a comparative study on simultaneous … comparative study on simultaneous saccharification and...
TRANSCRIPT

Chiang Mai J. Sci. 2013; 40(3) 307
Chiang Mai J. Sci. 2013; 40(3) : 307-320http://it.science.cmu.ac.th/ejournal/Contributed Paper
A Comparative Study on SimultaneousSaccharification and Fermentation of AgriculturalWastes to Bio-ethanol Using Two SaccharomycesStrainsPoludasu Rama Mohan [a], Sake Pradeep [b], and Obulam Vijaya Sarathi Reddy*[a][a] Department of Biochemistry, Sri Venkateswara University, Tirupati - 517 502, India.[b] Department of Microbiology, Yogi Vemana University, Kadapa - 516 003, India.*Author for correspondence; e-mail: [email protected]
Received: 23 February 2012Accepted:20 September 2012
ABSTRACTApplication of simultaneous saccharification and fermentation (SSF) technique for
the conversion of agricultural wastes to bio-ethanol would result in a more cost-effectiveprocess. Peracetic acid (PAA) pretreated various lignocellulosic substrates like sugarcane bagasse,rice straw and corn cobs were examined with solka floc as reference in SSF experiments forthe production of bio-ethanol using Asperigillus niger crude cellulase, Cladosporium sp. crudeCMCase (Carboxy Methyl Cellulase) and yeast cells. Fermentation temperature was selected as40 and 43°C for Saccharomyces cerevisiae NCIM 3215 and Saccharomyces bayanus respectively.Further SSF experiments were compared for ethanol production. The S. bayanus was foundto perform better and faster fermentation in the SSF process and yielded higher bio-ethanolin pretreated bagasse just after 48 h (17.03% w/v) than the S. cerevisiae that gave high yields ofbio-ethanol only after 72 h (13.44% w/v). Increased bio-ethanol yields were obtained whenthe cellulase was supplemented with CMCase of Cladosporium sp. The cell viability was lower inS. bayanus compared with S. cerevisiae. The bio-ethanol yields of the fermented substrates werein the following order: Solka floc> treated sugarcane bagasse> treated rice straw > treatedcorn cobs.
Keywords: simultaneous saccharification and fermentation (SSF), Cladosporium sp.,Asperigillus niger, lignocellulose, Saccharomyces cerevisiae, Saccharomyces bayanus,bio-ethanol
1. INTRODUCTIONProduction of renewable fuels, especially
bio-ethanol from lignocellulosics biomass,holds potential to meet the energy demandas well as to mitigate greenhouse gas emissionsfor a sustainable environment. At presenttechnologies to produce bio-ethanol largelydepend on sugarcane, starch based grains andtubers [1]. However, production of bio-
ethanol requires the development of aneconomically feasible and sustainable processutilizing renewable lignocellulosic materialsthat do not compete with food sources [2].
Agricultural residues (e.g., wheat straw,sugarcane bagasse, corn stover), forestproducts (hardwood and softwood), anddedicated crops (switchgrass, salix) are

308 Chiang Mai J. Sci. 2013; 40(3)
renewable sources of energy [3]. Theenzymatic hydrolysis of sugarcane bagassecould be greatly enhanced by peracetic acid(PAA) pretreatment [4], which was achievedmainly by delignification accompanyingwith the increase of the surface area andexposure of cellulose fibers [5].
Zayed and Meyer [6] carried out singlebatch bioconversion of wheat straw toethanol by employing the fungus Trichodermaviride and the yeast Pachysolen tannophilus.Miyomoto [7] used raw material likesugarcane bagasse, rice straw, and forestwastes in pilot scale unit for the productionof ethanol. Nakamura et al., [8] reported thatrice straw is a potential feed stock forproduction of power and fuel ethanol. Laseret al., [9] used sugarcane bagasse forbioconversion to ethanol.
During enzymatic hydrolysis, cellulaseactivity is severely inhibited by cellobioseand to a lesser extent by glucose [10]. Thecompetitive product inhibition of thecellobiose can be overcome to some extentby the addition of a surplus of β-glucosidaseactivity [11]. The cellulose fraction oflignocelluloses can be converted tobio-ethanol by either simultaneoussaccharification and fermentation (SSF) orseparate enzymatic hydrolysis andfermentation (SHF) processes. SSF is morefavoured because of its low potential costs[12]. Large number of publicationsindicated that the optimum temperature forenzymatic hydrolysis is at 40-50oC [13],while the microorganisms with goodethanol productivity and yield do notusually tolerate this high temperature.This problem has usually been tackled byapplying thermo-tolerant yeast [14] andS. bayanus [15,16].
Earlier we have reported the highgravity finger millet medium for ethanolproduction using S. bayanus [17]. Recently
we have reported the optimization of ethanolproduction from sugarcane bagasse [18].In the present study agricultural wasteswere considered as inexpensive, abundantand renewable sources for bio-ethanolproduction using S. cerevisiae and S.bayanus in SSF.
2. MATERIALS AND METHODS2.1 Fungal Strains and Their Maintenance
The Cladosporium sp. NCIM 901procured from National Collection ofIndustrial Microorganisms (NCIM), Pune,India was employed for CMCaseproduction. The Asperigillus niger soilisolate was employed for cellulase and β-glucosidase production. Both fungal strainswere grown and maintained on potatosoluble starch agar at 4oC as agar slantcultures. The actively growing fungalcultures were maintained as such byperiodic transfer into fresh potato dextroseagar (PDA) medium at every 2-3 months.
2.2 Preparation of Fungal InoculumBoth the fungal cultures were grown on
PDA slants at 32oC for 4 days and washedwith 5 ml sterile distilled water to prepare thespore suspension. Spores were filteredthrough Whatman No.1 filter paper andresuspended in sterile distilled water to get aspore density of 1 × 108 spores/ml with theaid of a Neubauer counting chamber. Theabove spore suspension was added (2 ml/g)as inoculum to SSF medium.
2.3 Solid State Fermentation for EnzymeProduction
It was carried individually in 250 mlErlenmeyer conical flasks that contained 1gmicro crystalline cellulose (solka floc) and 1gof carboxy methyl cellulose as carbon sourcefor A. niger and Cladosporium sp. NCIM 901respectively, and 5 ml of basal salt medium

Chiang Mai J. Sci. 2013; 40(3) 309
of following composition (g/l) NH4NO3, 5.0;Peptone, 5; NaCl, 5; Urea, 2; CaCl2,1; Tween80, 0.5 ml and trace elements: FeSO4⋅7H2O,0.005; MnSO4⋅7H2O, 0.001; ZnSO4⋅7H2O,0.001 and CoCl2,0.002 g. The initial moisturecontent of the medium was adjusted to70-75%. Prior to sterilization, the initial pHof the medium was adjusted to 5.5. Afterincubation, 10 ml sodium citrate buffer(50 mM, pH 6.0) was added to flasks, andthese flasks were kept at 50oC for 1 h undermild stirring. The fermented slurry was filteredthrough muslin cloth and centrifuged at8000 × g for 15 min. The filtrate wasconcentrated using Amicon cell fitted with aPM-10 membrane (10 kDa cut off) andused for β-glucosidase purification andcarboxymethyl cellulase purification [19].
