apresentação do powerpoint -...
TRANSCRIPT

01/04/2011
1
Neodarwinismo (ou Síntese Moderna)
1920-1950
Professor Fabrício R Santos [email protected]
Departamento de Biologia Geral, UFMG 2011
Gregor Mendel (1822-1884)
(1865)
Antes da Síntese Moderna (Neodarwinismo) 1- A história evolutiva era amplamente aceita, mas a hipótese de Seleção Natural foi compreendida por poucos e aceita por ainda menos pessoas. 2- O mecanismo de herança não era compreendido: acreditava-se na herança por mistura (caracteres eram a média dos pais). Um cientista chamado Jenkins demonstrou em 1867 que na herança por mistura os caracteres tendiam a se tornar homogêneos em algumas gerações nas populações. Se a Seleção Natural não gerava variabilidade, mas dependia da variação aleatória pré-existente, com a herança por mistura não haveria variantes suficientes para a Seleção atuar. 3- Acreditava-se também nas variações induzidas pelo ambiente que poderiam ser herdadas, e que também era uma crença de Darwin em algumas raras situações (ex: desuso de órgãos em animais de cavernas, produção de leite em bovinos) nos últimos anos de sua vida. Em 1883, August Weisman sugeriu que o plasma germinativo era completamente separado do resto do corpo (o soma), rejeitando a influência do ambiente sobre a hereditariedade. Na época, ele foi atacado veementemente por NeoLamarckistas, que estavam em ascensão no final do século XIX.
Geneticistas mutacionistas e neolamarckistas de 1900 a 1940 (a negação dos princípios da Seleção Natural de Darwin)
As primeiras tentativas de se associar teorias evolutivas aos princípios da genética foram amplamente desfavoráveis às ideias de Darwin. Essas divergências apareciam pela incompreensão da natureza dos genes e do fundamentalismo Mendeliano: 1- Apenas os caracteres de variação descontínua (cor de olho, forma lisa e rugosa) eram considerados genéticos. Caracteres contínuos (tamanho) não teriam componentes genéticos. Além disto, acreditavam que poucas mutações em caracteres descontínuos seriam suficientes para gerar uma nova espécie [Mutacionismo]. Deste modo a seleção natural e o gradualismo (acúmulo gradual de variações) eram desnecessários à evolução. 2- A ideia do Mutacionismo continuou até o princípio da síntese moderna, e em 1940, R. Goldschmidt publicou seu livro Material Basis of Evolution, invocando as mutações sistêmicas (que levam a grandes alterações) como a fonte de origem de novas espécies, uma teoria chamada de Saltacionismo. 3- No início do século XX também ressurgiram várias linhas de pensamento Neolamarckistas (ortogênese, características adquiridas etc). Ex: as pesquisas do austríaco Paul Kammerer nos seus trabalhos com anfíbios e a ortogênese do alemão Theodor Eimer.
Hugo de Vries (1900)
Hugo de Vries (1848–1935) The Mutation Theory [1901–1903]
A Teoria da Mutação
Prímula 2N Prímula 4N
Outros adeptos do Mutacionismo: Thomas Hunt Morgan, William Bateson etc. As ideias mutacionistas perduraram até 1940, com os trabalhos de Richard Goldschmidt.
Mutacionismo extremo: O Saltacionismo de Richard Goldschmidt
Sabe-se atualmente que a Evolução ocorre majoritariamente de forma gradual, mas existe a possibilidade de algumas raras características se originarem a partir de mudanças fenotípicas bruscas, geralmente envolvendo mutações pleiotrópicas.
Termitoxenia

01/04/2011
2
Alguns experimentos pareciam apoiar a Evolução Lamarckiana
• O herpetólogo austríaco Paul Kammerer (1920) fez experimentos com salamandras e sapos, cujos fenótipos pareciam ter sua herdabilidade alterada após exposição a diferentes ambientes.
• Nos estudos com o sapo (midwife toad), resultados surpreendentes foram publicados, mas Kammerer nunca deixava outros pesquisadores inspecionarem diretamente os animais, apenas as fotos. Em 1926, dois especialistas examinaram seu último e único espécime fixado e concluíram que este tinha sido injetado com tinta para reproduzir o aparecimento de uma pata nupcial. Seis semanas após a revelação da fraude, Kammerer cometeu suicídio.
