untersuchungen über die pathogenese der aleutenkrankheit der nerze viii. depression der...
TRANSCRIPT

Zbl. Vet. Med. B, 21, 467-479 (1974) @ 1974 Verlag Paul Parey, Berlin und Hamburg ISSN 0044-4294/ASTM-Coden: ZVRBAZ
Aus dem Institut fiir Pathologie der Tierarztlichen Hochschule Hannover Direktor: Prof. Dr. L.-Cl. Schulz
Abteilung fur Imrnunpathologie Leiter: Pro f . Dr. G. Trautwein
Untersuchungen iiber die Pathogenese der Aleutenkrankheit der Nerze
VIII. Depression der Antikorperbildung gegen Meerretticfi-Peroxydase
Von
G. TRAUTWEIN, P. SCHNEIDER und E. ERNST
Mit 8 Abbildungen und einer Tabelle
(Eingegangen am 25. Februar 1974)
Zu den friihesten Befunden bei der Aleutenkrankheit (A. K.) der Nerze gehort die Beobachtung, dai3 aleutenkranke homozygote Aleutennerze eine besondere Anfalligkeit gegeniiber bakteriellen Infektionen aufweisen. So konnte gezeigt werden, dai3 bei bestehender spontaner Aleutenkrankheit nach Sekundarinfektionen mit pyogenen Bakterien und Mykobakterien die Morta- litat ansteigt (HARTSOUGH u. GORHAM, 1956; HELMBOLDT u. JUNGHERR, 1958; PADGETT u. Mitarb., 1968). Die Ergebnisse experimenteller Unter- suchungen weisen darauf hin, dai3 es offenbar in bestimmten Phasen der A. K. zu einer Depression der Antikorperbildung gegen bestimmte Antigene wie Brucella-Antigen, Hamocyanin und bovines Gammaglobulin kommt (KENYON, 1966; LODMELL u. Mitarb., 1970; PORTER u. Mitarb., 1965).
In der vorliegenden experimentellen Studie wurde als Zweitantigen Meerrettich-Peroxydase eingesetzt und die Primarreaktion von Versuchsnerzen in verschiedenen Phasen der experimentellen A. K. naher untersucht. Die Verwendung dieses Antigens bietet den Vorteil, dai3 neben der serologischen Bestimmung des Antikorpertiters die intrazellulare Antikorperbildung mit einer kombinierten immunologisch-enzymhistochemischen Methode (AVRA- MEAS u. LEDUC, 1970) sichtbar gemacht werden kann. Mit Hilfe dieses Ver- fahrens liei3 sich eine Depression der Antikorperbildung gegen das Antigen Meerrettich-Peroxydose in bestimmten Stadien der Aleutenkrankheit sowohl serologisch als auch zytologisch-histologisch nachweisen.
*) Aus dem Sonderforschungsbereich 54 ,,Rheumatoide Krankheiten des Tieres".

468 G. TRAUTWEIN, P. SCHNEIDER und E. ERNST
Material und Methode Infektion rnit A. K.-Virus. Insgesamt 24, zwolf Monate alte, weibliche
und mannliche Saphir-Nerze wurden intraperitoneal mit jeweils 2 ml Organ- suspension des A. K.-Virusstammes SK/66, 6. Nerzpassage, gemag voraus- gehenden Versuchen (TRAUTWEIN, 1969, 1972) infiziert. Gruppen von jeweils 4 Nerzen wurden nach vorausgehender Peroxydase-Injektion 4, 8, 12, 16, 20 und 24 Wochen post infectionem getotet und serologisch, zytologisch, histolo- gisch sowie in Einzelfallen elektronenmikroskopisch untersucht.
Kontrollen. Insgesamt 12 Nerze wurden nicht rnit dem A. K.-Virus infi- ziert und erhielten nur Injektionen von Meerrettich-Peroxydase. Jeweils 2 Kontrollnerze wurden zum gleichen Zeitpunkt wie die A. K.-infizierten Nerze getotet.
Znjektion v o n Meerrettich-Peroxydase. 10 mg Meerrettich-Peroxydase (Nr. 15302 EPAB, Reinheitsgrad 11, lyophilisiert, Fa. C. F. Boehringer, Mann- heim) wurden in 1,0 ml physiolog. NaCl gelost und in 1,0 ml komplettem Freundschem Adjuvans emulgiert. Die Emulsion wurde in beide Fugballen der Hinterextremitaten eines Nerzes injiziert. Diese Injektionen wurden sowohl bei den rnit A. K.-Virus infizierten Nerzen als auch bei Kontrollnerzen ein- malig 24 Tage vor dem Totungstermin gegeben.
Serumuntersuchungen a ) Elektrophorese. Die y-Globulinspiegel im Blutserum wurden in 4wo-
chigen Abstanden sowie bei der Totung papierelektrophoretisch bestimmt (TRAUTWEIN, 1969).
b) Anti-Peroxydase-Antikorper, Die Antiperoxydastiter im Serum wur- den mit der Methode der passiven Hamagglutination nach AVRAMEAS und Mitarbeitern (1969) bestimmt. Hierzu wurde Peroxydase in Phosphatpuffer 1 : 1 (Gew./Vol.) gelost und mittels 2,5 O/oigem Glutaraldehyd an gewaschene Ziegenerythrozyten gebunden. Mit den beladenen Erythrozyten wurde die passive Hamagglutination in Mikrotiterplatten durchgefuhrt. Als Kontrollen fanden nichtsensibilisierte Erythrozyten und Anti-Peroxydaseserum bzw. Nor- malserum Verwendung.
c) Charakterisierung des Anti-Peroxydase-Antikorpers durch Gelfiltra- tion. Jeweils 2 ml eines 1 : 2 rnit Tris-HC1-Puffer, pH 8,O verdunnten Nerz- vollserums wurden uber Sephadex G 200 filtriert und die entstehenden 3 Hauptfraktionen mittels Fraktionssammler gesammelt. Nach Ankonzentrie- ren wurde in den Einzelfraktionen ebenfalls der Antiperoxydastiter bestimmt.
d ) Charakterisierung des Anti-Peroxydase-Antikorpers durch Immun- elektrophorese. Nerzseren wurden in 1,5 O/oiger Agarose elektrophoretisch aufgetrennt und die Prazipitate rnit polyvalentem Anti-Nerzserum von Ka- ninchen entwickelt (TRAUTWEIN, 1969). S ezifischer Antikorper wurde im
tischen Praparate mit Peroxydase (0,l mg Peroxydase/ml) nach den Angaben von AVRAMEAS und LEDUC (1970) nachgewiesen. Nach grundlichem Aus- waschen der Praparate wurden die an Immunglobulinfraktionen gebundene Peroxydase mit der gleichen Nachweismethode wie fur Zellen dargestellt.