2.4 Crude Enzyme PreparationCulture filtrate of the crude preparation
was precipitated by the addition of ammo-nium sulphate with 90% saturation. It waskept at 4oC for 24 h and the precipitatewas collected by using centrifugation (4000× g) at 4oC for 30 min. The precipitate wasre-dissolved in 10 ml of 0.02 M sodiumphosphate buffer and dialysed for 24 hagainst 0.02 M sodium phosphate buffer,pH 7.0 containing 1 mM EDTA and 1 mM2-mercaptoethanol [20]. The dialyzedsuspension was used for SSF experiments asenzyme source.
2.5 Substrates and PretreatmentRice straw was collected from local
paddy fields around Tirupati (India). Corncobs were collected in agricultural market,Tirupati. Sugarcane bagasse was collectedat Sri Venkateswara sugar Industries,Renigunta, Tirupati. The peracetic acid(PAA) (5% wt/vol) pretreatment [18] of thesubstrates were carried out at 90oC for 60min. Solid-liquid ratio was maintained at
10 g/100 ml. Solka floc SW-40 (micro-crystalline cellulose) was employed as thesubstrate of reference. The solid and liquidhydrolyzate was separated and used forfurther experiments. The liquid hydrolysatewas removed and used for sugar contentanalysis by Yang and Wyman [21] method.
2.6 Yeast Strains and Preparation ofInoculum
The Saccharomyces cerevisiae (NCIM3215) obtained from National Collectionof Industrial Microorganisms (NCIM),Pune, India and Saccharomyces bayanus waskindly provided by Dr.Roberto Ambrosoli,University of Turin, Italy. Both strainswere used in this study and the inoculumwas prepared by transferring the organismsmaintained on MGYP medium (malt extract,3; glucose, 10; yeast extract, 5; peptone, 5g/l) into 250 ml flask with 100 ml basalmedium having 36 g/l glucose. The yeastswere cultured at 33oC on an orbital shakerfor 12 h. The inoculum concentration wasabout 1.5 × 108 yeast cells/ml and theamount of inoculum added was 10% (v/v)of the SSF medium.
2.7 Simultaneous Saccharification andFermentation (SSF)
The reaction mixture contained PAApretreated substrate 10% (w/v), crudecellulase (16 FPU) and carboxymethylcellulase (15 IU), 10% (v/v) yeast inoculumand 87 ml of SSF basal medium [peptone(0.5%, w/v), yeast extract (0.3%, w/v), maltextract (0.3%, w/v) and Tween 80 (0.1%,w/v) supplemented with mineral salts (ing/l) (NH4)2SO4 1.4, KH2PO4 2.0, urea 0.3,CaCl2 2H2O 0.4, MgSO4 7H2O 0.3; and(in mg/l) FeSO4 7H2O 5.0, MnSO4 H2O 1.6,ZnSO4 7H2O 1.4, CoCl2 6H2O 3.7. The pHof the medium was adjusted to 4.8 using0.05 M acetate buffer] and final the volume

310 Chiang Mai J. Sci. 2013; 40(3)
made up to 100 ml [22]. The final pH wasadjusted to 5.1 with 0.05 M citrate buffer.Reactions were carried out in 250 mlErlenmeyer flasks with 100 ml workingvolume on an orbital shaker at 150 rpm.Samples were aseptically taken at regularintervals for the analysis of sugars, reducingsugars and ethanol content.
2.8 Assay MethodsThe components (cellulose, hemicellulose
and lignin) of pretreated and untreatedsubstrates were analyzed according to theprocedures of Goering and Van Soest [23].Total drymatter, moisture, ash content, crudefibre, crude fat and crude protein weredetermined by the procedures of AOAC [24].The amounts of total soluble sugar (TS) ofthe hydrolysates were tested according tophenol-sulfuric acid [25] method. Thereducing sugar concentration was estimatedusing 3, 5-dinitrosalicylic acid (DNS) method[26]. The xylose and arabinose content wasestimated by the method of Khabarov et al.,[27]. The reagents (solution 1, 2 and H2SO4)were prepared and mixed in test tubes.It was heated on a boiling water bath,the reaction mixture was transferred into a100 ml volumetric flask, and water wasadded to the mark. The absorption spectrawere recorded on a Spectrophotometer.The calibration plots were constructed andthe concentration of pentoses were expressedas mg/g.
Protein content was estimated bymethod of Lowry et al., [28]. Filter paperhydrolase (FPase) and carboxymethylcellulase (CMCase) activities were assayedas described by Mandels et al., [29] at pH5.0 and 50oC (60 min for FPase and 30 minfor CMCase). The β-glucosidase (EC3.2.1.21) activity was assayed according toDesrochers et al., [30] at pH 5.0 and 50oCfor 30 min. Glucose produced was assayed
by the method of Miller [26] using glucoseas the standard. Enzyme activity (U) wasexpressed either as μmol glucose (for β-glucosidase) or glucose equivalent (forFPase and CMCase) liberated per min.
2.9 Glucosamine DeterminationFor fungal chitin hydrolysis into N-acetyl
glucosamine, 20 mg dried biomass wasincubated with 2 ml of H2SO4 (72%) in atest tube. After continuous shaking (130rpm) for 60 min at 25°C, it was dilutedwith 3 ml of distilled water and autoclavedfor 2 h. The hydrolyzate was neutralizedto pH 7.0 with 10 M and then with 0.5 MNaOH and diluted to 100 ml. Finally,glucosamine was assayed by the colorime-tric method described by Tsuji et al., [31]and modified by Ride and Drysdale [32].Hydrolyzate (1 ml diluted) was mixed with1 ml of NaNO2 (5%) and 1ml of KHSO4
(5%) in a centrifuge tube. After shakingoccasionally for 15 min, it was centrifugedat 3500 rpm for 5 min; 2 ml of supernatantwas mixed with 0.67 ml of NH4SO3NH2
(12.5%) and shaken for 3 min. To themixture was added 0.67 ml of 3-methyl-2-benzothiazolinone hydrazone hydrochloride(MBTH; 0.5%, prepared daily), and thenthe mixture was boiled for 3 min andimmediately cooled to room temperature;0.67 ml of FeCl3 (0.5%, prepared within 3days) was added. After standing for 30 min,the absorbance at 650 nm was measuredspectrophotometrically. The chitin content(mg of glucosamine/mg of dry weight offungal biomass) was calculated in accordancewith a standard curve established bymeasuring the absorbance of knownamounts of glucosamine hydrochloride.
