biosensors and bioelectronics - kaistbel.kaist.ac.kr/extfiles/papers/20130515.pdf · biosensors and...
TRANSCRIPT

Biosensors and Bioelectronics 46 (2013) 142–149
Contents lists available at SciVerse ScienceDirect
Biosensors and Bioelectronics
0956-56
http://d
n Corr
Institut
Tel.: þ8nn Cor
sung Bi
Tel.: þ8
E-m1 Th
journal homepage: www.elsevier.com/locate/bios
Rapid detection of exon 2-deleted AIMP2 mutation as a potentialbiomarker for lung cancer by molecular beacons
Seong-Min Jo a,1, Youngwook Kim c,1, Young-Su Jeong a, Young Hee Oh a,Keunchil Park c,d,nn, Hak-Sung Kim a,b,n
a Department of Biological Sciences, Korea Advanced Institute of Science and Technology (KAIST), Daejeon, Republic of Koreab Graduate School of Nanoscience and Technology, Korea Advanced Institute of Science and Technology (KAIST), Daejeon, Republic of Koreac Medical Nano-Element Development Center, Samsung Biomedical Research Institute, Seoul, Republic of Koread Division of Hematology–Oncology, Department of Medicine, Samsung Medical Center, Sungkyunkwan University, Seoul, Republic of Korea
a r t i c l e i n f o
Article history:
Received 23 January 2013
Received in revised form
24 February 2013
Accepted 25 February 2013Available online 6 March 2013
Keywords:
Molecular beacon
AIMP2-DX2
Diagnosis
Biomarker
Lung cancer
63/$ - see front matter & 2013 Elsevier B.V. A
x.doi.org/10.1016/j.bios.2013.02.037
esponding author at: Department of Biologic
e of Science and Technology (KAIST), Daejeon
2 42 350 2616; fax: þ82 42 350 2610.
responding author at: Medical Nano-Elemen
omedical Research Institute, Seoul, Republic o
2 2 3410 3450; fax: þ82 2 3410 1754.
ail addresses: [email protected] (K. Park), hskim
ese authors equally contributed to this work
a b s t r a c t
Exon 2 deletion in aminoacyl tRNA synthetase complex-interacting multifunctional protein 2 (AIMP2)
has been suggested to be associated with the progression of various cancers such as lung and ovarian
cancers. However, few studies have been conducted regarding detection and relevance of exon
2-deleted AIMP2 (AIMP2-DX2) mutation to a specific cancer. Here, we demonstrate the rapid and simple
detection of the AIMP2-DX2 mutation by molecular beacons and its relation to lung cancer. Real-time
PCR with molecular beacons allowed a sensitive detection of the AIMP2-DX2 mutation as low as 0.3 pg
initial template. Dual-conjugated liposomes with folate and molecular beacon enabled fluorescence
imaging of cancer cells harboring the AIMP2-DX2 mutation with high resolution. Association of the
AIMP2-DX2 mutation with lung cancer was shown by analyzing tissue samples from lung cancer patients
using real-time PCR. Approximately, 60% of lung cancer patients harbored the AIMP2-DX2 mutation,
which implies a potential of the AIMP2-DX2 mutation as a prognostic biomarker for lung cancer.
Molecular beacon-based approaches will find applications in the simple and rapid detection of mutations
on nucleotides for diagnosing and monitoring the progression of relevant cancer.
& 2013 Elsevier B.V. All rights reserved.
1. Introduction
Cancer is one of the most common life-threatening diseasesworldwide (Siegel et al., 2012). Over the past decades, significantadvances have been made methods to treat cancers. Along withthe development of efficacious therapies, early detection anddiagnosis of relevant cancers are considered to be crucial forreducing the mortality rates. Many factors cause transformationof normal to cancer cells, and genetic mutations in normal cellsare the most deleterious causes. In this regard, analysis of cancer-related mutations has generally been accepted to be the mosteffective way of diagnosing cancers as early as possible.
Recently, exon 2 deletion in aminoacyl tRNA synthetasecomplex-interacting multifunctional protein 2 (AIMP2) has
ll rights reserved.
al Sciences, Korea Advanced
, Republic of Korea.
t Development Center, Sam-
f Korea.
[email protected] (H.-S. Kim).
.
