canadian journal of botany - natural resources...
TRANSCRIPT

Canadian Journal of Botany Published by THE NATIONAL RESEARCH COUNCIL OF CANADA
VOLUME 46 MARCH 1968 NUMBER 3
Kellermania and. its generic segregates
B. C. SUTTON Department of Forestry and Rural Development of Canada, Forest Research Laboratory, Winnipeg, Manitoba
Received September 11, 1967
The history of the generic names Kellermania Ell. & Ev., Clzaetoconis Clements, Scolecosporiella Petrak non Hahn., Amphorula Grove and Brencklea Petrak is reviewed. Two species are included in Kellermania, K. anomala (Cooke) Hahn., and K. major Dearn. & Barth., and one in Clzaetoconis, C. polygoni (Ell. & Ev.) Clements. Brencklea is reduced to synonymy with Scolecosporiella, and this genus enlarged to include three species, S. typhae (Oud.) Petrak, S. sisyrincllii (Ell. & Ev.) n. comb. (- Kellermania sisyrinchii Ell. & Ev.), and S. kranzii n. sp. Obstipipilus n. gen., type species O. malabaricus (T.S. & K. Ramakrishnan) n. comb. (= Kellermania malabarica T. S. & K. Ramakrishnan), is described. Six species described in or transferred to Kellermania are excluded from the genus.
Canadian Journal of Botany, 46, 181 (1968)
Introduction The generic name Kellermania, first published
by Ellis and Everhart (1885), was coined in honor of Dr. W. A. Kellerman. The following short generic description was given: "Perithecia immersed, membranaceous, ostiolate; stylospores cylindrical, large, septate, stipitate". The single species, K. yuccaegena Ell. & Ev., was described from leaves of Yucca angust!folia Pursh with the following diagnosis: "Perithecia membranaceous, about .2 mm in diameter, globose, buried in the substance of the leaf and only visible outwardly as small, dusky circles with a black spot in the centre, caused by the minute papilliform ostiolum barely visible through the slightly ruptured epidermis. Spores cylindrical, granular, 45-50 X
10-12 fJ., abruptly contracted below into a slender, stipe-like base, 18-25 fJ. long. The granular contents are divided by a septum across the middle with indications of becoming faintly multi septate" .
Ellis and Everhart (1886) added two new species, K. polygoni Ell. & Ev. and K. sisyrinchii Ell. & Ev. , and altered the spelling of the generic name to Kellermannia without explanation for the change. The later spelling has been adopted by some authors (Hohne! 1915; Clements and Shear 1931; Ainsworth and
Bisby 1961) but Cash (1953) listed the five species eventually named by Ellis and Everhart under the name Kellermania. The original spelling was derived correctly from its source (Lanjouw 1961, Art. 73, Recomm. 73B) and must be accepted as correct. The nature of the attenuated structure on the conidium was misinterpreted by Ellis and Everhart (1885) as being a basal stipe, and in 1886 they modified the generic characters to include conidia with an apical "awn-like appendage." The spores were described as sessile and "borne on slightly elongated cells of the proligerous layer." The type species, K. yuccaegena, was suggested as being identical with Discella anomala Cooke (1878) but the latter epithet was rejected as not applicable to this fungus although reasons for this decision were not given.
A fourth name K. mutica, was added by Ellis and Everhart in Langlois (1887) but this was published without an accompanying description and is considered a nomen nudum by Cash (1953). Ellis and Everhart (1900) considered 'K. mutica' to be identical with Septoria megaspermaSpeg.
Three additional species, K. alpina Ell. & Ev. , K. pruni McAlp. , and K. rumicis Fautr. & Lamb., were placed in Kellermania before Clements (1909) proposed the first revision of the

182 CANADIAN JOURNAL OF BOTANY. VOL. 46. 1968
genus. K. polygoni was made the type species of a new mono typic genus, Chaetoconis Clements. This generic name appeared first on page 125 but with no formal description apart from the positive and negative characters used to key out the genus from related ones. Despite this, the criteria distinguishing this genus from others were clearly enumerated in the key and the generic name is considered to have been validly published here. The type, and only species, was clearly indicated by reference to Saccardo (1892) and the new combination "Ch. polygoni (E. & Ev. )" was made on page 176 (Clements 1909) with a back reference to page 125; the identity of the basionym is considered unambiguous. Chaetoconis was characterized by separate, unbeaked, smoothwalled, erumpent pycnidia and dark, ovoid to oblong, I-septate conidia each with a cilium at the apex. It was distinguished from Kellermania, which was typified by globose pycnidia and hyaline, 2- to several-septate, oblong to fusoid conidia each with a seta at the apex. Later, Clements and Shear (1931) placed Chaetoconis, together with Amphorula Grove and Brencklea Petrak, in synonymy with Kellermania.
Hahnel (1915) produced the first detailed study of several Kellermania species, including K. polygoni, K. sisyrinchii, K. rumicis, K. alpina, K. pruni, and K. yuccaegena. The earlier epithet provided by Discella anomala for the type species was adopted and the new combination K. anomala (Cooke) HOlm. was formally proposed. Hahnel examined Ellis and Everhart's material and enlarged the description of the type species, including an account of pycnidium structure, which originally was not described in detail by Ellis and Everhart. He also criticized the erection of Chaefoconis Clements (1909), which he considered premature, and questioned the reduction of K. rumicis to synonymy with Heteropatella cercosperma (Rostr.) Lind by Lind (1913). Hahnel pointed out that K. sisyrinchii and K. pruni did not belong in Kellermania but offered no suggestion about where they should be placed. His proposal that K. alpina would be more satisfactorily accommodated in Heteropatella resulted in the formal combination eventually being made by W. B. Cooke apud Sprague and W. B. Cooke (1939).
Petrak (1923) made K. sisyrinchii the type
species of the monotypic genus Brencklea with a generic diagnosis as follows: "Ohne Stroma. Pykniden typisch, subepidermal eingewachsen, zerstreut, mit undeutlichem, durchbohrtem Ostiolum oder einfachem Porus. Membran weichhautig, faserig, undeutlich kleinzellig, nur urn den dunkelgefarbten Porus deutlich parenchymatisch. Konidien schmal keulig oder fast spindelig, hellfarbig, mehrzellig, an der Spitze sehr allmahlich in eine lange, hyaline, oft gekriimmte, diinne Zilie iibergehend. Konidientrager sehr kurz und undeutlich." Petrak gave little information about how Brencklea differed from related genera and merely cited Hohnel's initial observation that K. sisyrinchii was incorrectly placed in Kellermania. He stated that it "differed from K. anomala which was characterized by the structure of the pycnidial wall and the constantly 2-celled hyaline conidia."* In the taxonomic treatment presented in this paper, Brencklea is reduced to synonymy with Scolecosporiella so an historical account of this genus is pertinent.