2.10 Yeast Cell ViabilityThe yeast cells viability was determined
by the methylene blue staining technique [33].
Chiang Mai J. Sci. 2013; 40(3) 311
A 200 μl sterile solution of methylene blue(3.3 mM in 68 mM sodium citrate) wasmixed with 200 μl of yeast suspension anddiluted to reach an OD of 0.4-0.7 at 620nm. This mixture was shaken and after 5min incubation, placed in a Neubauercounting chamber. The number of stained(inactive) and unstained (active) cells werecounted in five different fields with a totalof 200-300 cells and the percentage of viable(active) cells was noted.
2.11 Ethanol Quantification Using GasChromatography
The Agilent systems model 6890 wasused and the conditions were as follows:Graphitized packed column 5% carbowax20M phase, matrix 80/120 carbopack-B,length 6 ft (1.83m) × 2 mm I.D. × 1/4" O.D.Nitrogen was used as carrier gas with flowrate of 20 ml/min and eluted compoundswere detected by flame ionization detector(FID). Hydrogen was used as fuel gas, withflow rate 40 ml/min, along with air at a flowrate of 400 ml/min and secondary butylacetate was used as internal standard [34].
3. RESULTS AND DISCUSSION3.1 Proximate Analysis of LignocellulosicSubstrates
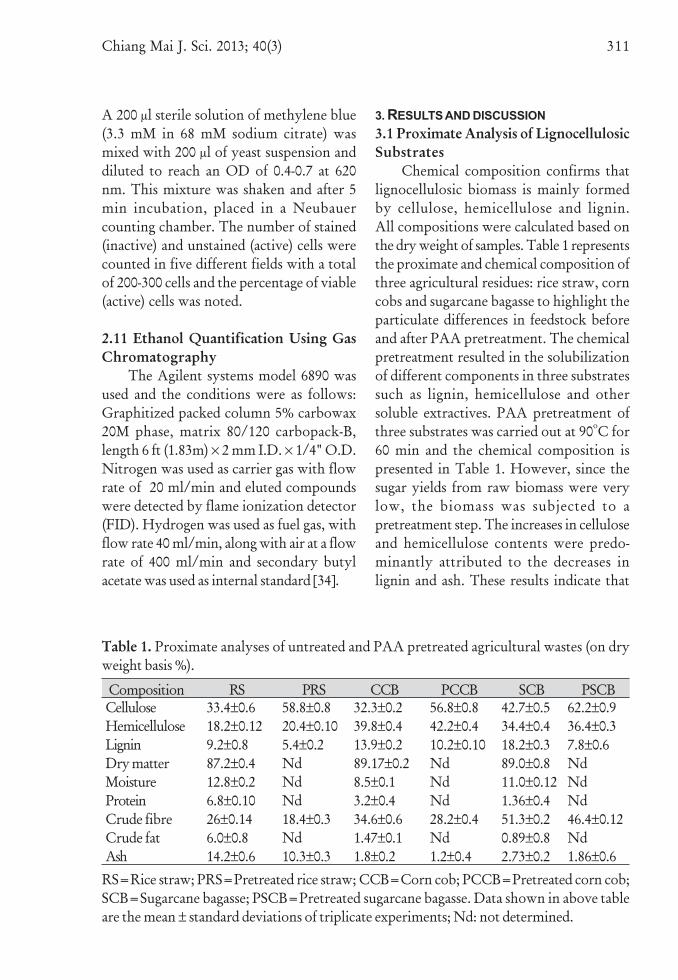
Chemical composition confirms thatlignocellulosic biomass is mainly formedby cellulose, hemicellulose and lignin.All compositions were calculated based onthe dry weight of samples. Table 1 representsthe proximate and chemical composition ofthree agricultural residues: rice straw, corncobs and sugarcane bagasse to highlight theparticulate differences in feedstock beforeand after PAA pretreatment. The chemicalpretreatment resulted in the solubilizationof different components in three substratessuch as lignin, hemicellulose and othersoluble extractives. PAA pretreatment ofthree substrates was carried out at 90oC for60 min and the chemical composition ispresented in Table 1. However, since thesugar yields from raw biomass were verylow, the biomass was subjected to apretreatment step. The increases in celluloseand hemicellulose contents were predo-minantly attributed to the decreases inlignin and ash. These results indicate that
Table 1. Proximate analyses of untreated and PAA pretreated agricultural wastes (on dryweight basis %).
RS=Rice straw; PRS=Pretreated rice straw; CCB=Corn cob; PCCB=Pretreated corn cob;SCB=Sugarcane bagasse; PSCB=Pretreated sugarcane bagasse. Data shown in above tableare the mean ± standard deviations of triplicate experiments; Nd: not determined.
Composition RS PRS CCB PCCB SCB PSCBCelluloseHemicelluloseLigninDry matterMoistureProteinCrude fibreCrude fatAsh
33.4±0.618.2±0.129.2±0.887.2±0.412.8±0.26.8±0.1026±0.146.0±0.814.2±0.6
58.8±0.820.4±0.105.4±0.2NdNdNd18.4±0.3Nd10.3±0.3
32.3±0.239.8±0.413.9±0.289.17±0.28.5±0.13.2±0.434.6±0.61.47±0.11.8±0.2
56.8±0.842.2±0.410.2±0.10NdNdNd28.2±0.4Nd1.2±0.4
42.7±0.534.4±0.418.2±0.389.0±0.811.0±0.121.36±0.451.3±0.20.89±0.82.73±0.2
62.2±0.936.4±0.37.8±0.6NdNdNd46.4±0.12Nd1.86±0.6

312 Chiang Mai J. Sci. 2013; 40(3)
PAA pretreatment could partially disrupt thelignin structure and expose more accessiblesurface area of cellulose to cellulase.Furthermore, lignin removal could also reduceunproductive binding of cellulase to lignin [35].Consequently, it improved enzymaticbiocatalysis and increased the yields of desiredproducts and recycled more cellulase. Thus,the costs associated with enzymaticsaccharification of biomass could remarkablybe reduced.
