the vascular endothelial growth factor (vegf)/ vegf ...€¦ · the vegf (vascular endothelial...
TRANSCRIPT

Clinical Science (2005) 109, 227–241 (Printed in Great Britain) doi:10.1042/CS20040370 227
R E V I E W
The vascular endothelial growth factor (VEGF)/VEGF receptor system and its role underphysiological and pathological conditions
Hiroyuki TAKAHASHI∗† and Masabumi SHIBUYA∗∗Division of Genetics, Institute of Medical Science, University of Tokyo, 4-6-1 Shirokane-dai, Minato-ku, Tokyo, 108-8639,Japan, and †Department of Internal Medicine, Graduate School of Medicine, University of Tokyo, 7-3-1 Hongo, Bunkyo-ku,Tokyo, 113-0033, Japan
A B S T R A C T
The VEGF (vascular endothelial growth factor) family and its receptors are essential regulatorsof angiogenesis and vascular permeability. Currently, the VEGF family consists of VEGF-A, PlGF(placenta growth factor), VEGF-B, VEGF-C, VEGF-D, VEGF-E and snake venom VEGF. VEGF-Ahas at least nine subtypes due to the alternative splicing of a single gene. Although the VEGF165
isoform plays a central role in vascular development, recent studies have demonstrated that eachVEGF isoform plays distinct roles in vascular patterning and arterial development. VEGF-A binds toand activates two tyrosine kinase receptors, VEGFR (VEGF receptor)-1 and VEGFR-2. VEGFR-2mediates most of the endothelial growth and survival signals, but VEGFR-1-mediated signallingplays important roles in pathological conditions such as cancer, ischaemia and inflammation. Insolid tumours, VEGF-A and its receptor are involved in carcinogenesis, invasion and distantmetastasis as well as tumour angiogenesis. VEGF-A also has a neuroprotective effect on hypoxicmotor neurons, and is a modifier of ALS (amyotrophic lateral sclerosis). Recent progress in themolecular and biological understanding of the VEGF/VEGFR system provides us with novel andpromising therapeutic strategies and target proteins for overcoming a variety of diseases.
INTRODUCTION
Angiogenesis and vasculogenesis are regulated pre-dominantly by several different growth factors and theirassociated RTKs (receptor tyrosine kinases). Foremost
among these is the VEGF (vascular endothelial growthfactor) family and VEGFRs (VEGF receptors). VEGF-A, also referred to as VPF (vascular permeability factor),an important regulator of endothelial cell physiology,was identified approx. 15 years ago [1,2] and has been
Key words: angiogenesis, inflammation, signal transduction, tumour, vascular endothelial growth factor (VEGF), vascularpermeability.Abbreviations: ALS, amyotrophic lateral sclerosis; DSCR1, Down syndrome critical region protein 1; ECM, extracellular matrix;ERK, extracellular signal-regulated kinase; FAK, focal adhesion kinase; HIF-1, hypoxia-inducible factor-1; HRE, hypoxia responseelement; HUVEC, human umbilical vein endothelial cell; LSEC, liver sinusoidal endothelial cell; mAb, monoclonal antibody;MAPK, mitogen-activated protein kinase; MMP, matrix metalloproteinase; NFATc, nuclear factor of activated T-cell; NO, nitricoxide; NOS; NO synthase; eNOS, endothelial NOS; NRP, neuropilin; PAIP2, polyadenylated-binding protein-interacting pro-tein 2; PDK1, phosphoinositide-dependent kinase 1; PI3K, phosphoinositide 3-kinase; PILSAP, puromycin-intensive leucyl-specificaminopeptidase; PKC, protein kinase C; PLC, phospholipase C; PlGF, placenta growth factor; pVHL, von Hippel–Lindau tumoursuppressor protein; RA, rheumatoid arthritis; RTK, receptor tyrosine kinase; S6K, S6 kinase; Tag, T antigen; TH2, T-helper type 2;UTR, untranslated region; VEGF, vascular endothelial growth factor; VEGFR, VEGF receptor; sVEGFR-1, soluble VEGFR-1;svVEGF, snake venom VEGF; Tf svVEGF, Trimeresurus flavoviridis svVEGF; VPF, vascular permeability factor.Correspondence: Professor Masabumi Shibuya (email [email protected]).
C© 2005 The Biochemical Society

228 H. Takahashi and M. Shibuya
recognized as the major growth factor that is relativelyspecific for endothelial cells. VEGF-A is a dimeric glyco-protein essential for many angiogenic processes in normaland abnormal states, such as tumour vascularization,mainly by interacting with two tyrosine kinase receptors,VEGFR-1 [also known as Flt-1 (Fms-like tyrosine kin-ase-1)] and VEGFR-2 [also known as Flk-1 (fetal liverkinase-1) and, in humans, as KDR (kinase insert domain-containing receptor)] [3–5]. VEGF-A exhibits two majorbiological activities: one is the capacity to stimulatevascular endothelial cell proliferation [1,6,7], and theother is the ability to increase vascular permeability [2,8].VEGF-A also promotes the survival and migration ofendothelial cells. In addition, recent studies have revealeda variety of biological functions and the precise molecularmechanisms of the VEGF/VEGFR system. In this review,we will discuss the recent advances in the basic biology ofthe VEGF/VEGFR system, which give insight into manyphysiological and pathological conditions.
VEGF AND VEGF FAMILY PROTEINS
Currently, the VEGF family includes VEGF-A, PlGF(placenta growth factor), VEGF-B, VEGF-C, VEGF-D,VEGF-E and svVEGF (snake venom VEGF). The mol-ecular and biological functions of each ligand have beenwell characterized.
VEGF-AStructurally, VEGF belongs to the VEGF/PDGF (plate-let-derived growth factor) supergene family. Among thegene products of this family, eight cysteine residues areconserved at the same positions. These products functionas a dimer, since two out of eight cysteine residues gen-erate intermolecular cross-linked S–S bonds [9]. Thehuman VEGF-A gene is organized into eight exons,separated by seven introns [10,11] and is located at 6p21.3[12].
Human VEGF-A has at least nine subtypes due to thealternative splicing of a single gene: VEGF121, VEGF145,VEGF148, VEGF162, VEGF165, VEGF165b, VEGF183,VEGF189 and VEGF206 [13,14] (Figure 1). VEGF165b is anendogenous inhibitory form of VEGF, which bindsVEGFR-2 with the same affinity as VEGF165, but doesnot activate it or stimulate downstream signalling path-ways [15]. VEGF is produced in endothelial cells, macro-phages, activated T-cells and a variety of other cell types[16–18]. Although virtually nothing is known abouthow VEGF isoform levels are regulated, most VEGF-producing cells appear to preferentially express VEGF121,VEGF165 and VEGF189. VEGF165, the predominant iso-form, is secreted as an approx. 46 kDa homodimer, whichhas a basic character and moderate affinity for heparin,owing to the presence of 15 basic amino acids within
the 44 residues encoded by exon 7 [1,2,7]. In contrast,VEGF121, which lacks the residues encoded by exons6 and 7, does not bind heparin and is freely releasedfrom the cell. VEGF189, which contains an additionalsequence encoded by exon 6, binds heparin strongly and iscompletely sequestered in the ECM (extracellular matrix)and to a lesser extent at the cell surface [16]. The ECM-bound isoforms can be released slowly by exposure toheparin and heparinases, or more rapidly released asbioactive fragments through cleavage by plasmin or uro-kinase at the C-terminus [19]. VEGF165 binds the co-receptors NRP-1 (neuropilin-1) [20] and NRP-2 (neuro-pilin-2), whereas VEGF145 binds only NRP-2 [21](Figure 2).
Approx. 50 % of mice expressing exclusively theVEGF120 isoform (murine VEGF is shorter by one aminoacid) die within a few hours after birth and the rest diewithin 14 days due to impaired myocardial angiogenesisand ischaemic cardiomyopathy [22]. VEGF120/120 micealso exhibit a specific decrease in capillary branch form-ation and the impairment of the directed extension ofendothelial cell filopodia during embryogenesis [23] aswell as severe defects in retinal vascular outgrowthand patterning [24], suggesting that the heparin-bindingVEGF isoforms provide spatially restricted stimulatorycues to initiate vascular branch formation. VEGF164/164
mice are normal and healthy, and have a normal retinalangiogenesis, whereas VEGF188/188 mice display normalvenular outgrowth but impaired arterial development inretinas as well as dwarfism, disrupted developmentof growth plates and secondary ossification centres, andknee joint dysplasia [25]. These findings suggest that thevarious VEGF isoforms play distinct roles in vascular pat-terning and arterial development, although the VEGF164
isoform plays a central role in vascular development.Gene expression of VEGF is regulated by a variety of
stimuli such as hypoxia, growth factors, transformation,p53 mutation, oestrogen, TSH (thyroid-stimulatinghormone), tumour promoters and NO (nitric oxide).Although all of the stimuli responsible for the up-regulation of the VEGF gene are quite interesting, hyp-oxia has been of particular interest because of its import-ance and the unique transcriptional regulation involved.It is now well established that HIF-1 (hypoxia-inducible factor-1) is a key mediator of hypoxic res-ponses. HIF-1 is a transcriptional activator composedof HIF-1α and HIF-1β subunits. Both HIF-1α andHIF-1β are constitutively expressed in various types oftumour. Under normal oxygenation conditions, HIF-1α is scarcely detectable because it is targeted for rapiddestruction by an E3 ubiquitin ligase containing pVHL(von Hippel–Lindau tumour suppressor protein). Theinteraction between pVHL and a specific domain ofthe HIF-1α subunit is regulated through hydroxylationof a proline residue (Pro564 in HIF-1α) by prolyl-4-hydroxylase, which requires molecular oxygen and
C© 2005 The Biochemical Society
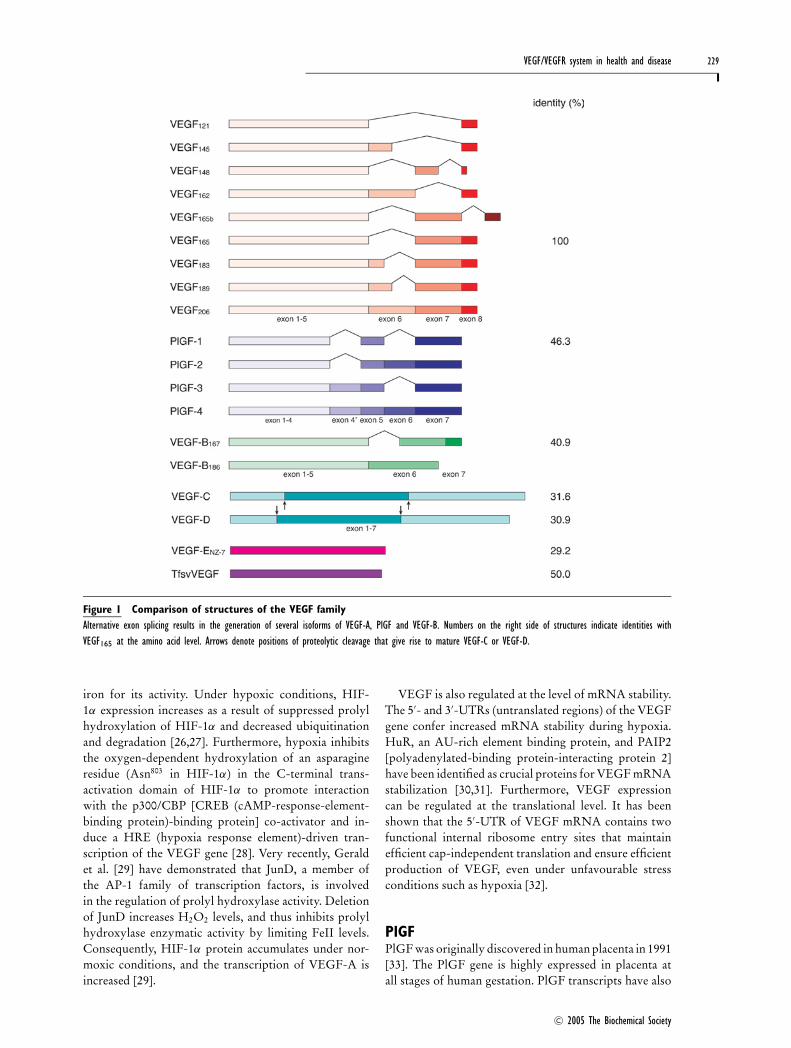
VEGF/VEGFR system in health and disease 229
Figure 1 Comparison of structures of the VEGF familyAlternative exon splicing results in the generation of several isoforms of VEGF-A, PlGF and VEGF-B. Numbers on the right side of structures indicate identities withVEGF165 at the amino acid level. Arrows denote positions of proteolytic cleavage that give rise to mature VEGF-C or VEGF-D.
