carlo trompetto dipartimento di neuroscienze, oftalmologia … trompetto... · carlo trompetto...
TRANSCRIPT

Voluntary movement
Carlo Trompetto Dipartimento di Neuroscienze, Oftalmologia e Genetica
Università di Genova

Voluntary movement
• Organized around the performance of a purposeful task
• Effectiveness improves with experience and learning
• Can be generated internally

Cortical organization of voluntary movement
Electrical stimulation of the frontal lobe (1870)

Primary motor cortex
•The contralateral precentral gyrus (area 4) is the area in which the lowest-intensity stimulation elicited movements.
•Simple movements of single joints (twitch)
•Activation of neurons near the electrode

Premotor cortical areas•Also the direct electrical stimulation of the premotor areas (area 6) is able to evoke movements (although the intensity of stimulation must be higher).
•Area 6 lies anterior to the precentral gyrus, on the lateral (lateral premotor area) and medial (supplementary motor area) surfaces of the cortex.
•More complex movements, involving multiple joints and resembling natural coordinated hand shaping or reaching movements
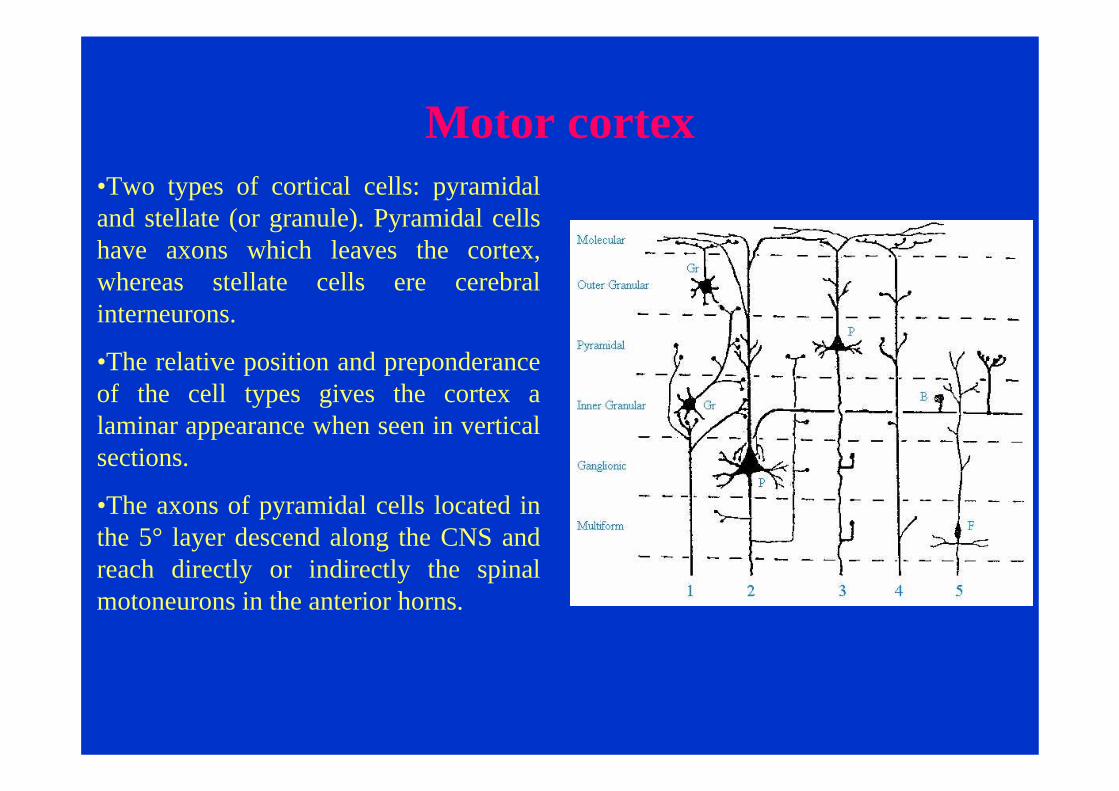
Motor cortex•Two types of cortical cells: pyramidal and stellate (or granule). Pyramidal cells have axons which leaves the cortex, whereas stellate cells ere cerebral interneurons.
•The relative position and preponderance of the cell types gives the cortex a laminar appearance when seen in vertical sections.
•The axons of pyramidal cells located in the 5° layer descend along the CNS and reach directly or indirectly the spinal motoneurons in the anterior horns.

Motor cortex
Two types of cortical cells: pyramidal and stellate (or granule). Pyramidal cells (3° and 5° layers) have axons which leaves the cortex, whereas stellate cells ere cerebral interneurons.
The pyramidal neurons in 5°layer represent the cortical moneurons and are the cells activated by direct electrical stimulation

Motor cortex: corticospinal pathway
•The primary motor cortex and the premotorareas project directly to the spinal cord through the corticospinal tract: all pyramidal tract fibers have cell bodies in the 5° layer.
•This descending fibers pass through the pyramids of the bulb; since the name of pyramidal tract.
•About one million fibers are found in the pyramidal tract. This is more than in any other animals.

Motor cortex: corticospinal pathwayOn leaving the cerebral cortex, the pyramidal tract axons descend in a small region of the posterior limb of the internal capsule. They then pass through the middle portion of the cerebral peduncle, and into pons. The majority of the pyramidal tract fibers decussate in the brain stem and reach the dorsolateral colums of the spinal cord (the lateral corticospinal tract). A small number of uncrossed fibers descend within the ventromedial part of the spinal cord (the ventral corticospinal tract)

Motor cortex: corticospinal pathwayIn the spinal cord, the motoneurons are arranged somatotopically. Those innervating the axial muscles lie most medially and their cell bodies form a ventromedial column in the grey matter. Motoneuronswhich innervate the extremities are situated in the lateral part of the ventral horn. They receive the fibersof the lateral corticospinaltract. The uncrossed fibers of the ventral corticospinal tract terminate in the ventromedialcolumn.
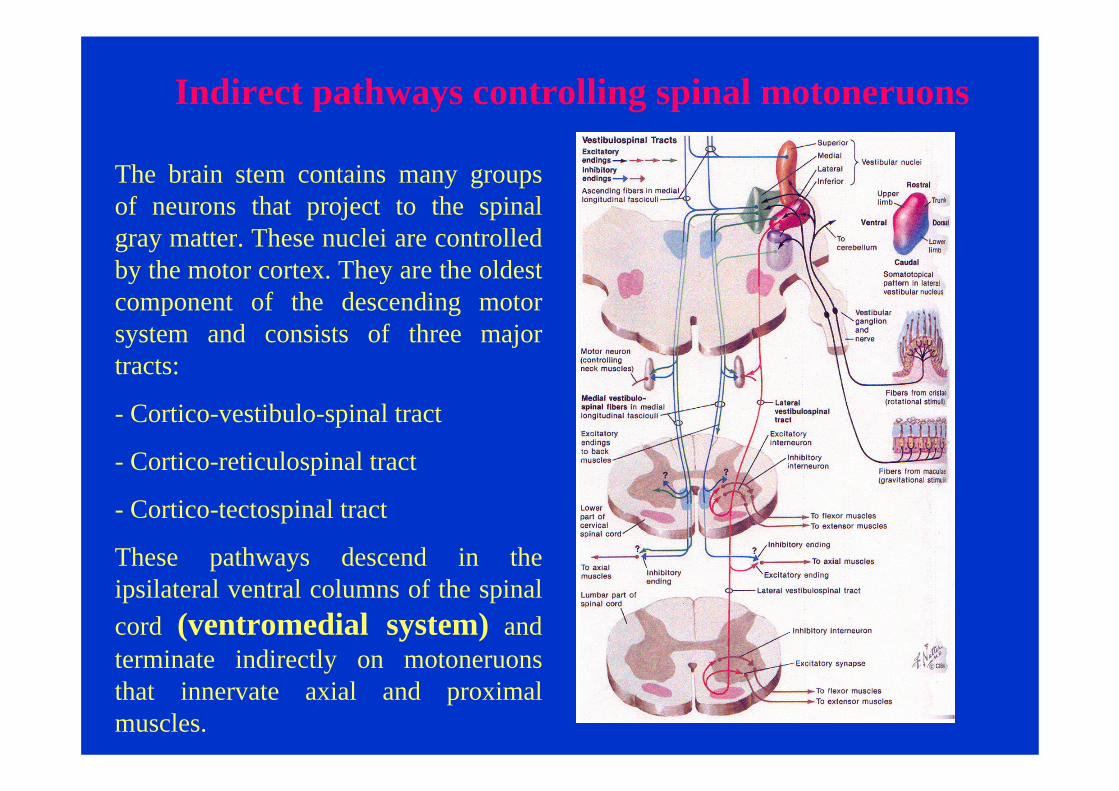
Indirect pathways controlling spinal motoneruons
The brain stem contains many groups of neurons that project to the spinal gray matter. These nuclei are controlled by the motor cortex. They are the oldest component of the descending motor system and consists of three major tracts:
- Cortico-vestibulo-spinal tract
- Cortico-reticulospinal tract
- Cortico-tectospinal tract
These pathways descend in the ipsilateral ventral columns of the spinal cord (ventromedial system) and terminate indirectly on motoneruonsthat innervate axial and proximal muscles.

