-helices(Kuhlbrandt 1994) It is interesting that three-helix Lhcproteins seem to be absent from extant cyanobacteria andchloroxybacteria (Green amp Kuhlbrandt 1995)
(ii) The five-helix rulePerhaps with the three-helix model of LHC II in mind
a variation of the hydrophobicity hypothesis is the empiri-cal lsquofive-helix rulersquo (Popot amp de Vitry 1990 Claros et al1995 Wollman et al 1999) which states that no intrinsicmembrane protein with five or more membrane-spanninghelices can be encoded in the nucleus Five is a good guessfor a threshold number of helices demanding organellar-encoding in the context of photosynthetic reaction centres(Heathcote et al 2002) where the earliest known struc-ture that of type II centres (Deisenhofer et al 1985) asin photosystem II (Zouni et al 2001) shows exactly fivehelices with type I centres containing 11 (Jordan et al2001) There are however many nuclearly encoded sub-units in both mitochondria and chloroplasts that havefewer than five membrane helices so the five-helix rulecan only describe a sufficient condition and not a neces-sary condition for nuclear encoding
It should also be noted that ADPndashATP carriers (AACs)are encoded in the nucleus AACs of the mitochondrialinner membrane have six transmembrane helices (van derGiezen et al 2002) while the those of the chloroplast innerenvelope (Winkler amp Neuhaus 1999) have 12 (Tjaden etal 1998)
With the counter-examples of nuclearly encoded AACswith six or 12 membrane-spanning helices many intrinsicmembrane proteins with fewer than five helices somenuclearly encoded and some not and with the RUBISCOlarge subunit as a major product of chloroplast geneexpression with no membrane domain at all the five-helixrule begins to seem rather like a case of special pleadingEven if there are indeed special mitigating circumstancesfor AACs (this argument is outlined later) the five-helixrule as with the hydrophobicity hypothesis of which it isa special case will disappear if it can be shown that pos-session of five or more membrane helices is required forsome other function such as that of CORR that deter-mines the organellar location of the gene concernedHydrophobicity is in some cases a feature of proteins thatare encoded in organelles but it is not in itself the issue
For chloroplasts the hydrophobicity hypothesis predictsretention of cyanobacterial genes in functional categoriesassociated with the chloroplast envelope and with trans-port just as much as with photosynthesis Table 1 showsthat this is not the case
A mechanism for the unimportability of hydrophobicproteins is rarely stated explicitly and it is rather left tothe imagination whether hydrophobic proteins might asit were get stuck halfway through the chloroplast envelopeor mitochondrial outer membrane whether transport can-not get started on a precursor protein that has alreadyspontaneously inserted into the membrane or whetherhydrophobic proteins will be unable to make the journey
Phil Trans R Soc Lond B (2003)
through the cytosol without precipitating there In thecytosol co-translational transport or membrane insertionwould in principle address the latter point but it seemsto be generally true that chloroplast and mitochondrialproteins are post-translationally inserted into the organel-lar outer membrane So the obvious ancillary questionabout unimportability is lsquowhy then cannot hydrophobicproteins be co-translationally imported into organellesrsquoThere is evidence in yeast for co-translational transportinto mitochondria of proteins thought to be derived fromthe original endosymbiont (Marc et al 2002) If there isany truth in hydrophobicity as a barrier to protein importthe CORR principle of unselective import (sect 3c) shouldbe easily refuted for example by a demonstration that noknown targeting presequence is capable of importing poly-phenylalanine encoded by polyU for in vitro import stud-ies
An additional proposal intended to account for theunimportability of some hydrophobic proteins and forimportability of others is suggested by A de Grey(personal communication) who argues that bacteriallyderived protein precursors imported into mitochondriacross the mitochondrial inner membrane completelybefore insertion into that membrane from the mitochon-drial matrix (the N-phase) This is held to be an obstacleto successful import of membrane-intrinsic proteins thatmight en route discharge the proton motive force By con-trast eukaryotically derived proteins can be inserted intothe mitochondrial inner membrane from the inter-mem-brane space (the P-phase) and then there is for somereason no similar uncoupling effect Thus adenylatetransporters or AACs can be nuclearly encoded whereascytochrome oxidase subunits for example cannot It isnot obvious why bacterially derived subunits such asthose of cytochrome oxidase did not acquire eukaryoticinsertion mechanisms Perhaps this acquisition is forbid-den by the need to conserve pre-existing routes ofassembly into multi-subunit complexes or perhaps theneed for insertion from the inside reflects a need to incor-porate a cofactor for example haem or ironndashsulphur clus-ters along the way
(iii) Unimportability of three dimensional structures andcofactors
Accumulating evidence on protein transport mech-anisms seems to make a specific obstacle to successfultransport of certain sorts of protein less and less plausibleespecially given the observation that precursors with N-terminal targeting sequences not mature functional poly-peptides themselves traverse membranes during biosynth-esis
Protein folding inside the living cell is invariably assistedby molecular chaperones (Gatenby amp Ellis 1990 Azem etal 1997 Komiya et al 1997 1990 Jackson-Constan et al2001) proteins that impede the formation of lsquoimproperrsquostructural interactions that will otherwise produce aggre-gation or other non-functional interactions Proteinimport mechanisms appear mostly to rely on specific mol-ecular chaperones for guided unfolding before membraneinsertion and refolding after translocation of the polypep-tide into its destined compartment (Gatenby amp Ellis 1990Azem et al 1997 Komiya et al 1997 Jackson-Constanet al 2001) Because polypeptides are mostly lsquothreadedrsquo
30 J F Allen Genomes in mitochondria and chloroplasts
through membranes no special constraint on transportshould be expected to arise from 3D structure or surfaceproperties of a protein
An exception to this unfolding rule may be providedby holoproteins containing cofactors that are transportedalong with the apoprotein presumably with a largely intact3D structure The lsquoTATrsquo (twin-arginine translocase) sys-tem (Bogsch et al 1998 Dalbey amp Robinson 1999 Berkset al 2000ab Robinson 2000 Robinson amp Bolhuis 2001Sargent et al 2002) works broadly along these linesthough its mechanism is unresolved TAT is relevant herebecause it adds transport of essentially complete holopro-teins to the cellrsquos repertoire of transport mechanisms andeven proteins that may not be allowed to unfold perhapsbecause of the toxicity of the free cofactor are not therebymade candidates for an unimportability rule One variantof the unimportability hypothesis is that transport canwork only by unfolding and dissociated cofactors may betoxic to the cell The bacterially derived TAT system(Bogsch et al 1998 Dalbey amp Robinson 1999 Berks etal 2000ab Sargent et al 2002) overcomes this argumentas would examples of cofactor-free polypeptides such as(C)Fo subunit c which are nevertheless generally encodedin organelles
The presence of a vesicular transport system in chloro-plasts (Westphal et al 2001) counts against the unim-portability hypothesis Vesicular transport between thechloroplast envelope and the thylakoid membrane shouldin principle allow any protein to be ferried across thechloroplast stroma Vesicles might allow transport inparticular of membrane proteins that would discharge theproton motive force if allowed to connect transiently theN and P aqueous phases on each side of a coupling mem-brane
Another observation that seems to count against unim-portability is the increased number of membrane barriersin plastids derived from secondary tertiary or higher-orderendosymbioses These plastids seem to have retained ifanything fewer genes rather than more as unimportabilitypredicts The nucleomorph the residual nucleus of theprimary host retains many genes for molecular chap-erones (Archibald et al 2001 Cavalier-Smith 2002) inagreement with the conclusion that protein import pro-ceeds by conserved and specialized mechanisms If eachnew import added to gain a higher-order endosymbiosisrequires additional targeting presequences then perhapsthere is an advantage in components of an import machin-ery not being imported themselves
It seems that unimportability is less easy an assumptionto make today than in previous years and the onus maynow be on its proponents to cite specific examples of pro-teins that can never be imported into chloroplasts or mito-chondria
(f ) Some genes cannot be moved(i) Differences in the mitochondrial genetic code
The genetic code is the specific relation between ribon-ucleotide triplets of codons and tRNAs that recognizethem and which thus determine which individual aminoacids if any will be incorporated into the growing poly-peptide chain Mitochondria show small departures fromthe otherwise universal genetic code (Attardi amp Schatz1988 Alberts et al 1994) UGA is a lsquostoprsquo codon in the
Phil Trans R Soc Lond B (2003)
universal code and in plant mitochondria but is the codonfor tryptophan in mitochondria of animals and fungiCUA means leucine everywhere except fungal mitochon-dria where it means threonine and AGA and AGU codefor arginine in the lsquouniversalrsquo code but serine in insectmitochondria and lsquostoprsquo in mammalian ones The signifi-cance of this heterogeneity in coding is most unlikely torest in the early origin of mitochondria as the lsquouniversalrsquocode is employed by bacteria and protists Furthermorethe diversity of departures from the lsquouniversalrsquo codeamong mitochondria of different eukaryotic lineages indi-cates they had independent origins A general rule formitochondrial genetic codes is that reliance on the thirdlsquowobblersquo position is decreased which allows all 20 proteinamino acids to be specified by fewer tRNAs (22 in animalmitochondria instead of 31 in the cytosol) This in turnmeans that mtDNA can specify more proteins for a givenlength of DNA or conversely that the mitochondrialgenome can be condensed or lsquostreamlinedrsquo Although thisarrangement does not explain the retention of mitochon-drial genomes and may be a secondary event in mitochon-drial evolution it almost certainly places an extra barrierin place of gene relocation As discussed previously how-ever there seems to be no single barrier that can explainthe retention of a relatively constant subset of mitochon-drial genes
There has clearly been a contraction of mitochondrialDNA especially in animals Animal mitochondrial DNAhas few if any non-coding sequences and animal mito-chondrial genetic codes require fewer tRNAs than the sin-gle code operating in the cytosol (Gray 1999 Gray et al1999 Selosse et al 2001) These may be devices todecrease the total frequency of mutations per mitochon-drial genome assuming a constant frequency per unitlength of mitochondrial DNA It has been proposed thata genersquos removal to the nucleus decreases its susceptibilityto free radical mutagenesis (Raven et al 1994ab Allen ampRaven 1996) If this is the case then the non-standardcode may be a further indication of an overriding need tomaintain certain genes within the mitochondrion What-ever the reason for the retention of genes in organelles ithad better be a good one bioenergetic organelles areclearly the wrong place to keep genetic information andgo to extraordinary lengths to offset the consequences ofit being there
(ii) Molecular refugeesA phenomenon related to differences in the genetic code
is seen in chloroplasts This is a bias in codon usage Thelsquomolecular refugeesrsquo hypothesis (Howe et al 2000) sug-gests that nuclear copies of what were once chloroplastgenes escape constraints on codon usage and thus thatwhat distinguishes chloroplast-encoded proteins is thattheir amino-acid composition is unusual enough for thebias in codon usage not to apply It is difficult to see whatphysico-chemical properties might be uniquely shared bythe amino acids of chloroplast-encoded proteins and inaddition there seem to be no lsquoforbiddenrsquo amino-acid resi-due in chloroplasts As with the departure from the univer-sal genetic code in mitochondria is seems likely that abias in codon usage reflects some secondary property ofchloroplast genomes and is not the primary reason fortheir retention In any case any restriction on gene move-
Genomes in mitochondria and chloroplasts J F Allen 31
ment will act in the wrong direction to explain why chloro-plasts have retained genomes at all provided there is noresulting obstacle to a chloroplast protein being success-fully encoded as a precursor in the nucleus
(iii) Genes are copied and replicated not relocated in onestep
If complete relocation is envisaged as proceeding by thesimultaneous appearance of a gene in one compartmentand its deletion from another then it is possible to imaginethat whatever the vector is there may be selectivity ingene transfer (Henze amp Martin 2001) For example ifviral incorporation replication and recombination in bothorganelle and nucleus were the mechanism of relocationthen successful gene transfer might depend on the abilityof the virus to complete the process in one step This inturn might depend on some property of the gene or ofthe protein it encodes if expression occurs at any stage inthe process of relocation An assumption of the CORRhypothesis favoured here is that genes are not movedphysically but merely copied after which either the copyin the original location or the copy in the new locationmay be lost as a result of selection A useful analogy maybe the action of copying files between computer storagevolumes In this analogy the copy is identical to the orig-inal and either may subsequently be deleted without lossof information
Because any gene that is not usefully expressed willundergo no selection to maintain a functional allele theprevailing location will be the one in which the functionalcopy of the gene resides The problem of which gene sur-vives is then subsumed into the problem of which is theselectively advantageous location for it It seems reason-able to assume that there wasmdashand ismdashan interim periodin which homologous genes in different compartments areessentially in competition to see which functions best Asoutlined above there is no obvious obstacle for copyingDNA sequences within the cell The physico-chemicalproperties of DNA are fairly uniform and not directlydependent on nucleotide sequence though organelles likeintra-cellular parasites seem to prefer A and T to G andC The answer to the question lsquowhich gene location func-tions bestrsquo may have nothing to do with the gene itselfnor directly with the structure and function of its geneproduct Instead the answer may have everything to dowith the required regulatory interaction between the geneproduct and expression of the gene that produces it
(g) CORRThis hypothesis has been propounded earlier (sect 3) and
will be summarized as follows Vectorial electron and pro-ton transfer exerts regulatory control over expression ofgenes encoding proteins directly involved in or affectingredox poise This regulatory coupling is indispensable foradaptation to environmental changes in chemical activitiesof oxygen light and CO2 and requires co-location of thesegenes with their gene products Organelles lsquomake theirown decisionsrsquo on the basis of environmental changesaffecting redox state The hypothesis is abbreviatedCORR for co-location for redox regulation or more com-pletely lsquoco-location (of gene and gene product) for(evolutionary) continuity of redox regulation of geneexpressionrsquo
Phil Trans R Soc Lond B (2003)
5 SUMMARY OF THE CURRENT STATUS OF THEHYPOTHESIS OF CORR
Q Why do mitochondria and chloroplasts have theirown genetic systems
A CORR co-location of genes and gene products per-mits direct and autonomous redox regulation of geneexpression
(a) Objections to CORR(i) There is no regulation of gene expression in chloroplasts
and mitochondriaThis is still commonly stated but incorrect See the prin-
ciple in sect 3d
(ii) There is no regulation of gene expression in chloroplastsand mitochondria at the level of transcription
For chloroplasts there is now clear evidence for a func-tionally intelligible redox control of transcription(Pfannschmidt et al 1999a) This invalidates the previouswidely held assumption that all control of chloroplast geneexpression is post-transcriptional (Gruissem amp Tonkyn1993 GoldschmidtClermont 1998) The evidence formitochondria is less clear but redox control of transcrip-tion certainly cannot be ruled out and there are indi-cations that it may occur (Pearson et al 1993Konstantinov et al 1995 Wilson et al 1996b) It mustalso be restated that transcriptional control is not neces-sary for CORR but serves as a compelling example of aconserved typically bacterial mode of response that oper-ates still in eukaryotic cytoplasmic organelles thus illus-trating the principle of continuity of redox control (sect 3d)Post-transcriptional redox regulation (Danon amp Mayfield1994 Kim amp Mayfield 1997 Brown et al 2001 Treb-itsh amp Danon 2001) is consistent with CORR and thereis nothing to forbid it being a primary mechanism thatsatisfies most or indeed all of the formal requirements ofthe hypothesis in particular cases
(iii) There are no redox signalling components encoded inchloroplast or mitochondrial DNA
This is what CORR predicts See the principle in sect 3j
(iv) There is redox control of nuclear gene expressionThis is correct and many examples are now known
especially for chloroplasts (Escoubas et al 1995 Kovacs etal 2000 Pfannschmidt et al 2001ab Surpin et al 2002)Redox signals from photosynthetic or respiratory electrontransport chains are not at all prohibited by CORR Theessence of CORR is that there is a subset of organellarproteins for which there is an overriding requirement fora redox regulatory control that is initiated within theorganelle and whose effects are exerted by the simplestand most direct route This redox signalling is envisagedto take place without a requirement for prior consultationand coordination with what may be regarded as a nucleo-cytoplasmic bureaucracy
(v) Nuclear genes can be expressed as rapidly as chloroplastand mitochondrial genes
Zerges (2002) for example cites the equal times takenfor complete expression of nuclear and chloroplast genesin Chlamydomonas as evidence against the CORR hypoth-
32 J F Allen Genomes in mitochondria and chloroplasts
esis However the question is not one of the time takento express a gene but the time taken to alter the rate ofits expression in response to a (redox) signal originatingwithin the organelle An equally important additional fac-tor may be the autonomy of the signalling pathway thatis its independence from other inputs as described in theprevious paragraph One might say that mitochondria andchloroplasts require a free hand
(vi) There is no evidence to support this hypothesisThe hypothesis is rich in predictions (sectsect 3 and 6) and
those that have been tested stringently are in agreementwith experimental results None of the predictions of thehypothesis has been found to be incorrect and none of theunderlying principles (sect 3) has been disproved Furtherpredictions will be the subject of future experiments
(b) Evidence for CORR(i) Chloroplasts have selectively retained cyanobacterial genes
for primary events in photosynthesisSee table 1
(ii) There is regulation of gene expression in chloroplastsThere is transcriptional regulation of gene expression
in chloroplasts (sect 3d) The case for mitochondria is notyet proven
(iii) There is redox regulation of gene expression inchloroplasts
There is transcriptional redox regulation of geneexpression in chloroplasts (sect 3d) The case for mitochon-dria is not yet proven though see (Allen et al 1995 Kon-stantinov et al 1995 Galvis et al 1998)
(iv) There are no redox signalling components encoded inchloroplast or mitochondrial DNA
Correct but with possible exceptions discussed in sect 3j
(v) There are lsquobacterialrsquo redox signalling components ineukaryotes
See sect 3j
(vi) Further predictions DNA-array results and phenotypesof allotopic mutants
A systematic test of CORR might be to make an oxid-ized-minus-reduced gene expression profile of the lsquotran-scriptomersquo of a cyanobacterium to see if the set of geneswhose transcription is under most rapid and direct redoxcontrol corresponds to the genes of the chloroplast gen-ome Laboratories with this experimental capability do notseem to have addressed this possibility (Hihara et al 2002Ikeuchi 2002) Likewise if a putative modern representa-tive of the proto-mitochondrion can be found and agreedeg Rhodospirillum rubrum or Paracoccus denitrificans thena similar experiment should reveal a subset of genes corre-sponding to mitochondrial genomes DNA-microarraysalso present the possibility of testing the relative rate ofredox response and dependency on other signallinginputs of transcription of eukaryotic genes To dateorganellar genes have not been included in such investi-gations (Ferea et al 1999) A further experimental possi-bility is to examine the redox regulatory phenotypes ofexisting allotopic mutants as outlined in sect 3e
Phil Trans R Soc Lond B (2003)
6 CONCLUSION AND FUTURE PROSPECTS
The CORR hypothesis states that the function of mito-chondrial and chloroplast genomes is to provide co-location (of gene and gene product) for (evolutionary)continuity of redox regulation of gene expression Thishypothesis is neither self-contradictory nor devoid of test-able predictions It is also consistent with available evi-dence and in principle makes sense of the otherwisepuzzling distribution of genes between compartments ineukaryotic cells The explanation that CORR offersapplies equally to mitochondria and chloroplasts andidentifies selective forces that still operate to determine thelocation of genes No selective barriers need be assumedto gene transfer protein import nor membrane insertionNor do we need to invoke a lsquofrozen accidentrsquo that some-thing happened to stop gene transfer for some reason atsome time in the remote and inaccessible past CORR alsoexplains the absence of genomes from organelles that havelost their primary bioenergetic functions CORR may actas a stimulus to a search for residual bioenergetic functionin those organelles that have lost aerobic respiration andphotosynthesis but where residual genomes remain
CORR also predicts regulatory properties of knowncomponents of chloroplasts and mitochondria that in turnindicate important but as yet uncharacterized flexibility inenergy transduction These components include the RUB-ISCO large subunit and the CFo and Fo subunits of coup-ling ATPase in both chloroplasts and mitochondriaRedox regulation of the relative stoichiometry of compo-nents as seen in regulation of chloroplast photosystemstoichiometry (Pfannschmidt et al 1999a) may extend to(C)Fo-(C)F1-ATPase indicating that the function ofretention of genes for one or more of these componentsis flexibility in the stoichiometries of H1 to e2 and H1 toATP in chemiosmotic coupling (Schemidt et al 1998Stock et al 2000 Allen 2002)
So far explicit tests of CORR have largely been con-fined to chloroplasts and to some extent mitochondria ofhigher plants but this merely reflects the expertise andinterests of the investigators Further attempted disproofof CORR is welcomed and encouraged Novel experi-mental tests can be devised as well as attempted refu-tation through analysis of existing data It is hoped thatexplicit attention will be given to the CORR hypothesis inlaboratories already suitably equipped and that resourcescan be allocated for this purpose in laboratories wherethere is the interest but not currently the technology orexpertise
The heuristic value of the hypothesis already seen inthe role of redox-regulated transcription of photosyntheticreaction centre genes in chloroplasts may be its most use-ful property The central proposal is that mitochondriaand chloroplasts contain genes for proteins whose functionin electron transfer demands rapid direct and uncon-ditional redox regulatory control of their biosynthesis Thefull implications of this hypothesis are very broad and onlyjust beginning to be explored
I thank Carol Allen Aubrey de Grey William Martin John ARaven and Francis-Andre Wollman for discussion and corre-spondence during the preparation of this paper and WilliamMartin for suggestions including table 1 Research grants from
Genomes in mitochondria and chloroplasts J F Allen 33
The Swedish Natural Sciences Research Council (NFR) andthe Crafoord Foundation are gratefully acknowledged
REFERENCES
Adams K L Daley D O Qiu Y L Whelan J amp PalmerJ D 2000 Repeated recent and diverse transfers of a mito-chondrial gene to the nucleus in flowering plants Nature408 354ndash357
Alberts B Bray D Lewis J Raff M Roberts K amp Wat-son J D 1994 The molecular biology of the cell New Yorkand London Garland Publishing
Allen C A Hakansson G amp Allen J F 1995 Redox con-ditions specify the proteins synthesized by isolated chloro-plasts and mitochondria Redox Rep 1 119ndash123
Allen J F 1992 Protein phosphorylation in regulation ofphotosynthesis Biochim Biophys Acta 1098 275ndash335
Allen J F 1993a Control of gene expression by redox poten-tial and the requirement for chloroplast and mitochondrialgenomes J Theor Biol 165 609ndash631
Allen J F 1993b Redox control of gene expression and thefunction of chloroplast genomes a hypothesis PhotosynthRes 36 95ndash102
Allen J F 1993c Redox control of transcription sensorsresponse regulators activators and repressors FEBS Lett332 203ndash207
Allen J F 1995 Thylakoid proteinndashphosphorylation state-1ndashstate-2 transitions and photosystem stoichiometry adjust-ment redox control at multiple levels of gene expressionPhysiol Plant 93 196ndash205
Allen J F 1996 Separate sexes and the mitochondrial theoryof ageing J Theor Biol 180 135ndash140
Allen J F 2002 Photosynthesis of ATP-electrons protonpumps rotors and poise Cell 110 273ndash276
Allen J F amp Allen C A 1999 A mitochondrial model forpremature ageing of somatically cloned mammals IUBMBLife 48 369ndash372
Allen J F amp Nilsson A 1997 Redox signalling and the struc-tural basis of regulation of photosynthesis by protein phos-phorylation Physiol Plant 100 863ndash868
Allen J F amp Pfannschmidt T 2000 Balancing the two photo-systems photosynthetic electron transfer governs transcrip-tion of reaction centre genes in chloroplasts Phil TransR Soc Lond B 355 1351ndash1357 (DOI 101098rstb20000697)
Allen J F amp Race H L 2002 Will the real LHC II kinaseplease step forward Sciencersquos STKE httpwwwstkeorgcgicontentfullsigtrans2002155pe43
Allen J F amp Raven J A 1996 Free-radical-induced mutationvs redox regulation costs and benefits of genes in organellesJ Mol Evol 42 482ndash492
Archibald J M Cavalier-Smith T Maier U amp Douglas S2001 Molecular chaperones encoded by a reduced nucleusthe cryptomonad nucleomorph J Mol Evol 52 490ndash501
Ashby M K amp Mullineaux C W 1999a Cyanobacterialycf27 gene products regulate energy transfer from phycobili-somes to photosystems I and II FEMS Microbiol Lett 181253ndash260
Ashby M K amp Mullineaux C W 1999b The role of ApcDand ApcF in energy transfer from phycobilisomes to PSI andPSII in a cyanobacterium Photosynth Res 61 169ndash179
Ashby M K Houmard J amp Mullineaux C W 2002 Theycf27 genes from cyanobacteria and eukaryotic algae distri-bution and implications for chloroplast evolution FEMSMicrobiol Lett 214 25ndash30
Attardi G amp Schatz G 1988 Biogenesis of mitochondria ARev Cell Biol 4 289ndash333
Azem A Oppliger W Lustig A Jeno P Feifel B SchatzG amp Horst M 1997 The mitochondrial hsp70 chaperone
Phil Trans R Soc Lond B (2003)
system effect of adenine nucleotides peptide substrate andmGrpE on the oligomeric state of mhsp70 J Biol Chem272 20 901ndash20 906
Barbrook A C Symington H Nisbet R E R LarkumA amp Howe C J 2001 Organisation and expression of theplastid genome of the dinoflagellate Amphidinium opercula-tum Mol Genet Genomics 266 632ndash638
Bauer C E Elsen S amp Bird T H 1999 Mechanisms forredox control of gene expression A Rev Microbiol 53495ndash523
Bauer C E Elsen S Swem L R Swem D L amp MasudaS 2003 Redox and light regulation of gene expression inphotosynthetic prokaryotes Phil Trans R Soc Lond B 358147ndash154 (DOI 101098rstb20021189)
Berks B C Sargent F De Leeuw E Hinsley A P Stan-ley N R Jack R L Buchanan G amp Palmer T 2000aA novel protein transport system involved in the biogenesisof bacterial electron transfer chains Biochim Biophys ActaBioenerg 1459 325ndash330
Berks B C Sargent F amp Palmer T 2000b The tat proteinexport pathway Mol Microbiol 35 260ndash274
Blackstone N W 1999 Redox control in development andevolution evidence from colonial hydroids J Exp Biol 2023541ndash3553
Blackstone N W 2000 Redox control and the evolution ofmulticellularity Bioessays 22 947ndash953
Blackstone N W 2001 Redox state reactive oxygen speciesand adaptive growth in colonial hydroids J Exp Biol 2041845ndash1853
Bogorad L 1975 Evolution of organelles and eukaryotic gen-omes Science 188 891ndash898
Bogsch E G Sargent F Stanley N R Berks B C Robin-son C amp Palmer T 1998 An essential component of a novelbacterial protein export system with homologues in plastidsand mitochondria J Biol Chem 273 18 003ndash18 006
Brown E C Somanchi A amp Mayfield S P 2001 Interor-ganellar crosstalk new perspectives on signaling from thechloroplast to the nucleus Genome Biol 2 10211ndash10214
Cavalier-Smith T 2002 Chloroplast evolution secondarysymbiogenesis and multiple losses Curr Biol 12 R62ndashR64
Chitnis P R amp Thornber J P 1988 The major light-har-vesting complex of photosystem 2 Aspects of its molecularand cell biology Photosynth Res 16 41ndash63
Chow W S Melis A amp Anderson J M 1990 Adjustmentsof photosystem stoichiometry in chloroplasts improve thequantum efficiency of photosynthesis Proc Natl Acad SciUSA 87 7502ndash7506
Claros M G Perea J Shu Y Samatey F A PopotJ L amp Jacq C 1995 Limitations to in vivo import of hydro-phobic proteins into yeast mitochondria The case of a cyto-plasmically synthesized apocytochrome b Eur J Biochem228 762ndash771
Dalbey R E amp Robinson C 1999 Protein translocation intoand across the bacterial plasma membrane and the plant thy-lakoid membrane Trends Biochem Sci 24 17ndash22
Daley D O Adams K L Clifton R Qualmann S MillarA H Palmer J D Pratje E amp Whelan J 2002 Genetransfer from mitochondrion to nucleus novel mechanismsfor gene activation from Cox2 Plant J 30 11ndash21
Danon A amp Mayfield S P 1994 Light-regulated translationof chloroplast messenger RNAs through redox potentialScience 266 1717ndash1719
de Grey A 2000 Mitochondrial gene therapy an arena for thebiomedical use of inteins Trends Biotechnol 18 394ndash399
Deisenhofer J Epp O Miki K Huber R amp Michel H1985 Structure of the protein subunits in the photosyntheticreaction center of Rhodopseudomonas viridis at 3Ac resolutionNature 318 618ndash624
34 J F Allen Genomes in mitochondria and chloroplasts
Delwiche C F 1999 Tracing the thread of plastid diversitythrough the tapestry of life Am Nat 154 S164ndashS177
Doolittle W F 1999 Phylogenetic classification and the uni-versal tree Science 284 2124ndash2129
Douglas S E 1998 Plastid evolution origins diversitytrends Curr Opin Genet Dev 8 655ndash661
Duby G amp Boutry M 2002 Mitochondrial protein importmachinery and targeting information Plant Sci 162 477ndash490
Eberhard S Drapier D amp Wollman F A 2002 Searchinglimiting steps in the expression of chloroplast encoded pro-teins relations between gene copy number transcriptiontranscript abundance and translation rate in the chloroplastof Chlamydomonas reinhardtii Plant J 31 149ndash160
El Bissati K amp Kirilovsky D 2001 Regulation of psbA andpsaE expression by light quality in Synechocystis species PCC6803 A redox control mechanism Plant Physiol 1251988ndash2000
Ellis R J 1984 The nuclear domination of chloroplast devel-opment Science Progress 69 129ndash142
Embley T M amp Martin W 1998 Molecular evolution ahydrogen-producing mitochondrion Nature 396 517ndash519
Ems S C Morden C W Dixon C K Wolfe K HdePamphilis C W amp Palmer J D 1995 Transcriptionsplicing and editing of plastid RNAs in the non-photosyn-thetic plant Epifagus virginiana Plant Mol Biol 29 721ndash733
Eraso J M amp Kaplan S 2000 From redox flow to gene regu-lation role of the PrrC protein of Rhodobacter sphaeroides241 Biochemistry 39 2052ndash2062
Eraso J M amp Kaplan S 2002 Redox flow as an instrumentof gene regulation Meth Enzymol 348 216ndash229
Escoubas J M Lomas M Laroche J amp Falkowski P G1995 Light-intensity regulation of cab gene-transcription issignaled by the redox state of the plastoquinone pool ProcNatl Acad Sci USA 92 10 237ndash10 241
Ferea T L Botstein D Brown P O amp Rosenzweig R F1999 Systematic changes in gene expression patterns follow-ing adaptive evolution in yeast Proc Natl Acad Sci USA96 9721ndash9726
Forsberg J Rosenquist M Fraysse L amp Allen J F 2001Redox signalling in chloroplasts and mitochondria genomicand biochemical evidence for two-component regulatory sys-tems in bioenergetic organelles Biochem Soc Trans 29403ndash407
Fujita Y 1997 A study on the dynamic features of photosys-tem stoichiometry accomplishments and problems forfuture studies Photosynth Res 53 83ndash93
Gabriel K Buchanan S K amp Lithgow T 2001 The alphaand the beta protein translocation across mitochondrial andplastid outer membranes Trends Biochem Sci 26 36ndash40
Galvis M L E Allen J F amp Hakansson G 1998 Proteinsynthesis by isolated pea mitochondria is dependent on theactivity of respiratory complex II Curr Genet 33 320ndash329
Gatenby A A amp Ellis R J 1990 Chaperone function theassembly of ribulose bisphosphate carboxylase-oxygenase ARev Cell Biol 6 125ndash149
GoldschmidtClermont M 1998 Coordination of nuclear andchloroplast gene expression in plant cells Int Rev CytolSurv Cell Biol 177 115ndash180
Gray M W 1992 The endosymbiont hypothesis revisited IntRev Cytol 141 233ndash357
Gray M W 1999 Evolution of organellar genomes CurrOpin Genet Dev 9 678ndash687
Gray M W 2000 Evolutionary biology mitochondrial geneson the move Nature 408 302ndash305
Gray M W amp Doolittle W F 1982 Has the endosymbionthypothesis been proven Microbiol Rev 46 1ndash42
Gray M W Burger G amp Lang B F 1999 Mitochondrialevolution Science 283 1476ndash1481
Phil Trans R Soc Lond B (2003)
Grebe T W amp Stock J B 1999 The histidine protein kinasesuperfamily Adv Microb Physiol 41 139ndash227
Green B R amp Kuhlbrandt W 1995 Sequence conservationof light-harvesting and stress-response proteins in relation tothe 3-dimensional molecular structure of Lhcii PhotosynthRes 44 139ndash148
Gruissem W amp Tonkyn J C 1993 Control mechanisms ofplastid gene expression Crit Rev Plant Sci 12 19ndash55
Hackstein J H P (and 11 others) 2001 Hydrogenosomesconvergent adaptations of mitochondria to anaerobicenvironments Zool Anal Complex Syst 104 290ndash302
Harrison M A Keen J N Findlay J B C amp Allen J F1990 Modification of a Glnb-like gene-product by photosyn-thetic electron-transport in the cyanobacterium Synechoc-occus 6301 FEBS Lett 264 25ndash28
Harvey A J Kind K L amp Thompson J G 2002 Redoxregulation of early embryo development Reproduction 123479ndash486
Heathcote P Fyfe P K amp Jones M R 2002 Reactioncentres the structure and evolution of biological solarpower Trends Biochem Sci 27 79ndash87
Henze K amp Martin W 2001 How do mitochondrial genesget into the nucleus Trends Genet 17 383ndash387
Herrmann R G amp Westhoff P 2001 Thylakoid biogenesis anddynamics the result of a complex phylogenetic puzzle InRegulation of photosynthesis vol 11 (ed E-M Aro amp BAndersson) pp 1ndash28 Dordrecht The Netherlands Kluwer
Hihara Y Sonoike K Kanehisa M amp Ikeuchi M 2002Search for redox responsive genes in Synechocystis sp PCC6803 by using DNA microarrays Plant Cell Physiol 43S178
Howe C J Barbrook A C amp Lockhart P J 2000 Organellegenes do they jump or are they pushed Trends Genet 1665ndash66
Ikeuchi M 2002 Gene expression and responses to light stressin cyanobacteria Plant Cell Physiol 43 S21
Iuchi S amp Lin E C C 1992 Mutational analysis of signaltransduction by Arcb a membrane sensor protein respon-sible for anaerobic repression of operons involved in the cen-tral aerobic pathways in Escherichia coli J Bacteriol 1743972ndash3980
Jackson-Constan D Akita M amp Keegstra K 2001 Molecu-lar chaperones involved in chloroplast protein importBiochim Biophys Acta Mol Cell Res 1541 102ndash113
Jarvis P amp Soll M 2001 Toc Tic and chloroplast proteinimport Biochim Biophys Acta Mol Cell Res 1541 64ndash79
Jordan P Fromme P Witt H T Klukas O SaengerW amp Krauss N 2001 Three-dimensional structure ofcyanobacterial photosystem I at 25 acngstrom resolutionNature 411 909ndash917
Kaneko T (and 23 others) 1996a Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regionsDNA Res 3 109ndash136
Kaneko T (and 23 others) 1996b Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regions[Supplement] DNA Res 3 185ndash209
Kanevski I amp Maliga P 1994 Relocation of the plastid rbcLgene to the nucleus yields functional ribulose-15-bisphosph-ate carboxylase in tobacco chloroplasts Proc Natl Acad SciUSA 91 1969ndash1973
Kim J amp Mayfield S P 1997 Protein disulfide isomerase as aregulator of chloroplast translational activation Science 2781954ndash1957
Genomes in mitochondria and chloroplasts J F Allen 35
Kirk J T O amp Tilney-Bassett R A E 1978 The plastidsTheir chemistry structure growth and inheritance Amster-dam Elsevier
Koehler C M Merchant S amp Schatz G 1999 How mem-brane proteins travel across the mitochondrial intermem-brane space Trends Biochem Sci 24 428ndash432
Komiya T Rospert S Schatz G amp Mihara K 1997 Bind-ing of mitochondrial precursor proteins to the cytoplasmicdomains of the import receptors Tom70 and Tom20 isdetermined by cytoplasmic chaperones FASEB J 11 1196
Konstantinov Y M Lutsenko G N amp Podsosonny V A1995 Genetic functions of isolated maize mitochondriaunder model changes of redox conditions Biochem MolBiol Int 36 319ndash326
Korab-Laskowska M Rioux P Brossard N LittlejohnT G Gray M W Lang B F amp Burger G 1998 Theorganelle genome database project (GOBASE) Nucleic AcidsRes 26 138ndash144
Kovacs L Wiessner W Kis M Nagy F Mende D ampDemeter S 2000 Short- and long-term redox regulation ofphotosynthetic light energy distribution and photosystemstoichiometry by acetate metabolism in the green alga Chla-mydobotrys stellata Photosynth Res 65 231ndash247
Kuhlbrandt W 1994 Structure and function of the plant light-harvesting complex LHC-II Curr Opin Struct Biol 4519ndash528
Lang B F Gray M W amp Burger G 1999 Mitochondrialgenome evolution and the origin of eukaryotes A RevGenet 33 351ndash397
Lemire B D Fankhauser C Baker A amp Schatz G 1989The mitochondrial targeting function of randomly generatedpeptide sequences correlates with predicted helical amphi-philicity J Biol Chem 264 20 206ndash20 215
Leuenberger D Bally N A Schatz G amp Koehler C M1999 Different import pathways through the mitochondrialintermembrane space for inner membrane proteins EMBOJ 18 4816ndash4822
Li H amp Sherman L A 2000 A redox-responsive regulator ofphotosynthesis gene expression in the cyanobacterium Syne-chocystis sp strain PCC 6803 J Bacteriol 182 4268ndash4277
Lin X Y (and 36 others) 1999 Sequence and analysis of chro-mosome 2 of the plant Arabidopsis thaliana Nature 402761ndash768
Lonsdale D M Hodge T P Howe C J amp Stern D B1983 Maize mitochondrial-DNA contains a sequence hom-ologous to the ribulose-15-bisphosphate carboxylase largesubunit gene of chloroplast DNA Cell 34 1007ndash1014
McFadden G I 1999 Endosymbiosis and evolution of theplant cell Curr Opin Plant Biol 2 513ndash519
McFadden G I 2001 Chloroplast origin and integrationPlant Physiol 125 50ndash53
Mahler H R amp Raff R A 1975 The evolutionary origin ofthe mitochondrion a nonsymbiotic model Int Rev Cytol43 1ndash124
Manfredi G Fu J Ojaimi J Sadlock J E Kwong J QGuy J amp Schon E A 2002 Rescue of a deficiency in ATPsynthesis by transfer of MTATP6 a mitochondrial DNA-encoded gene to the nucleus Nature Genet 30 394ndash399
Marc P Margeot A Devaux F Blugeon C Corral-Debrinski M amp Jacq C 2002 Genome-wide analysis ofmRNAs targeted to yeast mitochondria EMBO Rep 3159ndash164
Marienfeld J Unseld M amp Brennicke A 1999 The mito-chondrial genome of Arabidopsis is composed of both nativeand immigrant information Trends Plant Sci 4 495ndash502
Martin W 1999 A briefly argued case that mitochondria andplastids are descendants of endosymbionts but that thenuclear compartment is not Proc R Soc Lond B 2661387ndash1395 (DOI 101098rspb19990792)
Phil Trans R Soc Lond B (2003)
Martin W amp Herrmann R G 1998 Gene transfer fromorganelles to the nucleus how much what happens andwhy Plant Physiol 118 9ndash17
Martin W amp Muller M 1998 The hydrogen hypothesis forthe first eukaryote Nature 392 37ndash41
Martin W amp Russell M J 2003 On the origins of cells ahypothesis for the evolutionary transitions from abiotic geo-chemistry to chemoautotrophic prokaryotes and from pro-karyotes to nucleated cells Phil Trans R Soc Lond B 35859ndash85 (DOI 101098rstb20021183)
Martin W Stoebe B Goremykin V Hansmann S Hase-gawa M amp Kowallik K V 1998 Gene transfer to thenucleus and the evolution of chloroplasts Nature 393162ndash165
Martin W Hoffmeister M Rotte C amp Henze K 2001 Anoverview of endosymbiotic models for the origins of eukary-otes their ATP-producing organelles (mitochondria andhydrogenosomes) and their heterotrophic lifestyle BiolChem 382 1521ndash1539
Martin W Rujan T Richly E Hansen A Cornelsen SLins T Leister D Stoebe B Hasegawa M amp PennyD 2002 Evolutionary analysis of Arabidopsis cyanobacterialand chloroplast genomes reveals plastid phylogeny and thou-sands of cyanobacterial genes in the nucleus Proc NatlAcad Sci USA 99 12 246ndash12 251
Millen R S (and 12 others) 2001 Many parallel losses of infAfrom chloroplast DNA during angiosperm evolution withmultiple independent transfers to the nucleus Plant Cell 13645ndash658
Morse D Salois P Markovic P amp Hastings J W 1995 Anuclear-encoded form-II RUBISCO in dinoflagellatesScience 268 1622ndash1624
Murakami A Kim S J amp Fujita Y 1997 Changes in photo-system stoichiometry in response to environmental con-ditions for cell growth observed with the cyanophyteSynechocystis PCC 6714 Plant Cell Physiol 38392ndash397
Oh J I amp Kaplan S 2000 Redox signaling globalization ofgene expression EMBO J 19 4237ndash4247
Ozawa T 1995 Mitochondrial-DNA mutations associatedwith aging and degenerative diseases Exp Gerontol 30269ndash290
Palmer J D 1997 Organelle genomes going going goneScience 275 790ndash791
Pearson C K Wilson S B Schaffer R amp Ross A W 1993NAD turnover and utilization of metabolites for RNA syn-thesis in a reaction sensing the redox state of the cyto-chrome-b( 6 )f complex in isolated chloroplasts Eur JBiochem 218 397ndash404
Pfanner N amp Geissler A 2001 Versatility of the mitochon-drial protein import machinery Natl Rev Mol Cell Biol 2339ndash349
Pfanner N amp Wiedemann N 2002 Mitochondrial proteinimport two membranes three translocases Curr Opin CellBiol 14 400ndash411
Pfannschmidt T Nilsson A amp Allen J F 1999a Photosyn-thetic control of chloroplast gene expression Nature 397625ndash628
Pfannschmidt T Nilsson A Tullberg A Link G amp AllenJ F 1999b Direct transcriptional control of the chloroplastgenes psbA and psaAB adjusts photosynthesis to light energydistribution in plants IUBMB Life 48 271ndash276
Pfannschmidt T Allen J F amp Oelmuller R 2001a Prin-ciples of redox control in photosynthesis gene expressionPhysiol Plant 112 1ndash9
Pfannschmidt T Schutze K Brost M amp Oelmuller R2001b A novel mechanism of nuclear photosynthesis generegulation by redox signals from the chloroplast during pho-
36 J F Allen Genomes in mitochondria and chloroplasts
tosystem stoichiometry adjustment J Biol Chem 27636 125ndash36 130
Popot J L amp de Vitry C 1990 On the microassembly of inte-gral membrane proteins A Rev Biophys Biophys Chem 19369ndash403
Race H L Herrmann R G amp Martin W 1999 Why haveorganelles retained genomes Trends Genet 15 364ndash370
Raven J A Johnston A M Parsons R amp Kubler J 1994aThe influence of natural and experimental high O2 concen-trations on O2 evolving phototrophs Biol Rev CambridgePhil Soc 69 61ndash94
Raven J A Johnston A M Parsons R amp Kubler J 1994bThe occurrence and influence on photolithotrophs of highoxygen concentrations Proc R Soc Edinb B 102 193ndash201
Robinson C 2000 The twin-arginine translocation system anovel means of transporting folded proteins in chloroplastsand bacteria Biol Chem 381 89ndash93
Robinson C amp Bolhuis A 2001 Protein targeting by the twin-arginine translocation pathway Natl Rev Mol Cell Biol 2350ndash356
Rodermel S 2001 Pathways of plastid-to-nucleus signalingTrends Plant Sci 6 471ndash478
Roger A J 1999 Reconstructing early events in eukaryoticevolution Am Nat 154 S146ndashS163
Roger A J Svard S G Tovar J Clark C G SmithM W Gillin F D amp Sogin M L 1998 A mitochondrial-like chaperonin 60 gene in Giardia lamblia evidence thatdiplomonads once harbored an endosymbiont related to theprogenitor of mitochondria Proc Natl Acad Sci USA 95229ndash234
Roodyn D B amp Wilkie D 1968 The biogenesis of mitochondriaMethuenrsquos monographs on biological subjects London Methuen
Rujan T amp Martin W 2001 How many genes in Arabidopsiscome from cyanobacteria An estimate from 386 proteinphylogenies Trends Genet 17 113ndash120
Saccone C Gissi C Lanave C Larizza A Pesole G ampReyes A 2000 Evolution of the mitochondrial genetic sys-tem an overview Gene 261 153ndash159
Salzberg S L White O Peterson J amp Eisen J A 2001Microbial genes in the human genome lateral transfer orgene loss Science 292 1903ndash1906
Sargent F Berks B C amp Palmer T 2002 Assembly of mem-brane-bound respiratory complexes by the Tat protein-trans-port system Arch Microbiol 178 77ndash84
Schatz G 1998 Protein transport the doors to organellesNature 395 439ndash440
Schemidt R A Qu J Williams J R amp Brusilow W S1998 Effects of carbon source on expression of F0 genes andon the stoichiometry of the c subunit in the F1 F0 ATPaseof Escherichia coli J Bacteriol 180 3205ndash3208
Selosse M A Albert B R amp Godelle B 2001 Reducing thegenome size of organelles favours gene transfer to thenucleus Trends Ecol Evol 16 135ndash141
Stern D B amp Lonsdale D M 1982 Mitochondrial and chlo-roplast genomes of maize have a 12- kilobase DNA-sequencein common Nature 299 698ndash702
Stock D Gibbons C Arechaga I Leslie A G W ampWalker J E 2000 The rotary mechanism of ATP synthaseCurr Opin Struct Biol 10 672ndash679
Stoebe B amp Maier U G 2002 One two three naturersquos toolbox for building plastids Protoplasma 219 123ndash130
Stoebe B Martin W amp Kowallik K V 1998 Distributionand nomenclature of protein-coding genes in 12 sequencedchloroplast genomes Plant Mol Biol Rep 16 243ndash255
Stupar R M Lilly J W Town C D Cheng Z Kaul SBuell C R amp Jiang J M 2001 Complex mtDNA consti-tutes an approximate 620 kb insertion on Arabidopsis thali-ana chromosome 2 implication of potential sequencing
Phil Trans R Soc Lond B (2003)
errors caused by large-unit repeats Proc Natl Acad SciUSA 98 5099ndash5103
Surpin M Larkin R M amp Chory J 2002 Signal transduc-tion between the chloroplast and the nucleus Plant Cell 14S327ndashS338
Tjaden J Schwoppe C Mohlmann T Quick P W ampNeuhaus H E 1998 Expression of a plastidic ATPADPtransporter gene in Escherichia coli leads to a functional aden-ine nucleotide transport system in the bacterial cytoplasmicmembrane J Biol Chem 273 9630ndash9636
Tovar J Fischer A amp Clark C G 1999 The mitosome a novelorganelle related to mitochondria in the amitochondrial para-site Entamoeba histolytica Mol Microbiol 32 1013ndash1021
Trebitsh T amp Danon A 2001 Translation of chloroplastpsbA mRNA is regulated by signals initiated by both photos-ystems II and I Proc Natl Acad Sci USA 98 12 289ndash12 294
Tullberg A Alexciev K Pfannschmidt T amp Allen J F2000 Photosynthetic electron flow regulates transcription ofthe psaB gene in pea (Pisum sativum L) chloroplasts throughthe redox state of the plastoquinone pool Plant Cell Physiol41 1045ndash1054
Turmel M Lemieux C Burger G Lang B F Otis CPlante I amp Gray M W 1999 The complete mitochondrialDNA sequences of Nephroselmis olivacea and Pedinomonasminor two radically different evolutionary patterns withingreen algae Plant Cell 11 1717ndash1729
Unden G amp Bongaerts J 1997 Alternative respiratory path-ways of Escherichia coli energetics and transcriptional regu-lation in response to electron acceptors Biochim BiophysActa Bioenerg 1320 217ndash234
Unseld M Marienfeld J R Brandt P amp Brennicke A1997 The mitochondrial genome of Arabidopsis thaliana con-tains 57 genes in 366 924 nucleotides Nature Genet 1557ndash61
Van der Giezen M Slotboom D J Horner D S DyalP L Harding M Xue G P Embley T M amp KunjiE R S 2002 Conserved properties of hydrogenosomal andmitochondrial ADPATP carriers a common origin for bothorganelles EMBO J 21 572ndash579
Von Heijne G 1986 Why mitochondria need a genome FEBSLett 198 1ndash4
Westphal S Soll J amp Vothknecht U C 2001 A vesicle trans-port system inside chloroplasts FEBS Lett 506 257ndash261
Whitehouse H L K 1969 Towards an understanding of themechanism of heredity London Edward Arnold Ltd
Williams B A P Hirt R P Lucocq J M amp Embley T M2002 A mitochondrial remnant in the microsporidian Trachi-pleistophora hominis Nature 418 865ndash869
Wilson R J (and 10 others) 1996a Complete gene map of theplastid-like DNA of the malaria parasite Plasmodium falcipa-rum J Mol Biol 261 155ndash172
Wilson S B Davidson G S Thomson L M amp PearsonC K 1996b Redox control of RNA synthesis in potato mito-chondria Eur J Biochem 242 81ndash85
Winkler H H amp Neuhaus H E 1999 Non-mitochondrialATP transport Trends Biochem Sci 24 64ndash68
Wolfe K H Morden C W amp Palmer J D 1992 Small sin-gle-copy region of plastid DNA in the non-photosyntheticangiosperm Epifagus virginiana contains only two genes Dif-ferences among dicots monocots and bryophytes in geneorganization at a non-bioenergetic locus J Mol Biol 22395ndash104
Wollman F A Minai L amp Nechushtai R 1999 The biogen-esis and assembly of photosynthetic proteins in thylakoidmembranes Biochim Biophys Acta Bioenerg 1411 21ndash85
Yeliseev A A amp Kaplan S 2000 TspO of Rhodobactersphaeroides a structural and functional model for the mam-
Genomes in mitochondria and chloroplasts J F Allen 37
malian peripheral benzodiazepine receptor J Biol Chem275 5657ndash5667
Zerges W 2002 Does complexity constrain organelle evol-ution Trends Plant Sci 7 175ndash182
Zhang Z D Green B R amp Cavalier-Smith T 1999 Singlegene circles in dinoflagellate chloroplast genomes Nature400 155ndash159
Zhang Z D Cavalier-Smith T amp Green B R 2001 A fam-ily of selfish minicircular chromosomes with jumbled chloro-plast gene fragments from a dinoflagellate Mol Biol Evol18 1558ndash1565
Zouni A Witt H T Kern J Fromme P Krauss NSaenger W amp Orth P 2001 Crystal structure of photosys-tem II from Synechococcus elongatus at 38 acngstrom resol-ution Nature 409 739ndash743
DiscussionD S Horner (Dipartimento di Fisiologia e Biochimica
Generali University of Milan Milan Italy) Given thatthere are components of these membrane-bound energy-transduction pathways that are consistently nuclear enco-ded not the key ones you pointed out but others do youexpect perhaps a slower perhaps a more crude but yet aredox communication between the organelle andexpression of nuclear-encoded genes
J F Allen There is and there is no question about itI think that Bogoradrsquos lock-in hypothesis has somethingto say here and there are other people who say it is all todo with assembling multi-subunit complexes If you havecontrol over the initial construction of the core aroundwhich the whole complex is eventually built then redoxcontrol or some sort of control over synthesis of that corewill control construction of the whole thing eventuallyand I think that may well be largely correct I think it ispossible in principle for nuclear genes to make precursorsaccording to redox instructions but it is not really soimportant that those precursors are here and now presentin exactly the right quantities because the precursors canto a limited extent hang around and wait Actually thereis proteolytic degradation of unattached precursors and soon but I think the fact that these chloroplast and mito-chondrially DNA-encoded subunits are at the core ofthese energy-transducing complexes is the crucial issue
T Cavalier-Smith (Department of Zoology University ofOxford Oxford UK ) In the case of mitochondria the sub-set of genes that remains is much more variable than youimplied It runs from three proteins in the minimal casesup to I think over 90 That is a big difference and manyof the genes in different lineages of protists that remainhave nothing to do with redox control they are things likeribosomal proteins and so on So I think that even thoughwhat you say about redox control of certain genes mayactually be an important thing it is not the fundamentaloverall reason why the genomes are maintained It may bepart of the explanation for why certain genes are alwaysincluded in that subset but even in mitochondria I thinkthat cytochrome b is the only one that is always presentSo I think that both this great diversity of what has beenretained in different lineages and the nucleomorph caseindicate that evolution was able to do one thing in onelineage 500 million years ago and never got around todoing it for 500 million years in another lineage so thereis a lot more haphazardness and evolutionary accidentsand contingency than a straight functional interpretation
Phil Trans R Soc Lond B (2003)
there is a lot of just sheer lsquohappenedndashorndashdidnrsquotndashhappenrsquoor carryover
J F Allen I was not implying anything about the rangeof mitochondrial genomes I have a limited amount oftime so I just talked about what I could talk about withinthat limit Of course there will be a range and this willdepend upon the physiology and bioenergetics of theparticular organism But what I am trying to do is to seewhat the common properties of the irreducible core areof components that are always encoded in the organellesand cytochrome b is a perfect example because that reallyis the thing that binds these two all-important quinoneswithout which there is no Q cycle without which there isno proton pumping and bioenergetics and in Plasmodiumwhere the mitochondrion is reduced to this bare mini-mum there is still a mitochondrial gene for cytochromeb Now my argument about the ribosomal proteins andall the rest of it is that in order to be able to exert thisall-important redox control over gene expression of certaincore components you need an in situ genetic system andthis can vary in composition for reasons that I cannotdescribe Maybe you need RNA polymerase or maybeyou can import RNA polymerase In plastids you haveboth of these options You are going to need ribosomestransfer RNAs and the rest of the apparatus of geneexpression and that needs to be in situ but we are stillleft with the question why does there have to be any gen-etic system at all The mature tobacco or spinach chloro-plast seems to have an entire genetic system just to makeone protein D1 which is all it ever needs to resynthesizeI think that this is a clue It is worth having this very unec-onomical crazy system because there is an overridingrequirement to connect electron transfer with geneexpression directly and unambiguouslymdashwithout recourseto a bureaucracy if you like which is what I think thenucleus is
K Willison (Institute of Cancer Research Chester BeattyLaboratories London UK ) These photosynthetic reactioncomplexes are a sort of baroque mixture of polypeptidechains and cofactors and my understanding is that someof them in high photon flux can turn over within say 30minutes so I am wondering if there is a correlationbetween gene retention and the turnover rates of some ofthe components and the complexes it seems to me thatthat could really be a limiting factor and the reason thatthe proteins had to be encoded locally in order to get thecomplexes to be assembled rapidly
J F Allen I think that is right You can only alter thesteady-state concentration of a protein complex by affect-ing the synthesis and breakdown and therefore turnoverThere is an extreme example the D1 protein encoded bythe psbA gene of chloroplasts which is always in the chlor-oplast genome without exception High light will tend todegrade this protein and when the high light goes awaythere is an absolute need very quickly to bring it back upagain I think that is a very good example Whether thisargument applies to genes such as cytochrome oxidase Ido not know
C J Leaver (Department of Plant Sciences University ofOxford Oxford UK ) Your experiments tend to relatemainly to the gene expression side What about proteinRNA turnover the whole concept of redox control ofthatmdashdo we know anything
38 J F Allen Genomes in mitochondria and chloroplasts
J F Allen Well quite a lot is known about chloro-plasts There is redox control of messenger RNA stabilityfor this same D1 protein which has been described byStephen Mayfield and his co-workers The evidence is thatthe site of redox control is ferredoxin via thioredoxin andthe implication is that thioredoxin tells whether non-cyclicelectron transport is working at all When it is workingthere is a signal that says lsquoOK now continue with all theprotein synthesisrsquo It is not particularly specific but moreof a global effect I think this is what Mayfield argues butof course this is not inconsistent with my argument I haveconcentrated on quinone pools and medium-potentialredox regulatory control because that is where I can seesignals about the dangerous consequences of getting itwrong but there is control of other levels of geneexpression In mitochondria I am not so sure I wouldbounce the question back to you on that one
W Martin (Institute of Botany III Heinrich-Heine Univ-ersitat Dusseldorf Dusseldorf Germany) The nucleomorphgenome was cited as evidence in conflict with your modelbut is that true The nucleomorph genome is not in abioenergetic organelle in that sense It resides in the cyto-sol so does the size of the nucleomorph genome workagainst your model Is it really evidence against yourmodel
J F Allen I do not know why should it be evidenceI cannot explain why the nucleomorph has a genomeThat is why I clutched at the straw that it is storage fornucleic acid precursors
Phil Trans R Soc Lond B (2003)
T Cavalier-Smith The reason I introduced the nucleo-morph is not to say it is direct evidence against yourhypothesis but to show that there probably was no singlefunctional reason for the retention of every gene in everyorganelle It may be that there are specific physiologicalreasons for the retention of certain genes and thereforeyou might be right about a subset of genes but I verymuch doubt if there are specific physiological reasons forthe retention of every single gene that is retained differen-tially because odd things happen in evolution Somethings are easy and sometimes things are difficult Evol-ution is history
J F Allen Of course I agree with that I do not thinkI can explain everything but I do think I can explainsomething If I have done that then I am content
GLOSSARY
AAC ADPndashATP carrierCES control by epistasy of synthesisCORR co-location for redox regulationLHC light-harvesting complexPCC Pasteur culture collectionORF open reading frameRUBISCO ribulose-15-bisphosphate carboxylasendashoxygenaseTAT twin-arginine translocase
Visit httpwwwpubsroyalsocacuk to see an electronic appendix tothis paper

26 J F Allen Genomes in mitochondria and chloroplasts
existence of chloroplast and mitochondrial genomes thefocus of attention should therefore be on the relativelyconstant primary subset of bioenergetic genes (sect 3e)
(g) The principle of contemporary operation ofselection on gene location
For any species the distribution of genes between organelle(by the principle in sect 3e) and nucleus (by the principle in sect 3f)is the result of selective forces that continue to operate
This principle is essentially a statement contradictingboth the lsquofrozen accidentrsquo hypothesis and the hypothesisthat gene relocation is progressive predictable and stillincomplete (sect 4) In support of this principle it is becom-ing increasingly clear that mitochondria that lose theirfunction in aerobic respiration also lose their genomes asseen in the relict mitochondria of microsporidia(Embley amp Martin 1998 Van der Giezen et al 2002) andthe lsquomitosomersquo of Entamoeba histolytica (Tovar et al1999) In chloroplasts loss of photosynthesis results inloss of photosynthetic genes The examples of Epifagus (aparasitic higher plant) (Ems et al 1995 Wolfe et al 1992)and the residual apicomplexan plastids of Plasmodium(Wilson et al 1996a) and Toxoplasma where some plas-tidic genetic system is retained mean that some of theirgene products must be subject to redox regulation of geneexpression if this principle is correct Experimental test ofthis principle is possible using allotopic mutants as forthe predictions of the principle in sect 3f The evidence thatthe apicoplast (Wilson et al 1996a) is a plastid is largelygenetic the residual genome resembles that of photosyn-thetic plastids (chloroplasts) in gene organization ratherthan in content A plastid that has completely lost its gen-ome by virtue of having lost photosynthesis and all otherprocesses requiring redox regulation might be difficult todistinguish from an organelle derived in a similar way froma mitochondrion The persistence of targeted importedchaperones Hsp70 (Tovar et al 1999 Williams et al2002) and cp60 (Roger et al 1998) in residual organellesmight not distinguish a plastid ancestry from a mitochon-drial one
(h) The principle of primary involvement inenergy transduction
Those genes for which redox control is always vital to cellfunction have gene products involved in or closely connectedwith primary electron transfer These genes are always con-tained within the organelle
This principle would be violated and the CORRhypothesis refuted if any photosynthetic reaction centrecore subunit or respiratory chain subunit currentlyorganelle-encoded is found that functions in primary vec-torial electron transport and is nevertheless encoded in thenucleus and synthesized cytosolically as a precursor forimport all without any selective cost to the individualFrom the chloroplast transcriptional experiments ofPfannschmidt et al (1999ab) the chloroplast geneswhose transcription is regulated by the plastoquinoneredox state include psaAB (for photosystem I reactioncentre core polypeptides) psbA (for the photosystem IIpolypeptide D1) and rbcL (for the large subunit of RUB-ISCO interestingly following the same pattern of redoxcontrolmdashupregulated by oxidized plastoquinonemdashaspsbA) Transcription of the chloroplast genes petA
Phil Trans R Soc Lond B (2003)
(cytochrome f ) trnG (glycine tRNA) and rpoB (RNApolymerase) did not respond to plastoquinone redox statepetA for cytochrome f is predicted by this principle to beregulated at a different level of redox control or if at plas-toquinone at a different stage in gene expression Thisprediction might also apply to trnG and rpoB though thesequalify as members of the secondary subset that of geneticsystem genes to which the principle in sect 3i applies
(i) The principle of secondary involvement inenergy transduction
Genes whose products contribute to the organelle genetic sys-tem itself or whose products are associated with secondaryevents in energy transduction may be contained in theorganelle in one group of organisms but not in anotherdepending on the physiology and biochemistry of photosynthesisand respiration in the species concerned
Even phylogenetically moderately related species(within kingdoms) show differences in the location ofsome genes for example the ATP synthase subunits ofNeurospora and Saccharomyces (Attardi amp Schatz 1988)The CORR hypothesis also does not explicitly account forthe wide disparities in the distribution of genetic systemgenes for example the almost complete import of tRNAsin Chlamydomonas (GoldschmidtClermont 1998) Trans-port of nucleic acids across membranes is uncommon andwhy some RNAs should be organelle-encoded in somespecies but nuclearly encoded in others is not currentlyunderstood The only suggestion that can be made alongthe lines of CORR is that in some genetic systems genesmay need to be under the control of a regulatory systemthat responds to a signal arising within the organelle itselfThis signal could be the primary signal of altered redoxstate or some secondary signal Among the latter might befound signals that themselves report on the current stateof gene expression another piece of information that thenucleus might not have at its immediate disposal Suchsecondary signals could result at least in part fromCORR though other developmental and environmentalinputs may be imagined and would not detract fromCORR unless it could be shown that they provided a suf-ficient reason for the retention of the genetic system asa whole
(j) The principle of the nuclear encoding of redoxsignalling components
Components of the redox-signalling pathways upon whichCORR depends are themselves not involved in primary electrontransfer and so their genes have been relocated to the nucleusin accordance with sect 3f
This principle states that the redox signalling pathwaysthat are required for CORR should not themselves beexpected to utilize components whose biosynthesis mustbe under direct redox control Such components shouldtherefore fall into the major category of organellar pro-teins and be imported as precursors from the cytosol inline with the principle in sect 3f However the principleabove might be need to be modified in cases where ampli-fication of the redox signal is useful to the cell In thebacterial ntr system which was a precedent for CORR(Harrison et al 1990 Allen 1992) the histidine kinasentrB and aspartate response regulator ntrC form part ofthe gln operon whose transcription they control Odontella
Genomes in mitochondria and chloroplasts J F Allen 27
and Porphyra have plastids that encode more proteins thanthose of most chlorophytes (Stoebe et al 1998 Turmel etal 1999) and these proteins include two-component sig-nal transduction proteins prr and ycf27 (Stoebe et al1998) Extreme or extremely rapid environmental fluctu-ations might be catered for by amplification of signals thatare transmitted by signalling pathways whose componentsare under their own control It is not known whether ycf27and prr are in operons whose promoters their gene pro-ducts control It is also uncertain whether ycf27 is a redoxsignalling component and its distribution at least in itsplastidic location seems to be correlated with the presenceof a phycobilin-based light-harvesting system as found inred algae and cyanobacteria (Ashby amp Mullineaux1999ab Ashby et al 2002) The best guess of the originalstatement of the CORR hypothesis (Allen 1992 1993ac)was that bacterial two-component systems (Iuchi amp Lin1992 Allen 1993c Grebe amp Stock 1999) provide a plaus-ible means of exerting redox regulatory control over mito-chondrial and chloroplast gene expression andsubsequent research confirms the presence of two-compo-nent genes in genomes of Arabidopsis and yeast SeveralArabidopsis two-component genes are predicted to be tar-geted to mitochondria and chloroplasts (Forsberg et al2001) These must be considered as prime candidates forcomponents involved in redox regulation of chloroplasttranscription seen in the experiments of Pfannschmidt etal (1999a 2001a) The presumably homologous regulat-ory process serving to control photosystem stoichiometryin cyanobacteria the prr system (Li amp Sherman 2000) isnow characterized as a two-component system involvinga redox sensor and redox response regulator as originallydefined (Allen 1992 1993ac)
4 CONTRASTING HYPOTHESES CONCERNINGTHE RETENTION OF CYTOPLASMIC GENOMES
CORR is a recent addition to explanations that havebeen put forward to account for the persistence of gen-omes in organelles These models and their underlyingmechanisms are now discussed and compared
(a) There is no reason that is just how it isOne reaction to the problem of cytoplasmic genetic sys-
tems (sect 2) is that there is no problem and no particularreason for their retention However there is a very widephylogenetic distribution of a common core or primarysubset of organellar-encoded genes and one which haswithstood considerable genomic reorganization over evol-utionary time This constancy is seen for example in thesame genes being present in the chloroplasts of landplants with multiple copies of a single circular chloroplastchromosome and in dinoflagellate chloroplasts with acollection of different DNA minicircles each with essen-tially one gene (Zhang et al 1999 2001 Barbrook et al2001) Another suggested explanation of the retention ofmitochondrial and chloroplast genomes is simply torestate the endosymbiont hypothesis and to point out thatchloroplast and mitochondrial genes lsquocame inrsquo with thebacterial endosymbiont This suggestion however doesnot address the question of why some genes were retainedwhereas others were not It is also now clear that mostgenes that came in and still survive have nuclear and cyto-
Phil Trans R Soc Lond B (2003)
solic gene products whereas many chloroplast and mito-chondria proteins had a eukaryotic origin (Salzberg et al2001 Martin amp Russell 2003) The odds against a con-served pattern in gene location (eg table 1) arising bychance must be astronomical Thus there is a reason forthe retention of organellar genomes and a reason for theretention of the same primary subset of genes Becauseevolution proceeds by natural selection of chance vari-ation lsquoreasonrsquo in this context requires identification of thefunction of the location of a gene that is the selectivevalue of the phenotype produced by that location Evol-ution sometimes retains apparently functionless relics ofpreviously useful features but not for long especially ifthere is a cost
(b) The lock-in hypothesisThe lsquolock-inrsquo hypothesis of Bogorad (1975) was pro-
posed independently of the endosymbiont hypothesis anddoes not depend upon it According to the lock-in hypoth-esis core components of multi-subunit complexes must besynthesized de novo in the correct cellular compartmentotherwise photosystems or respiratory chain complexesfor example might assemble in the plasma membranevacuolar membrane or endoplasmic reticulum The lock-in hypothesis made no clear prediction about the conti-nuity of cytoplasmic genetic systems even though cyto-plasmic uniparental non-Mendelian inheritance for bothmitochondria (Roodyn amp Wilkie 1968) and plastids(Kirk amp Tilney-Bassett 1978) made it likely that organellarDNA the chondriome and the plastome replicated inde-pendently of nuclear DNA (Whitehouse 1969)
Mechanisms of protein import and targeting (Schatz1998 Koehler et al 1999 Leuenberger et al 1999 Gab-riel et al 2001 Jarvis amp Soll 2001 Pfanner amp Geissler2001 Duby amp Boutry 2002 Pfanner amp Wiedemann 2002)now effectively answer the problem of specifying the cor-rect locations for membrane protein complexes for whichthe lock-in hypothesis was proposed as a solution How-ever regulation of synthesis of an entire functional multi-subunit complex by control of synthesis of individual sub-units is still an important principle This seems to be afeature of the lsquoCESrsquo model (control by epistasy ofsynthesis) of Wollman (Wollman et al 1999 Eberhard etal 2002) which envisages nuclear control as all-embracingwith organellar genetic systems doing as it were whatthey are told In the hypothesis of CORR propoundedhere CES is equally important but the essence of CORRis that the organellar genetic system alone knows what arethe immediate and most important signals of environmen-tal change The organelle may keep the nucleus informedbut this is a lower priority than initiating an appropriateresponse CES and CORR agree that assembly of multi-subunit complexes is a complex business and requirescoordinated control in which no step is rate-limiting butmany may be obligatory
(c) Going going gone the evolutionary process oftransfer of genes from organelles to the
nucleus is still underway but incompleteThis hypothesis envisages that for most eukaryotes we
are now witnessing the last stages in a slow evolutionarytransfer of genes to the nucleus and this transfer may beexpected to proceed to completion in what is in evol-
28 J F Allen Genomes in mitochondria and chloroplasts
utionary terms the near future (Palmer 1997 Gray 19992000 Gray et al 1999 Adams et al 2000 Daley et al2002) A clear and compelling statement of this widelyheld position is that of Herrmann amp Westhoff (2001 p10)
hellipthe transfer of geneshelliptells that the partite plant gen-ome is not in a phylogenetic equilibrium All availabledata suggest that the ultimate aim of genome restructur-ing in the plant cell as in the eukaryotic cell in generalis the elimination of genome compartmentation whileretaining physiological compartmentation
One serious problem with this hypothesis is the dis-tinctly non-random sample of genes that remain to betransferred A relatively constant subset of genes is foundin bioenergetically functional mitochondria from fungi tomammals and angiosperms as in photosynthetic plastids(chloroplasts) from unicellular algae to angiosperms Thishypothesis therefore requires an additional ad hocassumption that some gene transfers are inherently moreprobable than others (this too is widely supposed seelater) and thus that gene transfer events proceed in thesame sequence in quite independent lineages Thereseems to be no evidence for this and the order of geneloss from mitochondria and chloroplasts has been used toinfer phylogeny (Martin et al 1998 Adams et al 2000Millen et al 2001) lsquoGoing going gonersquo (Palmer 1997)is difficult to refute categorically but it does not seem tobe consistent with recent evidence For example chromo-some two of Arabidopsis thaliana (Lin et al 1999) has asingle insertion encompassing with about 99 sequencesimilarity the complete Arabidopsis mitochondrial genome(Unseld et al 1997 Marienfeld et al 1999 Stupar et al2001) Unless this single transfer is a freak occurrencethis finding strongly indicates that gene transfer occurswith relative ease (Adams et al 2000 Henze amp Martin2001 Rujan amp Martin 2001 Salzberg et al 2001) Inaddition there is experimental evidence that randomlygenerated polypeptides are mitochondrially imported athigh frequency (Lemire et al 1989) The hypothesis thatwe now see the same few stragglers in diverse organellargenomes is not impossible but should be regarded as alast resort perhaps applicable only in extreme cases suchas the greatly reduced apicomplexan plastomes and chon-driomes Even there the hypothesis carries with it an obli-gation to explain why organisms with few organellar geneshave lsquoevolvedrsquo faster than those with many
