the overexpression of the putative gut stem cell marker...
TRANSCRIPT

3256 Research Article
IntroductionThe intestinal epithelium is characterized by rapid and continuousrenewal throughout life. This renewal process depends onmultipotent stem cells located in the intestinal crypts (Stappenbecket al., 1998). These somatic stem cells self-renew by asymmetricdivision and give rise to undifferentiated progenitors that proliferate,differentiate while migrating, and are eventually shed in the lumenafter apoptosis. In mice, this entire process lasts 3–4 days(Stappenbeck et al., 1998). The continuous cell renewal and thehomeostasis of the intestinal epithelium are regulated by severalsignaling pathways, such as the Wnt and Notch pathways (Korineket al., 1998; van Es et al., 2005). We have also shown that thethyroid hormone receptor TR1 participates in these processes(Plateroti et al., 2001; Kress et al., 2008). The integration of thesesignals at the level of crypt progenitors has been shown to play animportant role (Plateroti et al., 2006; Fre et al., 2009; Kress et al.,2009). However, it remains unclear whether these functionalinteractions also play a role in intestinal stem cell physiology,because only recently have robust markers for these cells beendescribed (Montgomery and Breault, 2008). Importantly, amongthem, Musashi-1 (Potten et al., 2003; Kayahara et al., 2003) is atthe crossroads of the Wnt, Notch and thyroid hormone signalingpathways.
Musashi-1 (Msi1) is an RNA-binding protein, which acts as atranslational regulator of target mRNAs, by competing with thetranslation initiation factor eIF4G (Kawahara et al., 2008). Severalstudies have linked Msi1 with both stem cell and tumor biology,through its functional relations with signaling pathways. In neuronalprecursors, Msi1 is involved in the control of ‘stemness’ byregulating the Notch pathway (Okano et al., 2005). Moreover,
some data reported that Msi1 is regulated in mouse intestine by theWnt pathway (Potten et al., 2003) and by thyroid hormones duringmetamorphosis in amphibians (Ishizuya-Oka et al., 2003), as wellas in rat neural precursors (Cuadrado et al., 2002). Its involvementin stem cell physiology was first discovered through pioneer studiesin Drosophila (Nakamura et al., 1994; Siddall et al., 2006; Okabeet al., 2001). In mammals, Msi1 has been described as a marker ofadult stem cells and progenitors in other tissues such as the centralnervous system (Kaneko et al., 2000), hair follicles (Sugiyama-Nakagiri et al., 2006), and mammary glands (Clarke et al., 2005).However, the role of Msi1 in the stem-cell-progenitor compartmenthas been described only in the nervous system (Sakakibara et al.,2002).
Until recently, only two mRNA targets of Msi1 were known:mNumb, an antagonist of the Notch pathway that has been studiedin neural progenitor cells (Imai et al., 2001), and p21/Waf1,described in the HEK293T cell line (Battelli et al., 2006). de SousaAbreu et al. (de Sousa Abreu et al., 2009) identified new mRNAtargets of Msi1 in HEK293T cells. These mRNA targets areimplicated in cell proliferation and protein modification, both ofwhich are involved in tumorigenesis. Additional data reported theinvolvement of Msi1 in inducing cell proliferation in a mammarygland cell line (Wang et al., 2008), as well as in the growth of acolon carcinoma cell line in xenograft experiments (Sureban et al.,2008).
Taken together, these data indicate that Msi1 is important forstem cell biology in several systems. However, Msi1 is also relatedto several signaling pathways (i.e. Wnt, Notch, thyroid hormones)important for progenitor cell homeostasis, which is affected inintestinal tumorigenesis.
The overexpression of the putative gut stem cellmarker Musashi-1 induces tumorigenesis through Wntand Notch activationAmelie Rezza1, Seham Skah1, Colette Roche2, Julien Nadjar1, Jacques Samarut1 and Michelina Plateroti1,*1Institut de Génomique Fonctionnelle de Lyon, Université de Lyon, Université Lyon 1, CNRS, INRA, Ecole Normale Supérieure de Lyon,46 Allée d’Italie, 69364 Lyon Cedex 07, France2Inserm, U865, Lyon, France*Author for correspondence ([email protected])
Accepted 11 June 2010Journal of Cell Science 123, 3256-3265 © 2010. Published by The Company of Biologists Ltddoi:10.1242/jcs.065284
SummaryThe RNA-binding protein Musashi-1 (Msi1) has been proposed as a marker of intestinal epithelial stem cells. These cells areresponsible for the continuous renewal of the intestinal epithelium. Although the function of Msi1 has been studied in several organsfrom different species and in mammalian cell lines, its function and molecular regulation in mouse intestinal epithelium progenitorcells are still undefined. We describe here that, in these cells, the expression of Msi1 is regulated by the canonical Wnt pathway,through a mechanism involving a functional Tcf/Lef binding site on its promoter. An in vitro study in intestinal epithelium primarycultures showed that Msi1 overexpression promotes progenitor proliferation and activates Wnt and Notch pathways. Moreover, Msi1-overexpressing cells exhibit tumorigenic properties in xenograft experiments. These data point to a positive feedback loop betweenMsi1 and Wnt in intestinal epithelial progenitors. They also suggest that Msi1 has oncogenic properties in these cells, probably throughinduction of both the Wnt and Notch pathways.
Key words: Intestinal epithelium, Intestinal tumorigenesis, Musashi-1, Stem cells, Wnt
Jour
nal o
f Cel
l Sci
ence
We analyzed the physiological significance of these regulatorypathways as well as the function of Msi1 in intestinal epithelialprogenitor cells. We show that the expression of Msi1 is regulatedin a cell-autonomous way by the Wnt pathway. Lentiviral-mediatedincrease in the expression of Msi1, in turn, stimulates the Wntpathway through the induction of Frat1, a potent activator of -catenin stabilization (Hay et al., 2005; van Amerongen et al.,2005). Finally, we also show that Msi1 overexpression has thepotential to induce tumor formation in a xenograft model.
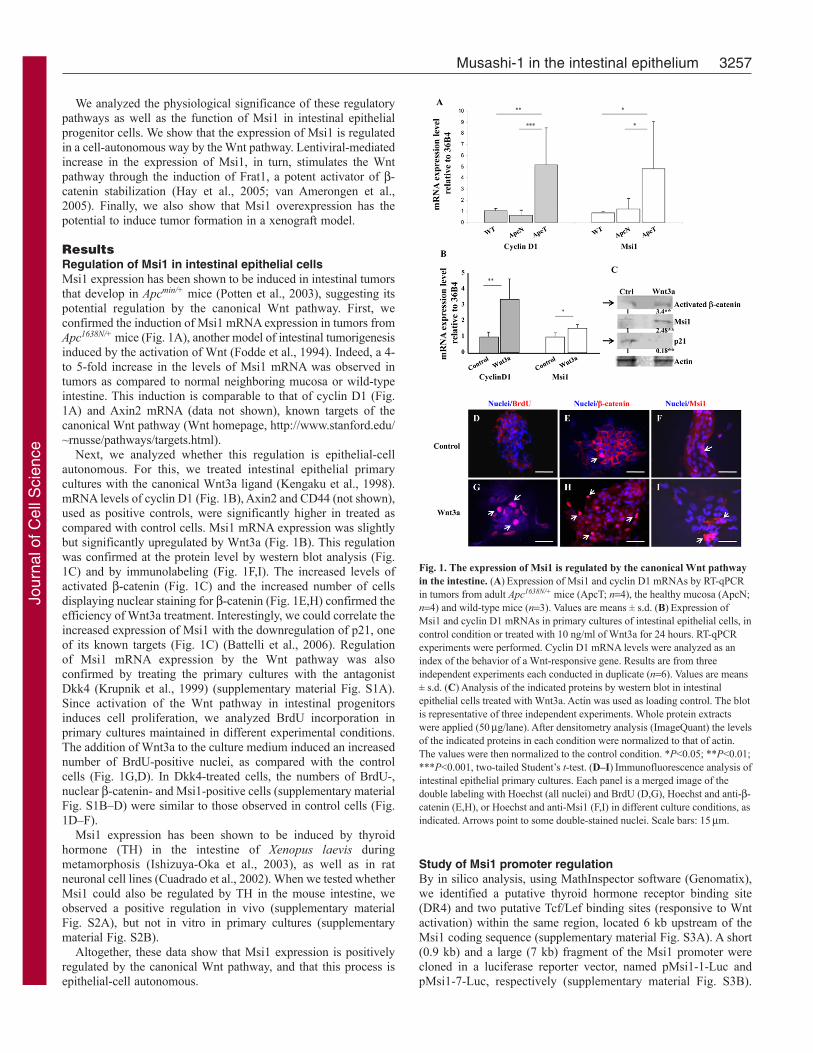
ResultsRegulation of Msi1 in intestinal epithelial cellsMsi1 expression has been shown to be induced in intestinal tumorsthat develop in Apcmin/+ mice (Potten et al., 2003), suggesting itspotential regulation by the canonical Wnt pathway. First, weconfirmed the induction of Msi1 mRNA expression in tumors fromApc1638N/+ mice (Fig. 1A), another model of intestinal tumorigenesisinduced by the activation of Wnt (Fodde et al., 1994). Indeed, a 4-to 5-fold increase in the levels of Msi1 mRNA was observed intumors as compared to normal neighboring mucosa or wild-typeintestine. This induction is comparable to that of cyclin D1 (Fig.1A) and Axin2 mRNA (data not shown), known targets of thecanonical Wnt pathway (Wnt homepage, http://www.stanford.edu/~rnusse/pathways/targets.html).
