zooplankton of turbid and hydrologically dynamic prairie ... · pdf filejames h. thorp* and...
TRANSCRIPT

Zooplankton of turbid and hydrologically dynamicprairie rivers
JAMES H. THORP* AND SARA MANTOVANI †
*Kansas Biological Survey and Department of Ecology and Evolutionary Biology, University of Kansas, Lawrence, KS, U.S.A.†Department of Biology, University of Ferrara, Ferrara, Italy
SUMMARY
1. Compared with rivers in more humid, forested ecoregions of eastern and midwestern
U.S.A., rivers in semi-arid grassland of the U.S. Great Plains tend to be relatively shallow,
more variable in discharge, and characterised by high suspended sediment loads.
Although critical life stages of fish in prairie rivers probably depend at least partially on
zooplanktonic food, data on community and distributional patterns of potamoplankton in
these widespread ecosystems are almost entirely absent.
2. We examined summer zooplankton distribution in five prairie rivers (Arkansas,
Kansas, Platte, Elkhorn, and Niobrara Rivers) spread over six degrees of latitude during
2003–2004. We compared our results from 126 samples with previously collected data from
the Ohio and St Lawrence Rivers in forested ecoregions and correlated differences with
abiotic environmental conditions.
3. The importance of hydrological retention zones to stream biota has been recently
demonstrated for rivers with quasi-permanent islands and slackwater regions, but the
importance of slackwaters formed by ephemeral sandbar islands in prairie rivers is
unknown. We evaluated the role of hydrological retention for planktonic rotifers,
cladocera, and copepods in the Kansas River during the summer of 2004.
4. Zooplankton assemblages were extremely similar among prairie rivers (Sorensen
Dissimilarity Index: mean ¼ 0.07) but moderately disparate for comparisons of prairie
versus forested-basin rivers (mean ¼ 0.50).
5. Total zooplankton densities in prairie rivers (approximately 81 L)1) were intermediate
between the Ohio (approximately 92 L)1) and St Lawrence Rivers (approximately 43 L)1),
but relative abundances were significantly different. Rotifers represented >99% of
zooplankton individuals in grassland rivers, but only approximately 37–68% in other
rivers. Rotifer species richness was lower in prairie rivers, but relative abundances of
common genera were much less skewed compared with eastern rivers where Polyarthra
dominated rotifer assemblages (41–73%).
6. For comparisons among rivers, rotifers were significantly more abundant in turbid
rivers, while microcrustaceans were less dense. However, for comparisons within the
Kansas River over time, rotifer densities were inversely related to turbidity. We
hypothesise that rotifers indirectly benefit from river turbidity because their food
competitors (cladocera) and predators (e.g. cyclopoid copepods and visually feeding fish)
are relatively more susceptible to suspended sediments.
7. Crustacean densities were positively related to the degree of hydrological retention
(negatively to current velocities) throughout the study, but rotifer densities were
significantly depressed by current velocities only when river discharge was high, making
Correspondence: James H. Thorp, Kansas Biological Survey, Higuchi Hall, University of Kansas, 2101 Constant Ave., Lawrence,
KS 66047-3759 U.S.A. E-mail: [email protected]
Freshwater Biology (2005) 50, 1474–1491 doi:10.1111/j.1365-2427.2005.01422.x
1474 � 2005 Blackwell Publishing Ltd

slackwaters that much more valuable. Ephemeral sandbars may not provide sufficient
hydrological retention in time and space to sustain viable crustacean populations, but they
are adequate to help sustain growth of rotifer populations.
Keywords: Great Plains, hydrologic retention, Kansas River, microcrustaceans, rotifers
Introduction
Rivers in the semi-arid Great Plains of North America
are often physically rigorous and ecologically
demanding habitats for planktonic and benthic organ-
isms because of unstable sand substrates and high
suspended sediment loads. Their hydrographs are
relatively dynamic and largely controlled by highly
variable, thunderstorm-precipitation events (cf.
Dodds et al., 2004). Ecological research on North
American prairie rivers is rare, despite their distribu-
tion in an area constituting roughly one-third of the
conterminous U.S.A. Moreover, scientific knowledge
of potamoplankton in these rivers is almost non-
existent, although studies of plankton in prairie
reservoirs are not uncommon. Indeed, a computer
literature search revealed no ecological or systematics
publication on zooplankton in prairie or grassland
rivers in the last two decades.
Zooplankton are critical links in riverine food webs
between phytoplankton and fish (Jack & Thorp, 2002;
Thorp & Casper, 2003). In prairie rivers they are
probably now a primary food source for larval and
some adult fish and were undoubtedly important long
before humans began building reservoirs. For exam-
ple, adults of the large and ancient paddlefish
Polyodon spathula (Walbaum), which have attained a
body weight of at least 36.7 kg in grassland rivers of
Kansas, feed almost exclusively on zooplankton
(Cross & Collins, 1995). Consequently, determining
the factors controlling zooplankton density, diversity,
and distribution should be an important step toward
understanding the ecology of prairie rivers.
Riverine zooplankton are controlled by a poorly
understood mixture of abiotic and biotic factors
varying seasonally among and within rivers (Thorp
& Casper, 2002). Abiotic factors include those influen-
cing the abundance and access to food, mechanics of
feeding, downstream transport versus temporary
retention, direct mortality (e.g. from ultraviolet radia-
tion), and thermal conditions. Biotic factors include
competition for food, parasitism, disease, and plank-
tivory by fish and both benthic and pelagic inverte-
brates.
The relative importance of various abiotic and biotic
factors to zooplankton assemblages are likely to vary
among species, seasons, and types of rivers. For
example, the substantial role of hydrological retention
for production of riverine zooplankton has been
demonstrated in large rivers (Thorp et al., 1994;
Schiemer et al., 2001; Hein et al., 2005; J.H. Thorp &
A.F. Casper, unpublished data for the St Lawrence
River). However, those studies have focused on rivers
with relatively stable hydrographs compared to those
in rivers of semi-arid ecoregions. Moreover, pre-
viously studied rivers usually featured relatively
permanent islands and slackwaters (¼shorelines,
embayments, and other areas outside the main chan-
nel where current velocities are substantially reduced,
e.g. below 0.1 m s)1). The scientific literature does not
indicate whether the more ephemeral slackwaters
associated with relatively unstable and ephemeral
sandbar islands in prairie rivers significantly affect
potamoplankton community diversity and produc-
tion.
In our potamoplanktonic study, we asked three
primary questions. First, do assemblages of rotifers,
copepods, and cladocera differ substantially among
Great Plains rivers and between those prairie streams
and rivers with forested watersheds in the midwes-
tern and north-eastern U.S.A.? In an initial step
examining this question, we compared potamoplank-
ton among: (i) five turbid, medium to large grassland
rivers (the Elkhorn, Niobrara, Arkansas, Platte, and
Kansas Rivers) whose hydrographs range from low to
high variability; (ii) the very large, constricted Ohio
River, which carries less sediment and has a moder-
ately variable hydrograph; and (iii) the very large
St Lawrence River, which is exceptionally clear and
hydrologically stable because of its origin primarily in
the Great Lakes (Thorp, Lamberti & Casper, 2005a).
Second, are differences among rivers in zooplankton
assemblages correlated with hydrological character-
istics, turbidity, water temperatures, or some other
Zooplankton in prairie rivers 1475
� 2005 Blackwell Publishing Ltd, Freshwater Biology, 50, 1474–1491

abiotic factor? Third, what is the importance of
hydrological retention for zooplankton in prairie
rivers with ephemeral sandbars? To answer this last
question, we sampled habitats with different current
velocities in the Kansas River.
Methods
Sample sites and habitat analyses
We sampled zooplankton from five prairie rivers in the
summers of 2003 and 2004 (Fig. 1a) and compared our
results with data collected in the summers of 1992–2000
from the Ohio and St Lawrence Rivers by JHT and other
researchers (especially additional Ohio River data from
Dr Debbie Guelda at Bemidji State University, Bemidji,
MN, U.S.A.). The last two rivers were selected in part
because zooplankton have been studied more exten-
sively in these rivers than in other U.S.A. and Canadian
rivers. We also collected miscellaneous environmental
data for all prairie rivers and current velocity by habitat
for the Kansas River during 2004. Finally, we analysed
rivers for discharge and turbidity using gauging data
from the US Geological Survey (USGS). Turbidity was
measured in nephelometric units (NTUs) because
USGS data bases were more extensive for NTUs than
for suspended sediment (mg L)1). The weakness of this
approach is that suspended sediments are not the only
factor contributing to an NTU level. Some pertinent
differences among these seven rivers are shown in
Table 1, and the distribution of rivers are plotted
(Fig. 2a) using a principal component analysis (PCA;
Clarke, 1993) of turbidity, mean discharge, and dis-
charge variability. This PCA plot is useful later in the
Results for interpreting differences in zooplankton
assemblages among rivers.
Zooplankton from prairie rivers were collected
from a single reach in the Arkansas, Elkhorn, Nio-
brara, and Platte Rivers and from multiple reaches in
the Kansas River (Fig. 1a). Samples were collected
in 2003–2004 from a single habitat type (near shore in
slackwater sites) from all five prairie rivers as part of a
larger watershed-river study sponsored by the US
Environmental Protection Agency (EPA); these are
referred to as EPA data in the Results. In addition,
zooplankton were more intensively sampled from
multiple habitats in several reaches of the Kansas
River (Fig. 1b) during July to September 2004. These
latter habitats varied substantially in current velocity
(all £ 0.51 m s)1, with mean ¼ 0.14 m s)1), tempera-
ture, and other environmental parameters. To be
conservative, we used the EPA samples of the Kansas
River when comparing among all seven rivers and the
non-EPA samples from the Kansas River in 2004 for
analyses of possible effects of hydrological retention.
All five prairie rivers are relatively wide for their
discharge, shallow (often <2 m deep), and turbid
compared with a typical river in a forested ecoregion.
