the regeneration of the crystalline lens lens fibers (figs. 4 and 4a)\ the early formation of...
TRANSCRIPT

The regeneration of the crystalline lens
L. S. Stone
A brief account is given of renewed interest in lens regeneration in rabbits from the epithelialcells along the equator of the lens after removal of the crystalline body from the lens cap-sule. This interest is due to the claims by some recent investigators that better lens re-generation takes place in the presence of implanted cytolyzed fetal tissue within the lenscapsule. However, overwhelming evidence has now accumulated to show that the presenceof cytolyzing cells does not favor lens regeneration over that of normal controls. The evidencefor lens regeneration after complete lentectomy is reviewed for all classes of vertebrates.After the original lens is removed from the eyes of several species of salamanders a new oneregenerates from a budding process along the pupillary margin of the dorsal iris. Except fortwo species of fishes and one species of frog-tadpoles, lens regeneration following lentectomyhas never been observed in any of the other vertebrates. Many experiments show that cellswith the potential for lens regeneration in the newt eye are confined to the dorsal part ofthe eye and are scattered over the dorsal iris and even among the retinal pigment cells inthe dorsal wall of the eye. Many experiments by the author on the eyes of adult newts arecited, dealing with the release of lens regeneration by a retinal factor and with the apparentinhibitor effect by living lens tissue.
T.-he focus of attention upon lens regen-eration is largely due to the fact that com-plete lentectomy in some members of onegroup of amphibians, namely, larval andadult urodeles, the salamanders, is fol-lowed by lens regeneration from the pupil-lary margin of the dorsal iris. This hasbeen demonstrated many times. One thenbecomes curious to know whether or notthe same phenomenon ever occurs in theeyes of other vertebrates.
Among the other groups of amphibians,the tailless anurans, there are a numberof species in which the lens does not re-generate.1 In others it is not yet proved
From the Anatomical Laboratory, Yale UniversitySchool of Medicine, Yale University, NewHaven, Conn.
Aided by Grant B-3444 from the National Insti-tute of Neurological Diseases and Blindness ofthe National Institutes of Health, United StatesPublic Health Service.
that a lens regenerates from the dorsal irisafter complete lentectomy because of thepossibility of incomplete lens removal inthe material of those who claim it. Thisgroup should be investigated morethoroughly.
Sato2 found no lens regenerated from theiris of young Japanese frog larvae afterlentectomy. However, his unusual findingwas that only when iris epithelium or reti-nal pigment epithelium was implanted intoa lentectomized eye did there appear inrare cases lens regenerates from the iso-lated epithelium. In his experiments theisolated epithelium most often gave riseto neural retina as in the case of implantedretinal pigment epithelium in the eyes ofurodeles.3' 4 Sato pointed out that the nu-tritional conditions of the lens and neuralretina are normally quite different. In-fluenced by this fact he concluded that thenutritional condition prevailing in the pig-mented epithelium may be one of the fac-
420
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932951/ on 11/05/2018

Volume 4Number 4
Regeneration of crystalline lens 421
tors which bring forth the capacity foreither lens or neural retina formation.
Sato further suggests that the factors in-hibiting lens regeneration in the tadpolesare probably the powerful proliferative ca-pacity of the iris and natural tendency ofthe internal layer of the iris to form neuralretina. These findings emphasize the im-portance of exploring further the condi-tions to be found in other anurans to un-cover any hidden capacity for lens regen-eration.
Sato5 also working on two species ofJapanese fresh water fishes has offered thebest evidence to date that lens regenera-tion takes place from the dorsal iris inthis group of vertebrates after lentectomy.
There is no evidence of lens regenera-tion after complete removal of a well-de-veloped lens in reptiles, birds, and mam-mals.
For a long time it has been known thatthe mammalian lens as well as the lens inother vertebrates is constantly suppliedby new fibers from the epithelial cellsalong the equator of the lens capsule. Coc-teau and d'Etoilles,G Middlemore,7 andMilliot8 found lens regeneration in rabbitsfollowing removal of the crystalline bodyfrom the lens capsule. Randolph9 in a fineseries of experiments confirmed these find-ings.
Stewart and 'Espenasse10 reported thattwo Russian investigators, Chanturishvili11
and Sicharulidze12 had concluded lens re-generation in rabbits was enhanced by im-planting sterile cytolyzed fetal tissue intothe lens capsule after removing the crystal-line lens. Stewart and 'Espenasse reportedthat they had repeated the Russians' ex-periments on some rabbits. After a fewweeks they examined by various tech-niques the living conditions of the regen-erating lenses in the eyes with cytolyzedtissue and compared them with controleyes with simple extracapsular lens extrac-tion. They concluded that, so far as theliving conditions could reveal, the implantsfavored larger and better lens regeneratesthan in those of the controls. A histological
study of regenerated rabbit lenses byStewart13 revealed that larger lenses withmore normal optic density regenerated ifcytolyzed fetal tissue was implanted afterextracapsular lens extraction.
Binder and associates14 and Agarwal andco-workers15'16 reported that implantedcytolyzed tissue did not induce larger lensregenerates in the rabbit. Agarwal, and as-sociates,17 in a few experiments on mon-keys, suggested that the implants mighthave induced larger lens regenerates. Pet-tit18 also studied lens regeneration and hesuggested that the implants with pro-longed irritation might have induced largerlens regenerates in rabbits.
My impression of the results of all ofthese investigators is that the larger lensregenerates depend upon the condition ofthe lens capsule. If it collapses, little or nolens tissue regenerates. If it remains openlarger lenses are regenerated. Perhaps inmany cases the introduction of cytolyzedtissue, which nearly always later degen-erates, helps keep the capsule open andhence in counting the number of cases inan experimental group there may appearlarger regenerates in those which havethe implants.
In studies of embryonic lens develop-ment the presence of a cytolyzed cell ortwo may have occasionally been cited. Ibelieve they have no significance withrespect to lens formation. A few cytolyzingcells can often be seen in various parts ofdeveloping embryos.