Next, we analyzed whether this regulation is epithelial-cellautonomous. For this, we treated intestinal epithelial primarycultures with the canonical Wnt3a ligand (Kengaku et al., 1998).mRNA levels of cyclin D1 (Fig. 1B), Axin2 and CD44 (not shown),used as positive controls, were significantly higher in treated ascompared with control cells. Msi1 mRNA expression was slightlybut significantly upregulated by Wnt3a (Fig. 1B). This regulationwas confirmed at the protein level by western blot analysis (Fig.1C) and by immunolabeling (Fig. 1F,I). The increased levels ofactivated -catenin (Fig. 1C) and the increased number of cellsdisplaying nuclear staining for -catenin (Fig. 1E,H) confirmed theefficiency of Wnt3a treatment. Interestingly, we could correlate theincreased expression of Msi1 with the downregulation of p21, oneof its known targets (Fig. 1C) (Battelli et al., 2006). Regulationof Msi1 mRNA expression by the Wnt pathway was alsoconfirmed by treating the primary cultures with the antagonistDkk4 (Krupnik et al., 1999) (supplementary material Fig. S1A).Since activation of the Wnt pathway in intestinal progenitorsinduces cell proliferation, we analyzed BrdU incorporation inprimary cultures maintained in different experimental conditions.The addition of Wnt3a to the culture medium induced an increasednumber of BrdU-positive nuclei, as compared with the controlcells (Fig. 1G,D). In Dkk4-treated cells, the numbers of BrdU-,nuclear -catenin- and Msi1-positive cells (supplementary materialFig. S1B–D) were similar to those observed in control cells (Fig.1D–F).
Msi1 expression has been shown to be induced by thyroidhormone (TH) in the intestine of Xenopus laevis duringmetamorphosis (Ishizuya-Oka et al., 2003), as well as in ratneuronal cell lines (Cuadrado et al., 2002). When we tested whetherMsi1 could also be regulated by TH in the mouse intestine, weobserved a positive regulation in vivo (supplementary materialFig. S2A), but not in vitro in primary cultures (supplementarymaterial Fig. S2B).
Altogether, these data show that Msi1 expression is positivelyregulated by the canonical Wnt pathway, and that this process isepithelial-cell autonomous.
3257Musashi-1 in the intestinal epithelium
Study of Msi1 promoter regulationBy in silico analysis, using MathInspector software (Genomatix),we identified a putative thyroid hormone receptor binding site(DR4) and two putative Tcf/Lef binding sites (responsive to Wntactivation) within the same region, located 6 kb upstream of theMsi1 coding sequence (supplementary material Fig. S3A). A short(0.9 kb) and a large (7 kb) fragment of the Msi1 promoter werecloned in a luciferase reporter vector, named pMsi1-1-Luc andpMsi1-7-Luc, respectively (supplementary material Fig. S3B).
Fig. 1. The expression of Msi1 is regulated by the canonical Wnt pathwayin the intestine. (A)Expression of Msi1 and cyclin D1 mRNAs by RT-qPCRin tumors from adult Apc1638N/+ mice (ApcT; n4), the healthy mucosa (ApcN;n4) and wild-type mice (n3). Values are means ± s.d. (B)Expression ofMsi1 and cyclin D1 mRNAs in primary cultures of intestinal epithelial cells, incontrol condition or treated with 10 ng/ml of Wnt3a for 24 hours. RT-qPCRexperiments were performed. Cyclin D1 mRNA levels were analyzed as anindex of the behavior of a Wnt-responsive gene. Results are from threeindependent experiments each conducted in duplicate (n6). Values are means± s.d. (C)Analysis of the indicated proteins by western blot in intestinalepithelial cells treated with Wnt3a. Actin was used as loading control. The blotis representative of three independent experiments. Whole protein extractswere applied (50g/lane). After densitometry analysis (ImageQuant) the levelsof the indicated proteins in each condition were normalized to that of actin.The values were then normalized to the control condition. *P<0.05; **P<0.01;***P<0.001, two-tailed Student’s t-test. (D–I)Immunofluorescence analysis ofintestinal epithelial primary cultures. Each panel is a merged image of thedouble labeling with Hoechst (all nuclei) and BrdU (D,G), Hoechst and anti--catenin (E,H), or Hoechst and anti-Msi1 (F,I) in different culture conditions, asindicated. Arrows point to some double-stained nuclei. Scale bars: 15m.
Jour
nal o
f Cel
l Sci
ence

Modulation of the transcriptional activity of both constructs byWnt signaling was tested in Cos7 cells. TopFlash and FopFlashreporter vectors were used as positive and negative controls,respectively. Cotransfection of TopFlash and expression vectorscoding for -catenin and Tcf1, nuclear effectors of the canonicalWnt pathway, lead to a 4-fold increase in luciferase activity (Fig.2A). As expected, Fopflash activity did not change in the differentexperimental conditions (Fig. 2B). Interestingly, cotransfection ofpMsi1-7-Luc together with -catenin and Tcf1 expression vectorsled to a 4-fold increase in luciferase activity, whereas pMsi1-1-Lucdid not respond to -catenin and Tcf1 overexpression (Fig. 2A).Moreover, a dominant negative form of Tcf1 (DNTcf), in thepresence of the wild-type Tcf1, abolished the induction of theluciferase activity on both TopFlash and pMsi1-7-Luc.
To identify the roles of the two putative Tcf/Lef binding sitespresent in the Msi1 promoter, we subcloned each of them upstreamof a luciferase reporter system. These constructs, as well as thespecific mutants, were used in cotransfection experiments (Fig.2C). Interestingly, only the site1 construct (Msi1-s1) was induced
3258 Journal of Cell Science 123 (19)
by cotransfection with -catenin and Tcf1 vectors. This regulationwas blunted when the same site was mutated (Msi1-s1m).
Taken together, these data show that Msi1 expression is regulatedby the Wnt–-catenin pathway in intestinal epithelial cells, likelythrough the functional Tcf/Lef binding site Msi1-s1 located in thepromoter region.
Similar experiments, aimed to evaluate the responsiveness ofthe Msi1 promoter to TH-TR1, clearly indicated that this promoteris unresponsive to the thyroid hormone-dependent signal(supplementary material Fig. S2C).
Overexpression of Msi1 cDNA in intestinal epithelialprimary culturesSeveral studies explored the function of Msi1 in tumor cell lines(Sureban et al., 2008; Wang et al., 2008). We decided to investigateits function in intestinal epithelial progenitor cells in a non-tumoralex vivo primary culture model. First, we overexpressed Msi1 inintestinal epithelial primary cultures using lentiviral infection.Intestinal epithelial cells were infected using GFP (I-GFP)- orMsi1 (I-Msi1)-coding viral particles. Msi1 overexpression wasconfirmed by RT-qPCR (Msi1CDS; Fig. 3A), immunofluorescence(Fig. 3C) and western blot analysis (Fig. 3F). It is worth noting thatoverexpression of Msi1 had no effect on the expression ofendogenous Msi1 mRNA (Msi1UTR; Fig. 3A).