The Platte, Kansas, and Elkhorn are in a high-
turbidity cluster (Table 1), whereas the Niobrara and
Arkansas carry only about half as much suspended
sediment (correlated with turbidity). However, the
mean of the latter group was still twice as high as the
average for the Ohio River and nearly 60 times higher
than the average turbidity of the St Lawrence River,
which is the clearest of the top 10 large rivers of the
world (Gleick, 1993). This permitted us to analyse
turbidity effects using average values for turbidity
rather than relying on highly variable, daily values
during sample dates.
Zooplankton are directly influenced by current
velocity and turbulence and only indirectly by river
discharge, but one can gain a perspective on differ-
ences among rivers in overall hydrological conditions
by examining discharge patterns. The St Lawrence
and Ohio represent two of the four largest rivers in
North America in mean discharge, and both are at
least 15 times larger than the biggest prairie river we
studied (the Kansas). Sampling from prairie rivers in
2003 and early summer of 2004 was during a severe,
multi-year drought, but the more intensive sampling
of the Kansas River associated with habitat analysis
occurred mostly in postdrought conditions (Fig. 3).
Indeed, the summer of 2004 was the third rainiest in
the lower basin of the Kansas River since 1939.
Consequently stream discharges were much higher
in 2004 than in either 2003 or the historical averages
(Fig. 3). Habitat conditions, such as the presence and
nature of sandbars, alter as a result of changes in river
stage and current velocity.
River flows are relatively dynamic for these prairie
rivers in both drought and flood years compared with
similar-order rivers from the more humid eastern
U.S.A. This results because prairie rivers drain semi-
arid watersheds and are largely fed by surface runoff
following thunderstorms (Dodds et al., 2004) and some
groundwater, with the proportions varying among
seasons and rivers. Our seven rivers fit within four
1476 J.H. Thorp and S. Mantovani
� 2005 Blackwell Publishing Ltd, Freshwater Biology, 50, 1474–1491
43°0'0"N
37°0'0N"N
102°0'0"W
95°0'0"W
MississippiRiver
MissouriRiver
OhioRiver
ArkansasRiver
Kansas River
PlatteRiver
Niobrara River Elkhorn River
*
**
**
*
(a)
(b)
Slack waters0 50 100 m
Temporarysand bar island
Agricultural field Riparian zone
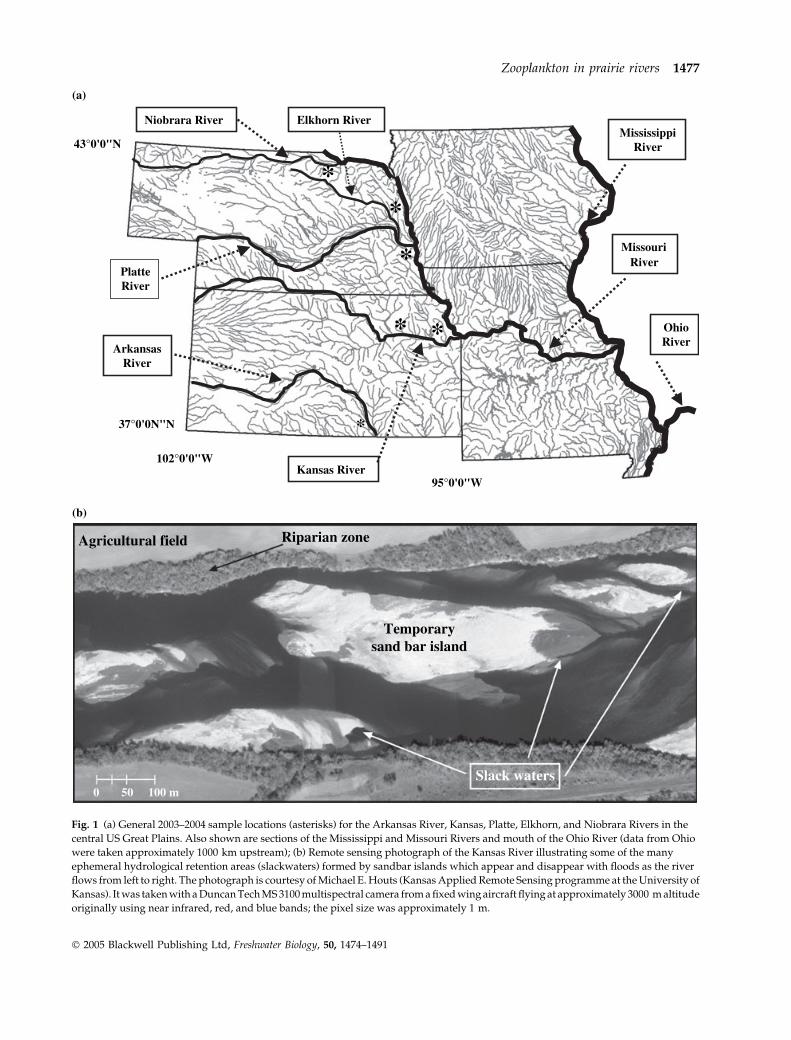
Fig. 1 (a) General 2003–2004 sample locations (asterisks) for the Arkansas River, Kansas, Platte, Elkhorn, and Niobrara Rivers in the
central US Great Plains. Also shown are sections of the Mississippi and Missouri Rivers and mouth of the Ohio River (data from Ohio
were taken approximately 1000 km upstream); (b) Remote sensing photograph of the Kansas River illustrating some of the many
ephemeral hydrological retention areas (slackwaters) formed by sandbar islands which appear and disappear with floods as the river
flows from left to right. The photograph is courtesy of Michael E. Houts (Kansas Applied Remote Sensing programme at the University of
Kansas). It was taken with a Duncan Tech MS 3100 multispectral camera from a fixed wing aircraft flying at approximately 3000 m altitude
originally using near infrared, red, and blue bands; the pixel size was approximately 1 m.
Zooplankton in prairie rivers 1477
� 2005 Blackwell Publishing Ltd, Freshwater Biology, 50, 1474–1491
arbitrary categories of discharge variability, as deter-
mined by the coefficient of variation (standard devi-
ation/mean) (Table 1): high (Arkansas, Kansas, and
Elkhorn), medium (Platte and Ohio), low (Niobrara),
and exceedingly low (St Lawrence). The Niobrara has
unusually low discharge variability for a prairie river
because groundwater constitutes a significant source of
its river water (Dodds, 2002). The St Lawrence may
have one of the least variable discharge patterns (0.15;
Table 1) of the large rivers of the world because >95%
of its discharge at a point several hundred kilometres
from its origin (where most of our zooplankton data
were obtained) (Thorp et al., 2005a) is derived from the
Laurentian Great Lakes.
The abundance of hydrological retention areas –
where current velocities and sometimes turbulence are
low – varies among these seven rivers. Slackwaters in
our prairie rivers are primarily formed by ephemeral
sandbars and a few more permanent forested islands.
Semi-permanent islands with trees are relatively rare
because sandbar islands often arise and disappear
periodically with floods. Consequently, slackwater
habitats formed next to these islands are much more
ephemeral than in rivers with more stable banks and
river beds. The predominant substrate in these prairie
rivers is coarse to fine sand and silt, with some pea-
sized gravel in faster flowing habitats and more silt in
low velocity habitats. While stone substrates are rare
except for some small gravel bars, the shallow nature
of these rivers and their presence in a generally windy
climatic zone increases turbulence over the relatively
homogenous sand substrates. In contrast, the Ohio
and St Lawrence Rivers contain a few to very many
relatively permanent, wooded islands; substrates are
typically much coarser on average. The mostly con-
stricted-channel Ohio River now has few islands and
embayments and is deeper than prairie rivers. The
upper two-thirds of the river (including areas sampled
for zooplankton) has many shoreline boulders and
cobble which promote some turbulence. The St Law-
rence is replete with semi-permanent slackwater areas
formed by forested islands and embayments, and
submerged rocks are common near shore. In addition
to natural turbulence, the last two rivers are also
heavily navigated by ships (mostly St Lawrence),
barges, and pleasure craft whose wakes disturb the
water to some extent, especially near shore.
Our seven rivers vary in the number and types of
dams present on their main channels and tributaries.
High dams are absent or rare in the main channels of all
five prairie rivers, but all contain one or more reser-
voirs on tributaries. The Kansas basin has the highest
proportion of dammed tributaries. Most samples from
the Kansas River were collected at sites approximately
20–60 km below a high-dam reservoir (Perry Lake) on
the Delaware River tributary. However, we also
sampled zooplankton in the Kansas 5–10 km upstream
from its confluence with the Delaware to account for
any potentially significant reservoir effects. The main
channel of the Ohio River has many low-head naviga-
tion dams. Tributaries upstream of the sample sites on
the Ohio include some low and high dams. The St
Lawrence River was sampled in the main channel and
slackwaters between Lake Ontario and the first hydro-
electric dam (at Massena, NY and Cornwall, Ontario,
hundreds of kilometres downstream from the Great
Table 1 Selected environmental characteristics of rivers in our study. Data from the US Geological Survey, a book on North American
rivers (Benke & Cushing, 2005), and other sources. Information reflects environmental characteristics near sites where we sampled.
Discharge records span 66–102 years and represent means of daily discharge values. Turbidity records cover at least 18 years but
were less frequently taken each year.
River Biome
Freshwater
ecoregion
River order
(approximate)
Discharge
mean (cm)
Coefficient of
variation
Turbidity
(NTUs)
Rivers in forested watersheds
St Lawrence Temperate deciduous and
boreal forests
Lower Saint
Lawrence
8 7565.3 0.15 1.16
Ohio Eastern deciduous forests Teays-Old Ohio 9 3491.3 1.00 31.09
Rivers in grassland watersheds
Niobrara Temperate grasslands Middle Missouri 4 51.2 0.55 69.97
Elkhorn Temperate grasslands Middle Missouri 4 40.7 1.68 109.29
Platte Temperate grasslands Middle Missouri 5 168.4 0.95 155.25
Kansas Temperate grasslands Middle Missouri 7 221.9 1.69 125.90
Arkansas Temperate grasslands Southern plains 6 57.7 1.89 67.54
1478 J.H. Thorp and S. Mantovani
� 2005 Blackwell Publishing Ltd, Freshwater Biology, 50, 1474–1491

Lakes). The main channels of all seven rivers are
flowing ecosystems lacking both significant vertical
stratification and other conditions that characterise
rivers with reservoirs.