Enzymzytochemie. Zum Nachweis von spezifischem Antikorper in lym- phatischen Zellen wurden unmittelbar nach der Totung der Nerze die Lnn. poplitei, inquinales und iliacales entnommen. Von der Schnittflache jeweils einer Lymphknotenhalfte wurden Tupfpraparate auf entfetteten Objekttragern angefertigt. Die zweite Halfte wurde in Isopentan und flussigem Stickstoff eingefroren und fur die Herstellung von Cryostatschnitten verwandt. Nach
Bereich der IgG- bzw. IgM-Linie durch In K ubation der immunelektrophore-

Untersuchungen iiber die Pathogenese der Aleutenkrankheit der Nerze 469
Trocknen der Tupfpraparate und Cryostatschnitten und Fixieren in Alkohol- Ather (60 : 40) wurden die Praparate 30 Min. in Peroxydaselosung (0,l mg Peroxydase/ml physiol. NaCl) inkubiert (AVRAMEAS u. LEDUC, 1970). Die Bin- dung des Enzyms an Peroxydase-Antikorper bildende Zellen wurde enzym- zytochemisch nachgewiesen (BLOOM u. Mitarb., 1970). Das Inkubationsmedium enthielt 49 ml 0,2 M Trispuffer, pH 7,6, 25 mg Diaminobenzidintetrachlorid (Fa. E. Merck) und 1 ml frisches 1 O/oiges H202. Nach Inkubation fur 15 Min. wurden die Praparate in aqua dest. gespult und mit Hamalaun gefarbt. Bei den Kontrollpraparaten wurde die Enzymreaktion ohne vorausgehende Inku- bation mit Peroxydase durchgefuhrt.
Elektronenmikroskopie. Die elektronenmikroskopische Darstellung Anti- Peroxydase bildender Zellen erfolgte entsprechend den Angaben von LEDUC und Mitarbeitern (1968; 1969). Nach Fixierung der Lymphknotenstucke in 1 O/oigem Formalin und weiterer Zerkleinerung wurde in Peroxydaselosung (1 mg/ml Puffer) 1 Std. lang bei Zimmertemperatur inkubiert. Daran schloi3 sich eine kurze Nachfixierung in 2,5 O/oigem Glutaraldehyd an. Die nach- folgende histochemische Reaktion zum Nachweis der an Zellen gebundenen Peroxydase erfolgte wie fur die Lichtmikroskopie. Nach einer Nachfixierung in 2 O/oigem OsO, wurden die Gewebsstucke nach Standardmethoden in Epon eingebettet.
Ergebnisse Makroskopische Befunde
Lymphknoten: Sowohl bei aleutenkranken als auch bei Kontrollnerzen finden sich 24 Tage nach Injektion von Peroxydase und Freundschem Adjuvans in die Fui3ballen der Hinterextremitaten mittel- bis hochgradige Vergroi3e- rungen der Lnn. poplitei, inquinales und ilici mediales.
Milz: Die Milz zeigt bei Kontrolltieren und aleutenkranken Nerzen das Bild einer mittel- bis hochgradigen hyperplastischen Schwellung, wobei die Milzfollikel deutlich hervortreten.
Lunge: Bei etwa der Halfte der Kontroll- und A. K.-Nerze sind makrosk- opisch miliare, subpleural und in der Tiefe des Lungenparenchyms gelegene grauweif3e bis gelblichweii3e Herde erkennbar.
Ubrige Organe a) Kontrollnerze: Neben Schwellung der Milz und der regionaren
Lymphknoten der Hinterextremitaten erweisen sich alle ubrigen Organe makroskopisch als frei von pathologischen Veranderungen.
b) Aleutenkranke Nerze: Die Intensitat der Veranderungen in Korper- und Organlymphknoten, Leber, Niere und gelegentlich im Magen-Darmtrakt steht in deutlicher Abhangigkeit von der Krankheitsdauer. Es lassen sich die bereits naher beschriebenen Stadien der Krankheit erkennen (TRAUTWEIN, 1970).