attracted considerable attention, because this mutation wassuggested to be associated with various cancers including lungand ovarian cancers (Choi et al., 2011, 2012; Kim et al., 2011).Wild-type AIMP2 composed of four exons was shown to play animportant role in cells as a tumor-suppressor, which have pro-apoptotic and anti-proliferative effects (Han et al., 2008; Choiet al., 2009). On the other hand, a splicing variant of AIMP2 withexon 2 deletion (AIMP2-DX2) occurs frequently not only invarious cancer cells, but also results in a loss of tumor-suppressor function. The AIMP2-DX2 mutant binds to TRAF2,which is a mediator for cell death signaling, but loses the role intriggering the interaction between TRAF2 and cIAP1, which is anubiquitin ligase, and consequently TNF-a-mediated cell death isinhibited. Unlike wild-type AIMP2, the AIMP2-DX2 mutant is notable to protect p53 from MDM2-mediated degradation, whichresults in the survival and growth of abnormal cells despiteserious damages or mutations. Therefore, the existence of theAIMP2-DX2 mutation provides important insight into the devel-opment and progression of cancerous cells as well as the status ofcancer patients. Nonetheless, no detailed studies have beenreported concerning analysis of the AIMP2-DX2 mutation andits relation with cancers. Direct sequencing has been mostcommonly used for detecting mutations, even though it is
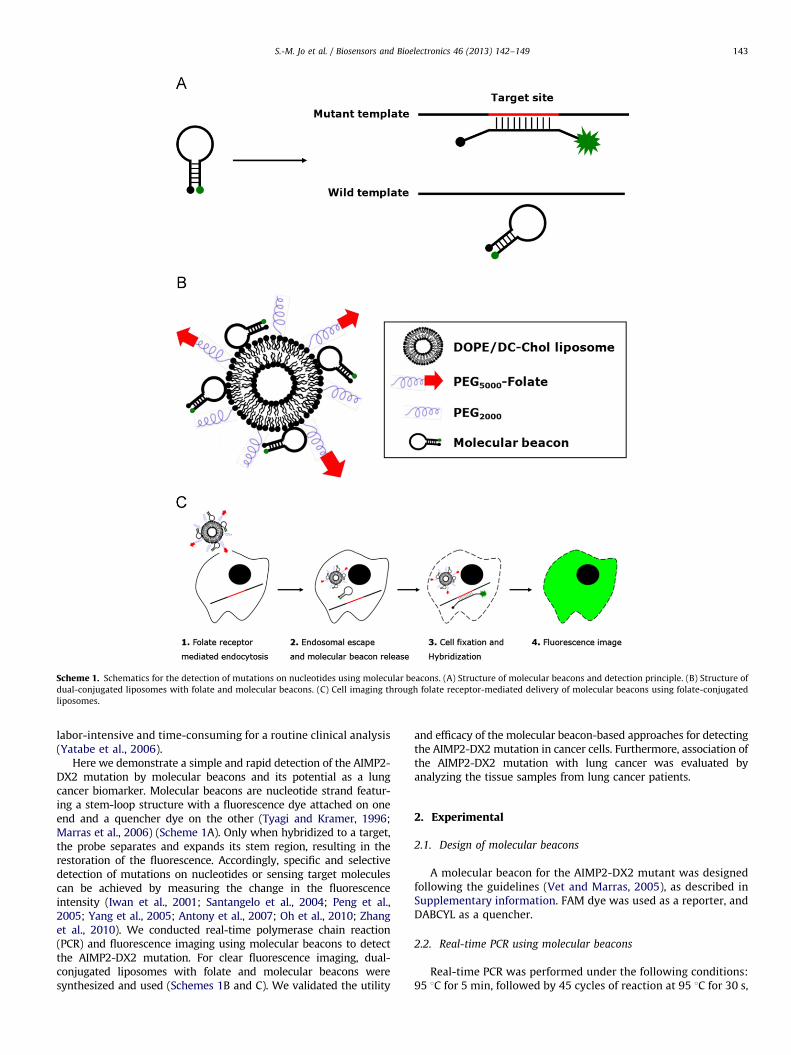
Scheme 1. Schematics for the detection of mutations on nucleotides using molecular beacons. (A) Structure of molecular beacons and detection principle. (B) Structure of
dual-conjugated liposomes with folate and molecular beacons. (C) Cell imaging through folate receptor-mediated delivery of molecular beacons using folate-conjugated
liposomes.
S.-M. Jo et al. / Biosensors and Bioelectronics 46 (2013) 142–149 143
labor-intensive and time-consuming for a routine clinical analysis(Yatabe et al., 2006).
Here we demonstrate a simple and rapid detection of the AIMP2-DX2 mutation by molecular beacons and its potential as a lungcancer biomarker. Molecular beacons are nucleotide strand featur-ing a stem-loop structure with a fluorescence dye attached on oneend and a quencher dye on the other (Tyagi and Kramer, 1996;Marras et al., 2006) (Scheme 1A). Only when hybridized to a target,the probe separates and expands its stem region, resulting in therestoration of the fluorescence. Accordingly, specific and selectivedetection of mutations on nucleotides or sensing target moleculescan be achieved by measuring the change in the fluorescenceintensity (Iwan et al., 2001; Santangelo et al., 2004; Peng et al.,2005; Yang et al., 2005; Antony et al., 2007; Oh et al., 2010; Zhanget al., 2010). We conducted real-time polymerase chain reaction(PCR) and fluorescence imaging using molecular beacons to detectthe AIMP2-DX2 mutation. For clear fluorescence imaging, dual-conjugated liposomes with folate and molecular beacons weresynthesized and used (Schemes 1B and C). We validated the utility
and efficacy of the molecular beacon-based approaches for detectingthe AIMP2-DX2 mutation in cancer cells. Furthermore, association ofthe AIMP2-DX2 mutation with lung cancer was evaluated byanalyzing the tissue samples from lung cancer patients.
2. Experimental
2.1. Design of molecular beacons
A molecular beacon for the AIMP2-DX2 mutant was designedfollowing the guidelines (Vet and Marras, 2005), as described inSupplementary information. FAM dye was used as a reporter, andDABCYL as a quencher.
2.2. Real-time PCR using molecular beacons
Real-time PCR was performed under the following conditions:95 1C for 5 min, followed by 45 cycles of reaction at 95 1C for 30 s,

S.-M. Jo et al. / Biosensors and Bioelectronics 46 (2013) 142–149144
66 1C for 30 s, and 72 1C for 30 s. The concentration of molecularbeacons was 160 nM, and the total cycling time was around 2 h(Supplementary information). The PCR products were subjectedto 2% agarose gel electrophoresis. Primers for amplification ofAIMP2 were used as follows: forward primer, 50-gcttcccacctgcatg-tacc-30; reverse primer: 50-ggtttgcgttgatcacgatgtc-30. For two-stepPCR, total RNA was extracted from cultured cells using a RNAextraction kit. Concentration of RNA was measured using a UVspectrometer (GeneQuant 1300, GE Healthcare, UK). Synthesis ofcomplementary DNA (cDNA) was carried out using dT15 primer,and the resulting cDNA was directly used for a two-step PCR.
2.3. In situ cell imaging with molecular beacons
H460 (human non-small-cell lung cancer) and WI-26 (humandiploid fibroblast from embryonic lung) cells were cultured onslides. Cells were fixed with cold acetone for 7 min. Fixed cellswere incubated with 200 nM of the molecular beacons solution at60 1C for 30 min. The slides were washed three times usingDulbecco’s phosphate-buffered saline (DPBS), and mountingmedia with 40,6-diamidino-2-phenylindole (DAPI) was added.Fluorescence images were obtained by a LSM 710 confocalmicroscope (Carl Zeiss, NY, USA) (Oh et al., 2010).