(d ) The frozen accidentWhat may be described as lsquothe frozen accident hypoth-
esisrsquo states that the evolutionary process of gene transferfrom organelle to nucleus was underway when somethinghappened that stopped it One example of the frozen acci-dent is a scenario (Von Heijne 1986) in which import intomitochondria of the precursor proteins arising from newlytranslocated genes proceeded for long enough after theprimordial endosymbiosis for most of the symbiont-derived genes to be moved to the cell nucleus The lsquoacci-dentrsquo that halted the process was the evolutionary originof exocytosis and protein secretion after which it becameimpossible for precursors properly destined for mitochon-drial import to be distinguished from those destined forexport from the cell As with the lsquolock-inrsquo hypothesis the
Phil Trans R Soc Lond B (2003)
lsquofrozen accidentrsquo seems incompatible with the remarkableprecision and specificity of protein targeting (Schatz 1998Koehler et al 1999 Leuenberger et al 1999 Gabriel etal 2001 Jarvis amp Soll 2001 Pfanner amp Geissler 2001Duby amp Boutry 2002 Pfanner amp Wiedemann 2002) Pro-tein sorting mechanisms do not seem to forbid controlledimport into any cellular compartment Another lsquoaccidentrsquothat may have impeded subsequent gene transfer in thecase of mitochondria is their departure from what is other-wise a universal genetic code Genetic code differences arediscussed later in the context of whether some genes can-not be moved
(e) Some proteins (with cofactors) cannot beimported
We may describe as the lsquounimportability hypothesisrsquo theproposal that some proteins are inherently unimportableand must therefore be synthesized de novo in the correctcompartment within the cell The unimportability hypoth-esis is most often proposed as its special case the hydro-phobicity hypothesis with hydrophobicity being thereason for unimportability
(i) HydrophobicityA commonly entertained notion is that proteins that are
encoded and synthesized within organelles comprise a pri-mary subset characterized by shared hydrophobicity allare intrinsic membrane proteins (Von Heijne 1986Popot amp de Vitry 1990 Claros et al 1995) As with theCORR hypothesis favoured here the primary subset thenrequires a secondary subset of genetic system componentsthe latter not themselves being hydrophobic but beingrequired for synthesis of the others Certainly the mito-chondrial inner membrane contains some intrinsic proteincomplexes whose subunits broadly satisfy the criterion ofhydrophobicity However the core subunits that are mito-chondrially encoded are also explained by the CORRhypothesis This would make hydrophobicity a require-ment for their function in vectorial electron and protontransfer and not a reason in itself for retention of theirgenes in the cytoplasm
The hydrophobicity hypothesis has some very clearcounter-examples in chloroplasts and cannot thereforestand if one wishes for a single principle that embracesboth types of bioenergetic organelle One clear counter-example is the large (L) subunit of the enzyme RUBISCOan abundant but entirely membrane-extrinsic protein ofca 55 kDa molecular mass The RUBISCO large subunitis generally chloroplast-encoded with the exception of thenuclearly encoded bacterial-type L subunit of L2 RUBI-SCO in peridinin-containing dinoflagellates (Morse et al1995) In most photosynthetic eukaryotes RUBISCO hasa chloroplast encoded large subunit and a nuclearly enco-ded small (S) subunit (Ellis 1984 Gatenby amp Ellis 1990)giving an L8S8 holoenzyme and this RUBISCO is widelytaken as a paradigm for the need for coordination ofnuclear and chloroplast gene expression (Rodermel 2001)
By complete contrast to the RUBISCO large subunitwhich is a chloroplast-encoded and water-soluble proteina second major violation of the hydrophobicity hypothesisis the existence of the universally nuclearly encoded butalways hydrophobic subunits of the chloroplast LHC IIand LHC I (Chitnis amp Thornber 1988) These apopro-
Genomes in mitochondria and chloroplasts J F Allen 29
teins of ca 27ndash30 kDa are quite water-insoluble and areusually fully extracted only in chloroformndashmethanol or inthe presence of detergents The Lhc gene family is diverse(Green amp Kuhlbrandt 1995) but most gene productsor Lhcp (light-harvesting chlorophyll protein) polypeptideshave three membrane-spanning hydrophobic a-helices(Kuhlbrandt 1994) It is interesting that three-helix Lhcproteins seem to be absent from extant cyanobacteria andchloroxybacteria (Green amp Kuhlbrandt 1995)
(ii) The five-helix rulePerhaps with the three-helix model of LHC II in mind
a variation of the hydrophobicity hypothesis is the empiri-cal lsquofive-helix rulersquo (Popot amp de Vitry 1990 Claros et al1995 Wollman et al 1999) which states that no intrinsicmembrane protein with five or more membrane-spanninghelices can be encoded in the nucleus Five is a good guessfor a threshold number of helices demanding organellar-encoding in the context of photosynthetic reaction centres(Heathcote et al 2002) where the earliest known struc-ture that of type II centres (Deisenhofer et al 1985) asin photosystem II (Zouni et al 2001) shows exactly fivehelices with type I centres containing 11 (Jordan et al2001) There are however many nuclearly encoded sub-units in both mitochondria and chloroplasts that havefewer than five membrane helices so the five-helix rulecan only describe a sufficient condition and not a neces-sary condition for nuclear encoding
It should also be noted that ADPndashATP carriers (AACs)are encoded in the nucleus AACs of the mitochondrialinner membrane have six transmembrane helices (van derGiezen et al 2002) while the those of the chloroplast innerenvelope (Winkler amp Neuhaus 1999) have 12 (Tjaden etal 1998)
With the counter-examples of nuclearly encoded AACswith six or 12 membrane-spanning helices many intrinsicmembrane proteins with fewer than five helices somenuclearly encoded and some not and with the RUBISCOlarge subunit as a major product of chloroplast geneexpression with no membrane domain at all the five-helixrule begins to seem rather like a case of special pleadingEven if there are indeed special mitigating circumstancesfor AACs (this argument is outlined later) the five-helixrule as with the hydrophobicity hypothesis of which it isa special case will disappear if it can be shown that pos-session of five or more membrane helices is required forsome other function such as that of CORR that deter-mines the organellar location of the gene concernedHydrophobicity is in some cases a feature of proteins thatare encoded in organelles but it is not in itself the issue
For chloroplasts the hydrophobicity hypothesis predictsretention of cyanobacterial genes in functional categoriesassociated with the chloroplast envelope and with trans-port just as much as with photosynthesis Table 1 showsthat this is not the case
A mechanism for the unimportability of hydrophobicproteins is rarely stated explicitly and it is rather left tothe imagination whether hydrophobic proteins might asit were get stuck halfway through the chloroplast envelopeor mitochondrial outer membrane whether transport can-not get started on a precursor protein that has alreadyspontaneously inserted into the membrane or whetherhydrophobic proteins will be unable to make the journey
Phil Trans R Soc Lond B (2003)
through the cytosol without precipitating there In thecytosol co-translational transport or membrane insertionwould in principle address the latter point but it seemsto be generally true that chloroplast and mitochondrialproteins are post-translationally inserted into the organel-lar outer membrane So the obvious ancillary questionabout unimportability is lsquowhy then cannot hydrophobicproteins be co-translationally imported into organellesrsquoThere is evidence in yeast for co-translational transportinto mitochondria of proteins thought to be derived fromthe original endosymbiont (Marc et al 2002) If there isany truth in hydrophobicity as a barrier to protein importthe CORR principle of unselective import (sect 3c) shouldbe easily refuted for example by a demonstration that noknown targeting presequence is capable of importing poly-phenylalanine encoded by polyU for in vitro import stud-ies
An additional proposal intended to account for theunimportability of some hydrophobic proteins and forimportability of others is suggested by A de Grey(personal communication) who argues that bacteriallyderived protein precursors imported into mitochondriacross the mitochondrial inner membrane completelybefore insertion into that membrane from the mitochon-drial matrix (the N-phase) This is held to be an obstacleto successful import of membrane-intrinsic proteins thatmight en route discharge the proton motive force By con-trast eukaryotically derived proteins can be inserted intothe mitochondrial inner membrane from the inter-mem-brane space (the P-phase) and then there is for somereason no similar uncoupling effect Thus adenylatetransporters or AACs can be nuclearly encoded whereascytochrome oxidase subunits for example cannot It isnot obvious why bacterially derived subunits such asthose of cytochrome oxidase did not acquire eukaryoticinsertion mechanisms Perhaps this acquisition is forbid-den by the need to conserve pre-existing routes ofassembly into multi-subunit complexes or perhaps theneed for insertion from the inside reflects a need to incor-porate a cofactor for example haem or ironndashsulphur clus-ters along the way
(iii) Unimportability of three dimensional structures andcofactors
Accumulating evidence on protein transport mech-anisms seems to make a specific obstacle to successfultransport of certain sorts of protein less and less plausibleespecially given the observation that precursors with N-terminal targeting sequences not mature functional poly-peptides themselves traverse membranes during biosynth-esis
Protein folding inside the living cell is invariably assistedby molecular chaperones (Gatenby amp Ellis 1990 Azem etal 1997 Komiya et al 1997 1990 Jackson-Constan et al2001) proteins that impede the formation of lsquoimproperrsquostructural interactions that will otherwise produce aggre-gation or other non-functional interactions Proteinimport mechanisms appear mostly to rely on specific mol-ecular chaperones for guided unfolding before membraneinsertion and refolding after translocation of the polypep-tide into its destined compartment (Gatenby amp Ellis 1990Azem et al 1997 Komiya et al 1997 Jackson-Constanet al 2001) Because polypeptides are mostly lsquothreadedrsquo
30 J F Allen Genomes in mitochondria and chloroplasts
through membranes no special constraint on transportshould be expected to arise from 3D structure or surfaceproperties of a protein
An exception to this unfolding rule may be providedby holoproteins containing cofactors that are transportedalong with the apoprotein presumably with a largely intact3D structure The lsquoTATrsquo (twin-arginine translocase) sys-tem (Bogsch et al 1998 Dalbey amp Robinson 1999 Berkset al 2000ab Robinson 2000 Robinson amp Bolhuis 2001Sargent et al 2002) works broadly along these linesthough its mechanism is unresolved TAT is relevant herebecause it adds transport of essentially complete holopro-teins to the cellrsquos repertoire of transport mechanisms andeven proteins that may not be allowed to unfold perhapsbecause of the toxicity of the free cofactor are not therebymade candidates for an unimportability rule One variantof the unimportability hypothesis is that transport canwork only by unfolding and dissociated cofactors may betoxic to the cell The bacterially derived TAT system(Bogsch et al 1998 Dalbey amp Robinson 1999 Berks etal 2000ab Sargent et al 2002) overcomes this argumentas would examples of cofactor-free polypeptides such as(C)Fo subunit c which are nevertheless generally encodedin organelles
The presence of a vesicular transport system in chloro-plasts (Westphal et al 2001) counts against the unim-portability hypothesis Vesicular transport between thechloroplast envelope and the thylakoid membrane shouldin principle allow any protein to be ferried across thechloroplast stroma Vesicles might allow transport inparticular of membrane proteins that would discharge theproton motive force if allowed to connect transiently theN and P aqueous phases on each side of a coupling mem-brane
Another observation that seems to count against unim-portability is the increased number of membrane barriersin plastids derived from secondary tertiary or higher-orderendosymbioses These plastids seem to have retained ifanything fewer genes rather than more as unimportabilitypredicts The nucleomorph the residual nucleus of theprimary host retains many genes for molecular chap-erones (Archibald et al 2001 Cavalier-Smith 2002) inagreement with the conclusion that protein import pro-ceeds by conserved and specialized mechanisms If eachnew import added to gain a higher-order endosymbiosisrequires additional targeting presequences then perhapsthere is an advantage in components of an import machin-ery not being imported themselves
It seems that unimportability is less easy an assumptionto make today than in previous years and the onus maynow be on its proponents to cite specific examples of pro-teins that can never be imported into chloroplasts or mito-chondria
(f ) Some genes cannot be moved(i) Differences in the mitochondrial genetic code
The genetic code is the specific relation between ribon-ucleotide triplets of codons and tRNAs that recognizethem and which thus determine which individual aminoacids if any will be incorporated into the growing poly-peptide chain Mitochondria show small departures fromthe otherwise universal genetic code (Attardi amp Schatz1988 Alberts et al 1994) UGA is a lsquostoprsquo codon in the
Phil Trans R Soc Lond B (2003)
universal code and in plant mitochondria but is the codonfor tryptophan in mitochondria of animals and fungiCUA means leucine everywhere except fungal mitochon-dria where it means threonine and AGA and AGU codefor arginine in the lsquouniversalrsquo code but serine in insectmitochondria and lsquostoprsquo in mammalian ones The signifi-cance of this heterogeneity in coding is most unlikely torest in the early origin of mitochondria as the lsquouniversalrsquocode is employed by bacteria and protists Furthermorethe diversity of departures from the lsquouniversalrsquo codeamong mitochondria of different eukaryotic lineages indi-cates they had independent origins A general rule formitochondrial genetic codes is that reliance on the thirdlsquowobblersquo position is decreased which allows all 20 proteinamino acids to be specified by fewer tRNAs (22 in animalmitochondria instead of 31 in the cytosol) This in turnmeans that mtDNA can specify more proteins for a givenlength of DNA or conversely that the mitochondrialgenome can be condensed or lsquostreamlinedrsquo Although thisarrangement does not explain the retention of mitochon-drial genomes and may be a secondary event in mitochon-drial evolution it almost certainly places an extra barrierin place of gene relocation As discussed previously how-ever there seems to be no single barrier that can explainthe retention of a relatively constant subset of mitochon-drial genes
There has clearly been a contraction of mitochondrialDNA especially in animals Animal mitochondrial DNAhas few if any non-coding sequences and animal mito-chondrial genetic codes require fewer tRNAs than the sin-gle code operating in the cytosol (Gray 1999 Gray et al1999 Selosse et al 2001) These may be devices todecrease the total frequency of mutations per mitochon-drial genome assuming a constant frequency per unitlength of mitochondrial DNA It has been proposed thata genersquos removal to the nucleus decreases its susceptibilityto free radical mutagenesis (Raven et al 1994ab Allen ampRaven 1996) If this is the case then the non-standardcode may be a further indication of an overriding need tomaintain certain genes within the mitochondrion What-ever the reason for the retention of genes in organelles ithad better be a good one bioenergetic organelles areclearly the wrong place to keep genetic information andgo to extraordinary lengths to offset the consequences ofit being there
(ii) Molecular refugeesA phenomenon related to differences in the genetic code
is seen in chloroplasts This is a bias in codon usage Thelsquomolecular refugeesrsquo hypothesis (Howe et al 2000) sug-gests that nuclear copies of what were once chloroplastgenes escape constraints on codon usage and thus thatwhat distinguishes chloroplast-encoded proteins is thattheir amino-acid composition is unusual enough for thebias in codon usage not to apply It is difficult to see whatphysico-chemical properties might be uniquely shared bythe amino acids of chloroplast-encoded proteins and inaddition there seem to be no lsquoforbiddenrsquo amino-acid resi-due in chloroplasts As with the departure from the univer-sal genetic code in mitochondria is seems likely that abias in codon usage reflects some secondary property ofchloroplast genomes and is not the primary reason fortheir retention In any case any restriction on gene move-
Genomes in mitochondria and chloroplasts J F Allen 31
ment will act in the wrong direction to explain why chloro-plasts have retained genomes at all provided there is noresulting obstacle to a chloroplast protein being success-fully encoded as a precursor in the nucleus
(iii) Genes are copied and replicated not relocated in onestep
If complete relocation is envisaged as proceeding by thesimultaneous appearance of a gene in one compartmentand its deletion from another then it is possible to imaginethat whatever the vector is there may be selectivity ingene transfer (Henze amp Martin 2001) For example ifviral incorporation replication and recombination in bothorganelle and nucleus were the mechanism of relocationthen successful gene transfer might depend on the abilityof the virus to complete the process in one step This inturn might depend on some property of the gene or ofthe protein it encodes if expression occurs at any stage inthe process of relocation An assumption of the CORRhypothesis favoured here is that genes are not movedphysically but merely copied after which either the copyin the original location or the copy in the new locationmay be lost as a result of selection A useful analogy maybe the action of copying files between computer storagevolumes In this analogy the copy is identical to the orig-inal and either may subsequently be deleted without lossof information
Because any gene that is not usefully expressed willundergo no selection to maintain a functional allele theprevailing location will be the one in which the functionalcopy of the gene resides The problem of which gene sur-vives is then subsumed into the problem of which is theselectively advantageous location for it It seems reason-able to assume that there wasmdashand ismdashan interim periodin which homologous genes in different compartments areessentially in competition to see which functions best Asoutlined above there is no obvious obstacle for copyingDNA sequences within the cell The physico-chemicalproperties of DNA are fairly uniform and not directlydependent on nucleotide sequence though organelles likeintra-cellular parasites seem to prefer A and T to G andC The answer to the question lsquowhich gene location func-tions bestrsquo may have nothing to do with the gene itselfnor directly with the structure and function of its geneproduct Instead the answer may have everything to dowith the required regulatory interaction between the geneproduct and expression of the gene that produces it
(g) CORRThis hypothesis has been propounded earlier (sect 3) and
will be summarized as follows Vectorial electron and pro-ton transfer exerts regulatory control over expression ofgenes encoding proteins directly involved in or affectingredox poise This regulatory coupling is indispensable foradaptation to environmental changes in chemical activitiesof oxygen light and CO2 and requires co-location of thesegenes with their gene products Organelles lsquomake theirown decisionsrsquo on the basis of environmental changesaffecting redox state The hypothesis is abbreviatedCORR for co-location for redox regulation or more com-pletely lsquoco-location (of gene and gene product) for(evolutionary) continuity of redox regulation of geneexpressionrsquo
Phil Trans R Soc Lond B (2003)
5 SUMMARY OF THE CURRENT STATUS OF THEHYPOTHESIS OF CORR
Q Why do mitochondria and chloroplasts have theirown genetic systems
A CORR co-location of genes and gene products per-mits direct and autonomous redox regulation of geneexpression
(a) Objections to CORR(i) There is no regulation of gene expression in chloroplasts
and mitochondriaThis is still commonly stated but incorrect See the prin-
ciple in sect 3d
(ii) There is no regulation of gene expression in chloroplastsand mitochondria at the level of transcription
For chloroplasts there is now clear evidence for a func-tionally intelligible redox control of transcription(Pfannschmidt et al 1999a) This invalidates the previouswidely held assumption that all control of chloroplast geneexpression is post-transcriptional (Gruissem amp Tonkyn1993 GoldschmidtClermont 1998) The evidence formitochondria is less clear but redox control of transcrip-tion certainly cannot be ruled out and there are indi-cations that it may occur (Pearson et al 1993Konstantinov et al 1995 Wilson et al 1996b) It mustalso be restated that transcriptional control is not neces-sary for CORR but serves as a compelling example of aconserved typically bacterial mode of response that oper-ates still in eukaryotic cytoplasmic organelles thus illus-trating the principle of continuity of redox control (sect 3d)Post-transcriptional redox regulation (Danon amp Mayfield1994 Kim amp Mayfield 1997 Brown et al 2001 Treb-itsh amp Danon 2001) is consistent with CORR and thereis nothing to forbid it being a primary mechanism thatsatisfies most or indeed all of the formal requirements ofthe hypothesis in particular cases
(iii) There are no redox signalling components encoded inchloroplast or mitochondrial DNA
This is what CORR predicts See the principle in sect 3j
(iv) There is redox control of nuclear gene expressionThis is correct and many examples are now known
especially for chloroplasts (Escoubas et al 1995 Kovacs etal 2000 Pfannschmidt et al 2001ab Surpin et al 2002)Redox signals from photosynthetic or respiratory electrontransport chains are not at all prohibited by CORR Theessence of CORR is that there is a subset of organellarproteins for which there is an overriding requirement fora redox regulatory control that is initiated within theorganelle and whose effects are exerted by the simplestand most direct route This redox signalling is envisagedto take place without a requirement for prior consultationand coordination with what may be regarded as a nucleo-cytoplasmic bureaucracy
(v) Nuclear genes can be expressed as rapidly as chloroplastand mitochondrial genes
Zerges (2002) for example cites the equal times takenfor complete expression of nuclear and chloroplast genesin Chlamydomonas as evidence against the CORR hypoth-
32 J F Allen Genomes in mitochondria and chloroplasts
esis However the question is not one of the time takento express a gene but the time taken to alter the rate ofits expression in response to a (redox) signal originatingwithin the organelle An equally important additional fac-tor may be the autonomy of the signalling pathway thatis its independence from other inputs as described in theprevious paragraph One might say that mitochondria andchloroplasts require a free hand
(vi) There is no evidence to support this hypothesisThe hypothesis is rich in predictions (sectsect 3 and 6) and
those that have been tested stringently are in agreementwith experimental results None of the predictions of thehypothesis has been found to be incorrect and none of theunderlying principles (sect 3) has been disproved Furtherpredictions will be the subject of future experiments
(b) Evidence for CORR(i) Chloroplasts have selectively retained cyanobacterial genes
for primary events in photosynthesisSee table 1
(ii) There is regulation of gene expression in chloroplastsThere is transcriptional regulation of gene expression
in chloroplasts (sect 3d) The case for mitochondria is notyet proven
(iii) There is redox regulation of gene expression inchloroplasts
There is transcriptional redox regulation of geneexpression in chloroplasts (sect 3d) The case for mitochon-dria is not yet proven though see (Allen et al 1995 Kon-stantinov et al 1995 Galvis et al 1998)
(iv) There are no redox signalling components encoded inchloroplast or mitochondrial DNA
Correct but with possible exceptions discussed in sect 3j
(v) There are lsquobacterialrsquo redox signalling components ineukaryotes
See sect 3j
(vi) Further predictions DNA-array results and phenotypesof allotopic mutants
A systematic test of CORR might be to make an oxid-ized-minus-reduced gene expression profile of the lsquotran-scriptomersquo of a cyanobacterium to see if the set of geneswhose transcription is under most rapid and direct redoxcontrol corresponds to the genes of the chloroplast gen-ome Laboratories with this experimental capability do notseem to have addressed this possibility (Hihara et al 2002Ikeuchi 2002) Likewise if a putative modern representa-tive of the proto-mitochondrion can be found and agreedeg Rhodospirillum rubrum or Paracoccus denitrificans thena similar experiment should reveal a subset of genes corre-sponding to mitochondrial genomes DNA-microarraysalso present the possibility of testing the relative rate ofredox response and dependency on other signallinginputs of transcription of eukaryotic genes To dateorganellar genes have not been included in such investi-gations (Ferea et al 1999) A further experimental possi-bility is to examine the redox regulatory phenotypes ofexisting allotopic mutants as outlined in sect 3e
Phil Trans R Soc Lond B (2003)
6 CONCLUSION AND FUTURE PROSPECTS
The CORR hypothesis states that the function of mito-chondrial and chloroplast genomes is to provide co-location (of gene and gene product) for (evolutionary)continuity of redox regulation of gene expression Thishypothesis is neither self-contradictory nor devoid of test-able predictions It is also consistent with available evi-dence and in principle makes sense of the otherwisepuzzling distribution of genes between compartments ineukaryotic cells The explanation that CORR offersapplies equally to mitochondria and chloroplasts andidentifies selective forces that still operate to determine thelocation of genes No selective barriers need be assumedto gene transfer protein import nor membrane insertionNor do we need to invoke a lsquofrozen accidentrsquo that some-thing happened to stop gene transfer for some reason atsome time in the remote and inaccessible past CORR alsoexplains the absence of genomes from organelles that havelost their primary bioenergetic functions CORR may actas a stimulus to a search for residual bioenergetic functionin those organelles that have lost aerobic respiration andphotosynthesis but where residual genomes remain
CORR also predicts regulatory properties of knowncomponents of chloroplasts and mitochondria that in turnindicate important but as yet uncharacterized flexibility inenergy transduction These components include the RUB-ISCO large subunit and the CFo and Fo subunits of coup-ling ATPase in both chloroplasts and mitochondriaRedox regulation of the relative stoichiometry of compo-nents as seen in regulation of chloroplast photosystemstoichiometry (Pfannschmidt et al 1999a) may extend to(C)Fo-(C)F1-ATPase indicating that the function ofretention of genes for one or more of these componentsis flexibility in the stoichiometries of H1 to e2 and H1 toATP in chemiosmotic coupling (Schemidt et al 1998Stock et al 2000 Allen 2002)
So far explicit tests of CORR have largely been con-fined to chloroplasts and to some extent mitochondria ofhigher plants but this merely reflects the expertise andinterests of the investigators Further attempted disproofof CORR is welcomed and encouraged Novel experi-mental tests can be devised as well as attempted refu-tation through analysis of existing data It is hoped thatexplicit attention will be given to the CORR hypothesis inlaboratories already suitably equipped and that resourcescan be allocated for this purpose in laboratories wherethere is the interest but not currently the technology orexpertise
The heuristic value of the hypothesis already seen inthe role of redox-regulated transcription of photosyntheticreaction centre genes in chloroplasts may be its most use-ful property The central proposal is that mitochondriaand chloroplasts contain genes for proteins whose functionin electron transfer demands rapid direct and uncon-ditional redox regulatory control of their biosynthesis Thefull implications of this hypothesis are very broad and onlyjust beginning to be explored
I thank Carol Allen Aubrey de Grey William Martin John ARaven and Francis-Andre Wollman for discussion and corre-spondence during the preparation of this paper and WilliamMartin for suggestions including table 1 Research grants from
Genomes in mitochondria and chloroplasts J F Allen 33
The Swedish Natural Sciences Research Council (NFR) andthe Crafoord Foundation are gratefully acknowledged
REFERENCES
Adams K L Daley D O Qiu Y L Whelan J amp PalmerJ D 2000 Repeated recent and diverse transfers of a mito-chondrial gene to the nucleus in flowering plants Nature408 354ndash357
Alberts B Bray D Lewis J Raff M Roberts K amp Wat-son J D 1994 The molecular biology of the cell New Yorkand London Garland Publishing
Allen C A Hakansson G amp Allen J F 1995 Redox con-ditions specify the proteins synthesized by isolated chloro-plasts and mitochondria Redox Rep 1 119ndash123
Allen J F 1992 Protein phosphorylation in regulation ofphotosynthesis Biochim Biophys Acta 1098 275ndash335
Allen J F 1993a Control of gene expression by redox poten-tial and the requirement for chloroplast and mitochondrialgenomes J Theor Biol 165 609ndash631
Allen J F 1993b Redox control of gene expression and thefunction of chloroplast genomes a hypothesis PhotosynthRes 36 95ndash102
Allen J F 1993c Redox control of transcription sensorsresponse regulators activators and repressors FEBS Lett332 203ndash207
Allen J F 1995 Thylakoid proteinndashphosphorylation state-1ndashstate-2 transitions and photosystem stoichiometry adjust-ment redox control at multiple levels of gene expressionPhysiol Plant 93 196ndash205
Allen J F 1996 Separate sexes and the mitochondrial theoryof ageing J Theor Biol 180 135ndash140
Allen J F 2002 Photosynthesis of ATP-electrons protonpumps rotors and poise Cell 110 273ndash276
Allen J F amp Allen C A 1999 A mitochondrial model forpremature ageing of somatically cloned mammals IUBMBLife 48 369ndash372
Allen J F amp Nilsson A 1997 Redox signalling and the struc-tural basis of regulation of photosynthesis by protein phos-phorylation Physiol Plant 100 863ndash868
Allen J F amp Pfannschmidt T 2000 Balancing the two photo-systems photosynthetic electron transfer governs transcrip-tion of reaction centre genes in chloroplasts Phil TransR Soc Lond B 355 1351ndash1357 (DOI 101098rstb20000697)
Allen J F amp Race H L 2002 Will the real LHC II kinaseplease step forward Sciencersquos STKE httpwwwstkeorgcgicontentfullsigtrans2002155pe43
Allen J F amp Raven J A 1996 Free-radical-induced mutationvs redox regulation costs and benefits of genes in organellesJ Mol Evol 42 482ndash492
Archibald J M Cavalier-Smith T Maier U amp Douglas S2001 Molecular chaperones encoded by a reduced nucleusthe cryptomonad nucleomorph J Mol Evol 52 490ndash501
Ashby M K amp Mullineaux C W 1999a Cyanobacterialycf27 gene products regulate energy transfer from phycobili-somes to photosystems I and II FEMS Microbiol Lett 181253ndash260
Ashby M K amp Mullineaux C W 1999b The role of ApcDand ApcF in energy transfer from phycobilisomes to PSI andPSII in a cyanobacterium Photosynth Res 61 169ndash179
Ashby M K Houmard J amp Mullineaux C W 2002 Theycf27 genes from cyanobacteria and eukaryotic algae distri-bution and implications for chloroplast evolution FEMSMicrobiol Lett 214 25ndash30
Attardi G amp Schatz G 1988 Biogenesis of mitochondria ARev Cell Biol 4 289ndash333
Azem A Oppliger W Lustig A Jeno P Feifel B SchatzG amp Horst M 1997 The mitochondrial hsp70 chaperone
Phil Trans R Soc Lond B (2003)
system effect of adenine nucleotides peptide substrate andmGrpE on the oligomeric state of mhsp70 J Biol Chem272 20 901ndash20 906
Barbrook A C Symington H Nisbet R E R LarkumA amp Howe C J 2001 Organisation and expression of theplastid genome of the dinoflagellate Amphidinium opercula-tum Mol Genet Genomics 266 632ndash638
Bauer C E Elsen S amp Bird T H 1999 Mechanisms forredox control of gene expression A Rev Microbiol 53495ndash523
Bauer C E Elsen S Swem L R Swem D L amp MasudaS 2003 Redox and light regulation of gene expression inphotosynthetic prokaryotes Phil Trans R Soc Lond B 358147ndash154 (DOI 101098rstb20021189)
Berks B C Sargent F De Leeuw E Hinsley A P Stan-ley N R Jack R L Buchanan G amp Palmer T 2000aA novel protein transport system involved in the biogenesisof bacterial electron transfer chains Biochim Biophys ActaBioenerg 1459 325ndash330
Berks B C Sargent F amp Palmer T 2000b The tat proteinexport pathway Mol Microbiol 35 260ndash274
Blackstone N W 1999 Redox control in development andevolution evidence from colonial hydroids J Exp Biol 2023541ndash3553
Blackstone N W 2000 Redox control and the evolution ofmulticellularity Bioessays 22 947ndash953
Blackstone N W 2001 Redox state reactive oxygen speciesand adaptive growth in colonial hydroids J Exp Biol 2041845ndash1853
Bogorad L 1975 Evolution of organelles and eukaryotic gen-omes Science 188 891ndash898
Bogsch E G Sargent F Stanley N R Berks B C Robin-son C amp Palmer T 1998 An essential component of a novelbacterial protein export system with homologues in plastidsand mitochondria J Biol Chem 273 18 003ndash18 006
Brown E C Somanchi A amp Mayfield S P 2001 Interor-ganellar crosstalk new perspectives on signaling from thechloroplast to the nucleus Genome Biol 2 10211ndash10214
Cavalier-Smith T 2002 Chloroplast evolution secondarysymbiogenesis and multiple losses Curr Biol 12 R62ndashR64
Chitnis P R amp Thornber J P 1988 The major light-har-vesting complex of photosystem 2 Aspects of its molecularand cell biology Photosynth Res 16 41ndash63
Chow W S Melis A amp Anderson J M 1990 Adjustmentsof photosystem stoichiometry in chloroplasts improve thequantum efficiency of photosynthesis Proc Natl Acad SciUSA 87 7502ndash7506
Claros M G Perea J Shu Y Samatey F A PopotJ L amp Jacq C 1995 Limitations to in vivo import of hydro-phobic proteins into yeast mitochondria The case of a cyto-plasmically synthesized apocytochrome b Eur J Biochem228 762ndash771
Dalbey R E amp Robinson C 1999 Protein translocation intoand across the bacterial plasma membrane and the plant thy-lakoid membrane Trends Biochem Sci 24 17ndash22
Daley D O Adams K L Clifton R Qualmann S MillarA H Palmer J D Pratje E amp Whelan J 2002 Genetransfer from mitochondrion to nucleus novel mechanismsfor gene activation from Cox2 Plant J 30 11ndash21
Danon A amp Mayfield S P 1994 Light-regulated translationof chloroplast messenger RNAs through redox potentialScience 266 1717ndash1719
de Grey A 2000 Mitochondrial gene therapy an arena for thebiomedical use of inteins Trends Biotechnol 18 394ndash399
Deisenhofer J Epp O Miki K Huber R amp Michel H1985 Structure of the protein subunits in the photosyntheticreaction center of Rhodopseudomonas viridis at 3Ac resolutionNature 318 618ndash624
34 J F Allen Genomes in mitochondria and chloroplasts
Delwiche C F 1999 Tracing the thread of plastid diversitythrough the tapestry of life Am Nat 154 S164ndashS177
Doolittle W F 1999 Phylogenetic classification and the uni-versal tree Science 284 2124ndash2129
Douglas S E 1998 Plastid evolution origins diversitytrends Curr Opin Genet Dev 8 655ndash661
Duby G amp Boutry M 2002 Mitochondrial protein importmachinery and targeting information Plant Sci 162 477ndash490
Eberhard S Drapier D amp Wollman F A 2002 Searchinglimiting steps in the expression of chloroplast encoded pro-teins relations between gene copy number transcriptiontranscript abundance and translation rate in the chloroplastof Chlamydomonas reinhardtii Plant J 31 149ndash160
El Bissati K amp Kirilovsky D 2001 Regulation of psbA andpsaE expression by light quality in Synechocystis species PCC6803 A redox control mechanism Plant Physiol 1251988ndash2000
Ellis R J 1984 The nuclear domination of chloroplast devel-opment Science Progress 69 129ndash142
Embley T M amp Martin W 1998 Molecular evolution ahydrogen-producing mitochondrion Nature 396 517ndash519
Ems S C Morden C W Dixon C K Wolfe K HdePamphilis C W amp Palmer J D 1995 Transcriptionsplicing and editing of plastid RNAs in the non-photosyn-thetic plant Epifagus virginiana Plant Mol Biol 29 721ndash733
Eraso J M amp Kaplan S 2000 From redox flow to gene regu-lation role of the PrrC protein of Rhodobacter sphaeroides241 Biochemistry 39 2052ndash2062
Eraso J M amp Kaplan S 2002 Redox flow as an instrumentof gene regulation Meth Enzymol 348 216ndash229
Escoubas J M Lomas M Laroche J amp Falkowski P G1995 Light-intensity regulation of cab gene-transcription issignaled by the redox state of the plastoquinone pool ProcNatl Acad Sci USA 92 10 237ndash10 241
Ferea T L Botstein D Brown P O amp Rosenzweig R F1999 Systematic changes in gene expression patterns follow-ing adaptive evolution in yeast Proc Natl Acad Sci USA96 9721ndash9726
Forsberg J Rosenquist M Fraysse L amp Allen J F 2001Redox signalling in chloroplasts and mitochondria genomicand biochemical evidence for two-component regulatory sys-tems in bioenergetic organelles Biochem Soc Trans 29403ndash407
Fujita Y 1997 A study on the dynamic features of photosys-tem stoichiometry accomplishments and problems forfuture studies Photosynth Res 53 83ndash93
Gabriel K Buchanan S K amp Lithgow T 2001 The alphaand the beta protein translocation across mitochondrial andplastid outer membranes Trends Biochem Sci 26 36ndash40
Galvis M L E Allen J F amp Hakansson G 1998 Proteinsynthesis by isolated pea mitochondria is dependent on theactivity of respiratory complex II Curr Genet 33 320ndash329
Gatenby A A amp Ellis R J 1990 Chaperone function theassembly of ribulose bisphosphate carboxylase-oxygenase ARev Cell Biol 6 125ndash149
GoldschmidtClermont M 1998 Coordination of nuclear andchloroplast gene expression in plant cells Int Rev CytolSurv Cell Biol 177 115ndash180
Gray M W 1992 The endosymbiont hypothesis revisited IntRev Cytol 141 233ndash357
Gray M W 1999 Evolution of organellar genomes CurrOpin Genet Dev 9 678ndash687
Gray M W 2000 Evolutionary biology mitochondrial geneson the move Nature 408 302ndash305
Gray M W amp Doolittle W F 1982 Has the endosymbionthypothesis been proven Microbiol Rev 46 1ndash42
Gray M W Burger G amp Lang B F 1999 Mitochondrialevolution Science 283 1476ndash1481
Phil Trans R Soc Lond B (2003)
Grebe T W amp Stock J B 1999 The histidine protein kinasesuperfamily Adv Microb Physiol 41 139ndash227
Green B R amp Kuhlbrandt W 1995 Sequence conservationof light-harvesting and stress-response proteins in relation tothe 3-dimensional molecular structure of Lhcii PhotosynthRes 44 139ndash148
Gruissem W amp Tonkyn J C 1993 Control mechanisms ofplastid gene expression Crit Rev Plant Sci 12 19ndash55
Hackstein J H P (and 11 others) 2001 Hydrogenosomesconvergent adaptations of mitochondria to anaerobicenvironments Zool Anal Complex Syst 104 290ndash302
Harrison M A Keen J N Findlay J B C amp Allen J F1990 Modification of a Glnb-like gene-product by photosyn-thetic electron-transport in the cyanobacterium Synechoc-occus 6301 FEBS Lett 264 25ndash28
Harvey A J Kind K L amp Thompson J G 2002 Redoxregulation of early embryo development Reproduction 123479ndash486
Heathcote P Fyfe P K amp Jones M R 2002 Reactioncentres the structure and evolution of biological solarpower Trends Biochem Sci 27 79ndash87
Henze K amp Martin W 2001 How do mitochondrial genesget into the nucleus Trends Genet 17 383ndash387
Herrmann R G amp Westhoff P 2001 Thylakoid biogenesis anddynamics the result of a complex phylogenetic puzzle InRegulation of photosynthesis vol 11 (ed E-M Aro amp BAndersson) pp 1ndash28 Dordrecht The Netherlands Kluwer
Hihara Y Sonoike K Kanehisa M amp Ikeuchi M 2002Search for redox responsive genes in Synechocystis sp PCC6803 by using DNA microarrays Plant Cell Physiol 43S178
Howe C J Barbrook A C amp Lockhart P J 2000 Organellegenes do they jump or are they pushed Trends Genet 1665ndash66
Ikeuchi M 2002 Gene expression and responses to light stressin cyanobacteria Plant Cell Physiol 43 S21
Iuchi S amp Lin E C C 1992 Mutational analysis of signaltransduction by Arcb a membrane sensor protein respon-sible for anaerobic repression of operons involved in the cen-tral aerobic pathways in Escherichia coli J Bacteriol 1743972ndash3980
Jackson-Constan D Akita M amp Keegstra K 2001 Molecu-lar chaperones involved in chloroplast protein importBiochim Biophys Acta Mol Cell Res 1541 102ndash113
Jarvis P amp Soll M 2001 Toc Tic and chloroplast proteinimport Biochim Biophys Acta Mol Cell Res 1541 64ndash79
Jordan P Fromme P Witt H T Klukas O SaengerW amp Krauss N 2001 Three-dimensional structure ofcyanobacterial photosystem I at 25 acngstrom resolutionNature 411 909ndash917
Kaneko T (and 23 others) 1996a Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regionsDNA Res 3 109ndash136
Kaneko T (and 23 others) 1996b Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regions[Supplement] DNA Res 3 185ndash209
Kanevski I amp Maliga P 1994 Relocation of the plastid rbcLgene to the nucleus yields functional ribulose-15-bisphosph-ate carboxylase in tobacco chloroplasts Proc Natl Acad SciUSA 91 1969ndash1973
Kim J amp Mayfield S P 1997 Protein disulfide isomerase as aregulator of chloroplast translational activation Science 2781954ndash1957
Genomes in mitochondria and chloroplasts J F Allen 35
Kirk J T O amp Tilney-Bassett R A E 1978 The plastidsTheir chemistry structure growth and inheritance Amster-dam Elsevier
Koehler C M Merchant S amp Schatz G 1999 How mem-brane proteins travel across the mitochondrial intermem-brane space Trends Biochem Sci 24 428ndash432
Komiya T Rospert S Schatz G amp Mihara K 1997 Bind-ing of mitochondrial precursor proteins to the cytoplasmicdomains of the import receptors Tom70 and Tom20 isdetermined by cytoplasmic chaperones FASEB J 11 1196
Konstantinov Y M Lutsenko G N amp Podsosonny V A1995 Genetic functions of isolated maize mitochondriaunder model changes of redox conditions Biochem MolBiol Int 36 319ndash326
Korab-Laskowska M Rioux P Brossard N LittlejohnT G Gray M W Lang B F amp Burger G 1998 Theorganelle genome database project (GOBASE) Nucleic AcidsRes 26 138ndash144
Kovacs L Wiessner W Kis M Nagy F Mende D ampDemeter S 2000 Short- and long-term redox regulation ofphotosynthetic light energy distribution and photosystemstoichiometry by acetate metabolism in the green alga Chla-mydobotrys stellata Photosynth Res 65 231ndash247
Kuhlbrandt W 1994 Structure and function of the plant light-harvesting complex LHC-II Curr Opin Struct Biol 4519ndash528
Lang B F Gray M W amp Burger G 1999 Mitochondrialgenome evolution and the origin of eukaryotes A RevGenet 33 351ndash397
Lemire B D Fankhauser C Baker A amp Schatz G 1989The mitochondrial targeting function of randomly generatedpeptide sequences correlates with predicted helical amphi-philicity J Biol Chem 264 20 206ndash20 215
Leuenberger D Bally N A Schatz G amp Koehler C M1999 Different import pathways through the mitochondrialintermembrane space for inner membrane proteins EMBOJ 18 4816ndash4822
Li H amp Sherman L A 2000 A redox-responsive regulator ofphotosynthesis gene expression in the cyanobacterium Syne-chocystis sp strain PCC 6803 J Bacteriol 182 4268ndash4277
Lin X Y (and 36 others) 1999 Sequence and analysis of chro-mosome 2 of the plant Arabidopsis thaliana Nature 402761ndash768
Lonsdale D M Hodge T P Howe C J amp Stern D B1983 Maize mitochondrial-DNA contains a sequence hom-ologous to the ribulose-15-bisphosphate carboxylase largesubunit gene of chloroplast DNA Cell 34 1007ndash1014
McFadden G I 1999 Endosymbiosis and evolution of theplant cell Curr Opin Plant Biol 2 513ndash519
McFadden G I 2001 Chloroplast origin and integrationPlant Physiol 125 50ndash53
Mahler H R amp Raff R A 1975 The evolutionary origin ofthe mitochondrion a nonsymbiotic model Int Rev Cytol43 1ndash124
Manfredi G Fu J Ojaimi J Sadlock J E Kwong J QGuy J amp Schon E A 2002 Rescue of a deficiency in ATPsynthesis by transfer of MTATP6 a mitochondrial DNA-encoded gene to the nucleus Nature Genet 30 394ndash399
Marc P Margeot A Devaux F Blugeon C Corral-Debrinski M amp Jacq C 2002 Genome-wide analysis ofmRNAs targeted to yeast mitochondria EMBO Rep 3159ndash164
Marienfeld J Unseld M amp Brennicke A 1999 The mito-chondrial genome of Arabidopsis is composed of both nativeand immigrant information Trends Plant Sci 4 495ndash502
Martin W 1999 A briefly argued case that mitochondria andplastids are descendants of endosymbionts but that thenuclear compartment is not Proc R Soc Lond B 2661387ndash1395 (DOI 101098rspb19990792)
Phil Trans R Soc Lond B (2003)
Martin W amp Herrmann R G 1998 Gene transfer fromorganelles to the nucleus how much what happens andwhy Plant Physiol 118 9ndash17
Martin W amp Muller M 1998 The hydrogen hypothesis forthe first eukaryote Nature 392 37ndash41
Martin W amp Russell M J 2003 On the origins of cells ahypothesis for the evolutionary transitions from abiotic geo-chemistry to chemoautotrophic prokaryotes and from pro-karyotes to nucleated cells Phil Trans R Soc Lond B 35859ndash85 (DOI 101098rstb20021183)
Martin W Stoebe B Goremykin V Hansmann S Hase-gawa M amp Kowallik K V 1998 Gene transfer to thenucleus and the evolution of chloroplasts Nature 393162ndash165
Martin W Hoffmeister M Rotte C amp Henze K 2001 Anoverview of endosymbiotic models for the origins of eukary-otes their ATP-producing organelles (mitochondria andhydrogenosomes) and their heterotrophic lifestyle BiolChem 382 1521ndash1539
Martin W Rujan T Richly E Hansen A Cornelsen SLins T Leister D Stoebe B Hasegawa M amp PennyD 2002 Evolutionary analysis of Arabidopsis cyanobacterialand chloroplast genomes reveals plastid phylogeny and thou-sands of cyanobacterial genes in the nucleus Proc NatlAcad Sci USA 99 12 246ndash12 251
Millen R S (and 12 others) 2001 Many parallel losses of infAfrom chloroplast DNA during angiosperm evolution withmultiple independent transfers to the nucleus Plant Cell 13645ndash658
Morse D Salois P Markovic P amp Hastings J W 1995 Anuclear-encoded form-II RUBISCO in dinoflagellatesScience 268 1622ndash1624
Murakami A Kim S J amp Fujita Y 1997 Changes in photo-system stoichiometry in response to environmental con-ditions for cell growth observed with the cyanophyteSynechocystis PCC 6714 Plant Cell Physiol 38392ndash397
Oh J I amp Kaplan S 2000 Redox signaling globalization ofgene expression EMBO J 19 4237ndash4247
Ozawa T 1995 Mitochondrial-DNA mutations associatedwith aging and degenerative diseases Exp Gerontol 30269ndash290
Palmer J D 1997 Organelle genomes going going goneScience 275 790ndash791
Pearson C K Wilson S B Schaffer R amp Ross A W 1993NAD turnover and utilization of metabolites for RNA syn-thesis in a reaction sensing the redox state of the cyto-chrome-b( 6 )f complex in isolated chloroplasts Eur JBiochem 218 397ndash404
Pfanner N amp Geissler A 2001 Versatility of the mitochon-drial protein import machinery Natl Rev Mol Cell Biol 2339ndash349
Pfanner N amp Wiedemann N 2002 Mitochondrial proteinimport two membranes three translocases Curr Opin CellBiol 14 400ndash411
Pfannschmidt T Nilsson A amp Allen J F 1999a Photosyn-thetic control of chloroplast gene expression Nature 397625ndash628
Pfannschmidt T Nilsson A Tullberg A Link G amp AllenJ F 1999b Direct transcriptional control of the chloroplastgenes psbA and psaAB adjusts photosynthesis to light energydistribution in plants IUBMB Life 48 271ndash276
Pfannschmidt T Allen J F amp Oelmuller R 2001a Prin-ciples of redox control in photosynthesis gene expressionPhysiol Plant 112 1ndash9
Pfannschmidt T Schutze K Brost M amp Oelmuller R2001b A novel mechanism of nuclear photosynthesis generegulation by redox signals from the chloroplast during pho-
36 J F Allen Genomes in mitochondria and chloroplasts
tosystem stoichiometry adjustment J Biol Chem 27636 125ndash36 130
Popot J L amp de Vitry C 1990 On the microassembly of inte-gral membrane proteins A Rev Biophys Biophys Chem 19369ndash403
Race H L Herrmann R G amp Martin W 1999 Why haveorganelles retained genomes Trends Genet 15 364ndash370
Raven J A Johnston A M Parsons R amp Kubler J 1994aThe influence of natural and experimental high O2 concen-trations on O2 evolving phototrophs Biol Rev CambridgePhil Soc 69 61ndash94
Raven J A Johnston A M Parsons R amp Kubler J 1994bThe occurrence and influence on photolithotrophs of highoxygen concentrations Proc R Soc Edinb B 102 193ndash201
Robinson C 2000 The twin-arginine translocation system anovel means of transporting folded proteins in chloroplastsand bacteria Biol Chem 381 89ndash93
Robinson C amp Bolhuis A 2001 Protein targeting by the twin-arginine translocation pathway Natl Rev Mol Cell Biol 2350ndash356
Rodermel S 2001 Pathways of plastid-to-nucleus signalingTrends Plant Sci 6 471ndash478
Roger A J 1999 Reconstructing early events in eukaryoticevolution Am Nat 154 S146ndashS163
Roger A J Svard S G Tovar J Clark C G SmithM W Gillin F D amp Sogin M L 1998 A mitochondrial-like chaperonin 60 gene in Giardia lamblia evidence thatdiplomonads once harbored an endosymbiont related to theprogenitor of mitochondria Proc Natl Acad Sci USA 95229ndash234
Roodyn D B amp Wilkie D 1968 The biogenesis of mitochondriaMethuenrsquos monographs on biological subjects London Methuen
Rujan T amp Martin W 2001 How many genes in Arabidopsiscome from cyanobacteria An estimate from 386 proteinphylogenies Trends Genet 17 113ndash120
Saccone C Gissi C Lanave C Larizza A Pesole G ampReyes A 2000 Evolution of the mitochondrial genetic sys-tem an overview Gene 261 153ndash159
Salzberg S L White O Peterson J amp Eisen J A 2001Microbial genes in the human genome lateral transfer orgene loss Science 292 1903ndash1906
Sargent F Berks B C amp Palmer T 2002 Assembly of mem-brane-bound respiratory complexes by the Tat protein-trans-port system Arch Microbiol 178 77ndash84
Schatz G 1998 Protein transport the doors to organellesNature 395 439ndash440
Schemidt R A Qu J Williams J R amp Brusilow W S1998 Effects of carbon source on expression of F0 genes andon the stoichiometry of the c subunit in the F1 F0 ATPaseof Escherichia coli J Bacteriol 180 3205ndash3208
Selosse M A Albert B R amp Godelle B 2001 Reducing thegenome size of organelles favours gene transfer to thenucleus Trends Ecol Evol 16 135ndash141
Stern D B amp Lonsdale D M 1982 Mitochondrial and chlo-roplast genomes of maize have a 12- kilobase DNA-sequencein common Nature 299 698ndash702
Stock D Gibbons C Arechaga I Leslie A G W ampWalker J E 2000 The rotary mechanism of ATP synthaseCurr Opin Struct Biol 10 672ndash679
Stoebe B amp Maier U G 2002 One two three naturersquos toolbox for building plastids Protoplasma 219 123ndash130
Stoebe B Martin W amp Kowallik K V 1998 Distributionand nomenclature of protein-coding genes in 12 sequencedchloroplast genomes Plant Mol Biol Rep 16 243ndash255
Stupar R M Lilly J W Town C D Cheng Z Kaul SBuell C R amp Jiang J M 2001 Complex mtDNA consti-tutes an approximate 620 kb insertion on Arabidopsis thali-ana chromosome 2 implication of potential sequencing
Phil Trans R Soc Lond B (2003)
errors caused by large-unit repeats Proc Natl Acad SciUSA 98 5099ndash5103
Surpin M Larkin R M amp Chory J 2002 Signal transduc-tion between the chloroplast and the nucleus Plant Cell 14S327ndashS338
Tjaden J Schwoppe C Mohlmann T Quick P W ampNeuhaus H E 1998 Expression of a plastidic ATPADPtransporter gene in Escherichia coli leads to a functional aden-ine nucleotide transport system in the bacterial cytoplasmicmembrane J Biol Chem 273 9630ndash9636
Tovar J Fischer A amp Clark C G 1999 The mitosome a novelorganelle related to mitochondria in the amitochondrial para-site Entamoeba histolytica Mol Microbiol 32 1013ndash1021
Trebitsh T amp Danon A 2001 Translation of chloroplastpsbA mRNA is regulated by signals initiated by both photos-ystems II and I Proc Natl Acad Sci USA 98 12 289ndash12 294
Tullberg A Alexciev K Pfannschmidt T amp Allen J F2000 Photosynthetic electron flow regulates transcription ofthe psaB gene in pea (Pisum sativum L) chloroplasts throughthe redox state of the plastoquinone pool Plant Cell Physiol41 1045ndash1054
Turmel M Lemieux C Burger G Lang B F Otis CPlante I amp Gray M W 1999 The complete mitochondrialDNA sequences of Nephroselmis olivacea and Pedinomonasminor two radically different evolutionary patterns withingreen algae Plant Cell 11 1717ndash1729
Unden G amp Bongaerts J 1997 Alternative respiratory path-ways of Escherichia coli energetics and transcriptional regu-lation in response to electron acceptors Biochim BiophysActa Bioenerg 1320 217ndash234
Unseld M Marienfeld J R Brandt P amp Brennicke A1997 The mitochondrial genome of Arabidopsis thaliana con-tains 57 genes in 366 924 nucleotides Nature Genet 1557ndash61
Van der Giezen M Slotboom D J Horner D S DyalP L Harding M Xue G P Embley T M amp KunjiE R S 2002 Conserved properties of hydrogenosomal andmitochondrial ADPATP carriers a common origin for bothorganelles EMBO J 21 572ndash579
Von Heijne G 1986 Why mitochondria need a genome FEBSLett 198 1ndash4
Westphal S Soll J amp Vothknecht U C 2001 A vesicle trans-port system inside chloroplasts FEBS Lett 506 257ndash261
Whitehouse H L K 1969 Towards an understanding of themechanism of heredity London Edward Arnold Ltd
Williams B A P Hirt R P Lucocq J M amp Embley T M2002 A mitochondrial remnant in the microsporidian Trachi-pleistophora hominis Nature 418 865ndash869
Wilson R J (and 10 others) 1996a Complete gene map of theplastid-like DNA of the malaria parasite Plasmodium falcipa-rum J Mol Biol 261 155ndash172
Wilson S B Davidson G S Thomson L M amp PearsonC K 1996b Redox control of RNA synthesis in potato mito-chondria Eur J Biochem 242 81ndash85
Winkler H H amp Neuhaus H E 1999 Non-mitochondrialATP transport Trends Biochem Sci 24 64ndash68
Wolfe K H Morden C W amp Palmer J D 1992 Small sin-gle-copy region of plastid DNA in the non-photosyntheticangiosperm Epifagus virginiana contains only two genes Dif-ferences among dicots monocots and bryophytes in geneorganization at a non-bioenergetic locus J Mol Biol 22395ndash104
Wollman F A Minai L amp Nechushtai R 1999 The biogen-esis and assembly of photosynthetic proteins in thylakoidmembranes Biochim Biophys Acta Bioenerg 1411 21ndash85
Yeliseev A A amp Kaplan S 2000 TspO of Rhodobactersphaeroides a structural and functional model for the mam-
Genomes in mitochondria and chloroplasts J F Allen 37
malian peripheral benzodiazepine receptor J Biol Chem275 5657ndash5667
Zerges W 2002 Does complexity constrain organelle evol-ution Trends Plant Sci 7 175ndash182
Zhang Z D Green B R amp Cavalier-Smith T 1999 Singlegene circles in dinoflagellate chloroplast genomes Nature400 155ndash159
Zhang Z D Cavalier-Smith T amp Green B R 2001 A fam-ily of selfish minicircular chromosomes with jumbled chloro-plast gene fragments from a dinoflagellate Mol Biol Evol18 1558ndash1565
Zouni A Witt H T Kern J Fromme P Krauss NSaenger W amp Orth P 2001 Crystal structure of photosys-tem II from Synechococcus elongatus at 38 acngstrom resol-ution Nature 409 739ndash743
DiscussionD S Horner (Dipartimento di Fisiologia e Biochimica
Generali University of Milan Milan Italy) Given thatthere are components of these membrane-bound energy-transduction pathways that are consistently nuclear enco-ded not the key ones you pointed out but others do youexpect perhaps a slower perhaps a more crude but yet aredox communication between the organelle andexpression of nuclear-encoded genes
J F Allen There is and there is no question about itI think that Bogoradrsquos lock-in hypothesis has somethingto say here and there are other people who say it is all todo with assembling multi-subunit complexes If you havecontrol over the initial construction of the core aroundwhich the whole complex is eventually built then redoxcontrol or some sort of control over synthesis of that corewill control construction of the whole thing eventuallyand I think that may well be largely correct I think it ispossible in principle for nuclear genes to make precursorsaccording to redox instructions but it is not really soimportant that those precursors are here and now presentin exactly the right quantities because the precursors canto a limited extent hang around and wait Actually thereis proteolytic degradation of unattached precursors and soon but I think the fact that these chloroplast and mito-chondrially DNA-encoded subunits are at the core ofthese energy-transducing complexes is the crucial issue
T Cavalier-Smith (Department of Zoology University ofOxford Oxford UK ) In the case of mitochondria the sub-set of genes that remains is much more variable than youimplied It runs from three proteins in the minimal casesup to I think over 90 That is a big difference and manyof the genes in different lineages of protists that remainhave nothing to do with redox control they are things likeribosomal proteins and so on So I think that even thoughwhat you say about redox control of certain genes mayactually be an important thing it is not the fundamentaloverall reason why the genomes are maintained It may bepart of the explanation for why certain genes are alwaysincluded in that subset but even in mitochondria I thinkthat cytochrome b is the only one that is always presentSo I think that both this great diversity of what has beenretained in different lineages and the nucleomorph caseindicate that evolution was able to do one thing in onelineage 500 million years ago and never got around todoing it for 500 million years in another lineage so thereis a lot more haphazardness and evolutionary accidentsand contingency than a straight functional interpretation
Phil Trans R Soc Lond B (2003)
there is a lot of just sheer lsquohappenedndashorndashdidnrsquotndashhappenrsquoor carryover
J F Allen I was not implying anything about the rangeof mitochondrial genomes I have a limited amount oftime so I just talked about what I could talk about withinthat limit Of course there will be a range and this willdepend upon the physiology and bioenergetics of theparticular organism But what I am trying to do is to seewhat the common properties of the irreducible core areof components that are always encoded in the organellesand cytochrome b is a perfect example because that reallyis the thing that binds these two all-important quinoneswithout which there is no Q cycle without which there isno proton pumping and bioenergetics and in Plasmodiumwhere the mitochondrion is reduced to this bare mini-mum there is still a mitochondrial gene for cytochromeb Now my argument about the ribosomal proteins andall the rest of it is that in order to be able to exert thisall-important redox control over gene expression of certaincore components you need an in situ genetic system andthis can vary in composition for reasons that I cannotdescribe Maybe you need RNA polymerase or maybeyou can import RNA polymerase In plastids you haveboth of these options You are going to need ribosomestransfer RNAs and the rest of the apparatus of geneexpression and that needs to be in situ but we are stillleft with the question why does there have to be any gen-etic system at all The mature tobacco or spinach chloro-plast seems to have an entire genetic system just to makeone protein D1 which is all it ever needs to resynthesizeI think that this is a clue It is worth having this very unec-onomical crazy system because there is an overridingrequirement to connect electron transfer with geneexpression directly and unambiguouslymdashwithout recourseto a bureaucracy if you like which is what I think thenucleus is
K Willison (Institute of Cancer Research Chester BeattyLaboratories London UK ) These photosynthetic reactioncomplexes are a sort of baroque mixture of polypeptidechains and cofactors and my understanding is that someof them in high photon flux can turn over within say 30minutes so I am wondering if there is a correlationbetween gene retention and the turnover rates of some ofthe components and the complexes it seems to me thatthat could really be a limiting factor and the reason thatthe proteins had to be encoded locally in order to get thecomplexes to be assembled rapidly
J F Allen I think that is right You can only alter thesteady-state concentration of a protein complex by affect-ing the synthesis and breakdown and therefore turnoverThere is an extreme example the D1 protein encoded bythe psbA gene of chloroplasts which is always in the chlor-oplast genome without exception High light will tend todegrade this protein and when the high light goes awaythere is an absolute need very quickly to bring it back upagain I think that is a very good example Whether thisargument applies to genes such as cytochrome oxidase Ido not know
C J Leaver (Department of Plant Sciences University ofOxford Oxford UK ) Your experiments tend to relatemainly to the gene expression side What about proteinRNA turnover the whole concept of redox control ofthatmdashdo we know anything
38 J F Allen Genomes in mitochondria and chloroplasts
J F Allen Well quite a lot is known about chloro-plasts There is redox control of messenger RNA stabilityfor this same D1 protein which has been described byStephen Mayfield and his co-workers The evidence is thatthe site of redox control is ferredoxin via thioredoxin andthe implication is that thioredoxin tells whether non-cyclicelectron transport is working at all When it is workingthere is a signal that says lsquoOK now continue with all theprotein synthesisrsquo It is not particularly specific but moreof a global effect I think this is what Mayfield argues butof course this is not inconsistent with my argument I haveconcentrated on quinone pools and medium-potentialredox regulatory control because that is where I can seesignals about the dangerous consequences of getting itwrong but there is control of other levels of geneexpression In mitochondria I am not so sure I wouldbounce the question back to you on that one
W Martin (Institute of Botany III Heinrich-Heine Univ-ersitat Dusseldorf Dusseldorf Germany) The nucleomorphgenome was cited as evidence in conflict with your modelbut is that true The nucleomorph genome is not in abioenergetic organelle in that sense It resides in the cyto-sol so does the size of the nucleomorph genome workagainst your model Is it really evidence against yourmodel
J F Allen I do not know why should it be evidenceI cannot explain why the nucleomorph has a genomeThat is why I clutched at the straw that it is storage fornucleic acid precursors
Phil Trans R Soc Lond B (2003)
T Cavalier-Smith The reason I introduced the nucleo-morph is not to say it is direct evidence against yourhypothesis but to show that there probably was no singlefunctional reason for the retention of every gene in everyorganelle It may be that there are specific physiologicalreasons for the retention of certain genes and thereforeyou might be right about a subset of genes but I verymuch doubt if there are specific physiological reasons forthe retention of every single gene that is retained differen-tially because odd things happen in evolution Somethings are easy and sometimes things are difficult Evol-ution is history
J F Allen Of course I agree with that I do not thinkI can explain everything but I do think I can explainsomething If I have done that then I am content
GLOSSARY
AAC ADPndashATP carrierCES control by epistasy of synthesisCORR co-location for redox regulationLHC light-harvesting complexPCC Pasteur culture collectionORF open reading frameRUBISCO ribulose-15-bisphosphate carboxylasendashoxygenaseTAT twin-arginine translocase
Visit httpwwwpubsroyalsocacuk to see an electronic appendix tothis paper

Genomes in mitochondria and chloroplasts J F Allen 27
and Porphyra have plastids that encode more proteins thanthose of most chlorophytes (Stoebe et al 1998 Turmel etal 1999) and these proteins include two-component sig-nal transduction proteins prr and ycf27 (Stoebe et al1998) Extreme or extremely rapid environmental fluctu-ations might be catered for by amplification of signals thatare transmitted by signalling pathways whose componentsare under their own control It is not known whether ycf27and prr are in operons whose promoters their gene pro-ducts control It is also uncertain whether ycf27 is a redoxsignalling component and its distribution at least in itsplastidic location seems to be correlated with the presenceof a phycobilin-based light-harvesting system as found inred algae and cyanobacteria (Ashby amp Mullineaux1999ab Ashby et al 2002) The best guess of the originalstatement of the CORR hypothesis (Allen 1992 1993ac)was that bacterial two-component systems (Iuchi amp Lin1992 Allen 1993c Grebe amp Stock 1999) provide a plaus-ible means of exerting redox regulatory control over mito-chondrial and chloroplast gene expression andsubsequent research confirms the presence of two-compo-nent genes in genomes of Arabidopsis and yeast SeveralArabidopsis two-component genes are predicted to be tar-geted to mitochondria and chloroplasts (Forsberg et al2001) These must be considered as prime candidates forcomponents involved in redox regulation of chloroplasttranscription seen in the experiments of Pfannschmidt etal (1999a 2001a) The presumably homologous regulat-ory process serving to control photosystem stoichiometryin cyanobacteria the prr system (Li amp Sherman 2000) isnow characterized as a two-component system involvinga redox sensor and redox response regulator as originallydefined (Allen 1992 1993ac)
4 CONTRASTING HYPOTHESES CONCERNINGTHE RETENTION OF CYTOPLASMIC GENOMES
CORR is a recent addition to explanations that havebeen put forward to account for the persistence of gen-omes in organelles These models and their underlyingmechanisms are now discussed and compared
(a) There is no reason that is just how it isOne reaction to the problem of cytoplasmic genetic sys-
tems (sect 2) is that there is no problem and no particularreason for their retention However there is a very widephylogenetic distribution of a common core or primarysubset of organellar-encoded genes and one which haswithstood considerable genomic reorganization over evol-utionary time This constancy is seen for example in thesame genes being present in the chloroplasts of landplants with multiple copies of a single circular chloroplastchromosome and in dinoflagellate chloroplasts with acollection of different DNA minicircles each with essen-tially one gene (Zhang et al 1999 2001 Barbrook et al2001) Another suggested explanation of the retention ofmitochondrial and chloroplast genomes is simply torestate the endosymbiont hypothesis and to point out thatchloroplast and mitochondrial genes lsquocame inrsquo with thebacterial endosymbiont This suggestion however doesnot address the question of why some genes were retainedwhereas others were not It is also now clear that mostgenes that came in and still survive have nuclear and cyto-
Phil Trans R Soc Lond B (2003)
solic gene products whereas many chloroplast and mito-chondria proteins had a eukaryotic origin (Salzberg et al2001 Martin amp Russell 2003) The odds against a con-served pattern in gene location (eg table 1) arising bychance must be astronomical Thus there is a reason forthe retention of organellar genomes and a reason for theretention of the same primary subset of genes Becauseevolution proceeds by natural selection of chance vari-ation lsquoreasonrsquo in this context requires identification of thefunction of the location of a gene that is the selectivevalue of the phenotype produced by that location Evol-ution sometimes retains apparently functionless relics ofpreviously useful features but not for long especially ifthere is a cost
(b) The lock-in hypothesisThe lsquolock-inrsquo hypothesis of Bogorad (1975) was pro-
posed independently of the endosymbiont hypothesis anddoes not depend upon it According to the lock-in hypoth-esis core components of multi-subunit complexes must besynthesized de novo in the correct cellular compartmentotherwise photosystems or respiratory chain complexesfor example might assemble in the plasma membranevacuolar membrane or endoplasmic reticulum The lock-in hypothesis made no clear prediction about the conti-nuity of cytoplasmic genetic systems even though cyto-plasmic uniparental non-Mendelian inheritance for bothmitochondria (Roodyn amp Wilkie 1968) and plastids(Kirk amp Tilney-Bassett 1978) made it likely that organellarDNA the chondriome and the plastome replicated inde-pendently of nuclear DNA (Whitehouse 1969)
Mechanisms of protein import and targeting (Schatz1998 Koehler et al 1999 Leuenberger et al 1999 Gab-riel et al 2001 Jarvis amp Soll 2001 Pfanner amp Geissler2001 Duby amp Boutry 2002 Pfanner amp Wiedemann 2002)now effectively answer the problem of specifying the cor-rect locations for membrane protein complexes for whichthe lock-in hypothesis was proposed as a solution How-ever regulation of synthesis of an entire functional multi-subunit complex by control of synthesis of individual sub-units is still an important principle This seems to be afeature of the lsquoCESrsquo model (control by epistasy ofsynthesis) of Wollman (Wollman et al 1999 Eberhard etal 2002) which envisages nuclear control as all-embracingwith organellar genetic systems doing as it were whatthey are told In the hypothesis of CORR propoundedhere CES is equally important but the essence of CORRis that the organellar genetic system alone knows what arethe immediate and most important signals of environmen-tal change The organelle may keep the nucleus informedbut this is a lower priority than initiating an appropriateresponse CES and CORR agree that assembly of multi-subunit complexes is a complex business and requirescoordinated control in which no step is rate-limiting butmany may be obligatory
(c) Going going gone the evolutionary process oftransfer of genes from organelles to the
nucleus is still underway but incompleteThis hypothesis envisages that for most eukaryotes we
are now witnessing the last stages in a slow evolutionarytransfer of genes to the nucleus and this transfer may beexpected to proceed to completion in what is in evol-
28 J F Allen Genomes in mitochondria and chloroplasts
utionary terms the near future (Palmer 1997 Gray 19992000 Gray et al 1999 Adams et al 2000 Daley et al2002) A clear and compelling statement of this widelyheld position is that of Herrmann amp Westhoff (2001 p10)
hellipthe transfer of geneshelliptells that the partite plant gen-ome is not in a phylogenetic equilibrium All availabledata suggest that the ultimate aim of genome restructur-ing in the plant cell as in the eukaryotic cell in generalis the elimination of genome compartmentation whileretaining physiological compartmentation
One serious problem with this hypothesis is the dis-tinctly non-random sample of genes that remain to betransferred A relatively constant subset of genes is foundin bioenergetically functional mitochondria from fungi tomammals and angiosperms as in photosynthetic plastids(chloroplasts) from unicellular algae to angiosperms Thishypothesis therefore requires an additional ad hocassumption that some gene transfers are inherently moreprobable than others (this too is widely supposed seelater) and thus that gene transfer events proceed in thesame sequence in quite independent lineages Thereseems to be no evidence for this and the order of geneloss from mitochondria and chloroplasts has been used toinfer phylogeny (Martin et al 1998 Adams et al 2000Millen et al 2001) lsquoGoing going gonersquo (Palmer 1997)is difficult to refute categorically but it does not seem tobe consistent with recent evidence For example chromo-some two of Arabidopsis thaliana (Lin et al 1999) has asingle insertion encompassing with about 99 sequencesimilarity the complete Arabidopsis mitochondrial genome(Unseld et al 1997 Marienfeld et al 1999 Stupar et al2001) Unless this single transfer is a freak occurrencethis finding strongly indicates that gene transfer occurswith relative ease (Adams et al 2000 Henze amp Martin2001 Rujan amp Martin 2001 Salzberg et al 2001) Inaddition there is experimental evidence that randomlygenerated polypeptides are mitochondrially imported athigh frequency (Lemire et al 1989) The hypothesis thatwe now see the same few stragglers in diverse organellargenomes is not impossible but should be regarded as alast resort perhaps applicable only in extreme cases suchas the greatly reduced apicomplexan plastomes and chon-driomes Even there the hypothesis carries with it an obli-gation to explain why organisms with few organellar geneshave lsquoevolvedrsquo faster than those with many
(d ) The frozen accidentWhat may be described as lsquothe frozen accident hypoth-
