the shb adapter protein is required for efficient multilineage differentiation of mouse embryonic...
TRANSCRIPT

The SHB adapter protein is required for efficient multilineagedifferentiation of mouse embryonic stem cells
Vitezslav Kriz,a Cecilia Anneren,b Cheng Lai,b,1 Jacob Karlsson,a Jaroslav Mares,c
and Michael Welsha,*a Department of Medical Cell Biology, Uppsala University, Box 571, Husargatan 3, 75123 Uppsala, Sweden
b Howard Hughes Medical Institute, Department of Molecular Cell Biology, Harvard University, Cambridge, MA, USAc Institute of Biology and Medical Genetics, Charles University, Prague, Czech Republic
Received 9 September 2002, revised version received 11 December 2002
Abstract
The SH2 domain-containing adapter protein SHB transmits signals from receptor tyrosine kinases regulating diverse processes such asapoptosis and differentiation. To elucidate a role for SHB in cell differentiation, wild-type and R522K (inactive SH2 domain-mutant) SHBwere transfected and expressed in mouse embryonic stem (ES) cells. Microarray analysis using Affymetrix U74A chips on undifferentiatedES cells and expression of selected differentiation markers after generation of embryoid bodies were subsequently assessed. Wild-type SHBaltered the expression of 16 genes in undifferentiated ES cells, many of which have been found to relate to neural cell function. R522K-SHBaltered the expression of 128 genes in undifferentiated ES cells, the majority of which were decreased, including several transcription factorsrelated to development. When grown as embryoid bodies, after 4 days R522K-SHB ES cells were already found to display a differentmorphological appearance, with an impaired cavity formation that occurred in the absence of altered OCT4 expression. This impairmentwas reversed by exogenous addition of Matrigel. In addition, R522K-SHB embryoid bodies displayed reduced mRNA contents of the liverprotein albumin, the pancreatic proteins amylase, glucagon and insulin after 20 days of differentiation. Matrigel did not restore the impairedexpression of albumin in the R522K-SHB cells. Expression of the mesodermal marker cardiac actin and the neural marker neurofilamentheavy chain � was not affected by wild-type or R522K-SHB overexpression. It is concluded that SHB is required for efficient differentiationof ES cells into embryoid bodies with normal cavities and cells belonging to endodermal lineages.© 2003 Elsevier Science (USA). All rights reserved.
Keywords: SHB; Embryonic stem cell; Differentiation; Tyrosine kinase signaling; Endoderm; Pancreas; Extracellular matrix proteins
Introduction
SHB is an adapter protein with proline-rich motifs in itsN terminus, a central phosphotyrosine-binding domain, sev-eral potential tyrosine phosphorylation sites, and a C-termi-nal SH2 domain [1–3]. The role of SHB is to generatesignaling complexes in response to tyrosine kinase activa-tion, and it has been found to operate downstream of plate-let-derived growth factor (PDGF) receptors, fibroblastgrowth factor receptor 1 (FGFR-1), T-cell receptor, and
nerve growth factor (NGF) receptor TRKA [2,4–6]. Cellu-lar responses to SHB overexpression include regulation ofapoptosis [7–9], cell proliferation [6,10], and differentiationof neuronal pheochromocytoma PC12 [5] and endothelial[11] cells. In addition, a transgenic mouse expressing SHBunder the control of the rat insulin 2 promoter in � cellsexhibits increased �-cell mass at birth, suggesting the pos-sibility that SHB may play a positive role in �-cell forma-tion during late fetal development subsequent to insulingene expression [9].
Expression of SHB with an inactive SH2 domain(R522K) interferes with wild-type SHB signaling in re-sponse to T-cell receptor activation [12] and FGFR-1 acti-vation [6] in a dominant-negative fashion. This supports an
* Corresponding author. Fax: �46-18-556401.E-mail address: [email protected] (M. Welsh).1 Present address: Curis Inc, Cambridge, MA, USA
R
Available online at www.sciencedirect.com
Experimental Cell Research 286 (2003) 40–56 www.elsevier.com/locate/yexcr
0014-4827/03/$ – see front matter © 2003 Elsevier Science (USA). All rights reserved.doi:10.1016/S0014-4827(03)00099-5

experimental design in which the normal role of SHB in acellular response is elucidated by ectopic expression of thisSHB mutant.
Mouse embryonic stem (ES) cells provide a powerfultool for investigating pathways of cell differentiation. Thesecells are pluripotent and have been found to differentiatewhen grown as embryoid bodies (EBs) into cells of ecto-dermal origin, such as neural cells and keratinocytes [13,14], and mesodermal origin, i.e., cardiac and skeletal myo-cytes, vascular endothelium, hematopoietic cells [15–18].Differentiation into cells of endodermal origin has beenfound to be more difficult to achieve, but expression ofalbumin, amylase, and insulin has been reported [19–22].
To address a role for SHB in cell differentiation, partic-ularly in the development of the pancreas, mouse ES cellswere transfected with wild-type SHB and the SHB mutantR522K-SHB, and the effects on gene expression and dif-ferentiation were investigated. The results suggest thatR522K-SHB modifies ES cell differentiation by reducingexpression of a number of genes. In addition, wild-typeSHB may influence the differentiation into specific lineagesunder certain conditions.
Materials and methods
Cell lines and culture conditions
ES cells 129-R1 with a PDX1-LacZ knock-in [23] werestably transfected by electroporation (240 mV using a Bio-Rad Gene Pulser, capacitance 0.5 mF, cuvette 0.4 cm) withthe wild-type SHB cDNA (SHB) or SHB with a pointmutation in the SH2 domain rendering it inactive (SHB-R522K) inserted in the pCAGGS expression vector usingthe cytomegalovirus (CMV) enhancer �-globin promoter todrive expression. Selection was carried out in the presenceof 0.2 mg/ml hygromycin B after co-transfection with pM-SCVhph [24] and one clone for each transfection conditionincluding control transfection with antibiotic resistance onlywas obtained. In addition, ES cells from 129-SvJ were culturedin the absence of feeders, and transfected with pMSCVhph orpMSCVhph � R522K-SHB in pCAGGS. After selection inhygromycin, one control clone (C2) and one clone overex-pressing R522K-SHB (R522K-SHB-2) were obtained.
ES cells (except C2 and R522K-SHB-2 that were cul-tured feeder-free) were cultured on mitotically inactivatedprimary mouse embryonic fibroblasts in ES medium com-posed of Dulbecco’s modified medium with glutamax-1(DMEM) (Gibco-BRL, Catalog No. 31966-031), supple-mented with 15% heat-inactivated fetal bovine serum(Gibco-BRL), 1� nonessential amino acids (Gibco-BRL),1000 U/ml recombinant human leukemia inhibitory factor(LIF; ESG1107, Chemicon), 5 �M �-mercaptoethanol(Gibco-BRL), PenStrep (Roche), and gentamycin (Gibco-BRL). ES medium was changed every day. Between 5 �106 and 1 � 107 cells were trypsinized 3 min at 37°C;
trypsin (1� trypsin–EDTA, Sigma) was inactivated by se-rum-containing medium. Cells were centrifuged 5 min at220g, and washed with phosphate-buffered saline (PBS;Medicago AB).
Microarray analysis
Cells (107) from control, wild-type, and R522K-SHB ESclones were collected for mRNA preparation using the Qia-gen total RNA preparation kit (RNeasy) prior to selectionfor poly(A)� RNA using the Oligotex kit. Approximately 1�g high-quality poly(A)� RNA was used for cDNA syn-thesis using an oligo(dT) primer with the T7 RNA poly-merase promoter sequence incorporated followed by first-and second- strand cDNA synthesis using standard proce-dures. The biotinylated RNA probes were then transcribedusing T7 RNA polymerase (Enzo; Bioarray High YieldRNA Biotinylation Kit). The yield of this reaction wasapproximately 50 �g biotinylated RNA for hybridization.
The generated probes were then hybridized to the Af-fymetrix U74A chips. The A chip is composed of spottedsequences derived from 6500 known murine cDNAs and6500 ESTs. The intensity of the probes binding to the spotswas assessed by scanning and computerized storage. Thefiles containing the comparisons were then analyzed withResolver software or Affyware and differences comparingSHB or SHB-R522K-1 with the control ES clone were de-termined. Fold changes and significance values after com-parison with internal controls were calculated using Re-solver software.
Isolation of RNA and semiquantitative RT-PCR analysis
For RNA isolation the Oligotex Direct mRNA Micro Kit(Qiagen, Catalog No. 72012) was used. For RT-PCR, the OneStep RT-PCR Kit (Qiagen, Catalog No. 210212) was used: 30min 50°C; 15 min 95°C, 1 min 95°C, 1 min 55 (57)°C, 1 min72°C, 30 (35) cycles; 72°C 10 min). For each gene (Table 1)a specific pair of primers were designed (www-genome.wi .mi t . edu /cg i -b in /pr imer /pr imer3 .cg i /primer3_www.cgi). To verify that equal amounts of RNA wereused, all samples were tested for the presence of the house-keeping gene glyceraldehyde 3-phosphate dehydrogenase(GAPDH). PCR products were electrophoretically separatedon 1.5–2% agarose gel. Gels were stained with SYBR Goldnucleic acid stain (Molecular Probes) and the amount of PCRproduct was quantified by Kodak Digital Science ID.
For real-time RT-PCR, a Roche Light Cycler was used.The RNA samples were analyzed using the SYBR GreenRT-PCR kit (Qiagen, Catalog No. 204243) under the fol-lowing conditions: Reverse transcription was performed at50°C for 20 min followed by an inactivation step at 95°C,for 15 min. PCR (50 cycles) was then carried out accord-ingly: 95°C 15 s, 55°C 10 s, and 72°C 19 s. Melting curveswere assessed between 65 and 95°C (0.1°C/s). Cross points
41V. Kriz et al. / Experimental Cell Research 286 (2003) 40–56

for the different PCR products were calculated using theLight Cycler software (Version 3.5).
