thymidine triphosphate : a-d-galactose l-phosphate
TRANSCRIPT

THE JOURNAL cm BIOLOGICAL CHEMISTRY Vol. 238, No. 10, October 1963
Printed in U.S.A.
Thymidine Triphosphate : a-D-Galactose l-Phosphate
Thymidylyltransferase from Streptococcus faecalis
Grown on D-Galactose*
JOHN H. PAZUR AND JOHN S. ANDERSON
From the Department of Biochemistry and Nutrition, the University of Nebraska, College of Agriculture, Lincoln S, Nebraska
(Received for publication, March 19, 1963)
In a preliminary communication (1) it was reported that Streptococcus faecalis grown on n-galactose elaborated an induci- ble enzyme, a thymidine triphosphate (dTTP) : a-D-galactose l-phosphate thymidylyltransferase (thymidine diphosphate galactose pyrophosphorylase) which effects the synthesis of thymidine diphosphate galactose from thymidine triphosphate and a-n-galactose-1-P. The suggestion was made that this enzyme was utilized for the synthesis of cellular constituents which can arise only from intact hexose moieties via the thymi- dine diphosphate hexose pathway. That D-galactose-1-V is indeed incorporated into cellular constituents without randomi- zation of the label has now been verified. D-Glucose, D-galactose, and L-rhamnose isolated from hydrolysates of cells grown on n-galactose-l-Cl4 as the sole carbohydrate source were labeled exclusively in the l-position. Of considerable interest was the finding that D-glucosamine isolated from the hydrolysate was also labeled in the l-position indicating that this hexosamine arises from an intact hexose moiety. Results of experiments on the stability of the transferases at 3” and on their paper elec- trophoretic mobility showed that the dTTP : cr-n-galactose-1-P thymidylyltransferase differs from the dTTP : a-n-glucose-l-P thymidylyltransferase and uridylyltransferases also produced by S. faecalis. The enzyme exhibited a narrow pH activity range with maximal activity at pH 7. From its sedimentation rate on density gradient centrifugation (2, 3) it was estimated that the molecular weight of the dTTP : or-n-galactose-1-P thymidylyl- transferase is in the range of 80,000 to 100,000.
EXPERIMENTAL PROCEDURE AND RESULTS
Estimation of Thymidylyltransferase and Uridylyltransferase ActivitiesS. faecalis was grown on nutrient broth containing 0.1 y0 D-glucose, n-galactose, L-rhamnose, or L-arabinose. En- zyme extracts buffered to pH 7 in 0.1 M potassium phosphate containing 0.01 M magnesium chloride were prepared from the cells from liter cultures by a published procedure (4). The pro- tein concentrations in the extracts as measured by the method of Lowry et al. (5) ranged from 15 to 20 mg per ml. These en- zyme extracts were examined immediately for thymidylyl- and uridylyltransferase activities by the following method. Assay tubes containing 1 mg of thymidine triphosphate and 1 mg of a-u-galactose-1-P in 0.05 ml of solution buffered to pH 7, and
* Published with the approval of the Director as Paper 1349, Journal Series, Nebraska Agricultural Experiment Station. Supported in part by a grant from the National Science Founda- tion.
0.05 ml of the enzyme extract were incubated at room tempera- ture for 4 hours. A O.Ol-ml aliquot of the reaction mixture and another of an enzyme blank were analyzed for new ultraviolet- absorbing compounds by paper chromatography (6). As judged by pHydrion paper, no detectable change in pH of the reaction mixture occurred. The amount of nucleotide diphosphate hexoses in each mixture was determined by elution of the com- pounds from the chromatogram with 3 ml of water and by meas- uring the optical density of the solution at 260 mp. The optical density values for the samples ranged from 0.150 to 0.350. A correction factor of from 0.025 to 0.040 optical unit was applied to these values and represents the ultraviolet-absorbing material in a blank of a comparable area of the paper. On the basis of the yield of nucleotide diphosphate hexoses, relative values for transferase activities were calculated. Uridine triphosphate and cr-D-glucose-l-P were also used as substrates for the enzyme preparations. Although for some of the transferases, assay methods coupled with NADP reduction could be used, all trans- ferases were assayed by the chromatographic technique in order for the values to be comparable. The large amounts of sub- strates per assay tube were required to facilitate detection of the compounds on the paper. In an experiment designed to check the reproducibility of the assay, values from eight assays agreed to within lo’%, indicating a reasonable reproducibility of the assay. In some experiments the amounts of nucleotide di- phosphate hexoses were estimated by comparison of the intensi- ties of the ultraviolet spots with that of known concentrations of the compounds.