Pycnidia of Hendersonia typhae Oudemans (1873), described from leaves of Typha angustifolia L., were characterized as black with a central protruding ostiole, and with spores formed from the inner wall of the pycnidium. The spores were described as being 7-septate, brown, fusiform, with an obtuse apex and an acute base. Hahnel (1902), found no evidence of a pycnidial wall and in a later paper (1909) transferred the name to Scolecosporium. Petrak (1921) compared S. typhae (Oud.) Hahnel with Scolecosporium Jagi Libert, the type species of the genus, and confirmed Hahnel's observation that there was no pycnidial wall in S. typhae, although he disagreed with its transfer to Scolecosporium which he considered to be a Hyphomycete genus. Petrak maintained that conidiophores were absent from the basal cushion of pseudoparenchyma in S. typhae. He suggested that this feature also separated the fungus from "H ender sonia auct.". Consequently Petrak proposed the monotypic genus, Scolecosporiella for H. typhae, with the following diagnosis: "Sporenlager subepidermal, scharf umgrenzt, niedergedriickt-kuglig, die Epidermis mit kriesrunder Offnung durchbohrend, mit dunner, fast hyaliner Basalschicht. Sporen schmal
*Translation from German.

SUTTON: KELLER MANIA 183
spindelformig, mit mehreren Querwanden, honiggelb bis braun. Sporentrager fehlen." Hohnel (1923) proposed the generic name Scolecosporiella, which was presumably based on a different fungus. This name is illegitimate, being a later homonym of Scolecosporiella Petrak (1921), and must therefore be rejected (Lanjouw 1961, Art. 64). Clements and Shear (1931) reduced Scolecosporiella Hohnel to synonymy with Scolecosporium Lib. and Scolecosporiella Petrak to synonymy with "Hendersonia Westd.". Ainsworth and Bisby (1961) considered Scolecosporiella Hohnel to be a nomen nudum. The generic name appeared in Hohnel's key but no species was mentioned or described. Webster (1955) showed Hendersonia typhae to be the conidial state of Leptosphaeria typharum (Desm.) Karst. and Webster and Hudson (1957) produced a similar, but not identical, conidial state from Ophiobolus herpotrichus (Fr.) Sacc., which they tentatively assigned to Urohendersoniella Petrak (Petrak 1955).
Grove (1922) proposed the new generic name, Amphorula, with a single species, A. sachalinensis Grove. The generic diagnosis was as follows: "Pycnidia immersa, solidiuscula, carbonacea. Sporulae ampulliformes, longirostratae, septatae, hyalinae. Genus Kellermaniae Ell. & Ev. affine, sed forma sporularum pycnidioque solidiore bene distinctum." Grove emphasized the resemblance between A. sachalinens is, K. polygoni, and K. rumicis in spore form, and reiterated this opinion in 1935. However, in the absence of any specimens or figures of these species, he came to no definite conclusions concerning their possible synonymy. A. sachalinensis was separated from K. yuccaegena by the nature of the conidial appendage and the fine structure of the pycnidia. Characters of the genus Amphorula are more readily deduced from the species diagnosis: "Pycnidiis sparsis, depresso-globosis vel placentiformibus, 300-750 j.1 diam. ; vix papillatis, immersis, epidermide tectis eique primo arcte adhaerantibus eandem postea poro orbiculari minutissimo deinde ampliore v. rimiformi penetrantibus, postremo epidermide emortua desiliente superficialibus aut subinde cum eadem dilapsis, diu astomis atris, peridio crasso opaco sed stratum versus proliferum pallidiore circummunitis. Sporulis elliptco-fusoideis, supierne in 10ngum
rostrum filiforme attentuatis, rostro incluso 40-60 j.1 longis, parte inferiore elliptica 15-22 X 2t-4 j.1, achrois, saepe guttulatis, dein tenuissime 1- septatis, sporophoris rectis brevibus suffultis."
Petrak (1959), in studies on A. sachalinensis and its correlated ascomycete state, Amphorulopsis polygonacearum Petrak, confirmed Clement's (1909) separation of K. polygoni from Kellermania. However, he regarded the generic name Chaetoconis as a "nomen subnudum", and therefore proposed the adoption of the generic name Amphorula for this fungus. Petrak formally emended Grove's generic description, the main points being that the fructifications were interpreted as stromata, sometimes imperfectly divided instead of simple carbonaceous pycnidia, and that the attenuated apices were interpreted as cilia instead of rostra. K. polygoni provided an earlier specific epithet for the type species, A. sachalinensis Grove, and accordingly the binomial A. polygoni (Ell. & Ev.) Petrak was published, with K. rumicis being reduced to synonymy.
Taxonomy
GENERIC KEY 1. Conidiophores annellate. Conidia pale brown,
I-septate with an apical eccentric setula. Fructifi-cations acervular ... .. . . . .. .. . . ... .. .. Obstipipilus
1. Conidiophores phialidic. Conidia hyaline, septate with apical cell drawn out into a filiform appendage. Fructifications pycnidial, scIerotioid divided ..................................... Chaetoconis
1. Conidiophores simple, blastic ... . .. . .... . . . .. ... 2. 2. Conidia septate, hyaline, with an extracellular
apical elongated setula. Fructifications scIerotioid, pycnidial .. .. ...... .... . ... . . .. . . . . Kellermania
2. Conidia transversely septate (sometimes longitudinally), pale brown, with apical cell drawn out into a filiform appendage. Fructifications thin-walled, pycnidiai. ..... .......... Scolecosporiella
CHAETOCONIS Clements, Genera of Fungi, p. 125, 1909.
=Amphorula Grove, J. Bot. Lond. 60, p. 82,1922.
Pycnidia immersed, solitary or confluent, glabrous, cavity unilocular or multilocular, opening by one or more distinct ostioles. Pycnidial wall several layers thick, composed of compact outer sclerotioid and inner pseudoparenchymatic tissue. Conidiophores lining the inner wall, hyaline, branched, septate, bearing single phialidic apertures at the apices of main
184 CANADIAN JOURNAL OF BOTANY. VOL. 46, 1968
20"
100!, ®
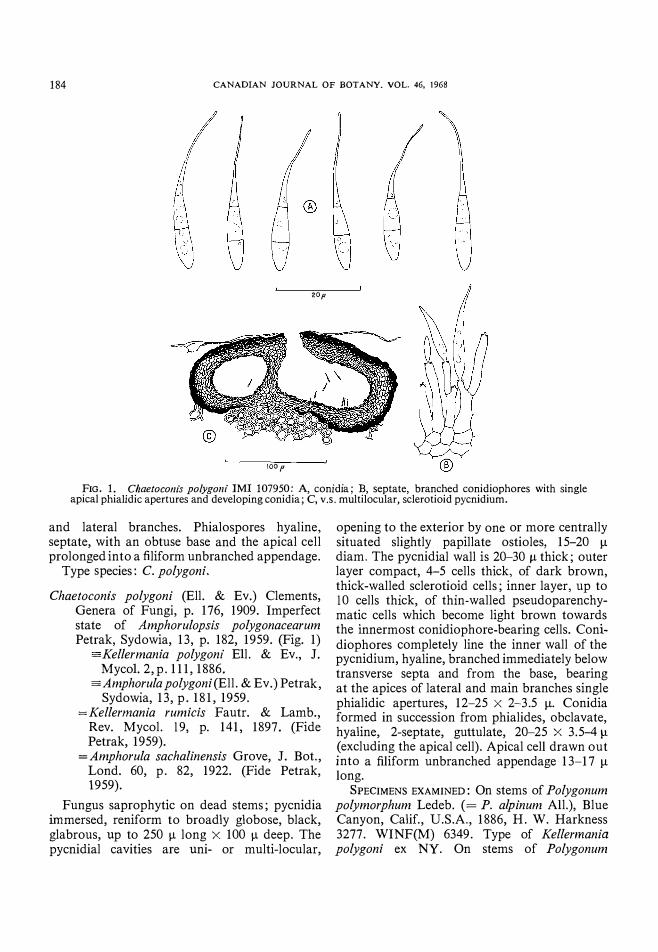
FIG. 1. Chaetocollis polygoni 1M1 107950: A, conidia; B, septate, branched conidiophores with single apical phialidic apertures and developing conidia; C, v.s. multilocular, sclerotioid pycnidium.
and lateral branches. Phialospores hyaline, septate, with an obtuse base and the apical cell prolonged into a filiform unbranched appendage.