Differentiation of EBs
For in vitro differentiation experiments, ES cells wereremoved from feeders as follows: confluent ES cells weretrypsinized (see above) and resuspended in EB medium (ESmedium without LIF) and plated for 30 min on tissueculture dishes; the step was repeated one more time. EScells from the suspension were centrifuged and then placedin Petri dishes (Day 0) at a concentration of 1–1.25 � 105
cells/cm2 and cultured as EBs for 4 or 20 days, changing themedium, every second day and subsequently collecting theEBs for histology or RNA isolation. In some experiments,Matrigel (Becton-Dickinson) was added at 4% (v/v) simul-taneously when ES cells were used for generating EBs.
Histology
Collected EBs were pelleted, washed twice with PBS,and fixed in 4% formaldehyde overnight. EBs were embed-ded in paraffin before hematoxylin–eosin staining.
Results
SHB overexpression in ES cells
Expression of SHB protein by Western blot analysis incontrol, SHB, SHB-R522K-1 and SHB-R522K-2 ES cells is
shown in Fig. 1. After transfection with SHB, only oneclone each was identified overexpressing p55 SHB. Theincrease in SHB expression was 2.2-fold after normalizationfor total ERK, and a similar degree of SHB overexpressionhas in other systems exerted effects on cell apoptosis anddifferentiation [5,7,11]. Two clones expressing R522K-SHB were obtained (R522K-SHB-1 and R522K-SHB-2).These clones exhibited increased SHB contents of 2.8-fold
Table 1PCR primers used to assess expression of the genes in ES cells and differentiated EBs
Genea Left primer Right primer Size (bp) Reference Annealing temperature(°C) (Q buffer)
GAPDH accacagtccatgccatcac tccaccaccctgttgctgta 452 Gualandris et al., 2000 [44] 55�-Globin gagaaggctgctgtctctt gaagttctcaggatccacat 291 GenBank:J00413 55Amylase atgactgggtctgtgaacat agcaggaagaccagtctgta 200 GenBank:M11895 55c-Actin aactacaggcattgttctgg ttatgtcacggacaatttca 197 GenBank:X03767 55Albumin tatccctgttgctgagactt actagaatggcattttggaa 199 GenBank:AJ011413 55PDX1 atgaaatccaccaaagctc gatgtgtctctcggtcaagt 207 GenBank:NM_008814 55 � Q bufferOCT-4 ggcgttctctttggaaaggtgttc ctcgaaccacatccttctct 312 GenBank:X52437 55Glucagon actcacagggcacattcacc ccagttgatgaagtccctgg 352 GenBank:AF276754 55Insulin ggctctctacctggtgtgt tgcagcactgatctacaatg 180 GenBank:X04724 57 � Q bufferNF-H aagccaaagaaagaggagat ctttctttctggtctgtgct 200 GenBank:M35131 55PAFAH agcagaagaagtagcaggtg ccgtctatatcaaggagctg 201 GenBank:U57747 55LKGRSD agcagctgagcaagaaatac ttcatggctggtgtaatgta 199 GenBank:AJ224761 55Twist gtctgaacactcgtttgtgtcc tgatcccagcgtttttatttct 197 GenBank:M63649 55PEG/MEST atcccggtgcttcttctc agagcaggggagagctga 365 GenBank:AF017994 55SOX17 gagctaagcaagatgctagg tacttgtagttggggtggtc 122 GenBank:D49474 55GL12 aggagacctgagcaacacta gatgccccttaatcttttct 174 GenBank:X99104 55KLF7 actgacaaacaaacagaccc acaagttgtagctcctggaa 118 GenBank:NM_033563 55FOXD3 actgataaccgaatttccaa aaaataaaaaccccaaaagg 129 GenBank:NM_010425 55Collagen agtcagacaacagatgaccc gatgttgcagaagagaaagg 167 GenBank J04694 55Laminin agacacatcctgatggagag taacactctggtgagctcct 279 GenBank NM_008480 55G-utrophin gctggatgaccttaatatgc caacaggttgtctgtgtttg 201 GenBank:X83506 55
a PAFAH, platelet-activating factor acetylhydrolase; LKGRSD, lysine-ketoglutarate reductase/saccharopine dehydrogenase; KLF7, Kruppel-like factor 7;FOXD3, forkhead box D3; collagen, �1 type IV collagen; SOX17, HMG box transcription factor.
Fig. 1. Expression of SHB in ES cells transfected with wild-type SHB orR522K-SHB by Western blot analysis. Cell lysates from control (C or C2),wild-type SHB (SHB), and two separate clones expressing R522K-SHB(R522K-SHB-1 and R522K-SHB-2) were dissolved in SDS-sample bufferand electrophoresed on 9% acrylamide gels prior to electrical transfer toImmobilon filters. These were then incubated with SHB or ERK antibodiesprior to incubation with secondary antibody and enhanced chemilumines-cence for specific protein determination. p55 SHB has been indicated, andexpression of ERK 1,2 is shown as a control of loading.
42 V. Kriz et al. / Experimental Cell Research 286 (2003) 40–56

and 3.5-fold, respectively. The amount of total ERK isshown as a control for loading.
Gene expression in undifferentiated ES cells
Microarray analysis was performed on undifferentiatedcontrol, SHB, and SHB-R522K-1 ES cells to understand therole of SHB in ES cell differentiation. The microarrays wereperformed twice, and effects after comparison with thecontrol clone that were larger than 1.5-fold in both experi-ments and significant at the P � 0.001 level in one exper-iment are listed. The analyses revealed 128 changes, ofwhich 107 were decreased gene expression in the SHB-R522K-1 cells (Table 2). Changes in expression of manytranscription factors that previously have been found tocontrol developmental processes were detected. These in-clude retinoic acid receptors, H19 mRNA, SOX17, GL12Kruppel family member, ID-2, GBX2, and Kruppel-likefactor 7 (Table 2). In addition, the expression of severalsignaling and extracellular matrix proteins was decreased,presumably as a consequence of the altered transcriptionfactor profile. All five genes that were independently testedby RT-PCR (GLI2, SOX17, Kruppel-like factor 7, collagentype IV, and laminin alpha 1) were also decreased by thisanalysis in SHB-R522K-1 (Table 2). In SHB-R522K-2 cells,all but GLI2 were decreased (Table 4). Appendix Table 1gives a more comprehensive view of the changes in geneexpression observed in SHB-R522K-1 ES cells, includingthe fold changes, P-values, and brief descriptions of knownfunctions of the protein products.
Wild-type SHB caused 16 changes in gene expression(Table 3). Expression of five of the genes that were notedthrough microarray analysis was independently assessed byRT-PCR, and of these five changes four were qualitativelyreproduced by RT-PCR. Many of the SHB-induced changesin gene expression are related to neural cell function, suchas connexin 43, Ca2�/calmodulin-dependent protein kinaseII, lysine-ketoglutarate reductase, G-utrophin, and platelet-activating factor acetylhydrolase [25–29]. In addition, threegenes belonging to the two-cell-stage gene family weredetected, which are considered to relate to early zygotedifferentiation. Twist is a transcription factor of importancefor mesodermal differentiation [30] and PEG1/MEST is adetoxification enzyme required for mesodermal cell survival[31]. Appendix Table 2 provides more information on thesemicroarray data.
Differentiating EBs derived from the SHB-R522K clonedisplay altered morphology after 4 days
The changes in undifferentiated ES cell gene expressionprofiles were next followed by assessing the morphology of4-day-old EBs during differentiation (Fig. 2). Both the con-trol and SHB clones developed cystic cavities (42 and 59%,respectively). The central cavity is smooth and regular andcovered with at least two layers of cells, an inner ectodermal
buyer and an outer endodermal layer. The ectodermal layerdifferentiates into pseudostratified columnar epithelium.The cavitation process was in our hands most developed ondays 4 and 5 after generation of EBs. ES cells of SHB-R522K-1 or SHB-R522K-2 clones (0 and 20%, respec-tively), however, exhibited reduced cavitation (Fig. 2). EBsderived from the SHB-R522K-1 clone failed to produce aregular cavity with a clear epithelial lining. In these, the twoepithelial layers are not easily visualized (Fig. 2). Also, theSHB-R522K-2 EBs exhibited fewer regular cavities than thecontrol cells. When the SHB-R522K-1 EBs were generatedin the presence of 4% Matrigel, cavitation was restored andthe morphological appearance of the EBs was similar to thatof the other clones (Fig. 2). An oblique arrow indicates acavity with an epithelial lining. Likewise, the SHB-R522K-2EBs produced cavities to a much larger extent when gen-erated in the presence of Matrigel (results not shown).
The OCT4 mRNA level in SHB-R522K-1 EBs was sim-ilar to that of the control EBs under these conditions(Fig. 2).
This suggests that expression of inactive SHB will havean impact on early ES cell differentiation in EBs and thatthis in part depends on altered synthesis of extracellularmatrix proteins.