The results showing the production of thymidine and uridine diphosphate hexoses with the enzyme preparations from cells grown on n-glucose and n-galactose are presented in Table I. It will be noted that the enzyme preparation from the cells adapted to n-glucose effected a rapid synthesis of UDP-glucose, UDP-galactose, and dTDP-glucose. A nucleotide diphosphate hexose was not detectable in the reaction mixture of thymidine triphosphate and a-n-galactose-1-P with this enzyme prepara- tion, even on prolonged incubation. The enzyme preparation from the cells grown on n-galactose produced the nucleotide diphosphate hexoses mentioned above from the appropriate substrates and in addition produced nucleotide diphosphate hexoses from thymidine triphosphate and cr-D-galactose-1-P. The new nucleotide material in the latter reaction mixture was isolated by paper chromatography (4). A sample of the ma- terial was hydrolyzed in dilute acid and the nature of the hy- drolytic products was determined. On the basis of the types of
3155
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

3156 dTTP: a-o-Galactose-1-P Thymidylyltransferase Vol. 238, No. 10
TABLE I Initial nucleotide diphosphate hexoses synthesized by enzyme extracts of S. jaecalis
Reaction mixture Enzyme extract from glucose-adapted cells Enzyme extract from galactose-adapted cells Rp values of nucleotide hexoses
UTP + glucose-l-P.. UDP-glucose UDP-glucose 0.27 UTP + galactose-1-P. UDP-galactose UDP-galactose 0.26 dTTP + glucose-l-P.. dTDP-glucose dTDP-glucose 0.38 dTTP + galactose-l-P.................... None dTDP-galactose 0.37
hydrolytic products obtained, this nucleotide fraction appeared to be a mixture of thymidine diphosphate galactose, thymidine diphosphate glucose, and thymidine diphosphate rhamnose. Apparently the cells grown on D-g&XtOSe produced an inducible enzyme, a thymidylyltransferase which effected the synthesis of thymidine diphosphate galactose from thymidine triphosphate and a-n-galactose-1-P. This enzyme was not detectable in enzyme extracts from cells grown on other carbohydrates such as n-glucose, L-rhamnose, or L-arabinose. The thymidine di- phosphate galactose was, in turn, converted to thymidine di- phosphate glucose and eventually to thymidine diphosphate rhamnose by the constitutive enzyme complex in the bacterial extracts. Dialysis of the crude enzyme preparation removed the cofactors necessary for the production of thymidine diphos- phate rhamnose but did not affect the production of thymidine diphosphate glucose from thymidine diphosphate galactose.
It is unlikely that the thymidine diphosphate hexoses in the reaction mixture with the enzyme extracts from n-galactose- adapted cells were produced via the uridine pathway coupled to the thymidine pathway for the following reasons. First, when UDP-glucose or UDP-galactose and thymidine triphosphate were incubated with the enzyme extract prepared from the D- galactose-adapted cells, thymidine diphosphate hexoses were not produced in either digest. Apparently under the conditions of the experiments, an interconversion of the uridine and thymi- dine compounds did not occur. Second, since both the extracts from the o-glucose- and the n-galactose-adapted cells possessed UDP-galactose 4-epimerase activity, thymidine diphosphate hexose nucleotides should have been obtained from thymidine triphosphate and a-n-galactose-1-P with both preparations if the uridine pathway were involved. As shown in Table I, a synthesis of thymidine diphosphate galactose was not obtained with the enzyme extracts from the cells grown on n-glucose. Finally, as described in a later section, a separation of the dTTP : galactose-1-P thymidylyltransferase from the other transferases has been achieved by paper electrophoresis.
Evidence for Thymidine Diphosphate Galactose-A reaction mixture of 30 mg of thymidine triphosphate and 30 mg of Q-D-
galactose-I-P in 2 ml of enzyme extract from the D-galactose- adapted cells was prepared and incubated for 6 hours at room temperature. The material in the reaction mixture which mi- grated at RF value of 0.37 in the solvent system of ethyl alcohol and 1 M ammonium acetate was extracted from the chromato- gram with water and concentrated to a small volume (0.4 ml) under vacuum. The over-all yield of the nucleotide was 30% based on the initial thymidine triphosphate employed. Hy- drolysis of an aliquot of the above sample in 0.1 N hydrochloric acid at 100” for 2 hours converted the product into thymidine monophosphate, inorganic phosphate and the reducing sugars, n-galactose, n-glucose, and L-rhamnose. The thymidine mono-
phosphate was identified by its RF values in several solvents and by its ultraviolet absorption spectra and the inorganic phos- phate was determined by a calorimetric procedure. The re- ducing compounds in the hydrolysate possessed apparent RF values of 0.59, 0.65, and 0.88 on paper after three ascents of the solvent system n-butyl alcohol-pyridine-water (45 : 25 : 35 by volume). These values are characteristic of n-galactose, D-
glucose, and L-rhamnose, respectively, in this solvent system. The presence of D-glucose and L-rhamnose in the hydrolysate was confirmed by use of the glucose oxidase reaction (7) and the Dische-Shettles calorimetric reaction (8).