Now to get back to the interesting prob-lems presented by the phenomenon of lensregeneration from the dorsal iris of certainsalamander eyes first revealed in. adultTriturus newts by Colucci19 in 1891. SinceI have worked extensively with the Ameri-can adult newt, Triturus viridescens, Ishall deal largely with the results fromseveral hundred of my own experimentsover many years. A review of the contri-butions of other investigators not men-tioned in the text will be found in the lit-erature cited, especially in the fine reviewby Reyer.20
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932951/ on 11/05/2018
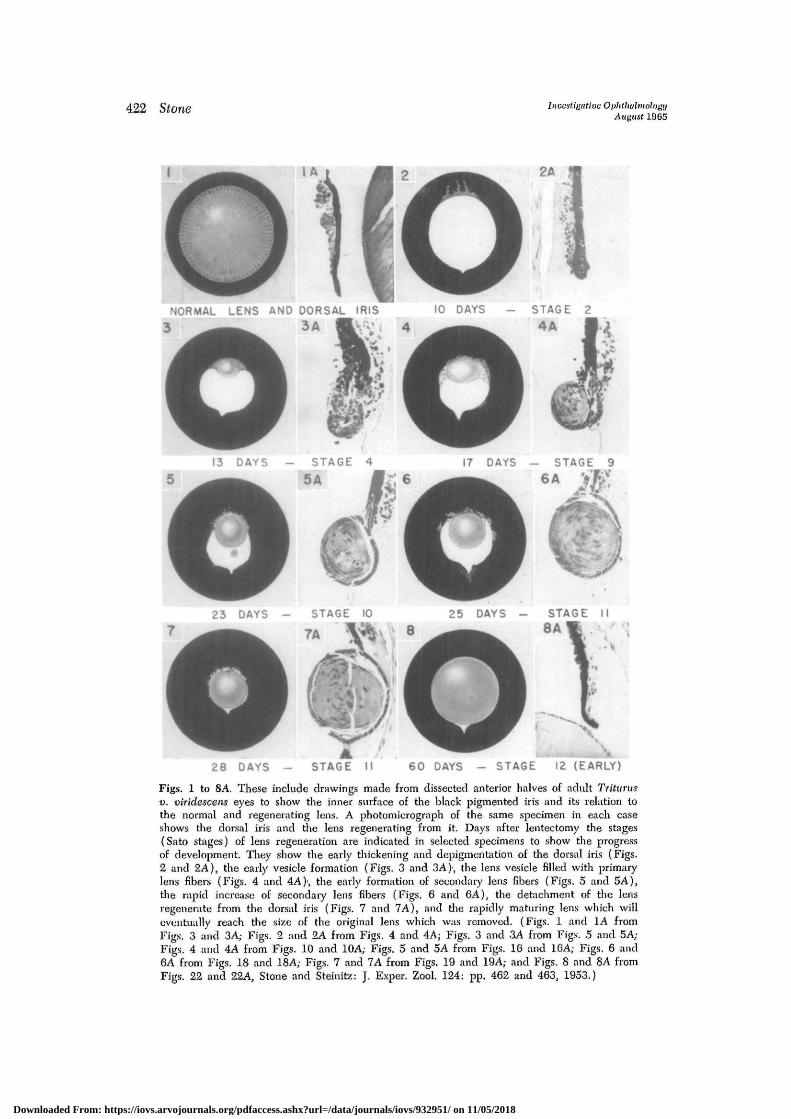
422 Stone Investigative OphthalmologyAugust 1965
10 DAYS - STAGE 2NORMAL LENS AND DORSAL IRIS
3A
17 DAYS — STAGE
23 DAYS - STAGE 10
28 DAYS - STAGE II 60 DAYS - STAGE 12 (EARLY)
Figs. 1 to 8A. These include drawings made from dissected anterior halves of adult Tritunisv. viridescens eyes to show the inner surface of the black pigmented iris and its relation tothe normal and regenerating lens. A photomicrograph of the same specimen in each caseshows the dorsal iris and the lens regenerating from it. Days after lentectomy the stages(Sato stages) of lens regeneration are indicated in selected specimens to show the progressof development. They show the early thickening and depigmentation of the dorsal iris (Figs.2 and 2A), the early vesicle formation (Figs. 3 and 3A), the lens vesicle filled with primarylens fibers (Figs. 4 and 4A)\ the early formation of secondary lens fibers (Figs. 5 and 5A),the rapid increase of secondary lens fibers (Figs. 6 and 6A), the detachment of the lensregenerate from the dorsal iris (Figs. 7 and 7A), and the rapidly maturing lens which willeventually reach the size of the original lens which was removed. (Figs. 1 and 1A fromFigs. 3 and 3A; Figs. 2 and 2A from Figs. 4 and 4A; Figs. 3 and 3A from Figs. 5 and 5A;Figs. 4 and 4A from Figs. 10 and 10A; Figs. 5 and 5A from Figs. 16 and 16A; Figs. 6 and6A from Figs. 18 and 18A; Figs. 7 and 7A from Figs. 19 and 19A; and Figs. 8 and 8A fromFigs. 22 and 22A, Stone and Steinitz: J. Exper. Zool. 124: pp. 462 and 463, 1953.)
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932951/ on 11/05/2018

Volumd 4Number 4
Regeneration of crystalline lens 423
Although the rate of lens regenerationfrom the dorsal iris may vary in some in-dividuals,-1 Figs. 1 to 8A illustrate in afew selected specimens what takes placein the iris after the original lens is excised.
The dorsal iris which was quite thin(Fig. 1A) begins to be depigmented andto thicken along the mid-dorsal margin ofthe pupil within 10 days (Figs. 2 and 2A).Cell proliferation takes place and a vesicledevelops during the second week (Figs. 3and 3A). The vesicle soon becomes filledwith primary lens fibers (Figs. 4 and 4A).Then the lens enlarges with the aquisitionof secondary lens fibers (Figs. 5 to 6A).Within 4 weeks the lens regenerate be-comes detached from its source of origin(Figs. 7 and 7A). During the secondmonth the lens increases rapidly in size bythe growth of many secondary lens fibers(Figs. 8 and 8A). The dorsal iris takes onthe normal appearance again. This growthcontinues for several months until the lensregenerate attains the size of the originalone.
The cells from which the lens regen-erates appear to lie only in the dorsal re-gion of the eye where they are widelyscattered. The ventral half of the iris pos-sesses no lens-forming cells.22 In the irisitself potential lens-forming cells are notconfined to the pupillary border. This canbe shown in the following experiments.