Type species: C. polygoni.
Chaetoconis polygoni (Ell. & Ev.) Clements, Genera of Fungi, p. 176, 1909. Imperfect state of Arnphorulopsis polygonacearurn Petrak, Sydowia, 13, p. 182, 1959. (Fig. 1)
-=Kellerrnania polygoni Ell. & Ev., J. Mycol. 2, p. 111, 1886.
-=Arnphorula polygoni (Ell. & Ev.) Petrak, Sydowia, 13, p. 181, 1959.
=Kellerrnania rurnicis Fautr. & Lamb., Rev. Mycol. 19, p. 141, 1897. (Fide Petrak, 1959).
=Arnphorula sachalinensis Grove, J. Bot., Lond. 60, p. 82, 1922. (Fide Petrak, 1959).
Fungus saprophytic on dead stems; pycnidia immersed, reniform to broadly globose, black, glabrous, up to 250 11 long X 100 11 deep. The pycnidial cavities are uni- or multi-locular,
opening to the exterior by one or more centrally situated slightly papillate ostioles, 15-20 11 diam. The pycnidial wall is 20-30 11 thick; outer layer compact, 4-5 cells thick, of dark brown, thick-walled sclerotioid cells; inner layer, up to 10 cells thick, of thin-walled pseudoparenchymatic cells which become light brown towards the innermost conidiophore-bearing cells. Conidiophores completely line the inner wall of the pycnidium, hyaline, branched immediately below transverse septa and from the base, bearing at the apices of lateral and main branches single phialidic apertures, 12-25 X 2-3.5 11. Conidia formed in succession from phialides, obclavate, hyaline, 2-septate, guttulate, 20-25 X 3.5-4 11 (excluding the apical cell). Apical cell drawn out into a filiform unbranched appendage 13-17 11 long.
SPECIMENS EXAMINED: On stems of Polygonurn polyrnorphurn Ledeb. (= P. alp in urn AIl.), Blue Canyon, Calif., U.S.A., 1886, H. W. Harkness 3277. WINF(M) 6349. Type of Kellerrnania polygoni ex NY. On stems of Polygonurn

SUTTON: KELLERMANIA 185
100!,
FiG. 2. Kellermallia allomala IMI 97350; A, conidia; B, doliform simple conidiophores with developing conidia; C, v.s. unilocular, th ick-walled pycnidium.
alpinum All., Oregon Caves, Josephine Co., Calif., U.S.A., 7-10-1936, H. E. Parks 5631a. WINF(M) 6350 ex NY. On stems of Polygonum cuspidatum Sieb. & Zucc., Ilam Park, Dovedale, Derbyshire, England, 5-5-1963, G. MorganJones. IMI 107950. On stems of Rumex acetosa L. , Malon, Nr. Bastad. Sweden, 6-7-1927, A. G. Eliasson. IMI 16634.
KELLERMANIA Ellis & Everhart, J. Mycol. 1, p. 153, 1885.
Pycnidia immersed, solitary or aggregated, globose, glabrous, cavity unilocular, opening by a single distinct ostiole. Pycnidia1 wall several cells thick, loosely composed of outer sclerotioid and inner pseudoparenchymatic tissue. Conidiophores lining the inner wall, small, hyaline, rarely branched, continuous or I-septate. Blastospores hyaline, cylindrical, septate, with an apical setula, and the remains of the conidiophore apex persisting at the base.
Type species: K. anomala.
KEY TO SPECIES
Conidia I-septate, 34-40 X 5.5-7 11, setulae 18-23 I.l long ..... ... .......... . . . . .. . ........ K. allomala
Conidia 2-septate , 50-68 X 8-11 11, setulae 14-21 11 long .......................................... K. major
Kellermania anomala (Cooke) Hahnel, Sitzb. k. Acad. Wiss. Wien, 124, p. 84, 1915. (Fig. 2)
== Discella anomala Cooke, Grevillea, 7, p. 11, 1878.
== Discula anomala (Cooke) Sacc., Sylloge Fungorum, 3, p. 677, 1884.
=Kellermania yuccaegena Ell. & Ev., J. Mycol. 1, p. 154, 1885.
Pycnidia are formed in cream-brown, irregular lesions with a raised reddish-brown margin, immersed, black, solitary or aggregated, globose, glabrous, 250-300 11 diam X 150 11 deep; wall 7-1 1 cells thick, 25 11 wide, composed of loosely aggregated, thick-walled, dark brown, sclerotioid hyphae which become lighter in texture and more pseudoparenchymatic towards the hyaline conidiophore-bearing region. Wall tissue
186 CANADIAN JOURNAL OF BOTANY. VOL. 46, 1968
!\
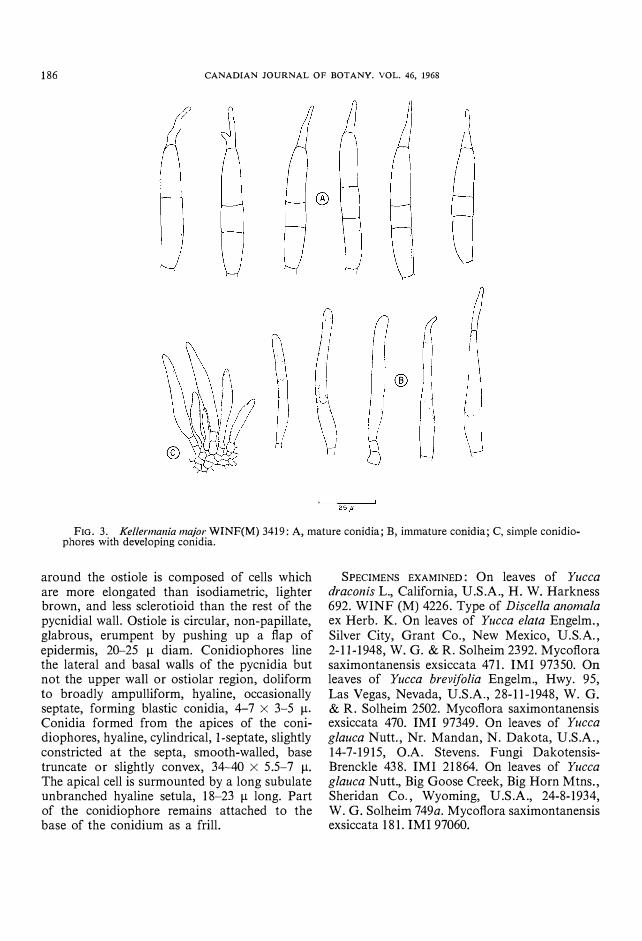
25 i'
FIG. 3. Kellermallia major WINF(M) 3419: A, mature conidia; B, immature conidia; C, simple conidiophores with developing conidia.
around the ostiole is composed of cells which are more elongated than isodiametric, lighter brown, and less sclerotioid than the rest of the pycnidial wall. Ostiole is circular, non-papillate, glabrous, erumpent by pushing up a flap of epidermis, 20-25 I-l diam. Conidiophores line the lateral and basal walls of the pycnidia but not the upper wall or ostiolar region, doliform to broadly ampulliform, hyaline, occasionally septate, forming blastic conidia, 4-7 X 3-5 I-l. Conidia formed from the apices of the conidiophores, hyaline, cylindrical, I-septate, slightly constricted at the septa, smooth-walled, base truncate or slightly convex, 34-40 X 5.5-7 I-l. The apical cell is surmounted by a long subulate unbranched hyaline setula, 18-23 I-l long. Part of the conidiophore remains attached to the base of the conidium as a frill.