Histologische Befunde Lymphknoten: Bei allen Nerzen finden sich in den Lnn. poplitei, inquina-
les und ilici deutliche Veranderungen, welche einmal auf die Wirkung der Peroxydase, zum anderen auf das komplette Freundsche Adjuvans zu beziehen sind. Die Follikel sind durch Bildun von groaen, mitosereichen Keimzentren
und inquinales stark verschmalert und enthalten reife Plasmazellen und Plas- mazellvorstufen (Abb. 3). In den oft hochgradig erweiterten Intermediar- und
stark vergroi3ert (Abb. 1). Die Mar a strange erscheinen in den Lnn. poplitei
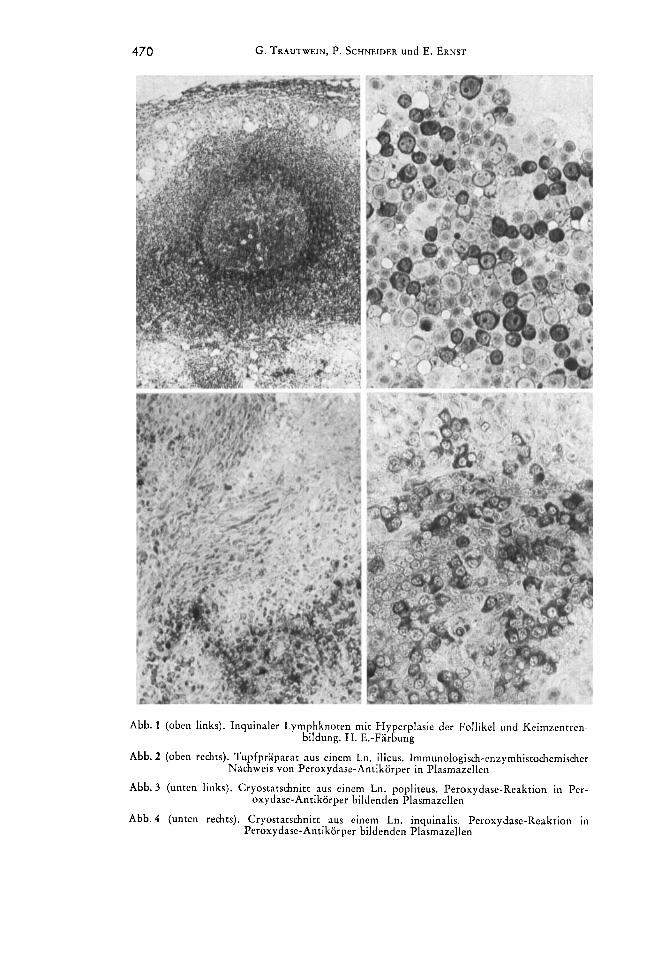
470 G. TRAUTWEIN, P. SCHNEIDER und E. ERNST
Abb. 1 (oben links). lnquinaler Lymphknoten mit Hyperplasie der Follikel und Keim7entren- bildung. H. E.-Farbung
Abb. 2 (oben rechts). Tupfpraparat aus einem Ln. ilicus. Immunologisch-enzymhistochcmischer Nachweis von Peroxydase-Antikorper in Plasmazellen
Abb. 3 (unten links). Cryostatschnitt aus einem Ln. popliteus. Peroxydase-Reaktion in Per- oxydase-Antikorper bildenden Plasmazellen
Abb. 4 (unten rechts). Cryostatschnitt aus einem Ln. inquinalis. Peroxydase-Reaktion in Peroxydase-Antikorper bildenden Plasmazellen

Untersuchungen iiber die Pathogenese der Aleutenkrankheit der Nerze 471
Versuc hs-
9ruPPe
A. K. I A. K. 11 A. K. 111 A. K. I V A. K. V A. K . V I Kontrolle
Randsinus sind lipidhaltige Makrophagen, Epitheloidzellen und Fibroblasten enthalten (Abb. 3). Aui3erdem finden sich groi3e Fettvakuolen, die stellenweise von neutrophilen Granulozyten umsaumt werden. In den Lnn. ilici mediales tritt die Ausfiillung der Sinus mit den beschriebenen Zellen zuriick. Demgegen- uber ist hier die Verbreiterung der Markstrange durch Plasmazellen deutlicher.
Zytologie und Irnrnunhistochernie des Lyrnphknotens: Bei dem kombi- nierten Verfahren zum direkten Nachweis von Peroxydase-Antikorper in Plasmazellen und lymphatischen Zellen wird zunachst Meerrettich-Peroxydase spezifisch an den Antikorper in antikorperbildenden Zellen gebunden (AVRAMEAS u. LEDUC, 1970; LEDUC u. Mitarb., 1968). Hierbei bleibt das Enzym selbst aktiv und kann nach Einwirkung auf das Substrat der Zellen nachgewiesen werden. Die Farbintensitat als Mai3 fur die Reaktionsstarke variiert zwischen hellbraun und dunkelbraun.
Wochen post infection
Ln. inquinalis Ln. ilicus
4 20,8 t 9,76 25,7 ? 17,90
12 32,9 2 15,13 31,l t 3,7L 8 25,4 t 13,96 26,l ? 8,SO
16 19,5 t 10,48 8,1 t 3,66 20 1,8 t 1,oo 4 5 t 4/45 24 4,2 2 3,24 13,O t 8,36
19,4 t 8,16 23,9 t 6,17
In Tupfpraparaten von Lymphknoten der Hinterextremitaten lassen sich antikorperbildende Zellen als Plasmoblasten, Proplasmozyten und reife Plas- mazellen bzw. lymphozytare Zellen differenzieren (Abb. 2 ) . Innerhalb der Plasmazellreihe weisen die reifen Plasmazellen die starkste Reaktion auf.
Bei der quantitativen Bestimmung antikorperbildender Zellen wurden alle Plasmazellen und Plasmazellvorstufen sowie lymphoide Zellen mit positi-
Zelkahl (O/O) 0 Ln. inquinalis
Kontr, 4 8 12 16 20 24 Wochen post infect,
Abb. 5. Zahl der peroxydase-psitiven Lymphozyten in zwei regionaren Lymphknoten (Ln. inquinalis, Ln. ilicus)

472 G. TRAUTWEIN, P. SCHNEIDER und E. ERNST
ver Peroxydase-Reaktion innerhalb einer Lymphknotenzellpopulation von 1.000 Zellen ausgezahlt. Das Ergebnis ist in Tabelle 1 und Abbildung 5 dar- gestellt. Es zeigt sich, dai3 die Zahl der Peroxydase-Antikorper bildenden Lymphknotenzellen 20 Wochen post infectionem auf den tiefsten Wert ab- sinkt. Danach deutet sich wieder ein leichter Anstieg antikorperbildender Zellen an.