iron for its activity. Under hypoxic conditions, HIF-1α expression increases as a result of suppressed prolylhydroxylation of HIF-1α and decreased ubiquitinationand degradation [26,27]. Furthermore, hypoxia inhibitsthe oxygen-dependent hydroxylation of an asparagineresidue (Asn803 in HIF-1α) in the C-terminal trans-activation domain of HIF-1α to promote interactionwith the p300/CBP [CREB (cAMP-response-element-binding protein)-binding protein] co-activator and in-duce a HRE (hypoxia response element)-driven tran-scription of the VEGF gene [28]. Very recently, Geraldet al. [29] have demonstrated that JunD, a member ofthe AP-1 family of transcription factors, is involvedin the regulation of prolyl hydroxylase activity. Deletionof JunD increases H2O2 levels, and thus inhibits prolylhydroxylase enzymatic activity by limiting FeII levels.Consequently, HIF-1α protein accumulates under nor-moxic conditions, and the transcription of VEGF-A isincreased [29].
VEGF is also regulated at the level of mRNA stability.The 5′- and 3′-UTRs (untranslated regions) of the VEGFgene confer increased mRNA stability during hypoxia.HuR, an AU-rich element binding protein, and PAIP2[polyadenylated-binding protein-interacting protein 2]have been identified as crucial proteins for VEGF mRNAstabilization [30,31]. Furthermore, VEGF expressioncan be regulated at the translational level. It has beenshown that the 5′-UTR of VEGF mRNA contains twofunctional internal ribosome entry sites that maintainefficient cap-independent translation and ensure efficientproduction of VEGF, even under unfavourable stressconditions such as hypoxia [32].
PlGFPlGF was originally discovered in human placenta in 1991[33]. The PlGF gene is highly expressed in placenta atall stages of human gestation. PlGF transcripts have also
C© 2005 The Biochemical Society

230 H. Takahashi and M. Shibuya
Figure 2 Schematic diagram illustrating the receptor-binding specificity of VEGF family members and the VEGFR-2 signallingpathwaysThe VEGF family of ligands and their receptor-binding patterns are shown at the top. Downstream VEGFR signalling pathways focusing on VEGFR-2 are shown at thebottom. Tyr1175 (Y1175) and Tyr1214 (Y1214) are the two major autophosphorylation sites in VEGFR-2. PLC-γ binds to Y1175, leading to the phosphorylation andactivation of this protein. Y1214 appears to be required to trigger the sequential activation of Cdc42 and p38 MAPK. Many proteins are activated by VEGFR-2 throughan unknown mechanism, including FAK, PI3K and Src. The activation of downstream signal transduction molecules leads to several different endothelial cell functionssuch as migration, vascular permeability, survival and proliferation.
been detected in the heart, lung, thyroid gland and skeletalmuscle [34]. PlGF binds VEGFR-1, but not VEGFR-2[35,36]. Alternative splicing of the human PlGF genegenerates four isoforms which differ in size and bindingproperties: PlGF-1 (PlGF131), PlGF-2 (PlGF152), PlGF-3(PlGF203) and PlGF-4 (PlGF224) [37–39] (Figure 1).PlGF-1 is the shortest isoform and a non-heparin bindingprotein. PlGF-2 is able to bind heparin and the co-receptors NRP-1 and NRP-2 due to the insertion of ahighly basic 21-amino acid sequence encoded by exonVI near the C-terminus [37] (Figure 2). PlGF-3, whichcontains an insertion of 216 nucleotides coding for a 72-amino acid sequence between exons 4 and 5 of the PlGFgene but lacks the coding sequence of exon 6, is unable tobind heparin [38]. PlGF-4 consists of the same sequence
of PlGF-3, plus a heparin-binding domain previouslythought to be present only in PlGF-2 [39].
The crystal structure of human PlGF-1 has shownthat this protein is structurally similar to VEGF-A [40].Furthermore, despite this moderate sequence conser-vation, PlGF and VEGF-A bind to the same bindinginterface of VEGFR-1 in a very similar fashion [41].However, recent studies have reported that, unlike inVEGF-A, N-glycosylation in PlGF plays an importantrole in VEGFR-1 binding [42].
Carmeliet et al. [43] have shown that a deficiency inPlGF (PlGF−/−) does not affect embryonic angiogenesisin mice. However, loss of PlGF impairs angio-genesis, plasma extravasation and collateral growth du-ring ischaemia, inflammation, wound healing and cancer,
C© 2005 The Biochemical Society

VEGF/VEGFR system in health and disease 231
indicating the importance of VEGFR-1 signalling inpathological conditions.
VEGF-BVEGF-B has a wide tissue distribution, but is particularlyabundant in the heart and skeletal muscle [44]. HumanVEGF-B has two isoforms generated by alternativesplicing: VEGF-B167 and VEGF-B186 (Figure 1). TheVEGF-B isoforms bind and activate VEGFR-1 and canalso bind to NRP-1 [44] (Figure 2).
Studies using VEGF-B knockout (VEGF-B−/−) micehave yielded slightly conflicting results regarding therole of VEGF-B in angiogenesis and the developmentof the cardiovascular system. VEGF-B−/− mice areviable and fertile; however, although Bellomo et al. [45]demonstrated that VEGF-B−/− mice had smaller hearts,dysfunctional coronary arteries and an impaired recoveryfrom experimentally induced myocardial ischaemia, Aaseet al. [46] claimed that these mice showed a subtle cardiacphenotype such as an atrial conduction abnormalitycharacterized by a prolonged PQ interval, and thatVEGF-B was not required for proper development ofthe cardiovascular system either during developmentor angiogenesis in adults. Recent studies using VEGF-B−/− mice have demonstrated the role of VEGF-Bin pathological vascular remodelling in inflammatoryarthritis [47] and protection of the brain from ischaemicinjury [48].
VEGF-C and VEGF-DVEGF-C contains a region sharing approx. 30 % aminoacid identity with VEGF165; however, it is more closelyrelated to VEGF-D by virtue of the presence of N- andC-terminal extensions that are not found in other VEGFfamily members [49] (Figure 1). Both VEGF-C andVEGF-D bind and activate VEGFR-3 (Flt-4; a member ofthe VEGFR family that does not bind VEGF-A) as wellas VEGFR-2, and are mitogenic for cultured endothelialcells. VEGF-C also binds to NRP-2 [49] (Figure 2). BothVEGF-C and VEGF-D are produced as a preproproteinwith long N- and C-terminal propeptides flanking theVEGF homology domain. Initial proteolytic cleavage ofthe precursor generates a form with a moderate affinityfor VEGFR-3, but a second proteolytic step is requiredto produce the fully processed form with a high affinityfor both VEGFR-2 and VEGFR-3 [49]. This activationof VEGF-C and VEGF-D by proteolytic cleavage is atleast partly regulated by the serine protease plasmin [50].
Overexpression of VEGF-C in the epidermis oftransgenic mice results in the development of a hyper-plastic lymphatic vessel network [51]. In vitro, VEGF-C and VEGF-D stimulate the migration and mitogenesisof cultured endothelial cells [49]. A recent study usingVEGF-C−/− mice has demonstrated that VEGF-C isrequired for the initial steps in lymphatic developmentand that both VEGF-C alleles are required for normal
lymphatic development [52]. Thus VEGF-C is the para-crine factor essential for lymphangiogenesis. Less isknown of the function of VEGF-D, but Stacker et al.[53] have revealed that VEGF-D induces the formation oflymphatics within tumours and promotes the metastasisof tumour cells.
VEGF-EHomologues of VEGF have also been identified in thegenome of the parapoxvirus Orf virus [54] and have beenshown to have VEGF-A-like activities. VEGF-E is thecollective term for a group of these proteins, includingVEGF-ENZ-2 (VEGF from Orf virus strain NZ-2)[55], VEGF-ENZ-7 (VEGF from Orf virus strain NZ-7)[56], VEGF-ENZ-10 (VEGF from Orf virus strain NZ-10) [57], VEGF-ED1701 (VEGF from Orf virus strainD1701) [58] and VEGF-EVR634 (VEGF from Pseudo-cowpox virus strain VR634) [57]. All VEGF-E variantsstudied bind and activate VEGFR-2, but not VEGFR-1or VEGFR-3. VEGF-ENZ-2, VEGF-ENZ-10 and VEGF-ED1701 can bind NRP-1. VEGF-ENZ-7 and VEGF-EVR634,however, are unable to bind NRP-1 (Figure 2). VEGF-Eseems to be as potent as VEGF165 at stimulating endo-thelial cell proliferation despite lacking a heparin-bindingbasic domain. K14-driven VEGF-ENZ-7 transgenic micehave shown a significant increase in angiogenesis at sub-cutaneous tissue without clear side effects [59].
svVEGFRecently, VEGF family proteins have been identified insnake venom, including svVEGF from Bothrops insularis[60] and Tf svVEGF (Trimeresurus flavoviridis svVEGF)[61] from pit vipers in addition to HF (hypotensivefactor) [62], ICPP (increasing capillary permeabilityprotein) [63] and vammin [64] from vipers. Takahashiet al. [61] have shown that snakes utilize these venom-specific VEGFs in addition to VEGF-A. svVEGFs func-tion as dimers and each chain comprises approx. 110–122 amino acid residues. The cysteine knot motif, acharacteristic of the VEGF family of proteins, is com-pletely conserved in svVEGFs and the sequence identitywith human VEGF165 is approx. 50 % (Figure 1).Vammin does not bind VEGFR-1 but binds VEGFR-2with high affinity as well as VEGF165 [64]. However,Tf svVEGF binds VEGFR-1 with high affinity andVEGFR-2 with low affinity compared with VEGF165,leading to a strong enhancement of vascular permeabilitybut weak stimulation of endothelial cell proliferation[61] (Figure 2). Both vammin and Tf svVEGF are unableto bind VEGFR-3 or NRP-1, but Tf svVEGF bindsheparin. svVEGFs may contribute to the enhancementof toxicity in envenomation, but they seem to have indi-vidual biological characteristics reflecting divergence inthe classification of the host snake.