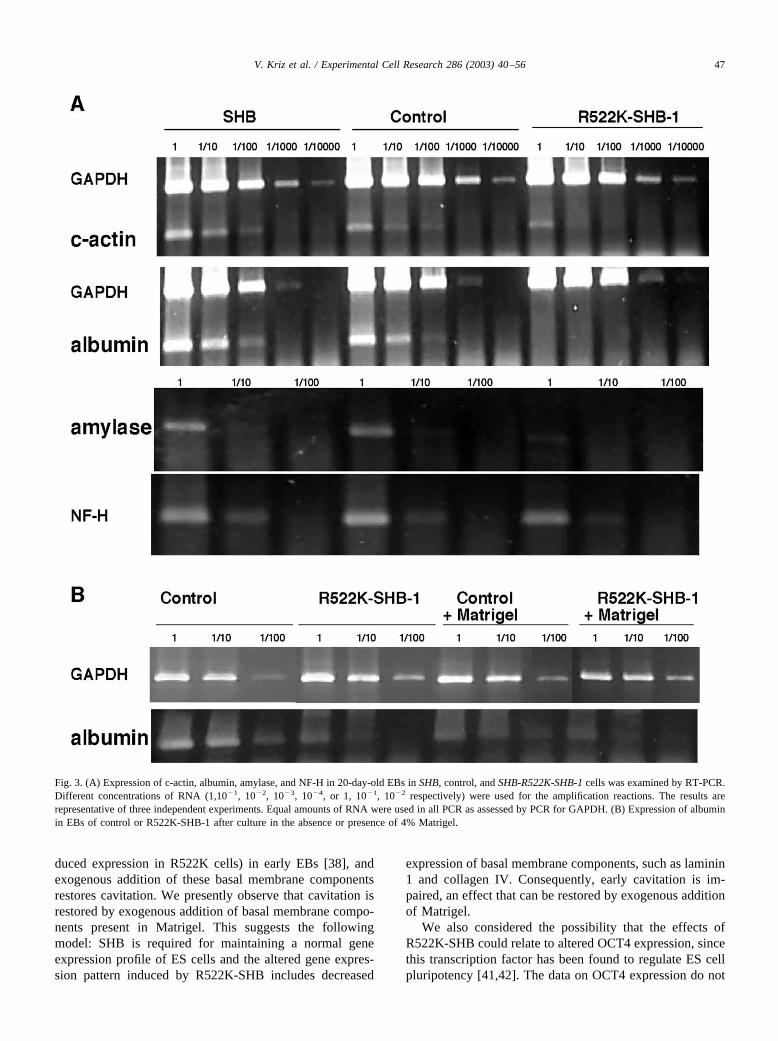
R522K-SHB negatively affected expression of some genescorresponding to differentiated functions in 20-day-oldEBs
We have performed semiquantitative RT-PCR analysison 20-day-old EBs derived from the different wild-type orR522K-SHB clones to study the subsequent consequencesof the altered gene expression profile and cavitation pattern.As we have previously shown that SHB is involved in thedifferentiation of neural PC12 cells, tube formation of en-dothelial cells, and T-cell signaling, we anticipated thatSHB might influence ES differentiation into cells express-ing markers for the three germ layers. Thus, we decided totest expression of one mesodermal gene, cardiac actin (c-actin), as a marker for heart muscle. The liver and exocrinepancreas markers albumin and amylase are of endodermalorigin and the brain marker neurofilament component(NF-H) is a marker for the ectoderm. All procedures wereperformed three times and related to expression of thehousekeeping gene GAPDH (Fig. 3). We could not detectany alterations in expression of NF-H, c-actin, albumin, oramylases in the SHB clone relative its control (Fig. 3).Overexpression of R522K-SHB negatively affected expres-sion of albumin and amylase in comparison with the controlor SHB cells in both R522K-SHB clones (Fig. 3, Table 4),albeit to a lesser extent in the R522K-SHB-2 clone. Theimpaired differentiation into cells expressing these genessuggests that SHB is required for these differentiation path-ways since R522K-SHB may function in a dominant-nega-tive fashion [2,6,12]. The effects of SHB may be direct orindirect via effects on cell survival. The exogenous addition
43V. Kriz et al. / Experimental Cell Research 286 (2003) 40–56

Table 2Microarray analysis comparing the R522K-SHB ES cells with control ES cells using U74A Affymetrix chipsa
Decreased gene expression
Transcription factors and DNA-binding proteins Signaling proteinsGLI-Kruppel family member GL12 (2.7; 5.5) Patched homolog 2 (Shh receptor) (3.3)Retinoic acid receptor, alpha (7.5) Disabled homolog 2, Drosophila (8.6)Retinoic acid receptor, gamma (4.0) FGF-18 (6.9)Kruppel-like factor 7 (ubiquitous) (4.2; 1.9) Proliferin 3 (6.5)SRY-box containing gene 17 (Sox17, 4.7; 7.4; 1.8) Protein kinase C, lambda (2.1)H19 (3.2) Lysophospholipase 1 (3.0; 6.9; 5.1)Gastrulation brain homeobox 2, Gbx2 (2.4) Phosphatidylinositol transfer protein, beta (8.1; 3.7)Helix–loop–helix ld2 (2.4) Inositol polyphosphate-5-phosphatase, 145 kDa (5.4)GA repeat binding protein, alpha (4.5) Pleckstrin homology, Sec7 and coiled/coil domains 3 (4.9)RIKEN cDNA E130112L17 gene (3.0) Transferrin receptors 1 and 2 (6.1, 5.3)General transcription factor II 1 (2.6) Casein kinase II, alpha 1 related sequence 4 (6.2)Nuclear transcription factor-Y gamma (13.7) Shc (SH2 domain-containing transforming protein C1)(8.0)Reduced expression 2, Rex2 (5.1) Rab23 (GTPase) (3.3)COP9 (constitutive photomorphogenic) homolog, subunit 5 (Arabidopsisthaliana) (2.1)
Gab1 (growth factor receptor-bound protein 2-associated protein 1) (5.2)LIM and SH3 protein 1 (3.3)
Forkhead box M1 (3.8) Protein-tyrosine phosphatase IF1 (2.6)TBP-interacting protein, Tp120a (2.7) Protein phosphatase 1, regulatory subunit 7 (2.3)Non-POU-domain-containing, octamer-binding protein (2.6) Casein Kinase 1 delta (12.8)Transcription factor S-II-related protein (2.1) Platelet-activating factor acetylhydrolase 1b,alpha 2 subunit (8.7)c-myc (2.6) Thymopoietin (2.6)DP1 (6.0) Small inducible cytokine A2 (4.4)Host cell factor C1 (6.9) Rho GTPase-activating protein 5 (3.7)MAD2 (mitotic arrest deficient, homolog)-like 1 (yeast) (2.2) 3-Monooxygenase/tryptophan 5-monooxygenase activation
DNA and RNA metabolism protein, gamma polypeptide (14-3-3) (6.9)RIKEN cDNA 2310005B10 gene, DNA helicase (4.7) Mts 1 (p161NK4A) (2.8)CUG triplet repeat, RNA binding protein 1 (8.3) Extracellular matrix and structural proteinsSignal recognition particle 9 kDa (2.5) Laminin, alpha 1 (3.0; 1.6)Eukaryotic translation initiation factor 2, subunit 3, structural gene Y-linked (6.8)
Procolliagen, type IV, alpha 1 (4.2; 1.9)Procollagen, type IV, alpha 2 (2.7)
Splicing factor, arginine/serine-rich 10 (1.8) Procolliagen, type V, alpha 1 (4.1)DEAD (aspartate–glutamate–alanine–aspartate) box polypeptide, Ychromosome (4.1)
Actin, alpha 2, smooth muscle, aorta (2.3)Enabled homolog (Drosophila) (3.2)
Signal recognition particle receptor alpha (11.7) Utrophin (4.3)Signal sequence receptor, gamma (2.9; 4.5) Proteoglycan, secretory granule (12)Ribosomal protein L27a (2.2) Biglycan, bone proteoglycan (2.5)
Others Lysosomal membrane glycoprotein 2 (8.4)CD84 antigen (7.4) Proteolytic degradationComplement receptor-related protein (5.1) serine proteinase inhibitor, clade E, member 1 (Serpine 1) (2.6)Uridine monophosphate synthetase (4.6) tripeptidyl peptidase II (3.5)Solute carrier family 2 member 3 (3.7) coagulation factor III (5.7)Adenylosuccinate synthase 2 (2.3) coagulation factor II, thrombin receptor (5.2)UDP-glucose ceramide glucosyltransferase (5.0) cathepsin H (8.8)Importin alpha 3 (Kpna3) (8.3) ring finger protein 2 (4.2)Adapter protein AP-1, gamma 1 subunit (4.8) calpain 12 (2.9)Anti-oxidant protein 2 (3.1) NPC-derived proline rich protein 1 (2.1)SEC23A (6.6)Glucose-6-phosphate dehydrogenase 2 (2.8)
Increased gene expression
Transcription factors and DNA-binding proteins OthersTranscription factor AP-2,� (2.5) Membrane-associated protein 17 (3.4)Ikappa B alpha (2.5) Rhesus blood group CE and D (4.9)
Signaling proteins Glutathione S-transferase, mu1 (2.5)PDGF-AA (2.1) Phenylalkylamine Ca2� antagonist binding protein (2.9)Gbp1 (GTPase) (4.7) Spermidine/spermine N1-acetyltransferase (2.2)Retinol-binding protein 1 (2.8) BCL2/adenovirus E1B 19-interacting protein 1 (2.9)ERBB-3 (2.4) Cysteine-rich intestinal protein (3.1)BMP4 (2.5) Amine oxidase 3 (4.3)
Apolipoprotein CI (3.5)Apolipoprotein E (1.7)
a Changes (fold differences in parentheses) at the P � 0.001 level for genes with presently known function are listed. Boldface numbers indicate changesdetected independently by RT-PCR. When more than one value is shown, several sequences for the same gene were on the chip.
44 V. Kriz et al. / Experimental Cell Research 286 (2003) 40–56

of Matrigel did not restore the decreased albumin expres-sion in the R522K-SHB cells (Fig. 3B), suggesting thatSHB, in addition to being necessary for the production ofextracellular matrix components, controls EB differentia-tion at later stages independently of the early cavitationprocess. This may relate to the effects on the expressionpattern of specific transcription factors noted in SHB-R522K.