Kryostatschnitte von Lymphknoten und immunologisch-enzymhistoche- mischer Nachweis von Peroxydase-Antikorper ermoglichen eine prazise Loka- lisation antikorperbildender Zellen im Lymphknoten (Abb. 3, 4). Hierbei sind antikorperhaltige Plasmazellen nur in geringer Zahl in den Markstrangen der Lnn. poplitei, jedoch reichlich in Markstrangen der Lnn. inquinales und ilici anzutreff en. Analog zu den Ergebnissen der quantitativen Bestimmung erscheint auch im Schnitt raparat 20 und 24 Wochen nach Infektion mit
besonderem Interesse ist, da13 die Keimzentren der Rindenfollikel inkonstant antikorperhaltige lymphoide Zellen mit schmalem, peroxydase-positivem Cy- toplasmasaum enthalten. Diese Zellelemente durften den in Lymphknoten- tupfpraparaten nachweisbaren, aus dem Zellverband herausgelosten lympha- tischen Zellen mit positiver Peroxydase-Reaktion entsprechen. Neben sicher intrazellularem Peroxydase-Antikorper kommt geringgradig auch Antikorper in interzellularer Lokalisation zur Beobachtung.
Elektronenmikroskopisch 1a13t sicht mit der Methode von LEDUC und Mit- arbeitern (1968, 1969) Peroxydase-Antikorper spezifisch in Plasmoblasten, Proplasmozyten und reifen Plasmazellen als dunkelgraues, z. T. schwarzes Reaktionsprodukt darstellen. Auf dem Hohepunkt der Antikorpersynthese kommt es in den reifen Plasmazellen zu deutlicher Ausweitung des endoplas- matischen Reticulum infolge Anreicherung von Peroxydase-Antikorper (Abb. 6). Haufig finden sich Plasmazellen rnit cysternenartiger Dilatation des endoplasmatischen Reticulum und kugeligen peroxydase-positiven Kondensa- ten, welche Russelschen Korperchen entsprechen.
Lunge: Die makroskopisch erkennbaren miliaren Herde erweisen sich bei der histologischen Untersuchung als Alveolenbezirke, die lipidhaltige Alveo-
dem A. K.-Virus die Zah f peroxydase-positiver Plasmazellen reduziert. Von
Abb. 6 . Plasmazelle aus einem Ln. inquinalis. Ausweitung des endoplasmatischen Reticulum
Untersuchungen iiber die Pathogenese der Aleutenkrankheit der Nerze 473
larmakrophagen enthalten. In diesen Bezirken sind die Alveolarsepten durch Infiltration mit Lymphozyten und Plasmazellen verbreitert.
Ubrige Organe: Die durch das Virus der Aleutenkrankheit induzierten histologischen Organveranderungen entsprechen den Organveranderungen, wie sie bereits in einer fruheren pathogenetischen Studie beschrieben wurden ( TRAUTWEIN, 1970).
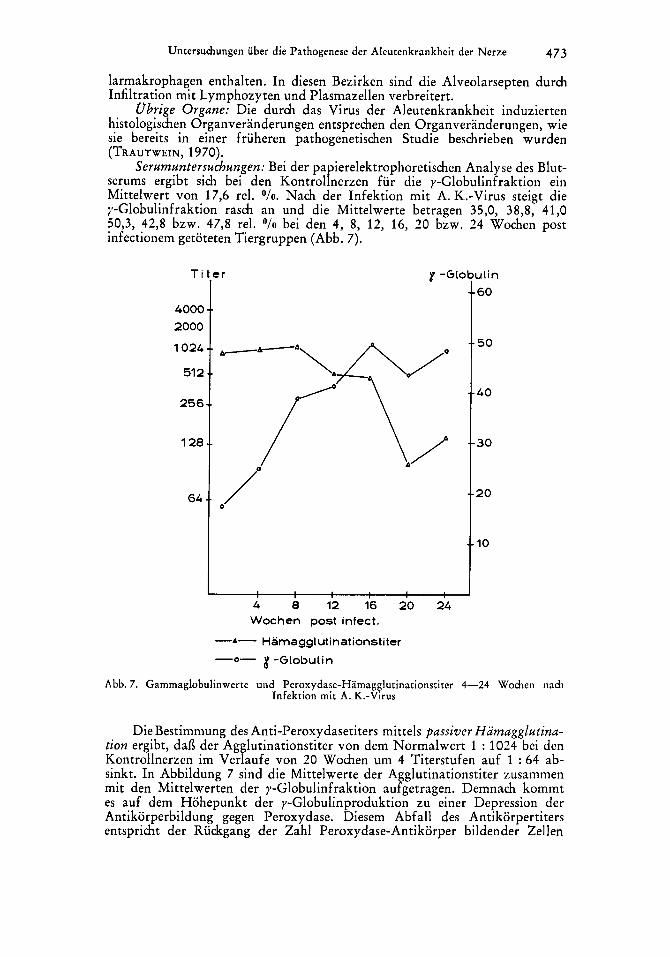
Serumuntersuchungen: Bei der papierelektrophoretischen Analyse des Blut- serums ergibt sich bei den Kontrollnerzen fur die y-Globulinfraktion ein Mittelwert von 17,6 rel. O / o . Nach der Infektion mit A. K.-Virus steigt die y-Globulinfraktion rasch an und die Mittelwerte betragen 35,0, 38,8, 41,O 50,3, 42,8 bzw. 47,8 rel. O / o bei den 4, 8, 12, 16, 20 bzw. 24 Wochen post infectionem getoteten Tiergruppen (Abb. 7).
T i
400C 2000
1024
51 2
256
1 28
64
er -GI0 ulin 60
50
40
30
20
10
4 8 12 16 20 24 Wochen post infect.