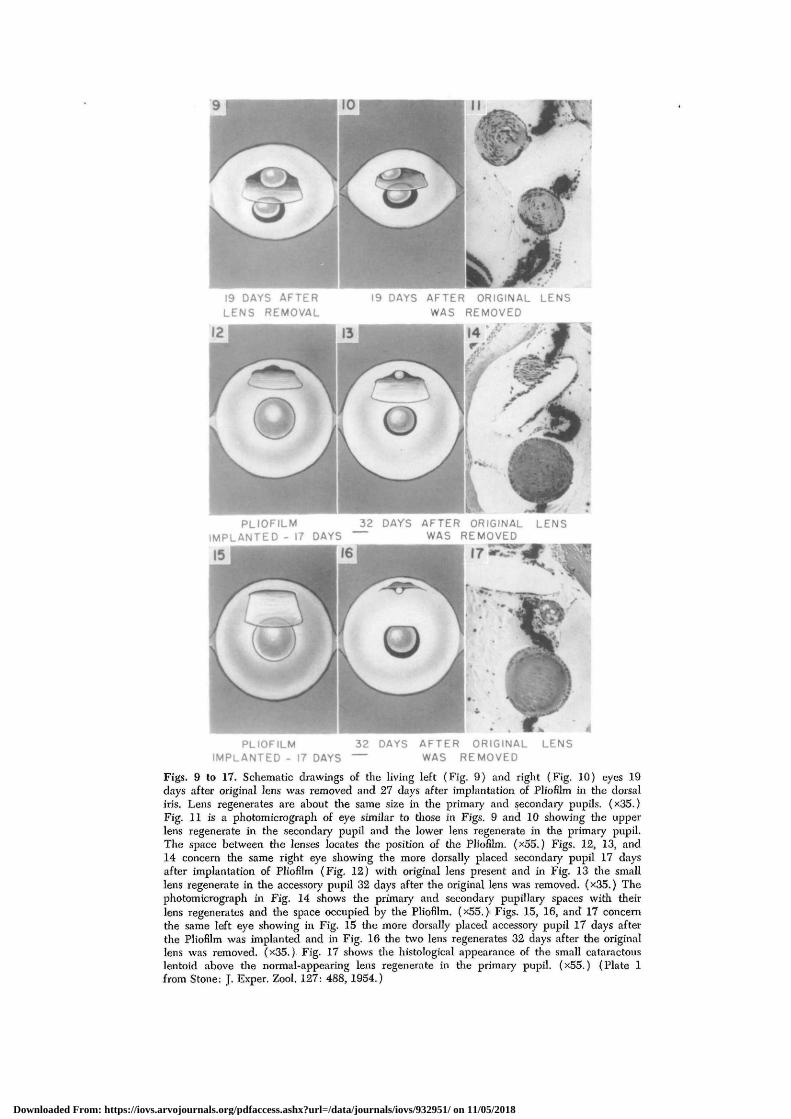
An incision is made in the cornea so thata comeal flap can be elevated to exposethe dorsal iris. Slits are then made in thelatter in various regions and into themsmall pieces of thin Pliofilm membranesare inserted to make permanent accessoiypupillary openings. The corneal flap is thenreplaced and allowed to heal. Later thelens is removed and then a lens regener-ates not only from the margin of the origi-nal pupil but in the accessory one as well.This is shown in 2 cases (Figs. 9 and 10).The histological appearance of the twolenses is shown in Fig. 11. The space be-tween the two was occupied by the Plio-film.
In Fig. 12 the accessory opening was
more dorsal in position and as long as theoriginal lens was present no lens regen-erated in the new opening. Fig. 13 showsthat, in the same eye, 32 days after lentec-tomy, a small lens had regenerated fromthe margin of the iris in the accessory pu-pil. A large lens regenerate lies in the nor-mal pupil as well. The photomicrographof the same eye in Fig. 14 shows the twolenses and the space occupied by the Plio-film between them.
In Figs. 15, 16, and 17 a similar sequenceof events is shown in another eye in whichthe accessory opening is near the borderof the retina. After lentectomy a small lensformed from the border of the iris in theaccessory opening below the Pliofilm mem-brane (Fig. 16). It is shown in Fig. 17 asa degenerating lentoid below the space oc-cupied by the implanted Pliofilm. A largenormal lens has regenerated in the primarypupil.
If permanent openings are made by in-serting small pieces of Pliofilm into thedorsal retinal wall of the eye, small lenseswill develop from retinal pigment cells atthe periphery of the openings.23 If piecesof retinal pigment epithelium are isolatedfrom the dorsal wall of the eyes of theadult newt and implanted into lentecto-mized eyes, they will give rise not onlyto neural retina tissue but also to smalllenses.3' 4
If all of the dorsal iris is removed, in-cluding some of the peripheral retinal wall,the dorsal iris regenerates from the retinalpigment cells. The regenerating iris givesrise to a lens regenerate as soon as theoriginal lens is removed.24 This is not sur-prising, for the regenerated iris was de-rived from retinal pigment cells whichwere shown in the experiment mentionedabove to possess cells with a capacity toform lenses.
The mid-central third of the dorsal irisis the most potent region for lens forma-tion. If it is excised, rotated 180 degrees,even inside out (Fig. 18, A), a lens willdevelop in a lentectomized eye (Fig. 18,B) from that portion of the iris now at the
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932951/ on 11/05/2018
19 DAYS AFTERLENS REMOVAL
19 DAYS AFTER ORIGINAL LENSWAS REMOVED
PLIOFILM 32 DAYS AFTER ORIGINAL LENSIMPLANTED - 17 DAYS WAS REMOVED
PLIOFILMIMPLANTFP
32 DAYS AFTER ORIGINAL LENS— WAS REMOVED
Figs. 9 to 17. Schematic drawings of tlie living left (Fig. 9) and right (Fig. 10) eyes 19days after original lens was removed and 27 days after implantation of Pliofilm in tlie dorsaliris. Lens regenerates are about tlie same size in tlie primary and secondary pupils. (x35.)Fig. 11 is a photomicrograph of eye similar to those in Figs. 9 and 10 showing tlie upperlens regenerate in the secondary pupil and die lower lens regenerate in tlie primary pupil.The space between the lenses locates the position of the Pliofilm. (x55.) Figs. 12, 13, and14 concern tlie same right eye showing the more dorsally placed secondary pupil 17 daysafter implantation of Pliofilm (Fig. 12) with original lens present and in Fig. 13 the smalllens regenerate in tlie accessory pupil 32 days after the original lens was removed. (x35.) Thephotomicrograph in Fig. 14 shows the primary and secondary pupillary spaces with theirlens regenerates and the space occupied by tlie Pliofilm. (x55. )• Figs. 15, 16, and 17 concernthe same left eye showing in Fig. 15 tlie more dorsally placed accessory pupil 17 days afterthe Pliofilm was implanted and in Fig. 16 tlie two lens regenerates 32 days after the originallens was removed. (x35.) Fig. 17 shows the histological appearance of the small cataractouslentoid above the normal-appearing lens regenerate in the primary pupil. (x55.) (Plate 1from Stone: J. Exper. Zool. 127: 488, 1954.)