Given that Msi1 is a putative marker of intestinal epithelial stemcells, we analyzed the effect of Msi1 overexpression on theexpression of Lgr5 and Bmi1, established markers of those cells(Barker et al., 2007; Sangiorgi and Capecchi, 2008). Interestingly,both mRNAs were significantly increased in Msi1-infected cellscompared with the control cells (supplementary material Fig. S4).Next, we wondered whether Msi1 overexpression could have aneffect on the expression of genes involved in stem cell physiology.For this investigation, we used the PCR array mouse stem cellapproach (SABiosciences), on RNA extracted from Msi1- or GFP-infected cells. The results of this analysis showed only a verylimited number of deregulated mRNAs in Msi1-overexpressingcells. However, we confirmed that the expression of cell cycleregulators can be affected by Msi1 (supplementary material TableS1), in accordance with a previous report (Wang et al., 2008).More importantly, our data also showed that Frat1 mRNA wassignificantly increased by Msi1 overexpression, as compared withthe level in the control cells. Interestingly, Frat1 is known to be apotent activator of the canonical Wnt pathway (Hay et al., 2005).This protein is dispensable with regard to Wnt–-catenin signaling,but it can enhance the accumulation of activated -catenin (vanAmerongen et al., 2005). We first validated Frat1 upregulation byconventional RT-qPCR on I-Msi1 as compared with control I-GFPcells (Fig. 3A). We also showed that Frat1 protein expression,analyzed by immunostaining and western blot, was clearlyincreased in Msi1-overexpressing cells (I-Msi1) as compared withcontrol cells (Fig. 3D–F).
Msi1 induces cell proliferation and activates Wnt–-catenin and Notch signaling pathwaysOur data on the stimulation of Msi1 expression by Wnt3a, togetherwith the results of the PCR arrays, clearly suggested a link betweenMsi1 and cell proliferation. We then quantified the proliferationrate of I-Msi1 as compared with the control I-GFP cells by countingBrdU-positive nuclei under the microscope (Fig. 4). Msi1-overexpressing cells showed a significant 3- to 4-fold increase inthe number of BrdU-positive nuclei as compared with I-GFP
Fig. 2. Identification of a functional Tcf/Lef binding site in the Msi1promoter. (A,B)Luciferase constructs were cotransfected with -catenin, Tcf1and a dominant negative form of Tcf1 (DNTcf) expression vectors in Cos7cells. In addition, Renilla luciferase expression vector (pRL-TK vector;Promega) was used to normalize the firefly luciferase content in each sample.TopFlash vector (Upstate) was used as a positive control for Tcf–-cateninactivation. FopFlash (B) vector (Upstate) was cotransfected into Cos7 cells asa negative control. Values are means ± s.d., from three independentexperiments, each conducted on six replicates (n18). (C)Cotransfectionexperiments with different luciferase constructs performed in Cos7 cells toidentify the functionality of the putative Tcf/Lef sites. Luciferase constructswith the Msi1 site1 (Msi1-s1) and Msi11 site2 (Msi1-s2), wild type andmutated (Msi1-s1m, Msi1-s2m) were used. TopFlash and FopFlash were usedas positive and negative controls, respectively. Values are means ± s.d., fromtwo independent experiments, each conducted on six replicates (n12)**P<0.01, ***P<0.001, by two-tailed Student’s t-test.
Jour
nal o
f Cel
l Sci
ence

control cells (Fig. 4C). Apoptosis, analyzed by immunostaining forcleaved caspase 3, did not vary among the different experimentalconditions (data not shown).
Msi1 was previously shown to activate the Notch pathway inneural precursors (Imai et al., 2001). Moreover, our data on Msi1-dependent induction of Frat1 expression led us to analyze the Wnt
3259Musashi-1 in the intestinal epithelium
and Notch pathways in intestinal epithelial primary cultures infectedwith Msi1. With regard to Notch, we observed that the intracellulardomain of the Notch-1 receptor (NICD), analyzed by western blot,was upregulated in Msi1-infected cells as compared with controlcells (Fig. 5A). Moreover, immunolabeling for Hes1, a knowntarget of NICD (Jarriault et al., 1995), showed a strong increase inthe number of cells displaying nuclear Hes1 in Msi1-overexpressingcells (Fig. 5D,C). Increased levels of both NICD and Hes1 reflectthe activation of Notch pathway. With regard to Wnt, infectionwith Msi1 clearly induced an increase in activated -catenincompared with the level in I-GFP cells (Fig. 5A). Immunostaininganalysis also showed an increased number of cells exhibitingnuclear -catenin in I-Msi1, as compared with control cells (Fig.5F,E). Interestingly, I-Msi1 cells also showed increased levels ofthe Wnt–-catenin targets Myc, cyclin D1 and cyclin D2 (Fig. 5B),all positive regulators of cell proliferation (He et al., 1998; Tetsuand McCormick, 1999; Kioussi et al., 2002). The increased numberof cyclin-D1-positive nuclei is also clearly illustrated byimmunofluorescence (Fig. 5H,G). Finally, reduced protein levelsof the cell cycle inhibitor p21 were also observed in I-Msi1 cells(Fig. 5B), probably due to the translational repression by Msi1(Battelli et al., 2006) and/or transcriptional repression by Myc(Gartel and Radhakrishnan, 2005).
To test the hypothesis that the Msi1-dependent increasedexpression of Frat1 was responsible for the Wnt–-cateninactivation, we used an approach of ShRNA to knockdown Frat1expression in I-Msi1 cells. The efficiency of two independentshort-hairpin constructs on Frat1 mRNA expression is illustratedin supplementary material Fig. S5A. This result was also confirmedat the protein level by the scoring of Frat1 immunolabeled cells in
Fig. 3. Overexpression of Msi1 in intestinal epithelial cells. (A)RT-qPCRanalysis to study specific mRNA expression, from intestinal epithelial cellsinfected with GFP (I-GFP) or Msi1 (I-Msi1)-expressing viral particles. TotalMsi1 (Msi1CDS), endogenous Msi1 (Msi1UTR) and Frat1 mRNA levels wereevaluated. Values are means ± s.d., from two independent experiments, eachconducted on triplicate (n6). *P<0.05, **P<0.01, by two-tailed Student’s t-test. (B–E)Immunostaining for GFP, Msi1, -catenin and Frat1 was performedon infected cells. Pictures show merged images of the double staining of nuclei(blue) and specific proteins (red) as indicated. Scale bars: (B–E) 15m;(higher magnification image of E) 8m. (F)Western blot analysis of theindicated proteins from infected cells. The blot is representative of threeindependent experiments, each conducted in duplicate. Actin was used as aloading control. After densitometry analysis (ImageQuant) the levels of theindicated proteins in each condition were normalized to that of actin. Thevalues were then normalized to the control condition. ***P<0.001; **P<0.01,n3, compared with control, by two-tailed Student’s t-test.
Fig. 4. Msi1 overexpression enhances cell proliferative capacity ofintestinal epithelial cells. (A,B)BrdU incorporation was analyzed in GFP-infected cells (I-GFP; A) and in Msi1-infected cells (I-Msi1; B). All nucleiwere labeled with Hoechst. Pictures show merged images of the doublestaining of nuclei (blue) and BrdU (green). Arrows point to some double-stained nuclei. (C)The percentage of BrdU-positive nuclei was determinedin GFP- and Msi1-infected cells. The histograms represent the summary(mean ± s.d.) of the scoring of specific immunolabeling in two independentexperiments, obtained by counting the BrdU-positive cells under themicroscope (300 cells per experimental condition). ***P<0.001, by two-tailedStudent’s t-test. Scale bars: 15m.
Jour
nal o
f Cel
l Sci
ence

different experimental conditions (Fig. 6A–C,J). Interestingly, weobserved that by infecting the I-Msi1 cells with ShRNAs againstFrat1, we reduced the number of cells expressing nuclear -cateninwhen compared with the cells infected only with Msi1 or with anShRNA negative control (Fig. 6D–F,K). Moreover, the decrease of-catenin-expressing nuclei was parallel to the decrease of cyclin-D1-expressing nuclei (Fig. 6G–I,L). Finally, the ShRNAs againstFrat1 had no effect on the number of Msi1- or Hes1-expressingcells (supplementary material Fig. S5B).