-1- Hamagglutinationstiter
0- 8 -Globulin - Abb. 7. Gammaglobulinwerte und Peroxydase-Hamagglutinationstiter 4-24 Wochen nach
Infektion rnit A. K.-Virus
Die Bestimmung des Anti-Peroxydasetiters mittels parsiver Hamagglutina- tion ergibt, daf3 der Agglutinationstiter von dem Normalwert 1 : 1024 bei den Kontrollnerzen im Verlaufe von 20 Wochen um 4 Titerstufen auf 1 : 64 ab- sinkt. In Abbildung 7 sind die Mittelwerte der Agglutinationstiter zusammen mit den Mittelwerten der y-Globulinfraktion aufgetragen. Demnach kommt es auf dem Hohepunkt der y-Globulinproduktion zu einer Depression der Antikorperbildung gegen Peroxydase. Diesem Abfall des Antikorpertiters entspricht der Ruckgang der Zahl Peroxydase-Antikorper bildender Zellen
474 G. TRAUTWEIN, P. SCHNEIDER und E. ERNST
in den Lymphknoten, wobei die tiefsten Werte bei Nerzen 20 Wochen nach Infektion mit dem A. K.-Virus gefunden werden.
Mittels Gelfiltration an Sephadex G 200 wurde das Vollserum von je- weils einem Nerz aus den insgesamt 6 Versuchsgruppen in die IgM- und die IgG-haltige Fraktion aufgetrennt. Die Bestimmung der Peroxydase-Hamag- glutinationstiter ergibt fur die IgG-Fraktion die gleichen Titerwerte wie fur Vollserum. In der IgM-Fraktion liegen die Titer aller untersuchten Seren urn durchschnittlich 4 Titerstufen niederer als in der IgG-Fraktion.
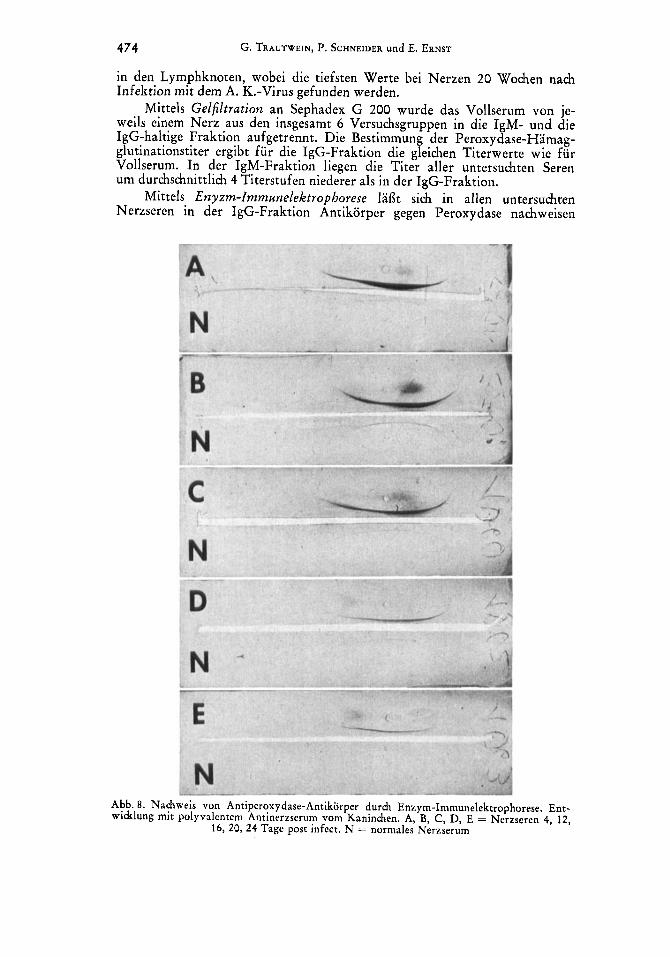
Mittels Enyzm-lmmunelektrophorese 1af3t sich in allen untersuchten Nerzseren in der IgG-Fraktion Antikorper gegen Peroxydase nachweisen
Abb. 8. Nachweis von Antiperoxydase-Antikorper durch Enzym-Immunelektrophorese. Ent- widdung mit polyvalentem Antinerzserum vom Kaninchen. A, B, C, D, E = Nerzseren 4, 12,
16, 20, 24 Tage post infect. N = normales Nerzserum

Untersuchungen uber die Pathogenese der Aleutenkrankheit der Nerze 475
(Abb. 8). Die IgG-Linie ist zumeist nicht einheitlich, sondern laflt unter- schiedlich starke Anteile einer schneller wandernden Igl- und einer langsamer wandernden Ig2-Fraktion erkennen. Unterschiedlich stark ist auch die Kon- zentration des Antiperoxydase-Antikorpers, was sich in der Lange der IgG- Linie und dem Grad des Antigenschleiers ausdruckt (Abb. 8). 20 und 24 Wochen post infectionem ist nur eine schmale, relativ kurze IgG-Linie nach- weisbar. Hinweise auf Antikorper in der IgM-Fraktion ergeben sich in Einzel- fallen im Bereich eines kathodenseitig und weit entfernt von der Antiserum- rinne gelegenen sog. ,,Depotfleckes".
Besprechung der Ergebnisse Das in den eigenen Untersuchungen gewahlte Antigen Meerrettich-Peroxy-
dase bietet grofle Vorteile gegenuber anderen, bislang beim Nerz gepriiften Antigenen. Mit dem kombinierten immunologisch-histochemischen Verfahren nach AVRAMEAS und LEDUC (1970) laflt sich neben dem serologischen Nach- weis des Peroxydase-Antikorpers die intrazellulare Bildung des Antikorpers in Lymphknoten sichtbar machen. Da bei der im ersten Schritt der Methode erfolgenden immunologischen Bindung der Peroxydase an antikorperbildende Zellen 98-100 O / o der Enzymaktivitat erhalten bleiben (HESS und SORDAT, 1970), konnen bei der nachfolgenden enzymhistochemischen Reaktion auch geringe Mengen von Peroxydase-Antikorper intrazellular nachgewiesen wer- den. Nach Auszahlung peroxydase-positiver Zellen laflt sich die Zahl antikor- perbildender Zellen mit dem Antikorpertiter des Serums in Beziehung setzen.