A Síntese Moderna (Neodarwinismo) - 1920 a 1950 Na ânsia de renovar as idéias de Darwin, surgiu o Neodarwinismo (ou a Síntese Moderna). Esta foi a reconciliação dos dados de biogeografia, paleontologia e sistemática utilizados por Darwin no livro “Origem das Espécies” com os princípios da genética.
Os principais avanços da Síntese Moderna foram: 1. demonstração de que caracteres adquiridos não eram herdados; 2. comprovação de que a variação contínua tem a mesma base mendeliana, mas depende da interação de muitos genes; 3. a variação dentro e entre regiões geográficas tem base genética, e algumas delas tem sentido adaptativo; 4. os sistematas demonstraram que as espécies não são tipos morfológicos, mas populações de formas variáveis e isoladas reprodutivamente de outras populações.
Genética populacional Hardy e Weinberg desenvolveram independentemente um modelo matemático representando a distribuição das frequências gênicas e genotípicas em populações. No entanto, três grandes matemáticos e pais da estatística atual: R.A. Fisher, J.B.S. Haldane e S. Wright desenvolveram este modelo e demonstraram a “ação” dos fatores evolutivos: Seleção Natural, Deriva Genética, Fluxo gênico, Endogamia, Mutações, etc. O mecanismo de Seleção Natural foi evidenciado como o mais efetivo e determinístico (pelo menos a curto prazo) nas alterações de frequências gênicas de uma geração para outra, e portanto o mecanismo principal de evolução. Em 1937, Theodosius Dobzhanski publicou “Genetics and the Origin of Species”, no qual foi feita a síntese completa da genética e evolução. Ele foi também um dos primeiros cientistas a relatar um evento de especiação (não induzida) em laboratório, a qual ele chamou de Drosophila paulistorun (porque foi coletada em São Paulo).
Equilíbrio de Hardy-Weinberg
p2 + 2pq + q2 = 1
John B.S. Haldane
Ronald Fisher
Sewall Wright
Fundadores da Síntese Moderna
Cálculos de diversidade genética Índices de endogamia (F) Análises de variância Efeito da Seleção (S e W) Testes de diferenciação populacional Interação deriva e seleção Estruturação populacional Cálculos estatísticos diversos
A Síntese Moderna (Neodarwinismo)
1937
Theodosius Dobzhansky

01/04/2011
3
Outros fundadores da Síntese Moderna
Julian Huxley – divulgação, Unesco, WWF Ernst Mayr - especiação
George G. Simpson - paleontologia George L. Stebbins - botânica
Contribuições da Genética de Laboratório 1. Existe uma diferença entre fenótipo (características físicas observadas) e genótipo (conjunto de genes do indivíduo), e as diferenças fenotípicas entre os organismos individuais são explicadas por diferenças genotípicas e influência ambiental.
2. Efeitos ambientais sobre o fenótipo individual não afetam os genes passados para a prole (caracteres adquiridos não são hereditários). No entanto, o ambiente pode afetar a expressão gênica (flexibilidade/plasticidade fenotípica, etc).
3. As variações hereditárias são baseadas em “partículas” – genes – que retêm sua identidade com o passar das gerações. Genes não se misturam com outros genes, mesmo no caso de herança poligênica.
4. Genes sofrem mutação para formas alternativas (alelos). O efeito fenotípico de tais mutações pode variar do inobservável ao muito grande.
5. Fatores ambientais (radioativos, químicos) podem afetar a taxa de mutação, mas eles não dirigem preferencialmente a produção de mutações que seriam favoráveis no ambiente específico do organismo.
Contribuições da Teoria Matemática da Genética de Populações 6. A mudança evolutiva é um processo populacional: há uma alteração na frequência relativa (proporção) de organismos individuais com diferentes genótipos (e, às vezes, com diferentes fenótipos) dentro de uma população.
