alternating attachment corrugated fibronectin - pnas.org ·...
TRANSCRIPT

Proc. Natl. Acad. Sci. USAVol. 77, No. 7, pp. 4108-4 112, July 1980Cell Biology
Corrugated attachment membrane in WI-38 fibroblasts: Alternatingfibronectin fibers and actin-containing focal contacts
(fluorescence microscopy/reflection contrast microscopy/transmembrane interaction/microfilaments/microinjection)
CARMEN BIRCHMEIER*, THOMAS E. KREISt, HANS M. EPPENBERGER*, KASPAR H. WINTERHALTERt, ANDWALTER BIRCHMEIERtT*Institut fur Zellbiologie, Eidgenossische Technische Hochschule Honggerberg, CH-8093 Zurich, Switzerland; and tLaboratorium fur Biochemie, EidgenossischeTechnische Hochschule Zentrum, CH-8092 Zurich, Switzerland
Communicated by Edmond H. Fischer, April 14, 1980
ABSTRACT The distributions of both fibronectin (LETS,CSP) fibers and focal contacts to the substratum, as viewed byfluorescence and reflection contrast microscopy, respectively,have been compared in freshly plated WI-38 human fibroblasts.Most frequently, the actual focal attachment plaques did notcontain fibronectin fluorescence and, furthermore, fibronectinspots and fibers often alternated with focal contacts. Overlap,however, was observed between focal contacts and the endingsof actin-containing stress fibers [see also Wehland, J., Osborn,M. & Weber, K. (1979) J. Cell Sci. 37,257-273]. Thus, the fibro-blast attachment membrane might best be described as a cor-rugated sheet that undulates between alternating microfila-ments and fibronectin fibers, at the points of closest and farthestdistance to the substratum, respectively.
Animal cells that attach and spread on artificial substrates havebeen widely used to examine interactions between the externalcell environment and the internal cytoskeleton. For coordinatedcell spreading to occur, signals must evidently be transmittedfrom the outside of the cell to the inside, and vice versa-i.e.,must traverse the plasma membrane. Furthermore, trans-membrane interaction seems to be essential for the expressionof complex properties of cells such as anchorage dependenceof growth, directed cell locomotion, or the integration of singlecells into tissues.
Closely associated with cell attachment and spreading arecomponents of the extracellular matrix-e.g., fibronectin andcollagen-as well as proteins of the intracellular stress fi-bers-e.g., actin, myosin, and a-actinin. This is indicated bya variety of observations: (i) transformed fibroblasts often ex-hibit a poor extracellular matrix and disorganized stress fibersand consequently show a reduced ability for attachment (cf.ref. I for a review); (ii) the drug cytochalasin B leads to thedissociation of the stress fibers, the rounding of the cells, andthe release of fibronectin into the medium (2, 3); (iii) treatmentwith proteases or chelating agents, which act on the extracellularmatrix, results in the detachment of the cells and the dissociationof the stress fibers (4).
At present, the detailed mechanism of such transmembraneinteractions is not known. It has recently been postulated,however, that integral membrane proteins (e.g., protein X)mediate a direct linkage between extra- and intracellularstructures (5). In fact, a variety of observations indicate that suchdirect linkage might occur in certain systems: (i) crosslinkedsurface receptors have been observed to redistribute concom-itantly with cytoskeletal elements (6, 7); (ii) surface receptorscan be copurified with actin-containing structures (8); (iii) in-terlocked microfilaments and fibronectin fibers at the upperfibroblast membrane have been identified in the electron mi-croscope (the fibronexus; cf. ref. 9).
In the present investigation we have analyzed the attachmentmembrane of WI-38 human fibroblasts by a combination ofreflection contrast and fluorescence microscopy. We arrive atthe conclusion that fibronectin spots and fibers do not overlap(cf. refs. 10 and 11) but alternate with microfilament endingsat the focal attachment plaques. Thus, attachment of fibroblaststo the substratum seems rather to be established by an indirectlinkage between the main extra- and intracellular fibers.