All prairie-river sample sites were in medium to
large rivers that drain tall to mostly mixed and short-
grass prairie ecoregions. The eastern portion of these
watersheds typically include extensive areas of row
crop agriculture and riparian forests, but their water is
also derived from the drier central to western regions
with cattle rangeland, row crops, and sparse riparian
forests. Direct or groundwater extraction of water for
this agriculture is a serious concern for most prairie
rivers. In contrast, the watersheds of the Ohio and St
Lawrence Rivers are naturally wooded (deciduous
and coniferous forests) but also contain row crop
agriculture [see environmental information on most of
these rivers in Benke & Cushing (2005)].
Collection and identification of zooplankton
Seventy-eight zooplankton samples from the shallow
Kansas River were collected in July to September 2004
from depths averaging approximately 32 cm using
multiple grab sampling with small buckets to obtain a
final volume of 21 L. A 1-L ‘rotifer sample’ was
immediately removed and filtered through a 20 lm
sieve, immersed in 95% ethyl alcohol (ETOH) for
approximately 30 s to kill rotifers quickly (preserving
body shape), and then preserved in 75% ETOH for
later identification. The remaining 20-L ‘microcrusta-
cean sample’ was filtered through a 106 lm sieve,
with the retained contents then preserved in 75%
ETOH. All rotifers and microcrustaceans (copepods
and cladocera only) in the 21-L sample were counted
and identified at least to genus using a Nikon TE 2000-
S inverted microscope for rotifers and a Nikon SMZ-
1500 stereomicroscope for microcrustaceans. Genera
were identified using taxonomic keys, illustrations,
and photographs in Stemberger (1979), Balcer, Korda
& Dodson (1984), and Thorp & Covich (2001).
Four to eight additional 20-L zooplankton samples
per river per year were collected in the summers of
2003–2004 from the Arkansas, Kansas, Platte, Elkhorn,
and Niobrara Rivers (total sample size ¼ 48).
Zooplankton were pumped (12-volt diaphragm
pump) from approximately 1 m depth into a 20-L
bucket and then poured through a 20 lm sieve. The
retained zooplankton were rapidly killed in 95%
ETOH before being preserved in 75% ETOH. Micro-
crustaceans were counted and identified from the
entire 20-L sample, but a subsample (approximately
1 L) was processed for rotifers.
Zooplankton were collected from the St Lawrence
Rivers using a high-speed diaphragm pump and
similar sieve sizes and from the Ohio River using
comparable sieves but either a manual (samples from
D. Guelda) or electric diaphragm pumps (samples
from JHT).
Statistical analyses
Communities of zooplankton in different rivers were
compared with techniques for multidimensional sca-
PC1 (77.9%)–2 –1 0 1 2 3 4
PC
2 (1
6.5%
)D
imen
sion
2
Dimension 1
–1.0
–0.5
0.0
0.5
1.0
1.5
Arkansas
Elkhorn
Kansas
Niobrara
Platte
Ohio
St. Lawrence
(a)
(b)
Principal component analysisof environmental conditions
–1.0 –0.5 0.0 0.5 1.0 1.5–1.0
–0.8
–0.6
–0.4
–0.2
0.0
0.2
0.4
0.6
0.8Multi-dimensional scaling analysisof rotifer diversity and density
Arkansas
Kansas
Niobrara
St. Lawrence
Elkhorn
PlatteOhio
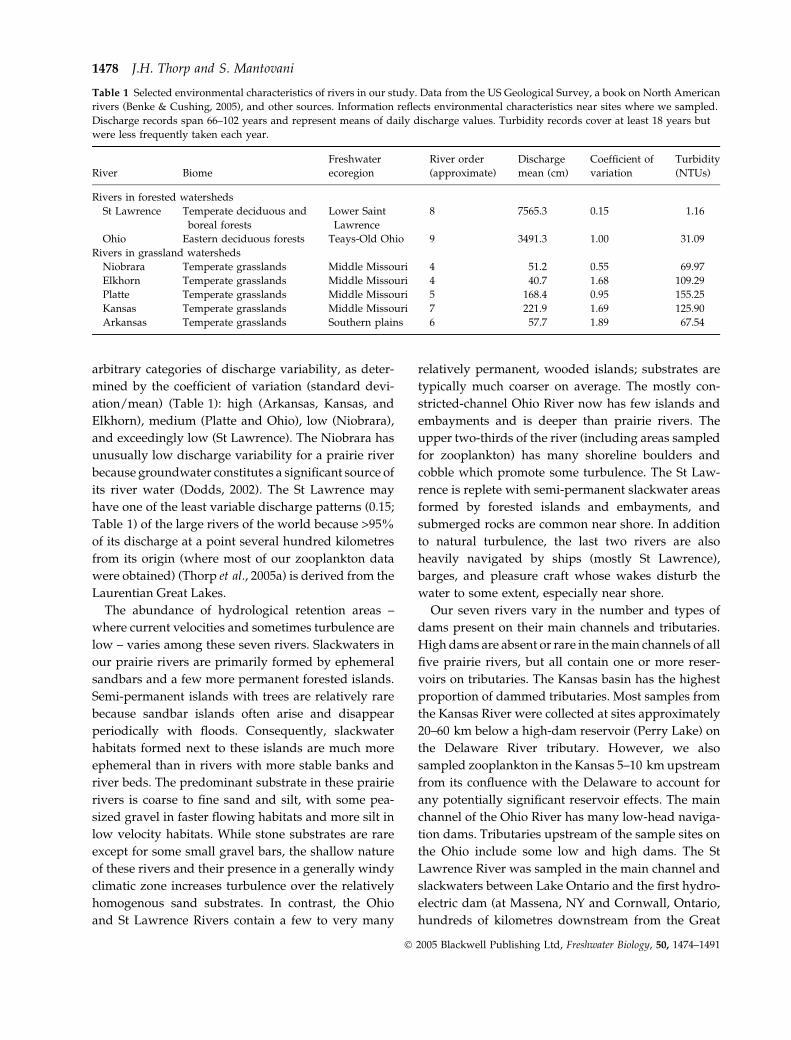
Fig. 2 (a) Principal component analysis based on environmental
differences among seven rivers. Mean river discharge, discharge
variability (SD/mean), and turbidity contributed 0.617, )0.574,
and )0.538, respectively to the PC1 axis, and )0.133, 0.598, and
)0.791 to the PC2 axis. Ovals link prairie rivers (on left) or the
two large rivers from eastern deciduous forest regions; (b)
Multidimensional scaling analysis based on densities of rotifer
genera in the seven rivers; The very low stress value (<0.001)
indicates that the plot was highly representative of the data.
Zooplankton in prairie rivers 1479
� 2005 Blackwell Publishing Ltd, Freshwater Biology, 50, 1474–1491

ling (MDS), PCA, a community similarity index, and
analysis of variance (ANOVAANOVA). Dissimilarities among
rivers in their zooplankton assemblage were quanti-
fied in part using the Sorensen index (DI):
DI ¼ 1 � ½2a=ðbþ cÞ�
where a and b equal the number of genera in river 1
and river 2, respectively, and c ¼ the number of genera
in all rivers (or separate groups of rivers); DI ¼ ranges
from 0 to 1 (i.e. minimum to maximum dissimilarity)
(Magurran, 1988). Community differences were also
analysed graphically with MDS techniques (Young &
Hamer, 1987) using densities of rotifer genera. ANOVAANOVA
tests were conducted on overall zooplankton densities,
species diversity/evenness (Shannon and Pielou
Indices), and abundance of the most abundant rotifer
species using transformed data [Log10 (x + 1)]. Results
were compared between rivers using post hoc Tukey
honestly significant difference (HSD) tests. We also
examined relationships between zooplankton densi-
ties and four environmental metrics (mean turbidity,
discharge, discharge variability, and current velocity)
using correlations (Pearson Index) and regressions
(examined with t-tests for significance).
Results
Zooplankton of rivers in grassland and forested
ecoregions
Principal component analysis of effects of turbidity,
mean discharge, and discharge variability demonstra-
ted that prairie rivers were in a group distinct from
Ohio and St Lawrence Rivers (Fig. 2a), and this pattern
was also apparent in their zooplankton assemblages.
An MDS plot of rotifer densities (based on the 11 most
common genera overall) clearly indicated that prairie
river zooplankton communities were different from
those in rivers of forested ecoregions (Fig. 2b). Ten
paired-river comparisons of zooplankton in the five
Great Plains rivers had an average Sorensen Index of
0.07 (range ¼ 0.03–0.11), indicating extremely high
similarity in their taxonomic composition. In contrast,
other paired comparisons showed much more dissimi-
lar communities for the Ohio versus St Lawrence (0.46),
Ohio versus prairie rivers as a group (0.51), St Lawrence
versus prairie rivers (0.49), and the two very large rivers
versus prairie rivers (0.42). These results also indicate
that zooplankton assemblages differed considerably
between the Ohio and St Lawrence Rivers, which is not
Fig. 3 May to September discharge (cm) patterns in the Kansas River during the study (2003–04) and averaged over a period of
85 years. Sample dates during our study are shown as asterisks.
1480 J.H. Thorp and S. Mantovani
� 2005 Blackwell Publishing Ltd, Freshwater Biology, 50, 1474–1491

surprising given their substantial differences in turbid-
ity, discharge variability, and watersheds (Table 1).
Decreasing water temperatures on a latitudinal basis
might also have caused zooplankton differences
between these two eastern rivers, but such biotic
differences were not evident along a similar gradient
for the prairie rivers.
Total zooplankton densities varied significantly
among rivers [ANOVAANOVA d.f. ¼ 6, 51, MS (error) ¼0.101, F ¼ 90 116.01, P < 0.01] (Fig. 4). Tukey HSD
tests demonstrated that densities of zooplankton in
the Ohio and St Lawrence Rivers were significantly
different (P < 0.05) from each other and from all
prairie rivers. Of 10 possible comparisons among
prairie rivers, however, only three were significantly
different, and two of these involved the more unique,
spring-fed Niobrara River (with the Kansas and Platte
Rivers). Mean zooplankton numbers for the Kansas
River were lowest among the seven rivers when using
2003–2004 EPA samples (Fig. 4), but were highest if
the larger data set from our 2004 hydrological retent-
ion study was used. There were no consistent patterns
in zooplankton densities to suggest that zooplankton
overall fared better or worse in prairie rivers com-
pared to eastern rivers.