Taken together, these data show that Msi1 overexpression inintestinal epithelial primary cultures enhances their proliferativestatus and induces both the Wnt–-catenin and Notch signalingpathways. We also demonstrated that the induction of Wnt dependson the previous upregulation of Frat1.
3260 Journal of Cell Science 123 (19)
Injection of IEC6-Msi1 cells in immunodeficient miceMsi1 is known to be overexpressed in different tumors, includingintestinal tumors in a mutated Apc background (Potten et al., 2003)(Fig. 1A). Moreover, its downregulation has been correlated witha reduction of tumor growth (Sureban et al., 2008). However, thesedata were related to established pathological conditions (i.e. tumorsor tumor cell lines). No studies have reported that Msi1 exhibitspotential tumorigenicity in normal intestinal epithelial cells.
To test this possibility, we used the rat intestinal crypt-derivedcell line IEC6, which are untransformed cells retaining most oftheir original characteristics (Quaroni et al., 1979). IEC6 cells
Fig. 5. Overexpression of Msi1 activates the Wnt and Notch pathways andcell cycle regulators. (A,B)Representative western blot analyses of theindicated proteins from GFP- or Msi1-infected cells. The blots arerepresentative of three independent experiments. Actin was used as loadingcontrol. After densitometry analysis (ImageQuant) the levels of the indicatedproteins in each condition were normalized to that of actin. The values werethen normalized to the control condition. ***P<0.001; **P<0.01, n3,compared with control by two-tailed Student’s t-test.(C–F)Immunofluorescence analysis of intestinal epithelial primary cultures,infected as indicated. Double-labeling for Hoechst (nuclei) and Hes1 (C,D),Hoechst and -catenin (E,F), Hoechst and cyclin D1 (G,H) are shown. Scalebars: (C–F) 5m; (G,H) 15m.
Fig. 6. Knockdown of Frat1 abolishes Msi1-dependent activation of theWnt pathway in intestinal epithelial cells. Cells were first infected with GFP(I-GFP) or Msi1 (I-Msi1); then a second infection was performed on I-Msi1cells to express Sh Sc (control Sh) or Sh 30 and Sh 32, both targeting Frat1mRNA. (A–I)Immunostaining for Frat1, -catenin and cyclin D1 of infectedcells under different experimental conditions, as indicated. Pictures showmerged images of the double staining of nuclei (blue) and specific proteins(red) as indicated. Scale bars: 10m. (J–L)Percentage of Frat1- (J), -catenin-(K) and cyclin D1- (L) positive cells per nuclei. This was determined ininfected cells under different experimental conditions, as indicated. Values aremeans ± s.d. of the scoring of specific immunolabeling obtained by countingthe positive cells under the microscope (100 cells per experimental condition);***P<0.001, compared with the I-GFP condition; $$P<0.01, compared withthe I-Msi1 or I-Msi1/I-Sh Sc conditions, using a two-tailed Student’s t-test.
Jour
nal o
f Cel
l Sci
ence

were stably transduced with GFP- or Msi1-coding viral particles.In those cells, as in the primary cultures, Msi1 overexpressionactivates Wnt and Notch pathways, as assessed by immunolabelingfor -catenin and Hes1 (supplementary material Fig. S6). Next,both cell lines were used for xenograft experiments on BALB-c/nude mice. Two independent series of cell injections, includingthree controls (IEC6-GFP), and injection of seven mice with IEC6-Msi1, were performed. For mice injected with control cells, we didnot detect any graft growth (example in Fig. 7A), in accordancewith the non-tumorigenic properties of this cell line (Buick et al.,1987). We also confirmed the absence of IEC6-GFP cells byrecovering some subcutaneous biopsies at the sites of the injectionsand at other areas (not shown). By contrast, injections of IEC6-Msi1 cells induced the development of xenografts growing at theoriginal site of injection in all mice (example in Fig. 7B–D). Thesesamples were collected and used for histological andimmunofluorescence analyses. Within the grafts, the IEC6-Msi1cells frequently organized around a lumen-like structure (Fig.8A–C). In some cases, polarized epithelial cells with basal nucleiwere clearly visible (Fig. 8D). As expected, these cells expressedMsi1, as revealed by immunofluorescence (Fig. 9A), and had anepithelial phenotype, as assessed by cytokeratin labeling (Fig.9D,E). Moreover, they expressed high levels of -catenin,sometimes with a clear nuclear localization (Fig. 9B,C). Thesecells also expressed cyclin D1 (Fig. 9F) and Hes1 (Fig. 9H,I),reflecting the activation of both Wnt and Notch pathways. Finally,the IEC6-Msi1 cells were actively proliferating, as assessed byKi67 immunolabeling (Fig. 9G).
DiscussionThe intestinal epithelial stem cells are responsible for the continuousand rapid renewal of this tissue. It is known that these cells arelocated in the crypts of Lieberkhün, but their identification andcharacterization have been controversial. However, it has becomeclear that there exist two populations of intestinal stem cells. Onepopulation is composed of columnar basal cells (CBC), at the verybottom of the crypts, between Paneth cells in the small intestine.This population cycles actively, specifically express the Lgr5marker and can give rise to the different cytotypes of the intestinalepithelium (Barker et al., 2007). The second population is composedof quiescent stem cells, located at the +4 position, expressingBmi1 (Sangiorgi and Capecchi, 2008). This population also givesrise to all epithelial cytotypes, but the cells are slow cycling, and
3261Musashi-1 in the intestinal epithelium
retain labeling. Interestingly, the RNA-binding protein Msi1 hasbeen described as a putative intestinal stem cell marker for bothpopulations (Potten et al., 2003; Kayahara et al., 2003). This proteinis known to be involved in the regulation of asymmetric division,cell fate decisions and stem cell maintenance in Drosophila(Nakamura et al., 1994; Okabe et al., 2001; Siddall et al., 2006).In mammals, Msi1 is an adult stem cell marker in different organs,suggesting its potential role in adult stem cell physiology. However,the function of Msi1 in somatic stem cells, apart from those of thenervous system (Imai et al., 2001), and in particular of the intestinalepithelium, remain undefined. We show here, in an in vitro model,that Msi1 is involved in the proliferative capacities of the intestinalepithelial progenitors. This occurs as a result of Msi1-dependentactivation of both Wnt and Notch pathways in Msi1-overexpressingcells, which probably maintains the cells in a ‘progenitor state’.This is in accordance with recent observations in the Drosophiladeveloping eye (Fre et al., 2009).
It is clear that Msi1 is at the crossroads of several signalingpathways (Okano et al., 2005) that are important in intestinaldevelopment and homeostasis. However, a detailed analysis ofthese multiple regulations in the same physiological model waslacking. We sought to construct a unified model. Indeed, we showedthat the thyroid hormones regulate the expression of Msi1 in themouse intestine, as previously demonstrated in amphibians(Ishizuya-Oka et al., 2003). In accordance with the paper byIshizuya-Oka et al. (Ishizuya-Oka et al., 2003), we also observedthat this regulation is not epithelial autonomous. Finally, given thatthe Msi1 promoter is not regulated by TH-TR1, we conclude thatthis control is indirect. However, supplementary studies arenecessary to clearly define the mechanism by which TH modulatesMsi1 levels. Msi1 expression is upregulated in intestinal tumorsfrom Apc-mutant mice (Potten et al., 2003), suggesting its possibleregulation by the Wnt pathway. We confirmed this finding in thepresent study and extended the observation by analyzing Msi1expression in intestinal tumors from the Apc1638N/+ mice, another
Fig. 7. Xenograft experiments in BALB-c/nude mice. (A–D)Nude mice thatreceived IEC6-GFP (A) or IEC6-Msi1 injections (B–D). Only in the case ofIEC6-Msi1 cells did we observe specific growth of xenografts (indicated bythe black dashed line).