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932951/ on 11/05/2018

Volume 4Number 4
Regeneration of crystalline lens 425
Fig. 18. Schematic drawings showing the mid-dorsal segment of the dorsal iris excised androtated 180 degrees and reimplanted inside out (A). Original lens was removed later (B).A lens regenerated eventually from the reversed pupillary margin (C and D) with lens fiberpole normally oriented toward the neural retina. Occasionally a lens regenerated from poten-tial lens-forming host iris on either side of the graft (D). (Fig. 4 from Stone: Anat. Rec.120: 611, 1954.)
pupillary margin (Fig. 18, C). In some in-stances a lens regenerate will also be re-leased on either side of the graft (Fig. 18,D). Two separated potential lens-formingregions can also act independently if apiece of ventral non-lens forming iris issubstituted for mid-dorsal iris.25
Wounds such as several slits made inthe dorsal iris do not initiate lens regenera-tion.-0 It follows after a complete removalof the original lens or a fully regeneratedone.
When the lens is removed from an eye,the loosened lens sometimes falls out ofsight into the ventral vitreous chamber.After some delay a lens, will regeneratefrom the dorsal iris and the dislocated lenswill be in various stages of cataract forma-tion. The cataractous lenses may be losingany inhibitory effect they may have pos-sessed.
Eguchi-7 noticed in the Japanese adultnewts that, when the ventrally displacedlenses in rhft niinilUirv cnafp W»r»amo r>n+n
performed. In a few instances when thedislocated lens remained intact a lens re-generated from the dorsal iris. He sug-gested that to inhibit lens regeneration anintact lens must be in contact with thedorsal iris.
Reyer,2S working on the eyes of larvalnewts, reports a few cases in which theimplanted intact lens was displaced ven-trally in the pupil with the result that alens regenerated from the dorsal iris onlyas far as a depigmented lens vesicle. Whenthe implanted lens remained intact and oc-cupied the normal position, it completelyblocked regeneration. An understanding ofthe role the presence of a lens plays whenlens regeneration is inhibited must awaitfurther study.
I— found that regenerating lenses im-planted in a lentectomized eye did not in-hibit lens regeneration until the regenerat-ing lens was 25 to 28 days old and detach-ing itself from the dorsal iris. It then pos-
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932951/ on 11/05/2018

426 Stone Investigative OphthalmologyAugust 1965
younger smaller regenerating lens andfound inhibition of lens regeneration. Heconcluded that a certain volume of im-planted lens tissue must be attained to pre-vent lens regeneration and that it was notnecessarily the age and quality of the im-planted regenerating lens which was in-volved.
Since the presence of a large lens, evenone from another species which does notregenerate lenses,30 is associated with inhi-bition of lens regeneration, I concluded thatsomething might be given off by the lensinto the aqueous humor in which the dor-sal iris was bathing. This appeared to besubstantiated by beginning on the secondday and injecting daily for a long time intolentectomized eyes aqueous humor fromeyes containing a normal lens.31' 32 Thebest eyes carried for many weeks failedto regenerate a lens while daily injectionsof saline solution failed to inhibit lens re-generation.
Takano and co-workers33 repeated thisexperiment using the Japanese newt Tri-turus pijrrhogaster beginning at the secondand seventh days after lentectomy. Theseeyes were followed for 20 to 35 days afteroperation in a few cases. There were alsoinjections of various extracts not concernedwith these comparisons. They often found,as I did, that the iris and retina were in-jured by this method. They reported lensregeneration as in controls in their bestcases and concluded that aqueous humorfrom normal eyes did not inhibit lens re-generation by their methods.
One has to be quite skilled in this tech-nique to control all phases of it, particu-larly in preventing leakage of the injectedfluid. It is essential that an amount ofaqueous humor be removed from the eyebefore an equal amount from the normaleye is injected. Otherwise the increasedintraocular pressure will be accompaniedby leakage of the injected material andinvalidate the experiment.
An easier and safer approach to thisproblem was devised.3" A right eye was ex-cised and a broad round opening cut in
its nasal half. It was then implanted intoa wound in the head so that the openingfused with a similar opening made in thetemporal half of the host's right eye. Botheye units were lentectomized. A number ofcases showed perfectly fused double eyeunits with a large common oval pupillaryspace in which both dorsal irises andlenses were bathed by the same aqueoushumor. After several weeks, two largelens regenerates were seen and one wasremoved. Animals killed 42 days latershowed that a lens had regenerated fromthe lentectomized unit in the presence ofthe other lens.
However, the fused eye units containedalmost double the amount of the neuralretina of the normal eye. It is known thata retinal factor initiates lens regeneration,and it is possible that the effect of astronger retinal factor may have overcomeany inhibiting capacity that the remaininglens unit might have possessed.
To clarify this, in a number of adultnewts the ventral half of the right eye wascut away and the dorsal half of a donoreye grafted to it. The lenses of both halveswere removed. Many of these half dorsalunits fused perfectly and in several weeksgave the appearance of a single normaleye. Two large lenses regenerated, onefrom each dorsal iris. After many weeksone large lens was removed. Those eyesnot injured by the second operationshowed no lens regeneration from thelentectomized half in the presence of theremaining lens. Is it possible that the in-hibiting influence involving the lens wasemanating from something present in theaqueous humor and potent enough toovercome in some way the retinal factor?