SHB-R522K decreases insulin and glucagon expression in20-day-old EBs
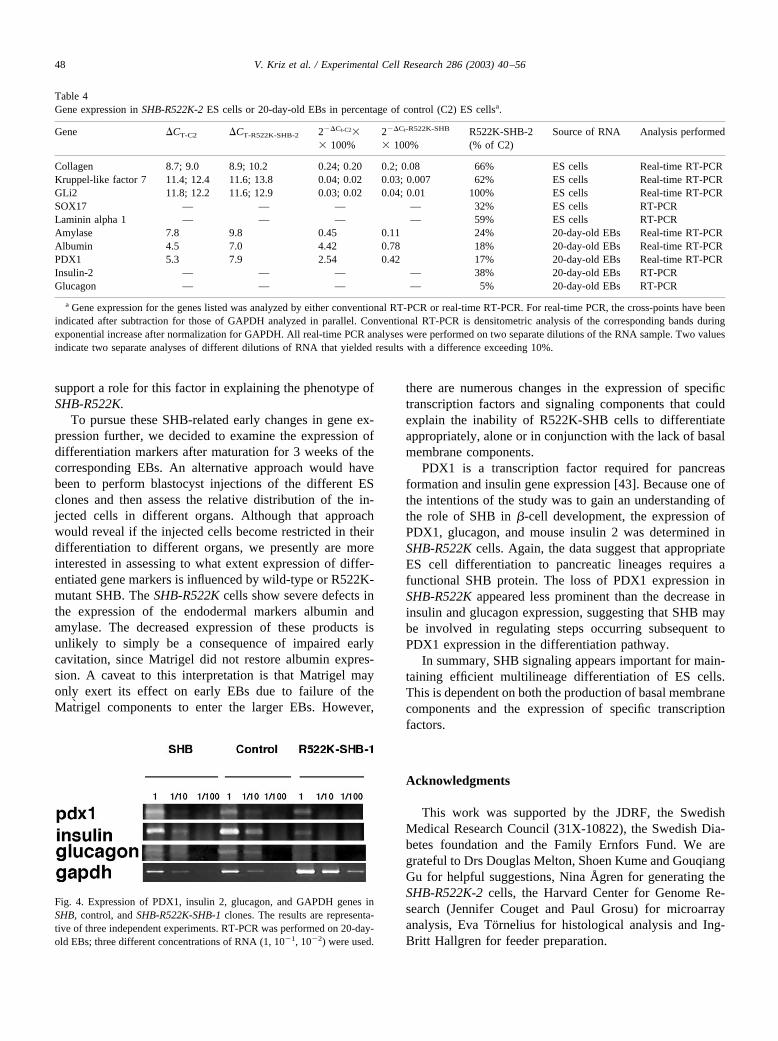
It was previously reported that a transgenic mouse ex-pressing SHB under the insulin promoter increased �-cellmass [9]. For this reason, we wished to study the expressionof PDXI, glucagon, and insulin in EBs. EBs from ES cellsexpressing wild-type SHB displayed contents of glucagonand insulin mRNA similar to those of their correspondingcontrols (Fig 4). Expression of both insulin 2 and glucagon,however, was decreased in SHB-R522K-1 and SHB-R522K-2 EBs (Fig. 4, Table 4). PDX1 expression was alsodecreased, but to a lesser extent. Thus a functional SHBprotein is required for appropriate expression of these en-docrine markers in differentiating EBs.
Discussion
The present investigation was conducted to assess therole for the SHB adapter protein during embryonic celldifferentiation. For this purpose, ES cells were transfectedwith wild-type SHB or SHB containing an SH2 domainmutation that renders it dominant-negative with respect toFGFR-1 and T-cell receptor signaling. A dominant-negativeapproach creates an experimental advantage over a nullmutation in case there are other SHB-family members withredundant function relative to SHB. Subsequent to SHBoverexpression, the gene expression profile in undifferenti-ated ES cells was assessed, since we argued that SHB mayalter the ensuing differentiation pattern as a consequence ofearly changes in gene expression. For wild-type SHB, mi-
croarray analysis revealed changes in the gene expressionpattern, including increased expression of two-cell-stagegenes and altered expression of several genes related toneural cell function or development. This altered expressionpattern does not, however, have any major impact on thedifferentiation of these SHB- overexpressing ES cells, sincethey exhibited no detectable morphological changes orchanges in the expression of markers for more differentiatedfunctions. However, our analysis of these cells was notexhaustive, and it is possible that more detailed assessmentof their characteristics could reveal changes, particularlywith respect to neural cell differentiation.
The undifferentiated SHB-R522K ES cells, on the otherhand, exhibited dramatic changes with respect to ES cellcharacteristics. The gene expression pattern was changedand most genes appeared downregulated. Among the genesthat were expressed at lower levels in the R522K cells werethose corresponding to the transcription factors retinoic acidreceptors, GLi2, Kruppel-like factor 7, and SOX17. Reti-noic acid receptors have been implicated in neuronal devel-opment and endodermal patterning [32] as has the Kruppel-like factor 3 which regulates the expression of the NGFreceptor TRKA (neural growth factor receptor 1) [33].SOX17 [34], has, in Drosophila, been reported to nega-tively regulate the WNT-�-catenin pathway, whereas GLI2supports sonic hedgehog (SHH) signaling [35]. Indeed, theSHH receptor Patch 2 was also decreased. GBX2 is aretinoic acid receptor-inducible gene [36], which was alsodownregulated. GBX2 has been suggested to maintain cellsin a pluripotent state [37]. In addition, numerous extracel-lular matrix and signaling genes were inhibited by theR522K mutation. The altered gene expression pattern has animmediate impact on the differentiation of the ES cells; theearly cavitation is impaired and the epithelial lining mor-phologically altered. The process of EB cavitation has beenshown to depend on FGFR-2 [38], Indian Hedgehog [39],and BMP [40] signaling. We have observed that SHB isrequired for FGFR-1 signaling in endothelial cells [6,11],but the role of SHB in FGFR-2 signaling is so far notknown. However, FGFR-2 signaling is required for laminin1 and collagen IV synthesis (two genes that exhibited re-
Table 3Affymetrix U74A microarray analysis comparing wild-type SHB ES cells with control ES cells at the P � 0.001 levela
Decreased gene expression Increased gene expression
X-linked lymphocyte-regulated 3b (2.6) Clone L2 uniform group of two-cell-stage gene family (5.2)Lactate dehydrogenase B (2.2) Clone L3 variable group of two-cell-stage gene family (6.9)Ca2�/calmodulin-dependent protein kinase II beta subunit (2.8) Clone L10 variable group of two-cell-stage gene family (4.7)G-utrophin (3.2; 1.6) Glutathione peroxidase 2, pseudogene 1 (3.2)Platelet-activating factor acetylhydrolase Apolipoprotein E (1.6)isoform lb beta subunit (2.6; 5) Connexin 43 (alpha-1 gap junction) (1.8)Peg1/MEST protein (6.9; 5.6) Lysine-ketoglutarate reductase/saccharopine dehydrogenase 3 (4.8; 1.4)H19 (1.8) Twist gene homolog, (Drosophila) (3; �2.8)
BMP4 (2.0)
a Fold changes compared with control are shown (means for two different microarrays). Boldface letters indicate the corresponding value by independentRT-PCR analysis.
45V. Kriz et al. / Experimental Cell Research 286 (2003) 40–56

Fig. 2. Histological examination of 4-day-old EBs from control, SHB, SHB-R522K-1, and SHB-R522K-SHB-2, ES cells. Also, the SHB-R522K-SHB-1 cellswere cultured for 4 days in the presence of Matrigel. Sections of EBs derived from the different ES clones harvested after 4 days in culture were stained withhematoxylin and eosin. Central cavities surrounded by epithelium have been indicated with the arrows. Representative EBs for each clone are shown. Thenumber of EBs with cavities has been indicated for each clone. Bars � 50 �m. Bottom: RT-PCR showing the expression of OCT4 and GAPDH in control,R522K-SHB-1, control � Matrigel, and R522K-SHB-1 � Matrigel EBs after 4 days. Three different concentrations of RNA were used.
duced expression in R522K cells) in early EBs [38], andexogenous addition of these basal membrane componentsrestores cavitation. We presently observe that cavitation isrestored by exogenous addition of basal membrane compo-nents present in Matrigel. This suggests the followingmodel: SHB is required for maintaining a normal geneexpression profile of ES cells and the altered gene expres-sion pattern induced by R522K-SHB includes decreased
expression of basal membrane components, such as laminin1 and collagen IV. Consequently, early cavitation is im-paired, an effect that can be restored by exogenous additionof Matrigel.
We also considered the possibility that the effects ofR522K-SHB could relate to altered OCT4 expression, sincethis transcription factor has been found to regulate ES cellpluripotency [41,42]. The data on OCT4 expression do not
Fig. 3. (A) Expression of c-actin, albumin, amylase, and NF-H in 20-day-old EBs in SHB, control, and SHB-R522K-SHB-1 cells was examined by RT-PCR.Different concentrations of RNA (1,10�1, 10�2, 10�3, 10�4, or 1, 10�1, 10�2 respectively) were used for the amplification reactions. The results arerepresentative of three independent experiments. Equal amounts of RNA were used in all PCR as assessed by PCR for GAPDH. (B) Expression of albuminin EBs of control or R522K-SHB-1 after culture in the absence or presence of 4% Matrigel.
47V. Kriz et al. / Experimental Cell Research 286 (2003) 40–56
support a role for this factor in explaining the phenotype ofSHB-R522K.
To pursue these SHB-related early changes in gene ex-pression further, we decided to examine the expression ofdifferentiation markers after maturation for 3 weeks of thecorresponding EBs. An alternative approach would havebeen to perform blastocyst injections of the different ESclones and then assess the relative distribution of the in-jected cells in different organs. Although that approachwould reveal if the injected cells become restricted in theirdifferentiation to different organs, we presently are moreinterested in assessing to what extent expression of differ-entiated gene markers is influenced by wild-type or R522K-mutant SHB. The SHB-R522K cells show severe defects inthe expression of the endodermal markers albumin andamylase. The decreased expression of these products isunlikely to simply be a consequence of impaired earlycavitation, since Matrigel did not restore albumin expres-sion. A caveat to this interpretation is that Matrigel mayonly exert its effect on early EBs due to failure of theMatrigel components to enter the larger EBs. However,
there are numerous changes in the expression of specifictranscription factors and signaling components that couldexplain the inability of R522K-SHB cells to differentiateappropriately, alone or in conjunction with the lack of basalmembrane components.