esisrsquo states that the evolutionary process of gene transferfrom organelle to nucleus was underway when somethinghappened that stopped it One example of the frozen acci-dent is a scenario (Von Heijne 1986) in which import intomitochondria of the precursor proteins arising from newlytranslocated genes proceeded for long enough after theprimordial endosymbiosis for most of the symbiont-derived genes to be moved to the cell nucleus The lsquoacci-dentrsquo that halted the process was the evolutionary originof exocytosis and protein secretion after which it becameimpossible for precursors properly destined for mitochon-drial import to be distinguished from those destined forexport from the cell As with the lsquolock-inrsquo hypothesis the
Phil Trans R Soc Lond B (2003)
lsquofrozen accidentrsquo seems incompatible with the remarkableprecision and specificity of protein targeting (Schatz 1998Koehler et al 1999 Leuenberger et al 1999 Gabriel etal 2001 Jarvis amp Soll 2001 Pfanner amp Geissler 2001Duby amp Boutry 2002 Pfanner amp Wiedemann 2002) Pro-tein sorting mechanisms do not seem to forbid controlledimport into any cellular compartment Another lsquoaccidentrsquothat may have impeded subsequent gene transfer in thecase of mitochondria is their departure from what is other-wise a universal genetic code Genetic code differences arediscussed later in the context of whether some genes can-not be moved
(e) Some proteins (with cofactors) cannot beimported
We may describe as the lsquounimportability hypothesisrsquo theproposal that some proteins are inherently unimportableand must therefore be synthesized de novo in the correctcompartment within the cell The unimportability hypoth-esis is most often proposed as its special case the hydro-phobicity hypothesis with hydrophobicity being thereason for unimportability
(i) HydrophobicityA commonly entertained notion is that proteins that are
encoded and synthesized within organelles comprise a pri-mary subset characterized by shared hydrophobicity allare intrinsic membrane proteins (Von Heijne 1986Popot amp de Vitry 1990 Claros et al 1995) As with theCORR hypothesis favoured here the primary subset thenrequires a secondary subset of genetic system componentsthe latter not themselves being hydrophobic but beingrequired for synthesis of the others Certainly the mito-chondrial inner membrane contains some intrinsic proteincomplexes whose subunits broadly satisfy the criterion ofhydrophobicity However the core subunits that are mito-chondrially encoded are also explained by the CORRhypothesis This would make hydrophobicity a require-ment for their function in vectorial electron and protontransfer and not a reason in itself for retention of theirgenes in the cytoplasm
The hydrophobicity hypothesis has some very clearcounter-examples in chloroplasts and cannot thereforestand if one wishes for a single principle that embracesboth types of bioenergetic organelle One clear counter-example is the large (L) subunit of the enzyme RUBISCOan abundant but entirely membrane-extrinsic protein ofca 55 kDa molecular mass The RUBISCO large subunitis generally chloroplast-encoded with the exception of thenuclearly encoded bacterial-type L subunit of L2 RUBI-SCO in peridinin-containing dinoflagellates (Morse et al1995) In most photosynthetic eukaryotes RUBISCO hasa chloroplast encoded large subunit and a nuclearly enco-ded small (S) subunit (Ellis 1984 Gatenby amp Ellis 1990)giving an L8S8 holoenzyme and this RUBISCO is widelytaken as a paradigm for the need for coordination ofnuclear and chloroplast gene expression (Rodermel 2001)
By complete contrast to the RUBISCO large subunitwhich is a chloroplast-encoded and water-soluble proteina second major violation of the hydrophobicity hypothesisis the existence of the universally nuclearly encoded butalways hydrophobic subunits of the chloroplast LHC IIand LHC I (Chitnis amp Thornber 1988) These apopro-
Genomes in mitochondria and chloroplasts J F Allen 29
teins of ca 27ndash30 kDa are quite water-insoluble and areusually fully extracted only in chloroformndashmethanol or inthe presence of detergents The Lhc gene family is diverse(Green amp Kuhlbrandt 1995) but most gene productsor Lhcp (light-harvesting chlorophyll protein) polypeptideshave three membrane-spanning hydrophobic a-helices(Kuhlbrandt 1994) It is interesting that three-helix Lhcproteins seem to be absent from extant cyanobacteria andchloroxybacteria (Green amp Kuhlbrandt 1995)
(ii) The five-helix rulePerhaps with the three-helix model of LHC II in mind
a variation of the hydrophobicity hypothesis is the empiri-cal lsquofive-helix rulersquo (Popot amp de Vitry 1990 Claros et al1995 Wollman et al 1999) which states that no intrinsicmembrane protein with five or more membrane-spanninghelices can be encoded in the nucleus Five is a good guessfor a threshold number of helices demanding organellar-encoding in the context of photosynthetic reaction centres(Heathcote et al 2002) where the earliest known struc-ture that of type II centres (Deisenhofer et al 1985) asin photosystem II (Zouni et al 2001) shows exactly fivehelices with type I centres containing 11 (Jordan et al2001) There are however many nuclearly encoded sub-units in both mitochondria and chloroplasts that havefewer than five membrane helices so the five-helix rulecan only describe a sufficient condition and not a neces-sary condition for nuclear encoding
It should also be noted that ADPndashATP carriers (AACs)are encoded in the nucleus AACs of the mitochondrialinner membrane have six transmembrane helices (van derGiezen et al 2002) while the those of the chloroplast innerenvelope (Winkler amp Neuhaus 1999) have 12 (Tjaden etal 1998)
With the counter-examples of nuclearly encoded AACswith six or 12 membrane-spanning helices many intrinsicmembrane proteins with fewer than five helices somenuclearly encoded and some not and with the RUBISCOlarge subunit as a major product of chloroplast geneexpression with no membrane domain at all the five-helixrule begins to seem rather like a case of special pleadingEven if there are indeed special mitigating circumstancesfor AACs (this argument is outlined later) the five-helixrule as with the hydrophobicity hypothesis of which it isa special case will disappear if it can be shown that pos-session of five or more membrane helices is required forsome other function such as that of CORR that deter-mines the organellar location of the gene concernedHydrophobicity is in some cases a feature of proteins thatare encoded in organelles but it is not in itself the issue
For chloroplasts the hydrophobicity hypothesis predictsretention of cyanobacterial genes in functional categoriesassociated with the chloroplast envelope and with trans-port just as much as with photosynthesis Table 1 showsthat this is not the case
A mechanism for the unimportability of hydrophobicproteins is rarely stated explicitly and it is rather left tothe imagination whether hydrophobic proteins might asit were get stuck halfway through the chloroplast envelopeor mitochondrial outer membrane whether transport can-not get started on a precursor protein that has alreadyspontaneously inserted into the membrane or whetherhydrophobic proteins will be unable to make the journey
Phil Trans R Soc Lond B (2003)
through the cytosol without precipitating there In thecytosol co-translational transport or membrane insertionwould in principle address the latter point but it seemsto be generally true that chloroplast and mitochondrialproteins are post-translationally inserted into the organel-lar outer membrane So the obvious ancillary questionabout unimportability is lsquowhy then cannot hydrophobicproteins be co-translationally imported into organellesrsquoThere is evidence in yeast for co-translational transportinto mitochondria of proteins thought to be derived fromthe original endosymbiont (Marc et al 2002) If there isany truth in hydrophobicity as a barrier to protein importthe CORR principle of unselective import (sect 3c) shouldbe easily refuted for example by a demonstration that noknown targeting presequence is capable of importing poly-phenylalanine encoded by polyU for in vitro import stud-ies
An additional proposal intended to account for theunimportability of some hydrophobic proteins and forimportability of others is suggested by A de Grey(personal communication) who argues that bacteriallyderived protein precursors imported into mitochondriacross the mitochondrial inner membrane completelybefore insertion into that membrane from the mitochon-drial matrix (the N-phase) This is held to be an obstacleto successful import of membrane-intrinsic proteins thatmight en route discharge the proton motive force By con-trast eukaryotically derived proteins can be inserted intothe mitochondrial inner membrane from the inter-mem-brane space (the P-phase) and then there is for somereason no similar uncoupling effect Thus adenylatetransporters or AACs can be nuclearly encoded whereascytochrome oxidase subunits for example cannot It isnot obvious why bacterially derived subunits such asthose of cytochrome oxidase did not acquire eukaryoticinsertion mechanisms Perhaps this acquisition is forbid-den by the need to conserve pre-existing routes ofassembly into multi-subunit complexes or perhaps theneed for insertion from the inside reflects a need to incor-porate a cofactor for example haem or ironndashsulphur clus-ters along the way
(iii) Unimportability of three dimensional structures andcofactors
Accumulating evidence on protein transport mech-anisms seems to make a specific obstacle to successfultransport of certain sorts of protein less and less plausibleespecially given the observation that precursors with N-terminal targeting sequences not mature functional poly-peptides themselves traverse membranes during biosynth-esis
Protein folding inside the living cell is invariably assistedby molecular chaperones (Gatenby amp Ellis 1990 Azem etal 1997 Komiya et al 1997 1990 Jackson-Constan et al2001) proteins that impede the formation of lsquoimproperrsquostructural interactions that will otherwise produce aggre-gation or other non-functional interactions Proteinimport mechanisms appear mostly to rely on specific mol-ecular chaperones for guided unfolding before membraneinsertion and refolding after translocation of the polypep-tide into its destined compartment (Gatenby amp Ellis 1990Azem et al 1997 Komiya et al 1997 Jackson-Constanet al 2001) Because polypeptides are mostly lsquothreadedrsquo
30 J F Allen Genomes in mitochondria and chloroplasts
through membranes no special constraint on transportshould be expected to arise from 3D structure or surfaceproperties of a protein
An exception to this unfolding rule may be providedby holoproteins containing cofactors that are transportedalong with the apoprotein presumably with a largely intact3D structure The lsquoTATrsquo (twin-arginine translocase) sys-tem (Bogsch et al 1998 Dalbey amp Robinson 1999 Berkset al 2000ab Robinson 2000 Robinson amp Bolhuis 2001Sargent et al 2002) works broadly along these linesthough its mechanism is unresolved TAT is relevant herebecause it adds transport of essentially complete holopro-teins to the cellrsquos repertoire of transport mechanisms andeven proteins that may not be allowed to unfold perhapsbecause of the toxicity of the free cofactor are not therebymade candidates for an unimportability rule One variantof the unimportability hypothesis is that transport canwork only by unfolding and dissociated cofactors may betoxic to the cell The bacterially derived TAT system(Bogsch et al 1998 Dalbey amp Robinson 1999 Berks etal 2000ab Sargent et al 2002) overcomes this argumentas would examples of cofactor-free polypeptides such as(C)Fo subunit c which are nevertheless generally encodedin organelles
The presence of a vesicular transport system in chloro-plasts (Westphal et al 2001) counts against the unim-portability hypothesis Vesicular transport between thechloroplast envelope and the thylakoid membrane shouldin principle allow any protein to be ferried across thechloroplast stroma Vesicles might allow transport inparticular of membrane proteins that would discharge theproton motive force if allowed to connect transiently theN and P aqueous phases on each side of a coupling mem-brane
Another observation that seems to count against unim-portability is the increased number of membrane barriersin plastids derived from secondary tertiary or higher-orderendosymbioses These plastids seem to have retained ifanything fewer genes rather than more as unimportabilitypredicts The nucleomorph the residual nucleus of theprimary host retains many genes for molecular chap-erones (Archibald et al 2001 Cavalier-Smith 2002) inagreement with the conclusion that protein import pro-ceeds by conserved and specialized mechanisms If eachnew import added to gain a higher-order endosymbiosisrequires additional targeting presequences then perhapsthere is an advantage in components of an import machin-ery not being imported themselves
It seems that unimportability is less easy an assumptionto make today than in previous years and the onus maynow be on its proponents to cite specific examples of pro-teins that can never be imported into chloroplasts or mito-chondria
(f ) Some genes cannot be moved(i) Differences in the mitochondrial genetic code
The genetic code is the specific relation between ribon-ucleotide triplets of codons and tRNAs that recognizethem and which thus determine which individual aminoacids if any will be incorporated into the growing poly-peptide chain Mitochondria show small departures fromthe otherwise universal genetic code (Attardi amp Schatz1988 Alberts et al 1994) UGA is a lsquostoprsquo codon in the
Phil Trans R Soc Lond B (2003)
universal code and in plant mitochondria but is the codonfor tryptophan in mitochondria of animals and fungiCUA means leucine everywhere except fungal mitochon-dria where it means threonine and AGA and AGU codefor arginine in the lsquouniversalrsquo code but serine in insectmitochondria and lsquostoprsquo in mammalian ones The signifi-cance of this heterogeneity in coding is most unlikely torest in the early origin of mitochondria as the lsquouniversalrsquocode is employed by bacteria and protists Furthermorethe diversity of departures from the lsquouniversalrsquo codeamong mitochondria of different eukaryotic lineages indi-cates they had independent origins A general rule formitochondrial genetic codes is that reliance on the thirdlsquowobblersquo position is decreased which allows all 20 proteinamino acids to be specified by fewer tRNAs (22 in animalmitochondria instead of 31 in the cytosol) This in turnmeans that mtDNA can specify more proteins for a givenlength of DNA or conversely that the mitochondrialgenome can be condensed or lsquostreamlinedrsquo Although thisarrangement does not explain the retention of mitochon-drial genomes and may be a secondary event in mitochon-drial evolution it almost certainly places an extra barrierin place of gene relocation As discussed previously how-ever there seems to be no single barrier that can explainthe retention of a relatively constant subset of mitochon-drial genes
There has clearly been a contraction of mitochondrialDNA especially in animals Animal mitochondrial DNAhas few if any non-coding sequences and animal mito-chondrial genetic codes require fewer tRNAs than the sin-gle code operating in the cytosol (Gray 1999 Gray et al1999 Selosse et al 2001) These may be devices todecrease the total frequency of mutations per mitochon-drial genome assuming a constant frequency per unitlength of mitochondrial DNA It has been proposed thata genersquos removal to the nucleus decreases its susceptibilityto free radical mutagenesis (Raven et al 1994ab Allen ampRaven 1996) If this is the case then the non-standardcode may be a further indication of an overriding need tomaintain certain genes within the mitochondrion What-ever the reason for the retention of genes in organelles ithad better be a good one bioenergetic organelles areclearly the wrong place to keep genetic information andgo to extraordinary lengths to offset the consequences ofit being there
(ii) Molecular refugeesA phenomenon related to differences in the genetic code
is seen in chloroplasts This is a bias in codon usage Thelsquomolecular refugeesrsquo hypothesis (Howe et al 2000) sug-gests that nuclear copies of what were once chloroplastgenes escape constraints on codon usage and thus thatwhat distinguishes chloroplast-encoded proteins is thattheir amino-acid composition is unusual enough for thebias in codon usage not to apply It is difficult to see whatphysico-chemical properties might be uniquely shared bythe amino acids of chloroplast-encoded proteins and inaddition there seem to be no lsquoforbiddenrsquo amino-acid resi-due in chloroplasts As with the departure from the univer-sal genetic code in mitochondria is seems likely that abias in codon usage reflects some secondary property ofchloroplast genomes and is not the primary reason fortheir retention In any case any restriction on gene move-
Genomes in mitochondria and chloroplasts J F Allen 31
ment will act in the wrong direction to explain why chloro-plasts have retained genomes at all provided there is noresulting obstacle to a chloroplast protein being success-fully encoded as a precursor in the nucleus
(iii) Genes are copied and replicated not relocated in onestep
If complete relocation is envisaged as proceeding by thesimultaneous appearance of a gene in one compartmentand its deletion from another then it is possible to imaginethat whatever the vector is there may be selectivity ingene transfer (Henze amp Martin 2001) For example ifviral incorporation replication and recombination in bothorganelle and nucleus were the mechanism of relocationthen successful gene transfer might depend on the abilityof the virus to complete the process in one step This inturn might depend on some property of the gene or ofthe protein it encodes if expression occurs at any stage inthe process of relocation An assumption of the CORRhypothesis favoured here is that genes are not movedphysically but merely copied after which either the copyin the original location or the copy in the new locationmay be lost as a result of selection A useful analogy maybe the action of copying files between computer storagevolumes In this analogy the copy is identical to the orig-inal and either may subsequently be deleted without lossof information
Because any gene that is not usefully expressed willundergo no selection to maintain a functional allele theprevailing location will be the one in which the functionalcopy of the gene resides The problem of which gene sur-vives is then subsumed into the problem of which is theselectively advantageous location for it It seems reason-able to assume that there wasmdashand ismdashan interim periodin which homologous genes in different compartments areessentially in competition to see which functions best Asoutlined above there is no obvious obstacle for copyingDNA sequences within the cell The physico-chemicalproperties of DNA are fairly uniform and not directlydependent on nucleotide sequence though organelles likeintra-cellular parasites seem to prefer A and T to G andC The answer to the question lsquowhich gene location func-tions bestrsquo may have nothing to do with the gene itselfnor directly with the structure and function of its geneproduct Instead the answer may have everything to dowith the required regulatory interaction between the geneproduct and expression of the gene that produces it
(g) CORRThis hypothesis has been propounded earlier (sect 3) and
will be summarized as follows Vectorial electron and pro-ton transfer exerts regulatory control over expression ofgenes encoding proteins directly involved in or affectingredox poise This regulatory coupling is indispensable foradaptation to environmental changes in chemical activitiesof oxygen light and CO2 and requires co-location of thesegenes with their gene products Organelles lsquomake theirown decisionsrsquo on the basis of environmental changesaffecting redox state The hypothesis is abbreviatedCORR for co-location for redox regulation or more com-pletely lsquoco-location (of gene and gene product) for(evolutionary) continuity of redox regulation of geneexpressionrsquo
Phil Trans R Soc Lond B (2003)
5 SUMMARY OF THE CURRENT STATUS OF THEHYPOTHESIS OF CORR
Q Why do mitochondria and chloroplasts have theirown genetic systems
A CORR co-location of genes and gene products per-mits direct and autonomous redox regulation of geneexpression
(a) Objections to CORR(i) There is no regulation of gene expression in chloroplasts
and mitochondriaThis is still commonly stated but incorrect See the prin-
ciple in sect 3d
(ii) There is no regulation of gene expression in chloroplastsand mitochondria at the level of transcription
For chloroplasts there is now clear evidence for a func-tionally intelligible redox control of transcription(Pfannschmidt et al 1999a) This invalidates the previouswidely held assumption that all control of chloroplast geneexpression is post-transcriptional (Gruissem amp Tonkyn1993 GoldschmidtClermont 1998) The evidence formitochondria is less clear but redox control of transcrip-tion certainly cannot be ruled out and there are indi-cations that it may occur (Pearson et al 1993Konstantinov et al 1995 Wilson et al 1996b) It mustalso be restated that transcriptional control is not neces-sary for CORR but serves as a compelling example of aconserved typically bacterial mode of response that oper-ates still in eukaryotic cytoplasmic organelles thus illus-trating the principle of continuity of redox control (sect 3d)Post-transcriptional redox regulation (Danon amp Mayfield1994 Kim amp Mayfield 1997 Brown et al 2001 Treb-itsh amp Danon 2001) is consistent with CORR and thereis nothing to forbid it being a primary mechanism thatsatisfies most or indeed all of the formal requirements ofthe hypothesis in particular cases
(iii) There are no redox signalling components encoded inchloroplast or mitochondrial DNA
This is what CORR predicts See the principle in sect 3j
(iv) There is redox control of nuclear gene expressionThis is correct and many examples are now known
especially for chloroplasts (Escoubas et al 1995 Kovacs etal 2000 Pfannschmidt et al 2001ab Surpin et al 2002)Redox signals from photosynthetic or respiratory electrontransport chains are not at all prohibited by CORR Theessence of CORR is that there is a subset of organellarproteins for which there is an overriding requirement fora redox regulatory control that is initiated within theorganelle and whose effects are exerted by the simplestand most direct route This redox signalling is envisagedto take place without a requirement for prior consultationand coordination with what may be regarded as a nucleo-cytoplasmic bureaucracy
(v) Nuclear genes can be expressed as rapidly as chloroplastand mitochondrial genes
Zerges (2002) for example cites the equal times takenfor complete expression of nuclear and chloroplast genesin Chlamydomonas as evidence against the CORR hypoth-
32 J F Allen Genomes in mitochondria and chloroplasts
esis However the question is not one of the time takento express a gene but the time taken to alter the rate ofits expression in response to a (redox) signal originatingwithin the organelle An equally important additional fac-tor may be the autonomy of the signalling pathway thatis its independence from other inputs as described in theprevious paragraph One might say that mitochondria andchloroplasts require a free hand
(vi) There is no evidence to support this hypothesisThe hypothesis is rich in predictions (sectsect 3 and 6) and
those that have been tested stringently are in agreementwith experimental results None of the predictions of thehypothesis has been found to be incorrect and none of theunderlying principles (sect 3) has been disproved Furtherpredictions will be the subject of future experiments
(b) Evidence for CORR(i) Chloroplasts have selectively retained cyanobacterial genes
for primary events in photosynthesisSee table 1
(ii) There is regulation of gene expression in chloroplastsThere is transcriptional regulation of gene expression
in chloroplasts (sect 3d) The case for mitochondria is notyet proven
(iii) There is redox regulation of gene expression inchloroplasts
There is transcriptional redox regulation of geneexpression in chloroplasts (sect 3d) The case for mitochon-dria is not yet proven though see (Allen et al 1995 Kon-stantinov et al 1995 Galvis et al 1998)
(iv) There are no redox signalling components encoded inchloroplast or mitochondrial DNA
Correct but with possible exceptions discussed in sect 3j
(v) There are lsquobacterialrsquo redox signalling components ineukaryotes
See sect 3j
(vi) Further predictions DNA-array results and phenotypesof allotopic mutants
A systematic test of CORR might be to make an oxid-ized-minus-reduced gene expression profile of the lsquotran-scriptomersquo of a cyanobacterium to see if the set of geneswhose transcription is under most rapid and direct redoxcontrol corresponds to the genes of the chloroplast gen-ome Laboratories with this experimental capability do notseem to have addressed this possibility (Hihara et al 2002Ikeuchi 2002) Likewise if a putative modern representa-tive of the proto-mitochondrion can be found and agreedeg Rhodospirillum rubrum or Paracoccus denitrificans thena similar experiment should reveal a subset of genes corre-sponding to mitochondrial genomes DNA-microarraysalso present the possibility of testing the relative rate ofredox response and dependency on other signallinginputs of transcription of eukaryotic genes To dateorganellar genes have not been included in such investi-gations (Ferea et al 1999) A further experimental possi-bility is to examine the redox regulatory phenotypes ofexisting allotopic mutants as outlined in sect 3e
Phil Trans R Soc Lond B (2003)
6 CONCLUSION AND FUTURE PROSPECTS
The CORR hypothesis states that the function of mito-chondrial and chloroplast genomes is to provide co-location (of gene and gene product) for (evolutionary)continuity of redox regulation of gene expression Thishypothesis is neither self-contradictory nor devoid of test-able predictions It is also consistent with available evi-dence and in principle makes sense of the otherwisepuzzling distribution of genes between compartments ineukaryotic cells The explanation that CORR offersapplies equally to mitochondria and chloroplasts andidentifies selective forces that still operate to determine thelocation of genes No selective barriers need be assumedto gene transfer protein import nor membrane insertionNor do we need to invoke a lsquofrozen accidentrsquo that some-thing happened to stop gene transfer for some reason atsome time in the remote and inaccessible past CORR alsoexplains the absence of genomes from organelles that havelost their primary bioenergetic functions CORR may actas a stimulus to a search for residual bioenergetic functionin those organelles that have lost aerobic respiration andphotosynthesis but where residual genomes remain
CORR also predicts regulatory properties of knowncomponents of chloroplasts and mitochondria that in turnindicate important but as yet uncharacterized flexibility inenergy transduction These components include the RUB-ISCO large subunit and the CFo and Fo subunits of coup-ling ATPase in both chloroplasts and mitochondriaRedox regulation of the relative stoichiometry of compo-nents as seen in regulation of chloroplast photosystemstoichiometry (Pfannschmidt et al 1999a) may extend to(C)Fo-(C)F1-ATPase indicating that the function ofretention of genes for one or more of these componentsis flexibility in the stoichiometries of H1 to e2 and H1 toATP in chemiosmotic coupling (Schemidt et al 1998Stock et al 2000 Allen 2002)
So far explicit tests of CORR have largely been con-fined to chloroplasts and to some extent mitochondria ofhigher plants but this merely reflects the expertise andinterests of the investigators Further attempted disproofof CORR is welcomed and encouraged Novel experi-mental tests can be devised as well as attempted refu-tation through analysis of existing data It is hoped thatexplicit attention will be given to the CORR hypothesis inlaboratories already suitably equipped and that resourcescan be allocated for this purpose in laboratories wherethere is the interest but not currently the technology orexpertise
The heuristic value of the hypothesis already seen inthe role of redox-regulated transcription of photosyntheticreaction centre genes in chloroplasts may be its most use-ful property The central proposal is that mitochondriaand chloroplasts contain genes for proteins whose functionin electron transfer demands rapid direct and uncon-ditional redox regulatory control of their biosynthesis Thefull implications of this hypothesis are very broad and onlyjust beginning to be explored
I thank Carol Allen Aubrey de Grey William Martin John ARaven and Francis-Andre Wollman for discussion and corre-spondence during the preparation of this paper and WilliamMartin for suggestions including table 1 Research grants from
Genomes in mitochondria and chloroplasts J F Allen 33
The Swedish Natural Sciences Research Council (NFR) andthe Crafoord Foundation are gratefully acknowledged
REFERENCES
Adams K L Daley D O Qiu Y L Whelan J amp PalmerJ D 2000 Repeated recent and diverse transfers of a mito-chondrial gene to the nucleus in flowering plants Nature408 354ndash357
Alberts B Bray D Lewis J Raff M Roberts K amp Wat-son J D 1994 The molecular biology of the cell New Yorkand London Garland Publishing
Allen C A Hakansson G amp Allen J F 1995 Redox con-ditions specify the proteins synthesized by isolated chloro-plasts and mitochondria Redox Rep 1 119ndash123
Allen J F 1992 Protein phosphorylation in regulation ofphotosynthesis Biochim Biophys Acta 1098 275ndash335
Allen J F 1993a Control of gene expression by redox poten-tial and the requirement for chloroplast and mitochondrialgenomes J Theor Biol 165 609ndash631
Allen J F 1993b Redox control of gene expression and thefunction of chloroplast genomes a hypothesis PhotosynthRes 36 95ndash102
Allen J F 1993c Redox control of transcription sensorsresponse regulators activators and repressors FEBS Lett332 203ndash207
Allen J F 1995 Thylakoid proteinndashphosphorylation state-1ndashstate-2 transitions and photosystem stoichiometry adjust-ment redox control at multiple levels of gene expressionPhysiol Plant 93 196ndash205
Allen J F 1996 Separate sexes and the mitochondrial theoryof ageing J Theor Biol 180 135ndash140
Allen J F 2002 Photosynthesis of ATP-electrons protonpumps rotors and poise Cell 110 273ndash276
Allen J F amp Allen C A 1999 A mitochondrial model forpremature ageing of somatically cloned mammals IUBMBLife 48 369ndash372
Allen J F amp Nilsson A 1997 Redox signalling and the struc-tural basis of regulation of photosynthesis by protein phos-phorylation Physiol Plant 100 863ndash868
Allen J F amp Pfannschmidt T 2000 Balancing the two photo-systems photosynthetic electron transfer governs transcrip-tion of reaction centre genes in chloroplasts Phil TransR Soc Lond B 355 1351ndash1357 (DOI 101098rstb20000697)
Allen J F amp Race H L 2002 Will the real LHC II kinaseplease step forward Sciencersquos STKE httpwwwstkeorgcgicontentfullsigtrans2002155pe43
Allen J F amp Raven J A 1996 Free-radical-induced mutationvs redox regulation costs and benefits of genes in organellesJ Mol Evol 42 482ndash492
Archibald J M Cavalier-Smith T Maier U amp Douglas S2001 Molecular chaperones encoded by a reduced nucleusthe cryptomonad nucleomorph J Mol Evol 52 490ndash501
Ashby M K amp Mullineaux C W 1999a Cyanobacterialycf27 gene products regulate energy transfer from phycobili-somes to photosystems I and II FEMS Microbiol Lett 181253ndash260
Ashby M K amp Mullineaux C W 1999b The role of ApcDand ApcF in energy transfer from phycobilisomes to PSI andPSII in a cyanobacterium Photosynth Res 61 169ndash179
Ashby M K Houmard J amp Mullineaux C W 2002 Theycf27 genes from cyanobacteria and eukaryotic algae distri-bution and implications for chloroplast evolution FEMSMicrobiol Lett 214 25ndash30
Attardi G amp Schatz G 1988 Biogenesis of mitochondria ARev Cell Biol 4 289ndash333
Azem A Oppliger W Lustig A Jeno P Feifel B SchatzG amp Horst M 1997 The mitochondrial hsp70 chaperone
Phil Trans R Soc Lond B (2003)
system effect of adenine nucleotides peptide substrate andmGrpE on the oligomeric state of mhsp70 J Biol Chem272 20 901ndash20 906
Barbrook A C Symington H Nisbet R E R LarkumA amp Howe C J 2001 Organisation and expression of theplastid genome of the dinoflagellate Amphidinium opercula-tum Mol Genet Genomics 266 632ndash638
Bauer C E Elsen S amp Bird T H 1999 Mechanisms forredox control of gene expression A Rev Microbiol 53495ndash523
Bauer C E Elsen S Swem L R Swem D L amp MasudaS 2003 Redox and light regulation of gene expression inphotosynthetic prokaryotes Phil Trans R Soc Lond B 358147ndash154 (DOI 101098rstb20021189)
Berks B C Sargent F De Leeuw E Hinsley A P Stan-ley N R Jack R L Buchanan G amp Palmer T 2000aA novel protein transport system involved in the biogenesisof bacterial electron transfer chains Biochim Biophys ActaBioenerg 1459 325ndash330
Berks B C Sargent F amp Palmer T 2000b The tat proteinexport pathway Mol Microbiol 35 260ndash274
Blackstone N W 1999 Redox control in development andevolution evidence from colonial hydroids J Exp Biol 2023541ndash3553
Blackstone N W 2000 Redox control and the evolution ofmulticellularity Bioessays 22 947ndash953
Blackstone N W 2001 Redox state reactive oxygen speciesand adaptive growth in colonial hydroids J Exp Biol 2041845ndash1853
Bogorad L 1975 Evolution of organelles and eukaryotic gen-omes Science 188 891ndash898
Bogsch E G Sargent F Stanley N R Berks B C Robin-son C amp Palmer T 1998 An essential component of a novelbacterial protein export system with homologues in plastidsand mitochondria J Biol Chem 273 18 003ndash18 006
Brown E C Somanchi A amp Mayfield S P 2001 Interor-ganellar crosstalk new perspectives on signaling from thechloroplast to the nucleus Genome Biol 2 10211ndash10214
Cavalier-Smith T 2002 Chloroplast evolution secondarysymbiogenesis and multiple losses Curr Biol 12 R62ndashR64
Chitnis P R amp Thornber J P 1988 The major light-har-vesting complex of photosystem 2 Aspects of its molecularand cell biology Photosynth Res 16 41ndash63
Chow W S Melis A amp Anderson J M 1990 Adjustmentsof photosystem stoichiometry in chloroplasts improve thequantum efficiency of photosynthesis Proc Natl Acad SciUSA 87 7502ndash7506
Claros M G Perea J Shu Y Samatey F A PopotJ L amp Jacq C 1995 Limitations to in vivo import of hydro-phobic proteins into yeast mitochondria The case of a cyto-plasmically synthesized apocytochrome b Eur J Biochem228 762ndash771
Dalbey R E amp Robinson C 1999 Protein translocation intoand across the bacterial plasma membrane and the plant thy-lakoid membrane Trends Biochem Sci 24 17ndash22
Daley D O Adams K L Clifton R Qualmann S MillarA H Palmer J D Pratje E amp Whelan J 2002 Genetransfer from mitochondrion to nucleus novel mechanismsfor gene activation from Cox2 Plant J 30 11ndash21
Danon A amp Mayfield S P 1994 Light-regulated translationof chloroplast messenger RNAs through redox potentialScience 266 1717ndash1719
de Grey A 2000 Mitochondrial gene therapy an arena for thebiomedical use of inteins Trends Biotechnol 18 394ndash399
Deisenhofer J Epp O Miki K Huber R amp Michel H1985 Structure of the protein subunits in the photosyntheticreaction center of Rhodopseudomonas viridis at 3Ac resolutionNature 318 618ndash624
34 J F Allen Genomes in mitochondria and chloroplasts
Delwiche C F 1999 Tracing the thread of plastid diversitythrough the tapestry of life Am Nat 154 S164ndashS177
Doolittle W F 1999 Phylogenetic classification and the uni-versal tree Science 284 2124ndash2129
Douglas S E 1998 Plastid evolution origins diversitytrends Curr Opin Genet Dev 8 655ndash661
Duby G amp Boutry M 2002 Mitochondrial protein importmachinery and targeting information Plant Sci 162 477ndash490
Eberhard S Drapier D amp Wollman F A 2002 Searchinglimiting steps in the expression of chloroplast encoded pro-teins relations between gene copy number transcriptiontranscript abundance and translation rate in the chloroplastof Chlamydomonas reinhardtii Plant J 31 149ndash160
El Bissati K amp Kirilovsky D 2001 Regulation of psbA andpsaE expression by light quality in Synechocystis species PCC6803 A redox control mechanism Plant Physiol 1251988ndash2000
Ellis R J 1984 The nuclear domination of chloroplast devel-opment Science Progress 69 129ndash142
Embley T M amp Martin W 1998 Molecular evolution ahydrogen-producing mitochondrion Nature 396 517ndash519
Ems S C Morden C W Dixon C K Wolfe K HdePamphilis C W amp Palmer J D 1995 Transcriptionsplicing and editing of plastid RNAs in the non-photosyn-thetic plant Epifagus virginiana Plant Mol Biol 29 721ndash733
Eraso J M amp Kaplan S 2000 From redox flow to gene regu-lation role of the PrrC protein of Rhodobacter sphaeroides241 Biochemistry 39 2052ndash2062
Eraso J M amp Kaplan S 2002 Redox flow as an instrumentof gene regulation Meth Enzymol 348 216ndash229
Escoubas J M Lomas M Laroche J amp Falkowski P G1995 Light-intensity regulation of cab gene-transcription issignaled by the redox state of the plastoquinone pool ProcNatl Acad Sci USA 92 10 237ndash10 241
Ferea T L Botstein D Brown P O amp Rosenzweig R F1999 Systematic changes in gene expression patterns follow-ing adaptive evolution in yeast Proc Natl Acad Sci USA96 9721ndash9726
Forsberg J Rosenquist M Fraysse L amp Allen J F 2001Redox signalling in chloroplasts and mitochondria genomicand biochemical evidence for two-component regulatory sys-tems in bioenergetic organelles Biochem Soc Trans 29403ndash407
Fujita Y 1997 A study on the dynamic features of photosys-tem stoichiometry accomplishments and problems forfuture studies Photosynth Res 53 83ndash93
Gabriel K Buchanan S K amp Lithgow T 2001 The alphaand the beta protein translocation across mitochondrial andplastid outer membranes Trends Biochem Sci 26 36ndash40
Galvis M L E Allen J F amp Hakansson G 1998 Proteinsynthesis by isolated pea mitochondria is dependent on theactivity of respiratory complex II Curr Genet 33 320ndash329
Gatenby A A amp Ellis R J 1990 Chaperone function theassembly of ribulose bisphosphate carboxylase-oxygenase ARev Cell Biol 6 125ndash149
GoldschmidtClermont M 1998 Coordination of nuclear andchloroplast gene expression in plant cells Int Rev CytolSurv Cell Biol 177 115ndash180
Gray M W 1992 The endosymbiont hypothesis revisited IntRev Cytol 141 233ndash357
Gray M W 1999 Evolution of organellar genomes CurrOpin Genet Dev 9 678ndash687
Gray M W 2000 Evolutionary biology mitochondrial geneson the move Nature 408 302ndash305
Gray M W amp Doolittle W F 1982 Has the endosymbionthypothesis been proven Microbiol Rev 46 1ndash42
Gray M W Burger G amp Lang B F 1999 Mitochondrialevolution Science 283 1476ndash1481
Phil Trans R Soc Lond B (2003)
Grebe T W amp Stock J B 1999 The histidine protein kinasesuperfamily Adv Microb Physiol 41 139ndash227
Green B R amp Kuhlbrandt W 1995 Sequence conservationof light-harvesting and stress-response proteins in relation tothe 3-dimensional molecular structure of Lhcii PhotosynthRes 44 139ndash148
Gruissem W amp Tonkyn J C 1993 Control mechanisms ofplastid gene expression Crit Rev Plant Sci 12 19ndash55
Hackstein J H P (and 11 others) 2001 Hydrogenosomesconvergent adaptations of mitochondria to anaerobicenvironments Zool Anal Complex Syst 104 290ndash302
Harrison M A Keen J N Findlay J B C amp Allen J F1990 Modification of a Glnb-like gene-product by photosyn-thetic electron-transport in the cyanobacterium Synechoc-occus 6301 FEBS Lett 264 25ndash28
Harvey A J Kind K L amp Thompson J G 2002 Redoxregulation of early embryo development Reproduction 123479ndash486
Heathcote P Fyfe P K amp Jones M R 2002 Reactioncentres the structure and evolution of biological solarpower Trends Biochem Sci 27 79ndash87
Henze K amp Martin W 2001 How do mitochondrial genesget into the nucleus Trends Genet 17 383ndash387
Herrmann R G amp Westhoff P 2001 Thylakoid biogenesis anddynamics the result of a complex phylogenetic puzzle InRegulation of photosynthesis vol 11 (ed E-M Aro amp BAndersson) pp 1ndash28 Dordrecht The Netherlands Kluwer
Hihara Y Sonoike K Kanehisa M amp Ikeuchi M 2002Search for redox responsive genes in Synechocystis sp PCC6803 by using DNA microarrays Plant Cell Physiol 43S178
Howe C J Barbrook A C amp Lockhart P J 2000 Organellegenes do they jump or are they pushed Trends Genet 1665ndash66
Ikeuchi M 2002 Gene expression and responses to light stressin cyanobacteria Plant Cell Physiol 43 S21
Iuchi S amp Lin E C C 1992 Mutational analysis of signaltransduction by Arcb a membrane sensor protein respon-sible for anaerobic repression of operons involved in the cen-tral aerobic pathways in Escherichia coli J Bacteriol 1743972ndash3980
Jackson-Constan D Akita M amp Keegstra K 2001 Molecu-lar chaperones involved in chloroplast protein importBiochim Biophys Acta Mol Cell Res 1541 102ndash113
Jarvis P amp Soll M 2001 Toc Tic and chloroplast proteinimport Biochim Biophys Acta Mol Cell Res 1541 64ndash79
Jordan P Fromme P Witt H T Klukas O SaengerW amp Krauss N 2001 Three-dimensional structure ofcyanobacterial photosystem I at 25 acngstrom resolutionNature 411 909ndash917
Kaneko T (and 23 others) 1996a Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regionsDNA Res 3 109ndash136
Kaneko T (and 23 others) 1996b Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regions[Supplement] DNA Res 3 185ndash209
Kanevski I amp Maliga P 1994 Relocation of the plastid rbcLgene to the nucleus yields functional ribulose-15-bisphosph-ate carboxylase in tobacco chloroplasts Proc Natl Acad SciUSA 91 1969ndash1973
Kim J amp Mayfield S P 1997 Protein disulfide isomerase as aregulator of chloroplast translational activation Science 2781954ndash1957
Genomes in mitochondria and chloroplasts J F Allen 35
Kirk J T O amp Tilney-Bassett R A E 1978 The plastidsTheir chemistry structure growth and inheritance Amster-dam Elsevier
Koehler C M Merchant S amp Schatz G 1999 How mem-brane proteins travel across the mitochondrial intermem-brane space Trends Biochem Sci 24 428ndash432
Komiya T Rospert S Schatz G amp Mihara K 1997 Bind-ing of mitochondrial precursor proteins to the cytoplasmicdomains of the import receptors Tom70 and Tom20 isdetermined by cytoplasmic chaperones FASEB J 11 1196
Konstantinov Y M Lutsenko G N amp Podsosonny V A1995 Genetic functions of isolated maize mitochondriaunder model changes of redox conditions Biochem MolBiol Int 36 319ndash326
Korab-Laskowska M Rioux P Brossard N LittlejohnT G Gray M W Lang B F amp Burger G 1998 Theorganelle genome database project (GOBASE) Nucleic AcidsRes 26 138ndash144
Kovacs L Wiessner W Kis M Nagy F Mende D ampDemeter S 2000 Short- and long-term redox regulation ofphotosynthetic light energy distribution and photosystemstoichiometry by acetate metabolism in the green alga Chla-mydobotrys stellata Photosynth Res 65 231ndash247
Kuhlbrandt W 1994 Structure and function of the plant light-harvesting complex LHC-II Curr Opin Struct Biol 4519ndash528
Lang B F Gray M W amp Burger G 1999 Mitochondrialgenome evolution and the origin of eukaryotes A RevGenet 33 351ndash397
Lemire B D Fankhauser C Baker A amp Schatz G 1989The mitochondrial targeting function of randomly generatedpeptide sequences correlates with predicted helical amphi-philicity J Biol Chem 264 20 206ndash20 215
Leuenberger D Bally N A Schatz G amp Koehler C M1999 Different import pathways through the mitochondrialintermembrane space for inner membrane proteins EMBOJ 18 4816ndash4822
Li H amp Sherman L A 2000 A redox-responsive regulator ofphotosynthesis gene expression in the cyanobacterium Syne-chocystis sp strain PCC 6803 J Bacteriol 182 4268ndash4277
Lin X Y (and 36 others) 1999 Sequence and analysis of chro-mosome 2 of the plant Arabidopsis thaliana Nature 402761ndash768
Lonsdale D M Hodge T P Howe C J amp Stern D B1983 Maize mitochondrial-DNA contains a sequence hom-ologous to the ribulose-15-bisphosphate carboxylase largesubunit gene of chloroplast DNA Cell 34 1007ndash1014
McFadden G I 1999 Endosymbiosis and evolution of theplant cell Curr Opin Plant Biol 2 513ndash519
McFadden G I 2001 Chloroplast origin and integrationPlant Physiol 125 50ndash53
Mahler H R amp Raff R A 1975 The evolutionary origin ofthe mitochondrion a nonsymbiotic model Int Rev Cytol43 1ndash124
Manfredi G Fu J Ojaimi J Sadlock J E Kwong J QGuy J amp Schon E A 2002 Rescue of a deficiency in ATPsynthesis by transfer of MTATP6 a mitochondrial DNA-encoded gene to the nucleus Nature Genet 30 394ndash399
Marc P Margeot A Devaux F Blugeon C Corral-Debrinski M amp Jacq C 2002 Genome-wide analysis ofmRNAs targeted to yeast mitochondria EMBO Rep 3159ndash164
Marienfeld J Unseld M amp Brennicke A 1999 The mito-chondrial genome of Arabidopsis is composed of both nativeand immigrant information Trends Plant Sci 4 495ndash502
Martin W 1999 A briefly argued case that mitochondria andplastids are descendants of endosymbionts but that thenuclear compartment is not Proc R Soc Lond B 2661387ndash1395 (DOI 101098rspb19990792)
Phil Trans R Soc Lond B (2003)
Martin W amp Herrmann R G 1998 Gene transfer fromorganelles to the nucleus how much what happens andwhy Plant Physiol 118 9ndash17
Martin W amp Muller M 1998 The hydrogen hypothesis forthe first eukaryote Nature 392 37ndash41
Martin W amp Russell M J 2003 On the origins of cells ahypothesis for the evolutionary transitions from abiotic geo-chemistry to chemoautotrophic prokaryotes and from pro-karyotes to nucleated cells Phil Trans R Soc Lond B 35859ndash85 (DOI 101098rstb20021183)
Martin W Stoebe B Goremykin V Hansmann S Hase-gawa M amp Kowallik K V 1998 Gene transfer to thenucleus and the evolution of chloroplasts Nature 393162ndash165
Martin W Hoffmeister M Rotte C amp Henze K 2001 Anoverview of endosymbiotic models for the origins of eukary-otes their ATP-producing organelles (mitochondria andhydrogenosomes) and their heterotrophic lifestyle BiolChem 382 1521ndash1539
Martin W Rujan T Richly E Hansen A Cornelsen SLins T Leister D Stoebe B Hasegawa M amp PennyD 2002 Evolutionary analysis of Arabidopsis cyanobacterialand chloroplast genomes reveals plastid phylogeny and thou-sands of cyanobacterial genes in the nucleus Proc NatlAcad Sci USA 99 12 246ndash12 251
Millen R S (and 12 others) 2001 Many parallel losses of infAfrom chloroplast DNA during angiosperm evolution withmultiple independent transfers to the nucleus Plant Cell 13645ndash658
Morse D Salois P Markovic P amp Hastings J W 1995 Anuclear-encoded form-II RUBISCO in dinoflagellatesScience 268 1622ndash1624
Murakami A Kim S J amp Fujita Y 1997 Changes in photo-system stoichiometry in response to environmental con-ditions for cell growth observed with the cyanophyteSynechocystis PCC 6714 Plant Cell Physiol 38392ndash397
Oh J I amp Kaplan S 2000 Redox signaling globalization ofgene expression EMBO J 19 4237ndash4247
Ozawa T 1995 Mitochondrial-DNA mutations associatedwith aging and degenerative diseases Exp Gerontol 30269ndash290
Palmer J D 1997 Organelle genomes going going goneScience 275 790ndash791
Pearson C K Wilson S B Schaffer R amp Ross A W 1993NAD turnover and utilization of metabolites for RNA syn-thesis in a reaction sensing the redox state of the cyto-chrome-b( 6 )f complex in isolated chloroplasts Eur JBiochem 218 397ndash404
Pfanner N amp Geissler A 2001 Versatility of the mitochon-drial protein import machinery Natl Rev Mol Cell Biol 2339ndash349
Pfanner N amp Wiedemann N 2002 Mitochondrial proteinimport two membranes three translocases Curr Opin CellBiol 14 400ndash411
Pfannschmidt T Nilsson A amp Allen J F 1999a Photosyn-thetic control of chloroplast gene expression Nature 397625ndash628
Pfannschmidt T Nilsson A Tullberg A Link G amp AllenJ F 1999b Direct transcriptional control of the chloroplastgenes psbA and psaAB adjusts photosynthesis to light energydistribution in plants IUBMB Life 48 271ndash276
Pfannschmidt T Allen J F amp Oelmuller R 2001a Prin-ciples of redox control in photosynthesis gene expressionPhysiol Plant 112 1ndash9
Pfannschmidt T Schutze K Brost M amp Oelmuller R2001b A novel mechanism of nuclear photosynthesis generegulation by redox signals from the chloroplast during pho-
36 J F Allen Genomes in mitochondria and chloroplasts
tosystem stoichiometry adjustment J Biol Chem 27636 125ndash36 130
Popot J L amp de Vitry C 1990 On the microassembly of inte-gral membrane proteins A Rev Biophys Biophys Chem 19369ndash403
Race H L Herrmann R G amp Martin W 1999 Why haveorganelles retained genomes Trends Genet 15 364ndash370
Raven J A Johnston A M Parsons R amp Kubler J 1994aThe influence of natural and experimental high O2 concen-trations on O2 evolving phototrophs Biol Rev CambridgePhil Soc 69 61ndash94
Raven J A Johnston A M Parsons R amp Kubler J 1994bThe occurrence and influence on photolithotrophs of highoxygen concentrations Proc R Soc Edinb B 102 193ndash201
Robinson C 2000 The twin-arginine translocation system anovel means of transporting folded proteins in chloroplastsand bacteria Biol Chem 381 89ndash93
Robinson C amp Bolhuis A 2001 Protein targeting by the twin-arginine translocation pathway Natl Rev Mol Cell Biol 2350ndash356
Rodermel S 2001 Pathways of plastid-to-nucleus signalingTrends Plant Sci 6 471ndash478
Roger A J 1999 Reconstructing early events in eukaryoticevolution Am Nat 154 S146ndashS163
Roger A J Svard S G Tovar J Clark C G SmithM W Gillin F D amp Sogin M L 1998 A mitochondrial-like chaperonin 60 gene in Giardia lamblia evidence thatdiplomonads once harbored an endosymbiont related to theprogenitor of mitochondria Proc Natl Acad Sci USA 95229ndash234
Roodyn D B amp Wilkie D 1968 The biogenesis of mitochondriaMethuenrsquos monographs on biological subjects London Methuen
Rujan T amp Martin W 2001 How many genes in Arabidopsiscome from cyanobacteria An estimate from 386 proteinphylogenies Trends Genet 17 113ndash120
Saccone C Gissi C Lanave C Larizza A Pesole G ampReyes A 2000 Evolution of the mitochondrial genetic sys-tem an overview Gene 261 153ndash159
Salzberg S L White O Peterson J amp Eisen J A 2001Microbial genes in the human genome lateral transfer orgene loss Science 292 1903ndash1906
Sargent F Berks B C amp Palmer T 2002 Assembly of mem-brane-bound respiratory complexes by the Tat protein-trans-port system Arch Microbiol 178 77ndash84
Schatz G 1998 Protein transport the doors to organellesNature 395 439ndash440
Schemidt R A Qu J Williams J R amp Brusilow W S1998 Effects of carbon source on expression of F0 genes andon the stoichiometry of the c subunit in the F1 F0 ATPaseof Escherichia coli J Bacteriol 180 3205ndash3208
Selosse M A Albert B R amp Godelle B 2001 Reducing thegenome size of organelles favours gene transfer to thenucleus Trends Ecol Evol 16 135ndash141
Stern D B amp Lonsdale D M 1982 Mitochondrial and chlo-roplast genomes of maize have a 12- kilobase DNA-sequencein common Nature 299 698ndash702
Stock D Gibbons C Arechaga I Leslie A G W ampWalker J E 2000 The rotary mechanism of ATP synthaseCurr Opin Struct Biol 10 672ndash679
Stoebe B amp Maier U G 2002 One two three naturersquos toolbox for building plastids Protoplasma 219 123ndash130
Stoebe B Martin W amp Kowallik K V 1998 Distributionand nomenclature of protein-coding genes in 12 sequencedchloroplast genomes Plant Mol Biol Rep 16 243ndash255
Stupar R M Lilly J W Town C D Cheng Z Kaul SBuell C R amp Jiang J M 2001 Complex mtDNA consti-tutes an approximate 620 kb insertion on Arabidopsis thali-ana chromosome 2 implication of potential sequencing
Phil Trans R Soc Lond B (2003)
errors caused by large-unit repeats Proc Natl Acad SciUSA 98 5099ndash5103
Surpin M Larkin R M amp Chory J 2002 Signal transduc-tion between the chloroplast and the nucleus Plant Cell 14S327ndashS338
Tjaden J Schwoppe C Mohlmann T Quick P W ampNeuhaus H E 1998 Expression of a plastidic ATPADPtransporter gene in Escherichia coli leads to a functional aden-ine nucleotide transport system in the bacterial cytoplasmicmembrane J Biol Chem 273 9630ndash9636
Tovar J Fischer A amp Clark C G 1999 The mitosome a novelorganelle related to mitochondria in the amitochondrial para-site Entamoeba histolytica Mol Microbiol 32 1013ndash1021
Trebitsh T amp Danon A 2001 Translation of chloroplastpsbA mRNA is regulated by signals initiated by both photos-ystems II and I Proc Natl Acad Sci USA 98 12 289ndash12 294
Tullberg A Alexciev K Pfannschmidt T amp Allen J F2000 Photosynthetic electron flow regulates transcription ofthe psaB gene in pea (Pisum sativum L) chloroplasts throughthe redox state of the plastoquinone pool Plant Cell Physiol41 1045ndash1054
Turmel M Lemieux C Burger G Lang B F Otis CPlante I amp Gray M W 1999 The complete mitochondrialDNA sequences of Nephroselmis olivacea and Pedinomonasminor two radically different evolutionary patterns withingreen algae Plant Cell 11 1717ndash1729
Unden G amp Bongaerts J 1997 Alternative respiratory path-ways of Escherichia coli energetics and transcriptional regu-lation in response to electron acceptors Biochim BiophysActa Bioenerg 1320 217ndash234
Unseld M Marienfeld J R Brandt P amp Brennicke A1997 The mitochondrial genome of Arabidopsis thaliana con-tains 57 genes in 366 924 nucleotides Nature Genet 1557ndash61
Van der Giezen M Slotboom D J Horner D S DyalP L Harding M Xue G P Embley T M amp KunjiE R S 2002 Conserved properties of hydrogenosomal andmitochondrial ADPATP carriers a common origin for bothorganelles EMBO J 21 572ndash579
Von Heijne G 1986 Why mitochondria need a genome FEBSLett 198 1ndash4
Westphal S Soll J amp Vothknecht U C 2001 A vesicle trans-port system inside chloroplasts FEBS Lett 506 257ndash261
Whitehouse H L K 1969 Towards an understanding of themechanism of heredity London Edward Arnold Ltd
Williams B A P Hirt R P Lucocq J M amp Embley T M2002 A mitochondrial remnant in the microsporidian Trachi-pleistophora hominis Nature 418 865ndash869
Wilson R J (and 10 others) 1996a Complete gene map of theplastid-like DNA of the malaria parasite Plasmodium falcipa-rum J Mol Biol 261 155ndash172
Wilson S B Davidson G S Thomson L M amp PearsonC K 1996b Redox control of RNA synthesis in potato mito-chondria Eur J Biochem 242 81ndash85
Winkler H H amp Neuhaus H E 1999 Non-mitochondrialATP transport Trends Biochem Sci 24 64ndash68
Wolfe K H Morden C W amp Palmer J D 1992 Small sin-gle-copy region of plastid DNA in the non-photosyntheticangiosperm Epifagus virginiana contains only two genes Dif-ferences among dicots monocots and bryophytes in geneorganization at a non-bioenergetic locus J Mol Biol 22395ndash104
Wollman F A Minai L amp Nechushtai R 1999 The biogen-esis and assembly of photosynthetic proteins in thylakoidmembranes Biochim Biophys Acta Bioenerg 1411 21ndash85
Yeliseev A A amp Kaplan S 2000 TspO of Rhodobactersphaeroides a structural and functional model for the mam-
Genomes in mitochondria and chloroplasts J F Allen 37
malian peripheral benzodiazepine receptor J Biol Chem275 5657ndash5667
Zerges W 2002 Does complexity constrain organelle evol-ution Trends Plant Sci 7 175ndash182
Zhang Z D Green B R amp Cavalier-Smith T 1999 Singlegene circles in dinoflagellate chloroplast genomes Nature400 155ndash159
Zhang Z D Cavalier-Smith T amp Green B R 2001 A fam-ily of selfish minicircular chromosomes with jumbled chloro-plast gene fragments from a dinoflagellate Mol Biol Evol18 1558ndash1565
Zouni A Witt H T Kern J Fromme P Krauss NSaenger W amp Orth P 2001 Crystal structure of photosys-tem II from Synechococcus elongatus at 38 acngstrom resol-ution Nature 409 739ndash743
DiscussionD S Horner (Dipartimento di Fisiologia e Biochimica
Generali University of Milan Milan Italy) Given thatthere are components of these membrane-bound energy-transduction pathways that are consistently nuclear enco-ded not the key ones you pointed out but others do youexpect perhaps a slower perhaps a more crude but yet aredox communication between the organelle andexpression of nuclear-encoded genes
J F Allen There is and there is no question about itI think that Bogoradrsquos lock-in hypothesis has somethingto say here and there are other people who say it is all todo with assembling multi-subunit complexes If you havecontrol over the initial construction of the core aroundwhich the whole complex is eventually built then redoxcontrol or some sort of control over synthesis of that corewill control construction of the whole thing eventuallyand I think that may well be largely correct I think it ispossible in principle for nuclear genes to make precursorsaccording to redox instructions but it is not really soimportant that those precursors are here and now presentin exactly the right quantities because the precursors canto a limited extent hang around and wait Actually thereis proteolytic degradation of unattached precursors and soon but I think the fact that these chloroplast and mito-chondrially DNA-encoded subunits are at the core ofthese energy-transducing complexes is the crucial issue
T Cavalier-Smith (Department of Zoology University ofOxford Oxford UK ) In the case of mitochondria the sub-set of genes that remains is much more variable than youimplied It runs from three proteins in the minimal casesup to I think over 90 That is a big difference and manyof the genes in different lineages of protists that remainhave nothing to do with redox control they are things likeribosomal proteins and so on So I think that even thoughwhat you say about redox control of certain genes mayactually be an important thing it is not the fundamentaloverall reason why the genomes are maintained It may bepart of the explanation for why certain genes are alwaysincluded in that subset but even in mitochondria I thinkthat cytochrome b is the only one that is always presentSo I think that both this great diversity of what has beenretained in different lineages and the nucleomorph caseindicate that evolution was able to do one thing in onelineage 500 million years ago and never got around todoing it for 500 million years in another lineage so thereis a lot more haphazardness and evolutionary accidentsand contingency than a straight functional interpretation
Phil Trans R Soc Lond B (2003)
there is a lot of just sheer lsquohappenedndashorndashdidnrsquotndashhappenrsquoor carryover
J F Allen I was not implying anything about the rangeof mitochondrial genomes I have a limited amount oftime so I just talked about what I could talk about withinthat limit Of course there will be a range and this willdepend upon the physiology and bioenergetics of theparticular organism But what I am trying to do is to seewhat the common properties of the irreducible core areof components that are always encoded in the organellesand cytochrome b is a perfect example because that reallyis the thing that binds these two all-important quinoneswithout which there is no Q cycle without which there isno proton pumping and bioenergetics and in Plasmodiumwhere the mitochondrion is reduced to this bare mini-mum there is still a mitochondrial gene for cytochromeb Now my argument about the ribosomal proteins andall the rest of it is that in order to be able to exert thisall-important redox control over gene expression of certaincore components you need an in situ genetic system andthis can vary in composition for reasons that I cannotdescribe Maybe you need RNA polymerase or maybeyou can import RNA polymerase In plastids you haveboth of these options You are going to need ribosomestransfer RNAs and the rest of the apparatus of geneexpression and that needs to be in situ but we are stillleft with the question why does there have to be any gen-etic system at all The mature tobacco or spinach chloro-plast seems to have an entire genetic system just to makeone protein D1 which is all it ever needs to resynthesizeI think that this is a clue It is worth having this very unec-onomical crazy system because there is an overridingrequirement to connect electron transfer with geneexpression directly and unambiguouslymdashwithout recourseto a bureaucracy if you like which is what I think thenucleus is
K Willison (Institute of Cancer Research Chester BeattyLaboratories London UK ) These photosynthetic reactioncomplexes are a sort of baroque mixture of polypeptidechains and cofactors and my understanding is that someof them in high photon flux can turn over within say 30minutes so I am wondering if there is a correlationbetween gene retention and the turnover rates of some ofthe components and the complexes it seems to me thatthat could really be a limiting factor and the reason thatthe proteins had to be encoded locally in order to get thecomplexes to be assembled rapidly
J F Allen I think that is right You can only alter thesteady-state concentration of a protein complex by affect-ing the synthesis and breakdown and therefore turnoverThere is an extreme example the D1 protein encoded bythe psbA gene of chloroplasts which is always in the chlor-oplast genome without exception High light will tend todegrade this protein and when the high light goes awaythere is an absolute need very quickly to bring it back upagain I think that is a very good example Whether thisargument applies to genes such as cytochrome oxidase Ido not know
C J Leaver (Department of Plant Sciences University ofOxford Oxford UK ) Your experiments tend to relatemainly to the gene expression side What about proteinRNA turnover the whole concept of redox control ofthatmdashdo we know anything
38 J F Allen Genomes in mitochondria and chloroplasts
J F Allen Well quite a lot is known about chloro-plasts There is redox control of messenger RNA stabilityfor this same D1 protein which has been described byStephen Mayfield and his co-workers The evidence is thatthe site of redox control is ferredoxin via thioredoxin andthe implication is that thioredoxin tells whether non-cyclicelectron transport is working at all When it is workingthere is a signal that says lsquoOK now continue with all theprotein synthesisrsquo It is not particularly specific but moreof a global effect I think this is what Mayfield argues butof course this is not inconsistent with my argument I haveconcentrated on quinone pools and medium-potentialredox regulatory control because that is where I can seesignals about the dangerous consequences of getting itwrong but there is control of other levels of geneexpression In mitochondria I am not so sure I wouldbounce the question back to you on that one
W Martin (Institute of Botany III Heinrich-Heine Univ-ersitat Dusseldorf Dusseldorf Germany) The nucleomorphgenome was cited as evidence in conflict with your modelbut is that true The nucleomorph genome is not in abioenergetic organelle in that sense It resides in the cyto-sol so does the size of the nucleomorph genome workagainst your model Is it really evidence against yourmodel
J F Allen I do not know why should it be evidenceI cannot explain why the nucleomorph has a genomeThat is why I clutched at the straw that it is storage fornucleic acid precursors
Phil Trans R Soc Lond B (2003)
T Cavalier-Smith The reason I introduced the nucleo-morph is not to say it is direct evidence against yourhypothesis but to show that there probably was no singlefunctional reason for the retention of every gene in everyorganelle It may be that there are specific physiologicalreasons for the retention of certain genes and thereforeyou might be right about a subset of genes but I verymuch doubt if there are specific physiological reasons forthe retention of every single gene that is retained differen-tially because odd things happen in evolution Somethings are easy and sometimes things are difficult Evol-ution is history
J F Allen Of course I agree with that I do not thinkI can explain everything but I do think I can explainsomething If I have done that then I am content
GLOSSARY
AAC ADPndashATP carrierCES control by epistasy of synthesisCORR co-location for redox regulationLHC light-harvesting complexPCC Pasteur culture collectionORF open reading frameRUBISCO ribulose-15-bisphosphate carboxylasendashoxygenaseTAT twin-arginine translocase
Visit httpwwwpubsroyalsocacuk to see an electronic appendix tothis paper

28 J F Allen Genomes in mitochondria and chloroplasts
utionary terms the near future (Palmer 1997 Gray 19992000 Gray et al 1999 Adams et al 2000 Daley et al2002) A clear and compelling statement of this widelyheld position is that of Herrmann amp Westhoff (2001 p10)
hellipthe transfer of geneshelliptells that the partite plant gen-ome is not in a phylogenetic equilibrium All availabledata suggest that the ultimate aim of genome restructur-ing in the plant cell as in the eukaryotic cell in generalis the elimination of genome compartmentation whileretaining physiological compartmentation
One serious problem with this hypothesis is the dis-tinctly non-random sample of genes that remain to betransferred A relatively constant subset of genes is foundin bioenergetically functional mitochondria from fungi tomammals and angiosperms as in photosynthetic plastids(chloroplasts) from unicellular algae to angiosperms Thishypothesis therefore requires an additional ad hocassumption that some gene transfers are inherently moreprobable than others (this too is widely supposed seelater) and thus that gene transfer events proceed in thesame sequence in quite independent lineages Thereseems to be no evidence for this and the order of geneloss from mitochondria and chloroplasts has been used toinfer phylogeny (Martin et al 1998 Adams et al 2000Millen et al 2001) lsquoGoing going gonersquo (Palmer 1997)is difficult to refute categorically but it does not seem tobe consistent with recent evidence For example chromo-some two of Arabidopsis thaliana (Lin et al 1999) has asingle insertion encompassing with about 99 sequencesimilarity the complete Arabidopsis mitochondrial genome(Unseld et al 1997 Marienfeld et al 1999 Stupar et al2001) Unless this single transfer is a freak occurrencethis finding strongly indicates that gene transfer occurswith relative ease (Adams et al 2000 Henze amp Martin2001 Rujan amp Martin 2001 Salzberg et al 2001) Inaddition there is experimental evidence that randomlygenerated polypeptides are mitochondrially imported athigh frequency (Lemire et al 1989) The hypothesis thatwe now see the same few stragglers in diverse organellargenomes is not impossible but should be regarded as alast resort perhaps applicable only in extreme cases suchas the greatly reduced apicomplexan plastomes and chon-driomes Even there the hypothesis carries with it an obli-gation to explain why organisms with few organellar geneshave lsquoevolvedrsquo faster than those with many
(d ) The frozen accidentWhat may be described as lsquothe frozen accident hypoth-
esisrsquo states that the evolutionary process of gene transferfrom organelle to nucleus was underway when somethinghappened that stopped it One example of the frozen acci-dent is a scenario (Von Heijne 1986) in which import intomitochondria of the precursor proteins arising from newlytranslocated genes proceeded for long enough after theprimordial endosymbiosis for most of the symbiont-derived genes to be moved to the cell nucleus The lsquoacci-dentrsquo that halted the process was the evolutionary originof exocytosis and protein secretion after which it becameimpossible for precursors properly destined for mitochon-drial import to be distinguished from those destined forexport from the cell As with the lsquolock-inrsquo hypothesis the
Phil Trans R Soc Lond B (2003)
lsquofrozen accidentrsquo seems incompatible with the remarkableprecision and specificity of protein targeting (Schatz 1998Koehler et al 1999 Leuenberger et al 1999 Gabriel etal 2001 Jarvis amp Soll 2001 Pfanner amp Geissler 2001Duby amp Boutry 2002 Pfanner amp Wiedemann 2002) Pro-tein sorting mechanisms do not seem to forbid controlledimport into any cellular compartment Another lsquoaccidentrsquothat may have impeded subsequent gene transfer in thecase of mitochondria is their departure from what is other-wise a universal genetic code Genetic code differences arediscussed later in the context of whether some genes can-not be moved
(e) Some proteins (with cofactors) cannot beimported
We may describe as the lsquounimportability hypothesisrsquo theproposal that some proteins are inherently unimportableand must therefore be synthesized de novo in the correctcompartment within the cell The unimportability hypoth-esis is most often proposed as its special case the hydro-phobicity hypothesis with hydrophobicity being thereason for unimportability
(i) HydrophobicityA commonly entertained notion is that proteins that are
encoded and synthesized within organelles comprise a pri-mary subset characterized by shared hydrophobicity allare intrinsic membrane proteins (Von Heijne 1986Popot amp de Vitry 1990 Claros et al 1995) As with theCORR hypothesis favoured here the primary subset thenrequires a secondary subset of genetic system componentsthe latter not themselves being hydrophobic but beingrequired for synthesis of the others Certainly the mito-chondrial inner membrane contains some intrinsic proteincomplexes whose subunits broadly satisfy the criterion ofhydrophobicity However the core subunits that are mito-chondrially encoded are also explained by the CORRhypothesis This would make hydrophobicity a require-ment for their function in vectorial electron and protontransfer and not a reason in itself for retention of theirgenes in the cytoplasm
The hydrophobicity hypothesis has some very clearcounter-examples in chloroplasts and cannot thereforestand if one wishes for a single principle that embracesboth types of bioenergetic organelle One clear counter-example is the large (L) subunit of the enzyme RUBISCOan abundant but entirely membrane-extrinsic protein ofca 55 kDa molecular mass The RUBISCO large subunitis generally chloroplast-encoded with the exception of thenuclearly encoded bacterial-type L subunit of L2 RUBI-SCO in peridinin-containing dinoflagellates (Morse et al1995) In most photosynthetic eukaryotes RUBISCO hasa chloroplast encoded large subunit and a nuclearly enco-ded small (S) subunit (Ellis 1984 Gatenby amp Ellis 1990)giving an L8S8 holoenzyme and this RUBISCO is widelytaken as a paradigm for the need for coordination ofnuclear and chloroplast gene expression (Rodermel 2001)
By complete contrast to the RUBISCO large subunitwhich is a chloroplast-encoded and water-soluble proteina second major violation of the hydrophobicity hypothesisis the existence of the universally nuclearly encoded butalways hydrophobic subunits of the chloroplast LHC IIand LHC I (Chitnis amp Thornber 1988) These apopro-
Genomes in mitochondria and chloroplasts J F Allen 29
teins of ca 27ndash30 kDa are quite water-insoluble and areusually fully extracted only in chloroformndashmethanol or inthe presence of detergents The Lhc gene family is diverse(Green amp Kuhlbrandt 1995) but most gene productsor Lhcp (light-harvesting chlorophyll protein) polypeptideshave three membrane-spanning hydrophobic a-helices(Kuhlbrandt 1994) It is interesting that three-helix Lhcproteins seem to be absent from extant cyanobacteria andchloroxybacteria (Green amp Kuhlbrandt 1995)
(ii) The five-helix rulePerhaps with the three-helix model of LHC II in mind
a variation of the hydrophobicity hypothesis is the empiri-cal lsquofive-helix rulersquo (Popot amp de Vitry 1990 Claros et al1995 Wollman et al 1999) which states that no intrinsicmembrane protein with five or more membrane-spanninghelices can be encoded in the nucleus Five is a good guessfor a threshold number of helices demanding organellar-encoding in the context of photosynthetic reaction centres(Heathcote et al 2002) where the earliest known struc-ture that of type II centres (Deisenhofer et al 1985) asin photosystem II (Zouni et al 2001) shows exactly fivehelices with type I centres containing 11 (Jordan et al2001) There are however many nuclearly encoded sub-units in both mitochondria and chloroplasts that havefewer than five membrane helices so the five-helix rulecan only describe a sufficient condition and not a neces-sary condition for nuclear encoding
It should also be noted that ADPndashATP carriers (AACs)are encoded in the nucleus AACs of the mitochondrialinner membrane have six transmembrane helices (van derGiezen et al 2002) while the those of the chloroplast innerenvelope (Winkler amp Neuhaus 1999) have 12 (Tjaden etal 1998)
With the counter-examples of nuclearly encoded AACswith six or 12 membrane-spanning helices many intrinsicmembrane proteins with fewer than five helices somenuclearly encoded and some not and with the RUBISCOlarge subunit as a major product of chloroplast geneexpression with no membrane domain at all the five-helixrule begins to seem rather like a case of special pleadingEven if there are indeed special mitigating circumstancesfor AACs (this argument is outlined later) the five-helixrule as with the hydrophobicity hypothesis of which it isa special case will disappear if it can be shown that pos-session of five or more membrane helices is required forsome other function such as that of CORR that deter-mines the organellar location of the gene concernedHydrophobicity is in some cases a feature of proteins thatare encoded in organelles but it is not in itself the issue
For chloroplasts the hydrophobicity hypothesis predictsretention of cyanobacterial genes in functional categoriesassociated with the chloroplast envelope and with trans-port just as much as with photosynthesis Table 1 showsthat this is not the case
A mechanism for the unimportability of hydrophobicproteins is rarely stated explicitly and it is rather left tothe imagination whether hydrophobic proteins might asit were get stuck halfway through the chloroplast envelopeor mitochondrial outer membrane whether transport can-not get started on a precursor protein that has alreadyspontaneously inserted into the membrane or whetherhydrophobic proteins will be unable to make the journey
Phil Trans R Soc Lond B (2003)
through the cytosol without precipitating there In thecytosol co-translational transport or membrane insertionwould in principle address the latter point but it seemsto be generally true that chloroplast and mitochondrialproteins are post-translationally inserted into the organel-lar outer membrane So the obvious ancillary questionabout unimportability is lsquowhy then cannot hydrophobicproteins be co-translationally imported into organellesrsquoThere is evidence in yeast for co-translational transportinto mitochondria of proteins thought to be derived fromthe original endosymbiont (Marc et al 2002) If there isany truth in hydrophobicity as a barrier to protein importthe CORR principle of unselective import (sect 3c) shouldbe easily refuted for example by a demonstration that noknown targeting presequence is capable of importing poly-phenylalanine encoded by polyU for in vitro import stud-ies