C© 2005 The Biochemical Society

232 H. Takahashi and M. Shibuya
VEGFRs
VEGFR-1VEGFR-1 is a 180 kDa high-affinity receptor for VEGF-A, VEGF-B, PlGF and Tf svVEGF. It is expressed invascular endothelial cells and a range of non-endo-thelial cells, including macrophages and monocytes [65],and haematopoietic stem cells [66]. The second Ig domainof VEGFR-1 is the major binding site for VEGF-Aand PlGF [16,41,67]. VEGFR-1 binds VEGF-A with atleast 10-fold higher affinity than VEGFR-2 (Kd = 10–30 pM) [16]; however, ligand binding results in a maximal2-fold increase in kinase activity. In many cases, theeffects of VEGFR-2 on endothelial cells, such as thoseon cell survival and proliferation, can be induced onlyweakly or slightly by treatment with VEGFR-1-specificligands. VEGFR-1 is a negative regulator of angiogenesisduring early development, but plays an important role inangiogenesis under pathological conditions (as describedbelow). VEGFR-1-blocking antibodies prevent themigration but not proliferation of HUVECs (humanumbilical vein endothelial cells) in response to VEGF-A,indicating the involvement of VEGFR-1 in endothelialcell migration [68]. VEGFR-1-mediated signalling ap-pears to preferentially modulate the reorganization ofactin via p38 MAPK (mitogen-activated protein kinase),whereas VEGFR-2 contributes to the re-organizationof the cytoskeleton by phosphorylating FAK (focaladhesion kinase) and paxillin (Figure 2), suggesting adifferent contribution of the two receptors to the chemo-tactic response. VEGFR-1 signalling is also involved inthe migration of monocytes/macrophages [65] and in thereconstitution of haematopoiesis by recruiting haemato-poietic stem cells [66].
An alternatively spliced form of VEGFR-1 that en-codes a soluble truncated form of the receptor, containingonly the first six Ig domains, has been cloned froma HUVEC cDNA library [16]. sVEGFR-1 (solubleVEGFR-1) inhibits VEGF-A activity by sequester-ing VEGF-A from signalling receptors and by formingnon-signalling heterodimers with VEGFR-2 [69]. Plasmalevels of sVEGFR-1 are elevated in individuals withcancer, ischaemia and pre-eclampsia [70–72]. A recentstudy has demonstrated that elevated levels of sVEGFR-1play an important role in pre-eclampsia [73]. Increasedcirculating levels of sVEGFR-1 in patients with pre-eclampsia are associated with decreased circulating levelsof free VEGF and PlGF, resulting in general endothelialdysfunction [73].
VEGFR-2VEGFR-2 is a 200–230 kDa high-affinity receptor forVEGF-A (Kd = 75–760 pM), VEGF-E and svVEGFs aswell as the processed form of VEGF-C and VEGF-D.The binding site for VEGF-A has been mapped to the
second and third Ig domains [74]. VEGFR-2 is ex-pressed in vascular and lymphatic endothelial cells, andother cell types such as megakaryocytes and haemato-poietic stem cells [75]. Tyrosine phosphorylation sitesin human VEGFR-2 bound to VEGF-A are Tyr951 andTyr996 in the kinase-insert domain, Tyr1054 and Tyr1059
in the kinase domain, and Tyr1175 and Tyr1214 in theC-terminal tail. Among them, Tyr1175 and Tyr1214 arethe two major VEGF-A-dependent autophosphorylationsites [76]. Tyr951 creates a binding site for the VEGFR-associated protein [77] and Tyr1175 creates a binding sitefor Sck [78], Shb [79] and PLC (phospholipase C)-γ [76].
VEGFR-2 is the major mediator of the mitogenic,angiogenic and permeability-enhancing effects of VEGF-A. Furthermore, recent studies have indicated thatthe activation of VEGFR-2 also promotes lymphan-giogenesis [80,81]. Survival signalling for endothelialcells from VEGFR-2 is reported to involve thePI3K (phosphoinositide 3-kinase)/Akt pathway [82,83](Figure 2). However, another pathway may be involved,since the signal to activate PI3K by VEGFR-2 is usuallynot very strong. Byzova et al. [84] have reported thatthe activation of VEGFR-2 by VEGF-A results in thePI3K/Akt-dependent activation of several integrins,leading to enhanced cell adhesion and migration. Thissynergic interaction with integrins is required forproductive signalling from VEGFR-2.
Very recently, a naturally occurring soluble truncatedform of VEGFR-2 has been detected in mouse andhuman plasma [85]. Similar to sVEGFR-1, sVEGFR-2(soluble VEGFR-2) may have regulatory consequenceswith respect to VEGF-mediated angiogenesis.
VEGFR-3VEGFR-3 is a 195 kDa high-affinity receptor for VEGF-C and VEGF-D. Unlike VEGFR-1 and VEGFR-2,VEGFR-3 is proteolytically cleaved within the fifthextracellular Ig loop into a 120 kDa and a 75 kDa formduring synthesis, and the two forms are linked bya disulphide bridge [49]. Overexpression of a solubleVEGFR-3 in the skin of mice inhibits fetal lymphan-giogenesis and induces a regression of already formedlymphatic vessels [86]. Furthermore, overexpression of aVEGFR-3-specific mutant of VEGF-C (VEGF-C 156S)in the skin induces the growth of lymphatic vesselswithout an influence on the blood vessel architecture[87], indicating that stimulation of VEGFR-3 alone issufficient to induce lymphangiogenesis. The stimulationof VEGFR-3 also protects the lymphatic endothelialcells from serum deprivation-induced apoptosis. Thephosphorylation of VEGFR-3 has been shown to leadto a PI3K-dependent activation of Akt and PKC (proteinkinase C)-dependent activation of p42/p44 MAPK [88].A recent study [89] has demonstrated that blockadeof VEGFR-3 signalling significantly suppresses corneal
C© 2005 The Biochemical Society

VEGF/VEGFR system in health and disease 233
dendritic cell trafficking to draining lymph nodes as wellas the induction of delayed-type hypersensitivity andrejection of corneal transplants, suggesting a role forVEGFR-3 in adaptive immunity.
NRP-1 and NRP-2NRP-1 is a 130–140 kDa cell-surface glycoprotein firstidentified as a semaphorin receptor involved in neuronalguidance [90] and subsequently found as an isoform-specific receptor for VEGF-A [20]. NRP-2 was identifiedby virtue of its sequence homology with NRP-1 andshares 44 % identity at the amino acid level with NRP-1[90]. NRP-1 is able to bind VEGF165, VEGF-B, PlGF-2and some VEGF-E variants, whereas NRP-2 can bindVEGF145, VEGF165, PlGF-2 and VEGF-C. The intra-cellular domains of NRPs are short and do not sufficefor the independent transduction of biological signalssubsequent to semaphorin or VEGF binding. It has beenshown that both NRPs can join with receptors belongingto the plexin family, and such plexin/NRP complexes areable to transduce signals as the physiological receptor ofclass-3 semaphorins [91,92]. The VEGF165-induced pro-liferation and migration of cells that express VEGFR-2are enhanced in the presence of NRP-1. Thus NRP-1also seems to function as an enhancer of VEGFR-2activity in the presence of VEGF165. Recent studies havedemonstrated that this effect is the result of the formationof a complex between VEGFR-2 and NRP-1 [93,94].
An in vivo study with transgenic mice has shown thatNRP-1 is important not only for neuronal development,but also for vascular formation [95]. NRP-1−/− micesuffer from severe defects in the cardiovascular system inaddition to a disorganized neural development, resultingin the death of homozygous embryos by embryonic day14 [96]. Defects in vessel formation include a failureof capillary ingrowth into the brain and the abnormalformation of aortic arches and the yolk-sac vasculature,suggesting the importance of NRP-1 in embryonic vesselformation. In contrast, NRP-2−/− mice show an absenceor severe reduction of small lymphatic vessels and capil-laries during development [97]. Arteries, veins and largercollecting lymphatic vessels develop normally, suggestingthat NRP-2 is selectively required for the formation ofsmall lymphatic vessels and capillaries.
VEGF/VEGFR SYSTEM IN PHYSIOLOGICALAND PATHOLOGICAL CONDITIONS
Physiological angiogenesisThe loss of a single VEGF allele is lethal in the mouse em-bryo between days 11 and 12 [98,99]. VEGF+/− embryosexhibit significant defects in the vasculature of severalorgans and a markedly reduced number of nucleatedred blood cells within the blood islands in the yolk sac.In addition, a 2- to 3-fold overexpression of VEGF-A
from its endogenous locus results in severe abnormalitiesin heart development and lethality at embryonic days12.5 and 14 [100]. These results demonstrate the im-portance of tightly regulating VEGF-A expressionduring embryonic development. Homozygous loss ofthe VEGFR-1 or VEGFR-2 gene results in embryoniclethality between days 8.5 and 9.5, indicating that theseVEGFRs play important roles in vasculogenesis andangiogenesis [101,102]. VEGFR-2−/− mice die due to alack of endothelial cell growth and blood vessel formationas well as extremely poor haematopoiesis. On the otherhand, VEGFR-1−/− mice die due to an overgrowth ofendothelial cells and disorganization of blood vessels.Furthermore, normal vascular development in micelacking the tyrosine kinase domain of VEGFR-1 [103]has indicated that VEGFR-2 is the major positive signaltransducer, whereas VEGFR-1 has a negative regulatoryrole in angiogenesis early in embryogenesis.
Takahashi et al. [76] have shown that Tyr1175 and Tyr1214
are two major VEGF-A-dependent autophosphorylationsites in VEGFR-2. However, only autophosphorylationof Tyr1175 is crucial for VEGF-dependent endothelial cellproliferation via the PLC-γ /PKC/Raf/MEK [MAPK/ERK (extracellular-signal-regulated kinase) kinase]/ERKpathway. An unusual feature of mitogenic signalling fromVEGFR-2 is the requirement for PKC but not Ras [104].Our recent study [105] using knockin mice substitutingTyr1173 (corresponding to Tyr1175 in human VEGFR-2)and Tyr1212 (Tyr1214 in human) of the VEGFR-2 genewith phenylalanine has revealed that the signalling viaTyr1173 of VEGFR-2 is essential for endothelial andhaematopoietic development during embryogenesis. Incontrast, the phosphorylation of Tyr1214 appears to berequired to trigger the sequential activation of Cdc42and p38 MAPK and to drive p38 MAPK-mediated actinremodelling in stress fibres in endothelial cells exposedto VEGF-A [106]. The activation of the PI3K/p70 S6K(S6 kinase) pathway by VEGFR-2 is also involved inVEGF-A-induced endothelial cell proliferation [107](Figure 2). PILSAP (puromycin-intensive leucyl-specificaminopeptidase) plays a crucial role in the activationof this pathway via the binding and modification ofPDK1 (phosphoinositide-dependent kinase 1) [108]. Inaddition, recent studies have revealed various down-stream mediators of VEGF-induced angiogenic signal-ling, such as diacylglycerol kinase α [109], SRF (serumresponse factor) [110], SREBP (sterol-regulatory-ele-ment-binding protein) [111] and IQGAP1 [112].
Studies using DNA microarrays have reported pos-sible endogenous feedback inhibitors for VEGF-inducedangiogenesis. Vasohibin and DSCR1 (Down syndromecritical region protein 1) are significantly induced byVEGF in endothelial cells [113,114]. Up-regulationof DSCR1 in endothelial cells inhibits the nuclearlocalization of NFAT (nuclear factor of activated T-cells),proliferation and tube formation [115].
