both α-haemolysin determinants contribute to full virulence of uropathogenic escherichia coli...
TRANSCRIPT

Microbes and Infection 8 (2006) 2006e2012www.elsevier.com/locate/micinf
Original article
Both a-haemolysin determinants contribute to full virulence ofuropathogenic Escherichia coli strain 536
Gabor Nagy a,b, Artur Altenhoefer b, Oliver Knapp c, Elke Maier c, Ulrich Dobrindt b,Gabriele Blum-Oehler b, Roland Benz c, Levente EmTdy a, Jorg Hacker b,*
a Department of Medical Microbiology and Immunology, University of Pecs, Szigeti ut 12, 7624 Pecs, Hungaryb Institut fur Molekulare Infektionsbiologie, Universitat Wurzburg, Rontgenring 11, 97070 Wurzburg, Germanyc Lehrstuhl fur Biotechnologie, Biozentrum der Universitat Wurzburg, Am Hubland, 97074 Wurzburg, Germany
Received 8 December 2005; accepted 27 February 2006
Available online 2 June 2006
Abstract
Uropathogenic Escherichia coli strain 536 possesses two intact copies of the a-haemolysin determinant localised on distinct pathogenicityislands. The coding regions of the two hlyCABD operons are conserved; however, upstream sequences are entirely dissimilar. Consequently,expression of the encoded toxin molecules in vitro is highly different. On the other hand, the contribution of the individual determinants tothe strain’s virulence is the same. Isogenic mutants lacking individual hly determinants have a similar increase in LD50 value in a mouse modelof urinary tract infection. Mouse lung toxicity as well as in vitro assays reveals a significant decrease in acute cytotoxicity of both mutants incomparison to the parent wild-type strain; however, the two hly mutants do not significantly differ from each other in these respects. Singlechannel recordings show no difference in electrophysiological characteristics of the pores formed by the individual HlyA molecules on syntheticplanar lipid membranes. Nor do the paralogues have any target cell preference in an in vitro cytotoxicity assay. Our data suggest that the two hlyparalogues encode identical toxin functions; however, due to different regulation of expression, they participate at distinct stages of the infectiousprocess. Interestingly, the unrelated uropathogenic E. coli strain J96 shares the same two hly alleles, suggesting that acquisition of the two pa-ralogues accorded a selective evolutionary advantage.� 2006 Elsevier SAS. All rights reserved.
Keywords: Uropathogenic Escherichia coli; a-Haemolysin; Gene duplication; Cytotoxicity; Virulence regulation; Pore formation
1. Introduction
The Escherichia coli pore-forming toxin a-haemolysin isthe best-characterised representative of the so-called RTXtoxin family. Although evidence has been shown that a-haemolysin production contributes to intestinal infections as
Abbreviations: bp, base pair; CFU, colony-forming unit; LB, Luriae
Bertani medium; LD50, 50% lethal dose; LDH, lactate dehydrogenase; MOI,
multiplicity of infection; OD, optical density; PAGE, polyacrylamide gel
electrophoresis; PAI, pathogenicity island; PCR, polymerase chain reaction;
TCA, trichloroacetic acid; UPEC, uropathogenic Escherichia coli.
* Corresponding author. Tel.: þ49 931 312575; fax: þ49 931 312578.
E-mail address: [email protected] (J. Hacker).
1286-4579/$ - see front matter � 2006 Elsevier SAS. All rights reserved.
doi:10.1016/j.micinf.2006.02.029
well [1], this toxin is mainly associated with E. coli strainscausing extraintestinal infections. While only 12% of com-mensal E. coli strains are haemolytic, this value is significantlyhigher among strains responsible for asymptomatic bacteriuria(20%), cystitis (40%), pyelonephritis (49%), and bacteriaemia(39%), indicating a prominent role of this toxin in extraintes-tinal infections [2].
The synthesis, activation and secretion of a-haemolysin areencoded on the hlyCABD operon. The unusual codon usage aswell as the low G-C content within the operon suggests thatthis determinant has been acquired by E. coli through horizon-tal gene transfer [3]. Indeed, animal pathogenic E. coli strainscarry the hly determinant on large conjugative plasmids,whereas human isolates have acquired them as parts of large

2007G. Nagy et al. / Microbes and Infection 8 (2006) 2006e2012
pathogenicity islands (PAIs) [4]. While the protein-coding se-quence within hly operons of different origin seems to be con-served, upstream regions can be highly variable [5]. Thedifferent promoter regions direct different toxin expressionand consequently various activity in vitro [6] and in vivo [7].
Both unrelated prototype uropathogenic E. coli (UPEC)strains 536 (O6:K15) and J96 (O4:K6) carry two paraloguesof hlyCABD. The hly determinants are not linked; they are car-ried on distinct PAIs of the two strains that do not, or only par-tially, resemble to each other [8e10]. We show below thatconserved pairs of hly determinants could be distinguishedin the two UPEC strains with respect to their upstream regions.In strain 536, both determinants contribute equally to thestrain’s virulence, suggesting that the two paralogues with dif-ferent conserved upstream regions supply selective evolution-ary advantage to E. coli pathogens.