An additional proposal intended to account for theunimportability of some hydrophobic proteins and forimportability of others is suggested by A de Grey(personal communication) who argues that bacteriallyderived protein precursors imported into mitochondriacross the mitochondrial inner membrane completelybefore insertion into that membrane from the mitochon-drial matrix (the N-phase) This is held to be an obstacleto successful import of membrane-intrinsic proteins thatmight en route discharge the proton motive force By con-trast eukaryotically derived proteins can be inserted intothe mitochondrial inner membrane from the inter-mem-brane space (the P-phase) and then there is for somereason no similar uncoupling effect Thus adenylatetransporters or AACs can be nuclearly encoded whereascytochrome oxidase subunits for example cannot It isnot obvious why bacterially derived subunits such asthose of cytochrome oxidase did not acquire eukaryoticinsertion mechanisms Perhaps this acquisition is forbid-den by the need to conserve pre-existing routes ofassembly into multi-subunit complexes or perhaps theneed for insertion from the inside reflects a need to incor-porate a cofactor for example haem or ironndashsulphur clus-ters along the way
(iii) Unimportability of three dimensional structures andcofactors
Accumulating evidence on protein transport mech-anisms seems to make a specific obstacle to successfultransport of certain sorts of protein less and less plausibleespecially given the observation that precursors with N-terminal targeting sequences not mature functional poly-peptides themselves traverse membranes during biosynth-esis
Protein folding inside the living cell is invariably assistedby molecular chaperones (Gatenby amp Ellis 1990 Azem etal 1997 Komiya et al 1997 1990 Jackson-Constan et al2001) proteins that impede the formation of lsquoimproperrsquostructural interactions that will otherwise produce aggre-gation or other non-functional interactions Proteinimport mechanisms appear mostly to rely on specific mol-ecular chaperones for guided unfolding before membraneinsertion and refolding after translocation of the polypep-tide into its destined compartment (Gatenby amp Ellis 1990Azem et al 1997 Komiya et al 1997 Jackson-Constanet al 2001) Because polypeptides are mostly lsquothreadedrsquo
30 J F Allen Genomes in mitochondria and chloroplasts
through membranes no special constraint on transportshould be expected to arise from 3D structure or surfaceproperties of a protein
An exception to this unfolding rule may be providedby holoproteins containing cofactors that are transportedalong with the apoprotein presumably with a largely intact3D structure The lsquoTATrsquo (twin-arginine translocase) sys-tem (Bogsch et al 1998 Dalbey amp Robinson 1999 Berkset al 2000ab Robinson 2000 Robinson amp Bolhuis 2001Sargent et al 2002) works broadly along these linesthough its mechanism is unresolved TAT is relevant herebecause it adds transport of essentially complete holopro-teins to the cellrsquos repertoire of transport mechanisms andeven proteins that may not be allowed to unfold perhapsbecause of the toxicity of the free cofactor are not therebymade candidates for an unimportability rule One variantof the unimportability hypothesis is that transport canwork only by unfolding and dissociated cofactors may betoxic to the cell The bacterially derived TAT system(Bogsch et al 1998 Dalbey amp Robinson 1999 Berks etal 2000ab Sargent et al 2002) overcomes this argumentas would examples of cofactor-free polypeptides such as(C)Fo subunit c which are nevertheless generally encodedin organelles
The presence of a vesicular transport system in chloro-plasts (Westphal et al 2001) counts against the unim-portability hypothesis Vesicular transport between thechloroplast envelope and the thylakoid membrane shouldin principle allow any protein to be ferried across thechloroplast stroma Vesicles might allow transport inparticular of membrane proteins that would discharge theproton motive force if allowed to connect transiently theN and P aqueous phases on each side of a coupling mem-brane
Another observation that seems to count against unim-portability is the increased number of membrane barriersin plastids derived from secondary tertiary or higher-orderendosymbioses These plastids seem to have retained ifanything fewer genes rather than more as unimportabilitypredicts The nucleomorph the residual nucleus of theprimary host retains many genes for molecular chap-erones (Archibald et al 2001 Cavalier-Smith 2002) inagreement with the conclusion that protein import pro-ceeds by conserved and specialized mechanisms If eachnew import added to gain a higher-order endosymbiosisrequires additional targeting presequences then perhapsthere is an advantage in components of an import machin-ery not being imported themselves
It seems that unimportability is less easy an assumptionto make today than in previous years and the onus maynow be on its proponents to cite specific examples of pro-teins that can never be imported into chloroplasts or mito-chondria
(f ) Some genes cannot be moved(i) Differences in the mitochondrial genetic code
The genetic code is the specific relation between ribon-ucleotide triplets of codons and tRNAs that recognizethem and which thus determine which individual aminoacids if any will be incorporated into the growing poly-peptide chain Mitochondria show small departures fromthe otherwise universal genetic code (Attardi amp Schatz1988 Alberts et al 1994) UGA is a lsquostoprsquo codon in the
Phil Trans R Soc Lond B (2003)
universal code and in plant mitochondria but is the codonfor tryptophan in mitochondria of animals and fungiCUA means leucine everywhere except fungal mitochon-dria where it means threonine and AGA and AGU codefor arginine in the lsquouniversalrsquo code but serine in insectmitochondria and lsquostoprsquo in mammalian ones The signifi-cance of this heterogeneity in coding is most unlikely torest in the early origin of mitochondria as the lsquouniversalrsquocode is employed by bacteria and protists Furthermorethe diversity of departures from the lsquouniversalrsquo codeamong mitochondria of different eukaryotic lineages indi-cates they had independent origins A general rule formitochondrial genetic codes is that reliance on the thirdlsquowobblersquo position is decreased which allows all 20 proteinamino acids to be specified by fewer tRNAs (22 in animalmitochondria instead of 31 in the cytosol) This in turnmeans that mtDNA can specify more proteins for a givenlength of DNA or conversely that the mitochondrialgenome can be condensed or lsquostreamlinedrsquo Although thisarrangement does not explain the retention of mitochon-drial genomes and may be a secondary event in mitochon-drial evolution it almost certainly places an extra barrierin place of gene relocation As discussed previously how-ever there seems to be no single barrier that can explainthe retention of a relatively constant subset of mitochon-drial genes
There has clearly been a contraction of mitochondrialDNA especially in animals Animal mitochondrial DNAhas few if any non-coding sequences and animal mito-chondrial genetic codes require fewer tRNAs than the sin-gle code operating in the cytosol (Gray 1999 Gray et al1999 Selosse et al 2001) These may be devices todecrease the total frequency of mutations per mitochon-drial genome assuming a constant frequency per unitlength of mitochondrial DNA It has been proposed thata genersquos removal to the nucleus decreases its susceptibilityto free radical mutagenesis (Raven et al 1994ab Allen ampRaven 1996) If this is the case then the non-standardcode may be a further indication of an overriding need tomaintain certain genes within the mitochondrion What-ever the reason for the retention of genes in organelles ithad better be a good one bioenergetic organelles areclearly the wrong place to keep genetic information andgo to extraordinary lengths to offset the consequences ofit being there
(ii) Molecular refugeesA phenomenon related to differences in the genetic code
is seen in chloroplasts This is a bias in codon usage Thelsquomolecular refugeesrsquo hypothesis (Howe et al 2000) sug-gests that nuclear copies of what were once chloroplastgenes escape constraints on codon usage and thus thatwhat distinguishes chloroplast-encoded proteins is thattheir amino-acid composition is unusual enough for thebias in codon usage not to apply It is difficult to see whatphysico-chemical properties might be uniquely shared bythe amino acids of chloroplast-encoded proteins and inaddition there seem to be no lsquoforbiddenrsquo amino-acid resi-due in chloroplasts As with the departure from the univer-sal genetic code in mitochondria is seems likely that abias in codon usage reflects some secondary property ofchloroplast genomes and is not the primary reason fortheir retention In any case any restriction on gene move-
Genomes in mitochondria and chloroplasts J F Allen 31
ment will act in the wrong direction to explain why chloro-plasts have retained genomes at all provided there is noresulting obstacle to a chloroplast protein being success-fully encoded as a precursor in the nucleus
(iii) Genes are copied and replicated not relocated in onestep
If complete relocation is envisaged as proceeding by thesimultaneous appearance of a gene in one compartmentand its deletion from another then it is possible to imaginethat whatever the vector is there may be selectivity ingene transfer (Henze amp Martin 2001) For example ifviral incorporation replication and recombination in bothorganelle and nucleus were the mechanism of relocationthen successful gene transfer might depend on the abilityof the virus to complete the process in one step This inturn might depend on some property of the gene or ofthe protein it encodes if expression occurs at any stage inthe process of relocation An assumption of the CORRhypothesis favoured here is that genes are not movedphysically but merely copied after which either the copyin the original location or the copy in the new locationmay be lost as a result of selection A useful analogy maybe the action of copying files between computer storagevolumes In this analogy the copy is identical to the orig-inal and either may subsequently be deleted without lossof information
Because any gene that is not usefully expressed willundergo no selection to maintain a functional allele theprevailing location will be the one in which the functionalcopy of the gene resides The problem of which gene sur-vives is then subsumed into the problem of which is theselectively advantageous location for it It seems reason-able to assume that there wasmdashand ismdashan interim periodin which homologous genes in different compartments areessentially in competition to see which functions best Asoutlined above there is no obvious obstacle for copyingDNA sequences within the cell The physico-chemicalproperties of DNA are fairly uniform and not directlydependent on nucleotide sequence though organelles likeintra-cellular parasites seem to prefer A and T to G andC The answer to the question lsquowhich gene location func-tions bestrsquo may have nothing to do with the gene itselfnor directly with the structure and function of its geneproduct Instead the answer may have everything to dowith the required regulatory interaction between the geneproduct and expression of the gene that produces it
(g) CORRThis hypothesis has been propounded earlier (sect 3) and
will be summarized as follows Vectorial electron and pro-ton transfer exerts regulatory control over expression ofgenes encoding proteins directly involved in or affectingredox poise This regulatory coupling is indispensable foradaptation to environmental changes in chemical activitiesof oxygen light and CO2 and requires co-location of thesegenes with their gene products Organelles lsquomake theirown decisionsrsquo on the basis of environmental changesaffecting redox state The hypothesis is abbreviatedCORR for co-location for redox regulation or more com-pletely lsquoco-location (of gene and gene product) for(evolutionary) continuity of redox regulation of geneexpressionrsquo
Phil Trans R Soc Lond B (2003)
5 SUMMARY OF THE CURRENT STATUS OF THEHYPOTHESIS OF CORR
Q Why do mitochondria and chloroplasts have theirown genetic systems
A CORR co-location of genes and gene products per-mits direct and autonomous redox regulation of geneexpression
(a) Objections to CORR(i) There is no regulation of gene expression in chloroplasts
and mitochondriaThis is still commonly stated but incorrect See the prin-
ciple in sect 3d
(ii) There is no regulation of gene expression in chloroplastsand mitochondria at the level of transcription
For chloroplasts there is now clear evidence for a func-tionally intelligible redox control of transcription(Pfannschmidt et al 1999a) This invalidates the previouswidely held assumption that all control of chloroplast geneexpression is post-transcriptional (Gruissem amp Tonkyn1993 GoldschmidtClermont 1998) The evidence formitochondria is less clear but redox control of transcrip-tion certainly cannot be ruled out and there are indi-cations that it may occur (Pearson et al 1993Konstantinov et al 1995 Wilson et al 1996b) It mustalso be restated that transcriptional control is not neces-sary for CORR but serves as a compelling example of aconserved typically bacterial mode of response that oper-ates still in eukaryotic cytoplasmic organelles thus illus-trating the principle of continuity of redox control (sect 3d)Post-transcriptional redox regulation (Danon amp Mayfield1994 Kim amp Mayfield 1997 Brown et al 2001 Treb-itsh amp Danon 2001) is consistent with CORR and thereis nothing to forbid it being a primary mechanism thatsatisfies most or indeed all of the formal requirements ofthe hypothesis in particular cases
(iii) There are no redox signalling components encoded inchloroplast or mitochondrial DNA
This is what CORR predicts See the principle in sect 3j
(iv) There is redox control of nuclear gene expressionThis is correct and many examples are now known
especially for chloroplasts (Escoubas et al 1995 Kovacs etal 2000 Pfannschmidt et al 2001ab Surpin et al 2002)Redox signals from photosynthetic or respiratory electrontransport chains are not at all prohibited by CORR Theessence of CORR is that there is a subset of organellarproteins for which there is an overriding requirement fora redox regulatory control that is initiated within theorganelle and whose effects are exerted by the simplestand most direct route This redox signalling is envisagedto take place without a requirement for prior consultationand coordination with what may be regarded as a nucleo-cytoplasmic bureaucracy
(v) Nuclear genes can be expressed as rapidly as chloroplastand mitochondrial genes
Zerges (2002) for example cites the equal times takenfor complete expression of nuclear and chloroplast genesin Chlamydomonas as evidence against the CORR hypoth-
32 J F Allen Genomes in mitochondria and chloroplasts
esis However the question is not one of the time takento express a gene but the time taken to alter the rate ofits expression in response to a (redox) signal originatingwithin the organelle An equally important additional fac-tor may be the autonomy of the signalling pathway thatis its independence from other inputs as described in theprevious paragraph One might say that mitochondria andchloroplasts require a free hand
(vi) There is no evidence to support this hypothesisThe hypothesis is rich in predictions (sectsect 3 and 6) and
those that have been tested stringently are in agreementwith experimental results None of the predictions of thehypothesis has been found to be incorrect and none of theunderlying principles (sect 3) has been disproved Furtherpredictions will be the subject of future experiments
(b) Evidence for CORR(i) Chloroplasts have selectively retained cyanobacterial genes
for primary events in photosynthesisSee table 1
(ii) There is regulation of gene expression in chloroplastsThere is transcriptional regulation of gene expression
in chloroplasts (sect 3d) The case for mitochondria is notyet proven
(iii) There is redox regulation of gene expression inchloroplasts
There is transcriptional redox regulation of geneexpression in chloroplasts (sect 3d) The case for mitochon-dria is not yet proven though see (Allen et al 1995 Kon-stantinov et al 1995 Galvis et al 1998)
(iv) There are no redox signalling components encoded inchloroplast or mitochondrial DNA
Correct but with possible exceptions discussed in sect 3j
(v) There are lsquobacterialrsquo redox signalling components ineukaryotes
See sect 3j
(vi) Further predictions DNA-array results and phenotypesof allotopic mutants
A systematic test of CORR might be to make an oxid-ized-minus-reduced gene expression profile of the lsquotran-scriptomersquo of a cyanobacterium to see if the set of geneswhose transcription is under most rapid and direct redoxcontrol corresponds to the genes of the chloroplast gen-ome Laboratories with this experimental capability do notseem to have addressed this possibility (Hihara et al 2002Ikeuchi 2002) Likewise if a putative modern representa-tive of the proto-mitochondrion can be found and agreedeg Rhodospirillum rubrum or Paracoccus denitrificans thena similar experiment should reveal a subset of genes corre-sponding to mitochondrial genomes DNA-microarraysalso present the possibility of testing the relative rate ofredox response and dependency on other signallinginputs of transcription of eukaryotic genes To dateorganellar genes have not been included in such investi-gations (Ferea et al 1999) A further experimental possi-bility is to examine the redox regulatory phenotypes ofexisting allotopic mutants as outlined in sect 3e
Phil Trans R Soc Lond B (2003)
6 CONCLUSION AND FUTURE PROSPECTS
The CORR hypothesis states that the function of mito-chondrial and chloroplast genomes is to provide co-location (of gene and gene product) for (evolutionary)continuity of redox regulation of gene expression Thishypothesis is neither self-contradictory nor devoid of test-able predictions It is also consistent with available evi-dence and in principle makes sense of the otherwisepuzzling distribution of genes between compartments ineukaryotic cells The explanation that CORR offersapplies equally to mitochondria and chloroplasts andidentifies selective forces that still operate to determine thelocation of genes No selective barriers need be assumedto gene transfer protein import nor membrane insertionNor do we need to invoke a lsquofrozen accidentrsquo that some-thing happened to stop gene transfer for some reason atsome time in the remote and inaccessible past CORR alsoexplains the absence of genomes from organelles that havelost their primary bioenergetic functions CORR may actas a stimulus to a search for residual bioenergetic functionin those organelles that have lost aerobic respiration andphotosynthesis but where residual genomes remain
CORR also predicts regulatory properties of knowncomponents of chloroplasts and mitochondria that in turnindicate important but as yet uncharacterized flexibility inenergy transduction These components include the RUB-ISCO large subunit and the CFo and Fo subunits of coup-ling ATPase in both chloroplasts and mitochondriaRedox regulation of the relative stoichiometry of compo-nents as seen in regulation of chloroplast photosystemstoichiometry (Pfannschmidt et al 1999a) may extend to(C)Fo-(C)F1-ATPase indicating that the function ofretention of genes for one or more of these componentsis flexibility in the stoichiometries of H1 to e2 and H1 toATP in chemiosmotic coupling (Schemidt et al 1998Stock et al 2000 Allen 2002)
So far explicit tests of CORR have largely been con-fined to chloroplasts and to some extent mitochondria ofhigher plants but this merely reflects the expertise andinterests of the investigators Further attempted disproofof CORR is welcomed and encouraged Novel experi-mental tests can be devised as well as attempted refu-tation through analysis of existing data It is hoped thatexplicit attention will be given to the CORR hypothesis inlaboratories already suitably equipped and that resourcescan be allocated for this purpose in laboratories wherethere is the interest but not currently the technology orexpertise
The heuristic value of the hypothesis already seen inthe role of redox-regulated transcription of photosyntheticreaction centre genes in chloroplasts may be its most use-ful property The central proposal is that mitochondriaand chloroplasts contain genes for proteins whose functionin electron transfer demands rapid direct and uncon-ditional redox regulatory control of their biosynthesis Thefull implications of this hypothesis are very broad and onlyjust beginning to be explored
I thank Carol Allen Aubrey de Grey William Martin John ARaven and Francis-Andre Wollman for discussion and corre-spondence during the preparation of this paper and WilliamMartin for suggestions including table 1 Research grants from
Genomes in mitochondria and chloroplasts J F Allen 33
The Swedish Natural Sciences Research Council (NFR) andthe Crafoord Foundation are gratefully acknowledged
REFERENCES
Adams K L Daley D O Qiu Y L Whelan J amp PalmerJ D 2000 Repeated recent and diverse transfers of a mito-chondrial gene to the nucleus in flowering plants Nature408 354ndash357
Alberts B Bray D Lewis J Raff M Roberts K amp Wat-son J D 1994 The molecular biology of the cell New Yorkand London Garland Publishing
Allen C A Hakansson G amp Allen J F 1995 Redox con-ditions specify the proteins synthesized by isolated chloro-plasts and mitochondria Redox Rep 1 119ndash123
Allen J F 1992 Protein phosphorylation in regulation ofphotosynthesis Biochim Biophys Acta 1098 275ndash335
Allen J F 1993a Control of gene expression by redox poten-tial and the requirement for chloroplast and mitochondrialgenomes J Theor Biol 165 609ndash631
Allen J F 1993b Redox control of gene expression and thefunction of chloroplast genomes a hypothesis PhotosynthRes 36 95ndash102
Allen J F 1993c Redox control of transcription sensorsresponse regulators activators and repressors FEBS Lett332 203ndash207
Allen J F 1995 Thylakoid proteinndashphosphorylation state-1ndashstate-2 transitions and photosystem stoichiometry adjust-ment redox control at multiple levels of gene expressionPhysiol Plant 93 196ndash205
Allen J F 1996 Separate sexes and the mitochondrial theoryof ageing J Theor Biol 180 135ndash140
Allen J F 2002 Photosynthesis of ATP-electrons protonpumps rotors and poise Cell 110 273ndash276
Allen J F amp Allen C A 1999 A mitochondrial model forpremature ageing of somatically cloned mammals IUBMBLife 48 369ndash372
Allen J F amp Nilsson A 1997 Redox signalling and the struc-tural basis of regulation of photosynthesis by protein phos-phorylation Physiol Plant 100 863ndash868
Allen J F amp Pfannschmidt T 2000 Balancing the two photo-systems photosynthetic electron transfer governs transcrip-tion of reaction centre genes in chloroplasts Phil TransR Soc Lond B 355 1351ndash1357 (DOI 101098rstb20000697)
Allen J F amp Race H L 2002 Will the real LHC II kinaseplease step forward Sciencersquos STKE httpwwwstkeorgcgicontentfullsigtrans2002155pe43
Allen J F amp Raven J A 1996 Free-radical-induced mutationvs redox regulation costs and benefits of genes in organellesJ Mol Evol 42 482ndash492
Archibald J M Cavalier-Smith T Maier U amp Douglas S2001 Molecular chaperones encoded by a reduced nucleusthe cryptomonad nucleomorph J Mol Evol 52 490ndash501
Ashby M K amp Mullineaux C W 1999a Cyanobacterialycf27 gene products regulate energy transfer from phycobili-somes to photosystems I and II FEMS Microbiol Lett 181253ndash260
Ashby M K amp Mullineaux C W 1999b The role of ApcDand ApcF in energy transfer from phycobilisomes to PSI andPSII in a cyanobacterium Photosynth Res 61 169ndash179
Ashby M K Houmard J amp Mullineaux C W 2002 Theycf27 genes from cyanobacteria and eukaryotic algae distri-bution and implications for chloroplast evolution FEMSMicrobiol Lett 214 25ndash30
Attardi G amp Schatz G 1988 Biogenesis of mitochondria ARev Cell Biol 4 289ndash333
Azem A Oppliger W Lustig A Jeno P Feifel B SchatzG amp Horst M 1997 The mitochondrial hsp70 chaperone
Phil Trans R Soc Lond B (2003)
system effect of adenine nucleotides peptide substrate andmGrpE on the oligomeric state of mhsp70 J Biol Chem272 20 901ndash20 906
Barbrook A C Symington H Nisbet R E R LarkumA amp Howe C J 2001 Organisation and expression of theplastid genome of the dinoflagellate Amphidinium opercula-tum Mol Genet Genomics 266 632ndash638
Bauer C E Elsen S amp Bird T H 1999 Mechanisms forredox control of gene expression A Rev Microbiol 53495ndash523
Bauer C E Elsen S Swem L R Swem D L amp MasudaS 2003 Redox and light regulation of gene expression inphotosynthetic prokaryotes Phil Trans R Soc Lond B 358147ndash154 (DOI 101098rstb20021189)
Berks B C Sargent F De Leeuw E Hinsley A P Stan-ley N R Jack R L Buchanan G amp Palmer T 2000aA novel protein transport system involved in the biogenesisof bacterial electron transfer chains Biochim Biophys ActaBioenerg 1459 325ndash330
Berks B C Sargent F amp Palmer T 2000b The tat proteinexport pathway Mol Microbiol 35 260ndash274
Blackstone N W 1999 Redox control in development andevolution evidence from colonial hydroids J Exp Biol 2023541ndash3553
Blackstone N W 2000 Redox control and the evolution ofmulticellularity Bioessays 22 947ndash953
Blackstone N W 2001 Redox state reactive oxygen speciesand adaptive growth in colonial hydroids J Exp Biol 2041845ndash1853
Bogorad L 1975 Evolution of organelles and eukaryotic gen-omes Science 188 891ndash898
Bogsch E G Sargent F Stanley N R Berks B C Robin-son C amp Palmer T 1998 An essential component of a novelbacterial protein export system with homologues in plastidsand mitochondria J Biol Chem 273 18 003ndash18 006
Brown E C Somanchi A amp Mayfield S P 2001 Interor-ganellar crosstalk new perspectives on signaling from thechloroplast to the nucleus Genome Biol 2 10211ndash10214
Cavalier-Smith T 2002 Chloroplast evolution secondarysymbiogenesis and multiple losses Curr Biol 12 R62ndashR64
Chitnis P R amp Thornber J P 1988 The major light-har-vesting complex of photosystem 2 Aspects of its molecularand cell biology Photosynth Res 16 41ndash63
Chow W S Melis A amp Anderson J M 1990 Adjustmentsof photosystem stoichiometry in chloroplasts improve thequantum efficiency of photosynthesis Proc Natl Acad SciUSA 87 7502ndash7506
Claros M G Perea J Shu Y Samatey F A PopotJ L amp Jacq C 1995 Limitations to in vivo import of hydro-phobic proteins into yeast mitochondria The case of a cyto-plasmically synthesized apocytochrome b Eur J Biochem228 762ndash771
Dalbey R E amp Robinson C 1999 Protein translocation intoand across the bacterial plasma membrane and the plant thy-lakoid membrane Trends Biochem Sci 24 17ndash22
Daley D O Adams K L Clifton R Qualmann S MillarA H Palmer J D Pratje E amp Whelan J 2002 Genetransfer from mitochondrion to nucleus novel mechanismsfor gene activation from Cox2 Plant J 30 11ndash21
Danon A amp Mayfield S P 1994 Light-regulated translationof chloroplast messenger RNAs through redox potentialScience 266 1717ndash1719
de Grey A 2000 Mitochondrial gene therapy an arena for thebiomedical use of inteins Trends Biotechnol 18 394ndash399
Deisenhofer J Epp O Miki K Huber R amp Michel H1985 Structure of the protein subunits in the photosyntheticreaction center of Rhodopseudomonas viridis at 3Ac resolutionNature 318 618ndash624
34 J F Allen Genomes in mitochondria and chloroplasts
Delwiche C F 1999 Tracing the thread of plastid diversitythrough the tapestry of life Am Nat 154 S164ndashS177
Doolittle W F 1999 Phylogenetic classification and the uni-versal tree Science 284 2124ndash2129
Douglas S E 1998 Plastid evolution origins diversitytrends Curr Opin Genet Dev 8 655ndash661
Duby G amp Boutry M 2002 Mitochondrial protein importmachinery and targeting information Plant Sci 162 477ndash490
Eberhard S Drapier D amp Wollman F A 2002 Searchinglimiting steps in the expression of chloroplast encoded pro-teins relations between gene copy number transcriptiontranscript abundance and translation rate in the chloroplastof Chlamydomonas reinhardtii Plant J 31 149ndash160
El Bissati K amp Kirilovsky D 2001 Regulation of psbA andpsaE expression by light quality in Synechocystis species PCC6803 A redox control mechanism Plant Physiol 1251988ndash2000
Ellis R J 1984 The nuclear domination of chloroplast devel-opment Science Progress 69 129ndash142
Embley T M amp Martin W 1998 Molecular evolution ahydrogen-producing mitochondrion Nature 396 517ndash519
Ems S C Morden C W Dixon C K Wolfe K HdePamphilis C W amp Palmer J D 1995 Transcriptionsplicing and editing of plastid RNAs in the non-photosyn-thetic plant Epifagus virginiana Plant Mol Biol 29 721ndash733
Eraso J M amp Kaplan S 2000 From redox flow to gene regu-lation role of the PrrC protein of Rhodobacter sphaeroides241 Biochemistry 39 2052ndash2062
Eraso J M amp Kaplan S 2002 Redox flow as an instrumentof gene regulation Meth Enzymol 348 216ndash229
Escoubas J M Lomas M Laroche J amp Falkowski P G1995 Light-intensity regulation of cab gene-transcription issignaled by the redox state of the plastoquinone pool ProcNatl Acad Sci USA 92 10 237ndash10 241
Ferea T L Botstein D Brown P O amp Rosenzweig R F1999 Systematic changes in gene expression patterns follow-ing adaptive evolution in yeast Proc Natl Acad Sci USA96 9721ndash9726
Forsberg J Rosenquist M Fraysse L amp Allen J F 2001Redox signalling in chloroplasts and mitochondria genomicand biochemical evidence for two-component regulatory sys-tems in bioenergetic organelles Biochem Soc Trans 29403ndash407
Fujita Y 1997 A study on the dynamic features of photosys-tem stoichiometry accomplishments and problems forfuture studies Photosynth Res 53 83ndash93
Gabriel K Buchanan S K amp Lithgow T 2001 The alphaand the beta protein translocation across mitochondrial andplastid outer membranes Trends Biochem Sci 26 36ndash40
Galvis M L E Allen J F amp Hakansson G 1998 Proteinsynthesis by isolated pea mitochondria is dependent on theactivity of respiratory complex II Curr Genet 33 320ndash329
Gatenby A A amp Ellis R J 1990 Chaperone function theassembly of ribulose bisphosphate carboxylase-oxygenase ARev Cell Biol 6 125ndash149
GoldschmidtClermont M 1998 Coordination of nuclear andchloroplast gene expression in plant cells Int Rev CytolSurv Cell Biol 177 115ndash180
Gray M W 1992 The endosymbiont hypothesis revisited IntRev Cytol 141 233ndash357
Gray M W 1999 Evolution of organellar genomes CurrOpin Genet Dev 9 678ndash687
Gray M W 2000 Evolutionary biology mitochondrial geneson the move Nature 408 302ndash305
Gray M W amp Doolittle W F 1982 Has the endosymbionthypothesis been proven Microbiol Rev 46 1ndash42
Gray M W Burger G amp Lang B F 1999 Mitochondrialevolution Science 283 1476ndash1481
Phil Trans R Soc Lond B (2003)
Grebe T W amp Stock J B 1999 The histidine protein kinasesuperfamily Adv Microb Physiol 41 139ndash227
Green B R amp Kuhlbrandt W 1995 Sequence conservationof light-harvesting and stress-response proteins in relation tothe 3-dimensional molecular structure of Lhcii PhotosynthRes 44 139ndash148
Gruissem W amp Tonkyn J C 1993 Control mechanisms ofplastid gene expression Crit Rev Plant Sci 12 19ndash55
Hackstein J H P (and 11 others) 2001 Hydrogenosomesconvergent adaptations of mitochondria to anaerobicenvironments Zool Anal Complex Syst 104 290ndash302
Harrison M A Keen J N Findlay J B C amp Allen J F1990 Modification of a Glnb-like gene-product by photosyn-thetic electron-transport in the cyanobacterium Synechoc-occus 6301 FEBS Lett 264 25ndash28
Harvey A J Kind K L amp Thompson J G 2002 Redoxregulation of early embryo development Reproduction 123479ndash486
Heathcote P Fyfe P K amp Jones M R 2002 Reactioncentres the structure and evolution of biological solarpower Trends Biochem Sci 27 79ndash87
Henze K amp Martin W 2001 How do mitochondrial genesget into the nucleus Trends Genet 17 383ndash387
Herrmann R G amp Westhoff P 2001 Thylakoid biogenesis anddynamics the result of a complex phylogenetic puzzle InRegulation of photosynthesis vol 11 (ed E-M Aro amp BAndersson) pp 1ndash28 Dordrecht The Netherlands Kluwer
Hihara Y Sonoike K Kanehisa M amp Ikeuchi M 2002Search for redox responsive genes in Synechocystis sp PCC6803 by using DNA microarrays Plant Cell Physiol 43S178
Howe C J Barbrook A C amp Lockhart P J 2000 Organellegenes do they jump or are they pushed Trends Genet 1665ndash66
Ikeuchi M 2002 Gene expression and responses to light stressin cyanobacteria Plant Cell Physiol 43 S21
Iuchi S amp Lin E C C 1992 Mutational analysis of signaltransduction by Arcb a membrane sensor protein respon-sible for anaerobic repression of operons involved in the cen-tral aerobic pathways in Escherichia coli J Bacteriol 1743972ndash3980
Jackson-Constan D Akita M amp Keegstra K 2001 Molecu-lar chaperones involved in chloroplast protein importBiochim Biophys Acta Mol Cell Res 1541 102ndash113
Jarvis P amp Soll M 2001 Toc Tic and chloroplast proteinimport Biochim Biophys Acta Mol Cell Res 1541 64ndash79
Jordan P Fromme P Witt H T Klukas O SaengerW amp Krauss N 2001 Three-dimensional structure ofcyanobacterial photosystem I at 25 acngstrom resolutionNature 411 909ndash917
Kaneko T (and 23 others) 1996a Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regionsDNA Res 3 109ndash136
Kaneko T (and 23 others) 1996b Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regions[Supplement] DNA Res 3 185ndash209
Kanevski I amp Maliga P 1994 Relocation of the plastid rbcLgene to the nucleus yields functional ribulose-15-bisphosph-ate carboxylase in tobacco chloroplasts Proc Natl Acad SciUSA 91 1969ndash1973
Kim J amp Mayfield S P 1997 Protein disulfide isomerase as aregulator of chloroplast translational activation Science 2781954ndash1957
Genomes in mitochondria and chloroplasts J F Allen 35
Kirk J T O amp Tilney-Bassett R A E 1978 The plastidsTheir chemistry structure growth and inheritance Amster-dam Elsevier
Koehler C M Merchant S amp Schatz G 1999 How mem-brane proteins travel across the mitochondrial intermem-brane space Trends Biochem Sci 24 428ndash432
Komiya T Rospert S Schatz G amp Mihara K 1997 Bind-ing of mitochondrial precursor proteins to the cytoplasmicdomains of the import receptors Tom70 and Tom20 isdetermined by cytoplasmic chaperones FASEB J 11 1196
Konstantinov Y M Lutsenko G N amp Podsosonny V A1995 Genetic functions of isolated maize mitochondriaunder model changes of redox conditions Biochem MolBiol Int 36 319ndash326
Korab-Laskowska M Rioux P Brossard N LittlejohnT G Gray M W Lang B F amp Burger G 1998 Theorganelle genome database project (GOBASE) Nucleic AcidsRes 26 138ndash144
Kovacs L Wiessner W Kis M Nagy F Mende D ampDemeter S 2000 Short- and long-term redox regulation ofphotosynthetic light energy distribution and photosystemstoichiometry by acetate metabolism in the green alga Chla-mydobotrys stellata Photosynth Res 65 231ndash247
Kuhlbrandt W 1994 Structure and function of the plant light-harvesting complex LHC-II Curr Opin Struct Biol 4519ndash528
Lang B F Gray M W amp Burger G 1999 Mitochondrialgenome evolution and the origin of eukaryotes A RevGenet 33 351ndash397
Lemire B D Fankhauser C Baker A amp Schatz G 1989The mitochondrial targeting function of randomly generatedpeptide sequences correlates with predicted helical amphi-philicity J Biol Chem 264 20 206ndash20 215
Leuenberger D Bally N A Schatz G amp Koehler C M1999 Different import pathways through the mitochondrialintermembrane space for inner membrane proteins EMBOJ 18 4816ndash4822
Li H amp Sherman L A 2000 A redox-responsive regulator ofphotosynthesis gene expression in the cyanobacterium Syne-chocystis sp strain PCC 6803 J Bacteriol 182 4268ndash4277
Lin X Y (and 36 others) 1999 Sequence and analysis of chro-mosome 2 of the plant Arabidopsis thaliana Nature 402761ndash768
Lonsdale D M Hodge T P Howe C J amp Stern D B1983 Maize mitochondrial-DNA contains a sequence hom-ologous to the ribulose-15-bisphosphate carboxylase largesubunit gene of chloroplast DNA Cell 34 1007ndash1014
McFadden G I 1999 Endosymbiosis and evolution of theplant cell Curr Opin Plant Biol 2 513ndash519
McFadden G I 2001 Chloroplast origin and integrationPlant Physiol 125 50ndash53
Mahler H R amp Raff R A 1975 The evolutionary origin ofthe mitochondrion a nonsymbiotic model Int Rev Cytol43 1ndash124
Manfredi G Fu J Ojaimi J Sadlock J E Kwong J QGuy J amp Schon E A 2002 Rescue of a deficiency in ATPsynthesis by transfer of MTATP6 a mitochondrial DNA-encoded gene to the nucleus Nature Genet 30 394ndash399
Marc P Margeot A Devaux F Blugeon C Corral-Debrinski M amp Jacq C 2002 Genome-wide analysis ofmRNAs targeted to yeast mitochondria EMBO Rep 3159ndash164
Marienfeld J Unseld M amp Brennicke A 1999 The mito-chondrial genome of Arabidopsis is composed of both nativeand immigrant information Trends Plant Sci 4 495ndash502
Martin W 1999 A briefly argued case that mitochondria andplastids are descendants of endosymbionts but that thenuclear compartment is not Proc R Soc Lond B 2661387ndash1395 (DOI 101098rspb19990792)
Phil Trans R Soc Lond B (2003)
Martin W amp Herrmann R G 1998 Gene transfer fromorganelles to the nucleus how much what happens andwhy Plant Physiol 118 9ndash17
Martin W amp Muller M 1998 The hydrogen hypothesis forthe first eukaryote Nature 392 37ndash41
Martin W amp Russell M J 2003 On the origins of cells ahypothesis for the evolutionary transitions from abiotic geo-chemistry to chemoautotrophic prokaryotes and from pro-karyotes to nucleated cells Phil Trans R Soc Lond B 35859ndash85 (DOI 101098rstb20021183)
Martin W Stoebe B Goremykin V Hansmann S Hase-gawa M amp Kowallik K V 1998 Gene transfer to thenucleus and the evolution of chloroplasts Nature 393162ndash165
Martin W Hoffmeister M Rotte C amp Henze K 2001 Anoverview of endosymbiotic models for the origins of eukary-otes their ATP-producing organelles (mitochondria andhydrogenosomes) and their heterotrophic lifestyle BiolChem 382 1521ndash1539
Martin W Rujan T Richly E Hansen A Cornelsen SLins T Leister D Stoebe B Hasegawa M amp PennyD 2002 Evolutionary analysis of Arabidopsis cyanobacterialand chloroplast genomes reveals plastid phylogeny and thou-sands of cyanobacterial genes in the nucleus Proc NatlAcad Sci USA 99 12 246ndash12 251
Millen R S (and 12 others) 2001 Many parallel losses of infAfrom chloroplast DNA during angiosperm evolution withmultiple independent transfers to the nucleus Plant Cell 13645ndash658
Morse D Salois P Markovic P amp Hastings J W 1995 Anuclear-encoded form-II RUBISCO in dinoflagellatesScience 268 1622ndash1624
Murakami A Kim S J amp Fujita Y 1997 Changes in photo-system stoichiometry in response to environmental con-ditions for cell growth observed with the cyanophyteSynechocystis PCC 6714 Plant Cell Physiol 38392ndash397
Oh J I amp Kaplan S 2000 Redox signaling globalization ofgene expression EMBO J 19 4237ndash4247
Ozawa T 1995 Mitochondrial-DNA mutations associatedwith aging and degenerative diseases Exp Gerontol 30269ndash290
Palmer J D 1997 Organelle genomes going going goneScience 275 790ndash791
Pearson C K Wilson S B Schaffer R amp Ross A W 1993NAD turnover and utilization of metabolites for RNA syn-thesis in a reaction sensing the redox state of the cyto-chrome-b( 6 )f complex in isolated chloroplasts Eur JBiochem 218 397ndash404
Pfanner N amp Geissler A 2001 Versatility of the mitochon-drial protein import machinery Natl Rev Mol Cell Biol 2339ndash349
Pfanner N amp Wiedemann N 2002 Mitochondrial proteinimport two membranes three translocases Curr Opin CellBiol 14 400ndash411
Pfannschmidt T Nilsson A amp Allen J F 1999a Photosyn-thetic control of chloroplast gene expression Nature 397625ndash628
Pfannschmidt T Nilsson A Tullberg A Link G amp AllenJ F 1999b Direct transcriptional control of the chloroplastgenes psbA and psaAB adjusts photosynthesis to light energydistribution in plants IUBMB Life 48 271ndash276
Pfannschmidt T Allen J F amp Oelmuller R 2001a Prin-ciples of redox control in photosynthesis gene expressionPhysiol Plant 112 1ndash9
Pfannschmidt T Schutze K Brost M amp Oelmuller R2001b A novel mechanism of nuclear photosynthesis generegulation by redox signals from the chloroplast during pho-
36 J F Allen Genomes in mitochondria and chloroplasts
tosystem stoichiometry adjustment J Biol Chem 27636 125ndash36 130
Popot J L amp de Vitry C 1990 On the microassembly of inte-gral membrane proteins A Rev Biophys Biophys Chem 19369ndash403
Race H L Herrmann R G amp Martin W 1999 Why haveorganelles retained genomes Trends Genet 15 364ndash370
Raven J A Johnston A M Parsons R amp Kubler J 1994aThe influence of natural and experimental high O2 concen-trations on O2 evolving phototrophs Biol Rev CambridgePhil Soc 69 61ndash94
Raven J A Johnston A M Parsons R amp Kubler J 1994bThe occurrence and influence on photolithotrophs of highoxygen concentrations Proc R Soc Edinb B 102 193ndash201
Robinson C 2000 The twin-arginine translocation system anovel means of transporting folded proteins in chloroplastsand bacteria Biol Chem 381 89ndash93
Robinson C amp Bolhuis A 2001 Protein targeting by the twin-arginine translocation pathway Natl Rev Mol Cell Biol 2350ndash356
Rodermel S 2001 Pathways of plastid-to-nucleus signalingTrends Plant Sci 6 471ndash478
Roger A J 1999 Reconstructing early events in eukaryoticevolution Am Nat 154 S146ndashS163
Roger A J Svard S G Tovar J Clark C G SmithM W Gillin F D amp Sogin M L 1998 A mitochondrial-like chaperonin 60 gene in Giardia lamblia evidence thatdiplomonads once harbored an endosymbiont related to theprogenitor of mitochondria Proc Natl Acad Sci USA 95229ndash234
Roodyn D B amp Wilkie D 1968 The biogenesis of mitochondriaMethuenrsquos monographs on biological subjects London Methuen
Rujan T amp Martin W 2001 How many genes in Arabidopsiscome from cyanobacteria An estimate from 386 proteinphylogenies Trends Genet 17 113ndash120
Saccone C Gissi C Lanave C Larizza A Pesole G ampReyes A 2000 Evolution of the mitochondrial genetic sys-tem an overview Gene 261 153ndash159
Salzberg S L White O Peterson J amp Eisen J A 2001Microbial genes in the human genome lateral transfer orgene loss Science 292 1903ndash1906
Sargent F Berks B C amp Palmer T 2002 Assembly of mem-brane-bound respiratory complexes by the Tat protein-trans-port system Arch Microbiol 178 77ndash84
Schatz G 1998 Protein transport the doors to organellesNature 395 439ndash440
Schemidt R A Qu J Williams J R amp Brusilow W S1998 Effects of carbon source on expression of F0 genes andon the stoichiometry of the c subunit in the F1 F0 ATPaseof Escherichia coli J Bacteriol 180 3205ndash3208
Selosse M A Albert B R amp Godelle B 2001 Reducing thegenome size of organelles favours gene transfer to thenucleus Trends Ecol Evol 16 135ndash141
Stern D B amp Lonsdale D M 1982 Mitochondrial and chlo-roplast genomes of maize have a 12- kilobase DNA-sequencein common Nature 299 698ndash702
Stock D Gibbons C Arechaga I Leslie A G W ampWalker J E 2000 The rotary mechanism of ATP synthaseCurr Opin Struct Biol 10 672ndash679
Stoebe B amp Maier U G 2002 One two three naturersquos toolbox for building plastids Protoplasma 219 123ndash130
Stoebe B Martin W amp Kowallik K V 1998 Distributionand nomenclature of protein-coding genes in 12 sequencedchloroplast genomes Plant Mol Biol Rep 16 243ndash255
Stupar R M Lilly J W Town C D Cheng Z Kaul SBuell C R amp Jiang J M 2001 Complex mtDNA consti-tutes an approximate 620 kb insertion on Arabidopsis thali-ana chromosome 2 implication of potential sequencing
Phil Trans R Soc Lond B (2003)
errors caused by large-unit repeats Proc Natl Acad SciUSA 98 5099ndash5103
Surpin M Larkin R M amp Chory J 2002 Signal transduc-tion between the chloroplast and the nucleus Plant Cell 14S327ndashS338
Tjaden J Schwoppe C Mohlmann T Quick P W ampNeuhaus H E 1998 Expression of a plastidic ATPADPtransporter gene in Escherichia coli leads to a functional aden-ine nucleotide transport system in the bacterial cytoplasmicmembrane J Biol Chem 273 9630ndash9636
Tovar J Fischer A amp Clark C G 1999 The mitosome a novelorganelle related to mitochondria in the amitochondrial para-site Entamoeba histolytica Mol Microbiol 32 1013ndash1021
Trebitsh T amp Danon A 2001 Translation of chloroplastpsbA mRNA is regulated by signals initiated by both photos-ystems II and I Proc Natl Acad Sci USA 98 12 289ndash12 294
Tullberg A Alexciev K Pfannschmidt T amp Allen J F2000 Photosynthetic electron flow regulates transcription ofthe psaB gene in pea (Pisum sativum L) chloroplasts throughthe redox state of the plastoquinone pool Plant Cell Physiol41 1045ndash1054
Turmel M Lemieux C Burger G Lang B F Otis CPlante I amp Gray M W 1999 The complete mitochondrialDNA sequences of Nephroselmis olivacea and Pedinomonasminor two radically different evolutionary patterns withingreen algae Plant Cell 11 1717ndash1729
Unden G amp Bongaerts J 1997 Alternative respiratory path-ways of Escherichia coli energetics and transcriptional regu-lation in response to electron acceptors Biochim BiophysActa Bioenerg 1320 217ndash234
Unseld M Marienfeld J R Brandt P amp Brennicke A1997 The mitochondrial genome of Arabidopsis thaliana con-tains 57 genes in 366 924 nucleotides Nature Genet 1557ndash61
Van der Giezen M Slotboom D J Horner D S DyalP L Harding M Xue G P Embley T M amp KunjiE R S 2002 Conserved properties of hydrogenosomal andmitochondrial ADPATP carriers a common origin for bothorganelles EMBO J 21 572ndash579
Von Heijne G 1986 Why mitochondria need a genome FEBSLett 198 1ndash4
Westphal S Soll J amp Vothknecht U C 2001 A vesicle trans-port system inside chloroplasts FEBS Lett 506 257ndash261
Whitehouse H L K 1969 Towards an understanding of themechanism of heredity London Edward Arnold Ltd
Williams B A P Hirt R P Lucocq J M amp Embley T M2002 A mitochondrial remnant in the microsporidian Trachi-pleistophora hominis Nature 418 865ndash869
Wilson R J (and 10 others) 1996a Complete gene map of theplastid-like DNA of the malaria parasite Plasmodium falcipa-rum J Mol Biol 261 155ndash172
Wilson S B Davidson G S Thomson L M amp PearsonC K 1996b Redox control of RNA synthesis in potato mito-chondria Eur J Biochem 242 81ndash85
Winkler H H amp Neuhaus H E 1999 Non-mitochondrialATP transport Trends Biochem Sci 24 64ndash68
Wolfe K H Morden C W amp Palmer J D 1992 Small sin-gle-copy region of plastid DNA in the non-photosyntheticangiosperm Epifagus virginiana contains only two genes Dif-ferences among dicots monocots and bryophytes in geneorganization at a non-bioenergetic locus J Mol Biol 22395ndash104
Wollman F A Minai L amp Nechushtai R 1999 The biogen-esis and assembly of photosynthetic proteins in thylakoidmembranes Biochim Biophys Acta Bioenerg 1411 21ndash85
Yeliseev A A amp Kaplan S 2000 TspO of Rhodobactersphaeroides a structural and functional model for the mam-
Genomes in mitochondria and chloroplasts J F Allen 37
malian peripheral benzodiazepine receptor J Biol Chem275 5657ndash5667
Zerges W 2002 Does complexity constrain organelle evol-ution Trends Plant Sci 7 175ndash182
Zhang Z D Green B R amp Cavalier-Smith T 1999 Singlegene circles in dinoflagellate chloroplast genomes Nature400 155ndash159
Zhang Z D Cavalier-Smith T amp Green B R 2001 A fam-ily of selfish minicircular chromosomes with jumbled chloro-plast gene fragments from a dinoflagellate Mol Biol Evol18 1558ndash1565
Zouni A Witt H T Kern J Fromme P Krauss NSaenger W amp Orth P 2001 Crystal structure of photosys-tem II from Synechococcus elongatus at 38 acngstrom resol-ution Nature 409 739ndash743
DiscussionD S Horner (Dipartimento di Fisiologia e Biochimica
Generali University of Milan Milan Italy) Given thatthere are components of these membrane-bound energy-transduction pathways that are consistently nuclear enco-ded not the key ones you pointed out but others do youexpect perhaps a slower perhaps a more crude but yet aredox communication between the organelle andexpression of nuclear-encoded genes
J F Allen There is and there is no question about itI think that Bogoradrsquos lock-in hypothesis has somethingto say here and there are other people who say it is all todo with assembling multi-subunit complexes If you havecontrol over the initial construction of the core aroundwhich the whole complex is eventually built then redoxcontrol or some sort of control over synthesis of that corewill control construction of the whole thing eventuallyand I think that may well be largely correct I think it ispossible in principle for nuclear genes to make precursorsaccording to redox instructions but it is not really soimportant that those precursors are here and now presentin exactly the right quantities because the precursors canto a limited extent hang around and wait Actually thereis proteolytic degradation of unattached precursors and soon but I think the fact that these chloroplast and mito-chondrially DNA-encoded subunits are at the core ofthese energy-transducing complexes is the crucial issue
T Cavalier-Smith (Department of Zoology University ofOxford Oxford UK ) In the case of mitochondria the sub-set of genes that remains is much more variable than youimplied It runs from three proteins in the minimal casesup to I think over 90 That is a big difference and manyof the genes in different lineages of protists that remainhave nothing to do with redox control they are things likeribosomal proteins and so on So I think that even thoughwhat you say about redox control of certain genes mayactually be an important thing it is not the fundamentaloverall reason why the genomes are maintained It may bepart of the explanation for why certain genes are alwaysincluded in that subset but even in mitochondria I thinkthat cytochrome b is the only one that is always presentSo I think that both this great diversity of what has beenretained in different lineages and the nucleomorph caseindicate that evolution was able to do one thing in onelineage 500 million years ago and never got around todoing it for 500 million years in another lineage so thereis a lot more haphazardness and evolutionary accidentsand contingency than a straight functional interpretation
Phil Trans R Soc Lond B (2003)
there is a lot of just sheer lsquohappenedndashorndashdidnrsquotndashhappenrsquoor carryover
J F Allen I was not implying anything about the rangeof mitochondrial genomes I have a limited amount oftime so I just talked about what I could talk about withinthat limit Of course there will be a range and this willdepend upon the physiology and bioenergetics of theparticular organism But what I am trying to do is to seewhat the common properties of the irreducible core areof components that are always encoded in the organellesand cytochrome b is a perfect example because that reallyis the thing that binds these two all-important quinoneswithout which there is no Q cycle without which there isno proton pumping and bioenergetics and in Plasmodiumwhere the mitochondrion is reduced to this bare mini-mum there is still a mitochondrial gene for cytochromeb Now my argument about the ribosomal proteins andall the rest of it is that in order to be able to exert thisall-important redox control over gene expression of certaincore components you need an in situ genetic system andthis can vary in composition for reasons that I cannotdescribe Maybe you need RNA polymerase or maybeyou can import RNA polymerase In plastids you haveboth of these options You are going to need ribosomestransfer RNAs and the rest of the apparatus of geneexpression and that needs to be in situ but we are stillleft with the question why does there have to be any gen-etic system at all The mature tobacco or spinach chloro-plast seems to have an entire genetic system just to makeone protein D1 which is all it ever needs to resynthesizeI think that this is a clue It is worth having this very unec-onomical crazy system because there is an overridingrequirement to connect electron transfer with geneexpression directly and unambiguouslymdashwithout recourseto a bureaucracy if you like which is what I think thenucleus is
K Willison (Institute of Cancer Research Chester BeattyLaboratories London UK ) These photosynthetic reactioncomplexes are a sort of baroque mixture of polypeptidechains and cofactors and my understanding is that someof them in high photon flux can turn over within say 30minutes so I am wondering if there is a correlationbetween gene retention and the turnover rates of some ofthe components and the complexes it seems to me thatthat could really be a limiting factor and the reason thatthe proteins had to be encoded locally in order to get thecomplexes to be assembled rapidly
J F Allen I think that is right You can only alter thesteady-state concentration of a protein complex by affect-ing the synthesis and breakdown and therefore turnoverThere is an extreme example the D1 protein encoded bythe psbA gene of chloroplasts which is always in the chlor-oplast genome without exception High light will tend todegrade this protein and when the high light goes awaythere is an absolute need very quickly to bring it back upagain I think that is a very good example Whether thisargument applies to genes such as cytochrome oxidase Ido not know
C J Leaver (Department of Plant Sciences University ofOxford Oxford UK ) Your experiments tend to relatemainly to the gene expression side What about proteinRNA turnover the whole concept of redox control ofthatmdashdo we know anything
38 J F Allen Genomes in mitochondria and chloroplasts
J F Allen Well quite a lot is known about chloro-plasts There is redox control of messenger RNA stabilityfor this same D1 protein which has been described byStephen Mayfield and his co-workers The evidence is thatthe site of redox control is ferredoxin via thioredoxin andthe implication is that thioredoxin tells whether non-cyclicelectron transport is working at all When it is workingthere is a signal that says lsquoOK now continue with all theprotein synthesisrsquo It is not particularly specific but moreof a global effect I think this is what Mayfield argues butof course this is not inconsistent with my argument I haveconcentrated on quinone pools and medium-potentialredox regulatory control because that is where I can seesignals about the dangerous consequences of getting itwrong but there is control of other levels of geneexpression In mitochondria I am not so sure I wouldbounce the question back to you on that one
W Martin (Institute of Botany III Heinrich-Heine Univ-ersitat Dusseldorf Dusseldorf Germany) The nucleomorphgenome was cited as evidence in conflict with your modelbut is that true The nucleomorph genome is not in abioenergetic organelle in that sense It resides in the cyto-sol so does the size of the nucleomorph genome workagainst your model Is it really evidence against yourmodel
J F Allen I do not know why should it be evidenceI cannot explain why the nucleomorph has a genomeThat is why I clutched at the straw that it is storage fornucleic acid precursors
Phil Trans R Soc Lond B (2003)
T Cavalier-Smith The reason I introduced the nucleo-morph is not to say it is direct evidence against yourhypothesis but to show that there probably was no singlefunctional reason for the retention of every gene in everyorganelle It may be that there are specific physiologicalreasons for the retention of certain genes and thereforeyou might be right about a subset of genes but I verymuch doubt if there are specific physiological reasons forthe retention of every single gene that is retained differen-tially because odd things happen in evolution Somethings are easy and sometimes things are difficult Evol-ution is history
J F Allen Of course I agree with that I do not thinkI can explain everything but I do think I can explainsomething If I have done that then I am content
GLOSSARY
AAC ADPndashATP carrierCES control by epistasy of synthesisCORR co-location for redox regulationLHC light-harvesting complexPCC Pasteur culture collectionORF open reading frameRUBISCO ribulose-15-bisphosphate carboxylasendashoxygenaseTAT twin-arginine translocase
Visit httpwwwpubsroyalsocacuk to see an electronic appendix tothis paper

Genomes in mitochondria and chloroplasts J F Allen 29
teins of ca 27ndash30 kDa are quite water-insoluble and areusually fully extracted only in chloroformndashmethanol or inthe presence of detergents The Lhc gene family is diverse(Green amp Kuhlbrandt 1995) but most gene productsor Lhcp (light-harvesting chlorophyll protein) polypeptideshave three membrane-spanning hydrophobic a-helices(Kuhlbrandt 1994) It is interesting that three-helix Lhcproteins seem to be absent from extant cyanobacteria andchloroxybacteria (Green amp Kuhlbrandt 1995)
(ii) The five-helix rulePerhaps with the three-helix model of LHC II in mind
a variation of the hydrophobicity hypothesis is the empiri-cal lsquofive-helix rulersquo (Popot amp de Vitry 1990 Claros et al1995 Wollman et al 1999) which states that no intrinsicmembrane protein with five or more membrane-spanninghelices can be encoded in the nucleus Five is a good guessfor a threshold number of helices demanding organellar-encoding in the context of photosynthetic reaction centres(Heathcote et al 2002) where the earliest known struc-ture that of type II centres (Deisenhofer et al 1985) asin photosystem II (Zouni et al 2001) shows exactly fivehelices with type I centres containing 11 (Jordan et al2001) There are however many nuclearly encoded sub-units in both mitochondria and chloroplasts that havefewer than five membrane helices so the five-helix rulecan only describe a sufficient condition and not a neces-sary condition for nuclear encoding
It should also be noted that ADPndashATP carriers (AACs)are encoded in the nucleus AACs of the mitochondrialinner membrane have six transmembrane helices (van derGiezen et al 2002) while the those of the chloroplast innerenvelope (Winkler amp Neuhaus 1999) have 12 (Tjaden etal 1998)
With the counter-examples of nuclearly encoded AACswith six or 12 membrane-spanning helices many intrinsicmembrane proteins with fewer than five helices somenuclearly encoded and some not and with the RUBISCOlarge subunit as a major product of chloroplast geneexpression with no membrane domain at all the five-helixrule begins to seem rather like a case of special pleadingEven if there are indeed special mitigating circumstancesfor AACs (this argument is outlined later) the five-helixrule as with the hydrophobicity hypothesis of which it isa special case will disappear if it can be shown that pos-session of five or more membrane helices is required forsome other function such as that of CORR that deter-mines the organellar location of the gene concernedHydrophobicity is in some cases a feature of proteins thatare encoded in organelles but it is not in itself the issue
For chloroplasts the hydrophobicity hypothesis predictsretention of cyanobacterial genes in functional categoriesassociated with the chloroplast envelope and with trans-port just as much as with photosynthesis Table 1 showsthat this is not the case
A mechanism for the unimportability of hydrophobicproteins is rarely stated explicitly and it is rather left tothe imagination whether hydrophobic proteins might asit were get stuck halfway through the chloroplast envelopeor mitochondrial outer membrane whether transport can-not get started on a precursor protein that has alreadyspontaneously inserted into the membrane or whetherhydrophobic proteins will be unable to make the journey
Phil Trans R Soc Lond B (2003)
through the cytosol without precipitating there In thecytosol co-translational transport or membrane insertionwould in principle address the latter point but it seemsto be generally true that chloroplast and mitochondrialproteins are post-translationally inserted into the organel-lar outer membrane So the obvious ancillary questionabout unimportability is lsquowhy then cannot hydrophobicproteins be co-translationally imported into organellesrsquoThere is evidence in yeast for co-translational transportinto mitochondria of proteins thought to be derived fromthe original endosymbiont (Marc et al 2002) If there isany truth in hydrophobicity as a barrier to protein importthe CORR principle of unselective import (sect 3c) shouldbe easily refuted for example by a demonstration that noknown targeting presequence is capable of importing poly-phenylalanine encoded by polyU for in vitro import stud-ies
An additional proposal intended to account for theunimportability of some hydrophobic proteins and forimportability of others is suggested by A de Grey(personal communication) who argues that bacteriallyderived protein precursors imported into mitochondriacross the mitochondrial inner membrane completelybefore insertion into that membrane from the mitochon-drial matrix (the N-phase) This is held to be an obstacleto successful import of membrane-intrinsic proteins thatmight en route discharge the proton motive force By con-trast eukaryotically derived proteins can be inserted intothe mitochondrial inner membrane from the inter-mem-brane space (the P-phase) and then there is for somereason no similar uncoupling effect Thus adenylatetransporters or AACs can be nuclearly encoded whereascytochrome oxidase subunits for example cannot It isnot obvious why bacterially derived subunits such asthose of cytochrome oxidase did not acquire eukaryoticinsertion mechanisms Perhaps this acquisition is forbid-den by the need to conserve pre-existing routes ofassembly into multi-subunit complexes or perhaps theneed for insertion from the inside reflects a need to incor-porate a cofactor for example haem or ironndashsulphur clus-ters along the way
(iii) Unimportability of three dimensional structures andcofactors
Accumulating evidence on protein transport mech-anisms seems to make a specific obstacle to successfultransport of certain sorts of protein less and less plausibleespecially given the observation that precursors with N-terminal targeting sequences not mature functional poly-peptides themselves traverse membranes during biosynth-esis
Protein folding inside the living cell is invariably assistedby molecular chaperones (Gatenby amp Ellis 1990 Azem etal 1997 Komiya et al 1997 1990 Jackson-Constan et al2001) proteins that impede the formation of lsquoimproperrsquostructural interactions that will otherwise produce aggre-gation or other non-functional interactions Proteinimport mechanisms appear mostly to rely on specific mol-ecular chaperones for guided unfolding before membraneinsertion and refolding after translocation of the polypep-tide into its destined compartment (Gatenby amp Ellis 1990Azem et al 1997 Komiya et al 1997 Jackson-Constanet al 2001) Because polypeptides are mostly lsquothreadedrsquo
30 J F Allen Genomes in mitochondria and chloroplasts
through membranes no special constraint on transportshould be expected to arise from 3D structure or surfaceproperties of a protein
An exception to this unfolding rule may be providedby holoproteins containing cofactors that are transportedalong with the apoprotein presumably with a largely intact3D structure The lsquoTATrsquo (twin-arginine translocase) sys-tem (Bogsch et al 1998 Dalbey amp Robinson 1999 Berkset al 2000ab Robinson 2000 Robinson amp Bolhuis 2001Sargent et al 2002) works broadly along these linesthough its mechanism is unresolved TAT is relevant herebecause it adds transport of essentially complete holopro-teins to the cellrsquos repertoire of transport mechanisms andeven proteins that may not be allowed to unfold perhapsbecause of the toxicity of the free cofactor are not therebymade candidates for an unimportability rule One variantof the unimportability hypothesis is that transport canwork only by unfolding and dissociated cofactors may betoxic to the cell The bacterially derived TAT system(Bogsch et al 1998 Dalbey amp Robinson 1999 Berks etal 2000ab Sargent et al 2002) overcomes this argumentas would examples of cofactor-free polypeptides such as(C)Fo subunit c which are nevertheless generally encodedin organelles
The presence of a vesicular transport system in chloro-plasts (Westphal et al 2001) counts against the unim-portability hypothesis Vesicular transport between thechloroplast envelope and the thylakoid membrane shouldin principle allow any protein to be ferried across thechloroplast stroma Vesicles might allow transport inparticular of membrane proteins that would discharge theproton motive force if allowed to connect transiently theN and P aqueous phases on each side of a coupling mem-brane
Another observation that seems to count against unim-portability is the increased number of membrane barriersin plastids derived from secondary tertiary or higher-orderendosymbioses These plastids seem to have retained ifanything fewer genes rather than more as unimportabilitypredicts The nucleomorph the residual nucleus of theprimary host retains many genes for molecular chap-erones (Archibald et al 2001 Cavalier-Smith 2002) inagreement with the conclusion that protein import pro-ceeds by conserved and specialized mechanisms If eachnew import added to gain a higher-order endosymbiosisrequires additional targeting presequences then perhapsthere is an advantage in components of an import machin-ery not being imported themselves
It seems that unimportability is less easy an assumptionto make today than in previous years and the onus maynow be on its proponents to cite specific examples of pro-teins that can never be imported into chloroplasts or mito-chondria
(f ) Some genes cannot be moved(i) Differences in the mitochondrial genetic code
The genetic code is the specific relation between ribon-ucleotide triplets of codons and tRNAs that recognizethem and which thus determine which individual aminoacids if any will be incorporated into the growing poly-peptide chain Mitochondria show small departures fromthe otherwise universal genetic code (Attardi amp Schatz1988 Alberts et al 1994) UGA is a lsquostoprsquo codon in the
Phil Trans R Soc Lond B (2003)
universal code and in plant mitochondria but is the codonfor tryptophan in mitochondria of animals and fungiCUA means leucine everywhere except fungal mitochon-dria where it means threonine and AGA and AGU codefor arginine in the lsquouniversalrsquo code but serine in insectmitochondria and lsquostoprsquo in mammalian ones The signifi-cance of this heterogeneity in coding is most unlikely torest in the early origin of mitochondria as the lsquouniversalrsquocode is employed by bacteria and protists Furthermorethe diversity of departures from the lsquouniversalrsquo codeamong mitochondria of different eukaryotic lineages indi-cates they had independent origins A general rule formitochondrial genetic codes is that reliance on the thirdlsquowobblersquo position is decreased which allows all 20 proteinamino acids to be specified by fewer tRNAs (22 in animalmitochondria instead of 31 in the cytosol) This in turnmeans that mtDNA can specify more proteins for a givenlength of DNA or conversely that the mitochondrialgenome can be condensed or lsquostreamlinedrsquo Although thisarrangement does not explain the retention of mitochon-drial genomes and may be a secondary event in mitochon-drial evolution it almost certainly places an extra barrierin place of gene relocation As discussed previously how-ever there seems to be no single barrier that can explainthe retention of a relatively constant subset of mitochon-drial genes
There has clearly been a contraction of mitochondrialDNA especially in animals Animal mitochondrial DNAhas few if any non-coding sequences and animal mito-chondrial genetic codes require fewer tRNAs than the sin-gle code operating in the cytosol (Gray 1999 Gray et al1999 Selosse et al 2001) These may be devices todecrease the total frequency of mutations per mitochon-drial genome assuming a constant frequency per unitlength of mitochondrial DNA It has been proposed thata genersquos removal to the nucleus decreases its susceptibilityto free radical mutagenesis (Raven et al 1994ab Allen ampRaven 1996) If this is the case then the non-standardcode may be a further indication of an overriding need tomaintain certain genes within the mitochondrion What-ever the reason for the retention of genes in organelles ithad better be a good one bioenergetic organelles areclearly the wrong place to keep genetic information andgo to extraordinary lengths to offset the consequences ofit being there
(ii) Molecular refugeesA phenomenon related to differences in the genetic code
is seen in chloroplasts This is a bias in codon usage Thelsquomolecular refugeesrsquo hypothesis (Howe et al 2000) sug-gests that nuclear copies of what were once chloroplastgenes escape constraints on codon usage and thus thatwhat distinguishes chloroplast-encoded proteins is thattheir amino-acid composition is unusual enough for thebias in codon usage not to apply It is difficult to see whatphysico-chemical properties might be uniquely shared bythe amino acids of chloroplast-encoded proteins and inaddition there seem to be no lsquoforbiddenrsquo amino-acid resi-due in chloroplasts As with the departure from the univer-sal genetic code in mitochondria is seems likely that abias in codon usage reflects some secondary property ofchloroplast genomes and is not the primary reason fortheir retention In any case any restriction on gene move-
Genomes in mitochondria and chloroplasts J F Allen 31
ment will act in the wrong direction to explain why chloro-plasts have retained genomes at all provided there is noresulting obstacle to a chloroplast protein being success-fully encoded as a precursor in the nucleus
(iii) Genes are copied and replicated not relocated in onestep
If complete relocation is envisaged as proceeding by thesimultaneous appearance of a gene in one compartmentand its deletion from another then it is possible to imaginethat whatever the vector is there may be selectivity ingene transfer (Henze amp Martin 2001) For example ifviral incorporation replication and recombination in bothorganelle and nucleus were the mechanism of relocationthen successful gene transfer might depend on the abilityof the virus to complete the process in one step This inturn might depend on some property of the gene or ofthe protein it encodes if expression occurs at any stage inthe process of relocation An assumption of the CORRhypothesis favoured here is that genes are not movedphysically but merely copied after which either the copyin the original location or the copy in the new locationmay be lost as a result of selection A useful analogy maybe the action of copying files between computer storagevolumes In this analogy the copy is identical to the orig-inal and either may subsequently be deleted without lossof information
Because any gene that is not usefully expressed willundergo no selection to maintain a functional allele theprevailing location will be the one in which the functionalcopy of the gene resides The problem of which gene sur-vives is then subsumed into the problem of which is theselectively advantageous location for it It seems reason-able to assume that there wasmdashand ismdashan interim periodin which homologous genes in different compartments areessentially in competition to see which functions best Asoutlined above there is no obvious obstacle for copyingDNA sequences within the cell The physico-chemicalproperties of DNA are fairly uniform and not directlydependent on nucleotide sequence though organelles likeintra-cellular parasites seem to prefer A and T to G andC The answer to the question lsquowhich gene location func-tions bestrsquo may have nothing to do with the gene itselfnor directly with the structure and function of its geneproduct Instead the answer may have everything to dowith the required regulatory interaction between the geneproduct and expression of the gene that produces it
(g) CORRThis hypothesis has been propounded earlier (sect 3) and
will be summarized as follows Vectorial electron and pro-ton transfer exerts regulatory control over expression ofgenes encoding proteins directly involved in or affectingredox poise This regulatory coupling is indispensable foradaptation to environmental changes in chemical activitiesof oxygen light and CO2 and requires co-location of thesegenes with their gene products Organelles lsquomake theirown decisionsrsquo on the basis of environmental changesaffecting redox state The hypothesis is abbreviatedCORR for co-location for redox regulation or more com-pletely lsquoco-location (of gene and gene product) for(evolutionary) continuity of redox regulation of geneexpressionrsquo
Phil Trans R Soc Lond B (2003)
5 SUMMARY OF THE CURRENT STATUS OF THEHYPOTHESIS OF CORR
Q Why do mitochondria and chloroplasts have theirown genetic systems
A CORR co-location of genes and gene products per-mits direct and autonomous redox regulation of geneexpression
(a) Objections to CORR(i) There is no regulation of gene expression in chloroplasts
and mitochondriaThis is still commonly stated but incorrect See the prin-
ciple in sect 3d
(ii) There is no regulation of gene expression in chloroplastsand mitochondria at the level of transcription
For chloroplasts there is now clear evidence for a func-tionally intelligible redox control of transcription(Pfannschmidt et al 1999a) This invalidates the previouswidely held assumption that all control of chloroplast geneexpression is post-transcriptional (Gruissem amp Tonkyn1993 GoldschmidtClermont 1998) The evidence formitochondria is less clear but redox control of transcrip-tion certainly cannot be ruled out and there are indi-cations that it may occur (Pearson et al 1993Konstantinov et al 1995 Wilson et al 1996b) It mustalso be restated that transcriptional control is not neces-sary for CORR but serves as a compelling example of aconserved typically bacterial mode of response that oper-ates still in eukaryotic cytoplasmic organelles thus illus-trating the principle of continuity of redox control (sect 3d)Post-transcriptional redox regulation (Danon amp Mayfield1994 Kim amp Mayfield 1997 Brown et al 2001 Treb-itsh amp Danon 2001) is consistent with CORR and thereis nothing to forbid it being a primary mechanism thatsatisfies most or indeed all of the formal requirements ofthe hypothesis in particular cases
(iii) There are no redox signalling components encoded inchloroplast or mitochondrial DNA
This is what CORR predicts See the principle in sect 3j
(iv) There is redox control of nuclear gene expressionThis is correct and many examples are now known