The results of an earlier experiment25
also bear upon the relation of lens andaqueous humor when lens regeneration isinhibited. One can insert a round thin discof Pliofilm through a corneal slit isolatingthe intact lens below it from the dorsal irisabove (Fig. 19, A). The disc sinks intothe retina medially and fits tightly againstthe cornea in front. This seals off the
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932951/ on 11/05/2018

Volume 4Number 4
Regeneration of crystalline lens 427
Fig. 19. Schematic representation of operation andresult showing implanted thin membrane of Plio-film isolating dorsal iris from lens and aqueoushumor bathing it (A). Lens regeneration is releasedfrom dorsal iris (B). (From Stone: Am. J. Ophth.36: 36, 1953.)
stretched dorsal iris above in a chamberby itself so that the aqueous humor aboveis separated from that around the intactlens below. A lens regenerated from theisolated dorsal iris (Fig. 19, B). Possiblythis is a result of the iris being removedfrom an inhibitory effect of something inthe aqueous humor emanating from theintact lens.
The role of the retina in lens regenera-tion can be established as follows: Whenan eye has been lentectomized and theneural retina has also been removed, lensregeneration is retarded until the retinais regenerating from the retinal pigmentcells.21
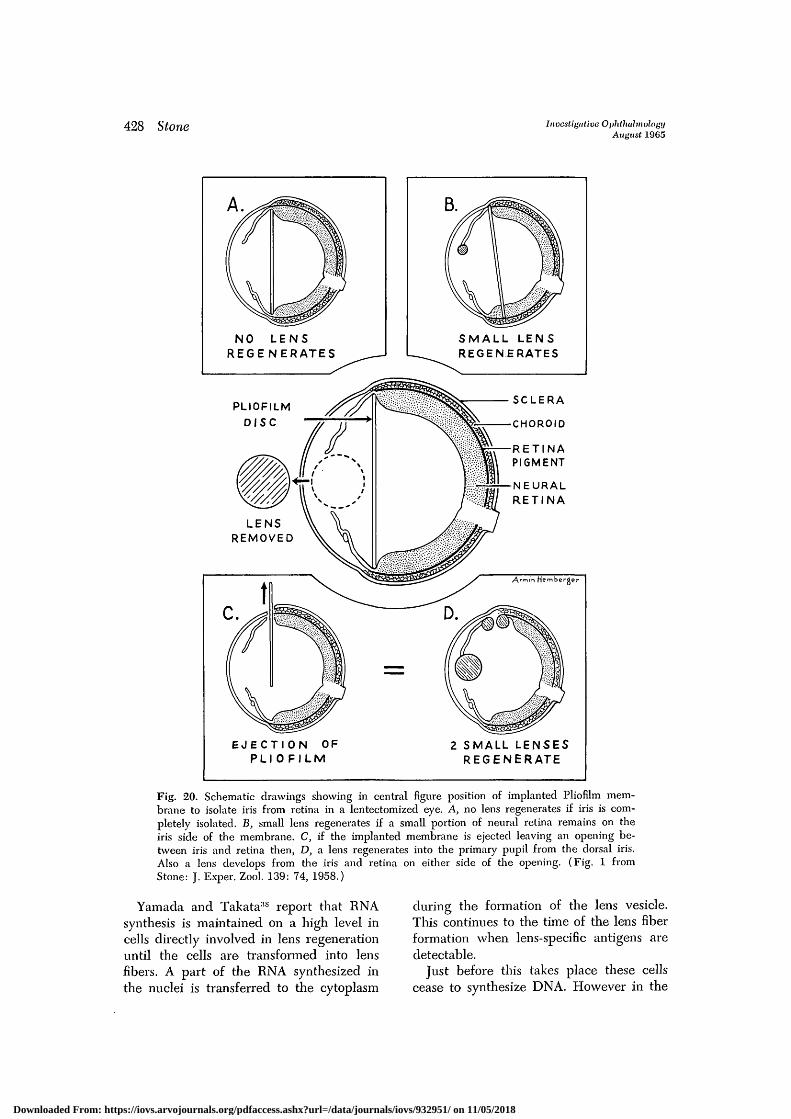
Lens regeneration is inhibited if the irisis isolated for a long time by the implant-ing of a plastic membrane between the irisand the neural retina (Fig. 20, A) in alentectomized eye. If there is a smallamount of neural retina on the iris sideof the chamber (Fig. 20, B) a small lensregenerates. This seems to indicate thatthe size of the lens regenerate may be de-termined by the amount of neural retina towhich the iris is exposed.
If the implanted membrane is extrudedand a small opening remains permanent atthe ora serrata, a small lens develops fromthe ciliary margin of the iris and anotherfrom the retinal pigment cells. When thereis failure to isolate the iris and retina, alens regenerates at the normal rate fromthe dorsal iris (Fig. 20, C and D).
When an implanted graft in a lentecto-mized eye partly encloses the pupillarymargin of the dorsal iris and shields it
from the neural retina no lens regenerates.3'1
The effect is the same as in those. experi-ments where the iris was isolated.