Significant differences among rivers were also
evident for species diversity [Shannon Index: ANOVAANOVA
d.f. ¼ 6, 51, MS (error) ¼ 0.002, F ¼ 1021.51, P < 0.01]
and evenness [Pielou Index: MS (error) ¼ 0.001, F ¼262.95, P < 0.01; Fig. 4]. As with overall densities,
both species diversity and evenness indices were
significantly different (Tukey HSD tests, P < 0.05)
when comparing both the Ohio versus the St
Lawrence River and each of those rivers versus any
prairie river. In contrast, only one of 10 possible
comparisons among prairie rivers were different for
the Shannon Index (Niobrara versus Arkansas), again
demonstrating similarity among zooplankton assem-
blages of Great Plains rivers. However, half the
comparisons among various prairie rivers for even-
ness showed significant differences (two involving the
Niobrara).
The most dramatic difference between prairie rivers
and the eastern rivers concerned the dominance of
rotifers (Appendix). On average, 99% of the zooplank-
ton fauna (exclusive of protozoa) in our five prairie
rivers were rotifers, compared with 68% in the St
Lawrence River and only 36% in the Ohio River.
(Data were derived from studies using slightly
different collection techniques, but the mesh sizes
Rivers
Div
ersi
ty in
dice
s
0.0
0.5
1.0
1.5
2.0
2.5
Zoo
plan
kton
den
sity
(no
. L–1
)
0
20
40
60
80
100
120
140Shannon
PielouDensity
AR KA PL EL NI Prairie OH SL
Fig. 4 Differences among rivers in density (line), generic diversity (Shannon index, black bars), and evenness (Pielou index, gray
diagonal bars); horizontal bars are mean±1 SE. River abbreviations are AR, Arkansas; KA, Kansas; PL, Platte; EL, Elkhorn; NI,
Niobrara; Prairie, those five previous rivers; OH, Ohio; and SL, St Lawrence.
Zooplankton in prairie rivers 1481
� 2005 Blackwell Publishing Ltd, Freshwater Biology, 50, 1474–1491

used for filtering water samples still collected roughly
the same percentage of rotifers.) The Ohio River was
much more balanced numerically among cladocera
(20%), copepods (44%, including nauplii), and roti-
fers, while the St Lawrence had more nearly equal
percentages of cladocera (14%) and copepods (18%).
Mean densities for all zooplankton (using EPA sam-
ples for all five prairie rivers) were greatest in the
Ohio River (approximately 93 animals L)1), medium
in the prairie rivers (approximately 81 L)1), and
lowest in the St Lawrence approximately 43 L)1),
despite the primary origin of the St. Lawrence in the
zooplankton-rich Great Lakes. However, such num-
bers will fluctuate over time, among habitats, and
with slightly different collection techniques. For
example, when we used samples from our 2004
hydrological retention study in the Kansas River as
a substitute for the 2003–2004 EPA samples from the
same river, the mean density for prairie rivers rose by
over 80% to 147 animals L)1. This dramatic rise was
caused primarily by the population explosion of one
genus (Proales), whose mean percentage of the total
rotifer density in Kansas River samples increased
from 9% to 51%.
More detailed taxonomic analyses also shows sig-
nificant differences among rivers in taxonomic com-
position. Crustaceans were very rare in channel and
slackwater (Fig. 1b) samples from prairie rivers but
were common in the Ohio and St. Lawrence Rivers
(Appendix). The primary cladoceran genera in the
Ohio River were Bosmina, Ceriodaphnia, and Diaphano-
soma. In the St. Lawrence, Bosmina and Daphnia were
the major cladocera throughout the international
section of the river on average; but after a hundred
river kilometres or so downstream from Lake Ontario,
Bosmina was the only common cladoceran. Calanoid
copepods were not common in any of the seven rivers,
but Eurytemora affinis – an estuarine invader – was the
overwhelming dominant calanoid in the Ohio and
St. Lawrence Rivers. Diacyclops was the most abun-
dant cyclopoid copepod in both eastern rivers; but
other genera, such as Mesocyclops, often were import-
ant in samples from experimental enclosure studies in
the Ohio and St Lawrence Rivers (Jack & Thorp, 2000,
2002; Thorp & Casper, 2002, 2003).
Significant differences (ANOVAANOVA; P < 0.01) existed
among the seven rivers in densities of 10 of the 11
most common rotifer genera, which together repre-
sented >90% of the rotifer densities. Moreover, the
11th genus, Keratella, was nearly different statistically
(P ¼ 0.062). The St Lawrence was strongly dominated
primarily by Polyarthra (approximately 73%) and
secondarily by Synchaeta (approximately 12%).
Polyarthra was also the dominant rotifer in the Ohio
River (41%), but three other genera represented at
least 9% of the rotifers (in order: Brachionus, Keratella,
and Synchaeta). In contrast, none of the rotifer dom-
inants of the Ohio and St Lawrence were important in
prairie rivers. About 78% of the zooplankton in these
Great Plains systems were composed of five genera
ranging in percentages from approximately 10–20%
(in order from most abundant: Monostyla, Notholca,
Gastropus, Tricocerca, and Proales). During our hydro-
logical retention study (discussed below), the domin-
ant rotifers in the Kansas River were Proales
(approximately 51%), Monostyla (approximately
19%), and Trichocerca (approximately 15%).
The role of water movement and turbidity among and
within rivers
We analysed relationships between zooplankton
densities and four environmental metrics for both
among- and within-river comparisons because these
potentially have different relationships to zooplank-
ton ecology. For comparisons among the seven rivers,
we evaluated effects on zooplankton from mean
turbidity, discharge, and discharge variability using
U.S.G.S. environmental data, our EPA zooplankton
data, and other data collected from the Ohio River
(data sets from Guelda, Thorp, and others) and the
St Lawrence River (by Thorp and Casper). For within-
river comparisons, however, we investigated the
relationships between zooplankton densities in the
Kansas River and both current velocity and turbidity
within specific habitats.
Relationships between zooplankton densities and
turbidity initially appeared inconsistent when making
among- and within-river comparisons, but these
differences may be explained by species adaptations
and/or biotic interactions (see Discussion). We found
a positive linear relationship between density of
rotifers and a river’s average turbidity (calculated
over many years) (Fig. 5a) and a negative regression
for crustaceans and turbidity (Fig. 5b); however, only
the first approached significance (d.f. ¼ 5, P < 0.10).
The linear regression R2-value for rotifers was mod-
erate (0.4965) when all seven rivers were considered,
1482 J.H. Thorp and S. Mantovani
� 2005 Blackwell Publishing Ltd, Freshwater Biology, 50, 1474–1491
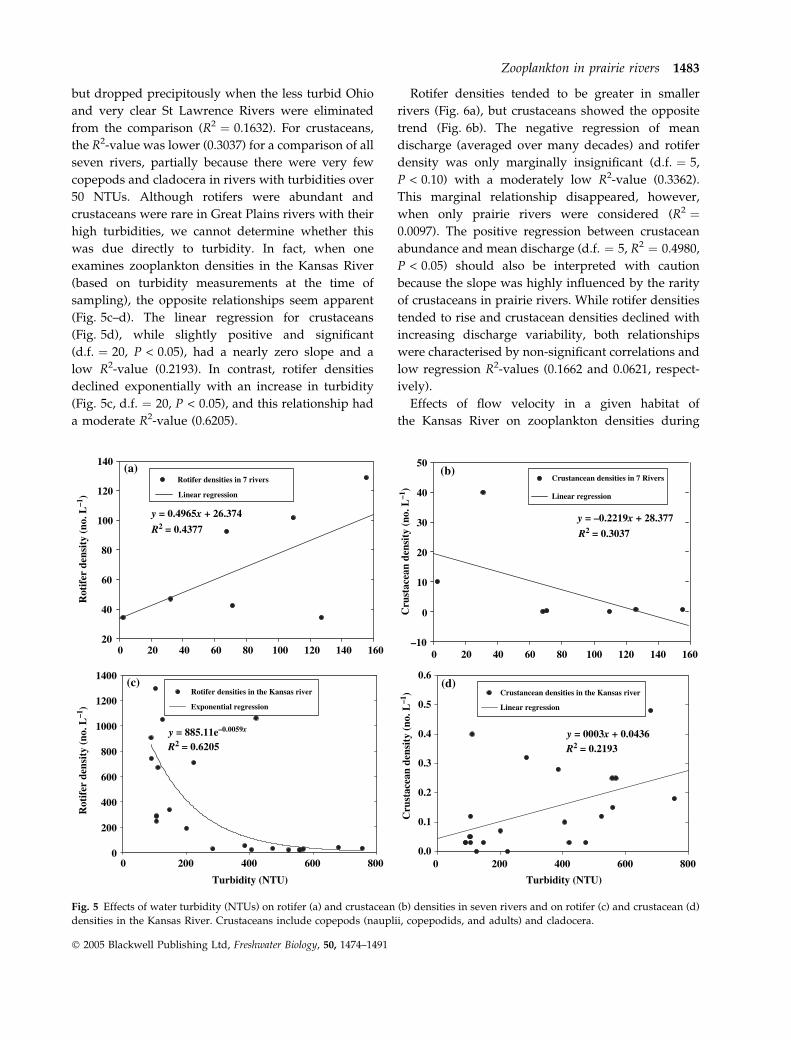
but dropped precipitously when the less turbid Ohio
and very clear St Lawrence Rivers were eliminated
from the comparison (R2 ¼ 0.1632). For crustaceans,
the R2-value was lower (0.3037) for a comparison of all
seven rivers, partially because there were very few
copepods and cladocera in rivers with turbidities over
50 NTUs. Although rotifers were abundant and
crustaceans were rare in Great Plains rivers with their
high turbidities, we cannot determine whether this
was due directly to turbidity. In fact, when one
examines zooplankton densities in the Kansas River
(based on turbidity measurements at the time of
sampling), the opposite relationships seem apparent
(Fig. 5c–d). The linear regression for crustaceans
(Fig. 5d), while slightly positive and significant
(d.f. ¼ 20, P < 0.05), had a nearly zero slope and a
low R2-value (0.2193). In contrast, rotifer densities
declined exponentially with an increase in turbidity
(Fig. 5c, d.f. ¼ 20, P < 0.05), and this relationship had
a moderate R2-value (0.6205).