2.4. Fabrication and characterization of dual-conjugated liposomes
Folate-conjugated liposomes were firstly prepared by mixingdioleoylphosphoethanolamine (DOPE), dimethylaminoethanecarbamoyl cholesterol (DC-Chol), distearoylphosphoethanolamine-methoxy polyethylene glycol2000 (DSPE-mPEG2000), and distearyl-phosphoethanolamine-polyethylene glycol5000-folate (DSPE-PEG5000-folate) solutions in chloroform so that the molar ratio of DOPE:DC-Chol: DSPE-mPEG2000: DSPE-PEG5000-folate is 6: 4: 0.5: 0.1.Chloroform was evaporated in vacuum oven, and dried lipid filmwas hydrated with HEPES buffer (10 mM, pH 7.2) by hand shaking.The crude mixture was then sonicated for 10 min with a bath-typesonicator (Branson, CT, USA). The resulting liposomal suspension waskept at room temperature for 2 h, and adjusted to 1.2 mg/mL. Forfabrication of dual-conjugated liposomes with folate and molecularbeacons, 80 mL of the folate-conjugated liposome suspension wasmixed with 10 mL of molecular beacon suspension (100 mM) and10 mL of 10�PBS (pH 7.4), followed by incubation for 30 min at roomtemperature. Free molecular beacons were removed by centrifugationwith a 300 K spin filter. Molecular beacons for the AIMP2-DX2 mutantwas used as a positive control, and random molecular beacon wasused as a negative control (Table S1). Size and zeta potential of theliposomes were measured using a Zetasizer Nano ZS (MalvernInstruments, UK). Morphology of liposomes was observed by trans-mission electron microscopy (JEOL, Tokyo, Japan) with negativestaining using 2% phosphotungstic acid.
2.5. Cell imaging with dual-conjugated liposomes
HeLa (cervical cancer) cells were cultured on slides, followedby addition of 10 mL of dual-conjugated liposomes in 150 mL offolate-free and serum-free media. The cells were incubated withthe media containing the liposome for 1 h at 37 1C, and washedthree times using DPBS. The cells were further incubated withfolate and serum free media for 3 h at 37 1C, and fixed with 4%paraformaldehyde for 15 min. The resulting cells were incubatedat 60 1C for 40 min, followed by addition of mounting mediacontaining DAPI. Fluorescence images were observed by confocalmicroscopy.
2.6. Analysis of tissue samples from lung cancer patients
Tissue samples of 75 lung cancer patients were obtained fromthe Samsung Medical Center and analyzed to detect the AIMP2-DX2 mutation by real-time PCR (Supplementary information).
3. Results
3.1. Design of molecular beacons for the AIMP2-DX2 mutant
We first designed a molecular beacon for the AIMP2-DX2mutant. A loop region of molecular beacons complementary tothe AIMP2-DX2 mutant was designed to hybridize with DNA ormRNA of the AIMP2-DX2 mutant comprising twelve nucleotidesof exon 1 and nine nucleotides of exon 3. The stem region of themolecular beacon has a GC-rich sequence of six base-pairs.To maintain the stem-loop structure, cytosine was insertedbetween the loops and the stem region, because the junctionsequence between exon 1 and exon 3 was shown to give rise to aself-formed stem structure. The designed molecular beacon spe-cifically recognizes the junction sequence between exon 1 andexon 3 of the AIMP2-DX2 mutant so that the molecular beaconscan be opened (Scheme 1A). Nucleotide sequences of the designedmolecular beacons for the AIMP2-DX2 mutant and their targetsare shown in Table S1.
To determine the optimal hybridization temperatures forthe molecular beacon targeting the AIMP2-DX2 mutation(Supplementary information), we examined the thermal profilesof the designed molecular beacons as described in Supplementaryinformation. The changes in fluorescence signals were measuredat temperatures ranging from 95 1C to 15 1C in the presence andabsence of the synthetic targets, namely a target template with ajunction sequence between exon 1 and exon 3, wild-type 1 with asequence between exon 1 and exon 2, and wild-type 2 with asequence between exon 2 and exon 3 (Fig. S1). No difference influorescence intensity was observed at temperatures rangingfrom 95 1C to 75 1C. At these high temperatures, the stem-loopstructure of the molecular beacons is hard to maintain, becauseDNA is denatured. At low temperatures between 60 1C and 15 1C,the fluorescence intensities in the presence of the target templateand wild-type 2 were almost identical. This is mainly due to thefact that some nucleotides are conserved in both the targettemplate and wild-type 1, and that the molecular beacons bindpartially to wild-type 1. On the other hand, the fluorescenceintensities in the presence of the target template were consider-ably higher than those of wild-type 1 and 2 at temperaturesranging from 72 1C to 60 1C. This result indicated that thespecificity of the designed molecular beacons for the target mRNAand the hybridization at this temperature range. These tempera-tures are the ‘window of discrimination’. Based on these results,fluorescence intensity was monitored at 66 1C during real-timePCR in this study.