From the results it is evident that inall substrates after pretreatment there wasincrease in cellulose and hemicellulosecontents and decrease in lignin content.The cellulose content of rice straw, corn coband bagasse increased after pretreatmentfrom 33.4±0.6, 32.3±0.2 and 42.7±0.5%to 58.8±0.8, 56.8±0.8 and 62.2±0.9%,respectively. The hemicellulose content wasslightly increased in three substrates afterpretreatment. Thus, increased conversion ofcellulose would increase the available sugarcontent in the hydrolyzate. Generally thebagasse contained relatively higher lignincontent (18.2±0.3%) than that of corn cob(13.9±0.2%) and rice straw (9.2±0.8%).Bagasse also contains a relatively highamount of lignin (20%) comparable to thatin wheat straw [36]. In general ligninstrengthens the bagasse structure andinfluences its suspension texture in water.Moreover, lignin hydrophobic characterconsiderably reduces the bagasse’s mois-turizing ability [37].
After pretreatment, the level of thecrude fiber was significantly reduced in allthe studied samples. Compositional changesobserved in three substrates before and afterpretreatment clearly showed that cellulose,hemicellulose content increased and ligninas well as ash content decreased in allsubstrates. The increases in cellulose werepredominantly attributed to the decreases
in lignin and ash. The ash content wasdecreased in pretreated samples comparedto raw untreated samples.
3.2 Production of Enzymes by Solid StateFermentation
The degradation of crystalline cellulosewas carried out by synergistic action ofFPase, CMCase and β-glucosidase. It hasbeen reported that optimum time for thehighest cellulase activity depended uponthe substrate and fungus used [38]. In thepresent study, pure commercial micro-crystalline cellulose and carboxy methylcellulose were used to obtain crude cellulaseand carboxymethyl cellulase from A. nigerand Cladosporium sp. respectively, by solidstate fermentation. Initially both fungalstrains were produced low activities of theenzyme but with the course of time, therewas gradual increase in enzyme synthesisand attained maximum activity on the 6th
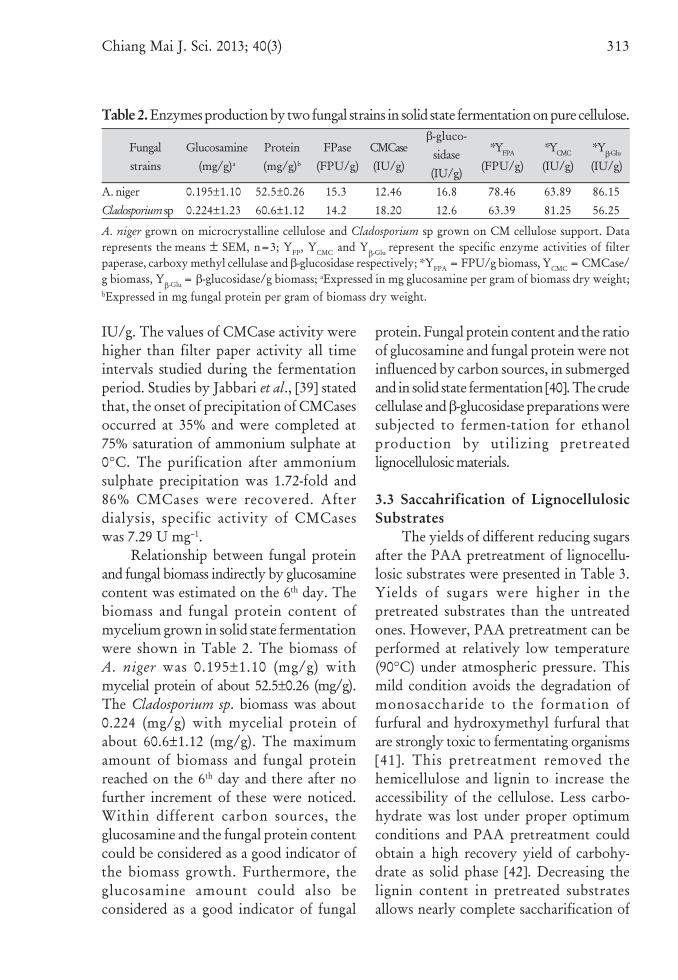
day of incubation in fermentation media(initial pH of 5.5). Further incubationresulted in the gradual decrease of enzymeactivity. This decreasing trend of enzymeactivity may be due to the reduction incellulose content and nutrients supplied inthe substrate. The decrease in cellulaseactivity may also be due to accumulationof cellobiose [38]. Cellulolytic enzyme(s)produced from the isolate A. niger andCladosporium sp. under solid state fermen-tation was precipitated by ammoniumsulphate. The precipitate obtained from A.niger isolate was having high FPase andβ-glucosidase activity (15.3 FPU/g and 16.8IU/g with relative specific activity of 78.46FPU/g and 86.15 IU/g, respectively) whichwas higher than the enzymatic activity ofCladosporium sp. (Table 2). The CMCaseactivity of Cladosporium sp. was higherthan the A. niger isolate, with activity of18.20 IU/g with specific activity of 81.25
Chiang Mai J. Sci. 2013; 40(3) 313
Table 2. Enzymes production by two fungal strains in solid state fermentation on pure cellulose.
A. niger grown on microcrystalline cellulose and Cladosporium sp grown on CM cellulose support. Datarepresents the means ± SEM, n=3; YFP, YCMC and Y
β-Glu represent the specific enzyme activities of filterpaperase, carboxy methyl cellulase and β-glucosidase respectively; *YFPA = FPU/g biomass, YCMC = CMCase/g biomass, Y
β-Glu = β-glucosidase/g biomass; aExpressed in mg glucosamine per gram of biomass dry weight;bExpressed in mg fungal protein per gram of biomass dry weight.
IU/g. The values of CMCase activity werehigher than filter paper activity all timeintervals studied during the fermentationperiod. Studies by Jabbari et al., [39] statedthat, the onset of precipitation of CMCasesoccurred at 35% and were completed at75% saturation of ammonium sulphate at0°C. The purification after ammoniumsulphate precipitation was 1.72-fold and86% CMCases were recovered. Afterdialysis, specific activity of CMCaseswas 7.29 U mg–1.