SPECIMENS EXAMINED: On leaves of Yucca draconis L., California, U.S.A., H. W. Harkness 692. WINF (M) 4226. Type of Discella anomala ex Herb. K. On leaves of Yucca elata Engelm. , Silver City, Grant Co. , New Mexico, U.S.A. , 2-11-1948, W. G. & R. Solheim 2392. Mycoflora saximontanensis exsiccata 471. IMI 97350. On leaves of Yucca brevifolia Engelm., Hwy. 95, Las Vegas, Nevada, U.S.A., 28-11-1948, W. G. & R. Solheim 2502. Mycoflora saximontanensis exsiccata 470. IMI 97349. On leaves of Yucca glauca Nutt. , Nr. Mandan, N. Dakota, U.S.A. , 14-7-1915, O.A. Stevens. Fungi DakotensisBrenckle 438. IMI 21864. On leaves of Yucca glauca Nutt., Big Goose Creek, Big Horn Mtns. , Sheridan Co., Wyoming, U.S.A., 24-8-1934, W. G. Solheim 749a. Mycoflora saximontanensis exsiccata 181. IMI 97060.

SUTTON: KELLER MANIA 187
© 50p
FIG. 4. Obstipipilus malabariclls IMI 110502: A, conidia; B, annellate conidiophores with developing conidia; C, v.s. acervulus.
Kellermania major Dearn. & Barth., Mycologia, 16, p. 163, 1924. (Fig. 3)
Pycnidia of similar size, shape, and structure and associated with identical symptoms shown by K. anomala. Conidiophores line the lateral and basal walls of the pycnidia, hyaline, occasionally septate, doliform to ampulliform, forming blastic conidia, 3-10 X 3-3.5 ).1. Conidia formed from the apices of the conidiophores, hyaline, cylindrical, 2-septate, slightly constricted at the septa, median cell much smaller than the cells either side, smooth-walled, base truncate or slightly convex, 50.5-68 X 8-11 ).1. The apical cell is surmounted by a short hyaline setula which is occasionally branched, 14-21 ).1 long. Part of the conidiophore remains attached to the conidium base as a frill.
SPECIMEN EXAMINED: On leaves of Yucca whipplei Torr., San Bernardino Co., California, U.S.A. 23-4-1923, E. T. Bartholomew. WINF (M) 3419. Type of Kellermania major ex Herb. DAOM.
OBSTIPIPILUS n. gen. Etym. L. obstipus-inclined to one side:
pilus-hair. Fungi Imperfecti, Melanconiales. Mycelium immersum, ex hyphis pallide
brunneis vel hyalinis, septatis, parietibus levibus compositum. Acervuli immersi, pallide brunnei, subglobosi, raro stromatici. Conidiophora ex cellulis superioribus formata, obclavata, hyalina vel pallide brunnea, aseptata, non ramosa, cum apicis proliferationibus. Conidia singulariter in apice conidiophori et dein proliferationis cujusque successivae oriunda, pallide brunnea, obovata, levia, l-septata; cellula basalis minuscula et truncata cum parte conidiophori affixa; cellula apicalis majuscula cum setula non ramosa, hyalin a, obstipa.
Sp. typ. : O. malabaricus. Immersed mycelium composed of pale brown
or hyaline, septate, smooth-walled hyphae. Acervuli immersed, pale brown, subglobose, rarely stromatic. Conidiophores formed from
188 CANADIAN JOURNAL OF BOTANY. VOL. 46, 1968
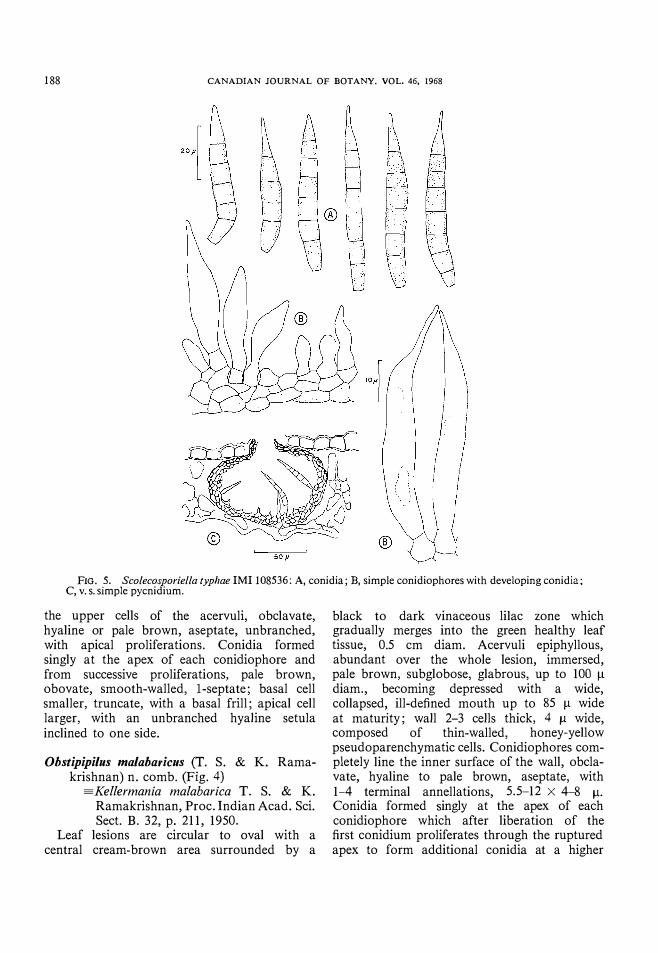
FIG. 5. Scolecosporiella typhae IMI 108536: A, conidia; B, simple conidiophores with developing conidia; C, v.s. simple pycnidium.
the upper cells of the acervuli, obclavate, hyaline or pale brown, aseptate, unbranched, with apical proliferations. Conidia formed singly at the apex of each conidiophore and from successive proliferations, pale brown, obovate, smooth-walled, I-septate; basal cell smaller, truncate, with a basal frill; apical cell larger, with an unbranched hyaline setula inclined to one side.
Ohstipipilus malaharicus (T. S. & K. Ramakrishnan) n. comb. (Fig. 4)
=.Kellermania malabarica T. S. & K. Ramakrishnan, Proc. Indian Acad. Sci. Sect. B. 32, p. 211, 1950.