2. Materials and methods
2.1. Bacterial strains, cell lines and culture conditions
Bacterial strains, plasmids and primers used in this studyare shown in Table 1. The wild-type strain 536 was isolatedfrom a patient suffering from acute pyelonephritis and hasbeen thoroughly investigated. This strain carries two intactcopies of hlyCABD, both of which are located on distinct path-ogenicity islands [10]. The spontaneous mutant 536-21 lacksboth PAIs carrying the hly determinants [11]. The isogenichly mutants were constructed using standard procedures[12]. Briefly, primer pair Dhly1-fw þ Dhly1-rev or Dhly2-fw þ Dhly2-rev was used to amplify a cat cassette fromtemplate plasmid pKD3. The resulting PCR products wereintegrated into the desired chromosomal sites by the l Redrecombinase encoded on a curable helper plasmid. In orderto create a double hly mutant, the cat cassette was excised[12] from its original insertional site (hlyI ) in order to allowselection for chloramphenicol, following integration into thesecond hly allele. In order to allow a high level of expressionof a-haemolysin, bacteria were cultured in 2xYT medium untillate-log phase of growth. Alternatively, for in vivo studies,overnight LB cultures were used. Media were supplementedwith ampicillin (100 mg/ml) or chloramphenicol (25 mg/ml)where appropriate. Cell lines were obtained from the stockof the Institut fur Molekulare Infektionsbiologie, Wurzburg,and were cultured following prescriptions by ATCC. Dendriticcells were generated from bone marrow-derived progenitorsobtained from BALB/c mice as described by Lutz et al. [13].
2.2. Cloning of hly determinants
Haemolysin determinants were amplified by long distancePCR (KlenTaq LA Polymerase, Sigma) using primer hlyrevtogether with either hly1ups or hly2ups containing the recog-nition sites for restriction endonucleases KpnI or XbaI (seeTable 1). After digestion and purification, the amplified prod-ucts were ligated into the corresponding sites of the cloningvector pUC18, giving rise to pGNH11 and pGNH22,
respectively. pGNH12 and pGNH21 were constructed frompGNH11 and pGNH22 by ligation of the 50 ends from onehaemolysin determinant with the 30 ends of the other corre-sponding operon. Fusion was achieved at the single BamHIsite within hlyC. Clones were verified by restriction mappingof the isolated plasmids.
For sequencing of the upstream region of hlyIJ96, PCRproducts using primer pairs HU1-fw þ HU1-rev or HU2-fw þ HU2-rev were amplified. The obtained overlapping frag-ments were cloned into pGEM T-Easy, giving rise to pGNHU1and pGNHU2. Sequencing of the inserts was performed onboth strands. The obtained sequence is available under acces-sion number: DQ397536.
2.3. RNA isolation and Northern blots
Late-log phase cultures (grown in 2xYT medium) of 536-21 (non-haemolytic derivative of wild-type strain 536 lackingboth PAIs encoding a-haemolysin) carrying either pGNH11(hlyI536 in pUC18) or pGNH22 (hlyII536 in pUC18) wereused. Total RNA was isolated using the RNeasy Mini Kit (Qia-gen, Germany). RNasin Ribonuclease Inhibitor (Promega,Germany) was applied from the first step of the isolation pro-cedure. Isolated RNA (10 mg per lane) was separated ona 1.2% agarose-formaldehyde gel and was transferred to a Bio-dyne B Transfer Membrane (Pall Ltd, UK) overnight by cap-illary blotting. A 600-bp-long DNA probe specific for hlyAgenerated by PCR was labelled with the ECL� direct label-ling system (Amersham-Pharmacia, Germany). Hybridisationwas carried out overnight at 42 �C as described by the manu-facturer. Before luminography, the filter was washed twice for15 min in 0.5� SSC, 0.4% SDS (50 �C), and then twice for5 min in 2� SSC (20 �C).
2.4. Western blots
Whole-cell extracts were obtained from late-log phase2xYT cultures, or alternatively, secreted a-haemolysin wasprecipitated from culture supernatants by the addition of10% trichloroacetic acid (TCA). Protein samples were sepa-rated on 10% SDSePAGE gels and were blotted onto nitrocel-lulose membranes. The blocked membranes were treated withspecific HlyA antisera (kindly provided by Ivaylo Gentschev)and secondary antibody (anti-rabbit IgGehorseradish peroxi-dase conjugate; Dako A/S, Denmark) diluted suitably inTBST containing 2% skimmed milk. The blot was developedusing ECL detection reagents (NEN Life Science, Boston,MA, USA) followed by luminography.
2.5. Animal experiments
Animal experiments were conducted according to the prin-ciples set forth in the Guide for the Care and Use of Labora-tory Animals in a Laboratory authorised by the Hungarianrules (Decree No. XXVII, 1998) and by subsequent regula-tions (Gov. Order No. 243/1998).