Nach dem Ergebnis der eigenen experimentellen Untersuchungen lai3t sich in bestimmten Stadien der Aleutenkrankheit eine Depression der Antikorper- bildung gegen das Modellantigen Meerrettich-Peroxydase nachweisen. Hierbei werden 20 Wochen nach experimenteller Infektion mit dem A. K.-Virus die niedersten Titer erreicht. Zum gleichen Zeitpunkt sinkt die Zahl der Anti- korper gegen Peroxydase bildende Lymphknotenzellen auf den tiefsten Wert ab.
Bei der von uns gewahlten Versuchsanordnung wurden sowohl die mit A. K.-Virus infizierten als auch die Kontrollnerze jeweils 24 Tage nach ein- maliger Stimulatin mit Peroxydase ausgewertet. Zu diesem Zeitpunkt der Primarreaktion lassen sich cytologisch in den regionaren Lymphknoten zwei Zellfamilien diff erenzieren. Zunachst sind dies der Plasmazellreihe zugehorige Zellen wie Plasmoblasten, Proplasmozyten und die typischen reifen Plasma- zellen. Der Antikorpergehalt scheint in diesen Zellen in der gleichen Reihen- folge zuzunehmen. Von dieser Zellgruppe sind klar abzugrenzen mittelgrofle lymphozytare Zellen mit dichtem Chromatingefiige und schmalem Cytoplas- masaum, der deutliche Peroxydase-Reaktion gibt und demnach spezifischen Antikorper enthalt. Dieser Zelltyp konnte den ,,Lymphoplasmozyten" ent- sprechen, die AVRAMEAS und LEDUC (1970) bei Mausen nach Stimulation mit Peroxydase iiberwiegend in den Sinus fanden; er laflt sich aber gleichfalls auch als Germinoblast der Keimzentren deuten.
Nach den elektronenmikroskopischen Untersuchungen von AVRAMEAS und LEDUC (1970) entwickeln sich sowohl die Plasmazellen wie auch die ,,Lymphoplasmazellen" iiber jeweils zwei Zwischenformen aus einer gemein- samen Stammzelle, dem Hamozytoblast; denn in dieser Zelle laflt sich wenige Tage nach Peroxydase-Injektion erstmalig Antikorper im perinuklearen Raum nachweisen. Von hier aus breitet sich bei beiden Zelltypen die Antikorper- bildung im Kanal- und Cysternensystem des rauhen endoplasmatischen Reti- culum aus.
Zbl. Vet. Med., Reihe B, Bd. 21, Heft 6 33

476 G. TRAUTWEIN, P. SCHNEIDER und E. ERNST
Topographisch kommen im Lymphknoten Peroxydase-Antikorper ent- haltende Zellen in zwei Hauptlokalisationen vor. Die Hauptmenge der Zelleri der Plasmazellreihe ist in den entsprechend dem jeweiligen Stadium der A. K. mai3ig oder stark verbreiterten Markstrangen anzutreff en, wahrend nur wenige Plasmazellen verstreut zwischen den Rindenfollikeln zu finden sind. Die zweite Gruppe Antikorper produzierender Zellen kommt als lympha- tische Zelle in einzelnen Keimzentren der Rindenfollikeln vor. Diese Beobach- tung stimmt gut mit Befunden von SORDAT und Mitarbeitern (1970) uberein, die bei der Primarreaktion der Maus zuerst auf das Vorkommen von Peroxy- dase-Antikorper in lymphoiden Zellen der Keimzentren hingewiesen haben. Spezifischer Antikorper wurde intrazellular und zwischen dentritischen reti- kularen Zellen 17-35 Tage na& einmaliger Stimulation mit Peroxydase ge- funden. Hierbei liei3en sich elektronenmikroskopisch Germinoblasten bzw. Germinozyten und eine zweite lymphoide Zelle mit spezifischem Antikorper in Cysternen des ER unterscheiden.
Bei den eigenen serologischen, elektrophoretischen und quantitativ-cytolo- gischen Befunden ist auffallig, dai3 es auf dem Hohepunkt der Gammaglobu- linproduktion zu einer Depression der Antikorperbildung gegen Peroxydase kommt. Diesem Abfall des spezifischen Antikorpertiters entspricht der Ruck- gang der Zahl Peroxydase-Antikorper bildender Zellen in den Lymphknoten. Diese Befunde lassen sich in der Weise deuten, dai3 auf dem Hohepunkt der Antikorperbildung gegen A. K.-Virus 4-5 Monate post infectionem eine Phase herabgesetzter Antikorperbildung gegen andere Antigene einsetzt. Hier- bei wurde uber einen nicht bekannten Mechanismus zu wenig Plasmazellklone fur die Bildung von Peroxydase-Antikorper zur Verfugung stehen.
In friiheren Untersuchungen konnte gezeigt werden, dai3 eine Korrelation zwischen der Zahl der in lymphatischen Organen proliferierten Plasmazellen und der Hohe des Gammaglobulinspiegels besteht (TRAUTWEIN, 1964). Bis vor kurzem war aber unklar, ob dem exzessiv gebildeten Gammaglobulin Antikorperspezifitat gegenuber dem A. K.-Virus zukommt. MCGUIRE und Mitarbeiter (1971) fanden bei aleutenkranken Nerzen hohe Titer komplement- bindender Antikorper gegen das A. K.-Virus. Weiterhin wurde mittels Immu- nodiff usion und Immunelektroosmophorese Antikorper gegen A. K.-Virus nachgewiesen (CHO u. INGRAM, 1973 a; 1973 b). Demnach darf als gesichert gelten, dai3 ein hoher Prozentsatz des bei A. K. gebildeten Gammaglobulins Antikorper gegen A. K.-Virus ist.