especially for chloroplasts (Escoubas et al 1995 Kovacs etal 2000 Pfannschmidt et al 2001ab Surpin et al 2002)Redox signals from photosynthetic or respiratory electrontransport chains are not at all prohibited by CORR Theessence of CORR is that there is a subset of organellarproteins for which there is an overriding requirement fora redox regulatory control that is initiated within theorganelle and whose effects are exerted by the simplestand most direct route This redox signalling is envisagedto take place without a requirement for prior consultationand coordination with what may be regarded as a nucleo-cytoplasmic bureaucracy
(v) Nuclear genes can be expressed as rapidly as chloroplastand mitochondrial genes
Zerges (2002) for example cites the equal times takenfor complete expression of nuclear and chloroplast genesin Chlamydomonas as evidence against the CORR hypoth-
32 J F Allen Genomes in mitochondria and chloroplasts
esis However the question is not one of the time takento express a gene but the time taken to alter the rate ofits expression in response to a (redox) signal originatingwithin the organelle An equally important additional fac-tor may be the autonomy of the signalling pathway thatis its independence from other inputs as described in theprevious paragraph One might say that mitochondria andchloroplasts require a free hand
(vi) There is no evidence to support this hypothesisThe hypothesis is rich in predictions (sectsect 3 and 6) and
those that have been tested stringently are in agreementwith experimental results None of the predictions of thehypothesis has been found to be incorrect and none of theunderlying principles (sect 3) has been disproved Furtherpredictions will be the subject of future experiments
(b) Evidence for CORR(i) Chloroplasts have selectively retained cyanobacterial genes
for primary events in photosynthesisSee table 1
(ii) There is regulation of gene expression in chloroplastsThere is transcriptional regulation of gene expression
in chloroplasts (sect 3d) The case for mitochondria is notyet proven
(iii) There is redox regulation of gene expression inchloroplasts
There is transcriptional redox regulation of geneexpression in chloroplasts (sect 3d) The case for mitochon-dria is not yet proven though see (Allen et al 1995 Kon-stantinov et al 1995 Galvis et al 1998)
(iv) There are no redox signalling components encoded inchloroplast or mitochondrial DNA
Correct but with possible exceptions discussed in sect 3j
(v) There are lsquobacterialrsquo redox signalling components ineukaryotes
See sect 3j
(vi) Further predictions DNA-array results and phenotypesof allotopic mutants
A systematic test of CORR might be to make an oxid-ized-minus-reduced gene expression profile of the lsquotran-scriptomersquo of a cyanobacterium to see if the set of geneswhose transcription is under most rapid and direct redoxcontrol corresponds to the genes of the chloroplast gen-ome Laboratories with this experimental capability do notseem to have addressed this possibility (Hihara et al 2002Ikeuchi 2002) Likewise if a putative modern representa-tive of the proto-mitochondrion can be found and agreedeg Rhodospirillum rubrum or Paracoccus denitrificans thena similar experiment should reveal a subset of genes corre-sponding to mitochondrial genomes DNA-microarraysalso present the possibility of testing the relative rate ofredox response and dependency on other signallinginputs of transcription of eukaryotic genes To dateorganellar genes have not been included in such investi-gations (Ferea et al 1999) A further experimental possi-bility is to examine the redox regulatory phenotypes ofexisting allotopic mutants as outlined in sect 3e
Phil Trans R Soc Lond B (2003)
6 CONCLUSION AND FUTURE PROSPECTS
The CORR hypothesis states that the function of mito-chondrial and chloroplast genomes is to provide co-location (of gene and gene product) for (evolutionary)continuity of redox regulation of gene expression Thishypothesis is neither self-contradictory nor devoid of test-able predictions It is also consistent with available evi-dence and in principle makes sense of the otherwisepuzzling distribution of genes between compartments ineukaryotic cells The explanation that CORR offersapplies equally to mitochondria and chloroplasts andidentifies selective forces that still operate to determine thelocation of genes No selective barriers need be assumedto gene transfer protein import nor membrane insertionNor do we need to invoke a lsquofrozen accidentrsquo that some-thing happened to stop gene transfer for some reason atsome time in the remote and inaccessible past CORR alsoexplains the absence of genomes from organelles that havelost their primary bioenergetic functions CORR may actas a stimulus to a search for residual bioenergetic functionin those organelles that have lost aerobic respiration andphotosynthesis but where residual genomes remain
CORR also predicts regulatory properties of knowncomponents of chloroplasts and mitochondria that in turnindicate important but as yet uncharacterized flexibility inenergy transduction These components include the RUB-ISCO large subunit and the CFo and Fo subunits of coup-ling ATPase in both chloroplasts and mitochondriaRedox regulation of the relative stoichiometry of compo-nents as seen in regulation of chloroplast photosystemstoichiometry (Pfannschmidt et al 1999a) may extend to(C)Fo-(C)F1-ATPase indicating that the function ofretention of genes for one or more of these componentsis flexibility in the stoichiometries of H1 to e2 and H1 toATP in chemiosmotic coupling (Schemidt et al 1998Stock et al 2000 Allen 2002)
So far explicit tests of CORR have largely been con-fined to chloroplasts and to some extent mitochondria ofhigher plants but this merely reflects the expertise andinterests of the investigators Further attempted disproofof CORR is welcomed and encouraged Novel experi-mental tests can be devised as well as attempted refu-tation through analysis of existing data It is hoped thatexplicit attention will be given to the CORR hypothesis inlaboratories already suitably equipped and that resourcescan be allocated for this purpose in laboratories wherethere is the interest but not currently the technology orexpertise
The heuristic value of the hypothesis already seen inthe role of redox-regulated transcription of photosyntheticreaction centre genes in chloroplasts may be its most use-ful property The central proposal is that mitochondriaand chloroplasts contain genes for proteins whose functionin electron transfer demands rapid direct and uncon-ditional redox regulatory control of their biosynthesis Thefull implications of this hypothesis are very broad and onlyjust beginning to be explored
I thank Carol Allen Aubrey de Grey William Martin John ARaven and Francis-Andre Wollman for discussion and corre-spondence during the preparation of this paper and WilliamMartin for suggestions including table 1 Research grants from
Genomes in mitochondria and chloroplasts J F Allen 33
The Swedish Natural Sciences Research Council (NFR) andthe Crafoord Foundation are gratefully acknowledged
REFERENCES
Adams K L Daley D O Qiu Y L Whelan J amp PalmerJ D 2000 Repeated recent and diverse transfers of a mito-chondrial gene to the nucleus in flowering plants Nature408 354ndash357
Alberts B Bray D Lewis J Raff M Roberts K amp Wat-son J D 1994 The molecular biology of the cell New Yorkand London Garland Publishing
Allen C A Hakansson G amp Allen J F 1995 Redox con-ditions specify the proteins synthesized by isolated chloro-plasts and mitochondria Redox Rep 1 119ndash123
Allen J F 1992 Protein phosphorylation in regulation ofphotosynthesis Biochim Biophys Acta 1098 275ndash335
Allen J F 1993a Control of gene expression by redox poten-tial and the requirement for chloroplast and mitochondrialgenomes J Theor Biol 165 609ndash631
Allen J F 1993b Redox control of gene expression and thefunction of chloroplast genomes a hypothesis PhotosynthRes 36 95ndash102
Allen J F 1993c Redox control of transcription sensorsresponse regulators activators and repressors FEBS Lett332 203ndash207
Allen J F 1995 Thylakoid proteinndashphosphorylation state-1ndashstate-2 transitions and photosystem stoichiometry adjust-ment redox control at multiple levels of gene expressionPhysiol Plant 93 196ndash205
Allen J F 1996 Separate sexes and the mitochondrial theoryof ageing J Theor Biol 180 135ndash140
Allen J F 2002 Photosynthesis of ATP-electrons protonpumps rotors and poise Cell 110 273ndash276
Allen J F amp Allen C A 1999 A mitochondrial model forpremature ageing of somatically cloned mammals IUBMBLife 48 369ndash372
Allen J F amp Nilsson A 1997 Redox signalling and the struc-tural basis of regulation of photosynthesis by protein phos-phorylation Physiol Plant 100 863ndash868
Allen J F amp Pfannschmidt T 2000 Balancing the two photo-systems photosynthetic electron transfer governs transcrip-tion of reaction centre genes in chloroplasts Phil TransR Soc Lond B 355 1351ndash1357 (DOI 101098rstb20000697)
Allen J F amp Race H L 2002 Will the real LHC II kinaseplease step forward Sciencersquos STKE httpwwwstkeorgcgicontentfullsigtrans2002155pe43
Allen J F amp Raven J A 1996 Free-radical-induced mutationvs redox regulation costs and benefits of genes in organellesJ Mol Evol 42 482ndash492
Archibald J M Cavalier-Smith T Maier U amp Douglas S2001 Molecular chaperones encoded by a reduced nucleusthe cryptomonad nucleomorph J Mol Evol 52 490ndash501
Ashby M K amp Mullineaux C W 1999a Cyanobacterialycf27 gene products regulate energy transfer from phycobili-somes to photosystems I and II FEMS Microbiol Lett 181253ndash260
Ashby M K amp Mullineaux C W 1999b The role of ApcDand ApcF in energy transfer from phycobilisomes to PSI andPSII in a cyanobacterium Photosynth Res 61 169ndash179
Ashby M K Houmard J amp Mullineaux C W 2002 Theycf27 genes from cyanobacteria and eukaryotic algae distri-bution and implications for chloroplast evolution FEMSMicrobiol Lett 214 25ndash30
Attardi G amp Schatz G 1988 Biogenesis of mitochondria ARev Cell Biol 4 289ndash333
Azem A Oppliger W Lustig A Jeno P Feifel B SchatzG amp Horst M 1997 The mitochondrial hsp70 chaperone
Phil Trans R Soc Lond B (2003)
system effect of adenine nucleotides peptide substrate andmGrpE on the oligomeric state of mhsp70 J Biol Chem272 20 901ndash20 906
Barbrook A C Symington H Nisbet R E R LarkumA amp Howe C J 2001 Organisation and expression of theplastid genome of the dinoflagellate Amphidinium opercula-tum Mol Genet Genomics 266 632ndash638
Bauer C E Elsen S amp Bird T H 1999 Mechanisms forredox control of gene expression A Rev Microbiol 53495ndash523
Bauer C E Elsen S Swem L R Swem D L amp MasudaS 2003 Redox and light regulation of gene expression inphotosynthetic prokaryotes Phil Trans R Soc Lond B 358147ndash154 (DOI 101098rstb20021189)
Berks B C Sargent F De Leeuw E Hinsley A P Stan-ley N R Jack R L Buchanan G amp Palmer T 2000aA novel protein transport system involved in the biogenesisof bacterial electron transfer chains Biochim Biophys ActaBioenerg 1459 325ndash330
Berks B C Sargent F amp Palmer T 2000b The tat proteinexport pathway Mol Microbiol 35 260ndash274
Blackstone N W 1999 Redox control in development andevolution evidence from colonial hydroids J Exp Biol 2023541ndash3553
Blackstone N W 2000 Redox control and the evolution ofmulticellularity Bioessays 22 947ndash953
Blackstone N W 2001 Redox state reactive oxygen speciesand adaptive growth in colonial hydroids J Exp Biol 2041845ndash1853
Bogorad L 1975 Evolution of organelles and eukaryotic gen-omes Science 188 891ndash898
Bogsch E G Sargent F Stanley N R Berks B C Robin-son C amp Palmer T 1998 An essential component of a novelbacterial protein export system with homologues in plastidsand mitochondria J Biol Chem 273 18 003ndash18 006
Brown E C Somanchi A amp Mayfield S P 2001 Interor-ganellar crosstalk new perspectives on signaling from thechloroplast to the nucleus Genome Biol 2 10211ndash10214
Cavalier-Smith T 2002 Chloroplast evolution secondarysymbiogenesis and multiple losses Curr Biol 12 R62ndashR64
Chitnis P R amp Thornber J P 1988 The major light-har-vesting complex of photosystem 2 Aspects of its molecularand cell biology Photosynth Res 16 41ndash63
Chow W S Melis A amp Anderson J M 1990 Adjustmentsof photosystem stoichiometry in chloroplasts improve thequantum efficiency of photosynthesis Proc Natl Acad SciUSA 87 7502ndash7506
Claros M G Perea J Shu Y Samatey F A PopotJ L amp Jacq C 1995 Limitations to in vivo import of hydro-phobic proteins into yeast mitochondria The case of a cyto-plasmically synthesized apocytochrome b Eur J Biochem228 762ndash771
Dalbey R E amp Robinson C 1999 Protein translocation intoand across the bacterial plasma membrane and the plant thy-lakoid membrane Trends Biochem Sci 24 17ndash22
Daley D O Adams K L Clifton R Qualmann S MillarA H Palmer J D Pratje E amp Whelan J 2002 Genetransfer from mitochondrion to nucleus novel mechanismsfor gene activation from Cox2 Plant J 30 11ndash21
Danon A amp Mayfield S P 1994 Light-regulated translationof chloroplast messenger RNAs through redox potentialScience 266 1717ndash1719
de Grey A 2000 Mitochondrial gene therapy an arena for thebiomedical use of inteins Trends Biotechnol 18 394ndash399
Deisenhofer J Epp O Miki K Huber R amp Michel H1985 Structure of the protein subunits in the photosyntheticreaction center of Rhodopseudomonas viridis at 3Ac resolutionNature 318 618ndash624
34 J F Allen Genomes in mitochondria and chloroplasts
Delwiche C F 1999 Tracing the thread of plastid diversitythrough the tapestry of life Am Nat 154 S164ndashS177
Doolittle W F 1999 Phylogenetic classification and the uni-versal tree Science 284 2124ndash2129
Douglas S E 1998 Plastid evolution origins diversitytrends Curr Opin Genet Dev 8 655ndash661
Duby G amp Boutry M 2002 Mitochondrial protein importmachinery and targeting information Plant Sci 162 477ndash490
Eberhard S Drapier D amp Wollman F A 2002 Searchinglimiting steps in the expression of chloroplast encoded pro-teins relations between gene copy number transcriptiontranscript abundance and translation rate in the chloroplastof Chlamydomonas reinhardtii Plant J 31 149ndash160
El Bissati K amp Kirilovsky D 2001 Regulation of psbA andpsaE expression by light quality in Synechocystis species PCC6803 A redox control mechanism Plant Physiol 1251988ndash2000
Ellis R J 1984 The nuclear domination of chloroplast devel-opment Science Progress 69 129ndash142
Embley T M amp Martin W 1998 Molecular evolution ahydrogen-producing mitochondrion Nature 396 517ndash519
Ems S C Morden C W Dixon C K Wolfe K HdePamphilis C W amp Palmer J D 1995 Transcriptionsplicing and editing of plastid RNAs in the non-photosyn-thetic plant Epifagus virginiana Plant Mol Biol 29 721ndash733
Eraso J M amp Kaplan S 2000 From redox flow to gene regu-lation role of the PrrC protein of Rhodobacter sphaeroides241 Biochemistry 39 2052ndash2062
Eraso J M amp Kaplan S 2002 Redox flow as an instrumentof gene regulation Meth Enzymol 348 216ndash229
Escoubas J M Lomas M Laroche J amp Falkowski P G1995 Light-intensity regulation of cab gene-transcription issignaled by the redox state of the plastoquinone pool ProcNatl Acad Sci USA 92 10 237ndash10 241
Ferea T L Botstein D Brown P O amp Rosenzweig R F1999 Systematic changes in gene expression patterns follow-ing adaptive evolution in yeast Proc Natl Acad Sci USA96 9721ndash9726
Forsberg J Rosenquist M Fraysse L amp Allen J F 2001Redox signalling in chloroplasts and mitochondria genomicand biochemical evidence for two-component regulatory sys-tems in bioenergetic organelles Biochem Soc Trans 29403ndash407
Fujita Y 1997 A study on the dynamic features of photosys-tem stoichiometry accomplishments and problems forfuture studies Photosynth Res 53 83ndash93
Gabriel K Buchanan S K amp Lithgow T 2001 The alphaand the beta protein translocation across mitochondrial andplastid outer membranes Trends Biochem Sci 26 36ndash40
Galvis M L E Allen J F amp Hakansson G 1998 Proteinsynthesis by isolated pea mitochondria is dependent on theactivity of respiratory complex II Curr Genet 33 320ndash329
Gatenby A A amp Ellis R J 1990 Chaperone function theassembly of ribulose bisphosphate carboxylase-oxygenase ARev Cell Biol 6 125ndash149
GoldschmidtClermont M 1998 Coordination of nuclear andchloroplast gene expression in plant cells Int Rev CytolSurv Cell Biol 177 115ndash180
Gray M W 1992 The endosymbiont hypothesis revisited IntRev Cytol 141 233ndash357
Gray M W 1999 Evolution of organellar genomes CurrOpin Genet Dev 9 678ndash687
Gray M W 2000 Evolutionary biology mitochondrial geneson the move Nature 408 302ndash305
Gray M W amp Doolittle W F 1982 Has the endosymbionthypothesis been proven Microbiol Rev 46 1ndash42
Gray M W Burger G amp Lang B F 1999 Mitochondrialevolution Science 283 1476ndash1481
Phil Trans R Soc Lond B (2003)
Grebe T W amp Stock J B 1999 The histidine protein kinasesuperfamily Adv Microb Physiol 41 139ndash227
Green B R amp Kuhlbrandt W 1995 Sequence conservationof light-harvesting and stress-response proteins in relation tothe 3-dimensional molecular structure of Lhcii PhotosynthRes 44 139ndash148
Gruissem W amp Tonkyn J C 1993 Control mechanisms ofplastid gene expression Crit Rev Plant Sci 12 19ndash55
Hackstein J H P (and 11 others) 2001 Hydrogenosomesconvergent adaptations of mitochondria to anaerobicenvironments Zool Anal Complex Syst 104 290ndash302
Harrison M A Keen J N Findlay J B C amp Allen J F1990 Modification of a Glnb-like gene-product by photosyn-thetic electron-transport in the cyanobacterium Synechoc-occus 6301 FEBS Lett 264 25ndash28
Harvey A J Kind K L amp Thompson J G 2002 Redoxregulation of early embryo development Reproduction 123479ndash486
Heathcote P Fyfe P K amp Jones M R 2002 Reactioncentres the structure and evolution of biological solarpower Trends Biochem Sci 27 79ndash87
Henze K amp Martin W 2001 How do mitochondrial genesget into the nucleus Trends Genet 17 383ndash387
Herrmann R G amp Westhoff P 2001 Thylakoid biogenesis anddynamics the result of a complex phylogenetic puzzle InRegulation of photosynthesis vol 11 (ed E-M Aro amp BAndersson) pp 1ndash28 Dordrecht The Netherlands Kluwer
Hihara Y Sonoike K Kanehisa M amp Ikeuchi M 2002Search for redox responsive genes in Synechocystis sp PCC6803 by using DNA microarrays Plant Cell Physiol 43S178
Howe C J Barbrook A C amp Lockhart P J 2000 Organellegenes do they jump or are they pushed Trends Genet 1665ndash66
Ikeuchi M 2002 Gene expression and responses to light stressin cyanobacteria Plant Cell Physiol 43 S21
Iuchi S amp Lin E C C 1992 Mutational analysis of signaltransduction by Arcb a membrane sensor protein respon-sible for anaerobic repression of operons involved in the cen-tral aerobic pathways in Escherichia coli J Bacteriol 1743972ndash3980
Jackson-Constan D Akita M amp Keegstra K 2001 Molecu-lar chaperones involved in chloroplast protein importBiochim Biophys Acta Mol Cell Res 1541 102ndash113
Jarvis P amp Soll M 2001 Toc Tic and chloroplast proteinimport Biochim Biophys Acta Mol Cell Res 1541 64ndash79
Jordan P Fromme P Witt H T Klukas O SaengerW amp Krauss N 2001 Three-dimensional structure ofcyanobacterial photosystem I at 25 acngstrom resolutionNature 411 909ndash917
Kaneko T (and 23 others) 1996a Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regionsDNA Res 3 109ndash136
Kaneko T (and 23 others) 1996b Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regions[Supplement] DNA Res 3 185ndash209
Kanevski I amp Maliga P 1994 Relocation of the plastid rbcLgene to the nucleus yields functional ribulose-15-bisphosph-ate carboxylase in tobacco chloroplasts Proc Natl Acad SciUSA 91 1969ndash1973
Kim J amp Mayfield S P 1997 Protein disulfide isomerase as aregulator of chloroplast translational activation Science 2781954ndash1957
Genomes in mitochondria and chloroplasts J F Allen 35
Kirk J T O amp Tilney-Bassett R A E 1978 The plastidsTheir chemistry structure growth and inheritance Amster-dam Elsevier
Koehler C M Merchant S amp Schatz G 1999 How mem-brane proteins travel across the mitochondrial intermem-brane space Trends Biochem Sci 24 428ndash432
Komiya T Rospert S Schatz G amp Mihara K 1997 Bind-ing of mitochondrial precursor proteins to the cytoplasmicdomains of the import receptors Tom70 and Tom20 isdetermined by cytoplasmic chaperones FASEB J 11 1196
Konstantinov Y M Lutsenko G N amp Podsosonny V A1995 Genetic functions of isolated maize mitochondriaunder model changes of redox conditions Biochem MolBiol Int 36 319ndash326
Korab-Laskowska M Rioux P Brossard N LittlejohnT G Gray M W Lang B F amp Burger G 1998 Theorganelle genome database project (GOBASE) Nucleic AcidsRes 26 138ndash144
Kovacs L Wiessner W Kis M Nagy F Mende D ampDemeter S 2000 Short- and long-term redox regulation ofphotosynthetic light energy distribution and photosystemstoichiometry by acetate metabolism in the green alga Chla-mydobotrys stellata Photosynth Res 65 231ndash247
Kuhlbrandt W 1994 Structure and function of the plant light-harvesting complex LHC-II Curr Opin Struct Biol 4519ndash528
Lang B F Gray M W amp Burger G 1999 Mitochondrialgenome evolution and the origin of eukaryotes A RevGenet 33 351ndash397
Lemire B D Fankhauser C Baker A amp Schatz G 1989The mitochondrial targeting function of randomly generatedpeptide sequences correlates with predicted helical amphi-philicity J Biol Chem 264 20 206ndash20 215
Leuenberger D Bally N A Schatz G amp Koehler C M1999 Different import pathways through the mitochondrialintermembrane space for inner membrane proteins EMBOJ 18 4816ndash4822
Li H amp Sherman L A 2000 A redox-responsive regulator ofphotosynthesis gene expression in the cyanobacterium Syne-chocystis sp strain PCC 6803 J Bacteriol 182 4268ndash4277
Lin X Y (and 36 others) 1999 Sequence and analysis of chro-mosome 2 of the plant Arabidopsis thaliana Nature 402761ndash768
Lonsdale D M Hodge T P Howe C J amp Stern D B1983 Maize mitochondrial-DNA contains a sequence hom-ologous to the ribulose-15-bisphosphate carboxylase largesubunit gene of chloroplast DNA Cell 34 1007ndash1014
McFadden G I 1999 Endosymbiosis and evolution of theplant cell Curr Opin Plant Biol 2 513ndash519
McFadden G I 2001 Chloroplast origin and integrationPlant Physiol 125 50ndash53
Mahler H R amp Raff R A 1975 The evolutionary origin ofthe mitochondrion a nonsymbiotic model Int Rev Cytol43 1ndash124
Manfredi G Fu J Ojaimi J Sadlock J E Kwong J QGuy J amp Schon E A 2002 Rescue of a deficiency in ATPsynthesis by transfer of MTATP6 a mitochondrial DNA-encoded gene to the nucleus Nature Genet 30 394ndash399
Marc P Margeot A Devaux F Blugeon C Corral-Debrinski M amp Jacq C 2002 Genome-wide analysis ofmRNAs targeted to yeast mitochondria EMBO Rep 3159ndash164
Marienfeld J Unseld M amp Brennicke A 1999 The mito-chondrial genome of Arabidopsis is composed of both nativeand immigrant information Trends Plant Sci 4 495ndash502
Martin W 1999 A briefly argued case that mitochondria andplastids are descendants of endosymbionts but that thenuclear compartment is not Proc R Soc Lond B 2661387ndash1395 (DOI 101098rspb19990792)
Phil Trans R Soc Lond B (2003)
Martin W amp Herrmann R G 1998 Gene transfer fromorganelles to the nucleus how much what happens andwhy Plant Physiol 118 9ndash17
Martin W amp Muller M 1998 The hydrogen hypothesis forthe first eukaryote Nature 392 37ndash41
Martin W amp Russell M J 2003 On the origins of cells ahypothesis for the evolutionary transitions from abiotic geo-chemistry to chemoautotrophic prokaryotes and from pro-karyotes to nucleated cells Phil Trans R Soc Lond B 35859ndash85 (DOI 101098rstb20021183)
Martin W Stoebe B Goremykin V Hansmann S Hase-gawa M amp Kowallik K V 1998 Gene transfer to thenucleus and the evolution of chloroplasts Nature 393162ndash165
Martin W Hoffmeister M Rotte C amp Henze K 2001 Anoverview of endosymbiotic models for the origins of eukary-otes their ATP-producing organelles (mitochondria andhydrogenosomes) and their heterotrophic lifestyle BiolChem 382 1521ndash1539
Martin W Rujan T Richly E Hansen A Cornelsen SLins T Leister D Stoebe B Hasegawa M amp PennyD 2002 Evolutionary analysis of Arabidopsis cyanobacterialand chloroplast genomes reveals plastid phylogeny and thou-sands of cyanobacterial genes in the nucleus Proc NatlAcad Sci USA 99 12 246ndash12 251
Millen R S (and 12 others) 2001 Many parallel losses of infAfrom chloroplast DNA during angiosperm evolution withmultiple independent transfers to the nucleus Plant Cell 13645ndash658
Morse D Salois P Markovic P amp Hastings J W 1995 Anuclear-encoded form-II RUBISCO in dinoflagellatesScience 268 1622ndash1624
Murakami A Kim S J amp Fujita Y 1997 Changes in photo-system stoichiometry in response to environmental con-ditions for cell growth observed with the cyanophyteSynechocystis PCC 6714 Plant Cell Physiol 38392ndash397
Oh J I amp Kaplan S 2000 Redox signaling globalization ofgene expression EMBO J 19 4237ndash4247
Ozawa T 1995 Mitochondrial-DNA mutations associatedwith aging and degenerative diseases Exp Gerontol 30269ndash290
Palmer J D 1997 Organelle genomes going going goneScience 275 790ndash791
Pearson C K Wilson S B Schaffer R amp Ross A W 1993NAD turnover and utilization of metabolites for RNA syn-thesis in a reaction sensing the redox state of the cyto-chrome-b( 6 )f complex in isolated chloroplasts Eur JBiochem 218 397ndash404
Pfanner N amp Geissler A 2001 Versatility of the mitochon-drial protein import machinery Natl Rev Mol Cell Biol 2339ndash349
Pfanner N amp Wiedemann N 2002 Mitochondrial proteinimport two membranes three translocases Curr Opin CellBiol 14 400ndash411
Pfannschmidt T Nilsson A amp Allen J F 1999a Photosyn-thetic control of chloroplast gene expression Nature 397625ndash628
Pfannschmidt T Nilsson A Tullberg A Link G amp AllenJ F 1999b Direct transcriptional control of the chloroplastgenes psbA and psaAB adjusts photosynthesis to light energydistribution in plants IUBMB Life 48 271ndash276
Pfannschmidt T Allen J F amp Oelmuller R 2001a Prin-ciples of redox control in photosynthesis gene expressionPhysiol Plant 112 1ndash9
Pfannschmidt T Schutze K Brost M amp Oelmuller R2001b A novel mechanism of nuclear photosynthesis generegulation by redox signals from the chloroplast during pho-
36 J F Allen Genomes in mitochondria and chloroplasts
tosystem stoichiometry adjustment J Biol Chem 27636 125ndash36 130
Popot J L amp de Vitry C 1990 On the microassembly of inte-gral membrane proteins A Rev Biophys Biophys Chem 19369ndash403
Race H L Herrmann R G amp Martin W 1999 Why haveorganelles retained genomes Trends Genet 15 364ndash370
Raven J A Johnston A M Parsons R amp Kubler J 1994aThe influence of natural and experimental high O2 concen-trations on O2 evolving phototrophs Biol Rev CambridgePhil Soc 69 61ndash94
Raven J A Johnston A M Parsons R amp Kubler J 1994bThe occurrence and influence on photolithotrophs of highoxygen concentrations Proc R Soc Edinb B 102 193ndash201
Robinson C 2000 The twin-arginine translocation system anovel means of transporting folded proteins in chloroplastsand bacteria Biol Chem 381 89ndash93
Robinson C amp Bolhuis A 2001 Protein targeting by the twin-arginine translocation pathway Natl Rev Mol Cell Biol 2350ndash356
Rodermel S 2001 Pathways of plastid-to-nucleus signalingTrends Plant Sci 6 471ndash478
Roger A J 1999 Reconstructing early events in eukaryoticevolution Am Nat 154 S146ndashS163
Roger A J Svard S G Tovar J Clark C G SmithM W Gillin F D amp Sogin M L 1998 A mitochondrial-like chaperonin 60 gene in Giardia lamblia evidence thatdiplomonads once harbored an endosymbiont related to theprogenitor of mitochondria Proc Natl Acad Sci USA 95229ndash234
Roodyn D B amp Wilkie D 1968 The biogenesis of mitochondriaMethuenrsquos monographs on biological subjects London Methuen
Rujan T amp Martin W 2001 How many genes in Arabidopsiscome from cyanobacteria An estimate from 386 proteinphylogenies Trends Genet 17 113ndash120
Saccone C Gissi C Lanave C Larizza A Pesole G ampReyes A 2000 Evolution of the mitochondrial genetic sys-tem an overview Gene 261 153ndash159
Salzberg S L White O Peterson J amp Eisen J A 2001Microbial genes in the human genome lateral transfer orgene loss Science 292 1903ndash1906
Sargent F Berks B C amp Palmer T 2002 Assembly of mem-brane-bound respiratory complexes by the Tat protein-trans-port system Arch Microbiol 178 77ndash84
Schatz G 1998 Protein transport the doors to organellesNature 395 439ndash440
Schemidt R A Qu J Williams J R amp Brusilow W S1998 Effects of carbon source on expression of F0 genes andon the stoichiometry of the c subunit in the F1 F0 ATPaseof Escherichia coli J Bacteriol 180 3205ndash3208
Selosse M A Albert B R amp Godelle B 2001 Reducing thegenome size of organelles favours gene transfer to thenucleus Trends Ecol Evol 16 135ndash141
Stern D B amp Lonsdale D M 1982 Mitochondrial and chlo-roplast genomes of maize have a 12- kilobase DNA-sequencein common Nature 299 698ndash702
Stock D Gibbons C Arechaga I Leslie A G W ampWalker J E 2000 The rotary mechanism of ATP synthaseCurr Opin Struct Biol 10 672ndash679
Stoebe B amp Maier U G 2002 One two three naturersquos toolbox for building plastids Protoplasma 219 123ndash130
Stoebe B Martin W amp Kowallik K V 1998 Distributionand nomenclature of protein-coding genes in 12 sequencedchloroplast genomes Plant Mol Biol Rep 16 243ndash255
Stupar R M Lilly J W Town C D Cheng Z Kaul SBuell C R amp Jiang J M 2001 Complex mtDNA consti-tutes an approximate 620 kb insertion on Arabidopsis thali-ana chromosome 2 implication of potential sequencing
Phil Trans R Soc Lond B (2003)
errors caused by large-unit repeats Proc Natl Acad SciUSA 98 5099ndash5103
Surpin M Larkin R M amp Chory J 2002 Signal transduc-tion between the chloroplast and the nucleus Plant Cell 14S327ndashS338
Tjaden J Schwoppe C Mohlmann T Quick P W ampNeuhaus H E 1998 Expression of a plastidic ATPADPtransporter gene in Escherichia coli leads to a functional aden-ine nucleotide transport system in the bacterial cytoplasmicmembrane J Biol Chem 273 9630ndash9636
Tovar J Fischer A amp Clark C G 1999 The mitosome a novelorganelle related to mitochondria in the amitochondrial para-site Entamoeba histolytica Mol Microbiol 32 1013ndash1021
Trebitsh T amp Danon A 2001 Translation of chloroplastpsbA mRNA is regulated by signals initiated by both photos-ystems II and I Proc Natl Acad Sci USA 98 12 289ndash12 294
Tullberg A Alexciev K Pfannschmidt T amp Allen J F2000 Photosynthetic electron flow regulates transcription ofthe psaB gene in pea (Pisum sativum L) chloroplasts throughthe redox state of the plastoquinone pool Plant Cell Physiol41 1045ndash1054
Turmel M Lemieux C Burger G Lang B F Otis CPlante I amp Gray M W 1999 The complete mitochondrialDNA sequences of Nephroselmis olivacea and Pedinomonasminor two radically different evolutionary patterns withingreen algae Plant Cell 11 1717ndash1729
Unden G amp Bongaerts J 1997 Alternative respiratory path-ways of Escherichia coli energetics and transcriptional regu-lation in response to electron acceptors Biochim BiophysActa Bioenerg 1320 217ndash234
Unseld M Marienfeld J R Brandt P amp Brennicke A1997 The mitochondrial genome of Arabidopsis thaliana con-tains 57 genes in 366 924 nucleotides Nature Genet 1557ndash61
Van der Giezen M Slotboom D J Horner D S DyalP L Harding M Xue G P Embley T M amp KunjiE R S 2002 Conserved properties of hydrogenosomal andmitochondrial ADPATP carriers a common origin for bothorganelles EMBO J 21 572ndash579
Von Heijne G 1986 Why mitochondria need a genome FEBSLett 198 1ndash4
Westphal S Soll J amp Vothknecht U C 2001 A vesicle trans-port system inside chloroplasts FEBS Lett 506 257ndash261
Whitehouse H L K 1969 Towards an understanding of themechanism of heredity London Edward Arnold Ltd
Williams B A P Hirt R P Lucocq J M amp Embley T M2002 A mitochondrial remnant in the microsporidian Trachi-pleistophora hominis Nature 418 865ndash869
Wilson R J (and 10 others) 1996a Complete gene map of theplastid-like DNA of the malaria parasite Plasmodium falcipa-rum J Mol Biol 261 155ndash172
Wilson S B Davidson G S Thomson L M amp PearsonC K 1996b Redox control of RNA synthesis in potato mito-chondria Eur J Biochem 242 81ndash85
Winkler H H amp Neuhaus H E 1999 Non-mitochondrialATP transport Trends Biochem Sci 24 64ndash68
Wolfe K H Morden C W amp Palmer J D 1992 Small sin-gle-copy region of plastid DNA in the non-photosyntheticangiosperm Epifagus virginiana contains only two genes Dif-ferences among dicots monocots and bryophytes in geneorganization at a non-bioenergetic locus J Mol Biol 22395ndash104
Wollman F A Minai L amp Nechushtai R 1999 The biogen-esis and assembly of photosynthetic proteins in thylakoidmembranes Biochim Biophys Acta Bioenerg 1411 21ndash85
Yeliseev A A amp Kaplan S 2000 TspO of Rhodobactersphaeroides a structural and functional model for the mam-
Genomes in mitochondria and chloroplasts J F Allen 37
malian peripheral benzodiazepine receptor J Biol Chem275 5657ndash5667
Zerges W 2002 Does complexity constrain organelle evol-ution Trends Plant Sci 7 175ndash182
Zhang Z D Green B R amp Cavalier-Smith T 1999 Singlegene circles in dinoflagellate chloroplast genomes Nature400 155ndash159
Zhang Z D Cavalier-Smith T amp Green B R 2001 A fam-ily of selfish minicircular chromosomes with jumbled chloro-plast gene fragments from a dinoflagellate Mol Biol Evol18 1558ndash1565
Zouni A Witt H T Kern J Fromme P Krauss NSaenger W amp Orth P 2001 Crystal structure of photosys-tem II from Synechococcus elongatus at 38 acngstrom resol-ution Nature 409 739ndash743
DiscussionD S Horner (Dipartimento di Fisiologia e Biochimica
Generali University of Milan Milan Italy) Given thatthere are components of these membrane-bound energy-transduction pathways that are consistently nuclear enco-ded not the key ones you pointed out but others do youexpect perhaps a slower perhaps a more crude but yet aredox communication between the organelle andexpression of nuclear-encoded genes
J F Allen There is and there is no question about itI think that Bogoradrsquos lock-in hypothesis has somethingto say here and there are other people who say it is all todo with assembling multi-subunit complexes If you havecontrol over the initial construction of the core aroundwhich the whole complex is eventually built then redoxcontrol or some sort of control over synthesis of that corewill control construction of the whole thing eventuallyand I think that may well be largely correct I think it ispossible in principle for nuclear genes to make precursorsaccording to redox instructions but it is not really soimportant that those precursors are here and now presentin exactly the right quantities because the precursors canto a limited extent hang around and wait Actually thereis proteolytic degradation of unattached precursors and soon but I think the fact that these chloroplast and mito-chondrially DNA-encoded subunits are at the core ofthese energy-transducing complexes is the crucial issue
T Cavalier-Smith (Department of Zoology University ofOxford Oxford UK ) In the case of mitochondria the sub-set of genes that remains is much more variable than youimplied It runs from three proteins in the minimal casesup to I think over 90 That is a big difference and manyof the genes in different lineages of protists that remainhave nothing to do with redox control they are things likeribosomal proteins and so on So I think that even thoughwhat you say about redox control of certain genes mayactually be an important thing it is not the fundamentaloverall reason why the genomes are maintained It may bepart of the explanation for why certain genes are alwaysincluded in that subset but even in mitochondria I thinkthat cytochrome b is the only one that is always presentSo I think that both this great diversity of what has beenretained in different lineages and the nucleomorph caseindicate that evolution was able to do one thing in onelineage 500 million years ago and never got around todoing it for 500 million years in another lineage so thereis a lot more haphazardness and evolutionary accidentsand contingency than a straight functional interpretation
Phil Trans R Soc Lond B (2003)
there is a lot of just sheer lsquohappenedndashorndashdidnrsquotndashhappenrsquoor carryover
J F Allen I was not implying anything about the rangeof mitochondrial genomes I have a limited amount oftime so I just talked about what I could talk about withinthat limit Of course there will be a range and this willdepend upon the physiology and bioenergetics of theparticular organism But what I am trying to do is to seewhat the common properties of the irreducible core areof components that are always encoded in the organellesand cytochrome b is a perfect example because that reallyis the thing that binds these two all-important quinoneswithout which there is no Q cycle without which there isno proton pumping and bioenergetics and in Plasmodiumwhere the mitochondrion is reduced to this bare mini-mum there is still a mitochondrial gene for cytochromeb Now my argument about the ribosomal proteins andall the rest of it is that in order to be able to exert thisall-important redox control over gene expression of certaincore components you need an in situ genetic system andthis can vary in composition for reasons that I cannotdescribe Maybe you need RNA polymerase or maybeyou can import RNA polymerase In plastids you haveboth of these options You are going to need ribosomestransfer RNAs and the rest of the apparatus of geneexpression and that needs to be in situ but we are stillleft with the question why does there have to be any gen-etic system at all The mature tobacco or spinach chloro-plast seems to have an entire genetic system just to makeone protein D1 which is all it ever needs to resynthesizeI think that this is a clue It is worth having this very unec-onomical crazy system because there is an overridingrequirement to connect electron transfer with geneexpression directly and unambiguouslymdashwithout recourseto a bureaucracy if you like which is what I think thenucleus is
K Willison (Institute of Cancer Research Chester BeattyLaboratories London UK ) These photosynthetic reactioncomplexes are a sort of baroque mixture of polypeptidechains and cofactors and my understanding is that someof them in high photon flux can turn over within say 30minutes so I am wondering if there is a correlationbetween gene retention and the turnover rates of some ofthe components and the complexes it seems to me thatthat could really be a limiting factor and the reason thatthe proteins had to be encoded locally in order to get thecomplexes to be assembled rapidly
J F Allen I think that is right You can only alter thesteady-state concentration of a protein complex by affect-ing the synthesis and breakdown and therefore turnoverThere is an extreme example the D1 protein encoded bythe psbA gene of chloroplasts which is always in the chlor-oplast genome without exception High light will tend todegrade this protein and when the high light goes awaythere is an absolute need very quickly to bring it back upagain I think that is a very good example Whether thisargument applies to genes such as cytochrome oxidase Ido not know
C J Leaver (Department of Plant Sciences University ofOxford Oxford UK ) Your experiments tend to relatemainly to the gene expression side What about proteinRNA turnover the whole concept of redox control ofthatmdashdo we know anything
38 J F Allen Genomes in mitochondria and chloroplasts
J F Allen Well quite a lot is known about chloro-plasts There is redox control of messenger RNA stabilityfor this same D1 protein which has been described byStephen Mayfield and his co-workers The evidence is thatthe site of redox control is ferredoxin via thioredoxin andthe implication is that thioredoxin tells whether non-cyclicelectron transport is working at all When it is workingthere is a signal that says lsquoOK now continue with all theprotein synthesisrsquo It is not particularly specific but moreof a global effect I think this is what Mayfield argues butof course this is not inconsistent with my argument I haveconcentrated on quinone pools and medium-potentialredox regulatory control because that is where I can seesignals about the dangerous consequences of getting itwrong but there is control of other levels of geneexpression In mitochondria I am not so sure I wouldbounce the question back to you on that one
W Martin (Institute of Botany III Heinrich-Heine Univ-ersitat Dusseldorf Dusseldorf Germany) The nucleomorphgenome was cited as evidence in conflict with your modelbut is that true The nucleomorph genome is not in abioenergetic organelle in that sense It resides in the cyto-sol so does the size of the nucleomorph genome workagainst your model Is it really evidence against yourmodel
J F Allen I do not know why should it be evidenceI cannot explain why the nucleomorph has a genomeThat is why I clutched at the straw that it is storage fornucleic acid precursors
Phil Trans R Soc Lond B (2003)
T Cavalier-Smith The reason I introduced the nucleo-morph is not to say it is direct evidence against yourhypothesis but to show that there probably was no singlefunctional reason for the retention of every gene in everyorganelle It may be that there are specific physiologicalreasons for the retention of certain genes and thereforeyou might be right about a subset of genes but I verymuch doubt if there are specific physiological reasons forthe retention of every single gene that is retained differen-tially because odd things happen in evolution Somethings are easy and sometimes things are difficult Evol-ution is history
J F Allen Of course I agree with that I do not thinkI can explain everything but I do think I can explainsomething If I have done that then I am content
GLOSSARY
AAC ADPndashATP carrierCES control by epistasy of synthesisCORR co-location for redox regulationLHC light-harvesting complexPCC Pasteur culture collectionORF open reading frameRUBISCO ribulose-15-bisphosphate carboxylasendashoxygenaseTAT twin-arginine translocase
Visit httpwwwpubsroyalsocacuk to see an electronic appendix tothis paper

30 J F Allen Genomes in mitochondria and chloroplasts
through membranes no special constraint on transportshould be expected to arise from 3D structure or surfaceproperties of a protein
An exception to this unfolding rule may be providedby holoproteins containing cofactors that are transportedalong with the apoprotein presumably with a largely intact3D structure The lsquoTATrsquo (twin-arginine translocase) sys-tem (Bogsch et al 1998 Dalbey amp Robinson 1999 Berkset al 2000ab Robinson 2000 Robinson amp Bolhuis 2001Sargent et al 2002) works broadly along these linesthough its mechanism is unresolved TAT is relevant herebecause it adds transport of essentially complete holopro-teins to the cellrsquos repertoire of transport mechanisms andeven proteins that may not be allowed to unfold perhapsbecause of the toxicity of the free cofactor are not therebymade candidates for an unimportability rule One variantof the unimportability hypothesis is that transport canwork only by unfolding and dissociated cofactors may betoxic to the cell The bacterially derived TAT system(Bogsch et al 1998 Dalbey amp Robinson 1999 Berks etal 2000ab Sargent et al 2002) overcomes this argumentas would examples of cofactor-free polypeptides such as(C)Fo subunit c which are nevertheless generally encodedin organelles
The presence of a vesicular transport system in chloro-plasts (Westphal et al 2001) counts against the unim-portability hypothesis Vesicular transport between thechloroplast envelope and the thylakoid membrane shouldin principle allow any protein to be ferried across thechloroplast stroma Vesicles might allow transport inparticular of membrane proteins that would discharge theproton motive force if allowed to connect transiently theN and P aqueous phases on each side of a coupling mem-brane
Another observation that seems to count against unim-portability is the increased number of membrane barriersin plastids derived from secondary tertiary or higher-orderendosymbioses These plastids seem to have retained ifanything fewer genes rather than more as unimportabilitypredicts The nucleomorph the residual nucleus of theprimary host retains many genes for molecular chap-erones (Archibald et al 2001 Cavalier-Smith 2002) inagreement with the conclusion that protein import pro-ceeds by conserved and specialized mechanisms If eachnew import added to gain a higher-order endosymbiosisrequires additional targeting presequences then perhapsthere is an advantage in components of an import machin-ery not being imported themselves
It seems that unimportability is less easy an assumptionto make today than in previous years and the onus maynow be on its proponents to cite specific examples of pro-teins that can never be imported into chloroplasts or mito-chondria
(f ) Some genes cannot be moved(i) Differences in the mitochondrial genetic code
The genetic code is the specific relation between ribon-ucleotide triplets of codons and tRNAs that recognizethem and which thus determine which individual aminoacids if any will be incorporated into the growing poly-peptide chain Mitochondria show small departures fromthe otherwise universal genetic code (Attardi amp Schatz1988 Alberts et al 1994) UGA is a lsquostoprsquo codon in the
Phil Trans R Soc Lond B (2003)
universal code and in plant mitochondria but is the codonfor tryptophan in mitochondria of animals and fungiCUA means leucine everywhere except fungal mitochon-dria where it means threonine and AGA and AGU codefor arginine in the lsquouniversalrsquo code but serine in insectmitochondria and lsquostoprsquo in mammalian ones The signifi-cance of this heterogeneity in coding is most unlikely torest in the early origin of mitochondria as the lsquouniversalrsquocode is employed by bacteria and protists Furthermorethe diversity of departures from the lsquouniversalrsquo codeamong mitochondria of different eukaryotic lineages indi-cates they had independent origins A general rule formitochondrial genetic codes is that reliance on the thirdlsquowobblersquo position is decreased which allows all 20 proteinamino acids to be specified by fewer tRNAs (22 in animalmitochondria instead of 31 in the cytosol) This in turnmeans that mtDNA can specify more proteins for a givenlength of DNA or conversely that the mitochondrialgenome can be condensed or lsquostreamlinedrsquo Although thisarrangement does not explain the retention of mitochon-drial genomes and may be a secondary event in mitochon-drial evolution it almost certainly places an extra barrierin place of gene relocation As discussed previously how-ever there seems to be no single barrier that can explainthe retention of a relatively constant subset of mitochon-drial genes
There has clearly been a contraction of mitochondrialDNA especially in animals Animal mitochondrial DNAhas few if any non-coding sequences and animal mito-chondrial genetic codes require fewer tRNAs than the sin-gle code operating in the cytosol (Gray 1999 Gray et al1999 Selosse et al 2001) These may be devices todecrease the total frequency of mutations per mitochon-drial genome assuming a constant frequency per unitlength of mitochondrial DNA It has been proposed thata genersquos removal to the nucleus decreases its susceptibilityto free radical mutagenesis (Raven et al 1994ab Allen ampRaven 1996) If this is the case then the non-standardcode may be a further indication of an overriding need tomaintain certain genes within the mitochondrion What-ever the reason for the retention of genes in organelles ithad better be a good one bioenergetic organelles areclearly the wrong place to keep genetic information andgo to extraordinary lengths to offset the consequences ofit being there
(ii) Molecular refugeesA phenomenon related to differences in the genetic code
is seen in chloroplasts This is a bias in codon usage Thelsquomolecular refugeesrsquo hypothesis (Howe et al 2000) sug-gests that nuclear copies of what were once chloroplastgenes escape constraints on codon usage and thus thatwhat distinguishes chloroplast-encoded proteins is thattheir amino-acid composition is unusual enough for thebias in codon usage not to apply It is difficult to see whatphysico-chemical properties might be uniquely shared bythe amino acids of chloroplast-encoded proteins and inaddition there seem to be no lsquoforbiddenrsquo amino-acid resi-due in chloroplasts As with the departure from the univer-sal genetic code in mitochondria is seems likely that abias in codon usage reflects some secondary property ofchloroplast genomes and is not the primary reason fortheir retention In any case any restriction on gene move-
Genomes in mitochondria and chloroplasts J F Allen 31
ment will act in the wrong direction to explain why chloro-plasts have retained genomes at all provided there is noresulting obstacle to a chloroplast protein being success-fully encoded as a precursor in the nucleus
(iii) Genes are copied and replicated not relocated in onestep
If complete relocation is envisaged as proceeding by thesimultaneous appearance of a gene in one compartmentand its deletion from another then it is possible to imaginethat whatever the vector is there may be selectivity ingene transfer (Henze amp Martin 2001) For example ifviral incorporation replication and recombination in bothorganelle and nucleus were the mechanism of relocationthen successful gene transfer might depend on the abilityof the virus to complete the process in one step This inturn might depend on some property of the gene or ofthe protein it encodes if expression occurs at any stage inthe process of relocation An assumption of the CORRhypothesis favoured here is that genes are not movedphysically but merely copied after which either the copyin the original location or the copy in the new locationmay be lost as a result of selection A useful analogy maybe the action of copying files between computer storagevolumes In this analogy the copy is identical to the orig-inal and either may subsequently be deleted without lossof information
Because any gene that is not usefully expressed willundergo no selection to maintain a functional allele theprevailing location will be the one in which the functionalcopy of the gene resides The problem of which gene sur-vives is then subsumed into the problem of which is theselectively advantageous location for it It seems reason-able to assume that there wasmdashand ismdashan interim periodin which homologous genes in different compartments areessentially in competition to see which functions best Asoutlined above there is no obvious obstacle for copyingDNA sequences within the cell The physico-chemicalproperties of DNA are fairly uniform and not directlydependent on nucleotide sequence though organelles likeintra-cellular parasites seem to prefer A and T to G andC The answer to the question lsquowhich gene location func-tions bestrsquo may have nothing to do with the gene itselfnor directly with the structure and function of its geneproduct Instead the answer may have everything to dowith the required regulatory interaction between the geneproduct and expression of the gene that produces it
(g) CORRThis hypothesis has been propounded earlier (sect 3) and
will be summarized as follows Vectorial electron and pro-ton transfer exerts regulatory control over expression ofgenes encoding proteins directly involved in or affectingredox poise This regulatory coupling is indispensable foradaptation to environmental changes in chemical activitiesof oxygen light and CO2 and requires co-location of thesegenes with their gene products Organelles lsquomake theirown decisionsrsquo on the basis of environmental changesaffecting redox state The hypothesis is abbreviatedCORR for co-location for redox regulation or more com-pletely lsquoco-location (of gene and gene product) for(evolutionary) continuity of redox regulation of geneexpressionrsquo
Phil Trans R Soc Lond B (2003)
5 SUMMARY OF THE CURRENT STATUS OF THEHYPOTHESIS OF CORR
Q Why do mitochondria and chloroplasts have theirown genetic systems
A CORR co-location of genes and gene products per-mits direct and autonomous redox regulation of geneexpression
(a) Objections to CORR(i) There is no regulation of gene expression in chloroplasts
and mitochondriaThis is still commonly stated but incorrect See the prin-
ciple in sect 3d
(ii) There is no regulation of gene expression in chloroplastsand mitochondria at the level of transcription
For chloroplasts there is now clear evidence for a func-tionally intelligible redox control of transcription(Pfannschmidt et al 1999a) This invalidates the previouswidely held assumption that all control of chloroplast geneexpression is post-transcriptional (Gruissem amp Tonkyn1993 GoldschmidtClermont 1998) The evidence formitochondria is less clear but redox control of transcrip-tion certainly cannot be ruled out and there are indi-cations that it may occur (Pearson et al 1993Konstantinov et al 1995 Wilson et al 1996b) It mustalso be restated that transcriptional control is not neces-sary for CORR but serves as a compelling example of aconserved typically bacterial mode of response that oper-ates still in eukaryotic cytoplasmic organelles thus illus-trating the principle of continuity of redox control (sect 3d)Post-transcriptional redox regulation (Danon amp Mayfield1994 Kim amp Mayfield 1997 Brown et al 2001 Treb-itsh amp Danon 2001) is consistent with CORR and thereis nothing to forbid it being a primary mechanism thatsatisfies most or indeed all of the formal requirements ofthe hypothesis in particular cases
(iii) There are no redox signalling components encoded inchloroplast or mitochondrial DNA
This is what CORR predicts See the principle in sect 3j
(iv) There is redox control of nuclear gene expressionThis is correct and many examples are now known
especially for chloroplasts (Escoubas et al 1995 Kovacs etal 2000 Pfannschmidt et al 2001ab Surpin et al 2002)Redox signals from photosynthetic or respiratory electrontransport chains are not at all prohibited by CORR Theessence of CORR is that there is a subset of organellarproteins for which there is an overriding requirement fora redox regulatory control that is initiated within theorganelle and whose effects are exerted by the simplestand most direct route This redox signalling is envisagedto take place without a requirement for prior consultationand coordination with what may be regarded as a nucleo-cytoplasmic bureaucracy
(v) Nuclear genes can be expressed as rapidly as chloroplastand mitochondrial genes
Zerges (2002) for example cites the equal times takenfor complete expression of nuclear and chloroplast genesin Chlamydomonas as evidence against the CORR hypoth-
32 J F Allen Genomes in mitochondria and chloroplasts
esis However the question is not one of the time takento express a gene but the time taken to alter the rate ofits expression in response to a (redox) signal originatingwithin the organelle An equally important additional fac-tor may be the autonomy of the signalling pathway thatis its independence from other inputs as described in theprevious paragraph One might say that mitochondria andchloroplasts require a free hand
(vi) There is no evidence to support this hypothesisThe hypothesis is rich in predictions (sectsect 3 and 6) and
those that have been tested stringently are in agreementwith experimental results None of the predictions of thehypothesis has been found to be incorrect and none of theunderlying principles (sect 3) has been disproved Furtherpredictions will be the subject of future experiments
(b) Evidence for CORR(i) Chloroplasts have selectively retained cyanobacterial genes
for primary events in photosynthesisSee table 1
(ii) There is regulation of gene expression in chloroplastsThere is transcriptional regulation of gene expression
in chloroplasts (sect 3d) The case for mitochondria is notyet proven
(iii) There is redox regulation of gene expression inchloroplasts
There is transcriptional redox regulation of geneexpression in chloroplasts (sect 3d) The case for mitochon-dria is not yet proven though see (Allen et al 1995 Kon-stantinov et al 1995 Galvis et al 1998)
(iv) There are no redox signalling components encoded inchloroplast or mitochondrial DNA
Correct but with possible exceptions discussed in sect 3j
(v) There are lsquobacterialrsquo redox signalling components ineukaryotes
See sect 3j
(vi) Further predictions DNA-array results and phenotypesof allotopic mutants
A systematic test of CORR might be to make an oxid-ized-minus-reduced gene expression profile of the lsquotran-scriptomersquo of a cyanobacterium to see if the set of geneswhose transcription is under most rapid and direct redoxcontrol corresponds to the genes of the chloroplast gen-ome Laboratories with this experimental capability do notseem to have addressed this possibility (Hihara et al 2002Ikeuchi 2002) Likewise if a putative modern representa-tive of the proto-mitochondrion can be found and agreedeg Rhodospirillum rubrum or Paracoccus denitrificans thena similar experiment should reveal a subset of genes corre-sponding to mitochondrial genomes DNA-microarraysalso present the possibility of testing the relative rate ofredox response and dependency on other signallinginputs of transcription of eukaryotic genes To dateorganellar genes have not been included in such investi-gations (Ferea et al 1999) A further experimental possi-bility is to examine the redox regulatory phenotypes ofexisting allotopic mutants as outlined in sect 3e
Phil Trans R Soc Lond B (2003)
6 CONCLUSION AND FUTURE PROSPECTS
The CORR hypothesis states that the function of mito-chondrial and chloroplast genomes is to provide co-location (of gene and gene product) for (evolutionary)continuity of redox regulation of gene expression Thishypothesis is neither self-contradictory nor devoid of test-able predictions It is also consistent with available evi-dence and in principle makes sense of the otherwisepuzzling distribution of genes between compartments ineukaryotic cells The explanation that CORR offersapplies equally to mitochondria and chloroplasts andidentifies selective forces that still operate to determine thelocation of genes No selective barriers need be assumedto gene transfer protein import nor membrane insertionNor do we need to invoke a lsquofrozen accidentrsquo that some-thing happened to stop gene transfer for some reason atsome time in the remote and inaccessible past CORR alsoexplains the absence of genomes from organelles that havelost their primary bioenergetic functions CORR may actas a stimulus to a search for residual bioenergetic functionin those organelles that have lost aerobic respiration andphotosynthesis but where residual genomes remain
CORR also predicts regulatory properties of knowncomponents of chloroplasts and mitochondria that in turnindicate important but as yet uncharacterized flexibility inenergy transduction These components include the RUB-ISCO large subunit and the CFo and Fo subunits of coup-ling ATPase in both chloroplasts and mitochondriaRedox regulation of the relative stoichiometry of compo-nents as seen in regulation of chloroplast photosystemstoichiometry (Pfannschmidt et al 1999a) may extend to(C)Fo-(C)F1-ATPase indicating that the function ofretention of genes for one or more of these componentsis flexibility in the stoichiometries of H1 to e2 and H1 toATP in chemiosmotic coupling (Schemidt et al 1998Stock et al 2000 Allen 2002)
So far explicit tests of CORR have largely been con-fined to chloroplasts and to some extent mitochondria ofhigher plants but this merely reflects the expertise andinterests of the investigators Further attempted disproofof CORR is welcomed and encouraged Novel experi-mental tests can be devised as well as attempted refu-tation through analysis of existing data It is hoped thatexplicit attention will be given to the CORR hypothesis inlaboratories already suitably equipped and that resourcescan be allocated for this purpose in laboratories wherethere is the interest but not currently the technology orexpertise
The heuristic value of the hypothesis already seen inthe role of redox-regulated transcription of photosyntheticreaction centre genes in chloroplasts may be its most use-ful property The central proposal is that mitochondriaand chloroplasts contain genes for proteins whose functionin electron transfer demands rapid direct and uncon-ditional redox regulatory control of their biosynthesis Thefull implications of this hypothesis are very broad and onlyjust beginning to be explored
I thank Carol Allen Aubrey de Grey William Martin John ARaven and Francis-Andre Wollman for discussion and corre-spondence during the preparation of this paper and WilliamMartin for suggestions including table 1 Research grants from
Genomes in mitochondria and chloroplasts J F Allen 33
The Swedish Natural Sciences Research Council (NFR) andthe Crafoord Foundation are gratefully acknowledged
REFERENCES
Adams K L Daley D O Qiu Y L Whelan J amp PalmerJ D 2000 Repeated recent and diverse transfers of a mito-chondrial gene to the nucleus in flowering plants Nature408 354ndash357
Alberts B Bray D Lewis J Raff M Roberts K amp Wat-son J D 1994 The molecular biology of the cell New Yorkand London Garland Publishing
Allen C A Hakansson G amp Allen J F 1995 Redox con-ditions specify the proteins synthesized by isolated chloro-plasts and mitochondria Redox Rep 1 119ndash123
Allen J F 1992 Protein phosphorylation in regulation ofphotosynthesis Biochim Biophys Acta 1098 275ndash335
Allen J F 1993a Control of gene expression by redox poten-tial and the requirement for chloroplast and mitochondrialgenomes J Theor Biol 165 609ndash631
Allen J F 1993b Redox control of gene expression and thefunction of chloroplast genomes a hypothesis PhotosynthRes 36 95ndash102
Allen J F 1993c Redox control of transcription sensorsresponse regulators activators and repressors FEBS Lett332 203ndash207
Allen J F 1995 Thylakoid proteinndashphosphorylation state-1ndashstate-2 transitions and photosystem stoichiometry adjust-ment redox control at multiple levels of gene expressionPhysiol Plant 93 196ndash205
Allen J F 1996 Separate sexes and the mitochondrial theoryof ageing J Theor Biol 180 135ndash140
Allen J F 2002 Photosynthesis of ATP-electrons protonpumps rotors and poise Cell 110 273ndash276
Allen J F amp Allen C A 1999 A mitochondrial model forpremature ageing of somatically cloned mammals IUBMBLife 48 369ndash372
Allen J F amp Nilsson A 1997 Redox signalling and the struc-tural basis of regulation of photosynthesis by protein phos-phorylation Physiol Plant 100 863ndash868
Allen J F amp Pfannschmidt T 2000 Balancing the two photo-systems photosynthetic electron transfer governs transcrip-tion of reaction centre genes in chloroplasts Phil TransR Soc Lond B 355 1351ndash1357 (DOI 101098rstb20000697)
Allen J F amp Race H L 2002 Will the real LHC II kinaseplease step forward Sciencersquos STKE httpwwwstkeorgcgicontentfullsigtrans2002155pe43
Allen J F amp Raven J A 1996 Free-radical-induced mutationvs redox regulation costs and benefits of genes in organellesJ Mol Evol 42 482ndash492
Archibald J M Cavalier-Smith T Maier U amp Douglas S2001 Molecular chaperones encoded by a reduced nucleusthe cryptomonad nucleomorph J Mol Evol 52 490ndash501
Ashby M K amp Mullineaux C W 1999a Cyanobacterialycf27 gene products regulate energy transfer from phycobili-somes to photosystems I and II FEMS Microbiol Lett 181253ndash260
Ashby M K amp Mullineaux C W 1999b The role of ApcDand ApcF in energy transfer from phycobilisomes to PSI andPSII in a cyanobacterium Photosynth Res 61 169ndash179
Ashby M K Houmard J amp Mullineaux C W 2002 Theycf27 genes from cyanobacteria and eukaryotic algae distri-bution and implications for chloroplast evolution FEMSMicrobiol Lett 214 25ndash30
Attardi G amp Schatz G 1988 Biogenesis of mitochondria ARev Cell Biol 4 289ndash333
Azem A Oppliger W Lustig A Jeno P Feifel B SchatzG amp Horst M 1997 The mitochondrial hsp70 chaperone
Phil Trans R Soc Lond B (2003)
system effect of adenine nucleotides peptide substrate andmGrpE on the oligomeric state of mhsp70 J Biol Chem272 20 901ndash20 906
Barbrook A C Symington H Nisbet R E R LarkumA amp Howe C J 2001 Organisation and expression of theplastid genome of the dinoflagellate Amphidinium opercula-tum Mol Genet Genomics 266 632ndash638
Bauer C E Elsen S amp Bird T H 1999 Mechanisms forredox control of gene expression A Rev Microbiol 53495ndash523
Bauer C E Elsen S Swem L R Swem D L amp MasudaS 2003 Redox and light regulation of gene expression inphotosynthetic prokaryotes Phil Trans R Soc Lond B 358147ndash154 (DOI 101098rstb20021189)
Berks B C Sargent F De Leeuw E Hinsley A P Stan-ley N R Jack R L Buchanan G amp Palmer T 2000aA novel protein transport system involved in the biogenesisof bacterial electron transfer chains Biochim Biophys ActaBioenerg 1459 325ndash330
Berks B C Sargent F amp Palmer T 2000b The tat proteinexport pathway Mol Microbiol 35 260ndash274
Blackstone N W 1999 Redox control in development andevolution evidence from colonial hydroids J Exp Biol 2023541ndash3553
Blackstone N W 2000 Redox control and the evolution ofmulticellularity Bioessays 22 947ndash953
Blackstone N W 2001 Redox state reactive oxygen speciesand adaptive growth in colonial hydroids J Exp Biol 2041845ndash1853
Bogorad L 1975 Evolution of organelles and eukaryotic gen-omes Science 188 891ndash898
Bogsch E G Sargent F Stanley N R Berks B C Robin-son C amp Palmer T 1998 An essential component of a novelbacterial protein export system with homologues in plastidsand mitochondria J Biol Chem 273 18 003ndash18 006
Brown E C Somanchi A amp Mayfield S P 2001 Interor-ganellar crosstalk new perspectives on signaling from thechloroplast to the nucleus Genome Biol 2 10211ndash10214
Cavalier-Smith T 2002 Chloroplast evolution secondarysymbiogenesis and multiple losses Curr Biol 12 R62ndashR64
Chitnis P R amp Thornber J P 1988 The major light-har-vesting complex of photosystem 2 Aspects of its molecularand cell biology Photosynth Res 16 41ndash63
Chow W S Melis A amp Anderson J M 1990 Adjustmentsof photosystem stoichiometry in chloroplasts improve thequantum efficiency of photosynthesis Proc Natl Acad SciUSA 87 7502ndash7506
Claros M G Perea J Shu Y Samatey F A PopotJ L amp Jacq C 1995 Limitations to in vivo import of hydro-phobic proteins into yeast mitochondria The case of a cyto-plasmically synthesized apocytochrome b Eur J Biochem228 762ndash771
Dalbey R E amp Robinson C 1999 Protein translocation intoand across the bacterial plasma membrane and the plant thy-lakoid membrane Trends Biochem Sci 24 17ndash22
Daley D O Adams K L Clifton R Qualmann S MillarA H Palmer J D Pratje E amp Whelan J 2002 Genetransfer from mitochondrion to nucleus novel mechanismsfor gene activation from Cox2 Plant J 30 11ndash21
Danon A amp Mayfield S P 1994 Light-regulated translationof chloroplast messenger RNAs through redox potentialScience 266 1717ndash1719
de Grey A 2000 Mitochondrial gene therapy an arena for thebiomedical use of inteins Trends Biotechnol 18 394ndash399
Deisenhofer J Epp O Miki K Huber R amp Michel H1985 Structure of the protein subunits in the photosyntheticreaction center of Rhodopseudomonas viridis at 3Ac resolutionNature 318 618ndash624
34 J F Allen Genomes in mitochondria and chloroplasts
Delwiche C F 1999 Tracing the thread of plastid diversitythrough the tapestry of life Am Nat 154 S164ndashS177
Doolittle W F 1999 Phylogenetic classification and the uni-versal tree Science 284 2124ndash2129
Douglas S E 1998 Plastid evolution origins diversitytrends Curr Opin Genet Dev 8 655ndash661
Duby G amp Boutry M 2002 Mitochondrial protein importmachinery and targeting information Plant Sci 162 477ndash490
Eberhard S Drapier D amp Wollman F A 2002 Searchinglimiting steps in the expression of chloroplast encoded pro-teins relations between gene copy number transcriptiontranscript abundance and translation rate in the chloroplastof Chlamydomonas reinhardtii Plant J 31 149ndash160
El Bissati K amp Kirilovsky D 2001 Regulation of psbA andpsaE expression by light quality in Synechocystis species PCC6803 A redox control mechanism Plant Physiol 1251988ndash2000
Ellis R J 1984 The nuclear domination of chloroplast devel-opment Science Progress 69 129ndash142
Embley T M amp Martin W 1998 Molecular evolution ahydrogen-producing mitochondrion Nature 396 517ndash519
Ems S C Morden C W Dixon C K Wolfe K HdePamphilis C W amp Palmer J D 1995 Transcriptionsplicing and editing of plastid RNAs in the non-photosyn-thetic plant Epifagus virginiana Plant Mol Biol 29 721ndash733
Eraso J M amp Kaplan S 2000 From redox flow to gene regu-lation role of the PrrC protein of Rhodobacter sphaeroides241 Biochemistry 39 2052ndash2062
Eraso J M amp Kaplan S 2002 Redox flow as an instrumentof gene regulation Meth Enzymol 348 216ndash229
Escoubas J M Lomas M Laroche J amp Falkowski P G1995 Light-intensity regulation of cab gene-transcription issignaled by the redox state of the plastoquinone pool ProcNatl Acad Sci USA 92 10 237ndash10 241
Ferea T L Botstein D Brown P O amp Rosenzweig R F1999 Systematic changes in gene expression patterns follow-ing adaptive evolution in yeast Proc Natl Acad Sci USA96 9721ndash9726
Forsberg J Rosenquist M Fraysse L amp Allen J F 2001Redox signalling in chloroplasts and mitochondria genomicand biochemical evidence for two-component regulatory sys-tems in bioenergetic organelles Biochem Soc Trans 29403ndash407
Fujita Y 1997 A study on the dynamic features of photosys-tem stoichiometry accomplishments and problems forfuture studies Photosynth Res 53 83ndash93
Gabriel K Buchanan S K amp Lithgow T 2001 The alphaand the beta protein translocation across mitochondrial andplastid outer membranes Trends Biochem Sci 26 36ndash40
Galvis M L E Allen J F amp Hakansson G 1998 Proteinsynthesis by isolated pea mitochondria is dependent on theactivity of respiratory complex II Curr Genet 33 320ndash329
Gatenby A A amp Ellis R J 1990 Chaperone function theassembly of ribulose bisphosphate carboxylase-oxygenase ARev Cell Biol 6 125ndash149
GoldschmidtClermont M 1998 Coordination of nuclear andchloroplast gene expression in plant cells Int Rev CytolSurv Cell Biol 177 115ndash180
Gray M W 1992 The endosymbiont hypothesis revisited IntRev Cytol 141 233ndash357
Gray M W 1999 Evolution of organellar genomes CurrOpin Genet Dev 9 678ndash687
Gray M W 2000 Evolutionary biology mitochondrial geneson the move Nature 408 302ndash305
Gray M W amp Doolittle W F 1982 Has the endosymbionthypothesis been proven Microbiol Rev 46 1ndash42
Gray M W Burger G amp Lang B F 1999 Mitochondrialevolution Science 283 1476ndash1481
Phil Trans R Soc Lond B (2003)
Grebe T W amp Stock J B 1999 The histidine protein kinasesuperfamily Adv Microb Physiol 41 139ndash227
Green B R amp Kuhlbrandt W 1995 Sequence conservationof light-harvesting and stress-response proteins in relation tothe 3-dimensional molecular structure of Lhcii PhotosynthRes 44 139ndash148
Gruissem W amp Tonkyn J C 1993 Control mechanisms ofplastid gene expression Crit Rev Plant Sci 12 19ndash55
Hackstein J H P (and 11 others) 2001 Hydrogenosomesconvergent adaptations of mitochondria to anaerobicenvironments Zool Anal Complex Syst 104 290ndash302
Harrison M A Keen J N Findlay J B C amp Allen J F1990 Modification of a Glnb-like gene-product by photosyn-thetic electron-transport in the cyanobacterium Synechoc-occus 6301 FEBS Lett 264 25ndash28
Harvey A J Kind K L amp Thompson J G 2002 Redoxregulation of early embryo development Reproduction 123479ndash486
Heathcote P Fyfe P K amp Jones M R 2002 Reactioncentres the structure and evolution of biological solarpower Trends Biochem Sci 27 79ndash87
Henze K amp Martin W 2001 How do mitochondrial genesget into the nucleus Trends Genet 17 383ndash387
Herrmann R G amp Westhoff P 2001 Thylakoid biogenesis anddynamics the result of a complex phylogenetic puzzle InRegulation of photosynthesis vol 11 (ed E-M Aro amp BAndersson) pp 1ndash28 Dordrecht The Netherlands Kluwer
Hihara Y Sonoike K Kanehisa M amp Ikeuchi M 2002Search for redox responsive genes in Synechocystis sp PCC6803 by using DNA microarrays Plant Cell Physiol 43S178
Howe C J Barbrook A C amp Lockhart P J 2000 Organellegenes do they jump or are they pushed Trends Genet 1665ndash66
Ikeuchi M 2002 Gene expression and responses to light stressin cyanobacteria Plant Cell Physiol 43 S21
Iuchi S amp Lin E C C 1992 Mutational analysis of signaltransduction by Arcb a membrane sensor protein respon-sible for anaerobic repression of operons involved in the cen-tral aerobic pathways in Escherichia coli J Bacteriol 1743972ndash3980
Jackson-Constan D Akita M amp Keegstra K 2001 Molecu-lar chaperones involved in chloroplast protein importBiochim Biophys Acta Mol Cell Res 1541 102ndash113
Jarvis P amp Soll M 2001 Toc Tic and chloroplast proteinimport Biochim Biophys Acta Mol Cell Res 1541 64ndash79
Jordan P Fromme P Witt H T Klukas O SaengerW amp Krauss N 2001 Three-dimensional structure ofcyanobacterial photosystem I at 25 acngstrom resolutionNature 411 909ndash917
Kaneko T (and 23 others) 1996a Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regionsDNA Res 3 109ndash136
Kaneko T (and 23 others) 1996b Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regions[Supplement] DNA Res 3 185ndash209
Kanevski I amp Maliga P 1994 Relocation of the plastid rbcLgene to the nucleus yields functional ribulose-15-bisphosph-ate carboxylase in tobacco chloroplasts Proc Natl Acad SciUSA 91 1969ndash1973
Kim J amp Mayfield S P 1997 Protein disulfide isomerase as aregulator of chloroplast translational activation Science 2781954ndash1957
Genomes in mitochondria and chloroplasts J F Allen 35
Kirk J T O amp Tilney-Bassett R A E 1978 The plastidsTheir chemistry structure growth and inheritance Amster-dam Elsevier
Koehler C M Merchant S amp Schatz G 1999 How mem-brane proteins travel across the mitochondrial intermem-brane space Trends Biochem Sci 24 428ndash432
Komiya T Rospert S Schatz G amp Mihara K 1997 Bind-ing of mitochondrial precursor proteins to the cytoplasmicdomains of the import receptors Tom70 and Tom20 isdetermined by cytoplasmic chaperones FASEB J 11 1196
Konstantinov Y M Lutsenko G N amp Podsosonny V A1995 Genetic functions of isolated maize mitochondriaunder model changes of redox conditions Biochem MolBiol Int 36 319ndash326
Korab-Laskowska M Rioux P Brossard N LittlejohnT G Gray M W Lang B F amp Burger G 1998 Theorganelle genome database project (GOBASE) Nucleic AcidsRes 26 138ndash144