PDX1 is a transcription factor required for pancreasformation and insulin gene expression [43]. Because one ofthe intentions of the study was to gain an understanding ofthe role of SHB in �-cell development, the expression ofPDX1, glucagon, and mouse insulin 2 was determined inSHB-R522K cells. Again, the data suggest that appropriateES cell differentiation to pancreatic lineages requires afunctional SHB protein. The loss of PDX1 expression inSHB-R522K appeared less prominent than the decrease ininsulin and glucagon expression, suggesting that SHB maybe involved in regulating steps occurring subsequent toPDX1 expression in the differentiation pathway.
In summary, SHB signaling appears important for main-taining efficient multilineage differentiation of ES cells.This is dependent on both the production of basal membranecomponents and the expression of specific transcriptionfactors.
Acknowledgments
This work was supported by the JDRF, the SwedishMedical Research Council (31X-10822), the Swedish Dia-betes foundation and the Family Ernfors Fund. We aregrateful to Drs Douglas Melton, Shoen Kume and GouqiangGu for helpful suggestions, Nina Ågren for generating theSHB-R522K-2 cells, the Harvard Center for Genome Re-search (Jennifer Couget and Paul Grosu) for microarrayanalysis, Eva Tornelius for histological analysis and Ing-Britt Hallgren for feeder preparation.
Table 4Gene expression in SHB-R522K-2 ES cells or 20-day-old EBs in percentage of control (C2) ES cellsa.
Gene �CT-C2 �CT-R522K-SHB-2 2��Ct-C2�� 100%
2��Ct-R522K-SHB
� 100%R522K-SHB-2(% of C2)
Source of RNA Analysis performed
Collagen 8.7; 9.0 8.9; 10.2 0.24; 0.20 0.2; 0.08 66% ES cells Real-time RT-PCRKruppel-like factor 7 11.4; 12.4 11.6; 13.8 0.04; 0.02 0.03; 0.007 62% ES cells Real-time RT-PCRGLi2 11.8; 12.2 11.6; 12.9 0.03; 0.02 0.04; 0.01 100% ES cells Real-time RT-PCRSOX17 — — — — 32% ES cells RT-PCRLaminin alpha 1 — — — — 59% ES cells RT-PCRAmylase 7.8 9.8 0.45 0.11 24% 20-day-old EBs Real-time RT-PCRAlbumin 4.5 7.0 4.42 0.78 18% 20-day-old EBs Real-time RT-PCRPDX1 5.3 7.9 2.54 0.42 17% 20-day-old EBs Real-time RT-PCRInsulin-2 — — — — 38% 20-day-old EBs RT-PCRGlucagon — — — — 5% 20-day-old EBs RT-PCR
a Gene expression for the genes listed was analyzed by either conventional RT-PCR or real-time RT-PCR. For real-time PCR, the cross-points have beenindicated after subtraction for those of GAPDH analyzed in parallel. Conventional RT-PCR is densitometric analysis of the corresponding bands duringexponential increase after normalization for GAPDH. All real-time PCR analyses were performed on two separate dilutions of the RNA sample. Two valuesindicate two separate analyses of different dilutions of RNA that yielded results with a difference exceeding 10%.
Fig. 4. Expression of PDX1, insulin 2, glucagon, and GAPDH genes inSHB, control, and SHB-R522K-SHB-1 clones. The results are representa-tive of three independent experiments. RT-PCR was performed on 20-day-old EBs; three different concentrations of RNA (1, 10�1, 10�2) were used.
48 V. Kriz et al. / Experimental Cell Research 286 (2003) 40–56

Appendix 1: R522K-SHB-induced changes in gene expression compared with control ES cells
DNA binding, transcription factors, RNA metabolism
Probe set ID Symbol Full name Function Foldchange
Pvalue
100395_at Gli2 GLI-Kruppel family member GLI2 Zinc finger transcription factor, skeletal development, �2.7 �0.001regulates sonic hedgehog pathway �3.7 �0.001
92901_at Rar_ Retinoic acid receptor, alpha Steroid hormone receptor, neuronal cell �7.5 �0.001differentiation, endodermal patterning in zebrafish �5.4 �0.001
102419_at Rar_ Retinoic acid receptor, gamma Steroid hormone receptor, neuronal cell �4.0 �0.001differentiation, endodermal patterning in zebrafish �2.8 �0.02
104645_at Klf7 Kruppel-like factor 7 (uniquitous) Transcription factor, regulates TrkA (NGF receptor) �4.2 �0.001expression �3.2 �0.5
92996_at Sox17 SRY-box containing gene 17 Transcription factor, negatively regulates WNT �- �4.7 �0.001catenin pathway in Xenopus, endodermal �2.5 �0.06
92997_g_at differentiation �7.4 �0.001�9.3 �0.04
93028_at H19 H19 fetal liver mRNA Transcription factor, activated on embryonic stem �3.3 �0.001cell implantation �3.2
94200_at Gbx2 Gastrulation brain homeobox 2 Transcription factor, retinoic acid-inducible gene, �2.4 �0.001role in maintaining cell pluripotency �1.6 �0.001
93013_at Helix–loop–helix Helix–loop–helix protein Id2 gene, NGF and IGF-1 receptor-responsive gene, regulates �2.4 �0.001protein Id2 transcription factor developmental processes and apoptosis �1.5 �0.05
94969_at Nfyc Nuclear transcription factor-Y gamma Transcription factor, responds to changes in cellular �13.7 �0.001redox potential �23.5 �0.001
104712_at c-myc c-myc gene exon 3 Proto-oncogene, transcription factor, regulates �2.6 �0.001proliferation, apoptosis, and differentiation �2.0 �0.001
100901_at Hcfc1 Host cell factor C1 Transcription factor, regulates cell cycle �6.9 �0.001�4.2 �0.02
97565_r_at DP1 Cell cycle regulatory transcription factor Forms heterodimeric complex with E2F transcription �6.0 �0.001DP1 (DP1) gene, promoter, and exon 1 factor, regulates cell cycle progression �8.5 �0.001
99632_at Mad2l1 MAD2 (mitotic arrest deficient, homolog)- Mitotic chromosome segregation �2.2 �0.001like 1 (yeast) �1.6 �0.001
98305_at Foxm1 Forkhead box M1 Transcription factor �3.8 �0.001�2.9 �0.46
99338_at Tp120a TBP-interactin protein TATA-binding protein, global transcription factor �2.7 �0.001�2.7 �0.001
93831_at Nono Non-POU-domain-containing, octamer-binding protein
Binds RNA and DNA, enhances the association ofmany DNA-binding proteins to their targets
�2.6�2.0
�0.001�0.001
104149_at Nfkbia 1 kappa B alpha Inhibitor of transcription factor nuclear factor kappaB, inflammatory response
�2.5 �0.001�2.2
95460_at Cops5 COP9 (constitutive photomorphogenic)homolog, subunit 5 (Arabidopsis thaliana)
Transcription factor, RNA initiation �2.1 �0.001�1.8 �0.001
101528_at Tcea1 Transcription factor S-II-related proteins Participates in RNA pol II transcription �2.1 �0.001101529_g_at �1.7 �0.001
�2.4 �0.001�2.1 �0.001
92275_at Tcfap2c Transcription factor AP-2, gamma Transcription factor �2.5 �0.001�2.6 �0.001
94296_s_at Gtf2i General transcription factor III Transcription factor �2.6 �0.001�2.6 �0.001
102981_at Gabp_ GA repeat-binding protein, alpha Transcription factor �4.5 �0.001�2.0 �0.07
95346_at Rex2 Reduced expression 2 DNA binding �5.1 �0.001�1.6 �0.36
103989_at E130112L17Rik RIKEN cDNA E130112L17 gene DNA binding �3.0 �0.001�4.0 �0.01
100037_at 2310005B1Rik RIKEN cDNA 2310005B10 gene DNA helicase �4.7 �0.001�4.2 �0.001
103842_at Dby DEAD (aspartate–glutamate–alanine–aspartate) box polypeptide, Y chromosome
DNA helicase �4.1 �0.001�5.3 �0.02
95791_s_at Sfrs10 Splicing factor, arginine/serine-rich 10 Splicing factor �1.8 �0.001�1.5 �0.001
93630_at Cugbp1 CUG triplet repeat, RNA binding protein1
Pre-mRNA splicing factor �8.3 �0.001�6.3 �0.19
94208_at Rpl27a Ribosomal protein L27a Protein translation �2.2 �0.001�2.5 �0.001
103674_f_at Eif2s3y Eukaryotic translation initiation factor 2,subunit 3, structural gene Y-linked
Translation elongation factor �6.8 �0.001�4.8 �0.01
(continued on next page)

Appendix 1 (continued)
Structural proteins
Probe set ID Symbol Gene Function Foldchange
Pvalue
103729_at Lam�1 Laminin, alpha 1 Extracellular matrix, adhesion, regulatesdifferentiation and FGF-dependent EBcavitation
�3.0�4.9
�0.001�0.001
101093_at Col4�1 Procollagen, type IV, alpha 1 Extracellular matrix, adhesion, regulatesdifferentiation and FGF-dependent EBcavitation
�4.2�3.9
�0.001�0.001
101039_at Col4�2 Procollagen, type IV, alpha 2 Extracellular matrix, adhesion, regulatesdifferentiation
�2.7�3.9
�0.001�0.001
92567_at Col5�2 Procollagen, type V, alpha 2 Extracellular matrix, adhesion, regulatesdifferentiation
�4.1�4.5
�0.001�0.001
93100_at Acta2 Actin, alpha 2, smooth muscle in aorta Actin cytosceleton, muscle development �2.3 �0.001�1.9 �0.001
93864_s_at Enah Enabled homolog (Drosophila) Actin filament organization �3.2 �0.001�2.8 �0.001
96049_s Bgn Biglycan Bone proteoglycan �2.5 �0.001�2.2
92508_s_at Utrn Utrophin Dystrophin-related protein particularlyexpressed in neural cells
�4.3 �0.001�8.4 �0.05
96935_at Map17-pending Membrane-associated protein 17 Transmembrane protein upregulated inhuman carcinomas; reduces tumorgrowth and cell proliferation.