Indirect pathways controlling spinal motoneruons
These indirect pathways provide the basic postural control system upon which the cortical motor areas can organize more highly differentiated movement.
These indirect pathways are bilaterally distributed.

Indirect pathways: latero-vestibulo-spinal tract
Lateral Vestibulospinal Tract. The lateral vestibulo spinal tract (LVST) arises primarily from neurons in the lateral and inferior vestibular nuclei and projects to all levels of the ipsilateral spinal cord.
Fibers of the LVST course through the anterior funiculus of the cord. Axons of many LVST neurons give off collaterals in different segments of the cord, thus ensuring that different muscle groups will be coordinated during postural control.
The coordinated actions of neurons that make up the LVST and provide postural stabilization are not completely understood. However, if a person begins tilting to the right, ipsilateral LVST fiberselicit extension of the left axial and limb muscu lature. Concurrently, right extensor muscles are inhibited. These actions stabilize the body’s centerof gravity and preserve upright posture.

Indirect pathways: medial-vestibulo-spinal tract
Medial Vestibulospinal Tract. These fibersoriginate primarily from the medial vestibularnucleus, although lesser projections arise from the inferior and lateral vestibular nuclei. Similar to LVST neurons, cells of the MVST receive input from vestibular receptors and the cerebellum, as well as somatosensoryinformation from the spinal cord. Fibers of the MVST descend bilaterally through the medial longitudinal fasciculus to terminate in the cervical spinal cord. These MVST fibers carry both excitatory and inhibitory signals, and they terminate on neck flexor and extensor motor neurons, as well as on proprio spinal neurons.
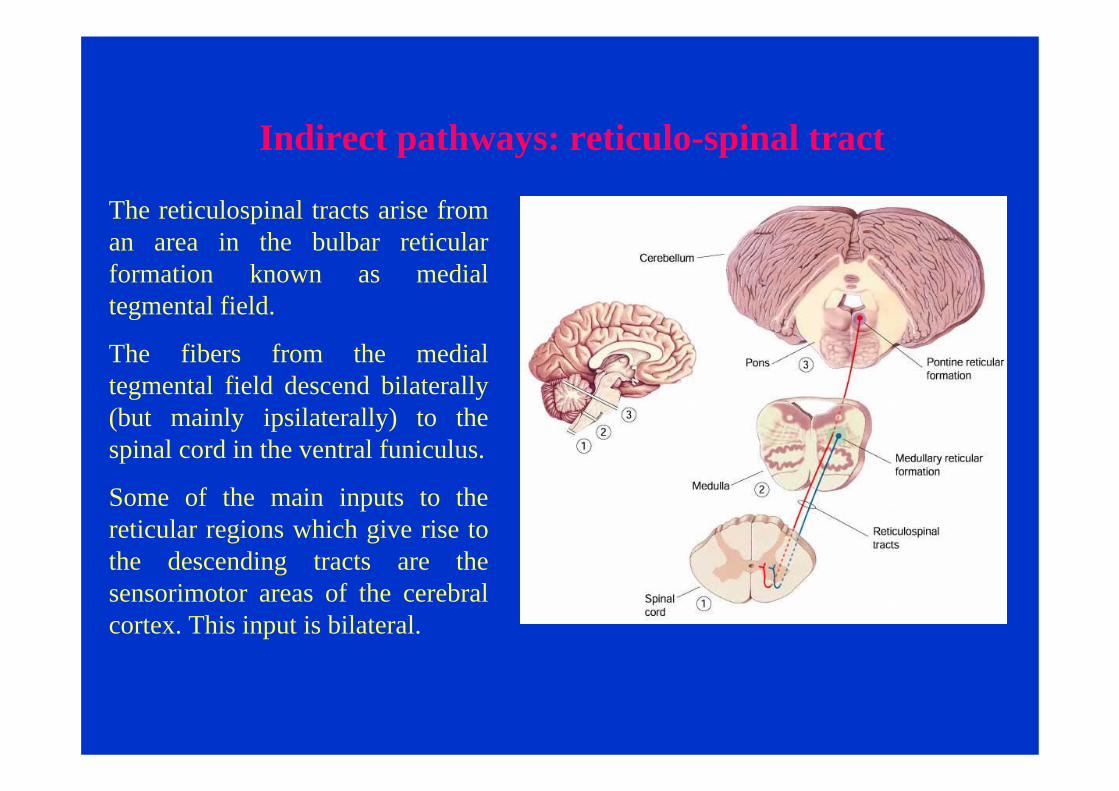
Indirect pathways: reticulo-spinal tract
The reticulospinal tracts arise from an area in the bulbar reticular formation known as medial tegmental field.
The fibers from the medial tegmental field descend bilaterally (but mainly ipsilaterally) to the spinal cord in the ventral funiculus.
Some of the main inputs to the reticular regions which give rise to the descending tracts are the sensorimotor areas of the cerebral cortex. This input is bilateral.

Indirect pathways: tecto-spinal tract
The tecto-spinal tract comes from cells in the deep layers of the superior colliculus. In the cord these fibres are found in the medial part of the ventral funiculus and travel only as far as cervical segments.
The main input to the superior colliculus in from visual cortical areas, and thus the tract may take part in head and neck orienting reactions to visual stimuli.

Motor maps
In the early twentieth century electrical stimulation of the primary motor cortex (area 4) was used to identify the specific motor effects of discrete sites in the frontal lobe. These experiments revealed an orderly arrangement along the precentral gyrus of control areas for the different body parts. This is the somatotopic orgsanization of the motor cortex.

Motor maps
Fingers, hands and face have disproportionately large representations in the motor areas of the cortex.
These parts of the body are used in tasks requiring the greatest precision and finest control from the cortex.