C© 2005 The Biochemical Society

234 H. Takahashi and M. Shibuya
Vascular permeabilityVEGF-A is known to increase the vascular permeabilityof microvessels to circulating macromolecules [14]. In-creased vascular permeability is often observed in areasof pathological angiogenesis in solid tumours, woundsand chronic inflammation. VEGF-A significantly accu-mulates in malignant ascites [116] and pleural effusion[117], suggesting that it plays a fundamental role in theaccumulation of malignant fluid through the enhance-ment of vascular permeability. Consistent with a role inthe regulation of vascular permeability, VEGF-A inducesendothelial fenestration in some vascular beds and incultured adrenal endothelial cells, the extravasation offerritin by way of the VVO (vesiculo-vacuolar organelle)[14], and disorganization of endothelial junctional pro-teins such as VE-cadherin and occludin [118]. VEGF-Aincreases vascular permeability in mesenteric microvess-els by activation of VEGFR-2 on endothelial cells andsubsequent activation of PLC. This causes increasedproduction of diacylglycerol that results in influx ofcalcium [14]. Other studies have also demonstrated thecrucial role of VEGFR-2 signalling in the enhancementof vascular permeability; however, our recent study [61]using Tf svVEGF has shown that the enhancement ofvascular permeability is intensified by the activationof VEGFR-1 more than the proliferation of endothelialcells under some active signalling from VEGFR-2. Thisfinding indicates the importance of VEGFR-1 signallingin vascular permeability.
An analysis of mice deficient in specific Src familykinases has demonstrated no decrease in VEGF-depen-dent neovascularization, but a complete ablation ofvascular permeability in Src−/− or Yes−/− mice, whereasFyn−/− mice show no such defect [119]. In addition,blockade of Src prevents the disassociation of a complexcomprising VEGFR-2, VE-cadherin and β-catenin withthe same kinetics with which it prevents VEGF-mediatedvascular permeability and oedema [120]. These findingsindicate that the activity of specific Src family kinases isessential for the VEGF-induced enhancement of vascularpermeability through the disruption of the VEGFR-2/cadherin/catenin complex.
VEGF-A can induce production of NO and endo-genous NO can increase vascular permeability [121].Among the three isoforms of NOS (NO synthase),eNOS (endothelial NOS) plays a predominant role inVEGF-induced angiogenesis and vascular permeability[122]. Furthermore, the activation of eNOS is regulatedby the PI3K/Akt pathway [123,124]. The small GTP-binding protein Rac, which is also activated by PI3K, hasbeen implicated in the regulation of vascular permeability[125]. A recent study [126] has shown that inhibition ofp38 MAPK activity abrogated VEGF-induced vascularpermeability in vivo and in vitro, suggesting the in-volvement of p38 MAPK in the control of vascularpermeability (Figure 2).
Solid tumoursNumerous studies have established VEGF-A as a keyangiogenic player in cancer. VEGF-A is expressed inmost tumours and its expression correlates with tumourprogression. In addition to tumour cells, tumour-asso-ciated stroma is also an important source of VEGF-A[127]. In the absence of access to an adequate vas-culature, tumour cells become necrotic and apoptotic,restraining the increase in tumour volume that shouldresult from continuous cell proliferation [128]. Theexpression of VEGF-A mRNA is highest in hypoxictumour cells adjacent to necrotic areas [16], indicatingthat the induction of VEGF-A by hypoxia in growingtumours can change the balance of inhibitors and activat-ors of angiogenesis, leading to the growth of new bloodvessels into tumour. Consistent with this hypothesis,capturing of VEGF or blocking of its signalling recep-tor VEGFR-2 by a VEGFR tyrosine kinase inhibitor,antisense oligonucleotides, vaccination or neutralizingantibodies reduced tumour angiogenesis and growth inpreclinical studies [129]. Unlike in physiological angio-genesis, VEGFR-1 signalling plays an important rolein angiogenesis under pathological conditions [43,130].Autiero et al. [131] have proposed that PlGF regulatesinter- and intra-molecular cross-talk between VEGFR-1and VEGFR-2, amplifying VEGF-driven angiogenesisthrough VEGFR-2.
Several studies also describe the role of VEGF incarcinogenesis [132]. Rip1–Tag2 (T-antigen 2) mice de-velop islet tumours of the pancreas by 12–14 weeks ofage as a result of expression of the SV40 Tag oncogenein insulin-producing β-cells. In this mouse, angiogenicactivity first appears in a subset of hyperplastic isletsbefore the onset of tumour formation. VEGF-A andVEGFRs are constitutively expressed in the islet vascu-lature before and after the initiation of angiogenesis(angiogenic switch) [133]; however, when VEGF-A isabsent from islet β-cells of Rip1–Tag2 mice, both angio-genic switching and carcinogenesis as well as tumourgrowth are severely disrupted [134], indicating thatVEGF-A plays a critical role in angiogenic switchingand carcinogenesis. Bergers et al. [135] have revealed thatMMP (matrix metalloproteinase)-9 is also a component ofthe angiogenic switch, as this proteinase makes VEGF-Aavailable for the interaction with its receptors by releasingsequestered VEGF-A.
VEGF-A impairs the endothelial barrier by disruptinga VE-cadherin/β-catenin complex via the activation ofSrc and facilitates tumour cell extravasation and meta-stasis [136]. VEGF-A also induces the disruption ofhepatocellular tight junctions, which may promotetumour invasion [137]. Pharmacological blockade ofVEGFR-2 stabilizes the endothelial barrier functionand suppresses tumour cell extravasation in vivo [136],suggesting the importance of VEGFR-2 signalling in thiskind of tumour invasion and metastasis. Hiratsuka et al.
C© 2005 The Biochemical Society

VEGF/VEGFR system in health and disease 235
[138] have shown that VEGFR-1 signalling is alsoinvolved in tumour metastasis, being linked to the induc-tion of MMP-9 in lung endothelial cells and to thefacilitation of lung-specific metastasis.
Recently, Hurwitz et al. [139] have shown that the ad-dition of bevacizumab (a humanized anti-VEGF mono-clonal antibody) to fluorouracil-based combinationchemotherapy results in statistically significant and clini-cally meaningful improvement in survival among patientswith metastatic colorectal cancer. Based on this result,bevacizumab (Avastin) was approved by the FDA(Food and Drug Administration) in February 2004 as afirst-line treatment for metastatic colorectal carcinoma.Besides bevacizumab, many other VEGF inhibitorsare being pursued clinically. These inhibitors includesmall-molecule RTK inhibitors such as PTK787, solublereceptors such as VEGF-Trap and anti-VEGFR-2 mAbs(monoclonal antibodies) [129].
Inflammatory diseasesVEGF acts as a pro-inflammatory cytokine by increasingthe permeability of endothelial cells, inducing the ex-pression of endothelial adhesion molecules and via itsability to act as a monocyte chemoattractant [140–142].VEGF is strongly expressed by epidermal keratinocytesin wound healing and psoriasis, conditions that arecharacterized by increased microvascular permeabilityand angiogenesis [16]. Transgenic mice that overexpressVEGF-A specifically in the epidermis exhibit an increaseddensity of tortuous cutaneous blood capillaries as wellas highly increased leucocyte rolling and adhesion inpostcapillary skin venules, suggesting that enhanced ex-pression of VEGF-A in epidermal keratinocytes is suffi-cient to develop psoriasis-like inflammatory skin lesions[143]. Moreover, heterozygous VEGF-A transgenic mice,which do not spontaneously develop inflammatoryskin lesions, are unable to down-regulate experimentallyinduced inflammation and exhibit a psoriasis-like pheno-type characterized by epidermal hyperplasia, the accumu-lation of lymphocytes, and lymphatic vessel proliferationand enlargement [144]. Transgenic overexpression ofPlGF-2 in epidermal keratinocytes also results in asignificantly increased inflammatory response, whereasa deficiency of PlGF results in a diminished and abbre-viated inflammatory response [145], suggesting the im-portance of VEGFR-1 signalling in chronic skin inflam-mation.
Local production of VEGF-A in arthritic synovialtissue has been documented [16] and appears to cor-relate with disease activity in humans. Subsequently,VEGF-A has been shown to be important in the patho-genesis of RA (rheumatoid arthritis) in animal models[146–148]. Treatment with anti-VEGFR-1 mAbs, butnot anti-VEGFR-2 mAbs, significantly reduces thearthritic destruction of joints by suppressing synovialinflammation and neovascularization, emphasizing the
importance of VEGFR-1 signalling in the destruction.The anti-inflammatory effects of anti-VEGFR-1 areattributable to a reduced mobilization of bone-marrow-derived myeloid progenitors into peripheral blood [147].The reduction of synovial inflammation in VEGF-B−/−
mice [47] also implies a critical role for VEGFR-1signalling in RA.
Exaggerated levels of VEGF-A have been detectedin tissues and biological samples from people withasthma, where these levels correlate directly with disease[149] and inversely with airway function [150]. VEGFhas been postulated to contribute to asthmatic tissueoedema through its effect on vascular permeability. Arecent study using lung-targeted VEGF165 transgenicmice has revealed a novel function of VEGF-A in allergicresponses. In these mice, VEGF-A induces asthma-like inflammation, airway and vascular remodelling, andairway hyper-responsiveness. VEGF-A also enhancesrespiratory sensitization to antigen as well as TH2(T-helper type 2) cell-mediated inflammation and in-creases the number of activated dendritic cells [151].Thus VEGF-A has a critical role in pulmonary TH2inflammation. Other studies have provided evidence fora role for VEGF-A as a pro-inflammatory mediator inallograft rejection [152] and neointimal formation [153].
Other pathological conditionsVEGF-A mRNA expression, not normally found in theadult mouse brain, is up-regulated after cerebral isch-aemia, and elevated VEGF-A levels can be detected asearly as 3 h after stroke with a peak between 12 and48 h [154]. Previous studies have demonstrated that theantagonism of VEGF-A results in reduced oedema andtissue damage after ischaemia implicating VEGF-A inthe pathophysiology of stroke [155]. Paul et al. [156]have reported that Src−/− mice are resistant to VEGF-A-induced vascular permeability and show decreasedinfarct volumes after stroke. Systemic application of aSrc inhibitor suppresses vascular permeability, protectingwild-type mice from ischaemia-induced brain damagewithout influencing VEGF-A expression. However, Sunet al. [157] have reported that intracerebroventricularadministration of VEGF-A reduces infarct size, improvesneurological performance and enhances the delayedsurvival of newborn neurons. These conflicting resultsappear to reflect dual roles of VEGF-A in stroke: neuro-protective and pro-inflammatory effects. In this context,when infused through the internal carotid artery, lowand intermediate doses of VEGF-A significantly promoteneuroprotection of the ischaemic brain, whereas a highdose of VEGF-A offers no neuroprotection to theischaemic brain or the damaged neurons of normal brain[158]. Further studies are required for the therapeuticapplication of VEGF-A against stroke.