�3.4 �0.001
�2.6 �0.39
99501_at Lamp2 Lysosomal membrane glycoprotein 2 Transmembrane lysosomalglycoprotein; mediates adhesion oftumor cells and lymphocytes tovascular endothelium
�8.4 �0.001�1.6 �0.64
94085_at Prg Proteoglycan, secretory granule Component of mast cell, basophilic andNK cell granules
�11.8 �0.001�12.2 �0.001
Signaling
Probe set ID Symbol Gene Function Foldchange
Pvalue
94932_at Pdgfa Platelet-derived growth factor, alpha Growth factor, developmentally regulatedpericyte formation
�2.1 �0.001�1.7 �0.001
93456_r_at Bmp4 Bone morphogenic protein 4 TGF�-family member, differentiation factor,regulates mesenchyme
�2.5 �0.001�2.0
98045_s_at DDab2 Disabled homolog 2 (Drosophila) Adapter protein that links TGF� receptor tothe Smads pathway, involved in embryonicdevelopment, tumor suppressor
�8.6 �0.001�3.3 �0.01
96557_at Ptch2 Patched homolog 2 Sonic hedgehog (Shh) receptor, involved inembryogenesis
�3.3 �0.001�2.1 �0.01
95316_at Fgf18 Fibroblast growth factor 18 Growth factor �6.9 �0.001�7.5 �0.06
101148_at Prkcl Protein kinase C, lambda Protein Ser/Thr kinase, involved in manysignaling responses
�2.1 �0.001�4.8 �0.03
101944_at Lypla1 Lysophospholipase 1 Lysophospholipase, phospholipid metabolism �3.0 �0.001�4.6 �0.001
101945_g_at �6.9 �0.001�5.0 �0.001�5.1 �0.001
101946_at �3.8 �0.001102696_s_at Pitpn � Phosphatidylinositol transfer protein, beta Phospholipid transporter �8.1 �0.001
�3.5 �0.001102697_at �3.7 �0.001
�5.4 �0.01102884_at Inpp5d Inositol polyphosphate-5-phosphatase, 145 kDa Phospholipid phosphatase �5.4 �0.001
�3.5 �0.001103433_at Pscd3 Pleckstrin homology, Sec7 and coiled/coil
domains 3Guanyl-nucleotide exchange factor �4.9 �0.001
�1.8 �0.51
50 V. Kriz et al. / Experimental Cell Research 286 (2003) 40–56
Probe set ID Symbol Gene Function Foldchange
Pvalue
102259_at Ywhag 3-Monooxygenase/tryptophan 5-monooxygenase activation protein, gammapolypeptide (14-3-3)
14-3-3, signal transduction protein, regulatesRaf and Bad
�6.3 �0.001�3.5 �0.25
93713_at Shc1 src homology 2 domain-containingtransforming protein C1
Adapter, intracellular signaling through ERKpathway, cell proliferation
�8.0 �0.001�6.0 �0.05
93902_at Gab1 Growth factor receptor-bound protein 2-associated protein 1
Docking protein downstrem of tyrosinekinase receptors
�5.2 �0.001�3.8 �0.001
104546_g_at Csnk2�1-rs4
Casein kinase II, alpha 1 related sequence 4 Protein Ser/Thr kinase �6.2 �0.001�2.6 �0.03
103078_at Trfr2 Transferrin receptor 2 Iron uptake �5.3 �0.001�3.1 �0.05
103957_at Trfr Transferrin receptor 2 Iron uptake �6.1 �0.001�5.9 �0.001
95974_at Gbp1 Guanylate nucleotide-binding protein 1 GTPase, immune response �4.7 �0.001�1.7 �0.45
96053_i_at Acp1 Protein-tyrosine phosphatase IF1 Protein-tyrosine phosphatase �2.6 �0.001�2.8 �0.001
97264_r_at Csnk1d Casein kinase 1, delta p53-targeted kinases, regulates DNA repair,chromosomal segregation and cell division
�12.8 �0.001�2.3 �0.13
97979_at Ppp1r7 Protein phosphatase 1, regulatory (inhibitor)subunit 7
Protein phosphatase �2.3 �0.001�2.1 �0.001
96771_at C76256 ERB3 receptor tyrosine kinase Receptor tyrosine kinase �2.4 �0.001�2.2
104716_at Rbp1 Retinol-binding protein 1 Retinol carrier �2.8 �0.001�2.4 �0.001
92247_at Arhgap5 Rho GTPase activating protein 5 Regulation of Rho activity, cytoskeleton �3.7 �0.001�3.9 �0.07
99051_at mts1(p16INK4A)
mts1 protein gene, exon 2. Cyclin-dependent kinase inhibitor (CDKI),negative regulator of the cell cycle
�2.8 �0.001�2.4 �0.01
92782_at Tmpo Thymopoietin Plays a role in nuclear envelope organizationand cell cycle control
�2.6 �0.001�3.5 �0.001
93929_s_at Mrp/plf3
Mitogen regulated protein/proliferin 3 Secreted protein of the same class as growthhormone, placental lactogen, and prolactin
�6.5�8.0
�0.001�0.001
Intracellular transport
Probe set ID Symbol Gene Function Foldchange
Pvalue
104248_at 0610038P07Rik Translocon-associated protein, gamma subunit Signal recognition particle receptor associated,component of the translocon
�2.9 �0.001104249_g_at �2.5 �0.001
�4.5 �0.001�4.5 �0.001
103680_at 1300011P19Rik Signal recognition particle receptor alphasubunit
Protein synthesis, docks the signal recognitionparticle arrested complex
�11.7 �0.001�9.6 �0.001
101579_at Srp9 Signal recognition particle 9 kDa Protein synthesis, confers elongation arrest andER targeting
�2.5 �0.001�1.8 �0.001
93930_at Lasp1 LIM and SH3 protein 1 Involved in regulation of membrane/vesicletrafficking
�3.3 �0.001�10.0 �0.02
93164_at Rnf2 Ring finger protein 2 Binds the ubiquitin-conjugating enzyme Hip-2,shows ubiquitin ligase (E3) activity in thepresence of Hip-2
�4.2 �0.001�4.6 �0.001
92288_at Ap1g1 Adapter protein complex AP-1, gamma 1subunit
Vesicle-mediated transport, Golgi �4.8 �0.001�6.0 �0.02
93717_at Rab23 RAB23, member RAS oncogene family Small monomeric GTPase, intracellulartransport
�3.3 �0.001�5.1 �0.001
93857_at Kpna3 Karyopherin (importin) alpha 3 Protein-nucleus transport �8.3 �0.001�4.4 �0.01
93711_at Sec23a SEC23A (S. cerevisiae) Component of the coat protein II (COTII)complex responsible for vesicle budding fromthe endoplasmic reticulum (ER)
�6.6 �0.001�9.9 �0.01
(continued on next page)
51V. Kriz et al. / Experimental Cell Research 286 (2003) 40–56
Appendix 1 (continued)
Appendix 1 (continued)
Immune response/defense mechanisms
Probe set ID Symbol Gene Function Foldchange
Pvalue
94061_at Crip Cysteine-rich intestinal protein LIM-family member, possibly involved in T-cell differentiation
�3.1 �0.001�2.1 �0.001
102736_at Scya2 Small inducible cytokine A2 Monocyte chemoattractant �4.4 �0.001�2.6 �0.01
100914_at Cd84 CD84 antigen Immunoglobulin gene family member of cellsurface molecules
�7.4 �0.001�3.0 �0.1
101879_s_at Crry Complement receptor-related protein Complement regulator �5.1 �0.001�2.6 �0.6
93543_f_at Gstm1 Glutathione S-transferase, mu 1 Glutathione S-transferase �2.5 �0.001�1.8 �0.001
96730_at Tpp2 Tripeptidyl peptidase II Cytoplasmic, high-molecular-weight protease,participates in the apoptotic pathway upstreamof caspase 1
�3.5 �0.001�2.7 �0.17
93836_at Bnip3 BCL2/adenovirus E1b 19-kDa-interactingprotein 1, NIP2
Regulates apoptosis �2.9 �0.001�2.0
100332_s_at Aop2-rs3 Antioxidant protein 2, related sequence 3 Mitochondrial antioxidant �3.1 �0.001�2.8 �0.001
97689_at F3 Coagulation factor III Blood coagulation factor �5.7 �0.001�4.5 �0.01
95313_at F2r Coagulation factor II (thrombin) receptor Receptor, involved in blood coagulation �5.2 �0.001�8.8 �0.001
94147_at Serpine 1 Serine (or cysteine) proteinase inhibitor, cladeE (nexin, plasminogen activator inhibitor type1), member 1
Serpin, serine protease inhibitor, involved inblood clotting cascade
�2.6�2.1
�0.001�0.01
95094_g_at Capn12 Calpain 12 Member of the calpain large subunit genefamily
�2.9 �0.001�2.2 �0.001
100472_at Ndpp1 NPC-derived proline-rich protein 1 Member of the human caspase recruitmentdomain (CARD) containing gene family and asurvival factor
�2.1 �0.001�1.6 �0.001
Metabolism, biosynthesis, catabolism
Probe set ID Symbol Gene Function Fold change P value
99023_at Pafah1b2 Platelet-activating factor acetylhydrolase,isoform 1b, alpha 2 subunit
2-Acetyl-1-alkylglycerophosphocholineesterase, lipid catabolism, regulates neuronalcell differentiation
�8.7�7.6
�0.001�0.001
94197_at Ugcg UDP-glucose ceramide glucosyltransferase Glucosylceramide synthase, important fordevelopment and differentiation
�5.0 �0.001�7.4 �0.02
101455_at Umps Uridine monophosphate synthetase Pyrimidine biosynthesis �4.6 �0.001�3.0 �0.44
92292_at Slc2a3 Solute carrier family 2 member 3 Facilitated glucose transporter �3.7 �0.001�3.1 �0.001
96627_at Ebp Phenylalkylamine Ca2� antagonist (emopamil)-binding protein
Cholesterol biosynthesis, hematopoiesis �2.9 �0.001�2.2 �0.001
99039_g_at Adss2 Adenylosuccinate synthetase 2, nonmuscle Purine nucleotide biosynthesis �2.3 �0.001�1.6 �0.001
94834_at Ctsh Cathepsin H Cysteine-type endopeptidase in lysosome �8.8 �0.001�10.0 �0.01
101294_g_at G6pd2 Glucose-6-phosphate dehydrogenase 2 Glucose metabolism �2.8 �0.001�2.5 �0.001
93354_at Apoc1 Apolipoprotein CI Associates with lipoproteins in plasma;involved in insulin-mediated glucose uptakeand free fatty acid metabolism.