The n-galactose was further identified in the hydrolysate by use of an enzyme specific for n-galactose, namely, galactose oxidase (9). In the procedure an O.l-ml aliquot of the neutral- ized hydrolysate was mixed with 2 ml of a peroxidase solution, buffered to pH 7 and containing 0.02% o-dianisidine, and with 0.2 ml of the galactose oxidase. Adequate controls of n-galac- tose, n-glucose, L-rhamnose, and blanks of reagents were also prepared in a similar manner. After incubation at 30” for 2 hours, a red color developed in the solution of n-galactose and the hydrolysate of the nucleotide fraction, establishing the pres- ence of n-galactose in the hydrolysate.
A preparation of pure thymidine diphosphate galactose has not yet been obtained in our laboratory because the enzyme preparations employed also contain the epimerase, which effects the conversion of thymidine diphosphate galactose to thymidine diphosphate glucose. A partial separation of the two nucleo- tides is, however, possible in the solvent system of ethyl alcohol and ammonium acetate. This difference may facilitate the preparation of pure thymidine diphosphate galactose. In this connection, it may be advantageous to utilize the thymidylyl- transferase from plant tissues for the preparation of thymidine diphosphate galactose since the latter enzyme is separable from the epimerase (10).
Isolation and Degradation of o-Gahtose, D-Glucose, and L-
Rhamnose from Cells Grown on &Gala&se-l -Cl-Several batches of CY4-labeled cells were obtained by culturing S. faecalti in 500 ml of nutrient broth solution containing 0.1 y. of n-galactose-l-C14 (total radioactivity, 0.1 mc) for 18 hours at 37”. Each batch of cells was collected separately by centrifugation, washed thor- oughly with water, and refluxed for 6 hours in 6 ml of 1.0 N
sulfuric acid. After neutralization of the acid in the reaction mixture with barium hydroxide and removal of insoluble ma- terial by centrifugation, the clear supernatant fluid was concen- trated to a volume of 2 ml. Aliquots of 0.1 ml were placed on large paper chromatograms (40 cm x 18 cm) and the carbo- hydrate components in the hydrolysate were separated by two ascents of the solvent systems of n-butyl alcohol-pyridine-water (45 :25 :35 by volume). For locating the radioactive com- pounds in the mixture, radioautograms of the chromatograms
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

October 1963 J. H. Pazur and J. S. Anderson 3157
were prepared. Two major radioactive components were found on the chromatogram at apparent RF values of 0.84 and 0.64 and minor bands were present, at apparent RF values of 0.73, 0.58, and 0.53. The former RF values are characteristic of L-rhamnose and n-glucose whereas the latter are characteristic of N-acetyl-n-glucosamine, n-galactose, and u-glucosamine. Additional evidence for these compounds was the typical color reaction with the aniline oxalate reagent (ll), and with the acetyl acetone-dimethylaminobenzaldehydc reagent (12). In addition the product at RF value 0.84 yielded a color complex with cysteine-sulfuric acid which had an absorption spectrum characteristic for the product from L-rhamnose (8), and the com- pound at RF 0.64 yielded a positive reaction with glucose oxidase (7); the further characterization of the minor components is described in a later section.
The individual radioactive compounds on the paper chromato- gram were extracted with water and concentrated to dryness. To the residues were added 200 mg of carrier carbohydrate (L-rhamnose, n-glucose, or n-galactose), 0.34 ml of concentrated hydrochloric acid, and 0.26 ml of ethyl mercaptan. Crystalline diethyl dithioacetals were obtained with all three compounds. These dithioacetals were recrystallized to constant specific ac- tivity and constant melting points; rhamnose derivative, 196 c.p.m. per pmole and m.p. 133”; glucose derivative, 305 c.p.m. per pmole and m.p. 114’; and the galactose derivative, 20 c.p.m. per pmole and m.p. 139”.
The dithioacetals were oxidized to the disulfones with perpro- pionic acid (13) and crystallized from isopropyl alcohol. The disulfone of rhamnose melted at 176179”, the disulfone of D-glu- case melted at 133-136”, and the disulfone of n-galactose melted over a wide range, 139-157”. Radioactivity measurements were also made on the crystalline disulfone derivatives and the specific activities were calculated. The disulfones were cleaved with dilute ammonium hydroxide to bis(ethylsulfonyl)methane and a pentosc (13). The bis(ethylsulfonyl)methane was extracted with chloroform and obtained in crystalline form, nip. 97-99”, from all three of the dithioacetals. The pentose fragment from the disulfones remained in the aqueous phase. This phase was concentrated to a small volume and the pentose was derivatized as the dithioacetal. The dithioacetal of 5-deoxy-L-arabinose, m.p. 101-103”, was obtained from the rhamnose disulfone whereas the dithioacetal of n-arabinose, m.p. 118-119”, and the dithioacetal of n-lyxose, m.p. 94-97”, were obtained from the glucose and galactose disulfones, respectively. The specific activities of the original hexose dithioacetals, the hexose disul- fones, the bis(ethylsulfonyl)methane, and the pentose dithio- acetals were determined in a Packard Tri-Carb liquid scintillation spectrometer. h sample of the original n-galactose-1-C” used as the carbohydrate source for the S. faecalis was also degraded by the above procedure and the specific activities of various derivatives and fragments were determined. The data are pre- sented in Table II.