Leaf lesions are circular to oval with a central cream-brown area surrounded by a
black to dark vinaceous lilac zone which gradually merges into the green healthy leaf tissue, 0.5 cm diam. Acervuli epiphyllous, abundant over the whole lesion, immersed, pale brown, subglobose, glabrous, up to 100 IJ. diam., becoming depressed with a wide, collapsed, ill-defined mouth up to 85 IJ. wide at maturity; wall 2-3 cells thick, 4 /1 wide, composed of thin-walled, honey-yellow pseudoparenchymatic cells. Conidiophores completely line the inner surface of the wall, obclavate, hyaline to pale brown, aseptate, with 1-4 terminal annellations, 5.5-12 X 4-8 /1. Conidia formed singly at the apex of each conidiophore which after liberation of the first conidium proliferates through the ruptured apex to form additional conidia at a higher

SUTTON: KELLERMANIA 189
FIG. 6. Scolecosporiella sisyrillchii IMI 21863: A, conidia; B, simple conidiophores with developing conidia; C, v.s. simple pycnidium.
level, honey-yellow, obovate, smooth-walled, I-septate with the lower cell smaller than the upper, 11.5-14 x 4-5.5 �; base truncate with a frill, apex furnished with a thin, unbranched, hair-like setula, eccentrically inserted, 4-5.5 � long.
SPECIMEN EXAMINED; On leaves of Anogeissus [alifolia Wall., Walayar, Kerala State, India, K. Ramakrishnan and G. S. Reddy 2638, 24 July 1949, IMI 110502. Type of Kellermania malabarica ex Herb. MH.
SCOLECOSPORIELLA Petrak, Ann. mycol., Berl. 19, p. 30, 1921. Non Scolecosporfella Hahne1, 1923.
=Brencklea Petrak, Ann. mycol., Berl. 21, p. 326, 1923.
Pycnidia immersed, solitary, glabrous, unilocular, opening by a single distinct ostiole;
wall up to 5 cells thick, composed of thin-walled, pseudoparenchymatic cells. Conidiophores lining the inner wall, hyaline, unicellular, ampulliform, bearing single conidia. Blastospores pale brown, smooth-walled, transversely and occasionally longitudinally septate, apical cell prolonged into a filiform appendage, base truncate with a frill.
Type species: S. typhae.
KEY TO SPECIES 1. Conidia often longitudinally septate, 48-70 X 12-14
II (excluding apical cell), appendage 15-20 Jl long . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S. kranzii
1. Conidia without longitudinal septa. . . . . . . . .. . .... 2 2. Conidia 4· 7-septate, 48-68 X 6.5-8.5 II (including
apical appendage), appendage up to 5 II long ... .... . . .. .. . ..... .. . . . . . ...... ... . S. typhae
2. Conidia 3-septate, 18-22 x 4-5 !l (excluding apical appendage), appendage 7-20 ilIon&; . .. ,' . ':
.
. . • . . . • . . . . . . . . . . . . . . . . . . . • . . . . .. S. SISYrlllChll

190 CANADIAN JOURNAL OF BOTANY. VOL. 46, 1968
FIG. 7. Scolecosporiella krallzii IMI 103919: A, conidia; B, simple conidioph ores with developing conidia; C, v.s. simple pycnidium.
Scolecosporiella typhae (Oud.) Petrak, Ann. mycoI., BerI. , 19, p. 30, 1921. Imperfect state of Leptosphaeria typharum (Desm.) Karst. , Bidr. FinI. Nat. Folk, p. 100, 1873. (Fig. 5)
=Hendersonia typhae Oud., Arch. need., Sci. , 8, p. 361, 1873.
=Scolecosporium typhae (Oud.) Hohnel, Sitzb. k. Akad. Wiss. Wien, 118, p. 405, 1909.
Pycnidia not produced in distinct lesions but formed on moribund leaves, interveinal, amphigenous, immersed, black, globose, glabrous, unilocular, 100-12011 diam; wall 2-3 cells thick, up to 12 11 wide, composed of thin-walled pseudoparenchymatic cells which are slightly darker on the outside and markedly so near
the ostiole; dehiscing through a slightly protruding, papillate, circular, ostiole which is up to 20 11 diam. Conidiophores undifferentiated from the inner cells of the pycnidial wall, simple, broadly ampulliform, hyaline, 0-1 septate, unbranched, producing blastic conidia, 4--7 X 3-5 11. Conidia almost navicular, very pale brown, smooth-walled, 4--7-septate, tapering to a truncate base with a frill, apical cell abruptly tapered or long acute, 48-68 X 6.5-8.5 11 (including the apical appendage).
SPECIMENS EXAMINED: On leaves of Typha angustifolia L. Type of Hendersonia typhae. IMI 108536 ex GRO. On leaves of Typha angustifolia L., Gros Skal, Germany, J. E. Kabat, 28 Sept. 1904. Fungi Bohemici (without number). IMI 108537 ex GRO. On leaves of Typha latifolia

SUTTON: KELLERMANIA 191
L., Studland Dunes, Studland, Dorset England, B. C. Sutton, 12 Aug. 1961. IMI 88870 b).
Scolecosporiella sisyrinchii (Ell. & Ev.) n. comb. (Fig. 6)
==.Kellermania sisyrinchii Ell. & Ev., J. Mycol. 2, p. 111, 1886.
==.Brencklea sisyrinchii (Ell. & Ev.) Petrak, Ann. mycol., Berl. , 21, p. 326, 1923.
Pycnidia formed on dead leaves and cu1ms, interveinal, amphigenous, immersed, black to dark brown, globose, glabrous, unilocular, 120-150 Il diam; wall 2-3 cells thick, up to 8 Il wide, composed of thin-walled very pale brown, pseudoparenchymatic cells which are darker and more sclerotioid around the ostiole and the sides of the pycnidium; dehiscing by an initially small but later a wide, slightly papillate ostiole up to 70 Il diam. Conidiophores undifferentiated from the inner cells of the pycnidia1 wall, absent from the ostiolar region, simple, aseptate, broadly ampulliform, hyaline, unbranched, forming blastic conidia, 2-5.5 X 2-4.5 Il. Conidia almost navicular, very pale brown, smooth-walled, 3-septate, lateral wall tapered slightly to a truncate base with a frill, 18-22 X 4-5 Il (without appendage). Apical cell attenuated into a long, filiform, unbranched appendage, 7-20 Il long.
SPECIMENS EXAMINED: On leaves and culms of Sisyrinchium bellum Wats., Berkeley, Calif., U.S.A., H. W. Harkness, 1886. Harkness No. 3017. Type of Kellermania sisyrinchii. IMI 108534 ex NY. On leaves of Sisyrinchium angustijolium Mill. , Near Kulm, N. Dakota, U.S.A., J. F. Brenckle, 10 June 1916. Fungi Dakotensis-Brenckle 383. IMI 21863.
SCOLECOSPORIELLA KRANZII sp. n. (Fig. 7) Pycnidia immersa in foliis mortuis, am
phigena, globosa, nigra, uniloculata, 95-110 Il diam. , cum ostiolo glabroso, medio-brunneo, 20-25 Il diam.; paries cum 1atitudine 2-4 cellularum pallide brunnearum pseudoparenchymaticarum et 12 Il crassus. Conidiophora hyalina, aseptata, ampulliformia, ex cellu1is parietis pycnidii interioribus nec distinctis oriunda, 2.5-5 X 2-4.5 Il. Conidia singula ex apice conidiophori oriunda, navicu1aria, pallide brunnea, parietibus levibus, 12- 15 septis transversis et 0- l3 septis 10ngitudina1ibus, basi truncata, cum parte conidiophori affixa, 48-70
X 12-14 Il. Cellula apicalis (appendix), filiformia, nonramosa, 15-20 Il 10nga.