2008 G. Nagy et al. / Microbes and Infection 8 (2006) 2006e2012
Table 1
Bacterial strain, plasmids and primers used in this study
Bacterial strain, plasmid or primer Relevant characteristics Reference
Strains
536 Wild-type UPEC strain (O6:K15:H31) [37]
536-21 Non-haemolytic spontaneous deletion mutant lacking both PAI I536 and PAI II536 [11]
536DH1 536DhlyI::cat This study
536DH2 536DhlyII::cat This study
536-HDM 536DhlyI, DhlyII::cat This study
J96 Wild-type UPEC strain (O4:K5) [38]
Plasmids
pKD3 Template plasmid for the amplification of a cat cassette, CmR, ApR [12]
pKD46 Curable helper plasmid encoding l Red recombinase, ApR [12]
pCP20 Curable helper plasmid encoding FLP required for excision of FRT-flanked resistance
cassettes, CmR, ApR[12]
pUC18 Cloning vector, ApR, oriColE1, lacZ [39]
pGEM-T Easy Cloning vector, ApR, orif1, lacZ Promega
pGNH11 hlyCABD of hlyI536 together with its 1500 bp of upstream region cloned into pUC18 This study
pGNH22 hlyCABD of hlyII536 together with its 1500 bp of upstream region cloned into pUC18 This study
pGNH12 50 region of hlyI536 fused to hlyII536 This study
pGNH21 50 region of hlyII536 fused to hlyI536 This study
pGNHU1 50 region of hlyIJ96 cloned into pGEM-T Easy This study
pGNHU2 50 region of hlyIJ96 cloned into pGEM-T Easy This study
Primers
Dhly1-fw GCAAGGTGCAGGAAATAAAAATTATCATTTCTTTATATGGTGTAGGCTGGAGCTGCTT
Dhly1-rev GAAGATATGAGCCGGATACCGCGGCTCTGAGACTTAACGCATATGAATATCCTCCTTAGTTCCTA
Dhly2-fw GAAGATATGAGCAGGATACCGCGGCTCTGAGACTTAACGGTGTAGGCTGGAGCTGCTT
Dhly2-rev ATTCAAGAATATAATTTAATTTATAGTGAGCTTATTATGCATATGAATATCCTCCTTAGTTCCTA
hly1ups-XbaI TTATCTAGAGGTGGTGATCCACATGAATTTGA
hly2ups-XbaI TTATCTAGAGGGTACTGGGAAGACCAGGGTTA
hlyrev-KpnI ATAGGTACCTTAACGCTCATGTAAACTTTCTGTT
HU1-fw GGGTACTGGGAAGACCAGGGTTA
HU1-rev AAAAGAAGGCAACTCTATTGGCGGG
HU2-fw CGGCTGACTGGTTAACTGGTC
HU2-rev GGGTTAATAAAGCATATTGGT
2.5.1. Ascending urinary tract infectionIntravesical infections of 3e4-day-old suckling NMRI
mice (Charles River, Hungary) were performed as describedby Allison et al. [14]. Briefly, bacteria were grown overnightat 37 �C in LB, harvested by centrifugation, washed once,and normalised to the required inoculum density (109 CFU/ml) in saline by adjusting the suspension to the appropriateOD600 value, which had been verified by viable counts. Groupsof 7e15 pups were infected by log dilutions of washed bacte-ria as described elsewhere in detail [15,16].
2.5.2. Intravenous infectionGroups of ten (5 and 5 each in two independent experi-
ments) 8-week-old female NMRI mice were infected intrave-nously. Threefold dilutions of washed bacterial suspensions(5 � 108e6.17 � 106 CFU) in 0.3 ml of saline were injectedthrough the tail veins. Mice were observed for 14 days; never-theless, deaths occurred within the first 72 h post-infection.
2.5.3. Lung toxicity assayThe lung toxicity assay was performed as described by
Hacker et al. [17]. Briefly, 4e5-week-old NMRI mice weigh-ing 10e12 g were instilled intranasally under superficial etheranaesthesia with 50 ml (w3 � 109 CFU/ml) of bacteria grown
overnight in LB medium. Twelve mice were subjected to eachstrain in two independent experiments. The animals were ob-served for 12 h and mortality was recorded hourly.
2.6. In vitro cytotoxicity assay
Toxicity in different cultured cell lines of strain 536 and itsmutants lacking individual or both hly determinants was mea-sured using a commercially available kit (Cytotox 96, Prom-ega). Monolayers of various cultured cell lines were infectedwith strain 536 or its derivatives. Bacteria grown until late-log phase were washed and diluted in the appropriate cell cul-ture media supplemented with 10% heat-inactivated serum.Cells (105) on 96-well tissue culture plates were infected byserial dilutions of bacteria (103e107) resulting in an initialMOI of 0.01e100. Lactate dehydrogenase (LDH) releaseupon cell death was determined from the supernatants follow-ing 3 h incubation at 37 �C in a CO2 thermostat.
2.7. Pore formation studies
Culture supernatants of hly mutants grown till late-logphase in 2xYT media were used for all measurements. Poreformation was tested on synthetic asolectin (a crude lipid

2009G. Nagy et al. / Microbes and Infection 8 (2006) 2006e2012
mixture from soybean) bilayer membranes using methods de-scribed elsewhere in detail [18,19]. Briefly, planar membraneswere formed across a 0.5-mm2 hole surrounded by aqueoussalt solution. The membrane current was measured witha pair of silver/silver chloride electrodes. In the single-channelrecordings, the electrometer was replaced by a current ampli-fier. The amplified signal was monitored with a storage oscil-loscope and recorded with a tape or a strip chart recorder. Theion selectivity of the channels formed was determined by zero-current membrane potential measurements in the presence ofa KCl gradient as described earlier [19].
2.8. Statistical analyses and software
LD50 values were calculated by using the method of Reedand Muench [20]. Log rank test (ManteleHaenszel test) wasused for statistical analysis of survival curves obtained fromthe animal experiments. KaplaneMeier survival curves werecreated and analysed using Prism 4.0 (GraphPad) software.Quantification of Western blots was performed with Bio-1Dþþ V.97 software.