3.2. Detection of the AIMP2-DX2 mutation with molecular beacons
To verify the performance of the designed molecular beacons,real-time PCR was carried out at varying amounts of targettemplates inserted in plasmids. As shown in Fig. 1A, changes influorescence intensity were dependent on the amount of initialtemplate, and the threshold cycles were inversely proportional tothe amount of initial templates (R2
¼0.9905) (Fig. 1B). Moreover, themolecular beacons could detect the AIMP2-DX2 mutation as low as0.3 pg of the mutant template. Amplicons of the wild-type AIMP2and AIMP2-DX2 mutant were located on 326 bp and 119 bp,respectively (Fig. S2). Conditions for real-time PCR, such as optimal
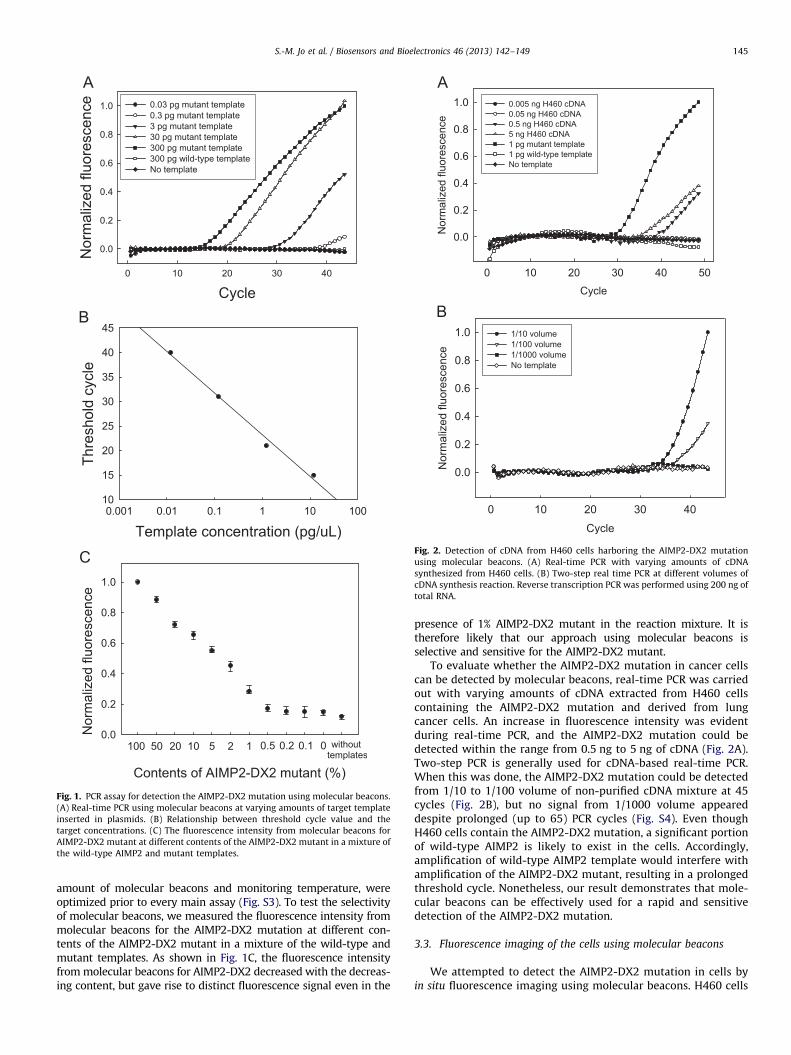
Fig. 1. PCR assay for detection the AIMP2-DX2 mutation using molecular beacons.
(A) Real-time PCR using molecular beacons at varying amounts of target template
inserted in plasmids. (B) Relationship between threshold cycle value and the
target concentrations. (C) The fluorescence intensity from molecular beacons for
AIMP2-DX2 mutant at different contents of the AIMP2-DX2 mutant in a mixture of
the wild-type AIMP2 and mutant templates.
Fig. 2. Detection of cDNA from H460 cells harboring the AIMP2-DX2 mutation
using molecular beacons. (A) Real-time PCR with varying amounts of cDNA
synthesized from H460 cells. (B) Two-step real time PCR at different volumes of
cDNA synthesis reaction. Reverse transcription PCR was performed using 200 ng of
total RNA.
S.-M. Jo et al. / Biosensors and Bioelectronics 46 (2013) 142–149 145
amount of molecular beacons and monitoring temperature, wereoptimized prior to every main assay (Fig. S3). To test the selectivityof molecular beacons, we measured the fluorescence intensity frommolecular beacons for the AIMP2-DX2 mutation at different con-tents of the AIMP2-DX2 mutant in a mixture of the wild-type andmutant templates. As shown in Fig. 1C, the fluorescence intensityfrom molecular beacons for AIMP2-DX2 decreased with the decreas-ing content, but gave rise to distinct fluorescence signal even in the
presence of 1% AIMP2-DX2 mutant in the reaction mixture. It istherefore likely that our approach using molecular beacons isselective and sensitive for the AIMP2-DX2 mutant.
To evaluate whether the AIMP2-DX2 mutation in cancer cellscan be detected by molecular beacons, real-time PCR was carriedout with varying amounts of cDNA extracted from H460 cellscontaining the AIMP2-DX2 mutation and derived from lungcancer cells. An increase in fluorescence intensity was evidentduring real-time PCR, and the AIMP2-DX2 mutation could bedetected within the range from 0.5 ng to 5 ng of cDNA (Fig. 2A).Two-step PCR is generally used for cDNA-based real-time PCR.When this was done, the AIMP2-DX2 mutation could be detectedfrom 1/10 to 1/100 volume of non-purified cDNA mixture at 45cycles (Fig. 2B), but no signal from 1/1000 volume appeareddespite prolonged (up to 65) PCR cycles (Fig. S4). Even thoughH460 cells contain the AIMP2-DX2 mutation, a significant portionof wild-type AIMP2 is likely to exist in the cells. Accordingly,amplification of wild-type AIMP2 template would interfere withamplification of the AIMP2-DX2 mutant, resulting in a prolongedthreshold cycle. Nonetheless, our result demonstrates that mole-cular beacons can be effectively used for a rapid and sensitivedetection of the AIMP2-DX2 mutation.