7. A taxa de mutação é muito baixa para levar uma população inteira de um genótipo para outro. A mudança na proporção do genótipo dentro de uma população pode ocorrer por deriva genética (fenômenos aleatórios/estocásticos) ou pelo valor adaptativo (fitness) de sobrevivência e/ou reprodução associado ao fenótipo e seu genótipo (seleção natural).
8. Mesmo uma leve intensidade da seleção natural pode (sob certas circunstâncias) levar a mudanças evolucionárias substanciais em um tempo relativamente curto.
Contribuições da Genética de Populações 9. A seleção natural pode alterar as populações entre gerações para além da extensão original da variação por incremento da proporção de alelos que, por recombinação com outros genes que afetam a mesma característica, dá surgimento a novos fenótipos.
10. As populações naturais são geneticamente variáveis.
11. Populações de uma espécie em diferentes regiões geográficas diferem em características que têm uma base genética: um genótipo que é raro em uma região pode ser predominante em outra.
12. A diferença em cada característica é freqüentemente baseada em diferenças em vários genes (poligenia), cada um dos quais tem um pequeno efeito genotípico.
13. Seleção Natural ocorre em populações naturais no tempo presente.
Contribuições da Sistemática 14. Diferenças entre populações geograficamente separadas de uma espécie são frequentemente adaptativas.
15. Organismos não pertencem necessariamente a espécies diferentes em função de diferenças em uma ou mais características fenotípicas.
16. No entanto, existe um continuum no grau de diferenciação das populações, com relação às diferenças fenotípicas e ao grau de isolamento reprodutivo, que vai de populações ligeiramente diferenciadas a espécies completamente distintas. Essa observação provê evidências de que uma espécie ancestral pode diferenciar-se em duas ou mais espécies diferentes por um acúmulo gradual de pequenas diferenças ao longo das gerações, envolvendo mutação, mas principalmente Deriva e Seleção Natural.
17. A especiação – origem de duas ou mais espécies a partir de um ancestral comum – normalmente ocorre através de diferenciação genética entre populações isoladas geograficamente (alopatria).
18. Os níveis taxonômicos mais altos surgem através de prolongado e sequencial acúmulo de pequenas diferenças, ao invés de uma origem mutacional súbita a partir de tipos drasticamente novos (Saltacionismo).
Contribuições da Paleontologia 19. O registro fóssil inclui muitas lacunas entre tipos totalmente diferentes de organismos, como também lacunas entre possíveis antepassados e descendentes. Tais lacunas podem ser explicadas pela incompletude do registro fóssil. Mas o registro fóssil também inclui vários exemplos de gradações do ancestral para o descendente. Junto com o argumento 18, isso nos leva à conclusão de que a evolução de grandes diferenças ocorre por pequenos passos. Dados do registro fóssil são muito importantes para detalhar a história evolutiva de alguns grupos específicos cuja fossilização foi favorecida, mas não há como utilizá-lo para o estudo detalhado de qualquer linhagem específica.
20. Consequentemente, todas as observações do registro fóssil são consistentes com os princípios precedentes da mudança evolutiva. Não há necessidade de invocar – e, aliás, existem evidências contra – qualquer hipótese não-darwiniana tais como mecanismos lamarckianos, teoria da ortogênese, vitalismo (“impulso interno”), ou origens abruptas por grandes mutações (mutacionismo e saltacionismo).