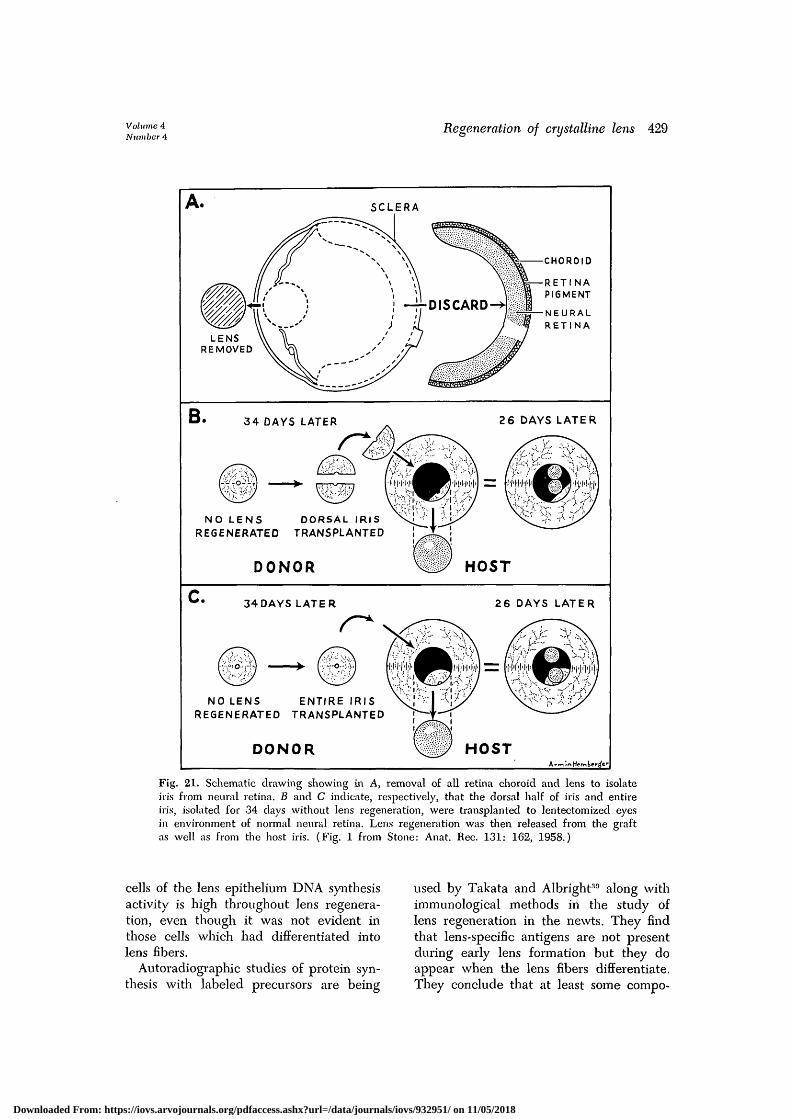
The importance of the presence of theneural retina during the release of lensregeneration can be demonstrated in an-other experiment.35 If the entire neuralretina and retina pigment cells in a lentec-tomized eye are excised, no neural retinacan regenerate (Fig. 21) because of thelack of the retinal pigment epithelium.The eye becomes small but the survivingiris does not regenerate a lens. If, later,the dorsal iris (Fig. 21, B) or the entireiris (Fig. 21, C) is transplanted to a len-tectomized eye in the environment of anormal neural retina, lens regenerationtakes place from the graft as well as fromthe host iris.
Takano and associates33 mentioned thatwhen retinal extracts were injected intolentectomized eyes the lens regeneratesseemed to surpass the controls slightlyboth in size and in differentiation.
There is also evidence that the neuralretina influences the formation of themediolateral axis of the lens. The fiber-forming pole of the regenerating lens wasalways directed toward the retina eventhough the mid-dorsal iris was rotated180 degrees inside out (Figs. 4 and 13,D).2G
In the experiment mentioned abovewhere there was a small segment of retinain the isolated chamber on the iris side ofthe implanted plastic membrane, the lens-fiber pole of the small regenerate usuallypointed directly toward the island of reti-nal tissue. This is an area of research whichshould be explored extensively and cor-related with histochemical techniques.
Takata3G working with adult newts andOgawa37 with larval newts observed thatRNase-sensitive basophilic substances andalkaline phosphatase increase markedlyduring rapid cell proliferation in the lensregenerate. These substances decrease asdifferentiation of the lens fibers takes placeand prior to the completion of regenera-tion.
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932951/ on 11/05/2018
428 Stone Investigative OphthalmologyAugust 1965
NO LENSREGENERATES
B.
S M A L L LENSREGENERATES
PLIOFILMDISC
E J E C T I O N OFPLI O F I L M
2 SMALL LENSESREGENERATE
Fig. 20. Schematic drawings showing in central figure position of implanted Pliofilm mem-brane to isolate iris from retina in a lentectomized eye. A, no lens regenerates if iris is com-pletely isolated. B, small lens regenerates if a small portion of neural retina remains on theiris side of the membrane. C, if the implanted membrane is ejected leaving an opening be-tween iris and retina then, D, a lens regenerates into the primary pupil from the dorsal iris.Also a lens develops from the iris and retina on either side of the opening. (Fig. 1 fromStone: J. Exper. Zool. 139: 74, 1958.)
Yamada and Takata38 report that RNAsynthesis is maintained on a high level incells directly involved in lens regenerationuntil the cells are transformed into lensfibers. A part of the RNA synthesized inthe nuclei is transferred to the cytoplasm
during the formation of the lens vesicle.This continues to the time of the lens fiberformation when lens-specific antigens aredetectable.
Just before this takes place these cellscease to synthesize DNA. However in the
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932951/ on 11/05/2018
Volume 4Number 4
Regeneration of crystalline lens 429
SCLERA
-CHOROID
-RETINAPIGMENT
-NEURALRETINA
LENSREMOVED
B. 34 DAYS LATER
NO LENS DORSAL IRISREGENERATED TRANSPLANTED
DONOR
2 6 DAYS LATER
HOST
C. 34 DAYS LATER 26 DAYS LATER
NO LENS ENTIRE IRISREGENERATED TRANSPLANTED
DONOR
Fig. 21. Schematic drawing showing in A, removal of all retina choroid and lens to isolateiris from neural retina. B and C indicate, respectively, that the dorsal half of iris and entireiris, isolated for 34 days without lens regeneration, were transplanted to lentectomized eyesin environment of normal neural retina. Lens regeneration was then released from the graftas well as from the host iris. (Fig. 1 from Stone: Anat. Rec. 131: 162, 1958.)
cells of the lens epithelium DNA synthesisactivity is high throughout lens regenera-tion, even though it was not evident inthose cells which had differentiated intolens fibers.
Autoradiographic studies of protein syn-thesis with labeled precursors are being
used by Takata and Albright30 along withimmunological methods in the study oflens regeneration in the newts. They findthat lens-specific antigens are not presentduring early lens formation but they doappear when the lens fibers differentiate.They conclude that at least some compo-
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932951/ on 11/05/2018

430 Stone Investigative OphthalmologyAugust 1965
nents of each of the major groups of lensprotein, alpha, beta, and gamma crystal-lins, make their appearance together in theyoungest primary lens fibers. This seemsto be a critical stage in the progress of lensregeneration. It may be possible that byexpanding these techniques in furtherstudies some light may be shed on themechanisms involved in the inhibition andrelease of lens regeneration.