3. Results
3.1. Sequence analysis of hly determinants
Both hly determinants carried by strain 536 as well as thehly operon carried on PAI II of strain J96 (hlyIIJ96) havebeen previously sequenced [3,10] and are available under ac-cession numbers AJ488511, AJ494981, and M10133. StrainJ96-M1, which has spontaneously lost PAI II, is still haemo-lytic [9], indicating the existence of a second functional hly pa-ralogue in strain J96 (hlyIJ96). Amplification of the 50 region ofthis hly allele was performed using primers deriving from thecorresponding region of hlyII536. Sequencing of the resultingPCR products revealed high homology (>98%) of this regionto that of hlyII536 (this sequence has been submitted to Gen-Bank and assigned accession number DQ397536). On theother hand, the upstream region of hlyI536 is homologous tothat of hlyIIJ96. Altogether, these data provide evidence thatboth strains have two hly paralogues, which differ in their 50
flanking regions including promoter sites and potential cis-acting regulatory sites. The different upstream regions are,however, conserved in the two strains, suggesting a potentialadvantage upon possession of both alleles.
The only similarity found between the two variants ofleader sequences was the 8-bp-long ops element. This cis-act-ing motif is essential for RfaH-dependent transcriptional anti-termination of hlyCABD [21e23]. As in hlyIIJ96, the ops ele-ment is located 334 bases upstream of the hlyC start codonin hlyI. Within the 50 region of hlyII, however, ops is situatedmore upstream (647 bases) of the hlyC start codon.
3.2. Expression of the two hly paralogues of strain 536
In order to investigate the expression of the two hlyparalogues originating from strain 536, both hlyCABD
determinants together with 1.5 kbp of their 50 flanking regionswere cloned into a conventional high copy number plasmidand were transformed into a non-haemolytic variant of thewild-type strain 536. Northern blots using a 600-bp probe spe-cific to hlyA revealed that expression of hlyI was considerablyhigher than that of hlyII (Fig. 1A). Western blotting with anHlyA-antiserum of total cellular proteins as well as TCA-pre-cipitated supernatants of late-log-phase cultures confirmed thisobservation at the protein level (Fig. 1B and C). Exchange ofupstream regions (up to the conserved BamHI site locatedwithin hlyC ) reversed expression rates (Fig. 1, lanes 3 and4) indicating that the difference observed in the amount of spe-cific mRNAs as well as encoded proteins is a consequence ofthe dissimilar 50 flanking regions. Quantification of Westernblots revealed that the hlyI-specific 50 region directed a 3.4-to 33-fold higher expression of HlyA regardless of the originof the coding genes.
Expression of the chromosomal hly determinants was alsoinvestigated in the wild-type background. Western blots wereperformed with precipitated supernatants obtained from iso-genic hly deletion mutants (Fig. 1D). The amount of secretedHlyA was 16% and 78% in 536DH1 (lacking hlyI ), and536DH2 (lacking hlyII ), respectively, relative to the wild-type strain. The observed 5-fold stronger expression of hlyIin comparison to hlyII confirmed our data with the recombi-nant hly determinants.
3.3. Contribution of individual hly alleles to virulence
To be able to assess the contribution of the two hly alleles tothe virulence of wild-type strain 536, isogenic mutants lackingindividual or both hly determinants were constructed. Viru-lence of the individual mutants as well as that of the doublehly mutant was compared to that of the parental wild-typestrain in the infant mouse model of urinary tract infection.
A
B
C
1
hly transcript
HlyA
HlyA
HlyAD
2 3 4
Fig. 1. Expression of individual hly determinants originating from E. coli strain
536. Both hlyI and hlyII together with 1.5 kbp of their 50 flanking regions were
cloned into vector pUC18 resulting in pGNH11 (lane 1) and pGNH22 (lane 2),
respectively. Exchange of the upstream regions gave rise to pGNH12 (up-
stream region of hlyI fused to hlyII; lane 3) and pGNH21 (upstream region
of hlyII fused to hlyI; lane 4). Expression was determined by Northern blotting
(A) of total RNA obtained from non-haemolytic strain 536-21 carrying the re-
combinant plasmids. HlyA protein amounts were detected by Western blots
from cell lysates (B) or TCA-precipitated culture supernatants (C). Alterna-
tively (D), the amount of HlyA was determined in culture supernatants of
wild-type strain 536 (lane 1) and its isogenic hly mutants 536DH1 (lane 2),
536DH2 (lane 3), and 536-HDM (lane 4).
2010 G. Nagy et al. / Microbes and Infection 8 (2006) 2006e2012
We determined a 67-fold and a 54-fold increase in LD50 of536DH1 (8.2 � 103 CFU) and 536DH2 (6.6 � 103 CFU), re-spectively, in comparison to the parental wild-type strain(1.22 � 102 CFU). The double hly mutant (536-HDM) ex-hibited a 3053-fold increase in LD50 value (3.7 � 105 CFU)in the same model.
Intravenous virulence of strain 536 as well as its isogenichly mutants was assessed in groups of ten 8-week-old NMRImice. Unlike the LD50 value of the double hly mutant(>5 � 108 CFU), that of neither 536DH1 (1.35 � 108 CFU)nor 536DH2 (9.98 � 107 CFU) differed significantly fromthe LD50 of the parental wild-type strain (9.33 � 107 CFU).However, statistical analysis by the log-rank test revealedthat the KaplaneMeier survival curves were significantly dif-ferent in case of both 536DH1 (P ¼ 0.0154) and 536DH2(P ¼ 0.0265) compared to that of the wild-type strain. The sur-vival curves of the two individual mutants, however, did notsignificantly differ (P ¼ 0.2025). This finding suggests thatthrough carrying both hly determinants, the wild type strainelicits a more expeditious in vivo toxicity (see below).