Kovacs L Wiessner W Kis M Nagy F Mende D ampDemeter S 2000 Short- and long-term redox regulation ofphotosynthetic light energy distribution and photosystemstoichiometry by acetate metabolism in the green alga Chla-mydobotrys stellata Photosynth Res 65 231ndash247
Kuhlbrandt W 1994 Structure and function of the plant light-harvesting complex LHC-II Curr Opin Struct Biol 4519ndash528
Lang B F Gray M W amp Burger G 1999 Mitochondrialgenome evolution and the origin of eukaryotes A RevGenet 33 351ndash397
Lemire B D Fankhauser C Baker A amp Schatz G 1989The mitochondrial targeting function of randomly generatedpeptide sequences correlates with predicted helical amphi-philicity J Biol Chem 264 20 206ndash20 215
Leuenberger D Bally N A Schatz G amp Koehler C M1999 Different import pathways through the mitochondrialintermembrane space for inner membrane proteins EMBOJ 18 4816ndash4822
Li H amp Sherman L A 2000 A redox-responsive regulator ofphotosynthesis gene expression in the cyanobacterium Syne-chocystis sp strain PCC 6803 J Bacteriol 182 4268ndash4277
Lin X Y (and 36 others) 1999 Sequence and analysis of chro-mosome 2 of the plant Arabidopsis thaliana Nature 402761ndash768
Lonsdale D M Hodge T P Howe C J amp Stern D B1983 Maize mitochondrial-DNA contains a sequence hom-ologous to the ribulose-15-bisphosphate carboxylase largesubunit gene of chloroplast DNA Cell 34 1007ndash1014
McFadden G I 1999 Endosymbiosis and evolution of theplant cell Curr Opin Plant Biol 2 513ndash519
McFadden G I 2001 Chloroplast origin and integrationPlant Physiol 125 50ndash53
Mahler H R amp Raff R A 1975 The evolutionary origin ofthe mitochondrion a nonsymbiotic model Int Rev Cytol43 1ndash124
Manfredi G Fu J Ojaimi J Sadlock J E Kwong J QGuy J amp Schon E A 2002 Rescue of a deficiency in ATPsynthesis by transfer of MTATP6 a mitochondrial DNA-encoded gene to the nucleus Nature Genet 30 394ndash399
Marc P Margeot A Devaux F Blugeon C Corral-Debrinski M amp Jacq C 2002 Genome-wide analysis ofmRNAs targeted to yeast mitochondria EMBO Rep 3159ndash164
Marienfeld J Unseld M amp Brennicke A 1999 The mito-chondrial genome of Arabidopsis is composed of both nativeand immigrant information Trends Plant Sci 4 495ndash502
Martin W 1999 A briefly argued case that mitochondria andplastids are descendants of endosymbionts but that thenuclear compartment is not Proc R Soc Lond B 2661387ndash1395 (DOI 101098rspb19990792)
Phil Trans R Soc Lond B (2003)
Martin W amp Herrmann R G 1998 Gene transfer fromorganelles to the nucleus how much what happens andwhy Plant Physiol 118 9ndash17
Martin W amp Muller M 1998 The hydrogen hypothesis forthe first eukaryote Nature 392 37ndash41
Martin W amp Russell M J 2003 On the origins of cells ahypothesis for the evolutionary transitions from abiotic geo-chemistry to chemoautotrophic prokaryotes and from pro-karyotes to nucleated cells Phil Trans R Soc Lond B 35859ndash85 (DOI 101098rstb20021183)
Martin W Stoebe B Goremykin V Hansmann S Hase-gawa M amp Kowallik K V 1998 Gene transfer to thenucleus and the evolution of chloroplasts Nature 393162ndash165
Martin W Hoffmeister M Rotte C amp Henze K 2001 Anoverview of endosymbiotic models for the origins of eukary-otes their ATP-producing organelles (mitochondria andhydrogenosomes) and their heterotrophic lifestyle BiolChem 382 1521ndash1539
Martin W Rujan T Richly E Hansen A Cornelsen SLins T Leister D Stoebe B Hasegawa M amp PennyD 2002 Evolutionary analysis of Arabidopsis cyanobacterialand chloroplast genomes reveals plastid phylogeny and thou-sands of cyanobacterial genes in the nucleus Proc NatlAcad Sci USA 99 12 246ndash12 251
Millen R S (and 12 others) 2001 Many parallel losses of infAfrom chloroplast DNA during angiosperm evolution withmultiple independent transfers to the nucleus Plant Cell 13645ndash658
Morse D Salois P Markovic P amp Hastings J W 1995 Anuclear-encoded form-II RUBISCO in dinoflagellatesScience 268 1622ndash1624
Murakami A Kim S J amp Fujita Y 1997 Changes in photo-system stoichiometry in response to environmental con-ditions for cell growth observed with the cyanophyteSynechocystis PCC 6714 Plant Cell Physiol 38392ndash397
Oh J I amp Kaplan S 2000 Redox signaling globalization ofgene expression EMBO J 19 4237ndash4247
Ozawa T 1995 Mitochondrial-DNA mutations associatedwith aging and degenerative diseases Exp Gerontol 30269ndash290
Palmer J D 1997 Organelle genomes going going goneScience 275 790ndash791
Pearson C K Wilson S B Schaffer R amp Ross A W 1993NAD turnover and utilization of metabolites for RNA syn-thesis in a reaction sensing the redox state of the cyto-chrome-b( 6 )f complex in isolated chloroplasts Eur JBiochem 218 397ndash404
Pfanner N amp Geissler A 2001 Versatility of the mitochon-drial protein import machinery Natl Rev Mol Cell Biol 2339ndash349
Pfanner N amp Wiedemann N 2002 Mitochondrial proteinimport two membranes three translocases Curr Opin CellBiol 14 400ndash411
Pfannschmidt T Nilsson A amp Allen J F 1999a Photosyn-thetic control of chloroplast gene expression Nature 397625ndash628
Pfannschmidt T Nilsson A Tullberg A Link G amp AllenJ F 1999b Direct transcriptional control of the chloroplastgenes psbA and psaAB adjusts photosynthesis to light energydistribution in plants IUBMB Life 48 271ndash276
Pfannschmidt T Allen J F amp Oelmuller R 2001a Prin-ciples of redox control in photosynthesis gene expressionPhysiol Plant 112 1ndash9
Pfannschmidt T Schutze K Brost M amp Oelmuller R2001b A novel mechanism of nuclear photosynthesis generegulation by redox signals from the chloroplast during pho-
36 J F Allen Genomes in mitochondria and chloroplasts
tosystem stoichiometry adjustment J Biol Chem 27636 125ndash36 130
Popot J L amp de Vitry C 1990 On the microassembly of inte-gral membrane proteins A Rev Biophys Biophys Chem 19369ndash403
Race H L Herrmann R G amp Martin W 1999 Why haveorganelles retained genomes Trends Genet 15 364ndash370
Raven J A Johnston A M Parsons R amp Kubler J 1994aThe influence of natural and experimental high O2 concen-trations on O2 evolving phototrophs Biol Rev CambridgePhil Soc 69 61ndash94
Raven J A Johnston A M Parsons R amp Kubler J 1994bThe occurrence and influence on photolithotrophs of highoxygen concentrations Proc R Soc Edinb B 102 193ndash201
Robinson C 2000 The twin-arginine translocation system anovel means of transporting folded proteins in chloroplastsand bacteria Biol Chem 381 89ndash93
Robinson C amp Bolhuis A 2001 Protein targeting by the twin-arginine translocation pathway Natl Rev Mol Cell Biol 2350ndash356
Rodermel S 2001 Pathways of plastid-to-nucleus signalingTrends Plant Sci 6 471ndash478
Roger A J 1999 Reconstructing early events in eukaryoticevolution Am Nat 154 S146ndashS163
Roger A J Svard S G Tovar J Clark C G SmithM W Gillin F D amp Sogin M L 1998 A mitochondrial-like chaperonin 60 gene in Giardia lamblia evidence thatdiplomonads once harbored an endosymbiont related to theprogenitor of mitochondria Proc Natl Acad Sci USA 95229ndash234
Roodyn D B amp Wilkie D 1968 The biogenesis of mitochondriaMethuenrsquos monographs on biological subjects London Methuen
Rujan T amp Martin W 2001 How many genes in Arabidopsiscome from cyanobacteria An estimate from 386 proteinphylogenies Trends Genet 17 113ndash120
Saccone C Gissi C Lanave C Larizza A Pesole G ampReyes A 2000 Evolution of the mitochondrial genetic sys-tem an overview Gene 261 153ndash159
Salzberg S L White O Peterson J amp Eisen J A 2001Microbial genes in the human genome lateral transfer orgene loss Science 292 1903ndash1906
Sargent F Berks B C amp Palmer T 2002 Assembly of mem-brane-bound respiratory complexes by the Tat protein-trans-port system Arch Microbiol 178 77ndash84
Schatz G 1998 Protein transport the doors to organellesNature 395 439ndash440
Schemidt R A Qu J Williams J R amp Brusilow W S1998 Effects of carbon source on expression of F0 genes andon the stoichiometry of the c subunit in the F1 F0 ATPaseof Escherichia coli J Bacteriol 180 3205ndash3208
Selosse M A Albert B R amp Godelle B 2001 Reducing thegenome size of organelles favours gene transfer to thenucleus Trends Ecol Evol 16 135ndash141
Stern D B amp Lonsdale D M 1982 Mitochondrial and chlo-roplast genomes of maize have a 12- kilobase DNA-sequencein common Nature 299 698ndash702
Stock D Gibbons C Arechaga I Leslie A G W ampWalker J E 2000 The rotary mechanism of ATP synthaseCurr Opin Struct Biol 10 672ndash679
Stoebe B amp Maier U G 2002 One two three naturersquos toolbox for building plastids Protoplasma 219 123ndash130
Stoebe B Martin W amp Kowallik K V 1998 Distributionand nomenclature of protein-coding genes in 12 sequencedchloroplast genomes Plant Mol Biol Rep 16 243ndash255
Stupar R M Lilly J W Town C D Cheng Z Kaul SBuell C R amp Jiang J M 2001 Complex mtDNA consti-tutes an approximate 620 kb insertion on Arabidopsis thali-ana chromosome 2 implication of potential sequencing
Phil Trans R Soc Lond B (2003)
errors caused by large-unit repeats Proc Natl Acad SciUSA 98 5099ndash5103
Surpin M Larkin R M amp Chory J 2002 Signal transduc-tion between the chloroplast and the nucleus Plant Cell 14S327ndashS338
Tjaden J Schwoppe C Mohlmann T Quick P W ampNeuhaus H E 1998 Expression of a plastidic ATPADPtransporter gene in Escherichia coli leads to a functional aden-ine nucleotide transport system in the bacterial cytoplasmicmembrane J Biol Chem 273 9630ndash9636
Tovar J Fischer A amp Clark C G 1999 The mitosome a novelorganelle related to mitochondria in the amitochondrial para-site Entamoeba histolytica Mol Microbiol 32 1013ndash1021
Trebitsh T amp Danon A 2001 Translation of chloroplastpsbA mRNA is regulated by signals initiated by both photos-ystems II and I Proc Natl Acad Sci USA 98 12 289ndash12 294
Tullberg A Alexciev K Pfannschmidt T amp Allen J F2000 Photosynthetic electron flow regulates transcription ofthe psaB gene in pea (Pisum sativum L) chloroplasts throughthe redox state of the plastoquinone pool Plant Cell Physiol41 1045ndash1054
Turmel M Lemieux C Burger G Lang B F Otis CPlante I amp Gray M W 1999 The complete mitochondrialDNA sequences of Nephroselmis olivacea and Pedinomonasminor two radically different evolutionary patterns withingreen algae Plant Cell 11 1717ndash1729
Unden G amp Bongaerts J 1997 Alternative respiratory path-ways of Escherichia coli energetics and transcriptional regu-lation in response to electron acceptors Biochim BiophysActa Bioenerg 1320 217ndash234
Unseld M Marienfeld J R Brandt P amp Brennicke A1997 The mitochondrial genome of Arabidopsis thaliana con-tains 57 genes in 366 924 nucleotides Nature Genet 1557ndash61
Van der Giezen M Slotboom D J Horner D S DyalP L Harding M Xue G P Embley T M amp KunjiE R S 2002 Conserved properties of hydrogenosomal andmitochondrial ADPATP carriers a common origin for bothorganelles EMBO J 21 572ndash579
Von Heijne G 1986 Why mitochondria need a genome FEBSLett 198 1ndash4
Westphal S Soll J amp Vothknecht U C 2001 A vesicle trans-port system inside chloroplasts FEBS Lett 506 257ndash261
Whitehouse H L K 1969 Towards an understanding of themechanism of heredity London Edward Arnold Ltd
Williams B A P Hirt R P Lucocq J M amp Embley T M2002 A mitochondrial remnant in the microsporidian Trachi-pleistophora hominis Nature 418 865ndash869
Wilson R J (and 10 others) 1996a Complete gene map of theplastid-like DNA of the malaria parasite Plasmodium falcipa-rum J Mol Biol 261 155ndash172
Wilson S B Davidson G S Thomson L M amp PearsonC K 1996b Redox control of RNA synthesis in potato mito-chondria Eur J Biochem 242 81ndash85
Winkler H H amp Neuhaus H E 1999 Non-mitochondrialATP transport Trends Biochem Sci 24 64ndash68
Wolfe K H Morden C W amp Palmer J D 1992 Small sin-gle-copy region of plastid DNA in the non-photosyntheticangiosperm Epifagus virginiana contains only two genes Dif-ferences among dicots monocots and bryophytes in geneorganization at a non-bioenergetic locus J Mol Biol 22395ndash104
Wollman F A Minai L amp Nechushtai R 1999 The biogen-esis and assembly of photosynthetic proteins in thylakoidmembranes Biochim Biophys Acta Bioenerg 1411 21ndash85
Yeliseev A A amp Kaplan S 2000 TspO of Rhodobactersphaeroides a structural and functional model for the mam-
Genomes in mitochondria and chloroplasts J F Allen 37
malian peripheral benzodiazepine receptor J Biol Chem275 5657ndash5667
Zerges W 2002 Does complexity constrain organelle evol-ution Trends Plant Sci 7 175ndash182
Zhang Z D Green B R amp Cavalier-Smith T 1999 Singlegene circles in dinoflagellate chloroplast genomes Nature400 155ndash159
Zhang Z D Cavalier-Smith T amp Green B R 2001 A fam-ily of selfish minicircular chromosomes with jumbled chloro-plast gene fragments from a dinoflagellate Mol Biol Evol18 1558ndash1565
Zouni A Witt H T Kern J Fromme P Krauss NSaenger W amp Orth P 2001 Crystal structure of photosys-tem II from Synechococcus elongatus at 38 acngstrom resol-ution Nature 409 739ndash743
DiscussionD S Horner (Dipartimento di Fisiologia e Biochimica
Generali University of Milan Milan Italy) Given thatthere are components of these membrane-bound energy-transduction pathways that are consistently nuclear enco-ded not the key ones you pointed out but others do youexpect perhaps a slower perhaps a more crude but yet aredox communication between the organelle andexpression of nuclear-encoded genes
J F Allen There is and there is no question about itI think that Bogoradrsquos lock-in hypothesis has somethingto say here and there are other people who say it is all todo with assembling multi-subunit complexes If you havecontrol over the initial construction of the core aroundwhich the whole complex is eventually built then redoxcontrol or some sort of control over synthesis of that corewill control construction of the whole thing eventuallyand I think that may well be largely correct I think it ispossible in principle for nuclear genes to make precursorsaccording to redox instructions but it is not really soimportant that those precursors are here and now presentin exactly the right quantities because the precursors canto a limited extent hang around and wait Actually thereis proteolytic degradation of unattached precursors and soon but I think the fact that these chloroplast and mito-chondrially DNA-encoded subunits are at the core ofthese energy-transducing complexes is the crucial issue
T Cavalier-Smith (Department of Zoology University ofOxford Oxford UK ) In the case of mitochondria the sub-set of genes that remains is much more variable than youimplied It runs from three proteins in the minimal casesup to I think over 90 That is a big difference and manyof the genes in different lineages of protists that remainhave nothing to do with redox control they are things likeribosomal proteins and so on So I think that even thoughwhat you say about redox control of certain genes mayactually be an important thing it is not the fundamentaloverall reason why the genomes are maintained It may bepart of the explanation for why certain genes are alwaysincluded in that subset but even in mitochondria I thinkthat cytochrome b is the only one that is always presentSo I think that both this great diversity of what has beenretained in different lineages and the nucleomorph caseindicate that evolution was able to do one thing in onelineage 500 million years ago and never got around todoing it for 500 million years in another lineage so thereis a lot more haphazardness and evolutionary accidentsand contingency than a straight functional interpretation
Phil Trans R Soc Lond B (2003)
there is a lot of just sheer lsquohappenedndashorndashdidnrsquotndashhappenrsquoor carryover
J F Allen I was not implying anything about the rangeof mitochondrial genomes I have a limited amount oftime so I just talked about what I could talk about withinthat limit Of course there will be a range and this willdepend upon the physiology and bioenergetics of theparticular organism But what I am trying to do is to seewhat the common properties of the irreducible core areof components that are always encoded in the organellesand cytochrome b is a perfect example because that reallyis the thing that binds these two all-important quinoneswithout which there is no Q cycle without which there isno proton pumping and bioenergetics and in Plasmodiumwhere the mitochondrion is reduced to this bare mini-mum there is still a mitochondrial gene for cytochromeb Now my argument about the ribosomal proteins andall the rest of it is that in order to be able to exert thisall-important redox control over gene expression of certaincore components you need an in situ genetic system andthis can vary in composition for reasons that I cannotdescribe Maybe you need RNA polymerase or maybeyou can import RNA polymerase In plastids you haveboth of these options You are going to need ribosomestransfer RNAs and the rest of the apparatus of geneexpression and that needs to be in situ but we are stillleft with the question why does there have to be any gen-etic system at all The mature tobacco or spinach chloro-plast seems to have an entire genetic system just to makeone protein D1 which is all it ever needs to resynthesizeI think that this is a clue It is worth having this very unec-onomical crazy system because there is an overridingrequirement to connect electron transfer with geneexpression directly and unambiguouslymdashwithout recourseto a bureaucracy if you like which is what I think thenucleus is
K Willison (Institute of Cancer Research Chester BeattyLaboratories London UK ) These photosynthetic reactioncomplexes are a sort of baroque mixture of polypeptidechains and cofactors and my understanding is that someof them in high photon flux can turn over within say 30minutes so I am wondering if there is a correlationbetween gene retention and the turnover rates of some ofthe components and the complexes it seems to me thatthat could really be a limiting factor and the reason thatthe proteins had to be encoded locally in order to get thecomplexes to be assembled rapidly
J F Allen I think that is right You can only alter thesteady-state concentration of a protein complex by affect-ing the synthesis and breakdown and therefore turnoverThere is an extreme example the D1 protein encoded bythe psbA gene of chloroplasts which is always in the chlor-oplast genome without exception High light will tend todegrade this protein and when the high light goes awaythere is an absolute need very quickly to bring it back upagain I think that is a very good example Whether thisargument applies to genes such as cytochrome oxidase Ido not know
C J Leaver (Department of Plant Sciences University ofOxford Oxford UK ) Your experiments tend to relatemainly to the gene expression side What about proteinRNA turnover the whole concept of redox control ofthatmdashdo we know anything
38 J F Allen Genomes in mitochondria and chloroplasts
J F Allen Well quite a lot is known about chloro-plasts There is redox control of messenger RNA stabilityfor this same D1 protein which has been described byStephen Mayfield and his co-workers The evidence is thatthe site of redox control is ferredoxin via thioredoxin andthe implication is that thioredoxin tells whether non-cyclicelectron transport is working at all When it is workingthere is a signal that says lsquoOK now continue with all theprotein synthesisrsquo It is not particularly specific but moreof a global effect I think this is what Mayfield argues butof course this is not inconsistent with my argument I haveconcentrated on quinone pools and medium-potentialredox regulatory control because that is where I can seesignals about the dangerous consequences of getting itwrong but there is control of other levels of geneexpression In mitochondria I am not so sure I wouldbounce the question back to you on that one
W Martin (Institute of Botany III Heinrich-Heine Univ-ersitat Dusseldorf Dusseldorf Germany) The nucleomorphgenome was cited as evidence in conflict with your modelbut is that true The nucleomorph genome is not in abioenergetic organelle in that sense It resides in the cyto-sol so does the size of the nucleomorph genome workagainst your model Is it really evidence against yourmodel
J F Allen I do not know why should it be evidenceI cannot explain why the nucleomorph has a genomeThat is why I clutched at the straw that it is storage fornucleic acid precursors
Phil Trans R Soc Lond B (2003)
T Cavalier-Smith The reason I introduced the nucleo-morph is not to say it is direct evidence against yourhypothesis but to show that there probably was no singlefunctional reason for the retention of every gene in everyorganelle It may be that there are specific physiologicalreasons for the retention of certain genes and thereforeyou might be right about a subset of genes but I verymuch doubt if there are specific physiological reasons forthe retention of every single gene that is retained differen-tially because odd things happen in evolution Somethings are easy and sometimes things are difficult Evol-ution is history
J F Allen Of course I agree with that I do not thinkI can explain everything but I do think I can explainsomething If I have done that then I am content
GLOSSARY
AAC ADPndashATP carrierCES control by epistasy of synthesisCORR co-location for redox regulationLHC light-harvesting complexPCC Pasteur culture collectionORF open reading frameRUBISCO ribulose-15-bisphosphate carboxylasendashoxygenaseTAT twin-arginine translocase
Visit httpwwwpubsroyalsocacuk to see an electronic appendix tothis paper

Genomes in mitochondria and chloroplasts J F Allen 31
ment will act in the wrong direction to explain why chloro-plasts have retained genomes at all provided there is noresulting obstacle to a chloroplast protein being success-fully encoded as a precursor in the nucleus
(iii) Genes are copied and replicated not relocated in onestep
If complete relocation is envisaged as proceeding by thesimultaneous appearance of a gene in one compartmentand its deletion from another then it is possible to imaginethat whatever the vector is there may be selectivity ingene transfer (Henze amp Martin 2001) For example ifviral incorporation replication and recombination in bothorganelle and nucleus were the mechanism of relocationthen successful gene transfer might depend on the abilityof the virus to complete the process in one step This inturn might depend on some property of the gene or ofthe protein it encodes if expression occurs at any stage inthe process of relocation An assumption of the CORRhypothesis favoured here is that genes are not movedphysically but merely copied after which either the copyin the original location or the copy in the new locationmay be lost as a result of selection A useful analogy maybe the action of copying files between computer storagevolumes In this analogy the copy is identical to the orig-inal and either may subsequently be deleted without lossof information
Because any gene that is not usefully expressed willundergo no selection to maintain a functional allele theprevailing location will be the one in which the functionalcopy of the gene resides The problem of which gene sur-vives is then subsumed into the problem of which is theselectively advantageous location for it It seems reason-able to assume that there wasmdashand ismdashan interim periodin which homologous genes in different compartments areessentially in competition to see which functions best Asoutlined above there is no obvious obstacle for copyingDNA sequences within the cell The physico-chemicalproperties of DNA are fairly uniform and not directlydependent on nucleotide sequence though organelles likeintra-cellular parasites seem to prefer A and T to G andC The answer to the question lsquowhich gene location func-tions bestrsquo may have nothing to do with the gene itselfnor directly with the structure and function of its geneproduct Instead the answer may have everything to dowith the required regulatory interaction between the geneproduct and expression of the gene that produces it
(g) CORRThis hypothesis has been propounded earlier (sect 3) and
will be summarized as follows Vectorial electron and pro-ton transfer exerts regulatory control over expression ofgenes encoding proteins directly involved in or affectingredox poise This regulatory coupling is indispensable foradaptation to environmental changes in chemical activitiesof oxygen light and CO2 and requires co-location of thesegenes with their gene products Organelles lsquomake theirown decisionsrsquo on the basis of environmental changesaffecting redox state The hypothesis is abbreviatedCORR for co-location for redox regulation or more com-pletely lsquoco-location (of gene and gene product) for(evolutionary) continuity of redox regulation of geneexpressionrsquo
Phil Trans R Soc Lond B (2003)
5 SUMMARY OF THE CURRENT STATUS OF THEHYPOTHESIS OF CORR
Q Why do mitochondria and chloroplasts have theirown genetic systems
A CORR co-location of genes and gene products per-mits direct and autonomous redox regulation of geneexpression
(a) Objections to CORR(i) There is no regulation of gene expression in chloroplasts
and mitochondriaThis is still commonly stated but incorrect See the prin-
ciple in sect 3d
(ii) There is no regulation of gene expression in chloroplastsand mitochondria at the level of transcription
For chloroplasts there is now clear evidence for a func-tionally intelligible redox control of transcription(Pfannschmidt et al 1999a) This invalidates the previouswidely held assumption that all control of chloroplast geneexpression is post-transcriptional (Gruissem amp Tonkyn1993 GoldschmidtClermont 1998) The evidence formitochondria is less clear but redox control of transcrip-tion certainly cannot be ruled out and there are indi-cations that it may occur (Pearson et al 1993Konstantinov et al 1995 Wilson et al 1996b) It mustalso be restated that transcriptional control is not neces-sary for CORR but serves as a compelling example of aconserved typically bacterial mode of response that oper-ates still in eukaryotic cytoplasmic organelles thus illus-trating the principle of continuity of redox control (sect 3d)Post-transcriptional redox regulation (Danon amp Mayfield1994 Kim amp Mayfield 1997 Brown et al 2001 Treb-itsh amp Danon 2001) is consistent with CORR and thereis nothing to forbid it being a primary mechanism thatsatisfies most or indeed all of the formal requirements ofthe hypothesis in particular cases
(iii) There are no redox signalling components encoded inchloroplast or mitochondrial DNA
This is what CORR predicts See the principle in sect 3j
(iv) There is redox control of nuclear gene expressionThis is correct and many examples are now known
especially for chloroplasts (Escoubas et al 1995 Kovacs etal 2000 Pfannschmidt et al 2001ab Surpin et al 2002)Redox signals from photosynthetic or respiratory electrontransport chains are not at all prohibited by CORR Theessence of CORR is that there is a subset of organellarproteins for which there is an overriding requirement fora redox regulatory control that is initiated within theorganelle and whose effects are exerted by the simplestand most direct route This redox signalling is envisagedto take place without a requirement for prior consultationand coordination with what may be regarded as a nucleo-cytoplasmic bureaucracy
(v) Nuclear genes can be expressed as rapidly as chloroplastand mitochondrial genes
Zerges (2002) for example cites the equal times takenfor complete expression of nuclear and chloroplast genesin Chlamydomonas as evidence against the CORR hypoth-
32 J F Allen Genomes in mitochondria and chloroplasts
esis However the question is not one of the time takento express a gene but the time taken to alter the rate ofits expression in response to a (redox) signal originatingwithin the organelle An equally important additional fac-tor may be the autonomy of the signalling pathway thatis its independence from other inputs as described in theprevious paragraph One might say that mitochondria andchloroplasts require a free hand
(vi) There is no evidence to support this hypothesisThe hypothesis is rich in predictions (sectsect 3 and 6) and
those that have been tested stringently are in agreementwith experimental results None of the predictions of thehypothesis has been found to be incorrect and none of theunderlying principles (sect 3) has been disproved Furtherpredictions will be the subject of future experiments
(b) Evidence for CORR(i) Chloroplasts have selectively retained cyanobacterial genes
for primary events in photosynthesisSee table 1
(ii) There is regulation of gene expression in chloroplastsThere is transcriptional regulation of gene expression
in chloroplasts (sect 3d) The case for mitochondria is notyet proven
(iii) There is redox regulation of gene expression inchloroplasts
There is transcriptional redox regulation of geneexpression in chloroplasts (sect 3d) The case for mitochon-dria is not yet proven though see (Allen et al 1995 Kon-stantinov et al 1995 Galvis et al 1998)
(iv) There are no redox signalling components encoded inchloroplast or mitochondrial DNA
Correct but with possible exceptions discussed in sect 3j
(v) There are lsquobacterialrsquo redox signalling components ineukaryotes
See sect 3j
(vi) Further predictions DNA-array results and phenotypesof allotopic mutants
A systematic test of CORR might be to make an oxid-ized-minus-reduced gene expression profile of the lsquotran-scriptomersquo of a cyanobacterium to see if the set of geneswhose transcription is under most rapid and direct redoxcontrol corresponds to the genes of the chloroplast gen-ome Laboratories with this experimental capability do notseem to have addressed this possibility (Hihara et al 2002Ikeuchi 2002) Likewise if a putative modern representa-tive of the proto-mitochondrion can be found and agreedeg Rhodospirillum rubrum or Paracoccus denitrificans thena similar experiment should reveal a subset of genes corre-sponding to mitochondrial genomes DNA-microarraysalso present the possibility of testing the relative rate ofredox response and dependency on other signallinginputs of transcription of eukaryotic genes To dateorganellar genes have not been included in such investi-gations (Ferea et al 1999) A further experimental possi-bility is to examine the redox regulatory phenotypes ofexisting allotopic mutants as outlined in sect 3e
Phil Trans R Soc Lond B (2003)
6 CONCLUSION AND FUTURE PROSPECTS
The CORR hypothesis states that the function of mito-chondrial and chloroplast genomes is to provide co-location (of gene and gene product) for (evolutionary)continuity of redox regulation of gene expression Thishypothesis is neither self-contradictory nor devoid of test-able predictions It is also consistent with available evi-dence and in principle makes sense of the otherwisepuzzling distribution of genes between compartments ineukaryotic cells The explanation that CORR offersapplies equally to mitochondria and chloroplasts andidentifies selective forces that still operate to determine thelocation of genes No selective barriers need be assumedto gene transfer protein import nor membrane insertionNor do we need to invoke a lsquofrozen accidentrsquo that some-thing happened to stop gene transfer for some reason atsome time in the remote and inaccessible past CORR alsoexplains the absence of genomes from organelles that havelost their primary bioenergetic functions CORR may actas a stimulus to a search for residual bioenergetic functionin those organelles that have lost aerobic respiration andphotosynthesis but where residual genomes remain
CORR also predicts regulatory properties of knowncomponents of chloroplasts and mitochondria that in turnindicate important but as yet uncharacterized flexibility inenergy transduction These components include the RUB-ISCO large subunit and the CFo and Fo subunits of coup-ling ATPase in both chloroplasts and mitochondriaRedox regulation of the relative stoichiometry of compo-nents as seen in regulation of chloroplast photosystemstoichiometry (Pfannschmidt et al 1999a) may extend to(C)Fo-(C)F1-ATPase indicating that the function ofretention of genes for one or more of these componentsis flexibility in the stoichiometries of H1 to e2 and H1 toATP in chemiosmotic coupling (Schemidt et al 1998Stock et al 2000 Allen 2002)
So far explicit tests of CORR have largely been con-fined to chloroplasts and to some extent mitochondria ofhigher plants but this merely reflects the expertise andinterests of the investigators Further attempted disproofof CORR is welcomed and encouraged Novel experi-mental tests can be devised as well as attempted refu-tation through analysis of existing data It is hoped thatexplicit attention will be given to the CORR hypothesis inlaboratories already suitably equipped and that resourcescan be allocated for this purpose in laboratories wherethere is the interest but not currently the technology orexpertise
The heuristic value of the hypothesis already seen inthe role of redox-regulated transcription of photosyntheticreaction centre genes in chloroplasts may be its most use-ful property The central proposal is that mitochondriaand chloroplasts contain genes for proteins whose functionin electron transfer demands rapid direct and uncon-ditional redox regulatory control of their biosynthesis Thefull implications of this hypothesis are very broad and onlyjust beginning to be explored
I thank Carol Allen Aubrey de Grey William Martin John ARaven and Francis-Andre Wollman for discussion and corre-spondence during the preparation of this paper and WilliamMartin for suggestions including table 1 Research grants from
Genomes in mitochondria and chloroplasts J F Allen 33
The Swedish Natural Sciences Research Council (NFR) andthe Crafoord Foundation are gratefully acknowledged
REFERENCES
Adams K L Daley D O Qiu Y L Whelan J amp PalmerJ D 2000 Repeated recent and diverse transfers of a mito-chondrial gene to the nucleus in flowering plants Nature408 354ndash357
Alberts B Bray D Lewis J Raff M Roberts K amp Wat-son J D 1994 The molecular biology of the cell New Yorkand London Garland Publishing
Allen C A Hakansson G amp Allen J F 1995 Redox con-ditions specify the proteins synthesized by isolated chloro-plasts and mitochondria Redox Rep 1 119ndash123
Allen J F 1992 Protein phosphorylation in regulation ofphotosynthesis Biochim Biophys Acta 1098 275ndash335
Allen J F 1993a Control of gene expression by redox poten-tial and the requirement for chloroplast and mitochondrialgenomes J Theor Biol 165 609ndash631
Allen J F 1993b Redox control of gene expression and thefunction of chloroplast genomes a hypothesis PhotosynthRes 36 95ndash102
Allen J F 1993c Redox control of transcription sensorsresponse regulators activators and repressors FEBS Lett332 203ndash207
Allen J F 1995 Thylakoid proteinndashphosphorylation state-1ndashstate-2 transitions and photosystem stoichiometry adjust-ment redox control at multiple levels of gene expressionPhysiol Plant 93 196ndash205
Allen J F 1996 Separate sexes and the mitochondrial theoryof ageing J Theor Biol 180 135ndash140
Allen J F 2002 Photosynthesis of ATP-electrons protonpumps rotors and poise Cell 110 273ndash276
Allen J F amp Allen C A 1999 A mitochondrial model forpremature ageing of somatically cloned mammals IUBMBLife 48 369ndash372
Allen J F amp Nilsson A 1997 Redox signalling and the struc-tural basis of regulation of photosynthesis by protein phos-phorylation Physiol Plant 100 863ndash868
Allen J F amp Pfannschmidt T 2000 Balancing the two photo-systems photosynthetic electron transfer governs transcrip-tion of reaction centre genes in chloroplasts Phil TransR Soc Lond B 355 1351ndash1357 (DOI 101098rstb20000697)
Allen J F amp Race H L 2002 Will the real LHC II kinaseplease step forward Sciencersquos STKE httpwwwstkeorgcgicontentfullsigtrans2002155pe43
Allen J F amp Raven J A 1996 Free-radical-induced mutationvs redox regulation costs and benefits of genes in organellesJ Mol Evol 42 482ndash492
Archibald J M Cavalier-Smith T Maier U amp Douglas S2001 Molecular chaperones encoded by a reduced nucleusthe cryptomonad nucleomorph J Mol Evol 52 490ndash501
Ashby M K amp Mullineaux C W 1999a Cyanobacterialycf27 gene products regulate energy transfer from phycobili-somes to photosystems I and II FEMS Microbiol Lett 181253ndash260
Ashby M K amp Mullineaux C W 1999b The role of ApcDand ApcF in energy transfer from phycobilisomes to PSI andPSII in a cyanobacterium Photosynth Res 61 169ndash179
Ashby M K Houmard J amp Mullineaux C W 2002 Theycf27 genes from cyanobacteria and eukaryotic algae distri-bution and implications for chloroplast evolution FEMSMicrobiol Lett 214 25ndash30
Attardi G amp Schatz G 1988 Biogenesis of mitochondria ARev Cell Biol 4 289ndash333
Azem A Oppliger W Lustig A Jeno P Feifel B SchatzG amp Horst M 1997 The mitochondrial hsp70 chaperone
Phil Trans R Soc Lond B (2003)
system effect of adenine nucleotides peptide substrate andmGrpE on the oligomeric state of mhsp70 J Biol Chem272 20 901ndash20 906
Barbrook A C Symington H Nisbet R E R LarkumA amp Howe C J 2001 Organisation and expression of theplastid genome of the dinoflagellate Amphidinium opercula-tum Mol Genet Genomics 266 632ndash638
Bauer C E Elsen S amp Bird T H 1999 Mechanisms forredox control of gene expression A Rev Microbiol 53495ndash523
Bauer C E Elsen S Swem L R Swem D L amp MasudaS 2003 Redox and light regulation of gene expression inphotosynthetic prokaryotes Phil Trans R Soc Lond B 358147ndash154 (DOI 101098rstb20021189)
Berks B C Sargent F De Leeuw E Hinsley A P Stan-ley N R Jack R L Buchanan G amp Palmer T 2000aA novel protein transport system involved in the biogenesisof bacterial electron transfer chains Biochim Biophys ActaBioenerg 1459 325ndash330
Berks B C Sargent F amp Palmer T 2000b The tat proteinexport pathway Mol Microbiol 35 260ndash274
Blackstone N W 1999 Redox control in development andevolution evidence from colonial hydroids J Exp Biol 2023541ndash3553
Blackstone N W 2000 Redox control and the evolution ofmulticellularity Bioessays 22 947ndash953
Blackstone N W 2001 Redox state reactive oxygen speciesand adaptive growth in colonial hydroids J Exp Biol 2041845ndash1853
Bogorad L 1975 Evolution of organelles and eukaryotic gen-omes Science 188 891ndash898
Bogsch E G Sargent F Stanley N R Berks B C Robin-son C amp Palmer T 1998 An essential component of a novelbacterial protein export system with homologues in plastidsand mitochondria J Biol Chem 273 18 003ndash18 006
Brown E C Somanchi A amp Mayfield S P 2001 Interor-ganellar crosstalk new perspectives on signaling from thechloroplast to the nucleus Genome Biol 2 10211ndash10214
Cavalier-Smith T 2002 Chloroplast evolution secondarysymbiogenesis and multiple losses Curr Biol 12 R62ndashR64
Chitnis P R amp Thornber J P 1988 The major light-har-vesting complex of photosystem 2 Aspects of its molecularand cell biology Photosynth Res 16 41ndash63
Chow W S Melis A amp Anderson J M 1990 Adjustmentsof photosystem stoichiometry in chloroplasts improve thequantum efficiency of photosynthesis Proc Natl Acad SciUSA 87 7502ndash7506
Claros M G Perea J Shu Y Samatey F A PopotJ L amp Jacq C 1995 Limitations to in vivo import of hydro-phobic proteins into yeast mitochondria The case of a cyto-plasmically synthesized apocytochrome b Eur J Biochem228 762ndash771
Dalbey R E amp Robinson C 1999 Protein translocation intoand across the bacterial plasma membrane and the plant thy-lakoid membrane Trends Biochem Sci 24 17ndash22
Daley D O Adams K L Clifton R Qualmann S MillarA H Palmer J D Pratje E amp Whelan J 2002 Genetransfer from mitochondrion to nucleus novel mechanismsfor gene activation from Cox2 Plant J 30 11ndash21
Danon A amp Mayfield S P 1994 Light-regulated translationof chloroplast messenger RNAs through redox potentialScience 266 1717ndash1719
de Grey A 2000 Mitochondrial gene therapy an arena for thebiomedical use of inteins Trends Biotechnol 18 394ndash399
Deisenhofer J Epp O Miki K Huber R amp Michel H1985 Structure of the protein subunits in the photosyntheticreaction center of Rhodopseudomonas viridis at 3Ac resolutionNature 318 618ndash624
34 J F Allen Genomes in mitochondria and chloroplasts
Delwiche C F 1999 Tracing the thread of plastid diversitythrough the tapestry of life Am Nat 154 S164ndashS177
Doolittle W F 1999 Phylogenetic classification and the uni-versal tree Science 284 2124ndash2129
Douglas S E 1998 Plastid evolution origins diversitytrends Curr Opin Genet Dev 8 655ndash661
Duby G amp Boutry M 2002 Mitochondrial protein importmachinery and targeting information Plant Sci 162 477ndash490
Eberhard S Drapier D amp Wollman F A 2002 Searchinglimiting steps in the expression of chloroplast encoded pro-teins relations between gene copy number transcriptiontranscript abundance and translation rate in the chloroplastof Chlamydomonas reinhardtii Plant J 31 149ndash160
El Bissati K amp Kirilovsky D 2001 Regulation of psbA andpsaE expression by light quality in Synechocystis species PCC6803 A redox control mechanism Plant Physiol 1251988ndash2000
Ellis R J 1984 The nuclear domination of chloroplast devel-opment Science Progress 69 129ndash142
Embley T M amp Martin W 1998 Molecular evolution ahydrogen-producing mitochondrion Nature 396 517ndash519
Ems S C Morden C W Dixon C K Wolfe K HdePamphilis C W amp Palmer J D 1995 Transcriptionsplicing and editing of plastid RNAs in the non-photosyn-thetic plant Epifagus virginiana Plant Mol Biol 29 721ndash733
Eraso J M amp Kaplan S 2000 From redox flow to gene regu-lation role of the PrrC protein of Rhodobacter sphaeroides241 Biochemistry 39 2052ndash2062
Eraso J M amp Kaplan S 2002 Redox flow as an instrumentof gene regulation Meth Enzymol 348 216ndash229
Escoubas J M Lomas M Laroche J amp Falkowski P G1995 Light-intensity regulation of cab gene-transcription issignaled by the redox state of the plastoquinone pool ProcNatl Acad Sci USA 92 10 237ndash10 241
Ferea T L Botstein D Brown P O amp Rosenzweig R F1999 Systematic changes in gene expression patterns follow-ing adaptive evolution in yeast Proc Natl Acad Sci USA96 9721ndash9726
Forsberg J Rosenquist M Fraysse L amp Allen J F 2001Redox signalling in chloroplasts and mitochondria genomicand biochemical evidence for two-component regulatory sys-tems in bioenergetic organelles Biochem Soc Trans 29403ndash407
Fujita Y 1997 A study on the dynamic features of photosys-tem stoichiometry accomplishments and problems forfuture studies Photosynth Res 53 83ndash93
Gabriel K Buchanan S K amp Lithgow T 2001 The alphaand the beta protein translocation across mitochondrial andplastid outer membranes Trends Biochem Sci 26 36ndash40
Galvis M L E Allen J F amp Hakansson G 1998 Proteinsynthesis by isolated pea mitochondria is dependent on theactivity of respiratory complex II Curr Genet 33 320ndash329
Gatenby A A amp Ellis R J 1990 Chaperone function theassembly of ribulose bisphosphate carboxylase-oxygenase ARev Cell Biol 6 125ndash149
GoldschmidtClermont M 1998 Coordination of nuclear andchloroplast gene expression in plant cells Int Rev CytolSurv Cell Biol 177 115ndash180
Gray M W 1992 The endosymbiont hypothesis revisited IntRev Cytol 141 233ndash357
Gray M W 1999 Evolution of organellar genomes CurrOpin Genet Dev 9 678ndash687
Gray M W 2000 Evolutionary biology mitochondrial geneson the move Nature 408 302ndash305
Gray M W amp Doolittle W F 1982 Has the endosymbionthypothesis been proven Microbiol Rev 46 1ndash42
Gray M W Burger G amp Lang B F 1999 Mitochondrialevolution Science 283 1476ndash1481
Phil Trans R Soc Lond B (2003)
Grebe T W amp Stock J B 1999 The histidine protein kinasesuperfamily Adv Microb Physiol 41 139ndash227
Green B R amp Kuhlbrandt W 1995 Sequence conservationof light-harvesting and stress-response proteins in relation tothe 3-dimensional molecular structure of Lhcii PhotosynthRes 44 139ndash148
Gruissem W amp Tonkyn J C 1993 Control mechanisms ofplastid gene expression Crit Rev Plant Sci 12 19ndash55
Hackstein J H P (and 11 others) 2001 Hydrogenosomesconvergent adaptations of mitochondria to anaerobicenvironments Zool Anal Complex Syst 104 290ndash302
Harrison M A Keen J N Findlay J B C amp Allen J F1990 Modification of a Glnb-like gene-product by photosyn-thetic electron-transport in the cyanobacterium Synechoc-occus 6301 FEBS Lett 264 25ndash28
Harvey A J Kind K L amp Thompson J G 2002 Redoxregulation of early embryo development Reproduction 123479ndash486
Heathcote P Fyfe P K amp Jones M R 2002 Reactioncentres the structure and evolution of biological solarpower Trends Biochem Sci 27 79ndash87
Henze K amp Martin W 2001 How do mitochondrial genesget into the nucleus Trends Genet 17 383ndash387
Herrmann R G amp Westhoff P 2001 Thylakoid biogenesis anddynamics the result of a complex phylogenetic puzzle InRegulation of photosynthesis vol 11 (ed E-M Aro amp BAndersson) pp 1ndash28 Dordrecht The Netherlands Kluwer
Hihara Y Sonoike K Kanehisa M amp Ikeuchi M 2002Search for redox responsive genes in Synechocystis sp PCC6803 by using DNA microarrays Plant Cell Physiol 43S178
Howe C J Barbrook A C amp Lockhart P J 2000 Organellegenes do they jump or are they pushed Trends Genet 1665ndash66
Ikeuchi M 2002 Gene expression and responses to light stressin cyanobacteria Plant Cell Physiol 43 S21
Iuchi S amp Lin E C C 1992 Mutational analysis of signaltransduction by Arcb a membrane sensor protein respon-sible for anaerobic repression of operons involved in the cen-tral aerobic pathways in Escherichia coli J Bacteriol 1743972ndash3980
Jackson-Constan D Akita M amp Keegstra K 2001 Molecu-lar chaperones involved in chloroplast protein importBiochim Biophys Acta Mol Cell Res 1541 102ndash113
Jarvis P amp Soll M 2001 Toc Tic and chloroplast proteinimport Biochim Biophys Acta Mol Cell Res 1541 64ndash79
Jordan P Fromme P Witt H T Klukas O SaengerW amp Krauss N 2001 Three-dimensional structure ofcyanobacterial photosystem I at 25 acngstrom resolutionNature 411 909ndash917
Kaneko T (and 23 others) 1996a Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regionsDNA Res 3 109ndash136
Kaneko T (and 23 others) 1996b Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regions[Supplement] DNA Res 3 185ndash209
Kanevski I amp Maliga P 1994 Relocation of the plastid rbcLgene to the nucleus yields functional ribulose-15-bisphosph-ate carboxylase in tobacco chloroplasts Proc Natl Acad SciUSA 91 1969ndash1973
Kim J amp Mayfield S P 1997 Protein disulfide isomerase as aregulator of chloroplast translational activation Science 2781954ndash1957
Genomes in mitochondria and chloroplasts J F Allen 35
Kirk J T O amp Tilney-Bassett R A E 1978 The plastidsTheir chemistry structure growth and inheritance Amster-dam Elsevier
Koehler C M Merchant S amp Schatz G 1999 How mem-brane proteins travel across the mitochondrial intermem-brane space Trends Biochem Sci 24 428ndash432
Komiya T Rospert S Schatz G amp Mihara K 1997 Bind-ing of mitochondrial precursor proteins to the cytoplasmicdomains of the import receptors Tom70 and Tom20 isdetermined by cytoplasmic chaperones FASEB J 11 1196
Konstantinov Y M Lutsenko G N amp Podsosonny V A1995 Genetic functions of isolated maize mitochondriaunder model changes of redox conditions Biochem MolBiol Int 36 319ndash326
Korab-Laskowska M Rioux P Brossard N LittlejohnT G Gray M W Lang B F amp Burger G 1998 Theorganelle genome database project (GOBASE) Nucleic AcidsRes 26 138ndash144
Kovacs L Wiessner W Kis M Nagy F Mende D ampDemeter S 2000 Short- and long-term redox regulation ofphotosynthetic light energy distribution and photosystemstoichiometry by acetate metabolism in the green alga Chla-mydobotrys stellata Photosynth Res 65 231ndash247
Kuhlbrandt W 1994 Structure and function of the plant light-harvesting complex LHC-II Curr Opin Struct Biol 4519ndash528
Lang B F Gray M W amp Burger G 1999 Mitochondrialgenome evolution and the origin of eukaryotes A RevGenet 33 351ndash397
Lemire B D Fankhauser C Baker A amp Schatz G 1989The mitochondrial targeting function of randomly generatedpeptide sequences correlates with predicted helical amphi-philicity J Biol Chem 264 20 206ndash20 215
Leuenberger D Bally N A Schatz G amp Koehler C M1999 Different import pathways through the mitochondrialintermembrane space for inner membrane proteins EMBOJ 18 4816ndash4822
Li H amp Sherman L A 2000 A redox-responsive regulator ofphotosynthesis gene expression in the cyanobacterium Syne-chocystis sp strain PCC 6803 J Bacteriol 182 4268ndash4277
Lin X Y (and 36 others) 1999 Sequence and analysis of chro-mosome 2 of the plant Arabidopsis thaliana Nature 402761ndash768
Lonsdale D M Hodge T P Howe C J amp Stern D B1983 Maize mitochondrial-DNA contains a sequence hom-ologous to the ribulose-15-bisphosphate carboxylase largesubunit gene of chloroplast DNA Cell 34 1007ndash1014
McFadden G I 1999 Endosymbiosis and evolution of theplant cell Curr Opin Plant Biol 2 513ndash519
McFadden G I 2001 Chloroplast origin and integrationPlant Physiol 125 50ndash53
Mahler H R amp Raff R A 1975 The evolutionary origin ofthe mitochondrion a nonsymbiotic model Int Rev Cytol43 1ndash124
Manfredi G Fu J Ojaimi J Sadlock J E Kwong J QGuy J amp Schon E A 2002 Rescue of a deficiency in ATPsynthesis by transfer of MTATP6 a mitochondrial DNA-encoded gene to the nucleus Nature Genet 30 394ndash399
Marc P Margeot A Devaux F Blugeon C Corral-Debrinski M amp Jacq C 2002 Genome-wide analysis ofmRNAs targeted to yeast mitochondria EMBO Rep 3159ndash164
Marienfeld J Unseld M amp Brennicke A 1999 The mito-chondrial genome of Arabidopsis is composed of both nativeand immigrant information Trends Plant Sci 4 495ndash502
Martin W 1999 A briefly argued case that mitochondria andplastids are descendants of endosymbionts but that thenuclear compartment is not Proc R Soc Lond B 2661387ndash1395 (DOI 101098rspb19990792)
Phil Trans R Soc Lond B (2003)
Martin W amp Herrmann R G 1998 Gene transfer fromorganelles to the nucleus how much what happens andwhy Plant Physiol 118 9ndash17
Martin W amp Muller M 1998 The hydrogen hypothesis forthe first eukaryote Nature 392 37ndash41
Martin W amp Russell M J 2003 On the origins of cells ahypothesis for the evolutionary transitions from abiotic geo-chemistry to chemoautotrophic prokaryotes and from pro-karyotes to nucleated cells Phil Trans R Soc Lond B 35859ndash85 (DOI 101098rstb20021183)
Martin W Stoebe B Goremykin V Hansmann S Hase-gawa M amp Kowallik K V 1998 Gene transfer to thenucleus and the evolution of chloroplasts Nature 393162ndash165
Martin W Hoffmeister M Rotte C amp Henze K 2001 Anoverview of endosymbiotic models for the origins of eukary-otes their ATP-producing organelles (mitochondria andhydrogenosomes) and their heterotrophic lifestyle BiolChem 382 1521ndash1539
Martin W Rujan T Richly E Hansen A Cornelsen SLins T Leister D Stoebe B Hasegawa M amp PennyD 2002 Evolutionary analysis of Arabidopsis cyanobacterialand chloroplast genomes reveals plastid phylogeny and thou-sands of cyanobacterial genes in the nucleus Proc NatlAcad Sci USA 99 12 246ndash12 251
Millen R S (and 12 others) 2001 Many parallel losses of infAfrom chloroplast DNA during angiosperm evolution withmultiple independent transfers to the nucleus Plant Cell 13645ndash658
Morse D Salois P Markovic P amp Hastings J W 1995 Anuclear-encoded form-II RUBISCO in dinoflagellatesScience 268 1622ndash1624
Murakami A Kim S J amp Fujita Y 1997 Changes in photo-system stoichiometry in response to environmental con-ditions for cell growth observed with the cyanophyteSynechocystis PCC 6714 Plant Cell Physiol 38392ndash397
Oh J I amp Kaplan S 2000 Redox signaling globalization ofgene expression EMBO J 19 4237ndash4247
Ozawa T 1995 Mitochondrial-DNA mutations associatedwith aging and degenerative diseases Exp Gerontol 30269ndash290
Palmer J D 1997 Organelle genomes going going goneScience 275 790ndash791
Pearson C K Wilson S B Schaffer R amp Ross A W 1993NAD turnover and utilization of metabolites for RNA syn-thesis in a reaction sensing the redox state of the cyto-chrome-b( 6 )f complex in isolated chloroplasts Eur JBiochem 218 397ndash404
Pfanner N amp Geissler A 2001 Versatility of the mitochon-drial protein import machinery Natl Rev Mol Cell Biol 2339ndash349
Pfanner N amp Wiedemann N 2002 Mitochondrial proteinimport two membranes three translocases Curr Opin CellBiol 14 400ndash411
Pfannschmidt T Nilsson A amp Allen J F 1999a Photosyn-thetic control of chloroplast gene expression Nature 397625ndash628
Pfannschmidt T Nilsson A Tullberg A Link G amp AllenJ F 1999b Direct transcriptional control of the chloroplastgenes psbA and psaAB adjusts photosynthesis to light energydistribution in plants IUBMB Life 48 271ndash276
Pfannschmidt T Allen J F amp Oelmuller R 2001a Prin-ciples of redox control in photosynthesis gene expressionPhysiol Plant 112 1ndash9
Pfannschmidt T Schutze K Brost M amp Oelmuller R2001b A novel mechanism of nuclear photosynthesis generegulation by redox signals from the chloroplast during pho-
36 J F Allen Genomes in mitochondria and chloroplasts
tosystem stoichiometry adjustment J Biol Chem 27636 125ndash36 130
Popot J L amp de Vitry C 1990 On the microassembly of inte-gral membrane proteins A Rev Biophys Biophys Chem 19369ndash403
Race H L Herrmann R G amp Martin W 1999 Why haveorganelles retained genomes Trends Genet 15 364ndash370
Raven J A Johnston A M Parsons R amp Kubler J 1994aThe influence of natural and experimental high O2 concen-trations on O2 evolving phototrophs Biol Rev CambridgePhil Soc 69 61ndash94
Raven J A Johnston A M Parsons R amp Kubler J 1994bThe occurrence and influence on photolithotrophs of highoxygen concentrations Proc R Soc Edinb B 102 193ndash201
Robinson C 2000 The twin-arginine translocation system anovel means of transporting folded proteins in chloroplastsand bacteria Biol Chem 381 89ndash93
Robinson C amp Bolhuis A 2001 Protein targeting by the twin-arginine translocation pathway Natl Rev Mol Cell Biol 2350ndash356
Rodermel S 2001 Pathways of plastid-to-nucleus signalingTrends Plant Sci 6 471ndash478
Roger A J 1999 Reconstructing early events in eukaryoticevolution Am Nat 154 S146ndashS163
Roger A J Svard S G Tovar J Clark C G SmithM W Gillin F D amp Sogin M L 1998 A mitochondrial-like chaperonin 60 gene in Giardia lamblia evidence thatdiplomonads once harbored an endosymbiont related to theprogenitor of mitochondria Proc Natl Acad Sci USA 95229ndash234
Roodyn D B amp Wilkie D 1968 The biogenesis of mitochondriaMethuenrsquos monographs on biological subjects London Methuen
Rujan T amp Martin W 2001 How many genes in Arabidopsiscome from cyanobacteria An estimate from 386 proteinphylogenies Trends Genet 17 113ndash120
Saccone C Gissi C Lanave C Larizza A Pesole G ampReyes A 2000 Evolution of the mitochondrial genetic sys-tem an overview Gene 261 153ndash159
Salzberg S L White O Peterson J amp Eisen J A 2001Microbial genes in the human genome lateral transfer orgene loss Science 292 1903ndash1906
Sargent F Berks B C amp Palmer T 2002 Assembly of mem-brane-bound respiratory complexes by the Tat protein-trans-port system Arch Microbiol 178 77ndash84
Schatz G 1998 Protein transport the doors to organellesNature 395 439ndash440
Schemidt R A Qu J Williams J R amp Brusilow W S1998 Effects of carbon source on expression of F0 genes andon the stoichiometry of the c subunit in the F1 F0 ATPaseof Escherichia coli J Bacteriol 180 3205ndash3208
Selosse M A Albert B R amp Godelle B 2001 Reducing thegenome size of organelles favours gene transfer to thenucleus Trends Ecol Evol 16 135ndash141
Stern D B amp Lonsdale D M 1982 Mitochondrial and chlo-roplast genomes of maize have a 12- kilobase DNA-sequencein common Nature 299 698ndash702
Stock D Gibbons C Arechaga I Leslie A G W ampWalker J E 2000 The rotary mechanism of ATP synthaseCurr Opin Struct Biol 10 672ndash679
Stoebe B amp Maier U G 2002 One two three naturersquos toolbox for building plastids Protoplasma 219 123ndash130
Stoebe B Martin W amp Kowallik K V 1998 Distributionand nomenclature of protein-coding genes in 12 sequencedchloroplast genomes Plant Mol Biol Rep 16 243ndash255
Stupar R M Lilly J W Town C D Cheng Z Kaul SBuell C R amp Jiang J M 2001 Complex mtDNA consti-tutes an approximate 620 kb insertion on Arabidopsis thali-ana chromosome 2 implication of potential sequencing
Phil Trans R Soc Lond B (2003)
errors caused by large-unit repeats Proc Natl Acad SciUSA 98 5099ndash5103
Surpin M Larkin R M amp Chory J 2002 Signal transduc-tion between the chloroplast and the nucleus Plant Cell 14S327ndashS338
Tjaden J Schwoppe C Mohlmann T Quick P W ampNeuhaus H E 1998 Expression of a plastidic ATPADPtransporter gene in Escherichia coli leads to a functional aden-ine nucleotide transport system in the bacterial cytoplasmicmembrane J Biol Chem 273 9630ndash9636
Tovar J Fischer A amp Clark C G 1999 The mitosome a novelorganelle related to mitochondria in the amitochondrial para-site Entamoeba histolytica Mol Microbiol 32 1013ndash1021
Trebitsh T amp Danon A 2001 Translation of chloroplastpsbA mRNA is regulated by signals initiated by both photos-ystems II and I Proc Natl Acad Sci USA 98 12 289ndash12 294
Tullberg A Alexciev K Pfannschmidt T amp Allen J F2000 Photosynthetic electron flow regulates transcription ofthe psaB gene in pea (Pisum sativum L) chloroplasts throughthe redox state of the plastoquinone pool Plant Cell Physiol41 1045ndash1054
Turmel M Lemieux C Burger G Lang B F Otis CPlante I amp Gray M W 1999 The complete mitochondrialDNA sequences of Nephroselmis olivacea and Pedinomonasminor two radically different evolutionary patterns withingreen algae Plant Cell 11 1717ndash1729
Unden G amp Bongaerts J 1997 Alternative respiratory path-ways of Escherichia coli energetics and transcriptional regu-lation in response to electron acceptors Biochim BiophysActa Bioenerg 1320 217ndash234
Unseld M Marienfeld J R Brandt P amp Brennicke A1997 The mitochondrial genome of Arabidopsis thaliana con-tains 57 genes in 366 924 nucleotides Nature Genet 1557ndash61
Van der Giezen M Slotboom D J Horner D S DyalP L Harding M Xue G P Embley T M amp KunjiE R S 2002 Conserved properties of hydrogenosomal andmitochondrial ADPATP carriers a common origin for bothorganelles EMBO J 21 572ndash579
Von Heijne G 1986 Why mitochondria need a genome FEBSLett 198 1ndash4
Westphal S Soll J amp Vothknecht U C 2001 A vesicle trans-port system inside chloroplasts FEBS Lett 506 257ndash261
Whitehouse H L K 1969 Towards an understanding of themechanism of heredity London Edward Arnold Ltd
Williams B A P Hirt R P Lucocq J M amp Embley T M2002 A mitochondrial remnant in the microsporidian Trachi-pleistophora hominis Nature 418 865ndash869
Wilson R J (and 10 others) 1996a Complete gene map of theplastid-like DNA of the malaria parasite Plasmodium falcipa-rum J Mol Biol 261 155ndash172
Wilson S B Davidson G S Thomson L M amp PearsonC K 1996b Redox control of RNA synthesis in potato mito-chondria Eur J Biochem 242 81ndash85
Winkler H H amp Neuhaus H E 1999 Non-mitochondrialATP transport Trends Biochem Sci 24 64ndash68
Wolfe K H Morden C W amp Palmer J D 1992 Small sin-gle-copy region of plastid DNA in the non-photosyntheticangiosperm Epifagus virginiana contains only two genes Dif-ferences among dicots monocots and bryophytes in geneorganization at a non-bioenergetic locus J Mol Biol 22395ndash104
Wollman F A Minai L amp Nechushtai R 1999 The biogen-esis and assembly of photosynthetic proteins in thylakoidmembranes Biochim Biophys Acta Bioenerg 1411 21ndash85
Yeliseev A A amp Kaplan S 2000 TspO of Rhodobactersphaeroides a structural and functional model for the mam-
Genomes in mitochondria and chloroplasts J F Allen 37
malian peripheral benzodiazepine receptor J Biol Chem275 5657ndash5667
Zerges W 2002 Does complexity constrain organelle evol-ution Trends Plant Sci 7 175ndash182
Zhang Z D Green B R amp Cavalier-Smith T 1999 Singlegene circles in dinoflagellate chloroplast genomes Nature400 155ndash159
Zhang Z D Cavalier-Smith T amp Green B R 2001 A fam-ily of selfish minicircular chromosomes with jumbled chloro-plast gene fragments from a dinoflagellate Mol Biol Evol18 1558ndash1565
Zouni A Witt H T Kern J Fromme P Krauss NSaenger W amp Orth P 2001 Crystal structure of photosys-tem II from Synechococcus elongatus at 38 acngstrom resol-ution Nature 409 739ndash743
DiscussionD S Horner (Dipartimento di Fisiologia e Biochimica
Generali University of Milan Milan Italy) Given thatthere are components of these membrane-bound energy-transduction pathways that are consistently nuclear enco-ded not the key ones you pointed out but others do youexpect perhaps a slower perhaps a more crude but yet aredox communication between the organelle andexpression of nuclear-encoded genes
J F Allen There is and there is no question about itI think that Bogoradrsquos lock-in hypothesis has somethingto say here and there are other people who say it is all todo with assembling multi-subunit complexes If you havecontrol over the initial construction of the core aroundwhich the whole complex is eventually built then redoxcontrol or some sort of control over synthesis of that corewill control construction of the whole thing eventuallyand I think that may well be largely correct I think it ispossible in principle for nuclear genes to make precursorsaccording to redox instructions but it is not really soimportant that those precursors are here and now presentin exactly the right quantities because the precursors canto a limited extent hang around and wait Actually thereis proteolytic degradation of unattached precursors and soon but I think the fact that these chloroplast and mito-chondrially DNA-encoded subunits are at the core ofthese energy-transducing complexes is the crucial issue
T Cavalier-Smith (Department of Zoology University ofOxford Oxford UK ) In the case of mitochondria the sub-set of genes that remains is much more variable than youimplied It runs from three proteins in the minimal casesup to I think over 90 That is a big difference and manyof the genes in different lineages of protists that remainhave nothing to do with redox control they are things likeribosomal proteins and so on So I think that even thoughwhat you say about redox control of certain genes mayactually be an important thing it is not the fundamentaloverall reason why the genomes are maintained It may bepart of the explanation for why certain genes are alwaysincluded in that subset but even in mitochondria I thinkthat cytochrome b is the only one that is always presentSo I think that both this great diversity of what has beenretained in different lineages and the nucleomorph caseindicate that evolution was able to do one thing in onelineage 500 million years ago and never got around todoing it for 500 million years in another lineage so thereis a lot more haphazardness and evolutionary accidentsand contingency than a straight functional interpretation
Phil Trans R Soc Lond B (2003)
there is a lot of just sheer lsquohappenedndashorndashdidnrsquotndashhappenrsquoor carryover
J F Allen I was not implying anything about the rangeof mitochondrial genomes I have a limited amount oftime so I just talked about what I could talk about withinthat limit Of course there will be a range and this willdepend upon the physiology and bioenergetics of theparticular organism But what I am trying to do is to seewhat the common properties of the irreducible core areof components that are always encoded in the organellesand cytochrome b is a perfect example because that reallyis the thing that binds these two all-important quinoneswithout which there is no Q cycle without which there isno proton pumping and bioenergetics and in Plasmodiumwhere the mitochondrion is reduced to this bare mini-mum there is still a mitochondrial gene for cytochromeb Now my argument about the ribosomal proteins andall the rest of it is that in order to be able to exert thisall-important redox control over gene expression of certaincore components you need an in situ genetic system andthis can vary in composition for reasons that I cannotdescribe Maybe you need RNA polymerase or maybeyou can import RNA polymerase In plastids you haveboth of these options You are going to need ribosomestransfer RNAs and the rest of the apparatus of geneexpression and that needs to be in situ but we are stillleft with the question why does there have to be any gen-etic system at all The mature tobacco or spinach chloro-plast seems to have an entire genetic system just to makeone protein D1 which is all it ever needs to resynthesizeI think that this is a clue It is worth having this very unec-onomical crazy system because there is an overridingrequirement to connect electron transfer with geneexpression directly and unambiguouslymdashwithout recourseto a bureaucracy if you like which is what I think thenucleus is
K Willison (Institute of Cancer Research Chester BeattyLaboratories London UK ) These photosynthetic reactioncomplexes are a sort of baroque mixture of polypeptidechains and cofactors and my understanding is that someof them in high photon flux can turn over within say 30minutes so I am wondering if there is a correlationbetween gene retention and the turnover rates of some ofthe components and the complexes it seems to me thatthat could really be a limiting factor and the reason thatthe proteins had to be encoded locally in order to get thecomplexes to be assembled rapidly
J F Allen I think that is right You can only alter thesteady-state concentration of a protein complex by affect-ing the synthesis and breakdown and therefore turnoverThere is an extreme example the D1 protein encoded bythe psbA gene of chloroplasts which is always in the chlor-oplast genome without exception High light will tend todegrade this protein and when the high light goes awaythere is an absolute need very quickly to bring it back upagain I think that is a very good example Whether thisargument applies to genes such as cytochrome oxidase Ido not know
C J Leaver (Department of Plant Sciences University ofOxford Oxford UK ) Your experiments tend to relatemainly to the gene expression side What about proteinRNA turnover the whole concept of redox control ofthatmdashdo we know anything
38 J F Allen Genomes in mitochondria and chloroplasts
J F Allen Well quite a lot is known about chloro-plasts There is redox control of messenger RNA stabilityfor this same D1 protein which has been described byStephen Mayfield and his co-workers The evidence is thatthe site of redox control is ferredoxin via thioredoxin andthe implication is that thioredoxin tells whether non-cyclicelectron transport is working at all When it is workingthere is a signal that says lsquoOK now continue with all theprotein synthesisrsquo It is not particularly specific but moreof a global effect I think this is what Mayfield argues butof course this is not inconsistent with my argument I haveconcentrated on quinone pools and medium-potentialredox regulatory control because that is where I can seesignals about the dangerous consequences of getting itwrong but there is control of other levels of geneexpression In mitochondria I am not so sure I wouldbounce the question back to you on that one
W Martin (Institute of Botany III Heinrich-Heine Univ-ersitat Dusseldorf Dusseldorf Germany) The nucleomorphgenome was cited as evidence in conflict with your modelbut is that true The nucleomorph genome is not in abioenergetic organelle in that sense It resides in the cyto-sol so does the size of the nucleomorph genome workagainst your model Is it really evidence against yourmodel
J F Allen I do not know why should it be evidenceI cannot explain why the nucleomorph has a genomeThat is why I clutched at the straw that it is storage fornucleic acid precursors
Phil Trans R Soc Lond B (2003)
T Cavalier-Smith The reason I introduced the nucleo-morph is not to say it is direct evidence against yourhypothesis but to show that there probably was no singlefunctional reason for the retention of every gene in everyorganelle It may be that there are specific physiologicalreasons for the retention of certain genes and thereforeyou might be right about a subset of genes but I verymuch doubt if there are specific physiological reasons forthe retention of every single gene that is retained differen-tially because odd things happen in evolution Somethings are easy and sometimes things are difficult Evol-ution is history
J F Allen Of course I agree with that I do not thinkI can explain everything but I do think I can explainsomething If I have done that then I am content
GLOSSARY
AAC ADPndashATP carrierCES control by epistasy of synthesisCORR co-location for redox regulationLHC light-harvesting complexPCC Pasteur culture collectionORF open reading frameRUBISCO ribulose-15-bisphosphate carboxylasendashoxygenaseTAT twin-arginine translocase
Visit httpwwwpubsroyalsocacuk to see an electronic appendix tothis paper

32 J F Allen Genomes in mitochondria and chloroplasts
esis However the question is not one of the time takento express a gene but the time taken to alter the rate ofits expression in response to a (redox) signal originatingwithin the organelle An equally important additional fac-tor may be the autonomy of the signalling pathway thatis its independence from other inputs as described in theprevious paragraph One might say that mitochondria andchloroplasts require a free hand
(vi) There is no evidence to support this hypothesisThe hypothesis is rich in predictions (sectsect 3 and 6) and
those that have been tested stringently are in agreementwith experimental results None of the predictions of thehypothesis has been found to be incorrect and none of theunderlying principles (sect 3) has been disproved Furtherpredictions will be the subject of future experiments
(b) Evidence for CORR(i) Chloroplasts have selectively retained cyanobacterial genes
for primary events in photosynthesisSee table 1
(ii) There is regulation of gene expression in chloroplastsThere is transcriptional regulation of gene expression
in chloroplasts (sect 3d) The case for mitochondria is notyet proven
(iii) There is redox regulation of gene expression inchloroplasts
There is transcriptional redox regulation of geneexpression in chloroplasts (sect 3d) The case for mitochon-dria is not yet proven though see (Allen et al 1995 Kon-stantinov et al 1995 Galvis et al 1998)
(iv) There are no redox signalling components encoded inchloroplast or mitochondrial DNA
Correct but with possible exceptions discussed in sect 3j
(v) There are lsquobacterialrsquo redox signalling components ineukaryotes
See sect 3j
(vi) Further predictions DNA-array results and phenotypesof allotopic mutants
A systematic test of CORR might be to make an oxid-ized-minus-reduced gene expression profile of the lsquotran-scriptomersquo of a cyanobacterium to see if the set of geneswhose transcription is under most rapid and direct redoxcontrol corresponds to the genes of the chloroplast gen-ome Laboratories with this experimental capability do notseem to have addressed this possibility (Hihara et al 2002Ikeuchi 2002) Likewise if a putative modern representa-tive of the proto-mitochondrion can be found and agreedeg Rhodospirillum rubrum or Paracoccus denitrificans thena similar experiment should reveal a subset of genes corre-sponding to mitochondrial genomes DNA-microarraysalso present the possibility of testing the relative rate ofredox response and dependency on other signallinginputs of transcription of eukaryotic genes To dateorganellar genes have not been included in such investi-gations (Ferea et al 1999) A further experimental possi-bility is to examine the redox regulatory phenotypes ofexisting allotopic mutants as outlined in sect 3e
Phil Trans R Soc Lond B (2003)
6 CONCLUSION AND FUTURE PROSPECTS
The CORR hypothesis states that the function of mito-chondrial and chloroplast genomes is to provide co-location (of gene and gene product) for (evolutionary)continuity of redox regulation of gene expression Thishypothesis is neither self-contradictory nor devoid of test-able predictions It is also consistent with available evi-dence and in principle makes sense of the otherwisepuzzling distribution of genes between compartments ineukaryotic cells The explanation that CORR offersapplies equally to mitochondria and chloroplasts andidentifies selective forces that still operate to determine thelocation of genes No selective barriers need be assumedto gene transfer protein import nor membrane insertionNor do we need to invoke a lsquofrozen accidentrsquo that some-thing happened to stop gene transfer for some reason atsome time in the remote and inaccessible past CORR alsoexplains the absence of genomes from organelles that havelost their primary bioenergetic functions CORR may actas a stimulus to a search for residual bioenergetic functionin those organelles that have lost aerobic respiration andphotosynthesis but where residual genomes remain
CORR also predicts regulatory properties of knowncomponents of chloroplasts and mitochondria that in turnindicate important but as yet uncharacterized flexibility inenergy transduction These components include the RUB-ISCO large subunit and the CFo and Fo subunits of coup-ling ATPase in both chloroplasts and mitochondriaRedox regulation of the relative stoichiometry of compo-nents as seen in regulation of chloroplast photosystemstoichiometry (Pfannschmidt et al 1999a) may extend to(C)Fo-(C)F1-ATPase indicating that the function ofretention of genes for one or more of these componentsis flexibility in the stoichiometries of H1 to e2 and H1 toATP in chemiosmotic coupling (Schemidt et al 1998Stock et al 2000 Allen 2002)
So far explicit tests of CORR have largely been con-fined to chloroplasts and to some extent mitochondria ofhigher plants but this merely reflects the expertise andinterests of the investigators Further attempted disproofof CORR is welcomed and encouraged Novel experi-mental tests can be devised as well as attempted refu-tation through analysis of existing data It is hoped thatexplicit attention will be given to the CORR hypothesis inlaboratories already suitably equipped and that resourcescan be allocated for this purpose in laboratories wherethere is the interest but not currently the technology orexpertise
The heuristic value of the hypothesis already seen inthe role of redox-regulated transcription of photosyntheticreaction centre genes in chloroplasts may be its most use-ful property The central proposal is that mitochondriaand chloroplasts contain genes for proteins whose functionin electron transfer demands rapid direct and uncon-ditional redox regulatory control of their biosynthesis Thefull implications of this hypothesis are very broad and onlyjust beginning to be explored
I thank Carol Allen Aubrey de Grey William Martin John ARaven and Francis-Andre Wollman for discussion and corre-spondence during the preparation of this paper and WilliamMartin for suggestions including table 1 Research grants from