Fig. 8. Histological analysis of the xenografts. (A–D)Hematoxylin–Eosinstaining of paraffin sections of xenograft growth after injection of IEC6-Msi1cells. Note that the cells organize around lumen-like structures. In some casethe cells are well polarized with basal nuclei, as indicated by the white dashedline in D. Scale bars: (A) 30m; (B) 15m; (C,D) 5m.
Jour
nal o
f Cel
l Sci
ence

Apc-mutant mouse model, and in an in vitro model of primarycultures. Furthermore, we dissected the mechanism responsible forthe activation of Msi1 expression by the Wnt pathway. In fact, weidentified a functional Tcf/Lef binding site located in the promoterof Msi1, at –6 kb from the start site. It is worth noting that previousstudies using 4.5 kb of the Msi1 mouse promoter failed todemonstrate its direct control by -catenin–Tcf (Keyoung et al.,2001; Kanai et al., 2006). This is not in contradiction with ourresults, because we showed that the Tcf/Lef binding site is outsidethe portion of the promoter that was used by these authors.
Msi1 is an RNA-binding protein that represses mRNA translationby binding specific consensus sites present in the 3�-UTR regionof its targets (Imai et al., 2001). In fact, Msi1 competes with thetranslation initiation factor eIF4G to bind PABP, preventing theinitiation of translation (Kawahara et al., 2008). A recent studyidentified new direct targets of Msi1 using a global RIP-Chipanalysis (de Sousa Abreu et al., 2009). The authors showed thatthese targets preferentially participate in two major processesinvolved in tumorigenesis: proliferation versus apoptosis, andprotein modifications. Another study described several mRNAsthat are deregulated by Msi1 overexpression, which are thereforeconsidered to be indirect targets (Wang et al., 2008). Interestingly,this study showed that the overexpression of Msi1 in a mammarycell line induces cell proliferation and activates Wnt and Notchpathways. Here we report similar results in a different cellularcontext. Indeed, we show that Msi1 overexpression increases the
3262 Journal of Cell Science 123 (19)
proliferative capacity of epithelial progenitors in vitro. Moreimportantly, and in accordance with the previous study on themammary cell line, we also described the joint induction of Wntand Notch pathways. These results, therefore, point to a generalmechanism of action of Msi1.
The mechanism at the basis of the Msi1-dependent activation ofthe Wnt pathway in our model system is probably due to enhancedexpression of Frat1. This is the first report describing the regulationof Frat1 by Msi1. Frat1 is known to be a potent activator of thecanonical Wnt signaling pathway, since it interacts with GSK3and Dvl proteins and enhances Lef-mediated transcription (Li etal., 1999). Accordingly, we showed that the increased expressionof Frat1 mediated by Msi1 stabilizes -catenin and increases levelsof its targets, cyclin D1 and Myc. It is worth noting that the studyon the mammary cell line described a different mechanism forMsi1 activation of the Wnt pathway, dependent on decreasedexpression of Dkk3 (Wang et al., 2008). Even if we can suppose ageneralized mechanism of action of Msi1 in stem and/or progenitorcells, our data suggest that cell-context specificities need to betaken into account. Finally, concerning Notch activation in Msi1-overexpressing cells, our results show increased levels of NotchNICD and Hes1. Because of the recent data describing Notchactivation by Wnt in colorectal cancer cells (Rodilla et al., 2009),it was possible that, in our model, the increased Notch activity wassecondary to that of Wnt. However, we have shown that this is notthe case. In fact, blocking the Wnt activation by Frat1 knockdown
Fig. 9. Immunolabeling analysis of the xenografts.(A–I)Immunolabeling for Msi1 (A), -catenin (B,C),cytokeratin (D,E), cyclin D1 (F), Ki67 (G) and Hes1 (H,I).Pictures show merged images of the double staining ofnuclei (blue) and specific proteins (red). White arrows in Cpoint to -catenin nuclear staining. Scale bars: (A–C, F–H)15m; (C,E,I) 10m.
Jour
nal o
f Cel
l Sci
ence

3263Musashi-1 in the intestinal epithelium
in I-Msi1 cells has no effect on Hes1 expression. These data clearlyshow that Notch activation in Msi1-overexpressing cells isindependent of Wnt activity. Hence, the pattern we observe mightbe due to the direct translational repression of mNumb, as has beenreported for neural precursors (Imai et al., 2001). Taken together,these results show that Msi1 overexpression enhances theproliferative capacities of intestinal progenitors throughconcomitant induction of Wnt and Notch pathways. Moreover, weidentified Frat1, an activator of -catenin signaling, as anintermediate target responsible for the Msi1-dependent activationof Wnt.
Different studies have shown that a deregulation of the canonicalWnt pathway leads to intestinal tumorigenesis (Moser et al., 1990;Fodde et al., 1994). Moreover, Wnt and Notch pathways act insynergy to promote colorectal cancer (Fre et al., 2009; Rodilla etal., 2009). Notably, increased levels of Msi1 have been clearlydemonstrated in several tumors, such as gliomas (Toda et al.,2001), endometrial tumors (Götte et al., 2008) and intestinal tumors(Potten et al., 2003). This observation suggests that Msi1overexpression, which induces both Wnt and Notch pathways,might have tumorigenic potential. In fact, we show here that Msi1overexpression induces subcutaneous tumor development. Thisinduction was clearly associated with increased proliferativecapacity within the xenografts. However, it is worth noting thatsuch a regulatory loop is achieved through high Msi1overexpression. Further investigation will be necessary to definewhether a precise correlation exists between Msi1-overexpressionlevels and specific cellular responses.
These data demonstrate the tumor-inducing potential of Msi1 innormal intestinal epithelial cells and suggest that deregulatedexpression of Msi1 could be an early event associated with intestinaltumorigenesis. In fact, it has been suggested that Frat1 could berecruited to activate -catenin–Tcf-dependent transcription when thephysiological balance between proliferation and differentiation isdisrupted (Ruiz et al., 2004). Increased expression of Frat1 in Msi1-overexpressing cells could then reflect a pathological state linked tothe tumor-inducer role of Msi1 described here. Our data are inagreement with recent reports that intestinal epithelial stem cells areat the origin of intestinal carcinogenesis (Sangiorgi and Capecchi,2008; Barker et al., 2009). Two previous studies linked the level ofMsi1 expression to human colorectal carcinomas in patients (Fan etal., 2009) and in an ex vivo study (Sureban et al., 2008). Our resultsconfirm this link and elucidate the function of Msi1 in early celltransformation events leading to tumorigenesis.
Materials and MethodsAnimals and tissue preparationWe used Apc1638N/+ and wild-type animals in this study. Mice were housed andmaintained with approval from the animal experimental committee of the EcoleNormale Supérieure de Lyon (Lyon, France) and in accordance with Europeanlegislation on animal care and experimentation. Thyroid hormone (TH) deficiencyin pups was induced by feeding the mothers a low-iodine diet supplemented with0.15% propylthiouracil (PTU; Harlan/Teklad) for 2 weeks. Hyperthyroidism wasinduced by intraperitoneal injections of a mixture of T4 and T3 (one daily injectionfor two days of 2.5 mg/kg T4 and 0.25 mg/kg T3 in 100 l of phosphate-bufferedsaline) to PTU-feed animals. Control animals were fed standard mouse chow.Animals were killed at the indicated ages, and the intestine was quickly removed.Samples were frozen in liquid nitrogen and used later for RNA extraction.
Primary cell cultures of intestinal epithelial cellsIntestinal epithelial primary cultures were derived from 4- to 6-day-old neonatalmice. Briefly, after sacrifice, the entire small intestine was removed. The epitheliumwas isolated as intact organoids by enzymatic dissociation using collagenase type XI(Sigma) and dispase (Boehringer Mannheim), followed by physical disaggregationand filtration on gauze. Organoids were plated in Dulbecco’s modified Eagle medium
(DMEM; Invitrogen) supplemented with 2.5% heat-inactivated fetal calf serum(Gibco), 20 ng/ml epidermal growth factor (Sigma), and insulin–transferrin–seleniumdiluted 1:100 (Sigma). Culture surfaces were coated with MatrigelTM BasementMembrane Matrix (BD Biosciences). For immunolabeling experiments, coverslipswere inserted in the wells before coating. For proliferation studies, 10 M BrdU wasadded to the culture medium for an overnight incubation. The purity of the epithelialcolonies was analyzed by immunolabeling for specific markers: anti-cytokeratins(ICN) for epithelial cells; anti-vimentin (Sigma) for fibroblasts; anti-smooth muscleactin (Sigma) for smooth muscle cells (supplementary material Fig. S7). For treatmentexperiments, after 2 days of culture, recombinant Wnt3a or Dkk4 was added to theculture medium for 24 hours.