Extensive evidence has suggested a causal role ofVEGF in several diseases of the human eye in which
C© 2005 The Biochemical Society

236 H. Takahashi and M. Shibuya
neovascularization and increased vascular permeabilityoccur. VEGF levels are increased in the vitreous andretina of patients and laboratory animals with active neo-vascularization from ischaemic retinopathies such asproliferative diabetic retinopathy, central retinal veinocclusion and retinopathy of prematurity. Subsequentstudies using various VEGF inhibitors have confirmedthat VEGF plays a central role in ischaemia-inducedintraocular neovascularization [159]. An anti-VEGFaptamer, pegaptanib (Macugen), has produced a stat-istically significant and clinically meaningful benefit inthe treatment of neovascular AMD (age-related maculardegeneration) [160], which is the leading cause of irre-versible severe loss of vision in people 50 years of age andolder in the developed world, and was approved by theFDA in December 2004.
Oosthuyse et al. [161] have reported that deletion ofthe HRE in the VEGF promoter reduces hypoxic VEGFexpression in the spinal cord and causes adult-onsetprogressive motor neuron degeneration, reminiscent ofALS (amyotrophic lateral sclerosis). VEGF165 promotessurvival of motor neurons during hypoxia throughbinding VEGFR-2 and NRP-1 [161]. A subsequent studyhas revealed that VEGF-A is a modifier associated withmotor neuron degeneration in human ALS and in a mousemodel of ALS [162]. VEGF-A treatment increases the lifeexpectancy of ALS mice without causing toxic side effects[163,164], indicating that VEGF-A has neuroprotectiveeffects on motor neurons, and treatment with VEGF-Acould be one of the most effective therapies for ALSreported so far.
LeCouter et al. [165] recently provided evidence for anovel function of VEGFR-1 in LSECs (liver sinusoidalendothelial cells). The activation of VEGFR-1 results inthe paracrine release of HGF (hepatocyte growth factor),IL-6 (interleukin-6) and other hepatotrophic moleculesby LSECs to the extent that hepatocytes are stimulated toproliferate when co-cultured with LSECs. VEGF-A hasno direct mitogenic effect on hepatocytes. A VEGFR-1agonist protected the liver from CCl4-induced damage, inspite of its inability to induce the proliferation of LSECs.
CONCLUSIONVEGF was originally described as a specific angiogenicand permeability-inducing factor and its function wasconsidered to be specific for endothelial cells. However,emerging evidence has revealed that the role of theVEGF/VEGFR system extends far beyond previousexpectations. First, a wide variety of VEGF family pro-teins and numerous splicing variants have been identifiedand found to play distinct but critical roles in variousconditions, including lymphangiogenesis. VEGF familyproteins have been utilized even in snake venoms andsome viruses. Secondly, several different VEGFRs havebeen shown to be essential, but the interaction between
these receptors has appeared to be complicated.VEGFR-1 has a negative regulatory role in embryonicangiogenesis, but functions as a positive signal transducerin some cases individually and sometimes synergisticallywith VEGFR-2 via the intra- and inter-molecular cross-talk between these two receptors. An association betweenVEGFR-2 and VEGFR-3 has also been reported [166].Thirdly, it has been shown that the VEGF/VEGFRsystem has multiple functions, such as the inductionof tumour metastasis, inflammation, neuroprotection,protection of liver and mobilization of marrow-derivedstem cells, as well as lymphangiogenesis. VEGF is alsoimportant for memory and learning [167]. Fourthly,numerous other molecules have been found to associatewith the VEGF/VEGFR system. Further studies arerequired to achieve a comprehensive understanding of theVEGF/VEGFR system; however, the recent progress inthe molecular and biological study of this system providesus with novel and promising therapeutic strategies forovercoming a variety of diseases.
ACKNOWLEDGMENTS
The authors’ work was supported by Grant-in-AidSpecial Project Research on Cancer-Bioscience 12215024from the Ministry of Education, Culture, Sports, Scienceand Technology of Japan, and grants for the program‘Research for the Future’ from the Japan Society forPromotion of Science, and for the program ‘Promotionof Fundamental Research in Health Sciences’ from theOrganization for Pharmaceutical Safety and Research.
REFERENCES
1 Leung, D. W., Cachianes, G., Kuang, W. J., Goeddel, D. V.and Ferrara, N. (1989) Vascular endothelial growth factoris a secreted angiogenic mitogen. Science 246, 1306–1309
2 Keck, P. J., Hauser, S. D., Krivi, G. et al. (1989) Vascularpermeability factor, an endothelial cell mitogen related toPDGF. Science 246, 1309–1312
3 Shibuya, M., Yamaguchi, S., Yamane, A. et al. (1990)Nucleotide sequence and expression of a novel humanreceptor-type tyrosine kinase gene (flt) closely related tothe fms family. Oncogene 5, 519–524
4 Matthews, W., Jordan, C. T., Gavin, M., Jenkins, N. A.,Copeland, N. G. and Lemischka, I. R. (1991) A receptortyrosine kinase cDNA isolated from a population ofenriched primitive hematopoietic cells and exhibitingclose genetic linkage to c-kit. Proc. Natl. Acad. Sci.U.S.A. 88, 9026–9030
5 Terman, B. I., Carrion, M. E., Kovacs, E., Rasmussen,B. A., Eddy, R. L. and Shows, T. B. (1991) Identificationof a new endothelial cell growth factor receptor tyrosinekinase. Oncogene 6, 1677–1683
6 Connolly, D. T., Heuvelman, D. M., Nelson, R. et al.(1989) Tumor vascular permeability factor stimulatesendothelial cell growth and angiogenesis. J. Clin. Invest.84, 1470–1478
7 Ferrara, N. and Henzel, W. J. (1989) Pituitary follicularcells secrete a novel heparin-binding growth factorspecific for vascular endothelial cells. Biochem. Biophys.Res. Commun. 161, 851–858
C© 2005 The Biochemical Society

VEGF/VEGFR system in health and disease 237
8 Senger, D. R., Galli, S. J., Dvorak, A. M., Perruzzi, C. A.,Harvey, V. S. and Dvorak, H. F. (1983) Tumor cells secretea vascular permeability factor that promotes accumulationof ascites fluid. Science 219, 983–985
9 Muller, Y. A., Li, B., Christinger, H. W., Wells, J. A.,Cunningham, B. C. and de Vos, A. M. (1997) Vascularendothelial growth factor: crystal structure and functionalmapping of the kinase domain receptor binding site.Proc. Natl. Acad. Sci. U.S.A. 94, 7192–7197
10 Houck, K. A., Ferrara, N., Winer, J., Cachianes, G., Li, B.and Leung, D. W. (1991) The vascular endothelial growthfactor family: identification of a fourth molecular speciesand characterization of alternative splicing of RNA.Mol. Endocrinol. 5, 1806–1814
11 Tischer, E., Mitchell, R., Hartman, T. et al. (1991) Thehuman gene for vascular endothelial growth factor.Multiple protein forms are encoded through alternativeexon splicing. J. Biol. Chem. 266, 11947–11954
12 Vincenti, V., Cassano, C., Rocchi, M. and Persico, G.(1996) Assignment of the vascular endothelial growthfactor gene to human chromosome 6p21.3. Circulation93, 1493–1495
13 Lange, T., Guttmann-Raviv, N., Baruch, L., Machluf, M.and Neufeld, G. (2003) VEGF162, a new heparin-bindingvascular endothelial growth factor splice form that isexpressed in transformed human cells. J. Biol. Chem. 278,17164–17169
14 Bates, D. O. and Harper, S. J. (2002) Regulation ofvascular permeability by vascular endothelial growthfactors. Vascul. Pharmacol. 39, 225–237
15 Woolard, J., Wang, W. Y., Bevan, H. S. et al. (2004)VEGF165b, an inhibitory vascular endothelial growthfactor splice variant: mechanism of action, in vivo effecton angiogenesis and endogenous protein expression.Cancer Res. 64, 7822–7835
16 Ferrara, N. and Davis-Smyth, T. (1997) The biology ofvascular endothelial growth factor. Endocr. Rev. 18, 4–25
17 Melter, M., Reinders, M. E., Sho, M. et al. (2000) Ligationof CD40 induces the expression of vascular endothelialgrowth factor by endothelial cells and monocytes andpromotes angiogenesis in vivo. Blood 96, 3801–3808
18 Freeman, M. R., Schneck, F. X., Gagnon, M. L. et al.(1995) Peripheral blood T lymphocytes and lymphocytesinfiltrating human cancers express vascular endothelialgrowth factor: a potential role for T cells in angiogenesis.Cancer Res. 55, 4140–4145
19 Plouet, J., Moro, F., Bertagnolli, S. et al. (1997)Extracellular cleavage of the vascular endothelial growthfactor 189-amino acid form by urokinase is required forits mitogenic effect. J. Biol. Chem. 272, 13390–13396
20 Soker, S., Takashima, S., Miao, H. Q., Neufeld, G. andKlagsbrun, M. (1998) Neuropilin-1 is expressed byendothelial and tumor cells as an isoform-specific receptorfor vascular endothelial growth factor. Cell 92, 735–745
21 Gluzman-Poltorak, Z., Cohen, T., Herzog, Y. andNeufeld, G. (2000) Neuropilin-2 and neuropilin-1 arereceptors for the 165-amino acid form of vascularendothelial growth factor (VEGF) and of placenta growthfactor-2, but only neuropilin-2 functions as a receptor forthe 145-amino acid form of VEGF. J. Biol. Chem. 275,18040–18045
22 Carmeliet, P., Ng, Y. S., Nuyens, D. et al. (1999) Impairedmyocardial angiogenesis and ischemic cardiomyopathy inmice lacking the vascular endothelial growth factorisoforms VEGF164 and VEGF188. Nat. Med. 5, 495–502
23 Ruhrberg, C., Gerhardt, H., Golding, M. et al. (2002)Spatially restricted patterning cues provided byheparin-binding VEGF-A control blood vessel branchingmorphogenesis. Genes Dev. 16, 2684–2698
24 Stalmans, I., Ng, Y. S., Rohan, R. et al. (2002) Arteriolarand venular patterning in retinas of mice selectivelyexpressing VEGF isoforms. J. Clin. Invest. 109, 327–336
25 Maes, C., Stockmans, I., Moermans, K. et al. (2004)Soluble VEGF isoforms are essential for establishingepiphyseal vascularization and regulating chondrocytedevelopment and survival. J. Clin. Invest. 113, 188–199