EXPERIMENTAL PROCEDURECells. WI-38 fibroblasts were grown on glass coverslips in
Dulbecco's modified Eagle's medium (12). At different stagesof spreading, usually 4-10 hr after subculture, the cells werewashed in phosphate-buffered saline (140 mM NaCl/10 mMNaPi, pH 7.4; same buffer used in all further steps), and fixedwith 1.5% (wt/vol) formaldehyde, pH 7.4, for 30 min (at roomtemperature, as in all the following incubations). After extensivewashing, which included one step of quenching residualformaldehyde reaction with 50 mM ammonium chloride, thecells were made permeable with 0.05% Triton X-100 for 10min, followed by washing. This treatment had no significanteffect upon the appearance of the cells as seen in the reflectioncontrast microscope (cf. also ref. 13).
Antibodies and Staining for Immunofluorescence Mi-croscopy. Affinity-purified rabbit anti-human plasma fibro-nectin IgG was a gift of M. Chiquet and R. Ehrismann of ourlaboratory (14). Rhodamine-labeled goat antibody to rabbit IgGwas obtained from Nordic Laboratories (Lausanne, Switzer-land), and affinity purified (courtesy of J. H. Lanz). TritonX-100-treated fibroblasts were incubated with appropriatelydiluted first antibody for 30 min, followed by extensive wash-ing. Incubation with the rhodamine-labeled second antibodywas performed under similar conditions. Cells were mountedin phosphate-buffered saline.
Microinjection of Rhodamitte-Labeled Actin. The proce-dure was identical to the one described earlier (12). After 1 hrof culture, the microinjected cells were observed in a nonfixedstate.
Optical Methods. A Leitz Diavert microscope equipped forboth reflection contrast and fluorescence microscopy was em-ployed (12). The precision of the Leitz Ploemopak allowed usto superimpose the fluorescence and reflection contrast patternson the same negative. Photographs were taken with an Or-thomat camera on Ilford HP-5 or Ektachrome color slide film(23 DIN, tungsten light; exposed at 29 DIN and developedcorrespondingly). Color prints of the slides were made on Ci-bachrome paper.
t To whom reprint requests should be addressed.
4108
The publication costs of this article were defrayed in part by pagecharge payment. This article must therefore be hereby marked "ad-vertisement" in accordance with 18 U. S. C. §1734 solely to indicatethis fact.

Proc. Natl. Acad. Sci. USA 77 (1980) 4109
I
FIG. 1. Relationship between fibronectin, actin, and the focal contacts in WI-38 fibroblasts. Fibronectin (a) was visualized by immunoflu-orescence using rhodamine-labeled second antibody; actin (d) was rhodamine-labeled and microinjected (cf. ref. 12 and also ExperimentalProcedure). (c and f) The corresponding reflection contrast images; (b and e) double exposures of both the fluorescence and reflection contrastpatterns. The arrows p and c indicate focal attachment plaques and extended focal contacts, respectively;§ arrows o indicate overlap betweenactin fluorescence and a focal attachment plaque. (X650 in a-c; X 500 in d-f.)
RESULTSThe distribution of fibronectin and actin in WI-38 fibroblastswas compared with the pattern of focal contacts to the sub-stratum as viewed by reflection contrast microscopy. § In par-ticular, double exposures of both fluorescence and corre-
§ Reflection contrast (interference reflection) microscopy identifiesthe sites of minimal distance between cell and substratum as darkareas (focal contacts; cf. refs. 15 and 16). The reflection contrastimage increases in brightness to a maximum where the distance be-tween cell membrane and substrate is ;100 nm. The term "focalcontact" is here used for the actual "attachment plaques" (16)-i.e.,the mostly triangular, darker structures near the cell periphery (pin Figs. 1-3), as well as for the thinner, elongated contacts of the at-tachment plane (c in Figs. 1-3).
sponding reflection contrast image were made on the samephotograph in order to closely visualize the relationship betweenthe two patterns. In the early stages of cell spreading, fibro-nectin, as viewed by immunofluorescence, showed both a spottyand fibrillar distribution upon focusing at the cell attachmentplane (red fluorescence on single exposure, Fig. la, and yellowafter double exposure, Fig. lb). The corresponding reflectioncontrast image was similar in some aspects such as in the overalldirection of the focal contacts (dark green on a light greenbackground, Fig. lc) but showed significant differences uponclose inspection. First, well-developed focal attachment plaques(p) near the cell periphery usually did not contain any fibro-nectin fluorescence; second, focal contacts (c) extending throughlarge parts of the cell often did not overlap but rather alternated
Cell Biology: Birchmeier et al.