3.3. Fluorescence imaging of the cells using molecular beacons
We attempted to detect the AIMP2-DX2 mutation in cells byin situ fluorescence imaging using molecular beacons. H460 cells
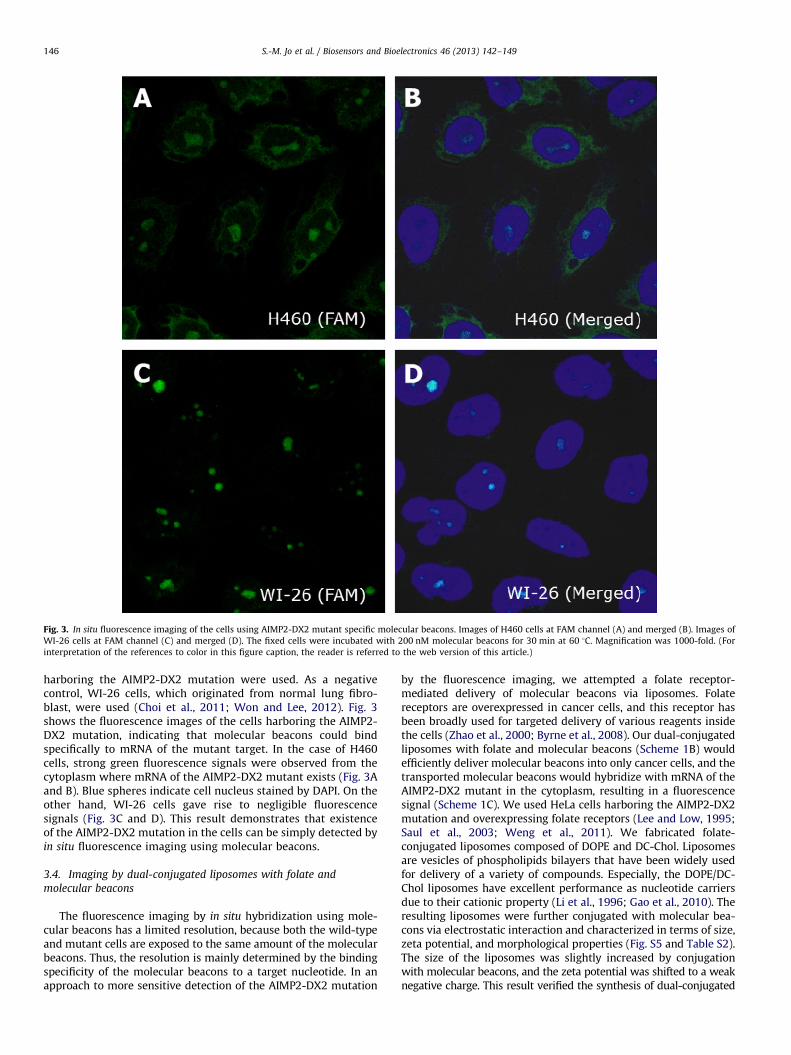
Fig. 3. In situ fluorescence imaging of the cells using AIMP2-DX2 mutant specific molecular beacons. Images of H460 cells at FAM channel (A) and merged (B). Images of
WI-26 cells at FAM channel (C) and merged (D). The fixed cells were incubated with 200 nM molecular beacons for 30 min at 60 1C. Magnification was 1000-fold. (For
interpretation of the references to color in this figure caption, the reader is referred to the web version of this article.)
S.-M. Jo et al. / Biosensors and Bioelectronics 46 (2013) 142–149146
harboring the AIMP2-DX2 mutation were used. As a negativecontrol, WI-26 cells, which originated from normal lung fibro-blast, were used (Choi et al., 2011; Won and Lee, 2012). Fig. 3shows the fluorescence images of the cells harboring the AIMP2-DX2 mutation, indicating that molecular beacons could bindspecifically to mRNA of the mutant target. In the case of H460cells, strong green fluorescence signals were observed from thecytoplasm where mRNA of the AIMP2-DX2 mutant exists (Fig. 3Aand B). Blue spheres indicate cell nucleus stained by DAPI. On theother hand, WI-26 cells gave rise to negligible fluorescencesignals (Fig. 3C and D). This result demonstrates that existenceof the AIMP2-DX2 mutation in the cells can be simply detected byin situ fluorescence imaging using molecular beacons.
3.4. Imaging by dual-conjugated liposomes with folate and
molecular beacons
The fluorescence imaging by in situ hybridization using mole-cular beacons has a limited resolution, because both the wild-typeand mutant cells are exposed to the same amount of the molecularbeacons. Thus, the resolution is mainly determined by the bindingspecificity of the molecular beacons to a target nucleotide. In anapproach to more sensitive detection of the AIMP2-DX2 mutation
by the fluorescence imaging, we attempted a folate receptor-mediated delivery of molecular beacons via liposomes. Folatereceptors are overexpressed in cancer cells, and this receptor hasbeen broadly used for targeted delivery of various reagents insidethe cells (Zhao et al., 2000; Byrne et al., 2008). Our dual-conjugatedliposomes with folate and molecular beacons (Scheme 1B) wouldefficiently deliver molecular beacons into only cancer cells, and thetransported molecular beacons would hybridize with mRNA of theAIMP2-DX2 mutant in the cytoplasm, resulting in a fluorescencesignal (Scheme 1C). We used HeLa cells harboring the AIMP2-DX2mutation and overexpressing folate receptors (Lee and Low, 1995;Saul et al., 2003; Weng et al., 2011). We fabricated folate-conjugated liposomes composed of DOPE and DC-Chol. Liposomesare vesicles of phospholipids bilayers that have been widely usedfor delivery of a variety of compounds. Especially, the DOPE/DC-Chol liposomes have excellent performance as nucleotide carriersdue to their cationic property (Li et al., 1996; Gao et al., 2010). Theresulting liposomes were further conjugated with molecular bea-cons via electrostatic interaction and characterized in terms of size,zeta potential, and morphological properties (Fig. S5 and Table S2).The size of the liposomes was slightly increased by conjugationwith molecular beacons, and the zeta potential was shifted to a weaknegative charge. This result verified the synthesis of dual-conjugated
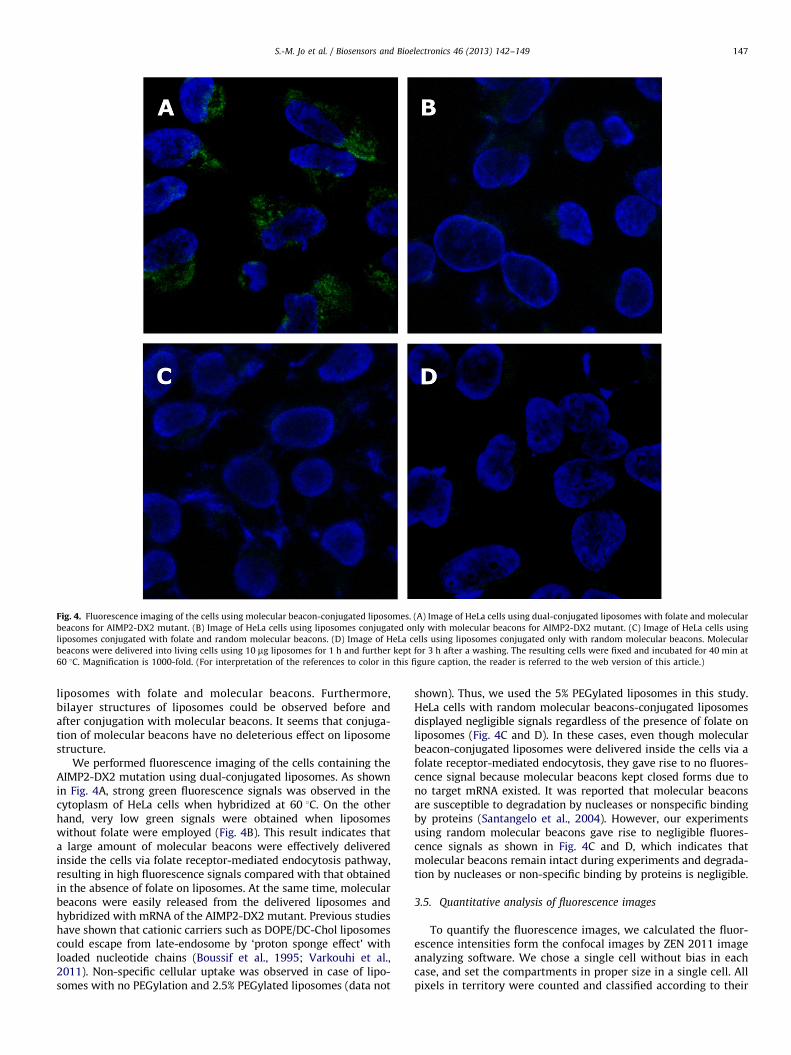
Fig. 4. Fluorescence imaging of the cells using molecular beacon-conjugated liposomes. (A) Image of HeLa cells using dual-conjugated liposomes with folate and molecular
beacons for AIMP2-DX2 mutant. (B) Image of HeLa cells using liposomes conjugated only with molecular beacons for AIMP2-DX2 mutant. (C) Image of HeLa cells using
liposomes conjugated with folate and random molecular beacons. (D) Image of HeLa cells using liposomes conjugated only with random molecular beacons. Molecular
beacons were delivered into living cells using 10 mg liposomes for 1 h and further kept for 3 h after a washing. The resulting cells were fixed and incubated for 40 min at