26 Ivan, M., Kondo, K., Yang, H. et al. (2001) HIFα targetedfor VHL-mediated destruction by proline hydroxylation:implications for O2 sensing. Science 292, 464–468
27 Jaakkola, P., Mole, D. R., Tian, Y. M. et al. (2001)Targeting of HIF-α to the von Hippel–Lindauubiquitylation complex by O2-regulated prolylhydroxylation. Science 292, 468–472
28 Lando, D., Peet, D. J., Whelan, D. A., Gorman, J. J. andWhitelaw, M. L. (2002) Asparagine hydroxylation of theHIF transactivation domain a hypoxic switch. Science295, 858–861
29 Gerald, D., Berra, E., Frapart, Y. M. et al. (2004) JunDreduces tumor angiogenesis by protecting cells fromoxidative stress. Cell 118, 781–794
30 Levy, N. S., Chung, S., Furneaux, H. and Levy, A. P.(1998) Hypoxic stabilization of vascular endothelialgrowth factor mRNA by the RNA-binding protein HuR.J. Biol. Chem. 273, 6417–6423
31 Onesto, C., Berra, E., Grepin, R. and Pages, G. (2004)Poly(A)-binding protein-interacting protein 2, a strongregulator of vascular endothelial growth factor mRNA.J. Biol. Chem. 279, 34217–34226
32 Xie, K., Wei, D., Shi, Q. and Huang, S. (2004)Constitutive and inducible expression and regulation ofvascular endothelial growth factor. Cytokine GrowthFactor Rev. 15, 297–324
33 Maglione, D., Guerriero, V., Viglietto, G., Delli-Bovi, P.and Persico, M. G. (1991) Isolation of a human placentacDNA coding for a protein related to the vascularpermeability factor. Proc. Natl. Acad. Sci. U.S.A. 88,9267–9271
34 Persico, M. G., Vincenti, V. and DiPalma, T. (1999)Structure, expression and receptor-binding properties ofplacental growth factor (PlGF). Curr. Top.Microbiol. Immunol. 237, 31–40
35 Park, J. E., Chen, H. H., Winer, J., Houck, K. A. andFerrara, N. (1994) Placenta growth factor. Potentiation ofvascular endothelial growth factor bioactivity, in vitro andin vivo, and high affinity binding to Flt-1 but not toFlk-1/KDR. J. Biol. Chem. 269, 25646–25654
36 Sawano, A., Takahashi, T., Yamaguchi, S., Aonuma, M.and Shibuya, M. (1996) Flt-1 but not KDR/Flk-1 tyrosinekinase is a receptor for placenta growth factor, which isrelated to vascular endothelial growth factor.Cell Growth Differ. 7, 213–221
37 Maglione, D., Guerriero, V., Viglietto, G. et al. (1993) Twoalternative mRNAs coding for the angiogenic factor,placenta growth factor (PlGF), are transcribed from asingle gene of chromosome 14. Oncogene 8, 925–931
38 Cao, Y., Ji, W. R., Qi, P. and Rosin, A. (1997) Placentagrowth factor: identification and characterization of anovel isoform generated by RNA alternative splicing.Biochem. Biophys. Res. Commun. 235, 493–498
39 Yang, W., Ahn, H., Hinrichs, M., Torry, R. J. and Torry,D. S. (2003) Evidence of a novel isoform of placentagrowth factor (PlGF-4) expressed in human trophoblastand endothelial cells. J. Reprod. Immunol. 60, 53–60
40 Iyer, S., Leonidas, D. D., Swaminathan, G. J. et al. (2001)The crystal structure of human placenta growth factor-1(PlGF-1), an angiogenic protein, at 2.0 A resolution.J. Biol. Chem. 276, 12153–12161
41 Christinger, H. W., Fuh, G., de Vos, A. M. and Wiesmann,C. (2004) The crystal structure of placental growth factorin complex with domain 2 of vascular endothelial growthfactor receptor-1. J. Biol. Chem. 279, 10382–10388
42 Errico, M., Riccioni, T., Iyer, S. et al. (2004) Identificationof placenta growth factor determinants for binding andactivation of Flt-1 receptor. J. Biol. Chem. 279,43929–43939
43 Carmeliet, P., Moons, L., Luttun, A. et al. (2001)Synergism between vascular endothelial growth factorand placental growth factor contributes to angiogenesisand plasma extravasation in pathological conditions.Nat. Med. 7, 575–583
44 Olofsson, B., Jeltsch, M., Eriksson, U. and Alitalo, K.(1999) Current biology of VEGF-B and VEGF-C. Curr.Opin. Biotechnol. 10, 528–535
45 Bellomo, D., Headrick, J. P., Silins, G. U. et al. (2000)Mice lacking the vascular endothelial growth factor-Bgene (Vegfb) have smaller hearts, dysfunctional coronaryvasculature, and impaired recovery from cardiac ischemia.Circ. Res. 86, E29–E35
C© 2005 The Biochemical Society

238 H. Takahashi and M. Shibuya
46 Aase, K., von Euler, G., Li, X. et al. (2001) Vascularendothelial growth factor-B-deficient mice display anatrial conduction defect. Circulation 104, 358–364
47 Mould, A. W., Tonks, I. D., Cahill, M. M. et al. (2003)Vegfb gene knockout mice display reduced pathology andsynovial angiogenesis in both antigen-induced andcollagen-induced models of arthritis. Arthritis Rheum. 48,2660–2669
48 Sun, Y., Jin, K., Childs, J. T., Xie, L., Mao, X. O. andGreenberg, D. A. (2004) Increased severity of cerebralischemic injury in vascular endothelial growthfactor-B-deficient mice. J. Cereb. Blood Flow Metab. 24,1146–1152
49 Lohela, M., Saaristo, A., Veikkola, T. and Alitalo, K.(2003) Lymphangiogenic growth factors, receptors andtherapies. Thromb. Haemostasis 90, 167–184
50 McColl, B. K., Baldwin, M. E., Roufail, S. et al. (2003)Plasmin activates the lymphangiogenic growth factorsVEGF-C and VEGF-D. J. Exp. Med. 198, 863–868
51 Jeltsch, M., Kaipainen, A., Joukov, V. et al. (1997)Hyperplasia of lymphatic vessels in VEGF-C transgenicmice. Science 276, 1423–1425
52 Karkkainen, M. J., Haiko, P., Sainio, K. et al. (2004)Vascular endothelial growth factor C is required forsprouting of the first lymphatic vessels from embryonicveins. Nat. Immunol. 5, 74–80
53 Stacker, S. A., Caesar, C., Baldwin, M. E. et al. (2001)VEGF-D promotes the metastatic spread of tumor cellsvia the lymphatics. Nat. Med. 7, 186–191
54 Lyttle, D. J., Fraser, K. M., Fleming, S. B., Mercer, A. A.and Robinson, A. J. (1994) Homologs of vascularendothelial growth factor are encoded by the poxvirus orfvirus. J. Virol. 68, 84–92
55 Wise, L. M., Veikkola, T., Mercer, A. A. et al. (1999)Vascular endothelial growth factor (VEGF)-like proteinfrom orf virus NZ2 binds to VEGFR2 and neuropilin-1.Proc. Natl. Acad. Sci. U.S.A. 96, 3071–3076
56 Ogawa, S., Oku, A., Sawano, A., Yamaguchi, S.,Yazaki, Y. and Shibuya, M. (1998) A novel type ofvascular endothelial growth factor, VEGF-E (NZ-7VEGF), preferentially utilizes KDR/Flk-1 receptor andcarries a potent mitotic activity without heparin-bindingdomain. J. Biol. Chem. 273, 31273–31282
57 Wise, L. M., Ueda, N., Dryden, N. H. et al. (2003) Viralvascular endothelial growth factors vary extensively inamino acid sequence, receptor-binding specificities, andthe ability to induce vascular permeability yet areuniformly active mitogens. J. Biol. Chem. 278,38004–38014
58 Meyer, M., Clauss, M., Lepple-Wienhues, A. et al. (1999)A novel vascular endothelial growth factor encoded byOrf virus, VEGF-E, mediates angiogenesis via signallingthrough VEGFR-2 (KDR) but not VEGFR-1 (Flt-1)receptor tyrosine kinases. EMBO J. 18, 363–374
59 Kiba, A., Sagara, H., Hara, T. and Shibuya, M. (2003)VEGFR-2-specific ligand VEGF-E inducesnon-edematous hyper-vascularization in mice.Biochem. Biophys. Res. Commun. 301, 371–377
60 Junqueira de Azevedo, I. L., Farsky, S. H., Oliveira, M. L.and Ho, P. L. (2001) Molecular cloning and expression ofa functional snake venom vascular endothelium growthfactor (VEGF) from the Bothrops insularis pit viper. Anew member of the VEGF family of proteins.J. Biol. Chem. 276, 39836–39842
61 Takahashi, H., Hattori, S., Iwamatsu, A., Takizawa, H.and Shibuya, M. (2004) A novel snake venom vascularendothelial growth factor (VEGF) predominantly inducesvascular permeability through preferential signaling viaVEGF receptor-1. J. Biol. Chem. 279, 46304–46314
62 Komori, Y., Nikai, T., Taniguchi, K., Masuda, K. andSugihara, H. (1999) Vascular endothelial growth factorVEGF-like heparin-binding protein from the venom ofVipera aspis aspis (Aspic viper). Biochemistry 38,11796–11803
63 Gasmi, A., Bourcier, C., Aloui, Z. et al. (2002) Completestructure of an increasing capillary permeability protein(ICPP) purified from Vipera lebetina venom. ICPP isangiogenic via vascular endothelial growth factor receptorsignalling. J. Biol. Chem. 277, 29992–29998
64 Yamazaki, Y., Takani, K., Atoda, H. and Morita, T. (2003)Snake venom vascular endothelial growth factors(VEGFs) exhibit potent activity through their specificrecognition of KDR (VEGF receptor 2). J. Biol. Chem.278, 51985–51988
65 Sawano, A., Iwai, S., Sakurai, Y. et al. (2001) Flt-1,vascular endothelial growth factor receptor 1, is a novelcell surface marker for the lineage ofmonocyte-macrophages in humans. Blood 97, 785–791
66 Hattori, K., Heissig, B., Wu, Y. et al. (2002) Placentalgrowth factor reconstitutes hematopoiesis by recruitingVEGFR1+ stem cells from bone-marrowmicroenvironment. Nat. Med. 8, 841–849
67 Wiesmann, C., Fuh, G., Christinger, H. W., Eigenbrot, C.,Wells, J. A. and de Vos, A. M. (1997) Crystal structure at1.7 A resolution of VEGF in complex with domain 2 ofthe Flt-1 receptor. Cell 91, 695–704
68 Kanno, S., Oda, N., Abe, M. et al. (2000) Roles of twoVEGF receptors, Flt-1 and KDR, in the signaltransduction of VEGF effects in human vascularendothelial cells. Oncogene 19, 2138–2146
69 Kendall, R. L., Wang, G. and Thomas, K. A. (1996)Identification of a natural soluble form of the vascularendothelial growth factor receptor, FLT-1, and itsheterodimerization with KDR. Biochem. Biophys.Res. Commun. 226, 324–328
70 Toi, M., Bando, H., Ogawa, T., Muta, M., Hornig, C. andWeich, H. A. (2002) Significance of vascular endothelialgrowth factor (VEGF)/soluble VEGF receptor-1relationship in breast cancer. Int. J. Cancer 98, 14–18
71 Scheufler, K. M., Drevs, J., van Velthoven, V. et al. (2003)Implications of vascular endothelial growth factor, sFlt-1,and sTie-2 in plasma, serum and cerebrospinal fluidduring cerebral ischemia in man. J. Cereb.Blood Flow Metab. 23, 99–110
72 Levine, R. J., Maynard, S. E., Qian, C. et al. (2004)Circulating angiogenic factors and the risk ofpreeclampsia. New Engl. J. Med. 350, 672–683