Rotifer densities tended to be greater in smaller
rivers (Fig. 6a), but crustaceans showed the opposite
trend (Fig. 6b). The negative regression of mean
discharge (averaged over many decades) and rotifer
density was only marginally insignificant (d.f. ¼ 5,
P < 0.10) with a moderately low R2-value (0.3362).
This marginal relationship disappeared, however,
when only prairie rivers were considered (R2 ¼0.0097). The positive regression between crustacean
abundance and mean discharge (d.f. ¼ 5, R2 ¼ 0.4980,
P < 0.05) should also be interpreted with caution
because the slope was highly influenced by the rarity
of crustaceans in prairie rivers. While rotifer densities
tended to rise and crustacean densities declined with
increasing discharge variability, both relationships
were characterised by non-significant correlations and
low regression R2-values (0.1662 and 0.0621, respect-
ively).
Effects of flow velocity in a given habitat of
the Kansas River on zooplankton densities during
0 20 40 60 80 100 120 140 160 0 20 40 60 80 100 120 140 160
Rot
ifer
den
sity
(no
. L–1
)
20
40
60
80
100
120
140
Rotifer densities in 7 rivers Linear regression
y = 0.4965x + 26.374
R2 = 0.4377
(a)
Cru
stac
ean
dens
ity
(no.
L–1
)
–10
0
10
20
30
40
50
y = –0.2219x + 28.377
R2 = 0.3037
(b)Crustancean densities in 7 Rivers
Linear regression
Turbidity (NTU)
Rot
ifer
den
sity
(no
. L–1
)
0 200 400 600 8000
200
400
600
800
1000
1200
1400
Exponential regression
y = 885.11e–0.0059x
R2 = 0.6205
(c)Rotifer densities in the Kansas river
Turbidity (NTU)
Cru
stac
ean
dens
ity
(no.
L–1
)
0 200 400 600 8000.0
0.1
0.2
0.3
0.4
0.5
0.6
y = 0003x + 0.0436 R2 = 0.2193
(d)Crustancean densities in the Kansas river
Linear regression
Fig. 5 Effects of water turbidity (NTUs) on rotifer (a) and crustacean (b) densities in seven rivers and on rotifer (c) and crustacean (d)
densities in the Kansas River. Crustaceans include copepods (nauplii, copepodids, and adults) and cladocera.
Zooplankton in prairie rivers 1483
� 2005 Blackwell Publishing Ltd, Freshwater Biology, 50, 1474–1491
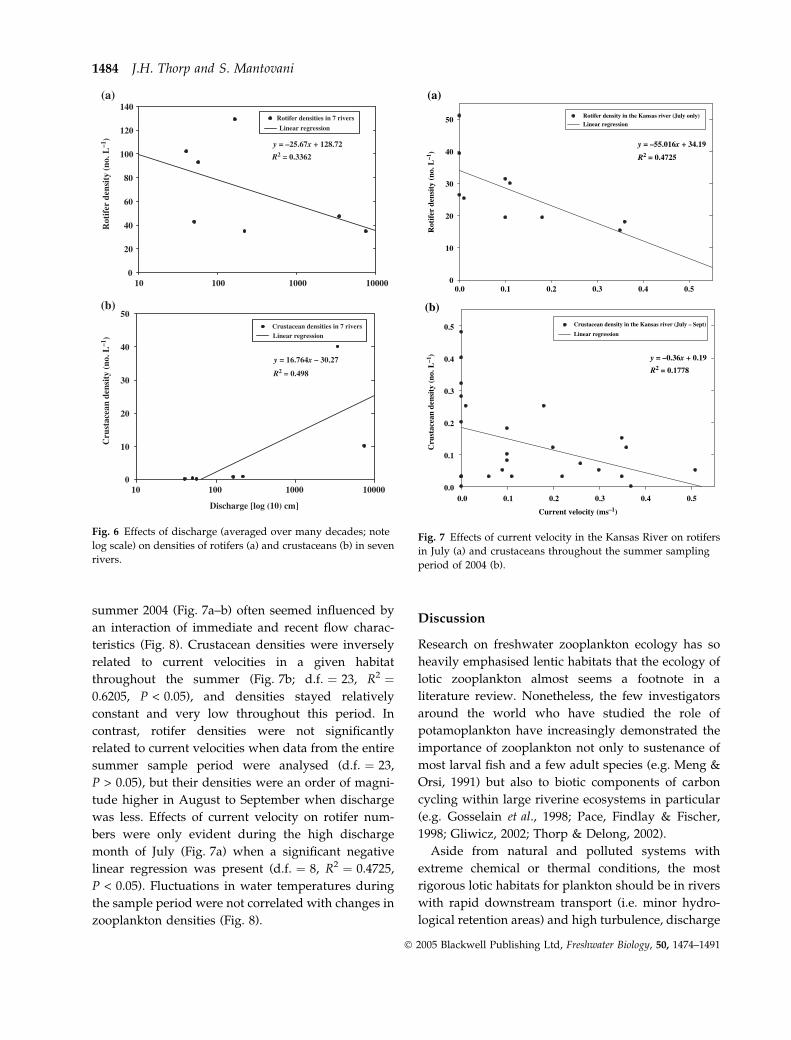
summer 2004 (Fig. 7a–b) often seemed influenced by
an interaction of immediate and recent flow charac-
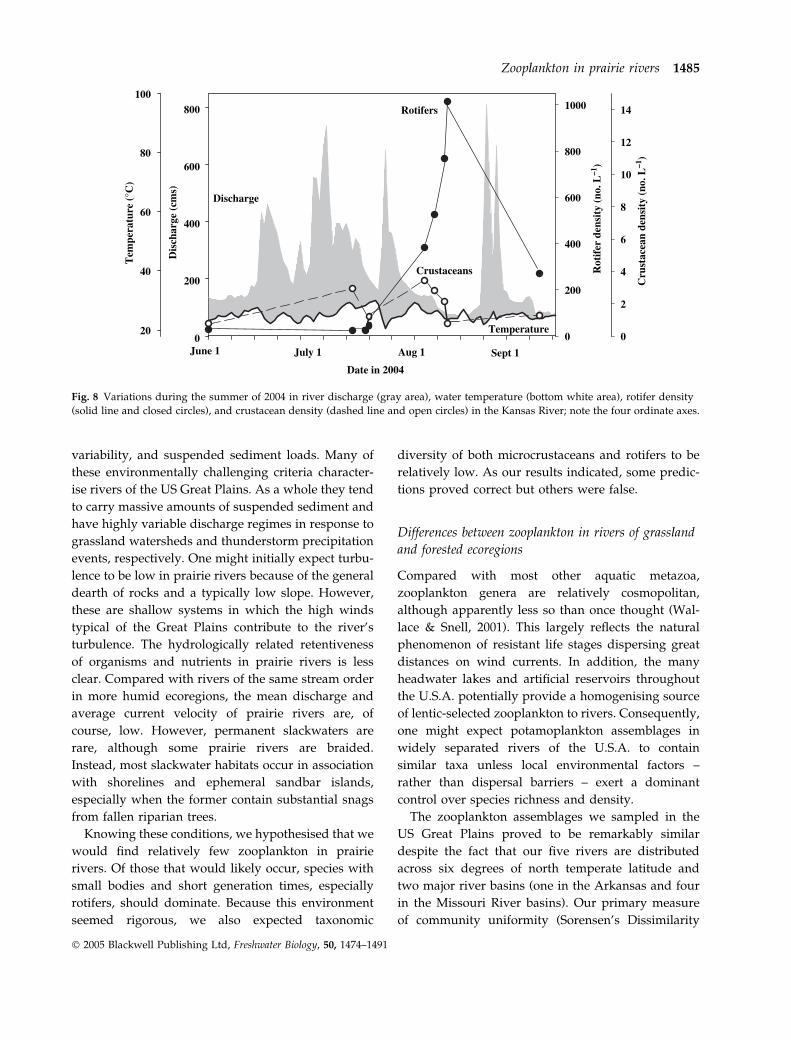
teristics (Fig. 8). Crustacean densities were inversely
related to current velocities in a given habitat
throughout the summer (Fig. 7b; d.f. ¼ 23, R2 ¼0.6205, P < 0.05), and densities stayed relatively
constant and very low throughout this period. In
contrast, rotifer densities were not significantly
related to current velocities when data from the entire
summer sample period were analysed (d.f. ¼ 23,
P > 0.05), but their densities were an order of magni-
tude higher in August to September when discharge
was less. Effects of current velocity on rotifer num-
bers were only evident during the high discharge
month of July (Fig. 7a) when a significant negative
linear regression was present (d.f. ¼ 8, R2 ¼ 0.4725,
P < 0.05). Fluctuations in water temperatures during
the sample period were not correlated with changes in
zooplankton densities (Fig. 8).
Discussion
Research on freshwater zooplankton ecology has so
heavily emphasised lentic habitats that the ecology of
lotic zooplankton almost seems a footnote in a
literature review. Nonetheless, the few investigators
around the world who have studied the role of
potamoplankton have increasingly demonstrated the
importance of zooplankton not only to sustenance of
most larval fish and a few adult species (e.g. Meng &
Orsi, 1991) but also to biotic components of carbon
cycling within large riverine ecosystems in particular
(e.g. Gosselain et al., 1998; Pace, Findlay & Fischer,
1998; Gliwicz, 2002; Thorp & Delong, 2002).