Relationship between fungal proteinand fungal biomass indirectly by glucosaminecontent was estimated on the 6th day. Thebiomass and fungal protein content ofmycelium grown in solid state fermentationwere shown in Table 2. The biomass ofA. niger was 0.195±1.10 (mg/g) withmycelial protein of about 52.5±0.26 (mg/g).The Cladosporium sp. biomass was about0.224 (mg/g) with mycelial protein ofabout 60.6±1.12 (mg/g). The maximumamount of biomass and fungal proteinreached on the 6th day and there after nofurther increment of these were noticed.Within different carbon sources, theglucosamine and the fungal protein contentcould be considered as a good indicator ofthe biomass growth. Furthermore, theglucosamine amount could also beconsidered as a good indicator of fungal
protein. Fungal protein content and the ratioof glucosamine and fungal protein were notinfluenced by carbon sources, in submergedand in solid state fermentation [40]. The crudecellulase and β-glucosidase preparations weresubjected to fermen-tation for ethanolproduction by utilizing pretreatedlignocellulosic materials.
3.3 Saccahrification of LignocellulosicSubstrates
The yields of different reducing sugarsafter the PAA pretreatment of lignocellu-losic substrates were presented in Table 3.Yields of sugars were higher in thepretreated substrates than the untreatedones. However, PAA pretreatment can beperformed at relatively low temperature(90°C) under atmospheric pressure. Thismild condition avoids the degradation ofmonosaccharide to the formation offurfural and hydroxymethyl furfural thatare strongly toxic to fermentating organisms[41]. This pretreatment removed thehemicellulose and lignin to increase theaccessibility of the cellulose. Less carbo-hydrate was lost under proper optimumconditions and PAA pretreatment couldobtain a high recovery yield of carbohy-drate as solid phase [42]. Decreasing thelignin content in pretreated substratesallows nearly complete saccharification of
Fungalstrains
Glucosamine(mg/g)a
Protein(mg/g)b
FPase(FPU/g)
CMCase(IU/g)
β-gluco-sidase(IU/g)
*YFPA
(FPU/g)*YCMC
(IU/g)*Y
β-Glu
(IU/g)
A. nigerCladosporium sp
0.195±1.100.224±1.23
52.5±0.2660.6±1.12
15.314.2
12.4618.20
16.812.6
78.4663.39
63.8981.25
86.1556.25

314 Chiang Mai J. Sci. 2013; 40(3)
Table 3. The yields (mg/g) of different sugars after PAA pretreatment.
the polysaccharides. A comparison of theresidual lignin content with glucose releasefor all samples during this study showedthat as lignin content dropped, glucoserelease increased.
The results presented in Table 3 showthat the initial raw untreated substratescontain low levels of total sugars andreducing sugars. After pretreatment, thereducing sugar concentration was higherin pretreated bagasse (42.8±0.2 mg/g)followed by rice straw (42.6±0.4 mg/g) andcorn cob (40.0±0.2 mg/g). The arabinosecontent was higher in rice straw samplescompared with bagasse and corncob. Thehigher cellulose content of bagasse resultedin increasing the reducing sugar content inpretreated bagasse than the other pretreatedsamples. Reducing sugar concentrationwas higher in rice straw (42.6±0.4 mg/g)than in corn cob (40.0±0.2 mg/g) which isdue to low lignin content of rice straw thancorn cob. The results showed that afterpretreatment the availability of reducingsugars from pretreated substrates wereincreased from their respective carbohydratecontent. In comparison the bagasse containedhigher cellulose content than the othersubstrates. The levels of reducing sugars inthe sample would determine to some extentthe percentage of ethanol that would beproduced in the fermenting medium. Yield
of sugars from their corresponding substrateswere presented in Table 3. Celluloseconversion of PAA pretreated samples after60 min were shown in Figure 1. Thehydrolysis of PAA pretreated bagassesamples resulted in the highest glucose fromglucan conversion at 35.97%, arabinosefrom arabinan conversion low at 22.22%compared with pretreated corn cob sample,higher than the pretreated rice strawsample.
The highest xylan and arabinanconversion rate at 36.50 and 31.11% wasobserved in pretreated corncob than theother substrates. The xylan and arabinanconversion rate at 27.90 and 21.42%respectively was noted in the pretreatedrice straw which is more than the pretreatedcorn cob.
3.4 Simultaneous Saccharification andFermentation (SSF)
The saccharification of pretreatedsubstrates by crude cellulase preparationsalong with fermentation process wasallowed to occur simultaneously for 96 hduration by employing S. cerevisiae and S.bayanus, which could be cultured very nearto the optimum temperature (50oC) forcellulase. Though 50oC was found to beoptimum, the range of 40-50oC did notaffect the yield significantly, and hence SSF
The values are based on dry-weight basis (%); Data represents the mean ± SEM, n=3.
Substrate Total sugarsReducing
sugars Glucose Xylose Arabinose
RSPRSSCBPSCBCCBPCCB
12.46±0.248.2±0.212.6±0.652.4±0.614.2±0.446.4±0.5
10.82±0.842.6±0.410.6±0.1242.8±0.212.4±0.640.0±0.2
7.4±0.230.5±0.18.2±0.232.8±0.48.6±0.326.9±0.2
2.8±0.18.6±0.25.4±0.210.6±0.62.4±0.312.6±0.3
1.6±0.34.2±0.13.2±0.13.6±0.42.5±0.24.5±0.2
Chiang Mai J. Sci. 2013; 40(3) 315
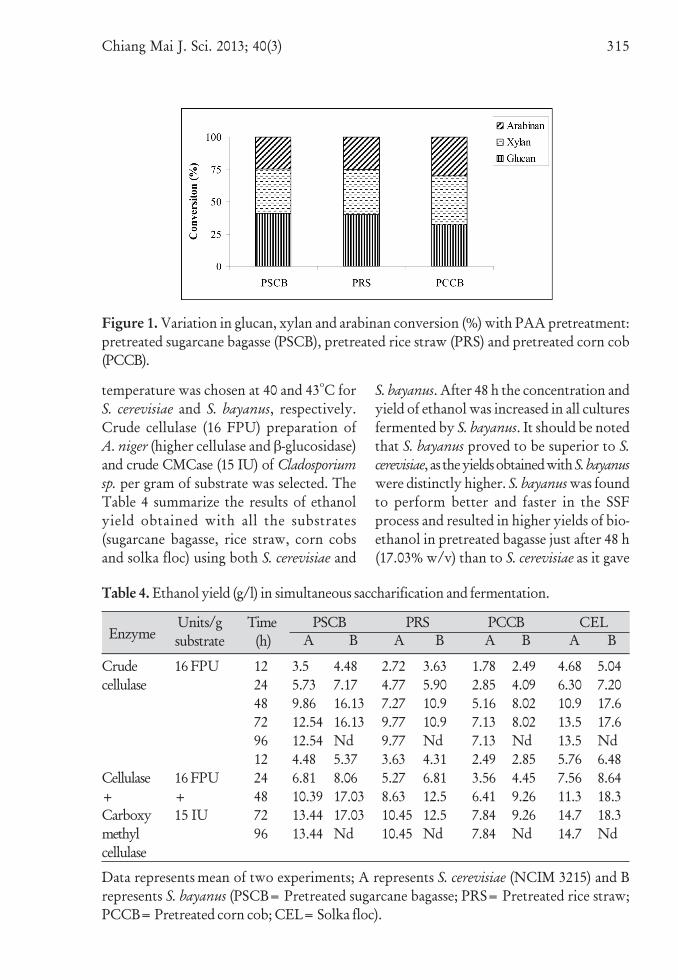
Table 4. Ethanol yield (g/l) in simultaneous saccharification and fermentation.