�3.5�3.4
�0.001�0.001
52 V. Kriz et al. / Experimental Cell Research 286 (2003) 40–56

Appendix 1 (continued)
Other
Probe set ID Symbol Gene Function Fold change P value
96657_at Sat Spermidine/spermine N1-acetyltransferase N-acetyltransferase in polyamine metabolism �2.2 �0.001�1.5 �0.001
103340_at Rhced Rhesus blood group CE and D Erythrocyte cell surface antigen �4.9 �0.001�2.4 �0.03
102327_at Aoc3 Amine oxidase, copper containing 3 Adipocyte oxidase �4.3 �0.001�2.2 �0.26
93916_at NN8-AG
Retinoic acid-responsive (NN8-4AG) Retinoic acid-responsive gene with unknownfunction
�5.8 �0.001�6.1 �0.03
95356_at Apoe Apolipoprotein E Apolipoprotein, major susceptibility factor forAlzheimer’s disease
�1.7 �0.001�1.6 �0.001
ESTs or genes with unknown function
Probe set ID Symbol Gene Function Foldchange
Pvalue
93909_f_at Ccr4 Carbon catabolite repression 4 homolog (S. cerevisiae) Unknown �3.6 �0.001�2.3 �0.001
94752_s_at Skir ski/sno related Unknown �9.8 �0.001�3.6 �0.001
100348_at EST �4.5 �0.001�4.5 �0.01
100920_at 2210039B01Rik RIKEN cDNA 2210039B01 gene �9.6 �0.001�7.5 �0.001
102349_at AI836109 Expressed sequence AI836109 �10.1 �0.001�2.8 �0.08
102875_at C80612 Expressed sequence C80612 �4.5 �0.001�2.1 �0.56
103746_at AU016842 Expressed sequence AU016842 �9.8 �0.001�7.6 �0.001
103768_at D19Ertd678e Expressed DNA segment, Chr 19, ERATO Doi 678 �6.2 �0.001�4.6 �0.04
103926_at 0610008L05Rik RIKEN cDNA 0610008L05 gene �4.6 �0.001�18.8 �0.001
104458_at EST �2.8 �0.001�2.7 �0.001
104584_f_at AU041752 Expressed sequence AU041752 �5.0 �0.001�6.1 �0.06
93201_at AU016353 Expressed sequence AU016353 �5.7 �0.001�5.4 �0.08
93742_at 5730449L18Rik RIKEN cDNA 5730449L18 gene �2.9 �0.001�3.2 �0.001
94238_at 2310046G15Rik RIKEN cDNA 2310046G15 gene 96%similarityto humanserineprotease
�3.1 �0.001�1.6 �0.38
95357_at 1200009124Rik RIKEN cDNA 1200009124 gene �3.9 �0.001�4.1 �0.001
95428_at D1Wsu40e DNA segment, Chr 1, Wayne State University 40,expressed
�2.4 �0.001�3.0 �0.001
95785_s_at Rab7-ps1 RAB7, member RAS oncogene family, pseudogene 1 �4.8 �0.001�4.3 �0.001
96886_at 2510015F01Rik RIKEN cDNA 2510015F01 gene �3.8 �0.001�2.7 �0.56
96907_at D8Wsu96e DNA segment, Chr 8, Wayne State University 96,expressed
�2.9 �0.001�2.6 �0.001
97131_at 1200014D22Rik RIKEN cDNA 1200014D22 gene �4.5 �0.001�3.4 �0.09
(continued on next page)
53V. Kriz et al. / Experimental Cell Research 286 (2003) 40–56

Probe set ID Symbol Gene Function Foldchange
Pvalue
97283_at EST �14 �0.001�8.6 �0.001
97352_f_at EST �2.6 �0.001�1.9 �0.001
98092_at D5Wsu111e DNA segment, Chr 5, Wayne State University 111,expressed
�2.5 �0.001�2.8 �0.001
99621_s_at D4Ertd314e DNA segment, Chr 4, ERATO Doi 314, expressed �2.7 �0.001�2.7 �0.001
99987_at EST �8.4 �0.001�7.5 �0.14
Note. Gene expression profiles were determined twice using the Affymetrix U74A chip. The Resolver software was used to calculate fold changes and Pvalues, after normalization for internal controls. The identities of the genes, names according to GenBank, and brief descriptions according to PubMed aregiven in the table.
Appendix 2: SHB-induced changes in gene expression compared with control ES cells
Probe set ID Symbol Full name Function Foldchange
Pvalue
96584_f_at AF067060 Clone L3 variable group of two-cell-stagegene family
Role in early development �4.8 �0.001�12.6 �0.001
97169_f_at AF067058 Clone L10 variable group of two-cell-stagegene family
Role in early development �4.5 �0.001�5.9 �0.001
102254_f_at AA289585 Clone L5 uniform group of two-cell-stagegene family
Role in early development �5.8 �0.001�2.7 �0.58
103389_at Lorsdh Lysine oxoglutarate reductase, saccharopinedehydrogenase
Enzyme in amino acid metabolism, role inneuronal cell development
�4.7 �0.001�5.3 �0.001
98028_at Twist Twist gene homolog, (Drosophila) Mesodermal differentiation �4.0 �0.001�2.6 �0.54
99810_at Gpx2-ps1 Glutathione peroxidase 2, pseudogene 1 �3.2 �0.001�2.0 �0.04
93456_r_at Bmp4 Bone morphogenic protein 4 TGF�-family member, differentiationfactor, regulates mesenchyme
�2.4 �0.001�1.6
100064_f_at Gja1 Gap junction membrane channel proteinalpha 1
Cell–cell communication, particularly inneuronal cells
�1.9 �0.001�1.6 �0.001
95356_at Apoe Apolipoprotein E Enzyme in lipid metabolism �1.8 �0.001�1.7 �0.001
92607_at Peg1/MEST Peg1/MEST protein (Peg1/MEST) gene Detoxification enzyme, involved in survivalof mesodermal cells
�7.5 �0.001�6.1 �0.001
92508_s_at Utrn Utrophin Dystrophin homologue, expressed mainlyin neuronal cells
�4.9 �0.001�1.7 �0.55
99023_at Pafah1b2 Platelet-activating factor acetylhydrolase,isoform
Enzyme in phospholipid metabolism,involved in neuronal cell differentiation
�3.8 �0.001�1.4 �0.57
1b, alpha2 subunit100453_at Camk2b Calcium/calmodulin-dependent protein
kinase II, betaCalcium signal transduction �3.4 �0.001
�1.5 �0.23101883_s_at Xlr3b X-linked lymphocyte-regulated 3b �2.6 �0.001
�2.8 �0.001101990_at Ldh2 Lactate dehydrogenase 2, B chain Glycolytic enzyme �2.2 �0.001
�2.4 �0.00193028_at H19 H19 fetal liver mRNA Transcription factor, activated on ES cell
implantation�1.7 �0.001�2.0
98092_at D5Wsu111e Expressed DNA segment, Chr 5, WayneState University 111
�2.8 �0.001�3.6 �0.001
Note. Affymetrix U74A chips were used, and fold changes and P values after normalization for internal control were calculated using the Resolver software.
54 V. Kriz et al. / Experimental Cell Research 286 (2003) 40–56
Appendix 1 (continued)

References
[1] M. Welsh, J. Mares, T. Karlsson, C. Lavergne, B. Breant, L. Claes-son-Welsh, Shb is a ubiquitously expressed Src homology 2 protein,Oncogene 9 (1994) 19–27.
[2] M. Welsh, Z. Songyang, J.D. Frantz, T. Trub, K.A. Reedquist, T.Karlsson, M. Miyazaki, L.C. Cantley, H. Band, S.E. Shoelson,Stimulation through the T cell receptor leads to interactions be-tween SHB and several signaling proteins, Oncogene 16 (1998)891–901.
[3] C.K. Lindholm, J.D. Frantz, S.E. Shoelson, M. Welsh, Shf, a Shb-likeadapter protein, is involved in PDGF-alpha-receptor regulation ofapoptosis, Biochem. Biophys. Res. Commun. 278 (2000) 537–543.
[4] T. Karlsson, Z. Songyang, E. Landgren, C. Lavergne, P. Di Fiore, M.Anafi, T. Pawson, L.C. Cantley, L. Claesson-Welsh, M. Welsh, Mo-lecular interactions of the Src homology 2 domain protein Shb withphosphotyrosine residues, tyrosine kinase receptors and Src homol-ogy 3 domain proteins, Oncogene 10 (1995) 1475–1483.
[5] T. Karlsson, K. Kullander, M. Welsh, The Src homology 2 domainprotein Shb transmits basic fibroblast growth factor- and nervegrowth factor-dependent differentiation signals in PC12 cells, CellGrowth Differ 9 (1998) 757–766.
[6] M. Cross, L. Lu, P. Magnusson, D. Nyqvist, K. Holmqvist, M. Welsh,L. Claesson-Welsh, The Shb adaptor protein binds tyrosine 766 in theFGFR-1 and regulates the Ras/MEK/MAPK pathway via FRS2 phos-phorylation in endothelial cells, Mol. Biol. Cell 13 (2002) 2881–2893.
[7] T. Karlsson, M. Welsh, Apoptosis of NIH3T3 cells overexpressingthe Src homology 2 domain protein Shb, Oncogene 13 (1996) 955–961.