60 1C. Magnification is 1000-fold. (For interpretation of the references to color in this figure caption, the reader is referred to the web version of this article.)
S.-M. Jo et al. / Biosensors and Bioelectronics 46 (2013) 142–149 147
liposomes with folate and molecular beacons. Furthermore,bilayer structures of liposomes could be observed before andafter conjugation with molecular beacons. It seems that conjuga-tion of molecular beacons have no deleterious effect on liposomestructure.
We performed fluorescence imaging of the cells containing theAIMP2-DX2 mutation using dual-conjugated liposomes. As shownin Fig. 4A, strong green fluorescence signals was observed in thecytoplasm of HeLa cells when hybridized at 60 1C. On the otherhand, very low green signals were obtained when liposomeswithout folate were employed (Fig. 4B). This result indicates thata large amount of molecular beacons were effectively deliveredinside the cells via folate receptor-mediated endocytosis pathway,resulting in high fluorescence signals compared with that obtainedin the absence of folate on liposomes. At the same time, molecularbeacons were easily released from the delivered liposomes andhybridized with mRNA of the AIMP2-DX2 mutant. Previous studieshave shown that cationic carriers such as DOPE/DC-Chol liposomescould escape from late-endosome by ‘proton sponge effect’ withloaded nucleotide chains (Boussif et al., 1995; Varkouhi et al.,2011). Non-specific cellular uptake was observed in case of lipo-somes with no PEGylation and 2.5% PEGylated liposomes (data not
shown). Thus, we used the 5% PEGylated liposomes in this study.HeLa cells with random molecular beacons-conjugated liposomesdisplayed negligible signals regardless of the presence of folate onliposomes (Fig. 4C and D). In these cases, even though molecularbeacon-conjugated liposomes were delivered inside the cells via afolate receptor-mediated endocytosis, they gave rise to no fluores-cence signal because molecular beacons kept closed forms due tono target mRNA existed. It was reported that molecular beaconsare susceptible to degradation by nucleases or nonspecific bindingby proteins (Santangelo et al., 2004). However, our experimentsusing random molecular beacons gave rise to negligible fluores-cence signals as shown in Fig. 4C and D, which indicates thatmolecular beacons remain intact during experiments and degrada-tion by nucleases or non-specific binding by proteins is negligible.
3.5. Quantitative analysis of fluorescence images
To quantify the fluorescence images, we calculated the fluor-escence intensities form the confocal images by ZEN 2011 imageanalyzing software. We chose a single cell without bias in eachcase, and set the compartments in proper size in a single cell. Allpixels in territory were counted and classified according to their

S.-M. Jo et al. / Biosensors and Bioelectronics 46 (2013) 142–149148
fluorescence intensities. Fig. S6A displays the quantified results ofimages obtained from H460 cells and WI-26 cells by in situ
hybridization, respectively. Much stronger green fluorescenceintensities were obtained from H460 cells, whereas negligiblegreen signals were observed from WI-26 cells. Most of pixelsshowing high fluorescence intensity were observed from H460cells (cancer cells), whereas WI-26 cells, non-tumorigenic lungfibroblast harboring normal levels of AIMP2-DX2 mutation, gen-erated few pixels exhibiting the intensity above 15. Therefore, thethreshold intensity distinguishing the tumorigenic cells fromnormal cells was determined to be 15. This result is coincidentwith transcription levels of mRNA in respective cell lines asobserved in PCR band (Choi et al., 2011; Won and Lee, 2012).The use of dual-conjugated liposomes for the fluorescence ima-ging of cells also led to stronger fluorescence intensities comparedwith that of only molecular beacon-conjugated liposomes (Fig.S6B). Imaging by dual-conjugated liposomes with folate andmolecular beacons only generated high intensity pixels. Thisresult indicates that folate receptor-mediated delivery of mole-cular beacons via liposomes is very effective for the fluorescenceimaging of the cells containing specific mutations.