Aside from natural and polluted systems with
extreme chemical or thermal conditions, the most
rigorous lotic habitats for plankton should be in rivers
with rapid downstream transport (i.e. minor hydro-
logical retention areas) and high turbulence, discharge
10 100 1000 10000
Rot
ifer
den
sity
(no
. L–1
)
0
20
40
60
80
100
120
140Rotifer densities in 7 rivers
Linear regression
(a)
y = –25.67x + 128.72
R2 = 0.3362
Discharge [log (10) cm]
10 100 1000 10000
Cru
stac
ean
dens
ity
(no.
L–1
)
0
10
20
30
40
50Crustacean densities in 7 riversLinear regression
(b)
y = 16.764x – 30.27
R2 = 0.498
Fig. 6 Effects of discharge (averaged over many decades; note
log scale) on densities of rotifers (a) and crustaceans (b) in seven
rivers.
0.0 0.1 0.2 0.3 0.4 0.5
Rot
ifer
den
sity
(no
. L–1
)
0
10
20
30
40
50Rotifer density in the Kansas river (July only) Linear regression
(a)
y = –55.016x + 34.19
R2 = 0.4725
Current velocity (ms–1)
0.0 0.1 0.2 0.3 0.4 0.5
Cru
stac
ean
dens
ity
(no.
L–1
)
0.0
0.1
0.2
0.3
0.4
0.5 Crustacean density in the Kansas river (July – Sept)
Linear regression
(b)
y = –0.36x + 0.19
R2 = 0.1778
Fig. 7 Effects of current velocity in the Kansas River on rotifers
in July (a) and crustaceans throughout the summer sampling
period of 2004 (b).
1484 J.H. Thorp and S. Mantovani
� 2005 Blackwell Publishing Ltd, Freshwater Biology, 50, 1474–1491
variability, and suspended sediment loads. Many of
these environmentally challenging criteria character-
ise rivers of the US Great Plains. As a whole they tend
to carry massive amounts of suspended sediment and
have highly variable discharge regimes in response to
grassland watersheds and thunderstorm precipitation
events, respectively. One might initially expect turbu-
lence to be low in prairie rivers because of the general
dearth of rocks and a typically low slope. However,
these are shallow systems in which the high winds
typical of the Great Plains contribute to the river’s
turbulence. The hydrologically related retentiveness
of organisms and nutrients in prairie rivers is less
clear. Compared with rivers of the same stream order
in more humid ecoregions, the mean discharge and
average current velocity of prairie rivers are, of
course, low. However, permanent slackwaters are
rare, although some prairie rivers are braided.
Instead, most slackwater habitats occur in association
with shorelines and ephemeral sandbar islands,
especially when the former contain substantial snags
from fallen riparian trees.
Knowing these conditions, we hypothesised that we
would find relatively few zooplankton in prairie
rivers. Of those that would likely occur, species with
small bodies and short generation times, especially
rotifers, should dominate. Because this environment
seemed rigorous, we also expected taxonomic
diversity of both microcrustaceans and rotifers to be
relatively low. As our results indicated, some predic-
tions proved correct but others were false.
Differences between zooplankton in rivers of grassland
and forested ecoregions
Compared with most other aquatic metazoa,
zooplankton genera are relatively cosmopolitan,
although apparently less so than once thought (Wal-
lace & Snell, 2001). This largely reflects the natural
phenomenon of resistant life stages dispersing great
distances on wind currents. In addition, the many
headwater lakes and artificial reservoirs throughout
the U.S.A. potentially provide a homogenising source
of lentic-selected zooplankton to rivers. Consequently,
one might expect potamoplankton assemblages in
widely separated rivers of the U.S.A. to contain
similar taxa unless local environmental factors –
rather than dispersal barriers – exert a dominant
control over species richness and density.
The zooplankton assemblages we sampled in the
US Great Plains proved to be remarkably similar
despite the fact that our five rivers are distributed
across six degrees of north temperate latitude and
two major river basins (one in the Arkansas and four
in the Missouri River basins). Our primary measure
of community uniformity (Sorensen’s Dissimilarity
Date in 2004
nJ nuJ nuJ nuJ nuJ lJ luJ luJ luJ guA guA guA guA guA eS peS
Dis
char
ge (
cms)
0
200
400
600
800
Tem
pera
ture
(°C
)
20
40
60
80
100
Cru
stac
ean
dens
ity
(no.
L–1
)
Rot
ifer
den
sity
(no
. L–1
)
0
2
4
6
8
10
12
14
0
200
400
600
800
1000
Discharge
Temperature
Rotifers
Crustaceans
June 1u July 1 Aug 1 Sept 1
Fig. 8 Variations during the summer of 2004 in river discharge (gray area), water temperature (bottom white area), rotifer density
(solid line and closed circles), and crustacean density (dashed line and open circles) in the Kansas River; note the four ordinate axes.
Zooplankton in prairie rivers 1485
� 2005 Blackwell Publishing Ltd, Freshwater Biology, 50, 1474–1491

Index) demonstrated unusually high similarity in
species composition (mean ¼ 0.07 on a scale from 0
to 1, with low values equal to high similarity).
Moreover, few differences were evident in species
diversity (Shannon Index), although half the compar-
isons of evenness (Pielou Index) revealed significant
differences.
Our prediction of the dominance of small zooplank-
ton in these rivers was verified. Microcrustaceans
(copepods and a few cladocera) were present in all
prairie rivers, but each river was dominated very
strongly by rotifers (99% numerically).
Predictions of relatively low zooplankton density in
grassland rivers proved incorrect however. Overall
zooplankton densities were on par with those in the
Ohio and St Lawrence Rivers, exceeding them in some
cases but trailing in others. A next research step
would be to compare secondary production of
zooplankton between prairie and forested rivers.
The zooplankton production in grassland rivers is
based on many small zooplankton (rotifers) with short
generation times, while zooplankton communities of
rivers in forested ecoregions have greater numbers of
large zooplankton with concomitant longer turnover
times. Environmental differences in thermal regime,
relative food availability, and predation could also
alter production comparisons among river types.
While we found minor differences among prairie
rivers, major differences existed between grassland
rivers as a whole and the two forested-basin rivers.
The most stark differences between those seven rivers
concerned the relative abundances of microcrusta-
ceans and rotifers. Zooplankton assemblages in the
two eastern rivers differed considerably from each
other, but they contained approximately 20–80 times
more microcrustaceans per litre than did prairie
rivers. Moreover, the ratio of microcrustacean density
to rotifer density was approximately 32–64 times
higher in the Ohio and St Lawrence Rivers compared
with prairie rivers. Every prairie river we studied
contained higher absolute densities of rotifers than the
eastern rivers, and the average difference was 2.5
times greater. In fact, rotifers constituted on average
99.46% of the metazoan zooplankton in these five
rivers. While 19 rotifer genera were recorded for the
Ohio and St Lawrence Rivers combined and only 11
genera were identified from the five prairie rivers,
the taxonomic evenness of rotifers in grassland rivers
was much higher. The eastern forested rivers were
dominated by very few genera (mostly Polyarthra at
41–73%), while nearly 80% of the rotifers in prairie
rivers were in five genera of roughly equal abun-
dance. Rotifers are typically the most abundant
zooplankton in rivers (e.g. Pace, Findlay & Lints,
1992; Thorp et al., 1994; Kobayashi et al., 1996; Kim &
Joo, 2000), but the extreme dominance of rotifers, as
shown in prairie rivers, is unusual.
Environmental regulation of zooplankton in prairie
rivers
Recognising patterns of zooplankton density and
diversity is relatively easy, but identifying proximate
and ultimate causes of these patterns is a more
arduous task. While some measure of biotic control
of potamoplankton has been demonstrated in the
Ohio and St Lawrence Rivers (Jack & Thorp, 2000,
2002; Thorp & Casper, 2002, 2003), no studies of biotic
control have been published for prairie rivers. Data
from the present study suggests, however, that abiotic
factors may strongly influence zooplankton commu-
nities in prairie rivers.
Five abiotic environmental factors should be espe-
cially important to lotic zooplankton: (i) turbidity
(especially from suspended sediment); (ii) water
turbulence; (iii) hydrological retention, which is
influenced by stream discharge and access to shel-
tered, low velocity sites (slackwaters); (iv) thermal
conditions; and (v) ultraviolet radiation.
Turbidity. Our data seem initially to imply a complex
relationships between turbidity and densities of rot-
ifers and microcrustaceans, but we suspect that a
simpler interaction is merely being altered by con-
comitant changes in both competition and predation.
When we compared a river’s average turbidity over
many years with recent zooplankton sampling data,
we found rotifers fared better and microcrustaceans
did worse in turbid rivers, such as those in the US
Great Plains. This is consistent with laboratory find-
ings (e.g. Kirk & Gilbert, 1990) that suspended clay
reduces population growth rates of cladocera much
more than it affects rotifers. It is also consistent with
findings in various field studies around the world
(e.g. Shiel, 1985; Pace et al., 1992; Thorp et al., 1994). In
contrast, densities of rotifers in the turbid Kansas
River rose by an order of magnitude from July 2004,
when turbidity and discharge were high, during
1486 J.H. Thorp and S. Mantovani
� 2005 Blackwell Publishing Ltd, Freshwater Biology, 50, 1474–1491

August to September, when the opposite conditions
prevailed. The answer to this apparent conundrum
may relate to biotic interactions. We suggest that two
direct effects of suspended sediments are to reduce
both overall zooplankton densities and the number of
species that can successfully colonise turbid environ-
ments once they disperse to them. This could account
for, or contribute to, the low numbers of microcrus-
taceans in prairie rivers. However, two countervailing
indirect effects of suspended sediments for rotifers are
a reduction in densities of cladocera, which are often
superior food competitors (Kirk & Gilbert, 1990), and
a decrease in both visually hunting fish planktivores
(cf. McCabe & O’Brien, 1983; Cuker, 1993) and
predatory cyclopoid copepods. A decrease in cyclo-
poids was linked experimentally to increased densi-
ties of rotifers in the St Lawrence River (Thorp &
Casper, 2003). Hence, rotifers probably do better in
turbid rivers not because this environmental condi-
tion favours them but rather because pernicious
effects of competition and predation are partially
alleviated by high suspended sediment loads.