Typus in fo1iis mortuis Andropogon tectorum Schum. & Thonn., IRF, Kindia, Guinea, J. Kranz 450, 5 Sept. 1963. IMI 103919.
Pycnidia formed in dead leaves and in lesions caused by other fungi, amphigenous, immersed, black, globose, glabrous, unilocular, 95-110 Il diam; wall 2-4 cells thick, up to 12 Il wide, composed of thin-walled, pale brown, pseudoparenchymatic cells which are slightly darker on the outside and near the ostiole; dehiscing by a minutely papillate, glabrous, circular ostiole, 20-25 Il diam. Conidiophores almost undifferentiated from the inner cells of the pycnidia1 wall, simple, aseptate, ampulliform, unbranched, forming blastic conidia, 2.5-5 X 2-4.5 Il. Conidia navicular, pale brown, smoothwalled, with 12- 15 transverse septa and a variable number of longitudinal septa (0- 13), tapered below to a truncate base with a frill, 48-70 X 12-14 Il (excluding apical cell). Apical cell attenuated into a filiform, pale brown, unbranched appendage, 18-20 Il long.
Excluded or Unexamined Kellermania Species
l . Kellermania alpina Ell. & Ev., Bull. Torrey bot. Cl. 27, p. 57, 1900.
Not examined. W. B. Cooke apud Sprague & W. B. Cooke
(1939) proposed the new combination Heteropatella alpina (Ell. & Ev.) W. B. Cooke but no mention was made that the type collection was examined. 2. K. caricis Golovin, Central Asian Univ.
Stud., n.s. , 14, 5, p. 37, 1950. Not examined. Vassilkov (1961), after an examination of the type, reduced this name to synonymy with Dilophospora alopecuri (Fr.) Fr.
3. K. cercosperma Rostrup, Micromycetes from N. E. Groenland, p. 157, 1910. Koebenhavn. Not examined. Lind (1913) made the combination into Heteropatella as H. cercosperma (Rostrup) Lind but later Lind (1926) reduced this binomial to synonymy with Heteropatella umbilicata (Fr.) Jaap (==. Peziza umbilicata Fr.). In the much-expanded synonymy, Kellermania rumicis Fautr. & Lamb. was reduced to synonymy, but this fungus has been shown to be identical with Chaetoconis

192 CANADIAN JOURNAL OF BOTANY. VOL. 46, 1968
>: �: ".' g.'.. . .. .. . . . ": . :i·:.:·:
FIG. 8. Pestalozzina thuemenii (= Kellermania pruni) IMI 112105: mature and immature conidia.
polygoni (Ell. & Ev.) Clements (as Amphorula polygoni (Ell. & Ev.) Petrak) by Petrak (1959).
4. K. gongrogena (Temme) Sacco & Trav., Sylloge Fungorum, 20, p. 1260, 1911.
== Pestalolia gongrogena Temme, Thiel's Landw. Jahrb. 16, p. 437, 1887.
Not examined. 5. K. Izordei Unamuno, Bol. Real Soc. Esp.
Hist. Nat. 30, p. 297, 1930. No trace of the fungus was found on the type collection borrowed from Herb. MA.
6. K. malabarica T. S. & K. Ramakrishnan, Proc. Indian Acad. Sci., Sect. B, 32, p. 211, 1950. Proposed as the type species of the new genus Obstipipilus Sutton (q.v.).
7. K. mutica Ell. & Ev. (nom. nud.), in Langlois, Cat. PI. Basse-La., p. 31, 1887. Not examined. The fungus was considered to be the same as Septoria megasperma Speg. by Ellis and Everhart (1900).
8. K. polygoni Ell. & Ev., J. Mycol. , 2, p. 111, 1886. See Chaetoconis polygoni (Ell. & Ev.) Petrak (q.v.).
9. K. pruni McAlpine, Fungus diseases of stone fruit trees, p. 104, 1902. Melbourne. (Fig. 8). McAlpine described the fungus as follows:
"Perithecia scattered, black, dark brown by transmitted light, depressed globose, membranaceous, with papillate mouth, 200-250 Il diam. Sporules chloro-hyaline, or yellowishhyaline, cylindrical or slightly fusoid, generally slightly curved, 3- to 4-septate, not constricted at septa, blunt at either end, apical segment perfectly hyaline, and bearing two to three diverging setae about 15-17 Il long, and borne by slender hyaline basidia, about 9 Il long, which are usually detached with the sporules, 28-34 X 4t-5 Il. Almond leaves kept moist. December, 1900. Stawell. Apricot twig living. July, 1900. Armadale. Plum leaves kept moist . March and April, 1901, Armadale. Peach and plum leaves. May, 1901. Armadale." In the discussion, McAlpine suggested that the genus was "practically a Pestalozzina, with a peri thecium." Two plates of illustrations accompany the description but there is no indication of which fungi were illustrated nor their host source. In Fig. 115 the conidia are 5-celled, the apical cell hyaline, the rest colored, with a single basal appendage and three apical appendages inserted at the tip of the terminal cell. The median cell was shown slightly smaller than the rest. In figure 196 the conidia are again 5-celled but none is colored. There is a single basal appendage and two or three apical appendages inserted laterally on the apical cell. The median cell is much smaller than the rest. Although the fungi depicted in the two figures do show some similarities, the insertion of the apical appendages indicates that two different species are illustrated.
Three collections borrowed from Herb. DAR were annotated as follows:
(1) Kellermannia pruni; plum leaf; Armadale, colI. D. McAlpine, 15.4.01. (IMI 112105).
(2) Kellermannia pruni?; plum and peach leaf; Armadale (Garden). 4.5.01. D. McAlpine. 256/01.
(3) Kellermannia pruni; peach leaf; Armadale, 4.5.01. D. McAlpine. No. (2090 + 1)
No fungus was found on collections (2) and (3), but a single fungus was present on (1). This corresponded with the fungus depicted in figure 196 except that the three median cells of the conidia are slightly colored and no basal appendage or conidiophore is attached to free conidia. The spores measure 23-34 X 4-4.5 Il with apical appendages up to l3 Il long. The

SUTTON: KELLERMANIA 193
important feature to note is that the appendages are inserted laterally on the apical cell. A strong argument can be made for amending the specific description of K. pruni, to limit its application to the fungus represented on IMI 112105. It is possible to select this element as a satisfactory lectotype; an alternative is to regard the name K. pruni McAlp. as a nomen confusum. McAlpine, in his original publication, and Saccardo (1906) suggest the fungus involved is a Pestalozzina. Observations on the specimen I chose as lectotype confirm this suspicion and the name is reduced to synonymy with Pestalozzina thuemenii (Speg.) Guba. 10. K. rumicis Fautrey & Lambotte, Rev. mycol.
19, p. 141, 1897. See Chaetoconis polygoni (Ell. & Ev.) Clements (q.v.).