Von vergleichendem Interesse sind ahnliche Untersuchungen anderer Autoren. KENYON (1966) immunisierte Nerze in verschiedenen Phasen der A. K. rnit Brucella-Antigen und fand eine deutliche Immundepression 3 Mo- nate post infectionem. Wurden die Versuchsnerze erst nach der Brucellen- Immunisierung rnit A. K.-Virus infiziert, entwickelten sich ahnlich hohe Agglu- tinationstiter wie bei normalen Kontrollnerzen. LODMELL und Mitarbeiter (1970) konnten rnit Hamocyanin a h Antigen ebenfalls zeigen, dai3 es bei aleutenkranken Nerzen zur Depression der Antikorperbildung bei der Pri- marreaktion kommt, hingegen bei Boosterreaktionen gute Immunreaktionen auftreten. Weiterhin fanden PORTER und Mitarbeiter (1965) bei aleutenkran- ken Tieren eine deutliche Abnahme der Antikorperbildung gegen Hamocyanin und bovines Gammaglobulin.
Der pathogene Mechanismus, der zur Depression der Antikorperbildung im Verlaufe der Aleutenkrankheit fuhrt, ist unklar. Immerhin lassen sich nach LODMELL und Mitarbeitern (1970) verschiedene Moglichkeiten der Entstehung einer Irnmundepression in Betracht ziehen. Zunachst konnte das bei A. K. fur die gesamte Krankheitsdauer exzessiv zirkulierende y-Globulin (Virusanti-

Untersuchungen iiber die Pathogenese der Aleutenkrankheit der Nerze 477
korper) mittels eines homoiostatischen Mechanismus im Sinne von CEGLOWSKI und FRIEDMAN (1968) die Bildung eines anderen Antikorpers blockieren. Eine weitere Moglichkeit ware, daf3 das A. K.-Virus direkt mit der immunologi- schen Reifung von Stammzellen der Immunozyten interferiert (CEGLOWSKI u. FRIEDMAN, 1968). Die monatelange Persistenz des A. K.-Virus in den Geweben ist durch verschiedene Untersucher wiederholt nachgewiesen worden. Weiter- hin erscheint die Kompetition von Virusantigen und unbelebten Antigenen um pluripotente Stammzellen denkbar (SIEGEL u. MORTON, 1966). Hierbei ware eine zu groi3e Zahl von Immunozyten durch die immunologische Reak- tion mit dem A. K.-Virus besetzt, und es kame nur zu einer schwachen Immun- reaktion gegen ein zweites Antigen. Die letzte ErklZrungsmoglichkeit wird in einer Storung der Makrophagentatigkeit gesehen (LODMELL u. Mitarb., 1970). Aus der Persistens des A. K.-Virus bzw. von A. K.-Virus-Antikorperkomple- xen (PORTER und LARSEN, 1968; PORTER u. Mitarb., 1969) in Makrophagen konnte eine Storung ihrer Phagozytenfunktion und ihrer Fahigkeit, Antigene zu verarbeiten, resultieren.
Zusammenfassung Insgesamt 24 Nerze wurden mit dem Virus der Aleutenkrankheit infi-
ziert und in Abstanden von 4 Wochen in Gruppen von 4 Nerzen getotet. Je- weils 24 Tage vor der Totung einer Gruppe erhielten die Tiere Meerrettich- Peroxydase in komplettem Freundschen Adjuvans plantar in die Hinterextre- mitaten injiziert. Auf dem Hohepunkt der fur die Aleutenkrankheit (A. K.) typischen Hypergammaglobulinamie kommt es zu einer Depression der Anti- korperbildung gegen Peroxydase. Diesem Abfall des Antikorpertiters ent- s richt der Ruckgang der Zahl Peroxydase-Antikorper bildender Zellen in &n Lymphknoten, wobei die tiefsten Werte bei Nerzen 20 Wochen nach In- fektion mit dem A. K.-Virus gefunden werden.
Summary Studies on the pathogenesis of Aleutian disease in mink
VIII. Depression of antibody formation against horseradish peroxidase A total of 24 mink were infected with the virus of Aleutian disease and
groups of 4 were sacrificed at intervals of 4 weeks. Each animal received a plantar injection into the hind limbs of horseradish peroxidase in complete Freund’s adjuvant 24 days before being destroyed. At the peak of the hyper- gammaglobulinaemia typical of Aleutian disease there occurred a depression of antibody formation against the peroxidase. This fall in antibody titre represents the reduction in the number of cells producing peroxidase antibody in the lymph nodes. The lowest antibody levels were found 20 weeks after infection with the Aleutian disease.
RCsum6 Recherches sur la pathogbnkse de la maladie alioutienne du vison
VIII. DCpression de la formation des anticorps contre une peroxydase (Meerrettich)
24 visons ont ktk infect& avec le virus de la maladie alkoutienne et sacrifiks par groupes de 4 toutes les 4 semaines. 24 jours avant le sacrifice d’un groupe, les animaux ont r e p une peroxydase (Meerrettich) injectke plantairement dans les pattes postkrieures avec un adjuvant complet de Freund. On a constatk une depression de la formation des anticorps contre la peroxydase au point culminant de l’hypergammaglobulinkmie typique de la maladie
33*

478 G. TRAUTWEIN, P. SCHNEIDER und E. ERNST
alkoutienne. Cette chute du titre des anticorps correspond la diminution du nombre des cellules formatrices d’anticorps contre la peroxydase dans les ganglions lymphatiques; les valeurs les plus basses ont k t k trouvkes chez des visions 20 semaines aprks l’infection avec le virus de la maladie alkoutienne.
Resumen Estudios sobre la patogenia de la enfermedad aleutiana del viscin europeo
VIII. Depresi6n de la formacicin de anticuerpos frente a la peroxidasa del r ibano rusticano
24 visones europeos en total se infectaron con el virus de la enfermedad aleutiana, y se mataron en plazos de 4 semanas en grupos de 4 animales. 24 dias cada vez antes del sacrificio de un grupo, se le inyectb a 10s visones peroxidasa del r ibano rusticano con el adyuvante cornpleto de FREUND por plantar en las extremidades posteriores. En el fastigio de la hipergamma- globulinemia tipica para la enfermedad aleutiana (EA) se produce una de- presi6n en la formacibn de anticuerpos frente a la peroxidasa. A este des- censo en el titulo de anticuerpos le corresponde la disminucibn de la cifra de cklulas formadoras de anticuerpos peroxidhsicos en 10s ganglios linfiticos, hallindose 10s valores minimos en visones a las 20 semanas tras infeccibn con el virus de la EA.