4110 Cell Biology: Birchmeier et al.
0
:1 :::
.,..
piiE 'sitS. ~Clo-+%,%ek; ; ; .:
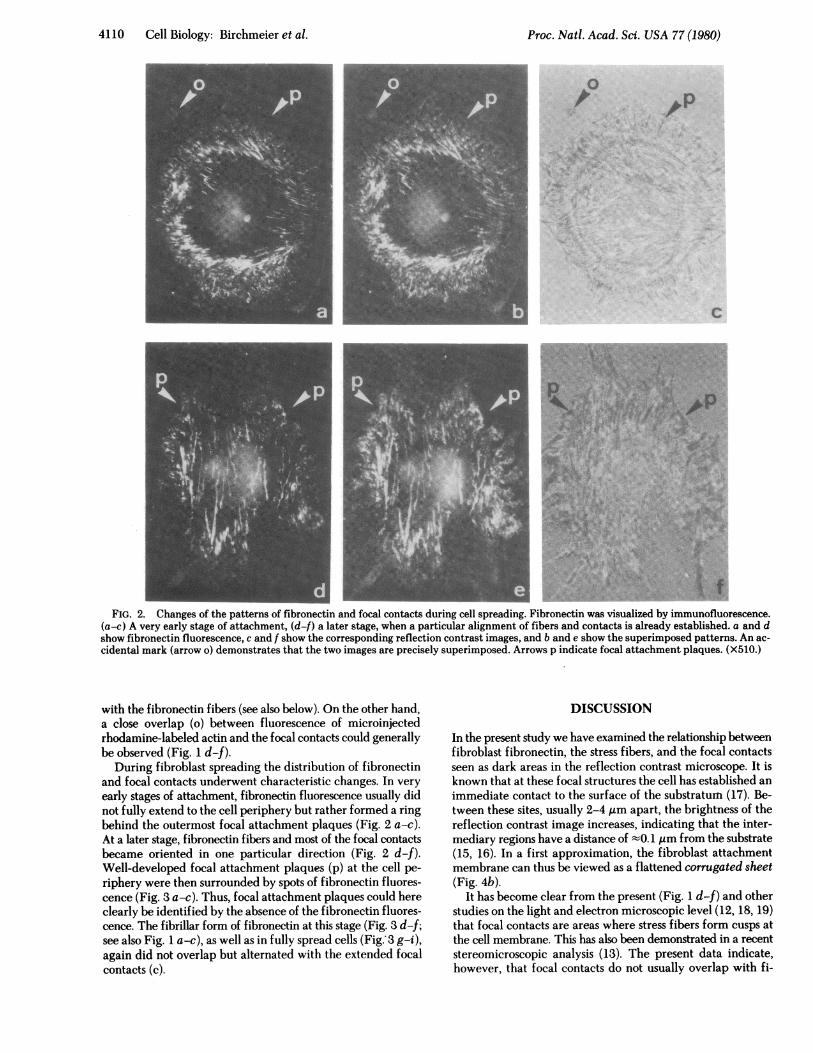
.t0t'X,,,;2*'sUIfs\;FIG. 2. Changes of the patterns of fibronectin and focal contacts during cell spreading. Fibronectin was visualized by immunofluorescence.
(a-c) A very early stage of attachment, (df) a later stage, when a particular alignment of fibers and contacts is already established. a and dshow fibronectin fluorescence, c and f show the corresponding reflection contrast images, and b and e show the superimposed patterns. An ac-cidental mark (arrow o) demonstrates that the two images are precisely superimposed. Arrows p indicate focal attachment plaques. (X510.)
with the fibronectin fibers (see also below). On the other hand,a close overlap (o) between fluorescence of microinjectedrhodamine-labeled actin and the focal contacts could generallybe observed (Fig. 1 d-f).