Motor maps
The motor cortex is not a switchboard where individual switches control individual muscles.
Studies performed with intracortical microstimulation(ICMS) showed a more complex organization.
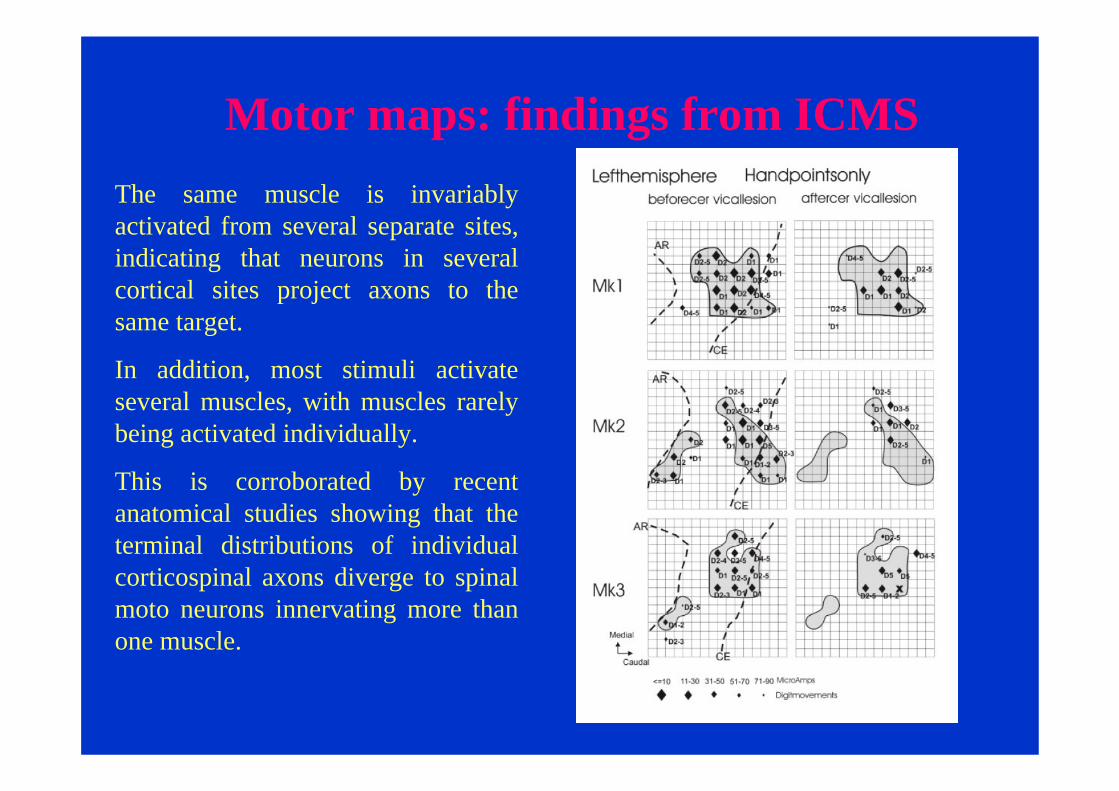
Motor maps: findings from ICMSThe same muscle is invariably activated from several separate sites, indicating that neurons in several cortical sites project axons to the same target.
In addition, most stimuli activate several muscles, with muscles rarely being activated individually.
This is corroborated by recent anatomical studies showing that the terminal distributions of individual corticospinal axons diverge to spinal moto neurons innervating more than one muscle.

Inputs to motor cortical areas
The primary motor cortex receives somatotopicallyorganized inputs from the primary somatosensory cortex (areas 1,2,and 3).
This means that neurons in the motor cortex have receptive fields in the periphery, receiving sensory inputs from the muscles to which they project.
Other sensory inputs come from the posterior parietal area 5, which is involved in integrating multiple sensory modalities for motor planning.

Inputs to motor cortical areas
The premotor areas receive major inputs from areas 5 and 7.
Each premotor area has its own pattern of inputs from distinct locations in areas 5 and 7.

Inputs to motor cortical areas
The premotor areas and the primary motor cortex also receive inputs from the basal ganglia and the cerebellum via different set of nuclei in the ventrolateral thalamus.

Premotor areas in planning movement
Although the outputs of the premotor areas and the primary motor cortex overlap in the spinal cord, the inputs to the premotor areas are quite different from those to the primary motor cortex.
Moreover, damage to premotor areas causes more complex motor impairments than does damage to primary motor cortex.
When a monkey with a large lesion of the premotor area is presented with food behind a transparent shield it will reach directly for the food and bump into the shield. Unlike a normal animal it is unable to incorporate visuospatial information about the shield into the kinematic plan for moving its hand.