Data represents mean of two experiments; A represents S. cerevisiae (NCIM 3215) and Brepresents S. bayanus (PSCB= Pretreated sugarcane bagasse; PRS= Pretreated rice straw;PCCB= Pretreated corn cob; CEL= Solka floc).
temperature was chosen at 40 and 43oC forS. cerevisiae and S. bayanus, respectively.Crude cellulase (16 FPU) preparation ofA. niger (higher cellulase and β-glucosidase)and crude CMCase (15 IU) of Cladosporiumsp. per gram of substrate was selected. TheTable 4 summarize the results of ethanolyield obtained with all the substrates(sugarcane bagasse, rice straw, corn cobsand solka floc) using both S. cerevisiae and
S. bayanus. After 48 h the concentration andyield of ethanol was increased in all culturesfermented by S. bayanus. It should be notedthat S. bayanus proved to be superior to S.cerevisiae, as the yields obtained with S. bayanuswere distinctly higher. S. bayanus was foundto perform better and faster in the SSFprocess and resulted in higher yields of bio-ethanol in pretreated bagasse just after 48 h(17.03% w/v) than to S. cerevisiae as it gave
Figure 1. Variation in glucan, xylan and arabinan conversion (%) with PAA pretreatment:pretreated sugarcane bagasse (PSCB), pretreated rice straw (PRS) and pretreated corn cob(PCCB).
EnzymeUnits/gsubstrate
Time (h)
PSCB PRS PCCB CELA B A B A BA B
Crudecellulase
Cellulase+Carboxymethylcellulase
16 FPU
16 FPU+15 IU
12244872961224487296
3.55.739.8612.5412.544.486.8110.3913.4413.44
4.487.1716.1316.13Nd5.378.0617.0317.03Nd
2.724.777.279.779.773.635.278.6310.4510.45
3.635.9010.910.9Nd4.316.8112.512.5Nd
1.782.855.167.137.132.493.566.417.847.84
2.494.098.028.02Nd2.854.459.269.26Nd
4.686.3010.913.513.55.767.5611.314.714.7
5.047.2017.617.6Nd6.488.6418.318.3Nd

316 Chiang Mai J. Sci. 2013; 40(3)
high yields of bio-ethanol only after 72 h(13.44% w/v). The time period 72 h wasrequired for S. cerevisiae to attain maximumethanol concentration with correspondingethanol yield. This result is attributed tothe thermo-tolerance of S. bayanus. But at50oC, the ethanol yield was found to bereduced with both the yeasts and significantamount of sugar was found unmetabolized.This was due to the thermo-lability of boththe yeasts at the higher temperature.However, S. bayanus generally tolerated ahigher ethanol and temperature thanS. cerevisiae. Addition of crude preparationof Cladosporium sp. was found to favorfaster reactions with both the yeasts. Thismight be due to low activities of CMCasein the crude preparation of A. niger andsupplementing it from external source gavebetter yields since the cellulose degradationwas initiated by the action of CMCase.Pretreatment of substrates has been foundto have significant influence on ethanolproduction. The levels of reducing sugarin the sample would determine to someextent the high percentage of ethanolproduced in the fermenting medium.The higher ethanol yield in the fermentedpretreated bagasse was due to high amountof reducing sugars than the others in thefermenting medium.
Thermo-tolerant yeasts capable ofgrowth and ethanol production at tem-peratures above 40oC have been investigatedin the past for suitability in SSF [43]. Theethanol yields of substrates under SSF byyeasts were in the order of: solka floc> sugarcane bagasse > rice straw >corncobs.
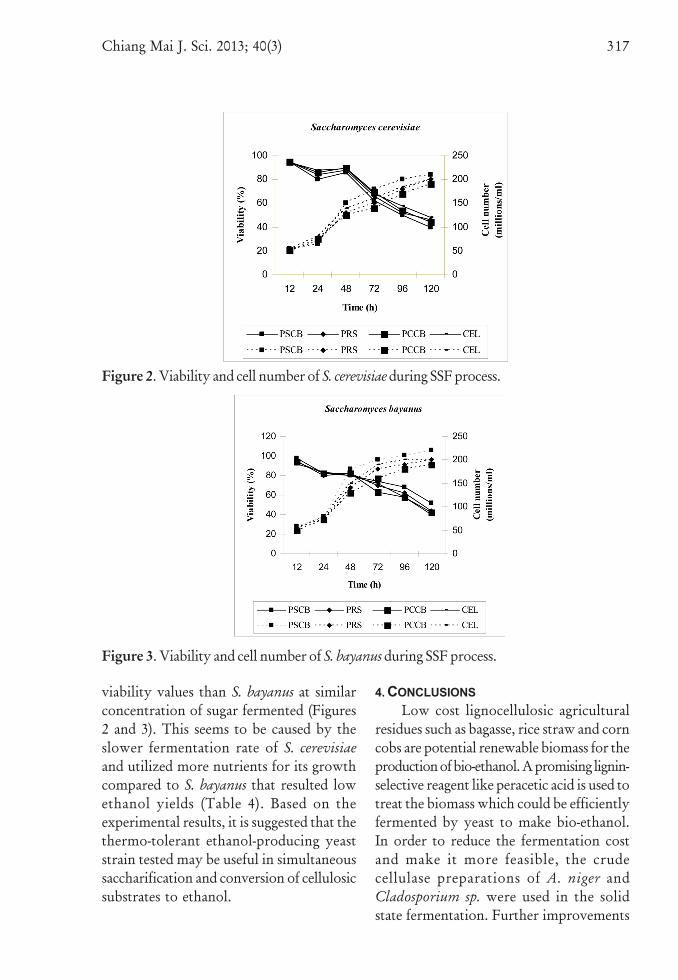
3.5 Yeast Cell Viability and Cell CountThe Figures 2 and 3 represent the yeast
cell (S. cerevisiae and S. bayanus) viability andtotal number of cells during fermentation of
pretreated substrates along with solka floc(reference culture medium). During initialstages of fermentation, there was no increasein yeast cell numbers, as they were in lag phase.After that there was a gradual increase incell numbers indicating that the yeast cellswere utilizing the sugars present in themedium. The cultures grown in pretreatedbagasse had higher cell numbers than theculture grown on other pretreated sub-strates. This is due to the availability of higherreducing sugars in the pretreated bagasseculture medium than the others. Generally,yeast performance may be affected both byvery low glucose concentration resultingin metabolic stress conditions and highethanol presence in fermenting media. Thelower viability shown by S. bayanus at samesugar concentrations seem to be due to afast accumulation of internal ethanol overshort fermentation periods. At 48 h, 80%viability of S. bayanus cells on bagasse mediumwas compared with 86% viability ofS. cerevisiae cells. At short fermentationperiods (48 h), S. bayanus has a greatfermentation activity. Despite the very lowviability of S. bayanus, it fermented glucoseat temperatures ≥ 42oC; increments inbiomass were obtained due to the contribu-tion of intact but non viable cells at theend of the fermentation. High values offinal viability were obtained due to the useof a high cell density. It has been reportedthat the maintenance of viability duringfermentation is favoured by high celldensities [44]. The limiting effect ofincreasing temperatures on the fermenta-tion capacity of S. bayanus is due to lossesin cell viability. Fast fermentation is oneof the causes for the drops in cell viability,due to a high internal ethanol concentrationthat reaches inside the cells at increasingsugar concentrations. On the other hand,S. cerevisiae was able to maintain higher
Chiang Mai J. Sci. 2013; 40(3) 317
Figure 2. Viability and cell number of S. cerevisiae during SSF process.