3.4. In vitro and in vivo toxicity encoded on theindividual hly alleles
In vitro cytotoxicity of hly mutants was tested and com-pared to that of the wild-type strain using different cell cul-tures as target cells. Table 2 shows that both mutants lackingindividual hly determinants were reduced in their toxicity to-wards all cell lines tested in comparison to the parentalwild-type strain. Nevertheless, no significant difference couldbe determined between 536DH1 and 536DH2 in cytotoxicityto any cell lines, indicating that no target specificity was en-coded in the individual hly determinants.
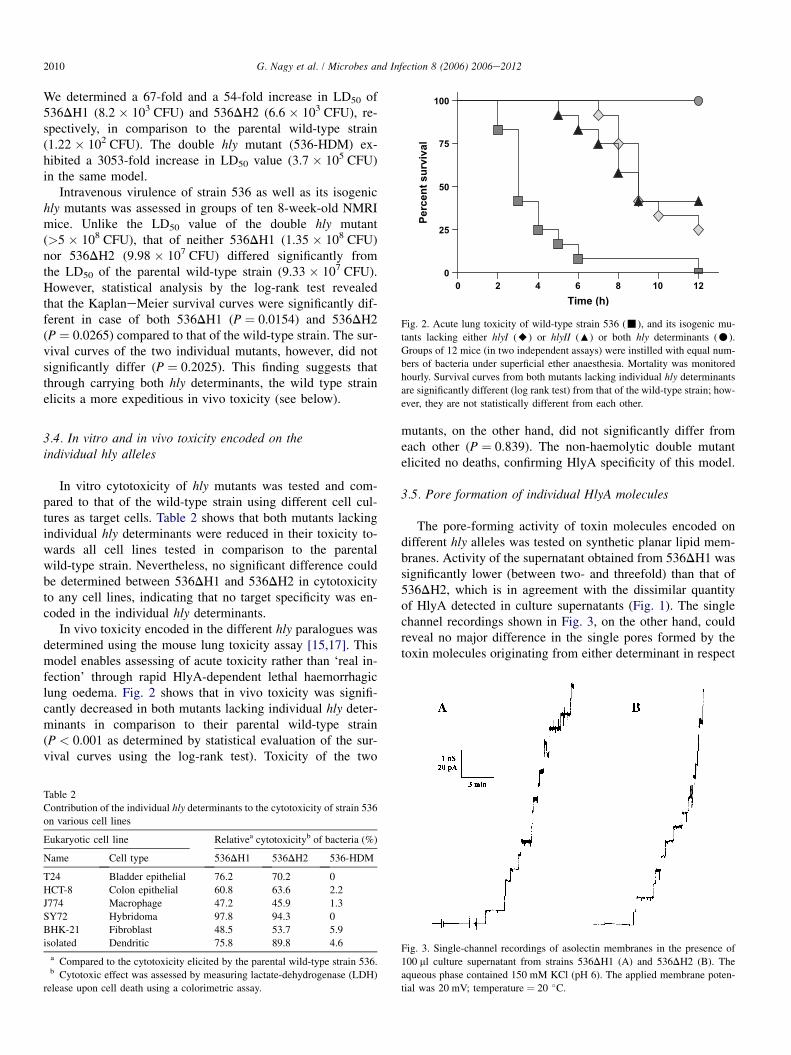
In vivo toxicity encoded in the different hly paralogues wasdetermined using the mouse lung toxicity assay [15,17]. Thismodel enables assessing of acute toxicity rather than ‘real in-fection’ through rapid HlyA-dependent lethal haemorrhagiclung oedema. Fig. 2 shows that in vivo toxicity was signifi-cantly decreased in both mutants lacking individual hly deter-minants in comparison to their parental wild-type strain(P < 0.001 as determined by statistical evaluation of the sur-vival curves using the log-rank test). Toxicity of the two
Table 2
Contribution of the individual hly determinants to the cytotoxicity of strain 536
on various cell lines
Eukaryotic cell line Relativea cytotoxicityb of bacteria (%)
Name Cell type 536DH1 536DH2 536-HDM
T24 Bladder epithelial 76.2 70.2 0
HCT-8 Colon epithelial 60.8 63.6 2.2
J774 Macrophage 47.2 45.9 1.3
SY72 Hybridoma 97.8 94.3 0
BHK-21 Fibroblast 48.5 53.7 5.9
isolated Dendritic 75.8 89.8 4.6
a Compared to the cytotoxicity elicited by the parental wild-type strain 536.b Cytotoxic effect was assessed by measuring lactate-dehydrogenase (LDH)
release upon cell death using a colorimetric assay.
mutants, on the other hand, did not significantly differ fromeach other (P ¼ 0.839). The non-haemolytic double mutantelicited no deaths, confirming HlyA specificity of this model.
3.5. Pore formation of individual HlyA molecules
The pore-forming activity of toxin molecules encoded ondifferent hly alleles was tested on synthetic planar lipid mem-branes. Activity of the supernatant obtained from 536DH1 wassignificantly lower (between two- and threefold) than that of536DH2, which is in agreement with the dissimilar quantityof HlyA detected in culture supernatants (Fig. 1). The singlechannel recordings shown in Fig. 3, on the other hand, couldreveal no major difference in the single pores formed by thetoxin molecules originating from either determinant in respect
Time (h)
0 2 4 6 8 10 12
0
25
50
75
100
Percen
t su
rvival
Fig. 2. Acute lung toxicity of wild-type strain 536 (-), and its isogenic mu-
tants lacking either hlyI (A) or hlyII (:) or both hly determinants (C).