II. K. sisyrinchii Ell. & Ev., J. Mycol. 2, p. 111, 1886. See Scolecosporiella sisyrinchii (Ell. & Ev.) Sutton (q.v.).
12. K. yuccaegena Ell. & Ev., J. Mycol. 1, p. 154, 1885. See K. anomala (Cooke) Hohn. (q.v.).
Discussion
There is an increasing tendency in current work on the Deuteromycotina (Fungi Imperfecti) to separate taxa by developmental characters of the conidiophores and conidia rather than exclusively morphological criteria (Ellis 1961; Hughes 1966; Kendrick 1961, 1962; Sutton 1964; Sutton and Sellar 1966 ; Tubaki 1958). This approach, originally suggested by Hughes (1953) for Hyphomycetes, is being adopted for other groups in the Fungi Imperfecti as more information about conidium formation becomes available. In the past, the dismemberment of Kellermania into a number of monotypic genera has been accomplished by emphasizing slight differences in spore morphology and their correlation with discernible variations in the type of fructification. Despite the fundamental similarity in spore morphology between the genera under consideration the use of developmental features of the sporogenous apparatus has made it possible to confirm the separation of Kellermania, Chaetoconis (=Amphorula), Scolecosporiella (=Brencklea), and Obstipipilus. Characters of the fructifications
have only been employed to emphasize the basic separation.
In previous work on Kellermania and related genera, little use, if any, has been made of the conidiophores in separating taxa at any level. The conidiophores of K. yuccaegena were not described by Ellis and Everhart (1885). In their later paper (Ellis and Everhart 1886), where the generic diagnosis was amended after an initial misinterpretation of conidium morphology, the conidia were referred to as sessile or at least borne on slightly elongated cells of the proligerous layer. Hohnel (1915) briefly dismissed the conidiophores as short. Oudemans (1873) made no mention of the conidiophores in Hendersonia typhae; however, his illustration shows conidia with obtuse apices, and acute tapered bases attached to the immersed pseudoparenchyma. Petrak (1921) referred to the conidiophores of Scolecosporiella as missing, whereas Webster (1955) described conidia arising by proliferation of the inner prismatic cells of the pycnidial wall. His illustrations indicate the mode of origin to be blastic, with the acute end of the conidium at the apex. This corrects the mistake concerning conidium orientation in Oudemans' work. For Brencklea, which is shown here to exhibit an identical mode of development, Petrak (1923) described the conidiophores as very short, indistinct, papillate, or rod-like. Information about the conidiophores of Amphorula is more complete than for other genera because the structures are larger and details have been more readily obtained. Petrak (1959) described them as formed from the whole inner surface of the loculus, very dense, simple, shortly filamentous or rod-like, and occasionally quite long. Unfortunately he made no comparisons with other genera. Clements (1909), Clements and Shear (1931), and Bender (1934) failed to take account of conidiophore morphology or development in their treatments of these genera.
Three different types of conidiophore have been recognized in the genera studied. In Chaetoconis, a plurality of conidia is formed in basipetal succession from single phialidic apertures at the apices of main and lateral branches without an increase in the length of the phialide itself. This corresponds with the type of development described by Hughes (1953) in his section IV. In Kellermania and Scolecosporiella solitary

194 CANADIAN JOURNAL OF BOTANY. VOL. 46, 1968
conidia are formed in a blastic manner from the innermost cells of the pycnidial wall terminating growth of the parent cell, whereas in Obstipipilus several blastic conidia develop in succession from a single conidiophore only by its proliferation through the conidial scar left by the previous conidium; a single conidium is produced at each level. Both types of development may be referred to Hughes' section III (1953). The truncate base of the conidium in Kellermania, Scolecosporiella, and Obstipipilus is encircled by a minute frill of variable length, whereas in Chaetoconis the conidium base is simply obtuse with no frill. The apical appendages in Chaetoconis, Kellermania, and Scolecosporiella have been the source of considerable variation in descriptive terminology. Grove (1922) maintained a difference in origin between the appendages of Amphorula (= Chaetoconis) and Kellermania in that the attenuated apex in Amphorula is merely a continuous prolongation of the spore which is not separable from it, whereas in Kellermania the subulate beak is a distinct and definite appendage to the spore similar to the appendages of Pestalotia deNot. Grove was correct in separating the two genera on this basis but the appendages of Amphorula (=Chaetoconis) and Scolecosporiella are closer in origin and development to those of Pestalotia deNot. The apex of the young conidium of Chaetoconis and Scolecosporiella becomes subulate and is eventually separated from the body of the conidium by a septum. The cellular appendages of Pestalotia and related genera are luminate but evidently devoid of stainable contents. It is conceivable in these genera that the cellular contents are retracted into the body of the conidium before or during transverse wall formation but this is purely conjecture. Amphorula and Scolecosporiella differ slightly from Pestalotia and related genera because stainable cellular contents are evident in immature and mature appendages. The apical setula of Kellermania is of quite a different origin and has little in common with the appendages of Pestalotia, Amphorula, or Scolecosporiella. Even in the most immature subulate conidia, the apical region is composed of unstainable material. As the conidium matures the apical region becomes increasingly elongated and a distinctly truncate wall separates the
unstainable material from the conidium protoplasm. Examination of mature conidia mounted in 10% KOH or 3 % erythrosin in 10% NH40H and viewed under phase contrast has shown three distinct retaining layers in the conidium. The protoplasm and its boundary membrane is enclosed by a thick cell wall which is not continuous with the apical setula. The latter is seen (at least at the tip of the apical cell) to be continuous with a very thin sheath completely enveloping the spore. The entire nature of the sheath is only clearly visible after immersion in the above reagents for 24 h or more. The sheath is similar to the mucilaginous cap described for conidia in Toxosporiopsis capitata Sutton and Sellar (1966), the origin and development of which was clearly described. The development of the eccentric setula in Obstipipilus is possibly similar. The mature setula shows affinities with those in the genera Dinemasporium Lev., Pseudolachnea Ranojevic, Stauronema Syd. & Butl., Chaetopatella Hino & Katumoto, and Menispora Pers. in that there is no definite separating septum and no direct continuity with the body of the conidium at maturity. In addition no setular lumen has been observed in Obstipipilus.
Apart from the appendages the other major means of separating these monotypic genera from Kellermania has been by the structure of the fructifications. Ellis and Everhart (1885) described K. yuccaegena with membranaceous globose pycnidia and later in 1886 differentiated between the thin-walled pycnidia of K. sisyrinchii ( == Scolecosporiella sisyrinchii) and the membranaceous coarsely cellular pycnidia of K. polygoni ( == Chaetoconis polygoni). No comparison was made with K. yuccaegena. Hahnel (1915) described the double-layered wall-structure, the complex arrangement of ostiolar tissue and the method of dehiscence more fully for K. yuccaegena, but failed to stress such features in the several other Kellermania species he studied. Petrak (1923) asserted a fundamental difference in pycnidial wall structure between Kellermania and Brencklea (=Scolecosporiella). For B. sisyrinchii he described the wall composed of simple, thin-walled pseudo parenchymatic cells, only darker round the ostiole. However, Scolecosporiella, which is shown to be identical with Brencklea in this study, was considered by Petrak (1921) to be

SUTTON: KELLERMANIA 195
like Hendersonia auct. but without any fruit body at all. The spore-bearing layer was described as subepidermal, depressed but globose, piercing the epidermis by a circular opening, with a flimsy basal layer from which spores are formed. Webster (1955), working with Scolecosporiella in culture, found the fructifications to be simple or compound pycnidia with a wall composed of up to five layers of olive-brown flattened cells lined by three to five layers of hyaline prismatic cells. Grove (1922, 1935) described the fructifications of Amphorula (= Chaetoconis) as immersed pycnidia with solid carbonaceous walls of hard close texture with dark brown pseudoparenchymatic cells on the outside, becoming paler towards the inside. Petrak (1959) preferred to interpret the fructifications as partially divided stromata but agreed with Grove in details of structure. Grove and Petrak separated Amphorula from Kellermania because Amphorula was typified by solid, thicker-walled fructifications.