Figure 3. Viability and cell number of S. bayanus during SSF process.
viability values than S. bayanus at similarconcentration of sugar fermented (Figures2 and 3). This seems to be caused by theslower fermentation rate of S. cerevisiaeand utilized more nutrients for its growthcompared to S. bayanus that resulted lowethanol yields (Table 4). Based on theexperimental results, it is suggested that thethermo-tolerant ethanol-producing yeaststrain tested may be useful in simultaneoussaccharification and conversion of cellulosicsubstrates to ethanol.
4. CONCLUSIONSLow cost lignocellulosic agricultural
residues such as bagasse, rice straw and corncobs are potential renewable biomass for theproduction of bio-ethanol. A promising lignin-selective reagent like peracetic acid is used totreat the biomass which could be efficientlyfermented by yeast to make bio-ethanol.In order to reduce the fermentation costand make it more feasible, the crudecellulase preparations of A. niger andCladosporium sp. were used in the solidstate fermentation. Further improvements

318 Chiang Mai J. Sci. 2013; 40(3)
should focus on increasing the viability rateof S. bayanus strain to obtain higher ethanolyield by adding the special nutrients like fingermillet [17] medium for rapid fermentation.
ACKNOWLEDGEMENTSWe are thankful to UGC, New Delhi
for providing funds. We also wish to thankDr. S.C. Basappa, former Deputy Directorand Scientist, Central Food TechnologicalResearch Institute (CFTRI), Mysore, forhis encouragement and critical commentson the manuscript.
REFERENCES
[1] Joshi B., Bhatt M.R., Sharma D.,Joshi J., Rajani M. and Lakshmaiah S.,Lignocellulosic ethanol production:Current practices and recentdevelopments, Biotechnol. Mol. Biol.Rev., 2011; 6(8): 172-182.
[2] Hayes D.J., An examination ofbiorefining processes, catalysts andchallenges, Catal. Today, 2009; 145:138-151.
[3] Yat S.C., Berger A., Shonnard D.R.,Kinetic characterization of dilutesurface acid hydrolysis of timbervarieties and switchgrass, BioresourceTechnol., 2008; 99: 3855-3863.
[4] Mosier N., Wyman C., Dale B., ElanderR., Lee Y.Y. and Holtzapple M., et al.Features of promising technologies forpretreatment of lignocellulosic biomass,Bioresource Technol., 2005; 96: 673-686.
[5] Teixeira L.C., Linden J.C. and SchroederH.A., Simultaneous saccharification andcofermentation of peracetic acid-pretreated biomass, Appl. Biochem.Biotechnol., 2000; 84-86: 111-27.
[6] Zayed G. and Meyer O., The single batchconversion of wheat straw to ethanolemploying the fungus Trichoderma viride
and the yeast Pachysolen tannophilus,Appl. Microbiol. Biotechnol., 1996; 45:551-555.
[7] Miyomoto K., Production of fuelalcohol from cellulosic biomass, FAOAgricultural Services Bulletin, 1997;128: 19-52.
[8] Nakamura Y., Sawada T. and InoueE., Enhanced ethanol production fromenzymatically treated steam explodedrice straw using extractive fermentation,J. Chem. Technol. Biotechnol., 2001; 76:879-884.
[9] Laser M., Schulman D., Allen S.G.,Lichwa J., Antal M.J. and Lynd L.R.,A comparison of liquid hot water andsteam pretreatments of sugarcanebagasse for bioconversion to ethanol,Bioresource Technol., 2002; 81: 33-44.
[10] Haki G.D. and Rakshit S.K.,Developments in industriallyimportant thermostable enzymes: Areview, Bioresource Technol., 2003; 89:17-34.
[11] Shen Y., Zhang Y., Ma T., Bao X.,Du F. and Qu Y., Simultaneoussaccharification and fermentation ofacid-pretreated corncobs with arecombinant Saccharomyces cerevisiaeexpressing beta-glucosidase, BioresourceTechnol., 2007; 99: 5099-5103.
[12] Wyman C.E., Ethanol fromlignocellulosic biomass: Technology,economics and opportunities, BioresourceTechnol., 1994: 50: 3-15.
[13] Qurat-ul-ain., Baig S. and Saleem M.,Production and characterization ofcellulases of Asperigillus niger by using ricehusk and saw dust as substrates, Pak. J.Bot., 2012; 44: 377-382.
[14] Spindler D.D., Wyman C.E.,Mohagheghi A. and Gorhmann K.,Thermotolerant yeast for simultaneoussaccharification and fermentation of

Chiang Mai J. Sci. 2013; 40(3) 319
cellulose to ethanol, Appl. Biochem.Biotechnol., 1988; 17: 279-293.
[15] Miller K.P., Biofuel Ethanol Productionby Saccharomyces bayanus, TheChampagne Yeast, Ph.D. thesis, Clemson,South Carolina. 2010.