Groups of 12 mice (in two independent assays) were instilled with equal num-
bers of bacteria under superficial ether anaesthesia. Mortality was monitored
hourly. Survival curves from both mutants lacking individual hly determinants
are significantly different (log rank test) from that of the wild-type strain; how-
ever, they are not statistically different from each other.
Fig. 3. Single-channel recordings of asolectin membranes in the presence of
100 ml culture supernatant from strains 536DH1 (A) and 536DH2 (B). The
aqueous phase contained 150 mM KCl (pH 6). The applied membrane poten-
tial was 20 mV; temperature ¼ 20 �C.

2011G. Nagy et al. / Microbes and Infection 8 (2006) 2006e2012
to pore stability and single channel conductance. The lifetimeof the pores (about 60 s) in both cases, nevertheless, appearedto be considerably longer than those reported in the plasmidencoded HlyA [24] and was very similar to that of anotherchromosome-encoded HlyA [25].
The channels formed by both mutants were cation selectivein a similar measure; the permeability ratio PK/PCl was 10.0and 8.5 for 536DH1 and 536DH2, respectively, indicating nosignificant difference.
4. Discussion
Gene duplication is the major source of genetic novelty anda driving force of evolution, since one of the duplicates mayfreely attain novel roles while the remaining copy still main-tains the original function [26]. At least 50% of prokaryoticgenes and even more eukaryotic genes are thought to be prod-ucts of gene duplication [27]. Duplication was recently shownto be more common among genes obtained by lateral genetransfer; nevertheless, few duplications survive subsequent de-letions [28]. The vast majority of gene duplicates are silencedunless one of the alleles acquires a novel beneficial functionthat might be preserved by natural selection [29]. Another pos-sible fate of gene duplicates is that they evolve expressionaldivergence. This mechanism has been shown to be a relativelycommon mechanism in the evolution of yeast genomes. In ad-dition to alterations in the upstream cis-acting regulatory mo-tifs [30], multiple trans-acting factors may influence theexpressional pattern of the newly evolved allele [31]. Further-more, evidence has been shown that expression of duplicategenes tends to evolve asymmetrically: namely, one alleleevolves rapidly, while the other copy maintains the ancestralexpression profile [32]. Gene duplication in regulatory net-work evolution has been shown to be an important mechanismnot only in yeast but in E. coli as well [27].
Pathogenic bacteria often carry multiple copies of toxinmolecules. Vibrio cholerae isolates with enhanced virulencewere selected in animal models in which tandem repeats ofcholera toxin genes were detected [33]. A clinical isolate ofE. coli was shown to harbour two identical copies of thegene encoding heat-labile enterotoxin on one single plasmid[34]. Duplication of the bifunctional (adenylate cyclase-hae-molysin) RTX toxin of Bordetella pertussis, which is closelyrelated to a-haemolysin, was observed in isolates with unusu-ally high haemolytic activity [35]. Gene duplication in thisstrain resulted in two identical copies including promoter re-gions and potential cis-acting regulatory sites. Increased in vi-tro haemolytic activity is paralleled neither with increased invivo virulence nor altered virulence properties associatedwith this toxin, suggesting that expression of one of the iden-tical copies is sufficient for the given virulence.
In contrast, we have shown that both hlyCABD determi-nants are required for full virulence of wild-type UPEC strain536. This appears to be not an additive effect since expression(at least in vitro) of one determinant (hlyII ) is infinitesimal incomparison to the other one (hlyI ) (Fig.1). In spite of theirhighly different expression, however, the two haemolysin
determinants seem to contribute equally to the virulence ofthe wild type strain. To be able to resolve this paradox, we pro-posed that the two toxin molecules participate at differentstages of the infectious process. This may materialise eitherby a variation in their toxic effect on target cells by HlyA mol-ecules encoded by the individual determinants, or their expres-sion at different periods during the course of infection. To testthe first possibility mutants lacking individual hly determi-nants were tested for in vitro cytotoxicity on different cell linesas well as for pore formation characteristics on artificial lipidmembranes. Our results showed that neither of the two a-hae-molysin variants have evolved a tropism to any cell types;moreover, they form identical pores on target membranes.These data are supported by the fact that the toxin moleculesoriginating from the individual hly alleles differ in as few as 12residues (out of 1024 amino acids). No relevant characteristicof the HlyA molecule encoded on hlyII536 was altered uponexchange of any of the differing residues (Nagy G., unpub-lished results).
Nevertheless, the (almost) identical coding regions of hlydeterminants are flanked by entirely dissimilar 50 regionswith presumably different expressional regulation. This sug-gests that the same cytotoxic effect expressed in diverse envi-ronments and/or stages of the infectious process may increasevirulence of the given pathogen. A similar explanation wasprovided in another study describing the prominent virulenceof a Vibrio harveyi isolate carrying two copies of haemolysingenes [36].
Interestingly, the only similarity found between the differ-ent upstream regions of hlyCABD operons is the 8-bp-longops element, required for the regulatory effect elicited by thetranscriptional anti-terminator RfaH [21e23]. Conservationof this cis-acting regulatory motif within the otherwise dissim-ilar regions suggests selective pressure for retaining RfaH-mediated transcriptional regulation. Indeed, a previous studyshowed that an isogenic rfaH mutant of strain 536 is non-haemolytic and severely attenuated [15].