Hydrological retention. Species diversity and density
vary significantly with current velocity throughout
river networks in general and are positively correlated
with hydrological retention within the riverscape of
larger rivers, except where taxa are restricted by other
abiotic environmental conditions (e.g. oxygen, tem-
perature, substrate type) (Thorp et al., 2005b). It is not
surprising, therefore, that lotic ecologists are increas-
ingly identifying hydrological retention in slackwaters
and floodplain lakes as a major factor influencing
potamoplankton production and diversity as well as
other structural (Thorp et al., 1994; Basu & Pick, 1996;
Reckendorfer et al., 1999; Schiemer et al., 2001) and
functional characteristics of rivers (Hein et al., 2005).
Based on these observations, we examined whe-
ther hydrological retention in prairie rivers played a
significant role in regulating zooplankton assem-
blages. Baranyi et al. (2002) found that rotifers
dominated the zooplankton community of periodic-
ally isolated channels of the River Danube during
periods of low to medium water age (i.e. periods of
isolation from the main channel) but gave way to
microcrustaceans with increasing water age. Could
this be true in prairie rivers where hydrological
retention results more from ephemeral sandbar
islands (Fig. 1b) than from the relatively permanent
slackwaters formed by forested islands in most other
studies?
Perhaps the most complex task in understanding
this relationship is interpreting interactions between
zooplankton densities and current velocity because
responses are influenced by proximate current velo-
cities, average current velocities in the river, and
recent patterns of discharge in the river. Mean river
discharge by itself was not a good predictor of
zooplankton densities in our study, but this hydro-
logical parameter must impinge on zooplankton
through current velocity, water depth, and turbu-
lence. Crustacean densities in the Kansas River were
positively related to hydrological retention through-
out the summer sample period, but rotifer densities
were significantly depressed by current velocities
only during July, when mean river discharge was
high. We hypothesise that ephemeral sandbars do
not provide sufficient hydrological retention in time
and space to sustain many if any viable populations
of microcrustaceans but that they are adequate to
help sustain growth of rotifer populations. This is
consistent with conclusions of other scientists that
rotifers require shorter water retention times in rivers
for somatic and reproductive growth than do micro-
crustaceans (e.g. Pace et al., 1992; Kobayashi, 1997).
The relatively importance of low hydrological retent-
ion versus high turbidities in affecting cladocera and
copepods is not known. We also hypothesise that
hydrological retention is relatively important to
rotifers in prairie rivers only when mean current
velocities in the river are high, making slackwaters
that much more valuable.
Although rotifer densities may not be strongly and
directly influenced by hydrological retention in the
Kansas River, this contrasts with concurrent studies
on benthic invertebrates (J. Kreft, S. Moore & J. Thorp,
unpublished data) and larval fish (S. Moore &
J. Thorp, unpublished data) which reveal strong
positive effects from hydrological retention. We sug-
gest that a complex system of channels and slackwa-
ters is directly beneficial to varying degrees to most
aquatic organisms in all rivers, but individual taxa
may suffer indirectly from concomitant increases in
competition and predation in these habitats.
Other abiotic factors. The shallow nature of prairie
rivers and their minimal canopy cover can produce
high water temperatures. During July to September
Zooplankton in prairie rivers 1487
� 2005 Blackwell Publishing Ltd, Freshwater Biology, 50, 1474–1491

habitat analysis in the Kansas River, the average water
temperature was 24.1 �C (range ¼ 16.4–31.3 �C); this
was close to a 30-year average for May to September of
24.5 �C (range ¼ 12–34 �C). It is difficult to obtain
comparable data from different rivers because sample
depth and period of record vary considerably (but see
data from USGS web sites and chapters in Benke &
Cushing, 2005). However, it appears that our five
prairie rivers have annual temperatures within about
1 �C of each other. The annual temperature of the much
deeper Ohio River seems to be no more than approxi-
mately 1–2 �C higher than our five prairie rivers; and in
shallow waters, its temperatures can occasionally
exceed 30 �C during the summer (Thorp, Alexander
& Cobbs, 2002). On the opposite extreme, the annual
temperature of the St Lawrence River is roughly 2 �Ccooler than our prairie rivers, and temperatures rarely
exceed 22 �C. The presence of microcrustaceans in the
slightly warmer Ohio River and slightly cooler St
Lawrence River and their occurrence in reservoirs
throughout the latitudinal range of the conterminous
U.S.A. suggest that water temperatures are not the
primary factor in prairie rivers contributing to low
numbers of microcrustaceans, especially cladocera.
However, warm temperatures in shallow habitats of
prairie rivers may enhance or diminish secondary
production of rotifers depending on the season.
Net primary productivity in rivers is limited in part
by the amount of time phytoplankton are swept below
the photic zone by the helical, downstream circulation
patterns of rivers and by the amount of bottom
surface area available for benthic microalgal produc-
tion. At least the former influences riverine zooplank-
ton production (Pace et al., 1992). High suspended
sediment loads decrease penetration of both photo-
synthetically active radiation (PAR) and ultraviolet
radiation (UVR), but the photons need only penetrate
a short distance to reach the bottom of prairie rivers
during much of the year. Rotifers, especially loricate
genera such as Keratella, are considered to be more
tolerant of UVR than microcrustaceans (Leech &
Williamson, 2000), but additional research is needed
to determine whether this factor significantly con-
tributes to the unusually high ratio of rotifers to
microcrustaceans in prairie rivers. Turbidity limita-
tions to total net autotrophic production and inter-
ference with feeding mechanisms in particular may
also contribute to the rarity of copepods and cladocera
in grassland rivers.
We lack sufficient information on wind-driven
turbulence in prairie rivers to begin evaluation of its
effects on zooplankton assemblages.
Biotic versus abiotic control of prairie river zooplankton
Although a widespread consensus exists that zoo-
plankton are regulated more by physical than biotic
factors (e.g. Hynes, 1970; Baranyi et al., 2002), this
conclusion is not founded on experimental evidence.
Although stochastic factors are probably more import-
ant overall and throughout the river network, signi-
ficant competitive and predator–prey interactions
involving zooplankton are likely to occur in lateral
slackwater habitats and in the main channel in areas
or times of minimal hydraulic stress (Thorp & Casper,
2003; Thorp et al., 2005b). In prairie rivers, the rarity of
competitive cladocera and predatory cyclopoid cope-
pods could contribute to the higher densities and
relative abundance of rotifers in comparison to rivers
we studied in eastern, forested ecoregions.
Acknowledgments
We appreciate the help of Stephanie Moore, Scott
Campbell, Leila Desotelle, and numerous undergrad-
uates (especially Jim Kreft, Uriah Price, and Jason
Robertson) in collecting zooplankton from five prairie
rivers in 2003–2004 and the help of Andy Casper in
collecting plankton from the St Lawrence and Ohio
Rivers. We are extremely grateful to Debbie Guelda
(with field assistance from Rick Koch and Tim Sellers)
for additional data on zooplankton from the Ohio
River, and we thank Edward Peters for environmental
data on the Platte River. Comments on an earlier draft
by Mike Delong, Jeff Jack, and two anonymous
reviewers improved the final manuscript. Collection
and processing of samples was supported both by a
S.T.A.R. grant to J. Thorp from the US Environmental
Protection Agency (RD-83059701) and by travel and
support for S. Mantovani from the Department of
Biology, University of Ferrara.
References
Balcer M.D., Korda N.L. & Dodson S.I. (1984) Zooplankton
of the Great Lakes: A guide to the identification and ecology
of the common crustacean species. University of Wiscon-
sin Press, Madison, WI, U.S.A.
1488 J.H. Thorp and S. Mantovani
� 2005 Blackwell Publishing Ltd, Freshwater Biology, 50, 1474–1491

Baranyi C., Hein T., Holarek C., Keckeis S. & Schiemer F.
(2002) Zooplankton biomass and community structure
in a Danube River floodplain system: effects of
hydrology. Freshwater Biology, 47, 473–482.
Basu B.K. & Pick F.R. (1996) Factors regulating phyto-
plankton and zooplankton biomass in temperate
rivers. Limnology and Oceanography, 41, 1572–1577.
Benke A.C. & Cushing C.E. (Eds) (2005) Rivers of North
America. Academic Press, San Diego, CA.
Clarke K.R. (1993) Non-parametric multivariate analysis
of changes in community structure. Australian Journal
of Ecology, 18, 117–143.
Cross F.B. & Collins J.T. (1995) Fishes in Kansas, 2nd edn.
University of Kansas Natural History Museum, Lawr-
ence, KS, U.S.A.
Cuker B.E. (1993) Suspended clays alter trophic interac-
tions in the plankton. Ecology, 74, 944–953.
Dodds W.K. (2002) Freshwater Ecology: Concepts and Envi-
ronmental Applications. Academic Press, San Diego, CA,
569 pp.
Dodds W.K., Gido K., Whiles M.R., Fritz K.M. &
Matthews M.J. (2004) Life on the edge: the ecology of
Great Plains prairie streams. BioScience, 54, 207–218.
Gleick P.H. (1993) Water in Crisis. A Guide to the World’s
FreshWaterResources. OxfordUniversity Press, New York.
Gliwicz Z.M. (2002) On the different nature of top-down
and bottom-up effects in pelagic food webs. Freshwater
Biology, 47, 2296–2312.
Gosselain V., Descy J.-P., Viroux L., Joaquim-Justo C.,
Hammer A., Metens A. & Schweitzer S. (1998) Grazing
by large river zooplankton? The case of the Meuse and
Moselle rivers in 1994 and 1995. Hydrobiologia, 369,
199–216.
Hein T., Reckendorfer W., Thorp J. & Schiemer F. (2005)
The role of slackwater areas and the hydrologic
exchange for biogeochemical processes in river corri-
dors: examples from the Austrian Danube. Archiv fur
Hydrobiologie, Suppl. 155 (Large Rivers 15), 425–442.
Hynes H.B.N. (1970) The Ecology of Running Waters.
University of Toronto Press, Toronto, 555 pp.
Jack J.D. & Thorp J.H. (2000) Effects of the benthic
suspension feeder Dreissena polymorpha on zooplank-
ton in a large river. Freshwater Biology, 44, 569–579.