Genomes in mitochondria and chloroplasts J F Allen 33
The Swedish Natural Sciences Research Council (NFR) andthe Crafoord Foundation are gratefully acknowledged
REFERENCES
Adams K L Daley D O Qiu Y L Whelan J amp PalmerJ D 2000 Repeated recent and diverse transfers of a mito-chondrial gene to the nucleus in flowering plants Nature408 354ndash357
Alberts B Bray D Lewis J Raff M Roberts K amp Wat-son J D 1994 The molecular biology of the cell New Yorkand London Garland Publishing
Allen C A Hakansson G amp Allen J F 1995 Redox con-ditions specify the proteins synthesized by isolated chloro-plasts and mitochondria Redox Rep 1 119ndash123
Allen J F 1992 Protein phosphorylation in regulation ofphotosynthesis Biochim Biophys Acta 1098 275ndash335
Allen J F 1993a Control of gene expression by redox poten-tial and the requirement for chloroplast and mitochondrialgenomes J Theor Biol 165 609ndash631
Allen J F 1993b Redox control of gene expression and thefunction of chloroplast genomes a hypothesis PhotosynthRes 36 95ndash102
Allen J F 1993c Redox control of transcription sensorsresponse regulators activators and repressors FEBS Lett332 203ndash207
Allen J F 1995 Thylakoid proteinndashphosphorylation state-1ndashstate-2 transitions and photosystem stoichiometry adjust-ment redox control at multiple levels of gene expressionPhysiol Plant 93 196ndash205
Allen J F 1996 Separate sexes and the mitochondrial theoryof ageing J Theor Biol 180 135ndash140
Allen J F 2002 Photosynthesis of ATP-electrons protonpumps rotors and poise Cell 110 273ndash276
Allen J F amp Allen C A 1999 A mitochondrial model forpremature ageing of somatically cloned mammals IUBMBLife 48 369ndash372
Allen J F amp Nilsson A 1997 Redox signalling and the struc-tural basis of regulation of photosynthesis by protein phos-phorylation Physiol Plant 100 863ndash868
Allen J F amp Pfannschmidt T 2000 Balancing the two photo-systems photosynthetic electron transfer governs transcrip-tion of reaction centre genes in chloroplasts Phil TransR Soc Lond B 355 1351ndash1357 (DOI 101098rstb20000697)
Allen J F amp Race H L 2002 Will the real LHC II kinaseplease step forward Sciencersquos STKE httpwwwstkeorgcgicontentfullsigtrans2002155pe43
Allen J F amp Raven J A 1996 Free-radical-induced mutationvs redox regulation costs and benefits of genes in organellesJ Mol Evol 42 482ndash492
Archibald J M Cavalier-Smith T Maier U amp Douglas S2001 Molecular chaperones encoded by a reduced nucleusthe cryptomonad nucleomorph J Mol Evol 52 490ndash501
Ashby M K amp Mullineaux C W 1999a Cyanobacterialycf27 gene products regulate energy transfer from phycobili-somes to photosystems I and II FEMS Microbiol Lett 181253ndash260
Ashby M K amp Mullineaux C W 1999b The role of ApcDand ApcF in energy transfer from phycobilisomes to PSI andPSII in a cyanobacterium Photosynth Res 61 169ndash179
Ashby M K Houmard J amp Mullineaux C W 2002 Theycf27 genes from cyanobacteria and eukaryotic algae distri-bution and implications for chloroplast evolution FEMSMicrobiol Lett 214 25ndash30
Attardi G amp Schatz G 1988 Biogenesis of mitochondria ARev Cell Biol 4 289ndash333
Azem A Oppliger W Lustig A Jeno P Feifel B SchatzG amp Horst M 1997 The mitochondrial hsp70 chaperone
Phil Trans R Soc Lond B (2003)
system effect of adenine nucleotides peptide substrate andmGrpE on the oligomeric state of mhsp70 J Biol Chem272 20 901ndash20 906
Barbrook A C Symington H Nisbet R E R LarkumA amp Howe C J 2001 Organisation and expression of theplastid genome of the dinoflagellate Amphidinium opercula-tum Mol Genet Genomics 266 632ndash638
Bauer C E Elsen S amp Bird T H 1999 Mechanisms forredox control of gene expression A Rev Microbiol 53495ndash523
Bauer C E Elsen S Swem L R Swem D L amp MasudaS 2003 Redox and light regulation of gene expression inphotosynthetic prokaryotes Phil Trans R Soc Lond B 358147ndash154 (DOI 101098rstb20021189)
Berks B C Sargent F De Leeuw E Hinsley A P Stan-ley N R Jack R L Buchanan G amp Palmer T 2000aA novel protein transport system involved in the biogenesisof bacterial electron transfer chains Biochim Biophys ActaBioenerg 1459 325ndash330
Berks B C Sargent F amp Palmer T 2000b The tat proteinexport pathway Mol Microbiol 35 260ndash274
Blackstone N W 1999 Redox control in development andevolution evidence from colonial hydroids J Exp Biol 2023541ndash3553
Blackstone N W 2000 Redox control and the evolution ofmulticellularity Bioessays 22 947ndash953
Blackstone N W 2001 Redox state reactive oxygen speciesand adaptive growth in colonial hydroids J Exp Biol 2041845ndash1853
Bogorad L 1975 Evolution of organelles and eukaryotic gen-omes Science 188 891ndash898
Bogsch E G Sargent F Stanley N R Berks B C Robin-son C amp Palmer T 1998 An essential component of a novelbacterial protein export system with homologues in plastidsand mitochondria J Biol Chem 273 18 003ndash18 006
Brown E C Somanchi A amp Mayfield S P 2001 Interor-ganellar crosstalk new perspectives on signaling from thechloroplast to the nucleus Genome Biol 2 10211ndash10214
Cavalier-Smith T 2002 Chloroplast evolution secondarysymbiogenesis and multiple losses Curr Biol 12 R62ndashR64
Chitnis P R amp Thornber J P 1988 The major light-har-vesting complex of photosystem 2 Aspects of its molecularand cell biology Photosynth Res 16 41ndash63
Chow W S Melis A amp Anderson J M 1990 Adjustmentsof photosystem stoichiometry in chloroplasts improve thequantum efficiency of photosynthesis Proc Natl Acad SciUSA 87 7502ndash7506
Claros M G Perea J Shu Y Samatey F A PopotJ L amp Jacq C 1995 Limitations to in vivo import of hydro-phobic proteins into yeast mitochondria The case of a cyto-plasmically synthesized apocytochrome b Eur J Biochem228 762ndash771
Dalbey R E amp Robinson C 1999 Protein translocation intoand across the bacterial plasma membrane and the plant thy-lakoid membrane Trends Biochem Sci 24 17ndash22
Daley D O Adams K L Clifton R Qualmann S MillarA H Palmer J D Pratje E amp Whelan J 2002 Genetransfer from mitochondrion to nucleus novel mechanismsfor gene activation from Cox2 Plant J 30 11ndash21
Danon A amp Mayfield S P 1994 Light-regulated translationof chloroplast messenger RNAs through redox potentialScience 266 1717ndash1719
de Grey A 2000 Mitochondrial gene therapy an arena for thebiomedical use of inteins Trends Biotechnol 18 394ndash399
Deisenhofer J Epp O Miki K Huber R amp Michel H1985 Structure of the protein subunits in the photosyntheticreaction center of Rhodopseudomonas viridis at 3Ac resolutionNature 318 618ndash624
34 J F Allen Genomes in mitochondria and chloroplasts
Delwiche C F 1999 Tracing the thread of plastid diversitythrough the tapestry of life Am Nat 154 S164ndashS177
Doolittle W F 1999 Phylogenetic classification and the uni-versal tree Science 284 2124ndash2129
Douglas S E 1998 Plastid evolution origins diversitytrends Curr Opin Genet Dev 8 655ndash661
Duby G amp Boutry M 2002 Mitochondrial protein importmachinery and targeting information Plant Sci 162 477ndash490
Eberhard S Drapier D amp Wollman F A 2002 Searchinglimiting steps in the expression of chloroplast encoded pro-teins relations between gene copy number transcriptiontranscript abundance and translation rate in the chloroplastof Chlamydomonas reinhardtii Plant J 31 149ndash160
El Bissati K amp Kirilovsky D 2001 Regulation of psbA andpsaE expression by light quality in Synechocystis species PCC6803 A redox control mechanism Plant Physiol 1251988ndash2000
Ellis R J 1984 The nuclear domination of chloroplast devel-opment Science Progress 69 129ndash142
Embley T M amp Martin W 1998 Molecular evolution ahydrogen-producing mitochondrion Nature 396 517ndash519
Ems S C Morden C W Dixon C K Wolfe K HdePamphilis C W amp Palmer J D 1995 Transcriptionsplicing and editing of plastid RNAs in the non-photosyn-thetic plant Epifagus virginiana Plant Mol Biol 29 721ndash733
Eraso J M amp Kaplan S 2000 From redox flow to gene regu-lation role of the PrrC protein of Rhodobacter sphaeroides241 Biochemistry 39 2052ndash2062
Eraso J M amp Kaplan S 2002 Redox flow as an instrumentof gene regulation Meth Enzymol 348 216ndash229
Escoubas J M Lomas M Laroche J amp Falkowski P G1995 Light-intensity regulation of cab gene-transcription issignaled by the redox state of the plastoquinone pool ProcNatl Acad Sci USA 92 10 237ndash10 241
Ferea T L Botstein D Brown P O amp Rosenzweig R F1999 Systematic changes in gene expression patterns follow-ing adaptive evolution in yeast Proc Natl Acad Sci USA96 9721ndash9726
Forsberg J Rosenquist M Fraysse L amp Allen J F 2001Redox signalling in chloroplasts and mitochondria genomicand biochemical evidence for two-component regulatory sys-tems in bioenergetic organelles Biochem Soc Trans 29403ndash407
Fujita Y 1997 A study on the dynamic features of photosys-tem stoichiometry accomplishments and problems forfuture studies Photosynth Res 53 83ndash93
Gabriel K Buchanan S K amp Lithgow T 2001 The alphaand the beta protein translocation across mitochondrial andplastid outer membranes Trends Biochem Sci 26 36ndash40
Galvis M L E Allen J F amp Hakansson G 1998 Proteinsynthesis by isolated pea mitochondria is dependent on theactivity of respiratory complex II Curr Genet 33 320ndash329
Gatenby A A amp Ellis R J 1990 Chaperone function theassembly of ribulose bisphosphate carboxylase-oxygenase ARev Cell Biol 6 125ndash149
GoldschmidtClermont M 1998 Coordination of nuclear andchloroplast gene expression in plant cells Int Rev CytolSurv Cell Biol 177 115ndash180
Gray M W 1992 The endosymbiont hypothesis revisited IntRev Cytol 141 233ndash357
Gray M W 1999 Evolution of organellar genomes CurrOpin Genet Dev 9 678ndash687
Gray M W 2000 Evolutionary biology mitochondrial geneson the move Nature 408 302ndash305
Gray M W amp Doolittle W F 1982 Has the endosymbionthypothesis been proven Microbiol Rev 46 1ndash42
Gray M W Burger G amp Lang B F 1999 Mitochondrialevolution Science 283 1476ndash1481
Phil Trans R Soc Lond B (2003)
Grebe T W amp Stock J B 1999 The histidine protein kinasesuperfamily Adv Microb Physiol 41 139ndash227
Green B R amp Kuhlbrandt W 1995 Sequence conservationof light-harvesting and stress-response proteins in relation tothe 3-dimensional molecular structure of Lhcii PhotosynthRes 44 139ndash148
Gruissem W amp Tonkyn J C 1993 Control mechanisms ofplastid gene expression Crit Rev Plant Sci 12 19ndash55
Hackstein J H P (and 11 others) 2001 Hydrogenosomesconvergent adaptations of mitochondria to anaerobicenvironments Zool Anal Complex Syst 104 290ndash302
Harrison M A Keen J N Findlay J B C amp Allen J F1990 Modification of a Glnb-like gene-product by photosyn-thetic electron-transport in the cyanobacterium Synechoc-occus 6301 FEBS Lett 264 25ndash28
Harvey A J Kind K L amp Thompson J G 2002 Redoxregulation of early embryo development Reproduction 123479ndash486
Heathcote P Fyfe P K amp Jones M R 2002 Reactioncentres the structure and evolution of biological solarpower Trends Biochem Sci 27 79ndash87
Henze K amp Martin W 2001 How do mitochondrial genesget into the nucleus Trends Genet 17 383ndash387
Herrmann R G amp Westhoff P 2001 Thylakoid biogenesis anddynamics the result of a complex phylogenetic puzzle InRegulation of photosynthesis vol 11 (ed E-M Aro amp BAndersson) pp 1ndash28 Dordrecht The Netherlands Kluwer
Hihara Y Sonoike K Kanehisa M amp Ikeuchi M 2002Search for redox responsive genes in Synechocystis sp PCC6803 by using DNA microarrays Plant Cell Physiol 43S178
Howe C J Barbrook A C amp Lockhart P J 2000 Organellegenes do they jump or are they pushed Trends Genet 1665ndash66
Ikeuchi M 2002 Gene expression and responses to light stressin cyanobacteria Plant Cell Physiol 43 S21
Iuchi S amp Lin E C C 1992 Mutational analysis of signaltransduction by Arcb a membrane sensor protein respon-sible for anaerobic repression of operons involved in the cen-tral aerobic pathways in Escherichia coli J Bacteriol 1743972ndash3980
Jackson-Constan D Akita M amp Keegstra K 2001 Molecu-lar chaperones involved in chloroplast protein importBiochim Biophys Acta Mol Cell Res 1541 102ndash113
Jarvis P amp Soll M 2001 Toc Tic and chloroplast proteinimport Biochim Biophys Acta Mol Cell Res 1541 64ndash79
Jordan P Fromme P Witt H T Klukas O SaengerW amp Krauss N 2001 Three-dimensional structure ofcyanobacterial photosystem I at 25 acngstrom resolutionNature 411 909ndash917
Kaneko T (and 23 others) 1996a Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regionsDNA Res 3 109ndash136
Kaneko T (and 23 others) 1996b Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regions[Supplement] DNA Res 3 185ndash209
Kanevski I amp Maliga P 1994 Relocation of the plastid rbcLgene to the nucleus yields functional ribulose-15-bisphosph-ate carboxylase in tobacco chloroplasts Proc Natl Acad SciUSA 91 1969ndash1973
Kim J amp Mayfield S P 1997 Protein disulfide isomerase as aregulator of chloroplast translational activation Science 2781954ndash1957
Genomes in mitochondria and chloroplasts J F Allen 35
Kirk J T O amp Tilney-Bassett R A E 1978 The plastidsTheir chemistry structure growth and inheritance Amster-dam Elsevier
Koehler C M Merchant S amp Schatz G 1999 How mem-brane proteins travel across the mitochondrial intermem-brane space Trends Biochem Sci 24 428ndash432
Komiya T Rospert S Schatz G amp Mihara K 1997 Bind-ing of mitochondrial precursor proteins to the cytoplasmicdomains of the import receptors Tom70 and Tom20 isdetermined by cytoplasmic chaperones FASEB J 11 1196
Konstantinov Y M Lutsenko G N amp Podsosonny V A1995 Genetic functions of isolated maize mitochondriaunder model changes of redox conditions Biochem MolBiol Int 36 319ndash326
Korab-Laskowska M Rioux P Brossard N LittlejohnT G Gray M W Lang B F amp Burger G 1998 Theorganelle genome database project (GOBASE) Nucleic AcidsRes 26 138ndash144
Kovacs L Wiessner W Kis M Nagy F Mende D ampDemeter S 2000 Short- and long-term redox regulation ofphotosynthetic light energy distribution and photosystemstoichiometry by acetate metabolism in the green alga Chla-mydobotrys stellata Photosynth Res 65 231ndash247
Kuhlbrandt W 1994 Structure and function of the plant light-harvesting complex LHC-II Curr Opin Struct Biol 4519ndash528
Lang B F Gray M W amp Burger G 1999 Mitochondrialgenome evolution and the origin of eukaryotes A RevGenet 33 351ndash397
Lemire B D Fankhauser C Baker A amp Schatz G 1989The mitochondrial targeting function of randomly generatedpeptide sequences correlates with predicted helical amphi-philicity J Biol Chem 264 20 206ndash20 215
Leuenberger D Bally N A Schatz G amp Koehler C M1999 Different import pathways through the mitochondrialintermembrane space for inner membrane proteins EMBOJ 18 4816ndash4822
Li H amp Sherman L A 2000 A redox-responsive regulator ofphotosynthesis gene expression in the cyanobacterium Syne-chocystis sp strain PCC 6803 J Bacteriol 182 4268ndash4277
Lin X Y (and 36 others) 1999 Sequence and analysis of chro-mosome 2 of the plant Arabidopsis thaliana Nature 402761ndash768
Lonsdale D M Hodge T P Howe C J amp Stern D B1983 Maize mitochondrial-DNA contains a sequence hom-ologous to the ribulose-15-bisphosphate carboxylase largesubunit gene of chloroplast DNA Cell 34 1007ndash1014
McFadden G I 1999 Endosymbiosis and evolution of theplant cell Curr Opin Plant Biol 2 513ndash519
McFadden G I 2001 Chloroplast origin and integrationPlant Physiol 125 50ndash53
Mahler H R amp Raff R A 1975 The evolutionary origin ofthe mitochondrion a nonsymbiotic model Int Rev Cytol43 1ndash124
Manfredi G Fu J Ojaimi J Sadlock J E Kwong J QGuy J amp Schon E A 2002 Rescue of a deficiency in ATPsynthesis by transfer of MTATP6 a mitochondrial DNA-encoded gene to the nucleus Nature Genet 30 394ndash399
Marc P Margeot A Devaux F Blugeon C Corral-Debrinski M amp Jacq C 2002 Genome-wide analysis ofmRNAs targeted to yeast mitochondria EMBO Rep 3159ndash164
Marienfeld J Unseld M amp Brennicke A 1999 The mito-chondrial genome of Arabidopsis is composed of both nativeand immigrant information Trends Plant Sci 4 495ndash502
Martin W 1999 A briefly argued case that mitochondria andplastids are descendants of endosymbionts but that thenuclear compartment is not Proc R Soc Lond B 2661387ndash1395 (DOI 101098rspb19990792)
Phil Trans R Soc Lond B (2003)
Martin W amp Herrmann R G 1998 Gene transfer fromorganelles to the nucleus how much what happens andwhy Plant Physiol 118 9ndash17
Martin W amp Muller M 1998 The hydrogen hypothesis forthe first eukaryote Nature 392 37ndash41
Martin W amp Russell M J 2003 On the origins of cells ahypothesis for the evolutionary transitions from abiotic geo-chemistry to chemoautotrophic prokaryotes and from pro-karyotes to nucleated cells Phil Trans R Soc Lond B 35859ndash85 (DOI 101098rstb20021183)
Martin W Stoebe B Goremykin V Hansmann S Hase-gawa M amp Kowallik K V 1998 Gene transfer to thenucleus and the evolution of chloroplasts Nature 393162ndash165
Martin W Hoffmeister M Rotte C amp Henze K 2001 Anoverview of endosymbiotic models for the origins of eukary-otes their ATP-producing organelles (mitochondria andhydrogenosomes) and their heterotrophic lifestyle BiolChem 382 1521ndash1539
Martin W Rujan T Richly E Hansen A Cornelsen SLins T Leister D Stoebe B Hasegawa M amp PennyD 2002 Evolutionary analysis of Arabidopsis cyanobacterialand chloroplast genomes reveals plastid phylogeny and thou-sands of cyanobacterial genes in the nucleus Proc NatlAcad Sci USA 99 12 246ndash12 251
Millen R S (and 12 others) 2001 Many parallel losses of infAfrom chloroplast DNA during angiosperm evolution withmultiple independent transfers to the nucleus Plant Cell 13645ndash658
Morse D Salois P Markovic P amp Hastings J W 1995 Anuclear-encoded form-II RUBISCO in dinoflagellatesScience 268 1622ndash1624
Murakami A Kim S J amp Fujita Y 1997 Changes in photo-system stoichiometry in response to environmental con-ditions for cell growth observed with the cyanophyteSynechocystis PCC 6714 Plant Cell Physiol 38392ndash397
Oh J I amp Kaplan S 2000 Redox signaling globalization ofgene expression EMBO J 19 4237ndash4247
Ozawa T 1995 Mitochondrial-DNA mutations associatedwith aging and degenerative diseases Exp Gerontol 30269ndash290
Palmer J D 1997 Organelle genomes going going goneScience 275 790ndash791
Pearson C K Wilson S B Schaffer R amp Ross A W 1993NAD turnover and utilization of metabolites for RNA syn-thesis in a reaction sensing the redox state of the cyto-chrome-b( 6 )f complex in isolated chloroplasts Eur JBiochem 218 397ndash404
Pfanner N amp Geissler A 2001 Versatility of the mitochon-drial protein import machinery Natl Rev Mol Cell Biol 2339ndash349
Pfanner N amp Wiedemann N 2002 Mitochondrial proteinimport two membranes three translocases Curr Opin CellBiol 14 400ndash411
Pfannschmidt T Nilsson A amp Allen J F 1999a Photosyn-thetic control of chloroplast gene expression Nature 397625ndash628
Pfannschmidt T Nilsson A Tullberg A Link G amp AllenJ F 1999b Direct transcriptional control of the chloroplastgenes psbA and psaAB adjusts photosynthesis to light energydistribution in plants IUBMB Life 48 271ndash276
Pfannschmidt T Allen J F amp Oelmuller R 2001a Prin-ciples of redox control in photosynthesis gene expressionPhysiol Plant 112 1ndash9
Pfannschmidt T Schutze K Brost M amp Oelmuller R2001b A novel mechanism of nuclear photosynthesis generegulation by redox signals from the chloroplast during pho-
36 J F Allen Genomes in mitochondria and chloroplasts
tosystem stoichiometry adjustment J Biol Chem 27636 125ndash36 130
Popot J L amp de Vitry C 1990 On the microassembly of inte-gral membrane proteins A Rev Biophys Biophys Chem 19369ndash403
Race H L Herrmann R G amp Martin W 1999 Why haveorganelles retained genomes Trends Genet 15 364ndash370
Raven J A Johnston A M Parsons R amp Kubler J 1994aThe influence of natural and experimental high O2 concen-trations on O2 evolving phototrophs Biol Rev CambridgePhil Soc 69 61ndash94
Raven J A Johnston A M Parsons R amp Kubler J 1994bThe occurrence and influence on photolithotrophs of highoxygen concentrations Proc R Soc Edinb B 102 193ndash201
Robinson C 2000 The twin-arginine translocation system anovel means of transporting folded proteins in chloroplastsand bacteria Biol Chem 381 89ndash93
Robinson C amp Bolhuis A 2001 Protein targeting by the twin-arginine translocation pathway Natl Rev Mol Cell Biol 2350ndash356
Rodermel S 2001 Pathways of plastid-to-nucleus signalingTrends Plant Sci 6 471ndash478
Roger A J 1999 Reconstructing early events in eukaryoticevolution Am Nat 154 S146ndashS163
Roger A J Svard S G Tovar J Clark C G SmithM W Gillin F D amp Sogin M L 1998 A mitochondrial-like chaperonin 60 gene in Giardia lamblia evidence thatdiplomonads once harbored an endosymbiont related to theprogenitor of mitochondria Proc Natl Acad Sci USA 95229ndash234
Roodyn D B amp Wilkie D 1968 The biogenesis of mitochondriaMethuenrsquos monographs on biological subjects London Methuen
Rujan T amp Martin W 2001 How many genes in Arabidopsiscome from cyanobacteria An estimate from 386 proteinphylogenies Trends Genet 17 113ndash120
Saccone C Gissi C Lanave C Larizza A Pesole G ampReyes A 2000 Evolution of the mitochondrial genetic sys-tem an overview Gene 261 153ndash159
Salzberg S L White O Peterson J amp Eisen J A 2001Microbial genes in the human genome lateral transfer orgene loss Science 292 1903ndash1906
Sargent F Berks B C amp Palmer T 2002 Assembly of mem-brane-bound respiratory complexes by the Tat protein-trans-port system Arch Microbiol 178 77ndash84
Schatz G 1998 Protein transport the doors to organellesNature 395 439ndash440
Schemidt R A Qu J Williams J R amp Brusilow W S1998 Effects of carbon source on expression of F0 genes andon the stoichiometry of the c subunit in the F1 F0 ATPaseof Escherichia coli J Bacteriol 180 3205ndash3208
Selosse M A Albert B R amp Godelle B 2001 Reducing thegenome size of organelles favours gene transfer to thenucleus Trends Ecol Evol 16 135ndash141
Stern D B amp Lonsdale D M 1982 Mitochondrial and chlo-roplast genomes of maize have a 12- kilobase DNA-sequencein common Nature 299 698ndash702
Stock D Gibbons C Arechaga I Leslie A G W ampWalker J E 2000 The rotary mechanism of ATP synthaseCurr Opin Struct Biol 10 672ndash679
Stoebe B amp Maier U G 2002 One two three naturersquos toolbox for building plastids Protoplasma 219 123ndash130
Stoebe B Martin W amp Kowallik K V 1998 Distributionand nomenclature of protein-coding genes in 12 sequencedchloroplast genomes Plant Mol Biol Rep 16 243ndash255
Stupar R M Lilly J W Town C D Cheng Z Kaul SBuell C R amp Jiang J M 2001 Complex mtDNA consti-tutes an approximate 620 kb insertion on Arabidopsis thali-ana chromosome 2 implication of potential sequencing
Phil Trans R Soc Lond B (2003)
errors caused by large-unit repeats Proc Natl Acad SciUSA 98 5099ndash5103
Surpin M Larkin R M amp Chory J 2002 Signal transduc-tion between the chloroplast and the nucleus Plant Cell 14S327ndashS338
Tjaden J Schwoppe C Mohlmann T Quick P W ampNeuhaus H E 1998 Expression of a plastidic ATPADPtransporter gene in Escherichia coli leads to a functional aden-ine nucleotide transport system in the bacterial cytoplasmicmembrane J Biol Chem 273 9630ndash9636
Tovar J Fischer A amp Clark C G 1999 The mitosome a novelorganelle related to mitochondria in the amitochondrial para-site Entamoeba histolytica Mol Microbiol 32 1013ndash1021
Trebitsh T amp Danon A 2001 Translation of chloroplastpsbA mRNA is regulated by signals initiated by both photos-ystems II and I Proc Natl Acad Sci USA 98 12 289ndash12 294
Tullberg A Alexciev K Pfannschmidt T amp Allen J F2000 Photosynthetic electron flow regulates transcription ofthe psaB gene in pea (Pisum sativum L) chloroplasts throughthe redox state of the plastoquinone pool Plant Cell Physiol41 1045ndash1054
Turmel M Lemieux C Burger G Lang B F Otis CPlante I amp Gray M W 1999 The complete mitochondrialDNA sequences of Nephroselmis olivacea and Pedinomonasminor two radically different evolutionary patterns withingreen algae Plant Cell 11 1717ndash1729
Unden G amp Bongaerts J 1997 Alternative respiratory path-ways of Escherichia coli energetics and transcriptional regu-lation in response to electron acceptors Biochim BiophysActa Bioenerg 1320 217ndash234
Unseld M Marienfeld J R Brandt P amp Brennicke A1997 The mitochondrial genome of Arabidopsis thaliana con-tains 57 genes in 366 924 nucleotides Nature Genet 1557ndash61
Van der Giezen M Slotboom D J Horner D S DyalP L Harding M Xue G P Embley T M amp KunjiE R S 2002 Conserved properties of hydrogenosomal andmitochondrial ADPATP carriers a common origin for bothorganelles EMBO J 21 572ndash579
Von Heijne G 1986 Why mitochondria need a genome FEBSLett 198 1ndash4
Westphal S Soll J amp Vothknecht U C 2001 A vesicle trans-port system inside chloroplasts FEBS Lett 506 257ndash261
Whitehouse H L K 1969 Towards an understanding of themechanism of heredity London Edward Arnold Ltd
Williams B A P Hirt R P Lucocq J M amp Embley T M2002 A mitochondrial remnant in the microsporidian Trachi-pleistophora hominis Nature 418 865ndash869
Wilson R J (and 10 others) 1996a Complete gene map of theplastid-like DNA of the malaria parasite Plasmodium falcipa-rum J Mol Biol 261 155ndash172
Wilson S B Davidson G S Thomson L M amp PearsonC K 1996b Redox control of RNA synthesis in potato mito-chondria Eur J Biochem 242 81ndash85
Winkler H H amp Neuhaus H E 1999 Non-mitochondrialATP transport Trends Biochem Sci 24 64ndash68
Wolfe K H Morden C W amp Palmer J D 1992 Small sin-gle-copy region of plastid DNA in the non-photosyntheticangiosperm Epifagus virginiana contains only two genes Dif-ferences among dicots monocots and bryophytes in geneorganization at a non-bioenergetic locus J Mol Biol 22395ndash104
Wollman F A Minai L amp Nechushtai R 1999 The biogen-esis and assembly of photosynthetic proteins in thylakoidmembranes Biochim Biophys Acta Bioenerg 1411 21ndash85
Yeliseev A A amp Kaplan S 2000 TspO of Rhodobactersphaeroides a structural and functional model for the mam-
Genomes in mitochondria and chloroplasts J F Allen 37
malian peripheral benzodiazepine receptor J Biol Chem275 5657ndash5667
Zerges W 2002 Does complexity constrain organelle evol-ution Trends Plant Sci 7 175ndash182
Zhang Z D Green B R amp Cavalier-Smith T 1999 Singlegene circles in dinoflagellate chloroplast genomes Nature400 155ndash159
Zhang Z D Cavalier-Smith T amp Green B R 2001 A fam-ily of selfish minicircular chromosomes with jumbled chloro-plast gene fragments from a dinoflagellate Mol Biol Evol18 1558ndash1565
Zouni A Witt H T Kern J Fromme P Krauss NSaenger W amp Orth P 2001 Crystal structure of photosys-tem II from Synechococcus elongatus at 38 acngstrom resol-ution Nature 409 739ndash743
DiscussionD S Horner (Dipartimento di Fisiologia e Biochimica
Generali University of Milan Milan Italy) Given thatthere are components of these membrane-bound energy-transduction pathways that are consistently nuclear enco-ded not the key ones you pointed out but others do youexpect perhaps a slower perhaps a more crude but yet aredox communication between the organelle andexpression of nuclear-encoded genes
J F Allen There is and there is no question about itI think that Bogoradrsquos lock-in hypothesis has somethingto say here and there are other people who say it is all todo with assembling multi-subunit complexes If you havecontrol over the initial construction of the core aroundwhich the whole complex is eventually built then redoxcontrol or some sort of control over synthesis of that corewill control construction of the whole thing eventuallyand I think that may well be largely correct I think it ispossible in principle for nuclear genes to make precursorsaccording to redox instructions but it is not really soimportant that those precursors are here and now presentin exactly the right quantities because the precursors canto a limited extent hang around and wait Actually thereis proteolytic degradation of unattached precursors and soon but I think the fact that these chloroplast and mito-chondrially DNA-encoded subunits are at the core ofthese energy-transducing complexes is the crucial issue
T Cavalier-Smith (Department of Zoology University ofOxford Oxford UK ) In the case of mitochondria the sub-set of genes that remains is much more variable than youimplied It runs from three proteins in the minimal casesup to I think over 90 That is a big difference and manyof the genes in different lineages of protists that remainhave nothing to do with redox control they are things likeribosomal proteins and so on So I think that even thoughwhat you say about redox control of certain genes mayactually be an important thing it is not the fundamentaloverall reason why the genomes are maintained It may bepart of the explanation for why certain genes are alwaysincluded in that subset but even in mitochondria I thinkthat cytochrome b is the only one that is always presentSo I think that both this great diversity of what has beenretained in different lineages and the nucleomorph caseindicate that evolution was able to do one thing in onelineage 500 million years ago and never got around todoing it for 500 million years in another lineage so thereis a lot more haphazardness and evolutionary accidentsand contingency than a straight functional interpretation
Phil Trans R Soc Lond B (2003)
there is a lot of just sheer lsquohappenedndashorndashdidnrsquotndashhappenrsquoor carryover
J F Allen I was not implying anything about the rangeof mitochondrial genomes I have a limited amount oftime so I just talked about what I could talk about withinthat limit Of course there will be a range and this willdepend upon the physiology and bioenergetics of theparticular organism But what I am trying to do is to seewhat the common properties of the irreducible core areof components that are always encoded in the organellesand cytochrome b is a perfect example because that reallyis the thing that binds these two all-important quinoneswithout which there is no Q cycle without which there isno proton pumping and bioenergetics and in Plasmodiumwhere the mitochondrion is reduced to this bare mini-mum there is still a mitochondrial gene for cytochromeb Now my argument about the ribosomal proteins andall the rest of it is that in order to be able to exert thisall-important redox control over gene expression of certaincore components you need an in situ genetic system andthis can vary in composition for reasons that I cannotdescribe Maybe you need RNA polymerase or maybeyou can import RNA polymerase In plastids you haveboth of these options You are going to need ribosomestransfer RNAs and the rest of the apparatus of geneexpression and that needs to be in situ but we are stillleft with the question why does there have to be any gen-etic system at all The mature tobacco or spinach chloro-plast seems to have an entire genetic system just to makeone protein D1 which is all it ever needs to resynthesizeI think that this is a clue It is worth having this very unec-onomical crazy system because there is an overridingrequirement to connect electron transfer with geneexpression directly and unambiguouslymdashwithout recourseto a bureaucracy if you like which is what I think thenucleus is
K Willison (Institute of Cancer Research Chester BeattyLaboratories London UK ) These photosynthetic reactioncomplexes are a sort of baroque mixture of polypeptidechains and cofactors and my understanding is that someof them in high photon flux can turn over within say 30minutes so I am wondering if there is a correlationbetween gene retention and the turnover rates of some ofthe components and the complexes it seems to me thatthat could really be a limiting factor and the reason thatthe proteins had to be encoded locally in order to get thecomplexes to be assembled rapidly
J F Allen I think that is right You can only alter thesteady-state concentration of a protein complex by affect-ing the synthesis and breakdown and therefore turnoverThere is an extreme example the D1 protein encoded bythe psbA gene of chloroplasts which is always in the chlor-oplast genome without exception High light will tend todegrade this protein and when the high light goes awaythere is an absolute need very quickly to bring it back upagain I think that is a very good example Whether thisargument applies to genes such as cytochrome oxidase Ido not know
C J Leaver (Department of Plant Sciences University ofOxford Oxford UK ) Your experiments tend to relatemainly to the gene expression side What about proteinRNA turnover the whole concept of redox control ofthatmdashdo we know anything
38 J F Allen Genomes in mitochondria and chloroplasts
J F Allen Well quite a lot is known about chloro-plasts There is redox control of messenger RNA stabilityfor this same D1 protein which has been described byStephen Mayfield and his co-workers The evidence is thatthe site of redox control is ferredoxin via thioredoxin andthe implication is that thioredoxin tells whether non-cyclicelectron transport is working at all When it is workingthere is a signal that says lsquoOK now continue with all theprotein synthesisrsquo It is not particularly specific but moreof a global effect I think this is what Mayfield argues butof course this is not inconsistent with my argument I haveconcentrated on quinone pools and medium-potentialredox regulatory control because that is where I can seesignals about the dangerous consequences of getting itwrong but there is control of other levels of geneexpression In mitochondria I am not so sure I wouldbounce the question back to you on that one
W Martin (Institute of Botany III Heinrich-Heine Univ-ersitat Dusseldorf Dusseldorf Germany) The nucleomorphgenome was cited as evidence in conflict with your modelbut is that true The nucleomorph genome is not in abioenergetic organelle in that sense It resides in the cyto-sol so does the size of the nucleomorph genome workagainst your model Is it really evidence against yourmodel
J F Allen I do not know why should it be evidenceI cannot explain why the nucleomorph has a genomeThat is why I clutched at the straw that it is storage fornucleic acid precursors
Phil Trans R Soc Lond B (2003)
T Cavalier-Smith The reason I introduced the nucleo-morph is not to say it is direct evidence against yourhypothesis but to show that there probably was no singlefunctional reason for the retention of every gene in everyorganelle It may be that there are specific physiologicalreasons for the retention of certain genes and thereforeyou might be right about a subset of genes but I verymuch doubt if there are specific physiological reasons forthe retention of every single gene that is retained differen-tially because odd things happen in evolution Somethings are easy and sometimes things are difficult Evol-ution is history
J F Allen Of course I agree with that I do not thinkI can explain everything but I do think I can explainsomething If I have done that then I am content
GLOSSARY
AAC ADPndashATP carrierCES control by epistasy of synthesisCORR co-location for redox regulationLHC light-harvesting complexPCC Pasteur culture collectionORF open reading frameRUBISCO ribulose-15-bisphosphate carboxylasendashoxygenaseTAT twin-arginine translocase
Visit httpwwwpubsroyalsocacuk to see an electronic appendix tothis paper

Genomes in mitochondria and chloroplasts J F Allen 33
The Swedish Natural Sciences Research Council (NFR) andthe Crafoord Foundation are gratefully acknowledged
REFERENCES
Adams K L Daley D O Qiu Y L Whelan J amp PalmerJ D 2000 Repeated recent and diverse transfers of a mito-chondrial gene to the nucleus in flowering plants Nature408 354ndash357
Alberts B Bray D Lewis J Raff M Roberts K amp Wat-son J D 1994 The molecular biology of the cell New Yorkand London Garland Publishing
Allen C A Hakansson G amp Allen J F 1995 Redox con-ditions specify the proteins synthesized by isolated chloro-plasts and mitochondria Redox Rep 1 119ndash123
Allen J F 1992 Protein phosphorylation in regulation ofphotosynthesis Biochim Biophys Acta 1098 275ndash335
Allen J F 1993a Control of gene expression by redox poten-tial and the requirement for chloroplast and mitochondrialgenomes J Theor Biol 165 609ndash631
Allen J F 1993b Redox control of gene expression and thefunction of chloroplast genomes a hypothesis PhotosynthRes 36 95ndash102
Allen J F 1993c Redox control of transcription sensorsresponse regulators activators and repressors FEBS Lett332 203ndash207
Allen J F 1995 Thylakoid proteinndashphosphorylation state-1ndashstate-2 transitions and photosystem stoichiometry adjust-ment redox control at multiple levels of gene expressionPhysiol Plant 93 196ndash205
Allen J F 1996 Separate sexes and the mitochondrial theoryof ageing J Theor Biol 180 135ndash140
Allen J F 2002 Photosynthesis of ATP-electrons protonpumps rotors and poise Cell 110 273ndash276
Allen J F amp Allen C A 1999 A mitochondrial model forpremature ageing of somatically cloned mammals IUBMBLife 48 369ndash372
Allen J F amp Nilsson A 1997 Redox signalling and the struc-tural basis of regulation of photosynthesis by protein phos-phorylation Physiol Plant 100 863ndash868
Allen J F amp Pfannschmidt T 2000 Balancing the two photo-systems photosynthetic electron transfer governs transcrip-tion of reaction centre genes in chloroplasts Phil TransR Soc Lond B 355 1351ndash1357 (DOI 101098rstb20000697)
Allen J F amp Race H L 2002 Will the real LHC II kinaseplease step forward Sciencersquos STKE httpwwwstkeorgcgicontentfullsigtrans2002155pe43
Allen J F amp Raven J A 1996 Free-radical-induced mutationvs redox regulation costs and benefits of genes in organellesJ Mol Evol 42 482ndash492
Archibald J M Cavalier-Smith T Maier U amp Douglas S2001 Molecular chaperones encoded by a reduced nucleusthe cryptomonad nucleomorph J Mol Evol 52 490ndash501
Ashby M K amp Mullineaux C W 1999a Cyanobacterialycf27 gene products regulate energy transfer from phycobili-somes to photosystems I and II FEMS Microbiol Lett 181253ndash260
Ashby M K amp Mullineaux C W 1999b The role of ApcDand ApcF in energy transfer from phycobilisomes to PSI andPSII in a cyanobacterium Photosynth Res 61 169ndash179
Ashby M K Houmard J amp Mullineaux C W 2002 Theycf27 genes from cyanobacteria and eukaryotic algae distri-bution and implications for chloroplast evolution FEMSMicrobiol Lett 214 25ndash30
Attardi G amp Schatz G 1988 Biogenesis of mitochondria ARev Cell Biol 4 289ndash333
Azem A Oppliger W Lustig A Jeno P Feifel B SchatzG amp Horst M 1997 The mitochondrial hsp70 chaperone
Phil Trans R Soc Lond B (2003)
system effect of adenine nucleotides peptide substrate andmGrpE on the oligomeric state of mhsp70 J Biol Chem272 20 901ndash20 906
Barbrook A C Symington H Nisbet R E R LarkumA amp Howe C J 2001 Organisation and expression of theplastid genome of the dinoflagellate Amphidinium opercula-tum Mol Genet Genomics 266 632ndash638
Bauer C E Elsen S amp Bird T H 1999 Mechanisms forredox control of gene expression A Rev Microbiol 53495ndash523
Bauer C E Elsen S Swem L R Swem D L amp MasudaS 2003 Redox and light regulation of gene expression inphotosynthetic prokaryotes Phil Trans R Soc Lond B 358147ndash154 (DOI 101098rstb20021189)
Berks B C Sargent F De Leeuw E Hinsley A P Stan-ley N R Jack R L Buchanan G amp Palmer T 2000aA novel protein transport system involved in the biogenesisof bacterial electron transfer chains Biochim Biophys ActaBioenerg 1459 325ndash330
Berks B C Sargent F amp Palmer T 2000b The tat proteinexport pathway Mol Microbiol 35 260ndash274
Blackstone N W 1999 Redox control in development andevolution evidence from colonial hydroids J Exp Biol 2023541ndash3553
Blackstone N W 2000 Redox control and the evolution ofmulticellularity Bioessays 22 947ndash953
Blackstone N W 2001 Redox state reactive oxygen speciesand adaptive growth in colonial hydroids J Exp Biol 2041845ndash1853
Bogorad L 1975 Evolution of organelles and eukaryotic gen-omes Science 188 891ndash898
Bogsch E G Sargent F Stanley N R Berks B C Robin-son C amp Palmer T 1998 An essential component of a novelbacterial protein export system with homologues in plastidsand mitochondria J Biol Chem 273 18 003ndash18 006
Brown E C Somanchi A amp Mayfield S P 2001 Interor-ganellar crosstalk new perspectives on signaling from thechloroplast to the nucleus Genome Biol 2 10211ndash10214
Cavalier-Smith T 2002 Chloroplast evolution secondarysymbiogenesis and multiple losses Curr Biol 12 R62ndashR64
Chitnis P R amp Thornber J P 1988 The major light-har-vesting complex of photosystem 2 Aspects of its molecularand cell biology Photosynth Res 16 41ndash63
Chow W S Melis A amp Anderson J M 1990 Adjustmentsof photosystem stoichiometry in chloroplasts improve thequantum efficiency of photosynthesis Proc Natl Acad SciUSA 87 7502ndash7506
Claros M G Perea J Shu Y Samatey F A PopotJ L amp Jacq C 1995 Limitations to in vivo import of hydro-phobic proteins into yeast mitochondria The case of a cyto-plasmically synthesized apocytochrome b Eur J Biochem228 762ndash771
Dalbey R E amp Robinson C 1999 Protein translocation intoand across the bacterial plasma membrane and the plant thy-lakoid membrane Trends Biochem Sci 24 17ndash22
Daley D O Adams K L Clifton R Qualmann S MillarA H Palmer J D Pratje E amp Whelan J 2002 Genetransfer from mitochondrion to nucleus novel mechanismsfor gene activation from Cox2 Plant J 30 11ndash21
Danon A amp Mayfield S P 1994 Light-regulated translationof chloroplast messenger RNAs through redox potentialScience 266 1717ndash1719
de Grey A 2000 Mitochondrial gene therapy an arena for thebiomedical use of inteins Trends Biotechnol 18 394ndash399
Deisenhofer J Epp O Miki K Huber R amp Michel H1985 Structure of the protein subunits in the photosyntheticreaction center of Rhodopseudomonas viridis at 3Ac resolutionNature 318 618ndash624
34 J F Allen Genomes in mitochondria and chloroplasts
Delwiche C F 1999 Tracing the thread of plastid diversitythrough the tapestry of life Am Nat 154 S164ndashS177
Doolittle W F 1999 Phylogenetic classification and the uni-versal tree Science 284 2124ndash2129
Douglas S E 1998 Plastid evolution origins diversitytrends Curr Opin Genet Dev 8 655ndash661
Duby G amp Boutry M 2002 Mitochondrial protein importmachinery and targeting information Plant Sci 162 477ndash490
Eberhard S Drapier D amp Wollman F A 2002 Searchinglimiting steps in the expression of chloroplast encoded pro-teins relations between gene copy number transcriptiontranscript abundance and translation rate in the chloroplastof Chlamydomonas reinhardtii Plant J 31 149ndash160
El Bissati K amp Kirilovsky D 2001 Regulation of psbA andpsaE expression by light quality in Synechocystis species PCC6803 A redox control mechanism Plant Physiol 1251988ndash2000
Ellis R J 1984 The nuclear domination of chloroplast devel-opment Science Progress 69 129ndash142
Embley T M amp Martin W 1998 Molecular evolution ahydrogen-producing mitochondrion Nature 396 517ndash519
Ems S C Morden C W Dixon C K Wolfe K HdePamphilis C W amp Palmer J D 1995 Transcriptionsplicing and editing of plastid RNAs in the non-photosyn-thetic plant Epifagus virginiana Plant Mol Biol 29 721ndash733
Eraso J M amp Kaplan S 2000 From redox flow to gene regu-lation role of the PrrC protein of Rhodobacter sphaeroides241 Biochemistry 39 2052ndash2062
Eraso J M amp Kaplan S 2002 Redox flow as an instrumentof gene regulation Meth Enzymol 348 216ndash229
Escoubas J M Lomas M Laroche J amp Falkowski P G1995 Light-intensity regulation of cab gene-transcription issignaled by the redox state of the plastoquinone pool ProcNatl Acad Sci USA 92 10 237ndash10 241
Ferea T L Botstein D Brown P O amp Rosenzweig R F1999 Systematic changes in gene expression patterns follow-ing adaptive evolution in yeast Proc Natl Acad Sci USA96 9721ndash9726
Forsberg J Rosenquist M Fraysse L amp Allen J F 2001Redox signalling in chloroplasts and mitochondria genomicand biochemical evidence for two-component regulatory sys-tems in bioenergetic organelles Biochem Soc Trans 29403ndash407
Fujita Y 1997 A study on the dynamic features of photosys-tem stoichiometry accomplishments and problems forfuture studies Photosynth Res 53 83ndash93
Gabriel K Buchanan S K amp Lithgow T 2001 The alphaand the beta protein translocation across mitochondrial andplastid outer membranes Trends Biochem Sci 26 36ndash40
Galvis M L E Allen J F amp Hakansson G 1998 Proteinsynthesis by isolated pea mitochondria is dependent on theactivity of respiratory complex II Curr Genet 33 320ndash329
Gatenby A A amp Ellis R J 1990 Chaperone function theassembly of ribulose bisphosphate carboxylase-oxygenase ARev Cell Biol 6 125ndash149
GoldschmidtClermont M 1998 Coordination of nuclear andchloroplast gene expression in plant cells Int Rev CytolSurv Cell Biol 177 115ndash180
Gray M W 1992 The endosymbiont hypothesis revisited IntRev Cytol 141 233ndash357
Gray M W 1999 Evolution of organellar genomes CurrOpin Genet Dev 9 678ndash687
Gray M W 2000 Evolutionary biology mitochondrial geneson the move Nature 408 302ndash305
Gray M W amp Doolittle W F 1982 Has the endosymbionthypothesis been proven Microbiol Rev 46 1ndash42
Gray M W Burger G amp Lang B F 1999 Mitochondrialevolution Science 283 1476ndash1481
Phil Trans R Soc Lond B (2003)
Grebe T W amp Stock J B 1999 The histidine protein kinasesuperfamily Adv Microb Physiol 41 139ndash227
Green B R amp Kuhlbrandt W 1995 Sequence conservationof light-harvesting and stress-response proteins in relation tothe 3-dimensional molecular structure of Lhcii PhotosynthRes 44 139ndash148
Gruissem W amp Tonkyn J C 1993 Control mechanisms ofplastid gene expression Crit Rev Plant Sci 12 19ndash55
Hackstein J H P (and 11 others) 2001 Hydrogenosomesconvergent adaptations of mitochondria to anaerobicenvironments Zool Anal Complex Syst 104 290ndash302
Harrison M A Keen J N Findlay J B C amp Allen J F1990 Modification of a Glnb-like gene-product by photosyn-thetic electron-transport in the cyanobacterium Synechoc-occus 6301 FEBS Lett 264 25ndash28
Harvey A J Kind K L amp Thompson J G 2002 Redoxregulation of early embryo development Reproduction 123479ndash486
Heathcote P Fyfe P K amp Jones M R 2002 Reactioncentres the structure and evolution of biological solarpower Trends Biochem Sci 27 79ndash87
Henze K amp Martin W 2001 How do mitochondrial genesget into the nucleus Trends Genet 17 383ndash387
Herrmann R G amp Westhoff P 2001 Thylakoid biogenesis anddynamics the result of a complex phylogenetic puzzle InRegulation of photosynthesis vol 11 (ed E-M Aro amp BAndersson) pp 1ndash28 Dordrecht The Netherlands Kluwer
Hihara Y Sonoike K Kanehisa M amp Ikeuchi M 2002Search for redox responsive genes in Synechocystis sp PCC6803 by using DNA microarrays Plant Cell Physiol 43S178
Howe C J Barbrook A C amp Lockhart P J 2000 Organellegenes do they jump or are they pushed Trends Genet 1665ndash66
Ikeuchi M 2002 Gene expression and responses to light stressin cyanobacteria Plant Cell Physiol 43 S21
Iuchi S amp Lin E C C 1992 Mutational analysis of signaltransduction by Arcb a membrane sensor protein respon-sible for anaerobic repression of operons involved in the cen-tral aerobic pathways in Escherichia coli J Bacteriol 1743972ndash3980
Jackson-Constan D Akita M amp Keegstra K 2001 Molecu-lar chaperones involved in chloroplast protein importBiochim Biophys Acta Mol Cell Res 1541 102ndash113
Jarvis P amp Soll M 2001 Toc Tic and chloroplast proteinimport Biochim Biophys Acta Mol Cell Res 1541 64ndash79
Jordan P Fromme P Witt H T Klukas O SaengerW amp Krauss N 2001 Three-dimensional structure ofcyanobacterial photosystem I at 25 acngstrom resolutionNature 411 909ndash917
Kaneko T (and 23 others) 1996a Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regionsDNA Res 3 109ndash136
Kaneko T (and 23 others) 1996b Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regions[Supplement] DNA Res 3 185ndash209
Kanevski I amp Maliga P 1994 Relocation of the plastid rbcLgene to the nucleus yields functional ribulose-15-bisphosph-ate carboxylase in tobacco chloroplasts Proc Natl Acad SciUSA 91 1969ndash1973
Kim J amp Mayfield S P 1997 Protein disulfide isomerase as aregulator of chloroplast translational activation Science 2781954ndash1957
Genomes in mitochondria and chloroplasts J F Allen 35
Kirk J T O amp Tilney-Bassett R A E 1978 The plastidsTheir chemistry structure growth and inheritance Amster-dam Elsevier
Koehler C M Merchant S amp Schatz G 1999 How mem-brane proteins travel across the mitochondrial intermem-brane space Trends Biochem Sci 24 428ndash432
Komiya T Rospert S Schatz G amp Mihara K 1997 Bind-ing of mitochondrial precursor proteins to the cytoplasmicdomains of the import receptors Tom70 and Tom20 isdetermined by cytoplasmic chaperones FASEB J 11 1196
Konstantinov Y M Lutsenko G N amp Podsosonny V A1995 Genetic functions of isolated maize mitochondriaunder model changes of redox conditions Biochem MolBiol Int 36 319ndash326
Korab-Laskowska M Rioux P Brossard N LittlejohnT G Gray M W Lang B F amp Burger G 1998 Theorganelle genome database project (GOBASE) Nucleic AcidsRes 26 138ndash144
Kovacs L Wiessner W Kis M Nagy F Mende D ampDemeter S 2000 Short- and long-term redox regulation ofphotosynthetic light energy distribution and photosystemstoichiometry by acetate metabolism in the green alga Chla-mydobotrys stellata Photosynth Res 65 231ndash247
Kuhlbrandt W 1994 Structure and function of the plant light-harvesting complex LHC-II Curr Opin Struct Biol 4519ndash528
Lang B F Gray M W amp Burger G 1999 Mitochondrialgenome evolution and the origin of eukaryotes A RevGenet 33 351ndash397
Lemire B D Fankhauser C Baker A amp Schatz G 1989The mitochondrial targeting function of randomly generatedpeptide sequences correlates with predicted helical amphi-philicity J Biol Chem 264 20 206ndash20 215
Leuenberger D Bally N A Schatz G amp Koehler C M1999 Different import pathways through the mitochondrialintermembrane space for inner membrane proteins EMBOJ 18 4816ndash4822
Li H amp Sherman L A 2000 A redox-responsive regulator ofphotosynthesis gene expression in the cyanobacterium Syne-chocystis sp strain PCC 6803 J Bacteriol 182 4268ndash4277
Lin X Y (and 36 others) 1999 Sequence and analysis of chro-mosome 2 of the plant Arabidopsis thaliana Nature 402761ndash768
Lonsdale D M Hodge T P Howe C J amp Stern D B1983 Maize mitochondrial-DNA contains a sequence hom-ologous to the ribulose-15-bisphosphate carboxylase largesubunit gene of chloroplast DNA Cell 34 1007ndash1014
McFadden G I 1999 Endosymbiosis and evolution of theplant cell Curr Opin Plant Biol 2 513ndash519
McFadden G I 2001 Chloroplast origin and integrationPlant Physiol 125 50ndash53
Mahler H R amp Raff R A 1975 The evolutionary origin ofthe mitochondrion a nonsymbiotic model Int Rev Cytol43 1ndash124
Manfredi G Fu J Ojaimi J Sadlock J E Kwong J QGuy J amp Schon E A 2002 Rescue of a deficiency in ATPsynthesis by transfer of MTATP6 a mitochondrial DNA-encoded gene to the nucleus Nature Genet 30 394ndash399
Marc P Margeot A Devaux F Blugeon C Corral-Debrinski M amp Jacq C 2002 Genome-wide analysis ofmRNAs targeted to yeast mitochondria EMBO Rep 3159ndash164
Marienfeld J Unseld M amp Brennicke A 1999 The mito-chondrial genome of Arabidopsis is composed of both nativeand immigrant information Trends Plant Sci 4 495ndash502
Martin W 1999 A briefly argued case that mitochondria andplastids are descendants of endosymbionts but that thenuclear compartment is not Proc R Soc Lond B 2661387ndash1395 (DOI 101098rspb19990792)
Phil Trans R Soc Lond B (2003)
Martin W amp Herrmann R G 1998 Gene transfer fromorganelles to the nucleus how much what happens andwhy Plant Physiol 118 9ndash17
Martin W amp Muller M 1998 The hydrogen hypothesis forthe first eukaryote Nature 392 37ndash41
Martin W amp Russell M J 2003 On the origins of cells ahypothesis for the evolutionary transitions from abiotic geo-chemistry to chemoautotrophic prokaryotes and from pro-karyotes to nucleated cells Phil Trans R Soc Lond B 35859ndash85 (DOI 101098rstb20021183)
Martin W Stoebe B Goremykin V Hansmann S Hase-gawa M amp Kowallik K V 1998 Gene transfer to thenucleus and the evolution of chloroplasts Nature 393162ndash165
Martin W Hoffmeister M Rotte C amp Henze K 2001 Anoverview of endosymbiotic models for the origins of eukary-otes their ATP-producing organelles (mitochondria andhydrogenosomes) and their heterotrophic lifestyle BiolChem 382 1521ndash1539
Martin W Rujan T Richly E Hansen A Cornelsen SLins T Leister D Stoebe B Hasegawa M amp PennyD 2002 Evolutionary analysis of Arabidopsis cyanobacterialand chloroplast genomes reveals plastid phylogeny and thou-sands of cyanobacterial genes in the nucleus Proc NatlAcad Sci USA 99 12 246ndash12 251
Millen R S (and 12 others) 2001 Many parallel losses of infAfrom chloroplast DNA during angiosperm evolution withmultiple independent transfers to the nucleus Plant Cell 13645ndash658
Morse D Salois P Markovic P amp Hastings J W 1995 Anuclear-encoded form-II RUBISCO in dinoflagellatesScience 268 1622ndash1624
Murakami A Kim S J amp Fujita Y 1997 Changes in photo-system stoichiometry in response to environmental con-ditions for cell growth observed with the cyanophyteSynechocystis PCC 6714 Plant Cell Physiol 38392ndash397
Oh J I amp Kaplan S 2000 Redox signaling globalization ofgene expression EMBO J 19 4237ndash4247
Ozawa T 1995 Mitochondrial-DNA mutations associatedwith aging and degenerative diseases Exp Gerontol 30269ndash290
Palmer J D 1997 Organelle genomes going going goneScience 275 790ndash791
Pearson C K Wilson S B Schaffer R amp Ross A W 1993NAD turnover and utilization of metabolites for RNA syn-thesis in a reaction sensing the redox state of the cyto-chrome-b( 6 )f complex in isolated chloroplasts Eur JBiochem 218 397ndash404
Pfanner N amp Geissler A 2001 Versatility of the mitochon-drial protein import machinery Natl Rev Mol Cell Biol 2339ndash349
Pfanner N amp Wiedemann N 2002 Mitochondrial proteinimport two membranes three translocases Curr Opin CellBiol 14 400ndash411
Pfannschmidt T Nilsson A amp Allen J F 1999a Photosyn-thetic control of chloroplast gene expression Nature 397625ndash628
Pfannschmidt T Nilsson A Tullberg A Link G amp AllenJ F 1999b Direct transcriptional control of the chloroplastgenes psbA and psaAB adjusts photosynthesis to light energydistribution in plants IUBMB Life 48 271ndash276
Pfannschmidt T Allen J F amp Oelmuller R 2001a Prin-ciples of redox control in photosynthesis gene expressionPhysiol Plant 112 1ndash9
Pfannschmidt T Schutze K Brost M amp Oelmuller R2001b A novel mechanism of nuclear photosynthesis generegulation by redox signals from the chloroplast during pho-
36 J F Allen Genomes in mitochondria and chloroplasts
tosystem stoichiometry adjustment J Biol Chem 27636 125ndash36 130
Popot J L amp de Vitry C 1990 On the microassembly of inte-gral membrane proteins A Rev Biophys Biophys Chem 19369ndash403
Race H L Herrmann R G amp Martin W 1999 Why haveorganelles retained genomes Trends Genet 15 364ndash370
Raven J A Johnston A M Parsons R amp Kubler J 1994aThe influence of natural and experimental high O2 concen-trations on O2 evolving phototrophs Biol Rev CambridgePhil Soc 69 61ndash94
Raven J A Johnston A M Parsons R amp Kubler J 1994bThe occurrence and influence on photolithotrophs of highoxygen concentrations Proc R Soc Edinb B 102 193ndash201
Robinson C 2000 The twin-arginine translocation system anovel means of transporting folded proteins in chloroplastsand bacteria Biol Chem 381 89ndash93
Robinson C amp Bolhuis A 2001 Protein targeting by the twin-arginine translocation pathway Natl Rev Mol Cell Biol 2350ndash356
Rodermel S 2001 Pathways of plastid-to-nucleus signalingTrends Plant Sci 6 471ndash478
Roger A J 1999 Reconstructing early events in eukaryoticevolution Am Nat 154 S146ndashS163
Roger A J Svard S G Tovar J Clark C G SmithM W Gillin F D amp Sogin M L 1998 A mitochondrial-like chaperonin 60 gene in Giardia lamblia evidence thatdiplomonads once harbored an endosymbiont related to theprogenitor of mitochondria Proc Natl Acad Sci USA 95229ndash234
Roodyn D B amp Wilkie D 1968 The biogenesis of mitochondriaMethuenrsquos monographs on biological subjects London Methuen
Rujan T amp Martin W 2001 How many genes in Arabidopsiscome from cyanobacteria An estimate from 386 proteinphylogenies Trends Genet 17 113ndash120
Saccone C Gissi C Lanave C Larizza A Pesole G ampReyes A 2000 Evolution of the mitochondrial genetic sys-tem an overview Gene 261 153ndash159
Salzberg S L White O Peterson J amp Eisen J A 2001Microbial genes in the human genome lateral transfer orgene loss Science 292 1903ndash1906
Sargent F Berks B C amp Palmer T 2002 Assembly of mem-brane-bound respiratory complexes by the Tat protein-trans-port system Arch Microbiol 178 77ndash84
Schatz G 1998 Protein transport the doors to organellesNature 395 439ndash440
Schemidt R A Qu J Williams J R amp Brusilow W S1998 Effects of carbon source on expression of F0 genes andon the stoichiometry of the c subunit in the F1 F0 ATPaseof Escherichia coli J Bacteriol 180 3205ndash3208
Selosse M A Albert B R amp Godelle B 2001 Reducing thegenome size of organelles favours gene transfer to thenucleus Trends Ecol Evol 16 135ndash141
Stern D B amp Lonsdale D M 1982 Mitochondrial and chlo-roplast genomes of maize have a 12- kilobase DNA-sequencein common Nature 299 698ndash702
Stock D Gibbons C Arechaga I Leslie A G W ampWalker J E 2000 The rotary mechanism of ATP synthaseCurr Opin Struct Biol 10 672ndash679
Stoebe B amp Maier U G 2002 One two three naturersquos toolbox for building plastids Protoplasma 219 123ndash130
Stoebe B Martin W amp Kowallik K V 1998 Distributionand nomenclature of protein-coding genes in 12 sequencedchloroplast genomes Plant Mol Biol Rep 16 243ndash255
Stupar R M Lilly J W Town C D Cheng Z Kaul SBuell C R amp Jiang J M 2001 Complex mtDNA consti-tutes an approximate 620 kb insertion on Arabidopsis thali-ana chromosome 2 implication of potential sequencing
Phil Trans R Soc Lond B (2003)
errors caused by large-unit repeats Proc Natl Acad SciUSA 98 5099ndash5103
Surpin M Larkin R M amp Chory J 2002 Signal transduc-tion between the chloroplast and the nucleus Plant Cell 14S327ndashS338
Tjaden J Schwoppe C Mohlmann T Quick P W ampNeuhaus H E 1998 Expression of a plastidic ATPADPtransporter gene in Escherichia coli leads to a functional aden-ine nucleotide transport system in the bacterial cytoplasmicmembrane J Biol Chem 273 9630ndash9636
Tovar J Fischer A amp Clark C G 1999 The mitosome a novelorganelle related to mitochondria in the amitochondrial para-site Entamoeba histolytica Mol Microbiol 32 1013ndash1021
Trebitsh T amp Danon A 2001 Translation of chloroplastpsbA mRNA is regulated by signals initiated by both photos-ystems II and I Proc Natl Acad Sci USA 98 12 289ndash12 294
Tullberg A Alexciev K Pfannschmidt T amp Allen J F2000 Photosynthetic electron flow regulates transcription ofthe psaB gene in pea (Pisum sativum L) chloroplasts throughthe redox state of the plastoquinone pool Plant Cell Physiol41 1045ndash1054
Turmel M Lemieux C Burger G Lang B F Otis CPlante I amp Gray M W 1999 The complete mitochondrialDNA sequences of Nephroselmis olivacea and Pedinomonasminor two radically different evolutionary patterns withingreen algae Plant Cell 11 1717ndash1729
Unden G amp Bongaerts J 1997 Alternative respiratory path-ways of Escherichia coli energetics and transcriptional regu-lation in response to electron acceptors Biochim BiophysActa Bioenerg 1320 217ndash234
Unseld M Marienfeld J R Brandt P amp Brennicke A1997 The mitochondrial genome of Arabidopsis thaliana con-tains 57 genes in 366 924 nucleotides Nature Genet 1557ndash61
Van der Giezen M Slotboom D J Horner D S DyalP L Harding M Xue G P Embley T M amp KunjiE R S 2002 Conserved properties of hydrogenosomal andmitochondrial ADPATP carriers a common origin for bothorganelles EMBO J 21 572ndash579
Von Heijne G 1986 Why mitochondria need a genome FEBSLett 198 1ndash4
Westphal S Soll J amp Vothknecht U C 2001 A vesicle trans-port system inside chloroplasts FEBS Lett 506 257ndash261
Whitehouse H L K 1969 Towards an understanding of themechanism of heredity London Edward Arnold Ltd
Williams B A P Hirt R P Lucocq J M amp Embley T M2002 A mitochondrial remnant in the microsporidian Trachi-pleistophora hominis Nature 418 865ndash869
Wilson R J (and 10 others) 1996a Complete gene map of theplastid-like DNA of the malaria parasite Plasmodium falcipa-rum J Mol Biol 261 155ndash172
Wilson S B Davidson G S Thomson L M amp PearsonC K 1996b Redox control of RNA synthesis in potato mito-chondria Eur J Biochem 242 81ndash85
Winkler H H amp Neuhaus H E 1999 Non-mitochondrialATP transport Trends Biochem Sci 24 64ndash68
Wolfe K H Morden C W amp Palmer J D 1992 Small sin-gle-copy region of plastid DNA in the non-photosyntheticangiosperm Epifagus virginiana contains only two genes Dif-ferences among dicots monocots and bryophytes in geneorganization at a non-bioenergetic locus J Mol Biol 22395ndash104
Wollman F A Minai L amp Nechushtai R 1999 The biogen-esis and assembly of photosynthetic proteins in thylakoidmembranes Biochim Biophys Acta Bioenerg 1411 21ndash85
Yeliseev A A amp Kaplan S 2000 TspO of Rhodobactersphaeroides a structural and functional model for the mam-
Genomes in mitochondria and chloroplasts J F Allen 37
malian peripheral benzodiazepine receptor J Biol Chem275 5657ndash5667
Zerges W 2002 Does complexity constrain organelle evol-ution Trends Plant Sci 7 175ndash182
Zhang Z D Green B R amp Cavalier-Smith T 1999 Singlegene circles in dinoflagellate chloroplast genomes Nature400 155ndash159
Zhang Z D Cavalier-Smith T amp Green B R 2001 A fam-ily of selfish minicircular chromosomes with jumbled chloro-plast gene fragments from a dinoflagellate Mol Biol Evol18 1558ndash1565
Zouni A Witt H T Kern J Fromme P Krauss NSaenger W amp Orth P 2001 Crystal structure of photosys-tem II from Synechococcus elongatus at 38 acngstrom resol-ution Nature 409 739ndash743
DiscussionD S Horner (Dipartimento di Fisiologia e Biochimica
Generali University of Milan Milan Italy) Given thatthere are components of these membrane-bound energy-transduction pathways that are consistently nuclear enco-ded not the key ones you pointed out but others do youexpect perhaps a slower perhaps a more crude but yet aredox communication between the organelle andexpression of nuclear-encoded genes
J F Allen There is and there is no question about itI think that Bogoradrsquos lock-in hypothesis has somethingto say here and there are other people who say it is all todo with assembling multi-subunit complexes If you havecontrol over the initial construction of the core aroundwhich the whole complex is eventually built then redoxcontrol or some sort of control over synthesis of that corewill control construction of the whole thing eventuallyand I think that may well be largely correct I think it ispossible in principle for nuclear genes to make precursorsaccording to redox instructions but it is not really soimportant that those precursors are here and now presentin exactly the right quantities because the precursors canto a limited extent hang around and wait Actually thereis proteolytic degradation of unattached precursors and soon but I think the fact that these chloroplast and mito-chondrially DNA-encoded subunits are at the core ofthese energy-transducing complexes is the crucial issue
T Cavalier-Smith (Department of Zoology University ofOxford Oxford UK ) In the case of mitochondria the sub-set of genes that remains is much more variable than youimplied It runs from three proteins in the minimal casesup to I think over 90 That is a big difference and manyof the genes in different lineages of protists that remainhave nothing to do with redox control they are things likeribosomal proteins and so on So I think that even thoughwhat you say about redox control of certain genes mayactually be an important thing it is not the fundamentaloverall reason why the genomes are maintained It may bepart of the explanation for why certain genes are alwaysincluded in that subset but even in mitochondria I thinkthat cytochrome b is the only one that is always presentSo I think that both this great diversity of what has beenretained in different lineages and the nucleomorph caseindicate that evolution was able to do one thing in onelineage 500 million years ago and never got around todoing it for 500 million years in another lineage so thereis a lot more haphazardness and evolutionary accidentsand contingency than a straight functional interpretation
Phil Trans R Soc Lond B (2003)
there is a lot of just sheer lsquohappenedndashorndashdidnrsquotndashhappenrsquoor carryover
J F Allen I was not implying anything about the rangeof mitochondrial genomes I have a limited amount oftime so I just talked about what I could talk about withinthat limit Of course there will be a range and this willdepend upon the physiology and bioenergetics of theparticular organism But what I am trying to do is to seewhat the common properties of the irreducible core areof components that are always encoded in the organellesand cytochrome b is a perfect example because that reallyis the thing that binds these two all-important quinoneswithout which there is no Q cycle without which there isno proton pumping and bioenergetics and in Plasmodiumwhere the mitochondrion is reduced to this bare mini-mum there is still a mitochondrial gene for cytochromeb Now my argument about the ribosomal proteins andall the rest of it is that in order to be able to exert thisall-important redox control over gene expression of certaincore components you need an in situ genetic system andthis can vary in composition for reasons that I cannotdescribe Maybe you need RNA polymerase or maybeyou can import RNA polymerase In plastids you haveboth of these options You are going to need ribosomestransfer RNAs and the rest of the apparatus of geneexpression and that needs to be in situ but we are stillleft with the question why does there have to be any gen-etic system at all The mature tobacco or spinach chloro-plast seems to have an entire genetic system just to makeone protein D1 which is all it ever needs to resynthesizeI think that this is a clue It is worth having this very unec-onomical crazy system because there is an overridingrequirement to connect electron transfer with geneexpression directly and unambiguouslymdashwithout recourseto a bureaucracy if you like which is what I think thenucleus is
K Willison (Institute of Cancer Research Chester BeattyLaboratories London UK ) These photosynthetic reactioncomplexes are a sort of baroque mixture of polypeptidechains and cofactors and my understanding is that someof them in high photon flux can turn over within say 30minutes so I am wondering if there is a correlationbetween gene retention and the turnover rates of some ofthe components and the complexes it seems to me thatthat could really be a limiting factor and the reason thatthe proteins had to be encoded locally in order to get thecomplexes to be assembled rapidly
J F Allen I think that is right You can only alter thesteady-state concentration of a protein complex by affect-ing the synthesis and breakdown and therefore turnoverThere is an extreme example the D1 protein encoded bythe psbA gene of chloroplasts which is always in the chlor-oplast genome without exception High light will tend todegrade this protein and when the high light goes awaythere is an absolute need very quickly to bring it back upagain I think that is a very good example Whether thisargument applies to genes such as cytochrome oxidase Ido not know
C J Leaver (Department of Plant Sciences University ofOxford Oxford UK ) Your experiments tend to relatemainly to the gene expression side What about proteinRNA turnover the whole concept of redox control ofthatmdashdo we know anything
38 J F Allen Genomes in mitochondria and chloroplasts
J F Allen Well quite a lot is known about chloro-plasts There is redox control of messenger RNA stabilityfor this same D1 protein which has been described byStephen Mayfield and his co-workers The evidence is thatthe site of redox control is ferredoxin via thioredoxin andthe implication is that thioredoxin tells whether non-cyclicelectron transport is working at all When it is workingthere is a signal that says lsquoOK now continue with all theprotein synthesisrsquo It is not particularly specific but moreof a global effect I think this is what Mayfield argues butof course this is not inconsistent with my argument I haveconcentrated on quinone pools and medium-potentialredox regulatory control because that is where I can seesignals about the dangerous consequences of getting itwrong but there is control of other levels of geneexpression In mitochondria I am not so sure I wouldbounce the question back to you on that one
W Martin (Institute of Botany III Heinrich-Heine Univ-ersitat Dusseldorf Dusseldorf Germany) The nucleomorphgenome was cited as evidence in conflict with your modelbut is that true The nucleomorph genome is not in abioenergetic organelle in that sense It resides in the cyto-sol so does the size of the nucleomorph genome workagainst your model Is it really evidence against yourmodel
J F Allen I do not know why should it be evidenceI cannot explain why the nucleomorph has a genomeThat is why I clutched at the straw that it is storage fornucleic acid precursors
Phil Trans R Soc Lond B (2003)
T Cavalier-Smith The reason I introduced the nucleo-morph is not to say it is direct evidence against yourhypothesis but to show that there probably was no singlefunctional reason for the retention of every gene in everyorganelle It may be that there are specific physiologicalreasons for the retention of certain genes and thereforeyou might be right about a subset of genes but I verymuch doubt if there are specific physiological reasons forthe retention of every single gene that is retained differen-tially because odd things happen in evolution Somethings are easy and sometimes things are difficult Evol-ution is history
J F Allen Of course I agree with that I do not thinkI can explain everything but I do think I can explainsomething If I have done that then I am content
GLOSSARY
AAC ADPndashATP carrierCES control by epistasy of synthesisCORR co-location for redox regulationLHC light-harvesting complexPCC Pasteur culture collectionORF open reading frameRUBISCO ribulose-15-bisphosphate carboxylasendashoxygenaseTAT twin-arginine translocase
Visit httpwwwpubsroyalsocacuk to see an electronic appendix tothis paper

34 J F Allen Genomes in mitochondria and chloroplasts
Delwiche C F 1999 Tracing the thread of plastid diversitythrough the tapestry of life Am Nat 154 S164ndashS177
Doolittle W F 1999 Phylogenetic classification and the uni-versal tree Science 284 2124ndash2129
Douglas S E 1998 Plastid evolution origins diversitytrends Curr Opin Genet Dev 8 655ndash661
Duby G amp Boutry M 2002 Mitochondrial protein importmachinery and targeting information Plant Sci 162 477ndash490
Eberhard S Drapier D amp Wollman F A 2002 Searchinglimiting steps in the expression of chloroplast encoded pro-teins relations between gene copy number transcriptiontranscript abundance and translation rate in the chloroplastof Chlamydomonas reinhardtii Plant J 31 149ndash160
El Bissati K amp Kirilovsky D 2001 Regulation of psbA andpsaE expression by light quality in Synechocystis species PCC6803 A redox control mechanism Plant Physiol 1251988ndash2000
Ellis R J 1984 The nuclear domination of chloroplast devel-opment Science Progress 69 129ndash142
Embley T M amp Martin W 1998 Molecular evolution ahydrogen-producing mitochondrion Nature 396 517ndash519
Ems S C Morden C W Dixon C K Wolfe K HdePamphilis C W amp Palmer J D 1995 Transcriptionsplicing and editing of plastid RNAs in the non-photosyn-thetic plant Epifagus virginiana Plant Mol Biol 29 721ndash733
Eraso J M amp Kaplan S 2000 From redox flow to gene regu-lation role of the PrrC protein of Rhodobacter sphaeroides241 Biochemistry 39 2052ndash2062
Eraso J M amp Kaplan S 2002 Redox flow as an instrumentof gene regulation Meth Enzymol 348 216ndash229
Escoubas J M Lomas M Laroche J amp Falkowski P G1995 Light-intensity regulation of cab gene-transcription issignaled by the redox state of the plastoquinone pool ProcNatl Acad Sci USA 92 10 237ndash10 241
Ferea T L Botstein D Brown P O amp Rosenzweig R F1999 Systematic changes in gene expression patterns follow-ing adaptive evolution in yeast Proc Natl Acad Sci USA96 9721ndash9726
Forsberg J Rosenquist M Fraysse L amp Allen J F 2001Redox signalling in chloroplasts and mitochondria genomicand biochemical evidence for two-component regulatory sys-tems in bioenergetic organelles Biochem Soc Trans 29403ndash407
Fujita Y 1997 A study on the dynamic features of photosys-tem stoichiometry accomplishments and problems forfuture studies Photosynth Res 53 83ndash93
Gabriel K Buchanan S K amp Lithgow T 2001 The alphaand the beta protein translocation across mitochondrial andplastid outer membranes Trends Biochem Sci 26 36ndash40
Galvis M L E Allen J F amp Hakansson G 1998 Proteinsynthesis by isolated pea mitochondria is dependent on theactivity of respiratory complex II Curr Genet 33 320ndash329
Gatenby A A amp Ellis R J 1990 Chaperone function theassembly of ribulose bisphosphate carboxylase-oxygenase ARev Cell Biol 6 125ndash149
GoldschmidtClermont M 1998 Coordination of nuclear andchloroplast gene expression in plant cells Int Rev CytolSurv Cell Biol 177 115ndash180
Gray M W 1992 The endosymbiont hypothesis revisited IntRev Cytol 141 233ndash357
Gray M W 1999 Evolution of organellar genomes CurrOpin Genet Dev 9 678ndash687
Gray M W 2000 Evolutionary biology mitochondrial geneson the move Nature 408 302ndash305
Gray M W amp Doolittle W F 1982 Has the endosymbionthypothesis been proven Microbiol Rev 46 1ndash42
Gray M W Burger G amp Lang B F 1999 Mitochondrialevolution Science 283 1476ndash1481
Phil Trans R Soc Lond B (2003)
Grebe T W amp Stock J B 1999 The histidine protein kinasesuperfamily Adv Microb Physiol 41 139ndash227
Green B R amp Kuhlbrandt W 1995 Sequence conservationof light-harvesting and stress-response proteins in relation tothe 3-dimensional molecular structure of Lhcii PhotosynthRes 44 139ndash148
Gruissem W amp Tonkyn J C 1993 Control mechanisms ofplastid gene expression Crit Rev Plant Sci 12 19ndash55
Hackstein J H P (and 11 others) 2001 Hydrogenosomesconvergent adaptations of mitochondria to anaerobicenvironments Zool Anal Complex Syst 104 290ndash302
Harrison M A Keen J N Findlay J B C amp Allen J F1990 Modification of a Glnb-like gene-product by photosyn-thetic electron-transport in the cyanobacterium Synechoc-occus 6301 FEBS Lett 264 25ndash28
Harvey A J Kind K L amp Thompson J G 2002 Redoxregulation of early embryo development Reproduction 123479ndash486