Isolation and Degradation of N-Scetgl-o-glucosamine from Cells Grown on o-Gala&se-l-C ILIt was indicated in the previ- ous section that the radioactive material at RF 0.73 on the chro- matogram exhibited some characteristics of N-acetyl-n-glucos- amine. Since n-ribose also migrated with the same RF value in this solvent and since it was present in small amounts in the hydrolysate, it was necessary to establish that radioactive N-acetyl-n-glucosamine was indeed present in the hydrolysate. The material at Rp 0.73 on the chromatogram was eluted and
TABLE 11
Specific activities of derivatives and fragments of hexoses obtained from cell wall of S. jaecalis grown on o-galactose-l -Cl4
I Smcific activities I
Galactose-1-W.. . Cell wall hexoses:
Rhamnose Glucose............. Galactose...........
c.p.m.//mk?1e % 467 468 467 1.1 99.8
196 187 198 0.2 99.9 305 282 289 1.5 99.5
20 20 21 0.1 99.6
concentrated to a small volume. Chromatography of an aliquot of this solution in a solvent system of phenol saturated with water and containing 0.570 ammonium hydroxide resulted in a separation of the N-acetyl-n-glucosamine (RF value, 0.72) and the n-ribose (RF value, 0.63). Most of the radioactivity on the chromatogram was found in the X-acetyl-n-glucosamine area. Acid hydrolysis of the preparation containing the N-acetyl- hexosamine and n-ribose removed the acetyl groups from the hexosamine. Chromatography of the hydrolysate by two ascents of the solvent system n-butyl alcohol-pyridine-water (45 :25 :35 by volume) separated the deacetylated n-glucosamine (RF value, 0.43) and the n-ribose (RF value, 0.69). However, in the hydrolysate a second radioactive component with an apparent RF value 0.39 was also detected. The characteriza- tion of the compound at RF 0.43 as n-glucosamine is described below; the characterization of the compound at RF value of 0.39 is still in progress. Evidence thus far indicates that the latter compound is n-galactosamine. Although some evidence for the presence of n-galactosamine in the cell wall of S. faecalis has been presented (14), additional evidence is desirable for the pres- ence of n-galactosamine in the cell wall and for the nature of the linkage by which the n-galactosamine is attached to other units of the cell wall polysaccharides.
The compound at RF 0.43 was eluted from the chromatogram and concentrated to 0.2 ml. The yield of compound was 60 pg and the specific activity was 33,600 c.p.m. per pmole. This value is lower than the specific activity (45,000 c.p.m. per pmole) of the original n-galactose employed in the experiment. The compound yielded a color reaction characteristic of hexosamine with the ninhydrin reagent, the silver nitrate reagent, and the Elson and Morgan reagent. Degradation of the n-glucosamine to a carbon 1 fragment and a pentose was achieved with the ninhydrin reaction (15). The pentose possessed an RF value identical with n-arabinose and yielded a typical color reaction for a pentose in the aniline oxalate reaction and the orcinol reac- tion. The area containing n-arabinose on a duplicate chromato- gram was eluted with water and concentrated to a syrup. The syrup was dissolved in 0.5 ml of ethyl alcohol and mixed with 15 ml of scintillation solution. The radioactivity of the sample was measured in a Tri-Carb liquid scintillation spectrometer. The specific activity of the material isolated from the chromato- gram was found to be 910 c.p.m. per pmole. However, since a complete separation of the n-arabinose from a n-glucosamine- ninhydrin reaction complex was difficult to achieve by the chromatographic technique employed, it is likely that much of the radioactivity in this fraction is due to the latter compound
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
3158 dTTP: a-o-Galactose-1-P Thymidylyltranqferase Vol. 238, so. 10
100 0 0
I------ \ l
\ 0 ---->--A
so ‘1 9
\ .
60 t\
-\ \
\
I\ \
70 \
\
2 \
\ 60
\ t
\ \
\ ‘\
‘.