Premotor areas in planning movement
The idea that premotorareas are involved in planning movements has received crucial support during the past 20 years from physiological, clinical and imaging studies of humans performing a variety of special tasks.
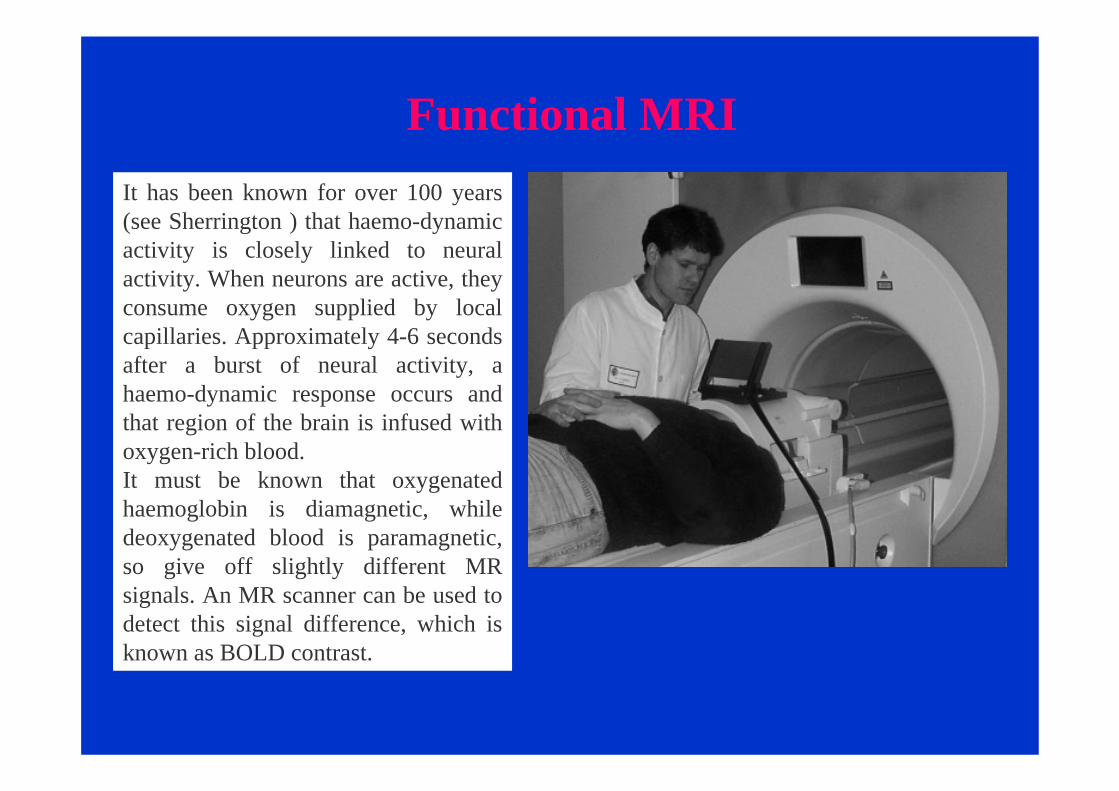
Functional MRIIt has been known for over 100 years (see Sherrington ) that haemo-dynamic activity is closely linked to neural activity. When neurons are active, they consume oxygen supplied by local capillaries. Approximately 4-6 seconds after a burst of neural activity, a haemo-dynamic response occurs and that region of the brain is infused with oxygen-rich blood. It must be known that oxygenated haemoglobin is diamagnetic, while deoxygenated blood is paramagnetic, so give off slightly different MR signals. An MR scanner can be used to detect this signal difference, which is known as BOLD contrast.
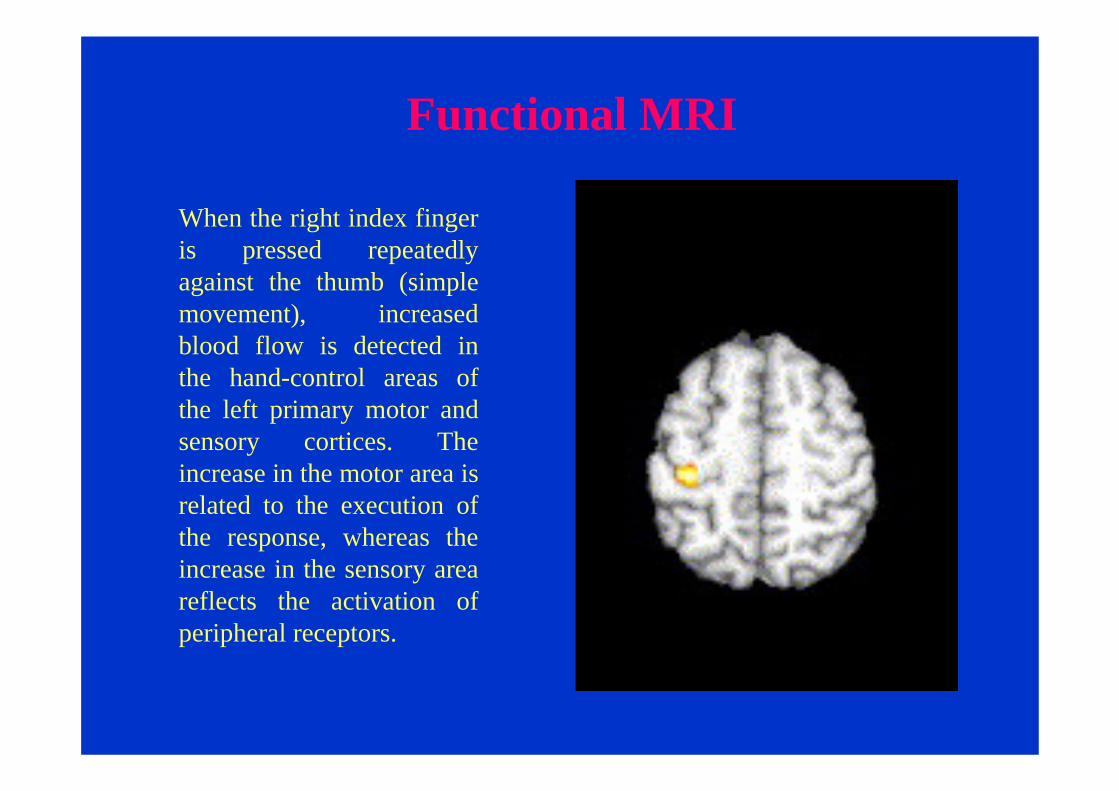
Functional MRI
When the right index finger is pressed repeatedly against the thumb (simple movement), increased blood flow is detected in the hand-control areas of the left primary motor and sensory cortices. The increase in the motor area is related to the execution of the response, whereas the increase in the sensory area reflects the activation of peripheral receptors.

Functional MRI
During a complex sequence of finger movements the increase in blood flow extends to the premotorareas, in particular into the supplementary motor area.
Complex movement sequences require more planning than do simple repetitive movements.

Functional MRI
Selection of a movement can be the result of internal reflection, which may involve evocation of mental imagery: self-paced movements.
More often, however, movements are responses to sensory cues (e.g., visual or auditory stimuli): external-triggered movements.
Self-paced
External-triggered

Functional MRI
Dick J. Veltman et al., 2003
Functional MRI during a movement triggered by a visual cue. Note the activation of lateral premotor cortex together with the visual areas in the occipital lobe

Transcranial magnetic stimulation
In the past decade it has become possible to stimulate motor cortical areas in alert humans by inducing electrical fields in the brain using a rapidly alternating magnetic fields produced by wire coils applied to the scalp.

Stimulating Stimulating the the BrainBrain !!!!
TMSTMS Barker et Barker et al., 1985al., 1985
NerveNerve
BrainBrain

TMS: how does it workTMS is based on the principle of electromagnetic induction described by Michael Faraday in 1831. This technique uses the rapid, time-varying, magnetic field provoked by a current pulse in a coil of wires. The magnetic field is unattenuated by the scalp and the skull and, with minimal or no activation of pain receptors, induces brief intracranial electrical currents that are in the same plane but largely opposite in direction to the current in the coil (Trompetto and Abbuzzese, 2002).
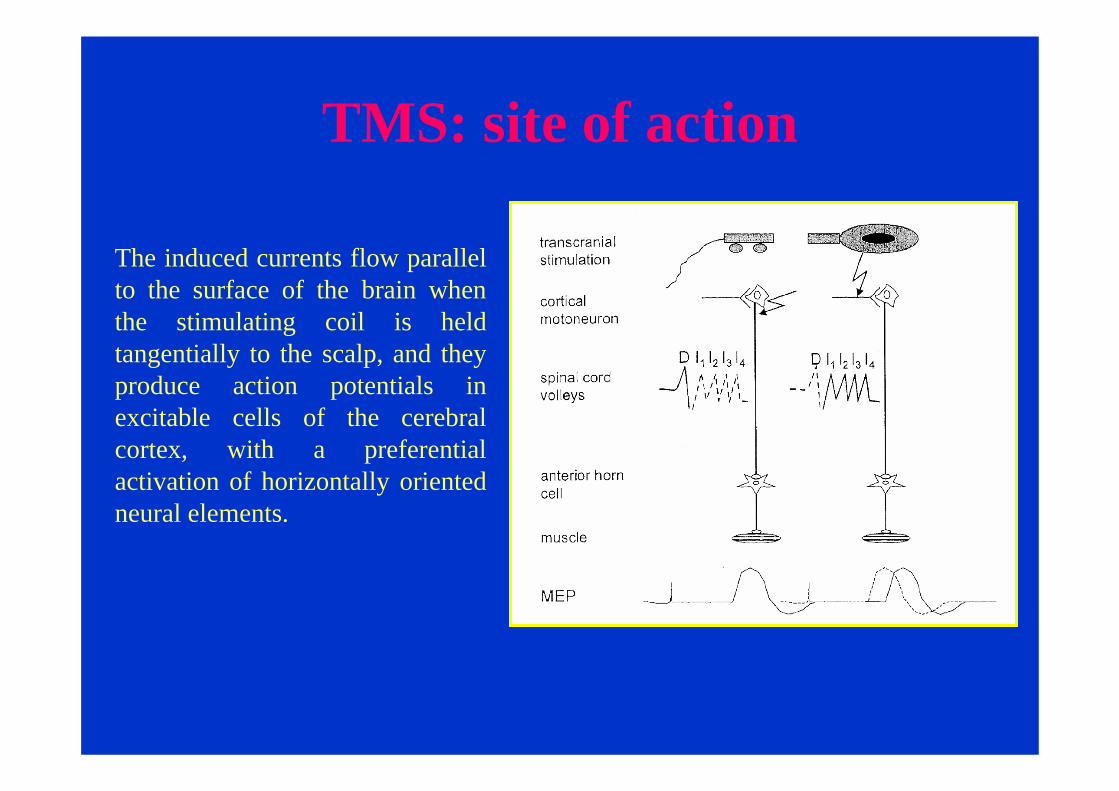
TMS: site of action
The induced currents flow parallel to the surface of the brain when the stimulating coil is held tangentially to the scalp, and they produce action potentials in excitable cells of the cerebral cortex, with a preferential activation of horizontally oriented neural elements.