It seems probable that duplication of the a-haemolysindeterminant did not occur in the recent strain, but rather, thetwo paralogues evolved in an ancestor strain and were inte-grated into different PAIs. The presence of the same two al-leles on distinct pathogenicity islands in the unrelated UPECstrain J96, nevertheless, suggests that parallel occurrence ofboth determinants may confer an evolutionary advantage onUPEC strains. The exact in vivo expressional regulation ofboth determinants, however, still needs to be elicited in orderto determine their exact contribution to urovirulence.
Acknowledgements
We thank I. Gentschev for the HlyA-antiserum, and C.Apetrei for isolated dendritic cells. This work was supportedby grants OTKA F04526 (to G.N.), T037833 (to L.E.), SFB479 (to J.H. and U.D.), SFB 487, project A5 (to R.B.), andthe Fonds der Chemischen Industrie. G.N. was supported byHumboldt and Bolyai fellowships.

2012 G. Nagy et al. / Microbes and Infection 8 (2006) 2006e2012
References
[1] S.J. Elliott, S. Srinivas, M.J. Albert, K. Alam, R.M. Robins-Browne,
S.T. Gunzburg, B.J. Mee, B.J. Chang, Characterization of the roles of
hemolysin and other toxins in enteropathy caused by alpha-hemolytic
Escherichia coli linked to human diarrhea, Infect. Immun. 66 (1998)
2040e2051.
[2] J.R. Johnson, Virulence factors in Escherichia coli urinary tract infection,
Clin. Microbiol. Rev. 4 (1991) 80e128.
[3] T. Felmlee, S. Pellett, R.A. Welch, Nucleotide sequence of an Escheri-
chia coli chromosomal hemolysin, J. Bacteriol. 163 (1985) 94e105.
[4] U. Dobrindt, B. Janke, K. Piechaczek, G. Nagy, W. Ziebuhr, G. Fischer,
A. Schierhorn, M. Hecker, G. Blum-Oehler, J. Hacker, Toxin genes
on pathogenicity islands: impact for microbial evolution, Int. J. Med.
Microbiol. 290 (2000) 307e311.
[5] S. Knapp, I. Then, W. Wels, G. Michel, H. Tschape, J. Hacker,
W. Goebel, Analysis of the flanking regions from different haemolysin
determinants of Escherichia coli, Mol. Gen. Genet. 200 (1985) 385e392.
[6] J. Hacker, C. Hughes, H. Hof, W. Goebel, Cloned hemolysin genes from
Escherichia coli that cause urinary tract infection determine different
levels of toxicity in mice, Infect. Immun. 42 (1983) 57e63.
[7] R.A. Welch, S. Falkow, Characterization of Escherichia coli hemolysins
conferring quantitative differences in virulence, Infect. Immun. 43 (1984)
156e160.
[8] D.L. Swenson, N.O. Bukanov, D.E. Berg, R.A. Welch, Two pathogenic-
ity islands in uropathogenic Escherichia coli J96: cosmid cloning and
sample sequencing, Infect. Immun. 64 (1996) 3736e3743.
[9] G. Blum, V. Falbo, A. Caprioli, J. Hacker, Gene clusters encoding the
cytotoxic necrotizing factor type 1, Prs- fimbriae and alpha-hemolysin
form the pathogenicity island II of the uropathogenic Escherichia coli
strain J96, FEMS Microbiol. Lett. 126 (1995) 189e195.
[10] U. Dobrindt, G. Blum-Oehler, G. Nagy, G. Schneider, A. Johann,
G. Gottschalk, J. Hacker, Genetic structure and distribution of four
pathogenicity islands (PAI I(536) to PAI IV(536)) of uropathogenic
Escherichia coli strain 536, Infect. Immun. 70 (2002) 6365e6372.
[11] G. Blum, M. Ott, A. Lischewski, A. Ritter, H. Imrich, H. Tschape,
J. Hacker, Excision of large DNA regions termed pathogenicity islands
from tRNA- specific loci in the chromosome of an Escherichia coliwild-type pathogen, Infect. Immun. 62 (1994) 606e614.
[12] K.A. Datsenko, B.L. Wanner, One-step inactivation of chromosomal
genes in Escherichia coli K-12 using PCR products, Proc. Natl. Acad.
Sci. U.S.A. 97 (2000) 6640e6645.
[13] M.B. Lutz, N. Kukutsch, A.L. Ogilvie, S. Rossner, F. Koch, N. Romani,
G. Schuler, An advanced culture method for generating large quantities
of highly pure dendritic cells from mouse bone marrow, J. Immunol.
Methods. 223 (1999) 77e92.
[14] C. Allison, L. Emody, N. Coleman, C. Hughes, The role of swarm cell
differentiation and multicellular migration in the uropathogenicity of
Proteus mirabilis, J. Infect. Dis. 169 (1994) 1155e1158.
[15] G. Nagy, U. Dobrindt, G. Schneider, A.S. Khan, J. Hacker, L. Emody,
Loss of regulatory protein RfaH attenuates virulence of uropathogenic
Escherichia coli, Infect. Immun. 70 (2002) 4406e4413.