[16] Barros M.R., Cabral J.M. and NovaisJ.M., Production of ethanol byimmobilized Saccharomyces bayanus inan extractive fermentation system,Biotechnol. Bioeng., 1987; 29(9): 1097-1104.
[17] Pradeep P., Krishna Goud G.K. andReddy O.V.S., Optimization of veryhigh gravity (VHG) finger millet (ragi)medium for ethanolic fermentation byyeast, Chiang Mai J. Sci. 2010; 37:116-123.
[18] Mohan P.R., Ramesh B. and O.V.S.Reddy., Production and optimizationof ethanol production from pretreatedsugarcane bagasse using Saccharomycesbayanus in simultaneous saccharifica-tion and fermentation, Microbiol. J.,2012; 2: 52-63.
[19] Kristian B.R.M.K., Biomass degradingenzymes from Penicillium-cloning andcharacterization, Ph.D. Thesis. 2008.
[20] Bakare M.K., Adewale I.O., Ajayi A.and Shonukan O.O., Purification andcharacterization of cellulase from thewild-type and two improved mutantsof Pseudomonas fluorescens, Afr. J.Biotechnol., 2005; 9: 898-904.
[21] Yang B. and Wyman C.E., Effect ofxylan and lignin removal by batch andflow through pretreatment on theenzymatic digestability of corn stovercellulose, Biotechnol Bioeng., 2004; 86:88-95.
[22] Takagi M., Abe S., Suzuki S., EmertG.H. and Yata N., A method forproduction of alcohol directly fromcellulose using cellulase and yeast. In
Ghose T.K. (ed.), Proceedings ofbioconversion of cellulosic substancesinto energy, chemicals and microbialprotein, I.I.T, New Delhi, 1997; 551-571.
[23] Goering H.K. and Van Soest P.J.,Forage fibre analysis. USDA-ARSagricultural hand book. Governmentprinting office, Washington DC, 1971.
[24] AOAC., Official methods of analysisof the association of official chemists,15th ed. In: Helrich K., ed., AOACInc., Arlington, VA., USA, 1990.
[25] Dubois M., Gills K.A., Hamilton J.A.and Rebers P.A., Colorimetricmethod for determination of sugarsand related substances, Anal. Chem.,1956; 28: 350-356.
[26] Miller G.L., Use of dinitrosalicylicacid reagent for determination ofreducing sugar, Anal. Chem., 1959; 31:426-428.
[27] Khabarov Y.G., Kamakina N.D.,Gusakov L.V. and Veshnyakov V.A.,New spectrophotometric method fordetermination of furfural andpentoses, Russ. J. Appl. Chem., 2006;79: 103-106.
[28] Lowry O.H., Rosenrough N.J., FarrA.L. and Randell R.J., Proteinmeasurement with the folin phenolreagent, J. Biol. Chem., 1951; 193: 265-275.
[29] Mandels M., Andreotti R. and RocheC., Measuring of saccharifyingcellulose, Biotechnol. Bioeng. Symp.,1976; 6: 21-33.
[30] Desrochers M., Jurasek L. andPaige M.G., Production of cellulase,β-glucosidase, and xylanase bySchizophyllum commune grown on acellulose-peptone medium, Dev. Ind.Microbiol., 1981; 22: 675-684.

320 Chiang Mai J. Sci. 2013; 40(3)
[31] Tsuji A., Kinoshita T. and HoshinoM., Analytical chemical studieson amino sugar, II. Determinationof hexosamines using 3-methyl-2-benzothiazolone hydrazone hydrochloride, Chem. Pharm. Bull., 1969;17: 1505-1510.
[32] Ride J.P. and Drysdale R.B., A rapidmethod for the chemical estimation offilamentous fungi in plant tissue,Physiol. Mol. Plant. Pathol., 1972; 2:7-15.
[33] Postgate J.P., Viable counts andviability, In: Norris, J.R., Ribbons,D.W. (Eds.), Methods in Microbiology,New York Academic Press, 1967:611-628.
[34] Anthony J.C., Malt beverages andmalt brewing materials: Gaschromatographic determination ofethanol in beer, J. Assoc. Anal. Chem.,1984; 67: 192-193.
[35] Lu C.G., Yang B., Gregg D., SaddlerJ.N. and Mansfield S.D., Cellulaseadsorption and an evaluation ofenzyme recycle during hydrolysis ofsteam-exploded softwood residues,Appl. Biochem. Biotechnol., 2002; 98:641-654.
[36] R mond-Zilliox C., Etude desm canismes de d gradation enzymatiquedes parois v g tales, Application l’hydrolyse de la paille de bl par uneendoxylanase, Ph.D. Thesis, Reims,France, 1996.
[37] Boussarsar H., Roge B. and MathlouthiM., Optimization of sugarcane bagasseconversion by hydrothermal treatmentfor the recovery of xylose, BioresourceTechnol., 2009; 100: 6537-6542.
[38] Singh A., Singh N. and Bishnoi N.R.,Production of cellulases by Aspergillusheteromorphus from wheat strawunder submerged fermentation, Int. J.Civil Environ. Eng., 2009; 1: 1-4.
[39] Jabbari A., Rashid M.H., MalanaM.A., Amanullah, Saleem M., NiazM. and Yasin M.Z., Production,partial purification and proteolysis ofcarboxymethyl cellulases fromArachniotus citrinus, Int. J. Agric.Biol., 2002; 6: 615-619.
[40] Chen Z., Wang Z. and Zhou A., Fungalprotein estimation of Aspergillusoryzae growing in solid statecultivation conditions, Can. J. PureAppl. Sci., 2010; 4: 1127-1131.
[41] Mussatto S.I. and Roberto I.C.,Alternatives for detoxification ofdiluted-acid lignocellulosic hydrolyzatesfor use in fermentative processes: Areview, Bioresource Technol., 2004; 93:1-10.
[42] Zhao X.B., Peng F., Cheng K.K. andLiu D.H., Enhancement of theenzymatic digestibility of sugarcanebagasse by alkali-peracetic acidpretreatment, Enz. Microb. Technol.,2009; 44: 156-164.
[43] Szezodrak J. and Targonski Z.,Selection of thermotolerant yeaststrains for simultaneous saccharifica-tion and fermentation of cellulose,Biotechnol Bioeng., 1988; 31: 300-303.
[44] Ueno R., Urano N. and Kimura S.,Effect of temperature and cell densityon ethanol fermentation by a thermo-tolerant aquatic yeast strain isolatedfrom a hot spring environment,Fisheries Sci., 2002; 68: 571.