TMS: how does it workThe first generation of stimulating coils were large, round, nonfocal coils, but nowadays figure-of-eight coils are available that allow a more focused stimulation of the brain with a spatial resolution of approximately 0.5 cm.
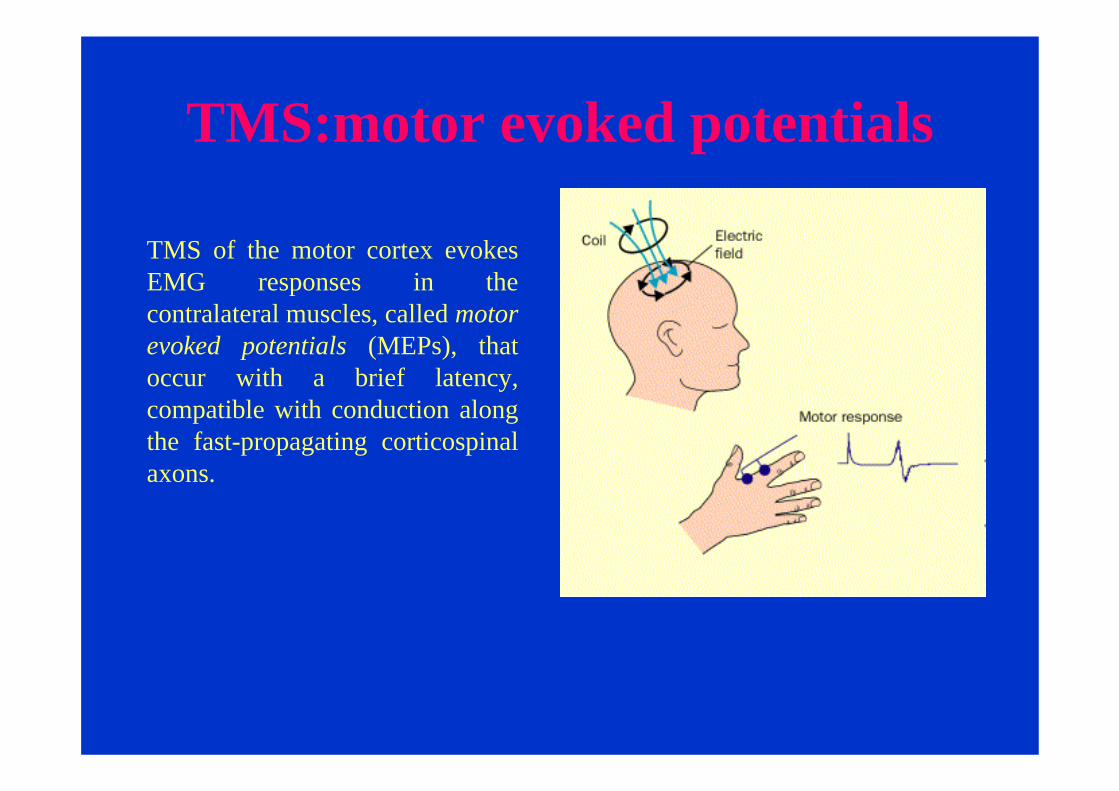
TMS:motor evoked potentials
TMS of the motor cortex evokes EMG responses in the contralateral muscles, called motor evoked potentials (MEPs), that occur with a brief latency, compatible with conduction along the fast-propagating corticospinalaxons.

TMS: voluntary facilitation
Trompetto et al., 2004
Voluntary contraction of the target muscle increases the MEP size. A period of EMG silence follows the MEP, which is called silent period.

TMS:motor mapsTMS delivered by focal coils can be used for noninvasive and painless mapping of the somatotopicrepresentation of muscles within the motor cortex. Cortical maps are constructed by stimulating different points on the scalp at a constant intensity and analyzing the number of sites from which MEPs can be elicited in the target muscle. These sites outline the motor area extension. Several types of data can be obtained with this technique—first, the location of the optimal position for stimulation (i.e., the scalp site from which MEPs of maximal amplitude and minimal latency can be elicited). These “hot spots”represent low threshold areas where the corticospinal cells impinging onto spinal motoneurons of the target muscle are mostly concentrated From Capaday et al., 2006

TMS:motor maps
Although cortical maps for intrinsic hand muscles are large and to some extent overlapping, generally the cortical representations of different muscles are clearly separate (Wilson et al., 1995) and confirm the traditional lateromedial sequence of face, arm, and leg muscles (Amassian et al., 1995).

TMS:motor maps
TMS studies showed that the somatotopic organization of the motor cortex is not fixed but can be altered following injury and during motor learning
For example, cortical motor representations of the muscles proximal to the stump have been found to be enlarged after limb amputation (Cohen et al., 1991;Fuhr et al., 1992). Similar changes are obtainable after ischemia (Brasil–Neto et al., 1993) and even during learning of new motor skills (Pascual–Leone et al., 1994a).

TMS:motor mapsStimulation of the cortical area previously devoted to the missing limb using TMS produces movements or muscle contractions of parts of the body that are represented in the cortical somatotopic map adjacent to the missing body part. For example, in the case of an above-elbow amputee, stimulating the presumed hand region in the motor cortex will produce muscle contractions in stump muscles such as the biceps and triceps and/or muscle contractions in the face This has been interpreted as an invasion of the armrepresentation into a region of the cortex that previously controlled the now-amputated hand (Roricht et al., 1999 ; Irlbacher et al., 2002 ).
Deltoid motor maps in a 21-year-old patient with a left upper extremity amputation at age 7. TMS revealed an enlarged cortical motor output area of the deltoid muscle at the amputation stump