Jack J.D. & Thorp J.H. (2002) Impacts of fish predation on
an Ohio River zooplankton community. Journal of
Plankton Research, 24, 119–127.
Kim H.-Y. & Joo G.-J. (2000) The longitudinal distribution
and community dynamics of zooplankton in a regu-
lated large river: a case study of the Nakdong River
(Korea). Hydrobiologia, 438, 171–184.
Kirk K.L. & Gilbert J.J. (1990) Suspended clay and the
population dynamics of planktonic rotifers and clado-
cerans. Ecology, 71, 1741–1755.
Kobayashi T. (1997) Associations between environmental
variables and zooplankton body masses in a regulated
Australian river. Marine and Freshwater Research, 48,
523–529.
Kobayashi T., Gibbs P., Dixon P.I. & Shiel R.J. (1996)
Grazing by a river zooplankton community: impor-
tance of microzooplankton. Marine and Freshwater
Research, 47, 1025–1036.
Leech D.M. & Williamson C.E. (2000) Is tolerance to UV
radiation in zooplankton related to body size, taxon, or
lake transparency?EcologicalApplications, 10, 1530–1540.
Magurran A.E. (1988) Ecological Diversity and Its Measure-
ment. Princeton University Press, New Jersey, 179 pp.
McCabe G.D. & O’Brien W.J. (1983) The effects of
suspended silt on feeding and reproduction of Daphnia
pulex. American Midland Naturalist, 110, 324–337.
Meng L. & Orsi J.J. (1991) Selective predation by larval
striped bass on native and introduced copepods.
Transactions of the American Fisheries Society, 120, 187–
192.
Pace M.L., Findlay S.E.G. & Fischer D. (1998) Effects of an
invasive bivalve on the zooplankton community of the
Hudson River. Freshwater Biology, 39, 103–116.
Pace M.L., Findlay S.E.G. & Lints D. (1992) Zooplankton
in advective environments: the Hudson River commu-
nity and a comparative analysis. Canadian Journal of
Fisheries and Aquatic Sciences, 49, 1060–1069.
Reckendorfer W., Keckeis H., Winkler G. & Schiemer F.
(1999) Zooplankton abundance in the River Danube,
Austria: the significance of inshore retention. Fresh-
water Biology, 41, 583–591.
Schiemer F., Keckeis H., Reckendorfer W. & Winkler G.
(2001) The ‘inshore retention concept’ and its signifi-
cance for large rivers. Archiv fur Hydrobiologie, Supple-
ment 135, Large Rivers, 12, 509–516.
Shiel R.J. (1985) Zooplankton of the Darling River system,
Australia. Verhandlungen. Internationale Vereinigung
Limnologie, 22, 2136–2140.
Stemberger R.S. (1979) A Guide to the Rotifers of the
Laurentian Great Lakes. EPA-600/4–79–021. US Environ-
mental Protection Agency, Cincinnati, OH, U.S.A.
Thorp J.H. & Casper A.F. (2002) Potential effects on
zooplankton from species shifts in mussel planktivory:
a field experiment in the St. Lawrence River. Freshwater
Biology, 47, 107–119.
Thorp J.H. & Casper A.F. (2003) Importance of biotic
interactions in large rivers: an experiment with
planktivorous fish, dreissenid mussels, and zooplank-
ton in the St. Lawrence. River Research and Applications,
19, 265–279.
Thorp J.H. & Covich A.P. (Eds) (2001) Ecology and
Classification of North American Freshwater Invertebrates.
2nd edn. Academic Press, San Diego, U.S.A.
Zooplankton in prairie rivers 1489
� 2005 Blackwell Publishing Ltd, Freshwater Biology, 50, 1474–1491
Thorp J.H. & Delong M.D. (2002) Dominance of
autochthonous autotrophic carbon in food webs of
heterotrophic rivers? Oikos, 96, 543–550.
Thorp J.H., Alexander J.E. Jr & Cobbs G.A. (2002) Coping
with warmer, large rivers: a field experiment on
potential range expansion of northern quagga mussels
(Dreissena bugensis). Freshwater Biology, 47, 1779–1790.
Thorp J.H., Lamberti G.A. & Casper A.F. (2005a)
Chapter. 21. St. Lawrence River. In: Rivers of North
America (Eds A.C. Benke & C.E. Cushing), pp. 983–
1028. Academic Press, San Diego, CA.
Thorp J.H., Thoms M.C. & Delong M.D. (2005b) The
riverine ecosystem synthesis: biocomplexity in river
networks across space and time. River Research and
Applications (in press).
Thorp J.H., Black A.R., Haag K.H. & Wehr J.D. (1994)
Zooplankton assemblages in the Ohio River: sea-
sonal, tributary, and navigation dam effects.
Canadian Journal of Fisheries and Aquatic Sciences, 51,
1634–1643.
Wallace R.L. & Snell T.W. (2001) Phylum Rotifera. In:
Ecology and Classification of North American Freshwater
Invertebrates, 2nd edn (Eds J.H. Thorp & A.P.
Covich), pp. 195–254. Academic Press, San Diego,
U.S.A.
Young F.W. & Hamer R.M. (1987) Multidimensional
Scaling: History, Theory and Applications. Erlbaum,
New York.
(Manuscript accepted 16 June 2005)
Appendix 1 Mean densities (no. L)1) of zooplankton genera collected from seven rivers
Taxa/Rivers AR KA PL EL NI OH SL
Branchiopoda
Acroperus 0.00 0.00 0.00 0.00 0.00 0.00 0.28
Alona 0.00 0.00 0.00 0.00 0.00 0.00 0.02
Bosmina 0.00 0.02 0.14 0.00 0.05 7.52 2.39
Bosminopsis 0.00 0.00 0.00 0.00 0.00 0.01 0.00
Ceriodaphnia 0.00 0.00 0.00 0.00 0.00 4.83 0.38
Chydorus 0.00 0.00 0.00 0.00 0.00 0.04 0.18
Daphnia 0.00 0.00 0.00 0.00 0.00 0.89 2.10
Diaphanosoma 0.00 0.40 0.21 0.00 0.07 4.83 0.03
Eubosmina 0.00 0.00 0.00 0.00 0.00 0.00 0.13
Holopedium 0.00 0.00 0.00 0.00 0.00 0.00 0.06
Polyphemus 0.00 0.00 0.00 0.00 0.00 0.00 0.26
Sida 0.00 0.00 0.00 0.00 0.00 0.00 0.05
Total cladocera 0.00 0.42 0.35 0.00 0.12 18.10 5.88
Copepoda
Calanoids (adults + copepodids) 0.01 0.02 0.04 0.01 0.01 0.68 0.63
Eurytemora 0.00 0.00 0.00 0.00 0.00 * 0.16
Leptodiaptomus 0.01 0.08 0.03 0.00 0.01 * 0.01
Cyclopoids (adults + copepodids) 0.01 0.12 0.28 0.02 0.16 20.00 1.11
Acanthocyclops 0.00 0.00 0.00 0.00 0.00 * 0.06
Diacyclops 0.01 0.06 0.17 0.01 0.00 * 0.25
Mesocyclops 0.00 0.01 0.00 0.00 0.00 * 0.01
Harpacticoids (adults + copepodids) 0.00 0.00 0.00 0.00 0.00 * 0.22
Nauplii 0.31 0.56 0.56 0.09 0.54 19.78 6.11
Total copepods 0.35 0.70 0.88 0.13 0.72 40.46 8.07
Rotifera
Ascomorpha 0.00 0.00 0.00 0.00 0.00 0.83 0.00
Asplanchna 0.00 0.00 0.13 0.00 0.00 0.99 0.11
Brachionus 4.13 0.44 16.38 4.25 1.63 7.43 0.00
Conchiloides 0.00 0.00 0.00 0.00 0.00 0.00 0.01
Encentrum 0.00 0.00 0.00 0.00 0.00 0.00 0.11
Euchlanis 12.38 1.56 0.63 3.13 0.50 0.00 0.64
Filinia 1.75 1.56 17.25 16.88 0.13 0.27 0.00
Gastropus 8.88 6.06 6.25 3.38 3.38 0.00 0.00
Kelicottia 0.00 0.00 0.00 0.00 0.00 0.01 0.00
Keratella 1.00 2.38 1.00 0.38 1.25 4.48 1.38
Lecane 0.00 0.00 0.00 0.00 0.00 0.21 0.04
1490 J.H. Thorp and S. Mantovani
� 2005 Blackwell Publishing Ltd, Freshwater Biology, 50, 1474–1491

Appendix 1 (Continued)
Taxa/Rivers AR KA PL EL NI OH SL
Lepadella 0.00 0.00 0.00 0.00 0.00 0.00 0.54
Monostyla 30.50 6.75 3.88 4.00 9.88 0.00 0.89
Notholca 10.25 6.19 44.50 12.50 1.50 0.06 0.06
Platyias 0.00 0.00 0.00 0.00 0.00 0.02 0.00
Ploesoma 7.38 1.44 21.00 0.00 1.75 2.38 0.08
Polyarthra 0.00 0.00 0.00 0.00 0.00 13.73 21.43
Proales 9.75 3.13 8.13 19.63 1.50 0.00 0.00
Synchaeta 0.00 0.00 0.00 0.00 0.00 3.02 3.41
Trichocerca 6.50 4.69 9.63 12.75 20.88 0.23 0.69
Trichotria 0.00 0.00 0.00 0.00 0.00 0.00 0.07
Total rotifers 92.50 34.38 128.75 101.63 42.38 33.65 29.46
*Taxa may be present but not tabulated in this data set. Based on other data sets, Eurytemora and Diacyclops thomasi were the dominant
calanoid and cyclopoid copepods in the Ohio River.
Mesocyclops was also common in the Ohio River. Other genera present at <0.01 animals per litre in all rivers were not included here.
Rivers: AR, KA, PL, EL, NI, OH, and SL ¼ Arkansas, Kansas, Platte, Elkhorn, Niobrara, Ohio, and St Lawrence Rivers, respectively.
Zooplankton in prairie rivers 1491
� 2005 Blackwell Publishing Ltd, Freshwater Biology, 50, 1474–1491