Lentiviral infectionsFor overexpression experiments, cells were infected with lentiviral particles. Lentiviralvectors expressing GFP or Msi1 were obtained from the Vectorology Facility atIFR128 Lyon-Biosciences. GAE–SFFV–GFP is an SIV-derived lentiviral vectorexpressing GFP (a gift from Els Verhoeyen, INSERM U758, Lyon, France). GAE–SFFV–Msi1 vector was obtained by cloning Msi1 cDNA into GAE–SFFV–GFPafter removal of GFP cDNA. After 2 days in culture, viral supernatant in epithelialmedium (1:1) was used to infect intestinal epithelial cells, for 48 hours or 96 hours.GAE–SFFV–GFP vector was used to produce control lentiviral particles. For ShRNAexperiments, primary cultures were first infected with GAE–SFFV–Msi1 or GAE–SFFV–GFP vectors. 48 hours after the first infection, cells were infected with Shviral supernatant in epithelial medium (3:2) for 96 hours. ShRNA lentiviral vectorswere Mission-shRNA (derived from pLKO.1-puro; Sigma). Lentiviral particlesexpressing Sh SC (scrambled control), Sh 30 and Sh 32 (targeting Frat1 mRNA)were obtained from the Vectorology Facility at IFR128 Lyon-Biosciences.
Cells were washed twice with phosphate-buffered saline and frozen at –80°C forRNA or protein extraction, or alternatively, fixed in 2% paraformaldehyde forimmunostaining experiments.
RNA extraction and analysisRNA was extracted from tissue samples using the Qiagen RNeasy kit (Qiagen). Toavoid the presence of contaminating DNA, DNase digestion was performed on allpreparations. Reverse transcription was performed using MuMLV reversetranscriptase (Promega) on 1 g of total RNA according to the manufacturer’sinstructions, using random hexanucleotide priming (Promega). For primary cultures,RNA was extracted using the Absolutely RNA Nanoprep Kit (Stratagene). Reversetranscription was performed using SuperScript III First-Strand Synthesis SuperMixfor qRT-PCR (Invitrogen) on 300 ng of total RNA. All cDNA samples were purifiedusing the Qiagen PCR purification kit (Qiagen) before use for qPCR experiments.qPCR analyses were performed with SYBR green PCR master mix (Qiagen) in anMxP3000 apparatus (Stratagene). The data from the qPCR were normalized to 36B4levels in each sample. Primer sequences are shown in supplementary material TableS2 or in van der Flier et al. (van der Flier et al., 2009) and Kress et al. (Kress et al.,2010).
Immunostaining and western blotImmunolabeling of primary intestinal epithelial cells was performed on 2%paraformaldehyde-fixed cultures. Paraffin sections (5 m thickness) of xenograftswere used for indirect immunostaining. Briefly, sections were deparaffinized inmethylcyclohexane for 10 minutes, hydrated in ethanol, and washed with PBS.Slides were then subjected to antigen retrieval, using microwave heating in 0.01 Mcitrate buffer (pH 6), and incubation for 1 hour at room temperature with blockingbuffer (10% normal goat serum, 1% BSA, and 0.02 Triton X-100 in PBS). The slidesor coverslips were incubated with primary antibodies overnight at 4°C, followed byincubation with fluorescent secondary antibodies (Jackson ImmunoResearchLaboratories, Inc., West Grove, PA, USA). All nuclei were stained with Hoechst(Sigma). Fluorescence and bright-field microscopy were performed on a ZeissAxioplan microscope (Carl Zeiss).
Protein extracts were obtained by homogenizing intestinal epithelial cells in RIPAbuffer (50 mM Tris, pH 7.4; 150 mM NaCl; 1 mM phenylmethylsulfonylfluoride; 1mM EDTA; 5 g/ml aprotinin; 5 g/ml leupeptin; 1% Triton X-100; 1% sodiumdeoxylate; 0.1% sodium dodecyl sulfate; 40 l/ml protease inhibitors). Proteins wereseparated on 8 or 10% acrylamide-bis acrylamide (29:1) gel and transferred to anitrocellulose membrane (Hybond ECL, Amersham) before saturation and incubationwith the first antibody. This was followed by incubation with secondary antibodies(Promega). The signal was analyzed using the enzymatic chemiluminescencedetection kit (LumiLight; Roche Clinical Laboratories).
We used the following primary antibodies: anti-activated -catenin (Upstate);anti--catenin (Santa Cruz); anti-Musashi-1 (Chemicon and Abcam); anti-Notch1-NICD (Cell Signaling); anti-p21 (Santa Cruz); anti-Frat1 (Santa Cruz); anti-cyclinD1(LabVision); anti-cyclinD2 (Sigma); anti-Ki67 (Labvision); anti-c-myc (Santa Cruz);anti-Hes1 (MBL).
Construction of pMsi1-Luc vectorsMsi1 promoters were amplified from BAC clone RP24-243N1 (BACPAC ResourcesCenter, CHORI, Oakland, CA). Briefly, a 900 bp fragment was amplified at the5� end of the Msi1 start codon and cloned in pGL3 vector (Promega) to construct
Jour
nal o
f Cel
l Sci
ence

3264 Journal of Cell Science 123 (19)
the pMsi1–1-Luc vector. A 6 kb distal fragment was added into pMsi1–1-Luc toobtain the pMsi1–7-Luc construct, using the SpeI site (scheme in supplementarymaterial Fig. S3). Msi1s1wt-Luc, Msi1s1Mut-Luc, Msi1s2wt-Luc and Msi1s2Mut-Luc constructs were obtained by cloning a 20-bp sequence corresponding,respectively, to Msi1–Tcf/Lef putative binding site1 wild-type (5�-TGGGAGTTC -TTTGAAACACACG-3�), site1 mutated (5�-TGGGAGTTCTTTGGGACACACG-3�), site2 wild-type (5�-TGGG CGGATCTTTGAATTCGAGG-3�), and site2 mutated(5�-TGGGCGGATCTTTG GGTTCGAGG-3�). These were cloned upstream of theTK minimal promoter of the TopFlash backbone (Upstate) using NdeI–BamHIrestriction enzymes, after removal of the Tcf/Lef sites.
Cell lines culture and transfection experimentsCos7 cells and IEC6 cells were cultured in DMEM supplemented with 5% heat-inactivated fetal calf serum. pGl2-DR4-Luc vector, TopFlash vector (Upstate),FopFlash vector (Upstate), pMsi1-Luc vectors, pCIneo--cateninXL vector, Evr2-Tcf1E vector or Evr2-DN-Tcf1E vector were transfected into Cos7 cells using theExgen transfection reagent (Euromedex). For T3 treatments, the cells weremaintained, after transfection, in thyroid hormone-depleted serum. T3 (10–7 M) orvehicle alone was added to the culture medium 24 hours before the end of theculture. Luciferase activity was measured 48 hours after transfection using theluciferase dual system (Promega). IEC6–GFP and IEC6–Msi1 cell lines were obtainedby lentiviral infection of IEC6 cells, using GAE–SFFV–GFP- or GAE–SFFV–Msi1-derived viral particles.
PCR arraysRNA extracted from cells that had been infected for 2 days were used with thePAMM-405A RT2 ProfilerTM PCR Array Mouse Stem Cell system (SABiosciences).Then 500 ng of RNA were retrotranscribed using the RT2 First Strand Kit(SABiosciences) according to the manufacturer’s protocol. RT-PCR array wasperformed using RT2 qPCR Master Mix (SABiosciences) in an Mx3000P apparatus(Stratagene). Results were analyzed with SABiosciences software available at:http://www.sabiosciences.com/rt_pcr_product/HTML/PAMM-405A.html.