3.6. Detection of the AIMP2-DX2 mutation in tissue samples from
lung cancer patients
To get some insight into a potential of the AIMP2-DX2mutation as a biomarker for lung cancer, we attempted to detectthis mutation in tissue samples from 75 lung cancer patientsby real-time PCR with molecular beacons (Supplementaryinformation). By using 200 ng of total RNA as an initial template,two-step PCR was performed under the same conditionsdescribed above, and the results were compared with end-pointPCR. As shown in Table 1, the AIMP2-DX2 mutation was detectedin 38 samples, and 26 samples had only wild-type AIMP2 (1sttrial). Three samples displayed discrepancy between PCR bandand molecular beacon signals, and eight samples resulted infailure of PCR assay. As a result, around 60% of lung cancerpatients was estimated to harbor the AIMP2-DX2 mutation.Additional experiment using another primer set (2nd trial) alsodisplayed a similar result, indicating that approximately 66.0% ofthe patients harbored the AIMP2-DX2 mutation. Correlationbetween first trial and second trials was 77.1% in overlapped 48valid cases. Small amount of the AIMP2-DX2 mRNA might bepresent in cancer cells, and the purity of clinical samples wouldbe considerably low. In addition, mRNAs from the wild-typeAIMP2 and AIMP2-DX2 mutant will compete for the sameprimers, because their binding regions are exactly same. In otherwords, amplification of wild-type AIMP2 mRNA is more dominantthan the AIMP2-DX2 mutant. These considerations could explainthe 22.9% discrepancy between the first and second trials, and thisdiscrepancy seemed not to be caused by malfunction of the
Table 1Real-time PCR of tissue samples from lung cancer patients using molecular
beacons.
Characteristic 1st Trial
(N¼75)
2nd Trial
(N¼75)
Valid cases 64 56
AIMP2-DX2 positive 38 (59.4%) 37 (66.1%)
AIMP2-DX2 negative 26 (40.6%) 19 (33.9%)
Overlapped 48
Correlation between 1st and 2nd
trial
37 (77.1%)
Inconsistence between band and signal 3 6
Failed PCR 8 13
molecular beacons. Our result indicates that the AIMP2-DX2mutation occurred at considerable rate in lung cancer patients,implying that this mutation has a great potential to be a lungcancer biomarker.
4. Discussion
We have demonstrated that molecular beacon-based approachesare very effective for detecting the AIMP2-DX2 mutation, and thatthis mutation is closely associated with lung cancer. The AIMP2-DX2mutation was shown to compromise the tumor suppressive abilityof the wild type AIMP2, and this mutation occurs at high rates inlung and ovarian cancers (Choi et al., 2011, 2012; Kim et al., 2011).In this regard, a simple and fast detection of the AIMP2-DX2mutation can be useful for prognostics of lung and ovarian cancers.Molecular beacons are versatile fluorescent probes, which can beused for detection of mutations in a target nucleotide instead ofdirect sequencing. We used FAM dye and DABCYL as a report dyeand a fluorescence quencher, respectively, to construct molecularbeacons. FAM dye has been widely used for many biological assayssuch as real-time PCR and in situ hybridization although its quantumyield was shown to decrease at high temperatures (Marras, 2006).In this study, however, no significant decrease in the fluorescenceintensity was observed, which implies that thermal decay of FAMdye is negligible during a short assay period.
By using real-time PCR and fluorescence imaging approaches,we could detect the AIMP2-DX2 mutation in different types ofcancer cells. Detection sensitivity of the AIMP2-DX2 mutationwas about 0.3 pg of the mutant template. Starting from cDNA,which corresponds to 2 ng of total RNA, we could identify theexistence of the AIMP2-DX2 mutation. The amount of RNA in asingle cell was estimated to range from 10 to 50 pg (Livesey,2003; White et al., 2011). Thus, the molecular beacon approachmight allow the detection of the AIMP2-DX2 mutation fromaround 100 cells. Real-time PCR is facile, sensitive, and widelyused in the general laboratory, and molecular beacon-based real-time PCR could be useful for detecting mutations clinically.Fluorescent imaging techniques could eventually lead to theidentification of the AIMP2-DX2 mutation in a single cell withoutan amplification step. Therefore, both approaches can be appliedto the detection of the mutations from cell-free nucleic acids andcirculating tumor cells due to their high sensitivity and require-ment of small amount of sample (Schwarzenbach et al., 2011).The ratio of mRNA between AIMP2-DX2 mutant and wild-typeAIMP2 would provide more insight into the development andprogression of related cancers. Folate receptor-mediated deliveryof molecular beacons via folate-conjugated liposomes was moreeffective for detecting the AIMP2-DX2 mutation in the cells byfluorescence imaging.
Our results show that the AIMP2-DX2 mutation can be devel-oped as a potential biomarker for lung cancer, because theoccurrence rate of the mutation was above 60% in a patient group.Previous study showed that 44% of non-small-cell lung cancerpatients have high occurrence rate of the AIMP2-DX2 mutation intumor tissues compared with normal tissues (Choi et al., 2011).Therefore, a high rate of the AIMP2-DX2 mutation is likely to beclosely associated with lung cancer. In addition, the AIMP2-DX2mutation was suggested to play important roles in tumor survivalwith poor prognosis. Thus, existence of the AIMP2-DX2 offers animportant guide for treating cancer patients. Expression of theAIMP2-DX2 mutant could be suppressed by specific small inter-fering RNA that blocks transcription of the target mRNAs (Choiet al., 2011). Moreover, a recent study revealed that chemo-sensitivities for paclitaxel and cisplatin are significantly increased

S.-M. Jo et al. / Biosensors and Bioelectronics 46 (2013) 142–149 149
when expression of the AIMP2-DX2 mutant is suppressed (Choiet al., 2012).