[8] J. Dixelius, H. Larsson, T. Sasaki, K. Holmqvist, L. Lu, A. Engstrom,R. Timpl, M. Welsh, L. Claesson-Welsh, Endostatin-induced tyrosinekinase signaling through the Shb adaptor protein regulates endothelialcell apoptosis, Blood 95 (2000) 3403–3411.
[9] M. Welsh, L. Christmansson, T. Karlsson, S. Sandler, N. Welsh,Transgenic mice expressing Shb adaptor protein under the control ofrat insulin promoter exhibit altered viability of pancreatic islet cells,Mol. Med. 5 (1999) 169–180.
[10] C. Anneren, Dual role of the tyrosine kinase GTK and the adaptorprotein SHB in beta-cell growth: enhanced beta-cell replication after60% pancreatectomy and increased sensitivity to streptozotocin, J.Endocrinol. 172 (2002) 145–153.
[11] L. Lu, K. Holmqvist, M. Cross, M. Welsh, Role of the Src homology2 domain-containing protein Shb in murine brain endothelial cellproliferation and differentiation, Cell Growth Differ 13 (2002) 141–148.
[12] C.K. Lindholm, E. Gylfe, W. Zhang, L.E. Samelson, M. Welsh,Requirement of the Src homology 2 domain protein Shb for T cellreceptor-dependent activation of the interleukin-2 gene nuclear factorfor activation of T cells element in Jurkat T cells, J. Biol. Chem. 274(1999) 28050–28057.
[13] A. Fraichard, O. Chassande, G. Bilbaut, C. Dehay, P. Savatier, J.Samarut, In vitro differentiation of embryonic stem cells into glialcells and functional neurons, J. Cell Sci. 108 (1995) 3181–3188.
[14] C. Bagutti, A.M. Wobus, R. Fassler, F.M. Watt, Differentiation ofembryonal stem cells into keratinocytes: comparison of wild-type andbeta 1 integrin-deficient cells, Dev. Biol. 179 (1996) 184–196.
[15] J.M. Metzger, W.I. Lin, LC. Samuelson, Transition in cardiac con-tractile sensitivity to calcium during the in vitro differentiation ofmouse embryonic stem cells, J. Cell Biol. 126 (1994) 701–711.
[16] J. Rohwedel, V. Maltsev, E. Bober, H.H. Arnold, J. Hescheler, A.M.Wobus, Muscle cell differentiation of embryonic stem cells reflectsmyogenesis in vivo: developmentally regulated expression of myo-genic determination genes and functional expression of ionic currents,Dev. Biol. 164 (1994) 87–101.
[17] U. Chen, Differentiation of mouse embryonic stem cells to lympho-hematopoietic lineages in vitro., Dev. Immunol. 2 (1992) 29–50.
[18] R. Wang, R Clark, V.L. Bautch, Embryonic stem cell-derived cysticembryoid bodies form vascular channels: an in vitro model of bloodvessel development, Development 114 (1992) 303–316.
[19] K. Abe, H. Niwa, K. Iwase, M. Takiguchi, M. Mori, S.I. Abe, K. Abe,K. Yamamura, Endoderm-specific gene expression in embryonicstem cells differentiated to embryoid bodies, Exp. Cell Res. 229(1996) 27–34.
[20] N. Lumelsky, O. Blondel, P. Laeng, I. Velasco, R. Ravin, R. McKay,Differentiation of embryonic stem cells to insulin-secreting structuressimilar to pancreatic islets, Science 292 (2001) 1389–1394.
[21] M. Schuldiner, O. Yanuka, J. Itskovitz-Eldor, D. A. Melton, N.Benvenisty, From the cover: effects of eight growth factors on thedifferentiation of cells derived from human embryonic stem cells,Proc. Natl. Acad. Sci. USA 97 (2000) 11307–11312.
[22] B. Soriat, A. Skoudy, F. Martin, From stem cells to beta-cells: newstrategies in cell therapy of diabetes mellitus, Diabetologia 44 (2001)407–415.
[23] M.F. Offield, T.L. Jetton, P.A. Labosky, M. Ray, R.W. Stein, M.A.Magnuson, B.L. Hogan, C.V. Wright, PDX-1 is required for pancre-atic outgrowth and differentiation of the rostral duodenum, Develop-ment 122 (1996) 983–995.
[24] R.G. Hawley, F.H. Lieu, A.Z. Fong, T.S. Hawley, Versatile retroviralvectors for potential use in gene therapy, Gene Ther. 1 (1994) 136–138.
[25] W.E. Li, P.A. Ochalski, E.L. Hertzberg, J.I. Nagy, Immunorecogni-tion, ultrastructure and phosphorylation status of astrocytic gap junc-tions and connexin43 in rat brain after cerebral focal ischaemia, Eur.J. Neurosci. 10 (1998) 2444–2463.
[26] V.V. Rao, X. Pan, Y.F. Chang, Developmental changes of L-lysine-ketoglutarate reductase in rat brain and liver, Comp. BiochemPhysiol. B. 103 (1992) 221–224.
[27] H. Donai, M. Nakamura, Y. Sogawa, J.K. Wang, M. Urushihara, T.Yamauchi, Involvement of Ca2� calmodulin-dependent protein ki-nase 11 in neurite outgrowth induced by cAMP treatment and serumdeprivation in a central nervous system cell line, CAD derived fromrat brain, Neurosci. Lett. 293 (2000) 111–114.
[28] U. Albrecht, R. Abu-Issa, B. Ratz, M. Hattori, J. Aoki, H. Arai, K.Inoue, G. Eichele, Platelet-activating factor acetylhydrolase expres-sion and activity suggest a link between neuronal migration andplatelet-activating factor, Dev. Biol. 180 (1996) 579–593.
[29] D.J. Blake, J.N. Schofield, R.A. Zuellig, D.C. Gorecki, S.R. Phelps,E.A. Bernard, Y.H. Edwards, K.E. Davies, G-utrophin, the autosomalhomologue of dystrophin Dp116, is expressed in sensory ganglia andbrain, Proc. Natl. Acad. Sci. USA 92 (1995) 3697–3701.
[30] C. Wolf, C. Thisse, C. Stoetzel, B. Thisse, P. Gerlinger, F. Perrin-Schmitt, The M-Twist gene of Mus is expressed in a subset ofmesodermal cells and is closely related to the Xenopus X-twi and theDrosophila twist genes, Dev. Biol. 143 (1991) 363–373.
[31] T. Kaneko-Ishino, Y. Kuriowa, N. Miyoshi, T. Kohda, R. Suzuki, M.Yokoyama, S. Viville, S.C. Barton, F. Ishino, M.A. Surani, PegI/Mestimprinted gene on chromosome 6 identified by cDNA subtractionhybridization, Nat. Genet. 11 (1995) 52–59.
[32] D. Stafford, V.E. Prince, Retinoic acid is required for a critical stepin zebrafish pancreatic development, Curr. Biol. 12 (2002) 1215–1220.
[33] L. Lei, L. Ma, S. Nef, T. Thai, L.F. Parada, mKlf7, a potentialtranscriptional regulator of TrkA nerve growth factor receptor expres-sion in sensory and sympathetic neurons, Development 128 (2001)1147–1158.
[34] M. Katoh, Molecular cloning and characterization of human SOX17,Int. J. Mol. Med. 9 (2002) 153–157.
[35] H. Sasaki, Y. Nishizaki, C. Hui, M. Nakafuku, H. Kondoh, Regula-tion of Gli2 and Gli3 activities by an amino-terminal repressiondomain: implication of Gli2 and Gli3 as primary mediators of Shhsignaling, Development 126 (1999) 3915–3924.
55V. Kriz et al. / Experimental Cell Research 286 (2003) 40–56

[36] P. Bouillet, C. Chazaud, M. Oulad-Abdelghani, P. Dolle, P. Cham-bon, Sequence and expression patter of the Stra7 (Gbx-2) homeobox-containing gene induced by retinoic acid in P19 embryonal carcinomacells, Dev. Dyn. 204 (1995) 372–382.
[37] G. Chapman, J.L. Remiszewski, G.C. Webb, T.C. Schultz, C.D.K.Bottema, P.D. Rathjen, The mouse homeobox gene, Gbx2: genomicorganization and expression in pluripotent cells in vitro and in vivo,Genomics 46 (1997) 223–233.
[38] X. Li, Y. Chen, S. Scheele, E. Arman, R. Haffner-Krausz, P. Ekblom,P. Lonai, Fibroblast growth factor signaling and basement membraneassembly are connected during epithelial morphogenesis of the em-bryoid body, J. Cell Biol. 153 (2001) 811–822.
[39] P. Maye, S Becker, E. Kasameyer, N, Byrd, L Grabel, Indianhedgehog signaling in extraembryonic endoderm and ectoderm differentia-tion in ES embryoid bodies, Mech Dev. 94 (2000) 117–132.
[40] E. Coucouvanis, G.R. Martin, BMP signaling plays a role in visceralendoderm differentiation and cavitation in the early mouse embryo,Development 126 (1999) 535–546.
[41] J. Rossant, Stem cell from mammalian blastocysts, Stem Cells 19(2001) 477–482.
[42] M. Boiani, S. Eckhardt, H.R. Scholer, K.J. McLaughlin, Oct4 distri-bution and level in mouse clones: consequences for pluripotency,Genes Dev. 16 (2002) 1209–1219.
[43] J. Jonsson, L. Carlsson, T. Edlund, H. Edlund, Insulin-promoter-factor 1 is required for pancreas development in mice, Nature 371(1994) 606–609.
[44] A. Gualandris, J.P. Annes, M. Arese, I. Noguera, V. Jurukovski, D.B.Rifkin, The latent transforming growth factor-�-binding protein-1promotes in vitro differentiation of embryonic stem cells into endo-thelium, Mol. Biol. Cell 11 (2000) 4295–4308.
56 V. Kriz et al. / Experimental Cell Research 286 (2003) 40–56