73 Maynard, S. E., Min, J. Y., Merchan, J. et al. (2003) Excessplacental soluble fms-like tyrosine kinase 1 (sFlt1) maycontribute to endothelial dysfunction, hypertension, andproteinuria in preeclampsia. J. Clin. Invest. 111, 649–658
74 Fuh, G., Li, B., Crowley, C., Cunningham, B. and Wells,J. A. (1998) Requirements for binding and signaling of thekinase domain receptor for vascular endothelial growthfactor. J. Biol. Chem. 273, 11197–11204
75 Katoh, O., Tauchi, H., Kawaishi, K., Kimura, A. andSatow, Y. (1995) Expression of the vascular endothelialgrowth factor (VEGF) receptor gene, KDR, inhematopoietic cells and inhibitory effect of VEGF onapoptotic cell death caused by ionizing radiation.Cancer Res. 55, 5687–5692
76 Takahashi, T., Yamaguchi, S., Chida, K. and Shibuya, M.(2001) A single autophosphorylation site on KDR/Flk-1is essential for VEGF-A-dependent activation of PLC-γand DNA synthesis in vascular endothelial cells.EMBO J. 20, 2768–2778
77 Wu, L. W., Mayo, L. D., Dunbar, J. D. et al. (2000) VRAPis an adaptor protein that binds KDR, a receptor forvascular endothelial cell growth factor. J. Biol. Chem. 275,6059–6062
78 Warner, A. J., Lopez-Dee, J., Knight, E. L., Feramisco,J. R. and Prigent, S. A. (2000) The Shc-related adaptorprotein, Sck, forms a complex with thevascular-endothelial-growth-factor receptor KDR intransfected cells. Biochem. J. 347, 501–509
79 Holmqvist, K., Cross, M. J., Rolny, C. et al. (2004) Theadaptor protein shb binds to tyrosine 1175 in vascularendothelial growth factor (VEGF) receptor-2 andregulates VEGF-dependent cellular migration.J. Biol. Chem. 279, 22267–22275
80 Nagy, J. A., Vasile, E., Feng, D. et al. (2002) Vascularpermeability factor/vascular endothelial growth factorinduces lymphangiogenesis as well as angiogenesis.J. Exp. Med. 196, 1497–1506
81 Hong, Y. K., Lange-Asschenfeldt, B., Velasco, P. et al.(2004) VEGF-A promotes tissue repair-associatedlymphatic vessel formation via VEGFR-2 and the α1β1and α2β1 integrins. FASEB J. 18, 1111–1113
C© 2005 The Biochemical Society

VEGF/VEGFR system in health and disease 239
82 Gerber, H. P., McMurtrey, A., Kowalski, J. et al. (1998)Vascular endothelial growth factor regulates endothelialcell survival through the phosphatidylinositol 3′-kinase/Akt signal transduction pathway. Requirement forFlk-1/KDR activation. J. Biol. Chem. 273, 30336–30343
83 Fujio, Y. and Walsh, K. (1999) Akt mediatescytoprotection of endothelial cells by vascular endothelialgrowth factor in an anchorage-dependent manner.J. Biol. Chem. 274, 16349–16354
84 Byzova, T. V., Goldman, C. K., Pampori, N. et al. (2000)A mechanism for modulation of cellular responses toVEGF: activation of the integrins. Mol. Cell 6, 851–860
85 Ebos, J. M., Bocci, G., Man, S. et al. (2004) A naturallyoccurring soluble form of vascular endothelial growthfactor receptor 2 detected in mouse and human plasma.Mol. Cancer Res. 2, 315–326
86 Makinen, T., Jussila, L., Veikkola, T. et al. (2001)Inhibition of lymphangiogenesis with resultinglymphedema in transgenic mice expressing soluble VEGFreceptor-3. Nat. Med. 7, 199–205
87 Veikkola, T., Jussila, L., Makinen, T. et al. (2001)Signalling via vascular endothelial growth factorreceptor-3 is sufficient for lymphangiogenesis intransgenic mice. EMBO J. 20, 1223–1231
88 Makinen, T., Veikkola, T., Mustjoki, S. et al. (2001)Isolated lymphatic endothelial cells transduce growth,survival and migratory signals via the VEGF-C/Dreceptor VEGFR-3. EMBO J. 20, 4762–4773
89 Chen, L., Hamrah, P., Cursiefen, C. et al. (2004) Vascularendothelial growth factor receptor-3 mediates inductionof corneal alloimmunity. Nat. Med. 10, 813–815
90 Neufeld, G., Cohen, T., Shraga, N., Lange, T., Kessler, O.and Herzog, Y. (2002) The neuropilins: multifunctionalsemaphorin and VEGF receptors that modulate axonguidance and angiogenesis. Trends Cardiovasc. Med. 12,13–19
91 Takahashi, T., Fournier, A., Nakamura, F. et al. (1999)Plexin-neuropilin-1 complexes form functionalsemaphorin-3A receptors. Cell 99, 59–69
92 Takahashi, T. and Strittmatter, S. M. (2001) Plexina1autoinhibition by the plexin sema domain. Neuron 29,429–439
93 Whitaker, G. B., Limberg, B. J. and Rosenbaum, J. S.(2001) Vascular endothelial growth factor receptor-2 andneuropilin-1 form a receptor complex that is responsiblefor the differential signaling potency of VEGF165 andVEGF121. J. Biol. Chem. 276, 25520–25531
94 Soker, S., Miao, H. Q., Nomi, M., Takashima, S. andKlagsbrun, M. (2002) VEGF165 mediates formation ofcomplexes containing VEGFR-2 and neuropilin-1 thatenhance VEGF165-receptor binding. J. Cell. Biochem. 85,357–368
95 Kitsukawa, T., Shimono, A., Kawakami, A., Kondoh, H.and Fujisawa, H. (1995) Overexpression of a membraneprotein, neuropilin, in chimeric mice causes anomalies inthe cardiovascular system, nervous system and limbs.Development 121, 4309–4318
96 Kawasaki, T., Kitsukawa, T., Bekku, Y. et al. (1999) Arequirement for neuropilin-1 in embryonic vesselformation. Development 126, 4895–4902
97 Yuan, L., Moyon, D., Pardanaud, L. et al. (2002)Abnormal lymphatic vessel development in neuropilin 2mutant mice. Development 129, 4797–4806
98 Carmeliet, P., Ferreira, V., Breier, G. et al. (1996)Abnormal blood vessel development and lethality inembryos lacking a single VEGF allele. Nature (London)380, 435–439
99 Ferrara, N., Carver-Moore, K., Chen, H. et al. (1996)Heterozygous embryonic lethality induced by targetedinactivation of the VEGF gene. Nature (London) 380,439–442
100 Miquerol, L., Langille, B. L. and Nagy, A. (2000)Embryonic development is disrupted by modest increasesin vascular endothelial growth factor gene expression.Development 127, 3941–3946
101 Fong, G. H., Rossant, J., Gertsenstein, M. and Breitman,M. L. (1995) Role of the Flt-1 receptor tyrosine kinase inregulating the assembly of vascular endothelium. Nature(London) 376, 66–70
102 Shalaby, F., Rossant, J., Yamaguchi, T. P. et al. (1995)Failure of blood-island formation and vasculogenesis inFlk-1-deficient mice. Nature (London) 376, 62–66
103 Hiratsuka, S., Minowa, O., Kuno, J., Noda, T. andShibuya, M. (1998) Flt-1 lacking the tyrosine kinasedomain is sufficient for normal development andangiogenesis in mice. Proc. Natl. Acad. Sci. U.S.A. 95,9349–9354
104 Takahashi, T., Ueno, H. and Shibuya, M. (1999) VEGFactivates protein kinase C-dependent, butRas-independent Raf-MEK-MAP kinase pathway forDNA synthesis in primary endothelial cells. Oncogene18, 2221–2230
105 Sakurai, Y., Ohgimoto, K., Kataoka, Y., Yoshida, N. andShibuya, M. (2005) Essential role of Flk-1 (vascularendothelial growth factor receptor-2) tyrosine residue1173 in vasculogenesis in mice. Proc. Natl. Acad.Sci. U.S.A. 102, 1076–1081
106 Lamalice, L., Houle, F., Jourdan, G. and Huot, J. (2004)Phosphorylation of tyrosine 1214 on VEGFR2 is requiredfor VEGF-induced activation of Cdc42 upstream ofSAPK2/p38. Oncogene 23, 434–445
107 Vinals, F., Chambard, J. C. and Pouyssegur, J. (1999) p70S6 kinase-mediated protein synthesis is a critical step forvascular endothelial cell proliferation. J. Biol. Chem. 274,26776–26782
108 Yamazaki, T., Akada, T., Niizeki, O., Suzuki, T.,Miyashita, H. and Sato, Y. (2004) Puromycin-insensitiveleucyl-specific aminopeptidase (PILSAP) binds andcatalyzes PDK1, allowing VEGF-stimulated activation ofS6K for endothelial cell proliferation and angiogenesis.Blood 104, 2345–2352
109 Baldanzi, G., Mitola, S., Cutrupi, S. et al. (2004)Activation of diacylglycerol kinase α is required forVEGF-induced angiogenic signaling in vitro. Oncogene23, 4828–4838
110 Chai, J., Jones, M. K. and Tarnawski, A. S. (2004) Serumresponse factor is a critical requirement for VEGFsignaling in endothelial cells and VEGF-inducedangiogenesis. FASEB J. 18, 1264–1266
111 Zhou, R. H., Yao, M., Lee, T. S., Zhu, Y.,Martins-Green, M. and Shyy, J. Y. (2004) Vascularendothelial growth factor activation of sterol regulatoryelement binding protein: a potential role in angiogenesis.Circ. Res. 95, 471–478
112 Yamaoka-Tojo, M., Ushio-Fukai, M., Hilenski, L. et al.(2004) IQGAP1, a novel vascular endothelial growthfactor receptor binding protein, is involved in reactiveoxygen species–dependent endothelial migration andproliferation. Circ. Res. 95, 276–283
113 Watanabe, K., Hasegawa, Y., Yamashita, H. et al. (2004)Vasohibin as an endothelium-derived negative feedbackregulator of angiogenesis. J. Clin. Invest. 114, 898–907
114 Hesser, B. A., Liang, X. H., Camenisch, G. et al. (2004)Down syndrome critical region protein 1 (DSCR1), anovel VEGF target gene that regulates expression ofinflammatory markers on activated endothelial cells.Blood 104, 149–158
115 Minami, T., Horiuchi, K., Miura, M. et al. (2004) Vascularendothelial growth factor- and thrombin-inducedtermination factor, down syndrome critical region-1,attenuates endothelial cell proliferation and angiogenesis.J. Biol. Chem. 279, 50537–50554
116 Luo, J. C., Yamaguchi, S., Shinkai, A., Shitara, K. andShibuya, M. (1998) Significant expression of vascularendothelial growth factor/vascular permeability factor inmouse ascites tumors. Cancer Res. 58, 2652–2660
117 Zebrowski, B. K., Yano, S., Liu, W. et al. (1999) Vascularendothelial growth factor levels and induction ofpermeability in malignant pleural effusions. Clin. CancerRes. 5, 3364–3368
118 Kevil, C. G., Payne, D. K., Mire, E. and Alexander, J. S.(1998) Vascular permeability factor/vascular endothelialcell growth factor-mediated permeability occurs throughdisorganization of endothelial junctional proteins. J. Biol.Chem. 273, 15099–15103
119 Eliceiri, B. P., Paul, R., Schwartzberg, P. L., Hood, J. D.,Leng, J. and Cheresh, D. A. (1999) Selective requirementfor Src kinases during VEGF-induced angiogenesis andvascular permeability. Mol. Cell 4, 915–924
C© 2005 The Biochemical Society

240 H. Takahashi and M. Shibuya
120 Weis, S., Shintani, S., Weber, A. et al. (2004) Src blockadestabilizes a Flk/cadherin complex, reducing edema andtissue injury following myocardial infarction.J. Clin. Invest. 113, 885–894