In this study it has also been possible to confirm a separation between genera on the basis of fructification structure. The simplest pycnidia are found in Scolecosporiella (=Brencklea) where the wall is relatively thin, and thickened pseudoparenchymatic cells only occur in the simple ostiolar region. Pycnidial walls in Kellermania are several cells thick and composed of loosely interwoven outer sclerotioid and inner pseudoparenchymatic tissue. The wall tissue around the ostiole is less sc1erotioid and the cells distinctly elongated rather than isodiametric. In Chaetoconis (=Amphorula) the pycnidial walls also are very thick but composed of compact pseudoparenchyma which is distinctly sclerotioid on the outside, becoming thin-walled towards the conidiophore-bearing region. Frequently the pycnidial cavity becomes multiloculate and more than one circular ostiole is formed. The fructifications of Obstipipilus are not considered as pycnidia because no definitie ostiole is formed. The acervuli are composed of an immersed, depressed layer of pale brown to hyaline smooth-walled hyphae and show no definite wall structure.
Acknowledgments
I am indebted to the following herbarium curators for freely lending collections in their
keeping: Dr. J. A. Parmelee (DAOM), Mr. J. Walker (DAR), Dr. R. Van der Wijk (GRO), Dr. M. B. Ellis (1M I), Dr. G. Milne-Redhead (K), Dr. E. Paunero (MA), Dr. H. Santapau, S.J. (MH), and Dr. C. T. Rogerson (NY). Thanks are also due to Mr. F. C. Deighton for his welcome advice on some nomenclatural points, to Dr. J. Reid for some helpful discussion, to Dr. J. Kranz for permission to publish the name Scolecosporiella kranzii, and to Mr. J. Grant for correcting the Latin diagnoses.
AINSWORTH, G. C. and BISBY, G. R. 1961. Dictionary of the fungi. 5th ed. Commonwealth Agricultural Bureaux, England.
BENDER, H. B. 1934. The fungi. imperfecti: order Sphaeropsidales. Tuttle, Morehouse and Taylor Co., Conn.
CASH, E. K. 1953. A record of the fungi named by J. B. Ellis, II, Hadl'otrichulIl-Pyrenophora. pp. 167-345.
CLEMENTS, F. E. 1909. Genera of fungi. H. W. Wilson Co., Minneapolis.
CLEMENTS, F. E. and SHEAR, C. L. 1931. The genera of fungi. Hafner Publishing Co., New York.
COOKE, M. C. 1878. Californian fungi. Grevillea, 7: 11-13.
ELLIS, J. B. and EVERHART, B. M. 1885. New fungi. J. Mycol. 1: 148-154.
-- 1886. Kellermannia, Ell. & Evrht. J. Mycol. 2: 111. -- 1900. New species of fungi from various localities.
Bull. Torrey Botan. Club, 27: 571-578. ELLIS, M. B. 1961. Dematiaceous hyphomycetes. III.
Mycol. Papers, 82: 1-55. GROVE, W. B. 1922. New or noteworthy fungi. VIII.
J. Botan., London, 60: 81-86. -- 1935. British stem- and leaf-fungi. Vol. 1. Cam
bridge University Press, England. HOHNEL, F. VON. 1902. Fragmente zur Mykologie, 1.
Mitteilung. Sitzb. k. Akad. Wiss. Wien, 111: 987-1056.
-- 1909. Fragmente zur Mykologie, VI. Mitteilung. Sitzb. k. Akad. Wiss. Wien, 118: 275-452.
--- 1915. Fragmente zur Mykologie. 900. Dber die Gattung Kellermannia Ellis et Everhart. Sitzb. k. Akad. Wiss. Wien, 124: 82-84.
-- 1923. System der fungi imperfecti Fuckel. Mykol. Untersuch. Ber. von R. Falck, 3: 301-369.
HUGHES, S. J. 1953. Conidiophores, conidia, and classification. Can. J. Botany, 31: 577-659.
-- 1966. New Zealand fungi 7. Capnocybe and Capnophialophora, new form genera of sooty moulds. N.Z. J. Botany, 4: 333-353.
KENDRICK, W. B. 1961. The Leptogl'aphium complex. Phialocephala gen. nov. Can. J. Botany, 39: 1079-1085.
--- 1962. The Leptographium complex. Verticicladiella Hughes. Can. J. Botany, 40: 771-797.
LANGLOIS, A. B. 1887. Catalogue Provisoir des plantes phanerogames et cryptogames de la Basse-Louisiane (Etats-Unis d'Amerique). Saint-Etienne.
LANJOUW, J. 1961. International code of botanical nomenclature. Utrecht, Netherlands.
LIND, J. 1913. Rostrup's Danish fungi. Copenhagen. -- 1926. Micromycetes from Northwestern Green
land found on plants collected during the Jubilee

196 CANADIAN JOURNAL OF BOTANY. VOL. 46, 1968
Expedition 1920-23. Medd. Gr0nland, 71: 161-179. OUDEMANS, C. A. J. A. 1873. Materiaux pour la flore
mycologique de la Neerlande. II. Arch. Neerl' Sci. 8: 343-416.
PETRAK, F. 1921. Mykologische Notizen II. Ann. Mycol., Berl. 19: 17-128.
--- 1923. Mykologische Notizen. Ann. Mycol., Berl. 21: 182-335.
--- 1955. Urohelldersolliella n. gen., eine neue Gattung der phaeophragmosporen Sphaeropsideen. Sydowia, 9: 571-573.
--- 1959. Uber die Gattung Amplzorll/a Grove und die zu ihr gehOrige Schlauchform. Sydowia, 13: 178-182.
SACCARDO, P. A. 1892. Sylloge Fungorum, Vol. 10, Padua.
-- 1906. SyUoge Fungorum. Vol. 18. Padua. SPRAGUE, R. and COOKE, W. B. 1939. Some fungi
imperfecti from the pacific northwest. Mycologia, 31: 43-52.
SUTION, B. C. 1964. Plzol11a and related genera. Trans. Brit. Mycol. Soc. 47: 497-509.
SUTION, B. C. and SELLAR, P. W. 1966. Toxosporiopsis n. gen., an unusual member of the Melanconiales. Can. J. Botany, 44: 1505-1513.
TUBAKI, K. 1958. Studies on the Japanese hyphomycetes. V. Leaf and stem group with a discussion of the classification of hyphomycetes and their perfect stages. J. Hattori Bot. Lab. 20: 142-244.
VASSILKOV, B. P. 1961. How not to describe new fungi species. Bull. Soc. Nat. Moscow, ser. bioI. 66(3): 93-100.
WEBSTER, J. 1955. Helldersollia typhae the conidial state of Leptosplz aeria typlzarllm. Trans. Brit. Mycol. Soc. 38: 405-408.
WEBSTER, J. and HUDSON, H. J. 1957. Graminicolous pyrenomycetes. VI. Conidia of Ophiobo/us herpotrichus, Leptosplzaeria /uctl/osa, L. lucke/ii, L. pollti/ormis and L. eustomoides. Trans. Brit. Mycol. Soc. 40: 509-522.