IO -
o- ’ ’ ’ ’ 0 2 4 6 6 IO 12 14 I6
TIME (DAYS)
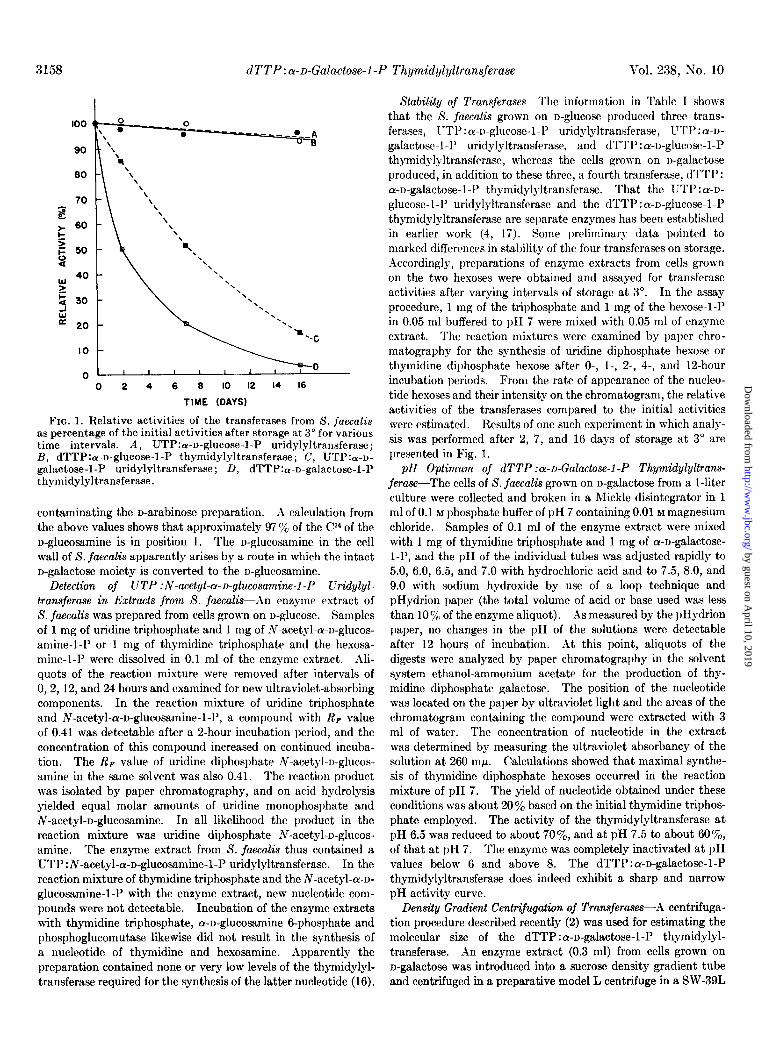
FIG. 1. Relative activities of the transferases from 8. fuecalis as percentage of the initial activities after storage at 3” for various time intervals. A, UTP:a-u-glucose-l-P uridylyltransferase; B, dTTP:a-n-glucose-l-P thymidylyltransferase; C, UTP:a-n- galactose-1-P uridylyltransferase; D, dTTP:a-n-galactose-1-P thymidylyltransferase.
contaminating the n-arabinose preparation. .I calculation from the above values shows that approximately 97 ye of the Cl4 of the n-glucosamine is in position 1. The n-glucosamine in the cell wall of S. faecalis apparently arises by a route in which the intact
n-galactose moiety is converted to the n-glucosamine. Detection of UTP :N-acetyt-a-o-glucosamine-1 -P Uridylyl-
transferase in Extracts from S. faecalti-An enzyme extract of S. faecalis was prepared from cells grown on n-glucose. Samples of 1 mg of uridine triphosphate and 1 mg of N-acetyl-cY-n-glucos- amine-l-P or 1 mg of thymidine triphosphate and the hexosa- mine-l-P were dissolved in 0.1 ml of the enzyme extract. Ali- quots of the reaction mixture were removed after intervals of 0, 2, 12, and 24 hours and examined for new ultraviolet-absorbing components. In the reaction mixture of uridine triphosphate
and N-acetyl-a-n-glucosamine-l-P, a compound with RF value of 0.41 was detectable after a 2-hour incubation period, and the concentration of this compound increased on continued incuba- tion. The Rp value of uridine diphosphate N-acetyl-n-glucos- amine in the same solvent was also 0.41. The reaction product was isolated by paper chromatography, and on acid hydrolysis yielded equal molar amounts of uridine monophosphate and N-acetyl-n-glucosamine. In all likelihood the product in the reaction mixture was uridine diphosphate N-acetyl-n-glucos- amine. The enzyme extract from S. faecalis thus contained a UTP :N-acetyl-a-n-glucosamine-1-P uridylyltransferase. In the reaction mixture of thymidine triphosphate and the N-acetyl-a-n- glucosamine-1-P with the enzyme extract, new nucleotide com- pounds were not detectable. Incubation of the enzyme extracts with thymidine triphosphate, a-n-glucosamine 6-phosphate and phosphoglucomutase likewise did not result in the synthesis of a nucleotide of thymidine and hexosamine. Apparently the preparation contained none or very low levels of the thymidylyl- transferase required for the synthesis of the latter nucleotide (16).