121 Fukumura, D., Yuan, F., Endo, M. and Jain, R. K. (1997)Role of nitric oxide in tumor microcirculation. Bloodflow, vascular permeability, and leukocyte-endothelialinteractions. Am. J. Pathol. 150, 713–725
122 Fukumura, D., Gohongi, T., Kadambi, A. et al. (2001)Predominant role of endothelial nitric oxide synthase invascular endothelial growth factor-induced angiogenesisand vascular permeability. Proc. Natl. Acad. Sci. U.S.A.98, 2604–2609
123 Fulton, D., Gratton, J. P., McCabe, T. J. et al. (1999)Regulation of endothelium-derived nitric oxideproduction by the protein kinase Akt. Nature (London)399, 597–601
124 Dimmeler, S., Fleming, I., Fisslthaler, B., Hermann, C.,Busse, R. and Zeiher, A. M. (1999) Activation of nitricoxide synthase in endothelial cells by Akt-dependentphosphorylation. Nature (London) 399, 601–605
125 Eriksson, A., Cao, R., Roy, J. et al. (2003) SmallGTP-binding protein Rac is an essential mediator ofvascular endothelial growth factor-induced endothelialfenestrations and vascular permeability. Circulation 107,1532–1538
126 Issbrucker, K., Marti, H. H., Hippenstiel, S. et al. (2003)p38 MAP kinase–a molecular switch betweenVEGF-induced angiogenesis and vascularhyperpermeability. FASEB J. 17, 262–264
127 Fukumura, D., Xavier, R., Sugiura, T. et al. (1998) Tumorinduction of VEGF promoter activity in stromal cells.Cell 94, 715–725
128 Holmgren, L., O’Reilly, M. S. and Folkman, J. (1995)Dormancy of micrometastases: balanced proliferation andapoptosis in the presence of angiogenesis suppression.Nat. Med. 1, 149–153
129 Ferrara, N., Hillan, K. J., Gerber, H. P. and Novotny, W.(2004) Discovery and development of bevacizumab, ananti-VEGF antibody for treating cancer. Nat. Rev. DrugDiscov. 3, 391–400
130 Hiratsuka, S., Maru, Y., Okada, A., Seiki, M., Noda, T.and Shibuya, M. (2001) Involvement of Flt-1 tyrosinekinase (vascular endothelial growth factor receptor-1) inpathological angiogenesis. Cancer Res. 61, 1207–1213
131 Autiero, M., Waltenberger, J., Communi, D. et al. (2003)Role of PlGF in the intra- and intermolecular cross talkbetween the VEGF receptors Flt1 and Flk1. Nat. Med. 9,936–943
132 Yoshiji, H., Kuriyama, S., Yoshii, J. et al. (2004) Haltingthe interaction between vascular endothelial growthfactor and its receptors attenuates liver carcinogenesis inmice. Hepatology 39, 1517–1524
133 Christofori, G., Naik, P. and Hanahan, D. (1995) Vascularendothelial growth factor and its receptors, flt-1 andflk-1, are expressed in normal pancreatic islets andthroughout islet cell tumorigenesis. Mol. Endocrinol. 9,1760–1770
134 Inoue, M., Hager, J. H., Ferrara, N., Gerber, H. P. andHanahan, D. (2002) VEGF-A has a critical, nonredundantrole in angiogenic switching and pancreatic β cellcarcinogenesis. Cancer Cell 1, 193–202
135 Bergers, G., Brekken, R., McMahon, G. et al. (2000)Matrix metalloproteinase-9 triggers the angiogenic switchduring carcinogenesis. Nat. Cell Biol. 2, 737–744
136 Weis, S., Cui, J., Barnes, L. and Cheresh, D. (2004)Endothelial barrier disruption by VEGF-mediated Srcactivity potentiates tumor cell extravasation andmetastasis. J. Cell Biol. 167, 223–229
137 Schmitt, M., Horbach, A., Kubitz, R., Frilling, A. andHaussinger, D. (2004) Disruption of hepatocellular tightjunctions by vascular endothelial growth factor (VEGF):a novel mechanism for tumor invasion. J. Hepatol. 41,274–283
138 Hiratsuka, S., Nakamura, K., Iwai, S. et al. (2002) MMP9induction by vascular endothelial growth factorreceptor-1 is involved in lung-specific metastasis. CancerCell 2, 289–300
139 Hurwitz, H., Fehrenbacher, L., Novotny, W. et al. (2004)Bevacizumab plus irinotecan, fluorouracil, and leucovorinfor metastatic colorectal cancer. New Engl. J. Med. 350,2335–2342
140 Melder, R. J., Koenig, G. C., Witwer, B. P., Safabakhsh,N., Munn, L. L. and Jain, R. K. (1996) Duringangiogenesis, vascular endothelial growth factor andbasic fibroblast growth factor regulate natural killer celladhesion to tumor endothelium. Nat. Med. 2,992–997
141 Kim, I., Moon, S. O., Kim, S. H., Kim, H. J., Koh, Y. S.and Koh, G. Y. (2001) Vascular endothelial growth factorexpression of intercellular adhesion molecule 1(ICAM-1), vascular cell adhesion molecule 1 (VCAM-1),and E-selectin through nuclear factor-κB activation inendothelial cells. J. Biol. Chem. 276, 7614–7620
142 Barleon, B., Sozzani, S., Zhou, D., Weich, H. A.,Mantovani, A. and Marme, D. (1996) Migration of humanmonocytes in response to vascular endothelial growthfactor (VEGF) is mediated via the VEGF receptor flt-1.Blood 87, 3336–3343
143 Detmar, M., Brown, L. F., Schon, M. P. et al. (1998)Increased microvascular density and enhanced leukocyterolling and adhesion in the skin of VEGF transgenic mice.J. Invest. Dermatol. 111, 1–6
144 Kunstfeld, R., Hirakawa, S., Hong, Y. K. et al. (2004)Induction of cutaneous delayed-type hypersensitivityreactions in VEGF-A transgenic mice results in chronicskin inflammation associated with persistent lymphatichyperplasia. Blood 104, 1048–1057
145 Oura, H., Bertoncini, J., Velasco, P., Brown, L. F.,Carmeliet, P. and Detmar, M. (2003) A critical role ofplacental growth factor in the induction of inflammationand edema formation. Blood 101, 560–567
146 Matsumoto, Y., Tanaka, K., Hirata, G. et al. (2002)Possible involvement of the vascular endothelial growthfactor-Flt-1-focal adhesion kinase pathway in chemotaxisand the cell proliferation of osteoclast precursor cells inarthritic joints. J. Immunol. 168, 5824–5831
147 Luttun, A., Tjwa, M., Moons, L. et al. (2002)Revascularization of ischemic tissues by PlGF treatment,and inhibition of tumor angiogenesis, arthritis andatherosclerosis by anti-Flt1. Nat. Med. 8, 831–840
148 De Bandt, M., Ben Mahdi, M. H., Ollivier, V. et al. (2003)Blockade of vascular endothelial growth factor receptor I(VEGF-RI), but not VEGF-RII, suppresses jointdestruction in the K/BxN model of rheumatoid arthritis.J. Immunol. 171, 4853–4859
149 Lee, Y. C. and Lee, H. K. (2001) Vascular endothelialgrowth factor in patients with acute asthma. J. AllergyClin. Immunol. 107, 1106
150 Hoshino, M., Nakamura, Y. and Hamid, Q. A. (2001)Gene expression of vascular endothelial growth factor andits receptors and angiogenesis in bronchial asthma.J. Allergy Clin. Immunol. 107, 1034–1038
151 Lee, C. G., Link, H., Baluk, P. et al. (2004) Vascularendothelial growth factor (VEGF) induces remodelingand enhances TH2-mediated sensitization andinflammation in the lung. Nat. Med. 10, 1095–1103
152 Reinders, M. E., Sho, M., Izawa, A. et al. (2003)Proinflammatory functions of vascular endothelialgrowth factor in alloimmunity. J. Clin. Invest. 112,1655–1665
153 Zhao, Q., Egashira, K., Hiasa, K. et al. (2004) Essentialrole of vascular endothelial growth factor and Flt-1signals in neointimal formation after periadventitialinjury. Arterioscler., Thromb., Vasc. Biol. 24,2284–2289
154 Marti, H. J., Bernaudin, M., Bellail, A. et al. (2000)Hypoxia-induced vascular endothelial growth factorexpression precedes neovascularization after cerebralischemia. Am. J. Pathol. 156, 965–976
155 van Bruggen, N., Thibodeaux, H., Palmer, J. T. et al.(1999) VEGF antagonism reduces edema formation andtissue damage after ischemia/reperfusion injury in themouse brain. J. Clin. Invest. 104, 1613–1620
C© 2005 The Biochemical Society

VEGF/VEGFR system in health and disease 241
156 Paul, R., Zhang, Z. G., Eliceiri, B. P. et al. (2001) Srcdeficiency or blockade of Src activity in mice providescerebral protection following stroke. Nat. Med. 7,222–227
157 Sun, Y., Jin, K., Xie, L. et al. (2003) VEGF-inducedneuroprotection, neurogenesis, and angiogenesis afterfocal cerebral ischemia. J. Clin. Invest. 111, 1843–1851
158 Manoonkitiwongsa, P. S., Schultz, R. L., McCreery, D. B.,Whitter, E. F. and Lyden, P. D. (2004) Neuroprotection ofischemic brain by vascular endothelial growth factor iscritically dependent on proper dosage and may becompromised by angiogenesis. J. Cereb. Blood FlowMetab. 24, 693–702
159 Campochiaro, P. A. and Hackett, S. F. (2003) Ocularneovascularization: a valuable model system. Oncogene22, 6537–6548
160 Gragoudas, E. S., Adamis, A. P., Cunningham, Jr, E. T.,Feinsod, M. and Guyer, D. R. (2004) Pegaptanib forneovascular age-related macular degeneration. New Engl.J. Med. 351, 2805–2816
161 Oosthuyse, B., Moons, L., Storkebaum, E. et al. (2001)Deletion of the hypoxia-response element in the vascularendothelial growth factor promoter causes motor neurondegeneration. Nat. Genet. 28, 131–138
162 Lambrechts, D., Storkebaum, E., Morimoto, M. et al.(2003) VEGF is a modifier of amyotrophic lateralsclerosis in mice and humans and protects motoneuronsagainst ischemic death. Nat. Genet. 34, 383–394
163 Azzouz, M., Ralph, G. S., Storkebaum, E. et al. (2004)VEGF delivery with retrogradely transported lentivectorprolongs survival in a mouse ALS model.Nature (London) 429, 413–417
164 Storkebaum, E., Lambrechts, D., Dewerchin, M. et al.(2005) Treatment of motoneuron degeneration byintracerebroventricular delivery of VEGF in a rat modelof ALS. Nat. Neurosci. 8, 85–92
165 LeCouter, J., Moritz, D. R., Li, B. et al. (2003)Angiogenesis-independent endothelial protection of liver:role of VEGFR-1. Science 299, 890–893
166 Dixelius, J., Makinen, T., Wirzenius, M. et al. (2003)Ligand-induced vascular endothelial growth factorreceptor-3 (VEGFR-3) heterodimerization withVEGFR-2 in primary lymphatic endothelial cellsregulates tyrosine phosphorylation sites. J. Biol. Chem.278, 40973–40979
167 Cao, L., Jiao, X., Zuzga, D. S. et al. (2004) VEGF linkshippocampal activity with neurogenesis, learning andmemory. Nat. Genet. 36, 827–835
Received 23 December 2004/5 April 2005; accepted 9 May 2005Published on the Internet 24 August 2005, doi:10.1042/CS20040370
C© 2005 The Biochemical Society