During fibroblast spreading the distribution of fibronectinand focal contacts underwent characteristic changes. In veryearly stages of attachment, fibronectin fluorescence usually didnot fully extend to the cell periphery but rather formed a ringbehind the outermost focal attachment plaques (Fig. 2 a-c).At a later stage, fibronectin fibers and most of the focal contactsbecame oriented in one particular direction (Fig. 2 d-f).Well-developed focal attachment plaques (p) at the cell pe-riphery were then surrounded by spots of fibronectin fluores-cence (Fig. 3 a-c). Thus, focal attachment plaques could hereclearly be identified by the absence of the fibronectin fluores-cence. The fibrillar form of fibronectin at this stage (Fig. 3 d-f;see also Fig. 1 a-c), as well as in fully spread cells (Fig. 3 g-i),again did not overlap but alternated with the extended focalcontacts (c).
DISCUSSION
In the present study we have examined the relationship betweenfibroblast fibronectin, the stress fibers, and the focal contactsseen as dark areas in the reflection contrast microscope. It isknown that at these focal structures the cell has established animmediate contact to the surface of the substratum (17). Be-tween these sites, usually 2-4 gm apart, the brightness of thereflection contrast image increases, indicating that the inter-mediary regions have a distance of t0. 1 tim from the substrate(15, 16). In a first approximation, the fibroblast attachmentmembrane can thus be viewed as a flattened corrugated sheet(Fig. 4b).
It has become clear from the present (Fig. 1 d-f) and otherstudies on the light and electron microscopic level (12, 18, 19)that focal contacts are areas where stress fibers form cusps atthe cell membrane. This has also been demonstrated in a recentstereomicroscopic analysis (13). The present data indicate,however, that focal contacts do not usually overlap with fi-
Proc. Natl. Acad. Sci. USA 77 (1980)

Proc. Natl. Acad. Sci. USA 77 (1980) 4111
AD4 .jS w a.
X., I,.. t ..t.,. .. ....t .ws i
^.a.*,.:e.: X #.. ,$. +tQ.R,.*f
$ ,,7;
s ..+n ye
.s J-,.Ps w
f;.'t A,
.*,
L (, At _ ,
_ - _-= '; *-W_lt~~~~~~~~~~-7I
FIG. 3. Enlargements of particular areas of the WI-38 attachment membrane. (a, d, and g) Fibronectin was visualized by immunofluorescence;(c, f, and i) the corresponding reflection contrast images; and (b, e, and h) the superimposed patterns. a-c is an enlargement of Fig. 2 d-f. Arrowsp indicate focal attachment plaques; arrows c indicate extended focal contacts. (X1070 in a-c; X700 in d-i.)
bronectin spots and fibers. In particular, the actual focal at-tachment plaques are characteristically free of fibronectinfluorescence (Figs. 2, 3 a-c). We further show that the bulk of
fibronectin in both the spotty early state as well as the fibrouslater state alternates with the focal contacts (Figs. 1 a-c and 3).This can be seen best in areas where both fibronectin and focal
Cell Biology: Birchmeier et al.

4112 Cell Biology: Birchmeier et al.
a;-, Membrane
Substrate
Microfilament
b_
Fibronectin
FIG. 4. Alternative models of attachment of WI-38 fibroblaststo glass surfaces. (a) Hypothesis of overlapping fibronectin fibers andstress fibers (cf. refs. 10 and 11); (b) model based upon the presentresults. For details see text.
contacts have such a characteristic arrangement that overlapcannot possibly be enforced. In older fibroblast cultures therelationship between the two patterns may become less clear,because at that time the cells have secreted large amounts offibronectin and, perhaps, have slightly moved from their initialattachment site.The distribution of fibronectin and actin fibers has previously
been analyzed by double-label immunofluorescence but notreflection contrast microscopy (10, 11). Because the two fibrousarrays showed some similarities, overlapping structures suchas depicted in Fig. 4a were proposed. The data presented in thisstudy, which takes advantage of reflection contrast microscopy,suggest the alternative model of Fig. 4b. According to thisscheme, the corrugated attachment membrane undulates be-tween stress fibers on the cytoplasmic side and alternating fi-bronectin and possibly collagen-containing fibers (20) on thecell outside.