5. Conclusion
In summary, we have demonstrated that the AIMP2-DX2mutation can be effectively detected by molecular beacons.Fluorescent imaging using dual-conjugated liposomes clearlyidentified the AIMP2-DX2 mutation in a single cell without anamplification step. Therefore, both approaches can be applied tothe detection of the mutations from cell-free nucleic acids andsingle cells. We believe that molecular beacon-based approachwill provide the information regarding the patient status andtherapeutic guideline.
Acknowledgments
This research was supported by the Korea Health 21C R&DProject (A040041) of Ministry for Health, Welfare and FamilyAffairs, and the Pioneer Research Program through the NationalResearch Foundation of Korea funded by the Ministry of Educa-tion, Science and Technology (20082000218).
Appendix A. Supporting information
Supplementary data associated with this article can be found inthe online version at http://dx.doi.org/10.1016/j.bios.2013.02.037.
References
Antony, K.C., Mark, A.B., Andrew, T., 2007. Nucleic Acids Research 35, e105.Boussif, O., Lezoualc’h, F., Zanta, M.A., Mergny, M.D., Scherman, D., Demeneix, B.,
Behr, J.P., 1995. Proceedings of the National Academy of Science 92,7297–7301.
Byrne, J.D., Betancourt, T., Brannon-Peppas, L., 2008. Advanced Drug DeliveryReviews 60, 1615–1626.
Choi, J.W., Kim, D.G., Lee, A.-E., Kim, H.R., Lee, J.Y., Kwon, N.H., Shin, Y.K., Hwang,S.-K., Chang, S.-H., Cho, M.-H., Choi, Y.-L., Kim, J., Oh, S.H., Kim, B., Kim, S.-Y.,
Jeon, S., Park, J.Y., Kang, H.P., Park, B.J., Han, J.M., Kim, S., 2011. PLoS Genetics 7,e1001351.
Choi, J.W., Kim, D.G., Park, M.C., Um, J.Y., Han, J.M., Park, S.G., Choi, E.-C., Kim, S.,2009. Journal of Cell Science 122, 2710–2715.
Choi, J.W., Lee, J.-W., Kim, J.K., Jeon, H.-K., Choi, J.-J., Kim, D.G., Kim, B.G., Nam, D.-H.,Kim, H.J., Yun, S.H., Kim, S., 2012. Journal of Molecular Cell Biology 4, 164–173.
Gao, J., Sun, J., Li, H., Liu, W., Zhang, Y., Li, B., Qian, W., Wang, H., Chen, J., Guo, Y.,2010. Biomaterials 31, 2655–2664.
Han, J.M., Park, B.-J., Park, S.G., Oh, Y.S., Choi, S.J., Lee, S.W., Hwang, S.-K., Chang, S.-H.,Cho, M.-H., Kim, S., 2008. Proceedings of the National Academy of Science 105,11206–11211.
Iwan, W., Sebastian, C.B., Fred, H., Nabil, D., Angela, V., Klaus, A., Sucharit, B., 2001.Proceedings of the National Academy of Science 98, 3185–3190.
Kim, S., You, S., Hwan, D., 2011. Nature Reviews Cancer 11, 708–718.Lee, R.J., Low, P.S., 1995. Biochimica et Biophysica Acta-Biomembranes 1233,
134–144.Li, S., Gao, X., Son, K., Sorgi, F., Hofland, H., Huang, L., 1996. Journal of Controlled
Release 39, 373–381.Livesey, F.J., 2003. Briefings in Functional Genomics 2, 31–36.Marras, S.A., Tyagi, S., Kramer, F.R., 2006. Clinica Chimica Acta 363, 48–60.Marras, S.A., 2006. in: Didenko, V.V., (Ed.) Methods in Molecular Biology, Humana
Press Inc., Totowa, NJ, Vol. 335, pp. 3–16.Oh, Y.H., Kim, Y., Kim, Y.P., Seo, S.W., Mitsudomi, T., Ahn, M.J., Park, K., Kim, H.S.,
2010. Journal of Molecular Diagnostics 12, 644–652.Peng, X.H., Cao, Z.H., Xia, J.T., Carlson, G.W., Lewis, M.M., Wood, W.C., Yang, L.,
2005. Cancer Research 65, 1909–1917.Santangelo, P.J., Nix, B., Tsourkas, A., Bao, G., 2004. Nucleic Acids Research 32, e57.Saul, J.M., Annapragada, A., Natarajan, J.V., Bellamkonda, R.V., 2003. Journal of
Controlled Release 92, 49–67.Schwarzenbach, H., Hoon, D.S.B., Pantel, K., 2011. Nature Reviews Cancer 11,
426–437.Siegel, R., Naishadham, D., Jemal, A., 2012. CA-Cancer Journal of Clinicians 62,
10–29.Tyagi, S., Kramer, F.R., 1996. Nature Biotechnology 14, 303–308.Varkouhi, A.K., Scholte, M., Storm, G., Haisma, H.J., 2011. Journal of Controlled
Release 151, 220–228.Vet, J.A., Marras, S.A., 2005. in: Herdewijn, P., (Ed.) Methods in Molecular Biology,
Humana Press Inc., Totowa, NJ, Vol. 288, pp. 273–289.Weng, J., Zhang, Z., Sun, L., Wang, J.A., 2011. Biosensors and Bioelectronics 26,
1847-1852.White, A.K., VanInsberghe, M., Petriv, O.I., Hamidi, M., Sikorski, D., Marra, M.A.,
Piret, J., Aparicio, S., Hansen, C.L., 2011. Proceedings of the National Academyof Science 108, 13999–14004.
Won, Y.S., Lee, S.W., 2012. Journal of Biotechnology 158, 44–49.Yang, L., Cao, Z., Li,n, Y., Wood, W.C., Staley, C.A., 2005. Cancer Biology and Therapy
4, 561–570.Yatabe, Y., Hida, T., Horio, Y., Kosaka, T., Takahashi, T., Mitsudomi, T., 2006. Journal
of Molecular Diagnostics 8, 335–341.Zhang, X.-B., Wang, Z., Xing, H., Xiang, Y., Lu, Y., 2010. Analytical Chemistry 82,
5005–5011.Zhao, X., Li, H., Lee, R.J., 2000. Advanced Drug Delivery Reviews 41, 147–162.