TMS:motor maps
TMS have demonstrated that motor skill training induces changes in the organization of movement representations in the primary motor cortex in the form of expansion and increased excitability of the cortical representation of specific muscles involved in the tasks (Pascual-Leone et al.).
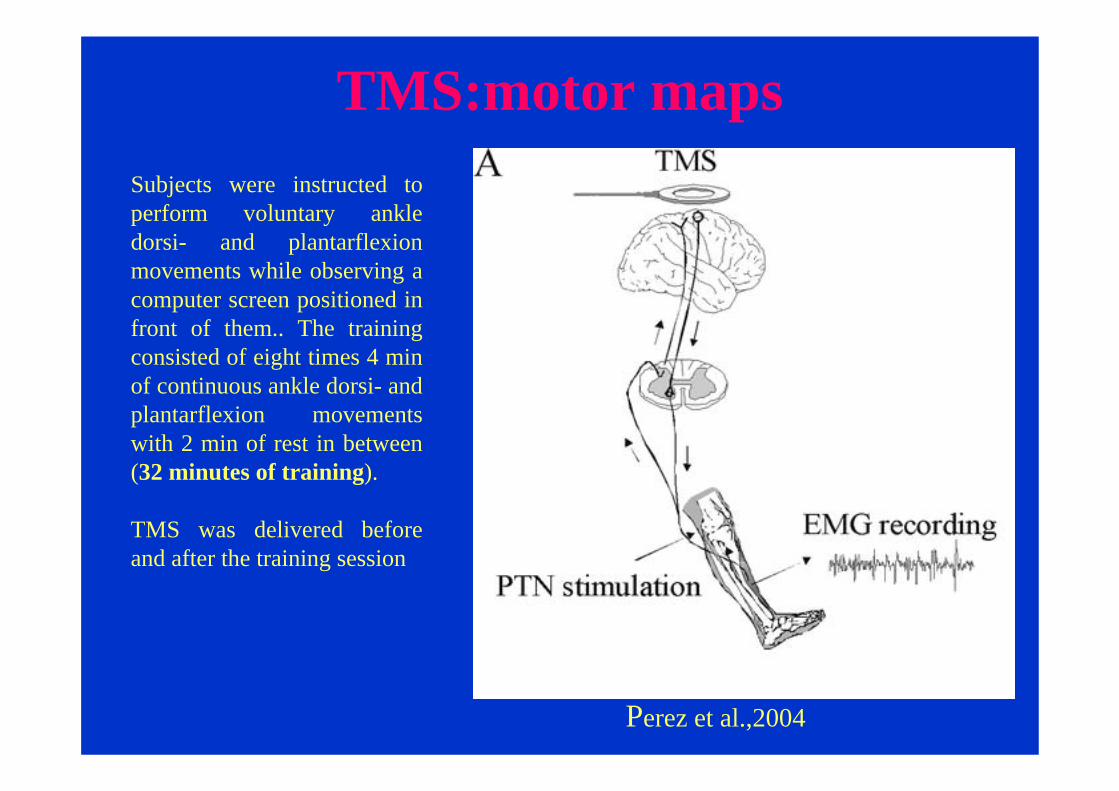
TMS:motor maps
Perez et al.,2004
Subjects were instructed to perform voluntary ankle dorsi- and plantarflexionmovements while observing a computer screen positioned in front of them.. The training consisted of eight times 4 min of continuous ankle dorsi- and plantarflexion movements with 2 min of rest in between(32 minutes of training).
TMS was delivered before and after the training session

TMS:motor maps
In this study, it has been showed that 32 min of motor skill training involving the ankle muscles increased MEPs elicited by TMS in Tibialis Anterior muscle(Perez et al., 2004).

Cortical organization of voluntary movement
The somatotopic organization of the motor cortex is not fixed but plastic.
The underlying neural mechanisms that accompany changes in cortical organization after motor skill training or injuries are unclear. Pharmacological studies have revealed that training-induced changes are related to a decreased GABAergic inhibition within intracortical circuits (Butefisch et al. 2000; Lech et al. 2001). In humans, this GABAergicinhibitory system can be tested using a paired-pulse TMS technique (Kujiraiet al. 1993), in which a conditioning TMS pulse below the threshold for eliciting a motor evoked potential (MEP) inhibits a suprathreshold test stimulus at short intervals (1–5 ms).

Paired-Pulse TMS investigate intracortical GABA-ergic circuits
• the intensity of both stimuli
• the interval between them (inter-stimulus interval, ISI)
• the presence (or absence) of voluntary muscle contraction
conditioning test

Paired-pulse testing with sub-threshold conditioning shocks: SICI
Kujirai, 1993

CSN
I3 I3 I3
I1I1
Level at which SICI occurs
I4I4
Inhibition acts on the motor cortex upstream to the cortico-spinal cell

Cortical organization of voluntary movement
•The observed decrease in SICI indicated that one of the mechanisms explaining the increased TA MEPs may be modulation of local intracortical circuits.•(Perez et al., 2004).
•SICI can be modulated by sensory stimulation
Perez et al.,2004

Cortical organization of voluntary movement
•The somatotopic organization of the motor cortex is not fixed but can change following injuries or after motor training.
•The somatotopic organization of the motor cortex is plastic.
•This plasticity is the theoretical basis for all the rehabilitation programs after injuriesof the nervous system.

Cerebellum and voluntary movement
The cerebellum is an important part of the motor system. It is composed of an outer mantle of gray matter (the cerebellar cortex), internal white matter and three pairs of deep nuclei: the fastigial, the interposed and the dentate. The cerebellum is connected to the dorsal aspect of the brain stem by three symmetrical pairs of tracts: the inferior, the middle and the superior cerebellarpeduncle.

Cerebellum and voluntary movementTwo longitudinal furrows distinguish three mediolateralregions that are important functionally. The furrows define an elevated ridge in the midline known as the vermis. On either side of the vermis are the cerebellar hemispheres, each of one is divided into intermediate and lateral regions. The three medio-lateral regions of the cerebellum (the vermis and intermediate and lateral parts of the hemispheres) receive different afferent inputs, project to different parts of the motor systems, and represent distinct functional subdivisions.

Cerebellum and voluntary movement
•The vermis ("worm"), located on the midline, receives visual, auditory and vestibular input as well as somatic sensory input from the head and proximal parts of the body. It projects by way of the fastigial nucleus to cortical and brain stem regions that give rise to the medial descending system that control proximal muscles of the body and limbs.
•The vermis governs posture and locomotion as well as gaze.

Cerebellum and voluntary movement•The intermediate zone projects via the thalamus to the primary motor cortex, thus controlling the lateral corticospinalsystem.
•This zone receives input from the corticopontocerebellar fibers that originate from the motor cortex. These fibers carry a duplicate of the information that was sent from the motor cortex to the spine in order to effect a movement. The intermediate zone also receives sensory feedback from the muscles.
•These two streams of information are integrated by this region, allowing for the feedback comparison of what the muscles are supposed to be doing with what they are actually doing.

Cerebellum and voluntary movement
The lateral parts of the hemispheres, which are phylogenetically more recent, are much larger in humans than in monkeys or cats. This region receives input exclusively from the cerebral cortex and it is thus called the cerebro-cerebellum. Its output is mediated by the dentate nucleus, which projects to motor, premotorand prefrontal cortices. Recent imaging data indicate that the cerebrocerebllum is involved in planning and mental rehearsal of complex motor actions.