A brief report by Karasaki40 has addedsome new knowledge about the ultrastruc-ture of the regenerating lens through theelectron microscope. Electron microscopicstudies on the regenerating lens revealedthat four days after lentectomy the medio-dorsal iris cells show an increase in thenumber of primary nucleoli. The nucleithen enlarge and become spherical. Promi-nent nucleoli appear which have a granu-lar cortex surrounded by a fibrous core.At this time the pigment granules of thedepigmenting iris cells are extruded intothe intercellular spaces where they aretaken up by leukocytes. The cytoplasticreticulum gradually disintegrates. Thenthe nucleoli become less prominent andthe ril^osomes increase in number in thecytoplasm. The cytoplasm of the lens fibersthen acquires fibrous elements of low den-sity corresponding to that of normal lensfibers.
Conclusion
Lens regeneration from the dorsal irisafter the original lens is removed is proba-bly limited to a few species among thelower vertebrates. This phenomenon is nowknown to occur in certain species of sala-manders and apparently in two species offishes, but in no other vertebrates.
Although lens regeneration from thepupillary margin of the dorsal iris has notyet been found in any anuran so far ex-amined, there is evidence in the tadpolestage of at least one of the anurans thatthe capacity for lens formation is appar-ently hidden in the pigment epithelium ofthe iris and retina.2 It has been revealedthat these pigment cells have a dual ca-
pacity to form either retinal tissue or occa-sionally lenses when transplanted into alentectomized eye. It may be possible thatin this case different nutritional conditionsin the environment or in the graft itselfdirect the type of tissue which emergesfrom these pigment cells.
Since in anurans so far examined lensregeneration does not take place from thedorsal iris as it does in certain newts, itremains to be shown by further experi-ments whether or not these findings2 arewidespread among other species of anu-rans. In fact, the same tests should be ap-plied to early stages in the growth of theeyes of other vertebrates in which a lensdoes not regenerate from the pupillarymargin of the dorsal iris after the originallens is removed.
In the experiments which are reviewedhere on the eyes of the newt there weremany which showed that the potentiallens-forming cells exist in various regionsof the dorsal half of the iris and also inthe retinal pigment cells of the dorsal reti-nal wall. It is important to note that inall other cases cited where potential lens-forming cells are present they are also lo-cated in the dorsal half of the eye. Fromearlier experiments of Sato'11 with rotatedembryonic eyes of the newt it is apparentthat the lens-forming area is polarized inthe dorsal half of the eye in an early opticvesicle stage under the influence of theventral fetal fissure. Previous to this, theentoderm of the archenteron and the pre-chordal mesoderm can already induce lensformation in the neighboring surface ecto-derm.42' 43 Then when the optic vesiclecontacts the surface ectoderm it carries onits part in the induction and differentiationof the lens. It would be interesting to knowthrough further experiments whether theinductive influence is centered in the dor-sal part of the optic vesicle at the timewhen its future lens-forming area is polar-ized.
However, the existence of an embryoniclens in the newt eye in the first place hasno role or influence in the development of
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932951/ on 11/05/2018

Volume 4Number 4
Regeneration of crystalline lens 431
the mechanism which incorporates thecapacity for lens formation later in thedorsal iris. Reyer44 has shown this con-clusively in his experiments on newt em-bryos where, in the absence of the em-bryonic lens, a lens regenerates from thedorsal iris as soon as the latter is formed.
In a previous part of this text many ex-periments on newt eyes were cited to showthat some retinal factor under favorableconditions induces lens regeneration fromthe dorsal iris. It may be a chemical factorbut the mechanism involved is unknown.
The role the presence of a living lensplays in inhibiting lens regeneration is stillnot clear. It is not certain whether undernormal conditions the lens shields the dor-sal iris from the retinal factor which is re-sponsible for releasing lens regeneration,whether it absorbs the retinal factor, orwhether the lens gives off some inhibitingsubstance in the aqueous humor to neu-tralize the retinal influence. Further ex-periments must be devised to find the an-swers to these questions.
REFERENCES1. Stone, L. S., and Sapir, P.: Experimental
studies on the regeneration of the lens in theeye of anurans, urodeles, and fishes, J. Exper.•Zool. 85: 71, 1940.
2. Sato, T.: t)ber die Ursachen des Ausbleibensder Linsen-Regeneration und zugleich iiberdie Linsen-Bildende Fahigkeit des Pigment-epithels bei den Anuren, Arch. Entwcklngs-mechn. Organ. 146: 487, 1953.
3. Sato, T.: t)ber die linsenbildende Fahigkeitdas Pigmentepithels bei Diemijctylus pyr-rhogaster. I. Pigmentepithel aus dorsalenAugenbereich, Embryologie 1: 21, 1951.
4. Stone, L. S., and Steinitz, H.: Regenerationof neural retina and lens from retina pigmentcell grafts in adult newts, J. Exper. Zool. 135:301, 1957.
5. Sato, T.: t)ber die Linsen-Regeneration beiden Cobitiden Fischen Misgurnus Anguil-licaudatus (Cantor), Embryologica 6: 251,1961.
6. Cocteau, I. T., and J. I. Leroy d'fitoilles:Experiences relatives a la reproduction ducrystallin, J. Physiol. Exper. 7: 30, 1827.
7. Middlemore, R.: On the reproduction of thecrystalline lens, London M. Gaz. 10: 344,.1832.
8. Milliot, V.: Experiment of the restoration of
a normal crystalline lens in some mammalsafter its removal, J. Anat. et physiol., Paris8: 1, 1872.
9. Randolph, R. L.: The regeneration of thecrystalline lens, Johns Hopkins Hosp. Rep. 9:•237, 1900.
10. Stewart, D. S., and 'Espinasse, P. C : Re-generation of the lens of the eye in the rab-bit, Nature 183: 1815, 1959.
11. Chanturishvili, P. S.: The role of ectodermin development of the crystalline lens, Tr.Ophth. Soc. United Kingdom 78: 411, 1958.
12. Sicharulidze, T. A.: The substitution of em-bryonal skin ectoderm for the lens of a rabbit,Bull. Acad. Sc. Georg. S.S.R. 14: 337, 1956.
13. Stewart, D. S.: Further observations on re-generated crystalline lenses in rabbits with'special reference to their refractive qualities,Trans. Ophth. Soc. 80: 357, 1960.