Xenograft experimentsWe used 4–5 week-old athymic (BALB/c nu/nu) female mice. Tumorigenicity wasassayed by subcutaneous dorsal injections of 3–4�106 IEC6–GFP or IEC6–Msi1cells resuspended in 50 l of IEC6 culture medium mixed with 50 l of matrigel(BD Biosciences). Animals were killed 6 weeks after cell inoculation, and graftswere removed, fixed in 4% paraformaldehyde and then embedded in paraffin forhistological analysis.
We gratefully acknowledge Nadine Aguilera for animal handling.We thank the Vectorology Facility at ENS-Lyon, as well as Els Verhoen,Didier Negre and Veronique Barateau for reagents and production oflentivirus. We are indebted to Fabrice Lavial and Marian Waterman forexpression vectors. This work was supported by the Agence Nationalepour la Recherche (grant ANR-06-BLAN-0232-01) and the InstitutNational du Cancer (grant INCA-2007-Col6). A.R. received supportfrom the Ligue Nationale contre le Cancer and the Fondation pour leRecherche Médicale.
Supplementary material available online athttp://jcs.biologists.org/cgi/content/full/123/19/3256/DC1
ReferencesBarker, N., van Es, J. H., Kuipers, J., Kujala, P., van den Born, M., Cozijnsen, M.,
Haegebarth, A., Korving, J., Begthel, H., Peters, P. J. et al. (2007). Identification ofstem cells in small intestine and colon by marker gene Lgr5. Nature 449, 1003-1007.
Barker, N., Ridgway, R. A., van Es, J. H., van de Wetering, M., Begthel, H., van denBorn, M., Danenberg, E., Clarke, A. R., Sansom, O. J. and Clevers, H. (2009).Crypt stem cells as the cells-of-origin of intestinal cancer. Nature 457, 608-611.
Battelli, C., Nikopoulos, G. N., Mitchell, J. G. and Verdi, J. M. (2006). The RNA-binding protein Musashi-1 regulates neural development through the translationalrepression of p21WAF-1. Mol. Cell. Neurosci. 31, 85-96.
Buick, R. N., Filmus, J. and Quaroni, A. (1987). Activated H-ras transforms rat intestinalepithelial cells with expression of alpha-TGF. Exp. Cell Res. 170, 300-309.
Clarke, R. B., Spence, K., Anderson, E., Howell, A., Okano, H. and Potten, C. S.(2005). A putative human breast stem cell population is enriched for steroid receptor-positive cells. Dev. Biol. 277, 443-456.
Cuadrado, A., Garcia-Fernandez, L. F., Imai, T., Okano, H. and Munoz, A. (2002).Regulation of tau RNA maturation by thyroid hormone is mediated by the neural RNA-binding protein musashi-1. Mol. Cell. Neurosci. 20, 198-210.
de Sousa Abreu, R., Sanchez-Diaz, P. C., Vogel, C., Burns, S. C., Ko, D., Burton, T.L., Vo, D. T., Chennasamudaram, S., Le, S.-Y., Shapiro, B. A. et al. (2009). Genomicanalyses of musashi1 downstream targets show a strong association with cancer-relatedprocesses. J. Biol. Chem. 284, 12125-12135.
Fan, L. F., Dong, W. G., Jiang, C. Q., Xia, D., Liao, F. and Yu, Q. F. (2009). Expressionof putative stem cell genes Musashi-1 and beta1-integrin in human colorectal adenomasand adenocarcinomas. Int. J. Colorectal Dis. 25, 17-23.
Fodde, R., Edelmann, W., Yang, K., van Leeuwen, C., Carlson, C., Renault, B.,Breukel, C., Alt, E., Lipkin, M., Khan, P. M. et al. (1994). A targeted chain-termination mutation in the mouse Apc gene results in multiple intestinal tumors. Proc.Natl. Acad. Sci. USA 91, 8969-8973.
Fre, S., Pallavi, S. K., Huyghe, M., Laè, M., Janssen, K.-P., Robine, S., Artavanis-Tsakonas, S. and Louvard, D. (2009). Notch and Wnt signals cooperatively controlcell proliferation and tumorigenesis in the intestine. Proc. Natl. Acad. Sci. USA 106,6309-6314.
Gartel, A. L. and Radhakrishnan, S. K. (2005). Lost in transcription: p21 repression,mechanisms, and consequences. Cancer Res. 65, 3980-3985.
Götte, M., Wolf, M., Staebler, A., Buchweitz, O., Kelsch, R., Schüring, A. N. andKiesel, L. (2008). Increased expression of the adult stem cell marker Musashi-1 inendometriosis and endometrial carcinoma. J. Pathol. 215, 317-329.
Hay, E., Faucheu, C., Suc-Royer, I., Touitou, R., Stiot, V., Vayssiere, B., Baron, R.,Roman-Roman, S. and Rawadi, G. (2005). Interaction between LRP5 and Frat1mediates the activation of the Wnt canonical pathway. J. Biol. Chem. 280, 13616-13623.
He, T.-C., Sparks, A. B., Rago, C., Hermeking, H., Zawel, L., da Costa, L. T., Morin,P. J., Vogelstein, B. and Kinzler, K. W. (1998). Identification of c-MYC as a target ofthe APC Pathway. Science 281, 1509-1512.
Imai, T., Tokunaga, A., Yoshida, T., Hashimoto, M., Mikoshiba, K., Weinmaster, G.,Nakafuku, M. and Okano, H. (2001). The neural RNA-binding protein Musashi1translationally regulates mammalian numb gene expression by interacting with itsmRNA. Mol. Cell. Biol. 21, 3888-3900.
Ishizuya-Oka, A., Shimizu, K., Sakakibara, S., Okano, H. and Ueda, S. (2003). Thyroidhormone-upregulated expression of Musashi-1 is specific for progenitor cells of theadult epithelium during amphibian gastrointestinal remodeling. J. Cell Sci. 116, 3157-3164.
Jarriault, S., Brou, C., Logeat, F., Schroeter, E. H., Kopan, R. and Israel, A. (1995).Signalling downstream of activated mammalian Notch. Nature 377, 355-358.
Kanai, R., Tomita, H., Shinoda, A., Takahashi, M., Goldman, S., Okano, H., Kawase,T. and Yazaki, T. (2006). Enhanced therapeutic efficacy of G207 for the treatment ofglioma through Musashi1 promoter retargeting of g34.5-mediated virulence. Gene Ther.13, 106-116.
Kaneko, Y., Sakakibara, S., Imai, T., Suzuki, A., Nakamura, Y., Sawamoto, K.,Ogawa, Y., Toyama, Y., Miyata, T. and Okano, H. (2000). Musashi1: an evolutionallyconserved marker for CNS progenitor cells including neural stem cells. Dev. Neurosci.22, 139-153.
Kawahara, H., Imai, T., Imataka, H., Tsujimoto, M., Matsumoto, K. and Okano, H.(2008). Neural RNA-binding protein Musashi1 inhibits translation initiation bycompeting with eIF4G for PABP. J. Cell Biol. 181, 639-653.
Kayahara, T., Sawada, M., Takaishi, S., Fukui, H., Seno, H., Fukuzawa, H., Suzuki,K., Hiai, H., Kageyama, R., Okano, H. et al. (2003). Candidate markers for stem andearly progenitor cells, Musashi-1 and Hes1, are expressed in crypt base columnar cellsof mouse small intestine. FEBS Lett. 535, 131-135.
Kengaku, M., Capdevila, J., Rodriguez-Esteban, C., De La Pena, J., Johnson, R. L.,Izpisua Belmonte, J. C. and Tabin, C. J. (1998). Distinct WNT pathways regulatingAER formation and dorsoventral polarity in the chick limb bud. Science 280, 1274-1277.
Keyoung, H. M., Roy, N. S., Benraiss, A., Louissaint, A., Jr, Suzuki, A., Hashimoto,M., Rashbaum, W. K., Okano, H. and Goldman, S. A. (2001). High-yield selectionand extraction of two promoter-defined phenotypes of neural stem cells from the fetalhuman brain. Nat. Biotechnol. 19, 843-850.
Kioussi, C., Briata, P., Baek, S. H., Rose, D. W., Hamblet, N. S., Herman, T., Ohgi, K.A., Lin, C., Gleiberman, A., Wang, J. et al. (2002). Identification of a Wnt/Dvl/[beta]-cateninrpitx2 pathway mediating cell-type-specific proliferation during development.Cell 111, 673-685.