Stability of Transferuses--The information in Table I shows that the S. faecalis grown on n-glucose produced three trans- ferases, I-Tl’:cr-r)-glucose-l-l’ uridylyltransferase, I,T'rl':a-~-
galactose-l-l’ uridylyltransferase, and dTTP : cy-D-glucose-l-P thymidylyltransferase, whereas the cells grown on n-galactose produced, in addition to these three, a fourth transferase, dTTP: a-n-galactose-1-P thymidylyltransferase. That the UTP : a-~- glucose-l-P uridylyltransferase and the dTTP:a-o-glucose-l-l’ thymidylyltransferase are separate enzymes has been established in earlier work (4, 17). Some preliminary data pointed to marked differences in stability of the four transferases on storage. A1ccordingly, preparations of enzyme extracts from cells grown on the two hexoses were obtained and assayed for transferase activities after varying intervals of storage at 3”. In the assay procedure, 1 mg of the triphosphate and 1 mg of the hexose-1-P in 0.05 ml buffered to pH 7 were mixed with 0.05 ml of enzyme extract. The reaction mixtures were examined by paper chro- matography for the synthesis of uridine diphosphate hexose or thymidine diphosphate hexose after 0-, I-, 2-, 4-, and 12-hour incubation periods. From the rate of appearance of the nucleo- tide hexoses and their intensity on the chromatogram, the relative activities of the transferases compared to the initial activities were estimated. Results of one such experiment in which analy- sis was performed after 2, 7, and 16 days of storage at 3” are presented in Fig. 1.
pH Optimum of dTTP :a-u-Gatactose-1 -P Thymidylyltrans- ferase-The cells of S. faecalis grown on n-galactose from a l-liter culture were collected and broken in a Mickle disintegrator in 1 ml of 0.1 M phosphate buffer of pH 7 containing 0.01 M magnesium chloride. Samples of 0.1 ml of the enzyme extract were mixed with 1 mg of thymidine triphosphate and 1 mg of cu-n-galactose- l-P, and the pH of the individual tubes was adjusted rapidly to
5.0, 6.0, 6.5, and 7.0 with hydrochloric acid and to 7.5, 8.0, and 9.0 with sodium hydroxide by use of a loop technique and pHydrion paper (the total volume of acid or base used was less than 10 y0 of the enzyme aliquot). As measured by the pHydrion paper, no changes in the pH of the solutions were detectable after 12 hours of incubation. At this point, aliquots of the digests were analyzed by paper chromatography in the solvent system ethanol-ammonium acetate for the production of thy- midine diphosphate galactose. The position of the nucleotide was located on the paper by ultraviolet light and the areas of the chromatogram containing the compound were extracted with 3 ml of water. The concentration of nucleotide in the extract was determined by measuring the ultraviolet absorbancy of the solution at 260 mp. Calculations showed that maximal synthe- sis of thymidine diphosphate hexoses occurred in the reaction mixture of pH 7. The yield of nucleotide obtained under these conditions was about 20 ‘% based on the initial thymidine triphos- phate employed. The activity of the thymidylyltransferase at pH 6.5 was reduced to about 70%, and at pH 7.5 to about SO%, of that at pH 7. The enzyme was completely inactivated at pH values below 6 and above 8. The dTTP:cu-n-galactose-1-P thymidylyltransferase does indeed exhibit a sharp and narrow pH activity curve.
Density Gradient Centtifugattin of Transfer-es-A centrifuga- tion procedure described recently (2) was used for estimating the molecular size of the dTTP : cY-n-galactose-1-P thymidylyl- transferase. An enzyme extract (0.3 ml) from cells grown on n-galactose was introduced into a sucrose density gradient tube and centrifuged in a preparative model L centrifuge in a SW-39L
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

October 1963 J. H. Pam- and J. X. Anderson 3159
swinging bucket rotor for 24 hours at 37,500 r.p.m. At the end of this time the tubes were analyzed in the fractionator (2) for ultraviolet-absorbing components and fractions of 0.3 ml of the effluent were collected. The individual tubes were assayed for uridylyltransferase and thymidylyltransferase activities by the methods described in a preceding section. The four types of transferase activity were found in tubes 6, 7, 8, and 9 in about the same relative concentrations. rZpparently the thymidylyl- and uridylyltransferases of S. juecalis are of approximately the same molecular size. From a comparison of the rates of sedi- mentation of the transferases and glucoamylase (2) and the relationship proposed by Martin and Ames (3), it was estimated that the molecular size of the transferases is in the 80,000 to 100,000 range.