Cerebellum and voluntary movement
Functional imaging of changes in cerebellar activity related to learning during a novel eye–hand tracking taskR. C. Miall1, 2, 2 and E. W. Jenkinson2
The lateral cerebellum appears to be important for learning both motor and cognitive tasks in which skilled responses are developed through repeated practice
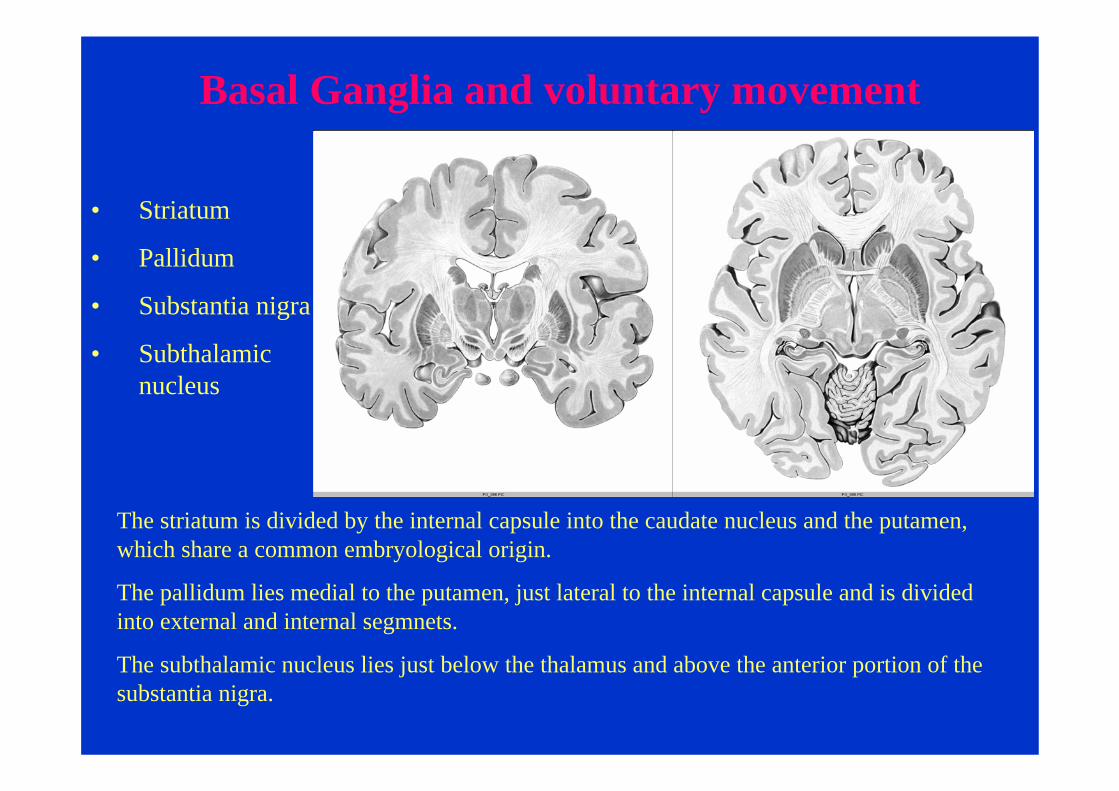
Basal Ganglia and voluntary movement
• Striatum
• Pallidum
• Substantia nigra
• Subthalamic nucleus
The striatum is divided by the internal capsule into the caudate nucleus and the putamen, which share a common embryological origin.
The pallidum lies medial to the putamen, just lateral to the internal capsule and is divided into external and internal segmnets.
The subthalamic nucleus lies just below the thalamus and above the anterior portion of the substantia nigra.

Basal Ganglia and voluntary movement
• Striatum
• Pallidum
• Substantia nigra
• Subthalamic nucleus
The substantia nigra lies in the midbrain on the medial side of the internal capsule.

Basal Ganglia and voluntary movement
• Striatum
• Pallidum
• Substantia nigra
• Subthalamic nucleus
The substantia nigra is formed by a reticular portion (pars reticulata) and a compact zone (pars compacta), which lies dorsal to the pars reticulata. The cells of the pars compacta are dopaminergic and contain neuromelanin, a dark pigment derived from polymerizeddopamine. Neuromelanin accounts for the dark discoloration of this structure. The cellbodies in the pars reticulata of the substantia nigra are similar to those forming the internal segment of the pallidum. The internal pallidal segment is related functionally to the pars reticulata of the substantia nigra.

Basal Ganglia and voluntary movement
The basal ganglia receive major input from the cerebral cortex and send their output back to the cortex (via the thalamus) and to the brain stem. Thus, the basal ganglia are major components of large cortical-subcortical reentrant circuits linking cortex and thalamus
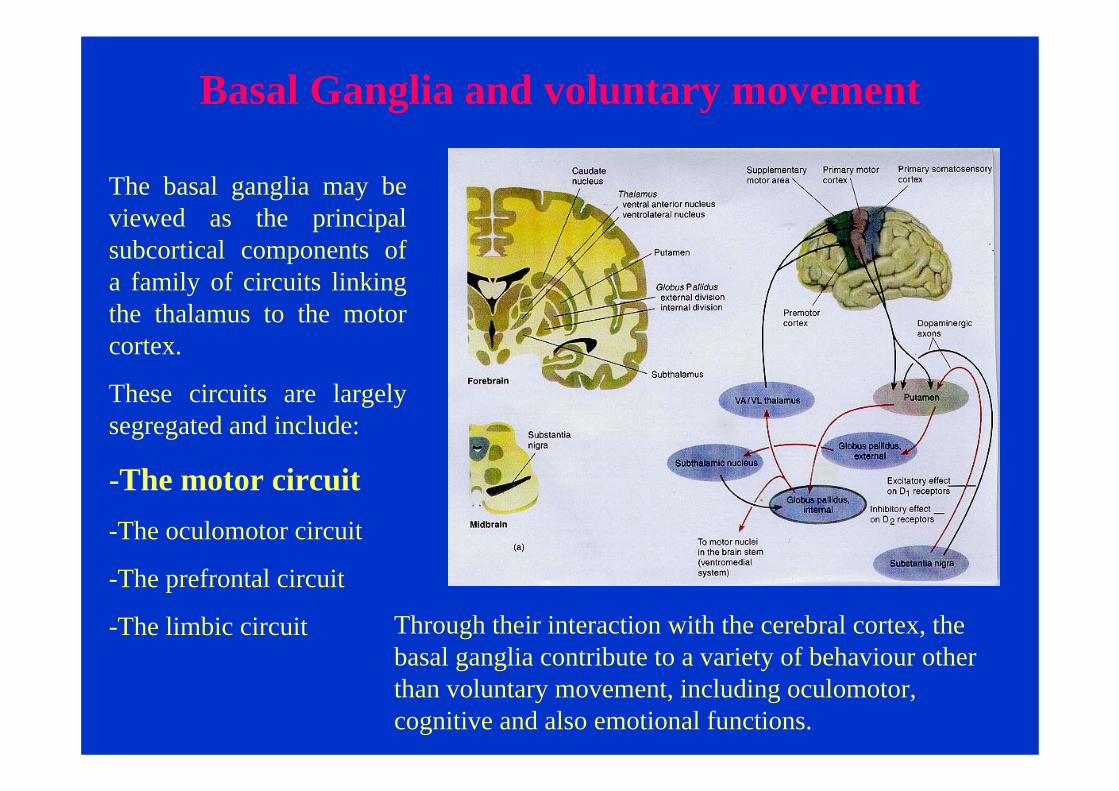
Basal Ganglia and voluntary movement
The basal ganglia may be viewed as the principal subcortical components of a family of circuits linking the thalamus to the motor cortex.
These circuits are largely segregated and include:
-The motor circuit-The oculomotor circuit
-The prefrontal circuit
-The limbic circuit Through their interaction with the cerebral cortex, the basal ganglia contribute to a variety of behaviour other than voluntary movement, including oculomotor, cognitive and also emotional functions.