Heathcote P Fyfe P K amp Jones M R 2002 Reactioncentres the structure and evolution of biological solarpower Trends Biochem Sci 27 79ndash87
Henze K amp Martin W 2001 How do mitochondrial genesget into the nucleus Trends Genet 17 383ndash387
Herrmann R G amp Westhoff P 2001 Thylakoid biogenesis anddynamics the result of a complex phylogenetic puzzle InRegulation of photosynthesis vol 11 (ed E-M Aro amp BAndersson) pp 1ndash28 Dordrecht The Netherlands Kluwer
Hihara Y Sonoike K Kanehisa M amp Ikeuchi M 2002Search for redox responsive genes in Synechocystis sp PCC6803 by using DNA microarrays Plant Cell Physiol 43S178
Howe C J Barbrook A C amp Lockhart P J 2000 Organellegenes do they jump or are they pushed Trends Genet 1665ndash66
Ikeuchi M 2002 Gene expression and responses to light stressin cyanobacteria Plant Cell Physiol 43 S21
Iuchi S amp Lin E C C 1992 Mutational analysis of signaltransduction by Arcb a membrane sensor protein respon-sible for anaerobic repression of operons involved in the cen-tral aerobic pathways in Escherichia coli J Bacteriol 1743972ndash3980
Jackson-Constan D Akita M amp Keegstra K 2001 Molecu-lar chaperones involved in chloroplast protein importBiochim Biophys Acta Mol Cell Res 1541 102ndash113
Jarvis P amp Soll M 2001 Toc Tic and chloroplast proteinimport Biochim Biophys Acta Mol Cell Res 1541 64ndash79
Jordan P Fromme P Witt H T Klukas O SaengerW amp Krauss N 2001 Three-dimensional structure ofcyanobacterial photosystem I at 25 acngstrom resolutionNature 411 909ndash917
Kaneko T (and 23 others) 1996a Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regionsDNA Res 3 109ndash136
Kaneko T (and 23 others) 1996b Sequence analysis of thegenome of the unicellular cyanobacterium Synechocystis spstrain PCC6803 II Sequence determination of the entiregenome and assignment of potential protein-coding regions[Supplement] DNA Res 3 185ndash209
Kanevski I amp Maliga P 1994 Relocation of the plastid rbcLgene to the nucleus yields functional ribulose-15-bisphosph-ate carboxylase in tobacco chloroplasts Proc Natl Acad SciUSA 91 1969ndash1973
Kim J amp Mayfield S P 1997 Protein disulfide isomerase as aregulator of chloroplast translational activation Science 2781954ndash1957
Genomes in mitochondria and chloroplasts J F Allen 35
Kirk J T O amp Tilney-Bassett R A E 1978 The plastidsTheir chemistry structure growth and inheritance Amster-dam Elsevier
Koehler C M Merchant S amp Schatz G 1999 How mem-brane proteins travel across the mitochondrial intermem-brane space Trends Biochem Sci 24 428ndash432
Komiya T Rospert S Schatz G amp Mihara K 1997 Bind-ing of mitochondrial precursor proteins to the cytoplasmicdomains of the import receptors Tom70 and Tom20 isdetermined by cytoplasmic chaperones FASEB J 11 1196
Konstantinov Y M Lutsenko G N amp Podsosonny V A1995 Genetic functions of isolated maize mitochondriaunder model changes of redox conditions Biochem MolBiol Int 36 319ndash326
Korab-Laskowska M Rioux P Brossard N LittlejohnT G Gray M W Lang B F amp Burger G 1998 Theorganelle genome database project (GOBASE) Nucleic AcidsRes 26 138ndash144
Kovacs L Wiessner W Kis M Nagy F Mende D ampDemeter S 2000 Short- and long-term redox regulation ofphotosynthetic light energy distribution and photosystemstoichiometry by acetate metabolism in the green alga Chla-mydobotrys stellata Photosynth Res 65 231ndash247
Kuhlbrandt W 1994 Structure and function of the plant light-harvesting complex LHC-II Curr Opin Struct Biol 4519ndash528
Lang B F Gray M W amp Burger G 1999 Mitochondrialgenome evolution and the origin of eukaryotes A RevGenet 33 351ndash397
Lemire B D Fankhauser C Baker A amp Schatz G 1989The mitochondrial targeting function of randomly generatedpeptide sequences correlates with predicted helical amphi-philicity J Biol Chem 264 20 206ndash20 215
Leuenberger D Bally N A Schatz G amp Koehler C M1999 Different import pathways through the mitochondrialintermembrane space for inner membrane proteins EMBOJ 18 4816ndash4822
Li H amp Sherman L A 2000 A redox-responsive regulator ofphotosynthesis gene expression in the cyanobacterium Syne-chocystis sp strain PCC 6803 J Bacteriol 182 4268ndash4277
Lin X Y (and 36 others) 1999 Sequence and analysis of chro-mosome 2 of the plant Arabidopsis thaliana Nature 402761ndash768
Lonsdale D M Hodge T P Howe C J amp Stern D B1983 Maize mitochondrial-DNA contains a sequence hom-ologous to the ribulose-15-bisphosphate carboxylase largesubunit gene of chloroplast DNA Cell 34 1007ndash1014
McFadden G I 1999 Endosymbiosis and evolution of theplant cell Curr Opin Plant Biol 2 513ndash519
McFadden G I 2001 Chloroplast origin and integrationPlant Physiol 125 50ndash53
Mahler H R amp Raff R A 1975 The evolutionary origin ofthe mitochondrion a nonsymbiotic model Int Rev Cytol43 1ndash124
Manfredi G Fu J Ojaimi J Sadlock J E Kwong J QGuy J amp Schon E A 2002 Rescue of a deficiency in ATPsynthesis by transfer of MTATP6 a mitochondrial DNA-encoded gene to the nucleus Nature Genet 30 394ndash399
Marc P Margeot A Devaux F Blugeon C Corral-Debrinski M amp Jacq C 2002 Genome-wide analysis ofmRNAs targeted to yeast mitochondria EMBO Rep 3159ndash164
Marienfeld J Unseld M amp Brennicke A 1999 The mito-chondrial genome of Arabidopsis is composed of both nativeand immigrant information Trends Plant Sci 4 495ndash502
Martin W 1999 A briefly argued case that mitochondria andplastids are descendants of endosymbionts but that thenuclear compartment is not Proc R Soc Lond B 2661387ndash1395 (DOI 101098rspb19990792)
Phil Trans R Soc Lond B (2003)
Martin W amp Herrmann R G 1998 Gene transfer fromorganelles to the nucleus how much what happens andwhy Plant Physiol 118 9ndash17
Martin W amp Muller M 1998 The hydrogen hypothesis forthe first eukaryote Nature 392 37ndash41
Martin W amp Russell M J 2003 On the origins of cells ahypothesis for the evolutionary transitions from abiotic geo-chemistry to chemoautotrophic prokaryotes and from pro-karyotes to nucleated cells Phil Trans R Soc Lond B 35859ndash85 (DOI 101098rstb20021183)
Martin W Stoebe B Goremykin V Hansmann S Hase-gawa M amp Kowallik K V 1998 Gene transfer to thenucleus and the evolution of chloroplasts Nature 393162ndash165
Martin W Hoffmeister M Rotte C amp Henze K 2001 Anoverview of endosymbiotic models for the origins of eukary-otes their ATP-producing organelles (mitochondria andhydrogenosomes) and their heterotrophic lifestyle BiolChem 382 1521ndash1539
Martin W Rujan T Richly E Hansen A Cornelsen SLins T Leister D Stoebe B Hasegawa M amp PennyD 2002 Evolutionary analysis of Arabidopsis cyanobacterialand chloroplast genomes reveals plastid phylogeny and thou-sands of cyanobacterial genes in the nucleus Proc NatlAcad Sci USA 99 12 246ndash12 251
Millen R S (and 12 others) 2001 Many parallel losses of infAfrom chloroplast DNA during angiosperm evolution withmultiple independent transfers to the nucleus Plant Cell 13645ndash658
Morse D Salois P Markovic P amp Hastings J W 1995 Anuclear-encoded form-II RUBISCO in dinoflagellatesScience 268 1622ndash1624
Murakami A Kim S J amp Fujita Y 1997 Changes in photo-system stoichiometry in response to environmental con-ditions for cell growth observed with the cyanophyteSynechocystis PCC 6714 Plant Cell Physiol 38392ndash397
Oh J I amp Kaplan S 2000 Redox signaling globalization ofgene expression EMBO J 19 4237ndash4247
Ozawa T 1995 Mitochondrial-DNA mutations associatedwith aging and degenerative diseases Exp Gerontol 30269ndash290
Palmer J D 1997 Organelle genomes going going goneScience 275 790ndash791
Pearson C K Wilson S B Schaffer R amp Ross A W 1993NAD turnover and utilization of metabolites for RNA syn-thesis in a reaction sensing the redox state of the cyto-chrome-b( 6 )f complex in isolated chloroplasts Eur JBiochem 218 397ndash404
Pfanner N amp Geissler A 2001 Versatility of the mitochon-drial protein import machinery Natl Rev Mol Cell Biol 2339ndash349
Pfanner N amp Wiedemann N 2002 Mitochondrial proteinimport two membranes three translocases Curr Opin CellBiol 14 400ndash411
Pfannschmidt T Nilsson A amp Allen J F 1999a Photosyn-thetic control of chloroplast gene expression Nature 397625ndash628
Pfannschmidt T Nilsson A Tullberg A Link G amp AllenJ F 1999b Direct transcriptional control of the chloroplastgenes psbA and psaAB adjusts photosynthesis to light energydistribution in plants IUBMB Life 48 271ndash276
Pfannschmidt T Allen J F amp Oelmuller R 2001a Prin-ciples of redox control in photosynthesis gene expressionPhysiol Plant 112 1ndash9
Pfannschmidt T Schutze K Brost M amp Oelmuller R2001b A novel mechanism of nuclear photosynthesis generegulation by redox signals from the chloroplast during pho-
36 J F Allen Genomes in mitochondria and chloroplasts
tosystem stoichiometry adjustment J Biol Chem 27636 125ndash36 130
Popot J L amp de Vitry C 1990 On the microassembly of inte-gral membrane proteins A Rev Biophys Biophys Chem 19369ndash403
Race H L Herrmann R G amp Martin W 1999 Why haveorganelles retained genomes Trends Genet 15 364ndash370
Raven J A Johnston A M Parsons R amp Kubler J 1994aThe influence of natural and experimental high O2 concen-trations on O2 evolving phototrophs Biol Rev CambridgePhil Soc 69 61ndash94
Raven J A Johnston A M Parsons R amp Kubler J 1994bThe occurrence and influence on photolithotrophs of highoxygen concentrations Proc R Soc Edinb B 102 193ndash201
Robinson C 2000 The twin-arginine translocation system anovel means of transporting folded proteins in chloroplastsand bacteria Biol Chem 381 89ndash93
Robinson C amp Bolhuis A 2001 Protein targeting by the twin-arginine translocation pathway Natl Rev Mol Cell Biol 2350ndash356
Rodermel S 2001 Pathways of plastid-to-nucleus signalingTrends Plant Sci 6 471ndash478
Roger A J 1999 Reconstructing early events in eukaryoticevolution Am Nat 154 S146ndashS163
Roger A J Svard S G Tovar J Clark C G SmithM W Gillin F D amp Sogin M L 1998 A mitochondrial-like chaperonin 60 gene in Giardia lamblia evidence thatdiplomonads once harbored an endosymbiont related to theprogenitor of mitochondria Proc Natl Acad Sci USA 95229ndash234
Roodyn D B amp Wilkie D 1968 The biogenesis of mitochondriaMethuenrsquos monographs on biological subjects London Methuen
Rujan T amp Martin W 2001 How many genes in Arabidopsiscome from cyanobacteria An estimate from 386 proteinphylogenies Trends Genet 17 113ndash120
Saccone C Gissi C Lanave C Larizza A Pesole G ampReyes A 2000 Evolution of the mitochondrial genetic sys-tem an overview Gene 261 153ndash159
Salzberg S L White O Peterson J amp Eisen J A 2001Microbial genes in the human genome lateral transfer orgene loss Science 292 1903ndash1906
Sargent F Berks B C amp Palmer T 2002 Assembly of mem-brane-bound respiratory complexes by the Tat protein-trans-port system Arch Microbiol 178 77ndash84
Schatz G 1998 Protein transport the doors to organellesNature 395 439ndash440
Schemidt R A Qu J Williams J R amp Brusilow W S1998 Effects of carbon source on expression of F0 genes andon the stoichiometry of the c subunit in the F1 F0 ATPaseof Escherichia coli J Bacteriol 180 3205ndash3208
Selosse M A Albert B R amp Godelle B 2001 Reducing thegenome size of organelles favours gene transfer to thenucleus Trends Ecol Evol 16 135ndash141
Stern D B amp Lonsdale D M 1982 Mitochondrial and chlo-roplast genomes of maize have a 12- kilobase DNA-sequencein common Nature 299 698ndash702
Stock D Gibbons C Arechaga I Leslie A G W ampWalker J E 2000 The rotary mechanism of ATP synthaseCurr Opin Struct Biol 10 672ndash679
Stoebe B amp Maier U G 2002 One two three naturersquos toolbox for building plastids Protoplasma 219 123ndash130
Stoebe B Martin W amp Kowallik K V 1998 Distributionand nomenclature of protein-coding genes in 12 sequencedchloroplast genomes Plant Mol Biol Rep 16 243ndash255
Stupar R M Lilly J W Town C D Cheng Z Kaul SBuell C R amp Jiang J M 2001 Complex mtDNA consti-tutes an approximate 620 kb insertion on Arabidopsis thali-ana chromosome 2 implication of potential sequencing
Phil Trans R Soc Lond B (2003)
errors caused by large-unit repeats Proc Natl Acad SciUSA 98 5099ndash5103
Surpin M Larkin R M amp Chory J 2002 Signal transduc-tion between the chloroplast and the nucleus Plant Cell 14S327ndashS338
Tjaden J Schwoppe C Mohlmann T Quick P W ampNeuhaus H E 1998 Expression of a plastidic ATPADPtransporter gene in Escherichia coli leads to a functional aden-ine nucleotide transport system in the bacterial cytoplasmicmembrane J Biol Chem 273 9630ndash9636
Tovar J Fischer A amp Clark C G 1999 The mitosome a novelorganelle related to mitochondria in the amitochondrial para-site Entamoeba histolytica Mol Microbiol 32 1013ndash1021
Trebitsh T amp Danon A 2001 Translation of chloroplastpsbA mRNA is regulated by signals initiated by both photos-ystems II and I Proc Natl Acad Sci USA 98 12 289ndash12 294
Tullberg A Alexciev K Pfannschmidt T amp Allen J F2000 Photosynthetic electron flow regulates transcription ofthe psaB gene in pea (Pisum sativum L) chloroplasts throughthe redox state of the plastoquinone pool Plant Cell Physiol41 1045ndash1054
Turmel M Lemieux C Burger G Lang B F Otis CPlante I amp Gray M W 1999 The complete mitochondrialDNA sequences of Nephroselmis olivacea and Pedinomonasminor two radically different evolutionary patterns withingreen algae Plant Cell 11 1717ndash1729
Unden G amp Bongaerts J 1997 Alternative respiratory path-ways of Escherichia coli energetics and transcriptional regu-lation in response to electron acceptors Biochim BiophysActa Bioenerg 1320 217ndash234
Unseld M Marienfeld J R Brandt P amp Brennicke A1997 The mitochondrial genome of Arabidopsis thaliana con-tains 57 genes in 366 924 nucleotides Nature Genet 1557ndash61
Van der Giezen M Slotboom D J Horner D S DyalP L Harding M Xue G P Embley T M amp KunjiE R S 2002 Conserved properties of hydrogenosomal andmitochondrial ADPATP carriers a common origin for bothorganelles EMBO J 21 572ndash579
Von Heijne G 1986 Why mitochondria need a genome FEBSLett 198 1ndash4
Westphal S Soll J amp Vothknecht U C 2001 A vesicle trans-port system inside chloroplasts FEBS Lett 506 257ndash261
Whitehouse H L K 1969 Towards an understanding of themechanism of heredity London Edward Arnold Ltd
Williams B A P Hirt R P Lucocq J M amp Embley T M2002 A mitochondrial remnant in the microsporidian Trachi-pleistophora hominis Nature 418 865ndash869
Wilson R J (and 10 others) 1996a Complete gene map of theplastid-like DNA of the malaria parasite Plasmodium falcipa-rum J Mol Biol 261 155ndash172
Wilson S B Davidson G S Thomson L M amp PearsonC K 1996b Redox control of RNA synthesis in potato mito-chondria Eur J Biochem 242 81ndash85
Winkler H H amp Neuhaus H E 1999 Non-mitochondrialATP transport Trends Biochem Sci 24 64ndash68
Wolfe K H Morden C W amp Palmer J D 1992 Small sin-gle-copy region of plastid DNA in the non-photosyntheticangiosperm Epifagus virginiana contains only two genes Dif-ferences among dicots monocots and bryophytes in geneorganization at a non-bioenergetic locus J Mol Biol 22395ndash104
Wollman F A Minai L amp Nechushtai R 1999 The biogen-esis and assembly of photosynthetic proteins in thylakoidmembranes Biochim Biophys Acta Bioenerg 1411 21ndash85
Yeliseev A A amp Kaplan S 2000 TspO of Rhodobactersphaeroides a structural and functional model for the mam-
Genomes in mitochondria and chloroplasts J F Allen 37
malian peripheral benzodiazepine receptor J Biol Chem275 5657ndash5667
Zerges W 2002 Does complexity constrain organelle evol-ution Trends Plant Sci 7 175ndash182
Zhang Z D Green B R amp Cavalier-Smith T 1999 Singlegene circles in dinoflagellate chloroplast genomes Nature400 155ndash159
Zhang Z D Cavalier-Smith T amp Green B R 2001 A fam-ily of selfish minicircular chromosomes with jumbled chloro-plast gene fragments from a dinoflagellate Mol Biol Evol18 1558ndash1565
Zouni A Witt H T Kern J Fromme P Krauss NSaenger W amp Orth P 2001 Crystal structure of photosys-tem II from Synechococcus elongatus at 38 acngstrom resol-ution Nature 409 739ndash743
DiscussionD S Horner (Dipartimento di Fisiologia e Biochimica
Generali University of Milan Milan Italy) Given thatthere are components of these membrane-bound energy-transduction pathways that are consistently nuclear enco-ded not the key ones you pointed out but others do youexpect perhaps a slower perhaps a more crude but yet aredox communication between the organelle andexpression of nuclear-encoded genes
J F Allen There is and there is no question about itI think that Bogoradrsquos lock-in hypothesis has somethingto say here and there are other people who say it is all todo with assembling multi-subunit complexes If you havecontrol over the initial construction of the core aroundwhich the whole complex is eventually built then redoxcontrol or some sort of control over synthesis of that corewill control construction of the whole thing eventuallyand I think that may well be largely correct I think it ispossible in principle for nuclear genes to make precursorsaccording to redox instructions but it is not really soimportant that those precursors are here and now presentin exactly the right quantities because the precursors canto a limited extent hang around and wait Actually thereis proteolytic degradation of unattached precursors and soon but I think the fact that these chloroplast and mito-chondrially DNA-encoded subunits are at the core ofthese energy-transducing complexes is the crucial issue
T Cavalier-Smith (Department of Zoology University ofOxford Oxford UK ) In the case of mitochondria the sub-set of genes that remains is much more variable than youimplied It runs from three proteins in the minimal casesup to I think over 90 That is a big difference and manyof the genes in different lineages of protists that remainhave nothing to do with redox control they are things likeribosomal proteins and so on So I think that even thoughwhat you say about redox control of certain genes mayactually be an important thing it is not the fundamentaloverall reason why the genomes are maintained It may bepart of the explanation for why certain genes are alwaysincluded in that subset but even in mitochondria I thinkthat cytochrome b is the only one that is always presentSo I think that both this great diversity of what has beenretained in different lineages and the nucleomorph caseindicate that evolution was able to do one thing in onelineage 500 million years ago and never got around todoing it for 500 million years in another lineage so thereis a lot more haphazardness and evolutionary accidentsand contingency than a straight functional interpretation
Phil Trans R Soc Lond B (2003)
there is a lot of just sheer lsquohappenedndashorndashdidnrsquotndashhappenrsquoor carryover
J F Allen I was not implying anything about the rangeof mitochondrial genomes I have a limited amount oftime so I just talked about what I could talk about withinthat limit Of course there will be a range and this willdepend upon the physiology and bioenergetics of theparticular organism But what I am trying to do is to seewhat the common properties of the irreducible core areof components that are always encoded in the organellesand cytochrome b is a perfect example because that reallyis the thing that binds these two all-important quinoneswithout which there is no Q cycle without which there isno proton pumping and bioenergetics and in Plasmodiumwhere the mitochondrion is reduced to this bare mini-mum there is still a mitochondrial gene for cytochromeb Now my argument about the ribosomal proteins andall the rest of it is that in order to be able to exert thisall-important redox control over gene expression of certaincore components you need an in situ genetic system andthis can vary in composition for reasons that I cannotdescribe Maybe you need RNA polymerase or maybeyou can import RNA polymerase In plastids you haveboth of these options You are going to need ribosomestransfer RNAs and the rest of the apparatus of geneexpression and that needs to be in situ but we are stillleft with the question why does there have to be any gen-etic system at all The mature tobacco or spinach chloro-plast seems to have an entire genetic system just to makeone protein D1 which is all it ever needs to resynthesizeI think that this is a clue It is worth having this very unec-onomical crazy system because there is an overridingrequirement to connect electron transfer with geneexpression directly and unambiguouslymdashwithout recourseto a bureaucracy if you like which is what I think thenucleus is
K Willison (Institute of Cancer Research Chester BeattyLaboratories London UK ) These photosynthetic reactioncomplexes are a sort of baroque mixture of polypeptidechains and cofactors and my understanding is that someof them in high photon flux can turn over within say 30minutes so I am wondering if there is a correlationbetween gene retention and the turnover rates of some ofthe components and the complexes it seems to me thatthat could really be a limiting factor and the reason thatthe proteins had to be encoded locally in order to get thecomplexes to be assembled rapidly
J F Allen I think that is right You can only alter thesteady-state concentration of a protein complex by affect-ing the synthesis and breakdown and therefore turnoverThere is an extreme example the D1 protein encoded bythe psbA gene of chloroplasts which is always in the chlor-oplast genome without exception High light will tend todegrade this protein and when the high light goes awaythere is an absolute need very quickly to bring it back upagain I think that is a very good example Whether thisargument applies to genes such as cytochrome oxidase Ido not know
C J Leaver (Department of Plant Sciences University ofOxford Oxford UK ) Your experiments tend to relatemainly to the gene expression side What about proteinRNA turnover the whole concept of redox control ofthatmdashdo we know anything
38 J F Allen Genomes in mitochondria and chloroplasts
J F Allen Well quite a lot is known about chloro-plasts There is redox control of messenger RNA stabilityfor this same D1 protein which has been described byStephen Mayfield and his co-workers The evidence is thatthe site of redox control is ferredoxin via thioredoxin andthe implication is that thioredoxin tells whether non-cyclicelectron transport is working at all When it is workingthere is a signal that says lsquoOK now continue with all theprotein synthesisrsquo It is not particularly specific but moreof a global effect I think this is what Mayfield argues butof course this is not inconsistent with my argument I haveconcentrated on quinone pools and medium-potentialredox regulatory control because that is where I can seesignals about the dangerous consequences of getting itwrong but there is control of other levels of geneexpression In mitochondria I am not so sure I wouldbounce the question back to you on that one
W Martin (Institute of Botany III Heinrich-Heine Univ-ersitat Dusseldorf Dusseldorf Germany) The nucleomorphgenome was cited as evidence in conflict with your modelbut is that true The nucleomorph genome is not in abioenergetic organelle in that sense It resides in the cyto-sol so does the size of the nucleomorph genome workagainst your model Is it really evidence against yourmodel
J F Allen I do not know why should it be evidenceI cannot explain why the nucleomorph has a genomeThat is why I clutched at the straw that it is storage fornucleic acid precursors
Phil Trans R Soc Lond B (2003)
T Cavalier-Smith The reason I introduced the nucleo-morph is not to say it is direct evidence against yourhypothesis but to show that there probably was no singlefunctional reason for the retention of every gene in everyorganelle It may be that there are specific physiologicalreasons for the retention of certain genes and thereforeyou might be right about a subset of genes but I verymuch doubt if there are specific physiological reasons forthe retention of every single gene that is retained differen-tially because odd things happen in evolution Somethings are easy and sometimes things are difficult Evol-ution is history
J F Allen Of course I agree with that I do not thinkI can explain everything but I do think I can explainsomething If I have done that then I am content
GLOSSARY
AAC ADPndashATP carrierCES control by epistasy of synthesisCORR co-location for redox regulationLHC light-harvesting complexPCC Pasteur culture collectionORF open reading frameRUBISCO ribulose-15-bisphosphate carboxylasendashoxygenaseTAT twin-arginine translocase
Visit httpwwwpubsroyalsocacuk to see an electronic appendix tothis paper

Genomes in mitochondria and chloroplasts J F Allen 35
Kirk J T O amp Tilney-Bassett R A E 1978 The plastidsTheir chemistry structure growth and inheritance Amster-dam Elsevier
Koehler C M Merchant S amp Schatz G 1999 How mem-brane proteins travel across the mitochondrial intermem-brane space Trends Biochem Sci 24 428ndash432
Komiya T Rospert S Schatz G amp Mihara K 1997 Bind-ing of mitochondrial precursor proteins to the cytoplasmicdomains of the import receptors Tom70 and Tom20 isdetermined by cytoplasmic chaperones FASEB J 11 1196
Konstantinov Y M Lutsenko G N amp Podsosonny V A1995 Genetic functions of isolated maize mitochondriaunder model changes of redox conditions Biochem MolBiol Int 36 319ndash326
Korab-Laskowska M Rioux P Brossard N LittlejohnT G Gray M W Lang B F amp Burger G 1998 Theorganelle genome database project (GOBASE) Nucleic AcidsRes 26 138ndash144
Kovacs L Wiessner W Kis M Nagy F Mende D ampDemeter S 2000 Short- and long-term redox regulation ofphotosynthetic light energy distribution and photosystemstoichiometry by acetate metabolism in the green alga Chla-mydobotrys stellata Photosynth Res 65 231ndash247
Kuhlbrandt W 1994 Structure and function of the plant light-harvesting complex LHC-II Curr Opin Struct Biol 4519ndash528
Lang B F Gray M W amp Burger G 1999 Mitochondrialgenome evolution and the origin of eukaryotes A RevGenet 33 351ndash397
Lemire B D Fankhauser C Baker A amp Schatz G 1989The mitochondrial targeting function of randomly generatedpeptide sequences correlates with predicted helical amphi-philicity J Biol Chem 264 20 206ndash20 215
Leuenberger D Bally N A Schatz G amp Koehler C M1999 Different import pathways through the mitochondrialintermembrane space for inner membrane proteins EMBOJ 18 4816ndash4822
Li H amp Sherman L A 2000 A redox-responsive regulator ofphotosynthesis gene expression in the cyanobacterium Syne-chocystis sp strain PCC 6803 J Bacteriol 182 4268ndash4277
Lin X Y (and 36 others) 1999 Sequence and analysis of chro-mosome 2 of the plant Arabidopsis thaliana Nature 402761ndash768
Lonsdale D M Hodge T P Howe C J amp Stern D B1983 Maize mitochondrial-DNA contains a sequence hom-ologous to the ribulose-15-bisphosphate carboxylase largesubunit gene of chloroplast DNA Cell 34 1007ndash1014
McFadden G I 1999 Endosymbiosis and evolution of theplant cell Curr Opin Plant Biol 2 513ndash519
McFadden G I 2001 Chloroplast origin and integrationPlant Physiol 125 50ndash53
Mahler H R amp Raff R A 1975 The evolutionary origin ofthe mitochondrion a nonsymbiotic model Int Rev Cytol43 1ndash124
Manfredi G Fu J Ojaimi J Sadlock J E Kwong J QGuy J amp Schon E A 2002 Rescue of a deficiency in ATPsynthesis by transfer of MTATP6 a mitochondrial DNA-encoded gene to the nucleus Nature Genet 30 394ndash399
Marc P Margeot A Devaux F Blugeon C Corral-Debrinski M amp Jacq C 2002 Genome-wide analysis ofmRNAs targeted to yeast mitochondria EMBO Rep 3159ndash164
Marienfeld J Unseld M amp Brennicke A 1999 The mito-chondrial genome of Arabidopsis is composed of both nativeand immigrant information Trends Plant Sci 4 495ndash502
Martin W 1999 A briefly argued case that mitochondria andplastids are descendants of endosymbionts but that thenuclear compartment is not Proc R Soc Lond B 2661387ndash1395 (DOI 101098rspb19990792)
Phil Trans R Soc Lond B (2003)
Martin W amp Herrmann R G 1998 Gene transfer fromorganelles to the nucleus how much what happens andwhy Plant Physiol 118 9ndash17
Martin W amp Muller M 1998 The hydrogen hypothesis forthe first eukaryote Nature 392 37ndash41
Martin W amp Russell M J 2003 On the origins of cells ahypothesis for the evolutionary transitions from abiotic geo-chemistry to chemoautotrophic prokaryotes and from pro-karyotes to nucleated cells Phil Trans R Soc Lond B 35859ndash85 (DOI 101098rstb20021183)
Martin W Stoebe B Goremykin V Hansmann S Hase-gawa M amp Kowallik K V 1998 Gene transfer to thenucleus and the evolution of chloroplasts Nature 393162ndash165
Martin W Hoffmeister M Rotte C amp Henze K 2001 Anoverview of endosymbiotic models for the origins of eukary-otes their ATP-producing organelles (mitochondria andhydrogenosomes) and their heterotrophic lifestyle BiolChem 382 1521ndash1539
Martin W Rujan T Richly E Hansen A Cornelsen SLins T Leister D Stoebe B Hasegawa M amp PennyD 2002 Evolutionary analysis of Arabidopsis cyanobacterialand chloroplast genomes reveals plastid phylogeny and thou-sands of cyanobacterial genes in the nucleus Proc NatlAcad Sci USA 99 12 246ndash12 251
Millen R S (and 12 others) 2001 Many parallel losses of infAfrom chloroplast DNA during angiosperm evolution withmultiple independent transfers to the nucleus Plant Cell 13645ndash658
Morse D Salois P Markovic P amp Hastings J W 1995 Anuclear-encoded form-II RUBISCO in dinoflagellatesScience 268 1622ndash1624
Murakami A Kim S J amp Fujita Y 1997 Changes in photo-system stoichiometry in response to environmental con-ditions for cell growth observed with the cyanophyteSynechocystis PCC 6714 Plant Cell Physiol 38392ndash397
Oh J I amp Kaplan S 2000 Redox signaling globalization ofgene expression EMBO J 19 4237ndash4247
Ozawa T 1995 Mitochondrial-DNA mutations associatedwith aging and degenerative diseases Exp Gerontol 30269ndash290
Palmer J D 1997 Organelle genomes going going goneScience 275 790ndash791
Pearson C K Wilson S B Schaffer R amp Ross A W 1993NAD turnover and utilization of metabolites for RNA syn-thesis in a reaction sensing the redox state of the cyto-chrome-b( 6 )f complex in isolated chloroplasts Eur JBiochem 218 397ndash404
Pfanner N amp Geissler A 2001 Versatility of the mitochon-drial protein import machinery Natl Rev Mol Cell Biol 2339ndash349
Pfanner N amp Wiedemann N 2002 Mitochondrial proteinimport two membranes three translocases Curr Opin CellBiol 14 400ndash411
Pfannschmidt T Nilsson A amp Allen J F 1999a Photosyn-thetic control of chloroplast gene expression Nature 397625ndash628
Pfannschmidt T Nilsson A Tullberg A Link G amp AllenJ F 1999b Direct transcriptional control of the chloroplastgenes psbA and psaAB adjusts photosynthesis to light energydistribution in plants IUBMB Life 48 271ndash276
Pfannschmidt T Allen J F amp Oelmuller R 2001a Prin-ciples of redox control in photosynthesis gene expressionPhysiol Plant 112 1ndash9
Pfannschmidt T Schutze K Brost M amp Oelmuller R2001b A novel mechanism of nuclear photosynthesis generegulation by redox signals from the chloroplast during pho-
36 J F Allen Genomes in mitochondria and chloroplasts
tosystem stoichiometry adjustment J Biol Chem 27636 125ndash36 130
Popot J L amp de Vitry C 1990 On the microassembly of inte-gral membrane proteins A Rev Biophys Biophys Chem 19369ndash403
Race H L Herrmann R G amp Martin W 1999 Why haveorganelles retained genomes Trends Genet 15 364ndash370
Raven J A Johnston A M Parsons R amp Kubler J 1994aThe influence of natural and experimental high O2 concen-trations on O2 evolving phototrophs Biol Rev CambridgePhil Soc 69 61ndash94
Raven J A Johnston A M Parsons R amp Kubler J 1994bThe occurrence and influence on photolithotrophs of highoxygen concentrations Proc R Soc Edinb B 102 193ndash201
Robinson C 2000 The twin-arginine translocation system anovel means of transporting folded proteins in chloroplastsand bacteria Biol Chem 381 89ndash93
Robinson C amp Bolhuis A 2001 Protein targeting by the twin-arginine translocation pathway Natl Rev Mol Cell Biol 2350ndash356
Rodermel S 2001 Pathways of plastid-to-nucleus signalingTrends Plant Sci 6 471ndash478
Roger A J 1999 Reconstructing early events in eukaryoticevolution Am Nat 154 S146ndashS163
Roger A J Svard S G Tovar J Clark C G SmithM W Gillin F D amp Sogin M L 1998 A mitochondrial-like chaperonin 60 gene in Giardia lamblia evidence thatdiplomonads once harbored an endosymbiont related to theprogenitor of mitochondria Proc Natl Acad Sci USA 95229ndash234
Roodyn D B amp Wilkie D 1968 The biogenesis of mitochondriaMethuenrsquos monographs on biological subjects London Methuen
Rujan T amp Martin W 2001 How many genes in Arabidopsiscome from cyanobacteria An estimate from 386 proteinphylogenies Trends Genet 17 113ndash120
Saccone C Gissi C Lanave C Larizza A Pesole G ampReyes A 2000 Evolution of the mitochondrial genetic sys-tem an overview Gene 261 153ndash159
Salzberg S L White O Peterson J amp Eisen J A 2001Microbial genes in the human genome lateral transfer orgene loss Science 292 1903ndash1906
Sargent F Berks B C amp Palmer T 2002 Assembly of mem-brane-bound respiratory complexes by the Tat protein-trans-port system Arch Microbiol 178 77ndash84
Schatz G 1998 Protein transport the doors to organellesNature 395 439ndash440
Schemidt R A Qu J Williams J R amp Brusilow W S1998 Effects of carbon source on expression of F0 genes andon the stoichiometry of the c subunit in the F1 F0 ATPaseof Escherichia coli J Bacteriol 180 3205ndash3208
Selosse M A Albert B R amp Godelle B 2001 Reducing thegenome size of organelles favours gene transfer to thenucleus Trends Ecol Evol 16 135ndash141
Stern D B amp Lonsdale D M 1982 Mitochondrial and chlo-roplast genomes of maize have a 12- kilobase DNA-sequencein common Nature 299 698ndash702
Stock D Gibbons C Arechaga I Leslie A G W ampWalker J E 2000 The rotary mechanism of ATP synthaseCurr Opin Struct Biol 10 672ndash679
Stoebe B amp Maier U G 2002 One two three naturersquos toolbox for building plastids Protoplasma 219 123ndash130
Stoebe B Martin W amp Kowallik K V 1998 Distributionand nomenclature of protein-coding genes in 12 sequencedchloroplast genomes Plant Mol Biol Rep 16 243ndash255
Stupar R M Lilly J W Town C D Cheng Z Kaul SBuell C R amp Jiang J M 2001 Complex mtDNA consti-tutes an approximate 620 kb insertion on Arabidopsis thali-ana chromosome 2 implication of potential sequencing
Phil Trans R Soc Lond B (2003)
errors caused by large-unit repeats Proc Natl Acad SciUSA 98 5099ndash5103
Surpin M Larkin R M amp Chory J 2002 Signal transduc-tion between the chloroplast and the nucleus Plant Cell 14S327ndashS338
Tjaden J Schwoppe C Mohlmann T Quick P W ampNeuhaus H E 1998 Expression of a plastidic ATPADPtransporter gene in Escherichia coli leads to a functional aden-ine nucleotide transport system in the bacterial cytoplasmicmembrane J Biol Chem 273 9630ndash9636
Tovar J Fischer A amp Clark C G 1999 The mitosome a novelorganelle related to mitochondria in the amitochondrial para-site Entamoeba histolytica Mol Microbiol 32 1013ndash1021
Trebitsh T amp Danon A 2001 Translation of chloroplastpsbA mRNA is regulated by signals initiated by both photos-ystems II and I Proc Natl Acad Sci USA 98 12 289ndash12 294
Tullberg A Alexciev K Pfannschmidt T amp Allen J F2000 Photosynthetic electron flow regulates transcription ofthe psaB gene in pea (Pisum sativum L) chloroplasts throughthe redox state of the plastoquinone pool Plant Cell Physiol41 1045ndash1054
Turmel M Lemieux C Burger G Lang B F Otis CPlante I amp Gray M W 1999 The complete mitochondrialDNA sequences of Nephroselmis olivacea and Pedinomonasminor two radically different evolutionary patterns withingreen algae Plant Cell 11 1717ndash1729
Unden G amp Bongaerts J 1997 Alternative respiratory path-ways of Escherichia coli energetics and transcriptional regu-lation in response to electron acceptors Biochim BiophysActa Bioenerg 1320 217ndash234
Unseld M Marienfeld J R Brandt P amp Brennicke A1997 The mitochondrial genome of Arabidopsis thaliana con-tains 57 genes in 366 924 nucleotides Nature Genet 1557ndash61
Van der Giezen M Slotboom D J Horner D S DyalP L Harding M Xue G P Embley T M amp KunjiE R S 2002 Conserved properties of hydrogenosomal andmitochondrial ADPATP carriers a common origin for bothorganelles EMBO J 21 572ndash579
Von Heijne G 1986 Why mitochondria need a genome FEBSLett 198 1ndash4
Westphal S Soll J amp Vothknecht U C 2001 A vesicle trans-port system inside chloroplasts FEBS Lett 506 257ndash261
Whitehouse H L K 1969 Towards an understanding of themechanism of heredity London Edward Arnold Ltd
Williams B A P Hirt R P Lucocq J M amp Embley T M2002 A mitochondrial remnant in the microsporidian Trachi-pleistophora hominis Nature 418 865ndash869
Wilson R J (and 10 others) 1996a Complete gene map of theplastid-like DNA of the malaria parasite Plasmodium falcipa-rum J Mol Biol 261 155ndash172
Wilson S B Davidson G S Thomson L M amp PearsonC K 1996b Redox control of RNA synthesis in potato mito-chondria Eur J Biochem 242 81ndash85
Winkler H H amp Neuhaus H E 1999 Non-mitochondrialATP transport Trends Biochem Sci 24 64ndash68
Wolfe K H Morden C W amp Palmer J D 1992 Small sin-gle-copy region of plastid DNA in the non-photosyntheticangiosperm Epifagus virginiana contains only two genes Dif-ferences among dicots monocots and bryophytes in geneorganization at a non-bioenergetic locus J Mol Biol 22395ndash104
Wollman F A Minai L amp Nechushtai R 1999 The biogen-esis and assembly of photosynthetic proteins in thylakoidmembranes Biochim Biophys Acta Bioenerg 1411 21ndash85
Yeliseev A A amp Kaplan S 2000 TspO of Rhodobactersphaeroides a structural and functional model for the mam-
Genomes in mitochondria and chloroplasts J F Allen 37
malian peripheral benzodiazepine receptor J Biol Chem275 5657ndash5667
Zerges W 2002 Does complexity constrain organelle evol-ution Trends Plant Sci 7 175ndash182
Zhang Z D Green B R amp Cavalier-Smith T 1999 Singlegene circles in dinoflagellate chloroplast genomes Nature400 155ndash159
Zhang Z D Cavalier-Smith T amp Green B R 2001 A fam-ily of selfish minicircular chromosomes with jumbled chloro-plast gene fragments from a dinoflagellate Mol Biol Evol18 1558ndash1565
Zouni A Witt H T Kern J Fromme P Krauss NSaenger W amp Orth P 2001 Crystal structure of photosys-tem II from Synechococcus elongatus at 38 acngstrom resol-ution Nature 409 739ndash743
DiscussionD S Horner (Dipartimento di Fisiologia e Biochimica
Generali University of Milan Milan Italy) Given thatthere are components of these membrane-bound energy-transduction pathways that are consistently nuclear enco-ded not the key ones you pointed out but others do youexpect perhaps a slower perhaps a more crude but yet aredox communication between the organelle andexpression of nuclear-encoded genes
J F Allen There is and there is no question about itI think that Bogoradrsquos lock-in hypothesis has somethingto say here and there are other people who say it is all todo with assembling multi-subunit complexes If you havecontrol over the initial construction of the core aroundwhich the whole complex is eventually built then redoxcontrol or some sort of control over synthesis of that corewill control construction of the whole thing eventuallyand I think that may well be largely correct I think it ispossible in principle for nuclear genes to make precursorsaccording to redox instructions but it is not really soimportant that those precursors are here and now presentin exactly the right quantities because the precursors canto a limited extent hang around and wait Actually thereis proteolytic degradation of unattached precursors and soon but I think the fact that these chloroplast and mito-chondrially DNA-encoded subunits are at the core ofthese energy-transducing complexes is the crucial issue
T Cavalier-Smith (Department of Zoology University ofOxford Oxford UK ) In the case of mitochondria the sub-set of genes that remains is much more variable than youimplied It runs from three proteins in the minimal casesup to I think over 90 That is a big difference and manyof the genes in different lineages of protists that remainhave nothing to do with redox control they are things likeribosomal proteins and so on So I think that even thoughwhat you say about redox control of certain genes mayactually be an important thing it is not the fundamentaloverall reason why the genomes are maintained It may bepart of the explanation for why certain genes are alwaysincluded in that subset but even in mitochondria I thinkthat cytochrome b is the only one that is always presentSo I think that both this great diversity of what has beenretained in different lineages and the nucleomorph caseindicate that evolution was able to do one thing in onelineage 500 million years ago and never got around todoing it for 500 million years in another lineage so thereis a lot more haphazardness and evolutionary accidentsand contingency than a straight functional interpretation
Phil Trans R Soc Lond B (2003)
there is a lot of just sheer lsquohappenedndashorndashdidnrsquotndashhappenrsquoor carryover
J F Allen I was not implying anything about the rangeof mitochondrial genomes I have a limited amount oftime so I just talked about what I could talk about withinthat limit Of course there will be a range and this willdepend upon the physiology and bioenergetics of theparticular organism But what I am trying to do is to seewhat the common properties of the irreducible core areof components that are always encoded in the organellesand cytochrome b is a perfect example because that reallyis the thing that binds these two all-important quinoneswithout which there is no Q cycle without which there isno proton pumping and bioenergetics and in Plasmodiumwhere the mitochondrion is reduced to this bare mini-mum there is still a mitochondrial gene for cytochromeb Now my argument about the ribosomal proteins andall the rest of it is that in order to be able to exert thisall-important redox control over gene expression of certaincore components you need an in situ genetic system andthis can vary in composition for reasons that I cannotdescribe Maybe you need RNA polymerase or maybeyou can import RNA polymerase In plastids you haveboth of these options You are going to need ribosomestransfer RNAs and the rest of the apparatus of geneexpression and that needs to be in situ but we are stillleft with the question why does there have to be any gen-etic system at all The mature tobacco or spinach chloro-plast seems to have an entire genetic system just to makeone protein D1 which is all it ever needs to resynthesizeI think that this is a clue It is worth having this very unec-onomical crazy system because there is an overridingrequirement to connect electron transfer with geneexpression directly and unambiguouslymdashwithout recourseto a bureaucracy if you like which is what I think thenucleus is
K Willison (Institute of Cancer Research Chester BeattyLaboratories London UK ) These photosynthetic reactioncomplexes are a sort of baroque mixture of polypeptidechains and cofactors and my understanding is that someof them in high photon flux can turn over within say 30minutes so I am wondering if there is a correlationbetween gene retention and the turnover rates of some ofthe components and the complexes it seems to me thatthat could really be a limiting factor and the reason thatthe proteins had to be encoded locally in order to get thecomplexes to be assembled rapidly
J F Allen I think that is right You can only alter thesteady-state concentration of a protein complex by affect-ing the synthesis and breakdown and therefore turnoverThere is an extreme example the D1 protein encoded bythe psbA gene of chloroplasts which is always in the chlor-oplast genome without exception High light will tend todegrade this protein and when the high light goes awaythere is an absolute need very quickly to bring it back upagain I think that is a very good example Whether thisargument applies to genes such as cytochrome oxidase Ido not know
C J Leaver (Department of Plant Sciences University ofOxford Oxford UK ) Your experiments tend to relatemainly to the gene expression side What about proteinRNA turnover the whole concept of redox control ofthatmdashdo we know anything
38 J F Allen Genomes in mitochondria and chloroplasts
J F Allen Well quite a lot is known about chloro-plasts There is redox control of messenger RNA stabilityfor this same D1 protein which has been described byStephen Mayfield and his co-workers The evidence is thatthe site of redox control is ferredoxin via thioredoxin andthe implication is that thioredoxin tells whether non-cyclicelectron transport is working at all When it is workingthere is a signal that says lsquoOK now continue with all theprotein synthesisrsquo It is not particularly specific but moreof a global effect I think this is what Mayfield argues butof course this is not inconsistent with my argument I haveconcentrated on quinone pools and medium-potentialredox regulatory control because that is where I can seesignals about the dangerous consequences of getting itwrong but there is control of other levels of geneexpression In mitochondria I am not so sure I wouldbounce the question back to you on that one
W Martin (Institute of Botany III Heinrich-Heine Univ-ersitat Dusseldorf Dusseldorf Germany) The nucleomorphgenome was cited as evidence in conflict with your modelbut is that true The nucleomorph genome is not in abioenergetic organelle in that sense It resides in the cyto-sol so does the size of the nucleomorph genome workagainst your model Is it really evidence against yourmodel
J F Allen I do not know why should it be evidenceI cannot explain why the nucleomorph has a genomeThat is why I clutched at the straw that it is storage fornucleic acid precursors
Phil Trans R Soc Lond B (2003)
T Cavalier-Smith The reason I introduced the nucleo-morph is not to say it is direct evidence against yourhypothesis but to show that there probably was no singlefunctional reason for the retention of every gene in everyorganelle It may be that there are specific physiologicalreasons for the retention of certain genes and thereforeyou might be right about a subset of genes but I verymuch doubt if there are specific physiological reasons forthe retention of every single gene that is retained differen-tially because odd things happen in evolution Somethings are easy and sometimes things are difficult Evol-ution is history
J F Allen Of course I agree with that I do not thinkI can explain everything but I do think I can explainsomething If I have done that then I am content
GLOSSARY
AAC ADPndashATP carrierCES control by epistasy of synthesisCORR co-location for redox regulationLHC light-harvesting complexPCC Pasteur culture collectionORF open reading frameRUBISCO ribulose-15-bisphosphate carboxylasendashoxygenaseTAT twin-arginine translocase
Visit httpwwwpubsroyalsocacuk to see an electronic appendix tothis paper

36 J F Allen Genomes in mitochondria and chloroplasts
tosystem stoichiometry adjustment J Biol Chem 27636 125ndash36 130
Popot J L amp de Vitry C 1990 On the microassembly of inte-gral membrane proteins A Rev Biophys Biophys Chem 19369ndash403
Race H L Herrmann R G amp Martin W 1999 Why haveorganelles retained genomes Trends Genet 15 364ndash370
Raven J A Johnston A M Parsons R amp Kubler J 1994aThe influence of natural and experimental high O2 concen-trations on O2 evolving phototrophs Biol Rev CambridgePhil Soc 69 61ndash94
Raven J A Johnston A M Parsons R amp Kubler J 1994bThe occurrence and influence on photolithotrophs of highoxygen concentrations Proc R Soc Edinb B 102 193ndash201
Robinson C 2000 The twin-arginine translocation system anovel means of transporting folded proteins in chloroplastsand bacteria Biol Chem 381 89ndash93
Robinson C amp Bolhuis A 2001 Protein targeting by the twin-arginine translocation pathway Natl Rev Mol Cell Biol 2350ndash356
Rodermel S 2001 Pathways of plastid-to-nucleus signalingTrends Plant Sci 6 471ndash478
Roger A J 1999 Reconstructing early events in eukaryoticevolution Am Nat 154 S146ndashS163
Roger A J Svard S G Tovar J Clark C G SmithM W Gillin F D amp Sogin M L 1998 A mitochondrial-like chaperonin 60 gene in Giardia lamblia evidence thatdiplomonads once harbored an endosymbiont related to theprogenitor of mitochondria Proc Natl Acad Sci USA 95229ndash234
Roodyn D B amp Wilkie D 1968 The biogenesis of mitochondriaMethuenrsquos monographs on biological subjects London Methuen
Rujan T amp Martin W 2001 How many genes in Arabidopsiscome from cyanobacteria An estimate from 386 proteinphylogenies Trends Genet 17 113ndash120
Saccone C Gissi C Lanave C Larizza A Pesole G ampReyes A 2000 Evolution of the mitochondrial genetic sys-tem an overview Gene 261 153ndash159
Salzberg S L White O Peterson J amp Eisen J A 2001Microbial genes in the human genome lateral transfer orgene loss Science 292 1903ndash1906
Sargent F Berks B C amp Palmer T 2002 Assembly of mem-brane-bound respiratory complexes by the Tat protein-trans-port system Arch Microbiol 178 77ndash84
Schatz G 1998 Protein transport the doors to organellesNature 395 439ndash440
Schemidt R A Qu J Williams J R amp Brusilow W S1998 Effects of carbon source on expression of F0 genes andon the stoichiometry of the c subunit in the F1 F0 ATPaseof Escherichia coli J Bacteriol 180 3205ndash3208
Selosse M A Albert B R amp Godelle B 2001 Reducing thegenome size of organelles favours gene transfer to thenucleus Trends Ecol Evol 16 135ndash141
Stern D B amp Lonsdale D M 1982 Mitochondrial and chlo-roplast genomes of maize have a 12- kilobase DNA-sequencein common Nature 299 698ndash702
Stock D Gibbons C Arechaga I Leslie A G W ampWalker J E 2000 The rotary mechanism of ATP synthaseCurr Opin Struct Biol 10 672ndash679
Stoebe B amp Maier U G 2002 One two three naturersquos toolbox for building plastids Protoplasma 219 123ndash130
Stoebe B Martin W amp Kowallik K V 1998 Distributionand nomenclature of protein-coding genes in 12 sequencedchloroplast genomes Plant Mol Biol Rep 16 243ndash255
Stupar R M Lilly J W Town C D Cheng Z Kaul SBuell C R amp Jiang J M 2001 Complex mtDNA consti-tutes an approximate 620 kb insertion on Arabidopsis thali-ana chromosome 2 implication of potential sequencing
Phil Trans R Soc Lond B (2003)
errors caused by large-unit repeats Proc Natl Acad SciUSA 98 5099ndash5103
Surpin M Larkin R M amp Chory J 2002 Signal transduc-tion between the chloroplast and the nucleus Plant Cell 14S327ndashS338
Tjaden J Schwoppe C Mohlmann T Quick P W ampNeuhaus H E 1998 Expression of a plastidic ATPADPtransporter gene in Escherichia coli leads to a functional aden-ine nucleotide transport system in the bacterial cytoplasmicmembrane J Biol Chem 273 9630ndash9636
Tovar J Fischer A amp Clark C G 1999 The mitosome a novelorganelle related to mitochondria in the amitochondrial para-site Entamoeba histolytica Mol Microbiol 32 1013ndash1021
Trebitsh T amp Danon A 2001 Translation of chloroplastpsbA mRNA is regulated by signals initiated by both photos-ystems II and I Proc Natl Acad Sci USA 98 12 289ndash12 294
Tullberg A Alexciev K Pfannschmidt T amp Allen J F2000 Photosynthetic electron flow regulates transcription ofthe psaB gene in pea (Pisum sativum L) chloroplasts throughthe redox state of the plastoquinone pool Plant Cell Physiol41 1045ndash1054
Turmel M Lemieux C Burger G Lang B F Otis CPlante I amp Gray M W 1999 The complete mitochondrialDNA sequences of Nephroselmis olivacea and Pedinomonasminor two radically different evolutionary patterns withingreen algae Plant Cell 11 1717ndash1729
Unden G amp Bongaerts J 1997 Alternative respiratory path-ways of Escherichia coli energetics and transcriptional regu-lation in response to electron acceptors Biochim BiophysActa Bioenerg 1320 217ndash234
Unseld M Marienfeld J R Brandt P amp Brennicke A1997 The mitochondrial genome of Arabidopsis thaliana con-tains 57 genes in 366 924 nucleotides Nature Genet 1557ndash61
Van der Giezen M Slotboom D J Horner D S DyalP L Harding M Xue G P Embley T M amp KunjiE R S 2002 Conserved properties of hydrogenosomal andmitochondrial ADPATP carriers a common origin for bothorganelles EMBO J 21 572ndash579
Von Heijne G 1986 Why mitochondria need a genome FEBSLett 198 1ndash4
Westphal S Soll J amp Vothknecht U C 2001 A vesicle trans-port system inside chloroplasts FEBS Lett 506 257ndash261
Whitehouse H L K 1969 Towards an understanding of themechanism of heredity London Edward Arnold Ltd
Williams B A P Hirt R P Lucocq J M amp Embley T M2002 A mitochondrial remnant in the microsporidian Trachi-pleistophora hominis Nature 418 865ndash869
Wilson R J (and 10 others) 1996a Complete gene map of theplastid-like DNA of the malaria parasite Plasmodium falcipa-rum J Mol Biol 261 155ndash172
Wilson S B Davidson G S Thomson L M amp PearsonC K 1996b Redox control of RNA synthesis in potato mito-chondria Eur J Biochem 242 81ndash85
Winkler H H amp Neuhaus H E 1999 Non-mitochondrialATP transport Trends Biochem Sci 24 64ndash68
Wolfe K H Morden C W amp Palmer J D 1992 Small sin-gle-copy region of plastid DNA in the non-photosyntheticangiosperm Epifagus virginiana contains only two genes Dif-ferences among dicots monocots and bryophytes in geneorganization at a non-bioenergetic locus J Mol Biol 22395ndash104
Wollman F A Minai L amp Nechushtai R 1999 The biogen-esis and assembly of photosynthetic proteins in thylakoidmembranes Biochim Biophys Acta Bioenerg 1411 21ndash85
Yeliseev A A amp Kaplan S 2000 TspO of Rhodobactersphaeroides a structural and functional model for the mam-
Genomes in mitochondria and chloroplasts J F Allen 37
malian peripheral benzodiazepine receptor J Biol Chem275 5657ndash5667
Zerges W 2002 Does complexity constrain organelle evol-ution Trends Plant Sci 7 175ndash182
Zhang Z D Green B R amp Cavalier-Smith T 1999 Singlegene circles in dinoflagellate chloroplast genomes Nature400 155ndash159
Zhang Z D Cavalier-Smith T amp Green B R 2001 A fam-ily of selfish minicircular chromosomes with jumbled chloro-plast gene fragments from a dinoflagellate Mol Biol Evol18 1558ndash1565
Zouni A Witt H T Kern J Fromme P Krauss NSaenger W amp Orth P 2001 Crystal structure of photosys-tem II from Synechococcus elongatus at 38 acngstrom resol-ution Nature 409 739ndash743
DiscussionD S Horner (Dipartimento di Fisiologia e Biochimica
Generali University of Milan Milan Italy) Given thatthere are components of these membrane-bound energy-transduction pathways that are consistently nuclear enco-ded not the key ones you pointed out but others do youexpect perhaps a slower perhaps a more crude but yet aredox communication between the organelle andexpression of nuclear-encoded genes
J F Allen There is and there is no question about itI think that Bogoradrsquos lock-in hypothesis has somethingto say here and there are other people who say it is all todo with assembling multi-subunit complexes If you havecontrol over the initial construction of the core aroundwhich the whole complex is eventually built then redoxcontrol or some sort of control over synthesis of that corewill control construction of the whole thing eventuallyand I think that may well be largely correct I think it ispossible in principle for nuclear genes to make precursorsaccording to redox instructions but it is not really soimportant that those precursors are here and now presentin exactly the right quantities because the precursors canto a limited extent hang around and wait Actually thereis proteolytic degradation of unattached precursors and soon but I think the fact that these chloroplast and mito-chondrially DNA-encoded subunits are at the core ofthese energy-transducing complexes is the crucial issue
T Cavalier-Smith (Department of Zoology University ofOxford Oxford UK ) In the case of mitochondria the sub-set of genes that remains is much more variable than youimplied It runs from three proteins in the minimal casesup to I think over 90 That is a big difference and manyof the genes in different lineages of protists that remainhave nothing to do with redox control they are things likeribosomal proteins and so on So I think that even thoughwhat you say about redox control of certain genes mayactually be an important thing it is not the fundamentaloverall reason why the genomes are maintained It may bepart of the explanation for why certain genes are alwaysincluded in that subset but even in mitochondria I thinkthat cytochrome b is the only one that is always presentSo I think that both this great diversity of what has beenretained in different lineages and the nucleomorph caseindicate that evolution was able to do one thing in onelineage 500 million years ago and never got around todoing it for 500 million years in another lineage so thereis a lot more haphazardness and evolutionary accidentsand contingency than a straight functional interpretation
Phil Trans R Soc Lond B (2003)
there is a lot of just sheer lsquohappenedndashorndashdidnrsquotndashhappenrsquoor carryover
J F Allen I was not implying anything about the rangeof mitochondrial genomes I have a limited amount oftime so I just talked about what I could talk about withinthat limit Of course there will be a range and this willdepend upon the physiology and bioenergetics of theparticular organism But what I am trying to do is to seewhat the common properties of the irreducible core areof components that are always encoded in the organellesand cytochrome b is a perfect example because that reallyis the thing that binds these two all-important quinoneswithout which there is no Q cycle without which there isno proton pumping and bioenergetics and in Plasmodiumwhere the mitochondrion is reduced to this bare mini-mum there is still a mitochondrial gene for cytochromeb Now my argument about the ribosomal proteins andall the rest of it is that in order to be able to exert thisall-important redox control over gene expression of certaincore components you need an in situ genetic system andthis can vary in composition for reasons that I cannotdescribe Maybe you need RNA polymerase or maybeyou can import RNA polymerase In plastids you haveboth of these options You are going to need ribosomestransfer RNAs and the rest of the apparatus of geneexpression and that needs to be in situ but we are stillleft with the question why does there have to be any gen-etic system at all The mature tobacco or spinach chloro-plast seems to have an entire genetic system just to makeone protein D1 which is all it ever needs to resynthesizeI think that this is a clue It is worth having this very unec-onomical crazy system because there is an overridingrequirement to connect electron transfer with geneexpression directly and unambiguouslymdashwithout recourseto a bureaucracy if you like which is what I think thenucleus is
K Willison (Institute of Cancer Research Chester BeattyLaboratories London UK ) These photosynthetic reactioncomplexes are a sort of baroque mixture of polypeptidechains and cofactors and my understanding is that someof them in high photon flux can turn over within say 30minutes so I am wondering if there is a correlationbetween gene retention and the turnover rates of some ofthe components and the complexes it seems to me thatthat could really be a limiting factor and the reason thatthe proteins had to be encoded locally in order to get thecomplexes to be assembled rapidly
J F Allen I think that is right You can only alter thesteady-state concentration of a protein complex by affect-ing the synthesis and breakdown and therefore turnoverThere is an extreme example the D1 protein encoded bythe psbA gene of chloroplasts which is always in the chlor-oplast genome without exception High light will tend todegrade this protein and when the high light goes awaythere is an absolute need very quickly to bring it back upagain I think that is a very good example Whether thisargument applies to genes such as cytochrome oxidase Ido not know
C J Leaver (Department of Plant Sciences University ofOxford Oxford UK ) Your experiments tend to relatemainly to the gene expression side What about proteinRNA turnover the whole concept of redox control ofthatmdashdo we know anything
38 J F Allen Genomes in mitochondria and chloroplasts
J F Allen Well quite a lot is known about chloro-plasts There is redox control of messenger RNA stabilityfor this same D1 protein which has been described byStephen Mayfield and his co-workers The evidence is thatthe site of redox control is ferredoxin via thioredoxin andthe implication is that thioredoxin tells whether non-cyclicelectron transport is working at all When it is workingthere is a signal that says lsquoOK now continue with all theprotein synthesisrsquo It is not particularly specific but moreof a global effect I think this is what Mayfield argues butof course this is not inconsistent with my argument I haveconcentrated on quinone pools and medium-potentialredox regulatory control because that is where I can seesignals about the dangerous consequences of getting itwrong but there is control of other levels of geneexpression In mitochondria I am not so sure I wouldbounce the question back to you on that one
W Martin (Institute of Botany III Heinrich-Heine Univ-ersitat Dusseldorf Dusseldorf Germany) The nucleomorphgenome was cited as evidence in conflict with your modelbut is that true The nucleomorph genome is not in abioenergetic organelle in that sense It resides in the cyto-sol so does the size of the nucleomorph genome workagainst your model Is it really evidence against yourmodel
J F Allen I do not know why should it be evidenceI cannot explain why the nucleomorph has a genomeThat is why I clutched at the straw that it is storage fornucleic acid precursors
Phil Trans R Soc Lond B (2003)
T Cavalier-Smith The reason I introduced the nucleo-morph is not to say it is direct evidence against yourhypothesis but to show that there probably was no singlefunctional reason for the retention of every gene in everyorganelle It may be that there are specific physiologicalreasons for the retention of certain genes and thereforeyou might be right about a subset of genes but I verymuch doubt if there are specific physiological reasons forthe retention of every single gene that is retained differen-tially because odd things happen in evolution Somethings are easy and sometimes things are difficult Evol-ution is history
J F Allen Of course I agree with that I do not thinkI can explain everything but I do think I can explainsomething If I have done that then I am content
GLOSSARY
AAC ADPndashATP carrierCES control by epistasy of synthesisCORR co-location for redox regulationLHC light-harvesting complexPCC Pasteur culture collectionORF open reading frameRUBISCO ribulose-15-bisphosphate carboxylasendashoxygenaseTAT twin-arginine translocase
Visit httpwwwpubsroyalsocacuk to see an electronic appendix tothis paper

Genomes in mitochondria and chloroplasts J F Allen 37
malian peripheral benzodiazepine receptor J Biol Chem275 5657ndash5667
Zerges W 2002 Does complexity constrain organelle evol-ution Trends Plant Sci 7 175ndash182
Zhang Z D Green B R amp Cavalier-Smith T 1999 Singlegene circles in dinoflagellate chloroplast genomes Nature400 155ndash159
Zhang Z D Cavalier-Smith T amp Green B R 2001 A fam-ily of selfish minicircular chromosomes with jumbled chloro-plast gene fragments from a dinoflagellate Mol Biol Evol18 1558ndash1565
Zouni A Witt H T Kern J Fromme P Krauss NSaenger W amp Orth P 2001 Crystal structure of photosys-tem II from Synechococcus elongatus at 38 acngstrom resol-ution Nature 409 739ndash743
DiscussionD S Horner (Dipartimento di Fisiologia e Biochimica
Generali University of Milan Milan Italy) Given thatthere are components of these membrane-bound energy-transduction pathways that are consistently nuclear enco-ded not the key ones you pointed out but others do youexpect perhaps a slower perhaps a more crude but yet aredox communication between the organelle andexpression of nuclear-encoded genes
J F Allen There is and there is no question about itI think that Bogoradrsquos lock-in hypothesis has somethingto say here and there are other people who say it is all todo with assembling multi-subunit complexes If you havecontrol over the initial construction of the core aroundwhich the whole complex is eventually built then redoxcontrol or some sort of control over synthesis of that corewill control construction of the whole thing eventuallyand I think that may well be largely correct I think it ispossible in principle for nuclear genes to make precursorsaccording to redox instructions but it is not really soimportant that those precursors are here and now presentin exactly the right quantities because the precursors canto a limited extent hang around and wait Actually thereis proteolytic degradation of unattached precursors and soon but I think the fact that these chloroplast and mito-chondrially DNA-encoded subunits are at the core ofthese energy-transducing complexes is the crucial issue
T Cavalier-Smith (Department of Zoology University ofOxford Oxford UK ) In the case of mitochondria the sub-set of genes that remains is much more variable than youimplied It runs from three proteins in the minimal casesup to I think over 90 That is a big difference and manyof the genes in different lineages of protists that remainhave nothing to do with redox control they are things likeribosomal proteins and so on So I think that even thoughwhat you say about redox control of certain genes mayactually be an important thing it is not the fundamentaloverall reason why the genomes are maintained It may bepart of the explanation for why certain genes are alwaysincluded in that subset but even in mitochondria I thinkthat cytochrome b is the only one that is always presentSo I think that both this great diversity of what has beenretained in different lineages and the nucleomorph caseindicate that evolution was able to do one thing in onelineage 500 million years ago and never got around todoing it for 500 million years in another lineage so thereis a lot more haphazardness and evolutionary accidentsand contingency than a straight functional interpretation
Phil Trans R Soc Lond B (2003)
there is a lot of just sheer lsquohappenedndashorndashdidnrsquotndashhappenrsquoor carryover
J F Allen I was not implying anything about the rangeof mitochondrial genomes I have a limited amount oftime so I just talked about what I could talk about withinthat limit Of course there will be a range and this willdepend upon the physiology and bioenergetics of theparticular organism But what I am trying to do is to seewhat the common properties of the irreducible core areof components that are always encoded in the organellesand cytochrome b is a perfect example because that reallyis the thing that binds these two all-important quinoneswithout which there is no Q cycle without which there isno proton pumping and bioenergetics and in Plasmodiumwhere the mitochondrion is reduced to this bare mini-mum there is still a mitochondrial gene for cytochromeb Now my argument about the ribosomal proteins andall the rest of it is that in order to be able to exert thisall-important redox control over gene expression of certaincore components you need an in situ genetic system andthis can vary in composition for reasons that I cannotdescribe Maybe you need RNA polymerase or maybeyou can import RNA polymerase In plastids you haveboth of these options You are going to need ribosomestransfer RNAs and the rest of the apparatus of geneexpression and that needs to be in situ but we are stillleft with the question why does there have to be any gen-etic system at all The mature tobacco or spinach chloro-plast seems to have an entire genetic system just to makeone protein D1 which is all it ever needs to resynthesizeI think that this is a clue It is worth having this very unec-onomical crazy system because there is an overridingrequirement to connect electron transfer with geneexpression directly and unambiguouslymdashwithout recourseto a bureaucracy if you like which is what I think thenucleus is
K Willison (Institute of Cancer Research Chester BeattyLaboratories London UK ) These photosynthetic reactioncomplexes are a sort of baroque mixture of polypeptidechains and cofactors and my understanding is that someof them in high photon flux can turn over within say 30minutes so I am wondering if there is a correlationbetween gene retention and the turnover rates of some ofthe components and the complexes it seems to me thatthat could really be a limiting factor and the reason thatthe proteins had to be encoded locally in order to get thecomplexes to be assembled rapidly
J F Allen I think that is right You can only alter thesteady-state concentration of a protein complex by affect-ing the synthesis and breakdown and therefore turnoverThere is an extreme example the D1 protein encoded bythe psbA gene of chloroplasts which is always in the chlor-oplast genome without exception High light will tend todegrade this protein and when the high light goes awaythere is an absolute need very quickly to bring it back upagain I think that is a very good example Whether thisargument applies to genes such as cytochrome oxidase Ido not know
C J Leaver (Department of Plant Sciences University ofOxford Oxford UK ) Your experiments tend to relatemainly to the gene expression side What about proteinRNA turnover the whole concept of redox control ofthatmdashdo we know anything
38 J F Allen Genomes in mitochondria and chloroplasts
J F Allen Well quite a lot is known about chloro-plasts There is redox control of messenger RNA stabilityfor this same D1 protein which has been described byStephen Mayfield and his co-workers The evidence is thatthe site of redox control is ferredoxin via thioredoxin andthe implication is that thioredoxin tells whether non-cyclicelectron transport is working at all When it is workingthere is a signal that says lsquoOK now continue with all theprotein synthesisrsquo It is not particularly specific but moreof a global effect I think this is what Mayfield argues butof course this is not inconsistent with my argument I haveconcentrated on quinone pools and medium-potentialredox regulatory control because that is where I can seesignals about the dangerous consequences of getting itwrong but there is control of other levels of geneexpression In mitochondria I am not so sure I wouldbounce the question back to you on that one
W Martin (Institute of Botany III Heinrich-Heine Univ-ersitat Dusseldorf Dusseldorf Germany) The nucleomorphgenome was cited as evidence in conflict with your modelbut is that true The nucleomorph genome is not in abioenergetic organelle in that sense It resides in the cyto-sol so does the size of the nucleomorph genome workagainst your model Is it really evidence against yourmodel
J F Allen I do not know why should it be evidenceI cannot explain why the nucleomorph has a genomeThat is why I clutched at the straw that it is storage fornucleic acid precursors
Phil Trans R Soc Lond B (2003)
T Cavalier-Smith The reason I introduced the nucleo-morph is not to say it is direct evidence against yourhypothesis but to show that there probably was no singlefunctional reason for the retention of every gene in everyorganelle It may be that there are specific physiologicalreasons for the retention of certain genes and thereforeyou might be right about a subset of genes but I verymuch doubt if there are specific physiological reasons forthe retention of every single gene that is retained differen-tially because odd things happen in evolution Somethings are easy and sometimes things are difficult Evol-ution is history
J F Allen Of course I agree with that I do not thinkI can explain everything but I do think I can explainsomething If I have done that then I am content
GLOSSARY
AAC ADPndashATP carrierCES control by epistasy of synthesisCORR co-location for redox regulationLHC light-harvesting complexPCC Pasteur culture collectionORF open reading frameRUBISCO ribulose-15-bisphosphate carboxylasendashoxygenaseTAT twin-arginine translocase
Visit httpwwwpubsroyalsocacuk to see an electronic appendix tothis paper

38 J F Allen Genomes in mitochondria and chloroplasts
J F Allen Well quite a lot is known about chloro-plasts There is redox control of messenger RNA stabilityfor this same D1 protein which has been described byStephen Mayfield and his co-workers The evidence is thatthe site of redox control is ferredoxin via thioredoxin andthe implication is that thioredoxin tells whether non-cyclicelectron transport is working at all When it is workingthere is a signal that says lsquoOK now continue with all theprotein synthesisrsquo It is not particularly specific but moreof a global effect I think this is what Mayfield argues butof course this is not inconsistent with my argument I haveconcentrated on quinone pools and medium-potentialredox regulatory control because that is where I can seesignals about the dangerous consequences of getting itwrong but there is control of other levels of geneexpression In mitochondria I am not so sure I wouldbounce the question back to you on that one
W Martin (Institute of Botany III Heinrich-Heine Univ-ersitat Dusseldorf Dusseldorf Germany) The nucleomorphgenome was cited as evidence in conflict with your modelbut is that true The nucleomorph genome is not in abioenergetic organelle in that sense It resides in the cyto-sol so does the size of the nucleomorph genome workagainst your model Is it really evidence against yourmodel
J F Allen I do not know why should it be evidenceI cannot explain why the nucleomorph has a genomeThat is why I clutched at the straw that it is storage fornucleic acid precursors
Phil Trans R Soc Lond B (2003)
T Cavalier-Smith The reason I introduced the nucleo-morph is not to say it is direct evidence against yourhypothesis but to show that there probably was no singlefunctional reason for the retention of every gene in everyorganelle It may be that there are specific physiologicalreasons for the retention of certain genes and thereforeyou might be right about a subset of genes but I verymuch doubt if there are specific physiological reasons forthe retention of every single gene that is retained differen-tially because odd things happen in evolution Somethings are easy and sometimes things are difficult Evol-ution is history
J F Allen Of course I agree with that I do not thinkI can explain everything but I do think I can explainsomething If I have done that then I am content
GLOSSARY
AAC ADPndashATP carrierCES control by epistasy of synthesisCORR co-location for redox regulationLHC light-harvesting complexPCC Pasteur culture collectionORF open reading frameRUBISCO ribulose-15-bisphosphate carboxylasendashoxygenaseTAT twin-arginine translocase
Visit httpwwwpubsroyalsocacuk to see an electronic appendix tothis paper