14. Binder, H. F., Binder, R. F., Wells, A. H.,and Katz, R. L.: Influence of embryonic im-plants upon lens regeneration in rabbits, Brit.J. Ophth. 46: 416, 1962.
15. Agarwal, L. P., Angra, S. K., Khosla, P.K., and Taudon, H. D.: Lens regeneration inmammals. I. Rabbits (after extracapsular ex-traction), Oriental Arch. Ophth. 2: 1, 1964.
16. Agarwal, L. P., Angra, S. K., Khosla, P.K., and Taudon, H. D.: Lens regeneration inmammals. III. Rabbits (after intracapsularlens extraction), Oriental Arch. Ophth. 2: 95,1964.
17. Agarwal, L. P., Angra, S. K., Khosla, P.•K., and Taudon, H. D.: Lens regeneration inmammals. II. Monkeys (after extracapsularextraction), Oriental Arch. Ophth. 2: 47, 1964.
18. Pettit, T. H.: A study of lens regeneration inthe rabbit, INVEST. OPHTH. 2: 243, 1963.
19. Colucci, V. I.: Sulla regenerazione parzialedell'occhio nei tritone. Istogenese e svilluppo.Studio sperimentale, Mem. r. Accad. Bolognaisez. sci. nat. (Ser. 5> 1: 167, 1891.
20. Reyer, R. W.: Regeneration of the lens in theamphibian eye, Quart. Rev. Biol. 29: 1, 1954.
21. Stone, L. S., and Steinitz, H.: The regenera-tion of lenses in eyes with intact and re-generating retina in adult Triturus v. virides-cens, J. Exper. Zool. 124: 435, 1953.
22. Stone, L. S.: An experimental study of theinhibition of release of lens regeneration inadult eyes of Triturus viridescens viridescens,J. Exper. Zool. 121: 181, 1952.
23. Stone, L. S.: Further experiments on lensregeneration from retina pigment cells in adultnewt eyes, J. Exper. Zool. 126: 75, 1957.
24. Stone, L. S.: Regeneration of the iris and lensfrom retina pigment cells in adult newt eyes,J. Exper. Zool. 129: 505, 1955.
25. Stone, L. S.: An experimental analysis of lensregeneration, Am. J. Ophth. 36: 31, 1953.
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932951/ on 11/05/2018

432 Stone Investigative OphthalmologyAugust 1965
26. Stone, L. S.: Further experiments in lens re-generation in eyes of the adult newt Triturusv. viriclescens, Anat. Rec. 120: 599, 1954.
27. Eguchi, G.: The inhibitory effect of the in-jured and displaced lens on the lens forma-tion in Triturus larvae, Embryologia vol. 6,Mangold Festschrift, p. 13, 1961.
28. Reyer, R. W.: Regeneration in the amphibianeye, in Rudmick, D., editor: Regeneration,New York, 1962, Ronald Press, pp. 211-265.
29. Frost, D.: Inhibition of lens regeneration byimplanted lenses in eyes of adult newtTriturus viriclescens, Devel. Biol. 3: 516,1961.
30. Stone, L. S.: Heteroplastic lens grafts relatedto factors inhibiting lens regeneration, Proc.Soc. Exper. Biol. & Med. 60: 10, 1945.
31. Stone, L. S., and Vultee, J. H.: Inhibitionand release of lens regeneration in the dorsaliris of Triturus v. viriclescens, Anat. Rec. 103:144, 1949.
32. Stone, L. S.: Experiments dealing with therole played by the aqueous humor and retinalin lens regeneration of adult newts. J. Exper.Zool. 153: 197, 1963.
33. Takano, K., Yoshida, Y., Ohash, T., Ogasa-wara, T., Takeuche, A., Masake, H., Miyazaki,A., and Mikami, Y.: Experimental analysisof the effect of lens upon the Wolffian lensregeneration in adults of the newt Trituruspyrrhogaster, Mie M. J. VII: 257, 1957.
34. Stone, L. S.: Unpublished data.35. Stone, L. S.: Inhibition of lens regeneration
in newt eyes by isolating the dorsal iris fromthe neural retina, Anat. Rec. 131: 151, 1958.
36. Takata, K.: Ribonucleic acid and lens-regen-eration, Experientia VIII: 217, 1952.
37. Ogawa, T.: Studies on the lens regenerationin the larval newt lacking pigment in theeye, Embryologia 7: 95, 1962.
38. Yamada, T., and Takata, C : An autoradio-vgraphic study of protein synthesis in regen-
erative tissue transformation of iris into lensin the newt, Devel. Biol. 8: 358, 1963.
39. Takata, K., and Albright, J. F.: Lens antigensin Wolffian regeneration, Abst. Conference onLens Differentiation held in Oak Ridge Na-tional Laboratory, March 20-21, 1964.
40. Karasaki, S.: Ultrastructural study of Wolf-fian lens regeneration in the adult newt, Abst.Conference on Lens Differentiation held inOak Ridge National Laboratory, March 20-21,1964.
41. Sato, T.: Beitrage zur Analyse der Wolff'schen Linsenregeneration. II, Arch. Entwck-lngsmechn. Organ. 130: 19, 1933.
42. Reyer, R. W.: Studies in lens induction inAmblystoma punctatum and Triturus v. viri-clescens. II. Transplants of prospective headectoderm, J. Exper. Zool. 139: 137, 1958.
43. Reyer, R. W.: Differentiation and growth ofthe embryonic nose, lens and corneal anlagenimplanted into the larval eye or dorsal fin in'Amblystoma punctatum, J. Exper. Zool. 151:123, 1962b.
44. Reyer, R. W.: An experimental study of lensregeneration in Triturus v. viriclescens. II.Lens development from the dorsal iris in theabsence of the embryonic lens, J. Exper. Zool.113: 317, 1950.
45. Stone, L. S.: Lens regeneration in secondarypupils experimentally produced in eyes of theadult newt, Triturus v. viriclescens, J. Exper.Zool. 127: 463, 1954.
46. Stone, L. S., and Gallagher, S. B.: Lens re-generation restored to iris membranes whengrafted to neural retina environment aftercultivation in vitro, J. Exper. Zool. 139: 247,1958.
47. Stone, L. S.: Lens regeneration in adult newteyes related to retina pigment cells and theneural retina factor, J. Exper. Zool. 139: 69,1958.
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932951/ on 11/05/2018