The striatum in the major receiver of inputs to the basal ganglia. These input come from the cerebral cortex(motor and sensory areas).
The neurons of the striatum project to the globus pallidusand substantia nigra pars reticulata. Together these two nuclei give rise to the major output projections from the basal ganglia.
Basal Ganglia and voluntary movement
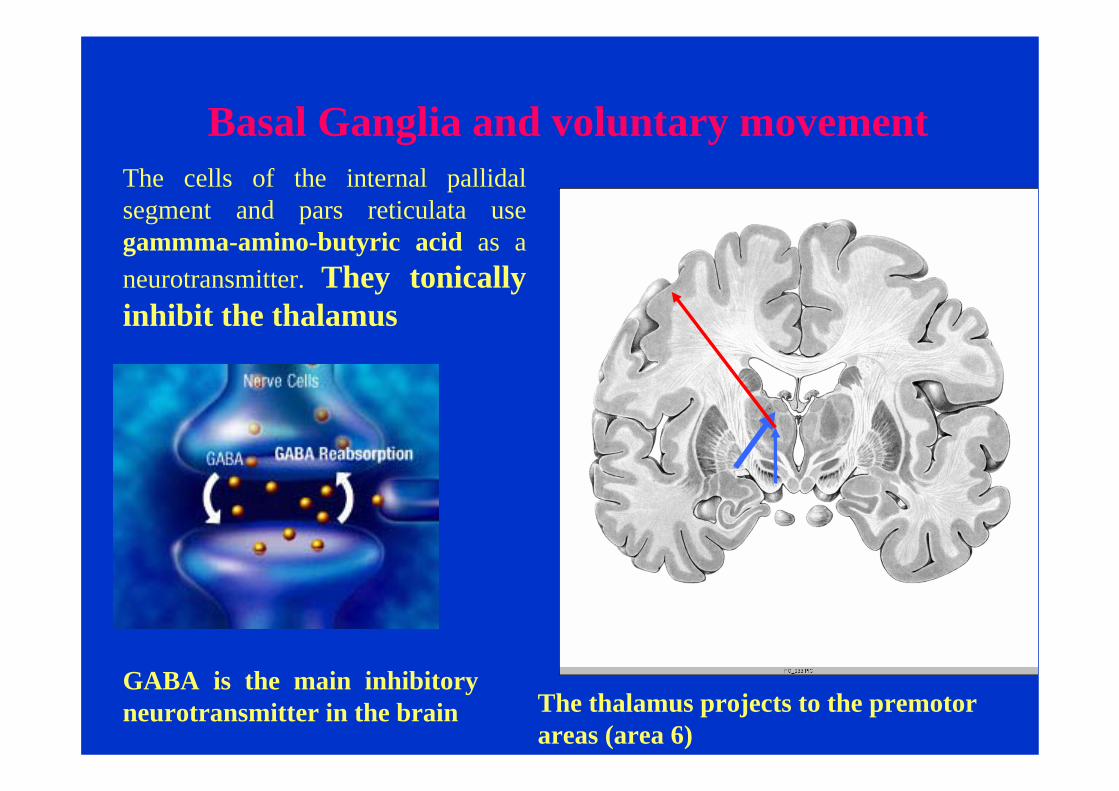
The cells of the internal pallidalsegment and pars reticulata use gammma-amino-butyric acid as a neurotransmitter. They tonically inhibit the thalamus
Basal Ganglia and voluntary movement
GABA is the main inhibitory neurotransmitter in the brain The thalamus projects to the premotor
areas (area 6)

The only excitatory projections of the basal ganglia are the glutamatergicic cells of the sub-thalamic nucleus.
Basal Ganglia and voluntary movement
Glutamate is the main excitatory neurotransmitter in the brain

Also the cells in the putamen and caudate use gammma-amino-butyric acid as a neurotransmitter.
Basal Ganglia and voluntary movement
GABA is the main inhibitory neurotransmitter in the brain

The two output nuclei of the basal ganglia are modulated by two parallel pathways that run from the striatum: the direct pathway and the indirect pathway.
Basal Ganglia and voluntary movement

The two output nuclei of the basal ganglia are modulated by two parallel pathways that run from the striatum: the direct pathway and the indirect pathway.
The direct pathway inhibits the output nuclei, thus activating the thalamus.
The indirect pathwayactivates the output nuclei, thus suppressing the firing of the thalamus.
Basal Ganglia and voluntary movement

Basal Ganglia and voluntary movement
GABA is the main inhibitory neurotransmitter in the brain
The neurons in the two output nuclei discharge tonically at high frequency.

Basal Ganglia and voluntary movement
When phasic excitatory inputs from the sensory-motor areas of the cortex transiently activate the direct pathway from the striatum to the pallidum, the tonicallyactive neurons in the pallidumare briefly suppressed, thus permitting the thalamus and ultimately the cortex to be activated.
The direct pathway activate the motor areas of the cortex and promote the movement
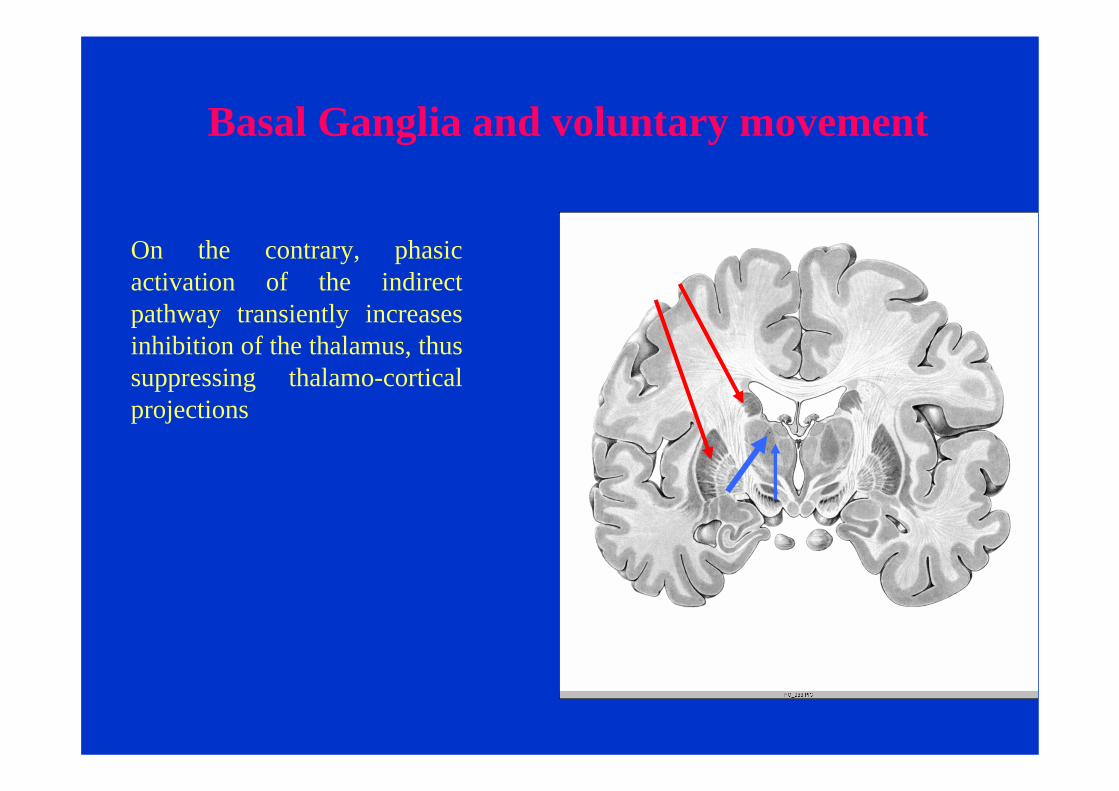
Basal Ganglia and voluntary movement
On the contrary, phasic activation of the indirect pathway transiently increases inhibition of the thalamus, thus suppressing thalamo-cortical projections

Activation of the direct pathway facilitates movement, whereas activation of the indirect pathway inhibits movement.
Basal Ganglia and voluntary movement

Basal Ganglia and voluntary movement
The dopaminergic neurons in the pars compacta of the substantia nigra project to the striatum.

GABA
EnkephalinGABA
Substantia P
D1 D2
D 1 + D 2 -
Basal Ganglia and voluntary movement
External segmentInternal segment
Substantia nigra
Direct pathway
Indirect
pathway
The two striatal output pathways are affected differently by the dopaminergic projection from the pars compacta. Striatal neurons that project directly to the two output nuclei have D1 dopamine receptors that facilitate transmission, while those that project indirectly have D2 receptors that reduce transmission.
Thus dopaminergic inputs reduce inhibition of the thalamo-cortical projections facilitating movements initiated in the cortex.

Parkinson Disease