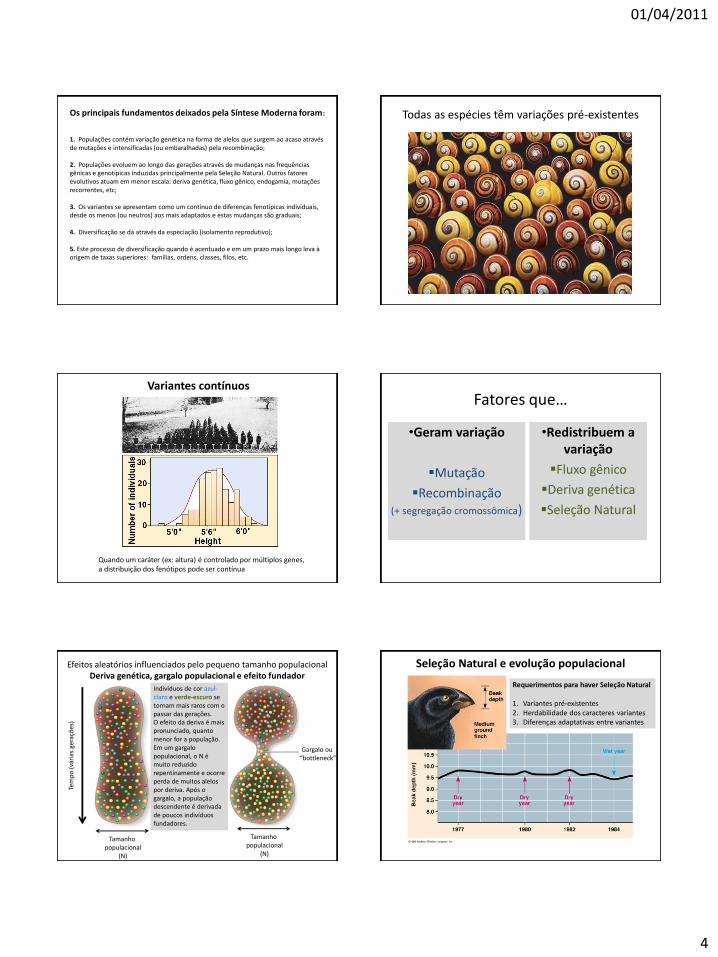
01/04/2011
4
Os principais fundamentos deixados pela Síntese Moderna foram: 1. Populações contém variação genética na forma de alelos que surgem ao acaso através de mutações e intensificadas (ou embaralhadas) pela recombinação; 2. Populações evoluem ao longo das gerações através de mudanças nas frequências gênicas e genotípicas induzidas principalmente pela Seleção Natural. Outros fatores evolutivos atuam em menor escala: deriva genética, fluxo gênico, endogamia, mutações recorrentes, etc; 3. Os variantes se apresentam como um contínuo de diferenças fenotípicas individuais, desde os menos (ou neutros) aos mais adaptados e estas mudanças são graduais; 4. Diversificação se dá através da especiação (isolamento reprodutivo); 5. Este processo de diversificação quando é acentuado e em um prazo mais longo leva à origem de taxas superiores: famílias, ordens, classes, filos, etc.
Todas as espécies têm variações pré-existentes
Variantes contínuos
Quando um caráter (ex: altura) é controlado por múltiplos genes, a distribuição dos fenótipos pode ser contínua
Fatores que…
•Geram variação
Mutação
Recombinação (+ segregação cromossômica)
•Redistribuem a variação
Fluxo gênico
Deriva genética
Seleção Natural
Efeitos aleatórios influenciados pelo pequeno tamanho populacional Deriva genética, gargalo populacional e efeito fundador
Tem
po
(vá
rias
ger
açõ
es)
Tamanho populacional
(N)
Tamanho populacional
(N)
Gargalo ou “bottleneck”
Indivíduos de cor azul-claro e verde-escuro se tornam mais raros com o passar das gerações. O efeito da deriva é mais pronunciado, quanto menor for a população. Em um gargalo populacional, o N é muito reduzido repentinamente e ocorre perda de muitos alelos por deriva. Após o gargalo, a população descendente é derivada de poucos indivíduos fundadores.
Seleção Natural e evolução populacional
Requerimentos para haver Seleção Natural 1. Variantes pré-existentes 2. Herdabilidade dos caracteres variantes 3. Diferenças adaptativas entre variantes

01/04/2011
5
Isolamento geográfico • Barreiras extrínsecas
(montanhas, rios, desertos etc) impedem o fluxo gênico entre populações de mesma espécie e promovem a diferenciação populacional e especiação.
Ammospermophilus harrisi A. leucurus
Especiação alopátrica
Camuflagem: uma consequência da seleção natural
(a) Um mantídeo floral da Malásia
(b) Mantídeo de Trinidad que mimetiza folhas
(c) Um mantídeo foliar na Costa Rica
Paleontologia e a história da vida no Planeta Terra
Mudança geológica e biológica (biogeografia histórica)
Figure 13.5
Descendência com modificação
Seleção artificial
Seleção natural