Paper Electrophoresis of Transferases--S. faecalis was grown in 5 liters of nutrient broth culture containing D-galactose. The cells were collected in a Sharples centrifuge and broken in 5 ml of phosphate buffer of pH 7 in a Rranson sonifier. An aliquot of the enzyme extract was placed on paper electrophoresis strips in a Spinco model R apparatus by means of the applicator pro- vided with the apparatus. The buffer used in the cell was 0.1 M phosphate buffer of pH 7.6 containing 0.01 M magnesium chloride and 0.01 M mercaptoethanol. Electrophoresis was per- formed at a potential of 120 direct current volts and a current of 15 ma in the cold room for periods of 2, 6, or 18 hours. At the end of these periods, the paper strips were removed and the 5-cm portion of the paper on either side of the origin was cut into 0.5-cm strips. Each strip was placed into separate tubes con- taining 0.1 ml of a 1 y0 solution of uridine or thymidine triphos- phate and appropriate hexose-l-P, buffered to pH 7.2. The mixtures were incubated at room temperatures for periods of 6 to 72 hours and examined periodically for nucleotide diphosphate hexoses by paper chromatography. From the intensity of the nucleotide diphosphate hexose spots on the chromatogram, the relative amounts of transferase in the various tubes were esti- mated. The results showed that after 2 hours of electrophoresis, the transferases had migrated towards the anode and were readily detectable by the assay procedure. After electrophoresis for 6 hours, the positions of the enzymes on the paper strips and the distribution of the protein components in the preparation were as shown in Fig. 2. The protein constituents were detected by staining the strip with bromphenol blue and measured by means of the Spinco Analytrol instrument. For detection of dTTP : a-D-gala&se-l-P thymidylyltransferase on these strips, it was necessary to use the long incubation period (72 hours) since apparently some inactivation of this transferase had occurred during electrophoresis. After electrophoresis for 18 hours, the UTP : a-D-glucose-l-P uridylyltransferase and dTTP : ar-D-glucose-l-P thymidylyltransferase were found at a distance of 1.5 cm from the origin. The two activities were distributed in an identical manner along the paper strip. The dTTP:cr-D- galactose-1-P thymidylyltransferase activity could not be de- tected on the paper strips after electrophoresis for 18 hours.
DISCUSSION
It can be seen in Table I that the enzyme preparation from S. faecalis grown on D-glucose contained the transferases re- sponsible for the synthesis of uridine diphosphate glucose, uridine diphosphate gala&se, and thymidine diphosphate glucose. The enzyme preparation from cells grown on D-galactose con- tained, in addition to the above, a thymidylyltransferase re-
-3-2-l I2 3456 789
FIG. 2. The distribution of the transferase activities and pro- tein components in extracts from S. jaecalis on paper electro- phoretic strips. A, UTP:a-D-glucose-l-P uridylyltransferase; B, dTTP:ar-D-glucose-l-P thymidylyltransferase; D, dTTP:a-D- galactose-1-P thymidylyltransferase.
sponsible for the synthesis of thymidine diphosphate galactose from thymidine triphosphate and ar-D-galactose-1-P. The thymidine diphosphate gala&se which was produced by this enzyme was converted rapidly to thymidine diphosphate glucose and thymidine diphosphate rhamnose by the multienzyme system in the S. fuecalis (4, 17, 18). The dTTP:cY-D-galactose-1-P thymidylyltransferase has not been detected in enzyme prepara- tions from cells grown on other carbohydrate sources such as D-glucose, n-rhamnose, or D-arabinose. Its formation in S. faecalis is apparently induced by some unique structural features of D-gala&se.
The results in Table I and Figs. 1 and 2 establish the separate existence of a dTTP : cY-D-galactose-1-P thymidylyltransferase. This enzyme diiers from the dTTP:cr-D-glucose-l-P thymidylyl- transferase and from the uridylyltransferases in its stability on storage and in its mobility on paper electrophoresis. The en- zyme is very sensitive to changes in pH and exhibits a sharp pH optimum curve with a maximum of pH 7. The molecular size of dTTP : ru-D-gala&se-l-P thymidylyltransferase as estimated by the density gradient centrifugation method (2, 3) is in the range of 80,000 to 100,000. The molecular sizes of the other thymidylyltransferases and the uridylyltransferases of S. faecalis are also in this range. Although prolonged electrophoresis of the dTTP :a-D-galactose-1-P thymidylyltransferase inactivated the enzyme, it may be possible to utilize short term electro- phoresis for the preparation of this enzyme free of the other thymidylyl- and uridylyltransferases in S. faecalis extracts. Such an enzyme preparation would be valuable for a variety of studies, including the synthesis of pure thymidine diphosphate gala&se, and studies on the mechanism of the conversion of D-galactose to D-rhamnose.
The earlier suggestion (1) that the dTTP : cY-D-galactose-1-P thymidylyltransferase was probably required for the synthesis of cell wall constituents which could arise from intact D-galactose via the thymidine pathway has been substantiated by the results of isotope experiments. ~-Glucose, D-gala&se, and L-rhamnose isolated from the S. fueculis cells grown on D-galactose-1-C” were labeled exclusively in position 1. The possibility of a conversion of D-gala&se to L-rhamnose by a pathway involving
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

3160 dTTP: a-D-Galactose-1 -P Thymidylyltransferase Vol. 238, No. 10
both uridine and thymidine nucleotides is unlikely since a direct vated on storage at 3”. The enzyme is probably utilized by the interconversion of these nucleotides was not demonstrable. As organism to synthesize cellular constituents which can arise it is well established that in bacteria L-rhamnose is synthesized only from the intact hesose units via the thymidine diphosphate from n-alucose via the thvmidine dinhosnhate hexose pathway hexose pathway. (4, 17, I$, it is most likely that in S.&c&s adapted to-n-gala;- tose, the L-rhamnose is also produced by this pathway. The dTTP : cr-n-galactose-1-P thymidylyltransferase is required to initiate the series of reactions resulting in the synthesis of thy- midine diphosphate galactose and eventually thymidine diphos- phate rhamnose.