[16] G. Schneider, U. Dobrindt, H. Bruggemann, G. Nagy, B. Janke, G. Blum-
Oehler, C. Buchrieser, G. Gottschalk, L. Emody, J. Hacker, The Pathoge-
nicity Island-Associated K15 Capsule Determinant Exhibits a Novel
Genetic Structure and Correlates with Virulence in Uropathogenic
Escherichia coli Strain 536, Infect. Immun. 72 (2004) 5993e6001.
[17] J. Hacker, H. Hof, L. Emody, W. Goebel, Influence of cloned Escherichia
coli hemolysin genes, S-fimbriae and serum resistance on pathogenicity
in different animal models, Microb. Pathog. 1 (1986) 533e547.
[18] R. Benz, A. Schmid, W. Wagner, W. Goebel, Pore formation by the Es-
cherichia coli hemolysin: evidence for an association-dissociation equilib-
rium of the pore-forming aggregates, Infect. Immun. 57 (1989) 887e895.
[19] A. Osickova, R. Osicka, E. Maier, R. Benz, P. Sebo, An amphipathic
alpha-helix including glutamates 509 and 516 is crucial for membrane
translocation of adenylate cyclase toxin and modulates formation and
cation selectivity of its membrane channels, J. Biol. Chem. 274 (1999)
37644e37650.
[20] L.J. Reed, H. Muench, A simple method of estimating fifty percent end
points, Am. J. Hyg. 27 (1938) 493e497.
[21] J.A. Leeds, R.A. Welch, RfaH enhances elongation of Escherichia colihlyCABD mRNA, J. Bacteriol. 178 (1996) 1850e1857.
[22] J.A. Leeds, R.A. Welch, Enhancing transcription through the Escherichia
coli hemolysin operon, hlyCABD: RfaH and upstream JUMPStart DNA
sequences function together via a postinitiation mechanism, J. Bacteriol.
179 (1997) 3519e3527.
[23] J.M. Nieto, M.J. Bailey, C. Hughes, V. Koronakis, Suppression of
transcription polarity in the Escherichia coli haemolysin operon by a short
upstream element shared by polysaccharide and DNA transfer determi-
nants, Mol. Microbiol. 19 (1996) 705e713.
[24] A. Ludwig, A. Schmid, R. Benz, W. Goebel, Mutations affecting pore
formation by haemolysin from Escherichia coli, Mol. Gen. Genet. 226
(1991) 198e208.
[25] R. Benz, A. Dobereiner, A. Ludwig, W. Goebel, Haemolysin of Escher-
ichia coli: comparison of pore-forming properties between chromosome
and plasmid-encoded haemolysins, FEMS Microbiol. Immunol. 5 (1992)
55e62.
[26] S.D. Hooper, O.G. Berg, On the nature of gene innovation: duplication
patterns in microbial genomes, Mol. Biol. Evol. 20 (2003) 945e954.
[27] S.A. Teichmann, M.M. Babu, Gene regulatory network growth by
duplication, Nat. Genet. 36 (2004) 492e496.
[28] S.D. Hooper, O.G. Berg, Duplication is more common among laterally
transferred genes than among indigenous genes, Genome Biol. 4
(2003) R48.
[29] M. Lynch, J.S. Conery, The evolutionary fate and consequences of dupli-
cate genes, Science 290 (2000) 1151e1155.
[30] B. Papp, C. Pal, L.D. Hurst, Evolution of cis-regulatory elements in
duplicated genes of yeast, Trends Genet. 19 (2003) 417e422.
[31] Z. Zhang, J. Gu, X. Gu, How much expression divergence after yeast
gene duplication could be explained by regulatory motif evolution?
Trends Genet. 20 (2004) 403e407.
[32] X. Gu, Z. Zhang, W. Huang, Rapid evolution of expression and
regulatory divergences after yeast gene duplication, Proc. Natl. Acad.
Sci. U.S.A. 102 (2005) 707e712.
[33] J.J. Mekalanos, Duplication and amplification of toxin genes in Vibrio
cholerae, Cell. 35 (1983) 253e263.
[34] G.L. Murphy, W.S. Dallas, Analysis of two genes encoding heat-labile
toxins and located on a single Ent plasmid from Escherichia coli, Gene
103 (1991) 37e43.
[35] K. Dalet, C. Weber, L. Guillemot, E. Njamkepo, N. Guiso, Characteriza-
tion of adenylate cyclase-hemolysin gene duplication in a Bordetella
pertussis isolate, Infect. Immun. 72 (2004) 4874e4877.
[36] X.H. Zhang, P.G. Meaden, B. Austin, Duplication of hemolysin genes in
a virulent isolate of Vibrio harveyi, Appl. Environ. Microbiol. 67 (2001)
3161e3167.
[37] H. Berger, J. Hacker, A. Juarez, C. Hughes, W. Goebel, Cloning of
the chromosomal determinants encoding hemolysin production and
mannose-resistant hemagglutination in Escherichia coli, J. Bacteriol.
152 (1982) 1241e1247.
[38] R.A. Hull, R.E. Gill, P. Hsu, B.H. Minshew, S. Falkow, Construction and
expression of recombinant plasmids encoding type 1 or D-mannose-
resistant pili from a urinary tract infection Escherichia coli isolate, Infect.
Immun. 33 (1981) 933e938.
[39] C. Yanisch-Perron, J. Vieira, J. Messing, Improved M13 phage cloning
vectors and host strains: nucleotide sequences of the M13mp18 and
pUC19 vectors, Gene 33 (1985) 103e119.