Korinek, V., Barker, N., Moerer, P., van Donselaar, E., Huls, G., Peters, P. J. andClevers, H. (1998). Depletion of epithelial stem-cell compartments in the small intestineof mice lacking Tcf-4. Nat. Genet. 19, 379-383.
Kress, E., Rezza, A., Nadjar, J., Samarut, J. and Plateroti, M. (2008). The thyroidhormone receptor-{alpha} (TR{alpha}) gene encoding TR{alpha}1 controlsdeoxyribonucleic acid damage-induced tissue repair. Mol. Endocrinol. 22, 47-55.
Kress, E., Rezza, A., Nadjar, J., Samarut, J. and Plateroti, M. (2009). The Frizzled-related sFRP2 Gene is a target of thyroid hormone receptor {alpha}1 and activates{beta}-catenin signaling in mouse intestine. J. Biol. Chem. 284, 1234-1241.
Kress, E., Skah, S., Sirakov, M., Nadjar, J., Gadot, N., Scoazec, J.-Y., Samarut, J. andPlateroti, M. (2010). Cooperation between the thyroid hormone receptor TR[alpha]1and the WNT pathway in the induction of intestinal tumorigenesis. Gastroenterology138, 1863-1874.
Krupnik, V. E., Sharp, J. D., Jiang, C., Robison, K., Chickering, T. W., Amaravadi,L., Brown, D. E., Guyot, D., Mays, G., Leiby, K. et al. (1999). Functional andstructural diversity of the human Dickkopf gene family. Gene 238, 301-313.
Li, L., Yuan, H., Weaver, C. D., Mao, J., Farr, G. H., 3rd, Sussman, D. J., Jonkers, J.,Kimelman, D. and Wu, D. (1999). Axin and Frat1 interact with dvl and GSK, bridgingDvl to GSK in Wnt-mediated regulation of LEF-1. EMBO J. 18, 4233-4240.
Montgomery, R. K. and Breault, D. T. (2008). Small intestinal stem cell markers. J.Anat. 213, 52-58.
Moser, A. R., Pitot, H. C. and Dove, W. F. (1990). A dominant mutation that predisposesto multiple intestinal neoplasia in the mouse. Science 247, 322-324.
Jour
nal o
f Cel
l Sci
ence

3265Musashi-1 in the intestinal epithelium
Nakamura, M., Okano, H., Blendy, J. A. and Montell, C. (1994). Musashi, a neuralRNA-binding protein required for Drosophila adult external sensory organ development.Neuron 13, 67-81.
Okabe, M., Imai, T., Kurusu, M., Hiromi, Y. and Okano, H. (2001). Translationalrepression determines a neuronal potential in Drosophila asymmetric cell division.Nature 411, 94-98.
Okano, H., Kawahara, H., Toriya, M., Nakao, K., Shibata, S. and Imai, T. (2005).Function of RNA-binding protein Musashi-1 in stem cells. Exp. Cell Res. 306, 349-356.
Plateroti, M., Gauthier, K., Domon-Dell, C., Freund, J. N., Samarut, J. and Chassande,O. (2001). Functional interference between thyroid hormone receptor alpha (TRalpha)and natural truncated TRDeltaalpha isoforms in the control of intestine development.Mol. Cell. Biol. 21, 4761-4772.
Plateroti, M., Kress, E., Mori, J. I. and Samarut, J. (2006). Thyroid hormone receptor{alpha}1 directly controls transcription of the {beta}-catenin gene in intestinal epithelialcells. Mol. Cell. Biol. 26, 3204-3214.
Potten, C. S., Booth, C., Tudor, G. L., Booth, D., Brady, G., Hurley, P., Ashton, G.,Clarke, R., Sakakibara, S. and Okano, H. (2003). Identification of a putative intestinalstem cell and early lineage marker; musashi-1. Differentiation 71, 28-41.
Quaroni, A., Wands, J., Trelstad, R. L. and Isselbacher, K. J. (1979). Epithelioid cellcultures from rat small intestine. Characterization by morphologic and immunologiccriteria. J. Cell Biol. 80, 248-265.
Rodilla, V., Villanueva, A., Obrador-Hevia, A., Robert-Moreno, A. l., Fernandez-Majada, V., Grilli, A., Lopez-Bigas, N., Bellora, N. S., Alba, M. M., Torres, F. et al.(2009). Jagged1 is the pathological link between Wnt and Notch pathways in colorectalcancer. Proc. Natl. Acad. Sci. USA 106, 6315-6320.
Ruiz, S., Segrelles, C., Santos, M., Lara, M. F. and Paramio, J. M. (2004). Functionallink between retinoblastoma family of proteins and the Wnt signaling pathway in mouseepidermis. Dev. Dyn. 230, 410-418.
Sakakibara, S., Nakamura, Y., Yoshida, T., Shibata, S., Koike, M., Takano, H., Ueda,S., Uchiyama, Y., Noda, T. and Okano, H. (2002). RNA-binding protein Musashifamily: roles for CNS stem cells and a subpopulation of ependymal cells revealed bytargeted disruption and antisense ablation. Proc. Natl. Acad. Sci. USA 99, 15194-15199.
Sangiorgi, E. and Capecchi, M. R. (2008). Bmi1 is expressed in vivo in intestinal stemcells. Nat. Genet. 40, 915-920.
Siddall, N. A., McLaughlin, E. A., Marriner, N. L. and Hime, G. R. (2006). The RNA-binding protein Musashi is required intrinsically to maintain stem cell identity. Proc.Natl. Acad. Sci. USA 103, 8402-8407.
Stappenbeck, T. S., Wong, M. H., Saam, J. R., Mysorekar, I. U. and Gordon, J. I.(1998). Notes from some crypt watchers: regulation of renewal in the mouse intestinalepithelium. Curr. Opin. Cell Biol. 10, 702-709.
Sugiyama-Nakagiri, Y., Akiyama, M., Shibata, S., Okano, H. and Shimizu, H. (2006).Expression of RNA-binding protein Musashi in hair follicle development and hair cycleprogression. Am. J. Pathol. 168, 80-92.
Sureban, S. M., May, R., George, R. J., Dieckgraefe, B. K., McLeod, H. L.,Ramalingam, S., Bishnupuri, K. S., Natarajan, G., Anant, S. and Houchen, C. W.(2008). Knockdown of RNA binding protein Musashi-1 Leads to tumor regression invivo. Gastroenterology 134, 1448-1458.
Tetsu, O. and McCormick, F. (1999). [beta]-Catenin regulates expression of cyclin D1in colon carcinoma cells. Nature, 398, 422-426.
Toda, M., Iizuka, Y., Yu, W., Imai, T., Ikeda, E., Yoshida, K., Kawase, T., Kawakami,Y., Okano, H. and Uyemura, K. (2001). Expression of the neural RNA-bindingprotein Musashi1 in human gliomas. Glia 34, 1-7.
van Amerongen, R., Nawijn, M., Franca-Koh, J., Zevenhoven, J., van der Gulden, H.,Jonkers, J. and Berns, A. (2005). Frat is dispensable for canonical Wnt signaling inmammals. Genes Dev. 19, 425-430.
van der Flier, L. G., van Gijn, M. E., Hatzis, P., Kujala, P., Haegebarth, A., Stange,D. E., Begthel, H., van den Born, M., Guryev, V., Oving, I. et al. (2009). Transcriptionfactor achaete scute-like 2 controls intestinal stem cell fate. Cell 136, 903-912.
van Es, J. H., van Gijn, M. E., Riccio, O., van den Born, M., Vooijs, M., Begthel, H.,Cozijnsen, M., Robine, S., Winton, D. J., Radtke, F. et al. (2005). Notch/gamma-secretase inhibition turns proliferative cells in intestinal crypts and adenomas intogoblet cells. Nature 435, 959-963.
Wang, X. Y., Yin, Y., Yuan, H., Sakamaki, T., Okano, H. and Glazer, R. I. (2008).Musashi1 modulates mammary progenitor cell expansion through proliferin-mediatedactivation of the Wnt and Notch pathways. Mol. Cell. Biol. 28, 3589-3599.
Jour
nal o
f Cel
l Sci
ence