It is generally assumed that the fibronectin involved in theformation of fibers between cell and substrate does not originatefrom fibronectin present in the medium but is rather synthe-sized and secreted in situ by the attached cells. It seems likelythat fibronectin secretion takes place at sites between the focalcontacts and that the spots seen in early stages of attachment(Fig. 3 a-c) represent fibronectin discharged by single secretoryvesicles.
Thus, the functional linkage between fibronectin and cy-toskeleton during fibroblast spreading (see Introduction) mustbe rather indirect-i.e., more components must be involved insignal transfer than is suggested by simple overlapping struc-tures. The complex gangliosides, which have recently beenimplicated in fibronectin binding to the cell membrane (21),might represent such a component.
This work was supported by Grants 3.196-0.77 (to W.B. and K.H.W.)and 3.187-0.77 (to H.M.E.) from the Swiss National Science Founda-tion, and Grant 0.330.079.76/5 (to W.B. and K.H.W.) from theEidgenossische Technische Hochschule Betriebsdirektion.1. Hynes, R. 0. (1976) Biochim. Biophys. Acta 458,73-107.2. Weber, K., Rathke, P. C., Osborn, M. & Franke, W. W. (1976)
Exp. Cell Res. 102, 285-297.3. Ali, I. U. & Hynes, R. 0. (1977) Biochim. Biophys. Acta 471,
16-24.4. Rees, D. A., Lloyd, C. W. & Thom, D. (1977) Nature (London)
267, 124-128.5. Singer, S. J., Ash, J. F., Bourguignon, L. Y. W., Heggeness, M. H.
& Louvard, D. (1978) J. Supramol. Struct. 9,373-389.6. Ash, J. F., Louvard, D. & Singer, S. J. (1977) Proc. Natl. Acad.
Sci. USA 74,5584-5588.7. Gabbiani, G., Chaponnier, C., Zumbe, A. & Vasalli, P. (1977)
Nature (London) 269,697-698.8. Flanagan, J. & Koch, G. L. E. (1978) Nature (London) 273,
278-281.9. Singer, I. I. (1979) Cell 16,675-685.
10. Heggeness, M. H., Ash, J. F. & Singer, S. J. (1978) Ann. NY Acad.Sci. 312, 414-417.
11. Hynes, R. 0. & Destree, A. T. (1978) Cell 15,875-886.12. Kreis, T. E., Winterhalter, K. H. & Birchmeier, W. (1979) Proc.
Natl. Acad. Sci. USA 76,3814-3818.13. Wehland, J., Osborn, M. & Weber, K. (1979) J. Cell Sci. 37,
257-273.14. Chiquet, M., Puri, E. C. & Turner, D. C. (1979) J. Biol. Chem.
254,5475-5482.15. Abercrombie, M. & Dunn, G. A. (1975) Exp. Cell Res. 92,57-
62.16. Izzard, C. S. & Lochner, L. R. (1976) J. Cell Sci. 21, 129-159.17. Bereiter-Hahn, J., Fox, C. H. & Thorell, B. (1979) J. Cell Biol.
82,767-779.18. Geiger, B. (1979) Cell 18, 193-205.19. Heath, V. P. & Dunn, G. A. (1978) J. Cell Sci. 29, 197-203.20. Vaheri, A., Kurkinen, M., Lehto, V.-P., Linder, E. & Timpl, R.
(1978) Proc. Natl. Acad. Sci. USA 75,4944-4948.21. Kleinman, H. K., Martin, G. R. & Fischman, P. H. (1979) Proc.
Natl. Acad. Sci. USA 76,3367-3371.
Proc. Natl. Acad. Sci. USA 77 (1980)