Literaturverzeichnis 1. AVRAMEAS, S., and E. H. LEDUC, 1970: Detection of simultaneous antibody synthesis in
plasma cells and specialized lymphocytes in rabbit lymph nodes. J. exp. Med. 131,
2. AVRAMEAS, S., B. TAUDOU and S. CHUILON, 1969: Glutaraldehyde, cyanuric chloride and tetraazotized o-dianisidine as coupling reagents in the passive hemagglutination test. Immunochemistry 6,67-76.
3. BLOOM, G. D., B. CARLSOO and A. KUMLIEN, 1970: Cytochemical localization of peroxi- dase activity in the submandibular salivary gland of the hamster. Histochemie 22,
4. CEGLOWSKI, W. S., and H. FRIEDMAN, 1968: Immunosuppressive effects of Friend and Rauscher leukemia disease viruses on cellular and humoral antibody formation. J. Nat. Cancer Inst. 40, 983-995.
5. CHO, H. J., and D. G. INGRAM, 1973 a: Isolation, purification and structure of Aleutian disease virus by immunological techniques. Nature New Biology 243, 174-176.
6 . CHO, H. J., and D. G. INGRAM, 1973 b: Antigen and antibody in Aleutian disease in mink. 11. The reaction of antibody with the Aleutian disease agent using immuno- diffusion and immunoelectroosmophoresis. Canad. J. comp. Med. 37, 21 7-223.
7. HARTSOUGH, G. R., and J. R. GORHAM, 1956: Aleutian disease in mink. Nat. Fur News
8. HELMBOLDT, C. F., and E. L. JUNGHERR, 1958: The pathology of Aleutian disease in mink. Am. J. Vet. Res. 19, 212-222.
9. HESS, M. W., and B. SORDAT, 1968: In : SORDAT, B., M. SORDAT, M. W. HESS, R. D. STONER and H. COTTIER, 1970: Specific antibody within lymphoid germinal center cells of mice after primary immunization with horseradish peroxidase: A light and electron microscopic study. J. exp. Med. 131, 77-91.
10. KENYON, A. J., 1966: Immunologic deficiency in Aleutian disease of mink. Am. J. Vet. Res. 27, 1780-1782.
11. LEDUC, E., S. AVRAMEAS and M. BOUTEILLE, 1968: Ultrastructural localization of anti- body in differentiating plasma cells. J. exp. Med. 127, 109-118.
12. LEDUC, E., G . B. SCOTT and S. AVRAMEAS, 1969: Ultrastructural localization of intra- cellular immune globulins in plasma cells and lymphoblasts by enzyme - labeled anti- bodies. J. Histochem. Cytochem. 17, 211-224.
13. LODMELL, D. L., W. J. HADLOW, J. J. MUNOZ and H. W. WHITFORD, 1970: Hcmagglutinin antibody response of normal and Aleutian disease - affected mink to keyhole limpet hemocyanin. J. Immunol. 104, 878-887.
1137-1 168.
294-301.
28, 10-11, 38.

Untersuchungen uber die Pathogenese der Aleutenkrankheit der Nerze 479
14. MCGUIRE, T. C., T. B. CRAWFORD, J. B. HENSON and J. R. GORHAM, 1971: Aleutian disease of mink: Detection of large quantities of complement-fixing antibody to viral antigen. J. Immunol. 107, 1481-1482.
15. PADGETT, G. A., C. W. REIQUAM, J. B. HENSON and J. R. GQRHAM, 1968: Comparative studies of susceptibility to infection in the Chediak-Higashi syndrome. J. Path. Bact.
16. PORTER, D. D., and A. E. LARSEN, 1968: Aleutian disease of mink: Infectious virus- antibody complexes in the serum. Proc. SOC. Exp. Biol. Med. 126, 680-682.
17. PORTER, D. D., F. J. DIXON and A. E. LARSEN, 1965: Metabolism and function of gamma globulin in Aleutian disease of mink. J. exp. Med. 121, 889-900.
18. PORTER, D. D., A. E. LARSEN and H. G. PORTER, 1969: The pathogenesis of Aleutian disease of mink. I. In vivo viral reDlication and the host antibody response to viral
95, 509-522.
I -
antigen. J. exp. Med. 130, 575-593. 19. SIEGEL. B. V., and 1. I. MORTON. 1966: Serum agglutinin levels to sheer, red blood cells
in mice infected &h Rauscher virus. Proc. SOC. E ip . Biol. Med. 123, 467-470. 20. SORDAT, B., M. SORDAT, M. W. HESS, R. D. STONER and H. COTTIER, 1970: Specific
antibody within lymphoid germinal center cells of mice after primary immunization with horseradish peroxidase: A light and electron microscopic study. J. exp. Med. 131,
21. TRAUTWEIN, G., 1964: Experimentelle Untersuchungen uber die Aleutenkrankheit (,,Aleu- tian disease") der Nerze. Arch. exp. Veterinarmed. 18, 287-395.
22. TRAUTWEIN, G., 1969: Untersuchungen uber die Pathogenese der Aleutenkrankheit der Nerze. I. Veranderungen der Serumproteine. Zbl. Vet. Med. B, 16, 738-752.
23. TRAUTWEIN, G., 1970: Untersuchungen uber die Pathogenese der Aleutenkrankheit der Nerze. 111. Pathomorphologie der Organveranderungen. Zbl. Vet. Med. B, 17, 262-285.
24. TRAUTWEIN, G., W. DROMMER und D. SEIDLER, 1972: Untersuchungen uber die Patho- genese der Aleutenkranliheit der Nerze. VI. Infektiositat von Zellfraktionen. Zbl. Vet. Med. B, 19, 158-165.
Anschrift des Verfassers: Prof. Dr. Trautwein, Institut fur Pathologie, 3 Hannover,
77-9 1.
Bischofsholer Damm 15.