REFEREh’CES
1. PAZUR, J. H., KLEPPE, K., AND CEPCRE, A., Biochem. and
2. Biophys. Research C&m&s., 7, 157 (19G2):
PAZ~R. J. H.. KLEPPE. K.. ANI) ANDERSON. J. S.. Biochim.
3. et Btophys. &a, 66, ‘369 i1962).
M.~RTI~V, R. G., AND AMES, B. K., J. Biol. Chem., 236, 1372 (1961). n-Clucosamine isolated from the S. juecalis cells grown on
n-galactose-l-Cl4 was labeled exclusively in position 1. This observation is indicative of a nucleotide pathway for the syn- thesis of n-glucosamine. Since S. jcu~aZis possesses the enzymes required for the synthesis of both uridine- and thymidine diphos- phate hexoses, the glucosamine may arrive by either or both pathways. The uridylyltransferase (19, 20) which effects the synthesis of uridine diphosphate-N-acetyl-n-glucosamine from uridine triphosphate and N-acetyl-n-glucosamine-1-P was found to be present in the extracts from S. jaecalis. Although the transfer of N-acetyl-n-glucosamine to cell wall material from uridine diphosphate N-acetyl-n-glucosamine has not yet been demonstrated in S. juec&s, such a transfer does occur with other bacteria (21) and also might be expected in S. jaecalis. A recent report implicates the thymidine diphosphate compounds as possible intermediates of synthesis of hexosamines in some species of bacteria (16) and the possibility of synthesis of D-&I-
cosamine in S. jaecalis via the thgmidine pathway cannot be ruled out at this time.
SUMMARY
Streptococcus jaeculis grown on n-galactose produces a thy- midine triphosphate : a-n-galactose l-phosphate thymidylyl- transferase which effects the synthesis of thymidine diphosphate galactose from thymidine triphosphate and cY-n-galactose l-phos- phate. The enzyme can be separated from the other thymidylyl- and uridylyltransferases of Streptococcus jaeculti by paper electrophoresis. It possesses a pH optimum of 7 and is inacti-
4. P~ZGR, J. H., .~XD SHCEY. E. W.. J. Biol. Chem.. 236. 1780
5.
6. 7. 8.
9.
10.
11.
12. 13.
14.
15.
16.
17.
18.
19.
20.
21.
(196i). LOWRY, 0. H., ROSEBROUGH, X. J., FARR, A. L., AND RANDAI,L,
R. J., J. Biol. Chem., 193, 265 (1951). PALADINI, A. C., .~ND LELOIR, L. F., Biochem. J., 61,426 (1952). WHITE, L: M., .~ND SECOR, G. E., Science, 126, 495 (195?). DISCHE, Z., AND SHETTLES, L. B., J. Biol. Chem., 1’76, 595
(1948). AVIGAD, G., AM.IR.IL, D., ASENSIO, C., ANI) HORECKER, B.
L., J. Biol. Chem., 237, 2736 (1962). NEUFELD, E. F., Biochem. and Biophys. Research Communs.,
7, 461 (1962). BLOCK, R. J., DURRUM, E. L., AND ZWEIG, G., Paper chroma-
tography and paper electrophoresis, Ed. 2, Academic Press, Inc., New York, 1958, p. 181.
PARTRIDGE, T. M., Mature, 164, 443 (1949). BARKER, R., ANI) MACDONALD, 1). L., J. ilm. Chem. Sot., 82,
2297 (1960). CUMMINS, C. S., AXD HARRIS, H., J. Gen. Microbial., 14, 583
(1956). STOFFYN, P. J., AND JEANLOZ. R. W., Arch. Biochem. Biophys.,
62, 37i (1954). . -
KORNFELD, S., .~ND GLASER, L., J. Biol. Chem., 237, 3052 (1962).
G~AsE~, L., AND KORNFELD, S., J. Biol. Chem., 236, 1795 (1961).
OKAZ~KI, R., OKAZ.IKI, T., STROMINGER, J. L., AND MICHEL- SON, A. M., J. Biol. Chem., 237, 3014 (1962).
SMITH, E. E. B., AND MILLS, G. T., Biochim. et Biophys. Acta, 13, 386 (1954).
STROMINGER, J. L., AND SMITH, M. S., J. Biol. Chem., 234, 1822 (1959).
NATHENSON, S. G., AND STROMINGER, J. L., J. Biol. Chem., 237, 3839 (1962).
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

John H. Pazur and John S. Anderson Grown on d-GalactoseStreptococcus faecalisfrom
-d-Galactose 1-Phosphate ThymidylyltransferaseαThymidine Triphosphate:
1963, 238:3155-3160.J. Biol. Chem.
http://www.jbc.org/content/238/10/3155.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/238/10/3155.citation.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from